
Steryl ethers in a Valanginian claystone: Molecular evidence for cooler waters in the central...
13
Steryl ethers in a Valanginian claystone: Molecular evidence for cooler waters in the central Pacific during the Early Cretaceous? Simon C. Brassell ⁎ Biogeochemical Laboratories, Department of Geological Sciences, Indiana University, Bloomington, IN 47405-1403, USA abstract article info Article history: Received 5 September 2008 Received in revised form 2 August 2009 Accepted 13 August 2009 Available online 27 August 2009 Keywords: Oceanic anoxic events Upwelling Paleoenvironment Biomarkers Biogeochemistry Zooplankton Coring on Shatsky Rise by Ocean Drilling Project Leg 198 recovered sediments rich in organic matter (>2.0% C org ) from three time intervals: the Early Aptian, corresponding to oceanic anoxic event (OAE 1a), the Valanginian and the Berriasian. Episodes of enhanced sequestration of organic matter during the Valanginian are well documented in the Tethyan realm, notably those associated with a positive δ 13 C isotopic excursion and with changes in plankton assemblages, including a demise in nannoconids. The δ 13 C excursion has been previously reported in the Pacific but the occurrence of Valanginian sediments rich in organic matter (> 1% C org ) is unprecedented. A distinctive feature of the biomarker composition of these Valanginian sediments at Shatsky Rise is the presence of a suite of steryl ethers, which extends the temporal record of these compounds from the Neogene to the Early Cretaceous. The biological source of steryl ethers remains enigmatic, although the sedimentary occurrences of these compounds in Quaternary and Neogene environments characterized by cool water, high seasonal productivity, and/or nutrient enrichment by upwelling, in combination with their liquid crystal properties, raise the possibility that they may represent storage lipids, akin to wax esters and triacylglycerols. Prior occurrences of steryl ethers suggest that their presence in the Valanginian may reflect an oceanic environment characterized by upwelling, consistent with the likely position of Shatsky Rise within the equatorial divergence zone of the Pacific during the Early Cretaceous. Alternatively, or in addition, the appearance of these compounds may represent a biological response to cooler temperatures, and thereby augment evidence for global cooling during the Early Valanginian, consistent with independent paleontological evidence for contemporaneous, upwelling- induced increases in productivity in the Western Atlantic. © 2009 Elsevier B.V. All rights reserved. 1. Introduction The first well documented global episode of enhanced burial and preservation of organic matter (OM) during the Cretaceous occurs in the early Aptian (e.g. Schlanger and Jenkyns, 1976; Arthur and Schlanger, 1979; Jenkyns, 1980; Arthur et al., 1987, 1990; Bralower et al., 1994; Leckie et al., 2002), hence its designation as oceanic anoxic event OAE- 1a (Arthur et al., 1990; Bralower et al., 1994). It is also characterized by changes in plankton assemblages and its onset marked by a negative δ 13 C excursion in carbonates best documented in the Tethyan realm (Menegatti et al., 1998; Hochuli et al., 1999; Bellanca et al., 2002; Bersezio et al., 2002; Danelian et al., 2004; Erba and Tremolada, 2004 Heimhofer et al., 2004; Herrle et al., 2004), but also manifest in the Tethys–Atlantic seaway (de Gea et al., 2003), in the Atlantic (Herrle et al., 2004; Bralower et al., 1999), and in terrestrial sequences (Gröcke et al., 1999; Ando et al., 2002). The worldwide extent (Leckie et al., 2002) for these changes in the global carbon cycle is related to emplacement of the Ontong–Java Plateau (Larson and Erba, 1999; Weissert and Erba, 2004) and potentially methane release from hydrates (Jahren et al., 2001; Beerling et al., 2002; Jenkyns, 2003). A comparable episode of OM sequestration in the Jurassic is observed in the Toarcian (Beerling et al., 2002; Jenkyns, 2003) and an earlier Cretaceous event with similarities to OAE-1a occurs in the Valanginian, which has been named the Weissert OAE (Erba et al., 2004). A positive δ 13 C excursion in carbonates (Lini et al., 1992; Weissert et al., 1998; Erba and Tremolada, 2004; Weissert and Erba, 2004; Erba et al., 2004) accompanies this event, which also shows a decline in nannoconids (Erba and Tremolada, 2004). Records of this event are best expressed in the Tethyan realm (Lini et al., 1992; Channell et al., 1993; Weissert et al., 1998; Gröcke et al., 2003; Erba et al., 2004), including OM-rich (>1.0% C org ) intervals in the Vocontian Basin (Hennig et al., 1999) and the Southern Alps (Bersezio et al., 2002); they also occur in the Pacific(Erba et al., 2004). In the Pacific, an OM-rich sediment corresponding to this event was first recovered from Shatsky Rise (Core 1213B-15R-1; 2.54% C org ; Brassell et al., 2004) during drilling by the Ocean Drilling Program (ODP) Leg 198 (Bralower et al., 2002), prompting investigation of the characteristics and sources of its OM that can provide evidence helpful in assessing its depositional environment. The comparatively shallow depth of burial (324 mbsf) of this interval and the resultant immaturity of its OM facilitate molecular analysis of Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57 ⁎ Fax: +1 812 855 7961. E-mail address: [email protected]. 0031-0182/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2009.08.009 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Steryl ethers in a Valanginian claystone: Molecular evidence for cooler waters in the central...

Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Steryl ethers in a Valanginian claystone: Molecular evidence for cooler waters in thecentral Pacific during the Early Cretaceous?
Simon C. Brassell ⁎Biogeochemical Laboratories, Department of Geological Sciences, Indiana University, Bloomington, IN 47405-1403, USA
⁎ Fax: +1 812 855 7961.E-mail address: [email protected].
0031-0182/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.palaeo.2009.08.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 5 September 2008Received in revised form 2 August 2009Accepted 13 August 2009Available online 27 August 2009
Keywords:Oceanic anoxic eventsUpwellingPaleoenvironmentBiomarkersBiogeochemistryZooplankton
Coring on Shatsky Rise by Ocean Drilling Project Leg 198 recovered sediments rich in organic matter (>2.0%Corg) from three time intervals: the Early Aptian, corresponding to oceanic anoxic event (OAE 1a), theValanginian and the Berriasian. Episodes of enhanced sequestration of organic matter during the Valanginianare well documented in the Tethyan realm, notably those associated with a positive δ13C isotopic excursionand with changes in plankton assemblages, including a demise in nannoconids. The δ13C excursion has beenpreviously reported in the Pacific but the occurrence of Valanginian sediments rich in organic matter (>1%Corg) is unprecedented. A distinctive feature of the biomarker composition of these Valanginian sediments atShatsky Rise is the presence of a suite of steryl ethers, which extends the temporal record of thesecompounds from the Neogene to the Early Cretaceous. The biological source of steryl ethers remainsenigmatic, although the sedimentary occurrences of these compounds in Quaternary and Neogeneenvironments characterized by cool water, high seasonal productivity, and/or nutrient enrichment byupwelling, in combination with their liquid crystal properties, raise the possibility that they may representstorage lipids, akin to wax esters and triacylglycerols. Prior occurrences of steryl ethers suggest that theirpresence in the Valanginian may reflect an oceanic environment characterized by upwelling, consistent withthe likely position of Shatsky Rise within the equatorial divergence zone of the Pacific during the EarlyCretaceous. Alternatively, or in addition, the appearance of these compounds may represent a biologicalresponse to cooler temperatures, and thereby augment evidence for global cooling during the EarlyValanginian, consistent with independent paleontological evidence for contemporaneous, upwelling-induced increases in productivity in the Western Atlantic.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
The first well documented global episode of enhanced burial andpreservation of organicmatter (OM)during the Cretaceousoccurs in theearly Aptian (e.g. Schlanger and Jenkyns, 1976; Arthur and Schlanger,1979; Jenkyns, 1980; Arthur et al., 1987, 1990; Bralower et al., 1994;Leckie et al., 2002), hence its designation as oceanic anoxic event OAE-1a (Arthur et al., 1990; Bralower et al., 1994). It is also characterized bychanges in plankton assemblages and its onset marked by a negativeδ13C excursion in carbonates best documented in the Tethyan realm(Menegatti et al., 1998; Hochuli et al., 1999; Bellanca et al., 2002;Bersezio et al., 2002; Danelian et al., 2004; Erba and Tremolada, 2004Heimhofer et al., 2004; Herrle et al., 2004), but also manifest in theTethys–Atlantic seaway (de Gea et al., 2003), in the Atlantic (Herrleet al., 2004; Bralower et al., 1999), and in terrestrial sequences (Gröckeet al., 1999;Andoet al., 2002). Theworldwideextent (Leckie et al., 2002)for these changes in the global carbon cycle is related to emplacement ofthe Ontong–Java Plateau (Larson and Erba, 1999; Weissert and Erba,
l rights reserved.
2004) and potentially methane release from hydrates (Jahren et al.,2001; Beerling et al., 2002; Jenkyns, 2003). A comparable episode of OMsequestration in the Jurassic is observed in the Toarcian (Beerling et al.,2002; Jenkyns, 2003) andanearlier Cretaceous eventwith similarities toOAE-1a occurs in the Valanginian, which has been named the WeissertOAE (Erba et al., 2004). A positive δ13C excursion in carbonates (Liniet al., 1992; Weissert et al., 1998; Erba and Tremolada, 2004; Weissertand Erba, 2004; Erba et al., 2004) accompanies this event, which alsoshows a decline in nannoconids (Erba and Tremolada, 2004). Records ofthis event are best expressed in the Tethyan realm (Lini et al., 1992;Channell et al., 1993;Weissert et al., 1998;Gröcke et al., 2003; Erba et al.,2004), including OM-rich (>1.0% Corg) intervals in the Vocontian Basin(Hennig et al., 1999) and the Southern Alps (Bersezio et al., 2002); theyalso occur in the Pacific (Erba et al., 2004). In the Pacific, an OM-richsediment corresponding to this event was first recovered from ShatskyRise (Core 1213B-15R-1; 2.54% Corg; Brassell et al., 2004) during drillingby the Ocean Drilling Program (ODP) Leg 198 (Bralower et al., 2002),prompting investigation of the characteristics and sources of its OM thatcan provide evidence helpful in assessing its depositional environment.The comparatively shallow depth of burial (324 mbsf) of this intervaland the resultant immaturity of its OM facilitate molecular analysis of

46 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
the OM within this interval. Indeed, prior investigations of similarlyimmature Cretaceous marine sediments have shown that they canpreserve diagnostic compounds either as intact biolipids (e.g. Brassellet al., 1987), or as components bound within insoluble OM (SinningheDamsté and de Leeuw, 1990) that are typically lost, modified, orobscured by diagenetic processes. It is therefore not surprising that theoldest occurrences of several intact functionalized biomarkers aredocumented in Cretaceous strata (e.g. Brassell et al., 1987, 1983, 2004),where their preservation permits use as paleoenvironmental andpaleoclimatic proxies (e.g. Didyk et al., 1978; Brassell and Eglinton,1986; Brassell et al., 1987; Sinninghe Damsté and de Leeuw, 1990;Kuypers et al., 2001; Dumitrescu and Brassell, 2005). Yet the range ofsedimentary biomarkers also includes components that possess generictraits characteristic of biosynthetic processes but await discovery asconstituents of extant organisms, and haveno obvious precursor amongknown biolipids (Ourisson et al., 1982; Brassell and Eglinton, 1986;Brassell et al., 1994). Over time the origin of most common biomarkershas been resolved, whereas other ‘orphan’ biomarkers, including chro-mans (Sinninghe Damsté et al., 1987) and steryl ethers (Brassell, 1994),still await discovery in contemporary biota or recognition of theirpathways for formation. These compounds present challenges in theinterpretation and assessment of the sources of sedimentary organicmatter, especially when they occur widely in both modern and ancientsediments (Rullkötter et al., 1982; Brassell and Eglinton, 1986).Elucidation of their biological origins or recognition ofmode of formationfrom known precursors in the environment helps to define the timing ofevolutionary processes coupled to the advent of biosynthetic pathways(Brassell et al., 1987; Sinninghe Damsté et al., 2004). Thus, the lack ofknown biological origins for sedimentary biomarkers, and whether theyare products of biosynthesis, herbivory or early diagenesis does notexclude the possibility to infer environmental conditions from theiroccurrence when extensive reports show that it is governed bysystematic controls or appears restricted to specific depositional settings.
