ELABORACION DE QUESO MOZZARELLA CON DIFERENTES PORCENTAJES DE GRASA EN LA LECHE DE VACA

Relative oxygenation of the Tithonian–Valanginian
Vaca Muerta–Chachao formations of the Mendoza Shelf,
Neuquen Basin, Argentina
P. DOYLE1, D. G POIRE2, L. A. SPALLETTI2, D. PIRRIE3,
P. BRENCHLEY4 & S. D. MATHEOS2
1Department of Earth Sciences, University College London, Gower Street,
London WC1E 6BT, UK2Centro de Investigaciones Geologicas, Universidad Nacional de La Plata-Conicet,
Calle 1 No. 644, 1900 La Plata, Argentina3Camborne School of Mines, University of Exeter, Redruth, Cornwall TR15 3SE, UK
4Department of Earth Sciences, University of Liverpool, 4 Brownlow Street,
Liverpool L69 3GP, UK
Abstract: Organic-rich sediments were deposited in the deeper sectors of the NeuquenBasin during the latest Jurassic and the Early Cretaceous. This paper presents the resultsof a detailed examination of these deposits in the northern-most extension of the basin, inthe Mendoza Province, and explores their wider significance for palaeo-oxygenation studies.
The Tithonian–Berriasian Vaca Muerta Formation, the primary source rock for theNeuquen Basin, comprises bituminous shales and interbedded limestones depositedduring a major transgression. In the Valanginian, the beginning of a regressive phase enabledthe development of shallow-marine carbonates to form the base of the Chachao Formation,which eventually led to extensive biohermal carbonates of the uppermost Chachao For-mation. Along the length of the narrow N–S-trending Mendoza Shelf of the NeuquenBasin both units are well exposed, permitting detailed study of the stratigraphy, sedimentol-ogy, ichnology and palaeoecology. The analysis of the Tithonian–Valanginian succession inthe Salado river valley shows that carbonate production increased up-section. Faunal associ-ations are mostly limited to poorly diverse epibenthos and pseudoplankton in the lower partof the section (Vaca Muerta Formation), with increased diversity in the lower Chachaosection, including shallow and deeper infaunal bivalves. A background level of laminatedshales to Chondrites bioturbation is typical of anoxic–suboxic conditions. Micritic lime-stones and carbonate sandstones throughout the section commonly show the developmentof Thalassinoides suevicus.
Relative oxygenation curves based on trace fossils and body fossils were developed andcompared. There was a primary substrate control on trace fossil diversity and occurrence,with a primary oxygenation signal provided by body fossil evidence. Interpretation of thepalaeo-oxygenation on the basis of trace fossil taxa alone, however, would lead to inaccurateresults. This study, therefore, demonstrates the importance of integrated trace and bodyfossil analysis in the fuller understanding of black shales.
Palaeoenvironmental analysis of oxygen-deficient basins has been the subject of manydetailed studies over the last 25 years, fuelledin part by the development of trace fossilmodels of relative palaeo-oxygenation followingthe initial seminal work by Rhoads & Morse(1971). In particular, Savrda & Bottjer (1986,1987), Ekdale & Mason (1988), Allison et al.(1995) and Savrda (1995) have developed andsynthesized models in which trace fossil diver-sity and burrow diameter are used as a proxy
for relative oxygen levels within specificbasins, with a direct relationship usually beingrecorded between increased diversity patterns/burrow diameter and increasing levels of dis-solved oxygen. This relationship has been testedin a number of ancient sedimentary basins follow-ing the development of the original Rhoads &Morse model, but little critical appraisal hasbeen made of it. This is particularly true for theupper, oxygenated conditions, where the occur-rence of wide burrow-diameter Thalassinoides
From: VEIGA, G. D., SPALLETTI, L. A., HOWELL, J. A. & SCHWARZ, E. (eds) 2005. The Neuquen Basin, Argentina: A CaseStudy in Sequence Stratigraphy and Basin Dynamics. Geological Society, London, Special Publications, 252, 185–206.0305-8719/05/$15.00 # The Geological Society of London 2005.

