
A dynamic concept for eastern Mediterranean circulation and oxygenation …
17
A dynamic concept for eastern Mediterranean circulation and oxygenation during sapropel formation J.S.L. Casford a;g; , E.J. Rohling a , R.H. Abu-Zied b , C. Fontanier c , F.J. Jorissen d , M.J. Leng e , G. Schmiedl f , J. Thomson a a Southampton Oceanography Centre (SOC), European Way, Southampton, SO14 3ZH, UK b Geology Department, Faculty of Science, El-Mansoura University, El-Mansoura 35516, Egypt c De ¤partement de Ge ¤ologie et Oce ¤anographie, l’Universite ¤ Bordeaux, 33405 Talence Cedex, France d Laboratoire de Ge ¤ologie, Fac. Sciences, Universite ¤ d’Angers, 49045 Angers Cedex, France e NERC Isotope Geoscience Laboratory (NIGL), Keyworth, Nottingham, UK f Universita «t Leipzig, Institut fu «r Geophysik und Geologie, TalstraMe 35, 04103 Leipzig, Germany g University of Durham, Science site, South Road, Durham, UK Received 19 March 2002; received in revised form 26 June 2002; accepted 18 October 2002 Abstract We propose that intermittent bottom water ventilation occurred throughout periods of sapropel deposition, and that the recently reported sapropel ‘interruptions’ represent centennial-scale episodes of enhanced frequency/intensity of that process. In essence, the modern high-frequency variability in deep water formation (affected by climatic variability over the northern basins on seasonal to longer time scales) prevailed also at times of sapropel deposition, although the overall ventilation state was much reduced. This concept is supported by: detailed multiple-species isotope records for three Aegean cores; the presence of abundant Globorotalia truncatulinoides within especially sapropels S7 and S8 in the western Levantine basin ; observations of three rapid benthic repopulations within sapropel S6 in the deep western Levantine basin; a report of continuous benthic presence through sapropel S1 at intermediate- deep locations offshore Libya; and further supporting information from the literature. In the Aegean records, concomitant abundance of low-oxygen tolerant benthic foraminifera and presence of the more oxyphilic benthic foraminifer Uvigerina mediterranea, with surface-similar N 13 C values, indicate repeated deep water re-oxygenation events throughout the deposition of S1. The observations of a continuous benthic presence through S1 (offshore from Libya) imply that no persistent anoxia developed at mid-depth levels in that region, which is far removed from direct deep ventilation influences. The abundance of deep mesopelagic G. truncatulinoides through several sapropels from the western Levantine basin also suggests the presence of bio-available oxygen at many hundreds of meters of depth. Moreover, the rapid/intermittent benthic repopulations within sapropels from the deep eastern Mediterranean imply that bottom water anoxia was spatially restricted and/or of a highly intermittent nature. The short time scales of these repopulation events are incompatible with titration of an extensively anoxic water column and subsequent re- establishment of water-column anoxia. We suggest that where anoxic/azoic conditions were present, they most likely were restricted to a veneer at the sediment/water interface. The extent of such an anoxic ‘blanket’ depends on the balance between advective oxygen supply into the deep sea, and biological and chemical oxygen demand. The demand functions imply a decoupling of oxygenation from water mass advection, allowing export production and C org posting 0031-0182 / 02 / $ ^ see front matter ȣ 2002 Elsevier Science B.V. All rights reserved. PII:S0031-0182(02)00601-6 * Corresponding author. E-mail address: [email protected] (J.S.L. Casford). PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 www.elsevier.com/locate/palaeo
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A dynamic concept for eastern Mediterranean circulation and oxygenation …

A dynamic concept for eastern Mediterranean circulation andoxygenation during sapropel formation
J.S.L. Casford a;g;�, E.J. Rohling a, R.H. Abu-Zied b, C. Fontanier c,F.J. Jorissen d, M.J. Leng e, G. Schmiedl f , J. Thomson a
a Southampton Oceanography Centre (SOC), European Way, Southampton, SO14 3ZH, UKb Geology Department, Faculty of Science, El-Mansoura University, El-Mansoura 35516, Egypt
c De¤partement de Ge¤ologie et Oce¤anographie, l’Universite¤ Bordeaux, 33405 Talence Cedex, Franced Laboratoire de Ge¤ologie, Fac. Sciences, Universite¤ d’Angers, 49045 Angers Cedex, France
e NERC Isotope Geoscience Laboratory (NIGL), Keyworth, Nottingham, UKf Universita«t Leipzig, Institut fu«r Geophysik und Geologie, TalstraMe 35, 04103 Leipzig, Germany
g University of Durham, Science site, South Road, Durham, UK
Received 19 March 2002; received in revised form 26 June 2002; accepted 18 October 2002
Abstract
We propose that intermittent bottom water ventilation occurred throughout periods of sapropel deposition, andthat the recently reported sapropel ‘interruptions’ represent centennial-scale episodes of enhanced frequency/intensityof that process. In essence, the modern high-frequency variability in deep water formation (affected by climaticvariability over the northern basins on seasonal to longer time scales) prevailed also at times of sapropel deposition,although the overall ventilation state was much reduced. This concept is supported by: detailed multiple-speciesisotope records for three Aegean cores; the presence of abundant Globorotalia truncatulinoides within especiallysapropels S7 and S8 in the western Levantine basin; observations of three rapid benthic repopulations within sapropelS6 in the deep western Levantine basin; a report of continuous benthic presence through sapropel S1 at intermediate-deep locations offshore Libya; and further supporting information from the literature. In the Aegean records,concomitant abundance of low-oxygen tolerant benthic foraminifera and presence of the more oxyphilic benthicforaminifer Uvigerina mediterranea, with surface-similar N
13C values, indicate repeated deep water re-oxygenationevents throughout the deposition of S1. The observations of a continuous benthic presence through S1 (offshore fromLibya) imply that no persistent anoxia developed at mid-depth levels in that region, which is far removed from directdeep ventilation influences. The abundance of deep mesopelagic G. truncatulinoides through several sapropels from thewestern Levantine basin also suggests the presence of bio-available oxygen at many hundreds of meters of depth.Moreover, the rapid/intermittent benthic repopulations within sapropels from the deep eastern Mediterranean implythat bottom water anoxia was spatially restricted and/or of a highly intermittent nature. The short time scales of theserepopulation events are incompatible with titration of an extensively anoxic water column and subsequent re-establishment of water-column anoxia. We suggest that where anoxic/azoic conditions were present, they most likelywere restricted to a veneer at the sediment/water interface. The extent of such an anoxic ‘blanket’ depends on thebalance between advective oxygen supply into the deep sea, and biological and chemical oxygen demand. The demandfunctions imply a decoupling of oxygenation from water mass advection, allowing export production and Corg posting
0031-0182 / 02 / $ ^ see front matter > 2002 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 0 2 ) 0 0 6 0 1 - 6
* Corresponding author. E-mail address: [email protected] (J.S.L. Casford).
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119
www.elsevier.com/locate/palaeo