One group of molecules that remain enigmatic is the suites of steryl(or sterol) ethers occasionally found inmarine sediments but unknownin any viable source organism. These compounds were first observedand characterized in Quaternary diatomaceous oozes from Walvis Bay(Boon and de Leeuw, 1979), and have subsequently been identified in anumber of othermarine sediments (Brassell 1980; Schouten et al., 2000,2005; Schefuß et al., 2001). Evaluation of the sedimentary occurrencesof steryl ethers can establish their relationship to depositionalenvironments and clarify their possible origins, which is the theme ofthis study. Moreover, the presence of a suite of these biomarkers in aValanginian immature sediment from the central Pacific promptsassessment of their potential as climate proxies.
2. Materials and methods
2.1. Lower Cretaceous sediments from Shatsky Rise
The Berriasian through Albian succession recovered at sites drilledon Shatsky Rise in the west–central Pacific during ODP Leg 198 waschert-dominated (Bralower et al., 2002). The detailed sedimentologyand stratigraphy of this succession is published elsewhere (Braloweret al., 2002). Samples for organic geochemical analyses were collectedwithin the Lower Cretaceous interval from various lithologies inter-bedded with the cherts, including claystones, calcareous claystones,chalks, radiolarites and porcellanites (Brassell et al., 2004; Dumitrescuand Brassell, 2005). Lower Aptian sediments corresponding to OAE-1awere recovered at three sites (Sites 1207, 1213and1214;Braloweret al.,2002; Brassell et al., 2004; Dumitrescu and Brassell, 2005, 2006). At Site1213 on the southern high of Shatsky Rise the recovery of organic-richLower Aptian sediments in Core 1213B-8R-1 (sampled at 47–48, 63–64,and 96–97 cm) at a depthof approximately 256 mbsf (meters below seafloor), was complemented by two dark, calcareous intervals fromdeeper sections (1213B-15R-1 5-7 cm, 324 mbsf; 1213B-19R-1 112-
113, 363 mbsf) that are assigned as Valanginian (Zone NK3a) andBerriasian (Zone NK2a), respectively, based on their calcareousnannoplankton assemblages (Bown, 2005). Plate reconstructionsplace Site 1213 in the central equatorial Pacific during the EarlyCretaceous (Fig. 1; Bralower et al., 2002; Dumitrescu and Brassell, 2005,2006) and microfossil assemblages from these strata suggest anintermediate water depth (>1.5 km; Bralower et al., 2002).
The analytical focus of this study is the molecular composition,notably the distribution of components with intermediate polarity, ofa single sample taken from the Valanginian organic-rich interval atSite 1213 (Bralower et al., 2002). A key objective is to explore specificdifferences in the biomarker distributions of this sample comparedwith the compositions of Lower Aptian samples from Shatsky Rise(Dumitrescu and Brassell, 2005), and the Berriasian interval from Site1213 (Bralower et al., 2002), especially molecular evidence that mightreflect the depositional conditions associated with enhanced burial oforganic matter in these sediments.
2.2. Quaternary and Tertiary sediments
During the past four decades the biomarker composition of anextensive range of modern and ancient marine sediments has beenexamined in efforts to determine the biological origins of sedimentaryorganic matter and controls on its supply and preservation. The focushere is on marine sediments from various locations (Fig. 2) that havebeen reported or found to contain steryl ethers (Table 1), typicallyduring examination of ketone fractions that contain alkenones(Marlowe et al., 1990; Brassell, 1993). Specifically, published reportsof steryl ethers in sediments (cf. Schouten et al., 2005) are comple-mented bypublisheddata for sediments fromthe Japan Trench (Brassellet al., 1980; Brassell et al., 1980) and previously unpublished data forsediments from the Iceland–Faeroe Ridge and the Falkland Plateau(Marlowe, 1984; Fig. 2). Steryl ethers were recognized in sedimentsfrom the last two locations during examination of alkenones in suites ofOligocene and Eocene sediments (Marlowe, 1984;Marlowe et al., 1990;Brassell, 1993). Steryl ethers were present in all the Oligocene samplesfrom these two sites, and in one Upper Eocene sample.
Consideration of steryl ether compositions for Quaternary throughEocene samples permits evaluation of consistencies and variations inthe distributions of these compounds through geologic time, andassessment of potential relationshipswith other steroidal componentswherein a broad range of samples is helpful in pattern recognition.
2.3. Analytical methodology
For molecular analyses sediment samples (<2 g) were lyophilized,and extracted ultrasonically using CH2Cl2 (10 mL) for 30 min. Theextract was reduced to dryness under N2 and eluted through silica in aPasteur pipette using hexane (4 mL) and CH2Cl2 (5 mL) as eluates tocollect fractions dominated by hydrocarbon and ketone constituents,respectively. Each eluant was taken to near dryness under N2 anddissolved in hexane (50 µL) for analysis by gas chromatography–massspectrometry (GC–MS) using a Hewlett-Packard 6973 system con-sisting of an HP 6890 GC with a Mass Selective Detector (MSD) and anHP 7683 Automatic Liquid Sampler (ALS). The GC was equipped withan EPC split–splitless injector and an HP capillary column (5% phenylmethyl siloxane; 30 m×0.25 µm) programmed from 40 °C to 130 °Cat 20 °C/min, then at 4 °C/min to 320 °C, and held isothermally at320 °C for 20 min. Heliumwas used as the carrier gas. The transfer linewas set at 280 °C and the source of the MS at 230 °C. The MSD wasscanned from m/z 27 to 550 and an HP MS Chemstation was used fordata acquisition and processing. The identity of individual componentswas determined from their mass spectral characteristics and retentiontimes by comparison with the literature.
The procedures used for extraction, fractionation, and separation ofbiomarkers in the Quaternary and Tertiary sediments that contain steryl

Fig. 1. Paleogeographic map indicating the locations of Lower Cretaceous sequences, including Site 1213 at Shatsky Rise, that are discussed in the text and/or considered in Fig. 5 (seereferences therein). Open stars represent the location of DSDP or ODP Sites with their site number; other sites are shownwith black stars, with that labeled Italy representing sites atBreggia, Polaveno, and Capriolo. Snowflake symbols depict the locations of ice-rafted deposits (Alaska, Siberia and central Australia).
47S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
ethers are described in the specific publications cited (Table 1). These arecoupled with the approaches outlined below for identification of sterylethers in lipid fractions wherein these compounds had not beenpreviously reported. For Oligocene and Eocene samples steryl etherswere recognized in total lipid extracts thatwere dominated by alkenones(Marlowe, 1984; Marlowe et al., 1984, 1990). However, only the majorbiomarker constituents of these extracts were examined in detail.
2.4. Biomarker identifications
Assignments of steryl ethers are based on interpretation of theirmass spectra by reference to previous reports (Boon and de Leeuw,
Fig. 2.Map of locations of sediment samples found to contain sterol ethers (Table 1; see refeshown with open stars.
1979; Brassell et al., 1980; Schouten et al., 2000, 2005), including datafor synthetic standards (Vill and Weber, 1991, 1994; Schouten et al.,2005). The position of the alkyl ether chain initially presumed to be atC-3 on biosynthetic grounds (Boon and de Leeuw, 1979; Brassell et al.,1980; Schouten et al., 2000) has been confirmed as 3β̃-alkoxy(Schouten et al., 2005). The absence of molecular ions in the massspectra of several components in the samples precluded directdetermination of their empirical formulae and likely structures, buttentative identifications were possible using the prominent fragmentions corresponding to the sterol side chain (e.g. m/z 127 for C9H19,nonyl) and to the sterol moiety with/without hydrogen transfer (e.g.m/z 368/370 for C27H44/C27H46). The features of the mass spectra
rences therein). The positions of the sampling sites discussed in the text and figures are
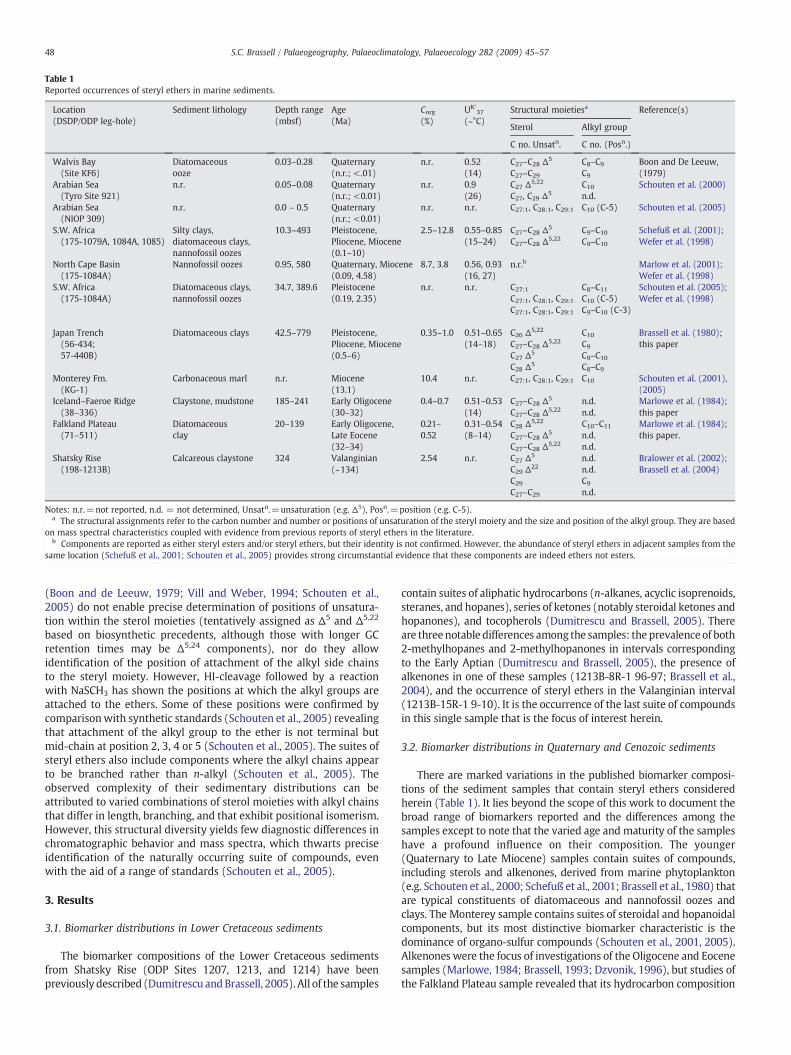
Table 1Reported occurrences of steryl ethers in marine sediments.
Location(DSDP/ODP leg-hole)
Sediment lithology Depth range(mbsf)
Age(Ma)
Corg(%)
UK′37
(~°C)Structural moietiesa Reference(s)
Sterol Alkyl group
C no. Unsatn. C no. (Posn.)
Walvis Bay(Site KF6)
Diatomaceousooze
0.03–0.28 Quaternary(n.r.; <.01)
n.r. 0.52(14)
C27–C28 Δ5
C27–C29C8–C9
C9
Boon and De Leeuw,(1979)
Arabian Sea(Tyro Site 921)
n.r. 0.05–0.08 Quaternary(n.r.; <0.01)
n.r. 0.9(26)
C27 Δ5,22
C27, C29 Δ5C10
n.d.Schouten et al. (2000)
Arabian Sea(NIOP 309)
n.r. 0.0 – 0.5 Quaternary(n.r.; <0.01)
n.r. n.r. C27:1, C28:1, C29:1 C10 (C-5) Schouten et al. (2005)
S.W. Africa(175-1079A, 1084A, 1085)
Silty clays,diatomaceous clays,nannofossil oozes
10.3–493 Pleistocene,Pliocene, Miocene(0.1–10)
2.5–12.8 0.55–0.85(15–24)
C27–C28 Δ5
C27–C28 Δ5,22C9–C10
C9–C10
Schefuß et al. (2001);Wefer et al. (1998)
North Cape Basin(175-1084A)
Nannofossil oozes 0.95, 580 Quaternary, Miocene(0.09, 4.58)
8.7, 3.8 0.56, 0.93(16, 27)
n.r.b Marlow et al. (2001);Wefer et al. (1998)
S.W. Africa(175-1084A)
Diatomaceous clays,nannofossil oozes
34.7, 389.6 Pleistocene(0.19, 2.35)
n.r. n.r. C27:1C27:1, C28:1, C29:1
C27:1, C28:1, C29:1
C8–C11
C10 (C-5)C9–C10 (C-3)
Schouten et al. (2005);Wefer et al. (1998)
Japan Trench(56-434;57-440B)
Diatomaceous clays 42.5–779 Pleistocene,Pliocene, Miocene(0.5–6)
0.35–1.0 0.51–0.65(14–18)
C26 Δ5,22
C27–C28 Δ5,22
C27 Δ5
C28 Δ5
C10
C9
C9–C10
C8–C9
Brassell et al. (1980);this paper
Monterey Fm.(KG-1)
Carbonaceous marl n.r. Miocene(13.1)
10.4 n.r. C27:1, C28:1, C29:1 C10 Schouten et al. (2001),(2005)
Iceland–Faeroe Ridge(38–336)
Claystone, mudstone 185–241 Early Oligocene(30–32)
0.4–0.7 0.51–0.53(14)
C27–C28 Δ5
C27–C28 Δ5,22n.d.n.d.
