A vaca leiteira do século 21: lições de metabolismo e nutrição
348
A vaca leiteira do século 21: lições de metabolismo e nutrição Faculdade de Veterinária Universidade Federal do Rio Grande do Sul Editor Félix H. D. González Coeditores: Ana C. R. Berreta Anne R. Guadagnin Pedro M. Mallmann Raquel Fraga e S. Raimondo Beatriz Riet-Correa Rivero Rodrigo Schallenberger Gonçalves André Dalto Porto Alegre - RS, Brasil 2021
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of A vaca leiteira do século 21: lições de metabolismo e nutrição

A vaca leiteira do século 21: lições de metabolismo e nutrição
Faculdade de Veterinária
Universidade Federal do Rio Grande do Sul
Editor Félix H. D. González
Coeditores: Ana C. R. Berreta
Anne R. Guadagnin Pedro M. Mallmann
Raquel Fraga e S. Raimondo Beatriz Riet-Correa Rivero
Rodrigo Schallenberger Gonçalves André Dalto
Porto Alegre - RS, Brasil 2021

2
CIP – CATALOGAÇÃO INTERNACIONAL DA PUBLICAÇÃO
A111 A vaca leiteira do século 21: lições de metabolismo e nutrição [livro
eletrônico] / Félix H. D. González, editor. – Porto Alegre : Universidade Federal do Rio Grande do Sul, Faculdade de Veterinária, 2021.
3800 Kb ; PDF , 348 p. : il.
Bibliografia no final dos capítulos Vários autores ISBN 978-65-5973-073-5
1. Medicina Veterinária: vacas leiteiras 2. Bovinos: nutrição. 3. Bovinos: metabolismo 4. Qualidade do leite I. González, Félix H. D. CDD 636.2
Catalogação na fonte: Ana Vera Finardi Rodrigues – CRB-10/884

3
Autores contribuintes
Álan Maia Borges. Departamento de Clínica e Cirurgia Veterinária – Universidade Federal
de Minas Gerais: [email protected]
André Thaler Neto. Centro de Ciências Agroveterinárias – Universidade do Estado de
Santa Catarina: [email protected]
Angélica Petersen Dias. Faculdade de Veterinária – Universidade Federal do Rio Grande
do Sul: [email protected]
Anne Rosi Guadagnin. Faculdade de Veterinária – Universidade Federal do Rio Grande do
Sul: [email protected]
Bolívar Nóbrega de Faria. Recursos Humanos no Agronegócio Ltda – Belo Horizonte:
Carla Bittar. Departamento de Zootecnia, ESALQ – Universidade de São Paulo:
Carlos Bondan. Serviço de Análises de Rebanhos Leiteiros – Universidade de Passo Fundo:
Enrico Lippi Ortolani. Faculdade de Medicina Veterinária e Zootecnia – Universidade de
São Paulo: [email protected]
Felipe Cardoso. Department of Animal Science – University of Illinois:
Fernando Wittwer. Facultad de Ciencias Veterinarias – Universidad Austral de Chile,
Valdivia, Chile: [email protected]
Giovani Noro. Tortuga Companhia Zootécnica Agrária: [email protected]
Jessica Karina Poncheki. Programa de Pós-Graduação em Zootecnia – Universidade
Federal do Paraná: [email protected]
Jorge Henrique Carneiro. Cooperativa Castrolanda Agroindustrial:
Júlio Viégas. Departamento de Zootecnia – Universidade Federal de Santa Maria:
Leopoldo Braz Los. Frísia Cooperativa Agroindustrial: [email protected]
Luis Barros (In memoriam). Facultad de Veterinaria – Universidad de la República,
Uruguay: [email protected]
Luis Albornoz. Facultad de Veterinaria – Universidad de la República, Uruguay:
Marcelo Cecim. Departamento de Clínica de Grandes Animais – Universidade Federal de
Santa Maria: [email protected]

4
Márcio Nunes Correa. Faculdade de Veterinária – Universidade Federal de Pelotas:
María Cecilia Cajarville. Facultad de Veterinaria – Universidad de la República, Uruguay:
Mirela Noro. Curso de Medicina Veterinária – Universidade Federal do Pampa:
Paula Montagner. Programa de Pós-graduação em Biotecnologia – Universidade Federal de
Pelotas:[email protected]
Rodrigo de Almeida. Departamento de Zootecnia – Universidade Federal do Paraná:
Vivian Fischer. Departamento de Zootecnia – Universidade Federal do Rio Grande do Sul:
Andressa Stein Maffi, Beatriz Riet-Correa, Ismael Mateus Cavazini, Marcelo Moreira
Antunes, Viviane R. Rabassa, Cássio C. Brauner, Camila Pizoni, Joabel Tonellotto dos
Santos, Patrícia Mattei, Rafael da Fonseca Prietsch, Uriel Secco Londero: Núcleo de
Pesquisa, Ensino e Extensão em Pecuária (NUPEEC) – Universidade Federal de Pelotas
Marcelo Tempel Stumpf, Alexandre Sausenbach Abreu, Elissa Forgiarini Vizzotto:
Departamento de Zootecnia – Universidade Federal do Rio Grande do Sul
Adriana Hauser, Marciel França, Veridiane Orsolin: Centro de Ciências Agroveterinárias –
Universidade do Estado de Santa Catarina
Telma da Mata Martins, Ana Carolina Leite, Philipe Pimenta Nunes: Programa de Pós-
Graduação em Ciência Animal – Universidade Federal de Minas Gerais
J. P. Albornoz, L.E Fidalgo: Universidade de Santiago de Compostela, Lugo, España.
M. Morales: Centro Diagnóstico Veterinario de Florida, Uruguay

5
Sumário
Página
Prefácio 6
Manejo nutricional pré e pós-parto para otimizar a
fertilidade em vacas de leite Felipe Cardoso 7
Como a dieta pode afetar a composição do leite Bolívar Nóbrega de Faria 17
Transtornos metabólicos da vaca leiteira no período
de transição Enrico Lippi Ortolani 35
Transtornos no período de transição da vaca leiteira
com ênfase nas enfermidades uterinas e da glândula
mamária
Márcio Nunes Correa et al. 47
Estresse calórico em vacas leiteiras: efeito sobre o
metabolismo e a qualidade do leite
Vivian Fischer et al.
61
Bem-estar de vacas leiteiras: melhor produção com a
vaca feliz Marcelo Cecim 74
Marcadores bioquímicos sanguíneos en el
diagnóstico y control de trastornos metabólicos en
vacas lecheras
Fernando Wittwer 79
Variações na qualidade composicional do leite no
Rio Grande do Sul Carlos Bondan 98
Acidose ruminal subaguda (SARA): monitoramento
e prevenção nos rebanhos leiteiros
Mirela Noro e Giovani Noro
118
Manejo nutricional da vaca leiteira para otimizar a
composição do leite Jessica Karina Poncheki et al. 136
Fundamentos y aplicaciones para la alimentación
proteica de vacas lecheras Cecília Cajarville 160
Cetose clínica e subclínica: manejo, diagnóstico e
efeitos no leite Marcio Nunes Correa et al. 169
Hipocalcemia e hipomagnesemia en vacas lecheras:
diagnóstico y control Luis Barros Vidal 181
Deslocamento de abomaso em vacas leiteiras:
ocorrência, manejo e indicadores diagnósticos Angélica P. Dias et al. 203
Alimentação e manejo de bezerras leiteiras Carla Maris Machado Bittar 215
Alimentação e manejo da novilha leiteira Júlio Viégas 235
Monitoramento de bem-estar e saúde em rebanhos
leiteiros Marcelo Cecim 253
Pontos críticos da qualidade do leite André Thaler Neto et al. 274
Relação entre metabolismo e reprodução em vacas
leiteiras Álan Maia Borges et al. 298
Prevenção das acidoses ruminais em rebanhos
leiteiros: novos conceitos Enrico Lippi Ortolani 311
Hipocalcemia puerperal bovina Luis Albornoz 318
Impacto da nutrição na composição e qualidade do
leite Leopoldo Braz Los 339

6
Prefácio
De 2014 a 2019 foram realizados anualmente seis Simpósios da Vaca Leiteira na Faculdade
de Veterinária da Universidade Federal do Rio Grande do Sul. Dos anais daqueles simpósios,
que podem ser consultados na página https://www.ufrgs.br/lacvet/extensao/, foram
selecionadas 22 palestras abrangendo exclusivamente temas de nutrição, metabolismo,
qualidade do leite e bem-estar animal. As palestras foram lecionadas por reconhecidos
professores e pesquisadores de várias universidades e de centros de pesquisa, do Brasil e do
exterior, engajados no estudo da clínica e da produção da vaca leiteira. Nosso mais sincero
agradecimento aos palestrantes, bem como aos nossos coeditores que colaboraram na
realização dos seis simpósios.
O editor
Félix Gonzalez
Porto Alegre, primavera de 2021

7
Manejo nutricional pré e pós-parto para otimizar a fertilidade em vacas
leiteiras
Felipe Cardoso
O sucesso reprodutivo é crítico para o sucesso econômico e para a sustentabilidade de
fazendas de leite em qualquer sistema de produção. O sucesso reprodutivo depende de uma
série de eventos fisiológicos coordenados, incluindo retorno da ciclicidade ovariana após o
parto, desenvolvimento e ovulação de um oócito viável, fertilização, involução uterina,
desenvolvimento e implantação embrionária e permanência da prenhez até a maturação fetal
(Butler, 2003; Garnsworthy et al., 2008). A formulação de dietas e o manejo da alimentação
durante o período seco, período do periparto e período imediato após o parto podem facilitar
ou interromper diversos dos passos citados acima antes da prenhez ser estabelecida e mantida
(Butler, 2003; Garnsworthy et al., 2008; Thatcher et al., 2011). As taxas de concepção para
vacas de alta produção que estão ciclando é alta (> 70%) mas muitas vacas (> 50%)
apresentam morte embrionária precoce e falham em manter a prenhez (Diskin and Morris,
2008). Os maiores pontos de ênfase relacionando nutrição e reprodução em vacas de leite
são a severidade e a duração do estado de nutrientes negativo, o chamado balanço energético
negativo (BEN), que no começo da lactação é fortemente influenciado pelo manejo
nutricional e do ambiente durante o período de transição desde o final da prenhez até o início
da lactação.
O período de 6 a 8 semanas em torno do parto é conhecido como o período de transição ou
peripartum, crítico para determinar o bem-estar e a lucratividade de vacas individualmente
durante a lactação seguinte (Drackley et al., 2005). Programas de manejo do período de
transição que não são adequados resultam em alta prevalência de doenças da producão (i.e.,
deslocamento de abomaso) em rebanhos leiteiros (Mulligan & Doherty, 2008). Distúrbios no
metabolismo durante o período de transição podem têr influência direta ou indireta na
fertilidade e transições difíceis têm impacto negativo posteriormente na reprodução
(Chapinal et al., 2012). Uma grande variedade de estratégias nutricionais para facilitar as
adaptações metabólicas e fisiológicas da gestação até o início da lactação têm sido propostas
(Friggens et al., 2004; Roche et al., 2013).

8
Os objetivos deste artigo são de revisar as causas do BEN e o impacto em distúrbios
metabólicos, de relacionar como estes distúrbios influenciam a reprodução e de avaliar como
diferentes estratégias nutricionais antes e depois do parto podem afetar o BEN e a adequação
dos nutrientes. O tema central do artigo é que o manejo para promover maior ingestão de
matéria seca (IMS) de dietas corretamente balanceadas após o parto é o fator unificador de
sucesso durante o período de transição para otimizar a reprodução.
Fertilidade, produção de leite e doenças do periparto
Uma informação que é aceita por todos é a de que a fertilidade de vacas de leite modernas
está piorando, especialmente para a genética Holandesa, pelo menos em parte pela
consequência indesejada de seleção continuada para alta produção de leite. Esta informação
já foi desafiada por LeBlanc (2010) e Bello et al. (2012). Existe uma grande distribuição do
sucesso reprodutivo tanto dentro do rebanho como entre rebanhos. Por exemplo, entre cinco
rebanhos na California totalizando 6.396 vacas, encontrou-se que vacas que estavam no mais
baixo quartil para produção de leite nos primeiros 90 dias após o parto (32,1 kg/dia) tiveram
menor probabilidade de retorno ao ciclo estral aos 65 dias pós-parto quando comparado a
vacas no 2º quartil (39,1 kg/dia), no 3º quartil (43,6 kg/dia) ou no 4º quartil (50,0 kg/dia), de
forma que a produção de leite não afetou a taxa de prenhez (Santos et al., 2009). Mudanças
de manejo e sistemas de manejo inadequados potencialmente são mais limitantes para a
fertilidade de vacas de leite modernas do que a sua genética per se.
Vacas de leite são suscetíveis a doenças da produção durante o periparto e início da lactação
(Mulligan et al., 2006; Ingvartsen & Moyes, 2013; Roche et al., 2013). Existe pouca
evidência de que a produção de leite per se contribua para maior ocorrência de doenças.
Entretanto, o pico de ocorrência de doenças logo após o parto corresponde com o momento
de maior BEN, com o pico de concentração sanguínea de ácidos graxos não esterificados
(AGNE) e com a maior aceleração na produção de leite (Ingvartsen et al., 2003). O pico na
produção de leite ocorre muitas semanas após. Doenças associadas com maior BEN no pós-
parto estão também relacionadas com subótima performance reprodutiva, incluindo fígado
gorduroso (Rukkwamsuk et al., 1999; Jorritsma et al., 2003) e cetose (Walsh et al., 2007;
McArt et al., 2012). Vacas que perderam mais de 1 unidade (escala de 1-5) no escore de
condição corporal (ECC) apresentaram maior incidência de metrite, retenção de placenta e

9
doenças metabólicas (deslocamento de abomaso, hipocalcemia e cetose) e também maior
intervalo do parto até a primeira cobertura do que vacas que perderam menos de 1 unidade
durante o período de transição.
Indicadores do BEN são altamente correlacionados com perda de produção de leite, aumento
da prevalência de doenças e redução na fertilidade (Ospina et al., 2010; Chapinal et al., 2012).
Entretanto, o grau no qual o BEN é o causador de doenças do periparto e não somente um
fenômeno associado, precisa ser examinado criticamente (Roche et al., 2013). Por exemplo,
a resposta inflamatória pode reduzir a IMS em vacas no período de transição, causando
alterações no metabolismo e predispor vacas a um maior BEN ou maior prevalência de
doenças (Bertoni et al., 2008; Graugnard et al., 2012, 2013; Ingvartsen & Moyes, 2013).
Aumento na concentração de AGNE no pós-parto imediato e redução na concentração de
glicose estão fortemente associados com a taxa de prenhez à primeira cobertura em um
programa de inseminação artificial em tempo fixo (IATF) (Garverick et al., 2013). Apesar
da concentração de AGNE não ter sido diferente entre vacas que ovularam ou não antes do
programa de IATF, a taxa de prenhez reduziu com a maior concentração de AGNE e
aumentou com a maior concentração de glicose 3 dias após o parto (Garverick et al., 2013).
McArt et al. (2012) concluíram que vacas com cetose subclínica diagnosticada entre 3 a 7
dias pós-parto tiveram 0,7 chances de serem classificadas como prenhes ao primeiro
serviço e 4,5 vezes mais chances de serem removidas do rebanho nos primeiros 30 dias
após o parto quando comparadas a vacas que desenvolveram cetose 8 dias após o parto ou
mais tardiamente. Vacas que conseguem se adaptar à subsequente lactação (Jorritsma et al.,
2003) e conseguem evitar problemas metabólicos (Ingvartsen et al., 2003) ou desbalanço
fisiológico (Ingvartsen and Moyes, 2013) são capazes de suportar alta produção de leite e
reproduzir com sucesso.
Balanço energético negativo (BEN) e fertilidade subótima
As vacas não têm a capacidade de consumir nutrientes energéticos através de IMS voluntária
após o parto para suprir os requerimentos para produção de leite. Consequentemente, o BEN
ocorre por um período de dias a semanas durante o começo da lactação. Apesar de estudos
não terem demostrado uma forte relação entre o grau de BEN e a fertilidade, o tempo para o
valor mínimo do BEN e a direção e/ou grau de mudança no BEN parecem ser fortes

10
indicadores (Whitaker et al., 1993; Butler, 2003; Reist et al., 2003). Os mecanismos
potencialmente envolvidos nos efeitos danosos do BEN na reprodução incluem: (1) lento
retorno da ciclicidade ovariana, (2) impacto na qualidade, viabilidade, ou função do ovócito
ou do corpo lúteo, e (3) desenvolvimento de lipidose hepática.
Em geral, vacas que ovulam mais cedo depois do parto tem maior fertilidade (Butler, 2003;
Bossaert et al., 2008; Galvão et al., 2010). A produção de estrógeno pelo folículo dominante,
a recuperação da secreção e pulsatilidade de LH, e a resposta do ovário ao LH vão determinar
o sucesso da ovulação. O estado do BEN no pós-parto é negativamente associado com a
performance reprodutiva em parte porque interrompe aqueles 3 fatores (Butler, 2003). A
insulina é o hormônio que normalmente reflete o estado energético e a adequação nutricional
à dieta, podendo ser a primeira ligação entre os sistemas metabólico e reprodutivo. Baixas
concentrações de insulina e IGF-I (insulin-like growth factor 1) foram associadas à baixa
qualidade do corpo lúteo e do ovócito (Jorritsma et al., 2003).
A lipidose hepática está negativamente associada com a fertilidade (Rukkwamsuk et al.,
1999), o que pode ser um efeito indireto do excessivo BEN em vacas. Entretanto, talvez
existam efeitos negativos diretos da infiltração hepática por lipídios e a reprodução. Um
grande número de funções vitais do fígado são negativamente afetadas por cetose e infiltração
hepática de lipídios no pós-parto imediato (Loor et al., 2007). Um exemplo é o aumento da
taxa de apoptose, identificada pela análise do fígado pela técnica de micro-arranjo (Loor et
al., 2007) e mais recentemente por métodos funcionais diretos (Tharwat et al., 2012).
Manejo nutricional para controlar o BEN e otimizar a fertilidade
O balanço energético negativo no pós-parto inicia com a síntese de leite, entretanto, a
severidade do BEN em vacas é fracamente associada com a produção de leite ou a secreção
energética de leite. O grau e duração do BEN está fortemente associado com IMS (Zurek et
al., 1995; Drackley et al., 2005). Consequentemente, alimentação e estratégias de manejo
para vacas no período seco, durante o período de transição ou no pós-parto imediato devem
buscar fornecer os nutrientes necessários, mas também promover o apetite e a vigorosa IMS
após o parto (Grummer et al., 2004).
Alguns fatores importantes a serem considerados para promover o apetite e a alta IMS após
o parto são: (1) reduzir fatores estressantes externos e manter as vacas confortáveis, (2) evitar

11
escore de condição corporal (ECC) excessivamente alto ou baixo, (3) prevenir o consumo
exagerado de energia durante o período seco, (4) assegurar a ingestão de forragens com alto
valor de fibra efetiva e evitar a ingestão excessiva de carboidratos rapidamente fermentáveis
na dieta após o parto, (5) reduzir a diferença cátion-ânion da dieta (DCAD) antes do parto
mas aumentar na dieta da vaca durante o pós-parto imediato, e (6) garantir suporte de
nutrientes para o sistema imunológico. Estratégias nutricionais para auxiliar vacas em fazer
estas adaptações foram revisados (Friggens et al., 2004; Beever, 2006; Ingvartsen, 2006;
Drackley & Dann, 2008; Lean et al., 2013a,b; Roche et al., 2013).
Bach et al. (2008) concluíram que o manejo não-nutricional representou mais de 50% da
variação média de produção de leite (20,6 a 33,8 kg/dia) entre 47 rebanhos que consumiam
exatamente a mesma dieta. Emmanuel et al. (2007, 2008) demonstraram que quantidades
crescentes de grão de cevada aumentaram as concentrações de endotoxina no fluído ruminal
e que, em situações de baixo pH no rúmen, aumentou a absorção ruminal de endotoxinas.
Fatores estressantes não-nutricionais podem reduzir a IMS e predispor vacas a problemas de
saúde no pós-parto. Vacas que desenvolveram metrite (Hammon et al., 2006; Huzzy et al.,
2007) ou cetose (Goldhawk et al., 2009) após o parto tiverem menor IMS ou comportamento
de consumo reduzido antes do parto.
Enquanto a utilização de dietas steam-up ou close-up antes do parto, recomendadas por
muitos anos, existe na literatura uma falta de fatores positivos associados ao seu uso com a
saúde, a produção, ou a reprodução de vacas. A estratégia mais simples e o princípio
nutricional mais facilmente defendido para a alimentação de vacas nos períodos seco e de
transição é o de alimentar vacas para atender, mas não exceder os requerimentos nutricionais
(Drackley & Dann, 2008). Diversos trabalhos do nosso grupo de pesquisa demonstraram que
dietas com energia controlada durante o período seco contribuem para um período de
transição de maior sucesso (Grum et al., 1996; Dann et al., 2005, 2006; Douglas et al., 2006;
Janovick et al., 2011; Graugnard et al., 2012, 2013; Ji et al., 2012). Nossos estudos indicam
que o consumo prolongado de energia durante o período seco pode reduzir a IMS após o
parto (Douglas et al., 2006; Dann et al., 2006; Janovick & Drackley, 2010, 2014). O consumo
excessivo de energia resulta em resposta negativa de indicadores metabólicos como altas
concentrações de AGNE e beta-hidroxibutirato (BHB) no sangue e mais triglicerídeos no
fígado após o parto (Douglas et al., 2006; Janovick et al., 2011). Alterações celulares e de

12
genes no fígado (Loor et al., 2006, 2007) e no tecido adiposo (Ji et al., 2012) potencialmente
podem explicar muitas das alterações que ocorrem na vaca. O consumo excessivo de energia
no pré-parto aumenta o “maquinário” enzimático no tecido adiposo para mobilização de
triglicerídeos após o parto, com alterações na transcrição de genes que levam à redução na
lipogênese, aumento da lipólise e redução na capacidade da insulina de inibir lipólise (Ji et
al., 2012). Controlar o consumo de energia no período seco também aumentou a função de
neutrófilos no pós-parto (Graugnard et al., 2012) ocasionando melhoria na função do sistema
imune.
Cardoso et al. (2013) analisaram as associações entre o regime energético de alimentação no
pré-parto e a performance reprodutiva de vacas de raça Holandesa. Dados individuais de 408
vacas foram utilizados. Tratamentos nutricionais foram divididos em vacas recebendo dieta
com energia controlada (CE; NEL = 13,7 Mcal/dia) ou vacas recebendo dieta com alta
energia (HE; NEL = 22,1 Mcal/dia) durante o período seco distante (FO, 60 até 28 dias antes
do parto) ou período seco próximo (CU, 28 dias antes do parto até o parto). O modelo COX
revelou uma diferença estatística em dias até a prenhez (DTP) entre HE e CE durante CU
(mediana = 167 e 157 dias, hazard ratio = 0,696; Figura 1). Vacas que foram alimentadas
com dietas HE durante CU perderam mais ECC (escala 1–5) durante as primeiras 6 semanas
pós-parto do que vacas alimentadas CE (−0,43 e −0,30, respectivamente). Vacas que foram
alimentadas com HE durante o período seco tiveram mais chances de apresentarem
deslocamento de abomaso ou cetose quando comparadas a vacas que receberam CE.
Conclusão
Formulação e alimentação de dietas apropriadas, que limitam o consumo total de energia
para suprir, e não exceder, os requerimentos de vacas durante o pré-parto podem auxiliar a
aliviar o BEN após o parto. Efeitos destas dietas em indicadores de metabolismo para saúde
são geralmente positivos, sugerindo o potencial para redução das doenças do periparto e
melhora da fertilidade. Estratégias para a formulação de dietas que estimulem a IMS e
diminuam o BEN no pós-parto imediato são menos pesquisadas. Entretanto, o balanço entre
fibra fisicamente efetiva e fermentação do amido nestas dietas é crítico.

13
Figura 1. Curvas de survival function para dias até a prenhez (DTP) para 332 vacas
Holstein alimentadas com dieta de energia controlada (CE = azul) ou alta energia (HE
= vermelho) durante as 4 últimas semanas antes do parto. Linhas azul e vermelha
representam a mediana dos valores para DTP onde 50% das vacas estavam prenhes.
Adaptado de Cardoso et al. (2013)
Referências
Bach A, Valls N, Solans A, Torrent T 2008. Association between nondietary factors and dairy herd
performance. Journal of Dairy Science 91, 3529-3267.
Beever DE 2006. The impact of controlled nutrition during the dry period on dairy cow health, fertility and
performance. Animal Reproduction Science 96, 212-226.
Bello NM, Stevenson JS,Tempelman RJ 2012. Invited review: Milk production and reproductive
performance: Modern interdisciplinary insights into an enduring axiom. Journal of Dairy Science 95, 5461-
5475.
Bertoni G, Trevisi E, Han X, Bionaz M 2008. Effects of inflammatory conditions on liver activity in
puerperium period and consequences for performance in dairy cows. Journal of Dairy Science 91, 3300-
3310.
Bossaert P, Leroy JLMR, De Vliegher S, Opsomer G 2008. Interrelations between glucose-induced insulin
response, metabolic indicators, and time of first ovulation in high-yielding dairy cows. Journal of Dairy
Science 91, 3363-3371.
Butler WR 2003. Energy balance relationships with follicular development, ovulation and fertility in
postpartum dairy cows. Livestock Production Science 83, 211-218.
Cardoso FC, LeBlanc SJ, Murphy MR, Drackley JK 2013. Prepartum nutritional strategy affects reproductive
performance in dairy cows. Journal of Dairy Science 96, 5859-5871.
Chapinal N, Leblanc SJ, Carson ME, Leslie KE, Godden S, Capel M, Santos JE, Overton MW, Duffield TF.
2012. Herd-level associations of serum metabolites in the transition period with disease, milk production, and
early lactation reproductive performance. Journal of Dairy Science 95, 5676-5682.

14
Dann HM, Litherland NB, Underwood JP, Bionaz M, D’Angelo A, McFadden JW, Drackley JK 2006. Diets
during far-off and close-up dry periods affect periparturient metabolism and lactation in multiparous cows.
Journal of Dairy Science 89, 3563-3577.
Dann HM, Morin DE, Murphy MR, Bollero GA, Drackley JK 2005. Prepartum intake, postpartum induction
of ketosis, and periparturient disorders affect the metabolic status of dairy cows. Journal of Dairy Science 88,
3249-3264.
Diskin MG, Morris DG 2008. Embryonic and early foetal losses in cattle and other ruminants. Reproduction
in Domestic Animals 43 (Suppl. 2), 260-267.
Douglas GN, Overton TR, Bateman HG, Dann HM, Drackley JK 2006. Prepartal plane of nutrition,
regardless of dietary energy source, affects periparturient metabolism and dry matter intake in Holstein cows.
Journal of Dairy Science 89, 2141-2157.
Drackley, JK, FC Cardoso 2014. Prepartum and postpartum nutritional management to optimize fertility in
high-yielding dairy cows in confined TMR systems. Animal, 8: S1, 5-14.
Drackley JK, Dann HM 2008. A scientific approach to feeding dry cows. In Recent advances in animal
nutrition – 2007. (Eds PC Garnsworthy and J Wiseman), pp. 43-74. Nottingham University Press,
Nottingham, UK.
Drackley JK, Dann HM, Douglas GN, Janovick Guretzky NA, Litherland NB, Underwood JP, Loor JJ 2005.
Physiological and pathological adaptations in dairy cows that may increase susceptibility to periparturient
diseases and disorders. Italian Journal of Animal Science 4, 323-344.
Emmanuel DG, Dunn SM, Ametaj BN 2008. Feeding high proportions of barley grain stimulates an
inflammatory response in dairy cows. Journal of Dairy Science 91, 606-614.
Emmanuel DG, Madsen KL, Churchill TA, Dunn SM, Ametaj BN 2007. Acidosis and lipopolysaccharide
from Excherichia coli B:055 cause hyperpermeability of rumen and colon tissues. Journal of Dairy Science
90, 5552-5557.
Friggens NC, Andersen JB, Larsen T, Aaes O, Dewhurst RJ 2004. Priming the dairy cow for lactation: a
review of dry cow feeding strategies. Animal Research 53, 453–473.
Galvão KN, Frajblat M, Butler WR, Brittin SB, Guard CL, Gilbert RO 2010. Effect of early postpartum
ovulation on fertility in dairy cows. Reproduction in Domestic Animals 45, e207-e211.
Garnsworthy PC, Sinclair KD, Webb R 2008. Integration of physiological mechanisms that influence fertility
in dairy cows. Animal 2, 1144-1152.
Garverick HA, Harris MN, Vogel-Bluel R, Sampson JD, Bader J, Lamberson WR, Spain JN, Lucy MC,
Youngquist RS 2013. Concentrations of nonesterified fatty acids and glucose in blood of periparturient dairy
cows are indicative of pregnancy success at first insemination. Journal of Dairy Science 96, 181-188.
Goldhawk C, Chapinal N, Veira DM, Weary DM, von Keyserlingk MA 2009. Prepartum feeding behavior is
an early indicator of subclinical ketosis. Journal of Dairy Science 92, 4971-4977.
Graugnard DE, Bionaz M, Trevisi E, Moyes KM, Salak-Johnson JL, Wallace RL, Drackley JK, Bertoni G,
Loor JJ 2012. Blood immunometabolic indices and polymorphonuclear leukocyt e function in peripartum
dairy cows are altered by level of dietary energy prepartum. Journal of Dairy Science 95, 1749-1758.
Graugnard DE, Moyes KM, Trevisi E, Khan MJ, Keisler D, Drackley JK, Bertoni G, Loor JJ 2013. Liver
lipid content and inflammometabolic indices in peripartal dairy cows are altered in response to prepartal
energy intake and postpartal intramammary inflammatory challenge. Journal of Dairy Science 96, 918-935.
Grum DE, Drackley JK, Younker RS, LaCount DW, Veenhuizen JJ 1996. Nutrition during the dry period
and hepatic lipid metabolism of periparturient dairy cows. Journal of Dairy Science 79, 1850-1864.
Grummer RR, Mashek DG, Hayirli A 2004. Dry matter intake and energy balance in the transition period.
Veterinary Clinics North America Food Animal Practice 20, 447-470.
Hammon DS, Evjen IM, Dhiman TR, Goff JP, Walters JL 2006. Neutrophil function and energy status in
Holstein cows with uterine health disorders. Veterinary Immunology and Immunopathology 113, 21-29.

15
Huzzey JM, Veira DM, Weary DM, von Keyserlingk MA 2007. Prepartum behavior and dry matter intake
identify dairy cows at risk for metritis. Journal of Dairy Science 90, 3220-3233.
Ingvartsen KL 2006. Feeding- and management-related diseases in the transition cow. Physiological
adaptations around calving and strategies to reduce feeding-related diseases. Animal Feed Science and
Technology 126, 175-213.
Ingvartsen KL, Dewhurst RJ, Friggens NC 2003. On the relationship between lactational performance and
health: is it yield or metabolic imbalance that cause production diseases in dairy cattle? A position paper.
Livestock Production Science 83, 277–308.
Ingvartsen KL, Moyes K 2013. Nutrition, immune function and health of dairy cattle. Animal 7 (Suppl. 1),
112-122.
Janovick NA, Drackley JK 2010. Prepartum dietary management of energy intake affects postpartum intake
and lactation performance by primiparous and multiparous Holstein cows. Journal of Dairy Science 93,
3086–3102.
Janovick NA, Boisclair YR, Drackley JK 2011. Prepartum dietary energy intake affects metabolism and
health during the periparturient period in primiparous and multiparous Holstein cows. Journal of Dairy
Science 94, 1385-1400.
Ji P, Osorio JS, Drackley JK, Loor JJ 2012. Overfeeding a moderate energy diet prepartum does not impair
bovine subcutaneous adipose tissue insulin transduction and induces marked changes in peripartal gene
network expression. Journal of Dairy Science 95, 4333-4351.
Jorritsma R, Wensing T, Kruip TAM, Vos PLAM, Noordhuizen JPTM 2003. Metabolic changes in early
lactation and impaired reproductive performance in dairy cows. Veterinary Research 34, 11-26.
Lean IJ, Van Saun R, Degaris PJ 2013a. Energy and protein nutrition management of transition dairy cows.
Veterinary Clinics North America Food Animal Practice 29, 337-366.
Lean IJ, Van Saun R, Degaris PJ 2013b. Mineral and antioxidant management of transition dairy cows.
Veterinary Clinics North America Food Animal Practice 29, 367-386.
LeBlanc, S 2010. Assessing the association of the level of milk production with reproductive performance in
dairy cattle. Journal of Reproduction and Development 56(Suppl.), S1-S7.
Loor JJ, Dann HM, Janovick Guretzky NA, Everts RE, Oliveira R, Green CA, Litherland NB, Rodriguez-Zas
SL, Lewin HA, Drackley JK 2006. Plane of nutrition prepartum alters hepatic gene expression and function
in dairy cows as assessed by longitudinal transcript and metabolic profiling. Physiological Genomics 27, 29-
41.
Loor JJ, Everts RE, Bionaz M, Dann HM, Morin DE, Oliveira R, Rodriguez-Zas SL, Drackley JK, Lewin HA
2007. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy
cows. Physiological Genomics 32, 105-116.
McArt JA, Nydam DV, Oetzel GR 2012. Epidemiology of subclinical ketosis in early lactation dairy cattle.
Journal of Dairy Science 95, 5056-5066.
Mulligan FJ, Doherty ML 2008. Production diseases of the transition cow. Veterinary Journal 176, 3-9.
Mulligan FJ, O’Grady L, Rice DA, Doherty ML. 2006. A herd health approach to dairy cow nutrition and
production diseases of the transition cow. Animal Reproduction Science 96, 331-353.
Ospina PA, Nydam DV, Stokol T, Overton TR 2010. Association between the proportion of sampled
transition cows with increased nonesterified fatty acids and β-hydroxybutyrate and disease incidence,
pregnancy rate, and milk production at the herd level. Journal of Dairy Science 93, 3595-3601.
Reist M, Erdin DK, von Euw D, Tschümperlin KM, Leuenberger H, Hammon HM, Morel C, Philipona C,
Zbinden Y, Künzi N, Blum JW 2003. Postpartum reproductive function: association with energy, metabolic
and endocrine status in high yielding dairy cows. Theriogenology 59, 1707-1723.
Roche JR, Bell AW, Overton TR, Loor JJ 2013. Nutritional management of the transition cow in the 21st
century – a paradigm shift in thinking. Animal Production Science 53, 1000-1023.

16
Rukkwamsuk T, Wensing T, Kruip TAM 1999. Relationship between triacylglycerol concentration in the
liver and first ovulation in postpartum dairy cows. Theriogenology 51, 1133-1142.
Santos JEP, Rutigliano HM, Sa Filho MF 2009. Risk factors for resumption of postpartum estrous cycles and
embryonic survival in lactating dairy cows. Animal Reproduction Science 110, 207-221.
Tharwat M, Takamizawa A, Hosaka YZ, Endoh D, Oikawa S. 2012. Hepatocyte apoptosis in dairy cattle
during the transition period. Canadian Journal of Veterinary Research 76, 241-247.
Thatcher W, Santos JEP, Staples CR 2011. Dietary manipulations to improve embryonic survival in cattle.
Theriogenology 76, 1619-1631.
Walsh RB, Walton JS, Kelton DF, LeBlanc SJ, Leslie KE, Duffield TF 2007. The effect of subclinical
ketosis in early lactation on reproductive performance of postpartum dairy cows. Journal of Dairy Science
90, 2788-2796.
Whitaker DA, Smith EJ, da Rosa GO, Kelly JM 1993. Some effects of nutrition and management on the
fertility of dairy cattle. Veterinary Record 133, 61-64.
Zurek E, Foxcroft GR, Kennelly JJ 1995. Metabolic status and interval to first ovulation in postpartum dairy
cows. Journal of Dairy Science 78, 1909-1920.

17
Como a dieta pode afetar a composição do leite
Bolivar Nóbrega de Faria
A composição e a porcentagem de proteína do leite de vacas têm recebido grande interesse
nos últimos anos, principalmente por influir diretamente no rendimento industrial. Uma
queda na concentração de proteína de 3,55% para 2,90% representa a necessidade de
aproximadamente 1.580 litros de leite adicionais para se produzir uma tonelada de queijo do
tipo Cheddar. Os resultados da alteração no teor de proteína do leite através da manipulação
da dieta giram em torno de 0,6 unidades percentuais (sendo mais prováveis 0,1 a 0,2
unidades). É modesto, se comparado ao potencial de alteração do teor de gordura do leite,
que gira em torno de 3,0 unidades percentuais. Porém, à medida que se aumenta o teor de
proteína do leite, geralmente também aumenta a produção de leite, o que já não ocorre com
a gordura (Waldner et al., 2004).
Além da nutrição, vários aspectos como estação do ano, composição racial, estádio de
lactação e doenças podem afetar o teor de proteína do leite. A sazonalidade relaciona-se com
a temperatura ambiente e disponibilidade de alimentos. Altas temperaturas e umidade
reduzem o teor de proteína e a produção de leite, pois o animal reduz o consumo de alimento.
Entre as raças leiteiras, a Holandesa possui a menor porcentagem de proteína no leite
enquanto a Jersey é a que possui a maior. A mudança na composição do leite utilizando a
seleção de animais é lenta, e não produz resultados imediatos, pois a herdabilidade das
características é baixa.
Os níveis de gordura do leite podem ser mais influenciados pela dieta. Maiores níveis de
gordura no leite também têm sido buscados pelos laticínios, uma vez que aumenta do
rendimento de alguns produtos nobres, como alguns tipos de queijo. Com isso, novas formas
de pagamento têm sido propostas se valorizando cada vez mais os constituintes do leite. Além
disso, concentrações baixas de gordura do leite têm sido relacionadas direta ou indiretamente
a várias doenças metabólicas, como o deslocamento de abomaso, laminite e, principalmente,
a acidose ruminal subaguda. Dessa forma, saber como a dieta afetas o perfil dos constituintes
do leite pode ser uma boa forma de se monitorar ou estimar a saúde dos animais.

18
A compreensão da composição do leite pode ser interessante ao produtor que precisa planejar
a lactação da vaca para maximizar os lucros. Isso envolve a compreensão do efeito da
alimentação, do manejo reprodutivo e da genética sobre a lactação. O conhecimento da
composição do leite também é importante para a indústria processadora, que depende da
manipulação das suas características físicas e químicas para a elaboração de diferentes
produtos lácteos, assim como para assegurar a sua qualidade.
O objetivo desta revisão é abordar os fatores nutricionais da vaca que interferem na
concentração de alguns componentes do leite.
Síntese dos carboidratos do leite
A lactose é o principal carboidrato encontrado no leite, sendo um dissacarídeo composto por
uma molécula de glicose ligada a uma molécula de galactose (Figura 1).
Figura 1. Fórmula estrutural de uma molécula de lactose e suas moléculas formadoras
A lactose possui um papel muito importante na síntese do leite. Sua função está relacionada
com a manutenção da osmolaridade do leite e nos processos de produção e secreção (Larson,
1995), ou seja, é o grande responsável por atrair a água para o leite (cerca de 50% da pressão
osmótica para a síntese do leite), sendo o principal responsável pelo volume do leite
produzido (Mühlbach, 2003). Cada grama de lactose do leite arrasta aproximadamente 10
vezes o mesmo peso em água. Devido à estreita relação entre a síntese de lactose e a
quantidade de água atraída para o leite, a lactose é o componente menos variável do leite
(Tabela 1). Segundo Hurley (2004) a lactose é relativamente insensível às mudanças na dieta

19
das vacas, no entanto, vacas subnutridas apresentam redução na produção de leite e na
percentagem de lactose, sendo que estes sintomas são revertidos quando dietas adequadas
são fornecidas.
Tabela 1. Variações nos constituintes de 257.540 amostras de leite nos
estados de São Paulo, Paraná e Santa Catarina (Ribas et al., 2004)
Constituinte do leite Concentração
média (%) Desvio padrão
Lactose 4,55 0,19
Proteína 3,24 0,24
Gordura 3,69 0,62
Sólidos totais 12,32 0,79
Compostos nitrogenados do leite
Como principais representantes das proteínas do leite estão as caseínas (α, β, κ), e as
lactoglobulinas (α e β), que representam mais de 90% da proteína total do leite, além de
albumina sérica e imunoglobulina G (Figura 2). A caseína constitui de 76 a 86% da proteína
presente no leite, sendo o componente determinante do rendimento industrial na produção de
queijo. É sintetizada pelas células secretoras, como resultado da expressão de quatro genes
(dois genes para α-caseína, s1 e s2; um para β-caseína e um para κ-caseína), sendo secretada
na forma de micelas, que são grupamentos de várias moléculas de caseína ligadas a íons
como o fosfato e o cálcio, e se encontram em suspensão no leite (De Peters & Cant, 1992).
Figura 2. Composição da fração proteica do leite de bovinos (Reis et al., 2008)

20
Ao contrário das caseínas, existem proteínas que se encontram em solução no leite, sendo
chamadas de proteínas do soro do leite. Estas proteínas possuem duas fontes principais,
podendo ser produzidas na glândula mamária, como a α-lactalbumina e β-lactoglobulina, ou
podem passar diretamente do sangue para a glândula mamária, como a albumina sérica e as
imunoglobulinas (De Peters & Ferguson, 1992). O leite possui ainda uma fração de
nitrogênio-não proteico (NNP), constituindo cerca de 5% da proteína bruta do leite, composta
principalmente de ureia (aproximadamente 48%) e em menor quantidade de creatinina,
amônia e outros compostos nitrogenados (De Peters & Ferguson, 1992) (Tabela 2).
Tabela 2. Fração de nitrogênio-não proteico do leite bovino (Bremel, 1995)
Componentes Concentração (mg N/dL)
Aminoácidos 3,7
Ureia 8,4
Creatina 1,2
Creatinina 0,2
Ácido úrico 0,8
Ácido orótico 1,2
Carnitina e acetil-carnitina 0,2
Gordura do leite
A gordura é o principal componente energético do leite, sendo responsável pelas
propriedades físicas, características industriais, qualidades organolépticas do leite e seus
derivados e, por isso, possuem um importante valor econômico (Bauman e Griinari, 2003).
Segundo Riel (1991), o tipo de gordura predominante no leite de vaca, com 97% a 99% dos
lipídios totais são os triglicerídeos. Os restantes são os fosfolipídios e esteróis, especialmente
o colesterol. Os triglicerídeos são constituídos principalmente de ácidos graxos, sendo na
gordura do leite constituídos em sua maioria de ácidos graxos de cadeia saturada (Tabela 3).
A gordura é o constituinte do leite mais sensível às variações, sendo possível observar
variações não somente em relação à raça, mas também ao longo da lactação (Figura 3).
Ultimamente várias pesquisas têm despertado interesse para o perfil de lipídios do leite. Isso
está mais relacionado aos seus efeitos na saúde humana, como é o interesse nas concentrações
de ácido linoleico conjugado (CLA) e não somente em relação aos seus efeitos sobre os

21
bovinos. Os CLA são compostos naturalmente encontrados na gordura de ruminantes que
exercem efeitos benéficos à saúde humana. Dentre os benefícios destacam-se efeitos anti-
carcinogênicos, alterações na partição de nutrientes e no metabolismo de lipídios,
propriedades antidiabéticas e antiaterogênicas, modulação da resposta imune, dentre outras
(Bauman et al., 2001). Portanto, seria desejável que as concentrações do CLA no leite fossem
aumentadas, já que são a maior fonte de CLA na dieta de humanos (Bauman et al., 2000).
Tabela 3. Composição de gordura do leite in natura ou em pó
(Adaptado de Jenkins e McGuire, 2006)
Constituinte do leite In natura Em pó
Água 88,32 2,47
Gordura 3,25 26,71
Ácidos graxos
(% do total)
Saturados 64,90 66,10
Monoinsaturados 28,30 31,30
Poliinsaturados 6,80 2,60
Figura 3. Variação do perfil da gordura do leite durante as primeiras semanas de lactação
em relação às concentrações obtidas na 16ª semana (Adaptado de Palmquist, 1993)

22
Com relação aos ruminantes, o foco tem sido dado a dois isômeros específicos, o cis-9 trans-
11 e o trans-10 cis-12 (Figura 4). O primeiro devido ao seu papel como um anti-carcinogênico
e o segundo, devido aos seus efeitos sobre o metabolismo de lipídios (Bauman et al., 2001).
Figura 4. Estrutura química do ácido linoléico e dos isômeros de CLA C18:2
cis-9, trans-11 e C18:2 trans-10, cis-12 (Bauman et al., 2001)
Síntese de carboidratos do leite
A produção de lactose no úbere está relacionada à síntese de propionato no rúmen e
disponibilidade de produtos gluconeogénicos. Quanto maior a disponibilidade de glicose
para a glândula mamária, maior será a possibilidade de produção de lactose (Figura 5) e,
consequentemente, de produção de leite.
UTP + glicose 1-P UDP-glicose + P-P (1)
UDP-glicose UDP-galactose (2)
UDP-galactose + glicose lactose + UDP (3)
UTP= uridina trifosfato; UDP= uridina difosfato; UDP-glicose= uridina difosforil glicose; UDP-galactose=
uridina difosforil galactose; P-P= pirofosfato; Enzimas: (1) uridina difosforil glicose pirofosfarilase; (2)
uridina difosforil galactose-4-epimerase; e (3) lactose sintetase (composta por galactosil transferase e α-
lactoalbumina).
Figura 5. Via metabólica de síntese de glicose na glândula mamária (Larson, 1995)
A glicose que o bovino necessita como precursor para a síntese de lactose na glândula
mamária é sintetizada no fígado via gliconeogênese, principalmente a partir do ácido

23
propiônico, sintetizado via degradação ruminal dos carboidratos oriundos da dieta. Outra
fonte importante de substrato para a gliconeogênese são os aminoácidos, oriundos da dieta
ou de mobilização corporal, principalmente no período pós-parto e/ou balanço energético
negativo (Figura 6). Após a absorção pelas células secretoras da glândula mamária, cerca de
60 a 70% da glicose é destinada para a síntese de lactose. No entanto, a glicose também tem
outras funções importantes na síntese dos constituintes do leite, sendo um importante
estimulador para a síntese de proteica, substrato para síntese de glicerol e da gordura do leite.
Figura 6. Fontes dos principais substratos utilizados para a síntese da lactose do leite
Síntese de compostos nitrogenados do leite
As proteínas do leite (caseínas e lactoglobulinas) são sintetizadas nas células secretoras da
glândula mamária a partir de aminoácidos provenientes do sangue ou transportados do
sangue (Figura 7). No entanto, alguns aminoácidos podem ser sintetizados na própria
glândula mamária através da utilização de precursores como a glicose e outros aminoácidos.
A síntese de proteína nas células secretoras ocorre no retículo endoplasmático rugoso. A
estrutura da proteína é pré-determinada pela informação genética contida no DNA presente
no núcleo da célula. O DNA serve de molde para a síntese de uma fita de RNA que contém
a informação sobre a sequência de aminoácidos para as proteínas. Vacúolos presentes no
citoplasma são formados para transporte da proteína sintetizada até o lúmen.
Algumas proteínas presentes no leite não são sintetizadas na glândula mamária e são
transportadas pelo sangue até entrarem no lúmen alveolar. A albumina encontrada no leite é

24
produzida no fígado e sua concentração no leite reflete a concentração no sangue. As
imunoglobulinas são transportadas para o leite via sangue e vasos linfáticos de origem do
baço e linfonodos. Linfócitos B produtores de anticorpos podem se alojar na glândula
mamária para auxiliar na síntese de imunoglobulinas do colostro (Larson, 1995).
Outro composto nitrogenado importante do leite, a ureia, não é produzido diretamente pela
glândula mamária. O nível de nitrogênio ureico no leite (MUN) está negativamente associado
à eficiência da utilização do nitrogênio da dieta para síntese proteica do leite. Como a ureia
se difunde livremente entre os fluidos do organismo animal, sua concentração na glândula
mamária reflete a concentração no sangue (Nousiainen et al., 2004). Qualquer fator que
interfira na eficiência da utilização da amônia presente no rúmen pelos microrganismos
ruminais irá resultar em aumento da quantidade de ureia no sangue. A baixa eficiência é
parcialmente devido à perda de nitrogênio amoniacal no rúmen. Estima-se que entre 40 a
80% do nitrogênio bacteriano tem origem na amônia ruminal (Hristov e Broderick, 1996), e
que a concentração ruminal de amônia é inversamente relacionada à disponibilidade de
carboidratos.
Figura 7. Fontes dos principais substratos utilizados para a síntese da proteína do leite

25
Síntese de gordura do leite
Segundo Harding (1995), a gordura do leite é sintetizada nas células epiteliais alveolares,
mais especificamente nas áreas do citoplasma ocupadas pelo retículo endoplasmático rugoso.
Os principais precursores da gordura do leite são ácidos graxos voláteis (AGV), o acetato e
o butirato, originados na fermentação ruminal de forragens e outros alimentos ricos em fibra,
e os ácidos graxos não esterificados da corrente sanguínea. Esses substratos irão dar origem
aos ácidos graxos, que posteriormente formarão os triglicerídeos do leite (Figura 8).
Figura 8. Fontes dos principais substratos utilizados para a síntese da gordura do leite
Ácidos graxos de cadeia curta, contendo entre 4 e 8 carbonos, e de cadeia média, com 10 a
14 carbonos, provem quase que exclusivamente da síntese “de novo”. Já os ácidos graxos de
cadeia longa, constituídos por cadeias superiores a 16 carbonos, são derivados da captação
direta dos lipídios da corrente sanguínea pela glândula mamária (Bauman e Griinari, 2003).
Os ácidos graxos com 16 carbonos podem ser obtidos através das duas fontes. Cerca de
metade dos ácidos graxos do leite são sintetizados pela própria glândula mamária, a partir da
síntese “de novo” tendo como principal fonte de carbono, principalmente, o acetato
produzido na fermentação ruminal (Bauman e Griinari, 2003).
Outra fonte de carbonos para a síntese “de novo” na glândula mamária é o β-hidroxibutirato
produzido pelo epitélio ruminal, a partir do butirato. Os ácidos graxos pré-formados captados
pela glândula mamária e diretamente usados para a síntese de gordura do leite são derivados
das lipoproteínas circulantes e dos ácidos graxos não esterificados originários da absorção

26
dos lipídios no trato gastrointestinal e da mobilização de reservas corporais, respectivamente.
A lipólise (mobilização de gordura corporal) é responsável por menos de 10% dos ácidos
graxos da gordura do leite, sendo a maior parte dos ácidos graxos de cadeia longa do leite
proveniente da absorção intestinal (Bauman e Griinari, 2003).
As gorduras de origem vegetal da dieta das vacas leiteiras são altamente insaturadas, desta
forma, essas gorduras quando ingeridas sofrem no rúmen a biohidrogenação, antes de serem
absorvidas pela corrente sanguínea na forma de triglicerídeos ligados a lipoproteínas. Esses
triglicerídeos são transportados pelo sangue até a glândula mamária, onde sofrem a quebra
em subunidades de glicerol e ácidos graxos livres que podem, então, ser absorvidos pelas
células da glândula mamária.
Fatores nutricionais que interferem nos compostos nitrogenados do leite
De forma geral, as estratégias nutricionais que visam o aumento da proteína do leite devem
ter como princípio o maior suprimento de aminoácidos e energia (glicose) para a glândula
mamária. A utilização de aminoácidos pela glândula mamária é dependente de uma série de
fatores como, irrigação sanguínea da glândula mamária, concentração arterial de
aminoácidos, eficiência no transporte e regulação intracelular das vias metabólicas (Bequette
et al. 1998). A eficiência na conversão de aminoácidos em proteína é baixa e inconstante (27
a 42%), e varia de acordo com o fornecimento de aminoácidos metabolizáveis (Doeppel et
al., 2004). Segundo Murphy e O’Mara, (1993), a limitação da produção de proteína pela
glândula mamária é devida principalmente aos aminoácidos essenciais, como a metionina,
lisina e histidina. De forma geral, lisina e metionina são consideradas aminoácidos limitantes
(Schwab et al., 1992).
Vários experimentos têm sido realizados na tentativa de se corrigir a relação lisina/metionina
através da suplementação desses aminoácidos na forma protegida, como hidroxi-análogos ou
precursores. No entanto, os resultados quanto aos amentos das concentrações de proteína no
leite são variáveis (Figura 9), uma vez que está resposta depende de outras variáveis
importantes como dieta base, ingestão de matéria seca, níveis energéticos e proteicos da dieta,
quantidade de fibra fisicamente efetiva, além do potencial genético do animal.
O aumento no fornecimento de aminoácidos para a glândula mamária pode ser obtido através
do aumento na quantidade de aminoácidos que chegam ao intestino delgado, ou seja, proteína

27
microbiana e proteína degradável no rúmen (PNDR). Com o maior suprimento de
aminoácidos absorvidos e disponíveis, além de um melhor perfil de aminoácidos não haveria
ou diminuiria a limitação dos principais aminoácidos essenciais para a síntese proteica
(Murphy e O’Mara, 1993). Além disso, os aminoácidos resultantes da mobilização proteica
endógena também colaborariam para esse melhor e maior pool.
Figura 9. Variação da proteína do leite (%) de vários experimentos realizados com
diferentes dietas, fontes e concentrações proteicas (Jenkins & McGuire, 2006)
Em dietas a base de silagem de milho e/ou suplementadas com milho, o aminoácido mais
limitante tende a ser a lisina. Em dietas ricas em soja e outras leguminosas pode haver
limitação de metionina, por haver grande suprimento de lisina (Tabela 4).
A utilização de aminoácidos protegidos, em especial lisina e metionina, tem apresentado
resultados modestos (Misciatteli et al., 2003). No entanto, há aumentos significativos na
eficiência de absorção dos aminoácidos em função da energia metabolizável da dieta. Mais
estudos são necessários para verificar outros aminoácidos possivelmente limitantes. Além
disso, o alto custo de suplementação dos aminoácidos protegidos tem limitado seu uso nos
países onde estes suplementos estão disponíveis.
O teor de proteína bruta da dieta possui efeito muito pequeno sobre o teor proteico do leite
(De Peters e Ferguson, 1992), relatou aumento de 0,02% para cada 1% de aumento na
2,6 2,7 2,8 2,9 3 3,1 3,2 3,3 3,4
Met (16,1)
F. Soja + Met (15,6)
Met (19,5)
Met (19,5)
Milho moído (15,7)
Soja espandida + Met (15,7)
Controle (18,8)
Controle (19,5)
Proteína do Leite (%)

28
proteína da dieta. No entanto, o fornecimento de dietas com deficiências de proteína pode
reduzir a concentração deste nutriente em 0,1 a 0,2 unidades percentuais (Sutton, 1989), além
de limitar a produção de leite. Pode-se dizer que a variação no teor proteico da dieta afeta
muito mais a produção de leite do que sua composição. A proteína bruta da dieta não está
correlacionada (p> 0,25) com a porcentagem de proteína no leite, mas fracamente
correlacionada com a produção de proteína no leite. Por outro lado, a proteína do leite
aumenta linearmente com o teor de PNDR na dieta (NRC, 2001).
Tabela 4. Composição de aminoácidos essenciais da proteína do leite (g/100 g aminoácidos), proteína
microbiana (g/100 g aminoácidos) e fontes de alimentos (g/100 g aminoácidos). Adaptado de Fonseca e
Santos (2000)
Fonte Arg His Ile Leu Lys Met Phe Thr
Leite 3,7 2,7 6,0 10,0 8,3 2,7 5,3 4,6
Proteína microbiana 5,1 2,0 5,7 8,1 7,9 2,6 5,1 5,8
Silagem de milho 1,7 0,8 2,8 6,5 1,8 0,8 3,0 2,5
Farelo de glúten de milho (21%) 4,2 2,9 2,5 8,0 2,5 2,1 3,3 3,8
Farelo de glúten de milho (60%) 2,8 1,8 3,4 14,1 1,5 2,8 5,7 3,0
Farelo de soja 6,9 2,2 5,1 6,9 5,9 1,3 4,5 3,5
No entanto, é muito difícil separar o efeito proveniente do aumento do teor da proteína da
dieta e o aumento no teor de energia ingerido pelo animal, tendo em vista que o teor de
proteína da dieta aumenta a eficiência microbiana, influenciando positivamente a ingestão de
matéria seca e a digestibilidade da dieta. A relação entre o aumento na ingestão de energia e
o aumento no teor de proteína do leite é mais importante que a relação entre o teor de proteína
ingerido na dieta e o aumento no teor de proteína do leite (Griinari et al., 1997). O aumento
da ingestão de energia por vacas em lactação aumenta a concentração de proteína no leite.
Estas mudanças na ingestão de energia são geralmente, obtidas por aumento da relação
concentrado/volumoso (Erro! Fonte de referência não encontrada.), por mudanças na fonte
de carboidratos ou por utilização de grãos processados.
Há limites para a elevação da produção de proteína do leite, com o aumento de concentrados
na dieta, que seria no ponto em que o pH cai sensivelmente e reduz a população celulolítica
do rúmen. Dietas com elevados teores energéticos costumam reduzir a gordura e aumentar a
proteína do leite. Pode-se aumentar potencialmente o fornecimento ou perfil de aminoácidos
disponíveis no intestino através da utilização de maiores quantidades de proteína não
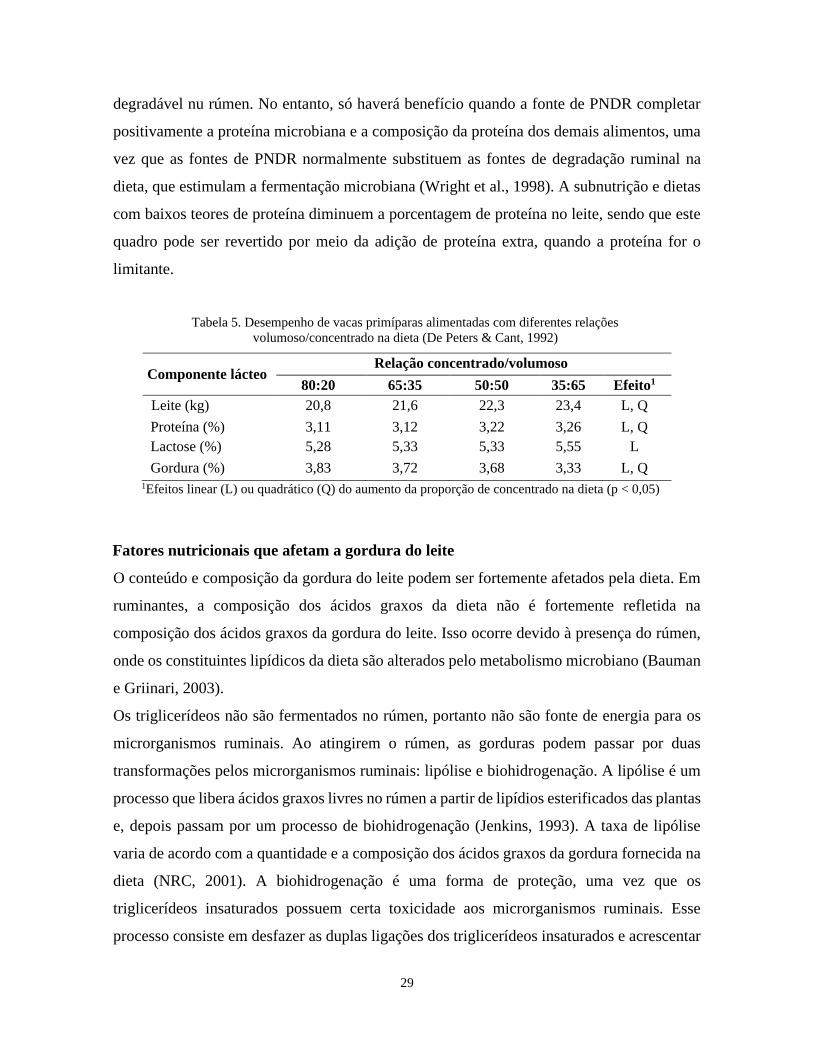
29
degradável nu rúmen. No entanto, só haverá benefício quando a fonte de PNDR completar
positivamente a proteína microbiana e a composição da proteína dos demais alimentos, uma
vez que as fontes de PNDR normalmente substituem as fontes de degradação ruminal na
dieta, que estimulam a fermentação microbiana (Wright et al., 1998). A subnutrição e dietas
com baixos teores de proteína diminuem a porcentagem de proteína no leite, sendo que este
quadro pode ser revertido por meio da adição de proteína extra, quando a proteína for o
limitante.
Tabela 5. Desempenho de vacas primíparas alimentadas com diferentes relações
volumoso/concentrado na dieta (De Peters & Cant, 1992)
Componente lácteo Relação concentrado/volumoso
80:20 65:35 50:50 35:65 Efeito1
Leite (kg) 20,8 21,6 22,3 23,4 L, Q
Proteína (%) 3,11 3,12 3,22 3,26 L, Q
Lactose (%) 5,28 5,33 5,33 5,55 L
Gordura (%) 3,83 3,72 3,68 3,33 L, Q 1Efeitos linear (L) ou quadrático (Q) do aumento da proporção de concentrado na dieta (p < 0,05)
Fatores nutricionais que afetam a gordura do leite
O conteúdo e composição da gordura do leite podem ser fortemente afetados pela dieta. Em
ruminantes, a composição dos ácidos graxos da dieta não é fortemente refletida na
composição dos ácidos graxos da gordura do leite. Isso ocorre devido à presença do rúmen,
onde os constituintes lipídicos da dieta são alterados pelo metabolismo microbiano (Bauman
e Griinari, 2003).
Os triglicerídeos não são fermentados no rúmen, portanto não são fonte de energia para os
microrganismos ruminais. Ao atingirem o rúmen, as gorduras podem passar por duas
transformações pelos microrganismos ruminais: lipólise e biohidrogenação. A lipólise é um
processo que libera ácidos graxos livres no rúmen a partir de lipídios esterificados das plantas
e, depois passam por um processo de biohidrogenação (Jenkins, 1993). A taxa de lipólise
varia de acordo com a quantidade e a composição dos ácidos graxos da gordura fornecida na
dieta (NRC, 2001). A biohidrogenação é uma forma de proteção, uma vez que os
triglicerídeos insaturados possuem certa toxicidade aos microrganismos ruminais. Esse
processo consiste em desfazer as duplas ligações dos triglicerídeos insaturados e acrescentar

30
um átomo de hidrogênio, formando uma ligação simples com o carbono (Figura 10). Isso é
nada mais que um processo de saturação do triglicerídeo.
Figura 10. Esquema da biohidrogenação ruminal
No processo de biohidrogenação, enzimas microbianas saturam o ácido linoleico (C18:2)
adicionando hidrogênio nas duplas ligações até que a molécula seja totalmente saturada e
transformada a ácido esteárico. No processo de formação do ácido esteárico, produtos
intermediários são formados, como os ácidos trans 18:1 e ácidos linoleicos conjugados
(CLA). Esses intermediários passam do rúmen ao intestino onde são absorvidos. Os maiores
substratos da biohidrogenação ruminal são o ácido linoleico e linolênico e a taxa de
biohidrogenação dos ácidos graxos é mais alta à medida que o grau de insaturação aumenta.
Para a maioria das dietas a taxa de biohidrogenação do ácido linoleico e linolênico é de 70-
95% e 85-100%, respectivamente (Doreau & Ferlay, 1994; Beam et al., 2000). Quando dietas
ricas em concentrado são fornecidas, a taxa de hidrogenação é reduzida, o que pode ser
atribuído à inibição da lipólise em pH ruminal baixo provocado por essas dietas (Van Nevel
& Demeyer, 1995; Van Nevel & Demeyer, 1996b). A hidrogenação também é afetada
quando uma quantidade excessiva de lipídios não protegidos está presente na dieta.
Dois importantes intermediários no processo de biohidrogenação são o trans-11 18:1 (ácido
trans-vaccênico) formados a partir de ácido linoleico e ácido linolênico e cis-9, trans-11

31
ácido linoleico conjugado (CLA) formado na biohidrogenação do ácido linoleico. Esses
intermediários estão presentes em quantidades apreciáveis na gordura dos ruminantes, em
uma relação de 3:1 (CLA: trans-vaccênico), mas no rúmen CLA cis-9, trans-11 é apenas um
intermediário transitório, enquanto o trans-vaccênico é que se acumula no rúmen.
Em condições de ácidos graxos insaturados em excesso no rúmen ou pH ruminal baixo,
alguns isômeros trans de ácidos graxos podem ser formados. Esses ácidos graxos estão
associados com depressão da gordura do leite (Griinari et al., 1998; Baumgard et al., 2000;
Bauman & Griinari, 2003). Portanto, dependendo de quanto os ácidos graxos saem do rúmen
na forma saturada, como ácido esteárico, ou na forma dos intermediários insaturados
influenciará em quanto de energia será disponibilizada para absorção e os efeitos dos ácidos
graxos na gordura do leite. Dessa forma, a dieta e alterações no ambiente ruminal podem
mudar as vias da biohidrogenação resultando em mudanças nos ácidos graxos
intermediários.
Outro aspecto importante que deve ser levado em consideração na dieta é a quantidade e
efetividade da fibra utilizada, o que reflete diretamente na condição ruminal e gordura do
leite (Tabela ). A fibra efetiva atua estimulando a ruminação e produção de saliva, o que
mantém o pH em níveis favoráveis para digestão da fibra, resultando em maior
disponibilidade de ácido acético, o principal precursor da gordura no leite.
Tabela 6. Ácidos graxos voláteis no rúmen e depressão da gordura do leite em vacas alimentadas com dieta
de baixa fibra, composta por alta quantidade de grãos e pouca forragem (Bauman & Griinari, 2003)
* p< 0,05
Parâmetro Dietas
Controle Baixa fibra
Leite (kg/dia) 19,1 20,9
Conteúdo de gordura (%) 3,6 1,7*
Produção de gordura (g/dia) 683 363*
Ácidos graxos voláteis no rúmen (% molar)
Acetato 67 46*
Propionato 21 46*
Butirato 11 9
Proporção acetato: propionato 3,2 1,0*
Produção no rúmen (moles/dia)
Acetato 29,4 28,1
Propionato 13,3 31,0*

32
Dietas com grandes quantidades de carboidratos prontamente fermentáveis e reduzida
quantidade de fibra, ou dietas com quantidade adequada de fibra, mas com fibra efetiva física,
possuem pouca habilidade de manutenção do funcionamento normal do rúmen, podendo
assim, levar a depressão na gordura do leite (Bauman e Griinari, 2003).
A inclusão de lipídios na dieta pode promover alterações sobre a gordura do leite, o que mais
uma vez é influenciado pelas alterações no ambiente ruminal (Palmquist et al., 1993). Os
efeitos observados, quando da adição de gordura à dieta, foram dependentes do tipo e
quantidade da gordura fornecida. O fornecimento de quantidades moderadas ou elevadas de
gordura insaturada reduz de maneira significativa o teor de gordura do leite, podendo esta
queda ser de até uma unidade percentual.
Griinari et al. (1998), confirmaram que dietas pobres em fibra e ricas em ácidos graxos
insaturados (óleo de milho) aumentam o conteúdo de C18:1 trans. Segundo estes mesmos
autores, este tipo de dieta está associado a uma diminuição significativa na produção e
conteúdo da gordura do leite, bem como nas concentrações de ácido esteárico, e menores
valores ruminais de pH. Isso provavelmente indica que houve uma redução das etapas finais
da biohidrogenação ruminal. O aumento da concentração intestinal de trans-vaccênico
diminui a capacidade lipogênica da glândula mamária (taxas de incorporação de acetato em
ácidos graxos) e a expressão de genes de enzimas relacionadas ao transporte de ácidos graxos
circulantes, diminui a síntese “de novo” de ácidos graxos, a dessaturação de ácidos graxos e
a formação de triglicerídeos. O mecanismo que provavelmente os esses ácidos graxos
insaturados diminuem a produção de gordura do leite envolve a redução na expressão do
mRNA de enzimas chave associadas à síntese de gordura do leite (Baumgard et al., 2002).
Kalscheur et al., (1997) observaram que vacas alimentadas com dietas ricas em concentrado
apresentaram diminuição da gordura do leite, acompanhada por aumento no conteúdo de C
18:1 trans.
Certos aspectos relacionados a formas de manejo de alimentação também alteram a ingestão
de fibra e, consequentemente, a gordura do leite. Pode-se destacar entre as deficiências no
manejo a baixa quantidade e qualidade de volumoso, sendo muitas vezes compensados pela
substituição por subprodutos fibrosos e concentrados; cochos mal dimensionados, o que
estimula a competição entre os animais, instalações inadequadas quanto ao conforto animal,
proporcionando menores tempos de ruminação e elevação do estresse.

33
Considerações finais
O aumento no teor de proteína do leite é interessante para as indústrias processadoras, pois
permite maior rendimento industrial, além da menor quantidade de água a ser transportada e
removida no beneficiamento. No entanto, para o produtor o aumento no teor de proteína do
leite só trará benefícios com a implementação de um sistema de pagamento por componentes,
onde o pagamento pela produção extra de proteína cobre os custos de produção. No entanto,
a opção por aumentar o teor de proteína do leite através da manipulação nutricional, deve
sempre avaliar a alternativa mais rentável disponível antes de proceder às alterações.
Referências
Bauman, D. E. et al. Update on conjugated linoleic acids (CLA). Proceedings of Cornell Nutrition Conference,
p.170-177, 2001.
Bauman, D. E.; et al. Technical note: production of butter with enhanced conjugated linoleic acid for use in
biomedical studies with animal models. J. Dairy Sci., v. 83, n. 11, p. 2422-2425, 2000.
Bauman, D. E.; Griinari, J. K. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr., v. 23, p. 203-227,
2003.
Baumgard, L. H. et al. Trans-10, cis-12 conjugated linoleic acid decreases lipogenic rates and expression of
genes involved in milk lipid synthesis in dairy cows. J. Dairy Sci., v. 85, p. 2155-2163, 2002.
Baumgard, L.H., Corl, B.A., Dwyer, D.A., Saebo, A. Bauman, D.E. Identification of the conjugated linoleic
acid isomer that inhibits milk fat synthesis. Am. J. Physiol., v. 278, p. R179-184, 2000.
Beam, T. M., Jenkins, T. C., Moate, P. J., Kohn, R. A., Palmquist, D. L. Effects of amount and source of fat on
the rates of lipolysis and biohydrogenation of fatty acids in ruminal contents. J. Dairy Sci. v. 83, p. 2564-2573,
2000.
Beckett, S. et al. Effects of monensin on the reproduction, health, and milk production of dairy cows J. Dairy
Sci., v. 81, p. 1563-1573, 1998.
De Peters, E. J.; Cant, J. P. Nutritional factors influencing the nitrogen composition of bovine milk: a review.
J. Dairy Sci. v. 75, p. 2043-2070, 1992.
De Peters, E. J.; Ferguson, J. D. Non nitrogen and protein distribution in the milk of cows. J. Dairy Sci. v. 75,
p. 3192-3209, 1992.
Doepel, L. et al. Milk protein synthesis as a function of amino acid supply. J. Dairy Sci. v. 87, p. 1279-1297,
2004.
Doreau, M., Ferlay, A. Digestion and utilization of fatty-acids by ruminants. Anim. Feed Sci. Technol., v. 45,
p. 379-396, 1994.
Fonseca, L. F. L; Santos, M. V. Conceitos básicos sobre composição de leite e métodos utilizados. 1° Curso on
line sobre qualidade do leite. Instituto Fernando Costa, Milkpoint. 2000.
Griinari, J. M. et al. Trans-Octadecenoic acids and milk fat depression in lactating dairy cows J. Dairy Sci., v.
81, p. 1251-1261,1998.
Griinari, J. M. McGuire, M. A. Dwyer, D. A. Bauman, D. E., Barbano, D. M. House, W. A. The role of insulin
in the regulation of milk protein synthesis in dairy cows. J. Dairy Sci. v. 80, p. 2361-2371, 1997.

34
Griinari, J.M., Dwyer, D.A., McGuire, M.A., Bauman, D.E., Palmquist, D.L., Nurmela, K.V.V.
Transoctadecenoic acids and milk fat depression in lactating dairy cows. J. Dairy Sci., v. 81, p. 1251-1261,
1998.
Harding, F. Milk quality. 1 ed., Glasgow: Blackie, p. 166, 1995.
Hristov, A.; Broderick, G. A. Synthesis of microbial protein in ruminally cannulated cows fed alfalfa silage,
alfalfa hay or corn silage. J. Dairy Sci., v. 79, p. 1627-1637, 1996.
Hurley, W. L. Nutritional factors affecting milk yield and composition. University of Illinois. 2004.
Jenkins, T. C., McGuire, M. A. Major Advances in Nutrition: Impact on Milk Composition, J. Dairy Sci., v. 89,
p. 1302–1310, 2006.
Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. v. 76, p. 3851-3863, 1993.
Kalscheur, K. F. B. et al. effect of dietary forage concentration and buffer addition on duodenal flow of trans C
18:1 fatty acids and milk fat production in dairy cows. J. Dairy Sci., v. 80, p. 2104-2114, 1997.
Larson, B. L. Biosynthesis & cellular secretion of milk. In: Larson, B. L. Lactation. Cap. 4. p.129-163. Iowa
State University Press. 1995.
Misciattelli, L. et al. Milk production, nutrient utilization, and endocrine responses to increased postruminal
lysine and meyhionine supply in dairy cows. J. Dairy Sci. v. 86, p 275-286, 2003.
Mühlbach, P.R.F. Nutrição da vaca em lactação e a qualidade do leite. In: Simpósio de Bovinocultura de Leite,
2003, Chapecó.
Murphy, J. J.; O’Mara, F. Nutritional manipulation of milk protein concentration and its impact on dairy
industry. Lives. Produc. Sci. v. 35, p 117-134, 1993.
National Research Council (NRC). Nutrient requirement of dairy cattle. Washington DC.: National Academy
Press. 2001.
Nousiainen, J. et al. Evaluation of milk urea as a diagnostic of protein feeding. J. Dairy Sci. v. 87, p. 386-398.
2004.
Nousiainen, J., Shingfield, K. J., Huhtanen, P. Evaluation of milk urea nitrogen as a diagnostic of protein
feeding. J. Dairy Sci. v. 87, p. 386–398, 2004.
Palmquist, D. L. et al. Adsa foundation symposium: milk fat synthesis and modification. J. Dairy Sci., v. 76, p.
1753-1771, 1993.
Reis, R. B., Glória, J. R., Vieira, L. R., Faria, B. N. Manipulação da composição do leite pela nutrição da vaca.
In Anais do Simpósio Mineiro de Nutrição, UFMG, 2008.
Ribas, N.P., Hartmann, W., Monardes, H.G., Andrade, U.V.C. Sólidos totais do leite em amostras de tanque
nos estados do Paraná, Santa Catarina e São Paulo. R. Bras. Zootec., v.33, n.6, p.2343-2350, 2004 (Supl. 3)
Riel, R. Composición y estructura físico-química de la leche. Ed. Acribia S.A.: Espanha, pp. 1-54, 1991.
Schwab, C.G., Bozak, C.K., Whitehouse, N.L., Mesbah, M.M.A. Amino acid limitation and flow to the
duodenum at four stages of lactation. I. Sequence of lysine and methionine limitation. J. Dairy Sci., v. 75, p.
3486–3502, 1992.
Sutton, J. D. Altering milk composition by feeding. J. Dairy Sci. v. 72, p. 2801-2814, 1989.
Van Nevel, C. J., Demeyer, D. I. Influence of pH on lipolysis and biohydrogenation of soybean oil by rumen
contents in vitro. Reprod. Nutr. Dev., v.36, p. 53-63, 1996
Van Nevel, C. J., Demeyer, D. I. Lipolysis and biohydrogenation of soybean oil in the rumen in vitro: Inhibition
by antimicrobials. J. Dairy Sci., v. 78, p. 2797-2806, 1995.
Waldner, D. N. et al. Managing milk composition: normal sources of variation. OSU: Extension facts. 2004.
Wright, T. C. et al. Effects of rumen-undegradable protein and feed intake on nitrogen balance and milk
production in dairy cows. J. Dairy Sci. v. 81, p 784-793, 1998.

35
Transtornos metabólicos da vaca leiteira no período de transição
Enrico Lippi Ortolani
Define-se período de transição como as quatro últimas e primeiras semanas em relação ao
parto. Nesse período a vaca leiteira está sujeita a ser acometida por várias enfermidades, com
destaque às de origem metabólica (Herdt, 2013). Várias condições predisponentes facilitam
o surgimento dessas enfermidades. No período pré-parto citam-se os seguintes fatores: (i) a
grande mobilização de nutrientes para o crescimento fetal, colostrogênese e o
desenvolvimento mamário; (ii) ajustes metabólicos necessários para favorecer a mobilização
de gordura e os teores de cálcio ionizável; e (iii) a representativa diminuição na ingestão de
matéria seca. No período pós-parto, embora ocorra uma lenta recuperação no apetite, o
animal deve subsistir a uma rápida mobilização e perda de nutrientes para atender a crescente
produção leiteira, em especial no concernente a gliconeogênese (Herdt, 2013).
Dentre as enfermidades metabólicas que se manifestam no período de transição, destacam-
se as seguintes: hipocalcemia acompanhada ou não de síndrome do animal caído; cetose com
presença ou não de esteatose hepática; edema mamário; deslocamento do abomaso e
hipomagnesemia (Herdt, 2013). Considerando a frequência e a relevância em nosso meio,
serão destacadas duas enfermidades: a hipocalcemia da vaca parturiente e a cetose da vaca
leiteira.
Hipocalcemia da vaca parturiente
Define-se a hipocalcemia como a dificuldade das vacas leiteiras em manter a homeostase do
cálcio livre plasmático, momentos antes ou em seguida ao parto, gerando inicialmente quadro
de tetania, seguido de paresia e paralisia, que se não foram tratadas levam frequentemente o
animal à morte (Ortolani, 1995b). A hipocalcemia tem as seguintes sinonímias: paresia da
parturiente, febre vitular ou febre do leite. Porém, essas duas últimas denominações são
incorretas, pois as vacas não têm síndrome febre e numa das fases clínicas podem até
apresentar hipotermia.