rates to the sea floor to delimit the extent of the anoxic blanket in both space and time. Low-productivity regionswould develop no anoxic blanket, allowing for the observed persistence of deep dwelling planktonic and bottomdwelling benthic faunas.> 2002 Elsevier Science B.V. All rights reserved.
Keywords: eastern Mediterranean; sapropel S1; benthic foraminifera
1. Introduction
The Mediterranean Sea is of particular interestto palaeoceanographers, as it possesses its ownthermohaline circulation with deep water beingformed in the northern basins (Bethoux et al.,1999). In the eastern Mediterranean, the AdriaticSea and the Aegean Sea are subject to orograph-ically channelled cold, dry air from high latitudes(Mariolopoulos, 1961; Poulos et al., 1997) similarto the mistral that a¡ects the western Mediterra-nean (Leaman and Schott, 1991). This drives deepoverturn that is known to respond rapidly to cli-matic variability in both the modern (Theocharis,1989; Roether et al., 1996) and palaeorecords(Casford et al., 2001; Rohling et al., 2002). Wepresent preliminary observations from RV Meteorcruise M51-3 (12 November^11 December 2001)and data from Aegean cores SL-11, SL-21 andSLA-9 (Fig. 1) that suggest that this high-fre-quency variability may also ¢nd expression duringtimes of sapropel formation.
Ever since the ¢rst discovery of sapropels ineastern Mediterranean sediments (Kullenberg,1952) they have been a source of controversy.Although the association between sapropel forma-tion and insolation-driven monsoon maxima iswell established (Rossignol-Strick, 1983, 1985),the role of anoxia has become a central issue inthe characterisation of processes that underliesapropel formation. Opinions are divided aboutthe relative importance of two possible processes,either a reduction in ventilation leading to anoxiaand hence organic carbon preservation (Olauson,1961; Cita et al., 1977; Vergnaud-Grazzini et al.,1977; Cita and Grignani, 1982; Vergnaud-Graz-zini, 1985), or an increase in primary productivity(DeLange and Ten Haven, 1983; Calvert, 1983;Boyle and Lea, 1989; Pederson and Calvert,1990; Van Os et al., 1994). Clearly, these scenar-
ios are not mutually exclusive (see Rohling andGieskes, 1989), as an increase in productivitywould enhance export of organic carbon (Corg)to the sediment surface and could utilise all theavailable oxygen, producing anything from dysox-ia to anoxia. Increased freshwater in£ux wouldenhance nutrient concentrations in the basin thatwould in turn enhance productivity, especiallywhen there appears to be a potential build-up ofnutrients over s 1000 years before utilisation(Casford et al., 2002). Observations of high pollenconcentrations (Cheddadi and Rossignol-Strick,1995) provide strong arguments that Corg preser-vation was much improved during sapropel times,compared with the present, and have been used tosuggest extensive anoxia. The upper depth limit ofsapropel occurrence at V300 m in the open east-ern Mediterranean (Rohling and Gieskes, 1989)and V125 m in the Aegean Sea (Perissoratisand Piper, 1992), has been used to suggest thatanoxia prevailed throughout the water column upto this depth. However, there are no observationsto eliminate the possibility that true anoxia onlydeveloped as a blanket limited to the sediment/water interface. Indications for this ‘Blanket Hy-pothesis’ have been seen in Pliocene sapropel C2from the Singa section of southern Italy (Rohlinget al., 1993). Calvert (1983) implies that low Corg
sapropels (i.e. those with 9 2% Corg) such as theHolocene S1 could be produced, without signi¢-cant increases in primary productivity, i.e. with-out the development of a Deep Chlorophyll Max-imum (DCM) (Rohling, 1994). Mercone et al.(2001) performed a combined micropalaeontolog-ical and geochemical study and infer that thepresence of an Oxygen De¢ciency Stress (ODS,sensu Rohling et al., 1997) assemblage of benthicforaminifera throughout S1 indicates either inter-mittent anoxia or continuous dysoxia. They fur-ther argue that uncertainties in the mechanism of
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119104

Mediterranean Sea
Location of Core
Rhodes
Crete
Cretan Sea
Mount Athos Basin
Greece
Turkey
Black Sea
Fig. 1. Map of the Mediterranean Sea showing locations of the cores.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 105

barium uptake in the water column and dissolu-tion processes leave the use of barium as an in-dicator of palaeoproductivity in S1 open to ques-tion. We here o¡er new observations within thecontext of previous work to assess whether, attimes of sapropel deposition, deep water forma-tion had ceased (or not) and to what extent dys-oxic and/or anoxic conditions prevailed. To ex-plain these observations, a new dynamic conceptof ventilation and oxygenation is presented.
2. Methods
We present: (1) initial observations from coringon RV Meteor cruise M51-3; (2) results from Ae-gean gravity cores SL-11 and SL-21 (North Ae-gean) and SLA-9 (South Aegean), which comple-ment previous work on southeast Aegean coreLC21 (Mercone et al., 2001; Casford et al.,2001; Rohling et al., 2002); (3) benthic foraminif-eral abundance data in sapropel S1 from multi-core 562MC o¡ the Gulf of Sirte; and (4) benthicforaminiferal abundance data from sapropel S6 ofODP Hole 971A in the open Mediterranean.These results are placed within the context of ob-servations from the literature. All cores investi-gated here consist of microfossil-rich hemipelagic
ooze, with clearly de¢ned darker bands of sapro-pel material. Cores SL-11, SL-21, SLA-9, and S6from ODP Hole 971A were sampled in continu-ous sequences of 0.5-cm intervals. Given the highsedimentation rates of our cores the 0.5-cm sam-ple interval gives a temporal resolution of between40 and 75 years (Table 1). Multi-core 562MC wassampled at a 1-cm resolution. These samples weredried, weighed and selected (weighed) subsampleswere disaggregated and wet sieved using demi-neralised water. The sieved fractions were col-lected on 600-, 150-, 125- and 63-Wm mesh sizes.The s 150-Wm fractions were subdivided using arandom splitter to provide an aliquot of about200 individual planktonic foraminifera (s 300for benthics). For multi-core 562MC benthic fau-na was analysed on the s 63-Wm fraction andbetween 50 and 100 individuals were countedwhere present. After sorting and counting, resultswere expressed as numbers g31 and percentages(see Abu-Zied, 2001; Casford et al., 2002 for theAegean faunal data). During RV Meteor cruiseM51-3, core SL97 was sampled to provide an ini-tial biostratigraphy. Several samples were takenfrom each sapropel and sieved over 150-Wmmesh. The faunas were qualitatively assessed onboard, using a binocular light microscope.
Detailed monospeci¢c stable oxygen and car-
Table 1Core location and type
Core Type of core Length Depth below sea level Sedimentation rates Co-ordinates(cm) (m) (cm kyr31)
LC-21a Piston ^ 1522 13.7 35‡40PN, 26‡35PESL-11 Gravity 209 258 9.5 39‡06PN, 25‡48PESL-21 Gravity 273 317 6.8 39‡01PN, 25‡25PESLA-9 Gravity 286 260 12.6 37‡31PN, 24‡33PE971A ODP 5880 2026 ^ 33‡42PN, 24‡43PESL97 Gravity 640 1879 ^ 33‡43PN, 23‡30PE562MC Multi-core 27 1390 ^ 32‡46PN, 19‡11PE
a Hayes et al. (1999); De Rijk et al. (1999); Mercone et al. (2000, 2001); Casford et al. (2001); Rohling et al. (2002).Sedimentation rates are derived from the GISP II tuned time framework of LC-21 and from Casford et al. (2002).
Fig. 2. Oxygen isotope data from cores SL-11, SL-21 and SLA-9. The main features of these records are summarised in the bot-tom panel. A solid bar indicates the extent of the sapropel. Explanation: (1) in£ection before the depletion into the Holocenefrom the Younger Dryas (Y.D.) as shown in the record of N
18OGlobigerinoides ruber ; (2) enrichment trend in the Neogloboquadrina pa-chyderma record and the lag in response of N
18ON: pachyderma compared with that of N18OG: ruber ; (3) in£ection in the benthic N
18Orecord, coinciding with the in£ection in N
18OG: ruber.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119106

PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 107

Fig. 3. Carbon isotope data from cores SL-11, SL-21 and SLA-9 showing the marked depletion in N13C across sapropel S1. The
main features of these records are summarised in the bottom panel with the extent of the sapropel indicated by a solid bar.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119108

bon isotope records were constructed for coresSL-11, SL-21 and SLA-9 with resolutions in theorder of 1 cm (Figs. 2 and 3). These were derivedfrom ‘alternate’ 0.5-cm samples and based onanalyses of at least 10 individuals for each chosenspecies per sample. These chosen species were: theepipelagic (6 50-m) species Globigerinoides ruber ;the mesopelagic species Neogloboquadrina pachy-derma (dextral) which thrives in the DCM nearthe base of the euphotic layer; and the epifau-nal/shallow infaunal benthics Uvigerina mediterra-nea (SL-21) and Uvigerina peregrina (SLA-9). Incore SLA-9 we also analysed the low-oxygen tol-erant benthic species Globobulimina a⁄nis, whichunder oxygenated conditions occupies a deep in-faunal habitat, at the Redox boundary. The selec-tion of planktonic and benthic species followsglobal and speci¢c Mediterranean habitat summa-ries in Hemleben et al. (1989), Rohling andGieskes (1989), Jorissen et al. (1993), Pujol andVergnaud-Grazzini (1995), Rohling et al. (1993,1995, 1997), De Rijk et al. (1999), Hayes et al.(1999), Jorissen (1999), Reiss et al. (2000), andAbu-Zied (2001). The analyses were performedat two separate inter-calibrated facilities : theEuropa Geo 20-20, with individual acid bathpreparation at the SOC, and the VG-Optimawith a common acid bath preparation at theNIGL. Isotope results are reported as x, stan-dardised to Vienna Pee Dee Belemnite. Externalprecision is in the order of 6 0.06x (std).
3. Observations
3.1. High-frequency events in the Aegean Sea
The fact that it is possible to pick mature, cleanUvigerina mediterranea in su⁄cient numbers fromall samples of cores SL-11 and SL-21 at a 1-cmresolution indicates that it occurs systematically(if rare) throughout S1 at this locale. This cannotbe explained by downslope transport since thisspecies was not found to be systematically associ-ated with other shallow water (sensu De Rijk etal., 1999) faunal elements (Abu-Zied, 2001). Inaddition, the ridge location of the core sitesmake downslope transport less likely. Detailed
abundance counts of benthic foraminifera in coresSL-31, SLA-9 and LC-21 show that U. mediterra-nea does not correlate with the known low-oxygentolerant ODS group (Abu-Zied, 2001) and thepresence of this group in itself suggests that, inthe Aegean at least, deep waters never becamepersistently anoxic during S1.
The oxygen isotope records (Fig. 2) show ob-vious depletions into the Holocene, shifting about32x in all species. This re£ects a combinationof global ice-volume reduction and changing re-gional climatic conditions as the eastern Mediter-ranean became warmer and more humid into theHolocene Climatic Optimum following the cooler,drier Younger Dryas (Rohling, 1999b). A clearo¡set can be seen, however, with the N
18O of theepipelagic species Globigerinoides ruber showingmore depleted values than the mesopelagic N. pa-chyderma, both in absolute values and generaltrends. The N
18O values of all benthic foraminif-era are similar to, or more enriched than,N
18ONeogloboquadrina pachyderma. This can be explainedin terms of the lower temperatures and highersalinities that characterise subsurface and deepwater, relative to the surface mixed layer. Signi¢-cantly, N
18OU : mediterranea (in cores SL-11 and SL-21) retains the heavy deep water values but in-£ects to lower values after the Younger Dryas atthe same time as the surface-water record ofG. ruber. This contrasts markedly with the laggedresponse of N
18ON: pachyderma that suggests an iso-lation of intermediate waters from surface pro-cesses (Casford et al., 2002). The combined dataportray a direct link between responses to region-al freshwater input in both surface and deepestwaters, which is not shared at intermediate levels.We view this as a re£ection of the proximity ofour cores to an area of active deep water forma-tion, with bottom waters penetrating intermit-tently below intermediate waters that continu-ously advect into the Aegean from a remotesource area with di¡erent N
18O characteristics(Casford et al., 2002).
The carbon isotope data (Fig. 3) allow furtherinsight into the habitat separations between spe-cies. The general N13C trend shows relatively con-stant values before and after periods of sapropeldeposition, with depletion in N
13C values over the
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 109
% Oxyphyllic% ODSbenthics g -1
10
9
8
7
6
5
4
3
2
1
% ODS % Oxyphyllic Numbers
30
20
10
0
% ODS
0 40 80 0 50 100 0 50 100 0 50 100 0 1 2 3 0 50 100 0 20 40 60 0 20 40 60
% Oxyphyllic
0 200 400 600
benthics g-1
LC-21 SLA-9 562MC
Tim
e (
un
ca
l ra
dio
ca
rbo
n c
on
ve
ntio
n k
y B
P)
De
pth
in
56
2M
C (
cm
)
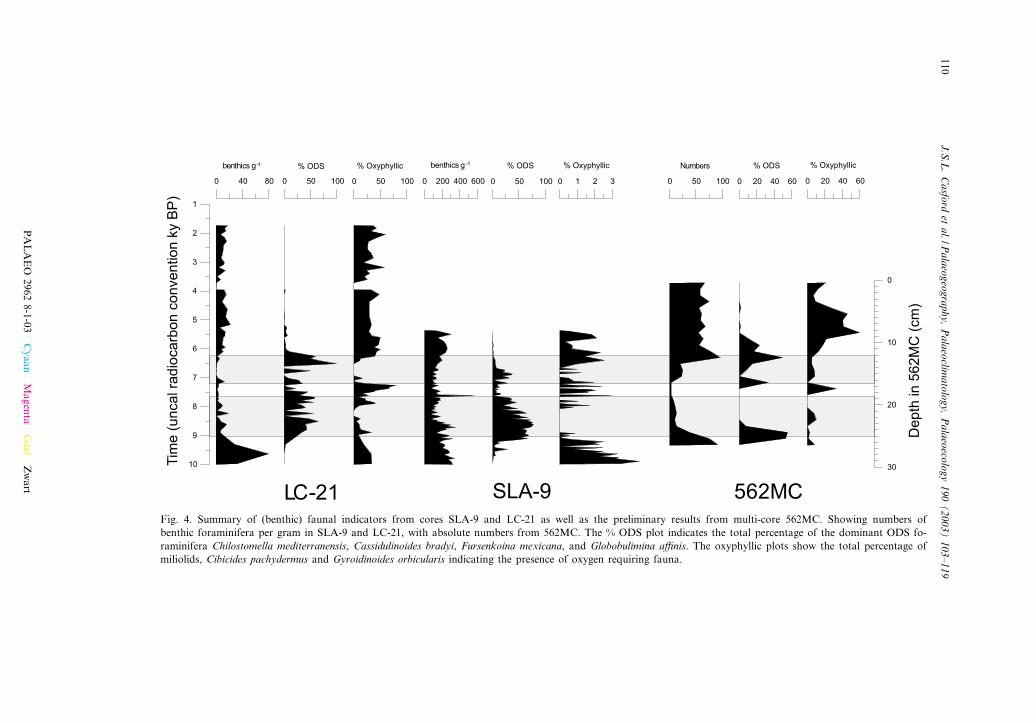
Fig. 4. Summary of (benthic) faunal indicators from cores SLA-9 and LC-21 as well as the preliminary results from multi-core 562MC. Showing numbers ofbenthic foraminifera per gram in SLA-9 and LC-21, with absolute numbers from 562MC. The % ODS plot indicates the total percentage of the dominant ODS fo-raminifera Chilostomella mediterranensis, Cassidulinoides bradyi, Fursenkoina mexicana, and Globobulimina a⁄nis. The oxyphyllic plots show the total percentage ofmiliolids, Cibicides pachydermus and Gyroidinoides orbicularis indicating the presence of oxygen requiring fauna.
PA
LA
EO
29628-1-03
Cyaan
Magenta
Geel
Zw
art
J.S.L.Casford
etal./P
alaeogeography,Palaeoclim
atology,Palaeoecology
190(2003)
103^119110