Marlowe et al. (1984);this paper
Falkland Plateau(71–511)
Diatomaceousclay
20–139 Early Oligocene,Late Eocene(32–34)
0.21–0.52
0.31–0.54(8–14)
C28 Δ5,22
C27–C28 Δ5
C27–C28 Δ5,22
C10–C11
n.d.n.d.
Marlowe et al. (1984);this paper.
Shatsky Rise(198-1213B)
Calcareous claystone 324 Valanginian(~134)
2.54 n.r. C27 Δ5
C29 Δ22
C29C27–C29
n.d.n.d.C9
n.d.
Bralower et al. (2002);Brassell et al. (2004)
Notes: n.r.=not reported, n.d. = not determined, Unsatn.=unsaturation (e.g. Δ5), Posn.=position (e.g. C-5).a The structural assignments refer to the carbon number and number or positions of unsaturation of the steryl moiety and the size and position of the alkyl group. They are based
on mass spectral characteristics coupled with evidence from previous reports of steryl ethers in the literature.b Components are reported as either steryl esters and/or steryl ethers, but their identity is not confirmed. However, the abundance of steryl ethers in adjacent samples from the
same location (Schefuß et al., 2001; Schouten et al., 2005) provides strong circumstantial evidence that these components are indeed ethers not esters.
48 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
(Boon and de Leeuw, 1979; Vill and Weber, 1994; Schouten et al.,2005) do not enable precise determination of positions of unsatura-tion within the sterol moieties (tentatively assigned as Δ5 and Δ5,22
based on biosynthetic precedents, although those with longer GCretention times may be Δ5,24 components), nor do they allowidentification of the position of attachment of the alkyl side chainsto the steryl moiety. However, HI-cleavage followed by a reactionwith NaSCH3 has shown the positions at which the alkyl groups areattached to the ethers. Some of these positions were confirmed bycomparisonwith synthetic standards (Schouten et al., 2005) revealingthat attachment of the alkyl group to the ether is not terminal butmid-chain at position 2, 3, 4 or 5 (Schouten et al., 2005). The suites ofsteryl ethers also include components where the alkyl chains appearto be branched rather than n-alkyl (Schouten et al., 2005). Theobserved complexity of their sedimentary distributions can beattributed to varied combinations of sterol moieties with alkyl chainsthat differ in length, branching, and that exhibit positional isomerism.However, this structural diversity yields few diagnostic differences inchromatographic behavior and mass spectra, which thwarts preciseidentification of the naturally occurring suite of compounds, evenwith the aid of a range of standards (Schouten et al., 2005).
3. Results
3.1. Biomarker distributions in Lower Cretaceous sediments
The biomarker compositions of the Lower Cretaceous sedimentsfrom Shatsky Rise (ODP Sites 1207, 1213, and 1214) have beenpreviously described (Dumitrescu andBrassell, 2005). All of the samples
contain suites of aliphatic hydrocarbons (n-alkanes, acyclic isoprenoids,steranes, and hopanes), series of ketones (notably steroidal ketones andhopanones), and tocopherols (Dumitrescu and Brassell, 2005). Thereare threenotable differences among the samples: the prevalenceof both2-methylhopanes and 2-methylhopanones in intervals correspondingto the Early Aptian (Dumitrescu and Brassell, 2005), the presence ofalkenones in one of these samples (1213B-8R-1 96-97; Brassell et al.,2004), and the occurrence of steryl ethers in the Valanginian interval(1213B-15R-1 9-10). It is the occurrence of the last suite of compoundsin this single sample that is the focus of interest herein.
3.2. Biomarker distributions in Quaternary and Cenozoic sediments
There are marked variations in the published biomarker composi-tions of the sediment samples that contain steryl ethers consideredherein (Table 1). It lies beyond the scope of this work to document thebroad range of biomarkers reported and the differences among thesamples except to note that the varied age and maturity of the sampleshave a profound influence on their composition. The younger(Quaternary to Late Miocene) samples contain suites of compounds,including sterols and alkenones, derived from marine phytoplankton(e.g. Schouten et al., 2000; Schefuß et al., 2001; Brassell et al., 1980) thatare typical constituents of diatomaceous and nannofossil oozes andclays. The Monterey sample contains suites of steroidal and hopanoidalcomponents, but its most distinctive biomarker characteristic is thedominance of organo-sulfur compounds (Schouten et al., 2001, 2005).Alkenones were the focus of investigations of the Oligocene and Eocenesamples (Marlowe, 1984; Brassell, 1993; Dzvonik, 1996), but studies ofthe Falkland Plateau sample revealed that its hydrocarbon composition

49S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
is dominated by C29 and C31 n-alkanes derived from higher plants (vonder Dick et al., 1983; Marlowe, 1984). A similar n-alkane distributionoccurred in the sediments from the Iceland–Faeroe Ridge (Marlowe,1984). Perhaps the most pertinent molecular traits from priorinvestigations of these samples (Schefuß et al., 2001; Brassell et al.,1980) are the carbon number distributions and the positions ofunsaturation of their constituent steryl moieties, whether present asalcohols, ketones, or hydrocarbons. Specifically, these characteristicsprovide a useful comparisonwith thedistributions of steryl ethers in thesame sequence of sediments.
3.3. Steryl ethers
The presence of a suite of steryl ethers (Fig. 3; Tables 1, 2 and 3) isthe most distinctive feature in the biomarker distributions of theValanginian interval from Site 1213 because these compounds werenot detected in any other Cretaceous intervals examined at Site 1213(Aptian, Berriasian) or in the Lower Aptian OM-rich sequencerecovered at Site 1207 (Dumitrescu and Brassell, 2005). GC–MSanalysis permitted partial characterization of six C27–C29 steryl ethers(Fig. 3; Table 3). It also enabled assignment of Δ5, Δ5,22 and Δ22
steroidal moieties, and recognition of a C9 side chain for some of thecompounds. These tentative structural identifications were aided bycomparison with the distributions and GC–MS characteristics (reten-tion times andmass spectra) of steryl ethers in othermarine sediment
Fig. 3. Partial GC–MS traces for fractions illustrating the distributions of sterol ethers in reprPleistocene (57-440B-3-5); (b) Iceland–Faeroe Ridge, Oligocene (38-336-15-5); (c) Shatsky Rassignments for tocopherols, steroidal ketones, hopanones, and alkenones (designated b(designated 1 to 14) are presented in Table 3.
samples of various ages (Fig. 3; Table 1), especially those previouslyanalyzed in detail (cf. Schouten et al., 2005). Each of the sedimentsamples found to contain steryl ethers tends to contain a distinct suiteof these compounds that often includes individual components notfound in other samples. These characteristics typify the diverse rangeof steryl ethers in sediments (Table 1), with the exception that asimilar variety of components occurs in the Eocene sample from theFalkland Plateau (and in Oligocene sediments from high latitudes inthe North and South Atlantic, respectively (Figs. 2, and 3; Tables 1and 2).
4. Discussion
4.1. Sedimentary occurrence and biological origins of steryl ethers
Recognition of steryl ethers in Cretaceous sediments is unprece-dented. Yet, the structural characteristics of steryl ethers in theValanginan sample from Shatsky Rise are broadly comparable to thosepreviously reported in marine sediments of various ages (Table 1) interms of the carbon number range (C26–C29) and unsaturation(prevalence of Δ5, Δ5,22; also perhaps Δ5,24) of their sterol moieties,the absence of 4-methylsteroids, and the size of their alkyl groups(C9–C11; Table 3). This structural consistency suggests that the originof these components is likely to be uniform throughout the temporal
esentative samples of different ages from four locations from Table 1. (a) Japan Trench,ise, Valanginian (198-1213B-15R-1); (d) Falkland Plateau, Eocene (71-511-16-1). Peak
y Greek symbols and letters) are given in Table 2. The assignments of steryl ethers

Table 2Peaks assignments for ketone fractions.
Peaka Assignment Abbrev.b
Tocopherolsβ/γ β- and γ-tocopherolα α-tocopherol
Steroidal ketones and hopanonesA UnknownB 5β(H)-cholestan-3-one C27 5βC 22,29,30-trisnor-17β(H)-hopan-21-one β C27
D 5α(H)-cholestan-3-one C27 5αE 24-methylcholest-22-en-3-one C28 Δ22
F 4-methyl-5α(H)-cholestan-3-one 4-Me C28
G cholest-4-en-3-one C27 Δ4
H 24-methyl-5α(H)-cholestan-3-one C28 5αI 24-methylcholesta-4,22-dien-3-one C28 Δ4,22
J 24-ethylcholest-22-en-3-one C29 Δ22
K 24-ethyl-5β(H)-cholestan-3-one C29 5βL 4,24-dimethyl-5α(H)-cholestan-3-one 4-Me C29
M 4,23,24-trimethylcholest-22-en-3-one 4-Me C30 Δ22
+ 24-methylcholest-4-en-3-one C28 Δ4
N 24-ethyl-5α(H)-cholestan-3-one C27 5αO 24-ethylcholest-4,22-dien-3-one C29 Δ4,22
P 4-methyl-24-ethyl-5α(H)-cholestan-3-one 4-Me C30
Q 24-ethylcholest-4-en-3-one C29 Δ4
R 4,23-dimethyl-24-ethylcholest-22-en-3-one 4-Me C31 Δ22
S 17β(H),21β(H)-hopanone ββ C30
Alkenonesa hexacontadien-3-one C36:2 Etb heptacontatrien-2-one C37:3 Mec heptacontadien-2-one C37:2 Med octacontatrien-3-one C38:3 Ete octacontatrien-2-one C38:3 Mef octacontadien-3-one C38:3 Etg octacontadien-2-one C38:2 Meh nonacontatrien-3-one C39:3 Eti nonacontadien-3-one C39:2 Etj nonacontadien-2-one C39:2 Me
a Peak designations in Fig. 3.b For sterones abbreviations denote 4-methyl substitution, carbon number, positions
of unsaturation and C-5 stereochemistry for sterones; for hopanones they denote carbonnumber and C-17 and C-21 stereochemistry; for alkenones they denote carbon number,degrees of unsaturation, and ketone designation based on the carbonyl group position.
Fig. 4. Stratigraphic record of sedimentary occurrences of steryl ethers in the samples(Table 1; Fig. 2).
50 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
record of their occurrence, which is here extended to the LowerCretaceous (Fig. 4).
The possibility that steryl ethers are diagenetic products derived insediments from sterol and/or steroidal ketone precursors can bediscounted because of the marked differences in the distributions of
Table 3Assignments and relative abundances of steryl ethers in sediments illustrated in Fig. 3.