36
Aspectos epidemiológicos
O primeiro estudo epidemiológico no Brasil detectou uma incidência de 4,25% em rebanhos
leiteiros do Vale do Paraíba (SP). Contudo, deve-se mencionar que os rebanhos retinham
vacas além da 6ª gestação, e que a partir da 3ª lactação ocorreu aumento exponencial na
incidência, atingindo na 12ª lactação 31% (Ortolani, 1995a). A letalidade foi alta (12,8%),
em especial em vacas velhas ou tratadas em fases adiantadas da doença. Outros estudos
nacionais identificaram incidências de 1,8% e 5,2% (Corassin, 2004; Coelho, 2004). Vacas
das raças Jersey podem ser mais acometidas que outras raças, ocorrendo o mesmo em vacas
gordas (> 3,5 de condição corporal com escores entre 1-5) no dia do parto em relação às
magras (Radostits et al., 2007).
De acordo com dados nacionais, a hipocalcemia é inicialmente notada nas primeiras 24 horas
pós-parto (64%), reduzindo-se no segundo dia (31%) e diminuindo muito no 3º dia a seguir
(5%) (Ortolani, 1995 a). Porém, também é detectada em algumas vacas nos últimos dias
anteriores ao parto (Radostits et al., 2007).
Prejuízos econômicos
Essa enfermidade provoca grandes prejuízos econômicos, pois além de diminuir a produção
leiteira (até 80 L), nas primeiras seis semanas de lactação, causa mortalidade (Ortolani,
1995a) e predispõem a chance do surgimento de outras doenças do período de transição, na
seguinte ordem: cetose (23,5x), distocia (7,3x quando ocorre no período pré-parto), mamite
(5,4x) , metrite (4,7x), retenção de secundinas (4,3x), menor eficiência reprodutiva (4,2 x) e
deslocamento do abomaso à esquerda (3x) (Corassin, 2004; Herdt, 2013). A hipocalcemia é,
depois da distocia, a segunda principal causa de decúbito patológico (doença da vaca caída),
sendo responsável pelo surgimento de 38% destes casos (Herdt, 2013).
Metabolismo do cálcio na vaca leiteira
No plasma se encontram duas frações principais de cálcio (Ca): o cálcio livre (Ca2+) ou
ionizável (48%) e o cálcio ligado às proteínas (52%). Quadros de hipocalcemia só ocorrem
quando a fração de cálcio ionizável se reduz abaixo dos 50%. Os teores de cálcio ionizável
são finamente mantidos dentro de valores específicos (1,0 a 1,25 mmol/L) por vários
mecanismos hormonais. Na hipocalcemia a paratireoide produz o hormônio da paratireoide

37
(PTH), que estimula a hidroxilação da molécula 25-hidroxicolecalciferol nos rins, formando
a vitamina D3, muito mais potente que o composto anterior. A vitamina D3 aumenta a
absorção intestinal de Ca pela síntese de proteínas específicas para carrear Ca dos intestinos.
Porém, há necessidade dessa vitamina se ligar a receptores específicos e aptos nos
enterócitos. Além disso, a vitamina D3 aumenta a ressorção de Ca nos ossos e a reabsorção
nos túbulos renais. O PTH pode per se ter uma ação semelhante à vitamina D3 nos ossos e
rins. Na hipercalcemia entra em ação a calcitonina, produzida pela tireoide, que excreta Ca
pela urina e promove deposição de cálcio nos ossos (Ortolani, 1995b).
Patogenia da hipocalcemia
A hipocalcemia é fruto do déficit entre a demanda de Ca dispendido e a quantidade deste
elemento obtido na absorção intestinal e reabsorção óssea no período do periparto. A maior
perda de Ca é para a produção de colostro, que contém o dobro deste elemento (2,2 g/L) em
comparação ao leite normal. Vacas entre a 3ª e 6ª lactações atingem o ápice da produção de
colostro. Vacas boas produtoras podem perder muito mais Ca pelo leite (23 g/dia) que para
formação do feto no final da gestação (11 g/dia). Superordenha do colostro nos primeiros
dois dias de lactação podem desencadear quadros de hipocalcemia.
Toda vaca no dia do parto apresenta uma diminuta hipocalcemia. Para reduzir este déficit de
Ca tanto os processos de ressorção óssea como de absorção intestinal necessitam estar
bastante ativos. Porém, exatamente nestes dias existe uma crise na obtenção de cálcio.
Quanto mais velha for a vaca menor é a capacidade de mobilizar Ca dos ossos devido ao
reduzido número de receptores para a vitamina D3 nos enterócitos. Vacas Jersey também têm
o mesmo problema em relação ao número de receptores. Vacas que, em sucessivas lactações,
têm hipocalcemia apresentam dificuldade de aumentar os receptores para vitamina D3 antes
do parto (Goff, 2008). Dietas muito ricas em Ca, P, Na e K no pré-parto interferem na
produção de vitamina D3, ocorrendo o contrário em relação às dietas pobres em Ca. Vacas
que recebem no final da gestação dietas ricas em sais catiônicos (Na+; K+) apresentam pH
sanguíneo superior a 7,35, o que parece interferir negativamente na ligação do PTH aos seus
receptores nos ossos, diminuindo a ressorção óssea, e nos rins interferindo na reabsorção
tubular de cálcio (Herdt, 2000; Goff, 2008, DeGaris e Lean, 2008).

38
Quadro clínico
A hipocalcemia pode apresentar três estágios clínicos, de acordo com o teor de Ca livre no
plasma (Ortolani, 1995b). No primeiro estágio, o animal mantém a consciência, mas
apresenta os seguintes sinais nervosos: excitamento, tremores de cabeça e tetania com
hipersensibilidade. Manifesta ranger de dentes e protrusão de língua, rigidez dos membros e
aspecto de cavalete, podendo ficar em decúbito lateral. A temperatura retal pode se elevar até
os 41º C. Esse estágio dura no máximo 8 h.
No segundo estágio, a consciência fica deprimida e o animal se torna sonolento, a tetania
desaparece e há paresia muscular, as pupilas se dilatam e ocorre insensibilidade cutânea. O
esfíncter anal se relaxa e as fezes se tornam secas. E comum a atitude de “auto-auscultação”.
As vacas apresentam hipotermia, esfriamento de extremidade e a pele e muflo ficam secos.
Chama a atenção a taquicardia com hipofonese (Barreto Jr. et al., 2011). O pulso pode ficar
imperceptível, o movimento de rúmen desaparece e sobrevêm o meteorismo gasoso e é
comum a instalação da síndrome do animal caído. A temperatura retal diminuí sensivelmente
apresentando quando de hipotermia.
No terceiro estágio, há perda de consciência, flacidez de musculatura e instalação de quadro
comatoso, com hipotermia (<36ºC). A taquicardia é destacada (> 130 bat./min). A ausência
de tratamento provoca iminente morte por falência cardiorrespiratória.
Em condições nacionais os quadros de hipocalcemia são acompanhados de hipofosfatemia e
normomagnesemia (Ortolani, 1995a).
Tratamento
Deve ser realizado com infusão lenta (IV) de soluções contendo cálcio. A reversão do quadro
está intimamente ligada ao grau de hipocalcemia. Geralmente, o déficit de Ca plasmático é
na ordem de 6 g e tratamentos com essa quantidade resolvem 70% dos casos, porém parte
deles necessitam quantidades superiores (Ortolani, 1995a). Doses excessivas de cálcio
provocam recorrência da hipocalcemia horas após a terapia, devido à liberação de
calcitonina. Intoxicações iatrogênicas pelo Ca podem ocorrer matando o animal por parada
cardíaca, mas podem ser revertidas com uso de sulfato de atropina (10 mg/animal). Vacas
em decúbito respondem bem ao tratamento com doses adicionais (9 g) de fósforo (IV).

39
Prevenção
A prevenção deve ser realizada quando a incidência for superior a 1%. Isso é feito
manipulando a dieta nos últimos 20 dias pré-parto. Deve-se evitar o oferecimento dietético
excessivo de potássio e fósforo, que devem ficar restritos a 15 e 35 g/vaca/dia,
respectivamente. Por outro lado, estimula-se a adição de cloreto e enxofre na dieta para gerar
um balanço ligeiramente aniônico. As dietas usuais a base de volumosos, concentrados e
silagem têm mais cátions que ânions, contendo uma diferença cátion-ânion da dieta (DCAD)
em média de +100 a +300 mEq/kg MS. Para tornar a dieta aniônica existem sais comerciais
específicos ricos em cloreto (NH4+Cl) e enxofre (SO4Ca.2H20), os quais são suplementados
no final de gestação, em torno de 120 a 200 g/vaca/dia, suprimindo-os no dia do parto. Essa
quantidade proporciona que o balanço permaneça entre -150 a 0 mEq/kg MS, onde se obtêm
os melhores resultados na prevenção. A constatação que a dieta aniônica está atuando é a
queda no pH urinário, que se deve situar-se em torno de 6,0 em vacas de raça Holandesa e
entre 5,5 e 6,0 em vacas Jersey (Goff, 2008; DeGaris e Lean, 2008).
A dieta aniônica provoca uma ligeira acidose metabólica sistêmica que por seu turno aumenta
a produção de vitamina D3 e de PTH, aumentando a ação dos osteoclastos na ressorção óssea,
na reabsorção urinária e na absorção intestinal de Ca. Essa acidose provoca também melhor
aderência do PTH aos seus receptores ósseos e renais. Recomenda-se ainda na dieta pré-parto
que a quantidades de Ca não ultrapasse 70 g/vaca/dia e ofereçam entre 40 a 50 g de Mg
(Ortolani, 1995b; Goff, 2008; DeGaris e Lean. 2008).
Cetose da vaca leiteira
A cetose ocorre no início da lactação, devido a um déficit no metabolismo energético, em
especial na gliconeogênese, causado pela alta demanda de nutrientes no pico da lactação
podendo levar a um quadro de hipoglicemia e acetonemia. Isso provoca diminuições na
produção leiteira e no apetite e o surgimento de sinais nervosos.
Aspectos epidemiológicos
Teoricamente, quanto mais alta for a produção leiteira maior é risco do surgimento de cetose.
Vários trabalhos nacionais já relataram a presença de cetose em nossos rebanhos, com uma
incidência acumulada de 13 % a 24 % (Coelho, 2004; Corassin; 2004; Garcia, 2010). Ela é

40
mais frequente em vacas no decorrer da 2ª a 6ª lactações, com maior incidência na 3ª e 4ª
lactações, quando a produção leiteira é mais alta. A cetose tipo I é mais frequente em vacas
que pariram em condições corporais menores (< 3,5), enquanto na cetose tipo II ocorre o
contrário (> 3,5); a cetose tipo III (secundária) pode acometer fêmeas com diferentes
condições corporais. O pico de ocorrência é mais tardio no tipo I (3ª semana pós-parto) do
que no tipo II (1ª semana), podendo ser variado no tipo III.
Prejuízos econômicos
Estudo nacional constatou que vacas cetóticas têm uma redução láctea durante a lactação de
427 L, ou seja, 4,11% inferior a vacas hígidas (Corassin, 2004). A cetose aumenta o
surgimento de risco de doenças na seguinte ordem: 6,17x de deslocamento do abomaso à
esquerda e 3,4x para metrite e mamite. Por outro lado, aumentam o risco de cetose as
seguintes enfermidades: hipocalcemia 23,6x; retenção de placenta 3x e pneumonia 4,75 x
(Corassin, 2004; Herdt, 2013; Ortolani, 2014).
Metabolismo da glicose e geração de corpos cetônicos na vaca
Nos monogástricos, cerca de 60% da glicose sistêmica é proveniente da absorção dos
carboidratos dietéticos, porém nos ruminantes estes são fermentados no rúmen. Assim, os
ruminantes têm que gerar glicose quase que unicamente pelo processo de gliconeogênese
hepática. Cerca de 60% da fonte de glicose é derivada do propionato produzido no rúmen,
30% dos aminoácidos, 5% do lactato e 5% do glicerol. Os requerimentos de glicose são
variáveis de acordo com a fase produtiva. Uma vaca prenhe no final de gestação necessita de
500 g de glicose/dia, porém no pico da lactação esta quantidade mais que triplica numa fêmea
com produção acima de 20 L de leite (Herdt, 1988).
Os ruminantes têm pequeno estoque de glicogênio hepático, sendo os depósitos de gordura a
principal forma de energia armazenada no organismo. A mobilização dessas gorduras é feita
pela ação das lipases. No superávit energético as gorduras são estocadas, ocorrendo o inverso
em casos de déficit. Assim, a alta secreção de insulina bloqueia a ação das lipases, porém o
glucagon, o hormônio do crescimento, a adrenalina, o balanço energético negativo (BEN), o
estresse e o jejum fortemente estimulam as lipases, que mobilizam os triglicerídeos e os
transformam em glicerol e ácidos graxos livres, também denominados de ácidos graxos não

41
esterificados (AGNE). Esses últimos são carreados aos hepatócitos para serem oxidados ou
esterificados.
No déficit energético os AGNE são grandemente oxidados gerando energia e acetil-coenzima
A (acetil-CoA). Em condições normais, esse composto se combina com o oxalacetato
formando o citrato, que entra no ciclo de Krebs, gerando ATP e podendo formar glicose.
Porém, quando a produção de acetil-CoA for superior ao oxalacetato disponível ele é
transformado em acetoacetato e este em acetona e β-hidroxibutirato (βHB), conhecidos como
corpos cetônicos. Isso ocorre abundantemente na cetose, devido à grande oxidação dos
AGNE. Excesso de corpos cetônicos provocam redução da disponibilidade de glicose para a
glândula mamária, menor proliferação de linfócitos e decréscimo de atividade dos
neutrófilos. Conjuntamente com os altos AGNE os corpos cetônicos deprimem o apetite e
aumentam o grau de resistência insulínica. O βHB pode ser transformado, em 5% das vacas
cetóticas em álcool isopropílico, que provoca o surgimento de vários sintomas nervosos.
Os AGNE podem também ser esterificados no citosol dos hepatócitos, se unindo ao glicerol
e sendo envelopados por colesterol, fosfolipídios e proteína formando lipoproteína (LDH-
VLDL) para ser utilizada nos tecidos extra-hepáticos. Porém, se a quantidade de AGNE no
citosol for muito grande superior a capacidade da esterificação, as moléculas de AGNE se
condensam e são depositadas em forma de gordura provocando quadro de esteatose hepática,
comum na cetose tipo II.
O principal fator predisponente para o surgimento da cetose é a depressão e lenta recuperação
do apetite a partir do parto, acompanhado do pico da lactação nesse período quando a
mobilização de nutrientes é máxima. Assim, a cetose prevalece quando o BEN é muito
significativo, quer seja por desbalanço nutricional ou por menor apetite, ou pelo fato que as
vacas gordas apresentam menor depressão no apetite pós-parto e maior mobilização de
gorduras que vacas magras (Herdt, 1988; 2000; 2013).
Categorias de cetose
Várias classificações de cetose foram propostas até hoje. Classicamente, classificava-se
como primária ou secundária (Herdt, 2000). A primária causada pela falta de carboidrato na
dieta no pós-parto para prover de glicose e manter a alta produção de leite e a secundária,
determinada pelo menor apetite causado por outra doença concorrente.

42
Nova proposta foi feita por Holtenius e Holtenius (1996), comparando-se a cetose à diabetes
mellitus humana. A cetose tipo I ocorre no pico da lactação sem outra doença concorrente e
com esteatose hepática, caracterizada por hipoglicemia e hipoinsulinemia em vacas que
receberam dietas pobres em energia no pós-parto, e que parem com condição corporal abaixo
de 3,5 semelhante ao descrito na forma primária clássica.
A cetose tipo II se manifesta nos primeiros 15 dias pós-parto, caracterizada por
hiperinsulinemia, hiperglicemia e alta resistência insulínica (estado que a insulina diminui a
sua ação biológica nos tecidos insulino-dependentes: por exemplo, musculatura e gordura),
em vacas superalimentadas no pré-parto e que pariram com alta condição corporal (> 3,5),
frequentemente com doenças concorrentes e esteatose hepática. Contudo, estudos posteriores
verificaram que a hiperinsulinemia e hiperglicemia eram pouco frequentes e que muitos
quadros inflamatórios poderiam incrementar a resistência insulínica (Herdt, 2013).
Uma classificação errônea divide a cetose em clínica e “subclínica” de acordo com os teores
de corpos cetônicos e a presença ou não de sintomas clínicos (Herdt, 2013) ignorando que o
diagnóstico clínico é firmado pela presença de sinais clínicos e/ou exame laboratorial
positivo. Além disso, a presença de sinais clínicos está ligada a capacidade das vacas
tolerarem corpos cetônicos, pois algumas se mantem assintomáticas com alta acetonemia, e
outras manifestam a doença com teores menores (Herdt, 2000).
Assim, é lógico classificar a cetose em três formas. A cetose tipo I causada por privação
alimentar no pós-parto; a cetose tipo II gerada por excesso de alimentação no pré-parto; e a
cetose tipo III, ou secundária, em vacas bem manejadas dieteticamente no pré e pós-parto,
mas acometidas por enfermidades concorrentes após o parto.
Quadro clínico
Embora etiologicamente existam três tipos de cetose, quanto ao quadro clínico pode-se
dividir em duas formas: a típica, presente em 95% dos casos, e a nervosa ou bizarra. A forma
típica tem uma evolução clínica média de 16 dias no tipo I e de 8 dias no tipo 2. Em ambos
os casos, é marcada por um tripé de sintomas: hipogalaxia, hipofagia e sintomas nervosos. A
hipogalaxia gira em torno de 30%, mas pode atingir até 80%. A hipofagia chega a 50%, sendo
mais destacada nos casos tipo II. O apetite pode ser seletivo e o animal priva-se de consumir
primeiro o concentrado, depois o volumoso, podendo desenvolver pica, ingerindo cama, areia

43
ou terra. Isso leva a uma perda evidente de peso corpóreo atingindo até 70 kg no primeiro
mês pós-parto. Os animais podem apresentar sonolência, olhar fixo, ataxia, pequenos
cambaleios e cegueira parcial. As grandes funções podem estar modificadas devido aos
quadros concorrentes. Devido à diminuição do apetite pode ocorrer afundamento na fossa
paralombar esquerda em 70% dos casos e constipação e fezes ressecadas em 50%. Muitas
vacas com cetose tipo I e III apresentam pelame eriçado. Cerca de 50% das fêmeas
manifestam odor cetótico pelo ar expirado.
O quadro de cetose nervosa ou bizarra surge subitamente. São comuns os sintomas de
hiperestesia, delírio, agressividade, intensa sialorreia, amaurose levando a ataxia, cambaleios
e quedas, e apoio da cabeça em obstáculos. Caso esses animais não sejam devidamente
tratados podem sucumbir após o surgimento de quadro depressivo (Radostits et al., 2007;
Herdt, 2013).
Diagnóstico
Como os sintomas nem sempre são tão evidentes assim, é fundamental o diagnóstico
confirmativo da enfermidade. Nova ferramenta foi desenvolvida, disponível e barata em
nosso meio, que permite o diagnóstico rápido ao “pé-do-animal” por meio da determinação
de βHB em sangue total mensurado em tiras reativas, por química seca (Precision Xtra), o
qual apresenta alta sensibilidade e especificidade (Araújo, 2014). Confirma-se o diagnóstico
com valores iguais ou superiores a 1,4 mmol/L de βHB, e suspeito entre 1,2 e 1,3 mmol/L.
No rebanho pode ser realizada essa prova de rotina para o diagnóstico de cetose,
estabelecendo exames na cetose tipo I no 15º, 21º e 28º, e na cetose tipo II no 4º, 7º e 12º dias
pós-parto.
Tratamento
O tratamento deve atingir as seguintes metas: diminuições da lipólise e da cetogênese, e
aumentos da glicemia e do consumo de matéria seca. Geralmente, 50% das vacas não tratadas
apresentam recuperação espontânea lenta. O arsenal de tratamento conta com diferentes tipos
de medicamentos, a base de glicose (1.500 mL solução 20% IV); precursores de glicose
(propileno-glicol 300 mL/dia por 5 ou mais dias ou glicerol/glicerina 300 mL/dia por quatro
dias via oral); corticosteroides (dose única de 20 mg de dexametasona, 1 g de cortisona ou 5

44
mg de flumetasona); e medicamentos contendo fósforo e vitamina B12 (Butafosfan 25
mL/dia, correspondente a 0,43 g de P e 1.250 µg de vitamina B12/dose), por 3 dias
consecutivos (Herdt, 2013).
Os tratamentos devem ser priorizados de acordo com as condições clínicas, assim: a glicose
deve ser administrada na cetose tipos I e III e nas formas bizarras; precursores de glicose em
todos os tipos de cetose; corticosteroides na cetose tipo II, devendo ser evitado seu uso em
animais com quadros inflamatórios; e os suplementos de fósforo e B12 na cetose tipo I. Nos
quadros mais rebeldes de cetose tipo II (glicemia > 2,2 mmol/L; βHB > 2,4 mmol/L e alta
resistência insulínica) deve-se empregar glicose, os precursores e principalmente
corticosteroides (Gordon, 2013).
Prevenção
Deve-se iniciar na 8ª semana pré-parto com a oferta de quantidades adequadas de nutrientes.
Daí até o parto a ingestão de matéria seca (MS) reduz-se sensivelmente (2% do peso vivo a
1,5% em kg de MS; ex. vaca de 500 kg passa da ingestão de 10 kg MS/dia para 7,5 kg),
principalmente nas últimas três semanas. Isso deve ser compensado com o aumento na
densidade energética nessas semanas finais. Da 8ª à 4ª semana pré-parto as vacas devem
receber dieta com 12 % de proteína bruta (PB), 50% de NDT e 40% de Fibra Detergente
Neutro (FDN), passando a oferecer, em seguida, 15% de PB, 60% de NDT e 35% de FDN.
Do ponto de vista prático, no primeiro período a dieta deve conter 10% de concentrados
energéticos e 90% de volumosos (50% de silagem e 40% de capins tenros e feno), recebendo,
em seguida, 30% de concentrados e 70% de volumosos.
Durante as últimas oito semanas pré-parto o ganho de peso diário deve atingir 0,2 kg/dia
fazendo com que a condição corporal permaneça entre 3,0 e 3,25. No primeiro mês pós-parto
a dieta deve ser fornecida com 19% de PB, 73% de NDT e 30% de FDN, mantendo-se a
relação volumoso: concentrado em 60:40. Nos meses posteriores a dieta conterá 18% de PB,
60% de NDT e 32 % de FDN. Do ponto de vista prático, a quantidade de concentrados
energéticos oferecido nos primeiros três meses pós-parto é baseado na produção láctea, ou
seja: vacas produzindo mais de 27 L recebem 1 kg de concentrado/2 L de leite; com produção
de 18 a 27 L recebem 1 kg de concentrado/2,5 L de leite e com produção entre 13 e 18 L
recebem 1 kg de concentrado/3 L CE leite.

45
A suplementação da dieta com 335 g/vaca/dia de monensina no último mês de gestação e no
decorrer do pós-parto aumentou a gliconeogênese, diminuiu os teores de βHB e aumentou a
produção láctea. O combate aos vários tipos de estresse às vacas no período de transição é
essencial, para não provocar reduções na ingestão da matéria seca e aumento na incidência
de cetose. A prevenção das várias enfermidades presentes no pós-parto é de grande
importância para diminuir o risco de ocorrência de cetose (Herdt, 2013).
Referências
Araújo, C.A.S.C. Perfil metabólico e hormonal de ovelhas superalimentadas submetidas a hipercetonemia e
hiperlipidemia, e tratadas com somatotropina recombinante bovina (r-bST). 114p. Tese (Doutorado). Faculdade
de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2014.
Barreto Júnior, R.A.; Minervino, A.H.H.; Rodrigues, F.A.M.L.; Meira Júnior, E.B.S.; Lima, A.S.; Mori, C.S.;
Barros, I.O.; Ortolani, E.L. Avaliação do quadro clínico e perfil bioquímico de bovinos durante a indução e
tratamento da hipocalcemia. Brazilian Journal Veterinary Research Animal Science, v. 48, p.192-199, 2011.
Coelho, K.O. Impacto de eventos ocorridos antes e após o parto sobre o desempenho produtivo e reprodutivo
na lactação atual e na posterior de vacas Holandesas. 70p. Dissertação (Mestrado), Escola Superior de
Agricultura “Luis de Queiroz”, Universidade de São Paulo, Piracicaba, 2004.
Corassin, C.H. Determinação e avaliação de fatores que afetam a produtividade de vacas leiteiras: Aspectos
sanitários e reprodutivos. 113p. Tese (Doutorado), Escola Superior de Agricultura “Luiz de Queiroz”,
Universidade de São Paulo, Piracicaba, 2004.
Degaris, P.J.; Lean, I.J. Milk fever in dairy cows: a review of pathophysiology and control principals. The
Veterinary Journal, v. 176, p.58-69, 2008.
Garcia, A.M.B. Avaliação metabólica de vacas leiteiras submetidas a diferentes estratégias de prevenção do
balanço energético negativo do pós-parto. 83p. Dissertação (Mestrado), Faculdade de Veterinária, Universidade
Federal do Rio Grande do Sul, Porto Alegre, 2010.
Goff, J.P. The monitoring, prevention and treatment of milk fever and subclinical hypocalcemia in dairy cows.
The Veterinary Journal, v.176, p.50-57, 2008.
Gordon, J. Factors risks and treatment for ketosis in lactating dairy cows. 162 p. Tese (Doutorado), Universidade
de Guelph, Guelph, 2013.
Herdt, T.H. Metabolic diseases of ruminant livestock. Veterinary Clinics North America – Food Animal
Practice, v.4, p. 213 - 439, 1988.
Herdt, T.H. Metabolic disorders of ruminants. Veterinary Clinics North America. – Food Animal. Practice,
v.16, p.215-408, 2000.
Herdt, T.H. Metabolic diseases of dairy cows. Veterinary Clinics North America. – Food Animal Practice, v.
29, p.267-468, 2013.
Holtenius, P.; Holtenius, K. New aspects of ketone bodies in energy metabolism of dairy cows: a review. Journal
Veterinary Medicine A, v.43, p. 579-587, 1996.
Ortolani, E.L. Aspectos clínicos, epidemiológicos e terapêuticos da hipocalcemia de vacas leiteiras. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v.47, p. 799-808, 1995a.
Ortolani, E.L. Hipocalcemia da vaca parturiente. Cadernos Técnicos Escola Veterinária UFMG, v.14: p. 59-71,
1995b.

46
Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.J. Veterinary Medicine: A textbook of the diseases
of cattle, horses, sheep, pigs and goats. 10 ed. Philadelphia, Saunders Elsevier, 2007, p. 2156.

47
Transtornos metabólicos no período de transição da vaca leiteira com
ênfase nas enfermidades uterinas e da glândula mamária
Marcio Nunes Corrêa
Andressa Stein Maffi
Beatriz Riet-Correa
Ismael Mateus Cavazini
Marcelo Moreira Antunes
Viviane R. Rabassa
Cássio C. Brauner
O período de transição em vacas leiteiras compreende as três semanas que antecedem e as
três que sucedem o parto (Goff & Horst, 1997). Doenças como a hipocalcemia, metrite,
cetose, deslocamento de abomaso e mastite geralmente ocorrem neste período (Ziguer et al.
2007). As doenças do periparto de vacas leiteiras constituem um complexo, de modo que
uma condição patológica predispõe a ocorrência de outra. Por exemplo, uma vaca que
apresenta hipocalcemia é 4 vezes mais susceptível à ocorrência de retenção de placenta e
16,4 vezes mais susceptível à ocorrência de cetose.
O início da produção de leite impõe grandes desafios para os mecanismos responsáveis pela
homeostase energética, proteica e mineral da vaca. Durante o início da lactação, fatores
homeorréticos são responsáveis pelo direcionamento de grande quantidade de energia para a
glândula mamária e síntese de leite. Nesta fase, a vaca acaba mobilizando reservas corporais
e passa a enfrentar um estado de balanço energético negativo (BEN). Neste período, as vacas
sofrem uma diminuição da resposta imunológica. Este estado de imunossupressão em que o
animal se encontra deve-se à diminuição da atividade dos neutrófilos e dos linfócitos (Kehrli
et al. 1989), decorrentes das alterações do padrão de consumo alimentar e das alterações
hormonais e metabólicas a que a vaca leiteira está sujeita durante o periparto. Todos esses
fatores predispõem a um aumento acentuado do risco de desenvolver doenças que
influenciam negativamente a resposta produtiva e reprodutiva (Youngquist & Threlfall,
2007). Assim, o BEN, a inflamação e a resposta imune estão diretamente relacionados,
podendo atuar como causa ou consequência de enfermidades comuns no periparto.
No periparto, a concentração de cálcio associa-se com o decréscimo na contração muscular
e com a imunocompetência. O cálcio é necessário para a realização de várias funções vitais,
na síntese de leite, na contração muscular e desempenhando funções como segundo

48
mensageiro ou como cofator em diversas vias metabólicas intracelulares (Bruno, 2010).
Durante o período seco, as necessidades de cálcio da vaca são baixas (manutenção e
desenvolvimento fetal), ocorrendo incremento acentuado após o parto para a síntese de leite.
Entretanto, os mecanismos de controle do organismo não conseguem mobilizar tamanha
exigência, levando a quadros de hipocalcemia subclínica e clínica.
A hipocalcemia na forma subclínica contribui para a diminuição da contração da musculatura
lisa, da motilidade do rúmen e abomaso, redução da ingestão de matéria seca e da produção
de leite no início da lactação, assim como predispõe ao desenvolvimento de outras doenças.
Esta desordem atinge cerca de 20 a 40% de vacas leiteiras no período pós-parto, e pode
persistir em até cerca de 22% dos animais até os 10 dias pós-parto (Charbonneau et al. 2008).
A diminuição das reservas de cálcio intracelular no sistema imune prejudica de forma direta
a resposta celular a um estímulo de ativação, contribuindo para um estado de supressão
imunitária. Diversos autores indicam que cortisol, hormônio do crescimento (GH) e fator de
crescimento semelhante a insulina (IGF) alteram a proliferação de linfócitos. Também, níveis
elevados de ácidos graxos não esterificados (AGNE) e de beta- hidroxibutirato (BHB) são
sugeridos como agentes imunossupressores. Estudos in vitro demonstram que a presença
desses metabólitos em níveis elevados reduz a atividade de polimorfonucleares e torna os
animais mais suscetíveis a doenças uterinas e a mastite.
Metabolismo e relação com enfermidades uterinas
Os casos de retenção de placenta, metrite e endometrite clínica e subclínica afetam a
regeneração do endométrio e o retorno a ciclicidade, resultando em atraso na primeira
inseminação, aumento no número de inseminações por concepção, aumento no intervalo
entre partos e redução da taxa de parição (Hussain & Daniel, 1991). Os tratamentos dessas
enfermidades resultam no aumento dos custos de produção.
Dentre os fatores decisivos para o sucesso de uma nova concepção no pós-parto recente de
vacas, estão o retorno à ciclicidade ovariana e o término da involução uterina de forma
fisiológica. Muitos são os fatores que influenciam o desempenho reprodutivo de bovinos de
leite, como a nutrição, o manejo, a detecção e sincronização de cio, e a correta inseminação
(Garnsworthy et al. 2008; Schefers et al. 2010). Além disso, algumas alterações ocorridas
durante o pré-parto, como a redução de ingestão de matéria seca e a maior demanda de

49
energia, podem comprometer o estado nutricional do animal e o seu sistema imunológico,
facilitando a ocorrência de doenças uterinas, afetando o desempenho reprodutivo (Sordillo,
2009).
Antes do parto o lúmen uterino é estéril, ocorrendo a contaminação do útero por
microrganismos durante o parto, os quais são eliminados durante o processo de involução
uterina. Quadros de infecção uterina ocorrem quando os mecanismos de defesa do organismo
não conseguem debelar a infecção havendo um desbalanço entre os mecanismos de defesa
local e as bactérias patogênicas. Animais que tiveram quadros de retenção de placenta,
distocias, partos gemelares, abortos e curtos períodos de gestação, têm mais risco ao
estabelecimento destas infecções (Sheldon et al. 2006; Benzaquen et al. 2007; Bell &
Roberts, 2007). O estabelecimento, a gravidade e a persistência dos diferentes tipos de
infecção são influenciadas basicamente pela condição do ambiente uterino, por fatores
genéticos, além da imunidade inata e adquirida (Willians et al. 2007). A expressão dos sinais
clínicos depende da interação entre resposta imunológica, quantidade e patogenicidade dos
agentes microbianos (Azawi, 2008).
Retenção de placenta
A retenção de placenta (RP) caracteriza-se pela falta de expulsão da placenta no período de
12 horas após o parto, sendo associado mais frequentemente com abortos, distocias e
gestações gemelares, mas apresentando também fatores de risco como idade avançada,
estresse, falhas de manejo, hipocalcemia e cetose, deficiências de vitaminas A e E,
deficiências minerais como iodo e selênio, intoxicações, distúrbios hormonais, doenças como
brucelose, leptospirose e rinotraqueite infecciosa bovina. A incidência de RP varia entre
rebanhos, sendo de 3 a 27%, podendo chegar a 46% em partos gemelares. Casos não
complicados não têm efeito significativo sobre a fertilidade e sobre o intervalo parto-
concepção, porém podem apresentar como sequela a ocorrência de metrite. Os prejuízos
causados pela retenção placentária podem ultrapassar 150 reais (em torno de U$68,00) por
caso tratado, e são devidos a perdas de leite, que alcançam 250 L em média (levando em
conta o leite descartado pelo uso de antibióticos mais o que a vaca deixou de produzir pela
enfermidade), gastos com tratamento e mão de obra, atraso de 15 dias para concepção,

50
descarte de 6% dos animais acometidos pela doença, além da mortalidade que apresenta
valores médios de 1,5% dos casos (Corrêa et al. 2010).
Além das consequências diretas da RP sobre a produção de leite, também é um fator de risco
para outras desordens reprodutivas e doenças como a cetose, laminite e mastite. Como
consequência, a RP afeta negativamente o desempenho reprodutivo, podendo ser observado
o aumento no intervalo entre o parto e a primeira ovulação, redução da taxa de concepção ao
primeiro serviço e menor taxa de prenhez (Fourichon et al. 1999; Bell & Roberts, 2007;
Tillard et al. 2008; Sartori et al. 2013). Relatos de ocorrência de RP em vários países incluem
os Estados Unidos com 7,8% dos animais, Coréia com 18,3% e Alemanha com 27,8%
(Drillich et al. 2006).
Estudos indicam a relação de RP com desbalanços de cálcio e fósforo (Pelissier, 1976). Vacas
hipocalcêmicas são mais pré-dispostas à ocorrência de RP, por via indireta, devido ao maior
risco de partos distócicos. Ademais, a imunossupressão gerada pelos baixos níveis de cálcio,
associada a quadros de RP, predispõem estes animais a um quadro clínico de endometrite.
Segundo Gunnink (1984), o sistema imune identifica a placenta como um corpo estranho.
Em seu estudo, avaliando nível de fagocitose dos leucócitos, observou que vacas com RP
tiveram menor capacidade fagocítica comparada à de vacas saudáveis. Esta menor atividade
de leucócitos foi observada antes, durante e após o parto em outros estudos (Gunnink, 1984a,
1984b, 1984c). Vacas com redução da atividade de leucócitos apresentam redução na
capacidade de ingestão de matéria seca duas semanas antes e duas semanas após o parto
(Romaniukowa, 1984). No estudo realizado por Kimura et al. (2002), vacas com RP
apresentavam baixa função de neutrófilos desde o pré-parto até duas semanas pós-parto.
Vacas com níveis elevados de AGNE, indicando alta lipomobilização, têm alteração tanto da
função dos linfócitos quanto da proliferação e secreção de imunoglobulina M e de interferon
(Lacetera et al. 2004), afetando também a capacidade oxidativa e viabilidade dos
polimorfonucleares (Scalia et al. 2006). Concentrações plasmáticas de AGNE pré e pós-parto
acima de 300 e 600 µmol/L, respectivamente, são consideradas fatores de risco para RP e
metrite (Ospina et al. 2010). A RP é um dos principais fatores predisponentes da metrite,
sendo que 20 a 25% das vacas com RP desenvolvem metrite (Dubuc et al. 2011).

51
Metrite
Os eventos do pós-parto, juntamente com vários mecanismos de defesa do útero, facilitam a
eliminação da flora bacteriana indesejável (Senger, 2005). As contrações do miométrio
permitem a eliminação de grande parte dos lóquios. O retorno à ciclicidade ovariana (estro)
é o fator natural mais importante, assim como o aumento das concentrações de estradiol, que
favorece a migração de leucócitos para o trato reprodutivo. A secreção ovárica de estrogênio
antes da primeira ovulação promove uma maior resistência uterina a infecções, possibilitando
o retorno precoce à atividade reprodutiva (Hafez & Hafez, 2000). Porém, casos de lipidose
hepática têm sido associados com uma extensão no período anovulatório no pós-parto (Reist
et al. 2000), reduzindo o desempenho reprodutivo de vacas leiteiras em lactação (Jorritsma
et al. 2000), e favorecendo a doenças uterinas, como a metrite.
A metrite séptica no pós-parto de vacas leiteiras caracteriza-se por toxemia grave e abundante
corrimento uterino com odor pútrido, acompanhado ou não de retenção de placenta, é uma
inflamação que atinge todas as camadas do útero. A sua etiologia é multifatorial, admitindo-
se uma combinação de fatores, como retenção de placenta, insuficiente involução uterina
pós-parto e infecção uterina ascendente. A ocorrência é maior no período de 2 a 4 dias pós-
parto, tendo como fatores predisponentes a ocorrência de distocias, retenção de placenta,
obesidade e debilidade orgânica.
A metrite puerperal aguda pode apresentar incidência variável entre 11 e 36%. As perdas
econômicas ocorrem de forma indireta, como a redução na produção de 266 L de leite até o
dia 119 de lactação, incluindo o leite descartado pelo tratamento. Além disso, prejuízos com
o aumento de 7% nas taxas de descarte e redução da vida útil da vaca em 6 a 8 meses. A
metrite ainda altera a involução uterina e reduz o desenvolvimento folicular pós-parto,
elevando o intervalo parto-concepção e comprometendo os resultados da inseminação
artificial (Corrêa et al. 2010).
A contaminação bacteriana do útero no pós-parto é comum, requerendo uma resposta imune
de forma rápida das células de defesa, principalmente neutrófilos. O recrutamento de
neutrófilos é iniciado por meio de citocinas pró-inflamatórias, geradas a partir do endométrio,
no momento de reconhecimento das bactérias. Produção de baixos níveis de citocinas pró-
inflamatórias, incluindo o fator necrose tumoral α (TNFa), IL-1β e IL-6, tem sido associada
com o desenvolvimento de metrite (Galvão et al. 2011). Vacas com atividade de leucócitos

52
reduzidos apresentam maior propensão a apresentarem quadros de metrite e mastite no pós-
parto (Cai et al. 1994). Além disso, a redução na ingestão de matéria seca que se inicia no
pré-parto e se estende até o pós-parto imediato, associada à grande demanda metabólica, gera
um déficit negativo de energia e, conforme abordado anteriormente, vacas com níveis
elevados de AGNE apresentam alteração da capacidade dos linfócitos de proliferar e secretar
imunoglobulina M e interferon (Lacetera et al. 2004), afetando também a capacidade
oxidativa e viabilidade dos polimorfonucleares (Scalia et al. 2006). Vacas que perderam entre
1 e 1,5 pontos de condição corporal entre o período seco e o pós-parto apresentam um balanço
energético negativo mais intenso e maior tempo para recuperação da condição corporal, entre
a primeira e quarta semana de lactação, do que as vacas que perderam 0,75 pontos de ECC.
Esses animais apresentam maior ocorrência de metrite (62% x 27%) e doenças metabólicas,
como cetose, deslocamento de abomaso e hipocalcemia (23% x 2%). Os animais com maior
perda de ECC apresentaram 103 dias em aberto, enquanto os animais com perda moderada
de ECC tiveram 87 dias em aberto (Kim & Suh, 2003).
Atualmente, vem-se estudando alguns marcadores que indicam o risco do animal apresentar
metrite antes da ocorrência de sinais clínicos. O quadro clínico de metrite é marcado por
alterações em proteínas hepáticas, conhecidas como proteínas de fase aguda, sendo a
haptoglobina uma proteína de fase aguda positiva e a paraoxanase e a albumina proteínas de
fase aguda negativa. Um estudo demonstrou aumento dos níveis de haptoglobina em torno
do parto, em vacas com metrite (Huzzey et al. 2009). Redução nos níveis de albumina
indicam alterações hepáticas e está relacionado com incidência de doenças uterinas (Bertoni
et al. 2008; Burke et al. 2010). No estudo realizado por Schneider et al. (2013) pode-se
observar que vacas com metrite apresentavam redução na atividade de paraoxanase no dia 7
pré-parto e de albumina aos 21 dias antes do parto. Neste estudo também foi possível
observar uma maior taxa de prenhez dos animais saudáveis aos 150 dias pós-parto,
comparado aos animais com metrite.
Endometrite
A endometrite é uma enfermidade caracterizada por uma inflamação do endométrio e sua
ocorrência se dá a partir dos 21 dias pós-parto, não estando associada com envolvimento
sistêmico. Esta enfermidade tem sido subdividida em endometrite clínica e endometrite

53
subclínica, sendo a endometrite clínica caracterizada pela presença de conteúdo purulento ou
mucopurulento dos 21 aos 26 dias pós-parto. O quadro de endometrite subclínica caracteriza-
se pela presença de mais de 18% de polimorfonucleares na citologia uterina no período de
21 aos 33 dias pós-parto, ou mais de 10% de polimorfonucleares dos 34 aos 47 dias pós-
parto. Vacas com quadros de endometrite subclínica não apresentam descarga de conteúdo
uterino, porém, apresentam efeito negativo na reprodução (Sheldon et al. 2006).
Um estudo recente realizado por Krause et al. (2014) demonstrou que vacas com ciclos estrais
normais tiveram menores níveis de células polimorfonucleares (26,3%) quando comparado
a vacas anovulatórias (53,4%). Estudos realizados por Sheldon et al. (2008, 2009)
demostraram que em torno de 40% dos animais de alta produção, mantidos em sistemas
intensivos, podem apresentar metrite nas três primeiras semanas após o parto, com
persistência da enfermidade na forma de endometrite clínica em até 20% dos animais. Assim
como na retenção de placenta e na metrite, os casos de endometrite estão associados com
uma redução da função imune. Kim et al. (2005) observaram que vacas com endometrite
apresentaram linfócitos com capacidade fagocítica reduzida já no pré-parto.
Há uma relação positiva entre endometrite, BEN (caracterizado por AGNE e BHB elevados)
e quadros de cetose (Hammon et al. 2006; Galvão et al. 2010). Demonstrou-se maior
incidência de endometrite em animais com esteatose hepática (Fronk et al. 1980) ou com
acetonemia durante o puerpério recente (Markusfeld, 1985).
Outro fator a ser levado em consideração é a maior tendência que animais com sobrepeso no
período seco têm de desencadearem quadros mais intensos de BEN e cetose no pós-parto,
por apresentarem maior perda de condição corporal e terem um maior atraso na recuperação
de condição corporal. É sabido que animais com sobrepeso no pré-parto tem maior tendência
à hiperlipomobilização no pós-parto, o que leva a quadros mais intensos de BEN e por
consequência acentuação da imunossupressão. Vacas com elevada condição corporal (acima
de 3,5) no pré-parto apresentam uma redução no metabolismo hepático, resultando em uma
menor atividade oxidativa quando desafiadas no pós-parto. A concentração elevada de
AGNE no fígado leva também ao aumento de proteínas de fase aguda, principalmente
haptoglobina, sendo possível detectar este perfil durante o pré-parto. Sua detecção pode ser
um bom indicador no diagnóstico precoce de enfermidades do periparto, uma vez que as

54
vacas com endometrite apresentam níveis mais elevados desta proteína já no pré-parto
(Schneider et al. 2013).
Metabolismo e relação com a mastite
Mastite é a inflamação da glândula mamária que afeta uma elevada proporção de vacas
(Seegers et al. 2003). Economicamente, é considerada a principal doença do gado leiteiro,
uma vez que apresenta elevada prevalência e os custos com perdas e tratamentos são
geralmente elevados (Seegers et al. 2003). Animais acometidos por mastite apresentam
alterações na glândula mamária (Schwegler et al. 2013), como edema, vermelhidão, dor;
alteração no aspecto e na composição do leite, como o aumento da contagem de células
somáticas (Hortet & Seegers, 1998), além de apresentarem uma queda crônica na produção
(Bar et al. 2007). Dependendo da severidade da mastite, alterações sistêmicas também podem
estar presentes e incluem a diminuição da ingestão de matéria seca e alterações dos padrões
fisiológicos durante o exame clínico. Em casos mais graves há severa desidratação e
endotoxemia. Além disso, a mastite clínica altera as concentrações metabólicas sanguíneas,
modificando assim o perfil hormonal da vaca, prejudicando sua fertilidade (Hockett et al.
2000; Lavon et al. 2010).
Dependendo do agente etiológico e das outras condições envolvidas, pode ocasionar a morte
do animal. Não é incomum a mastite ser um dos principais, senão o principal motivo de
descarte de vacas leiteiras nos rebanhos. Grande parte do que determina o animal a
desenvolver ou não a doença está relacionada com a capacidade do agente causador em
vencer as defesas inatas e adquiridas da glândula mamária e se multiplicar, assim como a
capacidade do animal em responder aos estímulos patogênicos e desenvolver uma resposta
imune de forma rápida e que seja eficaz.
Como já citado, durante o periparto, além do BEN, as vacas passam por uma fase de
imunossupressão, que pode ser definida como a baixa capacidade do organismo em formar
uma resposta imune rápida e eficaz para combater uma infecção. Durante esta fase, os
animais estão mais propensos ao surgimento de doenças infecciosas, entre elas a mastite.
Observa-se que aproximadamente 25% das mastites ocorrem durante as primeiras duas
semanas de lactação e que a maioria são causadas por agentes oportunistas, o que indica
influência da condição imune nesta fase.

55
Com o objetivo de avaliar se o BEN é o principal causador desses distúrbios, pesquisadores
induziram restrição alimentar em vacas no meio da lactação e procederam com infusões
intramamárias de endotoxinas, observando que o BEN neste momento não afetou
significativamente a expressão de moléculas de adesão sob a superfície de leucócitos bovinos
(Perkins et al. 2001; Moyes et al. 2009). Em outro estudo, inocularam quantidades
semelhantes de Escherichia coli em grupos de vacas no início ou meio da lactação,
identificando que as vacas periparturientes tiveram crescimento bacteriano mais rápido,
infecção mais severa e maior temperatura corporal do que as vacas no meio da lactação
(Shuster et al. 1996).
As principais células envolvidas no sistema imune da glândula mamária são os neutrófilos e
os linfócitos. Os neutrófilos são um dos principais tipos celulares do sistema imune, pois são
a primeira linha de defesa, respondendo de forma rápida (em minutos) e não precisam de
contato prévio com o agente. Uma de suas principais funções é a fagocitose. Os linfócitos
representam uma segunda linha de defesa, muito importante pelo fato da memória
imunológica. O estabelecimento de uma infecção em qualquer órgão ou tecido é dependente
de um balaço delicado entre os mecanismos de defesa nativos do organismo e a capacidade
da bactéria de resistir a condições desfavoráveis de crescimento. Quando há desequilíbrio,
como no caso da imunossupressão, a doença ocorre.
Com o objetivo de avaliar o efeito da produção de leite sobre a imunossupressão durante o
periparto, foram utilizadas um grupo de vacas com glândula mamária (inteiras) e outro com
vacas mastectomizadas, nas quais se avaliou a capacidade dos neutrófilos em eliminar
microrganismos (atividade da mieloperoxidade). As vacas mastectomizadas tiveram uma
rápida recuperação da habilidade fagocitária dos neutrófilos, enquanto que nos animais
inteiros foram necessárias cerca de três semanas após o parto para haver essa completa
recuperação (Kimura et al. 1999). Em virtude desse fato, alguns trabalhos vêm investigando
componentes metabólicos individuais associados ao BEN. A cetose é a elevação dos corpos
cetônicos na corrente sanguínea, urina ou leite, associado a um declínio na glicose sanguínea.
Os corpos cetônicos são resultado da excessiva mobilização lipídica que ocorre nas vacas em
BEN e da incapacidade do fígado em metabolizar o excesso de ácidos graxos (González et
al. 2000). Os corpos cetônicos são utilizados como energia alternativa por diversos tecidos.
Entretanto, muitas das células imunes são afetadas negativamente pelos níveis de metabólitos

56
típicos de um ambiente cetônico (baixa glicose, elevação nos corpos cetônicos e presença de
AGNE).
Dessa forma, diversos trabalhos vêm demonstrando que vacas cetônicas apresentam quadros
mais severos de mastite do que vacas saudáveis (Oltenacu & Ekesbo, 1994; Kremer et al.
1993). Linfócitos de vacas com cetose clínica e subclínica, cultivados in vitro, responderam
menos a desafios patogênicos, quando comparados com linfócitos de vacas saudáveis
(Kandefer & Szerszen et al. 1992). Ademais, houve uma diminuição na expressão de
moléculas de superfície associadas com a função de neutrófilos sanguíneos (Zerbe et al.
2000).
A excessiva mobilização lipídica no período de transição leva a um acúmulo de AGNE na
corrente sanguínea, o que causa efeitos maléficos sobre a função das células imunes. Estudos
in vitro demonstraram a diminuição da função, viabilidade e proliferação das células
mononucleares quando em contanto com AGNE (Scalia et al. 2006; Ster et al. 2012).
Portanto, níveis elevados de AGNE na proximidade do parto são preditivos para a
subsequente mastite (Moyes et al. 2009; Schwegler et al. 2013), além de outras desordens
como o deslocamento de abomaso, cetose clínica, metrite e retenção de placenta (Ospina et
al. 2010).
Além dos metabólicos envolvidos diretamente com o BEN (AGNE, corpos cetônicos) sabe-
se que o cálcio também exerce um papel importante sobre a ocorrência das mastites. A função
principal do cálcio está relacionada com a contração muscular. Dessa forma, em vacas
hipocalcêmicas o canal do teto da glândula mamária pode ficar aberto e favorecer a
penetração de patógenos ambientais para o interior da luz do órgão. Além disso, vacas caídas
e que ficam com os tetos expostos ao solo por longos períodos também tem chances maiores
de desenvolver mastites. A hipocalcemia também age como mais um fator estressante para a
vaca durante o pré-parto, já que os animais com hipocalcemia subclínica apresentam níveis
mais elevados nas concentrações plasmáticas de cortisol, quando comparados com vacas
saudáveis, prejudicando ainda mais o organismo já imunossuprimido (Horst & Jorgensen,
1982). Ademais, o cálcio também exerce influência sobre a imunocompetência celular, uma
vez que é importante na sinalização intracelular para a ativação das células imunes (Kimura
et al. 2006). Apesar dos maiores prejuízos serem observados com a forma clínica da doença,
é importante ressaltar que grande parte das vacas permanecem com hipocalcemia subclínica

57
durante as primeiras semanas após o parto, influenciando a função imune (Goff et al. 1996).
Foi demonstrado que as vacas com hipocalcemia clínica foram 8,1 vezes mais propensas a
desenvolverem mastite do que as vacas saudáveis (Curtis et al. 1983).
Considerações finais
O período de transição influencia a vaca leiteira em nível produtivo e reprodutivo no decorrer
da lactação, sendo por isso considerado um momento crucial no ciclo de produção da vaca.
A ocorrência de mastite e enfermidades uterinas são reconhecidas como os principais
problemas na sanidade dos rebanhos leiteiros e o metabolismo está diretamente relacionado
com a ocorrência dessas enfermidades, uma vez que fatores como o BEN, corpos cetônicos
e hipocalcemia influenciam diretamente a resposta imune dessas vacas nesse período.
Referências
Azawi, O.I. Postpartum uterine infection in cattle. Animal Reproduction Science, v. 105, p. 187-208, 2008.
Bar, D.; Gröhn, Y.T.; Bennett, G. et al. Effect of repeated episodes of generic clinical mastitis on milk yield in
dairy cows. Journal of Dairy Science, v. 90, p. 4643-4653, 2007.
Bell, M. J & Roberts, D. J. 2007. The impact of uterine infection on a dairy cow’s performance. Theriogenology
68: 1074-1079.
Benzaquen, M. E., Risco, C. A., Archbald, L.F. et al. 2007. Rectal temperature, calving-related factors, and the
incidence of puerperal metritis in postpartum dairy cows. Journal of Dairy Science 90: 2804-2814.
Bertoni, G., Trevisi, E., Han, X. et al. 2008. Effects of inflammatory conditions on liver activity in puerperium
period and consequences for performance in dairy cows. Journal of Dairy Science 91(9): 3300-3310.
Bruno, R. G. S. 2010. Nutrition and reproduction in modern dairy cows. Mid-south ruminant nutrition
conference, Texas, USA, 51-56.
Burke, C. R., Meier, S., McDougall, S. et al. 2010. Relationships between endometritis and metabolic state
during the transition period in pasture-grazed dairy cows. Journal of Dairy Science 93(11): 5363-5373.
Cai T. Q., Weston, P. G., Lund, L. A. et al. 1994. Association between neutrophil functions and periparturient
disorders in cows. American Journal of Veterinary Research 55: 934-943.
Charbonneau, E., Chouinard, P. Y., Tremblay, G. F. et al. 2008. Hay to reduce dietary cation-anion difference
of dairy cows. Journal of Dairy Science 91: 1585-1596.
Corrêa, M. N., González, F. H. D., Da Silva, S. C. 2010. Transtornos metabólicos nos animais domésticos.
Editora Universitária PREC-UFPel, Pelotas, RS, ed. 1.
Curtis, C. R., Erb, H. N., Sniffen, C. J. et al. 1983. Association of parturient hypocalcemia with eight
periparturient disorders in Holstein cows. Journal of the American Medical Association 183(5): 559-561.
Drillich, M. M., Mahlstedt, U., Reichert, B. A. et al. 2006. Strategies to improve the therapy of retained fetal
membranes in dairy cows. Journal of Dairy Science 89: 627-635.
Dubuc, J., Duffield, T. F., Leslie, K. E. et al. 2011. Randomized clinical trial of antibiotic and prostaglandin
treatments for uterine health and reproductive performance in dairy cows. Journal of Dairy Science 94: 1325-
1338.

58
Fourichon, C., Seegers, H., Bareille, N. et al. 1999. Effects of disease on milk production in the dairy cow: a
review. Preventive Veterinary Medicine 1: 1-35.
Fronk, T. J., Schultz, L. H., Hardie, A. R. 1980. Effect of period overconditioning on subsequent
metabolic disorders and performance of dairy cows. Journal of Dairy Science 63: 1080-1090.
Galvão, K. N., Flaminio, M. J., Brittin, S. B. et al. 2010. Association between uterine disease and indicators of
neutrophil and systemic energy status in lactating Holstein cows. Journal of Dairy Science 93: 2926-2937.
Galvão, K. N., Santos, N. R., Galvão, J. S. et al. 2011. Association between endometritis and endometrial
cytokine expression in postpartum Holstein cows. Theriogenology 76: 290-299.
Garnsworthy, P. C., Lock, A., Mann, G. E. et al. 2008. Nutrition, metabolism and fertility in dairy cows: Dietary
fat content and ovarian function. Journal of Dairy Science 91: 3824-3833.
Goff, J. P., Horst, R. P., Jardon, P. W. et al. 1996. Field trials of an oral calcium propionate paste as an AID to
prevent milk fever in periparturient dairy cows. Journal of Dairy Science 79: 378-383.
Goff, J. P. & Horst, R. L. 1997. Physiological changes at parturition and their relationship to metabolic
disorders. Journal of Dairy Science 80: 1260-1268.
González, F., Barcellos, J., Patiño, H. 2000 Perfil metabólico em ruminantes: seu uso em nutrição e doenças
nutricionais. Editora UFRGS, Porto Alegre, RS.
Gunnink, J. W. 1984a. Pre-partum leucocytic activity and retained placenta. Veterinary Quaterly 6(2): 52-54.
Gunnink, J. W. 1984b. Post-partum leucocytic activity and its relationship to caesarian section and retained
placenta. Veterinary Quaterly 6(2): 55-57.
Gunnink, J. W. 1984c. Retained placenta and leukocytic activity. Veterinary Quaterly 6(2): 49-51.
Hafez, E. S. E. & B. Hafez. 2000. Reproduction in Farm Animals. Lea and Febiger, Philadelphia, USA. ed. 7.
Hammon, D. S., Evjen, I. M., Dhiman, T. R., Goff, J. P. et al. 2006. Neutrophil function and energy status in
Holstein cows with uterine health disorders. Veterinary Immunology and Immunopathology 113: 21-29.
Hockett, M. E., Hopkins, F. M., Lewis, M. J. et al. 2000. Endocrine profile of dairy cows following
experimentally induced clinical mastitis during early lactation. Animal Reproduction Science 58: 241-251.
Horst, R. L. & Jorgensen, N. A. 1982. Elevated plasma cortisol during induced and spontaneous hypocalcemia
in ruminants. Journal of Dairy Science 65: 23-32.
Hortet, P. & Seegers, H. 1998. Review - Loss in milk yield and related composition changes resulting from
clinical mastitis in dairy cows. Preventive Veterinary Medicine 37: 1-20.
Hussain, A. M. & Daniel, R. C. W. 1991. Bovine endometritis: current and future alterative therapy. Journal of
the Veterinary Medicine 38: 641-651.
Huzzey, J. M., Duffield, T. F., Leblanc, S. J. et al. 2009. Short communication: Haptoglobin as an early indicator
of metritis. Journal of Dairy Science 92(2): 621-625.
Jorritsma, R., Jorritsma, H., Schukken, Y. H. et al. 2000. Relationships between fatty liver and fertility and
some periparturient diseases in commercial Dutch dairy herds. Theriogenology 54: 1065-1074.
Kandefer-Szerszen, M., Filar, J., Szuster-Ciesielska, A. et al. 1992. Suppression of interferon response of bovine
leukocytes during clinical and subclinical ketosis in lactating cows. Dtsch. Tierarztl. Wochenschr. 99: 440-443.
Kehrli, M. E. J. R., Nonnecke, B. J., Roth, J. A. 1989. Alterations in bovine peripheral blood lymphocyte
function during the periparturient period. American Journal of Veterinary Research 50: 215-220.
Kim I. H., Na, K. J., Yang M. P. 2005. Immune responses during the peripartum period in dairy cows with
postpartum endometritis. Journal of Reproduction and Development 51: 757-764.
Kim, I. H & Suh, G. H. 2003. Effect of the amount of body condition loss from the dry to near calving periods
on the subsequent body condition change, occurrence of postpartum diseases, metabolic parameters and
reproductive performance in Holstein dairy cows. Theriogenology 60: 1445-1456.

59
Kimura, K., Goff, J. P., Kehrli, M. E. 1999. Effects of the presence of the mammary gland on expression of
neutrophil adhesion molecules and myeloperoxidase activity in periparturient dairy cows. Journal of Dairy
Cows 82: 2385-2392.
Kimura, K., Goff, J. P., Kehrli, M. E. Jr. et al. 2002. Decreased neutrophil function as a cause of retained
placenta in dairy cattle. Journal of Dairy Science 85: 544-550.
Kimura, K., Reinhardt, T. A., Goff, J. P. 2006. Parturition and hypocalcemia blunts calcium signals in immune
cells of dairy cattle. Journal of Dairy Science. 89: 2588-2595.
Krause, A. R. T., Pfeifer, L. F. M., Montagner, P. et al. 2014. Associations between resumption of postpartum
ovarian activity, uterine health and concentrations of metabolites and acute phase proteins during the transition
periods in Holstein cows. Animal Reproduction Science 145: 8-14.
Kremer, W. D., Noordhuizen-Stassen, E. N., Grommers, F. J. et al. 1993. Severity of experimental Escherichia
coli mastitis in ketonemic and nonketonemic dairy cows. Journal of Dairy Science 76: 3428-3436.
Lacetera, N., Scalia, D., France, O. et al. 2004. Effects of nonesterified fatty acids on lymphocyte function in
dairy heifers. Journal of Dairy Science 87: 1012-1014.
Lavon, Y., Leitner, G., Voet, H. et al. 2010. Naturally occurring mastitis effects on timing of ovulation, steroid
and gonadotrophic hormone concentrations, and follicular and luteal growth in cows. Journal of Dairy Science
93: 911-921.
Markusfeld O. 1985. Relationship between overfeeding, metritis and ketosis in high yielding dairy cows.
Veterinary Record 116: 489-491.
Moyes, K. M., Drackley, J. K., Salak-Johnson, J. L. et al. 2009. Dietary-induced negative energy balance has
minimal effects on innate immunity during a Streptococcus uberis mastitis challenge in dairy cows during
midlactation. Journal of Dairy Science 92: 4301-4316.
Oltenacu, P. A. & Ekesbo, I. 1994. Epidemiological study of clinical mastitis in dairy cattle. Veterinary
Research 25: 208-212.
Ospina, P. A., Nydam, D. V., Stokol, T. et al. 2010. Evaluation of nonesterified fatty acids and beta-
hydroxybutyrate in transition dairy cattle in the northeastern United States: Critical thresholds for prediction of
clinical diseases. Journal of Dairy Science 93: 546-554.
Pelissier, C. L. 1976. Dary cattle breeding problems and their consequences. Theriogenology 6: 575-583.
Perkins, K. H., Vandehaar, M. J., Burton, J. L. et al. 2001. Negative energy balance does not decrease expression
of leukocyte adhesion or antigen-presenting molecules in cattle. Journal of Dairy Science 84: 421-428.
Reist, M., Koller, A., Busato, A. et al. 2000. First ovulation and ketone body status in the early postpartum
period of dairy cows. Theriogenology 54: 685-701.
Romaniukowa, K. 1984. Phagocytosis in the uterus. In: Proceedings of the 10th International Congress on
Animal Reproduction and AI. Urbana, Champaign, IV: XI, 30–33.
Sartori, R., Guardieiro, M. M., SURJUS, R. S. et al. 2013. Metabolic hormones and reproductive function in
cattle. Animal Reproduction 10: 199-205.
Scalia, D., Lacetera, N., Bernabucci, U. et al. 2006. In vitro effects of nonesterified fatty acids on bovine
neutrophils oxidative burst and viability. Journal of Dairy Science 89: 147-154.
Schefers J. M., Weigel, K. A., Rawson, C. L. et al. 2010. Management practices associated with conception
rate and service rate of lactating Holstein cows in large, commercial dairy herds. Journal of Dairy Science 93(4):
1459-1467.
Schneider A., Corrêa, M. N., Butler, W. R. 2013. Short communication: Acute phase proteins in Holstein cows
diagnosed with uterine infection. Research in Veterinary Science 95: 269-271.
Schwegler, E., Schneider, A., Montagner, P. et al. 2013. Predictive value of prepartum serum metabolites for
incidence of clinical and subclinical mastitis in grazing primiparous Holstein cows. Tropical Animal Health
and Production 45: 1549-1555.

60
Seegers, H., Fourichon, C., Beaudeau, F. 2003. Production effects related to mastitis and mastitis economics in
dairy cattle herds. Veterinary Research 34: 475-491.
Senger, P. L. 2005. Pathways to pregnancy and parturition. Current conceptions Inc., Pullan, WA, USA, ed. 2.
Sheldon, I. M., Cronin, J., Goetze, L. et al. 2009. Defining postpartum uterine disease and the mechanisms of
infection and immunity in the female reproductive tract in cattle. Biology of Reproduction 81: 1025-1032.
Sheldon, I. M., Lewis, G. S., Leblanc, S. et al. 2006. Defining postpartum uterine disease in cattle.
Theriogenology 65(8): 1516-1530.
Sheldon, I M., Williams, E. J., Miller, A. N. et al. 2008. Uterine diseases in cattle after parturition. Veterinary
Journal 176(1): 115-121.
Shuster, D. E., Lee, E. K. and Kehrli, M. E. Jr. 1996. Bacterial growth, inflammatory cytokine production, and
neutrophil recruitment during coliform mastitis in cows within ten days after calving, compared with cows at
midlactation. American Journal of Veterinary Research 57: 1569-1575.
Sordillo, L. M. 2009. Current concepts on immunity and mastitis. WCDS Advances in Dairy Technology 21:
111-119.
Ster, C., Loiselle, M. C., Lacasse, P. 2012. Effect of postcalving serum nonesterified fatty acids concentration
on the functionality of bovine immune cells. Journal of Dairy Science 95: 708-717.
Tillard, E., Humblot, P., Faye, B. et al. 2008. Postcalving factors affecting conception risk in Holstein dairy
cows in tropical and sub-tropical conditions. Theriogenology 69: 443-457.
Williams, E. J., Fisher, D. P., Noakes, D. E. et al. 2007. The relationship between uterine pathogen growth
density and ovarian function in the postpartum dairy cow. Theriogenology 68: 549-559.
Youngquist, R. S. & Threlfall, W. R. 2007. Current therapy in large animal theriogenology. WB Saunders,
USA, ed. 2.
Zerbe, H., Schneider, N. Schneider, W. et al. 2000. Altered functional and immunophenotypical properties of
neutrophilic granulocytes in postpartum cows associated with fatty liver. Theriogenology 54(5): 771-786.
Ziguer, E. A., Goulart, M. A. et al. 2007. Alterações clínicas em bovinos de leite na região centro-norte do Rio
Grande do Sul. A Hora Veterinária 156: 1-4.

61
Estresse calórico em vacas leiteiras: efeito sobre o metabolismo e a
qualidade do leite
Vivian Fischer
Marcelo Tempel Stumpf
Alexandre Sausenbach Abreu
Elissa Forgiarini Vizzotto
Cenário – por que o estresse térmico é importante?
Cenários mais pessimistas apontam para um inequívoco aumento nas médias de temperatura
na Terra em até 4°C até 2100 (IPCC, 2007). Os sistemas de produção animal, ao estarem
inseridos e serem diretamente dependentes das condições naturais vigentes, serão afetados
por tais modificações.
As condições climáticas afetam e desafiam o animal a se adaptar às condições ambientais
para manter a homeostase e o bem-estar. Nos trópicos e em condições de pastagem, o gado
é exposto ao sol, além de outros agentes meteorológicos como umidade, chuva e vento
durante várias horas por dia. A intensificação da atividade pecuária, reduzindo o espaço por
animal e modificando o ambiente pode dificultar ou mesmo impedir que animais encontrem
alternativas para se adaptarem a essas condições, tornando-os mais suscetíveis ao estresse,
resultando em mudanças fisiológicas que diminuem o seu desempenho produtivo
(Deitenbach et al., 2008).
Podem se esperar efeitos diretos em animais expostos a altas temperaturas, tais como
alterações comportamentais, como o aumento na ocorrência de interações agonísticas
(Vizzotto, 2014), maior tempo em ócio, redução tempo de pastejo diurno e aumento no
noturno, maior ingestão de água (West, 2003; Fischer et al., 2014); alterações fisiológicas,
como aumento nas frequências respiratória e cardíaca (Mitlöhner et al., 2001; Dalcin, 2013),
escore de ofegação e temperatura retal (McManus et al., 2009; Stumpf, 2014; Fischer ET
AL., 2014) e redução no consumo de alimento (West, 1994; Rhoads et al., 2009).
Mecanismos homeorréticos são acionados de forma a tornar o animal capaz de se adaptar e
se manter em um equilíbrio dinâmico com as novas contingências impostas. O animal
necessita dissipar calor ao meio em busca de manter sua homeostase, ou seja, manter a
temperatura interna dentro dos níveis normais de 38 a 39,5˚C (Stober, 1993). Mecanismos

62
de dissipação de calor requerem energia, a qual pode provir da sua partição em detrimento
da glândula mamária através de alterações hormonais acionadas e controladas pelo eixo
hipotálamo-hipófise-adrenal (Baumgard & Rhoads, 2013).
Os efeitos negativos do estresse térmico são mais evidentes em animais com elevada
produção devido à maior geração de calor metabólico (Kadzere et al., 2002). Animais
zebuínos, normalmente mais tolerantes ao calor, devido às suas adaptações de forma
corporal, superfície de pele, número de glândulas sudoríparas e características do pelame,
sofrem mais intensamente à medida que a sua produtividade aumenta. Entre as estratégias
dos animais para minimizarem o estresse térmico, se destacam a redução da carga térmica,
redução da produção de calor metabólico e aumento da dissipação de calor (Berman, 2012).
Na zona de conforto térmico, em torno de 75% da perda de calor ocorre por radiação,
convecção e/ou condução. No entanto, quando a temperatura ambiente excede a temperatura
crítica superior, o gradiente de temperatura torna-se pequeno para que ocorra resfriamento
por esses meios. Inicia-se então a termólise, especialmente por mecanismos evaporativos
para manter o balanço térmico e a evaporação, por meio da sudorese e/ou respiração,
tornando-se a principal via de dissipação de calor, responsável por 80% da perda de calor
corporal. Em condições de umidade relativa do ar aumentada, há dificuldade na evaporação
e o ambiente torna-se mais estressante para o animal (Collier et al., 2006).
Animais mantidos em ambientes com temperaturas acima da temperatura crítica superior
podem se considerar em estresse térmico, já que dependem de mecanismos de dissipação de
calor. A mediação de tais mecanismos se dá pelo eixo hipotálamo-hipófise-adrenal, o qual é
abastecido de informações provenientes das unidades termorreceptoras e termosensitivas no
sistema nervoso central (Baker, 1989). Uma das primeiras respostas do animal ao ambiente
estressante é a redução no consumo de alimentos (Castanheira, 2009), na busca de reduzir a
produção de calor proveniente de processos digestivos (calor metabólico) e maior consumo
de água, para repor as perdas promovidas pela evaporação. Estima-se aumento de consumo
de 1,2 kg de água para cada 1˚C de aumento na temperatura ambiental mínima (West, 2003).
Em adição, os animais aumentam suas exigências de mantença, necessárias para suportar as
mudanças fisiológicas para dissipação de calor: ofegacão, suor, aumento nas reações
químicas, síntese e mantença da funcionalidade das proteínas de choque térmico (Tomanek
et al., 2010), e aumento na atividade celular induzida pelos níveis elevados de adrenalina

63
(Gaffin & Hubbard, 1996). De acordo com o NRC (2001), estresse médio a severo pode
elevar entre 7 a 25% tais exigências, embora Fox & Tyluki (1998) sugiram aumentos de até
30%. Ocorre vasodilatação das vias sanguíneas e aumento do fluxo de sangue para as zonas
periféricas do animal, para que ocorra resfriamento por contato com o ar mais frio. A
elevação na frequência cardíaca promove aumento nesse fluxo sanguíneo, auxiliando o
processo. Na busca de elevar as perdas de calor por evaporação, os animais elevam também
a frequência respiratória, podendo prevenir o aumento da temperatura retal até índices de
temperatura e umidade (ITU) de 80 (Silanikove et al., 2000). O aumento nas perdas de gás
carbônico em virtude do aumento da frequência respiratória altera a proporção entre as
concentrações de CO2:HCO3 sanguíneos, a qual deve se encontrar por volta de 20:1, e é o
principal sistema de tamponamento do sangue (Kadzere et al., 2002). Para compensar essas
perdas, o CO2 diminui, levando a um quadro de alcalose respiratória e aumento do pH
sanguíneo. A redução no teor de CO2 força os rins a excretarem maior quantidade de
bicarbonato (tamponante), na tentativa de manter a proporção de 20:1 estável (Beatty et al.,
2006). Além disso, a reação CO2 ↔ HCO3 se desloca para a esquerda, para compensar as
perdas de gás carbônico. O menor teor de bicarbonato promove uma subsequente redução no
pH sanguíneo, provocando acidose metabólica compensatória.
Mecanismos de sobrevivência sobrepõem aqueles relacionados à produção de leite. Os
efeitos aditivos do menor consumo de alimentos, maiores exigências de energia para
mantença, possíveis distúrbios metabólicos e maior destinação de energia aos processos de
perda de calor geram as reduções nos níveis produtivos dos animais.
Efeitos do estresse calórico sobre produção de leite
Até recentemente se creditava a redução na produção de leite à diminuição no consumo de
alimento (Baccari, 2001). A redução da produção leiteira varia entre 20 e 40% em animais
severamente estressados (West, 2003; Abreu et al., 2011). Segundo Wheelock et al. (2010),
40 a 50% dessa redução se deve ao menor consumo de alimentos (efeito indireto), sendo o
restante em virtude de outros mecanismos induzidos pelo estresse (efeitos diretos). Além do
menor consumo, ocorre redução de até 35% na eficiência de utilização de energia para fins
produtivos (McDowell et al., 1976).

64
Silanikove et al. (2009) sugerem outro mecanismo de ação para explicar a redução na
produção leiteira. Sob estresse térmico agudo, ocorre aumento na concentração de um
peptídeo derivado da clivagem da β-caseína pela plasmina, o qual bloqueia os canais de
potássio da membrana apical das células epiteliais mamárias, promovendo despolarização
potencial da membrana, reduzindo o metabolismo celular, sua atividade secretória e, por
conseguinte, diminuindo a produção láctea. Os efeitos negativos do estresse térmico sobre a
produção leiteira (Figura 1) dependem do nível de produção antes do estresse (quanto maior
a produtividade, maior a redução na produção) e do estádio de produção (vacas no terço
médio reduzem mais a produção do que vacas ao início ou final da lactação (Bernabucci et
al., 2010).
Figura 1. Variação do consumo e da produção leiteira em vacas estressadas
termicamente (HS) e vacas com alimentação restrita (PF). Fonte: Rhoads et al. (2009)
Efeitos do estresse calórico sobre a composição do leite – características químicas
Vacas mantidas em ambientes com índice de temperatura e umidade (ITU) superiores a 72,
produziram leite com menores teores de proteína, alfa e beta caseínas. Todavia, os efeitos

65
sobre os teores de gordura foram contraditórios (Bernabucci et al., 2002). A principal causa
da redução nos teores de proteína nesse estudo foi a redução nos níveis de αs e β-caseínas,
provavelmente em virtude do menor aporte de energia e proteína. Essas caseínas são ricas
em grupos fosfato e são os componentes acídicos das micelas (Schmidt, 1980).
Reduções no teor de proteína no leite foram descritos em bovinos por Rhoads et al. (2009),
embora não tenham detectado diferenças nos teores de gordura e lactose (Figura 2). Ao
contrário do relatado por Abreu et al. (2011) os quais verificaram que vacas leiteiras sob
estresse térmico severo sem acesso a sombra produziram leite com menores teores de
proteína (2,75 vs 2,99%), porém maiores teores de gordura (3,61 vs 3,40%) e nitrogênio
ureico (23,70 vs 18,81 mg/dL), não encontrando diferença para os teores de lactose (4,37 vs
4,34).
Segundo Sevi & Caroprese (2012), a exposição ao sol em altas temperaturas pode promover
aumento na permeabilidade dos capilares, elevando a quantidade de enzimas proteolíticas no
leite, de forma a reduzir a concentração proteica. Em adição, a redução na oferta e consumo
de pasto nos meses mais quentes, bem como o menor aporte de energia e nitrogênio,
contribuem para os valores encontrados. O citado aumento na permeabilidade dos capilares
também promove aumento nas concentrações de enzimas lipolíticas, resultando em
decréscimo na síntese e alteração no perfil lipídico do leite (Sevi & Caroprese, 2012). Esses
autores observam que a menor taxa de passagem no rúmen aumenta o tempo de exposição
da digesta ao processo de biohidrogenação, de forma que os ácidos graxos insaturados são
convertidos em saturados e têm seus teores reduzidos no leite. O estresse térmico e a redução
na qualidade da forragem em virtude da maior atividade metabólica vegetal atuam em
conjunto diminuindo o consumo (Renna et al., 2011) e a produção de ácido acético no rúmen,
principal precursor da gordura láctea.
Os efeitos negativos do estresse térmico sobre os níveis de lactose no leite são decorrentes
do uso da glicose como fonte de energia ao animal, reduzindo o aporte à glândula mamária
e posterior síntese desse componente, além do efeito direto da queda no consumo de
alimentos. Alterações na concentração de lactose, por sua vez, são mais difíceis de ocorrer,
já que sua queda é acompanhada por menores níveis de produção de leite.

66
Figura 2. Variação da concentração de lactose, proteína e gordura
lácteas provenientes de vacas estressadas termicamente (HS) e vacas
com alimentação restrita (PF). Fonte: Rhoads et al. (2009)
Efeitos do estresse calórico sobre a composição do leite – características físicas
Em trabalhos realizados durante o verão (Mariani et al., 1994; Bernabucci & Calamari, 1998;
Calamari & Mariani, 1999), foram verificados valores de pH mais alto e de acidez titulável
mais baixa em animais em estresse térmico. No entanto, Abreu et al. (2011) relataram
menores valores de pH, crioscopia e densidade e maior valor de acidez titulável em vacas
estressadas termicamente sem acesso a sombra em relação àquelas com acesso a sombra.
Também, os autores encontraram que vacas estressadas termicamente produziram leite com
menor estabilidade (testes do álcool e do tempo de coagulação sob temperatura de 144°C,
Figura 3).

67
12
14
16
18
20
22
24
Produção de Leite (L)
62
64
66
68
70
72
74
76
78
80
Teste do Álcool (%etanol v/v)
0
50
100
150
200
250
300
Tempo de coagulação a 144°C (s)
0
5
10
15
20
25
Acidez titulável (°D)
Figura 3. Valores de produção de leite, estabilidade no teste do álcool e tempo de
coagulação e acidez titulável de vacas com (linha preta) e sem acesso a sombra (linha
vermelha). Fonte: Fischer et al. (2012)
Os efeitos do estresse térmico sobre a estabilidade do leite no teste do álcool podem ser
relacionados à redução no consumo de alimentos dos animais. De fato, a restrição de
consumo per se resultou em redução na estabilidade do leite (Zanela et al., 2006; Abreu et
al., 2011; Stumpf et al., 2013) e épocas do ano de menor oferta de alimentos promovem maior
incidência de leite instável (Ponce & Hernández, 2001; Marques, 2004).
Ademais, distúrbios metabólicos provenientes do estresse térmico, ao alterar de forma
significativa as condições fisiológicas do animal, apresentam grande potencial de reduzir a
estabilidade do leite, conforme relatado por Marques et al. (2011) ao induzirem acidose
metabólica através do fornecimento de sal aniônico em bovinos leiteiros.
Efeitos do estresse calórico sobre o metabolismo
O estresse térmico provoca mudanças na homeostase e na homeorrese das vacas lactantes.
Ocorrem alterações no perfil hormonal caracterizadas principalmente por um declínio e
aumento, respectivamente, naqueles com efeitos anabolizantes e catabolizantes, alterando o
metabolismo pós-absortivo da energia, lipídios e proteínas, com prejuízos à função hepática
e aumento do estresse oxidativo. Ocorre prejuízo à resposta imunológica e ao desempenho

68
reprodutivo. Os efeitos sobre o metabolismo podem ser quantificados pela mensuração das
variáveis fisiológicas, tais como a temperatura corporal, frequência respiratória,
concentrações hormonais e equilíbrio ácido-base e hidroeletrolítico (Bernabucci et al., 2010).
Assim, ocorre diminuição do hormônio do crescimento, catecolaminas e glicocorticoides.
Este estado endócrino reduz os níveis circulantes de tiroxina (T4) e triiodotironina (T3), e, em
consequência, a diminuição da taxa metabólica basal e da produção de calor (Johnson, 1980;
Yousef, 1987).
Vacas em balanço energético negativo decorrente de estresse térmico, apesar de ingerirem
menos, não apresentam reduções na secreção e na sensitividade à insulina (Wheelock et al.,
2010) e podem ter seus níveis de somatotropina reduzidos (Li et al., 2006). Dessa forma, as
vacas se tornam metabolicamente inflexíveis, uma vez que não podem oxidar ácidos graxos
para gerar energia, pois esse processo contribui ao incremento de calor metabólico
(Baumgard & Rhoads, 2013). Não se detectam elevações em ácidos graxos não esterificados
no sangue (Shwartz et al., 2009) e os animais passam a ser dependentes de glicose como
fonte de energia. Como consequência, o aporte de glicose à glândula mamária diminui, com
consequente redução da síntese de lactose, o que diminui a produção de leite.
Vacas sem acesso a sombra, mesmo em condição de estresse térmico leve, apresentaram
maiores valores de sódio, mas valores menores de hematócrito, PCO2, TCO2, HCO3, excesso
de base, e tenderam a apresentar hemoglobina baixa (Abreu, comunicação pessoal). Foram
observadas maiores concentrações de ureia, creatinina e albumina no sangue de vacas
severamente estressadas pelo calor (Abreu, comunicação pessoal). Devido às maiores perdas
renais de HCO3- e perda de saliva, pela redução da ruminação nos animais ofegantes no calor,
as vacas estressadas pelo calor são mais propensas a sofrer acidose ruminal e metabólica.
Estresse térmico em fases não produtivas (novilhas e período seco de vacas)
Vacas que sofreram estresse térmico durante o período seco apresentaram menor produção
leiteira durante 280 dias na lactação subsequente (28,9 vs 33,9 kg/dia), possivelmente devido
à menor proliferação celular observada no pré-parto, não sendo observados efeitos sobre a
apoptose (Tao et al., 2011). Também foi observado menor concentração de proteína láctea
(3,01 vs 2,87%). O estresse térmico durante o período seco reduz o consumo e debilita a
imunidade dos animais (Tal et al., 2013).