and other diagnostic traces have been recognizedas important indicators of increased oxygenlevels. Other studies have concentrated on analy-sis of the diversity of body fossil macrobenthosin low oxygen environments (e.g. Duff 1974;Wignall 1990; Etter 1995). Together, body andtrace fossil models have provided a usefulguide to dissolved oxygen levels in specificbasins.
Recent studies have shown that the diversityresponse is more complex and difficult to deter-mine solely with reference to oxygenation (e.g.Sageman & Bina 1997). Certain signals, suchas the common development of opportunisticspecies and the adoption by benthos of pseudo-planktonic habit, are common in black shales(e.g. Wignall & Simms 1990; Doyle &Whitham 1991; Wignall 1993; Etter 1996).Such palaeoecological responses may reflect aprimary low oxygen signal, as has been supposedin many studies. However, they may also indi-cate factors such as environmental instabilityand the presence of soupy substrates (Wignall1993; Sageman & Bina 1997) or the capabilityof mass flow and other environmental factors inintroducing oxygen-demanding trace makers into otherwise anoxic basins (e.g. Follmi &Grimm 1990).
The present study describes for the first timethe palaeoenvironments of an importantoxygen-deficient stratigraphic interval, the VacaMuerta–lowermost Chachao formations ofTithonian–Valanginian age, in the MendozaShelf of the Neuquen Basin (Mendoza Provinceof Argentina) (Fig. 1). The Vaca Muerta For-mation in particular is considered to be theprimary source rock for the important Neuquenoilfield in Argentina, and yet it has not, to date,received extensive study in this field. Previousworks on the palaeontology and stratigraphy ofthe Vaca Muerta Formation have been confinedto the stratigraphy, aspects of particular macroin-vertebrate and vertebrate fossil groups, anddetails of the sedimentology (e.g. Leanza et al.1977; Leanza 1981; Leanza & Zeiss 1990;Gasparini et al. 1997, 1999, 2002; Spallettiet al. 1999). The Chachao Formation, andparticularly its oxygen-deficient lower part, hasreceived similarly scant attention (Mombruet al. 1978; Legarreta et al. 1981; Legarreta &Kozlowski 1981). The aim of this paper is todescribe in detail the palaeoenvironments of theVaca Muerta and lower Chachao formationsfrom an examination of trace fossils andbenthic macropalaeontology, and to comparethe oxygenation signals provided by both datasources in order to test the veracity of the tracefossil oxygenation model.
Late Jurassic–Early Cretaceous
palaeogeography, stratigraphy and
sequence stratigraphy of the
Neuquen Basin
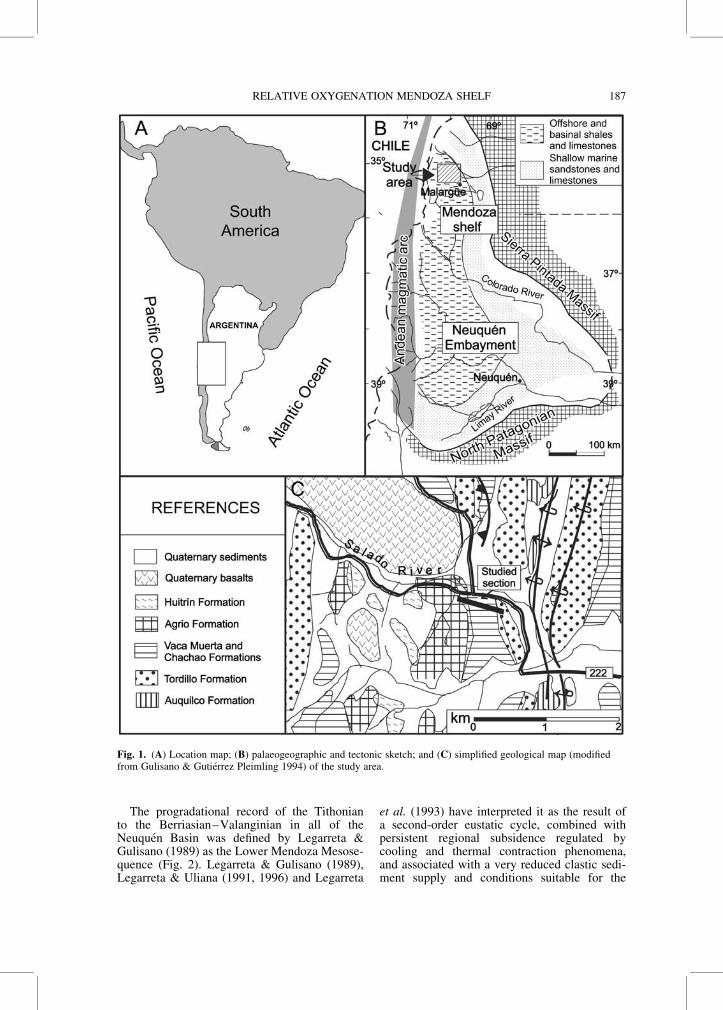
Two main depositional areas can be recognizedin the Neuquen Basin (Fig. 1): the NeuquenEmbayment (Bracaccini 1970) to the south, andthe narrow Mendoza Shelf (where the studiedsection is located) to the north. The NeuquenEmbayment was a wide subcircular bay or gulfthat developed behind the Andean magmaticarc. During Jurassic and Early Cretaceous timesthis region was limited by emergent areas tothe west, south and east; from all these pointsand to the interior of the depocentre, smoothdepositional ramp topographies converged. Thedeepest area of the basin was limited to anelongated strip whose greater extent was orientedNW–SE, oblique to the Andean magmatic arc,which extended north–south (Spalletti et al.2000). This obliquity between the arc andthe axis of the depocentre favoured thedevelopment of the Mendoza shelf to the north(Fig. 1). As shown previously by Mombruet al. (1978), Legarreta et al. (1993) andSpalletti et al. (2000), this area was charac-terized by a significant reduction of the ampli-tude of the basin, which in latitudes lower than348S was limited to a thin strip located in thesurroundings of the Jurassic–Cretaceousmagmatic arc.
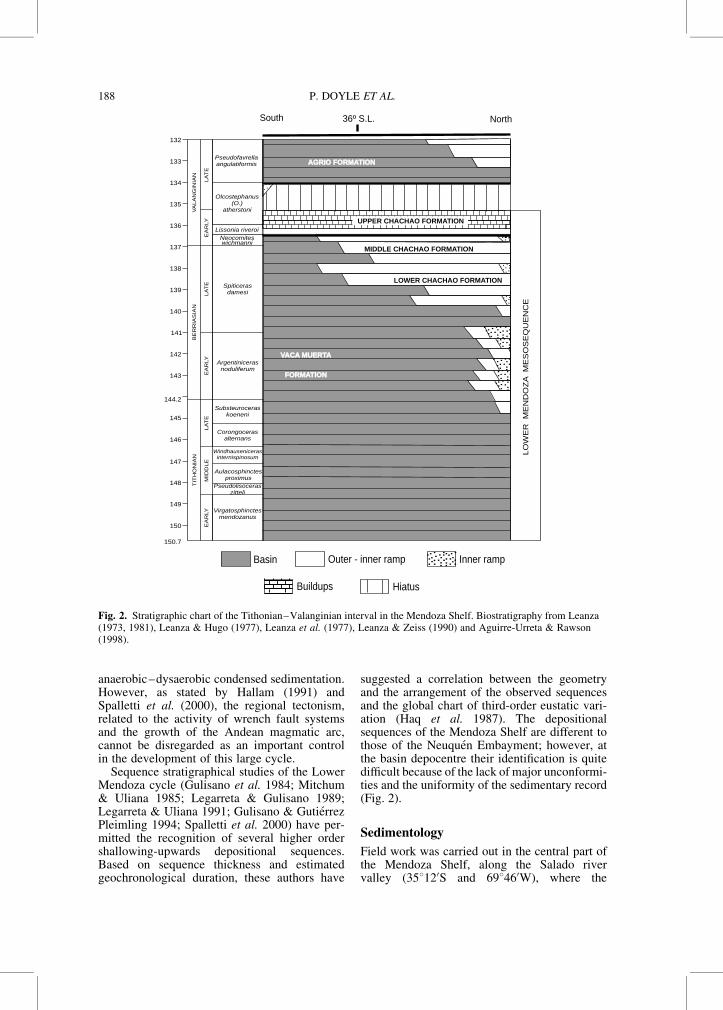
In most of the Neuquen Basin, the UpperJurassic–Lower Cretaceous record is characte-rized by the dark bituminous shales and marlsof the Vaca Muerta Formation (Weaver 1931).This unit concordantly overlies the clastic andcontinental deposits of the Tordillo Formation.The Tordillo–Vaca Muerta contact (Early Titho-nian) is an isochronous surface throughout thebasin and marks the begining of the marineTithonian transgression (Leanza 1981). The topof the Vaca Muerta Formation is diachronous(Leanza 1973, 1981; Leanza & Hugo 1977;Leanza et al. 1977) and progradational (Gulisanoet al. 1984; Mitchum & Uliana 1985; Legarreta& Gulisano 1989). It includes younger stageswhen passing from the southern (MiddleTithonian) to the central sector of the basin(Berriasian–Valanginian). In the MendozaShelf, the Vaca Muerta Formation is conform-ably overlain by the marine carbonates andshales of the Valanginian Chachao Formation(Fig. 2). The stratigraphy of these two units hasbeen summarized by Riccardi (1983, 1988),Legarreta et al. (1993) and Gulisano & GutierrezPleimling (1994).
P. DOYLE ET AL.186
The progradational record of the Tithonianto the Berriasian–Valanginian in all of theNeuquen Basin was defined by Legarreta &Gulisano (1989) as the Lower Mendoza Mesose-quence (Fig. 2). Legarreta & Gulisano (1989),Legarreta & Uliana (1991, 1996) and Legarreta
et al. (1993) have interpreted it as the result ofa second-order eustatic cycle, combined withpersistent regional subsidence regulated bycooling and thermal contraction phenomena,and associated with a very reduced clastic sedi-ment supply and conditions suitable for the
Fig. 1. (A) Location map; (B) palaeogeographic and tectonic sketch; and (C) simplified geological map (modifiedfrom Gulisano & Gutierrez Pleimling 1994) of the study area.
RELATIVE OXYGENATION MENDOZA SHELF 187
anaerobic–dysaerobic condensed sedimentation.However, as stated by Hallam (1991) andSpalletti et al. (2000), the regional tectonism,related to the activity of wrench fault systemsand the growth of the Andean magmatic arc,cannot be disregarded as an important controlin the development of this large cycle.
Sequence stratigraphical studies of the LowerMendoza cycle (Gulisano et al. 1984; Mitchum& Uliana 1985; Legarreta & Gulisano 1989;Legarreta & Uliana 1991; Gulisano & GutierrezPleimling 1994; Spalletti et al. 2000) have per-mitted the recognition of several higher ordershallowing-upwards depositional sequences.Based on sequence thickness and estimatedgeochronological duration, these authors have
suggested a correlation between the geometryand the arrangement of the observed sequencesand the global chart of third-order eustatic vari-ation (Haq et al. 1987). The depositionalsequences of the Mendoza Shelf are different tothose of the Neuquen Embayment; however, atthe basin depocentre their identification is quitedifficult because of the lack of major unconformi-ties and the uniformity of the sedimentary record(Fig. 2).
Sedimentology
Field work was carried out in the central part ofthe Mendoza Shelf, along the Salado rivervalley (358120S and 698460W), where the
36º S.L.
Basin Outer - inner ramp Inner ramp
Buildups Hiatus
UPPER CHACHAO FORMATION136
141
149
148
137
142
150
150.7
143
144.2
135
140
147
134
139
146
133
138
145
132
Olcostephanus(O.)
atherstoni
Pseudofavrellaangulatiformis
Neocomiteswichmanni
Lissonia riveroi
Spiticerasdamesi
Argentinicerasnoduliferum
Substeuroceraskoeneni
Corongocerasalternans
Windhausenicerasinternispinosum
Aulacosphinctesproximus
Pseudolisoceraszitteli
Virgatosphinctesmendozanus
EA
RLY
MID
DL
EL
AT
EL
AT
EL
AT
E
TIT
HO
NIA
NB
ER
RIA
SIA
NV
AL
AN
GIN
IAN
EA
RLY
EA
RLY
AGRIO FORMATIONAGRIO FORMATION
LOWER CHACHAO FORMATION
VACA MUERTA
FORMATION
VACA MUERTA
FORMATION
NorthSouth
MIDDLE CHACHAO FORMATION
LO
WE
RM
EN
DO
ZA
ME
SO
SE
QU
EN
CE
Fig. 2. Stratigraphic chart of the Tithonian–Valanginian interval in the Mendoza Shelf. Biostratigraphy from Leanza(1973, 1981), Leanza & Hugo (1977), Leanza et al. (1977), Leanza & Zeiss (1990) and Aguirre-Urreta & Rawson(1998).
P. DOYLE ET AL.188