Peaka Identityb Sterol moietyb Alkylb
1 C26 Δ5,22 C10 24-norcholesta-5,22-dienyl decyl2 C27 Δ5,22C9 cholesta-5,22-dienyl nonyl3 C27 Δ5C9 cholest-5-enyl nonyl4 C27 Δ5C9 cholest-5-enyl nonyl5 C28 Δ5,22C9 24-methylcholesta-5,22-dienyl nonyl6 C27 Δ5C10 cholest-5-enol decyl7 C28 Δ5,24C9 24-methylcholesta-5,24-dienyl nonyl8 C29 Δ5 24-ethylcholest-5-enyl ?9 C28 Δ5,22 C10 24-methylcholesta-5,22-dienyl decyl10 C29 Δ22 24-ethylcholest-22-enyl ?11 C29 Δ5 C9 24-ethylcholest-5-enyl nonyl12 C28 Δ5,22 C11 24-methylcholesta-5,22-dienyl undecyl13 C28 Δ5,22 norcholesta-5,22-dienyl ?14 C29 Δ5 C11 24-ethylcholest-5-enyl undecyl
a Peak designation in Fig. 3.b Assignment of the carbon number, positions of unsaturation and chain length of the alky
could not be determined from either the mass spectrum or the GC retention characteristicsc M+•=molecular ion; n.d. designates that the M+• ion was not detected.d Quat.=Quaternary (Japan Trench; Fig. 3a); Olig.=Oligocene (Iceland–Faeroe Ridge; Fige +, ++, and +++ represent approximate relative concentrations.
sterol moieties for the ethers versus their putative precursors amongother steroidal components (Brassell et al., 1980; Schouten et al.,2000; Schefuß et al., 2001; Brassell, 1993). Moreover, there are no
M+•c Quatd Oligd Eocd Vald
510 +e
510 +512 ++ ++512 + + +524 ++ +526 +++ + ++ +524 +n.d. +538 ++ ++ +++n.d. ++n.d. ++552 +n.d. +n.d. +
l moiety are determined from mass spectral data. ? indicates that the size of alkyl chain.
. 3b); Eoc.=Eocene (Falkland Plateau; Fig. 3d); Val.=Valanginian (Shatsky Rise; Fig. 3c).

51S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
readily available precursors for the 3-alkyl chains in the steryl ethersand their observed structural variety precludes formation by non-selective diagenetic reactions. Thus, a biological origin, directly orindirectly, for steryl ethers seem eminently more plausible. Theinference that steryl ethers might derive from diatoms (Boon and deLeeuw, 1979) was prompted by their occurrence in Cenozoic andQuaternary diatomaceous oozes (Table 1; Fig. 4). A diatom origin isplausible for the Valanginian since these algae originated in the earlyJurassic (Harwood et al., 2007; Sorhannus, 2007) and existed asseveral lineages prior to themajor diversification of marine diatoms inthe Lower Cretaceous (Sinninghe Damsté et al., 2004). However,steryl ethers have yet to be found in diatoms, or indeed any culturedalga (Schouten et al., 2005). The structural diversity of steryl ethersand their varied distributions seems more consistent with multiplecontrols (e.g. precursor compounds, biota, environment) on theirformation rather than production directly by biosynthesis by a specificclass of alga, whichwould be expected to yield amore restricted rangeof components in terms of both their steroidal nuclei and side chainmoieties. One likely candidate origin is formation by zooplanktoningestion that involves selective uptake of specific phytoplanktonsterols. The preferential metabolism of specific sterols by zooplankton(Harvey et al., 1987, 1989; Talbot et al., 2000) and their discriminationagainst 4-methylsterols can explain the compositional differencesbetween steryl ethers and co-occurring steroidal compounds, includ-ing the absence of 4-methylsterol moieties in the former (Brassellet al., 1980; Schouten et al., 2005), augmented by a prevalence of Δ5
and Δ5,22 steryl moieties. This interpretation parallels the processesrecognized in formation of chlorin steryl esters, namely that theyrepresent products from selective sterol assimilation by specificzooplankton (Talbot et al., 1999a,b, 2000; King and Wakeham, 1996;Pearce et al., 1998; Harradine et al., 1996; Chen et al., 2003).Formation of steryl ethers as byproducts of zooplankton grazing canalso explain why the distribution of steryl moieties present as ethersdiffers from that occurring as steroidal alcohols, hydrocarbons, and/orketones. Such differences can be attributed to the likelihood thatmultiple, distinct pools of sterols exist that experience varied fates inthe marine environment coupled to their exposure to predation,diagenetic alteration, or other depositional factors (cf. Schefuß et al.,2001). For example, 23,24-dimethylsteryl moieties, likely originatingfrom diatoms (Rampen et al., 2009), occur in several steroidalfractions (i.e. sterols, sterones, sterenes) for many of the samples,although they are not present in the Lower Cretaceous samples fromShatsky Rise (Bralower et al., 2002; Dumitrescu and Brassell, 2005).The apparent absence of this specific side chain among steryl ethers(Table 3) suggests that these compounds originate from discretebiosynthetic precursors via prescribed pathways rather than non-selective processes affecting all sterols. Recognition of a C26 sterylether in a Quaternary sediment from the Japan Trench (Table 1)suggests a planktonic origin for these compounds, or their precursors,given that C26 sterols are synthesized by diatoms and dinoflagellates(Rampen et al., 2007). Alternatively, the C26 component could beformed by dietary modification of the side chain of a higher sterolhomologue, a process that occurs in sponges (Kerr et al., 1992), whichare recognized as contributing organisms to the sterol distributions inthis environment (Brassell et al., 1980; Brassell and Eglinton, 1983).
These considerations strengthen the likelihood that steryl etherscould be formed as byproducts from phytoplankton grazing byzooplankton, wherein they may fulfill a role as storage lipids, akinto that of wax esters, including steryl esters, and triacylglycerols(Hagen et al., 1996; Campbell and Dowe, 2003). Individual species ofzooplankton exhibit compositional preferences (i.e. wax esters vs.triacylglycerols) in their storage lipids (Ju and Harvey, 2004), andperhaps steryl ethers play a similar role in other, as yet uncharacter-ized species. It is noteworthy that steryl ethers with alkyl side chainlengths comparable to those reported in sediments exhibit a smectic Aphase and possess liquid crystal properties (Vill and Weber, 1991,
1994). Such ‘stacking’ properties might be advantageous for storagelipids, which are accumulated for wintering by zooplankton speciesthat inhabit seasonally productive waters (e.g. Calanus spp.; Milleret al., 1998; Hagen and Auel, 2001). The timing of the development ofa biophysiology in zooplankton that facilitates this behavior isunknown, although the evolutionary record of these organismspredates the Cretaceous (Bradford-Grieve, 2002). Production ofstorage lipids, however, is not restricted to zooplankton as algaeand bacteria also synthesize wax esters and triacylglycerols for thispurpose (e.g. Sheia et al., 1991; Wälterman and Steinbüchel, 2005).However, the possibility of bacteria or Archaea as a source of sterylethers is precluded by the absence in organismswithin these domainsof the biosynthetic pathways that yield 4-desmethylsteroids (Ourissonet al., 1982). Thus, steryl ethers likely originate from eukaryoticphytoplankton or from zooplankton.
If steryl ethers do fulfill a role as zooplankton storage lipids theywould be expected to occur in marine environments characterized byseasonal productivity, perhaps coupled to periodic upwelling. Theymight also be associated with the species of zooplankton that surviveat depth in cold regions by adopting a seasonal dormant state calleddiapause (Hagen and Auel, 2001). Alternatively, algae may synthesizethem in regions where the seasonal supply of nutrients favorsorganisms that are capable of producing storage lipids. Such sourcesfor steryl ethers are speculative, but these compounds only occur inTertiary and Quaternary sediments deposited in marine settingscharacterized by cool waters or seasonal upwelling (Table 1). Thus,the environmental conditions conducive for production of sterylethers are constrained, even though their precise biological source isnot defined. This consideration prompts evaluation of the depositionalsetting of the Valanginian sediment that contains steryl ethers todetermine whether it also represents an environment characterizedby seasonal productivity or cooling.
4.2. Lower Cretaceous productivity
Lithologic, micropaleontological and geochemical evidence for theEarly Aptian OAE1a in several Tethyan records indicate that it was ahigh productivity event (Hochuli et al., 1999; Bersezio et al., 2002;Bellanca et al., 2002; Erba and Tremolada, 2004; Weissert and Erba,2004; Luciani et al., 2001; Price, 2003). Similarly, the Late Valanginianis recognized as a time interval characterized by enhanced primaryproductivity in Tethyan sequences (Bersezio et al., 2002; Erba andTremolada, 2004). For example, radiolaria provide evidence fornutrient-rich surface waters and upwelling (Weissert et al., 1998;Jud, 1994; Erba, 1994), although elsewhere stratigraphic variations inOM, and nannoflora suggest that such nutrification episodes wereintermittent (Reboulet et al., 2003), and inversely related to carbonateproduction (Reboulet et al., 2003; Wortmann and Weissert, 2000). Inthe Vocontian basin the enhanced fertility during the Valanginian isattributed to increased weathering and runoff (van de Schootbruggeet al., 2000; Duchamp-Alphonse et al., 2007), but this explanationcannot readily account for enhanced productivity in the mid-Pacific.However, recent investigationof nannofossil assemblages in Berriasian–Huterivian sequences from theWestern Atlantic reveals increases in theproportion of taxa associated with elevated nutrient levels during theLower Valanginian (zones NK2b/NK3a) that attest to enhancedproductivity (Fig. 5; Bournemann and Mutterlose, 2008). The increasein ocean productivity is attributed to intensified upwelling, perhapsassociated with the initial minor phase of Paraná–Etendeka volcanism(Fig. 5c; Bournemann and Mutterlose, 2008), clearly preceding thepositive carbon isotopic excursion associated with the Late Valanginian(Erba et al., 2004).
On Shatsky Rise at Site 1207 the accumulation rates for OM duringthe Early Aptian range from 0.1 to 2 g cm−2y−1, which are consistentwith intermittent high levels of plankton productivity (Dumitrescuand Brassell, 2005, 2006). Sediments rich in OM that correspond to

52 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57

53S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
OAE1a occur at Site 1213 as intervals interbedded with claystones,cherts and porcellanites. Geochemically they resemble the OAE1asequence recovered as a single unit at Site 1207 (Brassell et al., 2004),prompting a similar interpretation that they also reflect episodes ofenhanced productivity. At Site 1213 the organic carbon contents ofthe OM-rich Valanginian and Berriasian intervals fall within the lowerend of the range of values for the Lower Aptian sediments (Brassellet al., 2004). Sedimentation rates for these intervals are poorlyconstrained, with estimates for the Lower Hauterivian to Berriasianinterval of 12–16 mm y−1 based on the combined evidence ofcalcareous nannofossils, radiolarians, and planktonic foraminifers(Bralower et al., 2002). This range corresponds to an OM accumula-tion rate that exceeds 20 g cm−2y−1 for the OM-rich intervals,although their intermittent character suggests that such rates werenot sustained, and cannot preclude the possibility that sedimentaccumulation rates were significantly higher and/or lower during theepisodes of enhanced OM sequestration.
The nature of the OM in all of the OM-rich Cretaceous sediments atSite 1213 shows many similarities, exemplified by the distribution ofplanktonic biomarkers (Bralower et al., 2002; Dumitrescu andBrassell, 2005), although the Valanginian and Berresian intervals arecalcareous in contrast to the Lower Aptian samples. Thus, the generalcharacter of phytoplankton productivity recorded in intervals ofenhanced OM burial appears broadly consistent throughout thisstratigraphic section. However, there are differences in the relativecontributions from cyanobacteria (Dumitrescu and Brassell, 2005),and the appearance of steryl ethers in only the OM-rich Valanginianinterval sustains the presumption that their occurrence is associatedwith specific sources of OM, akin to the recognition of alkenonesderived from haptophyte algae in a single Lower Aptian sample(Brassell et al., 2004). Such differences in biomarker compositionssuggest temporal changes in biota likely coupled to environmentalvariations manifest at Shatsky Rise (e.g. Dumitrescu et al., 2006) thatmay relate to widespread, potentially global, perturbations of thebiogeochemical carbon cycle.
4.3. Global correlation of Early Cretaceous OM-rich intervals
Many OM-rich intervals have been reported in the EarlyCretaceous, although only those associated with OAE-1a and itsnegative 13C excursion have been proven to occur globally (e.g.Menegatti et al., 1998; Hochuli et al., 1999; Leckie et al., 2002;Dumitrescu and Brassell, 2006). In Pacific sequences, OM-richintervals earlier than OAE-1a are rare (e.g. Sliter, 1989), whichenhances the potential significance of correlations between theValanginian and Berriasian horizons recovered at Sites 1213 and thevarious OM-rich horizons of similar age in Tethyan and Atlanticsettings. Specifically, similarities in the timing or character of intervalsof enhanced OM sequestration in geographically disparate regionswould suggest that such episodes might have been favored by globalrather than local influences propitious to their formation.