69
Em comparação com as vacas estressadas termicamente durante o período seco, os animais
que ficaram sob resfriamento (ventiladores e aspersores) ganharam mais peso antes do parto,
mas perderam mais peso e condição corporal no início da lactação. As vacas com
resfriamento produziram mais leite do que vacas mantidas sob estresse térmico (34,0 vs 27,7
kg/dia), mas os tratamentos não afetaram a composição do leite (Figura 4). Os tratamentos
não influenciaram a insulina circulante e os metabólitos pré-parto, mas vacas sob
resfriamento apresentaram menor concentração plasmática de glicose, aumento dos ácidos
graxos livres, e tenderam a ter menor concentração de insulina no pós-parto em comparação
com vacas estressadas termicamente. O resfriamento das vacas no pré-parto não afetou as
respostas de insulina para o teste de tolerância à glicose e o teste de desafio à insulina antes
do parto e aos 28 após o parto (Tao et al., 2012).
Medidas mitigadoras de estresse térmico
Em termos de medidas de adaptação, é geralmente mais rápido alterar o ambiente para
melhorar o bem-estar, produção e reprodução (West, 2003; Renaudeau et al., 2012). Raças
de origem europeia sofrem mais o estresse térmico devido à sua alta produtividade, reduzindo
o seu limiar de conforto térmico (Silva et al., 2002). Por isso, os criadores brasileiros têm
procurado combinar as características desejáveis das raças europeias e zebuínas, através da
produção de animais mestiços, geralmente usando Holandês e Gir (Girolando). O rebanho
nacional é composto por aproximadamente 95% de animais mestiços (Martinez & Verneque,
2001), representando cerca de 70% da produção de leite (Alvim et al., 2005).
As raças diferentes reagem de forma diferente ao estresse térmico (McManus et al., 2009).
Além disso, os avanços na produção animais em áreas como a nutrição e a reprodução
resultaram em aumento significativo na produção de leite, o que resultou em uma maior taxa
de calor metabólico e, portanto, aumentando a necessidade de dissipação de calor (Silanikove
et al., 2000; Baumgard et al., 2007).
O cruzamento entre as raças Holandês e Gir é usado há décadas para obter animais mais
tolerantes ao calor. A comparação entre animais pertencentes aos grupos genéticos Holandês,
50% Holandês e 50% Gir e 75% holandês e 25% Gir, mostrou que as vacas 50% H 50% G
apresentaram menores frequências respiratórias e cardíacas, temperatura retal e escore de

70
ofegação comparadas ao grupo 100% Holandês, enquanto o grupo 75% H 25% G apresentou
valores intermediários (Stumpf, 2014).
Figura 4. Produção de leite de vacas sob estresse térmico com
(símbolo preto) ou sem resfriamento (símbolo vazado) durante o
período seco. Fonte: Tao et al. (2011)
Criar animais em um ambiente com conforto para proporcionar bem-estar, pode melhorar o
desempenho produtivo do animal. Portanto, para minimizar os efeitos nocivos do clima sobre
os animais em países tropicais é importante para mitigar o efeito negativo das variáveis
climáticas consideradas responsáveis pelo estresse por calor. Nos locais, caracterizado pelo
clima subtropical de altitude com umidade elevada, a velocidade do vento contribui para a
dissipação de calor dos processos de animais por convecção, mas a alta umidade reduz a
capacidade de dissipação de calor do corpo, por meio de evaporação (Azevedo et al., 2009).
O resfriamento evaporativo é a forma principal de perda de calor disponível para os animais
homeotérmicos quando a temperatura ambiente é superior à temperatura do corpo, e este
processo é mais eficiente quando a umidade relativa é baixa (Robinson, 2004).
O uso de sombra serve para atenuar o efeito da radiação solar, diminuindo a temperatura do
corpo (Mitlohner et al., 2001; Kendall et al., 2006; Tucker et al., 2007; Fischer et al., 2014).
No entanto, deve-se prover área em torno de 9 m2 por vaca para evitar competição entre os
animais pela sombra (Schutz et al., 2010).
O provimento de instalações como galpões dotados de aspersores na linha do cocho aliado a
ventiladores, tipo fixo ou tipo móvel, ou galpões com sistema de resfriamento no piso ou nas
camas, são ferramentas nos rebanhos comerciais dos países desenvolvidos. Existem

71
diferenças entre os sistemas quanto à capacidade de reduzir a temperatura corporal e a
frequência respiratória, aumentar o tempo em que a vaca fica deitada e aumentar o consumo
de alimentos. O provimento de conforto térmico durante o período seco exerce efeitos
positivos sobre o sistema imune, desenvolvimento placentário e desenvolvimento do terneiro,
com reflexos positivos sobre a produção de leite na lactação subsequente (Tao et al., 2013).
Referências
Abreu, A.S.; Fischer, V.; Kolling, G. et al. Estresse calórico induzido por privação de acesso à sombra em vacas
Holandesas reduz a produção leiteira e a estabilidade térmica do leite. Proceedings. 2ª Conferencia Internacional
de Leche Inestable, Colonia del Sacramento, Uruguay, 2011.
Alvim, M.J.; Paciullo, D.S.C.; Carvalho, M.M. et al. Sistema de produção de leite com recria de novilhas em
sistemas silvipastoris. 2005. EMBRAPA.
Azevêdo, D.M.R; Alves A.A. Bioclimatologia aplicada à produção de bovinos leiteiros nos trópicos. Embrapa
Meio-Norte, 83 p. Teresina. 2009.
Baker, M. A. Effect of dehydration and rehydration on thermoregulatory sweating in goats. Journal of
Physiology, v. 417, p. 421 – 435, 1989.
Baumgard, L.H.; Rhoads, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev.
Anim. Biosci. v.1, p. 311-333, 2013.
Beatty, D. T. et al. Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat
and humidity. J. Animal Sci., v. 84, p. 972-985, 2006.
Berman, A. From heat tolerance to heat stress relief: An evolution of notions in animal farming. In:
Environmental Physiology of Livestock. Ed. Collier, R.J. & Collier, J.L. 2012. p.1-16.
Bernabucci, U.; Lacetera, N.; Ronchi, B.; Nardone, A. Effects of the hot season on milk protein fractions in
Holstein cows. Anim. Res. v. 51, p. 25-33, 2002.
Bermanbucci, U.; Lacetera, N.; Baumgard, L.H. et al. Metabolic and hormonal acclimation to heat stress in
domesticated ruminants. Animal, v. 4, p. 1167–1183, 2010.
Castanheira, M. Análise multivariada de características que influenciam a tolerância ao calor de equinos, ovinos
e bovinos. 2009. 107 f. Tese (Doutorado em Ciência Animal). Universidade Federal de Goiás, Goiânia, 2009.
Cerutti, W.G.; Bermudes, R.F.; Viégas, J.; Martins, C.M.M. Physiological and productive responses of Holstein
cows in fosterage submitted or not to shading and aspersion in premilking. Rev. Bras. Saúde Prod. Anim., v.
14, p. 406-412, 2013.
Collier, R.J.; Dahl, G.E.; Van Baale, M.J. Major advances associated with environmental effects on dairy cattle.
J. Dairy Sci., v. 89, p. 1244-1253, 2006.
Costa, M.J.R. Ambiência na produção de bovinos de corte a pasto. Anais de Etologia, v. 18, p. 26-42, 2000.
Costa, M.J.R.; Cromberg, V.U. Alguns aspectos a serem considerados para melhorar o bem-estar de animais
em sistemas de pastejo rotacionado. In: Peixoto, A.M.; Moura, J.C.; Faria, V.P. Fundamentos do pastejo
rotacionado. Piracicaba: FEALQ, 1997, p. 327.
Dalcin, V.C. Parâmetros fisiológicos em bovinos leiteiros submetidos ao estresse térmico. Dissertação
(mestrado). PPG Zootecnia, UFRGS. Porto Alegre, 2013. 31 p.
Dietenbach, A.; Floriani, G.S.; Dubois, J.C.L et al. Manual agroflorestal para a Mata Atlântica. Ministério do
Desenvolvimento Agrário. Brasília, 2008.

72
Drackley, J.K. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci., v. 82, p.
2259–2273, 1999.
Fischer, V.; Vizzotto, E.F.; Abreu, A.S. et al. Behavior of lactating dairy cows under mild and severe heat stress
with free access or not to shade. In: Joint Annual Meeting, Kansas, MI, July 2014, Proceedings….
Fox, D.G.; Tylutki, T.P. Accounting for the effects of environment on the nutrient requirements of dairy cattle.
J. Dairy Sci., v. 81, p. 3085-3089, 1998.
Gaffin, S.L.; Hubbard, R. Experimental approaches to therapy and prophylaxis for heat stress and heatstroke.
Wilderness & Environmental Medicine, v. 7, p. 312-334, 1996.
Intergovernmental Panel on Climate Change (IPCC). 4th Assessment Report, 2007. Disponível em: <http://
www.ipcc.ch/publications_and_data/publications_and_data_reports.shtml#2>. Acesso em: 25 março, 2013.
Kadzere, C.T. et al. Heat stress in lactating dairy cows: a review. Livestock Production Science, v. 77, p. 59-
91, 2002.
Li, G.; Ali, I.; Currie, R.W. Insulin induces myocardial protection and Hsp70 localization to plasma
membranes in rat hearts. American Journal of Physiology. Heart and Circulatory Physiology, v. 291, p. 1709-
1721, 2006.
McDowell, R.E. et al. Effects of climate on performance of Holsteins in first lactation. J. Dairy Sci., v. 59, p.
965-973, 1976.
McManus, C.; Paludo, G.R.; Louvandini, H. et al. Heat tolerance in Brazilian sheep: physiological and blood
parameters. Tropical Animal Health and Production, v. 41, p. 95–101, 2009.
Marques, L.T.; Fischer, V.; Zanela, M.B. et al. Produção leiteira, composição do leite e perfil bioquímico
sanguíneo de vacas lactantes sob suplementação com sal aniônico. Rev. Bras. Zoot. v. 40, p. 1088-1094, 2011.
Mitlöhner, F.M. et al. Shade and water misting effects on behavior, physiology, performance, and carcass traits
of heat-stressed feedlot cattle. J. Anim. Sci., v. 79, p. 2327–2335, 2001.
National Research Council. Nutrient Requirement of Dairy Cattle. 7th ed. Washington: National Academic
Press, 2001. 408p.
Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livestock
Production Science, v. 67, p. 1-18, 2000.
Silanikove, N.; Shapiro, F.; Shinder, D. Acute heat stress brings down milk secretion in dairy cows by up-
regulating the activity of milk-borne negative feedback regulatory system. BMC Physiology, v. 9, p. 13-22,
2009.
Renna, M.; Lussiana, C.; Malfatto, V.; Mimosi, A.; Battaglini, L.M. Effect of exposure to heat stress conditions
on milk yield and quality of dairy cows grazing on Alpine pasture. Proceedings, 9th European IFSA
Symposium, Vienna, Austria, July 2011.
Renaudeau, D.; Collin, A.; Yahav, S.; De Basilio, V.; Gourdine, J.L.; Collier, R.J. Adaptation to hot climate
and strategies to alleviate heat stress in livestock production. Animal, v. 6, p. 707-728, 2012.
Rhoads, M.L.; Rhoads, R.P.; Van Baale, M.J. et al. Effects of heat stress and plane of nutrition on lactating
Holstein cows: I. Production, metabolism and aspects of circulating somatotropin. J. Dairy Sci., v. 92, p. 1986–
1997, 2009.
Robinson, N.E. Homeostase e termorregulação. In: Cunningham, J.G. Tratado de fisiologia veterinária. 3ª ed.
Rio de Janeiro: Guanabara Koogan, 2004. p. 550-561.
Sevi, A.; Annichiarico, G.; Albenzio, M. et al. Effects of solar radiation and feeding time on behavior, immune
response and production of lactating ewes under high ambient temperature. J. Dairy Sci., v. 84, p. 629-640,
2001.
Stober, M. Identificação, anamnese, regras básicas da técnica do exame clínico geral. In: Rosemberg, G. Exame
clínico dos bovinos. Rio de Janeiro: Guanabara Koogan, 1993. p. 419.

73
Stumpf, M.T.; Fischer, V.; McManus, C.M. et al. Severe feed restriction increases permeability of mammary
gland cell tight junctions and reduces ethanol stability of milk. Animal, v. 7, p. 1137-1142, 2013.
Tao, S.; Bubolz, J.W.; Amaral, B.C. et al. Effect of heat stress during the dry period on mammary gland
development. J. Dairy Sci., v. 94, p. 5976–5986, 2011.
Tao, S.; Thompson, I.M.; Monteiro, A.P. et al. Effect of cooling heat-stressed dairy cows during the dry period
on insulin response. J. Dairy Sci., v. 95, p. 5035-5046, 2012.
Tao, S.; Dahl, G.E. Invited review: Heat stress effects during late gestation on dry cows and their calves. J.
Dairy Sci., v. 96, p. 4079-4093, 2013.
Tomanek, L. Variation in the heat shock response and its implication for predicting the effect of global climate
change on species’ biogeographical distribution ranges and metabolic costs. Journal of Experimental Biology,
v. 213, p. 971-979, 2010.
Vizzotto, E.F. Comportamento animal e atributos fisiológicos de vacas leiteiras submetidas a ambientes com e
sem sombreamento durante a estação quente. Dissertação (mestrado). PPG Zootecnia, UFRGS, Porto Alegre,
2014.
West, J.W. Interactions of energy and bovine somatotropin with heat stress. J. Dairy Sci., v. 77, p. 2091–2102,
1994.
West, J.W. Physiological effects of heat stress on production and reproduction. In: Tri-State Dairy Nutrition
Conference, Fort Wayne. Proceedings, 2002.
West, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy Sci., v. 86, p. 2131-2144, 2003.
Wheelock, J.B.; Rhoads, R.P.; Van Baale, M.J. et al. Effects of heat stress on energetic metabolism in
lactating Holstein cows. J. Dairy Sci., v. 93, p.

74
Bem-estar de vacas leiteiras: melhor produção com a vaca feliz
Marcelo Cecim
Não há muito tempo atrás falar na preocupação com o conforto e o bem-estar de animais de
produção era um assunto que gerava risadas e desdém, pois acreditava-se que para um animal
ser lucrativo precisava apenas de boa sanidade, boa genética e boa nutrição. Hoje depois de
décadas de seleção, chegamos a um modelo de vaca leiteira que, apesar de produzir mais
leite, tem uma expectativa de vida cada vez menor e uma crescente infertilidade. Em termos
de evolução das espécies, uma raça que vive e se reproduz menos está fadada a extinção. No
entanto, insiste-se em chamar este fenômeno de "melhoramento genético".
É necessário entender que à medida que optamos por animais maiores e mais produtivos,
suas exigências serão maiores também. Muito trabalho tem sido direcionado no sentido de
reconhecer as demandas nutricionais crescentes nestes animais para poder produzir mais.
Infelizmente, isso não tem sido suficiente para mantê-las felizes. Ainda entendemos muito
pouco de necessidades sociais, de adaptação a técnicas de manejo e até mesmo da expectativa
individual de cada vaca. Em relação a isso, sempre vale lembrar o comentário de Louis
Patenaude, proprietário da recordista Gillette Smurf, uma vaca canadense que viveu 16 anos
e produziu mais de 225.000 kg de leite, quem disse: "Ela foi uma vaca que nunca precisou
ser mimada". Longevidade e fertilidade são indiscutivelmente a opinião das vacas sobre a
sua qualidade de vida. Em nossa realidade, o valor econômico da longevidade é enorme. A
maior parte das fazendas leiteiras no sul do Brasil está tentando aumentar o plantel, e cada
descarte involuntário atrasa o processo. Em rebanhos estáveis, cada descarte involuntário
precoce, representa uma novilha que o produtor precisa comprar de si mesmo. Com um custo
de recria da novilha oscilando entre R$ 850,00 e R$ 1.300,00 e um valor de mercado de
venda desta novilha entre R$ 4.500,00 e R$ 6.000,00, cada vez que isso acontece o produtor
faz uma operação comercial com algo próximo a 400% de prejuízo. Em um rebanho em
crescimento como o nosso, a venda de novilhas rende muito mais que a produção de leite.
Além disso, com um descarte menor, as novilhas selecionadas terão sem dúvida melhor
mérito genético. Também a lucratividade individual de uma vaca aumenta com a idade, pois
não só sua produção bruta aumenta até a quarta ou quinta lactação, mas quanto maior o

75
número de lactações menor será o custo proporcional da recria deste animal. O resultado final
é que animais com baixa vida produtiva, dificilmente serão lucrativos, mesmo que produzam
muito leite. Aí temos o impacto do bem-estar sobre a própria sustentabilidade do negócio.
Qual a expectativa de vida produtiva das vacas leiteiras ao redor do mundo? Estima-se que
esteja ao redor de 4,7 lactações no Reino Unido e 5,3 na Nova Zelândia. Um estudo avaliou
69 milhões de lactações de vacas de raça Holandesa em diferentes regiões dos EUA. A
expectativa de vida produtiva variou de 2,73 lactações na região nordeste até 1,94 lactações
na região sudeste. Este resultado assustador indica que se não fosse o advento da sexagem,
em breve haveria rebanhos em que cada vaca produziria menos de uma filha, e a raça entraria
em extinção. Qual a situação no sul do Brasil, onde a linhagem canadense é a mais utilizada?
Estes dados não existem. No entanto é bastante comum encontrarmos rebanhos onde 30 ou
até 35% dos animais em lactação são vacas de primeira cria. Nestes casos, a expectativa de
vida é menor que 3 lactações e é óbvio que a vida dos animais nestes rebanhos não é boa.
Existem diferentes formas para a vaca externar a sua insatisfação com o sistema, as mais
leves estão relacionadas a quadros depressivos e incluem comer menos, ficar menos tempo
deitada, estar mais tempo em pé, ter menor tempo de ruminação, ter cios mais curtos, etc.
Tais sinais, como são de difícil observação acabam passando despercebidos. Ou seja, como
elas não são escutadas, os transtornos tentam falar mais alto, deslocando abomaso, fazendo
cetose, metrite ou uma mastite séria. Quando nada funciona e elas de fato desistem, já que
vaca não foge, a solução é morrer. Um produtor ou técnico quando afirma que vai descartar
esta ou aquela vaca por problema de casco ou úbere, está no mínimo tendo uma atitude
covarde, jogando a culpa na vaca. Na verdade, é a vaca que está dizendo ao dono que não
quer mais viver ali, ou seja, ela o está descartando.
Hoje, na medicina humana, entendemos que prevenção e diagnóstico precoce são as melhores
estratégias de lidar com os nossos problemas de saúde. Em medicina de produção leiteira
temos a mesma situação, apenas chamamos de ambiência e monitoramento. Ambiência são
todos aqueles fatores pelos quais permitimos que as vacas expressem seu comportamento
normal, ou seja, ambiência são as ferramentas do bem-estar.
Se uma vaca pudesse de fato escolher o que fazer durante o dia, o que ela faria? A
distribuição do tempo das atividades é uma ferramenta básica na análise do bem-estar. Mais
especificamente, uma vaca gosta de deitar em cama seca por 12 a 14 horas por dia, a maior

76
parte deste tempo ruminando. Em torno de 5 horas são dedicadas à alimentação; 2 a 3 horas
elas passam bebendo caminhando e socializando; em torno de uma hora ruminando em pé;
até 3 horas por dia ela aceita estar presa ou na ordenha.
Aí iniciam os problemas: vacas pastando em piquetes com baixa oferta precisam comer por
cinco horas ou mais, vacas com calor deitam por menos tempo, duas ordenhas diárias mais o
tempo de espera normalmente passam de 4 horas, falta de cama seca aumenta o ócio em pé.
Este “tempo extra” acaba sendo retirado do tempo deitada, e ruminando, que é a grande
expressão de conforto da vaca. Todos estes fatos comuns no dia a dia de um rebanho são
sinais de que falta conforto e bem-estar para as vacas. No final das contas, prover conforto
para um rebanho leiteiro é um conjunto de atitudes, instalações e manejo que permitam que
o animal expresse o comportamento normal, em outras palavras, que façam o que uma vaca
feliz gostaria de fazer.
Reconhecer bem-estar de um rebanho leiteiro é algo que pode ser feito por qualquer um sem
nenhum equipamento especial, apenas precisa saber o que, quando e como observar,
quantificar os resultados e anotar. Para isso foram criadas classificações que são conhecidas
como escores de monitoramento, usando principalmente os escores de: consumo,
preenchimento de rúmen, uso do tempo, ambiência, interação social, conforto térmico,
consistência e composição de fezes, locomoção etc. Estes comportamentos representam a
opinião da vaca sobre as coisas que lhe são ofertadas. A análise conjunta de todos os escores
representa a análise do próprio sistema produtivo, mas, sob a ótica da vaca. Isto pode ser
usado com muita eficiência para a composição de um mapa de pontos de riscos e seguranças
do sistema, como uma análise FOFA (pontos fortes e oportunidades, pontos fracos e
ameaças). Esta é a estratégia básica das companhias de seguro na definição de risco do cliente
e do prêmio a ser pago. O problema inicia com o tempo que tudo isso consome, e, com a
capacidade do avaliador em entender a inter-relação dos escores e formular soluções práticas
e viáveis. É humanamente impossível acompanhar e anotar atividade de um rebanho 24 horas
por dia. Todo tratador experiente sabe quando uma vaca não está bem, muito embora ela não
esteja doente. Infelizmente esta informação, embora valiosa, é tardia. Se a vaca é reconhecida
como não estando bem, é porque ela não comeu, está deprimida, deu menos leite, andou mais
devagar etc. Tudo isso na verdade são sinais que a vaca “está mal” e o diagnóstico já é tardio,
em pelo menos 24 horas. Aí entram os sistemas de monitoramento de comportamento. São

77
ferramentas pelas quais a vaca pode dizer que não se sente bem, 1 ou 2 dias antes de deixar
de comer, 3 dias antes de diminuir a produção e 4 ou 5 dias antes de ficar doente, permitindo
desta forma uma ação proativa e não reativa ao problema. Entendemos que muito embora as
alterações de comportamento não digam qual é exatamente o problema da vaca, elas avisam
ao tratador que este animal requer uma olhada especial, ou seja, elas permitem um
diagnóstico precoce.
O uso do tempo e do consumo de alimento hoje são considerados os principais indicadores
de que um animal está bem. Portanto, cada vez que uma vaca altera estes parâmetros do seu
comportamento diário, sabemos que alguma coisa não vai bem, e que ela merece uma olhada
mais próxima. Esta é, sem dúvida, é a melhor medida preventiva que podemos usar. Os
pedômetros são usados essencialmente na identificação do cio. As coleiras que associam
deslocamento e ruminação tornam esta identificação bem mais precisa. A coleira brasileira
C-Tech, recentemente lançada no mercado, é a primeira a conter algoritmos relacionados ao
conforto, além da detecção de cio. Este sistema é provido de um acelerômetro que detecta a
posição da cabeça da vaca com 40 leituras por segundo, daí são formados pacotes de
comportamento a cada hora, onde são identificados minutos em: atividade (caminhando,
comendo, montando bebendo), minutos em ruminação e minutos em ócio. Estes pacotes de
dados são enviados automaticamente, 2 ou 3 vezes ao dia, para um programa que compara o
comportamento de cada vaca com ela mesma nos 15 dias anteriores e também com o resto
do rebanho. Cada vez que uma vaca aumenta seu tempo de atividade, diminuindo a
ruminação e o ócio, o programa reconhece como cio. Por outro lado, quando existe um
aumento no ócio com redução da ruminação e também da atividade, o sistema entende como
alteração de comportamento e o animal deve ser examinado. Ao final da ordenha, duas listas
são geradas contendo os brincos dos animais alterados. Se houver internet na fazenda, as
listas podem ser automaticamente enviadas por SMS para o celular do veterinário, por
exemplo. Também o comportamento individual e do lote a cada hora pode ser acompanhado
de qualquer lugar pelo gerente ou proprietário. É importante salientar, que este equipamento
à exceção de cio, não gera diagnósticos, mas reconhece que algo mudou para pior ou melhor
no dia de uma vaca em particular, ou no seu lote.
Classicamente, um bovino dedica seu dia a 3 atividades principais, de 8 horas cada:
ruminação, deslocamento e ócio. Hoje entende-se que a ruminação é uma variável mais

78
sensível que o consumo, pois ruminando menos, a taxa de esvaziamento cai e no dia seguinte
cai o consumo. O tempo de ruminação pode aumentar em consequência do aumento do
conforto, do consumo total e de fibra longa na dieta. Vai diminuir no estresse térmico, nos
quadros de dor, na redução de consumo ou simplesmente na falta de conforto. Os bovinos
preferem ruminar deitados, portanto a qualidade de cama também afeta a ruminação.
As primeiras observações de campo em rebanhos que utilizam esta tecnologia mostram
informações surpreendentes, como por exemplo, a alta frequência de cios curtos (5 ou 6
horas). A queda de minutos de ruminação é normal no dia do parto (até 50%), porém, vacas
que fazem uma boa transição, retornam aos 400-500 minutos de ruminação por dia, já no
quinto dia pós-parto. As vacas que apresentarão mastite, metrite ou cetose até o dia 21,
mostram uma ascendência da curva de ruminação bem mais lenta. As vacas que deslocam
abomaso entre o dia 14 e 21 mostram baixo tempo de ruminação entre os dias 0 e 7 depois
do parto.
Portanto, estamos aprendendo a escutar a opinião das vacas, principalmente durante uma fase
crítica que é a transição. Os próximos estudos devem direcionar-se no acúmulo destes dados
e a construção de uma base dados comportamentais das vacas em sistemas produtivos no
Brasil. Isto permitirá a criação de um programa de predição de desafios e doenças, que levará
ao diagnóstico precoce e, por fim, poderá diminuir o descarte involuntário através do
entendimento das necessidades individuais da vaca.
Referências
Caraviello, D.Z. et al. (2006). Survey on management practices on reproductive performance of dairy cattle
on large US commercial farms. J. Dairy Sci 89:4723-4735.
Cook, N. (2007). The dual roles of cow comfort in the ‘get lame – stay lame’ hypothesis. Sixth International
Dairy Housing Conference Proceedings16-18 June 2007, (Minneapolis, Minnesota, USA).
Cook, N., Norlund, K. (2009). The influence of the environment on dairy cow behavior, claw health and herd
lameness dynamics. The Veterinary Journal 179 (3):360-369
Haley, D.B., Rushen J. (2000). Behavioural indicators of cow comfort: activity and resting behaviour of dairy
cows in two types of housing. Canadian Journal of Animal Science 80(2): 257-263.
Hare, E., Norman, H.D., Wright, J.R. (2006). Survival rates and productive herd life of dairy cattle in the
United States J. Dairy Sci. 89:3713–3720.
Weary, D.M. & Tucker, C. (2003). The science of cow comfort. Proceedings of the Joint Meeting of the
Ontario Agri Business Association and the Ontario Association of Bovine Practitioners, Guelph, Ont., April,
2003.

79
Marcadores bioquímicos sanguíneos en el diagnóstico y control de
trastornos metabólicos en vacas lecheras
Fernando Wittwer
Enfermedades de la producción y estrés metabólico en vacas
La intensificación de los sistemas productivos junto a la selección genética de los animales
ha incrementado la producción animal. Paralelamente, se han impuesto mayores exigencias
metabólicas a los animales predisponiéndolos a desarrollar las enfermedades de la
producción. Éstas se producen debido a un desequilibrio entre los ingresos, circulación y
egresos de uno o más metabolitos en el organismo, alejando sus concentraciones de los
límites fisiológicos. En estas circunstancias se desarrollan alteraciones bioquímicas y
fisiológicas que inicialmente condicionan mermas productivas y de fertilidad en el rebaño
que culminan en trastornos clínicos e incluso la muerte de animales. Para prevenirlas es
fundamental mantener el equilibrio entre la cantidad de un nutriente que ingresa, es
absorbida, circula en la sangre, es depositada en los compartimientos u órganos de reserva y
egresa por conceptos de mantención y producción.
Los trastornos metabólicos en los rumiantes son en su mayoría relacionados con
desequilibrios nutricionales, incluidos las carencias nutricionales simples producto de mezcla
incompleta de la dieta o un manejo alimentario inadecuados, o bien por problemas más
complejos asociados a las interacciones entre la nutrición, el ambiente y el manejo (Cuadro
1). Su presentación es más frecuente en animales manejados en condiciones de pastoreo,
debido a la elevada variación en la disposición y contenido de nutrientes de los pastos
utilizados como forrajes, asociadas a características de suelo, composición botánica y estado
de desarrollo de los pastos y sus cambios estacionales y diarios. Los desequilibrios
nutricionales afectan a un grupo de animales en un rebaño, generalmente los
metabólicamente más susceptibles, que corresponden a los mayormente exigidos desde el
punto de vista productivo, vale decir vacas en el período de transición, ovejas al final de
gestación, animales en crecimiento, los que comúnmente cursan inicialmente con una
alteración en su salud de tipo subclínica o inaparente. Es en esta condición que se requiere
de un método de diagnóstico o evaluación de balance metabólico oportuno, antes que la
producción de los animales se vea afectada.

80
Cuadro 1. Alteraciones metabólicas en vacas
Carencia o disfunción Alteraciones metabólicas
Carencia de energía BEN, cetosis o acetonemia tipo 1 y 2, hígado graso
Desbalance de proteínas Desnutrición proteica, asincronía ruminal RDP/Eº
Disfunción ruminal Acidosis ruminal sub aguda “SARA”, acidosis láctica,
desvío a la izquierda del abomaso
Disfunción mineral Hipocalcemia, paresia puerperal, tetania hipomagnesémica
Carencia mineral Mg, P, Na, Se, Cu, Zn, Fe, Co, I
El período de transición de la vaca lechera (3 semanas preparto a 3 semanas posparto)
constituye el de mayor exigencia para mantener su homeostasia, producto de los cambios
fisiológicos, nutricionales, metabólicos e inmunes que se presentan en las 6 semanas
alrededor del parto. A ello hay que asociar los cambios de manejo (reagrupamiento,
ambientes diferentes) y alimentación (dietas preparto, lactancia) propios del fin de la
gestación, parto e inicio de la lactancia. En esta situación la capacidad homeostática se ve
sobrepasada produciéndose en la vaca el estrés metabólico, definido como la “incapacidad
de adaptación fisiológica al rápido crecimiento fetal, parto y alta demanda de energía para
lactancia, con consecuente alteración en la utilización de nutrientes esenciales, favoreciendo
la presentación de trastornos metabólicos, procesos inflamatorios y el estrés oxidativo”
(Figura 1).
Figura 1. Causa y consecuencias del estrés metabólico en vacas en transición
(Adaptado de Sordillo y Mavangira, 2014)

81
El estrés metabólico genera significativas pérdidas económicas en la industria ganadera,
limitando la productividad de las vacas y originando enfermedades. Al respecto, se describe
que el 30 al 50% de las vacas cursan con una enfermedad durante el período de transición
(Figura 2), constituyendo con ello preocupación no solo productiva sino también asociada al
bienestar animal. De allí que la determinación de la concentración de analitos cuyas
concentraciones son influidas por el estatus metabólico de los nutrientes en el organismo, se
realiza frecuentemente en muestras de sangre, leche y orina de vacas lecheras. Su
determinación permite evaluar la condición o balance metabólico nutricional de los animales
e identificar tempranamente los trastornos metabólicos que afectan de forma clínica o
subclínica los rebaños.
Figura 2. Enfermedades asociadas al estrés metabólico de vacas lecheras en sistemas
confinado de USA y Europa (Sordillo y Mavangira, 2014) y pastoril de Chile
(Sepúlveda-Varas et al . 2015)
Perfil metabólico
Análisis bioquímicos clínicos para individuos se utilizan comúnmente en las prácticas
veterinarias, es así que exámenes de campo o al lado de la vaca, como las pruebas rápidas
para la cetosis, hipocalcemia puerperal, pH ruminal, inmunoglobulinas en terneros,
constituyen un apoyo del laboratorio al diagnóstico de enfermedades metabólicas y
0 5 10 15 20 25 30 35
Distocia
Paresia puerperal
Hipocalcemia
Cetosis
DAI
Ret. Placenta
Metritis
Quiste ovárico
Mastitis clínica
Cojera
Casos (%)
Sistema confinado Sistema pastoril

82
alteraciones de salud de los animales. Sin embargo, hoy en día la medicina individual en
animales de producción tiene un uso limitado, mientras que los procedimientos de
diagnóstico de rebaño constituyen una herramienta útil para monitorear, diagnosticar y
controlar enfermedades que afectan a grupos de individuos.
Los marcadores bioquímicos corresponden a analitos que pueden ser cuantificados en
muestras de tejidos como sangre o fluidos corporales como leche u orina de un animal o en
un grupo en el rebaño, y que definen el grado de equilibrio metabólico logrado mediante la
“homeorresis”, que corresponde al mecanismo que regula y coordina los cambios en los
procesos metabólicos de tejidos del animal que son requeridos para sostener una condición o
carga fisiológica, como sucede en el período de transición, especialmente al inicio de la
producción láctea. Basado en este concepto y la idea que “lo que nos es medido difícilmente
puede ser corregido”, en los años 1970 se comenzó a utilizar en Europa el "perfil metabólico"
(PM), técnica que se ha ido adaptado a diferentes sistemas y especies productivas,
incorporando nuevos marcadores o formas de aproximación, acorde con las realidades
locales y los avances científicos y tecnológicos.
Un PM es definido como un conjunto de análisis bioquímicos realizados en un momento
definido con el propósito de monitorear la salud metabólica del rebaño y contribuir a
establecer las alteraciones metabólicas y enfermedades que afectan la producción. Es un
examen complementario utilizado en la evaluación y diagnóstico de las “enfermedades de la
producción” que se analiza, en uno o más grupos de animales representativos del rebaño, en
los que se determinan bio-marcadores indicadores del balance metabólico nutricional y del
estrés metabólico. Los resultados obtenidos se comparan con intervalos de referencia
poblacionales (IR) o umbrales críticos (cut-off), indicando así el grado de adecuación de las
principales vías metabólicas relacionadas con energía, proteína y minerales, la funcionalidad
de órganos vitales para la producción como el hígado o el grado de salud o bienestar de los
animales. Su mayor utilidad se ha centrado en el estudio de los desbalances metabólicos
nutricionales que se presentan en las vacas lecheras durante el período de transición e inicio
de la lactancia. Su empleo se ha promovido bajo dos sistemas, a) evaluación a nivel de
individuos como indicador para estimar el riesgo de presentación de trastornos metabólicos
e inflamatorios en las vacas en transición y b) a nivel de rebaño para hacer una prospección

83
de los desbalances metabólicos asociados a una inadecuada alimentación o manejo en lotes
de vacas agrupadas acorde a su condición fisiológica, productiva o sanitaria.
Estimación de riesgo
Este sistema emplea la determinación de bio-marcadores o analitos muy definidos durante el
período de transición de vacas lecheras con el propósito de establecer el riesgo que individuos
de un rebaño cursen posteriormente con enfermedades específicas. Los primeros trabajos en
esta línea fueron realizados por Sommer en Alemania quien describió la metafilaxis como
parte de la medicina preventiva de rebaños al demostrar que un diagnóstico temprano basado
en las determinaciones séricas de colesterol y AST en vacas preparto, permite identificar
vacas en riesgo, las que al ser tratadas preventivamente disminuye en ellas la presentación de
enfermedades en el posparto (Cuadro 2). Bajo este enfoque, se determina en algunos animales
la concentración sanguínea de un analito en un período definido y sus resultados se comparan
con un umbral crítico predefinido. Las vacas con resultados sobre o bajo los puntos de corte
definen el mayor riesgo que cursen posteriormente con una patología definida. Considerando
que la mayor presentación de trastornos metabólicos, como cetosis y desvío del abomaso e
inflamatorios como mastitis y metritis es al final del período de transición e inicio de la
lactancia, la estrategia de muestreo se concentra previo a su presentación por ello la mayor
carga de muestras se ubica en el preparto o inmediatamente después del parto.
Cuadro 2. Efecto de una metafilaxis basada en la determinación de colesterol y AST
séricos (hipocolesterolemia o AST aumentado) en la presentación de enfermedades en el
posparto de vacas (adaptado de Sommer, 1975)
Enfermedad Tratadas (n=202) Control (n=283)
Retención de placenta 5% 9%
Enfermedad metabólica 2% 12%
Endometritis 8% 25%
Disfunción ovárica 11% 19%
Mastitis 1% 8%
Esta técnica se ha empleado mayormente para definir las vacas que cursan con balance
energético negativo, BEN, y su mayor riesgo de enfermar en el posparto; con dicho propósito
se utiliza la determinación de ácidos grasos no esterificados, NEFA y ßOH-butirato (BHB)
en sangre, de igual manera se ha utilizado las determinaciones de colesterol sérico preparto

84
y posparto y de calcio sérico. Todas ellas se asocian con incremento en las presentaciones de
metritis y otras enfermedades del posparto. (Cuadro 3).
Cuadro 3. Asociaciones entre concentraciones séricas de ácidos grasos libres, (NEFA); ß-hidroxi-butirato
(BHB) y calcio (Ca), de vacas en transición con el riesgo de presentación de enfermedades al posparto
Analito Punto de corte
(mmol/L)
Días en relación
al parto
Enfermedad Riesgo Referencia
NEFA 1,1 -21 a -3 K+DA+RP+M 2,1 Van Saun (2006)
NEFA 0,5 3 a 21 K+DA+RP+M 4,2 Van Saun (2006)
NEFA 0,3 -14 a -3 K+DA+RP+M 1,6 Ospina et al. (2013)
NEFA 0,6 3 a 14 K+DA+RP+M 1,9 Ospina et al. (2013)
BHB 1,0 3 a 14 K+DA+RP+M 3,1 Ospina et al. (2013)
BHB 1,2 1 a 7 DA 8,0 LeBlanc et al. (2005)
BHB 1,2 1 a 7 M 3,3 Duffield et al. (2009)
BHB 1,0 3 a 14 K 2,8 Ospina et al. (2010)
Calcio 2,0 21 a 21 K+DA+RP+M 4,0 Van Saun (2006) *K= cetosis; DA= desvío de abomaso; RP= retención de placenta; M= metritis
En esta línea destacan los trabajos realizados por Ospina en Cornell-USA valorando BEN, a
nivel de individuos, en base al aumento de las concentraciones séricas de NEFA pre y
posparto y BHB posparto y su relación con la presentación de enfermedades en el posparto
(cetosis subclínica y clínica, desvío de abomaso, retención de placenta y metritis) (Cuadro
4). Se ha señalado que vacas con concentraciones de NEFA preparto de >0,3 y posparto
>0,7 mmol/L o de BHB posparto >1,0 mmol/L tienen una menor probabilidad de preñez y
su producción de leche se verá disminuida
Cuadro 4. Concentraciones séricas pre y posparto de ácidos grasos libres (NEFA) y ß-hidroxi-butirato (BHB)
como predictores de riesgo de enfermedades en el período de transición en vacas lecheras (adaptado de
Ospina et al., 2010)
Enfermedad
NEFA (mmol/L)
Preparto (-14 a -2 días) Posparto (3 a 14 días)
Punto de corte Riesgo Punto de corte Riesgo
Desvío de abomaso 0,3 2,0 0,7 9,7
Cetosis 0,3 1,8 0,6 5,0
Retención de placenta 0,4 2,2 - -
Metritis - - 0,4 17,0
Otras 0,3 1,8 0,6 4,4
BHB (mmol/L) Posparto (3 a 21 días)
Desvío de abomaso 1,0 6,9
Cetosis 1,0 4,9
Metritis 0,7 2,3
Otras 1,0 4,4

85
Los últimos trabajos en esta línea buscan proyectar los resultados de individuos a la salud del
rebaño, definiendo la asociación entre el porcentaje de vacas que en el período de transición
presentan valores alterados de un analito, con trastornos en la salud, producción o fertilidad
en el período de lactancia posterior. De esta forma constituye una alarma de futuras pérdidas
esperables de mantenerse dicha condición (Cuadro 5).
Cuadro 5. Asociación entre el porcentaje de vacas con concentraciones séricas aumentadas de NEFA y BHB
en el período de transición de vacas lecheras con pérdidas productivas y de fertilidad posterior
Porcentaje
de vacas
sobre punto
de corte
Analito
(mmol/L)
Días de
muestreo
referido al parto
Alteración Magnitud
≥ 15* NEFA ≥0,3 -3 a -14 Cetosis o DA 3,6 veces
≥ 15* NEFA ≥0,3 -3 a -14 Prod leche 305 días -282 kg/vaca/rebaño
≥ 15* NEFA ≥ 0,6 +3 a +14 Prod leche 305 días -288 kg/vaca/rebaño
≥15* BHB ≥ 1,2 +3 a +14 Prod leche 305 días -534 kg/vaca/rebaño
≥15* NEFA ≥0,3 -3 a -14 % preñez 70 días -1,2 veces
≥15* NEFA ≥ 0,7 +3 a +14 % preñez 70 días -0,9 veces
≥30** NEFA≥ 0,5 -1 a -7 Prod leche 1º control -3 kg/d/vaca
≥15** BHB ≥0,8 -1 a -7 Prod leche 1º control -4,4 kg/d/vaca
≥30** NEFA ≥ 1,0 +1 a +7 Preñez 1º servicio 0,6 Odds Ratio Datos de: *Ospina et al. (2013); **Chapinal et al. (2012)
El número de animales a muestrear varía acorde la precisión y exactitud deseada y de la
técnica analítica usada, la prevalencia esperada y el número de animales en riesgo. Así para
NEFA y BHB en un rebaño de 1000 vacas con 3 a 4 partos diarios, que tendría ±35 vacas en
riesgo, se deberían muestrear 14 vacas (con IC de 75% y error de ±10). Actualmente
aconsejan usar esta técnica determinando cada 2 semanas BHB en 20 vacas de 3 a 14 días
posparto (ejemplo de rebaño de 1000 vacas) y en caso de haber 15% a 40 % de positivas
(BHB ≥1,2 mmol/L), controlar todas las vacas en riesgo dos veces a la semana y tratar las
positivas con propileno-glicol, si la prevalencia es >40% se deben tratar todas las vacas en
riesgo desde el día 3 de lactancia por 5 días y continuar el monitoreo bisemanal.
Investigadores italianos (Bertoni et al 2013) enfatizan la importancia de centrar el uso de los
marcadores sanguíneos al periodo de transición. Para ello describen la utilidad de evaluar los
procesos inflamatorios que se presentan alrededor del parto mediante índices compuestos
basados en varios marcadores que denominan “Índice de funcionalidad hepática (LFI)”. Este

86
índice se obtiene de establecer los cambios en las concentraciones séricas de albumina,
colesterol y bilirrubina entre los días 3 a 28 posparto, para luego compararlo con un estándar
definido, obteniendo con ello una herramienta que permite medir la magnitud de la
inflamación en el periparto y su efecto en la producción, fertilidad y salud posterior de la
vaca.
Prospección diagnóstica
Constituye el enfoque tradicional de los PM descrito por Payne en los 1970 en UK, siendo a
la fecha la forma más utilizada en la evaluación de la condición metabólica de los rebaños
lecheros en Chile. En este examen se determinan varios analitos en muestras de sangre
obtenidas de uno o más grupos de vacas seleccionadas del rebaño y cuyos resultados se
comparan con sus respectivos IR. Se emplea como examen de aproximación diagnóstica para
evaluar el balance metabólico-nutricional e identificar los potenciales factores responsables
por la presentación de enfermedades. Este enfoque permite mayor flexibilidad para los
períodos de muestreo, siendo su limitante solo el costo de los análisis, situación superada en
la actualidad con el empleo de equipos de química clínica automatizados para análisis de
multi-analitos en corto tiempo en lotes de muestras y con bajo consumo de reactivos y costo.
El PM evalúa el balance o equilibrio metabólico, así como los cambios que se producen
durante el período de transición en la vaca, o de otros períodos críticos en grupos de animales.
La prueba tiene un enfoque amplio que incluye analitos que reflejan el balance de energía
(NEFA y BHB, colesterol), proteínas (urea y albúmina) y macro y micro minerales (Ca, P,
Mg, Na, Cu, Zn, Se). El panel de análisis es flexible pudiendo reducirse o ampliarse según
las circunstancias. Habitualmente se incluyen analitos para evaluar la condición de salud
hepática (AST, GMD, GGT) y actualmente se está avanzando en la incluir analitos asociados
a inflamación y bienestar animal (haptoglobina, globulinas).
La realización de un PM en un rebaño involucra cinco pasos: a) definir el objetivo que motiva
realizar el examen; b) definir los exámenes a solicitar, el o los grupos a ser muestreados y
obtener las muestras de los animales seleccionados; c) ejecutar los análisis requeridos; d)
obtener los resultados individuales y del grupo y comparar con IR y e) interpretar los
resultados.

87
Objetivo
El perfil metabólico fue diseñado con el propósito de monitorear la salud metabólica del
rebaño, identificar vacas metabólicamente superiores y contribuir a determinar las
alteraciones metabólicas y en general las enfermedades que afectan la producción.
Actualmente se considera que su objetivo es “obtener lo antes posible la opinión de un grupo
de animales sobre su condición metabólica nutricional y sanitaria”. Basado en este concepto
teórico general un PM está indicado cuando se pretende: a) evaluar y controlar el balance
metabólico de un grupo de animales acorde a sus ingresos (nutrición), circulación (capacidad
de movilización y regulación interna) y egresos (producción); b) diagnosticar la presencia de
trastornos metabólicos en un rebaño; c) establecer la presencia o ausencia de un factor
asociado a la presentación de enfermedades en el rebaño y d) Servir de instrumento de
evaluación metabólica en grupos de animales.
El PM no es un examen de evaluación de una dieta, sino como su nombre lo indica, de
evaluación del estado metabólico de un grupo de animales de un rebaño en respuesta a su
dieta. Se debe considerar que su correcta aplicación está determinada por la adecuada
selección de los animales muestreados y que es esencial integrar los resultados del PM con
la información obtenida a partir del análisis de los registros prediales. Bajo estas condiciones
es factible evaluar el estado nutricional del rebaño, detectar en forma temprana alteraciones
en su salud, identificar riesgos potenciales para la presentación de enfermedades y analizar
sus posibles causas orientando al médico veterinario en sus decisiones.
Considerando que el PM ha sido más utilizado en rebaños lecheros bovinos, los antecedentes
entregados a continuación hacen referencia mayormente a este tipo de animales,
empleándose en:
• Control del balance metabólico nutricional sanitario del rebaño.
• Sospecha de la presencia de trastorno metabólico de energía/proteínas en un grupo de
animales.
• Diagnóstico o evaluación de carencias minerales.
• Investigación de problemas de fertilidad, volumen o calidad de la producción de leche,
ganancia de peso en animales de carne.
• Evaluación de respuesta a intervenciones nutricionales.

88
Muestra
La muestra debe ser obtenida de grupos e individuos “representativos” de la población de
animales del rebaño. Es fundamental agrupar a los individuos en base a los factores
fisiológicos que afectan la concentración de los analitos a ser medidos, minimizando al
máximo las causas de variación de origen no nutricional. La selección de los animales acorde
con su etapa productiva permite observar tendencias, al asociar las concentraciones de los
componentes sanguíneos con la producción y comparar entre grupos en distintos estados
fisiológicos. El momento más apropiado para obtener las muestras es cuando los mecanismos
homeostáticos se encuentran fisiológicamente desafiados e inefectivos, siendo este el período
de transición que es coincidente con la mayor incidencia de enfermedades de la producción.
Por ello los grupos más analizados en un rebaño lechero son: a) vacas en transición pre-parto:
3 a 21 días preparto, idealmente 3 a 14 días; b) vacas en transición pos-parto: 3 a 30 días en
lactancia, idealmente 7 a 21 días y c) vacas en inicio de lactancia: 45 a 80 días de lactancia,
idealmente 50 a 70 días. De ellos con mayor frecuencia los veterinarios remiten muestras de
los grupos 1 y/o 2.
De cada grupo, independiente del tamaño, se seleccionan 6 a 14 individuos, comúnmente 8
vacas, de las cuales se obtendrán las muestras a analizar. En lecherías de gran tamaño se
recomienda sub dividir los grupos o bien obtener un número mayor de muestras por grupo.
Por ejemplo, con un lote de 200 vacas en lactancia se puede dividir en dos sub grupos por
producción o edad. De cada animal seleccionado se obtienen las muestras requeridas acorde
con los exámenes a solicitar. Habitualmente corresponden a muestras de sangre (±5 mL con
heparina y 5 mL para suero), las que deben ser obtenidas idealmente posterior al ordeño de
la mañana ya que las concentraciones de algunos metabolitos como BHB, urea, Pi varían
durante el día (Figura 3). Cuando el objetivo del PM es diagnosticar una enfermedad
subclínica, los animales seleccionados deben ser aquellos que se encuentran en riesgo de
presentar la enfermedad y aún no presentan signos clínicos. Para el diagnóstico de
enfermedades en que se presentan individuos con signos clínicos se requiere seleccionar
individuos que estén cursando con la enfermedad, pudiendo en este caso ser sólo 2 o 3
animales.
Junto a las muestras se debe completar una solicitud de examen que debe contener
información general del rebaño, motivo del examen, grupos de animales incluidos,
89
información de las vacas (producción, días respecto a parto, condición corporal, su manejo y
alimentación) y los análisis requeridos.
Figura 3. Variación diaria de las concentraciones plasmáticas (x ± EE) de ß-hidroxi-butirato, urea,
fosfato inorgánico (Pi) y triglicéridos en vacas lecheras a pastoreo (o) y suplementadas con
concentrado energético (■). (Noro et al., 2011)
Análisis
Los análisis que se realizan en el laboratorio cuantifican en las muestras los valores de los
analitos preseleccionados. La selección de los marcadores bioquímicos dependerá de los
requerimientos diagnósticos, las posibilidades de laboratorio, las muestras obtenidas y los
costos involucrados. Los analitos mayormente utilizados se presentan a continuación:
Energía (BHB, NEFA, glucosa, colesterol)
El BHB es un buen marcador de respuesta rápida del grado de síntesis de cuerpos cetónicos
asociado a la movilización de lípidos producto del BEN; es un analito de alta sensibilidad,

90
especialmente en vacas en lactancia, pero con limitada especificidad por la absorción de
butirato ruminal. El NEFA también es un marcador de respuesta rápida y específico para
movilización lipídica, siendo más sensible en vacas preparto, pero de costo analítico alto. Sus
concentraciones aumentan notoriamente con la movilización grasa de inicio de lactancia por
lo que se debe tener IR para pre y posparto. La glucemia es un marcador rápido del balance
de energía, pero de baja sensibilidad limitando su uso.
Junto a la obtención de la muestra se recomienda obtener la condición corporal (CC) de los
animales, que es un buen indicador de respuesta lenta del balance de energía, por lo que su
inclusión como analito en los resultados del PM amplía su utilidad. La colesterolemia es un
indicador de consumo de la ración y de aporte de fibra en la dieta, por lo que su
disminución se asocia a baja ingesta. Su concentración aumenta notoriamente con el
aumento de consumo en el posparto requiriéndose IR para pre y posparto.
Proteínas (urea y albúmina)
La uremia es un marcador de respuesta rápida, sensible y específico de la sincronía de la
proteína degradable (RDP) con la energía disponibles en el rumen (RDP:Eº). Su mayor
utilidad es en el diagnóstico de asincronía ruminal producto de una ingesta elevada de RDP
o escasa en energía, situaciones en que incrementa su concentración en sangre y asociado a
ello en la leche.
La albuminemia es un marcador de respuesta lenta (2 semanas) frente a una disminución en
la síntesis hepática de albúmina producto de carencias en la dieta o un desgaste por sostener
altas producciones lácteas. Su sensibilidad es limitada y su especificidad baja ya que otras
alteraciones orgánicas como la disfunción hepática cursan con hipoalbuminemia.
Minerales (Ca, Pi, Mg, Na, Cu, Zn, Se)
La calcemia es un analito de utilidad sólo frente a cuadros clínicos o subclínicos de
hipocalcemia, su intenso control hormonal tiende a mantener estable su concentración
sanguínea por lo que su sensibilidad para detectar desbalances nutricionales es baja, si bien
últimamente se ha descrito la importancia de controlar las hipocalcemias (Ca <2,0 mmol/L)
en el periodo de transición por su asociación con procesos inflamatorios al inicio de lactancia.
La fosfatemia es un marcador sensible y de respuesta rápida frente a situaciones de

91
desbalances nutricionales de P, siendo su especificidad limitada; a su vez, en muestras
antiguas o mantenidas en altas temperaturas sus concentraciones plasmáticas incrementan.
La magnesemia es un marcador sensible, específico y de respuesta rápida del balance
metabólico nutricional de Mg. La natremia es un analito de utilidad frente a desbalances
severos del Na, su control hormonal regula su concentración sanguínea, siendo por ello baja
su sensibilidad para detectar carencias nutricionales. La cupremia y zinquemia son
marcadores de respuesta lenta y de sensibilidad media frente a carencias de Cu y Zn además
este último es de baja especificidad. La determinación de la actividad sanguínea de GPx es
un marcador sensible y de respuesta lenta (1 mes) del balance metabólico nutricional de Se.
Salud y bienestar animal (globulinas, AST, GDH, GGT, hemoglobina, haptoglobina)
La globulinemia es un indicador de la presencia de cuadros infecciosos en el rebaño,
encontrándose aumentada en animales con mastitis, metritis y pododermatitis, entre otras
causas. Las actividades sanguíneas de las enzimas aspartato transaminasa (AST), glutamato
deshidrogenasa (GDH) y gama-glutamil transferasa (GGT) son marcadores de sensibilidad
moderada de daño hepático en cuyo caso se encuentran aumentadas. La hemoglobinemia es
un indicador sensible del estado general de salud de los animales y por ende de su bienestar,
ya que cuadros de anemia se presentan en animales con carencias nutricionales crónica
(proteínas, Fe, Cu, Co), cuadros tóxicos (exceso de brassicas) o alteraciones orgánicas
(hepatopatías, infecciones crónicas). Actualmente se plantea la determinación de proteínas
de fase aguda como indicadores de bienestar animal, fundamentalmente la haptoglobina en
bovinos, ya que sus concentraciones aumentan fuertemente en respuesta a inflamación
independiente de su origen, infeccioso o traumático.
Informe de resultados
Para cada grupo de animales se construye un cuadro que incluye para cada uno de los analitos
evaluados los valores obtenidos de cada vaca seguido de los resultados del grupo que
corresponden a la media (X), desviación estándar (DE), valores de “H” y “CD” y los
porcentajes de animales sobre y bajo los IR. El valor “H” representa la diferencia entre el
valor promedio del grupo con el promedio de referencia, expresado en DE. Vale decir, indica

92
la diferencia entre la media del grupo en estudio con respecto a la media de referencia
expresado en desvíos estándar, de acuerdo a la siguiente fórmula:
H= ( x del grupo – x de referencia) / DE de referencia
Cuadro 5. Marcadores bioquímicos sanguíneos utilizados en perfiles metabólicos
Variable M Intervalo d referencia Valoración T
Energía
Condición corporal
(1 a 5)
- 2,5 a 4,0 puntos
Según estado fisiológico o = acumulación o movilización de
reservas grasas
C
ß-OH-butirato Su Preparto: < 0,5 mmol/L
Lactancia: < 1,0 mmol/L = mayor síntesis de cuerpos cetónicos por
carencia de energía
A
NEFA
Su Preparto <0,4 mmol/L
Lactancia < 0,6 mmol/L = movilización grasa por carencia de
energía
A
Colesterol
Su Preparto: 1,7 a 4,3 mmol/L
Lactancia: 2,7 a 5,3 mmol/L = carencia de energía – fibra
= exceso de grasa, alteración hepática.
C
Proteínas
Albúmina
Su 30 – 41 g/L = Ingesta o síntesis hepática disminuida C
Urea
Su 2,5 a 7,0 mmol/L = Ingesta limitada de RDP
= Asincronía ruminal de RDP/energía
(exceso RDP, falta de energía)
A
Mineral
Calcio
Su 2,0 – 2,6 mmol/L Paresia hipocalcémica
(sensibilidad baja)
A
Fosfato
Su 1,1 – 2,3 mmol/L Deficiencia de P
(especificidad baja)
C
Magnesio
Su 0,7 – 1,1 mmol/L Hipomagnesemia A
Sodio
Su 134 – 154 mmol/L Carencia de Na C
Cobre Pl 10 – 22 µmol/L Carencia de Cu C
Zinc Su 8 – 24 µmol/L Carencia de Zn C
Selenio (GPx) Sa >130 U/g Hb Carencia de Se C
Salud y bienestar animal
Hemoglobina Sa 90 -125 g/L = Anemia C
AST Su < 110 U/L = Daño hepatocelular o muscular A
GDH Su < 30 U/L = Daño hepato-celular A
GGT Su < 40 U/L = Daño hepato-canalicular C
Globulinas Su < 50 g/L = Infección (bienestar animal) C
Haptoglobina Su - = Inflamación (bienestar animal) A
M: muestra, Su= suero, Pl=plasma, Sa=sangre; T= tiempo de respuesta: A= agudo; C= crónico

93
El “CD” o coeficiente de dispersión, compara la magnitud de la varianza de los valores
individuales del grupo con relación a la varianza de la población de referencia. En el Cuadro
6 se presenta como ejemplo un informe de resultado de un PM realizado a un grupo de 8
vacas en el cual se entregan los resultados de 9 analitos, además de la información de su
producción lechera y la condición corporal (CC).
Cuadro 6. Perfil metabólico de un grupo de vacas Holstein, al inicio de lactancia
Una alternativa, destinada a reducir el costo de análisis, es mezclar en volúmenes iguales, las
muestras de suero o plasma de las vacas de cada grupo, comúnmente 1 mL de cada muestra,
obteniendo así solo “1 pool” de 8 o más mL en la cual se realizan los análisis
correspondientes. Su interpretación se realiza de forma similar empleando el valor del pool
en reemplazo de la media del grupo ya que equivale al valor de H. Se ha definido que se
pueden agrupar hasta 20 muestras en un “pool” logrado de este un valor equivalente a la
media del grupo. La probabilidad de tener individuos con valores alterados en el rebaño es <
a 10% si el valor de “H” así calculado es < 0,5. Con este sistema se simplifica el trabajo y se
reducen los costos, pero se pierde la información referida a la varianza entre los individuos
del grupo, además de haber una pérdida de sensibilidad diagnóstica con el riesgo de no
identificar rebaños en riesgo.

94
Interpretación
Evaluar la condición metabólica y sanitaria de un grupo de animales constituye uno de los
aspectos más importantes y difíciles de realizar en el PM, requiriendo para ello tener un
adecuado conocimiento de los mecanismos fisiológicos y nutricionales que afectan la
concentración sanguínea de cada metabolito. Para la correcta interpretación de un PM se
requiere considerar el objetivo por el cual fue solicitado y las posibles causas de variación de
un analito: a) fisiológicas (preñez, lactancia, ambiente); b) analíticas (errores de muestra y
metrológicos) y patológicas (desbalances metabólicos nutricionales y sanitarios). Si el PM
considera más de un grupo se debe analizar primeramente los resultados de cada grupo
ordenados acorde con su estado productivo, transición preparto – transición posparto - inicio
de lactancia, y luego relacionarlos entre ellos.
La interpretación se basa en comparar la media y la dispersión de los datos (DE) de cada
variable, las que deben ser similares a las de la población de referencia que generó los IR.
Basado en ello se considera que el grupo es diferente a la población de referencia, por ende,
presenta una alteración cuando:
• El valor de “H” es mayor a 2 (positivo o negativo), lo que indica que la media del analito
difiere en más de ±2 DE a la media poblacional, valor superior a 2 positivo cuando está
aumentado o negativo cuando está disminuido.
• El valor de “CD” es mayor a 1, indica que la varianza entre los individuos del grupo es
mayor que la varianza de la población de referencia. Este hecho se asocia a un factor,
comúnmente de manejo, que provoca una diferencia no deseada entre los individuos del
grupo. También se aprecia cuando el porcentaje de individuos del grupo con valores sobre
a bajo el IR es mayor al 25%.
Los cambios en la concentración sanguínea de un marcador bioquímico son provocados no
solo por su balance de ingreso – egreso, sino también por otros nutrientes, existiendo una
relación entre ellos. Por ejemplo, el colesterol se asocia a la ingesta de materia seca, el ßHB
al uso de reservas de grasa y la urea a la relación RDP/energía en el rumen; por ello los tres
están asociados al balance de energía y mantienen se asocian entre ellos.
Cualquier alteración en los resultados del perfil metabólico debe ser analizada considerando
la estación del año, grupo (s) de animales afectados y las posibles causas que pueden provocar
variaciones en la concentración sanguínea de un analito. Información sumaria de la

95
valoración diagnóstica de los analitos utilizados en el PM se presenta en el Cuadro 5. El PM
del grupo de 8 vacas Holstein, al inicio de lactancia presentado en el Cuadro 6 corresponde
a un rebaño a pastoreo de praderas naturalizadas y suplementadas con 4 kg/d de concentrado
y que presenta índices reproductivos bajo lo esperado. De su análisis se aprecia que es un
grupo homogéneo, con adecuada condición corporal, pero con un fuerte balance negativo de
energía, BEN, (BHB =1,0 mmol/L, H =4,3) y asincronía ruminal de proteínas:energía (urea
= 9,7 mmol/L, H= 4,4). El balance mineral es adecuado y la condición general de salud del
rebaño no evidencia alteraciones trascendentes, si bien hay 2 posibles casos de lipidosis
hepática (GMD > 30 U/L) asociados al BEN y una vaca cursa un cuadro infeccioso crónico
con anemia (globulinas >52 g/L y hemoglobina < 90 g/L) que explican sus elevados valores
del CD del grupo.
El veterinario a cargo del rebaño debe juzgar la trascendencia de las alteraciones detectadas
en función de la magnitud del cambio y los antecedentes del rebaño, fundamentalmente
sanitarios, nutricionales, de producción y manejo. El empleo de los PM en el sur de Chile ha
permitido en numerosas oportunidades que el ganadero o el nutricionista, asesorado por el
médico veterinario, adopte las medidas correctivas que han permitido prevenir o superar
problemas productivos, de fertilidad y sanitarios en muchos rebaños. Es así que entre los años
1986 y 2010 se realizaron un promedio de 129 PM anuales en grupos de vacas lecheras
preparto e inicios de lactancia, en los cuales las alteraciones mayormente diagnosticadas
fueron: BEN, asincronía ruminal de proteínas degradables/energía, hiperfosfatemia,
hiponatremia, y carencia de selenio (Figura 4), con excepción de esta última, que disminuyó
durante el último decenio, las restantes aumentaron fuertemente en dicho período.

96
Figura 4. Porcentaje de rebaños con alteraciones metabólicas diagnosticadas mediante
perfiles metabólicos realizados en el sur de Chile a 3.216 grupos de vacas preparto e inicio
de lactancia
(Weschenfelder et al., 2010*; Noro et al., 2011**; Wagemann et al., 2014**)
Referencias
Bertoni G. 1999. Guida alla interpretazione dei profili metabolici. Univ Perugia.135p.
Bertoni G, E Trevisi. 2013. Use of the liver activity index and other metabolic variables in the assessment of
metabolic health in dairy herds. Vet Clin Food Anim 29: 413–431.
Chapinal N, SJ LeBlanc, ME Carson et al. 2012. Herd-level association of serum metabolites in the transition
period with disease, milk production, and early lactation reproductive performance. J Dairy Sci. 95:5676–82.
Duffield TF. 2007. Peripartum metabolic profiling. Proc Am Assoc Bov Pract. 40: 213-218.
Duffield TF, Lissemore KD, McBride BW, Leslie KE. 2009. Impact of hyperketonemia in early lactation
dairy cows on health and production. J Dairy Sci. 92: 571–580.
Herdt TH, Dart B, Neuder L. 2001. Will large dairy herds lead to the revival of metabolic profile testing? Proc
Am Assoc Bov Pract. 27 - 34.
LeBlanc SJ, Leslie KE, Duffield TF. 2005. Metabolic predictors of displaced abomasum in dairy cattle. J
Dairy Sci. 88: 159-170.
Melendez P, Marin MP, Robles J, Rios C, Duchens M, Archbald L. 2009. Relationship between serum
nonesterified fatty acids at calving and the incidence of periparturient diseases in Holstein dairy cows.
Theriogenology. 72: 826–833.
Noro M, J Borkert, GA Inostroza, R Pulido, F Wittwer. 2011. Variaciones diarias de metabolitos sanguíneos y
su relación con el comportamiento alimenticio en vacas lecheras a pastoreo primaveral. Revista Científica,
FCV-LUZ, 21: 125-130.
Noro M, R Chihuailaf, J Céspedes, F Wittwer. 2010. Acidosis subaguda (SARA) y alcalosis ruminal en
rebaños lecheros en pastoreo otoñal y primaveral. XXXV Congreso Soc. Chilena Prod. Animal. pp 193-194
Ospina PA, Nydam DV, Stokol T, Overton TR. 2010. Evaluation of nonesterified fatty acids and ß-
hydroxybutyrate in transition dairy cattle in the northeastern United States: Critical thresholds for prediction
of clinical diseases. J Dairy Sci. 93:546-554.
Ospina P, JA McArt, TR Overton, Stokol T, DV Nydam. 2013. Using nonesterified fatty acids and b-
hydroxybutyrate concentrations during the transition period for herd-level monitoring of increased risk of
disease and decreased reproductive and milking performance. Vet Clin Food Anim. 29: 387-412.
0 5 10 15 20 25 30 35
Daño hepático ( ↑AST o GGT)*
Carencia Se (↓GPx)***
Hipomagnesemia***
Hiperfosfatemia***
Hipocalcemia***
BEN (↑ßOHB)*
Porcentaje de rebaños
Lactancia Preparto

97
Ospina PA, Nydam DV, Stokol T, et al 2010. Associations of elevated nonesterified fatty acids and beta-
hydroxybutyrate concentrations with early lactation reproductive performance and milk production in
transition dairy cattle in the north-eastern United States. J Dairy Sci. 93:1596–603.
Oetzel GR. 2004. Monitoring and testing dairy herds for metabolic disease. Vet Clin Food Anim. 20: 651-
674.
Payne JM, Payne S. 1987. The metabolic profile test. Oxford: Oxford University Press. 179p.
Sepúlveda-Varas P, DM Weary, M Noro, MAG von Keyserlingk. 2015. Transition diseases in grazing dairy
cows are related to serum cholesterol and other analytes. PLoS ONE 10: 1-13.
Sommer H.1975. Preventive medicine in dairy cows. Vet Med Rev. 1:42-63.
Sordillo LM, V Mavangira. 2014. The nexus between nutrient metabolism, oxidative stress and inflammation
in transition cows. Animal Production Science. 54: 1204–1214.
Van Saun RJ. 2009. Metabolic profiling. In: Anderson DE, Rings DM, editors. Current Veterinary Therapy,
Food Anim Prac. WB Saunders Company. pp 153–164.
Wagemann, C., Chihuailaf, R., Wittwer, F., Noro, M. 2014. Estudio retrospectivo de la prevalencia de
desbalances minerales en grupos de vacas lecheras en el sur de Chile. Arch. Med. Vet. 363-373.
Weschenfelder, M., Barboza, C., Wagemann, C., Böhmwald, H., Chihuailaf, R., Wittwer, F., Noro, M. 2010.
Presentación de desbalances energéticos y alteraciones hepáticas en rebaños lecheros de Chile durante 1986 –
2010. XXXV Congreso Soc. Chilena Prod. Animal. pp 179-180.
Whitaker D, Kelly JM. 1995. Use and interpretation of metabolic profiles in dairy cows. Dept Vet Clin
Studies, Univ Edimburgh. 13p.
Wittwer F. 2000. Marcadores bioquímicos no controle de problemas metabólicos nutricionais em gado de
leite. In: González FHD, Barcellos JO, Ospina H, Ribeiro LAO, eds. Perfil metabólico em ruminantes: seu
uso em nutrição e doenças nutricionais. UFRGS, Brasil. pp53-62.
Wittwer F. 2012. Manual de patología clínica Veterinaria. Imprenta América, Valdivia. 200p.

98
Variações na qualidade composicional do leite no Rio Grande do Sul
Carlos Bondan
A produção brasileira de lácteos cresceu 57,5% de 2005 a 2015 e mesmo assim, a balança
comercial encontra-se historicamente deficitária. A melhoria do poder aquisitivo da
população brasileira tem contribuído para o aumento do consumo e a manutenção de preços
atraentes ao produtor, o que estimula o crescimento do setor lácteo no Brasil (Montoya et al.,
2014). Segundo o IBGE (2013) o Rio Grande do Sul (RS) está na segunda posição na
produção de leite, depois de Minas Gerais, e ocupa a primeira posição na produtividade entre
os estados brasileiros. Em 2012, a média de produção no RS foi 2.670 L/vaca/ano, bem acima
da média brasileira (1.417 L/vaca/ano). Segundo Montoya et al (2014) no período de 2001 a
2012 a produção no Rio Grande do Sul cresceu 82,24%, o número de vacas ordenhadas
aumentou 25,9% e a produtividade aumentou 44,7%, sendo um indicador da
profissionalização da produção leiteira no estado. O leite é um dos alimentos mais ricos
encontrados na natureza, composto por mais de 100.000 moléculas que colaboram com a
nutrição e a imunidade dos mamíferos, servindo também como matéria prima para
industrialização, possibilitando a produção de inúmeros derivados que servem como
alimento para todas as faixas etárias dos humanos (Bachman, 1992).
A glândula mamária tem um incrível nível de organização e uma notável capacidade de
converter nutrientes presentes na circulação sanguínea em componentes do leite. Sua síntese
necessita de intenso trabalho metabólico e sua composição é influenciada pela espécie, raça,
alimentação, estágio de lactação e estado sanitário das fêmeas. Nos bovinos, espécie mais
utilizada na exploração leiteira devido ao volume de leite produzido, a síntese de 1 L de leite
demanda aproximadamente 450 L de sangue, que ao percorrerem o tecido mamário,
entregam nutrientes para a síntese láctea (González, 2001).
A composição química do leite pode ser dividida em constituintes principais e secundários.
Como constituintes principais têm a água, gordura, proteínas e lactose e os secundários os
minerais, vitaminas, enzimas, além de células de descamação do epitélio mamário e
leucócitos (Dürr et al., 2000). O acompanhamento da composição do leite é importante para
a avaliação da dieta e do metabolismo das vacas em lactação, classificação do leite pelo seu

99
valor como matéria prima para a indústria processadora e verificação da integridade do leite
quanto à adição ou retirada de componentes (Dürr et al., 2001).
Sólidos não gordurosos (SNG)
A nutrição da vaca tem efeito marcante sobre a composição do leite, principalmente, no teor
de gordura. O valor de SNG do leite é composto por proteínas, lactose e minerais, e pode
variar em função de mudanças na dieta, porém em menor intensidade do que a gordura.
Outros fatores que influenciam o valor de SNG são a genética, as doenças, estádios de
lactação e as estações do ano. Mudanças que ocorrem no valor de SNG do leite são
primariamente devidas às mudanças na proteína e ocasionalmente ao teor de lactose. O
fornecimento de proteína na dieta, além do requerido conforme o NRC (2001) parece não
exercer efeito sobre o teor de SNG. Entretanto, o fornecimento extra de energia para vacas
de alta produção pode aumentar o valor de SNG em torno de 0,2%. Por outro lado, a redução
no suprimento energético para níveis inferiores aos recomendados pode resultar em queda de
0,2 a 0,5%. Adição de fontes de gordura (animal-vegetal) na dieta tem apresentado resultados
variados no valor de SNG.
Proteína
Atualmente a proteína é o nutriente mais valorizado nos atuais sistemas de pagamento por
qualidade do leite, sendo a caseína seu principal componente. Depois da gordura, a proteína
é o componente que mais varia em função de fatores ambientais, incluídos os nutricionais.
Contudo, o potencial de alteração do teor de proteína no leite através da nutrição não é muito
grande (em torno de 0,5%). Entretanto, à medida que aumenta o teor de proteína no leite,
como a melhoria na energia da dieta, geralmente aumenta a produção total, o que não ocorre
com a gordura (Carvalho, 2002).
As proteínas do leite (caseínas e lactoglobulinas) são sintetizadas nas células secretoras da
glândula mamária a partir de aminoácidos provenientes do sangue ou transportados do
sangue. No entanto, alguns aminoácidos podem ser sintetizados na própria glândula mamária,
através da utilização de precursores como a glicose, acetato e outros aminoácidos. A estrutura
da proteína é determinada pela informação genética contida no DNA da célula, servindo de
molde para uma fita de RNA, que contém a informação sobre a sequência de aminoácidos

100
para síntese das proteínas (Bequete et al., 1998). A síntese das proteínas ocorre no retículo
endoplasmático rugoso, sendo posteriormente secretadas para o lúmen das células através de
vacúolos. A caseína constitui de 76 a 86% do total de proteína láctea, sendo o componente
determinante do rendimento industrial na produção de queijo. É sintetizada pelas células
secretoras, como resultado da expressão de quatro genes (dois genes para α-caseína, s1 e s2;
um para β-caseína e um para κ-caseína), sendo secretada na forma de micelas, que são
grupamentos de várias moléculas de caseína ligadas a íons como o fosfato e o cálcio, e se
encontram em suspensão no leite (De Peters & Cant, 1992).
Ao contrário das caseínas, as proteínas do soro do leite, são aquelas em solução, produzidas
na glândula mamária (α-lactalbumina e β-lactoglobulina) e uma pequena porcentagem de
proteínas que podem passar diretamente do sangue para a glândula mamária (albumina sérica
e imunoglobulinas). O leite possui ainda uma fração de nitrogênio-não-proteico (NNP),
constituindo cerca de 5% da proteína bruta do leite, composto principalmente de ureia, cerca
de 48%, e em menor quantidade de creatinina, amônia e outros compostos nitrogenados (De
Peters & Cant, 1992). Uma equação para predizer a porcentagem de proteína verdadeira no
leite foi proposta pelo NRC (2001), sendo 93% do valor de proteína bruta do leite.
De forma geral, as estratégias nutricionais que visam o aumento da proteína do leite devem
ter como princípio o maior suprimento de aminoácidos e energia (glicose) para a glândula
mamária. A utilização de aminoácidos pela glândula mamária é dependente de uma série de
fatores como, irrigação sanguínea da glândula mamária, concentração arterial de
aminoácidos, eficiência no transporte e regulação intracelular das vias metabólicas (Bequette
et al., 1998). Segundo Murphy & O’Mara (1993), a limitação da produção de proteína pela
glândula mamária é devida principalmente aos seguintes aminoácidos: metionina, lisina,
fenilalanina, histidina ou treonina. O aumento no fornecimento de aminoácidos para a
glândula mamária pode ser obtido através do aumento na quantidade de aminoácidos que
chegam ao intestino delgado (proteína microbiana e proteína não degradada no rúmen),
resultando em maior quantidade de aminoácidos absorvidos e disponíveis. Aminoácidos
resultantes da mobilização proteica (endógena) também colaboram para esse pool. De forma
geral, lisina e metionina são consideradas aminoácidos limitantes.
O teor de proteína bruta da dieta possui efeito muito pequeno sobre o teor proteico do leite.
No entanto, o fornecimento de dietas com deficiências de proteína pode reduzir a

101
concentração deste nutriente em 0,1 a 0,2 unidades percentuais (Sutton, 1989), além de
limitar a produção de leite. Pode-se dizer que a variação no teor proteico da dieta afeta muito
mais a produção de leite do que sua composição.
Lactose
A lactose é um dissacarídeo formado por glicose e galactose com união α 1-4 e sintetizada
exclusivamente na glândula mamária ativa. A síntese é realizada no aparelho de Golgi das
células do epitélio mamário. As moléculas precursoras, glicose e galactose, provêm
principalmente da glicose sanguínea ou de substâncias rapidamente conversíveis em glicose,
através da via gliconeogênica como o propionato, o piruvato, o oxalacetato e os aminoácidos.
Metade da glicose que chega à glândula mamária é direcionada para a síntese de lactose, e a
outra metade para a formação de glicerol, necessário para a síntese dos triglicerídeos do leite
(González & Silva, 2006).
No leite dos bovinos, a lactose apresenta concentração de 4,6% e é considerado o constituinte
mais constante. Sua função está relacionada com a manutenção da osmolaridade e com os
processos de produção e secreção do leite (Larson, 1995). A lactose corresponde por
aproximadamente 50% da capacidade de atração de água para o interior do alvéolo mamário,
sendo responsável por sustentar a produção de leite. Essa é, provavelmente, a razão de ser o
componente com a menor variação (González et al., 2011).
Embora se considere que a lactose é o componente lácteo que apresenta menor variação,
trabalhos anteriores realizados no Brasil revelam diferenças significativas em relação a
variáveis ambientais. Fatores que rompem o equilíbrio metabólico da glândula mamária tais
como a mastite, podem diminuir o conteúdo de lactose no leite (González et al., 2011).
Assim, deve se esperar que aumentos na CCS estejam relacionados com menor teor de
lactose.
Segundo Hurley (2004) a lactose é relativamente insensível às mudanças na dieta das vacas,
no entanto, vacas subnutridas apresentam redução na produção de leite e na percentagem de
lactose, sendo que estes sintomas são revertidos quando dietas adequadas são fornecidas. Em
situações de balanço energético negativo (cetose), principalmente no pré ou pós-parto
imediato, em que não há pico de lactação, há diminuição no teor de lactose. A sua produção
no úbere está relacionada à produção de propionato no rúmen e disponibilidade de produtos

102
gliconeogênicos. Quanto mais lactose for produzida, maior é o volume de água arrastado para
dentro do alvéolo, aumentando o volume de leite (Mühlbach, 2003).
Existem também relatos de que a baixa relação forragem:concentrado na dieta pode aumentar
a lactose do leite e que a elevada suplementação com gorduras pode causar diminuição
(Sutton, 1989). Embora estas mudanças sejam estatisticamente significativas, elas são tão
pequenas que a lactose do leite não pode ser usada como um valor prático indicador do
“status” nutricional do animal.
Gordura
O componente lipídico do leite é formado por uma complexa mistura, sendo os triglicerídeos
os mais importantes. Estes são compostos de três ácidos graxos em ligação covalente a uma
molécula de glicerol por pontes de éster. A gordura do leite é secretada das células mamárias
na forma de glóbulos graxos, principalmente compostos de triglicerídeos rodeados de uma
dupla camada lipídica similar à membrana apical da célula epitelial. A fração de gordura do
leite serve de veículo para as vitaminas lipossolúveis (A, D, E, K), colesterol e outras
substâncias solúveis em gordura, como os carotenoides (provitamina A), que dão ao leite sua
cor amarelo-creme. A quantidade e a composição dos triglicerídeos do leite variam muito
entre as espécies. Nos ruminantes, a proporção de ácidos graxos de cadeia curta e insaturados
é bem maior que nos monogástricos (González et al., 2001; González & Silva, 2006).
Os ácidos graxos do leite são oriundos de duas fontes, da síntese “do novo” na glândula
mamária e da captação direta na circulação sanguínea. Ácidos graxos de cadeia curta,
contendo entre 4 e 8 carbonos, e de cadeia média, com 10 a 14 carbonos, provêm quase que
exclusivamente da síntese “do novo”. Os ácidos graxos de cadeia longa, constituídos por
cadeias superiores a 16 carbonos, são derivados da captação direta dos lipídios da corrente
sanguínea pela glândula mamária. Os ácidos graxos com 16 carbonos podem ser obtidos
através das duas fontes. Cerca de metade dos ácidos graxos do leite são sintetizados pela
própria glândula mamária, a partir da síntese “de novo”. Esta via utiliza como principal fonte
de carbono o acetato produzido na fermentação ruminal. Outra fonte para a síntese “de novo”
é o β-hidroxibutirato produzido pelo epitélio ruminal, a partir do butirato. Os ácidos graxos
pré-formados captados pela glândula mamária e diretamente usados para a síntese de gordura
do leite são derivados das lipoproteínas circulantes provenientes da mobilização de reservas

103
corporais e dos ácidos graxos não esterificados originários da absorção dos lipídios no trato
gastrointestinal. Em condições normais, a lipólise é responsável por menos de 10% dos
ácidos graxos da gordura do leite, sendo a maior parte dos ácidos graxos de cadeia longa
(AGCL) do leite proveniente da absorção intestinal. No entanto, animais em balanço
energético negativo, com grande mobilização de gordura corporal, têm a lipólise como fonte
de uma maior contribuição de gordura do leite (Bauman & Griinari, 2003).
A gordura é o principal componente energético do leite, sendo também responsável pelas
propriedades físicas às características industriais, e as qualidades organolépticas do leite e
seus derivados, por isso, a gordura láctea possui um importante valor econômico (Bauman &
Griinari, 2003).
No Brasil, ainda persiste o pagamento do leite pela produção total e o teor de gordura. Nos
últimos anos, diversos países têm dado maior ênfase para o teor de proteína, utilizando este
critério nos sistemas de pagamento por qualidade. Esta tendência se explica porque, enquanto
a gordura tem tido seu valor reduzido pelos hábitos de consumo da população, a proteína tem
sido valorizada por ser determinante do rendimento industrial de derivados lácteos
(Monardes, 1998). No Brasil a IN 51/2002 estabelece que o limite mínimo para a gordura no
leite cru refrigerado deve ser de 3%.
Vários aspectos exercem efeito sobre a concentração de gordura no leite. O fator racial e a
seleção genética, o estágio de lactação, a temperatura ambiente e as condições de estresse do
animal, a perda de condição corporal, a estação do ano, a contagem de células somáticas, a
saúde geral do animal, a manifestação de cio, a frequência e a técnica de ordenha e,
principalmente, os fatores nutricionais têm sido implicados como fatores que contribuem
com a variação da constituição lipídica do leite (González & Campos, 2003; Noro et al.,
2006; Barbano, 1990; Carvalho, 2002; Ribas et al., 2001; Marques et al., 2002).
Em ruminantes, a composição dos ácidos graxos da dieta não reflete a composição dos ácidos
graxos da gordura do leite, devido a que os constituintes lipídicos da dieta são alterados pelo
metabolismo microbiano (Bauman & Griinari, 2003). Os fatores nutricionais são os que
podem alterar de forma mais direta e com resultados mais rápidos e evidentes a gordura do
leite. No entanto, a manipulação da dieta visando alterações na gordura do leite demanda
conhecimento aprofundado, uma vez que esta manipulação afeta não somente a fermentação
ruminal, como também o metabolismo geral do animal (Mühlbach, 2003).