Tithonian–Valanginian succession (170 m) ispart of a large anticlinal structure oriented N–Sand accompanied by several N–S-orientedthrust faults (Fig. 1). The studied section hasbeen measured along the northern and southernmargins of the Salado river valley, across thewestern flank of the anticline. It comprises theupper part of the Vaca Muerta Formation (itslower section is not exposed) and the wholeChachao Formation up to its stratigraphiccontact with the overlying Agrio Formation(Figs 1 & 2).
Facies analysis
The Tithonian–Valanginian succession is domi-nated by fine-grained siliciclastic facies, nodularand bedded marls, and bioclastic carbonates(Fig. 3). The Vaca Muerta Formation is charac-terized by fine-grained sediments, mostly blackshales, with some dark-grey limy shales anddark marls, formed by suspension fall-out in geo-chemically anoxic–suboxic environments. Someof these fine-grained sediments are very rich inammonites and small bivalves. One of the mostcommon features of these deposits, already men-tioned by Gasparini et al. (1997, 1999), is thesmall–scale cyclicity consisting of 0.4–2 malternations of black shales and organic marls(Fig. 3). Nodular limestones (micrites andmarls) are associated with the fine-grained silici-clastic rocks; nodule concentrations are rhyth-mically spaced, and were mostly precipitatedon skeletal cores (small to large fragments).Very thin and episodic ash layers are relativelyfrequent, from 2 to 10 cm thick, occasionallyreaching 40 cm thick, and exhibiting sharpbases where the ashes overlie shales. Somemassive layers or those with well-developednormal gradation are interpreted as ash-falldeposits and/or volcaniclastic deposits reworkedby low-density or dilute gravitational flows.
Thin and isolated packstones and wackestonesappear within the fine-grained packages. Thesebeds usually exhibit an irregular and/or sharplower surface and slight normal gradation. Theycan be either massive, laminated or with sym-metrical or asymmetrical combined flow ripples.
In the Vaca Muerta Formation, two informal stra-tigraphical sections can be recognized (Fig. 4).The lowermost one is composed of 75 m ofblack bituminous shales and (massive and/ornodular) fetid marls and micrites, with subordi-nated intercalations of coarser grained bioclasticcarbonates, as well as thin and fine-grained ashlayers. The upper section is composed of a rhyth-mic succession (23 m) of grey limy shales andwackestones, with isolated intercalations of
packstone beds. Towards the top of this section,some coarsening- and fining-upwards smallcycles (1.5–3 m thick) were identified.
The Chachao Formation overlies gradationallythe Vaca Muerta Formation, and most of itsfacies are similar to those of the Vaca MuertaFormation. However, in the Chachao Formationmarly beds and bioclastic carbonates are morecommon. The Chachao Formation shows charac-teristically a series of wackestones and pack-stones with abundant molluscan (oyster-rich)shell debris. As discussed below, some of thesebeds show in situ deep- to shallow-burrowinginfaunal bivalves. Other beds are almost exclu-sively formed of abundant disarticulated andfragmentary bivalve debris. Commonly, theseshell beds are composed of a lower interval ofdeep-infaunal bivalves lying in life position,and a strongly reworked upper bioclasticinterval.
In the Salado river valley, the Chachao For-mation can be subdivided into three informal sec-tions (Figs 3 & 4). The lowermost one (36 mthick) is composed of dark-grey limy shalesand dark shales, heterolithic beds (thin alterna-tions of wave-rippled sandstones and mud-stones), marls and packstones showing primarystructures, such as planar bedding, wave-ripplelamination and hummocky cross-stratification(HCS), and deformational structures (water-escape features, ball-and-pillow). This sectionshows a well-developed parasequence stacking,composed of cycles averaging 4 m thick.
The middle section of the Chachao Formation(Fig. 4) is a 17 m-thick succession characterizedby alternating cycles of dark-grey shales andmarls, with isolated and thinly bedded, biotur-bated, bioclastic wackestones composed of frag-mented infaunal shells and commonly withconcentrations of the pectinid bivalve Entoliumconcentrations on top. However, for the mostpart this section, and especially its middle part,lacks diverse benthonic organisms and moreoften contains scattered planktonic and pseudo-planktonic remains. However, to the top of thesection, marly beds usually show a few infaunalbivalves and scattered semi-infaunal (Pinna)bivalves in life position.
The upper part of the Chachao Formation(23 m thick, Fig. 4) is composed of coarse-grained shell beds associated with less frequentmarls and dark shales (like those of the middlesection). In the lower part of this section, shellbeds comprise burrowed grainstones and pack-stones composed of fragmentary gastropods,some infaunal bivalves, including trigoniids,and oysters. As with the middle section, manyof these beds show concentrations of epifaunal
RELATIVE OXYGENATION MENDOZA SHELF 189

Fig.3.
Lit
host
rati
gra
phic
alco
lum
nfo
rth
eT
ith
on
ian
–V
alan
gin
ian
inte
r val
inth
eS
alad
oR
iver
sect
ion
.S
amp
len
um
ber
sar
eo
nth
ele
fto
fth
eco
lum
n.
P. DOYLE ET AL.190

Fig. 4. Simplified log of the Vaca Muerta and Chachao formations. Informal sections, facies and cycles. Samplenumbers are on the right of the column. CU, coarsening-upwards cycles; FU, fining-upwards cycles.
RELATIVE OXYGENATION MENDOZA SHELF 191

bivalves (e.g. Entolium) on top. The upper part ofthis section is, instead, dominated byautochthonous and para-autochthonous coarse-grained and massive (bioturbated) packstonesconsisting of amalgamated 0.3–0.6 m-thicktabular layers composed of bored ostreid oystershells. These organisms are associated with gas-tropods and epifaunal bivalves (Entolium), aswell as infaunal bivalves (trigoniids, Panopea,Protocardium), in life position. In some casesthese are found as articulated shells oriented par-allel to the bedding planes, even forming a morechaotic and finer grained matrix of bioclasticdebris.
Environmental interpretation
Based on the general geometry of the substrate,Mitchum & Uliana (1985) and Legarreta &Uliana (1991) have proposed a depositionalramp model for the Tithonian–Valanginianunits of the Neuquen Basin, therefore devoid ofsubmarine fan or apron deposits as well aslarge-scale reef structures. For the NeuquenEmbayment, Spalletti et al. (2000) showed asimple distribution of carbonate and siliciclasticfacies associations, and a gradual transitionfrom the shallow-marine areas to the deepestareas without a talus slope or a significant slopebreak. According to the conceptual model pro-posed by Burchette & Wright (1992), Spallettiet al. (2000) basinal, outer ramp, middle rampand inner ramp environments were identified.
In the Mendoza Shelf, the general patternof the Tithonian–Valanginian sedimentationseems to be slightly different. During the Valan-ginian the ramp geometry of the basin gave riseto a mixed siliciclastic–carbonate platform.The thick bioclastic carbonates of the upperChachao Formation (Mombru et al. 1978;Legarreta & Kozlowski 1981; Legarreta et al.1993) suggest the development of an almost con-tinuous build-up (reef structure) along the bound-ary between the narrow platform to the east andthe talus slope to the west and NW.
Most of the Vaca Muerta Formation in theSalado section is characterized by fine-grainedshaly and carbonate organic-rich sediments,often with a cyclic depositional arrangement.These deposits accumulated in a basinalenvironment, and represent the suspension sedi-mentation of siliciclastic and carbonateparticles above an anoxic–suboxic seafloor. Ashallower, but still basinal, marine environmentcan be proposed when the dark fine-grainedsediments show thin siliciclastic or carbonatesand intercalations. As described in previouspapers (Gasparini et al. 1997; Spalletti et al.
1999, 2000) these coarser-grained layers, withevidence of wave reworking and low-density tempestite flows, are interpreted as verydistal deposits produced by storm processes(Myrow & Southard 1991, 1996; Midtgaard1996).
A depositional setting representing the tran-sition between the offshore and the lower shore-face environments can be proposed for thedeposits of the lower Chachao Formation,where the bioclastic carbonate beds are more fre-quent and the parasequence stacking is clearlydeveloped. In the middle Chachao section,deeper basinal marine conditions, like those ofthe Vaca Muerta Formation, were re-established.
The oyster beds of the Upper ChachaoFormation represent a marginal sector of alarge ‘reef’ structure, probably developed bythe margin of the Mendoza Shelf. The shellbeds from the lower parts of the UpperChachao Formation were formed in shallowand high-energy environments under conditionsof reduced rates of terrigenous sediment accumu-lation (Abbott 1997). They represent a transitionzone between shoreface and offshore, with amacrofossil assemblage composed of mixedtaxa representing both nearshore and offshoreenvironments. The parasequences composed ofautochthonous and parautochthonous oyster-rich packstones, marls and dark shales in theuppermost Chachao Formation probably rep-resent a relatively quieter shelf environment.Although oyster beds have been associated withrestricted environments, we have also observedthe development of oyster banks in the openshallow-marine platform surrounding theMargarita Island (north Venezuela, 118Nlatitude). If this was the case in the upperChachao Formation, the vertical distribution ofshell beds can be interpreted in terms of a slightlyupwards-deepening succession, from the lowershoreface to the proximal offshore in an openshallow-marine mixed (carbonate–siliciclastic)platform. As such, it is more likely that the lowfaunal diversity of these beds may be the resultof reduced competition by other organisms forspace and nutrients (faunally restricted environ-ments) as well as a lack of predation (Glenn &Arthur 1990; Abed & Sadaqah 1998), ratherthan traditional interpretations of restrictedenvironments.
Ichnology
The Salado River section exhibits a poorlydiverse ichnological assemblage associatedwith a background of relatively fine-grained sub-strates (Fig. 5). The transition from the Vaca
P. DOYLE ET AL.192

1 1A 1B 1C 1D 1E 1F 2 3 4 5 6 7 8 9 10 11 12C
hach
ao F
m.
Vac
a M
uert
a F
m.
20 m
A B C D
1
85
138
230
86
139
231
385
Trace fossils Associations
Fig. 5. Trace fossil distribution. 1, Thalassinoides; (1A, type A; 1B, type B; 1C, type C; 1D, type D; 1E, type E; 1F,type F); 2, Arenicolites; 3, Chondrites; 4, Gordia; 5, Palaeophycus; 6, Planolites; 7, Phycodes; 8, Teichichnus; 9,Rhizocorallium; 10, small diameter burrows; 11, Gastrochaenolites; 12, Trypanites. A, laminated facies association; B,Chondrites association; C, Thalassinoides association; D, Arenicolites association.
RELATIVE OXYGENATION MENDOZA SHELF 193