Fig. 5. A compilation of paleoenvironmental and climatic evidence for part of the Lower Cderived from original publications have been reinterpreted to match the zones and stagecorresponding to the OM-rich interval within the Valanginian at Shatsky Rise is highlightedfrom Breggia, Capriolo and Polaveno (Lini et al., 1992) and SE France (Duchamp-AlphonseWestern N. Atlantic (redrawn by Lini et al., 1992 from others' data; Bournemann and MutterCO2: estimates through the early Cretaceous from the range calculated from Geocarb modelplus the range determined from δ13C values of porphyrins within an Upper Valanginian claysLarson, 1991a,b used by Mutterlose and Kessels, 2000) combined with the timing an(d) Nannoplankton and floral changes: widespread evidence of nannoconnid decrease (Bobservations of floral changes within the Tethyan and boreal realms (Melinte and Mutter(e) Productivity: assessments of changes in productivity based on the relative abundance of nAtlantic (Bournemann and Mutterlose, 2008). (f) Paleotemperatures: a compilation of temp2000), Svalbard (Ditchfield, 1997), the Vocontian Basin (van de Schootbrugge et al., 2000(Mutterlose and Kessels, 2000 after Podlaha et al., 1998). The temperature trends forWesternfor planktonic foraminifera (Barrera, 1994), respectively. Comparative temperatures for SE Frafted deposits: temporal range based on published compilations (Frakes and Francis, 1988
Calcareous nannoplankton assemblages place the intervalsenriched in OM in cores 15R and 19R at Site 1213 within ZonesNK3a and NK2, respectively, which constrains their ages as Valangi-nian and Berriasian (Bown, 2005). Additional OM-rich intervals mayexist within the Lower Cretaceous at Site 1213 that were not sampledbecause of poor core recovery. However, direct correlation betweeneither of the Valanginian and Berriasian OM-rich intervals in thePacific at Site 1213 and any OM-rich horizons occurring in Atlanticand Tethyan sequences are precluded by insufficient temporalresolution. Irrespective of this constraint it is significant that theassigned biostratigraphic zonation of core 15R (nannofossil zoneNK3a) at Site 1213 precedes both the Valanginian δ13C excursion(Channell et al., 1993; Erba et al., 2004; Gröcke et al., 2005; Fig. 5a)and themajority of associated OM-rich intervals (Bersezio et al., 2002;Erba et al., 2004) within the Tethyan realm. This temporal separationsuggests that oceanic conditions may have been conducive toincreased sequestration of OM prior to the positive excursion incarbon isotopic signals linked to perturbation of atmospheric CO2
levels by the Paraná–Etendeka volcanic event (Fig. 5c), although onecarbon cycle model suggests a gradual decrease through theValanginian (Tajika, 1999; Fig. 5b). However, the Lower ValanginianOM-rich interval at Site 1213 does appear to correspond with theupwelling-induced increase in productivity in the Western Atlantic(Bournemann and Mutterlose, 2008; Fig. 5e). Enhanced OM seques-tration within the Lower Valanginian is reported in the VocontianBasin (Reboulet et al., 2003), and in the Southern Alps (Bersezio et al.,2002; Erba et al., 2004; Erba and Tremolada, 2004) where OM-richBerriasian horizons also occur (Bersezio et al., 2002). Indeed, LowerCretaceous sequences at a number of locations in the Atlantic (DSDPSites 105, 391, 534, 535, and 603) contain intercalated OM-richclaystone and limestone intervals (e.g. Rullkötter andMukhopadhyay,1986; Rullkötter et al., 1986; van de Schootbrugge et al., 2000),including horizons assigned as Lower Valanginian or Upper Berriasian(i.e. within nannofossil zones NK2 or NK3a; e.g. Herbin et al., 1983,1984, 1987; Summerhayes andMasran, 1983; Katz, 1984; Patton et al.,1984; Rullkötter et al., 1984, 1987; Rullkötter and Mukhopadhyay,1986; Meyers, 1987). Thus, episodes of enhanced OM sequestrationare awidespread featurewithin these substages, typically occurring asdiscrete thin horizons intercalatedwith lithologies lean in OM, in starkcontrast to the more extensive sequences associated with OAE-1a.
The origin and nature of the OM in individual horizons andsequences can provide evidence of likely environmental or othercontrols on its sequestration. In Tethys the dominant source of OM formost of the Lower Valanginian samples is terrestrial (Bersezio et al.,2002), an assessment confirmed by the distributions of source-diagnostic biomarkers in sequences from the Southern Alps (Bersezioet al., 2002). Many Valanginian deposits from Atlantic continentalmargins are comprised of type III kerogen and contain series of higherplant n-alkanes that confirm the terrestrial affinities of their OM (e.g.Herbin et al., 1983, 1987). These deposits have been attributed toepisodes of increased runoff associated with an active hydrologic cycle
retaceous. The temporal trends and stratigraphic information for all of these data setsboundaries used in ODP Leg 198 (Bralower et al., 2002; Bown, 2005). The NK3a zone(checkered). (a) Carbon isotopes: δ13C values for carbonate from Tethyan sequenceset al., 2007), ocean drilling sites in the Gulf of Mexico, in the central Pacific, and thelose, 2008), and from the western equatorial Pacific (Erba et al., 2004). (b) Atmosphericing (Berner and Kothavala, 2001) and trends from a carbon cycle model (Tajika, 1999),tone (Popp et al., 1989). (c) Crust production: estimates of ocean crust production (fromd relative magnitude of Paraná flood basalts (van de Schootbrugge et al., 2000).ersezio et al., 2002; Erba et al., 2004; Erba and Tremolada, 2004) is accompanied bylose, 2001) and the presence of Tethyan taxa in Greenland (Mutterlose et al., 2003).annofossil taxa associated with higher nutrient levels in assemblages from theWesternerature estimates based on trends in δ18O for belemnites from Speeton clay (Price et al.,), Tethyan and Boreal realms (van de Schootbrugge et al., 2000), and high latitudesTethys and Shatsky Rise are based on δ18O values for fish teeth (Pucéat et al., 2003) and
rance and SE Spain are based on belemnite Mg/Ca ratios (McArthur et al., 2007). (g) Ice-; Price 1999; Mutterlose and Kessels, 2000).

54 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
and changes in sea level (van de Schootbrugge et al., 2000; Duchamp-Alphonse et al., 2007). Coaly material also occurs in some intervals,notably at Site 603 off Cape Hatteras (Rullkötter and Mukhopadhyay,1986; Meyers, 1987), whereas marine type II OM is prevalent furtherfrom land (notably Site 535; Herbin et al., 1984). Moreover, OM-richUpper Berriasian/Lower Valanginian samples from both the SouthernAlps (Bersezio et al., 2002) and Site 603 (Rullkötter et al., 1987) containamorphous OM coupled with suites of planktonic biomarkers thatresemble the distributions of these components associated withautochthonous marine OM in contemporaneous sediments from Site1213 (Bralower et al., 2002; Dumitrescu and Brassell, 2005).
The sequestration of OM during the Late Berriasian/Early Valangi-nian appears to reflect both geographic controls and levels of oceanproductivity. On continental margins enhanced OM results from aninflux of terrestrial OM without the need to invoke global changes inmarine productivity. However, there is evidence for enhancedphytoplankton productivity in locations remote from continentalinfluence based on evidence from OM composition and nannofossilassemblages (Bournemann and Mutterlose, 2008; Fig. 5e). However,increased Lower Valanginian productivity does not appear to beassociatedwith development of conditions favorable for specific biota,as exemplified by the prominence of cyanobacteria and nitrogenfixation during OAE-1a (Kuypers et al., 2004; Dumitrescu and Brassell,2005), and the expansion of Archaea during OAE-1b (Kuypers et al.,2001, 2002).
4.4. Valanginian climate
The association of steryl ethers with cooler waters or upwelling inthe Tertiary (Table 1) prompts consideration of indicators for similarclimatic or oceanographic conditions that might explain their occur-rence in theValanginian, in concertwithevidence relating increasedOMsequestration with global-scale perturbations of the carbon cycle.Sediment sequences from several regions provide evidence for EarlyCretaceous (Late Berriasian to Late Hauterivian) climate through avariety of measures – geochemical, paleontological, sedimentologicaland tectonic – that can be coupledwith global estimates of atmosphericCO2 levels derived from geochemical models (Fig. 5b).
Carbonates from the three major Cretaceous ocean basins(Atlantic, Tethys, and Pacific), belemnites from the Boreal–Arctic Sea(Price and Mutterlose, 2004), and terrestrial plants from Tethys(Gröcke et al., 2005) all exhibit a positive δ13C excursion within theValanginian (Fig. 5a; Lini et al., 1992; Hennig et al., 1999; van deSchootbrugge et al., 2000; Erba et al., 2004). This isotope excursionhas been linked to an increase in atmospheric CO2 levels attributed toenhanced volcanic activity (Lini et al., 1992) in turn associated withemplacement of the Paraná–Etendeka flood basalts (Fig. 5c; van deSchootbrugge et al., 2000). A reduced flux in nannofossil carbonateduring the Early Cretaceous is also attributed to enhanced pCO2 (Erbaand Tremolada, 2004). However, climate models based on geochem-ical data suggest a long-term progressive decrease in concentrationsof atmospheric CO2 during the Early Cretaceous (Fig. 5b; Berner andKothavala, 2001; Tajika, 1999), including little change during theValanginian (Wallmann, 2001; Hansen and Wallmann, 2003),although the temporal resolution of these models cannot discernshort-term fluctuations. The positive δ13C excursion in the UpperValanginian appears therefore to reflect an increase in the sequestra-tion of OM coupled to higher levels of primary productivity inducedby enhanced runoff, and associated with a general cooling trend(Weissert and Erba, 2004; Erba et al., 2004; Gröcke et al., 2005).Evidence for cooling during the Valanginian in various regions,including Tethys and high northern latitudes, is provided by severaltemperature proxies (i.e. δ18O values for foraminifera, belemnites andfish teeth, Mg/Ca ratios for belemnite calcite; Fig. 5f; Barrera, 1994;van de Schootbrugge et al., 2000; Price et al., 2000; Mutterlose andKessels, 2000; Pucéat et al., 2003; McArthur et al., 2007). Temperature
data derived from measurements of the varied substrates differ interms of the confidence afforded to the absolute values because ofconcerns regarding sample preservation (e.g. overgrowths on fora-minifera; Barrera, 1994). Similarly, variations in paleotemperaturesamong locations may, in part, reflect the imprecision of stratigraphiccorrelations compounded by use of different chronologies for theEarly Cretaceous. Yet, the temperature trends for different regions,including high latitudes and the record from Shatsky Rise (Barrera,1994), predominantly indicate cooling during the Valanginian withinnannofossil zone NK3a (Fig. 5f), although peak cooling has also beenplaced at the NK3b/NC4a boundary (Erba et al., 2004). Biotic changesalso occur during this biostratigraphic interval, especially the globaldecrease in nannoconnids that extends into nannofossil zone NC4a,which is attributed to a biocalcification crisis induced globally byelevated pCO2 (Fig. 5d; Bersezio et al., 2002; Erba et al., 2004; Erba andTremolada, 2004) and reinforced in Tethys by enhanced nutrientsupply triggered by increased terrestrial runoff (Duchamp-Alphonseet al., 2007). Other variations in floral and faunal assemblages havebeen observed at this time, notably changes in the paleobiogeogra-phical distributions of Boreal and Tethyan nannofossil taxa (Fig. 5d;Price et al., 2000; van de Schootbrugge et al., 2000; Melinte andMutterlose, 2001). The appearance of boreal species within theTethyan realm during the early Late Valanginian (NK3a; Fig. 5d) hasbeen attributed to changes in sea level that opened interconnectionsbetween floral provinces rather than a marked change in oceantemperatures (Melinte and Mutterlose, 2001). However, any rise insea level would likely reflect tectonic influences rather than anymelting episode given that the fauna changes coincide with evidencefor cooling (e.g. Gröcke et al., 2005). Indeed, the paleobiogeographicaldistributions of both foraminifera and calcareous nannofossils implypronounced latitudinal temperature gradients during the Valanginian(Mutterlose and Kessels, 2000; Mutterlose et al., 2003), consistentwith formation of deep water in the Arctic (Mutterlose et al., 2003)and glaciation (e.g. Gröcke et al., 2005). In addition, recognition of ice-rafted deposits throughout the Early Cretaceous (Mutterlose andKessels, 2000; Price, 1999, after Frakes and Francis, 1988) attests toglaciers in both northern and southern hemispheres (Fig. 5g).