104
A alimentação fornecida para as vacas leiteiras influencia o funcionamento normal do rúmen.
Para manter a ruminação e a consequente produção de saliva, a dieta deve apresentar uma
relação mínima de volumoso/concentrado de 50/50 para manutenção de um pH adequado,
acima de 6,0, quando a fermentação da fibra é favorecida, propiciando uma maior ingestão
de matéria seca e, consequentemente, maior produção de leite. A efetividade da fibra
utilizada também deve ser levada em conta, o que se reflete diretamente na condição ruminal
e na gordura do leite. A fibra efetiva atua estimulando a ruminação e a produção de saliva, o
que mantém o pH favorável para a digestão da fibra, resultando em maior disponibilidade de
ácido acético, o principal precursor da gordura no leite (Mühlbach, 2003).
O fornecimento de grandes quantidades de carboidratos prontamente fermentáveis e reduzida
quantidade de fibra, ou dietas com quantidade adequada de fibra, mas com fibra de pouca
efetividade, possuem pouca capacidade de manutenção do funcionamento normal do rúmen,
podendo assim, levar a depressão na gordura do leite (Bauman & Griinari, 2003). Quando o
consumo de concentrados ultrapassa 50% da matéria seca da dieta, ocorre depressão na
gordura do leite e mudanças também na composição dos ácidos graxos (Palmquist et al.,
1993).
A suplementação com óleos poli-insaturados, provenientes de fontes vegetais ou marinhas,
também reduzem a gordura do leite. O mesmo ocorre quando sementes de oleaginosas, ricas
em ácidos graxos poli-insaturados, são adicionadas em grande quantidade à dieta (Bauman
& Griinari, 2003). Griinari et al. (1998), confirmaram que dietas pobres em fibra e ricas em
ácidos graxos insaturados (óleo de milho) aumentam o conteúdo de C18:1 trans. Segundo
estes mesmos autores, este tipo de dieta está associado a uma diminuição significativa na
produção e conteúdo da gordura do leite.
A utilização de aditivos também interfere na síntese de gordura. É o caso dos tamponantes e
alcalinizantes que minimizam a queda do pH ruminal, favorecendo a digestão da fibra, e dos
manipuladores de fermentação (ionóforos) que alteram o perfil dos ácidos graxos voláteis
(AGV) no rúmen, reduzindo perdas com metano e gás carbônico e diminuindo a relação
acetato/propionato.
Em dietas com grandes quantidades de concentrados (acima de 50% da matéria seca), ou
grandes quantidades de alimentos fermentados, recomenda-se o uso de bicarbonato de sódio
ou suplementos minerais tamponantes para normalizar o ambiente ruminal e,

105
consequentemente, o teor de gordura do leite. Nestes casos, a ação dos tamponantes
alimentares compensa a menor ruminação, que por sua vez proporciona uma menor taxa de
salivação e tamponamento natural do rúmen. Como o bicarbonato de sódio é de curta ação
no rúmen, aditivos probióticos, como cepas específicas de leveduras vivas (Saccharomyces
cerevisiae) vêm sendo utilizadas como complemento aos sais tamponantes. Em face de sua
ação metabólica mais prolongada no rúmen, a levedura promove uma maior estabilidade do
rúmen, mantendo o ambiente ruminal mais favorável à digestão da celulose por elevar a
densidade populacional das bactérias celulolíticas e consumidoras de lactato (Mühlbach,
2003).
A adição de monensina à dieta de vacas leiteiras diminuiu a produção de metano e a ingestão
de matéria seca. A gordura total e a percentagem de gordura diminuíram temporariamente.
No entanto, a produção de leite e a quantidade de todos os ácidos graxos insaturados
aumentaram, incluindo o ácido linoleico conjugado (CLA), enquanto os saturados
diminuíram.
Várias teorias têm sido propostas na tentativa de se explicar a depressão da gordura do leite,
sendo a base para o desenvolvimento de todas as teorias os processos microbianos no rúmen
(Bauman & Griinari, 2003). Das diversas teorias propostas, três delas continuam tendo
suporte na literatura científica. A primeira delas sugere que o maior fornecimento de
concentrado na dieta, com elevação da proporção concentrado/volumoso, aumenta a
produção de ácidos no rúmen, o que promove a queda do pH ruminal. Em pH baixo, a
degradação da fibra é comprometida, promovendo alterações na fermentação ruminal,
resultando em inadequada produção de acetato e butirato, limitando sua utilização na síntese
da gordura do leite. Bauman & Griinari (2003) concluem que a variação na concentração de
gordura do leite é justificada pelas variações na proporção molar de AGV no rúmen. Em
outros estudos, observou-se que a produção de acetato não foi afetada significativamente, no
entanto, a relação molar de acetato/propionato do fluido ruminal apresentou forte queda. Essa
queda na relação acetato/propionato foi proporcionada, principalmente, pela elevação na
produção de propionato e não pela queda na produção de acetato (Bauman et al., 1971).
Apesar das evidencias de que em dietas com baixa fibra a produção de acetato e butirato não
são afetadas de forma apreciável, um déficit desses AGV é frequentemente considerada como
colaborador na redução da gordura do leite. Isto pode ser atribuído ao fato de que dietas com

106
baixa fibra normalmente reduzem o pH e isto afetará a taxa relativa de absorção individual
de ácidos graxos voláteis (Bauman & Griinari, 2003).
A segunda teoria ou teoria glicogênica-insulina é baseada na competição por nutrientes, entre
a glândula mamária e outros tecidos. Dietas com baixa fibra resultam em aumento da
produção de propionato no rúmen e elevação da taxa de gliconeogênese hepática. Além disto,
estas dietas resultam em melhor balanço energético devido a maior ingestão de energia e
redução na secreção da gordura do leite. Como consequência dessa combinação de fatores, a
concentração de insulina no sangue é aumentada, desviando nutrientes da glândula mamária.
Isto ocorre porque a insulina aumenta a utilização de acetato, β-hidroxibutirato e AGCL no
tecido adiposo, ou seja, há um aumento da lipogênese. Além disso, a insulina também reduz
a mobilização dos AGCL das reservas corporais. A teoria glicogênica tem sido avaliada por
meio de infusões exógenas de propionato e glicose. Os resultados encontrados de redução na
gordura do leite têm se mostrado altamente variáveis, entre 0 e 14% (Bauman & Griinari,
2003).
A terceira teoria é a que, na última década, tem recebido maior suporte dos pesquisadores.
Esta teoria sugere que certas dietas alteram a biohidrogenação dos ácidos graxos insaturados
no rúmen, produzindo ácido graxo intermediário denominado de ácido linoleico conjugado
(CLA) trans-10, cis-12. Estes AGCL teriam uma potente ação inibidora da síntese de gordura
no leite (Bauman & Griinari, 2003). O CLA trans-10, cis-12 diminui a capacidade lipogênica
da glândula mamária (taxas de incorporação de acetato em ácidos graxos) e a expressão de
genes de enzimas relacionadas ao transporte de ácidos graxos circulantes, diminui a síntese
“de novo” de ácidos graxos, a dessaturação de ácidos graxos e a formação de triglicerídeos.
Contagem de células somáticas (CCS)
Células somáticas são todas as células presentes no leite, que incluem as células originárias
da corrente sanguínea como leucócitos e células de descamação do epitélio glandular
secretor. Em casos de inflamação (mastite), há um aumento considerável na CCS,
principalmente por neutrófilos. É importante quantificar e qualificar os tipos celulares
presentes para conhecer o grau de inflamação e caracterizar se a doença é aguda ou crônica.
Além do aumento do número de células, a mastite provoca alterações nos três principais
componentes do leite, gordura, proteína e lactose. A extensão do aumento da CCS e as

107
mudanças na composição do leite estão diretamente relacionadas com a superfície do tecido
mamário atingido pela reação inflamatória. Portanto, há uma relação direta entre a CCS e a
concentração dos componentes do leite. Em relação às proteínas ocorre uma redução
naquelas sintetizadas na glândula mamária (α e β caseína, α-lactalbumina e β-lactoglobulina)
e aumento das proteínas de origem sanguínea (albumina sérica e imunoglobulinas), em
virtude do aumento de permeabilidade vascular secundário ao processo inflamatório. A
proteína total do leite tem pouca variação, mas a concentração de cada tipo de proteína varia
acentuadamente (Ribas, 1999).
A CCS no leite é uma ferramenta valiosa na avaliação e estimativa das perdas quantitativas
e qualitativas da produção do leite e derivados, como indicativo da quantidade do leite
produzido na propriedade e para estabelecer medidas de prevenção e controle da mastite.
Uma dessas medidas pode ser a implantação de protocolos de manejo de ordenha. Como
consequência de altos níveis de células somáticas, observam-se prejuízos tanto ao produtor
de leite quanto à indústria de laticínios. As maiores perdas causadas ao produtor estão
relacionadas à redução da produção. Consequentemente, esta redução gera problemas de
captação da matéria-prima para a indústria (Fonseca & Santos, 2000). As perdas sofridas pela
indústria são causadas pelas alterações químicas e microbiológicas do leite com alta CCS,
acarretando em diminuição do rendimento industrial e redução de sua qualidade final
(Fonseca & Santos, 2000).
Nas condições brasileiras, considera-se que, em animais sadios, a CCS individual no leite
deve ser menor de 300x103/mL (Fonseca & Santos, 2000), mas em países com maior
desenvolvimento esse limite chega a 200x103/mL (Philpot, 1998). A contagem de células
somáticas de tanque (CCST) possui limites legais maiores, sendo de 400x103/mL em países
da União Europeia e Austrália, de 500x103/mL no Canadá e de 750x103/mL nos EUA (Brito,
2003). No Brasil, a determinação legal para CCST (Instrução Normativa 62/2011) é de
500x103/mL, valor que deve diminuir para 400x103/mL a partir de primeiro de julho de 2016.
Variação da produção e composição do leite no Rio Grande do Sul
Para compreender os fatores que influenciam a produção e a composição do leite produzido
no Rio Grande do Sul foi realizado um estudo em 115 fazendas leiteiras que ordenhavam
vacas da raça Holandesa e realizavam controle leiteiro no período de janeiro de 2008 a

108
dezembro de 2013. Os efeitos do ano, das estações do ano, das etapas da lactação e do número
de lactações foram comparados com as variáveis: produção e composição do leite. A
produção média de leite no período foi de 25,54 ± 8,63 L/vaca/dia, superior às médias
brasileiras (4,64 L/vaca/dia), catarinenses (8,26 L/vaca/dia) e gaúchas (8,75 L/vaca/dia)
considerando lactações de 305 dias (Montoya et al., 2014) e superiores às produções das
vacas de raça Holandês descritas por Noro et al. (2006) no Rio Grande do Sul (19,36
L/vaca/dia), Nunes Jr et al. (2000) em Pernambuco (16,7 L/vaca/dia), Araújo et al. (2000)
em Minas Gerais (17,02 L/vaca/dia) e Bajaluk et al. (1999) no Paraná (24,77 L/vaca/dia).
Costa et al. (2013) estudaram a produção de leite em 24 rebanhos na região dos Campos
Gerais no estado do Paraná no período de janeiro de 2010 a dezembro de 2012 e encontraram
produção média de 30,9 ± 3,7 L/vaca/dia. Estas diferenças de produtividades estão
relacionadas com o balaço nutricional, características raciais e genéticas, assim como as
condições ambientais (Dürr et al., 2011). O efeito ano do ano sobre a produção e composição
láctea está apresentado na Tabela 1. Entre 2008 e 2010 ocorreu diminuição da produção
acompanhada pelo aumento do escore de células somáticas (ECS). Houve crescimento
progressivo da produção de leite de 2010 a 2013 em 8,20% enquanto e diminuição do ECS.
A gordura e os sólidos totais aumentaram progressivamente entre 2008 e 2013. Em 2010
foram observados os maiores níveis de gordura e sólidos totais, e nos anos de 2011 e 2012
não apresentaram diferenças entre si, sendo superiores aos anos anteriores. O maior
percentual de gordura e sólidos totais ocorreu em 2013. A proteína apresentou tendência de
aumento entre 2008 e 2013. O percentual de lactose foi maior em 2013. Nos anos de 2010,
2011 e 2012 não houve diferença nos percentuais de lactose e 2008 e 2009 apresentaram os
menores percentuais, respectivamente.
O aumento da produtividade e da composição química pode ser explicado pela relação preço
do leite e custo dos fatores de produção. O valor praticado na compra do leite cru no período
foi historicamente maior quando comparado ao custo de produção, tornando atrativos os
investimentos que melhoraram o desempenho produtivo das vacas (Montoya et al., 2014).

109
Tabela 1. Médias anuais (± desvio padrão) de produção, composição química e ECS entre o período de
janeiro de 2008 a dezembro de 2013 no Rio Grande do Sul
Ano Produção
(L/vaca/dia)
Gordura
(%)
Proteína
(%)
Lactose
(%)
Sólidos
totais (%) ECS#
2008 26,2 ± 9,08b 3,37 ± 0,65d 3,18 ± 0,38d 4,43 ± 0,25c 11,9 ± 0,94d 4,95 ± 2,01e
2009 25,2 ± 8,41d 3,38 ± 0,66d 3,17 ± 0,37e 4,41 ± 0,26d 11,9 ± 0,94d 5,26 ± 1,87b
2010 24,6 ± 8,14f 3,40 ± 0,68c 3,23 ± 0,35c 4,46 ± 0,25b 12,1 ± 1,02c 5,45 ± 1,61a
2011 25,0 ± 8,21e 3,50 ± 0,67b 3,26 ± 0,37b 4,46 ± 0,25b 12,2 ± 1,01b 5,30 ± 1,61b
2012 25,5 ± 8,53c 3,50 ± 0,69b 3,28 ± 0,39a 4,46 ± 0,25b 12,2 ± 0,99b 5,14 ± 1,70d
2013 26,8 ± 8,63a 3,52 ± 0,67a 3,24 ± 0,38c 4,49 ± 0,25a 12,2 ± 0,95a 5,19 ± 1,58c # Escore de células somáticas: logaritmo da contagem de células somáticas a, b, c, d, e indicam diferenças entre colunas
Comparando os achados de Noro et al. (2006) com as médias deste estudo, a produção de
leite apresentou crescimento de 24,07% e a constituição proteica 2,5%, enquanto a gordura
e lactose apresentaram queda de 3,38% e 0,66% respectivamente. Portanto, o aumento dos
componentes do leite não reflete melhorias contínuas nos últimos anos, pois os achados de
Noro et al. (2006) apresentam leite com qualidade composicional superior aos encontrados
neste estudo. O aumento na produtividade é consequências de um planejamento estratégico
onde o controle leiteiro é indispensável para a condução de manejos racionais nos rebanhos,
porém é necessário que o controle leiteiro também seja utilizado para melhorias na qualidade
do leite e não somente para aumento da produção e isto só será alcançado com a
conscientização de produtores e indústrias (Dürr et al. 2011). O comportamento estacional
da produção e composição do leite está apresentado na Tabela 2. A produção de leite foi
maior no inverno, decrescendo progressivamente na primavera, outono e verão. A gordura
foi maior no outono seguido pelo inverno, primavera e verão. A proteína foi maior no inverno
e outono, decrescendo no verão e primavera, respectivamente. O ECS foi maior no verão e
outono seguido pelo inverno, os menores ECS foram encontrados na primavera. A
concentração de lactose apresentou a mesma tendência que a produção de leite entre as
estações do ano, sendo maior no inverno, decrescendo progressivamente na primavera,
outono e verão.

110
Tabela 2. Médias (± desvio padrão) de produção, composição química e ECS nas diferentes estações do ano,
entre janeiro de 2008 a dezembro de 2013 no Rio Grande do Sul
Estação
do ano
Produção
(L/vaca/dia)
Gordura
(%)
Proteína
(%)
Lactose
(%)
Sólidos
totais (%) ECS#
Inverno 27,3 ± 9,16a 3,48 ± 0,69b 3,27 ± 0,36a 4,48 ± 0,25a 12,2 ± 0,98a 5,20 ± 1,71b
Primavera 25,8 ± 8,68b 3,40 ± 0,66c 3,17 ± 0,36c 4,46 ± 0,25b 12,0 ± 0,98b 5,16 ± 1,78c
Verão 24,2 ± 7,8d 3,37 ± 0,65d 3,18 ± 0,35b 4,42 ± 0,25d 11,9 ± 0,95c 5,27 ± 1,78a
Outono 24,4 ± 8,48c 3,53 ± 0,68a 3,28 ± 0,88a 4,43 ± 0,26c 12,2 ± 0,98a 5,26 ± 1,69a # Escore de Células Somáticas: logaritmo da contagem de células somáticas a ,b, c, d indicam diferenças entre colunas
O aumento da produção e da lactose durante o inverno seguido pela primavera corrobora com
os achados de Noro et al. (2006), quem atribuíram este aumento à melhor qualidade das
pastagens nestas estações do ano. Broderick (2003) observou que o aumento na porcentagem
de lactose do leite aumenta com maior valor energético da dieta. As forragens temperadas no
final do outono, inverno e início da primavera no Rio Grande do Sul, têm menor fibra em
detergente neutro (FDN) e maior concentração de carboidratos solúveis quando comparadas
as forrageiras tropicais, influenciando no desempenho produtivo dos animais (Fontaneli et
al., 2009). Os maiores percentuais de gordura e proteína ocorreram no outono e inverno.
Contudo, no outono, diferente do inverno, observou-se a menor produção de leite, embora os
níveis de sólidos não diferissem dos encontrados no inverno. Uma diminuição no volume de
leite pode trazer como consequência aumento nos teores de sólidos, principalmente gordura
e proteína, devido ao efeito concentrador do leite (Weiss et al., 2002). Não restam dúvidas
que as forrageiras temperadas, fornecem melhor equilíbrio nutricional o que explica em
partes a maior produção e as maiores proporções de gordura, proteína, lactose e sólidos totais
no inverno. Heck et al. (2009) estudaram as variações da composição do leite de vacas na
Holanda e atribuíram parte das variações dos componentes químicos a fatores sazonais.
Assim como no Rio Grande do Sul a gordura apresentou a maior variação seguida pela
proteína. A lactose foi o componente que apresentou menor variação. A explicação de Heck
et al. (2009) para a diminuição dos níveis de gordura está no pastejo de forragens frescas que
concentram ácidos graxos insaturados trans. Os ácidos graxos trans interfere na ação
enzimática da Δ9-desaturase, responsável pela síntese de ácidos graxos de cadeia curta e
média na glândula mamária que contribuem para a síntese do novo.

111
As condições de conforto térmico também interferem na produção e composição do leite.
Lambertz et al. (2013) concluíram que vacas no verão, com estresse térmico, diminuem
produção e composição de gordura e proteína e aumento no ECS, corroborando com os
resultados encontrados neste estudo onde o outono e verão apresentaram os maiores ECS.
A Tabela 3 apresenta as variações entre as etapas da lactação. As vacas em início da lactação
(6 a 60 DEL) apresentaram as maiores produções de leite, ocorrendo diminuição conforme o
avanço da lactação. O maior percentual de gordura foi observado em vacas com mais de 220
DEL, e o menor percentual foi observado no período de 6 a 60 e 61 a 120 DEL, sem
diferenças entre eles. O conteúdo de proteína láctea aumentou à medida que a lactação
avançou, com maiores valores nas vacas com mais de 200 DEL, e menores nas vacas entre
61 a 120 DEL.
Tabela 3. Médias (± desvio padrão) de produção, composição química, ECS e cociente entre G:P em
diferentes estádios de lactação ordenhadas entre janeiro de 2008 a dezembro de 2013 no Rio Grande do Sul
Etapa de
lactação
(dias)
Produção
(L/vaca/dia)
Gordura
(%)
Proteína
(%) G:P* Lactose (%)
Sólidos
totais (%)
ECS#
6 a 60 29,4 ± 8,72a 3,40 ± 0,65b 3,05 ± 0,36c 1,11 ± 0,22a 4,50 ± 0,23b 11,9 ± 0,93c 4,79 ± 1,90d
61 a 120 29,2 ± 8,66b 3,30 ± 0,66c 3,03 ± 0,31d 1,09 ± 0,21b 4,51 ± 0,23a 11,8 ± 0,92d 4,89 ± 1,90c
121 a 220 26,2 ± 8,01c 3,40 ± 0,66b 3,18 ± 0,32b 1,07 ± 0,19c 4,45 ± 0,24c 12,0 ± 0,94b 5,21 ± 1,75b
> a 220 22,0 ± 7,49d 3,55 ± 0,67a 3,41 ± 0,36a 1,04 ± 0,18d 4,40 ± 0,27d 12,4 ± 0,99a 5,53 ± 1,53a # Escore de células somáticas: logaritmo contagem de células somáticas
* G:P= cociente gordura:proteína a, b, c, d indicam diferenças entre colunas
O cociente G:P aumentou progressivamente com o avanço da lactação. O cociente
gordura:proteína (G:P) tem sido usada como indicador de lipomobilização em vacas com
balanço energético negativo (BEN). Considerando que o aumento de ácidos graxos livres,
originados do tecido adiposo, contribuem com o aumento do teor de gordura láctea é sugerido
que valores de G:P superiores a 1,25 em vacas de até 60 DEL sejam indicativos de BEN
moderado (DE ROOS et al., 2007). No presente trabalho, a G:P para vacas entre 6 a 60 DEL
foi de 1,11 ± 0,22, sendo que 25,72% apresentaram cociente ≥ 1,25. Cucunubo et al. (2013)
e Duffield et al (1997) encontraram baixa especificidade e sensibilidade na utilização dos
coeficientes de G:P de 1,25 como indicador de BEN e/ou cetose. Quando o cociente é

112
ajustado para ≥ 1,50, conferindo maior especificidade e sensibilidade, o percentual de vacas
com BEN e/ou cetose foi de 5,15% neste estudo.
O número de lactações influenciou a produção de leite (Tabela 4), de forma que vacas com
duas e três lactações tiveram as maiores produções, seguidas pelas vacas com quatro ou mais
lactações. A expressão máxima da produção leiteira de uma vaca ocorre quando ela atinge o
completo desenvolvimento da glândula mamária, o que ocorre na terceira lactação das vacas
Holandesas (Schutz et al., 1990). As vacas de primeira lactação apresentaram as menores
produções e as maiores concentrações de gordura, proteína, lactose e sólidos totais, resultados
semelhantes aos encontrados por Cunha et al. (2008). Este aumento pode ser atribuído ao
menor ECS nas vacas de primeira lactação e consequentemente menor dano as células
produtoras de leite e à menor produtividade quando comparado às multíparas, causando
efeito concentrador no leite (Auldist e Hubble, 1998; Weiss et al., 2002). A gordura, proteína,
lactose e sólidos totais diminuíram conforme aumentou o número de lactações. O ECS
também aumentou conforme aumentou o número de lactações. Schultz (1977) atribui o
aumento do ECS ao maior número de partos que como consequência causa perda de células
epiteliais secretoras, diminuído os componentes do leite. O cociente G:P foi maior em vacas
de primeira lactação seguido pelas vacas com mais de quatro lactações e duas e três lactações
respectivamente sugerindo que as vacas de primeira lactação estão mais predispostas a
ocorrência de transtornos metabólicos.
Tabela 4. Médias de produção (± desvio padrão), composição química, ECS e cociente entre G:P, distribuídas
conforme o número de lactações ordenhadas entre janeiro de 2008 a dezembro de 2013 no Rio Grande do Sul
Número
de
partos
Produção
(L/vaca/dia)
Gordura
(%)
Proteína
(%) G:P*
Lactose
(%)
Sólidos
totais (%) ECS#
1 23,9±7,72c 3,47±0,67a 3,24±0,37a 1,08±0,19a 4,54±0,23a 12,2±0,97a 4,83±1,73c
2-3 26,9±9,09a 3,43±0,68b 3,23±0,38b 1,07±0,20c 4,43±0,24b 12,1±0,98b 5,31±1,72b
≥ 4 26,1±8,86b 3,41±0,67c 3,19±0,37c 1,07±0,20b 4,32±0,26c 11,9±0,97c 5,84±1,62a # Escore de células somáticas: logaritmo da contagem de células somáticas
* G:P=cociente gordura:proteína a, b, c indicam diferenças entre colunas
A Tabela 5 apresenta resultados de correlação onde a produção de leite correlacionou-se
positivamente com os percentuais de lactose e o número de partos. Segundo Larson (1995),
a lactose aumenta a osmolaridade do leite e consequentemente a absorção de água para o

113
alvéolo, aumentando a produção. O aumento do ECS no leite teve impacto negativo sobre a
produção de leite, percentual de lactose e o cociente G:P. Hagnestam-Nielsen et al. (2009)
encontraram diminuição da produção de leite em vacas primíparas e multíparas com elevada
contagem de células somática (CCS). As perdas foram estimadas em 1,9 e 5,2% para vacas
primíparas e multíparas, respectivamente. Os mesmos autores encontraram as maiores perdas
de produção leiteira conforme avançou a lactação, sendo os coeficientes de regressão mais
negativos entre as semanas 33-44 independentemente do número de partos. Esses achados
estão de acordo com os resultados encontrados neste estudo, observando-se que à medida
que avança a lactação e o número de partos, maior é o valor de ECS e menor a produção de
leite.
Tabela 5. Correlações entre os dias em leite (DEL), número de partos, produção e composição láctea
DEL N° partos Produção
Gordura
(%)
Proteína
(%) G:P##
Lactose
(%)
Sólidos
(%)
Produção -0,341** 0,121** Gordura
(%) 0,125** -0,038** -0,226** Proteína
(%) 0,394** -0,041** -0,396** 0,400**
G:P -0,121** -0,013** 0,012** Lactose
(%) -0,164** -0,308** 0,200** -0,042** -0,085** 0,006* Sólidos
(%) 0,220** -0,131** -0,279** 0,879** 0,700** 0,486** 0,208**
ECS# 0,165** 0,214** -0,158** 0,076** 0,174** -0,028** -0,429** 0,016** ** Correlação significativa a nível de 0,01
*Correlação significativa a nível de 0,05
# Logaritmo da contagem de células somáticas
## G:P=cociente gordura:proteína
A diminuição dos níveis de lactose em vacas com elevado ECS também foi observado por
Prada e Silva et al. (2000), atribuindo 34% das perdas de lactose como sendo causadas pela
CCS. O ECS teve correlação positiva com os teores de gordura, proteína e sólidos, e com os
dias em lactação e número de partos, ao tempo que teve correlação negativa com a produção
de leite. Shutz et al. (1990) e Cunha et al. (2008) observaram correlações positivas entre o
ECS e os percentuais de gordura e proteína. Similarmente, Miller et al. (1983) analisando
leite individualizado por vaca e Mitchell et al. (1986) com leite de tanque de expansão,
verificaram um aumento da porcentagem de gordura no leite com alta CCS. Segundo Auldist

114
e Hubble (1998), não há consenso na literatura em relação ao aumento da porcentagem de
proteína total no leite de animais com alta CCS. Kitchen (1981) e Munro et al. (1984)
descrevem que o aumento da concentração de proteínas séricas no leite de vacas com mastite
subclínica pode ser atribuído ao aumento na permeabilidade vascular em consequência do
processo inflamatório. Em nosso estudo a correlação entre ECS e lactose foi de -0,429 (P<
0,01). Noro et al. (2006) no Rio Grande do Sul e Cunha et al. (2008) em Minas Gerais
encontraram as mesmas influencias do ECS sobre a produção e composição do leite,
indicando tratar-se de um problema persistente que afeta a produção e qualidade do leite.
Conclusão
A produtividade e composição do leite das fazendas que utilizam controle leiteiro no norte e
nordeste do Rio Grande do Sul têm melhorado nos últimos anos. É possível observar o efeito
da sazonalidade, onde no período de inverno ocorrem os maiores volumes de leite e de
sólidos totais. Vacas no início da lactação (6 a 60 DEL) e aquelas com dois e três partos
foram as mais produtivas. O maior desafio a ser vencido é a diminuição da contagem de
células somáticas que se correlacionaram negativamente com a produção e o percentual de
lactose.
Referências
Auldist, M. J., Hubble, I. B. Effects of mastitis on raw milk and dairy products. Australian Journal of Dairy
Technology, v. 53, p. 28-36, 1998.
Bachman, K. C. Managing milk composition. In: Van Horn, H., Wilcox, C. Large dairy herd management.
Champing: Dairy Science Association American, Dairy Science Association. 1992. p. 336-346.
Barbano, D.M. Seasonal and regional variation in milk composition in the US. In: Cornell Nutrition
Conference for Feed Manufacturers. 1990, Ithaca.
Bauman, D. E.; Griinari, J. K. Nutritional regulation of milk fat synthesis. Annual Review of Nutrition., v. 23,
p. 203-227, 2003.
Brito, J.R.F.; Portugal, J.A.B. Panorama da qualidade do leite na região Sudeste: Espírito Santo, Minas Gerais
e Rio de Janeiro. In: Diagnóstico da qualidade do leite, impacto para a indústria e a questão dos resíduos de
antibióticos. Juiz de Fora: Embrapa Gado de Leite, 2003. p.47-62.
Broderick, G. A. Effects of varying protein and energy level on the production of lactating dairy cows.
Journal of Dairy Science, v. 86, p. 1370-1381, 2003.
Beckett, S.; Lean, I.; Dyson, R.; Tranter, W.; Wade, L. Effects of monensin on the reproduction, health, and
milk production of dairy cows. Journal of Dairy Science, v. 81, p. 1563-1573, 1998.
Carvalho, G.F. et al. Milk yield, somatic cell count and physico-chemical characteristics of raw milk collected
from dairy cows in Minas Gerais State. In: Congresso Panamericano de Qualidade do Leite e Controle da
Mastite, 2002, Ribeirão Preto. Anais. Ribeirão Preto: FEPALE, 2002. 1 CD-ROM.

115
Cucunubo, L. G.; Strieder-Barboza, C.; Wittwer, F.; Noro, M. Diagnóstico de cetosis subclínica y balance
energético negativo en vacas lecheras mediante el uso de muestras de sangre, orina y leche. Revista
Científica, FCV-LUZ, v. 23, p. 111-119, 2013.
Cunha, R. P. L.; Molina, L. R.; Carvalho, A. U.; Facury Filho, E. J.; Ferreira, P. M.; Gentilini, M. B. Mastite
subclínica e relação da contagem de células somáticas com número de lactações, produção e composição
química do leite em vacas da raça Holandesa. Arquivos Brasileiros de Medicina Veterinária e Zootecnia, v.
60, p. 19-24, 2008.
De Peters, E. J.; Cant, J. P. Nutritional factors influencing the nitrogen composition of bovine milk: a review.
Journal of Dairy Science, v. 75, p. 2043-2070, 1992.
De Roos, A. P.; Van Den Bijgaart, H. J.; Horlyk, J.; De Long, G. Screening for subclinical ketosis in dairy
cattle by Fourier transform infrared spectrometry. Journal of Dairy Science, v. 90, p. 1761-1766, 2007.
Duffield, T. F.; Kelton, D. F.; Leslie, K. E.; Lissemore, K. D.; Lumsden, J. H., Use of test day milk fat and
milk protein to detect subclinical ketosis in dairy cattle in Ontario. Canadian Veterinary Journal, v. 38, p. 713-
718, 1997.
Dürr, J. W., Fontaneli, R. S., Burchard, J. F. (2000). Fatores que afetam a composição do leite. In: Cursos de
sistemas de produção para gado leiteiro baseado em pastagens sob plantio direto. Passo Fundo. Anais...
Embrapa-Trigo.
Dürr, J. W. Programa nacional de melhoria da qualidade do leite: uma oportunidade única. In: Dürr J. W.;
Carvalho M.P.; Santos, M.V. (eds.). O compromisso com a qualidade do leite no Brasil. Passo Fundo:
Universidade de Passo Fundo, Passo Fundo, 2004. p. 38-55.
Dürr, J. W.; Ribas, N. P.; Costa, C. N.; Horst, J. A.; Bondan, C. Milk recording as an indispensable procedure
to assure milk quality. Revista Brasileira de Zootecnia, v. 40, p. 76-81, 2011.
Fonseca, L. F. L.; Santos, M. V. Qualidade do leite e controle de mastite. São Paulo: Lemos Editorial, 2000.
p.39-141.
Fontaneli, R. S.; Santos, H. P.; Fontaneli, R. S. Forrageiras para integração lavoura-pecuária-floresta na região
sul-brasileira. Passo Fundo: Embrapa Trigo. 2009.
González, F. H. D. (2001). Composição do leite e hormônios da lactação. In: Uso do leite para monitorar a
nutrição e o metabolismo de vacas leiteiras. Gráfica da Universidade Federal do Rio Grande do Sul, Porto
Alegre.
González, F.H.D.; Campos, R. O leite como indicador metabólico-nutricional em vacas. A Hora Veterinária,
v.22, p.36-38, 2003.
González, F. H. D., Noro, G. (2011). Variação na composição do leite no subtrópico brasileiro. In: Qualidade
do leite bovino: variações no trópico e subtrópico. UPF Editora, Passo Fundo.
González, F. H. D., Silva. S. C. Introdução a bioquímica clínica. 2ª edição, Editora da UFRGS, Porto Alegre,
2006. p. 364
Griinari, J. M.; Dwyer, D. A.; McGuire, M. A.; Bauman, D. E.; Palmquist, D. L.; Nurmela, K.V.V. Trans-
Octadecenoic acids and milk fat depression in lactating dairy cows. Journal of Dairy Science, v. 81, p. 1251-
1261, 1998.
Hagnestam-Nielsen, C.; Emanuelson, U.; Berglund, B.; Strandberg, E. Relationship between somatic cell
count and milk yield in different stages of lactation. Journal of Dairy Science, v. 92, p. 3124-3133, 2009.
Heck, J. M.; Van Valenberg, H. J. F.; Dijkstra, J.; Van Hooijdonk, A. C. M. Seasonal variation in the Dutch
bovine raw milk composition. Journal of Dairy Science, v. 92, p. 4745-4755, 2009.
Hurley, W. L. Nutritional factors affecting milk yield and composition. University of Illinois. Disponível em:
www.classes.aces.uiuz.ede/ansci308/factorsnutritional.html Acessado em: 15 de maio de 2004.
IBGE – Instituto Brasileiro de Geografia e Estatística. Disponível em: http://www.sidra.ibge.gov.br. Acesso
em: 22 jun. 2013

116
Kitchen, B. J. Review of the progress of dairy science: bovine mastitis: milk compositional changes and
related diagnostic tests. Journal of Dairy Research, v. 48, p. 167-188, 1981.
Lambertz, C.; Sanker, C.; Gauly, M. Climatic effects on milk production traits and somatic cell score in
lactating Holstein-Friesian cows in different housing systems. Journal of Dairy Science. v. 97, p. 319-329,
2014.
Larson, B. L. Biosynthesis & cellular secretion of milk. In: LARSON, B. L. (ed), Lactation. Ames: Iowa State
University Press. 1995. p. 129-163.
Marques, L.T., Balbinotti, M.; Fischer, V. Variação da composição química do leite de acordo com a
contagem de células somáticas. In: Congresso Panamericano de Qualidade do Leite e Controle de Mastite, 2,
2002, Ribeirão Preto. Anais... Ribeirão Preto: FEPALE, 2002. 1 CD ROM.
Miller, R. H.; Emanuelsson, U.; Persson, E.; Brolund, L.; Philipsson, J.; Funke, H. Relationships of milk
somatic cell counts to daily milk yield and composition. Acta Agriculturae Scandinavica, v. 33, p. 209-223,
1983.
Mitchell, G. E.; Rogers, S. A.; Houlihan, D. B. The relationship between somatic cell count, composition and
manufacturing properties of bulk milk. 2. Composition of farm bulk milk. Australian Journal of Dairy
Technology, v. 41, p. 9-12, 1986.
Monardes, H. Programa de pagamento de leite por qualidade em Quebec, Canadá. In: Simpósio Internacional
sobre Qualidade do Leite, 1., 1998, Curitiba. Anais... Curitiba: Universidade Federal do Paraná, 1998. p.40-
43.
Montoya, M. A.; Pasqual, C. A.; Finamore, E. B. Panorama da produção leiteira no Rio Grande do Sul:
perspectivas e gestão nas propriedades no Corede Produção. In: Fontes de crescimento da produção de leite:
um enfoque no COREDE. Passo Fundo: Universidade de Passo Fundo, 2014. p. 25-38.
Munro, G. L.; Grieve, P. A.; Kitchen, B. J. Effects of mastitis on milk yield, milk composition, processing
properties and yield and quality of milk products. Australian Journal of Dairy Technology, v. 39, p. 7-16,
1984.
Noro, G.; González, F. H. D.; Campos, R.; Dürr, J. W. Fatores ambientais que afetam a produção e a
composição do leite em rebanhos assistidos por cooperativas no Rio Grande do Sul. Revista Brasileira de
Zootecnia, v. 35, p. 1129-1135, 2006.
Mühlbach, P. R. F. Nutrição da vaca em lactação e a qualidade do leite. In: Simpósio de Bovinocultura de
Leite, 2003, Chapecó. Sociedade Catarinense de Médicos Veterinários – Núcleo Oeste. Disponível em:
http://www.nucleovet.com.br/simposio.htm.
Murphy, J. J.; O’Mara, F. Nutritional manipulation of milk protein concentration and its impact on dairy
industry. Livestock Production Science v. 35, p 117-134, 1993.
National Research Council (NRC). Nutrient requirement of dairy cattle. Washington DC.: National Academy
Press. 2001.
Palmquist, D. L. et al. ADSA foundation symposium: milk fat synthesis and modification. Journal of Dairy
Science, v. 76, p. 1753-1771, 1993.
Philpot, W.N. Importância da contagem de células somáticas e outros fatores que afetam a qualidade do leite.
In: Simpósio Internacional sobre Qualidade do Leite, 1., 1998, Curitiba. Anais… Curitiba: UFPR, 1997. p.
28-35.
Prada E Silva, L. F.; Pereira, A. R.; Machado, P. F., Sarriés, G. A. Effects of somatic cell levels on milk
components II-lactose and total solids. Brazilian Journal of Veterinary Research and Animal Science, v. 37, p.
330-333, 2000.
Ribas, N.P.; Almeida, R.; Marcondes, E.A. Estudo de alguns fatores de meio sobre as produções de leite,
gordura e proteína em vacas da raça Jersey no Estado do Paraná. In: Reunião Anual da Sociedade Brasileira
de Zootecnia, 36, 1999, Porto Alegre. Anais... Porto Alegre: SBZ, 1999 p.159.

117
Ribas, N.P. et al. Produção diária de leite, porcentagens de gordura e proteína em vacas da raça Holandesa no
Estado do Paraná. Revista Batavo, v.8, p.26-33, 2001.
Ribas, N. P.; Hartmann, W.; Monardes, H. G.; Andrade, U. V. C de. Sólidos totais do leite em amostras de
tanque nos estados do Paraná, Santa Catarina e São Paulo. Revista Brasileira de Zootecnia, v. 33, p. 2343-
2350, 2004.
Schutz, M. M.; Hansen, L. B.; Steuernagel, G. R. Variation of milk, fat, protein and somatic cells for dairy
cattle. Journal of Dairy Science, v. 73, p. 484-493, 1990.
Sutton, J. D. Altering milk composition by feeding. Journal of Dairy Science. v. 72, p. 2801-2814, 1989.
Weiss, D.; Hilger, M.; Meyer, H. H. D.; Bruckmaier, R. M. Variable milking intervals and milk composition.
Milchwissenschaft, v. 57, p. 246-249, 2002.

118
Acidose ruminal subaguda: monitoramento e prevenção nos rebanhos
leiteiros
Mirela Noro
Giovani Noro
O rúmen é o compartimento anatômico do trato gastrointestinal responsável pelo processo
digestivo fermentativo, nos ruminantes. Nele os processos bioquímicos fermentativos são
realizados por uma ampla variedade de microrganismos, como protozoários, fungos, e
especialmente as bactérias. As enzimas presentes na parede e citoplasma bacteriano, assim
como aquelas liberadas no fluido ruminal, são as principais responsáveis pelos processos
fermentativos, dos quais são originados substratos energéticos, nitrogenados e algumas
vitaminas, que serão utilizados como nutrientes pelo hospedeiro. Desta forma, os
microrganismos ruminais vivem em simbiose com a vaca leiteira, possibilitando a
degradação de substratos que não poderiam ser degradados por mamíferos não-ruminantes.
Dos processos fermentativos ruminais são produzidas substâncias que baixam o pH ruminal
(caráter ácido), como os ácidos graxos voláteis (acetato, butirato, propionato, entre outros) e
ácido láctico; e substâncias alcalinas como os compostos nitrogenados (ex. amônia [NH4]).
Quando o pH do rúmen se acidifica (pH< 5,8) ocorrem alterações nos processos
fermentativos, e quando os valores baixam de pH< 5,5 ocorrem alterações não somente no
ambiente intrarruminal, como também na sua parede, assim como alterações sistêmicas, que
culminam no quadro de acidose ruminal subaguda (SARA). Este transtorno afeta uma alta
porcentagem de vacas nos sistemas produtivos leiteiros e ocasiona perdas produtivas e de
saúde animal, motivo pelo qual abordaremos as generalidades deste transtorno, seu impacto
nos sistemas produtivos leiteiros, assim como o seu monitoramento e prevenção a nível
populacional.
O que é o pH ruminal e como ele varia ao longo do dia na vaca leiteira
O pH, denominado potencial de hidrogênio, é a expressão para determinar concentração de
hidrogênio [H+] numa solução. Seus valores variam entre 0 a 14, sendo o valor pH 7,0 o
ponto neutro. À medida que o pH fica com valores inferiores a 7,0 ele tende a acidificação e
com valores maiores a 7,0 tende a alcalinização.

119
O pH pode ser calculado pela fórmula: pH= −log [H+]. Como sua fórmula considera o
logaritmo negativo na base 10 (log10), os valores altos de pH indicam que na solução temos
poucas moléculas de H+ e com valores baixos pH temos muitas moléculas de H+. Como
exemplo, quando no rúmen o valor de pH diminui em 1,0 ponto (ex. pH 7,0 quando diminui
a pH 6,0), temos 10 vezes mais H+, e quando diminuímos 2,0 pontos (ex. pH 7,0 para pH 5,0)
temos um aumento de 100 vezes na concentração de H+. Assim podemos inferir que uma
leve mudança no pH ruminal está associada a aumentos consideráveis na concentração de H+
no rúmen.
Diversos estudos têm proposto valores considerados “fisiológicos” para o pH ruminal, porém
mais que um valor em si o mais relevante é a estabilidade do pH ruminal ao longo do tempo.
Um rúmen saudável deve ter um pH > 5,8 até 7,0. Valores maiores que 7,0 seriam de alcalose
ruminal e menores a 5,6 de acidose ruminal. Valores de pH menores ou iguais a 5,8 afetam
a fermentação das bactérias celulolíticas, as quais trabalham eficazmente entre pH 6,2 a 6,8.
Deste modo, mesmo com valores de pH sendo considerados fisiológicos teríamos perdas
produtivas nas vacas por uma redução na fermentação da fibra dietética. Por outro lado, o pH
ruminal varia ao longo do dia associado ao comportamento ingestivo dos ruminantes,
composição e manejo dietéticos; e nas vacas a pastoreio associado às variações diárias na
composição da pastagem. Nos sistemas de produção em base a ração totalmente misturada
(TMR) observa-se uma redução nos valores de pH ruminal entre 3 a 4 horas após a ingestão
da ração, entretanto devido a que as vacas apresentam um comportamento ingestivo diurno,
existe uma tendência a que os valores de pH ruminal descendam com o transcurso do dia.
Portanto, encontraremos valores tendendo a neutralidade (pH 6,5-7,0) durante a madrugada
e período matutino, associado ao maior tempo de ruminação noturna; e tendendo a
acidificação (pH< 6,2) durante o período vespertino e anoitecer, associado as maiores taxas
fermentativas pelo maior consumo de alimento (Figura 1).
O que é a acidose ruminal subaguda
A acidose ruminal subaguda (SARA) é um transtorno digestivo subclínico ou inaparente
associado ao consumo de concentrados energéticos, alimentos altamente fermentáveis, ou
com baixa capacidade tampão (buffer). Está caracterizada por pH ruminal ácido ( 5,5), que
impacta negativamente na produção e saúde das vacas. Apresenta uma alta prevalência nos

120
sistemas de produção leiteira onde as vacas são suplementadas com alta proporção de
concentrados e baixa fibra efetiva (eF). Afeta principalmente as vacas de alta produção no
início da lactação, onde as papilas ruminais não se desenvolveram completamente, e no pico
do consumo de alimento, associada às altas taxas fermentativas. Também afeta rebanhos
onde ocorreram mudanças de uma dieta com baixa densidade energética para uma com alta
densidade e baixa eF. Tradicionalmente a SARA era reconhecida por afetar a rebanhos
estabulados em sistema de dieta com alto concentrado. Porém, a SARA tem sido
diagnosticada com alta incidência em rebanhos pastoris de diversas partes do mundo,
associada ao consumo de pastagens com baixo conteúdo de eF e/ou alto conteúdo de
carboidratos solúveis, como observado em gramíneas temperadas. Por este motivo, nos
sistemas produtivos leiteiros do sul do Brasil, durante o período de inverno e primavera onde
aumenta o uso dos sistemas pastoris, a prevalência de SARA pode ser elevada, associada ao
uso de pastagens de aveia e azevém. Também tem sido observada grande incidência de
SARA em rebanhos em pastagens tropicais, quando há suplementação de grandes
quantidades de concentrado (acima de 0,5% PV por refeição), principalmente em período
onde os animais estão sob stress calórico.
Figura 1. Variação (X ± DP) do pH ruminal ao longo do dia em vacas a
pastoreio de azevém perene (PB: 18,3% MS, EM: 2,7 Mcal/kg MS).
Adaptado de Scandolo et al., 2007 (Contreras & Noro, 2010)
Ainda não existe um consenso respeito aos valores de pH ruminal para diagnosticar SARA.
Alguns pesquisadores diagnosticam SARA quando os valores de pH ruminal são < 5,5
durante 4 ou mais horas por dia. Outros diagnosticam SARA quando o pH ruminal está

121
abaixo de 5,6 pelo menos por 3 horas ao dia. Enquanto outros são mais extremistas e
consideram SARA quando os valores de pH ruminal se encontram menores a 6,0 por mais
de 4 horas ao longo do dia, situação que inclui os animais com média de pH ruminal ao longo
do dia maiores a 6,25. Tomando por base a maioria dos estudos no tema, consideraremos
SARA quando os valores de pH ruminal encontrarem-se entre 5,0 a 5,5. Valores de pH entre
> 5,5 a 5,8 serão considerados marginais, e o pH “fisiológico” será considerado > 5,8 até 7,0.
Valores > 7,0 são considerados alcalinos, e podem ser observados em animais em jejum ou
anorexia. Os valores de pH ruminal < 5,0 são observados nos quadros clínicos de acidose
aguda ou hiperaguda conhecida como acidose láctica ruminal. Por ser um quadro subclínico,
a SARA é subdiagnosticada. Em rebanhos afetados por SARA podem observar-se quadros
de acidose crônica, os quais estão associados ao excessivo consumo de carboidratos por um
longo período de tempo, normalmente com adequada quantidade de eF, sendo assim uma
forma de acidose subaguda persistente. Neste quadro a população de bactérias celulolíticas
diminui e grandes quantidades de microrganismos utilizadores e produtores de lactato são
encontrados. Na acidose crônica a grande quantidade de ácidos produzidos no rúmen
estimula a proliferação das papilas ruminais, podendo resultar em paraqueratose, hiporexia e
hipomotilidade ruminal. A sobrecarga ácida persistente pode reduzir a eficiência metabólica
e o desempenho geral do animal. Outros achados no rebanho são a alta prevalência de
laminite e eventualmente poliencefalomalacia. Os fatores de risco para SARA estão
associados a fatores que alteram o padrão de fermentação ruminal como a dieta (quantidade
e tipo de carboidratos não fibrosos; quantidade e tipo de fibra, tamanho de partícula, seleção
da dieta), manejo dietético (tipo e frequência de alimentação), o ambiente (conforto,
temperatura e umidade ambiental), assim como fatores que afetem a absorção dos ácidos
graxos voláteis (AGV). Alguns destes fatores de risco serão abordados no capítulo de
monitoramento para prevenção de SARA.
A acifidicação do rúmen resulta de três processos que podem ou não estar integrados: (1)
Efeito químico/microbiológico, pelo alto consumo de carboidratos altamente fermentáveis,
alta taxa fermentativa e aumento de AGV no rúmen; (2) Efeito físico/químico, pela baixa
ingestão de eF, com baixa taxa de ruminação, que baixa a capacidade tampão proveniente
dos fosfatos e bicarbonatos da saliva; e (3) Efeito físico pela baixa capacidade de absorção
dos AGV pelas papilas ruminais em vacas não adaptadas a dieta com alta densidade

122
energética. Na forma clássica a SARA ocorre quando no rúmen, o amido e os açúcares da
dieta sofrem uma rápida fermentação, sem uma adequada capacidade tampão, aumentando a
taxa de crescimento das bactérias amilolíticas em relação às celulolíticas. Consequentemente
aumenta a produção dos AGV, diminuindo o pH ruminal. Na fase inicial da fermentação,
aumenta a produção do ácido propiônico (de 20% para aproximadamente 40%), e diminui a
do ácido acético associado à redução da degradação da fibra quando o pH baixa de 5,8. Nos
quadros de SARA e de acidose crônica, as proporções dos ácidos propiônico e butírico
aumentam e as de acético diminuem. É assim que no rúmen saudável a relação
acetato:propionato é de aproximadamente 2,2:1. Quando os valores de pH ruminal seguem
baixando pela maior produção de ácidos e disponibilidade de carboidratos, o Streptococus
bovis cresce exponencialmente convertendo o amido e a glicose em ácido láctico e outros
ácidos orgânicos, como valérico, fórmico e succínico. Este aumento dos ácidos acidifica o
pH ruminal a aproximadamente 5,4 que por si leva a uma anorexia, diminui o crescimento e
ocasiona a morte das bactérias lactolíticas que transformam o ácido láctico em propiônico,
assim como de outras populações microbianas. Com a morte das bactérias ruminais, que na
sua maioria são Gram-negativas, ocorre liberação de uma grande quantidade de endotoxinas
lipopolissacarídicas da parede celular ao ambiente ruminal. Estas endotoxinas quando
absorvidas produzem alterações hemodinâmicas e inflamatórias sistêmicas no animal. A
progressiva acidificação do rúmen, especialmente associada ao aumento dos AGV, leva a
hipotonia ruminal, sendo o butirato o principal responsável por este efeito, e quando o pH
atinge valores próximos a 5,0 ocorre uma atonia ruminal. Quando o ácido láctico produzido
pelo S. bovis acumula-se no rúmen, ocorre uma redução do pH ruminal a valores inferiores
a 5,0, com substituição da população de S. bovis por Lactobacillus, que são mais resistentes
a pH ácidos, caracterizando uma acidose láctica ruminal, transtorno clínico que não será
discutido neste trabalho. A hipomotilidade ruminal leva a uma diminuição da ruminação, e
menor produção de saliva, impedindo o efeito tampão. Por outro lado, quando ocorre
dissociação dos ácidos produzidos no rúmen, incrementa a osmolaridade ruminal e intestinal
a valores maiores que a do sangue ( 300 mOsm), o que se traduz em efeito osmótico com
passagem de água do sangue para o rúmen e intestino, gerando fezes desde mais fluidas até
diarreicas.

123
Sinais e impacto da SARA no sistema produtivo
A SARA impacta negativamente no rebanho por afetar aspectos produtivos e de saúde. Em
nível de rebanho a visualização de alguns achados como: (1) taxa de descarte anual maior a
30% (> 8% nos 60 dias pós-parto); (2) incidência de deslocamento de abomaso maior a 3%;
(3) baixo consumo de matéria seca no pós-parto; (4) escore de fezes heterogêneo entre vacas;
e (5) gordura láctea menor a 3,5%, sugerem alta prevalência de SARA. A seguir citaremos
alguns dos impactos da SARA no sistema produtivo.
Degradação da dieta
A acidificação ruminal (< 5,8) diminui a digestão da fibra e da proteína ruminal, por
detrimento das bactérias celulolíticas e proteolíticas. Como resultado ocorre perda de fibra
nas fezes e aumento de aporte de proteína passante, que pode ser absorvida a nível intestinal,
porém em detrimento da síntese de proteína microbiana.
Síndrome da baixa gordura láctea
Ocorre associada à diminuição da biohidrogenação de ácidos graxos insaturados presentes
na digesta e da síntese de acetato ruminal. A redução da biohidrogenação aumenta
percentualmente os ácidos graxos trans no sangue, inibindo a síntese do gordura láctea.
Muito comum onde dieta contém adição de óleos, principalmente de soja, monensina e baixa
eF.
Rumenite e paraqueratose
A exposição prolongada do epitélio ruminal ao baixo pH provoca rumenite, atrofia e necrose
das papilas ruminais e eventualmente hiperparaqueratose, impactando negativamente na
absorção dos AGV. Além dos ácidos orgânicos produzidos no rúmen, vários outros fatores
tóxicos (histamina, tiramina, etanol e endotoxinas) podem contribuir para a apresentação
desta afecção. Por outro lado, a rumenite é uma via de ingresso de bactérias e fungos para a
corrente circulatória.

124
Abscessos sistêmicos
Quando a mucosa ruminal perde sua integridade, também perde sua capacidade de atuar
como barreira entre o ambiente ruminal e o sangue. Isto permite que bactérias patogênicas
como Fusobacterium necrophorum penetrem no epitélio ruminal e, através da veia porta,
colonizem outros órgãos. Estas bactérias podem causar abscessos hepáticos, e em alguns
casos provocar peritonite localizada. Posteriormente, os abscessos podem ocorrer em outros
órgãos como pulmões, coração, rins, pele ou articulações.
Laminite
Nos quadros de acidose ruminal as bactérias que normalmente utilizam a histidina, estão
inibidas ou morrem. Nestes casos a Allinella histaminiformans, bactéria resistente a pH
ácidos, utiliza a histidina e libera histamina, que poderia contribuir na apresentação de
laminite. Entretanto, a etiologia deste quadro ainda não está completamente elucidada.
Porém, em rebanho com SARA se observa uma alta prevalência de claudicações.
Síndrome da veia cava caudal
Associada aos abscessos hepáticos, alguns animais podem apresentar congestão crônica
passiva na veia cava caudal com formação de trombos que provocam hipertensão pulmonar,
edema periférico e ascite. A formação dos trombos na veia cava cranial pode causar distensão
da jugular e edema local, que pode culminar no desenvolvimento de aneurismas que
ocasionam rupturas, provocando hemoptise aguda e morte.
Outros impactos
Outro achado nos rebanhos com acidose ruminal é o aumento das taxas de reposição por
aumento das taxas de descarte, a ocorrência de poliencefalomalacia por produção de
tiaminase pelas bactérias ruminais, e alta prevalência de deslocamento de abomaso. Além
disso, podem ocorrer quadros de morte súbita nos animais que cursaram com quadros prévios
de acidose, devido a uma hipersensibilidade anafilática as endotoxinas absorvidas.

125
Monitoramento do rebanho: medidas para o diagnóstico e prevenção
O monitoramento do rebanho para diagnóstico e prevenção da SARA deve ser realizado de
forma integral no sistema produtivo e considerando os diferentes espectros da produção e da
saúde populacional. Consequentemente, as medidas a serem tomadas impactam positiva ou
negativamente na produção e incidência de outros transtornos da saúde do rebanho. Neste
texto, abordaremos os principais aspectos que afetam a incidência de SARA, visando a
manutenção da produção e saúde populacional de forma integral.
No monitoramento do rebanho devemos levar em conta os aspectos associados diretamente
às vacas, aspectos de manejo e ambientais, e dar especial atenção à nutrição. Seguindo esta
lógica, abordaremos primeiro os aspectos relacionados ao diagnóstico da SARA,
visualizando aspectos observacionais nos animais, e metodologia analíticas de pH ruminal,
e logo abordaremos aspectos de manejo e nutrição, que são medidas associadas não só ao
diagnóstico, como à prevenção da afecção. Aspectos de ambiente e conforto como parte do
monitoramento e prevenção da SARA não serão abordados neste texto.
Observação das vacas
Em propriedades com dietas ou manejos com alto risco de SARA podemos observar animais
com quadros clínicos de acidose láctica ruminal, manifestada de forma hiperaguda e/ou
aguda, que ocorre em um número reduzido de vacas não adaptadas à dieta ou que consomem
uma grande quantidade de alimento concentrado por erro na distribuição nos cochos ou
homogeneização no mixer. Porém, na sua forma clássica, em rebanhos com SARA observam-
se sinais que podem ser sutis, como diminuição e flutuação no consumo de matéria seca
durante o dia e entre dias, situação bastante difícil de registrar na rotina de um tambo, sem o
uso de monitores comportamentais. Ademais, algumas vacas podem estar mais letárgicas e
apresentar grau variável de diarreia intermitente. No rebanho também podemos observar uma
queda na condição corporal, e a produção de leite tende a aumentar num primeiro momento
e logo pode diminuir. A incidência de deslocamento de abomaso aumenta durante o período
onde a SARA é mais incidente (> 3%), e após algumas semanas podem observar-se abscessos
subcutâneos, ou visualizar-se abscessos hepáticos nas necropsias. Porém, antes de qualquer
observação é necessário contar com os registros da propriedade, para identificar os grupos
de risco, motivo pelo qual é importante formar lotes por categoria fisiológica e nível de

126
produção, para o adequado monitoramento e para formular estratégias de prevenção. A seguir
serão abordados alguns aspectos observacionais nas vacas que indicam alta incidência de
SARA.
Problemas podais
Nos rebanhos com SARA pode-se evidenciar presença de sulcos nos dígitos, alta prevalência
de laminite, e após algumas semanas do quadro de laminite é frequente a manifestação de
úlceras e hematomas de sola e doença da linha branca. Como medida de monitoramento, é
recomendado realizar periodicamente a avaliação do escore de locomoção do rebanho. Ele
permite reconhecer a ocorrência de vacas com claudicação, que poderiam ser decorrentes de
uma laminite crônica.
Comportamento alimentar
Vacas com SARA apresentam um consumo de matéria seca flutuante, com diminuição
transitória, associado á diminuição da motilidade ruminal por aumento da osmolaridade
ruminal. Ademais, quando na dieta existe fibra longa (> 7 cm) especialmente proveniente de
silagens, é possível observar a seleção e rejeição da fibra no cocho. Nestes casos as vacas
selecionam e consomem as partículas mais finas, diminuindo consequentemente as taxas de
ruminação. Porém, quando o feno de qualidade está disponível ad libitum os animais tendem
a consumi-lo. Nestes rebanhos também é frequente ver-se alguns animais consumindo solo
e camas, e em locais com cochos com livre disposição de bicarbonato de sódio, se observa
um alto consumo (> 50 g/ vaca/dia).
Escore de ruminação
Rebanhos com SARA diminuem o tempo de ruminação, que pode indicar uma inadequada
quantidade de fibra na dieta. Normalmente as vacas leiteiras ruminam enquanto descansam
totalizando 6,7 horas de ruminação por dia, tempo que varia entre 3,4–14h dependendo do
tipo e manejo dietético. Normalmente ruminam quando estão deitadas (5,5 h) e menor tempo
quando estão em estação (1 h). Como as vacas de um mesmo lote tendem a sincronizar suas
atividades, o monitoramento da ruminação pode ser feito observando diariamente o rebanho
após a alimentação (40 a 70 minutos da entrega da ração), ocasião em que 50% ou mais das

127
vacas que estão descansando deveriam estar ruminando. No caso das vacas deitadas, mais
que 60% delas deveriam estar ruminando. Deve-se considerar que as vacas ruminam
principalmente durante a noite, porém quando a temperatura sobrepassa os 24ºC as vacas
tendem a alimentar-se pela noite e descansar durante o dia, o que altera o comportamento de
ruminação.
Avaliação visual das fezes
A observação das fezes das vacas entrega informação referente à interação entre a vaca e sua
dieta. Desta forma a avaliação das fezes serve para monitorar a dieta e sua digestibilidade.
Entre os aspectos que devem ser avaliados estão: 1. Avaliação visual e a homogeneidades
das fezes entre as vacas; 2. Escore de fezes; 3. Tamanho de partículas não digeridas nas fezes.
1. Avaliação visual e homogeneidade das fezes entre as vacas. Num grupo de vacas em dias
de lactação e dieta similares, as fezes deveriam ser homogêneas. Quando um grupo de vacas
apresenta fezes heterogêneas pode indicar a ocorrência de SARA e auxilia na melhora da
formulação da dieta e do manejo. Entre os aspectos a ser avaliados estão os “3 Cs”: cor,
consistência e conteúdo. Num rebanho com SARA é possível observar uma alta porcentagem
de vacas com perda da consistência fecal pelo aumento da osmolaridade ruminal e intestinal
(ver avaliação do escore de fezes); aparição de muco e fibrina, associado a lesão intestinal; e
presença de borbulhas, associada a fermentação do grão não digerido na parte posterior do
intestino.
2. Escore de fezes. O escore de fezes avalia a consistência fecal, observando as fezes no solo.
Este parâmetro avalia a qualidade da dieta em termos de carboidratos, fibra, proteína e água,
e sua digestibilidade. O escore 1 está associado a fezes líquidas verdes resultados do
consumo de pastagens de alta qualidade; porém também pode ser observado em vacas com
SARA e em vacas doentes. No escore 2 as fezes são semilíquidas com uma forma definida,
observadas em vacas recém paridas, em pastoreio e com SARA. No escore 3 as fezes têm
forma concêntrica, tipo um vulcão, sendo considerado o escore ideal. No escore 4 as fezes
são cônicas, tendendo a ser secas, sendo observadas em vacas secas com dietas com baixa
proteína e alta fibra. No escore 5 as fezes se apresentam bastante ressecadas, sendo
observadas em vacas doentes desidratadas ou alimentadas com dietas em base a forragens
secos e com muito baixa digestibilidade.

128
3. Avaliação do tamanho das partículas fecais. Indica a digestibilidade da dieta. Pode ser
realizada manualmente, porém idealmente com o uso do analisador de digestão (Nasco). O
método manual pode ser realizado coletando-se amostras de fezes ao pé da vaca com um
copo plástico. Deve-se avaliar amostras de um grupo homogêneo, de ao menos 8 vacas. As
fezes, de cada vaca individualmente, devem ser colocadas numa peneira de cozinha (18 cm
de diâmetro e 10 cm de profundidade), e com um fluxo constante de água lava-se o conteúdo
fecal até que a água filtrada fique translúcida. Após, observa-se a presença de partículas
indigeridas nas fezes, estimando a quantidade de partículas pequenas, médias e grandes,
como grãos não digeridos. A presença de grãos de milho, provenientes de silagem, pode
indicar que foi pobremente picada ou ensilada tardiamente. Grãos de milho com menos de
0,6 cm quando observados em conjunto com presença de fibras vegetais indica rápida taxa
de passagem, que pode estar associada a silagem muito jovem ou insuficiente eF na dieta.
Como diagnosticar a SARA: mensuração do pH ruminal
O único método direto, preciso e sensível para avaliar o grau de acidose ruminal no rebanho
é a determinação do pH no líquido ruminal. Entretanto, a obtenção de uma amostra
representativa é fundamental para que os valores sejam reais. A este respeito se sabe que
amostras obtidas mediante sondagem buço-esofágica ruminal podem sofrer contaminação
por saliva (2-30%), reduzindo a sensibilidade da técnica que consequentemente pode
aumentar o pH do líquido ruminal entre pH 0,6 a 1,7, produzindo falsos negativos de SARA.
Por este motivo as amostragens realizadas mediante ruminocentese são as indicadas para
monitoramento de SARA no rebanho. O volume a ser obtido depende da técnica para a
determinação do pH. Preferencialmente deve-se usar o pHmetro portátil com eletrodo fino,
que permita mensurar o pH em amostras de 3 a 5 mL obtidas em seringas de 5 ou 10 mL.
Quando a placa de leitura do pHmetro é larga o volume de líquido ruminal deve ser maior
para ser colocado num copo e permitir que o eletrodo seja submerso no líquido, necessitando
de volumes entre 5 a 10 mL.
Quando obter as amostras de líquido ruminal?
Devido ao comportamento ingestivo das vacas e as características de fermentação ruminal
ao longo dia, as amostras de líquido ruminal deveriam ser obtidas à tarde, 2 a 4 horas após a

129
ração vespertina com concentrado, ou 4 a 6 horas após o TMR, idealmente após a ordenha
da tarde (após 16:00), ocasião onde os valores de pH estão baixando, e que seguirão
baixando até o anoitecer. Amostras obtidas pela manhã tendem a neutralidade e geram
resultados falsos negativos de SARA.
Quais e quantos animais amostrar?
As vacas a serem monitorados são as que têm maior risco de SARA. Entre elas estão as
vacas em período de transição pós-parto (5 a 21 dias pós-parto) e as vacas no pico de
produção láctea e ingestão de alimento (50 a 120 pós-parto). Para fins de validade
diagnóstica em rebanhos grandes e médios deveriam coletar-se ao menos 20 vacas por
grupo, e em rebanhos pequenos pelo menos 8 vacas de cada grupo. Quando no rebanho o
número de vacas de cada grupo é menor a 8, recomenda-se coletar todas as vacas do grupo.
É importante que as vacas selecionadas para amostragem sejam homogêneas, e
representativas do grupo ao qual pertencem.
Como interpretar os resultados de pH ruminal?
Para o adequado diagnóstico da ocorrência de SARA no rebanho, deve-se calcular a
porcentagem de vacas com SARA, ou com valores marginais, em cada grupo de risco.
Consideram-se o grupo como positivo a SARA quando 25% ou mais das vacas apresentam
valores pH< 5,5, e marginais quando 33% ou mais das vacas tem valores pH< 5,8. Grupos
negativos são aqueles nos quais menos de 33% das vacas apresentam valores pH< 5,8. Esta
classificação serve para diagnosticar um rebanho com SARA ou risco de SARA (valores
marginais), assim como auxilia na triagem dos fatores que estão associados na ocorrência
de SARA no rebanho (Tabela 1).
Outros parâmetros a ser observados
A composição láctea em propriedades que fazem uso do controle leiteiro permite avaliar a
nutrição e o estado de saúde ruminal das vacas. Em rebanhos com SARA a gordura láctea
tende a diminuir (< 3,5%), e pode ocorrer inversão entre a gordura e a proteína lácteas.
Nestes rebanhos, mais de 10% das vacas apresenta gordura láctea 0,4% abaixo do valor da
proteína láctea. Alguns autores consideram o cociente entre gordura:proteína lácteas < 1,0

130
como indicativo de SARA, porém este ponto de corte deve ser validado nos nossos
rebanhos, visto que o sistema de amostragens realizado a nível de propriedade pode gerar
erros metrológicos.
Monitoramento do manejo e da dieta para atuar preventivamente
O monitoramento e prevenção da SARA está associado diretamente a aspectos que auxiliam
na manutenção do pH ruminal dentro de valores ótimos. Entre eles o de maior impacto é a
dieta. Porém, como o valor nutricional da dieta não é estático, e a dieta formulada
normalmente é diferente da dieta consumida, é muito difícil predizer em quais intervalos de
valores o pH das vacas se mantêm. Por este motivo é importante mensurar o pH do líquido
ruminal, com a finalidade de diagnosticar e prevenir os quadros de acidificação. Os valores
ótimos para a adequada fermentação da fibra e saúde ruminal se encontra entre 6,0 a 7,0.
Como medidas para manter o pH ruminal entre valores ótimos, estão a adoção de uma série
de estratégias, em conjunto, visto que cada uma de forma isolada não atingirá o objetivo de
manutenção do pH dentro de valores adequados, e estáveis ao longo do dia. Todos os aspectos
associados à manutenção da saúde e produção devem ser tomados considerando o estado
fisiológico e a produção das vacas. Assim, é necessário agrupar as vacas por estado
fisiológico e produção para que possam receber a dieta de acordo com seus requerimentos.
A seguir abordaremos alguns aspectos a ser considerados no manejo, na dieta e ao uso de
aditivos para prevenir quadros de SARA.
Tabela 1. Interpretação de valores alterados de pH ruminal em grupos de vacas
Vacas 50 – 120 DEL
+ -
Vacas < 20 DEL
+ Problemas com a ração e
/ou periparto
Problema no periparto
- Problema da ração Rebanho normal DEL= dias em lactação. Grupo positivo quando 25% ou mais das vacas com pH< 5,5
Grupo negativo quando menos de 33% das vacas com pH< 5,8 (Nordlund et al., 1995)
Dieta no período de transição pré-parto
A prevenção da SARA inicia desde o período de transição pré-parto (-21 dias até o parto).
Neste período as vacas devem ser adaptadas à dieta do pós-parto com dois objetivos

131
principais no que diz respeito a SARA: 1. Estimular o crescimento das papilas ruminais; 2.
Adaptação a dieta do pós-parto. As dietas com alta densidade energética devem ser ofertadas
de forma gradual e progressiva, especialmente a animais manejados de forma extensiva,
assim como as vacas secas. A dieta das vacas no período seco é de baixa qualidade, com
baixas taxas fermentativas, o que culmina numa redução do tamanho das papilas ruminais
em 50%. Desta forma, é fundamental incluir na dieta das vacas no período de transição
alimentos energéticos que aumentem as taxas fermentativas, para estimular o
desenvolvimento das papilas. O tamanho ótimo das papilas é atingido em 4 a 6 semanas após
a inclusão de dietas energéticas, e à medida que as papilas crescem ocorre um aumento da
capacidade de absorção dos AGV. Deste modo, a adaptação a uma nova dieta, como ocorre
no pós-parto, deve ser feita pelo menos por 15 dias prévios ao parto.
Composição da dieta. Para avaliar os fatores de risco para SARA, assim como formular
dietas para prevenção de SARA, é necessário contar com a informação das propriedades
químicas (MS, digestibilidade, energia, CNF, FDN) e físicas (tamanho partícula, tipo e grau
de processamento dos alimentos energéticos) da dieta, cociente forragem: concentrado, e
manejo dietético.
Manejo dietético: adaptar faz diferença. A priori, qualquer mudança da dieta deve ser feita
com um período de adaptação gradual a nova dieta, idealmente por 14 a 21 dias com
aumentos gradativos a cada 5 a 7 dias.
Maior número de rações diárias minimiza picos de acificação. O fracionamento da dieta
várias vezes ao dia, mínimo 2 a 3 rações diárias, além de estimular o consumo (quando o
alimento está disponível por mais de 20 horas ao dia), minimiza os picos de acidificação
ruminal por fracionar o consumo.
TMR: forma eficaz para entregar uma dieta balanceada. O uso da dieta totalmente misturada
(TMR) favorece em dois aspectos principais a saúde e produção da vaca leiteira: 1. Aumenta
o consumo de matéria seca; 2. Permite a incorporação de volumosos fibrosos na dieta, por
diminuir a seleção do concentrado pelas vacas, estimulando consequentemente a salivação,
e por consequência a manutenção do pH ruminal. Neste sistema é necessário monitorar a
ocorrência de seleção da dieta quanto a presença de partículas grosseiras de alimento.
O cocho é importante! Dois aspectos a respeito do cocho impactam no risco de SARA: (1)
Altura do cocho, pois vacas que ingerem alimento com a cabeça baixa, além de diminuir a

132
perda de alimento, produzem 17% mais saliva. Por este motivo os cochos deveriam estar
entre 5 e 15 cm da linha da sola do casco. (2) Espaço linear de cocho por vaca: para minimizar
os efeitos da dominância, em sistemas estabulados com uso de TMR o espaço linear por vaca
deve ser de 75 cm (linha unilateral), e quando o acesso à ração é limitado (baixo número de
rações por dia), é necessário contar com 80-100 cm linear/vaca.
Como minimizar altas taxas fermentativas por excesso de carboidratos não fibrosos (CNF)
de rápida degradabilidade? A dieta das vacas leiteiras deve permitir a expressão do mérito
genético, tendo em conta os desafios que as altas exigências trazem consigo. Uma dieta
adequada otimiza o cociente entre forragem e concentrado em termos de quantidade e
sincronismo de degradação. O nível de CNF da dieta varia com o desafio a que o sistema
quer impor a vaca leiteira, assim como da fonte e digestibilidade do carboidrato.
Normalmente o CNF da dieta varia entre 30-40% MS, com uma taxa máxima de 45-50%.
Valores ótimos são entre 38-40%, aceitáveis entre 35- 38% e mínimo entre 25-30%, estes
últimos apesar de não ser acidogênicos, não são adequados para que as vacas expressem seus
méritos genéticos, e podem gerar risco de transtornos energéticos em vacas em início de
lactação. Também é importante limitar o conteúdo de concentrados amiláceos de rápida
degradabilidade, pois são altamente acidogênicos e de difícil sincronização com a
degradabilidade da fibra dietética. Ademais, o processamento dos grãos impacta nos
processos fermentativos. A degradação dos grãos é em forma decrescente maior nos grãos
úmidos, que finamente moídos, que esmagados, que triturados, que inteiros secos. Desta
forma, deve-se evitar o uso de grãos finamente moídos (milho e sorgo são exceção), por
apresentarem rápida degradabilidade. Da mesma forma, o tipo de carboidrato afeta a
degradabilidade, sendo maior em açúcares, que amido, que pectina, que celulose. E o tipo de
grão utilizado, sendo maior a degradabilidade da aveia, que trigo, que cevada, que milho, que
sorgo.
Como estimular a produção de saliva (efeito buffer) pelo uso de fibra efetiva e tamanho de
partículas? O principal estimulo para produção de saliva nos ruminantes é o consumo de eF,
que estimula a ruminação. Em torno de 60% das vacas ruminam 2 horas após a ingestão da
ração, e quando existe uma adequada quantidade de fibra na dieta a produção de saliva supera
180 L ao dia. Para garantir uma adequada ruminação, a ração deve conter fibra em detergente
neutro (FDN) maior a 28% (27-33% MS, NRC 2001), com um mínimo 15% FDN

133
proveniente de forragem (70–80% da FDN). Idealmente, no início da lactação deveria ter um
19% FDN proveniente da forragem, com no mínimo FDN efetivo de 21% MS, considerando
que a eF do concentrado é de aproximadamente 0,33 e a eF da forragem é de 1,0. Porém,
deve-se ter cuidado com a limitação física que o consumo de fibra representa quando se
atinge 1,1 a 1,2% do peso vivo em FDN de forragem no caso de multíparas, e a 0,78 % PV
na 1º semana da lactação no caso das primíparas. Como o FDN é um parâmetro químico e
não físico, também é fundamental considerar o tamanho das partículas de forragem. As
partículas longas têm 3 funções principais: 1. Estimular a ruminação e consequentemente a
manutenção do pH ruminal; 2. Formar uma camada flutuante que retêm as partículas menores
no rúmen, melhorando a degradabilidade ruminal; e 3. Aderir a parede ruminal estimulando
a absorção dos AGV pelas papilas ruminais. Para que a fibra estimule a ruminação deve
conter partículas longas entre 2 cm e 7 cm, e por regra, um 20% das partículas da forragem
deveriam ter mais de 3,75 cm. Partículas maiores a 7 cm, estimulam a seleção por parte da
vaca.
Uma forma de avaliar o tamanho das partículas de forragem é mediante a o uso do separador
de partículas (peneira) Penn State. O objetivo da peneira é avaliar a quantidade partículas
longas assim como a distribuição das partículas da ração por tamanho, permitindo verificar
a eficiência do sistema de mistura do vagão de alimentação em dietas frescas. A peneira
também pode ser utilizada na avaliação da seletividade dos animais, quando usada para
avaliar as sobras do cocho em amostras pareadas com a dieta ofertada. Para o monitoramento
da dieta do rebanho, a avaliação da dieta com a peneira deveria ser realizada semanalmente,
após a distribuição da ração nos cochos (prévio a alimentação). Para avaliar o comportamento
de seletividade das vacas devem-se aferir os resíduos da ração às 4, 8, 12 horas depois da
alimentação. As duas bandejas superiores da peneira (19 e 8 mm de orifício) permitem
estimar a porcentagem de eF da dieta (Tabela 2).