Muerta to the Chachao Formation is marked bythe transition from black mud to carbonate-richand heterolithic intervals, and therefore theincreased possibility of substrate control havinga larger impact on the resultant biofacies devel-opment. This is particularly true where thereare periodic episodes of sand deposition, whichcontain typically sand-dominated traces such asArenicolites and Gordia, traces that are other-wise largely absent from the succession. Fourprincipal ichnological associations can berecognized (Fig. 5), which are discussed below.
Laminated facies (non-bioturbated
association)
This facies represents the basic background con-ditions within the basin. Laminated deposits,lacking bioturbation, are locally interbeddedwith the Chondrites bioturbated depositsdescribed below. This is a common feature ofmost black shale successions (Rhoads & Morse1971; Bromley & Ekdale 1984; Savrda &Bottjer 1986; Wignall 1993) and is clearlyrelated to low dissolved oxygen levels, laminatedfacies being largely a product of truly anoxicconditions (Wignall 1993).
Chondrites association
Chondrites is the commonest background-leveltrace fossil and its trace maker is considered tobe the primary bioturbator in homogenizinglaminated mud facies such as those in the VacaMuerta Formation. This is in keeping with therole for this trace fossil as the first to colonizeand the last to leave in oxygen-deficient blackshale sequences (Bromley & Ekdale 1984).However, thoughout the succession, Chondritesis poorly diverse, with small overall burrowdiameters that are on a millimetre scale. This isnothing like the large burrow diameter Chon-drites that developed in the oxygen-deficientLower Jurassic Posidonienschiefer of Germany,where burrow diameters commonly reach up to5 mm or more (Seilacher 1982a, b).
Thalassinoides association
Thalassinoides is relatively common and is mostfrequently associated with the carbonate andheterolithic facies within the Salado section.Thalassinoides is commonly considered as anindicator of oxic conditions, and is thereforean important oxic benchmark in the palaeo-oxygenation schemes of Savrda & Bottjer(1986). Furthermore, Savrda & Bottjer (1986,
1987, 1989) have suggested that the maximumdiameter of the burrows depends on the amountof oxygen in the substrate. The greater theoxygenation the larger the infaunal biotur-bation. From this relationship an upwardsincrease in oxic conditions is interpreted fromthe increase in the diameter of the Thalassinoidesburrows.
Thalassinoides is also a determinant of sub-strate firmness, as the complex galleries of theburrow system are not capable of remainingunsupported in a soupy substrate (Ekdale et al.1984). Within the Chachao section of theSalado River, at least six types have been recog-nized, A–F (Fig. 6). These forms are discussedbelow.
Thalassinoides types A–D. Thalassinoidestypes A–D conform to the ichnospecies Thalas-sinoides suevicus as currently defined (Ekdaleet al. 1984), characterized by regular galleriesand straight shafts (Fig. 6). They are developedwithin the carbonate units in the Salado Riversection, and the recognition of four separatemorphotypes (A–D) is dependent on the differ-entiation of burrow fill. Thalassinoides type Ais characterized by a bioclastic infill that dis-tinguishes it from its otherwise poorly bioclastichost sediment. In this case, the infill is interpretedas the by-product of feeding and predation by theThalassinoides trace-making organism, andperhaps represents active successive debrischamber infill by predators. Thalassinoidestype B displays a sand passive infill with somebioclastic pockets, the bioclasts again probablybeing derived from predation. Infill heremay conform to both passive and active infillingby the trace maker, and, dependent on the burrow,may be interpreted as a function of activities inthe range from both soft to firm grounds. Thalas-sinoides type C has a passive sand infill alone,with the sand filling the empty galleries withoutany intervention of the trace maker. Thalassi-noides type D is most probably the result of apost-depositional mud infill, in which the darkmud comes from the substrate above.
Thalassinoides types C and D are sometimesaccompanied by a background of small diameterburrows that form a conspicuous ichnofabric, inwhich it is impossible to recognize individualichnogenera. This bioturbated texture showsboth a very intense activity of the endobenthiccommunity and a cross-cutting of tiers.
Thalassinoides type E. This form correspondsto the ichnospecies Thalassinoides paradoxicus.It is associated only with coarse bioclasticfacies, and the form of the burrow shafts and
P. DOYLE ET AL.194

(A) Thalassinoidesbioclastic infillin limestone
(C) Thalassinoidessand infill inlimestone
(D)mud infill inlimestone
Thalassinoides
(B)sand infill withbioclastic pocketsin limestone
Thalassinoides
(E) Thalassinoide sandor coarser infill inbiogenic carbonate banks
(F) Any Thalassinoides typeexposed on the bedtopshowing erosive surfaces
Fig. 6. Sketches showing the six different types of Thalassinoides recognized in the studied section. Sample numbersare on the right of the column.
RELATIVE OXYGENATION MENDOZA SHELF 195

galleries are controlled largely by the occurrenceof shell debris, producing the contorted formscharacteristic of this ichnospecies, formingquite an irregular trace pattern. This is indicativeof primary substrate control on the assemblageand may be representative of a firm- to hard-ground environment, in this case the abundanceof bioclastic debris. In the same way, the oystershells of the beds bearing Thalassinoides typeE may show borings assignable to Trypanitesand Gastrochaenolites (Fig. 6).
Thalassinoides type F. This type is associatedwith eroded bed units, where Thalassinoidesgalleries are preserved upon the bedding-planesurface. This Thalassinoides type is scarce inthe Salado section but is very common in otherexposures. It developed when the upper part ofa substrate bearing Thalassinoides types A, Bor C was eroded and the trace level wasexhumed. The presence of Thalassinoides typeF suggests the development of a local discontinu-ity or hiatus between the bed with Thalassinoidesgalleries and the bed deposited above.
Arenicolites association
This is the rarest association and comprisesthe ichnogenera Arenicolites, occasionallyaccompanied by Gordia, Palaeophycus, Plano-lites, Phycodes, Rhizocorallium and Teichichnus.These traces are largely associated with sand-stone beds and even coarser-grained beds,which are uncommon in the Salado section.The Arenicolites association is interpreted asthe product of shallower marine conditionsand/or a higher level of kinetic energy in themarine substrate (Ekdale et al. 1984).
Macrofossil palaeoecology
The Vaca Muerta and lower Chachao formationscontain a relatively abundant macrofossil fauna,which is nevertheless restricted in diversity(Fig. 7). This comprises nektonic organisms,mostly ammonites, that form the backgroundfaunal signal, and a range of benthic faunalassociations, discussed below. The palaeoecolo-gical information provided was based on thestudy of bedding-plane associations exposedwithin the Salado section. Although not strictlyquantitative, this was taken to be significant,with due regard taken to the level of autochthonyof benthic assemblages.
Nektonic organisms
The Vaca Muerta and lower part of the Chachaoformations in the Rio Salado contain abundant
nekton, mostly in the form of ammonites,although there are rarer accumulations of fishdebris. These faunal components are discussedin the following paragraphs.
Ammonites. A range of ammonites are knownfrom the Vaca Muerta and lower Chachao for-mations, and have been illustrated by Riccardi(1983, 1988) amongst others. The majority ofammonites within are preserved flattened, andthe original aragonite dissolved. However,some preservation of siphuncles as phosphatictubes within the shells is observed. Some flat-tened ‘beermat’ preservation, with carbonatesintact, as well as three-dimensional preservationof ammonites is also recorded from some of thecarbonate concretionary horizons at the base ofthe section. The base of the Salado section(Vaca Muerta Formation) is particularly charac-terized by ammonites encrusted by a range ofbyssally attached and cemented bivalves.
At the top of the Salado section, within car-bonate facies of the Chachao Formation, the‘beermat’ preservation is fine enough to preserveammonite microconchs with intact lappets, andthe contents of body chambers, includingin situ aptychi, may be observed. This demon-strates little current activity or other shallow-marine processes that would easily destroy ordisturb such delicate skeletal features. Thereare few if any associated benthic bivalves ident-ifiable as either pseudoplankton or benthic colo-nies associated with these ammonites, and thismay indicate an inability of benthic bivalves tocolonize foundered ammonites on the seafloor,the most probable conclusion being through thelack of available dissolved oxygen in thebottom waters at this point.
The ammonite preservation encountered herehas also been identified in the Lower JurassicPosidonienschiefer by Seilacher et al. (1976),interpreted by him as the product of the early dis-solution of the skeletal aragonite, followed by thecollapse of the organic periostracum, whichforms an organic envelope for the ammonites,and there is no reason to doubt this interpretationhere. The presence of epibionts suggests that theammonites were either the hosts for pseudo-planktonic bivalves (e.g. Seilacher 1960,1982a, b; Wignall & Simms 1990; Doyle &Whitham 1991, Etter 1996) or were acting as‘benthic islands’ for epizoic colonization (e.g.Cope 1968; Kauffman 1982). Both are recordedfrom the Salado section.
Fish. That there were abundant fish within theNeuquen Basin is indicated through the scatteredpresence of scales and rare bones, including
P. DOYLE ET AL.196

jaws. In many cases these accumulations formcohesive ‘lumps’ or ‘blobs’ of bone materialthat may be safely interpreted as the by-productsof predation by other fish, reptiles or other largepredators. Elsewhere in the Neuquen Basin the
Vaca Muerta Formation contains abundant,well-preserved marine reptiles (Gasparini et al.1997) and, although no large vertebrate materialwas collected in the present study, this demon-strates the abundance of large nektonic
20 m
1
85
138
230
325
86
139
231
326
385
8761 52A 2B
2 46D 7A6A 6B 6C5C5A 7B5D
31B 1C 1D 4A 4B1A 5B
Cha
chao
Fm
.V
aca
Mue
rta
Fm
.
A B C D E F G
Macrofossils Associations
Fig. 7. Body fossil distribution. 1, Nekton (1A, ammonites; 1B, aptycus; 1C, belemnites; 1D, fish bones); 2,pseudoplankton (2A, bivalves byssally attached; 2B, oysters); 3, serpulids; 4, gastropods (4A, turretted; 4B,planktonic); 5, byssally attached bivalves (5A, oxitonids; 5B, pectinids; 5C, Entollium; 5D, inoceramids); 6, oysters(6A, Ceratostreon; 6B, Aetostreon; 6C, Deltoidion; 6D, indet.); 7, shallow infaunal bivalves (7A, thin valves; 7B, thickvalves); 8, deep infaunal bivalves. A, Nekton; B, pseudoplankton; C, serpulids; D, epibyssate bivalves; E: oysters; F,shallow infaunal bivalves; G, deep infaunal bivalves. Sample numbers are on the right of the column.
RELATIVE OXYGENATION MENDOZA SHELF 197