sapropel itself. The magnitude of this depletion isgreater in the mesopelagic Neogloboquadrina pa-chyderma than in the surface species Globigeri-noides ruber. The benthic species Globobuliminaa⁄nis and Uvigerina peregrina in SLA-9 (south-west Aegean) show an even stronger depletionthan N. pachyderma. Since 12C is preferentiallytaken up during photosynthesis, surface-waterDissolved Inorganic Carbon (DIC) attains rela-tively high N
13C values, while remineralisationcauses relatively light N
13CDIC values at depth.Hence, benthic and mesopelagic species wouldnormally be expected to show lighter N
13C valuesthan epipelagic species. Vital e¡ects might ac-count for constant o¡sets between the variousbenthic and deep planktonic species but the gen-eral trends in their records should be similar. Par-adoxically, the N
13C U : mediterranea signal in thenorthern Aegean cores SL-11 and SL-21 mimicsboth the trend and values of the epipelagic speciesG. ruber (Fig. 3). Note that SL-11 and SL-21 areboth from locations adjacent to an area of deepwater production (the Mount Athos basin) andare sited in relatively shallow water (Table 1).Hence, a likely explanation for the anomalousN
13CU : mediterranea signal is that Uvigerina mediterra-nea lived in newly advected deep water, with acarbon isotope signal that has very recently been‘set’ according to air^sea equilibrium, but retain-ing an oxygen isotope signal that re£ects the (cooland saline) deep water signature (Fig. 2). Thecombination of abundant ODS species (with car-bon isotope trends resembling that of N. pachy-derma) alongside U. mediterranea, with an ab-sence of comprehensive repopulation by otherspecies, suggests that advection events were inter-mittent and short-lived phenomena.
3.2. Centennial-scale sapropel interruptions
‘Interruptions’ of centennial-scale durationwithin sapropels have been reported from acrossthe Mediterranean, with examples from the Holo-cene to the Pliocene (e.g. Van Straaten, 1966,1972; Thunell et al., 1977; Stanley, 1978; Rossig-nol-Strick et al., 1982; Cita et al., 1984; Vismara-Schilling, 1984; Perissoratis and Piper, 1992;Rohling et al., 1993, 1997; Mercone et al.,
2001). It is important to establish the relationshipbetween the conjectured high-frequency advectionevents (above) and the longer-term ventilationchanges that caused these sapropel interruptions.
First of all, we note that the centennial-scale‘interruption’ of S1 in the Aegean core LC-21shows rapid benthic repopulation, from one sam-ple to the next (i.e. within V50 years; Fig. 4).SLA-9 also shows these marked shifts, alternatingbetween ODS species and less low-oxygen tolerantopportunists (Abu-Zied, 2001; Mercone et al.,2001); this is also the case in the Adriatic (Joris-sen et al., 1993; Rohling et al., 1997; Mercone etal., 2000). Here, we show similar rapid benthicrepopulations, bound by azoic intervals, withinS6 (Fig. 5). This is a signi¢cant observation be-cause the studied S6 comes from ODP Hole 971A,taken at a deep location (V2026 m) in the openeastern Mediterranean, rather than within a deepwater source region like the Adriatic or Aegean.Clearly deep ventilation improved su⁄ciently toallow the presence of bio-available oxygen at thesea £oor at this very deep site in the open basin,and it happened both repeatedly and abruptly.
Detailed study of the interruption in S1 haslinked it to climatic processes. Its benthic forami-niferal repopulations are indicative of re-oxygen-ation, related to improved deep water formationand while the entire interval lasted several centu-ries, its onset was very abrupt, within 50 years(Mercone et al., 2001; Rohling et al., 2002). Thereventilations were found to be related to ‘coldspells’ over northern basins of the Mediterranean(Rohling et al., 1997, 2002; Casford et al., 2001;Mercone et al., 2001), which are considered to beperiods of increased frequency or intensity of out-breaks of polar/continental northerly air massesfrom higher latitudes (Mariolopoulos, 1961; The-ocharis, 1989; Roether et al., 1996; Poulos et al.,1997). Individual outbreaks are seasonally depen-dent, normally lasting 1^5 days, concentrated inthe winter months, and interannually variable(Saaroni et al., 1996). Intense periods of concen-trated outbreaks can dominate over periods ofweeks to months; a recent episode started endNovember 2001 and lasted into January 2002(see on-board observations RV Meteor cruiseM51-3 for the ¢rst two weeks of the event; Fig.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 111