134
Tabela 2. Recomendações para o tamanho de partículas de forragem e TMR na peneira Penn State
(Hutiens, 2012)
Tamiz
Silagem de milho
(processada) 20
mm
Silagem de milho (não
processada) 10 mm
Silagem
pré-secada TMR
Peneira 19 mm > 10 < 5 > 20 > 10
Peneira 8 mm > 60 >50 40 a 50 40-50
Peneira 1,18 mm < 20 < 30 < 35 < 35
Fundo < 5 < 10 < 10 < 20
Quais aditivos nutricionais podem auxiliar na manutenção do pH ruminal? Em rebanhos
onde o uso de alimentos de alta densidade energética supera os 6 kg/vaca/dia, ou com
produções que superam os 20 L, faz-se necessário o uso de aditivos nutricionais que modulam
a fermentação e colaboram na manutenção do pH ruminal. Entre os aditivos nutricionais
temos os: 1. Químicos, como os buffers ruminais [NaHCO3] e alcalinizantes ruminais
[MgO]); 2. Antibióticos (ionóforos [monensina, lasalocida], virginamicina e tilosina); 3.
Microbianos, como os extratos de leveduras, leveduras vivas de Saccharomyces cerevisiae,
Aspergillus oryzae, probióticos (Selomonas ruminatium, Megasphera elsdenii, Enterococcus
faecium, Lactobacillus plantarum).
1. Químicos. Os tampões e alcalinizantes afetam a fermentação ruminal porque mantêm por
mais tempo ao longo do dia o pH ruminal a valores superiores a pH 6,0. Consequentemente
melhoram a degradação da fibra, especialmente em dietas com muita silagem (pH< 4,0),
dietas muito úmidas, forragens finamente picadas, alto uso de concentrado, grãos finamente
moídos, e concentrado fornecido em poucas refeições ao longo do dia. Normalmente tem-se
utilizado o bicarbonato de sódio associado ao óxido de magnésio, na proporção de 3:1, na
concentração de 2% no concentrado ou 1 % na dieta total. Recentemente têm surgido alguns
produtos tamponantes compostos por conchas marinhas.
2. Antibióticos. Entre eles temos os ionóforos (monensina, lasalocida) e a virginamicina. Eles
atuam modificando o transporte de íons na membrana celular das bactérias Gram-positivas,
como Streptococcus bovis e Lactobacillus sp, favorecendo as bactérias produtoras de
propionato. A ação deles permite melhorar a degradabilidade da fibra e auxiliam na
manutenção do pH ruminal, reduzindo o risco de acidose. Também favorecem o metabolismo
energético e nitrogenado, de modo que tendem a aumentar a produção láctea. Os resultados
dos antibióticos nas vacas dependem da dieta, e da forma da entrega deles na dieta, tendo

135
melhores resultados quando entregues não misturado no alimento (sobre o alimento) quando
comparado ao uso em TMR.
3. Probióticos. As leveduras vivas atuam mediante competição com a Streptococcus bovis
pela glicose, reduzindo a disponibilidade deste substrato, além consumir o O2 ruminal que é
tóxico para a flora fibrolíticas. Como efeito, diminuem a produção do ácido láctico,
auxiliando na manutenção do pH ruminal. Seus efeitos são observados especialmente quando
o conteúdo de carboidratos rapidamente fermentáveis da dieta é a alto.
Finalmente, apesar de na atualidade existir uma ampla gama de alternativas para modular a
fermentação ruminal, o uso delas depende do manejo, dieta e custo-benefício. Nenhuma
estratégia de prevenção é eficaz quando utilizada de forma isolada e sem monitoramento
constante e integral do sistema produtivo.
Referências
Bach A, C Iglesias, M Devant. 2007. Daily rumen pH pattern of loose-housed Dairy cattle as affected by
feeding pattern and live yeast supplementation. Anim. Feed Sci. & Tech. 136, 146-153.
Calsamiglia S, L Castillejos, M Busquet. 2005. Estrategias nutricionales para modificar la fermentación
ruminal en vacuno lechero. En: XXI Curso de Especialización FEDNA, Madrid, España.
Calsamiglia S, M Busquet, PW Cardozo, L Castillejos, A Ferret. 2007. Invited Review: Essential Oils as
Modifiers of Rumen Microbial Fermentation. J. Dairy Sci. 90, 2580-2595.
Contreras, PA, Noro, M. Rumen: Morfofisiología, trastornos y modulación de la actividad fermentativa.
Valdivia: América, 2010.
Krause KM, GR Oetzel. 2006. Understanding and preventing subacute ruminal acidosis in dairy herds: a
review. Anim. Feed Sci. Technol. 126, 215-236.
Kolver ES, MJ de Veth. 2002. Prediction of ruminal pH from pasture-based diets. J. Dairy Sci. 85, 1255-
1266.
Nordlund K, E Garrett, G Oetzel. 1995. Herd-based rumenocentesis: A clinical approach to the diagnosis of
subacute rumen acidosis. Comp. Cont. Edu.: Food Anim. S48, 48-56.
O’Grady L, ML Doherty, F Mulligan. 208. Subacute ruminal acidosis (SARA) in grazing Irish dairy cows.
The Veterinary J. 176, 44-49.
Varga G, E Kolver. 1997. Microbial and Animal Limitations to Fiber Digestion and Utilization. J. Nutr. 127,
819S-823S.

136
Manejo nutricional da vaca leiteira para otimizar a composição do leite
Jessica Karina Poncheki
Jorge Henrique Carneiro
Rodrigo de Almeida
Certamente a comercialização do leite é a principal fonte de renda da maioria das
propriedades leiteiras, sendo de suma importância o volume de leite produzido. Porém, assim
como já ocorreu em vários países de pecuária leiteira mais desenvolvida, há uma crescente
tendência pela remuneração por composição e qualidade do leite, e não apenas por volume
de leite produzido. Mesmo não sendo ainda uma realidade nacional, a remuneração com
bonificações e penalizações pelos teores de sólidos no leite (teores de gordura e proteína) e
sanidade (Contagem Bacteriana Total - CBT e Contagem de Células Somáticas - CCS) já é
uma realidade que ocorre, por exemplo, em vários laticínios e cooperativas progressistas nas
regiões Sul e Sudeste do Brasil. Atualmente, um bom produtor que entrega seu leite ao pool
ABC na região centro-oriental do Estado do Paraná, pode receber mais de R$0,30 de
bonificações por litro de leite sobre o preço base mensal (R$0,88 + R$0,30 = R$1,18/L).
Essas novas formas de remuneração seguem as demandas e exigências de mercado. O que se
tem observado em vários países, inclusive no Brasil, é um maior interesse dos consumidores
em adquirir produtos mais saudáveis. No que diz respeito à cadeia de lácteos, vem ocorrendo
a princípio uma crescente busca por produtos com menores teores de gordura e maiores teores
de proteína. Mesmo isto sendo verdade, é até curioso constatar que o consumidor de lácteos
(no mundo inteiro) é normalmente um pouco incoerente ou até hipócrita ao consumir leite e
derivados, pois se por um lado cada vez mais se consome leite fluido desnatado ou
semidesnatado, nunca se consumiu tanto queijo, sorvetes e outros lácteos com altos teores de
gordura. É por isso que muitos laticínios mantêm as premiações por gordura altas nos
sistemas vigentes de pagamento de leite por qualidade. Reconhecidamente, a gordura é o
sólido do leite que possui maior variabilidade, ou seja, é fortemente impactado pelas práticas
de alimentação e manejo do rebanho. As ocorrências de queda na gordura do leite, muito
comuns em todo o Brasil, particularmente em rebanhos de mediana ou alta produtividade
onde a inclusão de concentrado é mais alta, são tipicamente provocadas por falta de fibra
efetiva na dieta, excesso de amido, ou por ambos os fatores concomitantemente.

137
Tem-se observado uma crescente importância dos teores de proteína do leite, principalmente
em propriedades com sistemas de pagamento do leite por qualidade. Além da variabilidade
mediana, outra dificuldade de incrementar o teor de proteína do leite é a correlação negativa
com os teores de gordura do leite. Em outras palavras, práticas nutricionais e de manejo que
tipicamente aumentam os teores de proteína, diminuem os percentuais de gordura. Entre as
práticas mais relevantes neste incremento de proteína destacaremos o aumento nos teores de
amido da dieta. Outro fator de grande importância no monitoramento é a implantação de
análises rotineiras do leite, tanto amostras individuais de vacas, como amostras de tanque,
para o nitrogênio ureico do leite (NUL), popularmente conhecido como “ureia do leite”. Este
parâmetro nos dá uma ideia do sincronismo do metabolismo proteico e de carboidratos no
rúmen. O ideal são valores intermediários de ureia no leite, entre 10 a 14 mg/dL. Embora
alguns outros fatores ambientais possam afetar este parâmetro, tipicamente quando os valores
de NUL estão abaixo de 10 mg/dL, imediatamente pensamos numa dieta com carência de
proteína bruta (PB). Por outro lado, quando os valores de NUL estão acima de 14 mg/dL,
checamos se os níveis de proteína dietética não estão excessivos e/ou os níveis de amido da
dieta não estão baixos demais. Uma ferramenta adicional que pode auxiliar a identificação
de erros de manejo é a relação (%) gordura/proteína (G/P) do leite. Em toda a lactação,
quando a proporção de vacas com relação G/P invertida for alta (muitas vacas produzindo
menos gordura do que proteína), nos auxilia num provável diagnóstico de subacidose
ruminal. Por outro lado, principalmente nos primeiros 30 dias pós-parto, se a relação G/P for
muito alta (acima de 1,40 na raça Holandesa), isto é indicativo de cetose subclínica ou clínica,
pois neste caso, muito ácidos graxos oriundos da lipomobilização do tecido adiposo da vaca
recém parida são transferidos para o leite.
Genética, componentes e volume de leite
Mudanças na composição do leite através da genética são obtidas a longo prazo,
principalmente pelo longo intervalo de gerações na espécie bovina, se comparadas às
alterações produzidas por práticas de nutrição e de manejo, que geralmente ocorrem de forma
quase imediata. Por outro lado, as mudanças obtidas pela genética são permanentes, ao
contrário das alterações proporcionadas pelas práticas de nutrição, que são transitórias. Entre
os componentes do leite, a gordura apresenta a maior variabilidade, com mediana

138
variabilidade para a proteína e menor para a lactose. Segundo dezenas de publicações
científicas, as produções de gordura e de proteína apresentam medianas herdabilidades
(h2=0,25 a 0,30), enquanto que as porcentagens de gordura e de proteína apresentam altas h2
(0,45 a 0,50). Mas apesar das altas herdabilidades, selecionar para altas porcentagens de
gordura e de proteína pode causar problemas, em função das porcentagens serem
negativamente correlacionadas com o volume de leite.
Composição do leite e diferenças entre raças
A composição normal do leite bovino, usando como exemplo a típica composição do leite
da vaca Holandesa, contém cerca de 12,5% de sólidos (Tabela 1). A composição do leite
considerada normal para vacas leiteiras pode variar em função da raça dos animais. A raça
Holandesa apresenta menores percentuais de sólidos, mas maiores produções totais de
gordura e proteína. As raças Jersey e Pardo-Suíço, apesar dos altos percentuais de gordura e
proteína, apresentam produções totais destes componentes ligeiramente inferiores aos
encontrados na raça Holandesa. Além de características raciais e genéticas, as técnicas de
manejo, condições climáticas e principalmente a nutrição podem alterar a composição do
leite. Por se tratar de um fator econômico oneroso para produção de leite e causar grandes
impactos sob o volume de leite produzido e seus teores proteicos e lipídicos, a nutrição de
bovinos leiteiros é alvo de discussões já a longa data por pesquisadores, técnicos e
produtores.
Tabela 1. Composição normal do leite de vaca
Componentes % Subcomponentes
Água 87,5 Vitaminas hidrossolúveis
Lactose 4,7 Dissacarídeo não encontrado em outros
alimentos
Gordura 3,5 Mais de 400 ácidos graxos e vitaminas
lipossolúveis
Proteína 3,2 77% caseína, 17% proteínas do soro e 6%
NNP
Minerais 0,8 Macro e microminerais, com destaque para
o cálcio
Outros 0,2 Enzimas, hormônios e outros
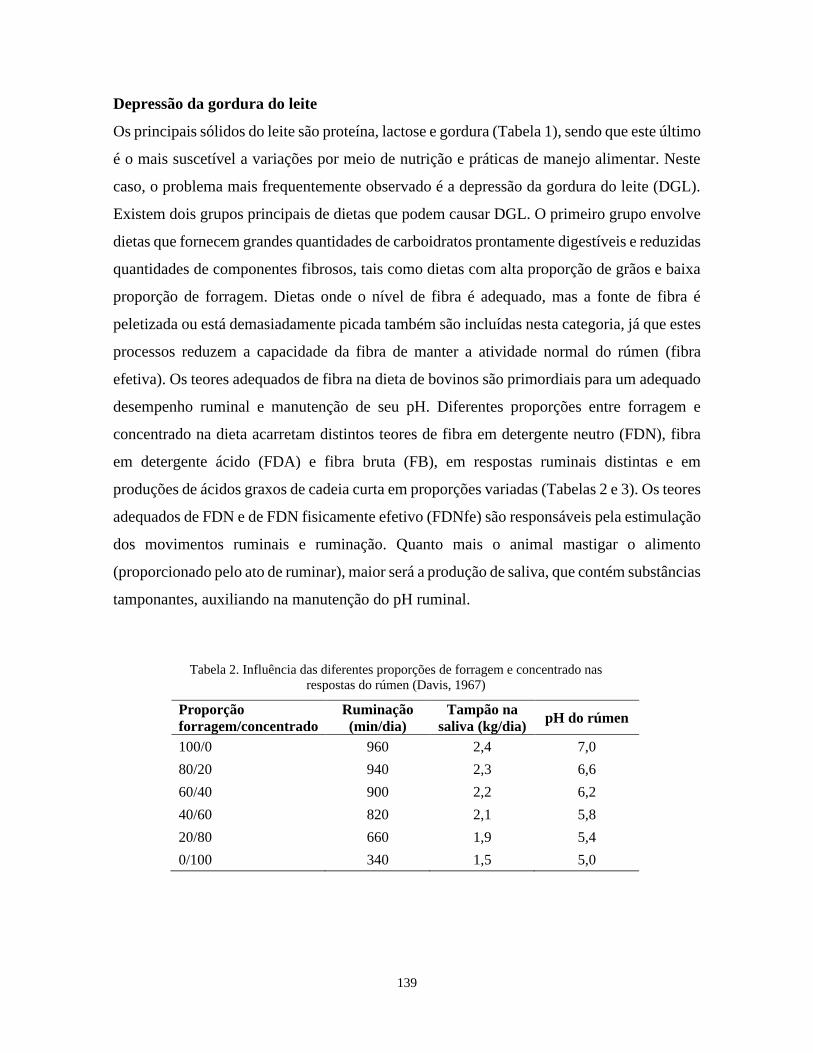
139
Depressão da gordura do leite
Os principais sólidos do leite são proteína, lactose e gordura (Tabela 1), sendo que este último
é o mais suscetível a variações por meio de nutrição e práticas de manejo alimentar. Neste
caso, o problema mais frequentemente observado é a depressão da gordura do leite (DGL).
Existem dois grupos principais de dietas que podem causar DGL. O primeiro grupo envolve
dietas que fornecem grandes quantidades de carboidratos prontamente digestíveis e reduzidas
quantidades de componentes fibrosos, tais como dietas com alta proporção de grãos e baixa
proporção de forragem. Dietas onde o nível de fibra é adequado, mas a fonte de fibra é
peletizada ou está demasiadamente picada também são incluídas nesta categoria, já que estes
processos reduzem a capacidade da fibra de manter a atividade normal do rúmen (fibra
efetiva). Os teores adequados de fibra na dieta de bovinos são primordiais para um adequado
desempenho ruminal e manutenção de seu pH. Diferentes proporções entre forragem e
concentrado na dieta acarretam distintos teores de fibra em detergente neutro (FDN), fibra
em detergente ácido (FDA) e fibra bruta (FB), em respostas ruminais distintas e em
produções de ácidos graxos de cadeia curta em proporções variadas (Tabelas 2 e 3). Os teores
adequados de FDN e de FDN fisicamente efetivo (FDNfe) são responsáveis pela estimulação
dos movimentos ruminais e ruminação. Quanto mais o animal mastigar o alimento
(proporcionado pelo ato de ruminar), maior será a produção de saliva, que contém substâncias
tamponantes, auxiliando na manutenção do pH ruminal.
Tabela 2. Influência das diferentes proporções de forragem e concentrado nas
respostas do rúmen (Davis, 1967)
Proporção
forragem/concentrado
Ruminação
(min/dia)
Tampão na
saliva (kg/dia) pH do rúmen
100/0 960 2,4 7,0
80/20 940 2,3 6,6
60/40 900 2,2 6,2
40/60 820 2,1 5,8
20/80 660 1,9 5,4
0/100 340 1,5 5,0

140
Tabela 3. Influência das diferentes proporções de forragem e concentrado na
produção de ácidos graxos voláteis (Davis, 1967)
Proporção
forragem/concentrado
Acetato
(%)
Propionato
(%)
Relação
acetato/propionato
100/0 70 18 3,9
80/20 67 20 3,4
60/40 64 22 2,9
40/60 58 28 2,1
20/80 48 34 1,4
0/100 36 45 0,8
Por décadas acreditou-se que a queda nos teores de gordura do leite era devida à escassez do
acetato ruminal, principal precursor da síntese de gordura do leite na glândula mamária. De
fato, quando analisamos em valores relativos, observa-se uma redução na quantidade de
acetato. Entretanto, vários trabalhos publicados na década passada, como Bauman & Griinari
(2003), aboliram essa teoria. Os autores avaliaram o fornecimento de dietas contendo baixa
fibra e seus efeitos sobre a produção de leite, gordura e ácidos graxos voláteis comparados a
um grupo controle. Os resultados demonstraram que houve redução na produção (g/dia) e no
teor (%) de gordura no leite, mas que a produção diária de acetato não foi alterada, ao
contrário do propionato que foi produzido em quantidades bem acima (13,3 vs. 31,0
moles/dia) dos níveis encontrados na dieta controle (Tabela 4).
Tabela 4. Ácidos graxos voláteis no rúmen e depressão da gordura do leite em dietas
de baixa fibra
Variáveis Controle Baixa fibra
Produção leite (kg/dia) 19,1 20,9
Produção gordura (g/dia) 683 363*
Gordura (%) 3,6 1,7*
Acetato (%) 67 46*
Propionato (%) 21 46*
Acetato (moles/dia) 29,4 28,1
Propionato (moles/dia) 13,3 31,0*
* Diferenças significativas entre controle e baixa fibra

141
As características físicas dos alimentos volumosos e concentrados, o modo de fornecimento
do alimento, a frequência de alimentação, bem como a competição dentro do lote, afetam
diretamente o consumo e o ambiente ruminal, que por sua vez pode refletir em uma redução
do teor (%) e da secreção (kg/dia) de gordura do leite. A fibra dietética deve conter um
tamanho mínimo para estimular a ruminação e consequentemente a produção de tamponantes
pelo animal. Caso este tamanho mínimo não seja atendido e a efetividade física da fibra esteja
comprometida, podemos ter problemas de DGL. Devido à importância deste assunto,
discutiremos o tema mais adiante separadamente. Características físicas e de processamento
dos alimentos concentrados também são relevantes para a manutenção da gordura do leite.
Processos como moagem fina ou a ensilagem de certos materiais promovem um aumento
expressivo na digestibilidade destes ingredientes, disponibilizando para o ambiente ruminal
carboidratos mais prontamente disponíveis e rapidamente fermentáveis, como por exemplo,
grão de milho moído (fubá) e silagem de grão úmido de milho. Situações em que sementes
de oleaginosas são processadas em demasia ou simplesmente processadas, ocasionam a
liberação do óleo contido na semente (muitas vezes rico em ácidos graxos poli-insaturados,
como o linoleico) dentro do rúmen, sendo um fator predisponente a DGL. Esta situação pode
ser observada ao processar soja integral excessivamente tostada, ou simplesmente pela
quebra do caroço de algodão antes do fornecimento aos animais.
O segundo grupo de dietas que induzem a DGL são suplementos dietéticos contendo óleos
poli-insaturados, tais como óleos de origem vegetal e de origem marinha. Como a
suplementação de óleos de origem marinha é restrita pela proibição imposta pelo Ministério
de Agricultura (MAPA) por conta da encefalite espongiforme bovina (BSE), nossa
preocupação deve ser direcionada aos óleos de origem vegetal e em particular às gorduras
com altas proporções do ácido linolénico (C18:2), tais como o óleo de soja e produtos
derivados. Em ruminantes, os lipídios presentes na dieta são extensivamente alterados pelos
microrganismos do rúmen (Jenkins, 1993). Esta alteração é uma consequência da
biohidrogenação dos ácidos graxos poli-insaturados presentes na dieta, um processo que
tende a tornar a gordura do leite mais saturada, mas que também resulta na formação e
secreção de inúmeros ácidos graxos do tipo trans no leite, com destaque aos isômeros de
CLA (ácido linoleico conjugado) cis-9 trans-11 e trans-10 cis-12. Devido a estas e outras

142
transformações que ocorrem no rúmen, mais de 400 tipos de ácidos graxos já foram
identificados na gordura do leite de bovinos. Os ácidos graxos presentes em maior
concentração são o ácido palmítico (C16:0) e o ácido oleico (C18:1 cis-9), sendo que a soma
dos dois representa cerca de 50% do total. Os ácidos graxos secretados no leite podem ter
duas origens: síntese de novo nas células epiteliais mamárias ou circulação sanguínea. Ácidos
graxos de cadeia curta (4-8 carbonos) e média (10-16 carbonos) são sintetizados quase que
exclusivamente pela síntese de novo na glândula mamária, predominantemente a partir do
acetato (produto da degradação de carboidratos estruturais no rúmen). Em contrapartida, os
ácidos graxos de cadeia longa, com 18 ou mais carbonos, são oriundos exclusivamente da
circulação (dieta ou lipomobilização). Várias teorias têm sido propostas para explicar a DGL
induzida pela dieta, e alterações nos processos microbianos do rúmen são a base de todas
estas teorias. A concentração molar de acetato diminui, a de propionato aumenta e, por
consequência, a relação acetato:propionato cai drasticamente (Tabela 4). Entretanto, embora
a proporção molar de acetato seja reduzida, a produção de acetato (moles/dia) não é afetada.
Portanto, a redução na relação acetato:propionato é uma consequência da maior produção de
propionato. Uma vez que a produção de acetato não é reduzida em dietas com baixa fibra,
esta teoria foi abolida, pois ela não poderia explicar a DGL.
A teoria mais aceita atualmente para explicar a DGL é conhecida como teoria da
biohidrogenação ou teoria dos ácidos graxos trans. Esta teoria postula que a DGL não é
causada pela escassez de precursores lipogênicos para a glândula mamária. Ela afirma que a
síntese mamária da gordura do leite é inibida diretamente por tipos específicos de ácidos
graxos, produzidos a partir da biohidrogenação parcial dos lipídios da dieta sob certas
condições ruminais. A queda no pH ruminal (proporcionada pelo fornecimento de dietas de
baixa fibra fisicamente efetiva ou excesso de carboidratos rapidamente fermentáveis) altera
as rotas de biohidrogenação ruminal, produzindo um ácido graxo específico, chamado de
CLA trans-10 cis-12, cuja concentração aumenta significativamente no leite de vacas com
DGL. Estudos posteriores, onde este isômero de CLA foi sintetizado industrialmente e
suplementado a vacas em lactação, comprovaram sua grande capacidade de reduzir a
secreção de gordura do leite, mas também de potencialmente incrementar o desempenho
reprodutivo (Bernal-Santos et al., 2003, Castañeda-Gutierrez et al., 2007). De grande
relevância, estudos recentes demonstraram que a formação do CLA trans-10 cis-12 no rúmen

143
só ocorre quando duas condições estão presentes: baixo pH ruminal (ex.: dietas de baixa
fibra) e presença de lipídios poli-insaturados na dieta (ex.: dietas suplementadas com grãos
de oleaginosas, óleo de milho, sais cálcicos de ácidos graxos a partir do óleo de soja, entre
outros). A ausência de qualquer uma dessas condições não resultará em DGL. Muitas
pesquisas têm enfocado no aumento da quantidade de ácido linoleico conjugado (CLA) e de
ácido transvaccênico (trans-11 C18:1) na gordura do leite pelos benefícios relatados na saúde
humana. Segundo Whitlock et al. (2002) certos isômeros de CLA, especialmente o cis-9,
trans-11, têm sido relacionados a propriedades anti-carcinogênicas, além de
antiaterosclerose, antitrombótico, hipocolesterolêmico, prevenir a diabetes mellitus e
apresentar efeito imunoestimulatório. Outros isômeros de CLA, incluindo o trans-10, cis-12,
parecem ainda possuir propriedades antiobesidade.
Monitoramento do FDN e do FDNfe da dieta
Respeitar as recomendações e exigências de FDN e FDNfe de vacas leiteiras em suas diversas
fases produtivas, são essenciais para a manutenção do pH ruminal, favorecendo assim a
fermentação microbiana e a biohidrogenação adequada de ácidos graxos poli-insaturados, ou
seja, sem a formação de CLA trans-10, cis-12, evitando a DGL. As recomendações do
penúltimo NRC para bovinos leiteiros (1989) sugeriam que a dieta de bovinos leiteiros
deveria conter no mínimo 28% de FDN e 21% de FDA. Para vacas de alta produção a
recomendação era de no mínimo 25% de FDN e 19% de FDA, e que 75% da FDN total da
dieta deveria ser proveniente de forragem. Por conta do uso crescente de subprodutos fibrosos
nas dietas de vacas leiteiras, Mertens (1997) e o NRC (2001) estabeleceram que o valor
efetivo médio do FDN não forragem é de 50% do FDN forragem. Desta forma, as
recomendações seguem a seguinte regra: para cada redução de 1% no FDN forragem abaixo
de 19%, a concentração do FDN total da dieta deve aumentar em 2%, enquanto que a %CNF
(carboidratos não fibrosos) deve diminuir em 2% (Tabela 5). Segundo as recomendações do
NRC (2001), os valores para FDN, FDA e CNF sugeridos são válidos quando três condições
específicas são observadas: uso de dieta total misturada (TMR), tamanho de partícula
adequado e o milho moído como a fonte predominante de amido utilizado na dieta. Em outras
palavras, o NRC (2001) sugere que a concentração de FDN na dieta deve ser
conservadoramente mais alto quando se usa forragem excessivamente picada ou processada

144
e em dietas que contenham altas proporções de subprodutos ricos em FDN não forragem, tais
como casquinha de soja, refinazil ou polpa cítrica. As recomendações de FDN podem sofrer
ajustes em função de aspectos qualitativos da dieta, tais como fontes de amido disponíveis,
tamanho de partícula da forragem, fibra efetiva da dieta, suplementação de tampões e do
manejo alimentar adotado.
Tabela 5. Exigências de carboidratos estruturais segundo o NRC (2001)
Mínimo
FDN
Forragem
Mínimo
FDN Dieta
Máximo
CNF Dieta
Mínimo
FDA Dieta
19 25 44 17
18 27 42 18
17 29 40 19
16 31 38 20
15 33 36 21
Tratando-se de diversas fontes de amido, as exigências de FDN aumentam quando a
disponibilidade do amido no rúmen aumenta. Essa maior ou menor disponibilidade de amido
no rúmen é relevante quando se compara fontes diferentes desse nutriente, como por
exemplo: grãos moídos vs. grãos quebrados, grãos floculados vs. grãos secos, grãos de alta
umidade vs. grãos secos, grãos de cereais de inverno vs. milho, e assim por diante. O conceito
de FDNfe surgiu para atender a exigência mínima de fibra que mantém a porcentagem de
gordura no leite em níveis adequados. No ambiente ruminal sabe-se que nem todo FDN
presente na dieta tem atividade efetiva no rúmen e no metabolismo animal, ou seja, podem
não apresentar características físicas desejadas (tamanho e densidade de partículas), estímulo
a atividade de mastigação e produção de saliva, manutenção de pH e neutralização de ácidos
produzidos no rúmen. Mertens (1997) propôs um método para calcular o FDNfe, o qual
considera a %FDN na dieta e a quantidade de partículas retidas em uma peneira de 1,18 mm
(FDNfe = %FDN x fef). O fator de efetividade física varia de 0 a 1. Não é incomum a
existência de alimentos com quantidades de FDN muito semelhantes, mas que apresentem
valores completamente diferentes de efetividade física. As exigências de FDNfe para bovinos

145
leiteiros não foram definidas pelo NRC (2001). Mertens (1997) estimou que a exigência de
FDNfe para vacas leiteiras é de 22%MS para manter um pH ruminal de 6,0 e de 20%MS para
manter o teor de gordura no leite em 3,4%. Na nossa experiência, ao formular dietas em
rebanhos leiteiros paranaenses, tentamos respeitar o limite de 21% de FDNfe para dietas de
vacas leiteiras de alta produção, desde que os níveis de amido não sejam excessivos (até
27%MS). Em uma Invited Review muito citada e até premiada de Zebeli et al. (2012),
publicada no Journal of Dairy Science, estes autores questionaram o uso da peneira de 1,18
mm na estimativa do FDN fisicamente efetivo, e sugeriram a peneira anterior de 8 mm.
Perante esta nova recomendação, Zebeli et al. (2012) recomendaram FDNfe > 8mm entre
18,5 (para não reduzir o pH ruminal aquém de 6,2) e 14,9% (para não limitar a ingestão de
MS). Avaliar o tamanho de partícula na dieta é fundamental para garantir a efetividade da
fibra e evitar que os animais selecionem alimentos mais concentrados. Muitas vezes a
preocupação por parte do nutricionista e do produtor, ou ambos, é tão grande para atingir
níveis mínimos de partículas fisicamente efetivas que acabam pecando pelo excesso.
Diversos trabalhos demonstram que vacas têm capacidade de selecionar partículas de
forragem maiores que 5 cm de comprimento. Quando a dieta total está muito “grosseira”, ou
seja, com fibras excessivamente longas, damos a possibilidade de os animais selecionarem o
alimento. O mais comumente observado na literatura e no campo é o comportamento de
seleção contra a fibra e a favor do concentrado. Quando este fato ocorre, o fluxo de nutrientes
para o rúmen passa a ser distinto ao longo do dia, sendo que no momento logo após o
fornecimento da dieta os animais ingerem grandes quantidades de concentrado, e a fibra
passa a ser ingerida apenas em períodos mais distantes. Quanto mais acentuada for esta
seleção, mais intensa será a queda do pH ruminal e consequentemente maior o risco de DGL.
Para reduzir este processo seletivo, algumas medidas devem ser tomadas, como por exemplo,
a picagem da forragem entre 2,5 e 5 cm. Dietas muito secas, acima de 50-55% MS, também
facilitam a seleção, já que as partículas estão mais soltas na mistura. A adição de água na
mistura pode ser um recurso para reduzir este comportamento, incluindo a água como
ingrediente na formulação de modo que as dietas tenham entre 40-45% MS. Esta prática
agrega mais partículas finas oriundas de ingredientes concentrados às forragens, dificultando
a triagem. O fornecimento da dieta na forma de TMR (ração total misturada) é uma das únicas
práticas nutricionais que permite o aumento concomitante da produção de leite, da

146
porcentagem de gordura e da porcentagem de proteína. Porém a homogeneidade da TMR
também é determinante para reduzir a seleção. Claramente, dietas pouco homogêneas são
mais facilmente processadas pelos animais.
Diversos pontos de manejo durante o preparo da dieta devem ser monitorados para um bom
resultado:
Tempo de mistura: tempos de mistura entre 3 e 8 minutos são suficientes para a promoção de
uma boa homogeneidade sem prejudicar em demasia o tamanho da fibra;
Ordem de carregamento dos ingredientes no misturador: como recomendação geral, fenos e
silagens pré-secadas devem ser incluídas primeiramente no vagão misturador, seguido de
concentrados e minerais, posteriormente a silagem de milho e, se houver, a adição de
líquidos;
Sobrecarregamento do equipamento: os misturadores são projetados para trabalhar com certa
carga, ou seja, carga acima ou abaixo do recomendado pode comprometer a eficiência do
vagão. Entre 60 e 80% da carga máxima permitida pelo fabricante para aquele modelo
específico deve ser preenchida para a obtenção de uma boa mistura.
Para determinar se o rebanho contém problemas de seleção, a utilização do conjunto de
peneiras Penn State pode ser utilizada como ferramenta. Basicamente, a distribuição das
partículas no conjunto deve ser obtida logo após a distribuição do alimento no cocho, sem
que os animais tenham tido acesso, e minutos antes do próximo trato ou da retirada da sobra
diária. Como indicativo de ausência de seleção excessiva, a diferença entre as proporções
retidas na peneira de 19 mm da dieta oferecida e das sobras não deve ser superior a 10%.
Fatores relacionados ao manejo alimentar como espaçamento de cocho, número de tratos
diários e aproximação da dieta têm sido relatados na literatura como possíveis impactantes
no teor de gordura do leite. Diversos estudos têm demonstrado que a redução do espaço
individual no cocho modifica completamente o comportamento alimentar dos animais,
aumentando a seleção do alimento, reduzindo o tempo de alimentação diária, o número de
visitas no cocho e principalmente o consumo de matéria seca. Em vacas de alta produção
onde o consumo de MS é determinante para a produção de leite, a redução do consumo e,
consequentemente, do aporte de nutrientes é completamente indesejável. Sova et al. (2013)
observaram em seu estudo com fazendas comerciais que a cada 10 cm a mais de espaço linear
de cocho houve um incremento de 0,06 pontos na porcentagem de gordura do leite. Fornecer

147
um espaçamento na linha de cocho de mínimo de 70 cm por animal, utilizar canzis ou algum
outro tipo de contenção podem ser boas práticas para reduzir a competição. É comum
observar em rebanhos que existe superlotação e consequentemente competição entre os
animais, que os teores individuais de gordura do leite dentro do mesmo lote são
extremamente variáveis, com vacas de percentual de gordura muito baixos (animais
dominantes que chegam primeiramente no cocho e se alimentam de uma dieta pouco
selecionada, ainda rica em concentrados), e outros com valores muito elevados (animais
inferiores na hierarquia do lote e que se alimentam por último, quando a quantidade de fibra
da dieta já é mais alto devido a seleção prévia dos outros animais). O aumento na frequência
de fornecimento da dieta aos animais tende a incrementar os teores de gordura do leite,
devido a maior constância de alimentação ao longo do dia, reduzindo momentos de queda de
pH ruminal muito acentuados. Ganhos significativos em produção de leite não têm sido
relatados na literatura ao aumentar o número de tratos além de duas vezes ao dia, obviamente
em situações que não haja limitação do consumo de matéria seca. Economicamente,
aproximar a dieta mais frequentemente pode ser mais interessante que realizar diversos tratos,
uma vez que o custo operacional é muito menor e os ganhos em produção são pouco
expressivos.
O quadro de subacidose é caracterizado por pH ruminal abaixo de 5,8 e uma das prováveis
causas é a ingestão excessiva de carboidratos de rápida fermentação, decorrente de
desbalanceamento da dieta ou seleção do alimento por parte do animal. Alguns sinais podem
facilitar a identificação de acidose ruminal: consumos de MS e produções de leite variando
muito de um dia para outro, esterco com aspecto inconsistente, falta de ruminação, consumo
voluntário de bicarbonato, acidose sistêmica provocando laminite e depressão de modo geral
das vacas, além de obviamente queda nos teores de gordura do leite.
Outra forma de diagnosticar a ocorrência da DGL em um rebanho é monitorar a relação
gordura:proteína do leite. Os teores de gordura no leite apresentam maiores variações que os
teores de proteína, além de ser um componente que permite maior manipulação,
principalmente através da dieta. Portanto quando se diz que houve uma inversão entre a
relação G/P, o que provavelmente ocorreu foi uma redução nos teores de gordura no leite e
não um aumento nos teores de proteína. A relação G/P no leite considerada ideal para a raça
Holandesa é de 1,1, mas valores entre 1,0 e 1,2 são aceitáveis. Se em um rebanho mais de

148
10% das vacas em lactação apresentarem 0,2% de gordura abaixo do teor de proteína
(exemplo 2,8%G e 3,0%P) isso pode ser um indício de acidose no rebanho e/ou
desbalanceamento da dieta.
Impacto do aumento dos níveis de PB da dieta e nitrogênio ureico no leite
A proteína é um nutriente essencial para a produção de leite e influencia a ingestão de
alimentos pelo animal. No leite de vacas, a proteína verdadeira constitui cerca de 94% do
nitrogênio total e o restante é representado pelo nitrogênio não proteico (NNP). O
monitoramento dos teores de nitrogênio ureico no leite (NUL) é uma ferramenta que tem se
tornado cada vez mais utilizada, por estar diretamente relacionada ao teor proteico e
energético da dieta e a excreção de N pelo animal. Tem-se buscado valores de NUL que
representem um ponto de equilíbrio entre ingestão de proteína e produção de leite sem
grandes perdas de N, ou seja, maior eficiência na utilização de N (EUN). Na última década,
a maioria das publicações norte-americanas passou a sugerir metas para NUL valores entre
8 e 12 mg/dL (Kohn et al., 2002; Rajala-Schultz & Saville, 2003), com algumas publicações
mais recentes sugerindo valores ainda menores, entre 7 a 10 mg/dL. No Brasil, nossa sugestão
é mais conservadora: valores entre 10 e 14 mg/dL parecem ser ainda os mais indicados
(Almeida, 2012; Doska et al., 2012). Embora vários rebanhos norte-americanos estejam
tendo sucesso em conciliar baixos níveis dietéticos de PB (inferiores a 16,0%PB) e baixos
valores de NUL (inferiores a 10 mg/dL) com altas produções de leite, acreditamos que no
Brasil a redução muito exagerada dos teores de PB das dietas poderão comprometer a
produção de leite. Esta distinção ocorre pela impossibilidade de uso de suplementos proteicos
de origem animal (naturalmente ricos em PNDR), pela não disponibilidade de suplementos
proteicos de origem vegetal com baixa degradabilidade ruminal, bem como pela não
popularidade na suplementação de aminoácidos sintéticos (metionina e lisina) em dietas
brasileiras.
O monitoramento mensal do NUL pode ser uma importante ferramenta no manejo de
rebanhos leiteiros, porque (1) o excesso no consumo de proteína (N) pode comprometer a
eficiência reprodutiva; (2) suplementos proteicos são ingredientes caros; e (3) excessos na
excreção de N têm um impacto ambiental negativo (Jonker et al., 1998). Entre as práticas
nutricionais que podem favorecer o aumento do NUL, podemos destacar: redução exagerada

149
dos níveis dietéticos de amido pela inclusão de fontes de carboidratos não amiláceos, como
casca de soja e polpa cítrica; o uso de silagem de milho “nova” (menos de três meses de
ensilagem); pastagens novas e muito adubadas, principalmente de inverno; pré-secado úmido
e com alto %PB; e fornecimento do grão de milho mais grosseiro (quebrado ao invés de
moído).
Impacto do estresse calórico na composição do leite
Animais da raça Holandesa são menos tolerantes a temperaturas mais elevadas que outras
raças leiteiras europeias, como Jersey e Pardo-Suíço, e muito menos tolerantes que
grupamentos raciais com sangue zebuíno, como Girolando e Gir Leiteiro. Vacas Holandesas
submetidas a estresse calórico reduzem sua produtividade de forma expressiva. Num
levantamento preliminar conduzido pelo nosso grupo de pesquisa (dados não publicados),
realizado em 26 rebanhos paranaenses em controle leiteiro oficial na região de Arapoti,
Paraná, a diferença entre o mês de maior produtividade média (setembro) e o mês de menor
produtividade média (fevereiro) alcançou 8 litros diários. O estresse calórico impacta
negativamente vários parâmetros na atividade leiteira, tais como produção de leite, qualidade
e composição do leite, saúde ruminal, crescimento e reprodução. St. Pierre et al. (2003)
estimaram as perdas da indústria leiteira norte-americana devido ao estresse calórico em 900
milhões de dólares anuais. Avanços no manejo com a adoção de práticas de resfriamento e
estratégias nutricionais podem amenizar alguns dos impactos negativos do estresse calórico
em bovinos leiteiros, mas as produções continuam a declinar nos meses mais quentes.
O mecanismo biológico pelo qual o estresse calórico impacta a produção e a reprodução é
parcialmente explicado pela depressão no consumo alimentar, mas também inclui outros
fatores tais como mudança no status endócrino, reduções na ruminação e na absorção de
nutrientes e aumento nas exigências de mantença. As reduções tanto nos teores de gordura
como nos de proteína verdadeira relatadas nas épocas mais quentes do ano pela indústria
leiteira norte-americana se situa entre 0,2 e 0,3%. Infelizmente desconhecemos a existência
de levantamentos similares pela indústria leiteira nacional. De acordo com Nayeri et al.
(2011) existem algumas estratégias que podem ser adotadas para minimizar o impacto do
estresse calórico na produção e composição do leite. Uma das mais tradicionais é aumentar
a densidade energética da dieta, seja pela redução de fibra, pelo aumento do concentrado ou

150
ainda pela suplementação de gorduras. Estas práticas devem ser adotadas com cuidado, pois
se aumenta a predisposição das vacas em estresse calórico à acidose ruminal. A manipulação
da Diferença Catiônica-Aniônica da Dieta (DCAD), buscando valores francamente positivos
durante a lactação (+20 a +30 meq/100 g MS), bem como o aumento nos níveis dietéticos do
mineral potássio no verão (1,4 a 1,6%MS) também têm sido sugeridos. Outras práticas
nutricionais e de manejo comumente recomendadas são o fornecimento preferencial de
alimento (60-70% da dieta) nas horas mais frescas do dia, a limpeza mais frequente dos
cochos com maior número de tratos diários, o fornecimento de concentrado mais parcelado
ao longo do dia e a inclusão de 2 a 2,5 kg de caroço de algodão.
Impacto de alguns aditivos nutricionais na composição do leite
Monensina
A monensina, assim como outros ionóforos, é um antibiótico produzido por fungos do gênero
Streptomyces cinnamonensis, com reconhecida eficácia nas explorações pecuárias. Os dados
de literatura são abundantes e extremamente consistentes para o efeito dos ionóforos, tanto
em bovinos de corte como em bovinos leiteiros. A maior parte dos experimentos foi
conduzida com monensina sódica. O efeito mais notório dos ionóforos é o aumento da
retenção de energia fermentada no rúmen devido a uma alteração no padrão de fermentação,
com maior produção de propionato (C3) em relação a acetato (C2) e decorrente diminuição
das perdas através do metano. Os ionóforos também parecem diminuir a degradação da
proteína ruminal e, portanto, reduzindo a degradação de peptídeos e aminoácidos e resultando
em menor produção de amônia. Por último, os ionóforos (particularmente a monensina)
diminuem a ocorrência de distúrbios metabólicos, como acidose e timpanismo, pela menor
concentração de ácido láctico e menor produção de mucopolissacarídeos que dão estabilidade
à espuma. Os efeitos nas produções de gordura e de proteína são heterogêneos e dependem
de fatores dietéticos. Em uma meta-análise que reuniu 36 artigos ou 77 experimentos
envolvendo monensina e dados de produção (Duffield et al., 2008), a suplementação com
monensina aumentou a produção de proteína e não apresentou efeito na produção de gordura.
Durante muito tempo atribuiu-se à menor produção de acetato no rúmen o efeito de redução
na gordura no leite, já que de fato o acetato é o principal precursor da gordura do leite. Mas

151
hoje se sabe que na realidade o efeito depressor da monensina na gordura do leite é explicado
pela diminuição das taxas de biohidrogenação dos ácidos graxos insaturados no rúmen, o que
contribui no aumento das concentrações do ácido linoleico conjugado trans-10 cis-12,
potente inibidor da síntese de gordura do leite na glândula mamária (Bauman & Griinari,
2003). Duffield et al. (2008) confirmaram que a suplementação com monensina aumenta a
proporção de ácido linoleico conjugado (CLA) no leite. Maiores consumos de fontes lipídicas
insaturadas na dieta e altas inclusões de monensina (ao redor de 400 ppm) exacerbam a
diminuição da gordura do leite com o tratamento com monensina. Dosagens recomendadas
de monensina para vacas em lactação estão entre 11-22 mg/kg de MS consumida, ou 250-
400 mg/vaca/dia (Hutjens, 2010). No Brasil, devido ao impacto negativo da monensina sobre
a gordura do leite e ao potencial decréscimo no consumo de ração, dosagens mais próximas
do limite inferior são sugeridas (11-15 mg/kg).
Leveduras
Leveduras vêm sendo fornecidas aos animais há mais de cem anos. Embora existam
aproximadamente 500 diferentes espécies de leveduras, a mais comum na suplementação dos
bovinos é a Saccharomyces cerevisiae. Segundo Santos & Greco (2012), de modo geral, o
efeito das leveduras vivas e cultura de leveduras se dá por alterações no ambiente ruminal,
com mudanças na população microbiana que favorecem o crescimento de microrganismos
celulolíticos e daqueles utilizadores do ácido láctico. De maneira geral, a adição de leveduras
a dieta de bovinos leiteiros resulta em aumento na ingestão de MS, estabilidade do pH
ruminal (utilização do ácido láctico), melhora a digestão de fibra no rúmen, aumenta a
produção de leite (entre 1 a 1,2 kg/dia), aumenta a concentração de gordura no leite e a
produção de componentes do leite (Santos & Greco, 2012). De acordo com os autores citados
anteriormente, a dose típica recomendada para bovinos leiteiros é de 1 a 2x1010 unidades
formadoras de colônias por dia para bovinos adultos. A conversão desses valores em
g/dia/animal depende da concentração de leveduras presentes no produto comercial, mas os
valores podem variar de 5 a 120 g/dia.

152
Aminoácidos metionina e lisina
A alimentação de vacas leiteiras especializadas baseia-se na oferta dos nutrientes exigidos
pelo animal de forma a atender suas necessidades adequadamente, sem que ocorra a falta ou
o excesso de algum nutriente. A proteína, quando em excesso na dieta, além de promover a
maior produção de fezes, também é responsável pela ineficiência de utilização do nitrogênio
para a produção de leite (Jonker et al., 2002). Com isso, torna-se imprescindível a utilização
de dietas adequadamente formuladas, principalmente quanto aos teores de proteína e seus
aminoácidos limitantes (metionina e lisina), que atendam às necessidades da vaca leiteira
especializada, resultando potencialmente em maior quantidade e qualidade de leite, redução
dos impactos negativos ao ambiente e diminuição dos custos com a alimentação (NRC,
2001). Robinson (2010) numa revisão de literatura que reuniu 54 experimentos comparando
a suplementação de lisina e/ou metionina com dietas controle, concluiu que a manipulação
das proporções de lisina e metionina da proteína duodenal pela suplementação de metionina
protegida, ou lisina combinada a metionina, apresentou efeitos positivos, mas modestos, na
melhoria do desempenho produtivo, bem como na redução do impacto ambiental de vacas
leiteiras. Robinson (2010) justificou os resultados argumentando que a contribuição da
proteína microbiana na proteína metabolizável é tão grande, e que o perfil de aminoácidos
desta proteína é tão similar com o perfil de aminoácidos das proteínas do leite, que mesmo
suplementando um aminoácido específico, o benefício gerado em resposta animal é
geralmente pequeno. Ainda assim, na meta-análise conduzida, o autor concluiu que a
suplementação com metionina aumentou em 1,3% a secreção de energia no leite, bem como
os teores de gordura (+1,1%) e de proteína (+2,3%) no leite, além de aumentar em 2,1% a
proporção de N dietético capturado como N no leite (eficiência de utilização do nitrogênio).
Em nossa opinião, há pelo menos três razões que limitam a inclusão de metionina análoga
em dietas de rebanhos leiteiros brasileiros: (1) a ainda restrita adoção de sistemas de
pagamento de leite por qualidade, (2) a falta de dados nacionais mostrando os benefícios da
inclusão dietética de metionina e (3) o pouco número de rebanhos de boa produtividade que
justificam a suplementação. Por outro lado, há no mínimo três razões que justificam a
inclusão da metionina análoga em rebanhos nacionais: (1) o fato de o farelo de soja (pobre
em metionina) ser o principal suplemento proteico em dietas de vacas leiteiras, (2) a proibição
(até exagerada) de suplementos proteicos de origem animal em dietas de ruminantes no Brasil

153
e (3) a limitada disponibilidade no mercado brasileiro de alimentos proteicos de origem
vegetal com baixa degradabilidade ruminal. Por conta da pequena disponibilidade de
suplementos proteicos com maiores valores de proteína não degradável no rúmen e da não
popularidade da suplementação de aminoácidos protegidos (metionina em particular), há no
Brasil uma cultura entre os nutricionistas de bovinos leiteiros que rebanhos ou lotes de vacas
de alta produção devem ser suplementados com altos teores de proteína bruta (ao redor de 17
a 18%PB). Não concordamos com esta excessiva margem de segurança, e acreditamos que
é possível trabalhar com valores menores (16 a 16,5%PB).
Bicarbonato de sódio e óxido de magnésio
Segundo o NRC (2001) compostos tais como bicarbonato de sódio e óxido de magnésio são
incorporados às dietas a fim de reduzir transtornos digestivos e manter o teor de gordura no
leite, principalmente quando as dietas são ricas em grãos (carboidratos de alta
fermentabilidade) e/ou pobres em ingredientes volumosos, fontes de fibra efetiva. Quando
corretamente suplementados, estes tampões estabilizam o pH ruminal em torno de 6,2
permitindo um máximo crescimento das bactérias celulolíticas. O óxido de magnésio, além
de ser fonte do macromineral magnésio, é um alcalinizante e parece atuar também na captura
de metabólitos sanguíneos pela glândula mamária, aumentando o teor de gordura no leite
produzido. A recomendação de inclusão do óxido de magnésio é de 1 parte deste alcalinizante
para cada 2-3 partes de bicarbonato de sódio. Assim, quando há suplementação de óxido de
magnésio, não se recomenda a exclusão do bicarbonato de sódio, mas sim a adição de ambos.
As recomendações de bicarbonato de sódio então entre 0,75-1,0% MS/dia, e de óxido de
magnésio em 0,25% MS/dia. Segundo Almeida & Ostrensky (2011), em rebanhos confinados
de mediana e alta produtividade, outra forma de suplementar bicarbonato de sódio às vacas
é fornecê-lo ad libitum, em cochos exclusivos, além do bicarbonato já fornecido às dietas.
Neste caso a função do bicarbonato ofertado nos cochos é de monitorar possíveis ocorrências
de acidose ruminal subaguda. Como animais sem acidose não consomem voluntariamente o
bicarbonato, quando isto acontece de maneira ávida por muitos animais, pode chamar a
atenção do produtor que a dieta está desequilibrada e que o nutricionista do rebanho precisa
reformular as dietas.

154
Gordura protegida
Gordura protegida ou gordura inerte ruminal é um sal cálcico de um ácido graxo (AG), ou
seja, um produto onde se combinam moléculas de AG com cálcio, para que o produto passe
pelo rúmen sem sofrer dano extensivo pela ação da flora ruminal e sem interferir com esta.
A ligação entre o cálcio e o AG não é quebrada por ação de enzimas microbianas e sim pelo
pH do meio. Segundo Almeida & Ostrensky (2011) para a produção deste sal cálcico de AG
podem ser utilizadas fontes de lipídios predominantemente saturados (como a gordura de
palma) ou insaturados (como os demais de origem vegetal, mas principalmente o de soja). O
efeito da suplementação de gordura protegida (na verdade, sais cálcicos de ácidos graxos)
para bovinos leiteiros demonstra resultados variáveis sobre a composição do leite,
dependendo da composição e da quantidade de gordura oferecida. A suplementação de
gorduras a base de ácidos graxos (AG) saturados é mais segura, pois estes são mais estáveis
no rúmen, portanto contribuem para a manutenção do teor de gordura no leite. As fontes ricas
em AG poli-insaturados são mais dependentes do pH ruminal para sua não dissociação e
consequente manutenção dos teores de gordura no leite. A explicação pela moderada
depressão da gordura do leite nos animais suplementados com sais cálcicos de AG de óleo
de soja é que os AG insaturados encontrados nesta fonte de gordura supostamente inerte não
são tão eficientemente protegidos contra a biohidrogenação parcial no rúmen, fato já
demonstrado por Chouinard et al. (1998). Mudanças na biohidrogenação destes AG
insaturados devem ter contribuído para a síntese do ácido linoleico conjugado trans-10 cis-
12, potente inibidor da síntese de gordura do leite na glândula mamária (Bauman & Griinari,
2003). Praticamente a totalidade dos experimentos consultados com suplementação de
gorduras resultou em redução na porcentagem de proteína (Rabiee et al., 2012),
principalmente da caseína. Esta redução pode ter impacto negativo nos atuais sistemas de
pagamento do leite por qualidade. Práticas para amenizar esta redução nos teores proteicos
do leite seria a suplementação com niacina, formulação de dietas com maiores níveis de
PNDR (proteína não degradável no rúmen) e suplementação de aminoácidos protegidos ou
análogos.

155
Somatotropina bovina
A condição nutricional das vacas tanto antes quanto durante a suplementação de
somatotropina bovina (bST) determina o efeito deste hormônio sobre as concentrações de
gordura e proteína no leite (NRC, 2001). Quando se faz uso de somatotropina bovina
associada a uma alimentação e manejo adequados e em balanço energético positivo, as
porcentagens de gordura, proteína, lactose, vitaminas e minerais não variam em animais
suplementados (St-Pierre et al., 2014). Porém, quando tais requerimentos não são atendidos
e o balanço energético negativo é prolongado, a porcentagem de gordura aumenta, a
porcentagem de proteína diminui e a composição das gorduras do leite é alterada, com maior
proporção de ácidos graxos de cadeia longa, oriundos da mobilização mais intensa das
reservas corporais (Bauman et al., 1989).
Relação gordura:proteína no leite e indicadores de cetose subclínica
Para Heuer et al. (1999) a relação gordura:proteína deve ser analisada a nível de rebanho; se
mais de 40% dos animais apresentarem valores abaixo de 1,0 ou acima de 1,4 o rebanho tem
grandes riscos de apresentar casos de acidose ou cetose, respectivamente. Nosso grupo de
pesquisa conduziu um estudo realizado em rebanhos paranaenses entre os anos de 2000 e
2013 (Poncheki, 2015). Foram avaliados 257.847 primeiros controles leiteiros mensais de
vacas que estavam em controle oficial. Este trabalho indicou que 63,5% das vacas no
primeiro controle leiteiro após o parto estavam com a relação gordura:proteína dentro dos
valores ideais. Valores abaixo de 1,0 sugerem uma depressão da gordura do leite, sendo que
neste banco de dados representaram 20,6% das avaliações. Quando nos deparamos com
valores acima de 1,4 refere-se a um aumento na proporção de gordura do leite, que pode ter
relação com quadros de cetose (clínica ou subclínica) e neste estudo representaram 15,9%
dos primeiros controles leiteiros após o parto avaliados. Estes quadros de relação
gordura:proteína acima de 1,4 podem ser mais evidenciados em situação de balanço
energético negativo (BEN), onde animais com BEN muito acentuado ou muito prolongado
mobilizam mais reservas corporais para manter a produção de leite. A gordura que é
mobilizada se direciona para a glândula mamária, onde irá compor a gordura do leite,
particularmente ácidos graxos de cadeia longa. Van Haelst et al. (2008) avaliaram se alguns
ácidos graxos específicos da gordura do leite podem ser usados na detecção precoce de cetose

156
subclínica. Vacas leiteiras recém paridas, diagnosticadas com cetose subclínica (BHB > 1,2
mmol/L), apresentaram elevadas proporções de C18:1 cis-9 na gordura do leite, conhecido
como ácido oleico. Ainda mais recentemente alguns serviços de controle leiteiro ao redor do
mundo, estão mensurando as concentrações de beta-hidroxibutirato (BHB) diretamente no
leite, em amostras de leite de vacas recém paridas (Van der Drift et al., 2012).
Resumo das práticas nutricionais e de manejo que afetam a composição do leite
Na Tabela 6 estão resumidas importantes práticas de nutrição e de manejo que podem afetar
tanto a produção de leite, como os teores de gordura e proteína.
Tabela 6. Práticas de nutrição e de manejo que podem afetar a produção de leite e seus principais
componentes (adaptado de Hutjens & Shanks, 1993)
Prática adotada kg leite % proteína % gordura
Maior consumo de MS ++ 0? 0?
Forragem, grãos - - +
Forragem, grãos + + -
Silagem de milho, pré-
secado + + -
Carboidratos não estruturais + + -
FDN/FDA - - +
Suplementos de gordura + - 0
Grãos mais frequentemente + + +
Proteína bruta mais alta + + 0
Proteína não degradável + + 0
Aminoácidos limitantes + + 0
Dieta total misturada (TMR) ++ + +
Grão úmido de milho + + -
Milho finamente moído + + -
Uso de tampões + 0 +
Conclusões
Nutrição, manejo e melhoramento genético são os principais meios de se alterar a
composição do leite. Mudanças na composição do leite através da genética são obtidas a
longo prazo quando comparadas às mudanças produzidas por práticas de nutrição e de

157
manejo, que geralmente ocorrem de forma quase imediata. Por outro lado, as mudanças
obtidas pela genética são permanentes, ao contrário das alterações proporcionadas pelas
práticas de nutrição, que são transitórias. Nutricionalmente, os teores de gordura e proteína
no leite são antagônicos. Assim, práticas nutricionais e de manejo que aumentam a produção
de leite também aumentam os teores de proteína do leite, mas por outro lado estas mesmas
práticas diminuem os teores de gordura do leite.
Considerações finais
O processo de valorização da qualidade do leite não pode ser negligenciado para uma data
futura. Se o sistema de pagamento aos produtores não for compatível com as exigências do
mercado consumidor, a sobrevivência de toda a cadeia produtiva do leite nacional estará
seriamente comprometida. A indústria de lácteos brasileira somente poderá competir com
outras bebidas (refrigerantes, sucos, água, cerveja, bebida de soja, etc.) se colocar à
disposição do mercado consumidor um produto seguro, nutritivo e saboroso.
Por fim, acreditamos piamente que os sistemas de pagamento de leite por qualidade vieram
para ficar. Cada vez mais, produtores que entregam leite com altos teores de gordura e de
proteína e baixas CCS e CBT, receberão premiações cada vez mais altas pelo seu leite. De
maneira oposta, produtores com leite de altas CCS e CBT e baixos teores de gordura e
proteína serão crescentemente penalizados. Por mais “cruel” que esta relação de mercado
pode parecer a alguns, esta tendência será positiva para o setor e principalmente para o
consumidor de lácteos brasileiro, pois neste momento “leite de qualidade” no Brasil passará
a ser a regra, e não mais a exceção.
Referências
Almeida, R.; Ostrensky, A. 2011. Aditivos para gado leiteiro. Manejo Alimentar de Bovinos. Editores C.M.M.
Bittar, F.A.P. Santos, J.C. Moura e V.P. Faria. Piracicaba: FEALQ, p.217-256.
Almeida, R. 2012. Nitrogênio ureico no leite como ferramenta para ajuste de dietas. In: II Simpósio
Internacional em Formulação de Dietas para Gado Leiteiro. Anais... Lavras, p.35-65.
Bauman, D.E.; Griinari, J.M. 2003. Nutritional regulation of milk fat synthesis. Annu. Ver. Nutr. 26, 203-227.
Bauman, D.E.; Hard, D.L.; Crooker, B.A.; Partridge, M.S. et al. 1989. Long-term evaluation of prolonged-
release formulation of N-methionyl bovine somatotropin in lactating dairy cows. Journal of Dairy Science 72,
642-651.
Bernal-Santos, G.; Perfield, J.W.; Barbano, D.M. et al. 2003. Production responses of dairy cows to dietary
supplementation with conjugated linoleic acid (CLA) during the transition period and early lactation. Journal
of Dairy Science 86, 3218-3228.

158
Castañeda-Gutierrez, E.; Benefield, B.C.; Veth, M.J. et al. 2007. Evaluation of the mechanism of action of
conjugated linoleic acid isomers on reproduction in dairy cows. Journal of Dairy Science 90, 4253-4264.
Chouinard, P.Y; Girard, V.; Brisson, G.J. 1998. Fatty acid profile and physical properties of milk fat from cows
fed calcium salts of fatty acids with varying unsaturation. Journal of Dairy Science 81,471-481.
Davis, C.L. 1967. Acetate production in the rumen of cows feed either control low-fiber, high-grain diets.
Journal of Dairy Science 50, 1621-1625.
Doska, M.C.; Silva, D.F.F.; Horst, J.A.; Valloto, A.A.; Rossi Jr, P.; Almeida, R. 2012. Sources of variation in
milk urea nitrogen in Paraná dairy cows. Revista Brasileira de Zootecnia 41, 692-697.
Duffield, T.F.; Rabiee, A.R.; Lean, I.J. 2008. A meta-analysis of the impact of monensin in lactating dairy
cattle. Part 2. Production effects. Journal of Dairy Science 91, 1347-1360.
French, N.; Kennelly, J.J. 1990. Effects of feeding frequency on ruminal parameters, plasma insulin milk yield,
and milk composition in Holstein cows. Journal of Dairy Science 73, 1857-1863.
Heuer, C.; Schukken, Y. H.; Dobbelaar, P. 1999. Postpartal body condition score and first milk test results as
predictors of disease, fertility, production, and culling in commercial dairy herds. Journal of Dairy Science 82,
295–304.
Hutjens, M.F.; Shanks, R.D. 1993. Feeding for milk composition. In: 1993 Tri-State Dairy Nutrition
Conference. Proceedings... Fort Wayne, p.162.
Hutjens, M.F. 2010. Sorting through your feed additive choices. Hoard’s Dairyman, September 25, p.620-621.
Jenkins, T.C. 1993. Lipid metabolism in the rumen. Journal of Dairy Science 76, 3851-3863.
Jonker, J.S.; Kohn, R.A.; Erdman, R.A. 1998. Using milk urea nitrogen to predict nitrogen excretion and
utilization efficiency in lactating dairy cows. Journal of Dairy Science 81, 2681-2692.
Jonker, J.S.; Kohn, R.A.; High, J. 2002. Dairy herd management practices that impact nitrogen utilization efficiency.
Journal of Dairy Science 85, 1218-1226.
Kohn, R.A.; Kalscheur, K.F.; Russek-Cohen, E. 2002. Evaluation of models to estimate urinary nitrogen and
expected milk urea nitrogen. Journal of Dairy Science 85, 227-233.
Mertems, D.R. 1997. Creating a system for meeting the fiber requirements of dairy cows. Journal of Dairy
Science 80, 1463-1481.
National Research Council. 1989. Nutrient requirements of dairy cattle. 6th ed. Washington, D.C.: National
Academy Press.
National Research Council. 2001. Nutrient requirements of dairy cattle. 7th ed. Washington, D.C.: National
Academy Press.
Nayeri, A.; Upah, N.C.; Sucu, E.; Pearce, S.C.; Fernandez, M.V. et al. 2011. Potential nutritional strategies to
mitigate the negative effects of heat stress. In: 2011 Four-State Dairy Nutrition and Management Conference.
Proceedings... Dubuque, p.128-133.
Poncheki, J.K. 2015. Avaliação do manejo de vacas no período de transição utilizando as informações do
primeiro controle leiteiro após o parto. Curitiba. Dissertação [Mestrado em Ciências Veterinárias].
Universidade Federal do Paraná.
Rabiee, A.R.; Breinhild, K.; Scott, W.; Golder, H.M. et al. 2012. Effect of fat additions to diets of dairy cattle
on milk production and components: A meta-analysis and meta-regression. Journal of Dairy Science 95, 3225-
3247.
Rajala-Schultz, P.J.; Saville, W.J.A. 2003. Sources of variation in milk urea nitrogen in Ohio dairy herds.
Journal of Dairy Science 86, 1653-1661.
Robinson, P.H. 2010. Impacts of manipulating ration metabolizable lysine and methionine levels on the
performance of lactating dairy cows: A systematic review of the literature. Livestock Science 127, 115-126.

159
Robinson, P.H. 1989. Dynamic aspects of feeding management for dairy cows. Journal of Dairy Science 72,
1197-1209.
Santos, J.E.P.; Greco, L.F. 2012, Leveduras vivas e cultivo de leveduras em dietas de bovinos leiteiros. II
Simpósio Internacional em Formulação de Dietas para Gado Leiteiro. Anais... Lavras, p.09-33.
Sova, A. D.; Leblanc, S. J.; McBride, B. W.; Devries, T. J. 2013. Associations between herd-level feeding
management practices, feed sorting, and milk production in freestall dairy farms. Journal of Dairy Science 96,
4759-4770.
St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. 2003. Economic losses from heat stress by US livestock industries.
Journal of Dairy Science 86, E52-E77.
Van der Drift, S.G.A.; Jorritsma, R.; Schonewille, J.T. et al. 2012. Routine detection of hyperketonemia in dairy
cows using Fourier transform infrared spectroscopy analysis of ß-hydroxybutyrate and acetone in milk in
combination with test-day information. Journal of Dairy Science 95, 4886-4898.
Van Haelst, Y.N.T.; Beeckman, A.; Van Knegsel, A.T.M.; Fievez, V. 2008. Elevated concentrations of oleic
acid and long-chain fatty acids in milk fat of multiparous subclinical ketotic cows. Journal of Dairy Science 91,
4683-4686.
Whitlock, L.A.; Schingoethe, D.J.; Hippen, A.R. et al. 2002. Fish oil and extruded soybeans fed in combination
increase conjugated linoleic acids in milk of dairy cows more than when fed separately. Journal of Dairy Science
85, 234-243.
Zebelli, Q.; Aschenbach, J.R.; Tafaj, M. et al. 2012. Invited review: Role of physically effective fiber and
estimation of dietary fiber adequacy in high-producing dairy cattle. Journal of Dairy Science 95, 1041-1056.

160
Fundamentos y aplicaciones para la alimentación proteica de vacas
lecheras
Cecilia Cajarville
La vaca lechera, y de manera más general, los rumiantes, poseen características digestivas
que hacen que el aprovechamiento de las proteínas de la dieta sea sustancialmente distinto
que en la mayoría de los animales. Los animales monogástricos y los humanos digieren la
proteína que consumen mediante enzimas presentes en el estómago y en el intestino delgado.
De esta forma, convierten la cadena proteica en subunidades más simples, los aminoácidos,
que son absorbidos en el intestino y son las unidades básicas que contribuyen a formar la
nueva proteína, que el animal utilizará para distintas funciones. Las enzimas presentes en el
tubo digestivo de los rumiantes son básicamente las mismas que en los monogástricos. El
rumiante también absorbe aminoácidos provenientes de la proteína digerida en el intestino,
y éstos son las unidades básicas para la formación de su propia proteína. Sin embargo, el
rumen, compartimento que retiene el flujo de los alimentos y que determina que todo lo que
ingresa al aparato digestivo sea sometido, en primer lugar, a un proceso de fermentación,
determina que las características de la digestión proteica sean radicalmente diferentes en los
rumiantes. En el compartimento ruminal se desarrolla una población microbiana (microbiota)
que mantiene con el rumiante una relación simbiótica y que le proporciona la oportunidad de
digerir la fibra de los alimentos. La microbiota ruminal es, en sí misma, la principal fuente
de proteína de alta calidad para los rumiantes. Una de sus características es la diversidad, y
está representada por bacterias (alrededor de 1010 UFC/ml) con más de 50 géneros, arqueas
(107-109 UFC/mL), protozoos (104-106/mL), hongos (103-105 zoosporas/mL) y bacteriófagos
(108-109/mL) (Kamra, 2005). El pasaje de las proteínas ingeridas por el rumen hace que éstas
sean sometidas a la acción de la microbiota, modificando sustancialmente su estructura y
determinando que la alimentación proteica de los rumiantes se realice sobre bases singulares
(Figura 1). La digestión de las proteínas en los rumiantes, incluye no sólo procesos de
degradación, sino también de formación de compuestos nuevos y síntesis. Conocer estos
procesos es útil para manipular la alimentación proteica de los rumiantes, por lo que a
continuación nos centraremos con más detalle en cada uno de ellos.

161
Figura 1. Esquema de la digestión de las proteínas por los rumiantes
Degradación de las proteínas en el rumen
Gran parte de la proteína que ingiere el rumiante se degrada a péptidos por acción de las
proteasas microbianas. La porción de proteína que no sufrió el ataque de los
microorganismos, seguirá su trayecto hasta el intestino delgado, donde será atacada por las
enzimas del animal, desdoblada hasta aminoácidos que serán absorbidos y utilizados para
formar las proteínas del animal y de los productos como la leche. Esta proteína es
denominada proteína de pasaje (by-pass). La cantidad de proteína de pasaje que proporciona
un alimento depende del tipo de proteína. Hay proteínas naturalmente más resistentes a la
degradación como las del maíz (zeína) y también la degradación depende de los tratamientos
a los que haya sido sometida esa proteína (Van Soest, 1994). En general, los tratamientos
térmicos (calor) que se realizan sobre los alimentos tienden a disminuir su degradación en el
rumen. A modo de ejemplo, las harinas animales, como la harina de pescado, tienen baja
degradabilidad ruminal, debido al tratamiento térmico recibido durante su elaboración
(FEDNA, 2010), por lo que se las considera una buena fuente de proteína de pasaje para los
rumiantes. La proteína que se degrada, lo hace primeramente a péptidos, que a su vez se
catabolizan formando aminoácidos libres. Esta degradación continúa y los microorganismos
desagregan los aminoácidos hasta amoníaco, ácidos grasos y CO2. El amoníaco formado es
utilizado por la población microbiana para sintetizar sus propias proteínas, aunque como se
verá luego, para que esta síntesis se produzca debe existir suficiente energía (carbonos), que
en general es aportada por carbohidratos. El amoníaco que no es captado por los

162
microorganismos se absorbe por las paredes del rumen, va al hígado y ahí es transformado
en urea. A su vez, parte de esa urea volverá al rumen a través principalmente de la saliva y
la que no se recicla se elimina por la orina y la leche. En nuestras latitudes, gran parte de la
producción lechera se realiza sobre pastoreo de praderas de gramíneas y leguminosas de
clima templado. Cuando son consumidas en el momento óptimo, tienen una composición
química caracterizada por niveles altos de proteína cruda (15-20%) y de fibras digestibles
(alrededor de 40 % de FND), pero que son de degradación relativamente lenta, y niveles de
carbohidratos de degradación rápida (azúcares) no muy altos (de 3-10%) (Sauvant et al.,
1995; Cajarville et al., 2007; Cajarville et al., 2015). A su vez, la proteína de estos forrajes,
es altamente soluble y de muy rápida degradación (Sauvant et al., 1995; Cohen, 2001;
Repetto et al., 2005), dando como resultado altas concentraciones instantáneas de amonio en
el rumen (Figura 2), que en buena parte son desaprovechadas y se excretan por orina y leche.
Debido a este desperdicio, actualmente se trabaja en la disminución de la degradación de los
componentes nitrogenados de las pasturas de alta calidad mediante el uso de compuestos
como los taninos (Kozloski et al., 2012). La alta degradación en rumen de los compuestos
nitrogenados de las pasturas hace que, en los sistemas pastoriles, o con altas ingestas de
forrajes de buena calidad, sea muy habitual observar altas concentraciones de urea en la leche
de las vacas, sin que ello implique la necesidad de ajustar las dietas.
Debido a este proceso de degradación hasta amoníaco, y la posibilidad de utilización de los
microorganismos de este compuesto, es que es conocida la habilidad de los rumiantes para
aprovechar como fuentes para la síntesis proteica materias nitrogenadas (MN) que no sean
proteínas verdaderas, es decir, fuentes de nitrógeno no proteico (NNP). El ejemplo más
conocido en este sentido es el uso de urea en las dietas como suplemento “proteico”. En el
rumen, la molécula de urea se desdobla muy rápidamente hasta amoníaco, y si no está
acompañada por carbohidratos de rápida fermentación (que aporten las cadenas carbonadas
para la síntesis microbiana) o es suministrada en exceso, la rápida absorción del amoníaco a
través de las paredes del rumen representa un potencial peligro de intoxicación para el animal.

163
Figura 2. Evolución del pH (A) y del amoníaco ruminal (B) a lo largo del día en vacas en
praderas templadas, suplementadas con trigo (cuadrados abiertos) o maíz (triángulos
negros) (medias ± error estándar). Las áreas en negro representan la proporción de vacas
pastoreando luego del período de confinamiento (suplementación). Las flechas indican el
momento de la suplementación; las líneas bajo las flechas indican los períodos de
confinamiento (Cajarville et al., 2006a)
Transformación de los compuestos nitrogenados en proteína microbiana
La masa microbiana que pasa junto con el bolo alimenticio a los tramos posteriores del
aparato digestivo, constituye el principal aporte de proteínas para el rumiante. Esta masa
microbiana contiene aproximadamente 50% de MN de las cuales el 80% se encuentra bajo
forma de proteína. La optimización en el crecimiento de la masa microbiana es uno de los
factores que deben preocupar al nutricionista. El término optimización de la masa microbiana
formada en el rumen implica dos conceptos:
1. Producción microbiana. Indica la generación de microorganismos en términos de
cantidades diarias. Se refiere a kg de masa microbiana (N microbiano) formado en el
rumen por día. Es variable con el tipo de animal, tamaño del rumen, cantidad de alimento
consumido, etc.

164
2. Eficiencia de producción de proteína microbiana. Se refiere a la cantidad de
microorganismos generados por unidad de alimento. Se expresa de diferentes formas:
masa microbiana (N microbiano) en función de la materia seca ingerida, de la materia
orgánica (MO) ingerida, o de la MO digerida o fermentada en el rumen (MODR o MOF,
respectivamente).
Ambos conceptos son importantes, pero si se trata de evaluar la efectividad de un alimento o
una dieta para proveer proteína microbiana, consideramos indicado trabajar en base a la
eficiencia de producción de proteína microbiana. Stern et al. (1994) calcularon la
contribución teórica de la proteína microbiana al total de la proteína que llega al duodeno
según la eficacia de síntesis. Cuando la eficacia de síntesis es baja, del orden de 20 g de N/kg
de materia orgánica verdaderamente fermentada en rumen (MOVF), la proteína microbiana
puede cubrir cerca del 50% de las necesidades proteicas de una vaca de 600 kg produciendo
25 litros de leche con 4% de grasa. Sin embargo, si la eficacia de síntesis fuera elevada (40
g N/kg MOVF) la proteína microbiana podría llegar a cubrir un 98% de las necesidades
proteicas del mismo animal. Por esta razón, las estrategias de alimentación en rumiantes
actualmente tienden a optimizar la producción microbiana a nivel ruminal (Shingoethe,
1996a; NRC, 2001). Para lograr la máxima eficiencia, el aporte de nutrientes para los
microorganismos, especialmente materias nitrogenadas y carbohidratos (o más
genéricamente materia orgánica fermentable en el rumen), debe ser el necesario (Russell et
al., 1992; Clark et al., 1992). Además, y aunque algunos autores ponen en duda la posibilidad
de manejar este concepto en las dietas (Valkeners et al., 2006; Hall y Huntington, 2008),
carbohidratos y materias nitrogenadas deberían estar disponibles en el rumen en forma
sincrónica, es decir, a la vez. Teóricamente, proporcionando a la microbiota ruminal las
fuentes de proteína y energía en forma simultánea y en las cantidades requeridas, se lograría
el máximo crecimiento microbiano y por lo tanto, el mayor aporte de nutrientes para el
animal.
Las materias nitrogenadas para los microorganismos ruminales provienen de la proteína y
del NNP de los alimentos que son degradados en el rumen, además de la urea recirculante
que ingresa por vía sanguínea o por la saliva. No aportan al crecimiento microbiano la
proteína indigestible (como la unida a la lignina o a algunos tipos de taninos) ni tampoco la
proteína de pasaje (Van Soest, 1994). Es necesario puntualizar, que no todos los compuestos

165
que tienen valor nutricional para el rumiante, son igualmente útiles para los
microorganismos. La materia orgánica utilizable por estos últimos, es la que tiene capacidad
de ser fermentada en el rumen (materia orgánica fermentable o fermentescible: MOF). Las
paredes celulares, el almidón y la proteína degradable en el rumen, junto con los azúcares,
son fuente de energía fermentescible formando MOF. Otros compuestos, a pesar de ser muy
energéticos para el animal, como los lípidos, ácidos orgánicos o el almidón no degradable en
rumen, así como la proteína no degradable, no forman parte de la MOF. Debido a su baja
fermentación ruminal, poco aportan para el crecimiento microbiano. Como se desprende de
lo anterior, la síntesis de proteína microbiana depende mucho de las cantidades de MOF en
el rumen. Ello hace que el nutricionista deba moverse en el difícil equilibrio entre salud y
enfermedad, considerando que altos niveles de MOF en el rumen siempre implican riesgo de
aparición de acidosis. De hecho, revisiones de trabajos publicados, reportan que las mayores
síntesis de proteína microbiana se observan a niveles de pH ruminal relativamente bajos,
incluso menores a 6,0 (Bach et al., 2005). Por otra parte, la fuente de carbohidrato y la cinética
de su disponibilidad en el rumen, son factores determinantes en el logro de la sincronización
de nutrientes (Lee et al., 2003). En este sentido, la disponibilidad de energía a nivel ruminal
puede ser muy distinta según la fuente de carbohidrato de que se trate, debido a las diferentes
velocidades de fermentación. La velocidad es muy rápida para los solubles como los azúcares
y muy lenta para los estructurales como la celulosa. En general, se acepta que una rápida
disponibilidad de energía a partir de carbohidratos es clave para lograr el crecimiento de la
masa microbiana (Henning et al., 1991). Sin embargo, hasta el momento, los modelos de
predicción de respuestas que se utilizan para la formulación de dietas (NRC, 2001) son
relativamente insensibles a cambios en las cinéticas de degradación de los carbohidratos
(Hall, 2004). De todas formas, más allá de las cinéticas de degradación per se de los
carbohidratos, existen otros factores como el pH ruminal, la cantidad de concentrado
suministrado, o la combinación de compuestos que modifican la cinética de degradación del
almidón y de los carbohidratos no amiláceos (Huntington, 1997). Todos estos aspectos deben
ser considerados ya que muchas veces son los que hacen los resultados no sean los previstos
cuando se formula una dieta. Las cantidades de energía y N disponible en el rumen son los
dos factores principales, pero no los únicos. Otros factores, como la disponibilidad de
minerales, sobre todo azufre, de oligoelementos y de vitaminas intervienen también.