organisms capable of living in a more oxic zonewithin the water column.
Planktonic–pseudoplanktonic associations
The planktonic and pseudoplanktonic associ-ations that have been recognized are predomi-nantly molluscan. These are described below.
Planktonic gastropod asssociation. Gastropodsoccur within several levels of the Vaca Muertaand lower Chachao formations. Local monospe-cific abundances or ‘gluts’ of small, rounded gas-tropods were also recorded. These are similar tothe gastropod associations recorded by Etter(1995) from the Middle Jurassic of Germanythat have been considered to be planktonic(Bandel & Hemleben 1987), but which werethought to be benthic by Etter. In either case,they represent opportunists, either as benthos ornekton, demonstrated by the monospecificnature of the assemblage, controlled as with allmonospecific assemblages encounted in theSalado section by primary oxygen levels.
Pseudoplanktonic association. This associa-tion comprises the close association of epibionticbivalves with ammonites, the bivalves beingfound attached to both flanks of the ammonitesobserved. This is possible because of the‘beermat’ preservation, which causes thethicker shelled bivalves to be pushed throughthe flanks of the ammonite test. The pseudo-plankton is restricted mostly to thin-shelledcementing oysters, displaying clear xenomorph-ism and to byssally attaching, oxytomid-typebivalves. The majority of these bivalves are con-centrated within the umbilical regions of theammonites studied. In most cases the frequencyof encrustation is intense, with a majority ofammonites infested, although it should be notedthat an accurate count is difficult to determinegiven there were no bedding-plane surfacessufficiently exposed to carry out a detailedpalaeoecological assessment. Infauna is other-wise restricted, although there are records ofsome thin-shelled individuals within thesection. The pseudoplanktonic association maybe distinguished from the otherwise similarbenthic island association by: (1) the presenceof bivalves on both sides of the ammonitesexamined; (2) the clustering of bivalves withinthe umbilical region of the ammonite, and theabsence of overgrowth; and (3) the absence ofother clusters of bivalves not associated with anammonite. The pseudoplanktonic associationwas recorded from several levels within thesection studied and, in most cases, no other
associated fauna were recorded from theselevels. Where found, associated fauna are gener-ally restricted to isolated bivalves of the samegroups as those attached to the ammonites.
This represents a classic pseudoplanktonicassociation given the frequency of the occurence,the presence of epibionts on both surfaces of theammonites and the overall absence of otherbenthos. It is similar to those described bySeilacher (1960, 1982a, b), Wignall & Simms(1990), Doyle & Whitham (1991) and Etter(1996), in which settings the presence of pseudo-planktonic host associations in the absence ofbenthic molluscs is considered typical of lowlevels of dissolved oxygen at the seabed.
Benthic associations
A number of benthic molluscan associations maybe recognized within the Salado section (Fig. 7).These are recurrent assemblages, and are autho-chonous or parautochthonous clusters that maybe interpreted as approximations of the originalskeletal macrofaunal benthos.
Benthic island association. The benthic islandassociation is represented by clusters of epifaunalpteriform and ostreid bivalves that are otherwisenot abundant. Usually the association is mono-specific, comprising clusters of a single epifaunalbivalve taxon typically with an ammonite host.The bivalves are not clustered in specific areasof the ammonite shell, and in the majority ofcases the bivalves are seen to be overgrowingthe shell margins of the ammonite. Bivalves areonly present on the upper surface of the shell.The benthic island association is restricted torelatively few beds within the Salado section,and is easily distinguished from pseudoplank-tonic associations that have a much moretightly constrained concentration of bivalves intheir umbilical regions.
The concept of benthic islands as a three-dimensional refuge for benthos, emergingabove the lowest levels of dissolved oxygen asdeveloped by Kauffman (1981) for the Posido-nienschiefer, is probably too sophisticated forthe Salado section, and the level of informationavailable is insufficient to suggest that theseislands provided small highs above a stratifiedlowest oxygen layer. Instead, it is probably suffi-cient to consider the association as representingmarginally increased oxygen conditions, and arelatively soft substrate (see also Wignall 1993).
Ammonite inquilinism. An association ofammonites and decapods is recorded on thebasis of two specimens of flattened ammonites
P. DOYLE ET AL.198

with fragmentary decapod carapaces within theirbody chambers in the upper part of the Saladosection. This is considered to be a primaryassociation for the following reasons: (1) thepreservation of the ammonites, which, althoughflattened, show delicate features such as lappetsintact; and (2) the absence of winnowing orother indications of current activity. Thissuggests a primary, rather than post-mortal,relationship with the decapods finding refugewithin the ammonite body chamber on theseabed. The presence of decapods within thebody chambers of the sunken ammonitessuggests that levels of dissolved oxygen weresufficient to permit life, and this association isin many ways equivalent to that of the benthicisland association already discussed.
This kind of association has been termedammonite inquilinism, the post-mortal associ-ation of decapod crustaceans within the bodychambers of large ammonites. It has beenrecorded from the Lower Jurassic of Englandand Germany (Fraaye & Jager 1995).
Rotularia association. Serpulids of the genusRotularia are common at certain levels in theSalado section. These are observed formingsmall monospecific clusters, although Rotulariais known occurring with other taxa, mostnotably fragmentary infauna, further up-section.Life orientation of this genus is with the elongatesiphonal tube vertical, the main part of the serpu-lid being oriented within the body of the sedi-ment, and this is the typical orientation ofRotularia within the Salado sediments.
Macellari (1984) has discussed the modeof life of Rotularia from the Cretaceous rocksof Antarctica. Three possible modes weresuggested: (1) epifaunal suspension-feeders; (2)infaunal deposit-feeders; and (3) infaunal sus-pension-feeders, with the position of the siphonaltube considered to be of paramount importancein the interpretation of the mode of life.
Macellari favoured an infaunal filter-feedingmode with the siphonal tube oriented verticallyin life, and this is supported by the presentassociation of rotularids. He also noted the corre-lation of serpulid abundance with increased mudlevels, and recorded the decline in abundancewith coarser grained facies. The present associ-ation is comparable in that the serpulids foundin the Salado section are most commonlylocated within black muds, with little otherassociated body fauna. The comparative glut ofserpulids associated with certain horizons andin small concentrations is consistent with the per-ceived characteristics of an opportunistic species(Levinton 1970). In this case, the presence of
appropriate mud facies is seen to be significant,although it is clear that without suitably oxyge-nated conditions, at least at dysaerobic levels, itis unlikely that the association would besuccessful.
Entolium association. Shell pavements com-prising disarticulated, but otherwise intact,valves of the free-living epifaunal bivalve Ento-lium are common in the upper part of the Saladosection. These epifaunal bivalves are commonlyfound associated with the tops of carbonate units,and form sparse shell pavements with little or noimbrication and with valves in both stable andunstable orientations. Valves are usually wellpreserved, but in some beds damage to thevalves has occurred, usually in the form ofdestruction of the wings or shell margin.
Entolium is usually interpreted as an free-living bivalve requiring a firm, or at least notsoupy, substrate (Wignall 1990). This interpret-ation is supported by the relatively high concen-tration of bioclastic debris within the carbonateunits of the Salado section. The concentrationof valves most probably represents a shell bedof para-autochthonous bivalves, indicating colo-nization of a relatively firm substrate with, onceagain, at least low levels of dissolved oxygen.Other bivalves adapted to softer substrates aretypically thin shelled, having adopted the snow-shoe approach (Thayer 1975; Wignall 1993;Etter 1996). It is more likely, given the free-living strategy of these bivalves, that the pre-sence of Entolium is indicative of a firmersubstrate.
Shallow infaunal suspension-feeding bivalveassociation. Small, shallow infaunal bivalvesare present towards the top of the Saladosection, comprising small, thin-shelled andmostly articulated valves of Aphrodina andrelated taxa. Typically, these taxa have relativelyshallow burrows, although they are not associ-ated with rapidly shifting substrates. Thickershelled Eriphyla, and trigoniids, particularlySteinmanella, are uncommon, but also form acomponent of the fauna.
All of these bivalves are interpreted as shallowinfaunal suspension-feeders. The presence ofEriphyla in several of the fine-grained mud-richor marl units demonstrates the capability ofthese taxa to inhabit fine-grained, althoughrelatively firm, substrata. The presence ofthicker shelled components towards the top ofthe sequence studied may be readily consideredas the product of normally oxygenated bottomwaters, and possibly even higher energeticconditions (Wignall 1993; Etter 1996). This
RELATIVE OXYGENATION MENDOZA SHELF 199