6), while a previous episode with particularly in-tense northerly cooling over the Aegean occurredin the late 1980s^early 1990s (Theocharis, 1989;Roether et al., 1996). Variations in the intensityand frequency of such outbreaks drive variabilityin the deep water formation rates (Theocharis,1989; Roether et al., 1996). The Holocene historyof centennial-scale cold spells has been linked tothe GISP2 ice-core based proxy for intensity ofthe Siberian High (Rohling et al., 2002). It wouldappear that the cold spells represent a longer-pe-riodicity ‘clustering’ of times with particularlyabundant and intense northerly outbreaks.
3.3. Other ventilation indicators
A key on-board observation from RV Meteorcruise 51-3 relevant to this study concerns thehigh relative abundances of both right and leftcoiling morphotypes of the planktonic foraminiferGloborotalia truncatulinoides, which we found in
several sapropels of core SL97 from the westernLevantine basin (especially S7 and S8). This spe-cies has been observed in the deepest tows in theMediterranean at V300 m (Pujol and Vergnaud-Grazzini, 1995), but is known from oceanic stud-ies to spend signi¢cant parts of its life cycle belowthese levels, down to V1000 m (Hemleben et al.,1989; Lohmann and Schweitzer, 1990). This un-precedented observation of G. truncatulinoidessuggests that the upper water column remainedregularly ventilated to a depth of many hundredsof meters in this region.
A further important observation concerns thepresence of benthic foraminiferal faunas reportedhere, throughout S1 at mid-depths (1391 m) o¡Libya in core 562MC (Table 1; Fig. 4), similar tosuch observations through much of S1 in thesouthern Aegean (Mercone et al., 2001) and theAdriatic (Jorissen et al., 1993; Rohling et al.,1997). Such continuations imply that either inter-mittent ventilation or continuous dysoxia pre-
Fig. 5. Core 971 scores on ¢rst-principle component for the benthic foraminifera abundance distributions through S6 in ODPHole 971A, illustrating three abrupt reventilation events in this deep location (2026 m).
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119112

vailed. The observations o¡ Libya are most sur-prising because of this area’s remote position rel-ative to likely deep ventilation sources.
4. Discussion
Although sample size even in high sedimenta-tion-rate areas limits the temporal resolution ofproxy records, some important conclusions canstill be drawn from the sedimentary record. Theoccurrence of Uvigerina mediterranea throughoutthe S1 in the Northern Aegean suggests that itrepopulated on a regular basis within our sampleresolution. Hence, ventilation must have occurredat least once and possibly several times within thetime span of each sample (V50 years). We in-
ferred that the resemblance of the trends inN
13CU : mediterranea to those of the epipelagic spe-cies Globigerinoides ruber suggests thatN
13CU : mediterranea (Fig. 3) re£ects DIC values thatwere recently ‘set’ at the surface, in agreementwith the core’s proximity to a sensitive key areaof regional deep water production. Hence, inter-mittent ventilation is suggested throughout S1within the Aegean (Fig. 7).
The benthic faunas present through S1 o¡ Lib-ya (Fig. 4) also suggest intermittent ventilation, orcontinuous dysoxia. Similarly, ventilation to adepth of many hundreds of meters is inferredfrom the abundance of Globorotalia truncatuli-noides in the open eastern Mediterranean (coreSL97) during S7 and S8, while discrete but sus-tained reventilation events are even observed in
-180 -90 0 90 180Wind direction (in degrees)
Me
an
air t
em
pe
ratu
re (
in d
eg
ree
s c
elc
ius)
-5
0
5
10
15
20
25
NE EN
Outbreak of cold air during transit of the Aegean sea,
shown in grey.
Fig. 6. Plot of wind direction vs. air temperature illustrating the northery £ow of the cool/continental air outbreak over the Ae-gean during RV Meteor cruise 51-3 (December 2001).
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 113

Subsurface O signature
subsurface C trend(i.e. ODS group/
δ δ
U.peregrina)
18
13
Subsurface O signature
surface similar C(
δ δ
U.mediterranea)
18
13
Anoxic/ azoic blanketSurface water
Intermediate water
Deep water (ODW - Old Deep Water NDW - New Deep Water)
SW
IW
DW
SW
ODW
IW
G.ruber
warm to cool,
high C
SW
δ13
'saline' O,δ18
N.pachyderma
cool
'saline' O
IW
δ18
low Cδ13
cool
'saline' O
ODW
δ18
low C δ13
SW
ODW
IW
Cooling Northerlies
SW
ODW
IW
N.pachyderma
cool
'saline' O
IW
δ18
low Cδ13
cool
'saline' O
ODW
δ18
low C δ13
G.ruber
warm to cool,
high C
SW
δ13
'saline' O,δ18
N.pachyderma
cool
'saline' O
IW
δ18
low Cδ13
cool
'saline' O
ODW
δ18
low C δ13
G.ruber
warm to cool,
high C
SW
δ13
'saline' O,δ18
N S N SN S
ND
Wcool-
'salin
e' δ
18O
, high'surface' δ13C
Fig. 7. Three-stage schematic representation of the inferred con¢guration of water masses before, during and after reventilation in the Aegean Sea. ODW indicatesthe presence of Old Deep Water; NDW represents the advection of New Deep Water.
PA
LA
EO
29628-1-03
Cyaan
Magenta
Geel
Zw
art
J.S.L.Casford
etal./P
alaeogeography,Palaeoclim
atology,Palaeoecology
190(2003)
103^119114