166
Adicionalmente, en la actualidad se le da especial importancia a alimentación con fuentes de
proteína verdadera, dado que se ha demostrado que los microorganismos necesitan de
algunos aminoácidos y péptidos preformados para un mejor desarrollo (Broderick y Reynal,
2009).
Digestión y absorción intestinal de la proteína
La proteína que llega al duodeno de un rumiante es principalmente la suma de la proteína
microbiana y la proteína de origen alimenticio que no ha sufrido degradación a nivel ruminal.
La digestión en el abomaso y en el intestino delgado, es muy similar a la de los
monogástricos. Las proteínas son hidrolizadas por enzimas pancreáticas en péptidos, y
posteriormente en oligopéptidos y aminoácidos. La absorción se realiza bajo forma de
aminoácidos en el intestino delgado. Esta digestión no es total, y una parte de las proteínas
alimenticias y microbianas que escapa a la misma pasa directamente a las heces. En el
intestino grueso las proteínas se degradan poco, ya que las bacterias del intestino grueso
utilizan principalmente la urea endógena para la síntesis de sus proteínas. En este paso es que
surge el concepto de Proteína Metabolizable, que emplean varios sistemas de alimentación
de los rumiantes. De acuerdo con el NRC (2001), la proteína metabolizable es la proteína
digerida luego del rumen, cuyos aminoácidos son absorbidos en el intestino. Está compuesta
por la sumatoria de la proteína microbiana digestible (más abundante), la proteína de pasaje
digestible, y una porción minoritaria compuesta por proteína endógena, proveniente de
proteínas de descamación y enzimas. Un aspecto a destacar, es que mientras que el valor
biológico de la proteína de origen alimenticio que llega al duodeno es muy variable, la
composición en aminoácidos de la proteína de origen microbiano es muy estable y de muy
alta calidad (Schingoethe, 1996b). Así, el contenido en lisina y metionina de los cuerpos
microbianos es casi tan alto como el de la carne o el pescado, por lo que la proteína
microbiana se encuentra dentro de las proteínas de mejor calidad (Verité y Peyraud, 1990).
Esta alta digestibilidad y excelente perfil aminoacídico es otra razón por la que el
nutricionista debe intentar, mediante el manejo de la alimentación, maximizar el aporte de
proteína microbiana al duodeno. La herramienta principal para ello será el logro de un buen
funcionamiento del rumen, considerando todos los aspectos que tiendan a mejorar la síntesis
de proteína microbiana.

167
Para concluir
A modo de resumen, es conveniente recordar que la síntesis de proteína microbiana en el
rumen es el mecanismo que permite la generación de proteína de alta calidad y la
optimización en el uso de los recursos nitrogenados por la vaca lechera, y por los rumiantes
en general. Por esta razón es que como nutricionistas nos interesa lograr altos niveles y
eficiencias de síntesis. Lo anterior se logrará manteniendo el ecosistema ruminal funcionando
al máximo, especialmente en lo que refiere al mantenimiento de niveles adecuados de
nitrógeno y de materia orgánica fermentable.
Referencias
Bach A., Calsamiglia S., Stern M. 2005. Nitrogen Metabolism in the Rumen. Journal of Dairy Sci. 88, E-
Supplement, E9-E21.
Broderick G., Reynal S. 2009. Effect of Source of Rumen-Degraded Protein on Production and Ruminal
Metabolism in Lactating Dairy Cows. Journal of Dairy Sci. 92, 2822-2834.
Cajarville C., Aguerre M., Repetto J.L. 2006a. Rumen pH, NH3-N concentration and forage degradation kinetics
of cows grazing temperate pastures and supplemented with different sources of grain. Animal Research. 55,
511-520.
Cajarville C., Pérez A., Aguerre M., Britos A., Repetto J.L. 2006b. Effect of the timing of cut on ruminal
environment of lambs consuming temperate pastures. Journal of Dairy Science 89, Suppl. 1, 103.
Cajarville C., Britos A., Caramelli A., Antúnez M., Zanoniani R., Boggiano P., Repetto J.L. 2007. El horario
de corte y el tipo de metabolismo fotosintético afectan la relación azúcares/nitrógeno de las pasturas. Archivos
Latinoamericanos de Producción Animal 15 Supl. 1, 408-409.
Cajarville C., Britos A., Errandonea N., Gutiérrez L., Cozzolino D., Repetto J.L. 2015. Diurnal changes in
water-soluble carbohydrate concentration in lucerne and tall fescue in autumn and the effects on in vitro
fermentation. New Zealand Journal of Agricultural Research. DOI: 10.1080/00288233.2015.1018391.
Clark J.H., Klusmeyer T.H., Cameron M.R. 1992. Microbial protein synthesis and flows of nitrogen fractions
to the duodenum of dairy cows. Journal of Dairy Science 75, 2304.
Cohen D.C. 2001. Degradability of crude protein from clover herbages used in irrigated dairy production
systems in Northern Victoria. Australian Journal of Agricultural Research. 52, 415-425.
FEDNA. 2010. Tablas FEDNA de composición y valor nutritivo de alimentos para la fabricación de piensos
compuestos (3º ed). C. de Blas, G.G. Mateos, P.G. Rebollar. Fundación Española para el Desarrollo de la
Nutrición Animal. Madrid, 502 p.
Hall M.B. 2004. Effect of carbohydrate fermentation rate on estimates of mass fermented and milk response.
Journal of Dairy Science 87, 1455-1456.
Hall M.B., Huntington G.B. 2008. Nutrient siynchrony: Sound in theory, elusive in practice. Journal of Animal
Science 86 (E. Suppl.), E287-E292.
Henning, P.H., Steyn D.G., Meissner. 1991. The effect of energy and nitrogen supply patterns on rumen
bacterial growth in vitro. Animal Production 53, 165-175.
Huntington G. B. 1997. Starch utilization by ruminants: from basics to the bunk. Journal of Animal Science 75,
852-867.

168
Kamra, D.N. 2005. Rumen microbial ecosystem. Current Science 89,124-135.
Kozloski G.V., Härter C.J., Hentz, F., de Avila S.C., Orlandi T., Stefanello, C.M. 2012. Intake, digestibility and
nutrients supply to wethers fed ryegrass and intraruminally infused with levels of Acacia mearnsii tannin
extract. Small Ruminant Research 106, 125-130.
Lee M.R.F., Merry R.J., Davies D.R., Moorby J.M., Humphreys M.O., Theodorou M.K., MacRae J.C., Scollan
N.D. 2003. Effect of increasing availability of water-soluble carbohydrates on in vitro rumen fermentation.
Animal Feed Science and Technology 104, 59-70.
NRC. 2001. Nutrient requirements of Dairy Cattle: 7th Revised Edition, 2001. National Academy Press,
Washington, DC.
Repetto J.L., Cajarville C., D’Alessandro J., Curbelo A., Soto C., Garín D. 2005. Effect of wilting and ensiling
on ruminal degradability of temperate grass and legume mixtures. Animal Research 54, 73-80.
Russell J. B., O'Connor J. D., Fox D. G., Van Soest P. J., Sniffen C. J. 1992. A net carbohydrate and protein
system for evaluating cattle diets: I. Ruminal fermentation. Journal of Animal Science 70, 3551-3561.
Sauvant D., Grenet E., M-Doreau B. 1995. Dégradation chimique des aliments dans le réticulo-rumen: cinétique
et importance. En: Nutrition des ruminants domestiques. Ed. INRA, Paris.
Schingoethe D.J. 1996a. Balancing the amino acids needs of dairy cows. Animal Feed Science and Technology
60, 153.
Schingoethe D.J. 1996b. Dietary influence on protein level in milk and milk yield in dairy cows. Animal Feed
Science and Technology 60, 181.
Stern M.D. Calsamiglia S., Endres M.I. 1994. Dinámica del metabolismo de los hidratos de carbono y del
nitrógeno en el rumen. En: Nuevos sistemas de valoración de alimentos y programas alimenticios para especies
domésticas. Ed. FEDNA, Madrid.
Valkeners D., Thewis A., Amant S., Beckers Y. 2006. Effect of various levels of imbalance between energy
and nitrogen release in the rumen on microbial protein synthesis and nitrogen metabolism in growing double-
muscled Belgian Blue bulls fed a corn silage-based diet. Journal of Animal Science 84, 877–885.
Van Soest P. 1994. Nutritional Ecology of the Ruminant (2nd edition). Cornell University Press, NY, USA.
Verité R., Peyraud J.L. 1990. Nutrición Nitrogenada. En: Alimentación de bovinos, ovinos y caprinos. Ed.
INRA.

169
Cetose clínica e subclínica: manejo, diagnóstico e efeitos no leite
Marcio Nunes Corrêa
Camila Pizoni
Joabel Tonellotto dos Santos
Patrícia Mattei
Rafael da Fonseca Prietsch
Uriel Secco Londero
Durante os últimos anos a produção de leite por vaca apresentou um aumento substancial,
resultado do aprimoramento das técnicas de seleção genética, melhora da nutrição e práticas
de manejo (Espósito et al., 2014). Entretanto, esse aumento trouxe também como
consequência uma maior ocorrência de transtornos metabólicos, que são mais frequentes
durante o período de transição, compreendido entre as três semanas pré e as três semanas
pós-parto (Drackley, 1999; Chapinal et al., 2011). Dentre os transtornos metabólicos destaca-
se a cetose, podendo se apresentar na forma subclínica ou clínica principalmente entre a
segunda e a sétima semana de lactação (Gonzalez et al., 2014). Essa enfermidade pode ser
definida como um desequilíbrio entre demanda energética e mobilização de gordura, levando
a um aumento da concentração de corpos cetônicos a níveis tóxicos nos fluidos corporais
(Garro et al., 2014). Sua incidência pode variar de 26 a 60% para a forma subclínica
(Simensen et al.,1990; Duffield et al., 1998; McArt et al., 2012a) e de 2 a 34% para a clínica
(Duffield, 2000; Gonzalez et al., 2014), pois sua ocorrência depende de diversos fatores,
como nível de produção, manejo, número de lactações e escore de condição corporal (ECC)
ao parto (Nielen et al., 1994; Duffield et al., 2009). As altas taxas de incidência, aliadas ao
período de maior ocorrência da cetose fazem com que a enfermidade apresente um
considerável interesse econômico, não apenas pelos gastos decorrentes do tratamento, mas
por perdas na produção leiteira e pela predisposição a outros transtornos (Duffield et al.,
2009; Ospina et al., 2010). Vacas que desenvolvem a doença na forma subclínica na primeira
semana de lactação apresentam um risco 6 vezes maior de desenvolver deslocamento de
abomaso, 4,5 vezes mais chance de serem descartadas do rebanho e produzem 2,2 kg de leite
a menos por dia nos primeiros trinta dias em lactação (DEL) (McArt et al., 2012a). Vacas
com quatro lactações ou mais são as mais atingidas pelo transtorno, com perdas na produção

170
leiteira que chegam a 353,4 kg de leite por lactação (Rajala-Schultz et al., 1999). A
manutenção da saúde e da produtividade de vacas leiteiras no período de transição é uma
tarefa desafiadora. Para cumprir com esse objetivo é necessário compreender os eventos
metabólicos que caracterizam esta fase e suas implicações. Diante desta problemática, serão
abordados alguns tópicos relacionados ao metabolismo da cetose, métodos diagnósticos e
implicações no leite, a fim de disponibilizar informações que possam ser utilizadas para uma
melhor compreensão, prevenção e o tratamento do transtorno.
Metabolismo energético durante o período de transição e sua relação com a cetose
A transição do final da gestação ao início da lactação é um período extremamente dinâmico
para a vaca leiteira (McArt al., 2013). As três semanas que antecedem o parto são marcadas
por um decréscimo de até 30% da ingestão de matéria seca (IMS), limitando a ingesta de
energia (Hayirli et al., 2002). Com o início da lactação e rápido aumento da produção de leite
após o parto, os requerimentos de proteínas, ácidos graxos e aminoácidos aumentam cerca
de 2 a 5 vezes (Bell, 1995; Bertoni et al., 2009), devido ao direcionamento desses nutrientes
para a glândula mamária, em detrimento de outros tecidos (Loor et al., 2013). Entretanto, o
organismo da vaca não é capaz de suprir essa alta demanda o que conduz a um desequilíbrio
energético denominado balanço energético negativo (BEN) (Drackley, 1999). O déficit
energético leva à mobilização de reservas corporais induzindo a lipólise, que ocasiona a
liberação de ácidos graxos não esterificados (AGNE) no sangue (Chapinal et al., 2012). Cerca
de 15 a 20% desses AGNE são removidos pelo fígado (Drackley & Andersen, 2006), onde
poderão seguir três destinos: (1) oxidação completa para fornecer energia ao próprio fígado,
(2) oxidação incompleta para produção de corpos cetônicos, que serão liberados no sangue
para prover energia a outros tecidos e (3) reconversão a triglicerídeos (TAG) para
armazenamento no tecido adiposo (Loor et al., 2013; McArt et al., 2013). Assim, vacas que
apresentam um BEN intenso, com uma alta taxa de lipomobilização experimentam uma
excessiva produção de AGNE e corpos cetônicos, predispondo o animal a quadros de
esteatose hepática e cetose (Loor et al., 2013).
Numa condição de hipoglicemia, como ocorre durante o periparto, a carência de propionato
(principal precursor gliconeogênico) induz a uma alta taxa de lipomobilização e uma alta
produção de acetil coenzima A (acetil-CoA), metabólito final da oxidação completa dos

171
AGNE. Essa produção excessiva do acetil-CoA supera a capacidade de sua utilização pelo
Ciclo de Krebs para a produção de energia, aumentando também a demanda de oxalacetato,
que deve ser condensado ao acetil-CoA para poder entrar no ciclo (Andrew et al., 2012). O
resultado deste acúmulo é o desvio do acetil-CoA para a produção de corpos cetônicos, do
qual os principais são acetoacetato, acetona e beta-hidroxibutirato (BHB) (Gonzalez & Silva,
2006). O acetoacetato é primeiro corpo cetônico, que apresenta elevada instabilidade reativa
(Duffield, 2000) e é então reduzido pela β-hidroxibutirato desidrogenase a BHB (Andrew et
al., 2012). A acetona é produzida em menores quantidades, e por ser volátil e tóxica para o
organismo, é excretada (Gonzalez et al., 2014). Assim, o papel principal dos corpos cetônicos
é a transferência da energia derivada dos lipídios do fígado para tecidos periféricos, para
suprirem as demandas energéticas não atendidas pela falta da glicose (Andrew et al., 2012).
Contudo, quando os níveis de corpos cetônicos no sangue ultrapassam a capacidade de
utilização por esses tecidos ocorre acúmulo, que é tóxico em determinados níveis,
ocasionando um quadro de cetose clínica ou subclínica (Risco, 1992; Duffield et al., 2009).
Em relação à etiologia, se reconhece a incidência da cetose como três diferentes síndromes.
A cetose tipo I ou espontânea ocorre quando a demanda de glicose supera a capacidade de
síntese pelo fígado através da gliconeogênese, mesmo quando esse mecanismo está ativado
ao máximo, e acomete vacas de alta produção. O quadro é caracterizado por hipoglicemia
seguida de lipólise, ocasionando um aumento nos níveis séricos de corpos cetônicos, mas
sem ocasionar a deposição lipídica no fígado (Herdt, 2000). A cetose tipo II ocorre quando
grandes quantidades de AGNE são depositadas no fígado, interferindo na capacidade máxima
da neoglicogênese hepática. Neste quadro, a glicemia não é tão baixa quanto na cetose tipo I
e está também associada à elevação das cetonas sanguíneas (Holtenius & Holtenius, 1996).
Por fim, a cetose alimentar ou toxicose butírica ocorre pela ingestão de silagem de má
qualidade, com excesso de butirato ou pelo feno mal conservado e em decomposição, pela
proliferação de fungos (Radostits et al. 2007).
Diagnóstico de cetose
Em rebanhos leiteiros a cetose é um dos principais transtornos metabólicos do pós-parto
recente, podendo ser dividida na forma clínica e subclínica, sendo a última a que ocasiona as
maiores perdas econômicas. Diante disso, seu diagnóstico se torna crucial para minimizar os

172
impactos negativos nos sistemas de produção. Para um diagnóstico satisfatório de cetose
clínica é essencial saber identificar seus sinais, que podem ser visualizados nos estágios
digestivo e nervoso. A forma digestiva é a mais comum entre os casos clínicos e apresenta
sinais inespecíficos, além de sinais envolvendo o sistema digestório como diarreia e redução
nos movimentos ruminais. Quando o quadro não é tratado rapidamente, torna-se perceptível
odor cetônico na respiração, urina e leite além de depressão e letargia seguida de decúbito.
O estágio nervoso é de menor ocorrência, porém de suma importância, pois faz parte da
evolução da enfermidade. Nesse estágio o animal demonstra mudanças no comportamento
como movimentos circulares, empurrar e lamber objetos, alteração da visão, além de
hiperestesia, tremores e incoordenação motora (Baird, 1982; Gordon et al., 2013). A forma
subclínica da doença é a mais incidente, porém de difícil diagnóstico por apresentar sinais
inespecíficos como inapetência, dorso arqueado, pele ressecada, redução da produção leiteira
e perda de condição corporal (Gonzalez et al., 2014). O diagnóstico é baseado no histórico
do animal, aliado ao exame clínico completo e confirmado através de exames
complementares realizados no sangue, urina, leite e líquido ruminal, que buscam elucidar o
estado metabólico do animal através da avaliação dos níveis de corpos cetônicos (Sato &
Shiogama, 2010; Gordon et al., 2013). Essa avaliação pode ser realizada principalmente
através de métodos cinéticos e colorimétricos. Dentre os métodos cinéticos
(predominantemente laboratoriais), a avaliação sérica de BHB tem se mostrado a mais
confiável forma de identificar o transtorno. Os níveis fisiológicos do metabólito para vacas
leiteiras estão abaixo de 1,0 mmol/L, entre 1,2 e 1,4 mmol/L para forma subclínica e acima
de 1,4 mmol/L para forma clínica (Leblanc, 2010). Também é possível detectar alterações no
metabolismo energético do animal através da presença de cetonas no leite (Wood et al.,
2004). Níveis superiores a 10 mg/dL (1,75 mmol/L) no leite indicam cetose.
A identificação dos casos a campo pode ser realizada através de métodos colorimétricos (fitas
reativas) para mensurar corpos cetônicos tanto na urina quanto no leite. O Keto-Test (Elanco
Animal Health, Sanwa Kagaku Kenkyusho) validado por Oetzel (2004), corresponde a uma
fita que altera sua coloração na presença de anormalidades na concentração de corpos
cetônicos no leite. Do mesmo modo, o Ketostix (Bayer Healthcare LLC), validado por
Carrier et al. (2004), também avalia as concentrações de corpos cetônicos, porém esta análise
é realizada na urina. Outro equipamento que pode ser utilizado a campo para identificação

173
do transtorno é o FreeStyle Optium (Abbott Laboratories) (McArt et al., 2013; Krempasky
et al., 2014; Weng et al., 2015), um equipamento de análise rápida (5 segundos) que avalia
os níveis séricos de BHB, apresentando alta sensibilidade e especificidade tanto para ovinos
quanto para bovinos (Voyvoda & Erdogan, 2010; Panousis et al., 2012). A utilização desses
métodos facilita a rápida detecção e consequente tratamento e/ou prevenção das formas de
cetose.
Efeitos da cetose no leite
O Brasil é o quarto maior produtor mundial de leite com produção anual de 34,2 bilhões de
litros (IBGE, 2014), tendo esta atividade uma significativa importância socioeconômica;
apesar disso a qualidade da matéria-prima ainda é um grande entrave ao desenvolvimento da
cadeia produtiva do leite (Borges, 2009). A qualidade do leite é determinada tanto pelo
processo de produção quanto pela sua composição (Rangel et al., 2009; Galvão et al., 2010).
A redução da IMS no pós-parto recente associada à produção de leite intensificam o BEN,
aumentando a lipomobilização e os níveis séricos de corpos cetônicos. Essa limitação
energética tende a aumentar o teor de gordura e reduzir a proteína no leite, sendo a razão
entre a porcentagem de gordura e proteína usada para monitorar a prevalência de cetose
subclínica no rebanho (Duffield, 2000; Gantner et al., 2009). A relação gordura:proteína
superior a 1,5 na primeira ordenha do dia aponta déficit energético, indicando um quadro de
cetose (Robinson et al., 2003). A associação que existe entre o aumento da gordura no leite
e a hipercetonemia deve-se presumivelmente ao aumento na disponibilidade do BHB e ácidos
graxos para a síntese da gordura do leite (Duffield, 2006; Gonzalez, 2009) provenientes da
β-oxidação do tecido adiposo (Reist et al., 2002). Quanto mais intensa a lipomobilização,
maior será o valor de gordura no leite e maior o risco de distúrbios metabólicos severos no
animal (cetose clínica) (Gonzalez, 2004).
O aumento de corpos cetônicos está diretamente ligado a uma redução da produção de leite,
que atinge valores de 1 a 1,5 litros por dia em animais com cetose subclínica (Duffield, 2000).
As perdas seguem aumentando à medida que os valores de BHB aumentam, havendo perdas
de até 4 kg de leite por dia (Duffield, 1998). A cetose subclínica também diminui o
desempenho reprodutivo (Walsh et al., 2007), que é decorrente do retardo na reativação da

174
função ovariana com aumento no intervalo do primeiro serviço pós-parto, aumento do
número de serviços e maior intervalo parto-concepção (Ingvartsen et al., 2004).
Prevenção, formas de controle e tratamento da doença
A detecção precoce de alterações na saúde da vaca leiteira permite que seja realizada
intervenção antes que os níveis de produção dos animais, ou até mesmo sua sobrevivência
esteja ameaçada (Weng et al., 2015). A prevenção da cetose está relacionada ao manejo do
periparto, mais especificamente com a nutrição adequada (Dann et al., 2006). A verificação
do ECC para monitorar a eficácia do programa nutricional tem efeito significativo sobre o
equilíbrio energético no início da lactação (Wathes et al., 2007) e é uma medida bastante
simples, mas que pode auxiliar muito na prevenção da doença, já que a ocorrência de cetose
tem sido associada à maior lipomobilização no pós-parto (Loor et al., 2013). Vacas com ECC
elevado (> 3,5) no pós-parto recente apresentam um BEN mais exacerbado do que vacas com
ECC adequado (2,5 a 3,5), predispondo o animal a desenvolver cetose (Allbrahim et al.,
2010; Shin et al., 2015). Outra estratégia de reduzir o risco da doença é disponibilizar mais
energia para as vacas no pós-parto recente através do uso de propilenoglicol e gordura
protegida. O propilenoglicol dá origem a grandes quantidades de propionato e glicerol no
rúmen, aumentando os níveis de glicose e insulina e reduzindo AGNE e BHB séricos nesses
animais (Gordon et al., 2013). O uso de gordura protegida com sais de cálcio eleva a
densidade energética da dieta (Ganjkhanlou et al., 2009) sem aumentar a ingestão de
carboidratos ou comprometer a ingestão de fibras, favorecendo um maior aporte de energia
para a síntese do leite e de seus componentes (Desnoyers, 2009). Além disso, a gordura
protegida pode contribuir para a redução da queda abrupta da IMS estabilizando os níveis de
cetonas sanguíneas (Schein, 2012).
A diminuição da IMS reduz a síntese de proteína microbiana limitando a oferta de
aminoácidos essenciais, responsáveis pela formação de precursores gliconeogênicos e
proteínas do leite (Drackley et al., 1999). O uso de aminoácidos protegidos também tem sido
utilizado como estratégia para prevenir a cetose. O aminoácido essencial metionina tem um
papel importante, pois reduz a metanogênese ruminal, desviando átomos de C e H+ do
metano. Esse desvio faz com que haja maior produção de propionato, aumentando a
eficiência energética da dieta em até 4% e reduzindo a cetogênese (Gonzalez et al., 2014).

175
Outros dois aminoácidos importantes são a niacina e a colina, sintetizados pelos
microrganismos ruminais. A niacina é necessária para síntese de compostos NAD+ e NADP+,
coenzimas essenciais no metabolismo de carboidratos, lipídios e proteínas. Sua utilização
ocasiona a redução da lipidose e redução do fluxo de AGNE no sangue (Yuanet al., 2012).
A colina tem uma importante ação na formação de fosfolipídios que estruturam lipoproteínas,
essenciais para o transporte dos TAG (Zom et al., 2011). Sua suplementação na forma
protegida tem se mostrado eficaz na redução da cetogênese, através da diminuição da
esterificação de ácidos graxos e do aumento na secreção de lipoproteínas (Piepenbrink &
Overton, 2003), além da redução de esteatose hepática (Cooke et al., 2007).
Como alternativa preventiva e/ou de tratamento o uso de butafosfan, um composto a base de
fósforo orgânico associado à cianocobalamina tem se mostrado eficiente (Fürll et al., 2010;
Rollin et al., 2010; Nuber et al., 2015), visto que age no ciclo de Krebs disponibilizando mais
energia e reduzindo a expressão de genes relacionados à cetogênese e oxidação de ácidos
graxos (Kreipe et al., 2011). Além disso, ele reduz a intensidade do BEN, através da
diminuição das concentrações plasmáticas de AGNE e BHB, aumentando a produção de leite
(Pereira et al., 2013). A utilização de cápsulas de liberação controlada de monensina também
são eficientes na prevenção de cetose clínica e subclínica, aumentando a produção ruminal
de ácido propiônico e reduzindo as concentrações de ácido acético e butírico, reduzindo a
formação de corpos cetônicos (Melendez et al., 2006).
Existem diferentes terapias associadas ao tratamento da cetose que se baseiam em aumentar
a glicose sanguínea, disponibilizando precursores gliconeogênicos e reduzindo a formação
de corpos cetônicos (Andrews et al., 2004). O tratamento mais comumente utilizado é a
administração endovenosa de glicose 50% com uma dosagem que varia de acordo com a
lactose eliminada no leite (Radostits et al., 2007); entretanto, o incremento energético dessa
estratégia é transitório (duas a três horas) (Gordon et al., 2013) e se não forem tomadas outras
medidas o quadro clínico pode retornar. Para tanto, utiliza-se a associação de glicose 50% ao
uso de glicocorticoides para manter a glicemia elevada por mais tempo (oito a dez horas)
(Schein, 2012). A insulina também pode ser utilizada juntamente com a glicose 50%, tendo
efeito anabólico que irá aumentar a síntese de gordura e o uso dos corpos cetônicos como
fonte energética. Seu uso pode ser importante em casos em que o animal não responde ao
tratamento com glicose ou glicocorticoides (Hayirli, 2006; Smith, 2008). No entanto, o início

176
da lactação, que é o período de maior ocorrência da doença, é marcado por uma resistência à
insulina transitória, que pode afetar negativamente esse protocolo de tratamento, além de ele
ser pouco explorado pelo elevado custo do produto (Gordon et al., 2013).
Outra estratégia comumente utilizada é a administração via oral de drench, que tem como
base o propilenoglicol. Ao chegar ao rúmen esse composto é diretamente absorvido, sendo
utilizado no ciclo de Krebs para aumentar a oxidação de acetil-CoA e estimular a
gliconeogênese ou é convertido em propionato, que também pode ser direcionado para esta
via e ainda estimula a liberação de insulina (Gordon et al., 2013). Este composto, utilizado
em vacas com cetose subclínica pode reduzir os casos de deslocamento de abomaso,
aumentar a taxa de concepção ao primeiro serviço (McArt et al., 2012b), aumentar a produção
de leite (Nielsen & Ingvartsen, 2004) e prevenir a cetose clínica (McArt et al., 2011). Além
disso, seu uso pode ser associado a glicocorticoides, que aumentam indiretamente a glicemia
através do bloqueio dos efeitos da insulina, sem influenciar a lipólise e diminuindo as
concentrações de BHB (Drift et al., 2015).
Considerações finais
Vacas que apresentam BEN mais acentuado são mais susceptíveis a desenvolverem cetose.
A prevenção, diagnóstico e controle desse transtorno são essenciais para evitar ou minimizar
os efeitos negativos na produção e qualidade do leite, reduzindo as perdas econômicas e
possibilitando ao animal expressar sua maior capacidade produtiva.
Referências
Allbrahim, R.M.; Crowe, M.A.; Duffy, P. et al. 2010. The effect of body condition at calving and
supplementation with Saccharomyces cerevisiae on energy status and some reproductive parameters in early
lactation dairy cows. Animal Reproduction Science 121, 63-71.
Andrew, P.; McPherson, C.; McEneny, J. 2012. The biochemistry of ketogenesis and its role in weight
management, neurological disease and oxidative stress. Journal of Physiology Biochemistry 68, 141-151.
Baird, G.D. 1982. Primary ketosis in the high-producing dairy cow: clinical and subclinical disorders, treatment,
prevention, and outlook. Journal of Dairy Science 65, 1-10.
Bell, A.W. 1995. Regulation of organic nutrient metabolism during transition from late pregnancy to early
lactation. Journal of Animal Science 73, 2804–2819.
Bertoni, G., Trevisi, E.; Lombardelli, R. 2009. Some new aspects of nutrition, health conditions and fertility of
intensively reared dairy cows. Italian Journal of Animal Science 8, 491–518.
Borges, K.A.; Reichert, S.; Zanela, A M.B.; Fischer, V. 2009. Avaliação da qualidade do leite de propriedades
da região do Vale do Taquari no estado do Rio Grande do Sul. Acta Scientiae Veterinariae 37, 39-44.

177
Carrier, J.; Stewart, S., Godden, S. et al. 2004. Evaluation and use of three cowside tests for detection of
subclinical ketosis in early postpartum cows. Journal of Dairy Science 87, 3725-3735.
Chapinal, N.; Carson, M.E.; Duffield, T.F. et al. 2011. The association of serum metabolites with clinical
disease during the transition period. Journal of Dairy Science 94, 4890-4903.
Chapinal, N.; Carson, M.E.; Leblanc, S.J. et al. 2012. The association of serum metabolites in the transition
period with milk production and early-lactation reproductive performance. Journal of Dairy Science 95, 130-
1309.
Cooke, R. F.; Silva, D.R.N.; Caraviello, D.Z. et al. 2007. Supplemental choline for prevention and alleviation
of fatty liver in dairy cattle. Journal of Dairy Science 90, 2413–2418.
Dann, H.M.; Litherland, N.B.; Underwood, J.P. et al. 2006. Diets during far-off and close-up dry periods affect
periparturient metabolism and lactation in multiparous cows. Journal of Dairy Science 89, 3563-3577.
Desnoyers, M.; Giger-Reverdin, S.; Bertini, G. et al. 2009. Meta-analysis of the influence of Saccharomyces
cerevisiae supplementation on ruminal parameters and milk production of ruminants. Journal of Dairy Science
92, 1620-1632.
Drackley, J.K. 1999. Biology of dairy cows during the transition period: the final frontier? Journal of Dairy
Science 82, 2259–2273.
Drackley, J.K.; Andersen, J.B. 2006. Splanchnic metabolism of long-chain fatty acids in ruminants. In:
Ruminant Physiology: Digestion, Metabolism and Impact of Nutrition on Gene Expression, Immunology and
Stress. Academic Publishers, Wageningen, The Netherlands, p. 199–224.
Duffield, T. F.; Sandals, D.; Leslie, K.E. et al. 1998. Efficacy of monensin for prevention of subclinical ketosis
in lactating dairy cows. Journal of Dairy Science 81, 2866–2873.
Duffield, T. 2000. Subclinical ketosis in lactating dairy cattle. Veterinary Clinics North America Food Animal
Practice 16, 231–253.
Duffield, T. 2006. Impact of Subclinical Ketosis on Health and Performance. Proceedings of the North
American Veterinary Conference. p. 1-5, Florida.
Duffield, T.F.; Lissemore, K.D.; McBride, B.W.; Leslie, K.E. 2009. Impact of hyperketonemia in early lactation
dairy cows on health and production. Journal of Dairy Science 92, 571–580.
Espósito, G; Irons, P.C.; Webb, E.C; Chapwanya, A. 2014. Interactions between negative energy balance,
metabolic diseases, uterine health and immune response in transition dairy cows. Animal Reproduction Science
144, 60-71.
Fürll, M.; Deniz, A.; Westphal, B. et al. 2010. Effect of multiple intravenous injections of butaphosphan and
cyanocobalamin on the metabolism of periparturient dairy cows. Journal of Dairy Science 93, 4155-4164.
Galvão Jr, J.G.B.; Rangel, A.H.N.; Medeiros, H.R. et al. 2010. Efeito da produção diária e da ordem de parto
na composição físico-química do leite de vacas de raças zebuínas. Acta Veterinaria Brasilica 4, 25-30.
Ganjkhanlou, M.; Rezayazdi, K.; Ghorbani, G. R. et al. 2009. Effects of protected fat supplements on production
of early lactation Holstein cows. Animal Feed Science and Technology 154, 276-283.
Gantner, V.; Potocnik, K.; Jovanovac, S. 2009. Test-day records as a tool for subclinical ketosis detection. Acta
Veterinaria (Beograd) 59, 85-191.
Garro, C.J.; Mian, L.; Roldán, M.C. 2014. Subclinical ketosis in dairy cows: prevalence and risk factors in
grazing production system. Journal of Animal Physiology and Animal Nutrition 98, 838–844.
Gonzalez, F.H.D. 2004. Pode o leite refletir o metabolismo da vaca? In: Congresso Brasileiro De Qualidade Do
Leite. Passo Fundo, RS, Anais-CR/ROM.
Gonzalez, F.H.D.; Silva, S.C. 2006. Introdução à bioquímica clínica veterinária. Porto Alegre: Editora
Universidade Federal do Rio Grande do Sul.
Gonzalez, F.H.D.; Correa, M.N.; Silva, S. C. 2014. Transtornos Metabólicos nos animais domésticos; 2ª ed.
Porto Alegre: Editora UFRGS.

178
Gonzalez, F.H.D.; Muiño, R.; Pereira, V. et al. 2009. Indicadores sanguíneos de lipomobilização e função
hepática no início da lactação em vacas leiteiras de alta produção. Ciência Animal Brasileira – Suplemento 1,
Anais do VIII Congresso Brasileiro de Buiatria.
Gordon, J.L.; Leblanc, S.J.; Duffield, T.F. 2013. Ketosis treatment in lactating dairy cattle. The Veterinary
Clinics of North America Food Animal Practice 29, 433-445.
Hayirli, A.; Grummer, R.R.; Nordheim, E.V.; Crump, P.M. 2002. Animal and dietary factors affecting feed
intake during the pre-fresh transition period in Holsteins. Journal of Dairy Science 85, 3430-3443.
Hayirli, A. 2006. The role of exogenous insulin in the complex of hepatic lipidosis and ketosis associated with
insulin resistance phenomenon in postpartum dairy cattle. Veterinary Research Communications 30, 749-774.
Herdt, T.H. 2000. Ruminant adaptation to negative energy balance. Influences on the etiology of ketosis and
fatty liver. The Veterinary Clinics of North America. Food Animal Practice, 16, 215-230.
Holtenius, P.; Holtenius, K. 1996. New aspects of ketone bodies in energy metabolism of dairy cows: a review.
Zentralblatt Für Veterinärmedizin 43, 579-587.
IBGE. 2014. Estatística da produção pecuária, Rio de Janeiro. 49 p. Disponível em:
<http://ibge.gov.br/home/estatistica/indicadores/ agropecuaria/producaoagropecuaria/abate-leite-
couroovos_201401_publ_completa.pdf>. Acesso em: 02 set.2015.
Ingvartsen, K.L.; Nielsen, N.I. 2004. Propylene glycol for dairy cows: A review of the metabolism of propylene
glycol and its effects on physiological parameters, feed intake, milk production and risk of ketosis. Animal Feed
Science and Technology 115, 191- 213.
Kreipe, L.; Deniz, A.; Bruckmaier, R.M.; Van Dorland, H.A. 2011. First report about the mode of action of
combined butaphosphan and cyanocobalamin on hepatic metabolism in non ketotic early lactating cows. Journal
of Dairy Science 94, 4904–4914.
Krempasky, M.; Maskalova, I.; Bujnak, L.; Vajda, V. 2014. Ketone bodies in blood of dairy cows: Prevalence
and monitoring of subclinical ketosis. Acta Veterinaria Brno 83, 411-416.
Leblanc, S. 2010. Monitoring metabolic health of dairy cattle in the transition period. Journal of Reproduction
and Development 56, 29-35.
Loor, J.J; Bionaz, M.; Drackley, J.K. 2013. Systems physiology in dairy cattle: nutritional genomics and
beyond. Annual Reviews of Animal Biosciences 1, 365-392.
McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. 2011. A field trial on the effect of propylene glycol on milk yield
and resolution of ketosis in fresh cows diagnosed with subclinical ketosis. Journal of Dairy Science 94, 6011-
6020.
McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. 2012a. Epidemiology of subclinical ketosis in early lactation dairy
cattle. Journal of Dairy Science 95, 5056–5066.
McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. 2012b. A field trial on the effect of propylene glycol on displaced
abomasum, removal from herd, and reproduction in fresh cows diagnosed with subclinical ketosis. Journal of
Dairy Science, 95, 2505–2512.
McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. et al. 2013. Elevated non-esterified fatty acids and β-
hydroxybutyrate and their association with transition dairy cow performance. The Veterinary Journal, 198, 560–
570.
Melendez, P.; Goff, J.P.; Risco, C.A. et al. 2006. Incidence of subclinical ketosis in cows supplemented with a
monensin controlled-release capsule in Holstein cattle. Preventive Veterinary Medicine 73, 33–42.
Nielen, M.; Aarts, M.G.; Jonkers, A.G.; Scwukken, Y.H. 1994. Evaluation of two cowside tests for the detection
of subclinical ketosis in dairy cows. Canadian Veterinary Journal 35, 229-232.
Nielsen, N.I.; Ingvartsen, K.L. 2004. Propylene glycol for dairy cows. A review of the metabolism of propylene
glycol and its effects on physiological parameters, feed intake, milk production and risk of ketosis. Animal Feed
Science and Technology 115, 191-213.

179
Nuber, U.; Van Dorland, H.A.; Bruckmaier, R.M. 2015. Effects of butafosfan with or without cyanocobalamin
on the metabolism of early lactating cows with subclinical ketosis. Journal of Animal Physiology and Animal
Nutrition 99, 1-10.
Oetzel, G.R. 2004. Monitoring and testing dairy herds for metabolic disease. Veterinary Clinics of North
America: Food Animal Practice 20, 651-674.
Panousis, N.; Brozos, C.H.; Karagiannis, I. et al. 2012. Evaluation of Precision Xceed meter for on-site
monitoring of blood β-hydroxybutyric acid and glucose concentrations in dairy sheep. Research in Veterinary
Science 93, 435-439.
Pereira, R.A.; Silveira, P.A.S.; Montagner, P. et al. 2013. Effect of butaphosphan and cyanocobalamin on
postpartum metabolism and milk production in dairy cows. Animal 7, 1143-1147.
Piepenbrink, M.S.; Overton, T.R. 2003. Liver metabolism and production of cows fed increasing amounts of
rumen-protected choline during the periparturient period. Journal of Dairy Science 86, 1722-1733.
Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W. et al. 2007. Veterinary Medicine: A textbook of the diseases of
cattle, horses, sheep, pigs and goats.10th edition., Philadelphia: Elsevier, 2156 p.
Rajala-Schultz, P.J.; Gro, Y.T.; McCulloch, C.E. 1999. Effects of milk fever, ketosis, and lameness on milk
yield in dairy cows. Journal of Dairy Science 82, 288-294.
Rangel, A.H.N.; Medeiros, H.R.; Silva, J.B. et al. 2009. Correlação entre a contagem de células somáticas
(CCS) e o teor de gordura, proteína, lactose e extrato seco desengordurado do leite. Revista Verde de
Agroecologia e Desenvolvimento Sustentável 4, 57- 60.
Reist, M.; Erdin, D.K.; Voneuw, D. et al. 2002. Estimation of energy balance at the individual and herd level
using blood and milk traits in high-yielding dairy cows. Journal of Dairy Science 85, 3314- 3327.
Risco, C.A. 1992. Calving related disorders. In: Van Horn, H.H.; Wilcox, C.J. Large dairy herd management.
Savoy, American Dairy Science Association, p. 193-198.
Robinson, W.F.; Huxtable, C.R.R. 2003. Clinicopathologic Principles for Veterinary Medicine, Cambridge
University Press.
Rollin, R.; Berghaus, R.D.; Rapnicki, P. et al. 2010. The effect of injectable butaphosphan and cyanocobalamin
on postpartum serum beta-hydroxybutyrate, calcium, and phosphorus concentrations in dairy cattle. Journal of
Dairy Science 93, 978-987.
Sato, H.; Shiogama, Y. 2010. Acetone and isopropanol in ruminal fluid and feces of lactating dairy cows.
Journal of Veterinary Medical Science 72, 297-300.
Schein, I.H. 2012. Cetose dos ruminantes. Disciplina de transtornos metabólicos dos animais domésticos.
Universidade Federal do Rio Grande do Sul, UFRGS, Porto Alegre, RS. 35p.
Simensen, E.; Halse, K.; Gillund, P.; Lutnaes, B. 1990. Ketosis treatment and milk yield in dairy cows related
to milk acetoacetate levels. Acta Veterinaria Scandinavica 31, 433–440.
Shin, E.K.; Jeong, J.K.; Choi, I.S. et al. 2015. Relationships among ketosis, serum metabolites, body condition,
and reproductive outcomes in dairy cows. Theriogenology 84, 252–260.
Smith, B.P. 2008. Large Animal Internal Medicine - Text and Veterinary Consult Package, 4ª ed. Editora
Mosby, EUA. 1872p.
Voyvoda, H.; Erdogan, H. 2010. Use of a hand-held meter for detecting subclinical ketosis in dairy cows.
Research in Veterinary Science 89, 344-351.
Wathes, D.C., Bourne, N., Cheng, Z. et al. 2007. Multiple correlation analyses of metabolic and endocrine
profiles with fertility in primiparous and multiparous cows. Journal of Dairy Science 90, 1310–1325.
Walsh, R.B., Walton, J.S.; Kelton, D. F. et al. 2007. The Effect of Subclinical Ketosis in Early Lactation on
Reproductive Performance of Postpartum Dairy Cows. Journal of Dairy Science 90, 2788–2796.
Wood, G.M., Boettcher, P.J.; Kelton, D.F.; Jansen, G.B. 2004. Phenotypic and genetic influences on test-day
measures of acetone concentration in milk. Journal of Dairy Science 87, 1108-1114.

180
Weng, X.; Chen, L.; Neethirajan, S.; Duffield, T. 2015. Development of quantum dots-based biosensor towards
on-farm detection of subclinical ketosis. Biosensors and Bioelectronics 72, 140-147.
Yuan, K.; Shaver, R.D.; Bertics, S.J. et al. 2012. Effect of rumen-protected niacin on lipid metabolism, oxidative
stress, and performance of transition dairy cows. Journal of Dairy Science 95, 2673–2679.
Zom, R.L.; Baal, J.; Goselink, R.M. et al. 2011. Effect of rumen-protected choline on performance, blood
metabolites, and hepatic triacylglycerols of periparturient dairy cattle. Journal of Dairy Science 94, 4016–4027.

181
Hipocalcemia e hipomagnesemia en vacas lecheras: diagnóstico y control
Luis Barros Vidal
(In memoriam)
La hipocalcemia y la hipomagnesemia son enfermedades tradicionales de las vacas lecheras,
ya consideradas metabólicas desde el siglo XIX. La hipocalcemia es una enfermedad
metabólica en el sentido que existe un mecanismo de regulación para su homeostasis, y
aunque no sucede lo mismo con la hipomagnesemia continúa siendo clasificada como tal. La
intensificación de la producción lechera en los últimos decenios y en particular la selección
genética para el incremento de la producción láctea por vaca individual, provoca un efecto
negativo sobre la estabilidad metabólica de los animales. Se comprueba en diferentes países
una prevalencia relativamente elevada de casos clínicos por hipocalcemia y particularmente
alta por hipocalcemia subclínica. La prevalencia de la hipomagnesemia es mayormente
dependiente del aporte proveniente de las pasturas mejoradas y de su control. La
intensificación de la producción láctea con altos rendimientos viene acompañada por
elevados costos para el mantenimiento de la homeostasis de esos animales, que son sometidos
a unas enormes y estresantes demandas metabólicas durante el período de lactancia. Esas
demandas se acrecientan con la edad del animal considerando que la producción por lactancia
se incrementa en cada año sucesivo, mientras que la habilidad de respuesta de las vacas tiende
a disminuir de la misma manera que sus reservas corporales, particularmente si la dieta no es
lo suficientemente balanceada para cubrir las necesidades en los diferentes momentos del
ciclo reproductivo-productivo. La capacidad de movilización mineral también disminuye con
la edad: un animal joven puede intercambiar un 17 a 20% de su calcio (Ca) óseo, mientras
que una vaca mayor a 10 años lo hará en un 2 a 5%.
De manera general, los períodos de producción deben contemplar las pérdidas minerales por
producción de leche, así como los períodos de recuperación metabólica, principalmente en el
período seco, con la finalidad de prevenir el desbalance del inicio de lactación, período donde
ocurre la mayor frecuencia de alteraciones metabólicas de hipocalcemia clínica y de otras
enfermedades asociadas a la hipocalcemia subclínica. La hipomagnesemia puede acompañar
este período, pero su prevalencia está mayormente vinculada con el pico de lactación y con
los ciclos de las pasturas.

182
En el desbalance mineral intervienen varios eventos metabólicos concurrentes, tales como la
disminución del apetito en los períodos de mayores demandas; la distensión ruminal; la
frecuencia y volumen de la materia seca consumida determinando la saciedad y el
comportamiento alimentario; la influencia del propio estrés del período parto-inicio de
lactancia; los factores ambientales de confort en el establecimiento; el comportamiento social
por dominancia, agrupamiento y atestamiento de los animales; las enfermedades
inflamatorias del periparto asociadas a la disminución de las defensas del aparato inmunitario
provocado por el estrés, además del estricto balance nutricional aportado por la dieta.
Durante el período de transición, léase de las tres o cuatro semanas anteriores y posteriores
al parto, el momento del periparto es aquel de mayores riesgos en la salud de las vacas
lecheras y es donde ocurre la mayor frecuencia de enfermedades del año productivo. La
adaptación metabólica de la vaca al crecimiento fetal al final de la gestación; a las
necesidades obligatorias de la glándula mamaria para la calostrogénesis; al inicio y
mantenimiento de la lactancia; a las exigencias hormonales de recuperación uterina; a las
demandas energéticas y proteicas para la producción de leche; al desbalance entre
necesidades alimentarias y la capacidad volumétrica del rumen; a la adaptación hormonal al
desbalance energético obligatorio del inicio de lactancia u homeorresis; son algunos de los
factores que tienen más relevancia por sus efectos en el metabolismo general y en la
estabilidad del sistema inmunitario. El estrés metabólico y los factores ambientales y sociales
de los animales están relacionados con una significativa incidencia de enfermedades
inflamatorias e infecciosas que afectan principalmente al aparato reproductivo, tales como:
el retraso en la involución uterina, las metritis o endometritis, las retenciones de placenta o
las mastitis por coliformes. El incremento de los procesos inflamatorios está relacionado con
la disminución de la respuesta inmune por cambios en los patrones de respuesta de las
proteínas de fase aguda, que provocan una inefectiva acción de los leucocitos,
particularmente los neutrófilos y los monocitos sobre los agentes patógenos. Los procesos
inflamatorios inducidos por las proteínas de fase aguda: citoquinas (tumor necrosis factor a
(TNF-a), interleucina 1 (IL-1), e interleucina 6 (IL-6) y eicosanoides son acompañados de
procesos inflamatorios agudos con hipertermia, taquicardia y anorexia o disorexia (Sordillo,
2013).

183
Las exigencias energéticas de esos períodos se manifiestan por una elevada lipomovilización
y una cetosis subclínica que inducen a modificaciones histopatológicas del parénquima
hepático con esteatosis, la que puede presentarse de discreta a severa, provocando una
consecuente disfuncionalidad del sistema digestivo y hormonal que agrega un efecto
negativo a la funcionalidad metabólica que ya se encuentra comprometida por los factores
mencionados anteriormente. Así, la condición corporal sufre las consecuencias producto del
catabolismo energético y proteico con disminución del estado corporal por pérdida de la masa
muscular y de la reserva grasa. Las consecuencias hormonales en el metabolismo de la
glucosa están relacionadas con la secreción de la insulina por el páncreas y su sensibilidad
en el músculo esquelético, en el tejido adiposo y en el hígado afectando la salud metabólica
del animal. Esas modificaciones se revelan como variaciones de la concentración de los
ácidos grasos libres y del beta-hidroxibutirato (BHB) induciendo una respuesta de insulino-
resistencia en esos órganos durante la preñez y la lactancia. A su vez, también existe una
relación entre el metabolismo energético y el metabolismo mineral (Lean, 2006). Los tenores
en minerales: calcio, magnesio, fósforo, la diferencia iónica y los niveles de potasio tienen
influencia en el balance del metabolismo mineral y en la fisiopatología del síndrome de vaca
caída y particularmente en el cuadro clínico y subclínico de la hipocalcemia y de la
hipomagnesemia de la vaca lechera. La importancia de estas enfermedades metabólicas está
en el efecto directo sobre la vida productiva de las vacas, pero aún más en su salud por las
alteraciones y afecciones asociadas que ocurren en el periparto y el puerperio como las
mastitis, las distocias, las retenciones de placenta, las infecciones uterinas, y los
desplazamientos de abomaso, entre otras.
Hipocalcemia en vacas lecheras
Antecedentes
La demanda por aumentar la producción de leche para consumo humano trajo aparejado la
selección genética y la mejora en la alimentación de la vaca lechera, resultando en un
significativo incremento de la secreción láctea por animal y por lactancia. Ese incremento de
la producción fue acompañado por una mayor exigencia metabólica, cuyo natural equilibrio
en la evolución de la especie no ha sido compensado por la ganancia genética, inducida por

184
el hombre en la última centuria. Es así que una enfermedad como la hipocalcemia, con una
prevalencia muy poco común, comienza a incrementarse. Hutrya et al. (1947) en su libro
sobre “Patologías especiales y terapéuticas de los animales domésticos” indican que un
primer caso de hipocalcemia se registró en 1793, coincidiendo con la introducción en Europa
de la selección genética orientada a buscar animales de alta producción bajo un limitado nivel
de intensificación agrícola. Diversas hipótesis sobre la etiología surgieron para esa
enfermedad que se presentaba en las horas próximas al parto, comprendiendo la apoplejía, la
trombosis, la infección del útero o de las mamas. Señala Jack Payne (1981) que un aporte
importante al conocimiento fue la teoría del danés Jurgens Schmidt que en 1897 sugirió que
la enfermedad era debida a una infección de la ubre y que la infusión intramamaria de ioduro
de potasio constituía un buen tratamiento, terapia que generó buenos resultados con una
reducción de la mortalidad desde un 15% a un 70%. Luego se observó que la infusión de
agua intramamaria era igualmente efectiva y que el insuflado de la ubre era incluso más
efectivo (Greig, 1930 in Payne, 1981). El segundo gran avance en el diagnóstico llega en
1925 cuando Dryerre y Greig enunciaron una teoría asociando la enfermedad con la
disminución del Ca en sangre, argumentando que tenía su génesis en el pasaje del mineral a
la mama en animales con deficiencia de hormonas paratiroideas. La confirmación de la teoría
que la hipocalcemia era la etiología de la paresia puerperal, fue dada por los mismos Dryerre
y Greig en 1930 de Edinburgo, empleando gluconato de Ca por vía parenteral para remitir
los síntomas clínicos. Es interesante notar que desde 1934, cuando Hayden sintetizó el
borogluconato de calcio para el tratamiento endovenoso, hoy en día continúa siendo el
tratamiento empleado y efectivo. Los mayores avances científicos y técnicos en la resolución
del problema clínico ha sido la prevención, con cambios en la alimentación y en el manejo
de las vacas pre-parturientas. Se han tomado en cuenta los cambios metabólicos del período
de transición y del posparto para controlar el metabolismo mineral que hace eclosión en el
periparto. Desde el punto de vista clínico, la hipocalcemia es un caso individual, pero las
medidas preventivas se toman para el manejo colectivo de la enfermedad, considerando su
importancia económica a nivel del establecimiento lechero y que más del 80% de esas vacas
se encuentra en hipocalcemia subclínica. Históricamente, la enfermedad clínica se describió
con diferentes sinonimias: hipocalcemia puerperal, paresia puerperal, fiebre vitular,

185
eclampsia posparto, fiebre de la leche, paresia posparto, paresia de la parturienta e
hipocalcemia de la vaca lechera.
Descripción
La hipocalcemia es una enfermedad metabólica que se presenta principalmente en la vaca
lechera durante el período periparto, manifestándose por una sintomatología nerviosa, de tipo
depresivo, con paresia y cuyo curso agudo evoluciona a la muerte si el animal no es tratado.
La etiopatogenia está determinada por una alteración del balance diario de Ca y por la
incapacidad de los mecanismos de regulación para mantener la homeostasis frente a pérdidas
enormes y súbitas, inducidas por el pasaje masivo del Ca plasmático hacia la glándula
mamaria en el inicio de lactancia. Las pérdidas de Ca pueden ser de 25 g o más para una
producción de al menos 10 L de calostro. La leche contiene 1,2 g de Ca por L, lo que permite
calcular una pérdida de 30 g para una vaca buena productora con necesidades para
mantenimiento sólo de 12 g por día.
La estabilidad de la calcemia es vital para la normal homeostasis de la mayoría de los órganos
y funciones del organismo. El pool de Ca es calculado en 14 g/kg PV en el bovino, estando
distribuido en un 99% en el esqueleto y el resto en otros órganos. Se destaca el espacio
extracelular dividido en extravascular e intersticial con 5 a 6 g, donde ocurren funciones de
intercambio iónico, siendo el mineral disponible para cubrir exigencias metabólicas
inmediatas. El Ca es transportado por el espacio intravascular como calcemia en sus dos
fases: el Ca no difusible, ligado y transportado por proteínas y el libre o iónico (40 a 50% del
Ca sanguíneo) necesario para las funciones celulares como: la permeabilidad de membrana,
la coagulación sanguínea, la excitabilidad neuromuscular, la contracción muscular y también
la acción como cofactor enzimático u hormonal, dependiendo su disponibilidad del pH
sanguíneo. El balance del mineral debe estar equilibrado entre la absorción intestinal
(alrededor del 68%), la eliminación endógena por recambio óseo y las necesidades para
gestación y para lactación con el Ca aportado por la dieta.
Los lentos mecanismos homeostáticos de la acción de la parathormona (PTH), la vitamina D3
(1,25 di-hidroxicolecalciferol) y la calcitonina, no permiten la adaptación del intestino, el
hueso y el riñón a las obligatorias y fuertes demandas de la calostrogénesis y la lactancia.
Particularmente, las dietas alcalinas en base a pasturas son un factor coadyuvante de la mayor

186
importancia, ya que afectan el equilibrio ácido-básico sanguíneo. El desequilibrio hacia la
dieta alcalina induce cambios en la receptividad de los receptores a la PTH bloqueando la
acción hormonal y, por lo tanto: disminuye la movilización del Ca proveniente de la resorción
osteoclástica de la hidroxiapatita de los huesos; disminuye la síntesis del 1,25 di-
hidroxicolecalciferol por el riñón y el hígado, resultando con una disminución en la respuesta
para mantener la homeostasis de la calcemia. Un factor muy destacado en la literatura
científica, como importante en el desequilibrio del mineral, es el alto contenido de potasio en
los forrajes provenientes de una intensa fertilización del suelo para mejorar los rendimientos
en materia seca. Este elemento fue reconocido en la patogenia de la hipomagnesemia por
Kemp y 't Hart en 1957, y ahora se le reconoce ampliamente en su papel de factor de riesgo
para la hipocalcemia al relacionarlo con el balance iónico de la dieta y en particular con la
modificación del equilibrio ácido-base sanguíneo. El potasio propende a la alcalinización
sanguínea que actúa reduciendo la movilización del Ca óseo y la disminución del Ca iónico
en sangre, factor principal para desencadenar el cuadro clínico.
El desbalance en la homeostasis cálcica desemboca con la presentación de la enfermedad,
cuya sintomatología se caracteriza por un cuadro neurológico que cursa con tres etapas: una
primera, fase breve de excitación; una segunda de postración con decúbito esternal y una final
con decúbito lateral, previo a la muerte. El caso clínico se presenta como una hipocalcemia
aguda posparto entre las 24 horas preparto y las 72 horas después. Desde el punto de vista
bioquímico se caracteriza por un descenso brusco del Ca iónico sérico y desde el punto de
vista clínico por un cuadro nervioso depresivo con persistencia del decúbito sin posibilidades
de recuperar la estación. Esta patología se manifiesta en vacas lecheras de mediana y elevada
producción, con mayor incidencia en animales que están entre su tercera y cuarta lactación,
siendo las vacas multíparas más susceptibles y las primíparas prácticamente no se enferman.
La susceptibilidad individual es un importante factor. Las vacas que han tenido hipocalcemia
clínica en un año y se recuperaron, tienen altas probabilidades que en el siguiente parto se
manifieste nuevamente la enfermedad. El cuadro clínico puede presentarse en todas las
épocas del año en que se produzcan los partos. Las razas tienen influencia en la prevalencia,
siendo las Jersey más susceptibles que las Holstein y éstas más que las razas de carne donde
la fiebre de la leche ocurre en forma muy poco frecuente. El diagnóstico se establece en base
a la sintomatología, en particular por el cuadro nervioso depresivo en el periparto, a los

187
exámenes colaterales y a la respuesta terapéutica al Ca. En la patología clínica se comprueban
valores de calcemia muy por debajo de los normales (referencia: 9 a 11 mg/dL), entre 3,0 y
4,8 mg/dL (0,75-1,2 mmol/L), que dan nombre a la afección. La anatomía patológica no
revela signos de relevancia para el diagnóstico.
Diagnóstico clínico
Clásicamente se definen tres fases o etapas de la hipocalcemia, una inicial con sintomatología
leve, otra denominada de decúbito esternal con auto-auscultación y otra final con decúbito
lateral con coma y muerte. La primera fase puede ser detectada por el tambero, ya que es
práctica de manejo común que las vacas en el período preparto final sean aproximadas hacia
el lugar donde serán ordeñadas para acostumbrarlas a la futura rutina del ordeño; en otras
ocasiones son alojadas en lugares o galpones cercanos. En la etapa inicial, en los dos o tres
días antes del parto, puede observarse a la inspección algunos espasmos musculares en la
región de los hombros, pero preferentemente la masa muscular del tren posterior es la que
presenta contracciones de tipo fasciculares en grupos de fibras musculares que son bien
visibles y que se evidencian claramente por la palpación colocando la mano sobre la piel de
la masa muscular. Estas contracciones pueden presentarse como mioclonias, particularmente
en los músculos estriados semimembranoso y semitendinoso. Se evidencian como temblores
en la estación y por una ligera dismetría en la marcha, pudiendo presentarse cierto grado de
rigidez de los miembros posteriores en movimiento, como consecuencia de la alteración de
la transmisión del impulso neuronal por un aumento de la irritabilidad neuromuscular. Puede
haber una ligera hiperexcitabilidad e hiperreflexia, con leves temblores en cabeza y dorso
pudiendo sufrir el animal grados leves de parestesia. La vaca procura no moverse, y en esta
etapa de corta duración puede caer con facilidad al caminar sobre un piso duro resbaladizo o
mojado, pero no es frecuente que el animal se mantenga en decúbito. La lengua puede
presentar cierto grado de protrusión y haber rechinamiento de dientes. En la musculatura lisa
se manifiestan síntomas digestivos de constipación con heces secas y duras, el apetito es
normal o puede comenzar una disorexia por la disminución en las contracciones ruminales
que más cerca del parto se profundizan. Puede presentarse cierto grado de distensión del
rumen con ligera deformación del perfil abdominal, principalmente del lado izquierdo. La
frecuencia de la rumia puede también estar disminuida. En la glándula mamaria hay

188
relajación del esfínter del pezón que permite la salida de calostro en goteo, a veces en forma
continua, mostrando una glándula muy distendida. El sensorio es normal y el animal responde
a los estímulos visuales y externos demostrando una expresión alerta. La temperatura en este
período es normal, así como la frecuencia respiratoria y el pulso. Por paraclínica se puede
constatar una discreta hipocalcemia con valores que pueden oscilar entre 7,5 y 8,5 mg/dL
(1,87 a 2,12 mmol/L). Hay que hacer notar que existe una hipocalcemia fisiológica del parto,
pero ésta no tiene manifestaciones clínicas, como en este caso.
En la segunda fase, los síntomas son claramente visibles, el cuadro predominante es un
cuadro de sensorio deprimido, con posición de decúbito esternal o ventral. El animal tiende
a no desplazarse y quedar en decúbito esternal con una posición de la columna vertebral
ligeramente encorvada en forma de S, apoyándose principalmente sobre las articulaciones
radio-cubitales de las patas delanteras, colocando la cabeza hacia adelante y abajo con
aspecto somnoliento, con los párpados semicerrados y dando respuesta a estímulos visuales
en forma lenta o ausente. Evoluciona hacia una paresia del tren posterior con tendencia a la
flaccidez, síntomas que se conocen como paresia puerperal dentro del síndrome de vaca
caída. En esta etapa de parálisis fláccida muscular, particularmente de los miembros
posteriores, pueden presentarse algunas contracciones musculares pasajeras de tipo
tetaniforme. La cabeza se vuelca hacia el flanco, en la llamada “posición de auto-
auscultación”, con el cuello en extensión hacia atrás y debajo como manifestación de la
disminución del sensorio, mostrando una ausencia o disminución muy marcada de respuesta
a los estímulos externos. Las acciones de tracción sobre la cabeza y el cuerpo, las vivas voces
o ligeros golpes pueden obligar al animal a incorporarse, aunque lo hará con dificultad o no
podrá hacerlo. La permanencia en la estación tiende a ser breve, aunque a veces permanece
de pie a pesar de la debilidad de los miembros posteriores que pueden presentar temblores
por el intento de mantener el aplomo y el equilibrio posterior, estirando su cabeza y el cuello
para equilibrar el peso corporal. Los reflejos de sensibilidad superficial en las extremidades
están disminuidos, particularmente en miembros posteriores, que responden lentamente al
pellizcamiento o punción de la piel o en el espacio interdigital. Igualmente ocurre con los
reflejos de sensibilidad profunda en tendones. Las pupilas están dilatadas y es lento el reflejo
pupilar a la luz; las córneas están secas y puede aparecer cierto grado de enoftalmia. Puede
percibirse el reflejo anal disminuido, con semidilatación del esfínter anal y la aparición de

189
materia fecal en escasa cantidad de color oscuro y seco al lado del animal. El esfínter del
pezón también está semidilatado permitiendo la salida de poca cantidad de calostro por efecto
del rebozamiento del canal del pezón. La temperatura corporal disminuye, con una
hipotermia discreta (36 a 38°C), pudiendo percibirse por palpación las orejas frías en su base,
así como las extremidades y la piel en general. Las mucosas están ligeramente pálidas y el
morro seco. La frecuencia del pulso aumenta por encima de 70 a 80 por minuto con
pulsaciones débiles al tacto. La frecuencia cardíaca aumenta y pueden auscultarse
alteraciones del ritmo, con arritmias de tipo bloqueo aurículo-ventricular como consecuencia
de la alteración en la conducción del impulso eléctrico a través del miocardio, con
alargamiento del intervalo Q-T del proceso de conducción y repolarización en la carga
eléctrica del corazón después de cada latido, que puede evidenciarse mediante un ECG. Este
cambio eléctrico está asociado al efecto de la hipocalcemia y particularmente al bajo Ca
ionizado. En las grandes funciones se comprueba que hay anorexia, la glándula mamaria está
plena y muy distendida ya que la hipocalcemia se presenta principalmente en vacas muy
buenas productoras y con más de tres o cuatro partos, en el período de mayor producción de
las lecheras. La bioquímica clínica indica valores subnormales de la calcemia (< 7,5 mg/dL
o < 1,87 mmol/L), así como del valor del Ca ionizado (Ca2+ por debajo del 40%). Puede
haber hipoalbuminemia (< 30 g/L) que contribuye con el déficit de transporte del Ca ligado
y los valores de fósforo y magnesio pueden estar disminuidos para el primero (< 4,5 mg/dL)
y aumentados para el segundo (> 2,7 mg/dL).
A partir de esta fase, se pasa gradualmente a la tercera, caracterizada por la depresión
profunda del sensorio, el decúbito lateral y la muerte en coma en el 80 % de los casos sin
tratamiento. En esta etapa el animal permanece en decúbito, primero ventral y luego se coloca
de lado con el cuello y la cabeza extendidos. No realiza movimientos y la flaccidez muscular
es particularmente evidente. Es interesante mencionar que la parálisis en los bovinos es
fláccida, probablemente porque la baja concentración extracelular de Ca reduce la liberación
de acetilcolina en la placa neuromuscular, disminuyendo el trofismo o fuerza de contracción,
cuando en otras especies (cerdo, perro, caballo) es mayormente espástica o tetaniforme, en
la llamada eclampsia. En el humano la espasticidad es similar y aparecen el signo de
Trousseau -espasmo visible y doloroso del carpo y mano al comprimirse el brazo-, y el de
Chvöstek -espasmo facial y de la comisura labial al percutir por delante de la oreja en la rama

190
del nervio facial-. En la vaca hay flaccidez de la musculatura estriada e hiporreflexia
generalizada con pérdida del reflejo pupilar, del palpebral, con protrusión del tercer párpado,
abolición del reflejo anal y del patelar, con ausencia de respuesta a las pruebas de sensibilidad
superficial y profunda. La depresión del sensorio es severa con pérdida de la conciencia y
con ausencia de respuesta a los estímulos externos manifestando gran somnolencia, sopor y
coma. El decúbito lateral induce a un meteorismo gaseoso por falta de eructación y debido a
la posición del cuerpo, que se recupera en parte si es posible colocar la vaca en posición
ventral, la administración de Ca endovenoso induce inmediatamente al inicio de la
eructación, abolida en esta etapa. Las contracciones ruminales están ausentes, puede haber
defecación con heces secas y duras por relajación del esfínter anal. No orina y si lo hace es
por rebosamiento, la orina es escasa y de color oscuro. La temperatura es francamente
hipotérmica con valores cercanos a 35-36°C, las extremidades, orejas y la piel están frías. La
respiración es superficial pudiendo auscultarse u oírse cerca de las narinas un ronquido
producido por parálisis de velo del paladar.
En la fase terminal hay taquicardia con una frecuencia cardíaca superior a 100-120
pulsaciones por minuto, puede auscultarse arritmia, el pulso es débil y filiforme pudiendo no
percibirse a la palpación, puede haber ingurgitación de las venas yugulares, las mucosas están
congestivas y el cuadro termina en colapso circulatorio. El análisis de laboratorio indica
valores de calcemia que pueden bordear los 5-6 mg/dL (< 1.25 a 1,5 mmol/L) y el valor del
Ca2+ por debajo del 30%, de un valor normal entre 42 y 47% de la calcemia. La fosfatemia
puede alcanzar valores disminuidos de 3 a 4 mg/dL y la magnesemia aumentada puede estar
con valores superiores a 3.2 mg/dL. La muerte del animal sobreviene en pocas horas sin
tratamiento, con una mortalidad superior al 80% en vacas multíparas de alta producción. La
recuperación en fases tempranas luego de la perfusión de Ca es superior al 90%, debiendo
preverse posibles recaídas en las 12 a 18 horas posteriores. En la etapa terminal, la
recuperación clínica puede ser del 50-60%.
Hipocalcemia subclínica
Cuando se estudia la hipocalcemia de la vaca lechera, debe ser tenido en cuenta un capítulo
especial, que es la hipocalcemia clínica, porque la casuística indica que tiene una incidencia
mayor cada vez que años precedentes. Esto es debido a la mayor tecnificación agrícola con

191
incremento de la producción de leche por animal y por el mayor control de otras
enfermedades intercurrentes. El empleo de medidas de vigilancia más eficaces a nivel
colectivo ha permitido controlar la prevalencia de la enfermedad clínica, pero aún la baja
frecuencia de casos individuales continúa siendo de importancia, particularmente por el valor
económico de estas vacas de alta producción y por la merma en la producción que provoca
la muerte de esos animales.
A pesar que los casos clínicos de vacas individuales están más fiscalizados, la hipocalcemia
continúa siendo un problema sanitario y productivo como consecuencia de la prevalencia de
la hipocalcemia subclínica que continúa aumentando y que a la vista del productor no siempre
es bien percibida. La importancia radica no solamente en los animales directamente
afectados, sino por las alteraciones concomitantes por la disfunción de los diferentes sistemas
del organismo dependientes de la homeostasis del Ca. Como producto de las alteraciones en
la musculatura lisa aumenta la susceptibilidad a otras patologías, como el menor tono del
esfínter del pezón, la mastitis a coliformes -con una tasa de riesgo 8 a 9 veces mayor en vacas
que sufrieron hipocalcemia previa-, la menor contractilidad uterina, las distocias, la retención
de placenta, el retraso de la involución uterina, las metritis o endometritis -con retraso en el
reinicio de la actividad reproductiva-, los desplazamientos de abomaso, la disminución de la
funcionalidad ruminal, la reducción del consumo voluntario, la cetosis por balance energético
negativo, la disminución de la producción láctea, la menor tasa de remodelación ósea, las
enfermedades podales y la disminución de la respuesta inmune entre otros trastornos
conocidos que tienen un alto impacto sobre la economía del establecimiento lechero.
La importancia de la hipocalcemia subclínica es que el período no se limita al periparto, sino
que abarca el preparto temprano y puede ir bastante más lejos en el tiempo que del pico de
lactancia cuando comienza a equilibrarse la pérdida mineral por la leche con el aporte por la
dieta. Los costos agregados en un establecimiento con alta prevalencia de hipocalcemia
subclínica son mucho mayores que los provocados por un bajo porcentaje de vacas con curso
clínico agudo.
Formas combinadas de hipocalcemia
La hipocalcemia clásica y más frecuente es la descrita con valores en sangre bajos en
calcemia, bajos en fosfatemia y altos en magnesemia, pero pueden existir otras

192
combinaciones patológicas. Una de ellas es la hipocalcemia con hipomagnesemia que tiene
un cuadro clínico similar en cuanto al cuadro de vaca caída pero la sintomatología nerviosa
tiene manifestaciones musculares con mayor espasticidad, con contracciones tónico-clónicas,
pudiendo haber hipertermia y muerte súbita. El tratamiento clásico no es tan efectivo a menos
que se agregue magnesio en la fórmula endovenosa. Otra manifestación de vaca caída en el
periparto es por la hipofosfatemia que presenta un cuadro también de posición de decúbito
con parálisis en miembros posteriores, permaneciendo en estado de alerta y que no responde
al tratamiento con Ca, el aporte de fósforo endovenoso es medianamente eficaz. Estos dos
últimos casos pueden cursar con hipercalemia.
Tratamiento
El tratamiento de la hipocalcemia clínica aguda con la infusión endovenosa de borogluconato
de Ca ya es un clásico, como fue mencionado anteriormente practicado desde Dryerre y Greig
en 1930 (in Payne, 1981). La opción para el tratamiento clínico de la fiebre de la leche debe
incluir inevitablemente el tratamiento IV con soluciones de Ca y considerar la propuesta más
actual de incluir en el momento del periparto la administración de un gel oral de Ca para
prevenir las recaídas del cuadro clínico que puede rondar el 25 al 40% luego de la
administración de borogluconato de Ca (Queen, 1993). Algunos veterinarios administran
simultáneamente a la infusión endovenosa alguna dosis subcutánea de soluciones cálcicas,
pero esta práctica tiene los inconvenientes de que la absorción no puede asegurarse, que la
dosis no es lo precisa que puede pretenderse, que depende de una buena perfusión y que
puede ocasionar reacciones subcutáneas adversas. Las dosis de borogluconato de Ca IV en
solución al 23% aportan 8 a 14 g de Ca en 500 mL, con recomendación de dosis única y
suministrada en forma lenta evitando el riesgo de paro cardíaco durante la infusión. La dosis
recomendada es hasta 20 g por vez. Cada dosis suple el equivalente de 4 g en la sangre.
Algunas vacas hipocalcémicas en recuperación requieren dos o más dosis intravenosas, ya
que la deficiencia se estima en 10 a 20 g de Ca. Un signo del efecto favorable durante o al
final de la perfusión es la respuesta de eructación indicando el reinicio de las contracciones
ruminales.
Las vacas caídas en la fase 2 generalmente tienen una recuperación inmediata luego de la
infusión de Ca, pero en la fase 3 puede ser necesario una segunda dosis en un volumen de

193
200 mL y algunas ya en coma no responden a tiempo. En casos extremos de debilidad con
alteración cardíaca por arritmia puede administrarse sulfato de atropina al 1% vía
endovenosa.
Los geles, en presentaciones como cloruro de Ca, o como propionato, se administran vía oral
en geles de dosis única, generalmente en 300-400 mL. La absorción es rápida, el AUC
máximo es a los 30 minutos y el efecto puede durar unas 6 horas. La recomendación es
proporcionar un gel de Ca (un frasco) vía oral en el día del parto, que cumple una función
preventiva y curativa. Se puede repetir la administración una segunda dosis a las 12 o 24
horas posparto (Oetzel, 2013). La administración de sales de Ca de buena biodisponibilidad
como carbonato de Ca en la ración puede ser un coadyuvante, aunque no se manifiesta en la
calcemia. Los animales en ese período se encuentran con disorexia o anorexia, y además
estas sales tienen baja palatabilidad, por lo que el consumo no será muy elevado. Su
administración es más conveniente para los períodos de recuperación mineral durante la
lactancia. Hubo en el pasado diferentes combinaciones para compensar las variaciones de la
calcemia, tales como el uso de vitamina D3 o de otros metabolitos como 1-calciferol, o el
empleo de la hormona paratiroidea (PTH), pero su eficacia, su disponibilidad en el mercado
o el precio no resultan de conveniencia al momento actual.
El animal caído y en alerta debe colocarse en un ambiente limpio, con sombra, con agua y
comida, previniendo que, si permanece mucho tiempo en decúbito esternal, hay que ayudarlo
a moverse de lugar varias veces al día, al menos 4 a 6 veces, porque el decúbito persistente
entumece y provoca lesiones severas en sus músculos, por lo que ponerse de pie es una
dificultad que a veces la vaca no puede superar por la debilidad del tren trasero. Ayudarlo a
ponerse de pie en forma mecánica con un aparato levanta-vacas y una cincha ancha con la
que pueda ser colgada desde arriba, inclusive con un engranaje palanca que pueda tener un
anclaje posterior y otro anterior (lo que parecería un exceso), ayuda al animal a levantarse y
estar de pie un tiempo entre 30 y 60 minutos cada vez, que puede ser la diferencia entre
recuperarse o ser sacrificado. Es recomendable masajear los miembros posteriores. La
complicación es mayor si la hipocalcemia ocurre en el preparto inmediato lo que es muy
frecuente.