Thus, collectively, critical aspects of Valanginian paleoclimate,specifically indicators of increased nutrient supply (e.g. Erba et al.,2004; Duchamp-Alphonse et al., 2007) combined with the potentialoutflow of Arctic deep water into the Pacific (Mutterlose et al., 2003),and with evidence for cooling from paleotemperature proxies (van deSchootbrugge et al., 2000; Price et al., 2000; Mutterlose and Kessels,2000; Pucéat et al., 2003; McArthur et al., 2007; Fig. 5f), providestrong support for the contention that production of steryl etherspotentially represents a biogeochemical response to cool waters.Moreover, Shatsky Rise lay within the equatorial divergence zoneduring this interval of the early Cretaceous (Fig. 1), which suggeststhat the occurrence of steryl ethers may be attributed to intensifiedupwelling in the equatorial Pacific, and more rigorous circulationpotentially triggered by developing ice-house conditions. Indepen-dent evidence for increased productivity attributed to upwelling atthis time is provided by temporal variations in nannofossil assem-blages in the Western Atlantic (Bournemann and Mutterlose, 2008;Fig. 5e). The synchroneity of these events in the Atlantic and Pacificsuggests that global changes in oceanic circulation may have occurredduring the Early Valanginian.
5. Conclusions
A series of steryl ethers occurs among the biomarkers identified ina Valanginian horizon from a sequence of Cretaceous sediments fromShatsky Rise that contain exceptionally well-preserved OM. Thesource for sedimentary steryl ethers remains enigmatic, but appearsbiological rather than diagenetic. The possibility of steryl ethergeneration by zooplankton in a manner akin to production of chlorin

55S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
steryl esters merits further assessment. The substantial extension tothe earliest occurrence of steryl ethers furthers their evolutionaryrecord, which may reflect a biological adaptation linked to energystorage in environments prone to seasonal variation in nutrientsupply. Moreover, if steryl ethers act as storage lipids it seemsplausible that increased seasonality during the early Cretaceous(Steuber et al., 2005) may have been a factor contributing to thisbiosynthetic innovation within the oceanic realm.
Irrespective of their biological origins and synthesis, the sedimen-tary occurrences of steryl ethers demonstrate that their production isconstrained by oceanic conditions, as these components only occur inenvironments characterized by cool waters or upwelling. Evidence foran enhancement of nannofossils associated with higher productivitylevels during Zone NK3a in the Western Atlantic supports thisassertion. The presence of steryl ethers in an Early Valanginiansediment from the Pacific is consistent with this assessment,augmenting evidence for enhanced upwelling during this interval ofthe Lower Cretaceous.
Acknowledgements
I should like to specially thank Anne Pimmel and Tim Bronk for theirassistance with geochemical analyses in the chemistry laboratoryonboard ODP Leg 198 and all R/V JOIDES Resolution marine techniciansfor their contributions to the successful cruise. I thank Stefan Schouten(NIOZ) for sharing insights on the structures of steryl ethers and formercolleagues at the University of Bristol where this study began. Iappreciate thehelpful comments and suggestionsof SebastiaanRampenand another anonymous reviewer that improved this contribution. Thisresearch used samples and data provided by theOceanDrilling Program(ODP). ODP was sponsored by the U.S. National Science Foundation(NSF) and participating countries under management of Joint Ocean-ographic Institutions (JOI), Inc. I am grateful to USSSAC and thePetroleumResearch Fund of theAmerican Chemical Society forfinancialsupport, and to the Hanse Wissenschaftskolleg for a fellowship.
References
Ando, A., Kakegawa, T., Takashima, R., Saito, T., 2002. New perspective on Aptian carbonisotope stratigraphy: data from δ13C records of terrestrial organic matter. Geology30, 227–230.
Arthur, M.A., Brumsack, H.-J., Jenkyns, H.C., Schlanger, S.O., 1990. Stratigraphy,geochemistry, and paleoceanography of organic carbon-rich Cretaceous intervals.In: Ginsburg, R.N., Beaudoin, B. (Eds.), Cretaceous Resources, Events and Rhythms.Kluwer, Dordrecht, pp. 75–119.
Arthur, M.A., Schlanger, S.O., 1979. Cretaceous oceanic anoxic events as causal factors indevelopment of reef-reservoired giant oil-fields. Bulletin of the AmericanAssociation of Petroleum Geologists 63, 870–885.
Arthur, M.A., Schlanger, S.O., Jenkyns, H.C., 1987. The Cenomanian–Turonian oceanicanoxic event, II. Palaeoceanographic controls on organic-matter production andpreservation. In: Brooks, J., Fleet, A.J. (Eds.), Marine Petroleum Source Rocks:Geological Society, London, Special Publications, vol. 26, pp. 401–420.
Barrera, E., 1994. Global changes preceding the Cretaceous–Tertiary boundary; Early–late Maastrichtian transition. Geology vol. 22, 877–880.
Beerling, D.J., Lomas, M.R., Gröcke, D.R., 2002. On the nature of methane gas-hydratedissociation during the Toarcian and Aptian oceanic anoxic events. AmericanJournal of Science 302, 28–49.
Bellanca, A., Erba, E., Neri, R., Premoli Silva, I., Sprovieri, M., Tremolada, F., Verga, D.,2002. Palaeoceanographic significance of the Tethyan ‘Livello Selli’ (Early Aptian)from the Hybla Formation, northwestern Sicily: biostratigraphy and high-resolution chemostratographic records. Palaeogeography, Palaeoclimatology,Palaeoecology 185, 175–196.
Berner, R.A., Kothavala, Z., 2001. GEOCARB III: a revised model of atmospheric CO2 overPhanerozoic time. American Journal of Science 301, 182–204.
Bersezio, R., Erba, E., Gorza, M., Riva, A., 2002. Berriasian–Aptian black shales of theMaiolica formation (Lombardian Basin, Southern Alps, Northern Italy): local andglobal events. Palaeogeography, Palaeoclimatology, Palaeoecology 180, 253–275.
Boon, J.J., de Leeuw, J.W., 1979. The analysis of wax esters, very long mid-chain ketonesand sterol ethers isolated fromWalvis Bay diatomaceous ooze. Marine Chemistry 7,117–132.
Bournemann, A., Mutterlose, J., 2008. Calcareous nannofossil and δ13C records from theEarly Cretaceous of the western Atlantic Ocean: evidence for enhanced fertilizationacross the Berriasian–Valanginian transition. Palaios 23, 821–832.
Bown, P., 2005. Early to mid-Cretaceous calcareous nannoplankton from the northwestPacific Ocean, Leg 198, Shatsky Rise. In: Bralower, T.J., Premoli Silva, I., Malone, M.J.(Eds.), Proceedings of the Ocean Drilling Program: Scientific Results, vol. 198.http://www-odp.tamu.edu/publications/198_SR/103/103.htm.
Bradford-Grieve, J.M., 2002. Colonization of the pelagic realm by calanoid copepods.Hydrobiologia 485, 223–244.
Bralower, T.J., Arthur, M.A., Leckie, R.M., Sliter, W.V., Allard, D.J., Schlanger, S.O., 1994.Timing and paleoceanography of oceanic dysoxia/anoxia in the late Barremian toearly Aptian. Palaios 9, 335–369.
Bralower, T.J., CoBabe, E., Clement, B., Sliter, W.V., Osburn, C.L., Longoria, J., 1999. Therecord of global change in mid-Cretaceous (Barremian–Albian) sections from theSierra Madre, northeastern Mexico. Journal of Foraminiferal Research 29, 418–437.
Bralower, T.J., Premoli Silva, I., Malone, M.J., et al. (Eds.), 2002. Proceedings of the OceanDrilling Program, Initial Reports, vol. 198.
Brassell, S.C. 1980. The origin and fate of lipids in the Japan Trench. Ph.D. thesis,University of Bristol, U.K., 254 pp.
Brassell, S.C., 1993. Applications of biomarkers for delineating marine paleoclimatefluctuations during the Quaternary. In: Engel, M.H., Macko, S.A. (Eds.), OrganicGeochemistry. Plenum, New York, pp. 699–738.
Brassell, S.C., 1994. Isopentenoids and geochemistry. In: Nes, W.D. (Ed.), Isopentenoidsand Other Natural Products: Evolution and Function: American Chemical SocietySymposium Series, vol. 562, pp. 2–30.
Brassell, S.C., Eglinton, G., 1983. Steroids and triterpenoids in deep sea sediments asenvironmental and diagenetic indicators. In: Bjorøy, M., et al. (Ed.), Advances inOrganic Geochemistry 1981. Wiley, Chichester, pp. 684–697.
Brassell, S.C., Eglinton, G., 1986. Molecular geochemical indicators in sediments. In:Sohn, M. (Ed.), Organic Marine Geochemistry: American Chemical SocietySymposium Series, vol. 305, pp. 10–32.
Brassell, S.C., Comet, P.A., Eglinton, G., Isaacson, P.J., McEvoy, J., Maxwell, J.R., Thomson,I.D., Tibbetts, P.J.C., Volkman, J.K., 1980. The origin and fate of lipids in the JapanTrench. In: Douglas, A.G., Maxwell, J.R. (Eds.), Advances in Organic Geochemistry1979. Pergamon, Oxford, pp. 375–392.
Brassell, S.C., Dumitrescu, M., ODP Leg 198 Shipboard Science Party, 2004. Recognitionof alkenones in a lower Aptian porcellanite from the west–central Pacific. OrganicGeochemistry 35, 181–188.
Brassell, S.C., Eglinton, G., Howell, V.J., 1987. Palaeoenvironmental assessment formarine organic-rich sediments usingmolecular organic geochemistry. In: Brooks, J.,Fleet, A.J. (Eds.), Marine Petroleum Source Rocks: Geological Society, London,Special Publications, vol. 26, pp. 79–98.
Brassell, S.C., Eglinton, G., Maxwell, J.R., 1983. The geochemistry of terpenoids andsteroids. Biochemical Society Transactions 11, 575–586.
Campbell, R.W., Dowe, J.F., 2003. Role of lipids in the maintenance of neutral buoyancyby zooplankton. Marine Ecology Progress Series 263, 93–99.
Channell, J.E.T., Erba, E., Lini, A., 1993. Magnetostratigraphic calibration of the LateValanginian carbon isotope event in pelagic limestones from Northern Italy andSwitzerland. Earth and Planetary Science Letters 118, 145–166.
Chen, N.H., Bianchi, T.S., Bland, J.M., 2003. Novel decomposition products of chlorophylla in continental shelf (Louisiana shelf) sediments: formation and transformation ofcarotenol chlorin esters. Geochimica et Cosmochimica Acta 67, 2027–2042.
Danelian, T., Tsikos, H., Gardin, S., Baudin, F., Bellier, J.-P., Emmanuel, L., 2004. Globaland regional palaeoceanographic changes as recorded in the mid-Cretaceous(Aptian–Albian) sequence of the Ionian zone (NW Greece). Journal of theGeological Society (London) 161, 703–709.
deGea,G.A., Castro, J.M.,Aguado, R., Ruiz-Ortiz, P.A., Company,M., 2003. LowerAptian carbonisotope stratigraphy from a distal carbonate shelf setting: the Cau section, Prebetic zone,SE Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 200, 207–219.
Didyk, B.M., Simoneit, B.R.T., Brassell, S.C., Eglinton, G., 1978. Organic geochemicalindicators of palaeoenvironmental conditions of sedimentation. Nature 272, 216–222.
Ditchfield, P.W., 1997. High northern palaeolatitude Jurassic–Cretaceous palaeotem-perature variation: new data from Kong Karls Land, Svalbard. Palaeogeography,Palaeoclimatology, Palaeoecology 130, 163–175.
Duchamp-Alphonse, S., Gardin, S., Fiet, N., Bartolini, A., Blamart, D., Pagel, M., 2007.Fertilization of the northwestern Tethys (Vocontian basin, SE France) during theValanginian carbon isotope perturbation: evidence from calcareous nannofossils andtraceelementdata. Palaeogeography, Palaeoclimatology, Palaeoecology243, 132–151.
Dumitrescu, M., Brassell, S.C., 2005. Biogeochemical assessment of sources of organicmatter and paleoproductivity during the Early Aptian oceanic anoxic event atShatsky Rise, ODP 198. Organic Geochemistry 36, 1002–1022.