the deep water S6 deposit of ODP Hole 971A(Fig. 5).
While persistent ventilation events in small con-vective regions such as the northern Aegean basinare relatively easy to envision, rapid and sustainedrepopulations in deep open settings are muchmore di⁄cult to explain. Such rapid events wouldrequire abrupt oxygenation of the bottom waters.This cannot be explained by di¡usive oxygentransport, since di¡usive mixing operates ontime scales s 1000 years (V450 years e-foldingdi¡usive time scale (Casford et al., 2002). Conse-quently, the rapid reventilations must have re-sulted from direct advection of newly formed bot-tom waters. This advection must have been strongenough to overcome the titration of any reducedchemical species (e.g. Fe2þ, Mn2þ) both in thewater column and at the sediment/water interface.If the entire water column were anoxic, then avery considerable reservoir of such chemical spe-cies would need to be titrated. In addition, anyBiological Oxygen Demand (BOD) in the watercolumn would need to be overcome, before a ¢-nite amount of bio-available oxygen could bepresent at the sea £oor to sustain viable benthicpopulations.
Using values summarised in Rohling (1994),modern BOD rates alone would require on theorder of 640 years to establish water-column an-oxia below 1500 m if advective oxygen supplyfailed completely. More sophisticated model re-sults (Stratford et al., 2000) suggest that it wouldtake nearer to V1500 years to establish anoxia.Today advective oxygen supply outstrips this de-mand by around ¢ve times (V2.67 mol O2 m33
y31, below 1500 m), so that a re-establishment oftoday’s intensity of advective supply after an an-oxic phase would re-oxygenate the water columnbelow 1500 m as rapidly as 1^2 centuries. This isin agreement with Roether and Well (2001) whocalculate a ¢gure of 150 years to replace existingdeep water. Following the above values for estab-lishment of anoxia and subsequent re-oxygena-tion, it would require around 740^1700 yearswhich far exceeds the rapid variability observed.Note that this ignores the potential build-up ofreduced chemical species in the water column,which would considerably lengthen the calculated
recovery period. BOD and chemical titrationwould remove oxygen from the new deep waterresulting in a spatial separation between the oxy-genation front and the penetration of the newdeep water. Furthermore, these controls will alsovary with the strength of export productivity andconcentration of reduced species (a function ofthe degree of anoxia) both of which are likely toshow regional and/or spatial variability.
To address the much faster response times ob-served it would appear much more likely that anyanoxia was limited to a thin ‘blanket’ at the sedi-ment/water interface. Such a ‘blanket’ would re-sult from remineralisation of organic componentsthat were rapidly posted to the sea £oor (e.g. dia-tom mats; Kemp et al., 1999), and so would re-£ect the patchy nature of the spatially and tem-porally variable export productivity distribution.
Observations of deep-living planktonic species(e.g. Globorotalia truncatulinoides) and benthicsthroughout sapropels suggest that they are mostcommon in the more oligotrophic areas of theeastern Mediterranean, e.g. o¡ the Gulf of Sirte.Might this re£ect spatial di¡erences in the exportproductivity levels? Poor ventilation would createdysoxic conditions everywhere, and regions ofhigh export £ux developed anoxic ‘blankets’whereas oligotrophic regions did not. The patchynature of such ‘blankets’ may also allow refugiafor certain benthic foraminifera to develop lo-cally, promoting more rapid repopulation.
In addition, there is a clear increase in sedimen-tary Corg concentrations with water depth (Muratand Got, 2000), that appears to con£ict with thenormal expectations of a mid-depth remineralisa-tion maximum/oxygen minimum. The observedincrease with depth may re£ect that a remineral-isation maximum instead prevailed at the sea£oor, presumably due to rapid export/dumpingof Corg, supporting our proposal that anoxiawas constrained to a blanket at the sediment/water interface.
Murat and Got (2000) observations of increas-ing organic carbon concentrations with depth sug-gest that deep water isolation also increases withdepth. In addition, the probability of signi¢cantventilation in a density-strati¢ed basin decreaseswith depth. As the indicators, reported here, of
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 115

(intermittent) bio-available oxygen within sapro-pels do not include observations from below 2026m, we cannot exclude the possibility that abyssaldepths developed more widespread and persistentanoxia. This uncertainty calls for high-resolutionstudies in abyssal sites of sapropels that haveknown reventilation histories in shallower locales.
5. Summary and conclusions: A dynamicventilation concept
Integrating the various strands of evidence dis-cussed above, we propose a dynamic concept for
the relationship between deep water ventilationand oxygenation at sapropel times. The ventila-tion state we propose is in fact similar to that oftoday, in that it remained characterised by inter-mittent climate-related maxima of new deep waterformation that were associated with cooling byinterannually variable northerly outbreaks of po-lar/continental air. However, at sapropel times,the ventilation was weakened overall, due to theadditional buoyancy gain imposed by the en-hanced freshwater in£ux into the basin. Our dy-namic concept of ventilation decouples watermass advection ^ driven by new formation andinternal mixing ^ from the oxygen concentrations
Northern Basins
Off Israel
Off the Gulf of Sirte
Cooling
Northerlies
B
B
B
B
Low export productionHigh export production
G.t
SW
IW
DW
N.pachyderma. ODS group/ U.peregrina
U.mediterranea
Dysoxic/ poorly ventilated water
Anoxic/ azoic blanket
Export production B
Rapid postingmechanisms (mats?)[Sancetta 1994, Pearce et al. 1998,Kemp et al. 1999]
Benthic foraminifera SW
DW
IW
Surface water
Intermediate water
Deep water
Fig. 8. Schematic 2-D summary of the discussed aspects of deep water advection and oxygenation, based on a compilation ofseveral sapropels. This is intended to represent a hypothetical north^south transect and is therefore not intended to de¢ne speci¢cspatial gradients. The letter B in the key represents benthic foraminifera in both the continuous benthic presence o¡ Israel andthe rapid repopulation S6. This exempli¢es the restricted nature of the anoxic blanket in both space and time.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119116