assemblage is therefore indicative of normal,unrestricted and relatively firm (i.e. non-soupy)substrate conditions.
Deep infaunal suspension-feeding bivalve asso-ciation. Deep-infaunal bivalves are almostwholly associated with the upper part of thesection, where they were mostly observed inlife position. Commonly, taxa such as Lucinia,Panopea and Thracia occur at the base of car-bonate units that often exhibit a bioclastic andstrongly reworked top. This is interpreted as theproduct of strong in situ reworking of moreshallow burrowing tiers of the carbonate unit,the level of reworking not tapping into thelower levels inhabited by the deeper burrowingbivalves.
Infaunal deposit-feeding association. This is arare component of the fauna, but, wherepresent, it comprises abundant infaunal deposit-feeders composed primarily of small nuculidbivalves. According to Morris (1980), deposit-feeders are often associated with dysaerobic con-ditions, but, as pointed out by Wignall (1993),this may be a function of softer substrates.
Exogyrid association. This comprises exogyridoysters of the genera Ceratostreon and Aetos-treon. These associations are common in theupper part of the succession and regularlydisplay a dense concentration of these taxa.These are usually not disarticulated and formbeds of up to 1 m in thickness. At their thickestthese shell beds are equivalent in facies tothose exhibited in the basin central facies of theChachao Formation, where great thicknesses ofalternating Ceratostreon and Aetostreon arefound.
Ceratostreon is also present as a minor com-ponent of the black muds occurring lower downthe succession, and, in some cases, they arefound with associated serpulids of the genusRotularia. In this case, small ‘clusters’ of Cera-tostreon are found in an assumed autochthonousassociation with a ‘shell pavement’ produced bythe concentration of serpulids.
This represents a modification of an otherwisesoft substrate by the concentration of serpulids,indicating a two-stage colonization process ana-logous to that demonstrated by Doyle &Whitham (1991) and discussed by Wignall(1993), with similar associations in the JurassicOxford Clay of the UK where low-diversityGryphaea assemblages were considered to bethe result of low oxygen conditions, in whichan increase in soft substrates combined toreduce the diversity still further. In the case of
the Chachao associations the increased import-ance of soft substrates is more likely to be afactor where Ceratostreon is found in isolatedclusters, rather than in shell beds.
Relative oxygenation curves
Following the methodology of Savrda & Bottjer(1986), it is possible to develop a trace fossilmodel for relative oxygenation of black shalesthrough the relative diversity of trace fossilassemblages. This technique has been reappliedby other authors (e.g. Doyle & Whitham 1991),and has been tested and reviewed by others(e.g. Allison et al. 1995; Etter 1995; Doyleet al. 1998; Spalletti et al. 2001), and is generallyheld to be robust. However, the close associationof low oxygen conditions with mud facies has ledto difficulties in the recognition of the dominantcontrolling factor, as outlined by Wignall(1993) and Goldring (1995). Two approachesare used here, that of the standard relativeoxygenation curve based on trace fossil assem-blages developed by Savrda & Bottjer (1986),and a consideration of relative oxygenationbased on body fossil assemblages (Doyle &Whitham 1991; Wignall & Hallam 1991;Wignall 1994).
Relative oxygenation from the ichnofauna
Following Savrda & Bottjer (1986), two basiccharacteristics were employed in constructingrelative oxygenation curves: (1) the diversity ofthe assemblage, which for the most part ispoorly diverse; and (2) the burrow diameter,including burrow fill. Increasing diversity andburrow diameter equating with increased relativeoxygenation. The relative oxygenation curvederived from this approach for the intervalstudied here is presented in Figure 8.
The most significant issue is the transitionfrom laminated to bioturbated shales, with Chon-drites being present in each bioturbated interval.Alternation between laminated and Chondrites-bearing sediments can be taken as the nominalboundary between anoxic and oxic conditions;effectively the anaerobic–exaerobic/dysaerobicboundary, as defined by Savrda & Bottjer(1986, 1987). Commonly, Chondrites isaccompanied by Thalassinoides with smallburrow diameters. This is commonest in theblack-mud-dominated part of the sequence, theVaca Muerta and lowermost Chachao formationsas developed in the Salado section. Largerburrow-diameter Thalassinoides occur morecommonly in the carbonate-dominated parts ofthe section (Chachao Formation), with a variety
P. DOYLE ET AL.200

of fills as described above. This is generallyindicative of increasing oxygenation of the basinup-section, consistent with the current models,with a transition from anoxic, through fluctuating
dysaerobic to oxic conditions determined bythe simple relationship of: laminated shales–Chondrites/small burrow-diameter Thalassi-noides–large burrow-diameter Thalassinoides.
Cha
chao
Fm
.V
aca
Mue
rta
Fm
.
Lam
inat
ed
Cho
ndrit
es
Sm
all
Tha
lass
inoi
des
Larg
eT
hala
ssin
oide
s
Are
nico
lites
Tha
lass
inoi
des
bioc
last
icin
fill
TRACE FOSSILS
RelativeO2
- +
RelativeO2
- +
Nek
ton
Pse
udop
lank
ton
Ser
pulid
s
Epi
byss
ate
biva
lves
Oys
ters
Dee
pin
faun
al
Sha
llow
infa
unal
Combinedcurves
- +
MACRO FOSSILS
20 m
A
C
B
D
E
Fig. 8. Oxygen indicators and relative oxygenation curves. A, B and C are suggested oxygenation cycles for the VacaMuerta Formation. D and E are suggested oxygenation cycles for the Chachao Formation.
RELATIVE OXYGENATION MENDOZA SHELF 201

In the lower part of the section, at the tran-sition between the Vaca Muerta and Chachaoformations, relatively large burrow-diameterThalassinoides conforming to types A–Ddescribed above occur in carbonate-rich levelsbetween black shales lacking pervasive biotur-bation, with the whole part of the sectionlacking infaunal bivalve associations. This rep-resents a paradox, for, as explained above, thepresence of Thalassinoides has been bench-marked in most palaeo-oxygenation studies asan indicator of oxic conditions, at odds withthe absence of other benthos and a backgroundof pseudoplanktonic bivalves attached tonektonic ammonites.
Relative oxygenation from body fauna
Benthic and pseudoplanktonic fauna are a valu-able proxy for the development of relativeoxygenation, although not as widely employedas the trace fossil models. In general, therelationship of abundance and diversity is seenas critical, as with most assemblages it is thetransition from opportunistic to equilibriumspecies (Levinton 1970; Morris 1980; Doyle &Whitham 1991; Wignall & Hallam 1991). Thepresence of benthos with a pseudoplanktonichabit is also significant (Wignall & Simms1987; Doyle & Whitham 1991).
In the Salado section the presence of nekton –ammonites – is pretty well universal, with ammo-nites reasonably abundant throughout thesection, declining to the top of the section. Pseudo-planktonic bivalves are similarly distributed, butwith a greater abundance in the mid-part of thesection.
It is the relationship between the epifaunalmolluscan fauna – epibyssate bivalves andoysters – and the infaunal components that isseen to be significant in terms of relativeoxygenation. Epifaunal assemblages include thebenthic island associations, and the concen-trations of gastropods and Entolium. Similarly,isolated clusters of exogyrid oysters are signi-ficant, and may represent increasing oxygen(dysaerobic conditions) and fluctuation in sub-strate from soft to firmer. The serpulid Rotulariais taken to represent an opportunistic speciesand is common at the base of the succession,occurring beneath the first common pseudo-planktonic development on the nektonicelements. It may well also carry a primary lowoxygen signal, but it is thought to prefer softersubstrates.
The vast majority of the benthic molluscspresent are suspension-feeders, with relativelyfew deposit-feeding bivalves present. Like that
of the serpulid Rotularia, the palaeoenviron-mental signal from these deposit-feeders isambiguous, with low oxygen being overprintedby a preference for soft substrates.
Epifaunal bivalves unattached to nekton occuronly at the base and very top of the succession,and are generally exclusive of the infaunalbivalves. Shallow infaunal bivalves are seenwithin all the major facies, and in some casesoccur exclusive of epifaunal bivalves. Thepresence of deep infaunal bivalves is taken torepresent the maximum oxygenation state present,and these molluscs are common only in thecarbonate-rich basal Chachao Formation(Fig. 8). As described by Wignall (1993)increasing diversity and burrow depth can be afunction of both increasing oxygenation, andincreasing substrate suitability, and therefore dif-ficult to disintangle. Despite this, the overallsignal is in line and in keeping with whatcould be expected from the lithofacies and theichnological information – increasing oxygen-ation up-section, associated with the increase incarbonate facies (Fig. 8).
Discussion
In the main, the relative oxygenation curves pro-duced by both datasets agree well, particularly inthe mid-upper part of the succession, character-ized by an increase in carbonate units. Thebasic trend in this upper part of the successionis a cyclical arrangement with increasing oxy-genation of bottom waters indicated by thecycle: laminated Chondrites–small-diameterThalassinoides–large-diameter Thalassinoides–Arenicolites–Thalassinoides with bioclasticinfill. This ichnological cycle is matched with asimilar cycle of body fossil occurrence: pseudo-plankton–epifaunal bivalves–shallow infaunalbivalves. This is, in itself, relatively unre-markable, demonstrating a cycle of increasingoxygenation of bottom waters, allowing for amore prolonged colonization window. Thecycles themselves overprint the pattern ofmarl–packe/wackestones observed, with no sig-nificant relationship observed between increasedfaunal diversity and increasing bioclastic carbon-ate. However, on the broad scale, the observedcycles are mostly associated with the uppercarbonate-rich facies, overlying the otherwiselow-oxygen black-shale-dominated facies at thebase of the section. At least three of thesecycles can be recognized in the carbonate-richsection (C–E in Fig. 8).
The basal black shales do, however, showsome exception to the general pattern, and, insome cases, there appears to be conflict
P. DOYLE ET AL.202