194
Prevención
La prevención de las enfermedades del período de transición de la vaca lechera incluye en
particular la hipocalcemia debido a la importancia económica y productiva en el
establecimiento, así como en el mantenimiento de la salud y de la sobrevida de los animales
sometidos a un continuo estrés para cumplir con las demandas productivas. El enfoque de la
prevención del período de transición es importante para disminuir la incidencia de casos
clínicos y de los efectos secundarios de la hipocalcemia subclínica, pero debe tenerse
presente que la recuperación mineral y metabólica de las vacas lecheras incluye el período
de lactancia tardío donde las demandas para producción disminuyen y donde el balance
energético y mineral es positivo y el animal recupera su peso vivo, su masa ósea y su
equilibrio metabólico, previo al fin de la gestación y del nuevo período de lactancia.
La alimentación y la nutrición son los elementos clave en el período de recuperación y
preparación para la nueva lactancia. La estrategia más aceptada para la prevención de la
hipocalcemia se refiere: al balance catión-anión, a dietas limitadas en Ca en el período seco,
a la administración de Ca oral al parto y al aporte balanceado de Ca durante la lactancia.
Dietas bajas en Ca durante las últimas semanas de gestación (menos de 20 g Ca/día en el
período seco) seguida de una dieta alta en Ca durante el período de lactación, reducen
manifiestamente la incidencia de la fiebre puerperal. Se propone utilizar dietas preparto bajas
en Ca para crear un balance negativo de manera de estimular los mecanismos de regulación
mediante la secreción de PTH que estimula la resorción ósea por los osteoclastos y la
producción renal de 1,25-(OH)2D3 incrementando su absorción intestinal para prevenir el
momento de máximas necesidades de Ca en el periparto. Se ha demostrado que la
hipocalcemia es principalmente causada por una alcalosis metabólica inducida por dietas de
pasturas altas en potasio. Si los niveles de potasio exceden el 2% debe considerarse
suplementar las vacas secas con sales aniónicas. El Ca ingerido durante el período seco puede
restringirse reemplazando total o parcialmente la alfalfa, conteniendo un 2% de Ca por henos
o silajes de forraje o de maíz con contenidos entre 0,4% y 0,8% del mineral.
Con respecto a las sales aniónicas, existe un acuerdo general basado en extensos trabajos
científicos que las dietas elevadas en Na+ y K+ y bajas en Cl- y SO42- tienen una influencia
negativa sobre la calcemia, mientras que proporciones inversas resultan favorables como
prevención. Los cationes monovalentes Na+ y K+, los bivalentes Ca2+ y Mg2+ y los aniones

195
SO42- y NaHCO3
tienen una influencia negativa sobre la hipocalcemia. Se han desarrollado
múltiples fórmulas para establecer la dieta con el mejor equilibrio en la homeostasis de los
minerales, calculadas sobre la base [Na+ + K+] - [Cl- + S2-]. La diferencia en la dieta de
cationes-aniones (DCAD) para prevenir la fiebre de la leche está basada en un modelo de
iones fuertes del balance ácido-base sanguíneo en el cual el pH plasmático está determinado
por cuatro factores independientes: pCO2; solubilidad del CO2 en el plasma; la diferencia de
iones y la concentración total de moléculas buffer débiles no volátiles del plasma,
principalmente albúmina, globulinas y fosfato. La base para establecer estos equilibrios
desde el punto de vista práctico es el empleo en la dieta de sales con los cationes: cloruro de
Ca (CaCl2), sulfato de Ca (CaSO4), cloruro de magnesio (MgCl2), sulfato de magnesio
(MgSO4), cloruro de amonio (NH4Cl), sulfato de amonio [(NH4)2SO4] o también los ácidos
clorhídrico (HCl) o sulfúrico (H2SO4). La absorción en el tracto digestivo del diferencial a
favor de cloruros y sulfatos con relación a los iones Ca, magnesio y amonio disminuyen el
pH plasmático. Esa inducción a la acidosis metabólica reduce el riesgo a la hipocalcemia
clínica basado en el metabolismo del Ca y la mayor disponibilidad y concentración del Ca
ionizado. Se han encontrado efectos favorables a las dietas de DCAD por: menor sensibilidad
a la PTH, aumento de síntesis de vitamina D3 por el riñón, mayor absorción de Ca en el
intestino e incremento de la resorción ósea. Como comprobación de la efectividad de las sales
para la acidificación sanguínea se recomienda su control con la medida del pH urinario,
debido a sus altos grado de correlación, donde los valores de pH 5,5–6,2 indican una correcta
acción acidificante aumentando la concentración sanguínea de Ca2+.
Finalmente, una dieta baja en Ca debe aportar mucho menos del requerimiento de la vaca
para ser capaz de preparar los mecanismos de regulación. Por ejemplo, una vaca de 600 kg
PV consumiendo 13 kg de materia seca debe ingerir una dieta que provea menos de 1,5 g/kg
de Ca, o sea menos de 20 g de Ca disponible por día. Es importante tener en cuenta que luego
del parto el animal debe ser cambiado a una dieta alta en Ca para cubrir los requerimientos
de mantenimiento y lactación, sabiendo que cada litro de leche contiene 1,2 g de Ca total y
que una vaca Holstein con 500 kg PV, con una producción de leche de 30 L/d, tiene
necesidades de al menos 52,5 g Ca/d (NRC, 2001).
En conclusión, la hipocalcemia clínica de la vaca lechera debe enfocarse con el tratamiento
medicamentoso en el caso individual, pero que es más importante el control de todo el rodeo,

196
comenzando desde el período de lactación, siguiendo por el período seco y por el período de
transición, con el manejo de la alimentación y el empleo de sales aniónicas, considerando
que todas las vacas del tambo estarán sometidas a las mismas condiciones ambientales y que
fisiológicamente esos animales presentarán un alto porcentaje de hipocalcemia subclínica,
antesala de trastornos de la salud que influirán negativamente en la producción y en la
economía del productor.
Hipomagnesemia
Antecedentes
La hipomagnesemia es una enfermedad metabólica clásica de los rumiantes, reconocida
desde 1929 por Sjollema y Seekles (in Payne, 1981), ocasionada por el descenso del
magnesio (Mg) en la sangre y caracterizada por un cuadro nervioso agudo con muerte en
tetania. Se le conoce también como tetania de los pastos, tetania de los avenales y tetania del
transporte. La alteración está determinada en primer lugar por el aporte deficiente del
mineral, y existen un par de teorías que explicarían su patogenia. Una de ellas, la más antigua,
está relacionada con la ingesta de vacas pastoreando por dos o tres semanas en gramíneas
jóvenes de rápido crecimiento o verdeos de cereales (avena) de invierno conteniendo bajos
tenores en Mg (< 0,20% MS). También es característica de las plantas jóvenes el contenido
de baja materia seca o sea de altos contenidos de agua, alto tenor en proteínas solubles y en
potasio que ingresadas al tubo digestivo aumentarían el tránsito intestinal, modificando las
fermentaciones ruminales y provocando diarreas que inducirían a una baja digestibilidad,
disminuyendo la absorción de un contenido ya de por sí bajo en Mg. Esta situación se da en
primavera u otoño que corresponde con la puesta en pasturas de las vacas para cubrir los
momentos de mayores requerimientos para mantener el pico de producción lechera. Otra
patogenia propuesta explicaría situaciones de estrés que desencadenan el cuadro clínico en
bovinos y en ovinos como ocurre en el ayuno, en adversidades atmosféricas con frío, viento
y lluvia y con el transporte. Esta respuesta está relacionada con la movilización grasa de los
adipocitos del tejido de reserva por efecto de la adrenalina, que prepara a las células para una
demanda energética incrementada para responder con el músculo esquelético a las exigencias
para la lucha o la huida, ante ese estrés. El adipocito utiliza Mg a través de su membrana para

197
realizar la lipólisis intracelular provocando una redistribución obligatoria del mineral. El
potasio podría utilizar ese mismo mecanismo, ya que es sabido que tenores elevados de
potasio son factores predisponentes que interfieren la absorción del mineral provocando la
deficiencia del balance y desarrollando la hipomagnesemia. Se han señalado como causas de
la hipomagnesemia las pérdidas del mineral por la leche particularmente durante el pico de
lactancia; el poco aporte del mineral en la ración o las pasturas; el aporte excesivo en el suelo
y las pasturas del ion K+ y la movilización de grasas que provoca la redistribución del Mg en
el organismo. Hay que señalar que contribuye con las situaciones deficitarias que las sales de
Mg de la dieta tienen un coeficiente de absorción intestinal más bajo que otros minerales con
valores de 29% del aporte. El balance nutricional de Mg y su concentración en el líquido
extracelular (2–3 g de Mg) dependen de la absorción activa y pasiva del Mg desde el rumen
y el intestino; de los requerimientos del animal para mantenimiento y producción; de la
excreción endógena del organismo y de las pérdidas por leche.
El Mg es un mineral mayor que comprende el 0,05% del peso corporal y que en una vaca de
500 kg PV hay unos 170 g de Mg en el tejido óseo, 70 g en el espacio intracelular, 2,5 g en
fluidos extracelulares y 0,7 g en sangre (Mayland, 1987, in NRC, 2001). Su forma química
se encuentra en forma libre o ionizada en el 55% o unido a proteínas en el 32% o formando
complejos con citratos y fosfatos en un 13% del total (Mayland, 1988), o expresado de otra
manera: 60% en hueso, 39% en tejidos blandos y 1% en el líquido extracelular (Contreras,
2003). El Mg participa en múltiples funciones del organismo, como activador o cofactor de
reacciones enzimáticas; en la síntesis de proteínas; en la regulación de la permeabilidad de
membranas celulares; en la movilización del Ca; en el tono y la contracción muscular y en la
regulación de la bomba de Ca2+ en el sarcoplasma; en la regulación de acetilcolina en la placa
motora; en la regulación de la estructura de RNA y DNA ribosomal, y por ello regula el
crecimiento celular en la síntesis de anticuerpos y linfocitos (Kaneko et al., 1997). Las
necesidades para una vaca de 500 kg PV con una producción de 20 L/d son: pérdidas por
leche (0,13 g/L) 2,6 g; pérdida fecal (3 mg/kg/d) 1,5 g y pérdida urinaria de 1,0 g. Por tanto,
sus necesidades de absorción son de 5,1 g/d. Ha sido propuesto que la relación K+/Ca2++Mg2+
sea < 2,2-2,5 (Kemp y 't Hart, 1957). Esos requerimientos mantendrían los valores de Mg en
sangre de 2,2 mg/dL (0,92 mmol/L). Hay varios factores de la dieta y fisiológicos que
influyen en la absorción del Mg: fundamentalmente la relación K/Mg; la relación Ca/Mg; la

198
proporción del Mg en el rumen; la relación Na+/K+ en el rumen (el aumento de K+ altera el
diferencial de potencial transmural celular y la actividad de la Na/K-ATPasa); las altas
concentraciones de amonio ruminal que actúa como quelante; la deficiencia de energía por
la microflora ruminal; el exceso de fosfatos (relación Mg/PO4) en la ración formando sales
insolubles y la edad de los animales y el pH intestinal.
Diagnóstico clínico
La hipomagnesemia se puede presentar bajo una forma subclínica y otra clínica con cuadro
nervioso de hiperexcitabilidad, tetania y muerte. La forma subclínica con ausencia de
síntomas clínicos presenta algunas manifestaciones como la disminución de la producción
láctea, la disminución de la conversión de los alimentos que puede pasar totalmente
inaparente y descubrirse solamente por la bioquímica o por la respuesta a la adición de Mg
en la dieta. La forma clínica se presenta como consecuencia de la disminución del Mg en
sangre, en el compartimento extracelular y en el líquido cefalorraquídeo afectando la
conductividad eléctrica de las terminaciones nerviosas y musculares, controlando la
excitabilidad neuronal y la transmisión de los impulsos nerviosos en las uniones
neuromusculares. Su acción actúa estabilizando las membranas biológicas en el transporte
de energía comportándose como agente de relajación muscular por ese motivo su
disminución se manifiesta por sintomatología nerviosa de excitación y parálisis espástica.
En el cuadro inicial hay disminución del apetito, disminución de la producción láctea
(Kronfeld, 1980), pero luego rápidamente aparecen cambios en el comportamiento,
volviéndose agresivo, pudiendo atacar a las personas u otros bovinos (Hicks y Pauli, 1976).
Continúan los signos de hiperexcitabilidad con respuesta exagerada a los estímulos externos
como los sonidos, hay hiperreflexia e hiperestesia, con temblores musculares en las grandes
masas musculares y cuello. La cabeza puede estar estirada y la boca abierta por dificultad
respiratoria con movimientos superficiales de mayor frecuencia, por la contractura de los
músculos intercostales. En esta etapa hay hipertermia, anorexia e intranquilidad con
defecaciones frecuentes con heces muy líquidas. Las micciones son frecuentes con orina de
color normal. Las orejas están erguidas y el reflejo anal y el palpebral aumentados, los
párpados se abren y se cierran, mostrando un nistagmus. Los síntomas neuromusculares
aumentan de intensidad presentándose astasia en la estación con rigidez en extensión de los

199
miembros, evolucionando a una incoordinación motora con ataxia que lleva a la caída del
animal en paresia espástica, instaurándose convulsiones tónico-clónicas para finalizar con un
cuadro de opistótonos, movimientos de pedaleo, rechinar de dientes, tetania y coma. Pueden
ocurrir algunos episodios de convulsiones tónico-clónicas con períodos cortos de
recuperación que se reiteran desencadenados por sonidos o simples movimientos externos,
pero finalmente se desencadena el cuadro final de tetania y muerte. Puede ocurrir que la única
manifestación visible sea la muerte súbita. A la forma clínica crónica, con desmedro del
animal, con baja producción láctea y con alteraciones óseas es difícil de reconocer como
hipomagnesemia, salvo por exámenes rutinarios del tipo de los perfiles metabólicos.
La bioquímica clínica es concluyente para confirmar el diagnóstico, con el descenso del Mg
en el suero (<1,2 mg/dL), en el líquido cefalorraquídeo (<1,45 mg/dL) y en el cadáver por la
concentración de Mg en el humor vítreo (< 1,8 mg/dL) y en la orina (< 2,5 mg/dL). Los
valores de referencia son 2,0 a 3,0 mg/dL (0,82 a 1;23 mmol/l) en suero y 2,0 mg/dL en
líquido cefalorraquídeo (Hunt, 1996). La anatomía patológica no presenta signos relevantes.
El diagnóstico se establece en base a la sintomatología clínica por el cuadro nervioso con
hiperexcitabilidad y tetania, a la patología clínica y a la respuesta al tratamiento.
Tratamiento
El tratamiento y el control se enfocan a elevar los niveles del Mg en el organismo. Para el
tratamiento se pueden usar soluciones de sulfato de Mg al 20% en dosis de 250-300 mL por
vía intravenosa lenta, teniendo en cuenta que la magnesemia no dura estable más de 24 a 48
horas ya que el bovino no tiene capacidad de almacenar el mineral y el exceso lo excretará
por vía urinaria. La formulación de Mg para administrar vía parenteral puede ser
glutamato/aspartato de Mg (Cseh, 2012). Es aconsejable aportar también óxido de Mg en
forma oral en una mezcla mineral a razón de 60 g/animal/día, mezclado con concentrado,
con melaza, disperso sobre el heno humedecido o como aditivo en ensilaje (1 kg/ton) o
mezclado con sal 20:80 (Contreras, 2009).
Prevención
Se puede recomendar para la prevención y control suministrar mezclas minerales con niveles
adecuado de Mg. En establecimientos con problemas se puede suministrar oralmente óxido

200
de Mg a dosis de 28-56 g/animal/día. Se puede fertilizar con óxido de Mg (85 kg/ha/año) o
esparcirlo sobre la pastura (28-32 kg/ha) y promover el desarrollo de leguminosas en la
pastura. Evitar desbalance de exceso de potasio en el forraje (> 2,0% MS) y el exceso de
proteína degradable o nitrógeno en la ración. Una forma práctica de administración es el
suministro de bloques para lamer o, mejor aún, de sales en bateas de libre disponibilidad.
Otra forma es suministrar fardos o rollos de forraje espolvoreados con óxido de Mg. También
puede ser práctico y aconsejable mezclar con maíz molido, afrechillo o melaza para mejorar
el gusto amargo del Mg. Otra alternativa es agregar 1,5 g de cloruro de Mg por L de agua en
los bebederos (Cseh, 2012). El Mg puede ser suministrado como bolos de Mg intrarruminales
o en sales como sulfato de Mg o cloruro de Mg; se debe tener precaución con la forma de
cloruro, porque éste es poco palatable o desagradable para los animales y no lo consumirán.
Las leguminosas tienden a acumular una mayor concentración de Mg por lo que es
recomendable el uso de asociaciones gramíneas-leguminosas para incrementar el consumo
diario del mineral contribuyendo en la prevención de la hipomagnesemia (Goff, 1998; NRC,
2001). En suma, se debería cubrir nutricionalmente las necesidades para mantenimiento y
producción, realizando análisis preventivos de suelo, alimento y animales en los períodos de
riesgo, con la precaución de manejar alternativas de control y prevención en los
establecimientos con alto riesgo de hipomagnesemia clínica.
Referencias
Cseh S, Rodríguez García M, Sciotti A, Campero C. 2012. Efecto de la suplementación con magnesio sobre
diversos parámetros en vacas con restricción alimentaria. Archivos de Zootecnia 61, 525-536.
Contreras P. 2002. Hipomagnesemia: efectos y procedimientos de prevención en los rebaños. In: Alonso Díaz,
A.J., González Montaña, J.R., Rejas López, J. Actas del Congreso de la Sociedad Española de Medicina Interna
Veterinaria. Universidad de León. España. pp. 20-29.
Contreras PA, F Wittwer, H Böhmwald. 1990. Concentraciones de calcio, fósforo y magnesio en suero
sanguíneo de bovinos de leche en 40 predios lecheros de la X Región, Chile. Arch. Med. Vet. 22, 185-189.
De Garis P, Lean I. 2008. Milk fever in dairy cows: a review of pathophysiology and control principles. Vet J.
176, 58-69.
Drackley J. 1999. Biology of dairy cows during the transition period: the final frontier. J. Dairy Sci. 82, 2259–
2273.
Dryerre H, Graig J. 1925. Milk fever: its possible association with derangements in the internal secretions. Can.
Vet. Rec. 5, 225-231.
Goff J, Horst R. 1997. Physiological changes at parturition and their relationship to metabolic disorders. J.
Dairy Sci. 80,1260–1268.

201
Goff J. 1998. Ruminant hypomagnesemic tetanies. Current Veterinary Therapy: Food Animal Practice. 4th Ed.
Howard J. W.B. Saunders. Philadelphia, USA. pp. 1-9.
Goff J. 2006. Macromineral physiology and application to the feeding of the dairy cow for prevention of milk
fever and other periparturient mineral disorders. Anim. Feed Sci. Technol. 126, 237-257.
Goff J. 2008. The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in dairy
cows. Vet. J. 176, 50-57.
Goff J. 2012. El magnesio y otros minerales en la vaca en transición. XVII Congreso Internacional ANEMBE
de Medicina Bovina. Santander: 124-128.
http://web.unillanos.edu.co/docus/libro%20ponencias%20ANEMBE%202012%281%29.pdf
Hayden C. 1934. A method for making concentrated calcium gluconate solutions. Cornell Vet, 24, 66.
Hicks J, Pauli, J. 1976. Chronic udder oedema: clinical aspects of the syndrome and its connection with
hypomagnesaemia and anaemia. N. Z. Vet. J. 24, 255-228.
Horst R, Goff J, Reinhardt T, Buxton D. 1997. Strategies for preventing milk fever in dairy cattle J. Dairy Sci.
80, 1269-1280.
Kaneko J, Harvey J, Bruss M (eds.). 1997. Clinical Biochemistry of Domestic Animals. 5a. ed., New York:
Academic Press. p. 674-687.
Kemp A, 't Hart M. 1957. Grass tetany in grazing milk cows. Neth. J. Agr. Sci. 5, 4-7.
Kronfeld D, Donoghue S, Naylor J, Johnson K, Bradley C. 1980. Metabolic effects of feeding protected tallow
to dairy cows. J. Dairy Sci. 63, 545-552.
Laporte J, Sykes A. 2004. Intestinal magnesium absorption in ruminant compensates by reduction ruminant
absorption. XI Congress International Society of Animal Clinical Biochemistry. Valdivia. Chile. p. 47.
Lean I, Van Saun R, DeGaris P. 2013. Mineral and Antioxidant Management of Transition Dairy Cows. In:
Metabolic Diseases of Dairy Cattle. Vet. Clin. Food Anim. 29, 367-386.
Mayland H. 1988. Grass tetany. In: Church (Ed.) The Ruminant Animal: Digestive Physiology and Nutrition.
Prentice Hall, Englewond Cliffs, NJ. pp 511-523.
National Research Council (NRC). 2001. Nutrient Requirements of Dairy Cattle. 7th revised edition.
Washington: National Academy Press. 381p.
Oetzel GR, MJ Fettman, DW Hamar, JD Olson. 1991. Screening of anionic salts for palatability, effects on
acid-base status and urinary calcium excretion in dairy cows. J. Dairy Sci. 74, 965-971.
Oetzel G. 2002. Milk Fever. In Encyclopedia Dairy Science. v. 2. Editorial Academic Press. p. 824-830.
Biblioteca de la División Académica de Ciencias Agropecuarias de la Universidad Juárez Autónoma de
Tabasco. México.
Oetzel G. 2013. Oral calcium supplementation in peripartum dairy cows. In: Metabolic Diseases of Dairy Cattle.
Vet. Clin. Food Anim. 29, 447.
Queen G. 1993. Effects of oral administration of a calcium-containing gel on serum calcium concentration in
postparturient dairy cows. JAVMA 202, 607-609.
Payne J. 1981. Enfermedades metabólicas de los animales zootécnicos. Ed. Acriba, 219 p.
Radostits O, Gay C, Blood D, Hinchcliff K. 1999. Medicina Veterinaria, Vol. II. Novena Edición McGaw-
Hill.Interamericana. p. 2159-2161.
Ramirez C, Tittarelli C, Mattioli G, Lasta G. 1998. Empleo del humor vítreo para la estimación postmortem de
la magnesemia en bovinos. Arch. Med. Vet. 30, 1.
Sordillo L, Raphael W. 2013. Significance of Metabolic Stress, Lipid Mobilization, and Inflammation on
Transition Cow Disorders. In: Metabolic Diseases of Dairy Cattle. Vet. Clin. Food Anim. 29, 267-278.
Underwood E, Suttle N. 1992. The Mineral Nutrition of Livestock. CABI Publishing. 3ªed. 614 p.

202
Underwood E. 1977. Trace elements in human and animal nutrition, 4ª ed. New York, Academic Press, 545 p.
Underwood, E. 1987. Los Minerales en la nutrición del ganado, 2ª ed. Zaragoza, Acribia, 210 p.

203
Deslocamento de abomaso em vacas leiteiras: ocorrência, manejo e
indicadores diagnósticos
Angélica Petersen Dias
Anne Rosi Guadagnin
Felipe Cardoso
A ocorrência do deslocamento de abomaso à esquerda ou à direita é comumente observada
em bovinos de grande porte e de alta produção leiteira após o parto, sendo que
aproximadamente 90% dos casos ocorrem até seis semanas após o parto (Radostits et al.,
2000). A prevalência desta doença varia de rebanho para rebanho dependendo da localização
geográfica, práticas de manejo e clima, dentre outros fatores. Nos EUA a incidência da
afecção está associada aos meses de inverno, provavelmente devido à maior inatividade dos
animais e à maior concentração de partos (Radostits et al., 2000). Detilleux et al. (1997)
relacionaram que as perdas econômicas relacionadas a esta doença estão na queda da
produção de leite durante o período de convalescença e o alto custo do tratamento. Estes
autores relataram que desde o parto até 60 dias após o diagnóstico, as vacas leiteiras com este
transtorno produziram 557 kg de leite a menos do que animais sadios, sendo que 30% das
perdas ocorreram antes do diagnóstico em estudo realizado com 12.572 vacas da raça
Holandesa. Raizman e Santos (2002) afirmam que a perda de produção de leite ocorre nos
primeiros 4 meses de lactação. Afirmam, também, que vacas acometidas por deslocamento
de abomaso à esquerda apresentam maior intervalo do parto até primeira inseminação
artificial e são duas vezes mais suscetíveis a outras doenças que os animais sem problemas
de saúde.
No estado do Rio Grande do Sul (sul do Brasil), este distúrbio é encontrado em bacias
leiteiras de alta produção. Inexiste, entretanto, um estudo de prevalência da doença no Brasil.
O avanço do conhecimento técnico para o diagnóstico do deslocamento de abomaso
provavelmente foi o responsável pelo aumento do número de casos observados no Estado.
Segundo relato de médicos veterinários, com experiência nesta afecção, a ocorrência é maior
no período inicial de inverno e final de primavera. Uma das possíveis explicações para esta
observação é que nestes períodos as pastagens de inverno e verão, respectivamente, ainda
não estão estabelecidas gerando um déficit de fibra na dieta destes animais. Nesse contexto,

204
é necessário o conhecimento dos eventos metabólicos que ocorrem antes e depois da
apresentação do transtorno, principalmente em sua relação com o manejo alimentar dos
animais e o envolvimento de patologias associadas, para que se possam diminuir as perdas
econômicas através de um diagnóstico eficaz e um monitoramento adequado do tratamento.
Os objetivos deste artigo são revisar as causas de deslocamento de abomaso, sua patogenia e
o impacto em distúrbios metabólicos associados e avaliar como diferentes estratégias
nutricionais antes e depois do parto podem afetar esta patogenia através da adequação dos
nutrientes. O tema central do artigo é o manejo nutricional para promover maior ingestão de
matéria seca (IMS) após o parto, e menor redução de IMS no pré-parto, que são um fator
unificador para o sucesso no período de transição e a prevenção do deslocamento de
abomaso.
Etiologia e epidemiologia
O deslocamento de abomaso (DA) é uma doença multifatorial relacionada com o manejo
alimentar que afeta vacas leiteiras de alta produção, principalmente durante o início da
lactação. A atonia abomasal é um pré-requisito para a sua ocorrência. O deslocamento de
abomaso para esquerda (DAE) é predominante em 85 a 96% das ocorrências (Trent, 1990).
O fornecimento de altos níveis de concentrado (grãos) na dieta aumenta a taxa de passagem
do alimento pelo rúmen, causando um aumento na concentração de ácidos graxos voláteis
(AGV) que pode inibir a motilidade do abomaso. O grande volume de metano e dióxido de
carbono encontrado no abomaso após a ingestão de grãos pode ser responsável pela sua
distensão e deslocamento (Van Winden et al., 2002). Em uma vaca saudável, a produção, a
difusão e o transporte de gás estão em equilíbrio, portanto não ocorre o acúmulo (Figura 1).
Uma concentração de fibra bruta menor que 16% é considerado um fator predisponente para
o DA. O fornecimento de uma dieta experimental completamente peletizada resultou em um
aumento na incidência de DA em 17%, enquanto uma dieta contendo feno de alfafa, silagem
de sorgo e concentrado com 18% de proteína bruta causou uma incidência de 1,6% (Dawson
et al., 1992).
Sugere-se que no estágio avançado de gestação, o volume uterino aumentado ocupa uma
porção do espaço do rúmen, e a retração uterina logo após o parto leva a uma predisposição
anatômica de deslocamento do abomaso (Goff & Horst, 1997). O DAE ocorre principalmente

205
no período de duas a oito semanas pós-parto. Wolf et al. (2001) verificaram que mais de 75%
dos casos de DA ocorrem nos primeiros 30 dias pós-parto. Tabeleao et al. (2005), em estudo
realizado no Paraná, encontraram maior ocorrência desse transtorno nos 10 dias após o parto.
Shaver (1997) relatou que 80 a 90% dos casos de DAE são diagnosticados no primeiro mês
pós-parto.
Figura 1. A) Vista lateral esquerda do aparelho digestório de um bovino normal. B) Vista lateral esquerda do
aparelho digestório do bovino com DAE (Turner & Mcllwrath, 1985)
Segundo Cardoso et al. (2008), o deslocamento de abomaso à esquerda é uma afecção com
distribuição mundial, com comprovada incidência na Região do Planalto do Rio Grande do
Sul. Em animais com DAE, a produção de leite, o peso e o escore de condição corporal
encontram-se diminuídos. A fita reagente indicativa de pH é uma boa ferramenta a campo
para a determinação do pH ruminal. O tempo de redução do azul de metileno (TRAM) é
significativamente menor no DAE, indicando que um dos fatores a ser corrigido no
tratamento pós-cirúrgico é a reposição do equilíbrio ruminal. Os valores séricos de lactato,
β-hidroxibutirato (BHB) e AST estão aumentados nos animais com DAE, sendo indicativos
para o diagnóstico auxiliar desta afecção. Dentro dos parâmetros utilizados, TRAM,
produção de leite, hematócrito, ureia, escore de condição corporal e BHB mostraram-se como
os mais eficazes na caracterização desta afecção.

206
Importância econômica
Detilleux et al. (1997) concluíram que as perdas econômicas relacionadas com essa doença
estão na queda na produção de leite durante o período de convalescência e o alto custo do
tratamento. Segundo esses autores, as vacas leiteiras com esse transtorno produziram 557 kg
de leite a menos em comparação a animais sadios, desde o parto até 60 dias após o
diagnóstico. Segundo Raizman e Santos (2002), a perda de produção de leite acontece nos
primeiros 4 meses de lactação e as vacas acometidas por DAE apresentam maior intervalo
do parto até a primeira inseminação artificial. Outros custos eventuais resultantes são o custo
referente à perda do animal, caso venha a óbito, e custos referentes às doenças secundárias
ao DAE. Segundo Bartlett et al. (1995), as despesas com cada caso de DAE variam entre
US$200 e US$400 (Bartlett et al., 1995) envolvendo custos cirúrgicos, desempenho da
lactação, custo de reposição da vaca e valor de abate do animal. Baseado nos valores descritos
por Bartlett et al. (1995), Andrade (2005) estimou os custos de dois casos de DAE que
ocorreram na região do Planalto Riograndense. Para tal, estimou gráficos de produção
esperada de leite para cada animal e calculou as despesas com medicamentos e custos
cirúrgicos. Os animais analisados encontravam-se na 1ª e 3ª lactação quando da ocorrência
de DAE, de modo que a produção esperada de leite foi calculada através da média de
produção de animais em 1ª e 3ª lactação no rebanho, respectivamente (Figura 2). Observa-
se que a queda na produção de leite de animais acometidos por DAE pode perdurar entre 3 a
6 meses, sendo um prejuízo considerável em termos financeiros. Os animais foram
submetidos à correção cirúrgica do deslocamento de abomaso, de modo que somado ao custo
decorrente da queda na produção leiteira está o custo do procedimento cirúrgico e os
medicamentos utilizados (Tabela 1). As perdas totais decorrentes da doença foram de US$
496,63. Desta forma os valores encontrados ultrapassaram os descritos por Bartlett et al.
(1995), demonstrando a importância da prevenção dessa doença. Convém ressaltar que não
estão incluídos nestes cálculos os custos referentes à eventual perda de animais decorrentes
da doença ou custos com doenças secundárias ao DAE, bem como os custos referentes ao
deslocamento do médico veterinário até a propriedade. Fleischer (2001) cita que o
deslocamento de abomaso está relacionado com a produção de leite: quanto maior a
produção, maior o risco de desenvolver DA. Geralmente, as vacas que desenvolvem

207
deslocamento de abomaso são animais de alta produção, mas devido ao DA o animal
apresenta um baixo desempenho em produção leiteira na lactação corrente.
Patogenia
Existem doenças associadas que predispõem o DA, como aquelas que resultam em anorexia
e inapetência, devido a uma diminuição do volume ruminal. Uma menor ingestão de matéria
seca (IMS), especialmente no período de transição, é um dos principais fatores de risco para
o desenvolvimento do DA, devido ao menor volume ruminal. Radostits et al. (2006) relatam
que dietas pré-parto com alta densidade energética como preparação para o início da lactação,
aumentam os fatores de risco para o DA. Logo, o consumo de fibra efetiva é importante para
o ruminante e não deve ser menosprezado no momento das formulações da dieta. A cetose
comumente diagnosticada antes do DA está fortemente associada com o problema, uma vez
que reduz o consumo de matéria seca e o preenchimento ruminal, diminuindo a motilidade
dos demais estômagos e, potencialmente, a motilidade do abomaso (Van Winden et al.,
2003b). Cardoso et al. (2008) encontraram valores de BHB de 1,14 ± 1,1 mmol/L para
animais com DA e de 0,69 ± 0,28 mmol/L no grupo controle, relacionando os valores
aumentados de BHB ao maior risco de DA, e confirmando os dados encontrados por Van
Winden et al (2002). Um estudo desenvolvido com 528 vacas de leite indicou que vacas com
valores de BHB >1,6 mmol/L durante as primeiras duas semanas pós-parto tem 6,9 vezes
mais chance de desenvolver DA quando comparadas com vacas que possuem menores
concentrações de BHB sanguíneo (Suthar et al., 2013).
A hipocalcemia também é uma patologia predisponente ao DA. Os níveis sanguíneos de
cálcio afetam diretamente a motilidade do abomaso, de forma que em concentrações abaixo
de 4,8 mg/dL não há motilidade. Massey et al. (1993) conduziram um estudo com 510 vacas
leiteiras e concluíram que todos os animais diagnosticados com hipocalcemia antes do parto
(< 7,9 mg/dL) tiveram 4,8 vezes mais chances de desenvolver DA. Com o intuito de prevenir
a hipocalcemia e consequentemente o DA, a diferença cátion-aniônica da dieta (DCAD) pode
ser utilizada durante as três últimas semanas pré-parto. Em alguns casos, a porção
deslocada do abomaso fica presa entre o retículo e o diafragma, o que resulta em um estado
de inanição severa e compromete a digestão e o movimento da ingesta. Uma alcalose
metabólica leve com hipocloremia e hipocalemia são comuns, devido provavelmente à atonia

208
abomasal e contínua secreção de ácido clorídrico com prejuízo no fluxo de alimento para o
duodeno (Howard & Smith, 1999).
Figura 2. O gráfico acima compara a produção de leite de uma vaca com DAE em 1ª lactação em
comparação com a produção esperada. O gráfico embaixo compara a produção de leite de uma vaca
com DAE em 3ª lactação com a produção esperada (Andrade, 2005)
Tabela 1. Estimativa de despesas totais decorrentes
de DAE em bovinos leiteiros (Andrade, 2005)
Item US$
Perdas em produção de leite 265,75
Terapia medicamentosa 35,58
Tratamento cirúrgico 196,30
Total 496,63
Nos casos de DA à direita, pode haver um comprometimento maior do abomaso devido à
possibilidade de ocorrência de vólvulo em diferentes graus, podendo chegar até 360°. Este
loop ocorre no sentido anti-horário tendo como ponto de visão o posterior do animal ou o

209
lado direito. A ingestão diminuída de líquido e o sequestro de grandes volumes de ácido
clorídrico no abomaso levam à desidratação e volemia (Gesihauser & Schemann, 1998). O
vólvulo pode provocar uma obstrução do fluxo sanguíneo através do abomaso. Este fato pode
levar a congestão, edema, e, eventualmente, necrose da parede do abomaso (Habel et al.,
1981).
Cardoso et al. (2008) realizaram um estudo sobre as variações do hemograma e perfil
bioquímico sanguíneo em vacas com DAE. Concluíram que em vacas acometidas por esse
transtorno a produção de leite, o peso e o escore de condição corporal diminuem. O tempo
de redução do azul de metileno (TRAM) é significativamente menor em animais com DAE.
Os valores séricos de lactato, BHB, ácidos graxos livres, ureia e AST apresentam-se
aumentados, enquanto os de proteína total, albumina e colesterol diminuem nos animais com
DAE. Portanto, esses metabólitos são indicativos para o prognóstico e tratamento desta
afecção.
Diagnóstico
Rosemberger (1990) descreveu os sinais observados no DA. Frequentemente os animais
apresentam uma queda brusca no consumo de grãos enquanto ainda continuam consumindo
forragens. As fezes apresentam-se moles e reduzidas sendo que períodos de diarreia quase
sempre ocorrem. A temperatura retal e as frequências cardíaca e respiratória encontram-se
normais na maioria dos casos. Os movimentos ruminais apresentam-se diminuídos na sua
frequência e intensidade. Animais com um quadro agudo de vólvulo ficam deitados 24 horas
após o episódio e podem vir a óbito dentro de 48 a 96 horas devido ao choque e desidratação.
Nos casos de DAE, o diagnóstico pode ser realizado através da auscultação e percussão do
flanco esquerdo localizando-se o som metálico característico de “ping” (Rosenberger, 1990).
A maioria dos deslocamentos pode ser encontrada no meio de uma linha imaginária
estabelecida entre a tuberosidade coxal esquerda e o cotovelo esquerdo. O “ping” pode estar
localizado desde a nona costela até a fossa paralombar esquerda. Caso exista dúvida na
origem do “ping” entre rúmen ou abomaso, pode-se realizar uma aspiração do líquido
presente na região de gás e aferir o pH, que deve diferenciar entre rúmen (pH 6-7) e abomaso
(pH 2-3) (Rosenberger, 1990). Nos casos de DA à direita as técnicas de diagnóstico são as
mesmas do DAE. Deve-se ter o cuidado de diferenciar quaisquer outras patologias que

210
possam provocar o “ping” no flanco direito. A mais comum é a dilatação e/ou torção de ceco
que pode ser diferenciada através de palpação retal (Howard & Smith, 1999).
Fatores genéticos
Fatores genéticos têm sido aceitos como um dos fatores predisponentes para a ocorrência de
DA. A herdabilidade (h2) foi estimada entre 0,15 a 0,3 para DA em geral (Uribe et al., 1995;
Zwald et al., 2004) e até 0,53 para DAE (Hamann et al., 2004). Vários loci de características
quantitativas (QTL) para DA têm sido identificados em vacas Holandesas (Mömke et al.,
2008), entre eles, o locus proximal no cromossomo 23 (BTA23). Uma vez que o DA é
precedido por uma diminuição da motilidade do abomaso, o gene motilina (MLN) localizado
proximalmente no BTA23, que codifica um pequeno hormônio regulador da contração
gastrointestinal, é um dos genes importantes e necessita ser estudado. Mömke et al. (2012)
identificaram um polimorfismo de nucleotídeo único (SNP) que afeta o sítio de ligação
NKX2-5 dentro do gene bovino MLN. Esse polimorfismo mostrou estar correlacionado com
uma diminuição significativa na expressão do MLN e com um significante aumento da
incidência de DAE em vacas Holandesas. Esse é o primeiro polimorfismo demonstrando uma
associação com o DAE e pode ser utilizado como teste de suscetibilidade nessa raça. Zwald
et al. (2004) encontraram uma correlação genética moderada entre DA e cetose (0,45 ± 0.16)
na primeira lactação de vacas Holandesas nos Estados Unidos. Neuenschwander et al. (2012)
encontraram uma alta correlação genética (0,58 ± 0.13) entre DAE e cetose com vacas
Holandesas no Canadá.
Tratamento
Os principais objetivos do tratamento do DA para esquerda ou para a direita ou vólvulo são
de recolocar o abomaso em sua posição original ou aproximada, criar uma ligação
permanente nesta posição, corrigir o balanço eletrolítico e a desidratação do animal e
providenciar tratamento apropriado para doenças associadas. Nenhuma das técnicas permite
a identificação exata do local de fixação do abomaso e existe a possibilidade de vazamento
de líquido abomasal no abdômen, ocasionando peritonite. Podem ocorrer complicações,
como a fixação de outras estruturas (como o rúmen e o intestino) ou fixar o abomaso em uma
posição equivocada (Fubini & Ducharme, 2004). A escolha da técnica deve ser aquela em

211
que o cirurgião esteja mais habituado e em que se sinta mais confortável realizando, uma vez
que todas elas apresentam resultados e períodos de recuperação semelhantes.
A abomasopexia pelo flanco esquerdo é utilizada para visualizar uma porção do abomaso
com o animal em pé. A colocação das suturas na região paramediana ventral direita deve ser
feita cuidadosamente para evitar a fixação de outras estruturas ao mesmo tempo na sutura
(Howard & Smith, 1999). A omentopexia pelo flanco direito é uma técnica muito bem aceita,
mas deve-se ter cuidado, pois muita força é disposta sobre o omento no momento da sutura
(Fubini & Ducharme, 2004). Todos os animais com DA ou vólvulo apresentam algum
distúrbio eletrolítico. A composição do fluido administrado pode ser ajustada conforme o
perfil bioquímico destes animais. Soluções isotônicas salinas e Ringer são comumente
utilizadas e apresentam bons resultados (Huhn & Nelson, 1995). O volume de líquido a ser
administrado depende do grau de desidratação do animal. A hidratação oral pode ser
utilizada, mas não é substituível à administração endovenosa, quando o animal apresenta um
grau de desidratação igual ou maior que 8%. A utilização de antimicrobiano fica a critério
do médico veterinário que deve levar em consideração o tempo do procedimento, assepsia
do tratamento cirúrgico e a manipulação que foi realizada no procedimento (Howard &
Smith, 1999).
Controle
Por se tratar de uma doença multifatorial, a identificação dos fatores predisponentes é a
melhor forma de prevenção. A diminuição na ingestão de matéria seca no pré-parto e o
gradual aumento na ingestão no pós-parto são fatores de risco, pois causam menor
preenchimento ruminal e aumento na incidência de doenças relacionadas ao período pós-
parto. Retenção de membranas fetais, metrite, cetose clínica ou subclínica e hipocalcemia
são fatores de risco prováveis ao desencadeamento de DAE. Da mesma forma, quantidades
excessivas de concentrado ou aumento repentino na quantidade de concentrado fornecido
durante o período pós-parto aumentam o risco de ocorrer DAE, pois a maior concentração de
AGV no conteúdo abomasal leva à diminuição da motilidade abomasal e do seu
esvaziamento, com consequente excesso de gás no abomaso (Radostits et al., 2006). A
alimentação e o manejo adequados impedem distúrbios no período pré e pós-parto, reduzindo
o risco de DA. Alguns princípios são importantes na prevenção da doença, como o ajuste da

212
dieta durante o período seco para evitar que as vacas estejam obesas no momento do parto,
facilitar o processo de adaptação das vacas em início de lactação (tanto nutricional quanto
socialmente), otimizar a IMS nos períodos pré e pós-parto imediato, garantir água e
alimentação palatável à vontade para as vacas em período periparturiente e não exceder de
1,65 Mcal EL/Kg de MS em densidade energética da dieta no período próximo ao parto. A
utilização de feno como fonte de fibra durante o período seco é essencial para o enchimento
do rúmen, fermentação ruminal e motilidade do trato gastrointestinal (Goff & Horst, 1997).
Uma dieta rica em fibras causa uma expansão física do rúmen, promovendo uma barreira
contra a migração do abomaso. Uma característica importante é que a fonte de fibra fornecida
durante o período seco deve ser a mesma fornecida durante o período inicial da lactação, pois
isso permite uma adaptação mais rápida da fermentação ruminal (Curtis et al., 1985).
Fornecer uma dieta rica em forragens é uma das mais comuns e mais eficazes formas de
manejo estratégico para minimizar a ocorrência de DAE durante o período periparturiente.
Isto implica em garantir no mínimo um teor de 17% de fibra na dieta (Radostitis, 2007).
O fornecimento de propilenoglicol resulta numa menor incidência de DA quando utilizada
como tratamento metafilático no pós-parto imediato. Esse fato se deve às propriedades
gliconeogênicas que incrementam as concentrações plasmáticas de glicose e insulina e
diminuem as concentrações de ácidos graxos não esterificados e de BHB (Studer et al., 1993).
Uma alternativa é utilizar monensina na forma de cápsulas de liberação controlada, visto que
a monensina é um antibiótico ionôforo que altera a produção de AGV em favor da produção
de propionato, o maior precursor de glicose no ruminante. Desta forma, os efeitos da
monensina são mediados por melhora no balanço energético e consequente diminuição na
incidência de cetose subclínica, DAE e outras doenças metabólicas. A administração de
monensina CRC (controlled-release capsule) 3 semanas antes do parto diminui as
concentrações de ácidos graxos não esterificados e BHB e aumenta a concentração de
colesterol e ureia sérico na semana imediatamente após o parto (Duffield et al., 2005). Isto
indica um metabolismo energético mais efetivo em vacas que recebem monensina CRC, o
que é importante para a prevenção de retenção de placenta e cetose clínica, além da prevenção
de DAE. Pode-se esperar por uma redução de 40% na ocorrência de DAE e cetose clínica
com a administração pré-parto de monensina CRC e 25% menos ocorrências de retenção de
placenta (Radostitis et al., 2006).

213
Conclusão
Pelas perdas e incidência apresentadas por esta patologia, fica claro que é um problema
extremamente importante na atividade leiteira, e que deve receber especial atenção no que
diz respeito à prevenção. Cerca de 30% destas perdas ocorrem antes do diagnóstico e, além
das perdas em produção leiteira, terapêutica medicamentosa e procedimento cirúrgico,
devem-se considerar as eventuais perdas de animais, sejam decorrentes ou secundárias desta
patologia, e o fato que estes animais ficam mais susceptíveis a outras enfermidades. Entre
as diversas causas da inapetência, algumas devem receber atenção especial, tais como, a
nutrição [relação energia x fibra (FDN e FDA)], o estresse ambiental (calor, confinamento,
etc.), e todas as situações que possam desencadear a febre (doenças, parto, lesões de cascos,
etc.). A cetose é um destes prognósticos mais comuns, devido ao balanço energético negativo
que se instala, justamente pela inapetência neste período. Manter a ingestão normal durante
o período de transição, não só evita a mobilização de gordura como fonte energética, como
também, preserva a barreira ruminal ao DAE, pelo consumo de matéria seca.
Referências
Andrade, E.L.C. 2005. Deslocamento de abomaso à esquerda em bovinos. Monografia (Graduação), Curso de
Medicina Veterinária, Universidade Federal do Rio Grande do Sul, Porto Alegre, 56 p.
Bartlett, P. C. et al. 1995. Economic comparison of the pyloro-omentopexy vs the roll-and-toggle procedure for
treatment of left displacement of the abomasum in dairy cattle. Journal of the American Veterinary Medical
Association 206, 1156-1162.
Cardoso, F.C. et al. 2008. Hematological, biochemical and ruminant parameters for diagnosis of left
displacement of the abomasum in dairy cows from Southern Brazil. Pesq. Agropec. Bras. 43, 141-147.
Curtis, C.R. et al. 1985. Path analysis of dry period nutrition, postpartum metabolic and reproductive disorders
and mastitis in Holstein cows. Journal of Dairy Science 68, 2347-2360.
Dawson, L. et al. 1992. Influence of fiber form in a complete mixed ration on incidence of left displacement
abomasum in postpartum dairy cows. Journal of the American Veterinary Medical Association 200, 1989-1992.
Detilleux, J.C.; Grohn, Y.T.; Eicker, S.W.; Quaas, R.L. 1997. Effects of left displaced abomasum on test day
milk of Holstein cows. Journal of Dairy Science 80, 121-126.
Duffield, T.F. et al. 2005. The impact of controlled release capsules of monensin on postcalving haptoglobin
concentrations in dairy cattle. Canadian Journal of Veterinary Research 69, 208-214.
Fleischer P. et al. 2001. The relationship between milk yield and the incidence of some diseases in dairy cows.
Journal of Dairy Science 84, 2025-2035.
Fubini, S.L.; Ducharme, N.G. 2004. Farm Animal Surgery. St. Louis: W.B. Saunders, 607 p.
Gesihauser, T.; Reiche, D.; Schemann, M. 1998. In vitro motility disorders associated with displaced abomasum
in dairy cows. Neurogastroenterology and Motility 10, 395-401.

214
Goff, J.P.; Horst, L.R. 1997. Physiological changes at parturition and their relationship to metabolic disorders.
Journal of Dairy Science 80, 1260-1268.
Habel, R.E. et al. 1981. Volvulus of the bovine abomasum and omasum. Journal of the American Veterinary
Medical Association 179, 447-455.
Hamann, H. et al. 2004. Relationships between lactational incidence of displaced abomasum and milk
production traits in German Holstein cows. Journal of Veterinary Medicine Series A 51, 203-208.
Howard, J.L.; Smith, R.A. 1999. Current veterinary therapy: Food Animal Practice. 4th ed. Edinburg: W.B.
Saunders, 766 p.
Huhn, J.C.; Nelson, D.R. 1995. Right-sided abomasal problems in dairy cattle. Veterinary Medicine 90, 1169-
1174.
Massey, C.D. et al. 1993. Hypocalcemia at parturition as a risk factor for left displacement of the abomasum in
dairy cows. Journal of the American Veterinary Medical Association 203, 852-853.
Mömke, S. et al. 2008. Mapping quantitative trait loci for milk performance traits in German Holstein cows
affected by left-side displaced abomasum (LDA). Journal of Dairy Science 91, 4383-4392.
Mömke, S. et al. 2012. Transcription factor binding site polymorphism in the motilin gene associated with left-
sided displacement of the abomasum in German Holstein cattle. PLoS ONE, 7, n. 4.
Neuenschwander, TF. et al. 2012. Genetic parameters for producer-recorded health data in Canadian Holstein
cattle. International Journal of Animal Bioscience 6, 571-578.
Radostits, O.M. et al. 2000. Veterinary Medicine, 9th ed. Edinburg: W.B. Saunders, 1877 p.
Radostits, O.M. et al. 2006. Veterinary Medicine, 10th ed. Edinburg: W. B. Saunders, 2156 p.
Raizman, E.A.; Santos, J.P. 2002. The effect of left displacement of abomasum corrected by toggle-plin suture
on lactation, reproduction, and health of Holstein dairy cows. Journal of Dairy Science 85, 1157-1164.
Rosenberger, G. 1990. Exame clínico dos bovinos. 3ª ed. Rio de Janeiro: Guanabara Koogan, p. 193-200.
Shaver, R.D. 1997. Nutritional risk factors in the etiology of left displaced abomasum in dairy cows: a review.
Journal of Dairy Science 80, 2449-2453.
Suthar, VS. et al. 2013. Prevalence of subclinical ketosis and relationships with postpartum diseases in
European dairy cows. Journal of Dairy Science 96, 2925-2938.
Studer, V.A.; Grummer, R.R.; Bertics, S.J. 1993. Effect of prepartum propylene glycol administration
periparturient fatty liver in dairy cows. Journal of Dairy Science 76, 2931-2939.
Tabeleao, V.C. et al. 2005. Ocorrência de deslocamento de abomaso em rebanhos leiteiros na região Centro-
Sul do Paraná. In: Congresso de Iniciação Científica, 14. Anais. Pelotas: Universidade Federal de Pelotas.
Trent, A.M. 1990. Surgery of the bovine abomasum. Veterinary Clinics of North America: Food Animal
Practice 6, 399-448.
Turner, A.S.; McIlwrath, C.W. 1985. Cirurgia gastrointestinal do bovino. In: Técnicas cirúrgicas em animais
de grande porte. São Paulo: Roca, p. 235-262.
Uribe, H.A. et al. 1995. Genetic parameters for common health disorders of Holstein cows. Journal of Dairy
Science 93, 3561-3568.
Van Winden et al. 2002. Studies on the pH value of abomasal contents in dairy cows during the first 3 weeks
after calving. Journal of Veterinary Medicine Series A 49, 157-160.
Wolf, V. et al. 2001. Influences on the occurrence of abomasal displacements in German Holstein cows.
Deutsche Tierarztliche Wochenschrift 108, 403-408.
Zwald, N.R. et al. 2004. Genetic selection for health traits using producer-recorded data. II. Genetics
correlations, disease probabilities, and relationships with existing traits. Journal of Dairy Science 87, 4295-
4302.

215
Alimentação e manejo de bezerras leiteiras
Carla Maris Machado Bittar
O manejo e a alimentação de bezerras são determinantes das taxas de morbidade e
mortalidade, assim como do desempenho animal durante esta fase e a fase subsequente, tendo
forte impacto na planilha de custo de animais de reposição. Os sistemas criação são bastante
variados e parte do rebanho nacional ainda cria bezerros ao pé da vaca, com aleitamento
natural; enquanto outra fração realiza o aleitamento artificial e utiliza tecnologias que
aumentam a eficiência do sistema (Santos & Bittar, 2015). O manejo alimentar de bezerras
tem início no fornecimento de colostro e culmina com o processo de desaleitamento dos
animais, importante para a manutenção do desempenho de animais recém desaleitados.
Cuidados com a recém-nascida
Os cuidados com as bezerras recém-nascidas começam ainda antes de seu nascimento,
quando vacas pré-parto são vacinadas contra patógenos que acometem animais jovens.
Devido ao tipo de placenta dos bovinos, os bezerros recém-nascidos são desprovidos de
imunoglobulinas (Ig) circulantes, dependendo do consumo de colostro para aquisição da
chamada imunidade passiva (Davis & Drackley, 2002). Dessa forma, dos cuidados com o
bezerro recém-nascido, o fornecimento de colostro é o mais importante para a redução nas
taxas de mortalidade e morbidade (Besser and Gay, 1994), assim como o desempenho
durante sua vida (Faber et al., 2005).
O sucesso da colostragem depende basicamente de três fatores: 1) tempo para fornecimento;
2) qualidade do colostro (concentração de Ig); e 3) volume de fornecimento (Quigley, 1996).
A concentração de imunoglobulinas no colostro tem sido tradicionalmente utilizada como
sinônimo de qualidade, no entanto, a carga bacteriana e a presença de patógenos como
também devem ser consideradas (Stewart et al., 2005). O colostro tem composição um pouco
diferente do leite, apresentando menores teores de lactose, mas maiores teores de gordura,
sólidos totais, minerais e vitaminas, e principalmente proteína (Tabela 1). O maior teor de
proteína do colostro se deve principalmente ao maior teor de Ig. Conforme as ordenhas, a
composição do colostro vai sendo alterada até a secreção ser considerada como leite.

216
Tabela 1. Alteração na composição de colostro conforme as ordenhas (adaptado de Foley & Otterby, 1978)
Parâmetro Colostro (ordenha pós-parto)
1 2 3 Leite
Gravidade especifica 1,056 1,040 1,035 1,032
Sólidos, % 23,9 17,9 14,1 12,9
Proteína, % 14,0 8,4 5,1 3,1
Caseína, % 4,8 4,3 3,8 2,5
IgG, mg/mL 48,0 25,0 15,0 0,6
Gordura, % 6,7 5,4 3,9 3,7
Lactose, % 2,7 3,9 4,4 5,0
A relação linear entre gravidade específica do colostro e concentração de anticorpos permite
o uso de um densímetro, chamado de colostrômetro, para o monitoramento da qualidade de
colostro (Figura 1). Entretanto, como a gravidade específica é dependente da temperatura, as
leituras devem seguir as recomendações do fabricante, de modo a não super ou subestimar a
qualidade. O colostro pode ser classificado em três faixas, de acordo com a concentração de
Ig: 1) Baixa qualidade: < 22 mg/mL; 2) Média qualidade: 22-50 mg/mL; e 3) Alta qualidade:
> 50 mg/mL. Outra ferramenta que pode ser utilizada para avaliação da qualidade do colostro
é o refratômetro de Brix (Figura 1), o qual tem uma menor dependência da temperatura para
avaliação. Quando utilizamos o refratômetro, consideramos o colostro como de alta
qualidade quando o mesmo apresenta leituras superiores a 22% (Quigley et al., 2013).
Para se garantir uma adequada transferência de imunidade, o colostro de alta qualidade deve
ser fornecido logo após o nascimento ou o mais rápido possível, uma vez que a absorção de
Ig é reduzida com o passar do tempo, não ocorrendo após 24h de nascimento (Godden, 2008).
Assim, a transferência de imunidade passiva é dependente da capacidade de absorção de Ig
(fator tempo) e da quantidade de Ig ingerida (fator qualidade). A adequada transferência de
imunidade passiva adequada ocorre quando o animal apresenta às 48 horas de vida,
concentração maior ou igual a 10 mg/mL, sendo 15 mg/mL o ideal, o que é garantido pelo
fornecimento de colostro de boa qualidade (> 50 mg/mL) nas primeiras horas de vida.
Quando estes valores não são alcançados, as taxas de mortalidade são significativamente
aumentadas.

217
Figura 1. Colostrômetro e refratômetro utilizados na avaliação de qualidade de colostro
A recomendação é que os animais recebam 10% de seu peso ao nascer nas primeiras 6 h após
o nascimento, sendo importante que a primeira refeição ocorra o mais cedo possível.
Considerando que os animais devem apresentar concentração sérica de IgG> 10 mg/mL, que
o volume de plasma de bezerros (~6,5% PV) e a que a eficiência aparente de absorção de
IgG é de 25%, obtém-se uma massa de IgG de 104 g, sendo esta a dose necessária no
fornecimento fornecida (Davis & Drackley, 1998). Assim, de acordo com a qualidade do
colostro, o volume fornecido deve ser alterado para que o consumo final seja de
aproximadamente 100 g de IgG. Para garantir este consumo, quando o animal não mama
voluntariamente o colostro, o uso de sonda esofágica é recomendado.
A aquisição de imunidade passiva pelos animais pode ser monitorada através de leituras com
refratômetro de proteína ou de Brix do soro destes animais até por volta das 48 h de vida.
Existe alta correlação de proteína sérica e de Brix com a concentração de Ig no soro de
bezerros (Deelen et al., 2014), o que permite inferir sobre o sucesso da colostragem. Quando
se utiliza o refratômetro de proteína os seguintes valores de leitura são considerados: 1)
transferência passiva adequada: > 5,5 g/ dL; transferência passiva moderada: 5,0-5,4 g/dL;
transferência passiva insuficiente: < 5,0 g/dL (Quigley, 2001). Quando o refratômetro de Brix
é utilizado, considera-se adequada transferência quando se obtém leituras > 8,4% (Deelen et
al., 2014).
A formação de um banco de colostro na propriedade garante a disponibilidade de colostro de
boa qualidade e em quantidade suficiente para todos os recém-nascidos. Alguns trabalhos
mostram que a conservação em geladeira pode ser feita durante até uma semana, enquanto a
conservação em freezer, mais utilizada em fazendas leiteiras, pode ser feita por até um ano
sem redução na qualidade (Davis & Drackley, 1998). O colostro deve ser congelado em

218
porções que facilitem seu descongelamento, que deve ser em banho-maria com temperatura
de até 50C (Elizondo-Salazar et al., 2010), para que não tenha sua qualidade reduzida.
Além de prover anticorpos aos animais até que estes tenham seu sistema imune ativo, o
consumo de colostro tem impacto na vida futura. Faber et al. (2005) mostraram que animais
colostrados com maiores volumes de colostro de alta qualidade apresentaram maior taxa de
crescimento e maiores produções de leite na primeira e segunda lactação. Embora a
colostragem seja a atividade mais importante a ser realizada com a recém-nascida, outras
práticas de manejo também são necessárias no primeiro dia de vida. A cura do umbigo deve
ser realizada ao nascer, e repetida pelo menos duas vezes ao dia, utilizando-se solução de
iodo 5-7%. A cura deve ser feita por imersão do cordão, de preferência com o animal em pé,
de forma que partículas e sujidades sejam lavadas com o excesso de iodo. Em torno de 3-4
dias o cordão já deve estar completamente mumificado e ter se destacado do animal. A cura
inadequada do umbigo é uma das causas de morte de animais jovens em decorrência da
formação de abscessos e de entrada de patógenos na circulação do animal.
A identificação e a pesagem do animal ao nascer são importantes para o acompanhamento
da vida produtiva do animal. Através da identificação e da anotação de data de nascimento e
peso, uma agenda pode ser elaborada para cada animal contendo as datas de desaleitamento,
vacinação, vermifugação, etc. Além disso, a pesagem do animal ao nascer permitirá a
avaliação do manejo alimentar durante o aleitamento.
Manejo alimentar
Os sistemas de aleitamento podem ser divididos em natural e artificial. O aleitamento natural
ainda ocorre em propriedades leiteiras, geralmente quando animais não especializados para
a produção de leite são utilizados, não havendo possibilidade de se ordenhar as vacas sem a
presença do bezerro. Um levantamento nacional mostra que ainda em torno de 35% das
propriedades leiteiras (Paraná, São Paulo e Minas Gerais) ainda tem bezerros mamando
diretamente de suas mães (Santos & Bittar, 2015). O sistema natural de aleitamento não é o
mais adequado do ponto de vista de manejo de bezerros, pois não permite o conhecimento
do volume de leite consumido. Este sistema resulta em grande variação no desempenho de
animais contemporâneos em resposta a variação na produção de leite de suas mães, maior ou
menor volume disponibilizado ao animal, e a consequente variação no consumo de

219
concentrado, como mostrou o trabalho de Campos et al. (1993). Já no sistema de aleitamento
artificial, os animais recebem volumes conhecidos e controlados de dieta líquida através de
mamadeiras, bibeirões, baldes, containers ou até mesmo aleitadores automáticos. Os
utensílios utilizados para o fornecimento são igualmente eficientes e resultam em mesmo
desempenho animal (Otterby & Linn, 1981), desde que a higiene dos mesmos seja adequada.
Caso isso não ocorra, maior frequência de diarreia e redução no desempenho pode ocorrer,
principalmente com o uso de utensílios com bicos. Enquanto o balde tem a vantagem de ser
de simples higienização, reduzindo os problemas com diarreias, tem a necessidade de
treinamento dos animais. Já as mamadeiras, não exigem treinamento, atendem parte da
necessidade comportamental de mamar e resultam em maior secreção de saliva e enzimas
digestivas. No entanto, estão normalmente associadas a maior ocorrência de diarreias devido
a resíduos da dieta líquida. Outra desvantagem das mamadeiras é maior tempo para
alimentação. No entanto, como frisado por Davis & Drackley (1998) as mamadeiras com
capacidade para 2 L fazem com que o produtor muitas vezes entenda que este volume é
suficiente para os animais, independentemente do sistema de aleitamento.
Independentemente do tipo de utensílio utilizado para o aleitamento, fornecer dieta líquida
para um grande número de animais é uma tarefa laboriosa e demorada. O aleitador
automático, que pode ou não ter o fornecimento de concentrado acoplado, é provido de um
único bico, sendo a dieta liberada após a identificação do bezerro por sistema eletrônico. O
uso do aleitador automático permite que animais em programas de alimentação intensiva
realizem um maior número de refeições, reproduzindo o comportamento natural de
alimentação como mostrou o trabalho de Jensen (2009). No caso do uso de baldes para
aleitamento intensivo, o maior número de alimentações aumenta o custo com mão de obra.
O aleitamento coletivo, sem alimentador automático, mas com contêineres com bicos de
mamadeira também tem sido empregado por alguns produtores, sendo mais indicado quando
o fornecimento é à vontade. Embora este sistema reduza de forma marcante o tempo gasto
com o aleitamento, reduzindo custos com mão de obra, requer atenção especial para a
formação de lotes homogêneos do ponto de vista de peso e tamanho dos animais, reduzindo
problemas de dominância. Da mesma forma, é importante que o número de bezerros no lote
seja menor que o número de bicos disponíveis, reduzindo problemas de dominância.

220
A temperatura da dieta líquida a ser fornecido deve ser próxima à temperatura corporal do
animal, sendo isso ainda mais importante para regiões de clima frio. Quando a dieta é
fornecida fria pode haver recusa e também menor desempenho animal devido a menor
secreção de enzimas. Já a frequência de alimentação deve ser de pelo menos duas vezes ao
dia, considerando-se o volume fornecido. Em situações naturais estes animais realizam várias
mamadas, consumindo até 12 L por dia.
O leite é um dos componentes que mais onera o custo de criação de bezerras leiteiras. Dessa
forma, o desaleitamento precoce ou a adoção de dieta líquida de menor custo, pode reduzir o
custo final da novilha de reposição. No Brasil, a maior parte das fazendas fornece leite ou
leite descarte, proveniente de vacas com mastite e/ou resíduo de antibióticos e menos de 15%
dos produtores adotam sucedâneos como dieta líquida (Santos & Bittar, 2015).
A oferta de leite descarte nas propriedades tem sido um obstáculo para o fornecimento de
dieta líquida de melhor qualidade para bezerros leiteiros. Muitos produtores não consideram
as taxas aceitáveis de vacas com mastite no rebanho (1,0%, Santos & Fonseca, 2007) e nem
que estes animais reduzem sua produção em torno de 10%, pelo fato de que este leite tem um
destino: o bezerreiro. Assim, transferem o problema da sala de ordenha para o bezerreiro
quando fornecem este leite com alta carga bacteriana, resíduo de antibióticos e composição
nutricional variada aos bezerros em aleitamento. Além de perderem com a venda deste leite
e com a queda na produção das vacas, tem perdas econômicas relacionadas ao menor
desempenho dos bezerros, normalmente devido ao aumento na ocorrência de diarreias, e
consequentemente com maior gasto com medicamentos e tempo de mão de obra para o
atendimento de animais doentes.
Entre as vantagens do uso de sucedâneos no aleitamento de bezerros estão, além da
economia, devido ao menor preço quando comparado com o leite integral, a possibilidade de
aumento na quantidade de leite a ser comercializada pelo produtor, o fornecimento de dieta
líquida com composição sempre constante e a independência do aleitamento com relação aos
horários de ordenha. Entretanto, a qualidade do sucedâneo, principalmente a fonte proteica,
é o fator determinante para a obtenção de resultados semelhantes aos observados com o
fornecimento de leite integral (NCR, 2001).
Durante as primeiras semanas de vida, o fornecimento de dieta líquida de qualidade é
essencial para garantir desempenho satisfatório dos animais. Durante a fase de aleitamento,

221
70% do custo total com alimentação e manejo estão relacionados ao fornecimento de leite
(Bittar, 2007), o que faz com que produtores busquem alternativas de dieta líquida de menor
custo. De acordo com Heinrichs et al. (1995), diversos sistemas e práticas de manejo de
animais em aleitamento vêm sendo adotados com o objetivo de reduzir o custo com a criação
destes animais. A decisão no uso de sucedâneo deve se basear em seu custo por litro diluído,
comparado ao preço do leite vendido à indústria, mas principalmente em sua composição.
O mercado tem uma grande variedade de produtos disponíveis para o aleitamento de
bezerros, sendo importante o entendimento de que estes produtos não são igualmente
recomendados para bezerros de todas as idades e sistemas de aleitamento. Uma vez que
bezerros jovens não tem aparato enzimático para digestão de fontes de proteína ou
carboidrato de origem vegetal até por volta da terceira semana de vida, é importante que o
sucedâneo tenha basicamente ingredientes de origem láctea. Os teores de proteína vão variar
conforme o sistema de aleitamento (de 20 até 28% na MS), enquanto os teores de gordura
são menos variáveis (entre 16 e 18% na MS). Muito embora as fontes lácteas sejam mais
recomendadas, proteína isolada de soja pode também ser utilizada com algum sucesso. A
quantidade de fibra de uma formulação é um bom indicativo da inclusão de fontes de proteína
de origem vegetal. Quanto maior a inclusão destas fontes, maior será o teor de fibra da
fórmula. Os sucedâneos para bezerros com menos de 3 semanas de idade não devem
apresentar mais que 0,15% de fibra bruta em sua composição. Embora os altos teores de fibra
bruta indiquem a inclusão de proteína de origem vegetal, valores menores que 0,15% não
garantem sua ausência. Isso ocorre devido às tecnologias para retirada de carboidratos
solúveis e também fibras da proteína da soja, por exemplo. Outro aspecto interessante com o
fornecimento de sucedâneos é a possibilidade de inclusão de aditivos que tenham ação
profilática contra diarreias, como os probióticos, prebióticos e acidificantes, resultando em
melhor aproveitamento da dieta líquida.
Em relação ao volume de dieta fornecida, podemos ter dois sistemas de aleitamento:
convencional e intensivo. O sistema de aleitamento convencional consiste no fornecimento
de dieta líquida no volume de 10% do peso vivo (PV) da bezerra (Jasper & Weary, 2002), o
que normalmente representa 4 L diários. Quando a dieta líquida é o sucedâneo, este deve
conter 20-22% de PB e 15-20% de gordura, sendo reconstituído a 12,5% de sólidos (Cowles
et al., 2006). Este sistema tem como objetivo estimular o consumo de concentrado para

222
favorecer o desenvolvimento das papilas do rúmen permitindo o desaleitamento precoce,
reduzir o risco de enfermidades e os gastos de alimentação e manejo. Entretanto, esta
quantidade padronizada de dieta líquida fornecida aos animais geralmente atende pouco mais
que as exigências de mantença, inviabilizando altas taxas de crescimento (Flower & Weary,
2001). Ainda assim, este sistema de aleitamento é adequado para alguns sistemas de
produção, com taxas de crescimento em torno de 400 g/d, sendo o mais utilizado no Brasil
(Santos & Bittar, 2015). Já o aleitamento intensivo tenta imitar o comportamento alimentar
natural do bezerro, preconizando o fornecimento de dieta líquida em volumes acima de 20%
do PV do bezerro, normalmente representando 8 L/d. No entanto, o número de refeições nem
sempre é maior do que dois, o que não representa o comportamento de mamada dos animais
quando estão com suas mães. Este tipo de sistema em geral utiliza sucedâneos com um
conteúdo de proteína bruta igual ou superior a 25%, gordura em quantidades semelhantes ao
sistema convencional (15-20%), e sólidos totais entre 12,5-17,5% (Cowles et al., 2006).
Aumentando as quantidades de dieta líquida fornecidas para os bezerros, aumenta-se a taxa
de crescimento (Jasper & Weary, 2002; Borderas et al., 2009), e o potencial de produção
futura destes animais (Soberon et al., 2012). Dentro do conceito de aleitamento intensivo
podemos aleitar bezerros de três diferentes formas. No sistema ad libitum os animais tem
acesso ilimitado a dieta líquida, a qual deverá estar acidificada ou ser fornecida através de
aleitadores que preparem o sucedâneo no momento da manada. Neste sistema, embora haja
um programa de alimentação fixando volumes máximos liberados em um determinado
período de tempo, o animal decide em que momento deseja mamar, fazendo com que o
aleitamento seja mais próximo do que ocorreria naturalmente. No sistema intensivo
propriamente, o animal recebe entre 15 e 20% do seu PV em dieta líquida, em duas ou mais
refeições. Já no sistema programado (step-up/step-down), o volume de dieta líquida e
variável, sendo normalmente reduzido (step-down) no final do período de aleitamento como
estratégia para o aumento no consumo de concentrado. Embora o custo de produção possa
ser reduzido com o fornecimento de menor quantidade de dieta líquida, volumes inferiores a
4 L não fornecem nutrientes suficientes para desempenho adequado devido ao baixo
consumo de energia e proteína (Tabela 2).
Além de resultar em maiores taxas de crescimento e, portanto, bezerras mais pesadas ao
desaleitamento, o fornecimento de maiores volumes de dieta líquida pode aumentar o

223
potencial de produção de leite futuro destes animais. Vários estudos mostram que o potencial
de produção de leite pode estar relacionado a efeitos do consumo controlado ou alimentação
ad libitum do nascimento até os 42 ou 56 dias de vida (Foldager & Krohn, 1994; BarPeled et
al., 1997; Foldager et al., 1997).
Tabela 2. Potencial de ganho de acordo à quantidade de leite fornecida e ao consumo
de energia metabólica e proteína bruta, baseado em equações do NRC (2001)
Leite
fornecido
(L/dia)
Consumo
MS (g)
Consumo
de EM
(Mcal)
Ganho
permitido
pela EM (g)
Consumo
de PB (g)
Ganho
permitido
pela PB (g)
2 250 1,34 -- 65,5 139
4 500 2,68 354 127 380
6 750 4,03 756 190 627
8 1000 5,37 1050 254 868 MS: matéria seca, EM: energia metabólica, PB: proteína bruta
De acordo com estes estudos, o aumento no consumo de nutrientes antes dos 56 dias de vida
resultou em aumento da produção de leite durante a primeira lactação, que variaram de 450
a 1400 kg a mais, quando comparados com bezerras alimentadas com dieta mais restrita
durante o mesmo período (Tabela 3). O estudo de Soberon et al. (2012) mostra a consistência
do efeito do fornecimento de maiores volumes de dieta líquida no potencial de produção de
leite futuro dos animais. Avaliando quase 1900 dados de um rebanho comercial e do rebanho
da Universidade de Cornell, estes autores mostraram que para cada 1 kg de ganho de peso
diário a mais, houve aumento de 970 kg de produção na primeira lactação. No entanto, estes
resultados foram observados em animais aleitados com sucedâneos contendo 28% de PB e
15 ou 20% de gordura. Os dados corroboram as sugestões de Van Amburgh & Drackley
(2005) para alterações nas recomendações para a nova edição do NRC, com maiores
exigências de proteína para animais em maiores taxas de crescimento.
Da mesma forma que ocorre para animais pré-púberes, quando em crescimento acelerado,
bezerras leiteiras devem receber dieta com maior relação proteína:energia de forma que a
composição do ganho não seja afetada de forma negativa (Blome et al., 2003). Assim,
sucedâneos com maiores teores de PB devem ser fornecidos em sistemas de aleitamento
intensivo para que o potencial de produção de leite destes animais seja aumentado. Soberon
et al. (2012) concluem que a taxa de crescimento de bezerras responde por 22% da variação

224
na produção de leite na 1ª lactação. No entanto, Van Amburgh e Drackley (2005) já haviam
sugerido que 20% da variação na produção de leite na primeira lactação poderiam ser
explicados pela taxa de crescimento até ao desaleitamento. De acordo com estes autores, as
bezerras devem duplicar o seu peso ao nascer ou crescer a uma taxa que lhes permita duplicar
o seu peso ao nascer até o desaleitamento. Assim, vários resultados de pesquisa comprovam
que o aleitamento intensivo pode ter efeitos em longo prazo sobre o desempenho dos animais,
podendo ser uma boa estratégia de alimentação e grande oportunidade de aumentar o
potencial de produção dos animais, contrapondo o sistema de desaleitamento precoce
tradicionalmente utilizado.
Tabela 3. Produção de leite de vacas com consumo de nutrientes 50% superior que
o recomendado durante o período de aleitamento
Estudo Diferença em relação ao
controle (kg)
Foldager & Krohn, 1991 1405s
Foldager et al., 1997 519t
BarPeled et al., 1998 453t
Ballard et al., 2005 700s
Drackley et al., 2007 835s
Raeth-Knight et al., 2009 718ns
Terre et al., 2009 624ns
Morrison et al., 2009 0ns
Rincker et al., 2011 416ns
Soberon et al., 2012 552s s Significativo; t tendência; ns não significativo
Por outro lado, pesquisas mostram que o aleitamento intensivo pode reduzir o consumo de
concentrado, e consequentemente retardar o desenvolvimento ruminal, uma vez que o
consumo de concentrado está negativamente relacionado ao volume de leite fornecido. O
desaleitamento no momento em que animal apresenta o rúmen parcialmente desenvolvido é
essencial para que o desempenho após o desaleitamento não seja prejudicado.
Um dos objetivos na fase de aleitamento é estimular o consumo de concentrado. Durante esta
fase o animal desenvolverá o sistema de digestão próprio de ruminantes e, ao final, deverá

225
estar apto a sobreviver e crescer apenas se alimentando de dieta sólida composta de
concentrado e volumoso. A fase de transição de pré-ruminante para ruminante está
relacionada ao desenvolvimento do rúmen, onde se estabelecerão bactérias amilolíticas num
primeiro momento, e depois celulolíticas e metanogênicas. Além do estabelecimento de
bactérias, que cumprirão o papel de fermentadores, o animal deve ter estruturas capazes de
absorver e metabolizar os produtos finais dessa fermentação, ou seja, um rúmen funcional,
com papilas desenvolvidas. Na fase pré-ruminante, a dieta é basicamente líquida, o principal
órgão digestivo é o abomaso, a fonte de energia é principalmente glicose e a proteica
totalmente proveniente da dieta. No ruminante, a dieta está na forma sólida, as fontes de
energia utilizadas pelo animal são os ácidos graxos de cadeia curta (AGCC) e glicose
proveniente de digestão intestinal, e a fonte proteica é composta de proteína microbiana e
proteína sobrepassante. Durante a fase de transição estas duas situações se misturam e o
manejo alimentar será determinante de uma transição mais lenta ou mais precoce.
A colonização microbiana, disponibilidade de água de bebida, além do desenvolvimento da
capacidade absortiva são fatores importantes e sinais de ocorrência de desenvolvimento
ruminal. Entretanto, o fator determinante para que isto ocorra é o consumo de alimento sólido
o mais cedo possível. Devido à alta taxa de fermentação de grãos, com grande quantidade de
carboidratos fermentáveis até ácidos propiônico e butírico, estes são os principais promotores
do desenvolvimento ruminal precoce. Por outro lado, diversos trabalhos mostram que
carboidratos estruturais originados de forragens são fermentados até ácido acético,
contribuindo pouco para o desenvolvimento de papilas ruminais.
O crescimento normal e desenvolvimento do trato digestório do ruminante podem ser
alterados pelo consumo de alimentos concentrados ou volumosos, níveis de inclusão, sua
forma física, entre outros fatores (Tabela 4). Porém, a composição química e os produtos
finais resultantes da fermentação é que tem mostrado maior influência no desenvolvimento
do epitélio ruminal (Nocek et al., 1984; Khan et al., 2016). Assim, o concentrado será
fornecido desde os primeiros dias de vida e deverá ser formulado com teores de FDN que
permitam manutenção de pH adequados.
Segundo o NRC (2001), o concentrado deve apresentar teores aproximados de 18% de
proteína bruta (PB) na matéria original, 80% de nutrientes digestíveis totais (NDT), e níveis
de FDA entre 6 e 20% e FDN entre 15 e 25%.

226
O teor de proteína no concentrado de bezerros leiteiros recomendado pelo NRC vem sendo
questionado por pesquisadores. Segundo Drackley (2003), bezerros consumindo concentrado
com 22% de PB apresentaram maior eficiência. No entanto, vários trabalhos mostram que o
teor de 18% de PB no concentrado é adequado para bezerros em aleitamento (Luchini et al.,
1991; Akayezu et al., 1994; Hill et al., 2001). Desta forma, tem sido sugerido que
concentrados para bezerros contenham 18% de PB na matéria original, utilizando-se de
preferência farelo de soja como fonte principal.
Tabela 4. Efeito de alimentos concentrados ou de forragem em parâmetros de desenvolvimento
ruminal (adaptado de Khan et al., 2016)
Parâmetro Concentrado Forragem
Peso do rúmen + ++
Volume do rúmen + ++
Diferenciação/crescimento de papilas ++ +
Cetogênese/concentração de BHB + +
Motilidade ruminal/taxa de passagem + ++
Microrganismos ruminais
Bactérias Amilolíticos Celulolíticos
Protozoários - +
Ácidos orgânicos (lactato, C2, C3, C4) ++ +
Acetato:propionato - +
Butirato ++ +
Lactato + -
pH ruminal - +
Capacidade tampão/ruminação - +
Saúde ruminal/paraqueratose - +
Os teores da fração fibra recomendados pelo NRC (2001) devem ser respeitados. Valores
superiores às recomendações indicam a inclusão de ingredientes de menor digestibilidade
para animais com o rúmen em desenvolvimento. Por outro lado, teores de FDN ou FDA
abaixo da recomendação podem resultar em problemas como acidose ruminal e
paraqueratose.
A forma física do concentrado inicial pode afetar o consumo, sendo de interesse econômico
o fornecimento de concentrados sob formas físicas que estimulem o consumo precocemente.
Segundo Coverdale et al. (2004), o tamanho de partícula da ração também pode afetar o
ambiente ruminal, a produção de AGCC, além da estrutura e a função das papilas ruminais.
Rações finamente moídas reduzem o pH ruminal, principalmente devido a menor ruminação

227
e menor fluxo de saliva (Santini et al., 1983), reduzindo consequentemente a população de
bactérias celulolíticas (Beharka et al., 1998). Assim, Warner et al. (1973) sugerem que pelo
menos 50% das partículas que compõem o concentrado inicial sejam maiores que 1,19 cm.
Os maiores efeitos do tamanho de partícula no desempenho animal são observados no
consumo de concentrado, no ganho de peso e no desenvolvimento ruminal. Alguns trabalhos
recentes avaliaram o efeito da forma física do concentrado inicial e não foram observadas
diferenças no consumo ou no ganho de peso quando compararam concentrados peletizados
com farelados para bezerros em aleitamento (Franklin et al., 2003; Ziegler et al., 2006; Bittar
et al., 2009).
Independentemente do fornecimento de leite, o animal deve receber água de boa qualidade
já na primeira semana de vida. O fornecimento de água no mesmo horário do fornecimento
do leite pode levar ao consumo descontrolado de água, afetando a formação do coágulo no
abomaso. A disponibilidade de água está diretamente relacionada ao consumo de
concentrado e à recuperação de quadros de diarreia. A falta de acesso à água pode levar a
reduções no consumo de concentrado e ganho de peso da ordem de 30% (Jenny et al., 1978).
Desaleitamento
O desaleitamento precoce é uma ferramenta de manejo muito importante do ponto de vista
econômico na produção de fêmeas de reposição. O custo de alimentação dos animais é
reduzido com a interrupção no fornecimento da dieta líquida, a manutenção de misturas
concentradas como dieta principal e a introdução de volumosos. Adicionalmente, o menor
tempo demandado para a alimentação desses animais reduz o custo com mão-de-obra.
Segundo Quigley (1996), o animal está pronto, do ponto de vista fisiológico, quando atinge
o consumo de 700 g/d de concentrado durante três dias consecutivos. Produtores tem
utilizado três diferentes critérios, ou uma combinação destes para desaleitamento: 1)
consumo de concentrado; 2) idade do animal; e/ou 3) peso do animal. Tradicionalmente o
desaleitamento vinha sendo realizado aos 60 dias, idade que os animais alcançam consumo
adequado para que o desaleitamento possa ser realizado sem prejuízos no ganho de peso.
Entretanto, este consumo é alcançado por animais em aleitamento convencional nesta idade.
No caso de aleitamento intensivo, menor consumo será observado de acordo com o volume
de dieta líquida fornecida. Assim, a idade não é o critério mais adequado quando utilizado

228
sem considerar o consumo. Por outro lado, uma vez que existem diferenças de peso ao nascer,
utilizar um consumo fixo como critério para o desaleitamento implica em bezerros mais leves
devendo ter um consumo maior em porcentagem de peso vivo. Assim, Greenwood et al.
(1997) sugerem que o consumo adequado para o aleitamento deva ser de 1,5% do peso ao
nascer dos animais. Dessa forma, o ideal é que o produtor utilize uma combinação de critérios
e realize o desaleitamento quando o animal apresentar maturidade anatômica e metabólica,
mantendo suas taxas de ganho de peso na fase subsequente.
A adaptação do animal ao alimento sólido é fundamental para que o desaleitamento ocorra
com sucesso. O desaleitamento é um fator de estresse para o bezerro o qual é forçado a várias
mudanças: 1) sua principal fonte de nutrientes muda da forma liquida para a forma sólida; 2)
a quantidade de matéria seca que o animal recebe é diminuída com o não fornecimento do
leite; 3) o bezerro deve se adaptar ao tipo de digestão e fermentação própria de ruminantes;
4) mudanças de manejo e instalações geralmente ocorrem juntamente com o desaleitamento
(Quigley, 1996). A adaptação do animal à fermentação é essencial para que a taxa de
crescimento do animal não seja afetada. Para isso, o animal deve ter o rúmen parcialmente
desenvolvido e capaz de absorver e metabolizar produtos finais da fermentação antes do
desaleitamento.
Com base em resultados da literatura, diversos autores recomendam o desaleitamento dos
animais de forma abrupta em relação ao sistema de desaleitamento gradual realizado por
alguns produtores (Otterby & Linn, 1981; Davis & Drackley, 1998). O desaleitamento
gradual é dificultado, devido à falta de operacionalidade do processo, principalmente em
grandes rebanhos. Por outro lado, o desaleitamento de forma gradual reduz o estresse dos
animais e ainda estimula o consumo de concentrado.
Instalações e conforto
Durante o período de aleitamento as bezerras são constantemente desafiadas pelo ambiente.
Muitos fatores contribuem para o bem-estar de bezerros em fazendas leiteiras, incluindo:
instalações e ambiente, manejo nutricional e sanitário, manipulação e interação com o
tratador, dinâmica de rebanho, além de práticas comuns como transporte, descorna, remoção
de tetos.