that are in addition a¡ected by chemical and bio-logical constraints, with their own temporal andspatial variability. This allows for contrastingoxygen concentrations within single water masses,both temporally and spatially.
Fig. 8 aims to summarise these key aspects in avery schematic 2-D representation, which obvi-ously cannot capture all observed 3-D and tem-poral variability. In addition, this ¢gure does notaccount for any possible anoxia deeper than 2000m, as mentioned above. Intermittent ventilation isshown in the north of the pro¢le, as determinedthroughout S1 in both the Aegean and Adriaticfrom the presence of benthic faunas and their iso-topic composition, in particular contrasting thelow-oxygen tolerant species Globobulimina a⁄nisand the more oxyphilic Uvigerina mediterranea.This deep water ventilation must occur at leastonce within every sample interval (V50 years),and most probably even more frequently. Thegraded grey ¢ll illustrates the inferred oxygenationgradient within both deep water and intermediatewater masses, and highlights the dissociation ofintermediate water from the surface and deepwaters in the Aegean Sea (see Casford et al.,2002). A blanket of anoxia develops under areasof higher export production, with the presence ofbenthic foraminifera in cores from even the deep,southern Mediterranean suggesting the limited ex-tent of these truly anoxic conditions in both spaceand time. This blanket hypothesis for the develop-ment of anoxia removes the requirement to titratethe complete water column upon reventilation,allowing very rapid events of re-oxygenation andreturn to anoxic conditions seen in the proxy re-cords.
Acknowledgements
We acknowledge the contributions of the scien-ti¢c team of RV Meteor cruise M51-3 (Novem-ber^December 2001), in particular the chief scien-tist Professor C. Hemleben. We thank LarryPeterson and Anne Murat for their invaluablecomments during review. Additional thanks arealso due to S. Cooke and H. Sloane for theirtechnical assistance.
References
Abu-Zied, R.H., 2001. High Resolution LGM-Present Pale-oceanography of the N.E. Mediterranean: A Benthic Per-spective. Ph.D. Thesis.
Bethoux, J.P., Gentili, B., Morin, P., Nicolas, E., Pierre, C.,Ruiz-Pino, D., 1999. The Mediterranean Sea: a miniatureocean for climatic and environmental studies and a key forthe climatic functioning of the North Atlantic. Prog. Ocean-ogr. 44, 131^146.
Boyle, E.A., Lea, D.W., 1989. Cd and Ba in planktonic fora-minifera from the Eastern Mediterranean: evidence for riverout£ow and enriched nutrients during sapropel production.Trans. Am. Geophys. Union 70, 101^134.
Calvert, S.E., 1983. Geochemistry of Pleistocene sapropels andassociated sediments from the eastern Mediterranean. Oce-anol. Acta 6, 255^267.
Casford, J.S.L., Rohling, E.J., Abu-Zied, R., Cooke, S., Fon-tanier, C., Leng, M., Lykousis, V., 2002. Circulationchanges and nutrient concentrations in the Late QuaternaryAegean Sea: A non-steady state concept for sapropel for-mation. Paleoceanography (in press).
Casford, J.S.L., Abu-Zied, R., Rohling, E.J., Cooke, S., Boes-senkool, K.P., Brinkhuis, H., DeVries, C., Wefer, G., Ge-raga, M., Papatheodorou, G., Croudace, I., Thomson, J.,Lykousis, V., 2001. Mediterranean climate variability duringthe Holocene. Mediterranean Mar. Sci. 2, 45^55.
Cheddadi, R., Rossignol-Strick, M., 1995. Improved preserva-tion of organic-matter and pollen in eastern Mediterraneansapropels. Paleoceanography 10, 301^309.
Cita, M.B., Grignani, D., 1982. Nature and origin of LateNeogene Mediterranean sapropels. In: Schlanger, S.O.,Cita, M.B. (Eds.), The Nature and Origin of CretaceousCarbon-Rich Facies. Academic Press, London, pp. 165^196.
Cita, M.B., Beghi, C., Camerlenghi, A., Kastens, K.A.,McKoy, F.W., Nosetto, A., Parisi, E., Scolari, F., Tomadin,L., 1984. Turbidites and megaturbidites from the Herodotusabyssal plain (Eastern Mediterranean) unrelated to seismicevents. Mar. Geol. 55, 79^101.
Cita, M.B., Vergnaud-Grazzini, C., Robert, C., Chamley, H.,Ciaran¢, N., d’Onofrio, S., 1977. Paleoclimatic record of along deep sea core from the eastern Mediterranean. Quat.Res. 8, 205^235.
DeLange, G.J., Ten Haven, H.L., 1983. Recent sapropel for-mation in the eastern Mediterranean. Nature 305, 797^798.
De Rijk, S., Rohling, E.J., Hayes, A., 1999. Onset of climaticdeterioration in the eastern Mediterranean around 7 ky BP;micropalaeontological data from Mediterranean sapropel in-terruptions. Mar. Geol. 153, 337^3432.
Hayes, A., Rohling, E.J., DeRijk, S., Kroon, D., Zachariasse,W.J., 1999. Mediterranean planktonic foraminifera faunasduring the last glacial cycle. Mar. Geol. 153, 239^252.
Hemleben, C., Spindler, M., Anderson, O.R., 1989. ModernPlanktonic Foraminifera. Springer, New York, 363 pp.
Jorissen, F.J., 1999. Benthic foraminiferal successions acrossLate Quaternary Mediterraenan sapropels. Mar. Geol. 153,91^101.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 117