between trace and body fossil oxygenationsignals. Within the black shales there is atleast one, and possibly two, furtherlaminated–Chondrites/small Thalassinoides–large Thalassinoides cycles (A and B in Fig. 8)that may possibly be substrate controlled, inthat the large Thalassinoides assemblages areseen associated with thicker carbonate layers.The basal cycle B is not in any way mirroredby the body fauna, which lacks either epi- orinfaunal bivalves (Fig. 8). The cycle below (A)is associated with common serpulids and, inter-estingly, the colonization of the carbonatecomponents with shallow, suspension-feeding,infaunal bivalves. This is consistent with well-oxygenated bottom waters, and is inconsistentwith the relatively low diversity of the tracefauna.
As previously recognized by Wignall (1993),and underlined by this study, disentangling thesubstrate and limited oxygentation signals inblack shale facies can be a difficult task,especially where there is a definite preferencefor softer substrates (e.g. Rotularia, Chondrites,deposit-feeding bivalves) that could be misinter-preted as a low oxygen signal, where diversitiesare low. As such, this study underlines theimportance of the comparison of more than onedataset in the interpretation of relative oxygen-ation. Clearly, the discordance between bodyfossil and ichnological factors demonstratesthat substrate can be a limiting factor asimportant as that of dissolved oxygen levels.
Conclusions
The Tithonian–Valanginian succession exposedin the Salado valley shows the transition fromblack shale to carbonate facies, although thelater biohermal carbonates are limited and largelyundeveloped. Our analysis of this section illus-trates the following important points: (1) thatcarbonate production increases up-section; (2)that faunal associations are mostly limited topoorly diverse epibenthos and pseudoplanktonin the lower part, with increased diversity inthe lower Chachao section, including shallowand deeper infaunal bivalves; (3) that there is abackground level of laminated shales to Chon-drites bioturbation typical of anoxic–suboxicconditions; (4) that Thalassinoides suevicus iscommonly developed in micritic limestone andcarbonated sands units throughout the section.
Interpretation of the palaeo-oxygenation ofthis section on the basis of trace fossil taxaalone (cf. Savrda & Bottjer 1986) would givean inaccurate interpretation because of: (1)widespread Thalassinoides burrows of more
than 1 cm in diameter at the base of the sectionsuggesting at least upper dysaerobic facies,whilst the coeval, greatly reduced, and probablyopportunistic fauna of rotularids and pseudo-planktonic bivalves on ammonite hosts is indica-tive of lower dysaerobic or exaerobic facies; (2)the almost exclusive development of Thalassi-noides in limestones and sandstones, whileassociated body fauna are not as facies depen-dent. We believe that there was a primarysubstrate control on trace fossil diversity andoccurrence, with a primary oxygenation signalprovided by body fossil evidence. This studydemonstrates the importance of integratedtrace and body fossil analysis in the fuller under-standing of black shales. When comparing thecurves inferred from fossils and trace fossils inthe Vaca Muerta and Chachao formationsthe mismatch between the palaeo-oxygenationcurves inferred sedimentologically and thoseobtained exclusively from fossil traces (Savrda& Bottjer 1986, 1987, 1989) indicates that tracefossils should not be used in isolation.
This work was funded by a CONICET–Royal Societyexchange grant, the CONICET Research Project PID858/98 and SETCYP Project PICT 07-08451, which wegratefully acknowledged.
References
ABBOTT, S.T. 1997. Mid-cycle shellbeds from mid-Pleistocene cyclothems, New Zealand: impli-cations for sequence architecture. Sedimentology,44, 805–824.
AGUIRRE-URRETA, M.B. & RAWSON, P.F. 1998.The Early Cretaceous (Valanginian) ammoniteChacantuceras gen. nov. – a link between theNeuquen and Austral Basins. AsociacionGeologica Argentina, Revista, 53, 354–3654.
ALLISON, P.A., WIGNALL, P.B. & BRETT, C.E. 1995.Palaeo-oxygenation: effects and recognition. In:BOSENCE, D.W.J. & ALLISON, P.A. (eds) MarinePalaeoenvironmental Analysis From Fossils.Geological Society, London, Special Publications,83, 97–112.
BANDEL, K. & HEMLEBEN, C. 1987. Jurassic hetero-pods and their modern counterparts (planktonicGastropoda, Mollusca). Neues Jahrbuch fur Geolo-gie und Palaontologie, Abhandlungen, 174, 1–22.
BRACACCINI, O.I. 1970. Cuenca del Salado. In:LEANZA, A.F. (ed.) Geologıa Regional Argentina,407–417. Academia Nacional Ciencias, Cordoba.
BROMLEY, R.G. & EKDALE, A.A. 1984. Chondrites: atrace fossil indicator of anoxia in sediments.Science, 224, 872–874.
BURCHETTE, T.P. & WRIGHT, V.P. 1992. Carbonateramp depositional systems. Sedimentary Geology,79, 3–57.
COPE, J.C.W. 1968. Epizoic oysters on Kimmeridgianammonites. Palaeontology, 11, 19–20.
RELATIVE OXYGENATION MENDOZA SHELF 203

DOYLE, P. & WHITHAM, A.G. 1991. Palaeoenviron-ments of the Nordenskjold Formation: an AntarcticLate Jurassic–Early Cretaceous black shale-tuffsequence. In: TYSON, R.V. & PEARSON, T.H.(eds) Modern and Ancient Continental ShelfAnoxia. Geological Society, London, SpecialPublications, 58, 397–414.
DOYLE, P., POIRE, D.G., SPALLETTI, L.A., PIRRIE, D.,BRENCHLEY, P. & MATHEOS, S.D. 1998. Palaeoen-vironmental analysis of the Vaca Muerta andChachao Formation of the Neuquen Basin inwest-central Argentina: implications for palaeo-oxygenation studies. 14th International Sedimento-logical Congress, Alicante, Abstracts, H-2,303–304.
DUFF, K.L. 1974. Palaeoecology of a bituminousshale: the Lower Oxford Clay of central England.Palaeontology, 18, 443–482.
EKDALE, A.A. & MASON, T.R. 1988. Characteristictrace-fossil associations in oxygen-poorsedimentary environments. Geology, 16, 720–723.
EKDALE, A.A., BROMLEY, R.G. & PEMBERTON, S.G.1984. Ichnology. Trace Fossils in Sedimentologyand Stratigraphy. SEPM Short Course, 15.
ETTER, W. 1995. Benthic diversity patterns in oxyge-nation gradients: an example from the MiddleJurassic of Switzerland. Lethaia, 28, 259–270.
ETTER, W. 1996. Pseudoplanktonic and bethicinvertebrates in the Middle Jurassic OpalinumClay, northern Switzerland. Palaeogeography,Palaeoclimatology, Palaeoecology, 126, 325–341.
FOLLMI, K.B & GRIMM, K.A. 1990. Doomed pioneers:gravity-flow deposition and bioturbation in marineoxygen-deficient environments. Geology, 18,1069–1072.
FRAAYE, R, & JAGER, M. 1995. Decapods in ammoniteshells: examples of inquilinism from the Jurassic ofEngland and Germany. Palaeontology, 38, 63–75.
GASPARINI, Z., SPALLETTI, L. & DE LA FUENTE, M.1997. Tithonian marine reptiles of the westernNeuquen Basin, Argentina. Facies and palaeoenvir-onments. Geobios, 30, 701–712.
GASPARINI, Z., SPALLETTI, L., FERNANDEZ, M. &DE LA FUENTE, M. 1999. Tithonian marine reptilesfrom the Neuquen Basin: diversity and paleo-environments. Revue Paleobiologie Geneve, 18,335–345.
GASPARINI, Z., SPALLETTI, L., MATHEOS, S. &FERNANDEZ, M. 2002. Reptiles marinos ypaleoambiente del Jurasico Superior–CretacicoInferior en la Yesera del Tromen (Neuquen, Argen-tina): un caso de estudio. 158 Congreso GeologicoArgentino, Actas, II, 473–478.
GLENN, C.R. & ARTHUR, M.A. 1990. Anatomy andorigin of a Cretaceous phosphorite–green sandgiant, Egypt. Sedimentology, 37, 123–148.
GOLDRING, R. 1995. Organisms and the substrate:response and effects. In: BOSENCE, D.W. &ALLISON, P.A. (eds), Marine PalaeoenvironmentalAnalysis from Fossils. Geological Society, London,Special Publications 83, 151–180.
GULISANO, C.A. & GUTIERREZ PLEIMLING, A.R.1994. The Jurassic of the Neuquen Basin,
Neuquen Province. Asociacion GeologicaArgentina Series, E2.
GULISANO, C.A., GUTIERREZ PLEIMLING, A.R. &DIGREGORIO, R.E. 1984. Analisis estratigraficodel intervalo Tithoniano-Valanginiano (Forma-ciones Vaca Muerta, Quintuco y Mulichinco) enel Suroeste de la Provincia del Neuquen. NovenoCongreso Geologico Argentino, Actas, 1,221–235.
HALLAM, A. 1991. Relative importance of regionaltectonics and eustacy for the Mesozoic of theAndes. In: MACDONALD, D.I.M. (ed.) Sea LevelChanges at Active Plate Margins: Processand Product. International Association ofSedimentologists, Special Publications, 12,189–200.
HAQ, B.U., HARDENBOL, J. & VAIL, P.R. 1987. Chron-ology of fluctuating sea levels since the Triassic.Science, 235, 1156–1167.
KAUFFMAN, E.G. 1981. Ecological reappraisal of theGerman Posidonienschiefer (Toarcian) and thestagnant basin model. In: GRAY, J., BOUCOT, A.J.& BERRY, W.B.N. (eds) Communities of the Past.Hutchinson Ross, Stroudsberg, 311–381.
KAUFFMAN, E.G. 1982. The community structure of‘shell islands’ on oxygen-depleted substratesin dark shales and laminated carbonates. In:EINSELE, G. & SEILACHER, A. (eds) Cyclic andEvent Stratigraphy. Springer, Berlin, 502–503.
LEANZA, H.A. 1973. Estudio sobre los cambiosfaciales de los Estratos limıtrofes Jurasico-Cretacicos entre Loncopue y Picun Leufu,Provincia de Neuquen, Republica Argentina.Asociacion Geologica Argentina, Revista, 28,97–132.
LEANZA, H.A. 1981. Faunas de ammonites del Jurasicosuperior y del Cretacico inferior de America delSur, con especial consideracion de la Argentina.Cuencas Sedimentarias del Jurasico y Cretacicode America del Sur, 2, 559–597.
LEANZA, H.A. & HUGO, C.A. 1977. Sucesion deammonites y edad de la Formacion Vaca Muertay sincronicas entre paralelos 358 y 408 L.S.,Cuenca Neuquina–Mendocina. AsociacionGeologica Argentina, Revista, 32, 248–264.
LEANZA, H.A. & ZEISS, A. 1990. Upper Jurassiclithographic limestones from Argentina (NeuquenBasin): Stratigraphy and fossils. Facies, 22,169–186.
LEANZA, H.A., MARCHESE, H. & RIGGI, J.C. 1977.Estratigrafıa del Grupo Mendoza con especialreferencia a la Formacion Vaca Muerta entre losparalelos 358 y 408 LS, Cuenca neuquina-mendocina. Asociacion Geologica Argentina,Revista, 32, 190–208.
LEGARRETA, L. & KOZLOWSKI, E. 1981. Estratigrafıay sedimentologıa de la Formacion Chachao, Pro-vincia de Mendoza. Octavo Congreso GeologicoArgentino, Actas, 2, 521–543.
LEGARRETA, L. & GULISANO, C.A. 1989. Analisisestratigrafico secuencial de la Cuenca Neuquina(Triasico superior–Terciario inferior, Argentina).In: CHEBLI, G. & SPALLETTI, L.A. (eds) CuencasSedimentarias Argentinas, Serie Correlacion
P. DOYLE ET AL.204