Dumitrescu, M., Brassell, S.C., 2006. Compositional and isotopic characteristics oforganic matter for the Early Aptian oceanic anoxic event at Shatsky Rise, ODP Leg198. Palaeogeography, Palaeoclimatology, Palaeoecology 235, 168–191.
Dumitrescu, M., Brassell, S.C., Schouten, S., Hopmans, E.C., Sinninghe Damsté, J.S., 2006.Instability in tropical Pacific sea surface temperatures during the Early Aptian.Geology 34, 833–836.
Dzvonik, J.P., 1996. Alkenones as records of oceanic paleotemperatures: studies of Eoceneand Oligocene sediments from the north, south, and equatorial Atlantic. M.S. thesis,Indiana University. 83 pp.
Erba, E., 1994. Nannofossils and superplumes: the early Aptian “nannoconid crisis”.Paleoceanography 9, 483–501.
Erba, E., Bartolini, A., Larson, R.L., 2004. The Valanginain Weissert OAE. Geology 32,149–152.
Erba, E., Tremolada, F., 2004. Nannofossil carbonate fluxes during the early Cretaceous:phytoplankton response to nutrification episodes, atmospheric CO2, and anoxia.Paleoceanography 19, 2003PA000884.
Frakes, L.A., Francis, J.E., 1988. A guide to Phanerozoic cold polar climates from highlatitude ice-rafting in the Cretaceous. Nature 333, 547–549.

56 S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
Gröcke, D.R., Hesselbo, S.P., Jenkyns, H.C., 1999. Carbon-isotope composition of LowerCretaceous fossil wood: ocean–atmosphere chemistry and relation to sea-levelchange. Geology 27, 155–158.
Gröcke, D.R., Price, G.D., Ruffell, A.H., Mutterlose, J., Baraboshkin, E., 2003. Isotopicevidence for Late Jurassic–Early Cretaceous climate change. Palaeogeography,Palaeoclimatology, Palaeoecology 202, 97–118.
Gröcke, D.R., Price, G.D., Robinson, S.A., Baraboshkin, E., Mutterlose, J., Ruffell, A.H.,2005. The Upper Valanginian (Early Cretaceous) positive carbon-isotope eventrecorded in terrestrial plants. Earth and Planetary Science Letters 240, 495–509.
Hagen, W., Van Vleet, E.S., Kattner, G., 1996. Seasonal lipid storage as overwinteringstrategy of Antarctica krill. Marine Ecology Progress Series 134, 85–89.
Hagen, W., Auel, H., 2001. Seasonal adaptation and the role of lipids in oceaniczooplankton. Zoology Analysis of Complex Systems 104, 313–326.
Hansen, K.W., Wallmann, K., 2003. Cretaceous and Cenozoic evolution of seawatercomposition, atmospheric O2 and CO2: a model perspective. American Journal ofScience 303, 94–148.
Harradine, P.J., Harris, P.G., Head, R.N., Harris, R.P., Maxwell, J.R., 1996. Steryl chlorinesters are formed by zooplankton herbivory. Geochimica et Cosmochimica Acta 60,2265–2270.
Harvey, H.R., Eglinton, G., O'Hara, S.C.M., Corner, E.D.S., 1987. Biotransformation andassimilation of dietary lipids by Calanus feeding on a dinoflagellate. Geochimica etCosmochimica Acta 51, 3030–3041.
Harvey, H.R., O'Hara, S.C.M., Eglinton, G., Corner, E.D.S., 1989. The comparative fate ofdinosterol and cholesterol in copepod feeding—implications for a conservativemolecularbiomarker in themarinewater column.OrganicGeochemistry 14, 635–641.
Harwood, D.M., Nikolaev, V.A., Winter, D.M., 2007. Cretaceous records of diatomevolution, radiation, and expansion. Paleontological Society Paper 13, 33–59.
Heimhofer, U., Hochuli, P.A., Herrle, J.O., Andersen, N., Weissert, H., 2004. Absence ofmajor vegetation and paleoatmospheric pCO2 changes associated with oceanicanoxic event 1a (Early Aptian, SE France). Earth and Planetary Science Letters 223,303–318.
Hennig, S., Weissert, H., Bulot, L., 1999. C-isotope stratigraphy, a calibration toolbetween ammonite- and magnetostratigraphy: the Valanginian–Hauteriviantransition. Geologica Carpathica 50, 91–99.
Herbin, J.P., Deroo, G., Roucaché, J., 1983. Organic geochemistry in the Mesozoic andCenozoic formations of Site 534, Leg 76, Blake–Bahama Basin, and comparison withSite 391, Leg 44. Initial Reports of the Deep Sea Drilling Project, 76. U.S. GovernmentPrinting Office, Washington, D.C., pp. 481–493.
Herbin, J.P., Deroo, G., Roucaché, J., 1984. Organic geochemistry of Lower Cretaceoussediments from Site 535, Leg 77, Florida Straits. Initial Reports of the Deep Sea DrillingProject, 77. U.S. Government Printing Office, Washington, D.C., pp. 459–475.
Herbin, J.P., Masure, E., Roucaché, J., 1987. Cretaceous formations from the lowercontinental rise off Cape Hatteras: organic geochemistry, dinoflagellate cysts, andthe Cenomanian/Turonian boundary event at Sites 603 (Leg 93) and 105 (Leg 11).Initial Reports of the Deep Sea Drilling Project, 93 (Pt. 2). U.S. Government PrintingOffice, Washington, D.C., pp. 1139–1162.
Herrle, J.O., Kößler, P., Friedrich, O., Erlenkeuser, H., Hemleben, C., 2004. High-resolution carbon isotope records of the Aptian to Lower Albian from SE France andthe Mazagan Plateau (DSDP Site 545): a stratigraphic tool for paleoceanographicand paleobiologic reconstruction. Earth and Planetary Science Letters 218,149–161.
Hochuli, P.A., Menegatti, A.P., Weissert, H., Riva, A., Erba, E., Premoli Silva, I., 1999.Episodes of high productivity and cooling in the early Aptian Alpine Tethys.Geology 27, 657–660.
Jahren, A.H., Arens, N.C., Sarmiento, G., Guerrero, J., Amundson, R., 2001. Terrestrialrecord of methane hydrate dissociation in the Early Cretaceous. Geology 29,159–162.
Jenkyns, H.C., 1980. Cretaceous oceanic events: from continents to oceans. Journal ofthe Geological Society (London) 137, 171–188.
Jenkyns, H.C., 2003. Evidence for rapid climate change in the Mesozoic–Palaeogenegreenhouse world. Philosophical Transactions of the Royal Society of London A 361,1885–1916.
Ju, S.J., Harvey, H.R., 2004. Lipids as markers of nutritional condition and diet in theAntarctic krill Euphausia superba and Euphausia crystallorophias during australwinter. Deep-Sea Research Part II 51, 2199–2214.
Jud, R., 1994. Biochronology and systematics of Early Cretaceous radiolarian of theWestern Tethys. Mémoires de Geologie (Lausanne) 19, 1–147.
Katz, B.J., 1984. Source quality and richness of Deep Sea Drilling Project Site 535sediments, southeastern Gulf of Mexico. Initial Reports of the Deep Sea DrillingProject, 77. U.S. Government Printing Office, Washington, D.C., pp. 445–450.
Kerr, R.G., Kerr, S.L., Malik, S., Djerassi, C., 1992. Biosynthetic studies of marine lipids 38.Mechanism and scope of sterol side-chain dealkylation in sponges: evidence forconcurrent alkylation and dealkylation. Journal of the American Chemical Society114, 299–303.
King, L.L., Wakeham, S.G., 1996. Phorbin steryl ester formation by macrozooplankton inthe Sargasso Sea. Organic Geochemistry 24, 581–585.
Kuypers, M.M.M., Blokker, P., Erbacher, J., Kinkel, H., Pancost, R.D., Schouten, S.,Sinninghe Damsté, J.S., 2001. Massive expansion of marine Archaea during a mid-Cretaceous oceanic anoxic event. Science 293, 92–94.
Kuypers, M.M.M., Blokker, P., Hopmans, E.C., Kinkel, H., Pancost, R.D., Schouten, S.,Sinninghe Damsté, J.S., 2002. Archaeal remains dominate marine organic matterfrom the early Albian oceanic anoxic event 1b. Palaeogeography, Palaeoclimatol-ogy, Palaeoecology 185, 211–234.
Kuypers, M.M.M., van Breugal, Y., Schouten, S., Erba, E., Sinninghe Damsté, J.S., 2004. N2-fixing cyanobacteria supplied nutrient N for Cretaceous oceanic anoxic events.Geology 32, 853–856.
Larson, R.L., 1991a. Latest pulse of Earth: evidence for a mid-Cretaceous superplume.Geology 19, 547–550.
Larson, R.L., 1991b. Geological consequences of superplumes. Geology 19, 963–966.Larson, R.L., Erba, E., 1999. Onset of the mid-Cretaceous greenhouse in the Barremian–
Aptian: igneous events and the biological, sedimentary, and geochemicalresponses. Paleoceanography 14, 663–678.
Leckie, R.M., Bralower, T.J., Cashman, R., 2002. Oceanic anoxic events and planktonevolution: biotic response to tectonic forcing during the mid-Cretaceous.Paleoceanography 17, 1–29.
Lini, A., Weissert, H., Erba, E., 1992. The Valanginian carbon isotope event: a firstepisode of greenhouse climate conditions during the Cretaceous. Terra Nova 4,374–384.
Luciani, V., Cobianchi, M., Jenkyns, H.C., 2001. Biotic and geochemical response toanoxic events: the Aptian pelagic succession of the Gargano Promontory (southernItaly). Geological Magazine 138, 277–298.
Marlow, J.R., Farrimond, P., Rosell-Melé, A., 2000. Analysis of lipid biomarkers insediments from the Benguela Current coastal upwelling system (Site 1084). In:Wefer, G., Berger, W.H., Richter, C. (Eds.), Proceedings of the Ocean DrillingProgram, Scientific Results, vol. 175, pp. 1–26.
Marlowe, I.T. 1984. Lipids as palaeoclimatic indicators. Ph.D. Thesis, University ofBristol, 273 pp.
Marlowe, I.T., Brassell, S.C., Eglinton, G., Green, J.C., 1984. Long chain unsaturatedketones and esters in living algae and marine sediments. Organic Geochemistry 6,135–141.
Marlowe, I.T., Brassell, S.C., Eglinton, G., Green, J.C., 1990. Long-chain alkenones andalkyl alkenoates and the fossil coccolith record of marine sediments. ChemicalGeology 88, 349–375.
McArthur, J.M., Janssen, N.M.M., Reboulet, S., Leng, M.J., Thirlwall, M.F., van deSchootbrugge, B., 2007. Palaeotemperatures, polar ice-volume, and isotope stratigra-phy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): the Early Cretaceous (Berriasian, Valanginian,Hauterivian). Palaeogeography, Palaeoclimatology, Palaeoecology 248, 391–430.
Melinte, M., Mutterlose, J., 2001. A Valanginian (Early Cretaceous) ‘boreal nannoplank-ton excursion’ in sections from Romania. Marine Micropaleontology 43, 1–25.
Menegatti, A.P., Weissert, H., Brown, R.S., Tyson, R.V., Farrimond, P., Strasser, A., Caron,M., 1998. High-resolution δ13C stratigraphy through the early Aptian “Livello Selli”of the Alpine Tethys. Paleoceanography 13, 530–545.
Meyers, P.A., 1987. Organic carbon content of sediments and rocks from Deep SeaDrilling Project Sites 603, 604, and 605, western margin of the North Atlantic. InitialReports of the Deep Sea Drilling Project, 93 (Pt. 2). U.S. Government Printing Office,Washington, D.C, pp. 1187–1194.
Miller, C.B., Morgan, C.A., Prahl, F.G., Sparrow, M.A., 1998. Storage lipids of the copepodCalanus jinmarchicus from Georges Bank and the Gulf of Maine. Limnology andOceanography 43, 488–497.
Mutterlose, J., Kessels, K., 2000. Early Cretaceous calcareous nannofossils from highlatitudes: implications for palaeobiogeography and palaeoclimate. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology. 160, 347–372.