Jorissen, F.J., Asioli, A., Borsetti, A.M., Capotondi, L., de-Visser, J.P., Hilgen, F.J., Rohling, E.J., vanderBorg, K.,Vergnaud-Grazzini, C., Zachariasse, W.J., 1993. Late Qua-ternary central Mediterranean biochronology. Mar. Micro-paleontol. 21, 169^189.
Kemp, A.E.S., Pearce, R.B., Koizumi, I., Pike, J., Rance, S.J.,1999. The role of mat-forming diatoms in the formation ofMediterranean sapropels. Nature 398, 57^61.
Kullenberg, B., 1952. On the salinity of the water contained inmarine sediments. Medd. Oceanogr. Inst. Go«teborg 21, 1^38.
Leaman, K.D., Schott, F.A., 1991. Hydrographic structure ofthe convection regime in the Gulf of Lions Winter 1987.J. Phys. Oceanogr. 21, 575^598.
Lohmann, G.P., Schweitzer, P.N., 1990. Globorotalia trunca-tulinoides’ growth and chemistry as probes of the past ther-mocline: 1. Shell size. Paleoceanography 5, 55^75.
Mariolopoulos, E.G., 1961. An Outline of the Climate ofGreece. Publ. Meteorol. Inst. Univ. Athens 6, 51 pp.
Mercone, D., Thomson, J., Abu-Zied, R.H., Croudace, I.W.,Rohling, E.J., 2001. High-resolution geochemical and micro-palaeontological pro¢ling of the most recent eastern Medi-terranean sapropel. Mar. Geol. 177, 25^44.
Mercone, D., Thomson, J., Croudace, I.W., Siani, G., Paterne,M., Troelstra, S., 2000. Duration of S1, the most recentsapropel in the eastern Mediterranean Sea, as indicated byaccelerator mass spectrometry radiocarbon and geochemicalevidence. Paleoceanography, 15, 336^347.
Murat, A., Got, H., 2000. Organic carbon variations of theeastern Mediterranean Holocene sapropel: a key for under-standing formation processes. Palaeogeogr. Palaeoclimatol.Palaeoecol. 158, 241^257.
Olauson, E., 1961. Studies of deep-sea cores. Rep. Swed. DeepSea Exped. 8, 335^391.
Pedersen, T.F., Calvert, S.E., 1990. Anoxia vs. productivity ^what controls the formation of organic-carbon-rich sedi-ments and sedimentary-rocks. AAPG Bull. 74, 454^466.
Perissoratis, C., Piper, J.W., 1992. Age, regional variation, andshallowest occurrence of S1 sapropel in the Northern Ae-gean Sea. Geo-Mar. Lett. 12, 49^53.
Poulos, S.E., Drakopoulos, P.G., Collins, M.B., 1997. Season-al variability in sea surface oceanographic conditions in theAegean Sea (eastern Mediterranean) an overview. J. Mar.Syst. 13, 225^244.
Pujol, A., Vergnaud-Grazzini, C., 1995. Distribution patternsof live planktic foraminifera as related to regional hydrog-raphy and productive systems of the Mediterranean Sea.Mar. Micropaleontol. 25, 187^217.
Reiss, Z., Halicz, E., Luz, B., 2000. Late-Holocene foraminif-era from the SE Levantine Basin. Isr. J. Earth Sci. 48, 1^27.
Roether, W.H., Manca, B.B., Klein, B., Bregant, D., Georgo-poulos, D., Beitzel, V., Kovacevic, V., Luchetta, A., 1996.Recent changes in eastern Mediterranean deep waters. Sci-ence 271, 333^335.
Roether, W., Well, R., 2001. Oxygen consumption in the East-ern Mediterranean. Deep-Sea Res. I 48, 1535^1551.
Rohling, E.J., Mayewski, P.A., Abu-Zied, R.H., Casford,J.S.L., Hayes, A., 2002. Holocene atmosphere^ocean inter-actions: records from Greenland and The Aegean Sea. Clim.Dyn. 18, 587^593.
Rohling, E.J., 1999b. Environmental controls on salinity andN
18O in the Mediterranean. Paleoceanography 14, 706^715.
Rohling, E.J., Jorissen, F.J., DeStigter, H.C., 1997. 200 yearinterruption of Holocene sapropel formation in the AdriaticSea. J. Micropaleontol. 16, 97^108.
Rohling, E.J., Den Dulk, M., Pujol, C., Vergnaud-Grazzini,C., 1995. Abrupt hydrographic change in the Alboran Sea(western Mediterranean) around 8000 yrs BP. Deep-Sea Res.I 42, 1609^1619.
Rohling, E.J., 1994. Review and new aspects concerning theformation of eastern Mediterranean sapropels. Mar. Geol.122, 1^28.
Rohling, E.J., Jorissen, F.J., Vergnaud-Grazzini, C., Zacha-riasse, W.J., 1993. Northern Levantine and Adriatic Quater-nary planktic foraminifera; Reconstruction of paleoenviron-mental gradients. Mar. Micropaleontol. 21, 191^218.
Rohling, E.J., Gieskes, W.W.C., 1989. Late Quaternarychanges in Mediterranean intermediate water density andformation rate. Paleoceanography 4, 531^545.
Rossignol-Strick, M., 1985. Mediterranean Quaternary sapro-pels: an immediate response of the African monsoon tovariation of insolation. Palaeogeogr. Palaeoclimatol. Palaeo-col. 49, 237^265.
Rossignol-Strick, M., 1983. African monsoons, an immediateresponse to orbital insolation. Nature 304, 46^49.
Rossignol-Strick, M., Nestero¡, W., Olive, P., Vergnaud-Graz-zini, C., 1982. After the deluge: Mediterranean stagnationand sapropel formation. Nature 295, 105^110.
Saaroni, H., Bitan, A., Alpert, P., Ziv, B., 1996. ContinentalPolar outbreaks into the Levant and Eastern Mediterranean.Int. J. Climatol. 16, 1175^1191.
Stanley, D.J., 1978. Ionian Sea sapropel distribution and LateQuaternary palaeoceanography in the Eastern Mediterra-nean. Nature 274, 149^152.
Stratford, K., Williams, R.G., Myers, P.G., 2000. Impact ofthe circulation on sapropel formation in the eastern Medi-terranean. Glob. Biogeochem. Cycles 14, 683^695.
Theocharis, A., 1989. Deep water formation and circulation inthe Aegean Sea. In: Charnock, H. (Ed.), Reports in Mete-orology and Oceanography, 40, I, pp. 335^359.
Thunell, R.C., Williams, D.F., Kennett, J.P., 1977. Late Qua-ternary paleoclimatology, stratigraphy, and sapropel historyin eastern Mediterranean deep-sea sediments. Mar. Micro-paleontol. 2, 371^388.
Van Os, B.J.H., Lourens, L.J., Hilgen, F.J., Delange, G.J.,Beaufort, L., 1994. The formation of Pliocene sapropelsand carbonate cycles in the Mediterranean ^ diagenesis, di-lution, and productivity. Paleoceanography 9, 601^617.
Van Straaten, R.M.V.U., 1966. Micro-malacological investiga-tion of cores from the southeastern Adriatic Sea. Proc. Kon.Ned. Akad. Wet. 69, pp. 429^445.
Van Straaten, R.M.V.U., 1972. Holocene stages of oxygen
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119118

depletion in waters of the Adriatic. In: Stanley, D.J. (Ed.),The Mediterranean Sea: A Natural Sedimentation Labora-tory. Dowden, Hutchinson and Ross, Stroudsburg, PA, 631pp.
Vergnaud-Grazzini, C., 1985. Mediterranean late Cenozoic sta-ble isotope record: Stratigraphic and paleoclimatic implica-tions. In: Stanley, D.J., Wezel, F.C. (Eds.), Geological Evo-lution of the Mediterranean Basin. Springer, New York, pp.413^451.
Vergnaud-Grazzini, C., Ryan, W.B.F., Cita, M.B., 1977. Sta-ble isotope fractionation, climate change and episodic stag-nation in the Eastern Mediterranean during the Late Qua-ternary. Mar. Micropaleontol. 2, 353^370.
Vismara-Schilling, A., 1984. Holocene stagnation event in theeastern Mediterranean. Evidence from deep-sea benthic fo-raminifera in Calabria and Western Mediterranean ridges.Benthos ’83,Second Int. Symp. Benthic Foraminifera, Pau,April 1983, pp. 585^596.
PALAEO 2962 8-1-03 Cyaan Magenta Geel Zwart
J.S.L. Casford et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 190 (2003) 103^119 119




