Geologica. Universidad Nacional de Tucuman, 6,221–243.
LEGARRETA, L. & ULIANA, M.A. 1991. Jurassic–Cretaceous marine oscillations and geometry ofback arc basin fill, Central Argentine Andes. In:MACDONALD, D.I.M. (ed.) Sea Level Changes atActive Plate Margins: Process and Product. Inter-national Association of Sedimentologists, SpecialPublications 12, 429–450.
LEGARRETA, L. & ULIANA, M.A. 1996. La sucesionjurasica en el centro-oeste de Argentina. Arregloestratigrafico, secuencias y evolucion paleogeogra-fica. Boletın de Informaciones Petroleras, 45,66–78.
LEGARRETA, L., GULISANO, C.A. & ULIANA, M.A.1993. Las secuencias sedimentarias jurasico-creta-cicas. Relatorio Geologıa y Recursos Naturales deMendoza, XII8 Congreso Geologico Argentino y II8Congreso de Exploracion de Hidrocarburos, 1,87–114.
LEGARRETA, L., KOZLOWSKI, E. & BOLL, A. 1981.Esquema estratigrafico y distribucion de faciesdel Grupo Mendoza en el ambito submendocinode la Ceunca Neuquina, Argentina. Octavo Con-greso Geologico Argentino, Actas, 3, 389–409.
LEVINTON, J.S. 1970. The paleoecological significanceof opportunistic species. Lethaia, 3, 69–78.
MACELLARI, C.E. 1984. Revision of serpulids of thegenus Rotularia (Annelida) at Seymour Island(Antarctic Peninsula) and their value in strati-graphy. Journal of Paleontology, 58, 1098–1116.
MIDTGAARD, H.H. 1996. Inner-shelf to lower-shore-face hummocky sandstone bodies with evidencefor geostrophic influenced combined flow,Lower Cretaceous, West Greenland. Journal ofSedimentary Research, 66, 343–353.
MITCHUM, R. & ULIANA, M. 1985. Seismic stratigra-phy of carbonate depositional sequences, UpperJurassic–Lower Cretaceous, Neuquen Basin,Argentina. In: BERG, B.R. & WOOLVERTON, D.G.(eds) Seismic Stratigraphy II: An IntegratedApproach to Hydrocarbon Analysis. AAPGMemoirs, 39, 255–274.
MOMBRU, C.A., ULIANA, M.A. & BERCOWSKI, F.1978. Estratigrafia y sedimentologıa de las acumu-laciones biocarbonaticas del Cretacico inferiorsurmendocino. Septimo Congreso GeologicoArgentino, Actas, 1, 685–700.
MORRIS, K.A. 1980. Comparison of major sequencesof organic-rich mud deposition in the British Juras-sic. Journal of the Geological Society, London,137, 157–170.
MYROW, P.M. & SOUTHHARD, J.B. 1991. Combinedflow model for vertical stratification sequences inshallow marine storm-deposited beds. Journal ofSedimentary Petrology, 61, 202–210.
MYROW, P.M. & SOUTHHARD, J.B. 1996. Tempestitedeposition. Journal of Sedimentary Research, 66,875–887.
RHOADS, D.C. & MORSE, J.M. 1971. Evolutionary andecologic significance of oxygen-deficient marinebasins. Lethaia, 4, 413–428.
RICCARDI, A. 1983. The Jurassic of Argentina andChile. In: MOULLADE, M. & NAIRN, A.E.M. (eds)
The Phanerozoic Geology of the World II, Meso-zoic, Vol. B8. Elsevier, Amsterdam, 201–263.
RICCARDI, A. 1988. The Cretaceous System ofSouthern South America. Geological Society ofAmerica Memoirs, 168, 1–161.
SAGEMAN, B.B. & BINA, C.R. 1997. Diversity andspecies abundance patterns in Late Cenomanianblack shale biofacies, Western Interior, U.S.Palaios, 12, 449–466.
SAVRDA, C.E. 1995. Ichological applications in paleo-ceanographic, paleoclimatic and sea-level studies.Palaios, 10, 565–577.
SAVRDA, C.E. & BOTTJER, D.J. 1986. Trace-fossilmodel for reconstruction of paleo-oxygenation inbottom waters. Geology, 14, 3–6.
SAVRDA, C.E. & BOTTJER, D.J. 1987. The exaerobiczone, a new oxygen-related marine biofacies.Nature, 327, 54–56.
SAVRDA, C.E. & BOTTJER, D.J. 1989. Anatomy andimplications of bioturbated beds in ‘black shale’sequences: examples from the Jurassic Posido-nienschiefer (Southern Germany). Palaios, 4,330–342.
SEILACHER, A. 1960. Epizoans as a key to ammoniteecology. Journal of Paleontology, 34, 189–193.
SEILACHER, A. 1982a. Ammonites as habitats in thePosidonia Shales of Holzmaden – floats orbenthic islands? Neues Jahrbuch fur Geologieund Palaontologie, Monatshefte, 2, 98–114.
SEILACHER, A. 1982b. Posidonia shales (Toarcian,S. Germany) – stagnant basin model revalidated.In: GALLITELLI, E.M. (ed.) Palaeontology, Essen-tial of Historical Geology, Modena Press,Modena, 25–55.
SEILACHER, A., ANDALIB, F., DIETL, G. & GOCHT, H.1976. Preservational history of compressed Juras-sic ammonites from southern Germany. NeuesJahrbuch fur Geologie und PalaontologieAbhandlungen, 152, 307–356.
SPALLETTI, L.A., FRANZESE, J.R., MATHEOS, S.D. &SCHWARZ, E. 2000. Sequence stratigraphy of atidaly-dominated carbonate-siliciclastic ramp; theTthonian of the southern Neuquen Basin, Argen-tina. Journal of the Geological Society, London,157, 433–446.
SPALLETTI, L.A., GASPARINI, Z., VEIGA, G.,SCHWARZ, E., FERNANDEZ, M. & MATHEOS, S.1999. Facies anoxicas, procesos deposicionales yherpetofauna de la rampa marina titoniano-ber-riasiana en la Cuenca Neuquina (Yesera delTromen), Neuquen, Argentina. Revista Geologicade Chile, 26, 109–123.
SPALLETTI, L.A., POIRE, D.G., SCHWARZ, E. &VEIGA, G.D. 2001. Sedimentologic and sequencestratigraphic model of a neocomian marine carbon-ate siliciclastic ramp: Neuquen Basin, Argentina.Journal of South American Earth Sciences, 14,609–624.
THAYER, C.W. 1975. Morphologic adaptations ofbenthic invertebrates to soft substrates. Journal ofMarine Research, 33, 177–189.
WEAVER, V. 1931. Paleontology of the Jurassic andCretaceous Central Argentina. University ofWashington, Memoirs, 1, 1–496.
RELATIVE OXYGENATION MENDOZA SHELF 205

WIGNALL, P.B. 1990. Benthic Palaeoecology of theLate Jurassic Kimmeridge Clay of England.Special Papers in Palaeontology, 43, 1–74.
WIGNALL, P.B. 1993. Distinguishing between oxygen andsubstrate control in fossil benthic communities. Journalof the Geological Society, London, 150, 193–196.
WIGNALL, P.B. 1994. Black Shales. Oxford Mono-graphs on Geology and Geophysics, 30. ClarendonPress, Oxford.
WIGNALL, P.B. & HALLAM, A. 1991. Biofacies,stratigraphic distribution and depositional modelsof British onshore Jurassic black shales. In:TYSON, R.V. & PEARSON, T.H. (eds) Modern andAncient Continental Shelf Anoxia. GeologicalSociety, London, Special Publications, 58,291–309.
WIGNALL, P.B. & SIMMS, M.J. 1990. Pseudoplankton.Palaeontology, 33, 359–378.
P. DOYLE ET AL.206
Copyright © 2022 FDOKUMEN