Mutterlose, J., Brumsack, H., Flögel, S., Hay, W., Klein, C., Langrock, U., Lipinski, M., Ricken,W., Söding, E., Stein, R., Swientek, O., 2003. The Greenland–Norwegian Seaway: a keyarea for understanding Late Jurassic to Early Cretaceous paleoenvironments.Paleoceanography 18, 2001PA000625.
Ourisson, G., Albrecht, P., Rohmer, M., 1982. Predictive microbial biochemistry—frommolecular fossils to prokaryotic membranes. Trends in Biochemical Sciences 7,236–239.
Patton, J.W., Choquette, P.W., Guennel, G.K., Kaltenback, A.J., Moore, A., 1984. Organicgeochemistry and sedimentology of lower to mid-Cretaceous deep-sea carbonates,site 535 and 540, Leg 77. Initial Reports of the Deep Sea Drilling Project, 77. U.S.Government Printing Office, Washington, D.C., pp. 417–443.
Pearce, G.E.S., Harradine, P.J., Talbot, H.M., Maxwell, J.R., 1998. Sedimentary sterols andsteryl chlorin esters: distribution differences and significance. Organic Geochem-istry 28, 3–10.
Podlaha, O.G., Mutterlose, J., Veizer, J., 1998. Preservation of δ18O and δ13C in belemniterostra from the Jurassic/Early Cretaceous succession. American Journal of Science298, 324–347.
Popp, B.N., Takigiku, R., Hayes, J.M., Louda, J.W., Baker, E.W., 1989. The post-Paleozoicchronology and mechanism of 13C depletion in primary marine organic matter.American Journal of Science 289, 436–454.
Price, G.D., 1999. The evidence and implications of polar-ice during the Mesozoic. EarthScience Reviews 48, 183–210.
Price, G.D., 2003. New constraints upon isotope variation during the early Cretaceous(Barremian–Cenomanian) from thePacific Ocean.GeologicalMagazine 140, 513–522.
Price, G.D., Mutterlose, J., 2004. Isotopic signals from late Jurassic–early Cretaceous(Volgian–Valanginian) subArctic Belemnites, Yatria River, Western Siberia. Journalof the Geological Society, London 161, 959–968.
Price, G.D., Ruffell, A.H., Jones, C.J., Kalin, R.M., Muttterlose, J., 2000. Isotopic evidence fortemperature variation during the early Cretaceous (late Ryazanian–mid-Hauter-ivian). Journal of the Geological Society, London 157, 335–343.
Pucéat, E., Lécuyer, C., Sheppard, S.M.F., Dromart, G., Reboulet, S., Grandjean, P., 2003.Thermal evolution of Cretaceous Tethyan marine waters inferred from oxygenisotope composition of fish tooth enamels. Paleoceanography 18, 2002PA000823.
Rampen, S.W., Schouten, S., Abbas, B., Panoto, F.E., Muyzer, G., Campbell, C.N., Fehling, J.,Sinninghe Damsté, J.S., 2007. On the origin of 24-norcholestanes and their use asage-diagnostic biomarkers. Geology 35, 419–422.
Rampen, S.W., Schouten, S., Hopmans, E.C., Abbas, B., Noordeloos, A.A.M., van Bleijswijk,J.D.L., Geenevasen, J.A.J., Sinninghe Damsté, J.S., 2009. Diatoms as a source for 4-desmethyl-23, 24-dimethyl steroids in sediments and petroleum. Geochimica etCosmochimica Acta 73, 377–387.

57S.C. Brassell / Palaeogeography, Palaeoclimatology, Palaeoecology 282 (2009) 45–57
Reboulet, S., Mattioli, E., Pittet, B., Baudin, F., Olivero, D., Proux, O., 2003. Ammonoid andnannoplankton abundance in Valanginian (early Cretaceous) limestone–marlsuccessions from the southeast France Basin: carbonate dilution or productivity?Palaeogeography, Palaeoclimatology, Palaeoecology 201, 113–139.
Rullkötter, J., Mukhopadhyay, P.K., 1986. Comparison of Mesozoic carbonaceousclaystones in the western and eastern North Atlantic (DSDP Legs 76, 79 and 93).In: Summerhayes, C.P., Shackleton, N.J. (Eds.), North Atlantic Palaeoceanography:Geological Society, London, Special Publications, vol. 21, pp. 377–387.
Rullkötter, J., Leythaeuser, D., Wendisch, D., 1982. Novel 23, 28-bisnorlupanes inTertiary sediments—widespread occurrence of nuclear demethylated triterpanes.Geochimica et Cosmochimica Acta 46, 2501–2509.
Rullkötter, J., Mukhopadhyay, P.K., Hartung, B., Welte, D.H., 1984. Geochemistry andpetrography of organic matter in Cretaceous sediments from the southeastern Gulfof Mexico, Deep Sea Drilling Project Hole 535—preliminary results. Initial Reports ofthe Deep Sea Drilling Project 77. U.S. Government Printing Office, Washington, D.C.,pp. 489–493.
Rullkötter, J., Mukhopadhyay, P.K., Disko, U., Schaefer, R.G., Welte, D.H., 1986. Facies anddiagenesis of organic matter in deep sea sediments from the Blake Outer Ridge and theBlake Bahama Basin,Western Atlantic. In: Degens, E.T., Meyers, P.A., Brassell, S.C. (Eds.),Biogeochemistry of Black Shales: Mitteilungen der Geologisch-PaläontologischesInstitut der Universitat Hamburg, pp. 179–203.
Rullkötter, J., Mukhopadhyay, P.K., Welte, D.H., 1987. Geochemistry and petrography oforganic matter from Deep Sea Drilling Project Site 603, lower continental rise offCape Hatteras. Initial Reports of the Deep Sea Drilling Project 93 (Pt. 2). U.S.Government Printing Office, Washington, D.C., pp. 1163–1176.
Schlanger, S.O., Jenkyns, H.C., 1976. Cretaceous oceanic anoxic events: causes andconsequences. Geologie en Mijnbouw 55, 179–184.
Schefuß, E., Versteegh, G.J.M., Jansen, J.H.F., Sinninghe Damsté, J.S., 2001. Marine andterrigenous lipids in southeast Atlantic sediments (Leg 175) as paleoenvironmentalindicators: initial results. In: Wefer, G., Berger, W.H., Richter, C. (Eds.), Proceedingsof the Ocean Drilling Program, Scientific Results, vol. 175, pp. 1–34.
Schouten, S., Hoefs, M.J.L., Sinninghe Damsté, J.S., 2000. A molecular and stable carbonisotopic study of lipids in late Quaternary sediments from the Arabian Sea. OrganicGeochemistry 31, 509–521.
Schouten, S., De Loureiro, M.R.B., Sinninghe Damsté, J.S., de Leeuw, J.W., 2001.Molecular biogeochemistry of Monterey sediments (Naples beach, USA) I:distributions of hydrocarbons and organic sulphur compounds. In: Isaacs, C.M.,Rullkötter, J. (Eds.), The Monterey Formation: from Rocks to Molecules, Columbia,pp. 150–174.
Schouten, S., Rampen, S.W., Geenevasen, J.A.J., Sinninghe Damsté, J.S., 2005. Structuralidentification of steryl alkyl ethers in marine sediments. Organic Geochemistry 36,1323–1333.
Sheia, J., Brassell, S.C., Ward, D.M., 1991. Comparative analysis of extractable lipids inhot spring microbial mats and their component photosynthetic bacteria. OrganicGeochemistry 17, 309–319.
Sinninghe Damsté, J.S., de Leeuw, J.W., 1990. Analysis, structure and geochemicalsignificance of organically-bound sulfur in the geosphere—state of the art andfuture research. Organic Geochemistry 16, 1077–1101.
Sinninghe Damsté, J.S., Kock vanDalen, A.C., de Leeuw, J.W., Schenck, P.A., Sheng, Guoying,Brassell, S.C., 1987. The identificationofmono-, di-, and trimethyl 2-methyl-2-(4,8,12-trimethyltridecyl) chromans and their occurrence in the geosphere. Geochimica etCosmochimica Acta 51, 2393–2400.
Sinninghe Damsté, J.S., Muyzer, G., Abbas, B., Rampen, S.W., Massé, G., Allard, G., Belt, S.T.,Robert, J.-M., Rowland, S.J., Moldowan, J.M., Barbanti, S.M., Fago, F.J., Denisevich, P.,
Dahl, J., Trindade, L.A.F., Schouten, S., 2004. The rise of the rhizosolenid diatoms.Science 304, 584–587.
Sliter, W.V., 1989. Aptian anoxia in the Pacific Basin. Geology 17, 909–912.Sorhannus, U., 2007. A nuclear-encoded small-subunit ribosomal RNA timescale for
diatom evolution. Marine Micropaleontology 65, 1–12.Steuber, T., Rauch, M., Masse, J.P., Graff, J., Malkoc, M., 2005. Low-latitude seasonality of
Cretaceous temperatures in warm and cold episodes. Nature 437, 1341–1344.Summerhayes, C.P., Masran, T.C., 1983. Organic facies of Cretaceous and Jurassic
sediments from Deep Sea Drilling Project Site 534 in the Blake–Bahama Basin,western North Atlantic. Initial Reports of the Deep Sea Drilling Project, 76. U.S.Government Printing Office, Washington, D.C., pp. 469–480.
Tajika, E., 1999. Carbon cycle and climate change during the Cretaceous inferred from abiogeochemical carbon cycle model. Island Arc 8, 293–303.
Talbot, H.M., Head, R.N., Harris, R.P., Maxwell, J.R., 1999a. Steryl esters of pyrophaeo-phorbide b: a sedimentary sink for chlorophyll b. Organic Geochemistry 30,1403–1410.
Talbot, H.M., Head, R.N., Harris, R.P., Maxwell, J.R., 1999b. Distribution and stability ofsteryl chlorin esters in copepod faecal pellets from diatom grazing. OrganicGeochemistry 30, 1163–1174.
Talbot, H.M., Head, R.N., Harris, R.P., Maxwell, J.R., 2000. Discrimination against 4-methyl sterol uptake during steryl chlorin ester production by copepods. OrganicGeochemistry 31, 871–880.
van de Schootbrugge, B., Föllmi, K., Bulot, L.G., Burns, S.J., 2000. Paleoceanographicchanges during the early Cretaceous (Valanginian–Hauterivian): evidence fromoxygen and carbon stable isotopes. Earth and Planetary Science Letters 181, 15–31.
Vill, V., Weber, N., 1991. Steroidal ether lipids with liquid crystalline properties.Chemistry and Physics of Lipids 58, 105–110.
Vill, V., Weber, N., 1994. Liquid crystalline cholestanyl and cholesteryl ether lipids.Molecular Crystals and Liquid Crystals Science and Technology Section A—Molecular Crystals and Liquid Crystals 250, 73–83.
Von der Dick, H., Rullkötter, J., Welte, D.H., 1983. Content, type, and thermal evolutionof organic matter in sediments from the eastern Falkland Plateau, Deep Sea DrillingProject, Leg 71. In: ILudwig, W.J., Krasheninnikov, V.A., et al. (Eds.), Initial Reports ofthe Deep Sea Drilling Project, 71. U.S. Government Printing Office, Washington D.C.,pp. 1015–1032.
Wallmann, K., 2001. Controls on the Cretaceous and Cenozoic evolution of seawatercomposition, atmospheric CO2 and climate. Geochimica et Cosmochimica Acta 65,3005–3025.
Wälterman, M., Steinbüchel, A., 2005. Neutral lipid bodies in prokaryotes: recentinsights into structure, formation, and relationship to eukaryotic lipid depots.Journal of Bacteriology 187, 3607–3619.
Wefer, G., Berger, W.H., Richter, C., et al. (Eds.), 1998. Proceedings of the Ocean DrillingProgram, Initial Reports, vol. 175.
Weissert, H., Erba, E., 2004. Volcanism, CO2 and palaeoclimate: a Late Jurassic–EarlyCretaceous carbon and oxygen isotope record. Journal of the Geological Society,London 161, 695–702.
Weissert, H., Lini, A., Föllmi, K.B., Kuhn, O., 1998. Correlation of Early Cretaceous carbonisotope stratigraphy and platform drowning events: a possible link? Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 137, 189–203.
Wortmann, U.G., Weissert, H., 2000. Tying platform drowning to perturbations of theglobal carbon cycle with a δ13C-curve from the Valanginian of DSDP Site 416. TerraNova 12, 289–294.




