229
Os objetivos gerais das instalações para bezerros são a proteção dos extremos térmicos e
climáticos, acesso adequado ao alimento, garantir a segurança no que diz respeito a
ferimentos e controlar a saúde e bem-estar dos bezerros. Tanto os sistemas de instalação
individual quanto em grupo podem ser projetados para atender a todas estas necessidades.
No entanto, muitos tipos de instalação podem atender todas estas premissas em relação ao
bem-estar, mas o sucesso ainda depende de gestão adequada. Assim, quando se pensa em um
abrigo para melhor alojar bezerras, existem quatro requisitos fundamentais que devem ser
considerados: 1) ventilação; 2) isolamento; 3) conforto; 4) economia.
No mundo todo existem variadas formas de criação de bezerras em aleitamento –criação em
abrigos individuais, baias coletivas ou individuais, construções fechadas ou abertas–
variando de acordo com o local da exploração, o sistema de produção e, principalmente, o
custo para sua construção. Ambientes satisfatórios para bezerros recém-nascidos e em
crescimento devem proporcionar conforto físico, térmico, psicológico e comportamental.
Cada uma dessas áreas pode ser uma fonte de estresse para os bezerros, que posteriormente
podem predispor os animais a comprometimento de sua resposta imunitária, das taxas de
crescimento, e finalmente do bem-estar propriamente dito. Embora o conforto térmico e
físico do ambiente para bezerros tenha sido amplamente avaliado, apenas recomendações
gerais foram desenvolvidas para satisfazer as necessidades comportamentais específicas de
bezerros leiteiros. As necessidades comportamentais em um ambiente incluem a ausência de
frustração, o sentimento de segurança e ausência de possibilidade de lesão, comportamento
social de rebanho e interações com o tratador adequadas.
A individualização tem como objetivo principal a redução na disseminação de doenças, muito
embora existam desvantagens do ponto de vista comportamental. Em um levantamento sobre
os sistemas de criação brasileiros, as diarreias foram apontadas como o principal problema
de saúde de bezerras, seguidas pelos problemas respiratórios (Santos & Bittar, 2015). Embora
estas duas doenças tenham forte relação com falhas no programa de colostragem, estão
também fortemente relacionadas com as instalações e o manejo das mesmas. Como a
transmissão dos principais patógenos que causam doenças em bezerros é do tipo oral-fecal,
seja através do contato entre animais ou uso de utensílios (baldes, cochos) com limpeza
inadequada, a individualização entre os animais é considerada um dos princípios
fundamentais de um bom sistema de criação. A individualização dos animais também facilita

230
a alimentação, evitando problemas com dominância, e permite um controle mais rígido do
consumo individual, tanto de concentrado quanto de água, e da saúde do animal. Em
levantamento nacional, 45% das propriedades cria bezerras leiteiras de forma
individualizada, mas este percentual cresce para 64% quando se avalia somente propriedades
com produção acima de 700 L/d (Santos & Bittar, 2015). Existem diferentes sistemas
individualizados como o sistema tie-stall, o sistema de baias individuais, mas o mais
comumente utilizado são os abrigos. Nestes sistemas, os animais normalmente têm acesso
individualizado a água e ao concentrado em baldes ou cochos, o que permite controle de
consumo. Ainda, o aleitamento é realizado de forma individual sendo utilizados baldes,
mamadeiras ou ainda bibeirões (baldes com bico). Assim, o controle da nutrição é também
individualizado, de forma que se pode avaliar o manejo alimentar de acordo com os ganhos
obtidos. Já a criação de bezerras em sistemas coletivos se baseia no princípio de que os
bezerros leiteiros são animais de rebanho (gregários) e o alojamento em grupo permite o
desenvolvimento de comportamento social. O alojamento coletivo permite a manifestação de
comportamentos lúdicos, ou seja, o exercício e o jogo entre bezerros dentro do grupo. Assim,
a criação de animais em lotes tem sido considerada como mais adequada do ponto de vista
de bem-estar e comportamento animal por alguns pesquisadores. No entanto, é sabido que
este sistema resulta em maior disseminação de doenças, além dos problemas associados à
mamada-cruzada e falta de controle de consumo individual de dieta líquida ou sólida,
dependendo do sistema de alimentação. Os animais podem ser criados em piquetes, em
galpões abertos com pisos ripados ou não e ainda em galpões fechados. Independentemente
do tipo de alojamento, é importante que o ambiente seja ventilado e com áreas de sombra
disponível. Quando os animais são criados em piquetes, a área deve ser bem drenada
impedindo a formação de barro na época das chuvas e a sombra pode ser natural ou artificial.
Uma desvantagem deste tipo de alojamento é a possibilidade de desenvolvimento de mamada
cruzada, associado ou não ao hábito de beber urina, comportamentos considerados
problemáticos. Além de problemas como traumas e inflamações de úbere, podem ocorrer
problemas no umbigo ou na orelha dos animais, regiões com preferência de mamada.
Os sistemas de criação coletiva podem ser manejados para aleitamento também coletivo,
quando se usam containers. Neste sistema é de extrema importância a homogeneidade do
tamanho do lote, de forma a reduzir problemas de competição e variado volume de dieta

231
líquida consumida. A grande desvantagem desse sistema é o fato de que não se tem controle
algum sobre o consumo de dieta líquida ou sólida, parâmetros importantes para a tomada de
decisão do desaleitamento, por exemplo. Este problema pode ser resolvido com sistemas de
aleitamento individual para animais criados em lotes, com a utilização de containers com
divisões internas, que separe o volume de dieta líquida para cada animal. A adoção de canzil
para contenção dos animais no horário de alimentação também é uma alternativa. Este
sistema tem a vantagem de permitir que os animais sejam mantidos por um período de tempo
após o consumo da dieta líquida, até que percam o estímulo da mamada, reduzindo a
ocorrência de mamada cruzada. Já com uso de aleitador automático existe a possibilidade
fornecimento de dieta líquida ad libitum ou com quantidade programada e controlada por
computador de acordo com a idade do animal e manejo nutricional. Embora muitos trabalhos
venham mostrando benefício deste tipo de alimentação por ser semelhante à maneira como
o animal se alimentaria normalmente, estes sistemas podem ter várias desvantagens.
Diarreias e doenças respiratórias podem se espalhar mais rapidamente quando este tipo de
alojamento e de alimentação é adotado.
Assim como nos sistemas individualizados, nos sistemas coletivos o treinamento do tratador
é decisivo para o sucesso da criação de bezerros, com altas taxas de crescimento e baixas
taxas de morbidade e mortalidade. Ainda mais importante que nos sistemas individualizados,
tratadores com atitudes positivas trazem grandes benefícios ao sistema de criação. A
manifestação de comportamentos lúdicos depende do atendimento de necessidades básicas
como alimentação adequada, acesso a sombra e água, conforto e sensação de segurança.
Animais assistidos por tratadores positivos são menos reativos e mais ativos na expressão de
comportamentos lúdicos, o que acaba refletindo também no melhor desempenho e menor
frequência de enfermidades (Schuetz et al., 2012). Abordar e manipular o animal para
práticas de manejo como pesagem, aplicação de vacinas, diagnóstico de doenças ou uma
simples medida de temperatura é mais difícil em sistemas coletivos onde o animal está solto
e misturado a outros animais. Métodos rápidos para diagnóstico de doenças têm sido
estudados no que se refere a alterações no comportamento ou no consumo da dieta líquida.

232
Considerações finais
A criação de bezerras é ainda um dos gargalos nos sistemas de produção de leite,
principalmente devido às taxas de mortalidade e o impacto que estas têm nas planilhas de
custo. Além disso, reduzidas taxas de crescimento devido a falhas na colostragem, associada
a manejo alimentar e alojamento inadequados também trazem prejuízos ao sistema de
produção. O manejo alimentar é determinante do desempenho dos animais durante a fase de
crescimento e pode ainda afetar a produção futura de bezerras leiteiras, sendo uma boa
oportunidade de aumentar a produtividade do rebanho.
Referências
Akayezu, J. M., J. G. Linn, D. E. Otterby, W. P. Hansen, and D. G. Johnson. 1994. Evaluation of calf starters
containing different amounts of crude protein for growth of holstein calves. Journal of Dairy Science 77: 1882-
1889.
Ballard, C. et al. 2005. The effect of feeding three milk replacer regimens preweaning on first lactation
performance of Holstein dairy cattle. Journal of Dairy Science 88: 22-22.
BarPeled, U. et al. 1997. Increased weight gain and effects on production parameters of Holstein heifer calves
that were allowed to suckle from birth to six weeks of age. Journal of Dairy Science 80: 2523-2528.
Beharka, A. A., T. G. Nagaraja, J. L. Morrill, G. A. Kennedy, and R. D. Klemm. 1998. Effects of form of the
diet on anatomical, microbial, and fermentative development of the rumen of neonatal calves. Journal of Dairy
Science 81: 1946-1955.
Besser, T. E., and C. C. Gay. 1994. The importance of colostrum to the health of the neonatal calf. Veterinary
Clinics of North America-Food Animal Practice 10: 107-117.
Bittar, C. M. M. Aleitar uma ou duas vezes ao dia? DBO Mundo do Leite, p. 14 - 16, 01 jul. 2007.
Bittar, C. M. M., L. S. Ferreira, F. A. P. Santos, and M. Zopollatto. 2009. Performance and ruminal development
of dairy calves fed starter concentrate with different physical forms. Revista Brasileira De Zootecnia-Brazilian
Journal of Animal Science 38: 1561-1567.
Blome, R. M., J. K. Drackley, F. K. McKeith, M. F. Hutjens, and G. C. McCoy. 2003. Growth, nutrient
utilization, and body composition of dairy calves fed milk replacers containing different amounts of protein.
Journal of Animal Science 81: 1641-1655.
Borderas, T. F., A. M. B. de Passille, and J. Rushen. 2009. Feeding behavior of calves fed small or large amounts
of milk. Journal of Dairy Science 92: 2843-2852.
Campos, O. F.; Lizieire, R. S.; Deresz, F.; Matos, L. L. Sistemas de aleitamento natural controlado ou artificial.
II Efeitos na performance de bezerros mestiços HZ. Revista Brasileira de Zootecnia, v. 22, n. 3, p. 413-422,
1993.
Coverdale, J. A., H. D. Tyler, J. D. Quigley, and J. A. Brumm. 2004. Effect of various levels of forage and form
of diet on rumen development and growth in calves. Journal of Dairy Science 87: 2554-2562.
Cowles, K. E., R. A. White, N. L. Whitehouse, and P. S. Erickson. 2006. Growth characteristics of calves fed
an intensified milk replacer regimen with additional lactoferrin. Journal of Dairy Science 89: 4835-4845.
Davis, C.L.; Drackley, J. K. The development, nutrition, and management of the young calf. Ames: Iowa State
University Press, p.339, 1998.

233
Deelen, S. M., T. L. Ollivett, D. M. Haines, and K. E. Leslie. 2014. Evaluation of a Brix refractometer to
estimate serum immunoglobulin G concentration in neonatal dairy calves. Journal of Dairy Science 97: 3838-
3844.
Dos Santos, G., and C. M. M. Bittar. 2015. A survey of dairy calf management practices in some producing
regions in Brazil. Revista Brasileira De Zootecnia-Brazilian Journal of Animal Science 44: 361-370.
Drackley, J. K. 2003. Nutritional strategies to promote calf health during the liquid feeding period.
Drackley, J. K., B. C. Pollard, H. M. Dann, and J. A. Stamey. 2007. First-lactation milk production for cows
fed control or intensified milk replacer programs as calves. Journal of Animal Science 85: 614-614.
Elizondo-Salazar, J. A., B. M. Jayarao, and A. J. Heinrichs. 2010. Effect of heat treatment of bovine colostrum
on bacterial counts, viscosity, and immunoglobulin G concentration. Journal of Dairy Science 93: 961-967.
Flower, F. C., and D. M. Weary. 2001. Effects of early separation on the dairy cow and calf: 2. Separation at 1
day and 2 weeks after birth. Applied Animal Behaviour Science 70: 275-284.
Foldager, J., Krohn, C.C. Level of milk for female calves affects their milk production in first lactation. Proc.
European Assoc. Animal Prod. 48th Annual Meeting, 1997.
Foldager, J.; Krohn C.C. Heifer calves reared on very high or normal levels of whole milk from birth to 6-8
weeks of age and their subsequent milk production. Proc. Soc. Nutr. Physiol. v.3, 1994.
Foley, J. A., and D. E. Otterby. 1978. Availability, storage, treatment, composition, and feeding value of surplus
colostrum - review. Journal of Dairy Science 61: 1033-1060.
Franklin, S. T., D. M. Amaral-Phillips, J. A. Jackson, and A. A. Campbell. 2003. Health and performance of
Holstein calves that suckled or were hand-fed colostrum and were fed one of three physical forms of starter.
Journal of Dairy Science 86: 2145-2153.
Godden, S. 2008. Colostrum management for dairy calves. Veterinary Clinics of North America-Food Animal
Practice 24: 19-+.
Greenwood, R. H., J. L. Morrill, and E. C. Titgemeyer. 1997. Using dry feed intake as a percentage of initial
body weight as a weaning criterion. Journal of Dairy Science 80: 2542-2546.
Heinrichs, A. J., S. J. Wells, and W. C. Losinger. 1995. Study of the use of milk replacers for dairy calves in
the United States. Journal of Dairy Science 78: 2831-2837.
Hill, T. M.; Aldrich, J. M.; Proeschel, A. J.; Schlotterbeck, R. L. Protein levels for neonatal calf starters. J.
Dairy Sci., v.84 (Suppl. 1), p.265, 2001.
Jasper, J., and D. M. Weary. 2002. Effects of ad libitum milk intake on dairy calves. Journal of Dairy Science
85: 3054-3058.
Jenny, B. F., S. E. Mills, W. E. Johnston, and G. D. Odell. 1978. Effect of fluid intake and dry-matter
concentration on scours and water-intake in calves fed once daily. Journal of Dairy Science 61: 765-770.
Jensen, M. B. 2009. Milk meal pattern of dairy calves is affected by computer-controlled milk feeder set-up.
Journal of Dairy Science 92: 2906-2910.
Khan, M. A., A. Bach, D. M. Weary, and M. A. G. von Keyserlingk. 2016. Invited review: Transitioning from
milk to solid feed in dairy heifers. Journal of Dairy Science 99: 885-902.
Luchini, N. D., S. F. Lane, and D. K. Combs. 1991. Evaluation of starter diet crude protein level and feeding
regimen for calves weaned at 26 days of age. Journal of Dairy Science 74: 3949-3955.
Morrison, S. J. et al. 2009. Effects of feeding level and protein content of milk replacer on the performance of
dairy herd replacements. Animal 3: 1570-1579.
National Research Council. Nutrient Requirements of Dairy Cattle. Seventh Ed., Natl. Acad. Sci., Washington,
D. C., 2001.
Nocek, J. E., C. W. Heald, and C. E. Polan. 1984. Influence of ration physical form and nitrogen availability on
ruminal morphology of growing bull calves. Journal of Dairy Science 67: 334-343.

234
Otterby, D. E., and J. G. Linn. 1981. Advances in nutrition and management of calves and heifers. Journal of
Dairy Science 64: 1365-1377.
Quigley III, J.D. Feeding prior to Weaning. In: Calves, heifers and dairy profitability. National Conference,
Pennsylvania, 1996. Proceedings. Ithaca: Northeast Regional Agricultural Engineering Service Cooperative
Extension, p.245-255, 1996.
Quigley, J. Calf age, total protein and FPT in calves. Calf Notes, p.5, 2001.
Quigley, J. D. 1996. Influence of weaning method on growth, intake, and selected blood metabolites in Jersey
calves. Journal of Dairy Science 79: 2255-2260.
Quigley, J. D., A. Lago, C. Chapman, P. Erickson, and J. Polo. 2013. Evaluation of the Brix refractometer to
estimate immunoglobulin G concentration in bovine colostrum. Journal of Dairy Science 96: 1148-1155.
Raeth-Knight, M. et al. 2009. Impact of conventional or intensive milk replacer programs on Holstein heifer
performance through six months of age and during first lactation. Journal of Dairy Science 92: 799-809.
Rincker, L. E. D. et al. 2011. Effect of intensified feeding of heifer calves on growth, pubertal age, calving age,
milk yield, and economics. Journal of Dairy Science 94: 3554-3567.
Santini, F. J., A. R. Hardie, N. A. Jorgensen, and M. F. Finner. 1983. Proposed use of adjusted intake based on
forage particle length for calculation of roughage indexes. Journal of Dairy Science 66: 811-820.
Santos, M.V.; Fonseca, L.F.L. Estratégias para controle de mastite e melhoria da qualidade do leite. São
Paulo: Manole, 2007. 313p.
Schuetz, K. E. et al. 2012. Effects of human handling during early rearing on the behaviour of dairy calves.
Animal Welfare 21: 19-26.
Soberon, F., E. Raffrenato, R. W. Everett, and M. E. Van Amburgh. 2012. Preweaning milk replacer intake and
effects on long-term productivity of dairy calves. Journal of Dairy Science 95: 783-793.
Stewart, S. et al. 2005. Preventing bacterial contamination and proliferation during the harvest, storage, and
feeding of fresh bovine colostrum. Journal of Dairy Science 88: 2571-2578.
Terre, M., C. Tejero, and A. Bach. 2009. Long-term effects on heifer performance of an enhanced-growth
feeding programme applied during the preweaning period. Journal of Dairy Research 76: 331-339.
Van Amburgh, M.E.; Drackley, J. K. Current perspectives on the energy and protein requirements of the pre-
weaned calf. Chap. 5. In: Calf and heifer rearing: Principles of rearing the modern dairy heifer from calf to
calving. Nottingham Univ. Press. P.C. Garnsworthy. 2005.
Warner, R.G.; Proter, J.C.; Slack, T.S. Calf starter formulation for neonatal calves fed no hay. In: Cornell
Nutrition Conference, 1973, Ithaca. Proceedings... Ithaca: Cornell University, 1973. p.116-122.
Ziegler, B. et al. 2006. Pre- and post weaning performance of dairy heifer calves fed texturized or pelleted calf
starters with or without intake enhancing flavors. Journal of Animal Science 84: 365-365.

235
Alimentação e manejo da novilha leiteira
Julio Viégas
Em boa parte das propriedades leiteiras do Brasil o manejo de novilhas é a atividade que mais
tem comprometido a viabilidade da atividade, contribuindo para o aumento dos custos de
produção e queda na eficiência produtiva. Isso decorre da falta de atenção por parte do
produtor a essa categoria animal tão importante em todo o processo produtivo. A origem da
pouca relevância é devida ao fato de ser uma categoria não produtiva e que, aparentemente,
não traz benefícios diretos ao produtor. Este erro grosseiro compromete a possibilidade do
melhoramento contínuo do rebanho e igualmente a possibilidade de incrementar a produção
leiteira, visto que as novilhas serão os animais de reposição.
Propriedades que não cuidam de maneira adequada de seus animais jovens acabam
apresentando índices zootécnicos, como por exemplo, a idade ao primeiro parto (IPP), muito
aquém do ideal. Como consequência o produtor irá pagar um preço elevado por este descuido
devido ao aumento no tempo de permanência de animais não produtivos no rebanho e
também devido à diminuição do número de animais para a comercialização, visto que nestas
propriedades a mortalidade, morbidade e o descarte involuntário são também muito elevados.
A desatenção inicia, frequentemente, a partir do desaleitamento ficando mais visível a partir
dos seis meses de idade, quando ocorre a interrupção do fornecimento do alimento
concentrado e a novilha passa a ser alimentada preferencialmente com volumosos. Apesar da
alimentação e manejo de novilhas ser uma das atividades mais fáceis de ser realizada,
considerando as demais categorias, é necessário redobrar os cuidados com a velocidade de
crescimento dos animais e avaliar se essa alimentação está de acordo com a raça em criação.
Somente um crescimento adequado irá permitir que as novilhas alcancem aos quinze meses
de idade o peso ideal para a concepção e que aos vinte e quatro meses de idade tenham o seu
primeiro bezerro. Ter como meta destes dois momentos na vida da novilha é sinônimo de
eficiência técnica e de maior rentabilidade na atividade.
A primeira parição aos 24 meses é viável, pois apresenta uma série de vantagens, sendo que
o manejo de novilhas, na maioria das situações, é uma questão de vigilância e bom senso de
técnicos e produtores.

236
Todo o planejamento de gestão e manejo alimentar do rebanho jovem da propriedade leiteira
deve partir do estabelecimento de metas bem claras como a idade pretendida para o primeiro
parto. O produtor deve ter em mente que ao optar por 24, 30 ou 36 meses como idade ao
primeiro parto (IPP) para as suas novilhas estará optando por níveis diferentes de
investimento e, consequentemente, por benefícios distintos.
Na medida em que a propriedade leiteira se aproxima de um índice de 24 meses para a IPP
estará alcançando o máximo de eficiência técnica, sem descuidar, na sequência, que o
intervalo entre partos (IEP) se aproxime de 12 meses. Não existe nenhum impedimento
biológico que determine que uma fêmea não possa parir com 24 meses de idade tendo
alcançado um peso adequado, e que mantenha uma vida produtiva e saudável.
Novilhas que parem mais precocemente também são mais rentáveis para a propriedade, não
somente devido ao número de bezerras produzidas e à produção de leite que aumentam, mas
igualmente pela possibilidade de ganho genético entre gerações. É esperado que com a
diminuição do intervalo entre gerações ocorra um ganho genético expresso por maior
produção de leite e de seus componentes. Se isto não está ocorrendo o programa de
melhoramento genético da propriedade deve ser revisado com urgência. Na medida em que
diminui a IPP o ganho genético será acelerado, desde que a escolha dos touros melhoradores
esteja sendo feita de maneira correta, ao mesmo tempo, esta mudança deverá estar em
consonância com a taxa de descarte e reposição das vacas leiteiras.
São exatamente os rebanhos em que as primíparas parem precocemente os que apresentam a
produção total de leite e de seus componentes mais elevada, independente de considerar
somente o lote de primíparas ou de todo o rebanho (Lefebvre et al., 2002). A lógica é a mesma
em relação aos animais que apresentam maior peso ao primeiro parto. A questão da
longevidade de animais que parem precocemente poderia ser questionada, entretanto, as
primíparas que parem tardiamente são aquelas que apresentam maior risco de serem
descartadas antes do tempo. Para que os objetivos da criação de novilhas sejam atingidos
basta um manejo apropriado e um programa alimentar bem equilibrado para manter uma taxa
de desenvolvimento corporal aceitável.
A redução da IPP acarreta em redução dos custos fixos e variáveis na medida em que as
novilhas passam a produzir leite mais cedo, amortizando mais rapidamente os gastos com as
fases de cria e recria. Uma novilha que venha a parir pela primeira vez com 36 meses de

237
idade permanece mais um ano dentro da propriedade se alimentando sem trazer retorno
econômico imediato. Caso isto ocorra com uma novilha de alto padrão genético, devido a
algum problema sanitário, metabólico ou de manejo, a propriedade até poderá absorver este
custo, entretanto, se esta é a situação real, e a propriedade possui um lote significativo de
animais entre 24 e 36 meses sem produzir, o prejuízo será muito maior.
À medida que ocorre atraso na IPP estão sendo geradas novas categorias de novilhas que não
produzem leite. Isto é antieconômico, pois aumenta os custos de produção e diminui a renda
do produtor. Na medida em que o produtor consegue reduzir a idade ao primeiro parto de
suas novilhas de 36 para 24 meses os custos fixos diminuem em torno de 40%. Assim, é
possível concluir que o ponto mais positivo da redução da IPP é exatamente a redução do
número de novilhas na propriedade. Como consequência mais atenção e espaço é
proporcionado para as demais novilhas permitindo maior aporte de alimento com a finalidade
de manter uma taxa de crescimento adequada, necessária para atingir a meta de peso para a
cobertura aos 15 meses de idade.
Para facilitar a gestão da atividade o produtor rural deve estabelecer, juntamente com o
técnico que lhe dá suporte metas claras e factíveis conforme a saúde financeira da
propriedade. Os indicadores que devem ser visualizados como metas para a categoria de
novilhas são os que seguem: idade a cobertura de 13 a 15 meses; idade ao primeiro parto de
22 a 24 meses; peso vivo após o parto próximo a 550 kg; altura ao parto de 143 cm; escore
de condição corporal ao 1º cio de 3,0 a 3,5; escore de condição corporal ao parto de 3,0 a 3,5;
ganho de peso de 3 a 12 meses de idade, preferencialmente entre 0,750 e 0,850 kg/dia (não
superior a 0,900 kg/d).
Para que se consiga manter as metas previamente estabelecidas é fundamental que os animais
mantem um ritmo de crescimento constante. Evidente que o ritmo de crescimento dos
bovinos jovens é determinado pelo padrão genético da raça e/ou linhagem escolhida, ou seja,
cada animal tem o seu potencial de crescimento que é delimitado pela sua herança genética,
mas que pode ser limitado fortemente pelo ambiente onde se encontra. De nada adianta o
produtor investir em animais de alto mérito genético se o sistema alimentar não atende as
exigências mínimas para mantença e crescimento. Importância similar tem as instalações e o
manejo reprodutivo e sanitário que também influenciam o crescimento das novilhas. A
exploração do potencial genético, a partir do fornecimento de um ambiente adequado que

238
permita a expressão deste potencial, é de fundamental importância para a obtenção de bons
rendimentos na atividade leiteira.
Considerando que a novilha tenha passado por uma fase de cria adequada, recebendo o
colostro em quantidade adequada logo nas primeiras horas de vida, o que lhe confere a
imunidade passiva, e que tenha apresentado crescimento adequado, ou seja, o mais próximo
possível de 0,500 kg/dia, seguramente será obtido um animal saudável para prosseguir a
recria. É fundamental que nos primeiros quatro meses da recria a bezerra ainda continue
recebendo alimento concentrado, a fim de garantir um bom aporte de matéria seca e
consequentemente suportar uma taxa de crescimento aceitável, (0,600 a 0,650 kg/dia) mesmo
após o estresse do desaleitamento.
A produção de leite da futura vaca é função do número de células secretoras que compõem
o parênquima das glândulas mamárias, da taxa de secreção de cada célula secretora e ou
alvéolo (aqui entendido, como a unidade fundamental de secreção) e do aporte de nutrientes
para os alvéolos. Qualquer situação que venha a restringir a máxima expressão de um destes
fatores trará como consequência um impacto negativo na produção, neste sentido o ambiente
tem um papel fundamental. Toda e qualquer condição ambiental que represente uma fonte
de estresse para a novilha irá determinar, igualmente uma redução na máxima expressão de
um dos fatores acima citados.
A literatura é farta em informações relacionadas à necessidade de limitação do ganho médio
diário de peso vivo (GMD) das novilhas até o estabelecimento da puberdade (Van Ambrurgh
et al., 1998; Radcliff et al., 2000). Como a puberdade não ocorre em um momento fixo da
vida da novilha estipulou-se que entre os três e doze meses de idade o GMD deve ser mantido
em no máximo 0,750 kg/dia, momento no qual o tecido mamário apresenta crescimento
alométrico. Esta limitação visa evitar o excesso de deposição de gordura na glândula
mamária, o qual ocorreria em detrimento da adequada formação do tecido secretor de leite,
ocasionando redução na produção de leite ao longo da vida produtiva da futura vaca. Assim,
ganhos superiores a estes, proporcionados por dietas com maior densidade energética, seriam
possíveis somente após a novilha ter atingido a puberdade.
Se o bezerro nascer com um bom peso ao parto (próximo de 40 kg, considerando raças
pesadas) e apresentar um bom GMD (mínimo 0,500 kg/dia) nos primeiros três meses, não há
efetiva necessidade de ganhos superiores a 0,750 kg/dia. A novilha terá capacidade de chegar

239
aos 15 meses de idade pesando 360 kg de PV, estando apta para a reprodução. Por outro lado,
estudos têm demonstrado que ganhos de peso superiores a 0,750 kg/dia não induzem,
necessariamente, a redução na quantidade de tecido parenquimático e consequente menor
produção de leite (Silva et al., 2002a; Silva et al., 2002b). Novilhas que apresentam
crescimento mais rápido em um determinado grupo de animais são aquelas com menor
deposição de gordura corporal e, portanto, menor quantidade de gordura corporal nas
glândulas mamárias. Desta maneira entre distintas novilhas, aquela que apresenta rápido
crescimento e destina a energia para ganho muscular, não apresentando ganho excessivo de
condição corporal, certamente será uma vaca com maior produção leiteira.
Uma boa forma de verificar esta situação é por meio da avaliação da altura em conjunto com
a condição corporal, ou seja, serão animais com menor escore de condição corporal e,
portanto, mais altos. Assim, a novilha atingir os 140 cm de altura se reveste de importância.
Esta velocidade de crescimento certamente está associada a maior produção natural de
somatotropina, com efeito benéfico sobre a repartição de nutrientes e no crescimento do
tecido mamário.
Aparentemente, a leptina tem um papel fundamental no crescimento mamário. A leptina é
um hormônio produzido pelo tecido adiposo, em condições de dietas ricas em energia.
Conforme Silva et al. (2002b), animais obesos, apresentam níveis de leptina mais elevados,
e quando o hormônio foi adicionado em meio de cultura com células epiteliais em
proliferação a síntese de DNA foi reduzida em 25%.
Em várias propriedades no Brasil, mais notadamente na Região Sul é ainda comum observar
a preferência por animais jovens com condição corporal mais elevada. Neste sentido, o
produtor deveria evitar realizar a avaliação de condição corporal do seu próprio rebanho,
devido à tendência de subestimar o escore de seus animais. Observam-se em algumas
propriedades animais jovens atingindo 400 kg de PV já aos 10 meses de idade, o que
configura uma situação de custo de alimentação elevado e prejuízo certo no futuro. Esse peso
é excessivo até mesmo para animais com 15 meses de idade. Entretanto, mesmo que
determinadas novilhas tenham um metabolismo para rápido crescimento e maior deposição
de tecido muscular, muitos outros fatores ambientais podem determinar o acúmulo excessivo
de gordura e a consequente redução na produção de leite da futura vaca. Dietas como as
baseadas em silagem de milho, tipicamente utilizadas, na fase de recria, favorecem maior

240
deposição de gordura e consequentemente limitações de ganho de peso devem ser impostas.
Como ainda não existe alguma forma prática de avaliação do crescimento do tecido mamário,
passível de ser utilizado em nível de propriedade, ganhos de peso mais conservadores ainda
são recomendados, evitando-se ganhos próximos ou acima de 0,900 kg/dia.
Neste sentido a Figura 1 ilustra bem a relação entre GMD de novilhas leiteiras e a produção
estimada de leite na primeira lactação. A máxima produção é obtida quando o GMD está
próximo de 0,800 kg/dia, com uma amplitude aceitável de GMD entre 0,700 e 0,900 kg/dia.
Importante verificar que não somente ganhos elevados, como ganhos reduzidos acabam por
promover perdas na produção de leite da futura vaca.
Figura 1. Relação entre ganho de peso vivo em novilhas pré-puberes e a
produção média de leite na primeira lactação, estudo de meta-análise (Fonte:
Zanton & Heinrichs, 2005)
Na Figura 2 são apresentadas diferentes simulações do desenvolvimento ponderal de
novilhas leiteiras. A novilha um é a que está mais próxima do “ideal” para o desenvolvimento
de um bom animal leiteiro de raças de grande porte, com a cobertura sendo realizada aos 15
meses (357 kg de peso vivo) e o parto ocorrendo aos 24 meses de idade (500 kg de peso
vivo). Esta novilha nasceu com 45 kg de peso vivo e apresentou os seguintes ganhos de peso
conforme cada etapa de vida: 0,450 kg/dia do nascimento ao desaleitamento; 0,650 kg/dia do
desaleitamento aos 6 meses de vida; 0,750 kg/dia dos 6 aos 12 meses de vida; 0,750 kg/dia
dos 12 aos 15 meses (cobertura); 0,522 kg/dia desde a cobertura até os 24 meses (parto).
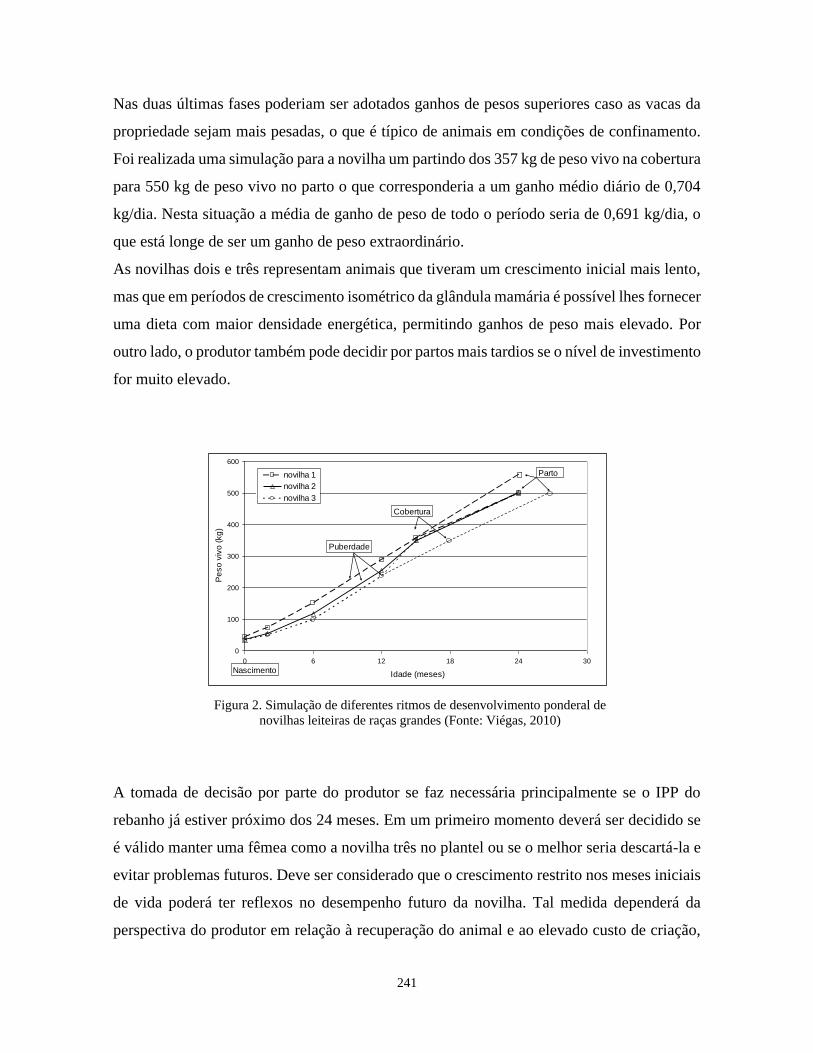
241
Nas duas últimas fases poderiam ser adotados ganhos de pesos superiores caso as vacas da
propriedade sejam mais pesadas, o que é típico de animais em condições de confinamento.
Foi realizada uma simulação para a novilha um partindo dos 357 kg de peso vivo na cobertura
para 550 kg de peso vivo no parto o que corresponderia a um ganho médio diário de 0,704
kg/dia. Nesta situação a média de ganho de peso de todo o período seria de 0,691 kg/dia, o
que está longe de ser um ganho de peso extraordinário.
As novilhas dois e três representam animais que tiveram um crescimento inicial mais lento,
mas que em períodos de crescimento isométrico da glândula mamária é possível lhes fornecer
uma dieta com maior densidade energética, permitindo ganhos de peso mais elevado. Por
outro lado, o produtor também pode decidir por partos mais tardios se o nível de investimento
for muito elevado.
Figura 2. Simulação de diferentes ritmos de desenvolvimento ponderal de
novilhas leiteiras de raças grandes (Fonte: Viégas, 2010)
A tomada de decisão por parte do produtor se faz necessária principalmente se o IPP do
rebanho já estiver próximo dos 24 meses. Em um primeiro momento deverá ser decidido se
é válido manter uma fêmea como a novilha três no plantel ou se o melhor seria descartá-la e
evitar problemas futuros. Deve ser considerado que o crescimento restrito nos meses iniciais
de vida poderá ter reflexos no desempenho futuro da novilha. Tal medida dependerá da
perspectiva do produtor em relação à recuperação do animal e ao elevado custo de criação,
0
100
200
300
400
500
600
0 6 12 18 24 30
Idade (meses)
Pe
so
viv
o (
kg
)
novilha 1
novilha 2
novilha 3
Parto
Cobertura
Puberdade
Nascimento

242
já que será necessário maior aporte de proteína e energia, ou seja, de alimentos de alta
qualidade. Em um segundo momento, sendo a decisão de manter a novilha, deve ser definida
a taxa de crescimento futuro, ou seja, entre um ganho de mais de 1,0 kg/dia com elevado
custo ou um ganho de peso mais modesto, mesmo comprometendo a IPP. O problema de
ganhos de peso muito elevados e repentinos após a cobertura, apesar de plenamente possíveis,
é a probabilidade de um acúmulo importante de gordura no aparelho reprodutivo da novilha
levando a complicações no momento do parto.
Conforme a Figura 3 é possível observar que taxas de crescimento muito baixas levam ao
atraso da maturidade sexual, ou seja, a puberdade é alcançada acima dos 18 meses quando as
novilhas já deveriam estar gestando. Consequentemente, um crescimento lento afeta não
somente a idade à puberdade, mas também a IPP. Ao contrário, novilhas que apresentam
crescimento muito acelerado poderão atingir a puberdade antes dos nove meses de idade, o
que também não é adequado.
Figura 3. Taxa de crescimento de novilhas e performance reprodutiva (Fonte: Wattiaux, 2004)
O limite mínimo de 13 meses de idade deve ser estabelecido para a realização da primeira
cobertura, o que está próximo da idade normalmente recomendada para raças de pequeno
porte, as quais são mais precoces. Desta forma preservamos a integridade da novilha e
conferimos um bom crescimento, sem comprometer a gestação. Novilhas que ganham muito

243
peso sem crescerem rapidamente além de apresentarem maior deposição de gordura na
glândula mamária certamente apresentarão menor porte. Isto por que, novilhas que atingem
a puberdade muito cedo (antes dos nove meses) e são cobertas somente próximo aos 15 meses
de idade passarão por vários ciclos estrais sofrendo por longo tempo o efeito dos hormônios
da reprodução, o que impede um crescimento mais adequado. Com altos níveis nutricionais,
principalmente energia, a puberdade é alcançada com peso vivo menor do que com um nível
nutricional mais baixo ou adequado.
O que determina o exato momento da cobertura é o desenvolvimento corporal da novilha e
não a idade. A novilha deve atingir a puberdade quando apresentar de 40 a 50% do peso
corporal adulto, o que é esperado, se o crescimento for adequado, que ocorra entre os 9 a 10
meses de idade. A primeira cobertura, ou inseminação, deve ocorrer quando a novilha atingir
de 50 a 60% do peso adulto, devendo ocorrer entre os 13 e 16 meses de idade. A cobertura
somente será realizada a partir do terceiro cio, permitindo assim, melhor desenvolvimento do
trato reprodutivo da fêmea. Por fim, a novilha deverá apresentar entre 80 a 85% do peso
corporal adulto no momento do primeiro parto, devendo ocorrer, preferencialmente, aos 24
meses. Na Tabela 1 são apresentados os pesos esperados ao nascimento, puberdade, cobertura
e parto para as diferentes raças.
O desenvolvimento das novilhas deve ser medido não somente pelo peso do animal, mas
também pela sua estatura. Estimar somente o peso da fêmea pode ser um erro grave na recria
das fêmeas jovens, pois não permite a distinção entre o desenvolvimento do esqueleto e de
depósitos de gordura. O tamanho da novilha tem grande importância em sua morfologia e
pode afetar práticas de manejo e a futura capacidade de produção de leite. Vacas mais altas
e longilíneas apresentam normalmente maior profundidade de corpo o que é desejável pelo
reflexo sobre as capacidades de ingestão, respiratória e circulatória. Igualmente, o úbere e
tetos ficam mais afastados do solo permitindo uma ordenha mais confortável um úbere mais
limpo e menor probabilidade de lesões.
Na Figura 4 é apresentada a expectativa em relação à evolução da condição corporal para
novilhas leiteiras. A condição corporal de novilhas não segue os mesmos princípios que as
vacas adultas e em produção, pois estas já possuem uma estrutura corporal bem formada. As
novilhas na etapa inicial do seu desenvolvimento têm por prioridade o crescimento da
estrutura de sustentação, ou seja, a formação do tecido ósseo. Somente a partir dos 9 meses,

244
quando a novilha deve estar atingindo a puberdade, é que é observada uma aceleração no
acúmulo de reservas corporais que é, exatamente, o que pode ser medido via a utilização de
um escore de condição corporal.
Tabela 1. Peso vivo de novilhas leiteiras de diferentes raças ao nascimento, puberdade, cobertura e parto
Raça Peso vivo (kg)
Nascimento Puberdade Cobertura Parto
Holandesa 38-45 270-280 350-380 (400) 500-550 (625)
Ayrshire 35-40 240-245 275-310 450-500
Guernsey 35-40 240-245 275-310 450-500
Jersey 20-25 180-210 230-260 380-420
Girolando 33-38 280-300 320-350 400-450
Pardo-Suíço 38-45 270-280 350-380 (400) 500-550 (625)
Valores entre parênteses representam animais em condições de confinamento
A novilha no momento do primeiro parto deverá apresentar um escore de condição corporal
semelhante ao de vacas adultas em início de lactação, ou seja, uma pontuação de 3,5 (Figura
4). Alguns técnicos consideram que esta pontuação é ainda muito elevada para novilhas, as
quais ainda não teriam terminado o seu crescimento, e desta forma propõem um escore de no
máximo 3,0. Outros técnicos preferem as novilhas com um escore superior entre 3,75 e 4,0.
Independentemente do valor pretendido a avaliação da condição corporal é um instrumento
prático e expedito que deve ser utilizado por todos os produtores para a avaliação do status
energético de seus animais e, igualmente, para avaliar a qualidade e adequação da dieta
fornecida às novilhas.
Várias são as alternativas de espécies forrageiras que podem garantir ganhos de peso da
ordem de 0,750 kg/dia desde que o comprometimento entre quantidade e qualidade seja
respeitado, o que depende do entendimento da fisiologia das plantas forrageiras. Espécies
temperadas como a aveia, o azevém e outros cereais de inverno além dos trevos e suas
consorciações são exemplos de excelentes espécies forrageiras. Algumas espécies tropicais
como as cultivares de panicum, capim elefante, principalmente o capim elefante anão cv.
Mott e com mais destaque para as diferentes cultivares de capim bermuda e quicuio, também
representam excelente opção forrageira. Somente nos momentos de menor disponibilidade

245
de forragem (na época da seca no Sudeste ou no vazio forrageiro de outono-inverno na
Região Sul) ou de baixa qualidade (final de crescimento) poderá haver a necessidade de
alguma suplementação proteica e/ou energética. Cabe salientar que, mesmo o campo nativo
do sul do país, em seu pico de crescimento na primavera tem condições de suportar ganhos
de peso próximos a 500 g/dia, o que pode ser adequado para as novilhas em determinada fase
do crescimento.
Figura 4. Evolução recomendada da condição corporal para novilhas leiteiras de
raças grandes em diferentes idades (adaptado de Hoffman, 1997)
A taxa de crescimento adequada para as novilhas dependerá do estabelecimento de um plano
de forrageamento detalhado e criterioso que leve em conta a sucessão de culturas e a
indisponibilidade, em quantidade e/ou qualidade, momentânea de forragem no campo
(Viégas, 2010).
Deve ser aceito que na maioria dos casos em que ocorrem falhas no processo contínuo de
crescimento das novilhas, essas falhas estão intimamente relacionadas com o manejo das
pastagens como fonte de alimento. Pastagens de qualidade e abundantes são a fonte mais
econômica de nutrientes para uma novilha em crescimento, entretanto, tal situação pode ser
esquecida pelos produtores e o resultado é o insucesso no ritmo de crescimento dos animais.
Neste contexto, o sistema de pastejo é o que apresenta o menor impacto sobre o sucesso ou
não do manejo desta fonte natural de nutrientes. Para a atividade leiteira o sistema de pastejo
rotativo tem a vantagem de manter os animais sobre uma vigilância mais restrita, permitindo,
0
0,5
1
1,5
2
2,5
3
3,5
4
3 6 9 12 15 18 21 24
Idade (meses)
Co
nd
içã
o c
orp
ora
l

246
de maneira mais rápida e eficaz a detecção do cio e de qualquer problema de ordem sanitária
ou comportamental. Tal situação também permite uma maior interação entre o tratador e os
animais o que é importante para a redução do estresse no manejo diário.
O uso de pastagens será sempre a forma mais econômica de fornecer os nutrientes necessários
ao crescimento das novilhas, até por que estes animais não estão produzindo leite. Contudo,
em situações limitantes ao crescimento das pastagens (secas, geadas) ou à sua qualidade
(final da estação de crescimento) é obrigatório o uso de forragem conservada para manter o
ritmo de crescimento das novilhas. O problema reside muitas vezes na igualmente baixa
qualidade do alimento conservado, ou na coincidência com um período de elevada demanda
por parte dos animais. Neste caso, o uso de um concentrado energético e/ou proteico é
imprescindível. Maior será a necessidade de concentrado quanto menor for a qualidade e/ou
quantidade da forragem utilizada, entretanto, a quantidade utilizada deverá ser limitada a, no
máximo, 5 kg/dia em situações extremas.
Para que a forragem proveniente de áreas de pastagem atenda as exigências das novilhas para
crescimento, deverá ter os seguintes teores: 60 a 75% de nutrientes digestíveis totais (NDT);
> 13% de proteína bruta (PB); 0,3 a 0,65% de cálcio (Ca); 0,22 a 0,35 de fósforo (P). Com a
finalidade de verificar os níveis destes nutrientes, a realização de análises bromatológicas de
todos os alimentos utilizados na propriedade deve ser uma regra para todo o produtor que
quiser bem balancear a dieta de seus animais. Os valores que se encontram em tabelas servem
como referência, mas como foram obtidos de plantas forrageiras e ingredientes de lugares
distintos à propriedade, sofreram influência de condições edafoclimáticas e de manejo
completamente diferentes.
Com relação ao manejo, as novilhas devem ser agrupadas em lotes de 6 a 8 animais com peso
razoavelmente idêntico, o que facilita a observação diária e rotineira e também o
fornecimento de alimento mais próximo das exigências individuais para crescimento.
Estudos sobre o comportamento de bovinos jovens têm demonstrado que novilhas que são
reagrupadas de maneira mais sistemática apresentam relações mais estáveis, competindo
menos pelo espaço relacionado à alimentação além de estabelecerem relações de dominância
mais rapidamente do que outras novilhas que não passaram por esta experiência. Esta prática
poderá ser importante no momento de agregar as novilhas ao lote de vacas secas.

247
A observação constante do comportamento das novilhas é ponto chave para o bom manejo
desta categoria. Esta observação auxilia, não somente na detecção de cio, bem como na busca
do bem-estar animal. Os animais estão em comunicação frequente com o tratador, basta
somente saber escutá-los. Observar é sempre o melhor remédio e pode prever uma série de
problemas. O bom observador conseguirá identificar rapidamente o animal que está apático
e que apresenta desconforto. Estas situações acarretam em diminuição da ingestão do
alimento e consequentemente a perda de peso e retardo na IPP. Em boa parte das situações o
desconforto é causado pela falta de uma prática simples na propriedade: disponibilizar para
qualquer categoria sombra e água fresca nas áreas de pastagem e de descanso dos animais.
(Viégas, 2010).
Na situação de oferta de alimento no cocho, quando, por exemplo, é utilizada forragem
conservada e alimento concentrado, ou em situações de confinamento quando a ração total
misturada (RTM) é comumente utilizada, o produtor deve estar sempre atento às informações
que está recebendo dos animais. Entre elas, por exemplo: está havendo muita sobra de
alimento? quanto tempo ao longo do dia o cocho fica vazio? os alimentos estão com aspecto
e cheiro normal? a sobra é idêntica ao que foi ofertado? quanto tempo os animais
permanecem no cocho? a água está sendo realmente consumida? O entendimento destes
sinais auxilia ao produtor a compreender se os alimentos usados e o regime alimentar estão
adequados, evitando que haja excesso de seleção do alimento ofertado o que poderá causar
um desbalanço no aporte de nutrientes. O escore de cocho na pecuária leiteira é instrumento
válido e fornece uma informação consistente.
O uso de RTM em situações de confinamento total em várias refeições ao longo do dia é
positivo ao permitir um bom balanceamento de nutrientes, que serão fornecidos todos no
mesmo momento. Esta prática evita tanto grandes alterações do pH ruminal quanto a
diminuição da digestibilidade dos alimentos e por consequência melhora o desempenho
animal.
Em termos da condução do rebanho, recomenda-se que a partir de 4 meses antes da data
provável de parto as novilhas passem a integrar o lote de vacas secas com a finalidade de
interagir com vacas mais velhas e estabelecer novas relações de dominância. Colocar as
novilhas junto com as vacas secas somente dois meses antes do parto não é recomendável,
pois, este período coincide com o maior desenvolvimento do feto e estresse sofridos pela

248
agressividade de outras vacas poderão ter efeito importante, principalmente, pela redução do
consumo voluntário de alimento no final da gestação.
A passagem da novilha, 2 a 3 semanas antes do parto, pela sala de ordenha é manejo
fundamental, para evitar que passe pela primeira vez por este estresse logo após o parto. Este
estresse pode ocasionar quedas no consumo de alimento e, consequentemente, na produção
de leite no início da primeira lactação. A novilha deve entrar juntamente com as vacas em
ordenha para se acostumar previamente com o ambiente (luzes, ruídos, cheiros) e com o
tratador, o qual deverá também realizar o processo de limpeza dos tetos, sem realizar,
evidentemente, a ordenha para que não haja perda de colostro. Após a passagem pela sala de
ordenha as novilhas devem receber alimento concentrado na quantidade de 3 a até 5 kg/dia,
fornecimento este que tem várias finalidades, como o de estabelecer a rotina do “prêmio”
pela ordenha realizada.
O aporte de alimento concentrado no final da gestação irá garantir um bom desenvolvimento
do feto bem como o crescimento da própria novilha. No final da gestação o feto está
ocupando um espaço muito grande da cavidade abdominal e, portanto o consumo voluntário
de alimento diminui podendo levar ao início do balanço energético ou proteico negativo,
desencadeando um quadro de cetose, antes do início da lactação o que é danoso para a saúde
da novilha. O início gradativo do fornecimento do concentrado no pré-parto, por modular a
microflora do rúmen à nova dieta rica em carboidratos, previne o aparecimento de quadros
de acidose ruminal.
A adição de sais aniônicos como o cloreto de amônia e o sulfato de amônia na dieta pré-
parto diminuem os riscos de desordens metabólicas no pós-parto como a febre do leite, pouco
comum em primíparas, e do edema de úbere, que causa bastante desconforto para as novilhas
resultando em diminuição na ingestão de alimento. As dietas aniônicas estão baseadas no
balanço entre íons de carga positiva, ou cátions, no caso o sódio e o potássio e íons de carga
negativa, ou ânions, sendo de interesse o cloro e o enxofre. O objetivo é fazer com que o pH
do sangue se torne levemente mais ácido e com isto ter uma resposta do organismo com o
aumento, ou ativação, da ressorção de cálcio dos ossos na busca de reestabelecer este
equilíbrio. A principal dificuldade no uso dos sais aniônicos é a sua baixa palatabilidade, o
que torna obrigatório o uso de palatabilizantes no concentrado como, por exemplo, o melaço.

249
São vários os relatos de diminuição dos casos de retenção de placenta nas propriedades que
utilizam as dietas aniônicas. Vários são os fatores que podem levar a retenção de placenta,
entre eles: parto prematuro ou gestações prolongadas; infecções; doenças metabólicas, como
a hipocalcemia; desequilíbrio hormonal; mastites. Uma das causas possíveis é a deficiência
em vitamina E e selênio. Para que o produtor se assegure da situação de seus animais é
recomendável que ele proceda à análise dos alimentos utilizados para verificar a carência de
um destes dois nutrientes. A recomendação sem análise ou baseada em tabelas é temerária,
visto a toxicidade do selênio, que pode causar abortos e até a morte do animal.
É fundamental reagir rapidamente a problemas de ordem metabólica ou reprodutiva, pois
quanto mais tempo durar o início do tratamento maiores serão os prejuízos, os quais são
devidos não somente pelo maior tempo necessário para a recuperação, mas, também, em
termos de redução na produção de leite e no aumento dos dias em aberto, o que acarreta em
aumento do IEP e consequentemente uma diminuição na rentabilidade da atividade. O
produtor deve ter em mente que o animal que está bem nutrido, o que não é sinônimo de
animal com excesso de reservas corporais (supercondicionado), dificilmente apresentará
problemas de ordem sanitária ou metabólica.
A importância do alimento concentrado está não somente no aporte de energia e/ou proteína,
mas também em fornecer parte das exigências de vitaminas e minerais. Os minerais são,
geralmente, negligenciados pelos produtores que acabam pagando um preço elevado pelos
problemas reprodutivos causados, principalmente pelas deficiências bastante comuns de
cobre e cobalto.
Por outro lado, o aporte de proteína na dieta de novilhas leiteiras deve ser observado com
muito cuidado, principalmente se é utilizada a ureia para o complemento proteico. Nas
regiões Sudeste e Centro-Oeste do Brasil é bastante comum o uso de cana-de-açúcar
associada à ureia para os períodos de escassez de alimento. Até mesmo na Região Sul tal
prática tem sido difundida. Existem muitos relatos sobre novilhas que, mesmo ingerindo os
níveis recomendados de ureia, acabam apresentando problemas reprodutivos.
Observando a Tabela 2 é possível constatar que efetivamente o excesso de proteína
prontamente degradável no rúmen leva à redução da fertilidade das novilhas e reduz de
maneira expressiva a composição iônica do fluído uterino. Quando é realizada a divisão dos
animais por níveis de ureia plasmática, é marcante a sua influência sobre a fertilidade.

250
Contudo, não há alteração no desempenho dos animais quanto ao ganho de peso, mas, apesar
das fêmeas atingirem peso adequado aos 15 meses, devido aos problemas de fertilidade
haverá um aumento considerável da IPP. O nível de ureia tanto antes quanto após a refeição
é mais elevado para os animais que receberam maior aporte de proteína degradável no rúmen.
Os animais com elevada ingestão de proteína apresentaram ciclo estral mais prolongado após
a inseminação e também possuíam a concentração de progesterona elevada. As informações
indicam que o excesso de proteína degradável no rúmen ao alterar o pH intrauterino
ocasionam alterações na atividade secretória do útero, entretanto, o mecanismo ainda não
está suficientemente esclarecido. Para evitar tais problemas deve sempre ser verificado o
balanço entre proteína e energia e também utilizar alguma fonte de proteína não degradável
no rúmen.
Tabela 2. Taxa de concepção ao primeiro serviço, duração do ciclo estral e ureia plasmática
para novilhas leiteiras alimentadas com níveis normais ou elevados de proteína (adaptado de
Elrod & Butler (1993)
Variável Proteína normal
(15,5%)
Proteína elevada
(21,8%)
Taxa concepção (%) 82 61
Duração ciclo estral antes
da inseminação (dias) 20,1
20,4
Duração ciclo estral após a
inseminação (dias) 21,9
24,4
Ganho médio diário de
peso (g) 623
Ureia plasmática antes da
alimentação (mg/dL) 10,2
14,8
Ureia plasmática após
alimentação (mg/dL) 17,5
23,6
Grupos por nível de ureia
plasmática (mg/dL) < 9,9 9,9-16 > 16
Taxa de concepção (%) 87,5 72,5 42,8
Evidente que o descarte de uma vaca é decisão única e exclusiva do produtor que deve se
basear nos controles zootécnicos realizados desde o nascimento da fêmea para decidir qual o
melhor momento para a venda do animal. Vacas de grande mérito produtivo e morfológico
permanecem mais tempo em produção com a finalidade de obter maior número de crias com
elevado potencial genético.

251
Em resumo, cuidados simples, e que não representam custos adicionais, permitem a melhoria
dos índices reprodutivos (IPP e IEP) e queda na mortalidade de animais jovens maximizando
o retorno financeiro, pelo maior número de animais obtidos para a reposição. Assim, não
somente será possível realizar uma seleção adequada das fêmeas que irão substituir as vacas
de descarte, garantindo ganho em produção de leite futuro, mas também a obtenção de mais
novilhas excedentes para a comercialização.
Referências
Bouissou, M.F., Boissy, A.; Lê Neindre, P.; Veissier, I. The social behaviour of cattle. In: KEELING, L.J.;
GONYOY, H.W. Social behaviour of farm animals. Wallingford: CABI, 1975. p113-145.
Bouissou, M.F.; Boissy, A. Le comportement social des bovins et ses conséquences en élevage. Production
Animales. INRA, Paris, v.18, n.2, p.87-99., 2005.
Campos, O.F.; Lizieire, R.S. Novilhas: elas também merecem sua atenção. Coronel Pacheco: EMBRAPA-
CNPGL, Circular Técnica, 36, 1995. 18p.
Campos, O.F.; Lizieire, R.S. Estratégias para obtenção de fêmeas de reposição em rebanhos leiteiros. In: 10º
Simpósio sobre Produção Animal: Planejamento da exploração leiteira, 1998, Piracicaba. Anais… Piracicaba:
FEALQ, 1998. p.215-255.
Cauty, I.; Perreau, J.M. La conduite du troupeau laitier. Paris: Éditions France Agricole, 2003, 288p.
Edmonson. A. J., Lean, J., Weaver, L.D., Farver, T., Webster G. A body condition scoring chart for Holstein
dairy cows. Journal of Dairy Science, v.72, p.68-75, 1989.
Elrod, C.C.; Butler, W.R. Reduction of fertility and alteration or uterine pH in Heifers fed excess ruminally
degradable protein. Journal of Animal Science. Savoy, v.71, p.694-701, 1993.
Heinrichs, A.J.; Hargrove, G.L. Standards of weight and height for Holstein, heifers. Journal of Dairy Science.
v.70, n.3, p.653-660, 1987.
Heinrichs, A.J.; Losinger, W.C. Growth of Holstein dairy heifers in the United States. Journal of Animal
Science. Savoy, v.76, p.1254-1260, 1998.
Hoffman, P. C. Optimum Body Size of Holstein Replacement Heifers. Journal of Animal Science. Savoy, v.75,
p.836–845, 1997.
Leaver, J.D. Producción lechera: Ciencia y práctica. Buenos Aires: Editorial Hemisfério Sur S.A., 1991, 172p.
Lefebvre, D.; Brisson, J.; Gosselin, B. Pour une production laitière supérieure: un vêlage à 24 mois au poids
optimal. Le producteur de lait québécois. p. 24-27, 2002
Lucci, C.S. Bovinos leiteiros jovens: nutrição, manejo e doenças. São Paulo: Nobel/Editora da Universidade de
São Paulo. 1989. 371p.
Ministère de l’Agriculture de l’Alimentation et des Affaires rurales, Ontario, Canada. Utilisation de la note
d’état corporel dans la conduite du troupeau laitier. Disponível em:
<http://www.omafra.gov.on.ca/french/livestock/dairy/facts/94-054.htm> Acesso em: 18 de setembro de 2006
Ministère de l’Agriculture de l’Alimentation et des Affaires rurales, Ontario, Canada. Utilisation Courbes de
croissance des génisses. Disponível em: <http://www.omafra.gov.on.ca/french/livestock/dairy/facts/86-
102.htm> Acesso em: 18 de setembro de 2006
Moura, J.C.; Faria, V.P.; Mattos, W.R.S. (Eds) Conceitos modernos de exploração leiteira. 2º Congresso
Brasileiro de Gado Leiteiro, 1995, Piracicaba: Anais... Piracicaba: FEALQ, 1996. 270p.

252
Neiva, R.S. Produção de Bovinos Leiteiros: Planejamento, Criação e Manejo. Lavras: UFLA, 2ª ed., 2000.
514p.
Phillips, C.J.C. Avances de la ciencia de la producción lechera. Zaragoza: Editorial Acribia, S.A. 1996. 417p.
Radcliff, R.P., M.J. Vandehaar, L.T. Chapin, T.E. Pilbeam, D.K. Beede, E.P. Stanisiewski, H.A. Tucker. Effects
of diet and injection of bovine somatotropin on prepubertal growth and first-lactation milk yields of Holstein
cows. Journal of Dairy Science, v.83, p.23-29, 2000
Santos, G.T.; Cavalieri, F.L.B; Massuda, E.M. Alguns aspectos econômicos e de manejo na criação de novilhas
leiteiras. Balde Branco, p.56-60, 2001
Santos, G.T.; Damasceno, J.C.; Massuda, E.M.; Cavalieri, F.L.B. Importância do manejo e considerações
econômicas na criação de bezerras e novilhas. In: II Sul-Leite: Simpósio sobre sustentabilidade da pecuária
leiteira na região sul do Brasil, 2002, Toledo: Anais… Maringá: UEM/CCA/DZO-NUPEL, 2002. p. 239-267.
Santos, J.E.P.; Santos, F.A. Novas estratégias no manejo e alimentação de vacas pré-parto. In: 10º Simpósio
sobre Produção Animal: Planejamento da exploração leiteira, 1998, Piracicaba. Anais…Piracicaba: FEALQ,
1998. p.165-214.
Silva, L.F.P.; Vandehaar, M.J.; Radcliff, R.P.; Tucker, H.A. Relationship between body growth and mammary
development in dairy heifers. Journal of Dairy Science, v.85, p.2600-2602, 2002a
Silva, L.F.P.; Vandehaar, M.J.; Nielsen, M.S.W.; Smith, G.W. Evidence for a local effect of leptin in bovine
mammary gland. Journal of Dairy Science, v.85, p.3277-3286, 2002b
Van Amburgh, M.E., Galton, D.M., Bauman, D.E., Everett, R.W., Fox, D.G., Chase, L.E., Erb, H.N. Effects of
three prepubertal body growth rates on performance of Holstein heifers during first lactation. Journal of Dairy
Science v.81, p. 527–538, 1998
Viégas, J. Manejo de novilhas leiteiras, em busca da eficiência técnica. In: Geraldo Tadeu dos Santos; Ely Mitie
Massuda; Daniele Cristina da Silva Kazama; Clóves Cabreira Jobim; Antônio Ferriani Branco. (Org.).
Bovinocultura Leiteira: Bases Zootécnicas, Fisiológicas e de Produção. 1ed. Maringá: EDUEM, 2010, p. 79-
107.
Wattiaux, M.A. In: Essentiels Laitiers, Institut Babcock. Disponível em:
<http://babcock.cals.wisc.edu/publications/overview/de.fr.lasso>. Acesso em: 03 setembro 2004
Zanton, G. I., A. J. Heinrichs. 2005. Meta-analysis to assess effect of prepubertal average daily gain of Holstein
heifers on first-lactation production. Journal of Dairy Science, v.88, p.3860–3867, 2005.

253
Monitoramento de bem-estar e saúde em rebanhos leiteiros
Marcelo Cecim
A discussão sobre o bem-estar de animais de produção está presente nos mais variados
círculos da sociedade mundial. Desde os grupos de fanáticos que se julgam protetores de
animais, mas nunca desenvolveram intimidade alguma com eles, até os cientistas de
formação produtivista que são tidos como autoridades, mas na maior parte das vezes nunca
trabalharam em um sistema produtivo ao lado dos animais e não viveram a experiência de
passar "um dia de vaca". No meio destes dois grupos está a população consumidora que
houve suas opiniões extremadas. Existe, no entanto, outro grande grupo de pessoas que
trabalham diretamente com animais de produção para os quais a mídia não dá tanta atenção.
Estes vão desde tratadores, ordenhadores, proprietários e técnicos que entendem que a
lucratividade de um sistema leiteiro está intimamente ligada à saúde e bem-estar de suas
vacas. Bem-estar é condição sine qua non para a saúde, assim como esta o é para
lucratividade. Esta filosofia evolucionista de produção animal, desafia o dogma produtivista
que assume que o homem define o que é melhor para um animal simplesmente avaliando sua
performance produtiva em um determinado momento. No evolucionismo, assumimos que as
vacas têm todas as respostas sobre o que é melhor para elas, e o esforço está no sentido de
entendê-las.
Animais são indiscutivelmente seres sencientes, mas não são, pelo menos até onde
entendemos, seres conscientes. Uma vaca não sabe o que ela representa no mundo ou o
porquê que é criada. É exatamente esta falta de consciência delas que nos impõe a
responsabilidade moral de seus cuidados, de dar-lhes uma vida digna e de buscar entender
seus desejos e sentimentos. O objetivo do presente texto é resumir o que reconhecemos hoje
como a linguagem das vacas. A forma com que elas todos os dias nos dizem como se sentem,
e de que forma esta informação pode ser usada para garantir respeito e um melhor conforto
e saúde para estes animais.
Da domesticação à vaca leiteira moderna
Os fósseis mais antigos de bovinos datam de 2 milhões de anos. Os bovinos modernos
apareceram em torno de 500.000 anos atrás (Albright & Arave, 1997). Estes animais eram

254
vistos inicialmente como presas, e por vezes como predadores. Somente por volta de 6000
AC iniciou a domesticação dos bovinos na região onde hoje é a Turquia. Todo o processo
de domesticação foi direcionado a produzir animais de tração, pois a humanidade iniciava a
produção de grãos e o uso destes animais tornava mais eficiente o preparo da terra. Não fazia
sentido criar um animal para carne, pois estes podiam ser simplesmente abatidos na natureza.
Isto tem uma implicação enorme no comportamento dos bovinos modernos, que são oriundos
destes ancestrais. A mansidão e a resistência são características essenciais para um animal de
tração, ou seja, capacidade de suportar sofrimento e dor sem maior reação. Toda seleção
iniciada mais tarde para produção de carne ou leite teve como base este pool genético pré-
selecionado para complacência, resistência e baixa reação a estímulos adversos.
Com isso, acabamos por criar animais que muito pouco expressam dor, descontentamento,
angústia, medo, frustração e tédio. Mas com certeza os sentem, como qualquer um de nós.
A International Dairy Federation (2008) listou os quatro estádios de domesticação de gado
leiteiro e como estes afetaram seu bem-estar:
1- Em um estado natural apesar de plena liberdade, o bem-estar dos animais era
comprometido por predadores, fome, doenças e clima.
2- À medida que foram domesticados, seu bem-estar melhorou, pois as necessidades básicas
de saúde, proteção e alimentação passaram a ser atendidas pelos humanos. É provável, que
esta época foi quando os bovinos tiveram a melhor qualidade de vida.
3- A partir deste ponto, a seleção genética criou animais mais exigentes por serem mais
produtivos, e o desconhecimento e não atendimento destas necessidades acabou por diminuir
sua qualidade de vida.
4- Hoje estamos em um ponto onde a demanda produtiva é tamanha que o bem-estar da vaca
leiteira é seriamente comprometido, externando-se como reduzida fertilidade e reduzida
expectativa de vida produtiva.
Vale ressaltar, que na natureza, espécies que têm menor fecundidade e menor tempo de vida
reprodutiva estão invariavelmente fadadas à extinção. Infelizmente, há quem diga que estas
duas características em gado leiteiro são resultado de um "melhoramento genético".
Acredita-se que o Bos taurus teve sua origem no leste do Mediterrâneo e rapidamente ocupou
regiões da Ásia formando o Bos taurus indicus e da Europa formando o Bos taurus taurus.
Na Europa se distribuíram buscando regiões de transição entre floresta e campo, visto que

255
este ambiente permite um consumo rápido de forragem e um local abrigado para deitar e
ruminar. Na natureza, o dia de uma vaca é distribuído em três períodos de 8 horas, dedicados
ao consumo, atividades e ruminação.
A principal característica comportamental da intensificação da produção leiteira é a perda de
liberdade pelo confinamento. Neste, os animais perdem em parte a possibilidade de escolher
onde se posicionar em relação ao grupo ou ao ambiente. Por exemplo, não poder procurar
um lugar mais fresco no calor, ser obrigado a comer ou beber próximo a animais dominantes,
caminhar e ficar em contato próximo a outras como na sala de espera. Muito embora entre
vacas não ocorram disputas físicas maiores para estabelecimento da Ordem de Dominância
Social, a imposição hierárquica ocorre a todo momento. A postura e o olhar ameaçador de
uma vaca dominante sobre uma subordinada é bastante sutil, mas o suficiente para levar a
segunda a uma ação evasiva. Por menor que seja, a ativação de mecanismos de proteção e
medo sempre irá refletir negativamente sobre saúde e produção. A habilidade do tratador de
assumir a posição de "animal alfa" no grupo leva os animais a segui-lo. Existe evidência que
uma pessoa com autoconfiança e atitudes consistentes torna o manejo dos animais mais fácil.
Muito daquele conhecimento e intimidade com seu rebanho, característico dos pastores da
antiguidade se perdeu na produção moderna. No entanto, quanto mais automatizado for o
sistema de produção, menor será contato direto entre pessoa-vaca e, portanto, se faz
necessária uma sensibilidade mais aguçada em entender as necessidades das vacas. Este é
um trabalho em que o homem jamais será substituído por uma máquina.
Etologia bovina
Todo e qualquer comportamento exibido por um bovino é resultado de 3 fatores ou de suas
interações. O instinto, como levantar e mamar ao nascer, ou ainda sentar-se e resistir sempre
que for puxado para frente. Os fatores sensoriais como cheiro, som e visão que levam o
animal a aproximar-se ou afastar-se do estímulo. O condicionamento ou experiência são
coisas que o animal aprende, como beber leite em um balde, comer concentrados, entrar na
ordenha e "baixar o leite". Um exemplo de interação destes fatores é a cópula onde
inicialmente o touro tem o instinto de montar, mas sua capacidade de monta melhora com
experiência.

256
Alguns vícios comportamentais ou estereótipos surgem como uma alternativa do animal em
conviver com um estímulo negativo. Um exemplo é o enrolar ou brincar com a língua que
algumas terneiras aprendem a fazer como forma de amenizar sua frustração pelo isolamento,
falta de contato direto com outras ou simplesmente tédio.
Bovinos são animais sociais e gregários, seu isolamento é sempre estressante. Sempre terão
medo de algo novo, mas, se o estímulo não parecer ameaçador, o medo é substituído por
curiosidade até que o estímulo se torne rotina e deixe de provocar reação. Vacas têm
excelente memória, o tom de voz de um tratador que as maltrata leva a reação de fuga ou
desconforto; se existe uma experiência positiva com o tratador elas naturalmente se
aproximam. Vacas leiteiras são animais de comportamento plácido, mas quando forçadas a
fazer o que não querem apresentam reações de fuga, confusão e desorientação, criando perigo
para si e para pessoas. O tratador precisa prever a reação e a solução, sempre evitando
enfrentamento direto. Quase todos os confrontos diretos entre vaca e homem são vencidos
pela vaca. Os confrontos entre animais são sempre resultado de alguma disputa, seja por
espaço de cocho, de cama, de sombra, etc. Confrontos são sempre disputados em pé, animais
deitados ou ruminando não estão participando naquele momento de questões hierárquicas ou
estressantes. Quanto maior o número de animais ruminando deitados em um determinado
momento, maior a estabilidade e menor o desafio hierárquico e comportamental do lote. Hoje
entendemos que a ruminação em grupo representa um momento de satisfação e bem-estar
onde os fatores estressantes impostos pelo sistema produtivo estão minimizados. A
importância do monitoramento das variações individuais da ruminação como parâmetro
precoce de detecção de desafios e quebra de bem-estar será discutida no final deste capítulo.
Indicadores de desconforto
Animais mantidos em liberdade a campo sempre têm menores problemas comportamentais
que animais em sistemas confinados, provavelmente pelo tédio e pela impossibilidade de
satisfazer desejos comportamentais. Esta quebra de bem-estar é um fator de grande
importância na produção leiteira intensificada, porque a reação natural do animal é estimular
mecanismos de proteção que sempre são contraproducentes.
A sensibilidade à dor é similar entre humanos e bovinos, no entanto, a expressão de dor em
todo o animal presa é diminuída, pois esta fraqueza é facilmente reconhecida por predadores.

257
A expressão da dor é maior em um animal isolado que em um animal dentro de seu grupo.
Esta pode aparecer na forma de vocalizações, coices ou sacudir a cola para estímulos
dolorosos agudos ou ainda na ausência de ruminação, ranger de dentes ou deitar em posições
incomuns. Dorso arqueado e passo encurtado são característicos de dores crônicas.
Em animais presa, medo é sempre a reação inicial ao desconhecido como forma de proteção.
Rotina e consistência de manejo são as melhores estratégias para evitar esta reação. Sempre
que expostos a uma cerca, tronco ou piso novo deve-se dar tempo para o medo ser substituído
pela curiosidade, que é o início da construção da rotina. Vacas que defecam ou urinam na
sala de ordenha estão expressando medo. Uma vocalização de dor faz todo um grupo desviar
atenção. Sons altos e agudos causam reação de fuga, que, por sua vez quando em piso de
concreto pode causar acidentes. A ordenha deve ser um momento de relaxamento para a vaca.
De uma maneira geral, espera-se que pelo menos 40% das vacas ruminem em pé durante a
ordenha, indicando que o procedimento não é estressante. Ordenhadores calmos "fazem"
vacas calmas, vacas calmas produzem mais leite.
A demonstração de comportamentos estereotipados (atitudes repetidas, sem nenhum
propósito ou benefício aparente) é aceita como resposta animal à frustração de repetidas
vezes ser exposto a um problema ou estímulo negativo. Quanto maior a intensificação do
sistema, maior o aparecimento de estereótipos. Estereótipos orais como morder ou lamber
barras de ferro, expor e enrolar a língua (linguateio) lamber e beber urina são oriundos da
necessidade de pastar em animais que recebem dieta total. Entrar na cama do free-stall e não
deitar é uma atitude de fuga em animais que têm medo, ou de dor em animais com dificuldade
de deitar. Entrar na cama com os membros anteriores e parar com as pernas no corredor
(perching) também é um estereótipo associado a acidose ruminal.
Qualquer situação de isolamento leva a reações, em manejos como aplicação de
medicamentos, inseminação, etc. A vaca alvo deve ser contida junto ou próximo a mais
animais do mesmo grupo. A comunicação visual é a mais comum entre bovinos, postura da
cabeça é usada como forma de desafio e imposição social e a fuga como resposta, expressões
faciais não são importantes. A posição elevada da cola está presente nos confrontos, cio,
curiosidade. A cola caída representa calma e relaxamento, enquanto o posicionamento desta
por entre as pernas indica medo ou frio.

258
A introdução de uma vaca diferente no grupo leva a uma redefinição de ordem de dominância
social caracterizada por enfrentamentos que duram até 24 h, a mistura de lotes mantém esta
agitação por até 72 h, podendo afetar produção leiteira. Em grupos maiores que 100 animais,
os confrontos são mais comuns, pois as vacas têm dificuldade em lembrar sobre quem são
dominantes ou a quem são submissas. Na natureza, os rebanhos tendem a se dividir quando
ultrapassam o número de 150 vacas, sugerindo que este é a sua capacidade de memória social.
A relação entre tratador/ordenhador e suas vacas tem sido objeto de vários estudos.
Aparentemente, vacas respondem melhor a pessoas introvertidas, mas autoconfiantes.
Pessoas com tolerância, paciência, dificuldade de relacionamento e convívio interpessoal
pessoal, desconfiadas, e de poucas palavras são indivíduos que estabelecem melhores
relações com vacas leiteiras. Esta facilidade de relacionamento é estabelecida pela distância
mínima de fuga e pode ir desde zero (quando a vaca deixa ser tocada) até 6 m, e isto pode
afetar a produção leiteira em até 20%. Vacas têm plena capacidade de diferenciar pessoas
familiares e estranhos, pessoas com atitude positiva ou não.
O termo "cowpersonship" tem sido usado para definir o conjunto de habilidades que permite
a uma pessoa reconhecer mudanças no comportamento de cada uma de suas vacas, ser
sensível às condições ambientais em que vivem os animais, conduzir a movimentação de
animais sob o ponto de vista das vacas, não o seu; e, se reconhecem algo que incomoda os
animais, procurar corrigir.
Com a intensificação dos sistemas produtivos ocorrida nos últimos 50 anos, uma série de
novos comportamentos e problemas têm sido identificados em animais confinados que até
então eram praticamente desconhecidos em animais livres. Baixa expressão de cio, ou cio
silencioso é uma situação onde vacas em cio não são montadas e, portanto, não identificadas.
Uma série de fatores contribui para tal, desde estresse térmico, umidade excessiva, pisos de
concreto, vacas maiores e com problemas podais, vacas dominantes que não são montadas
por submissas, etc. Também a incidência de cistos foliculares e por vezes ninfomania é maior
em animais confinados. Recém-nascidos, principalmente filhos de novilhas, que não querem
mamar o colostro são comuns e vacas que se negam a usar as camas e deitam no corredor.
Outros comportamentos anormais incluem atirar comida para cima, ou jogar para o chão de
um cocho elevado; bater com a língua e lamber a água, ao invés de introduzir o focinho e
sugar; aumento na defecação; diminuição de ócio deitado e aumento de ócio em pé; hesitação

259
com movimentos de vai e vem na hora de deitar ou levantar; fazer posição de cão sentado. O
aparecimento, mesmo que esporádico, destes comportamentos são primeiros indicativos de
falta de conforto no sistema. Muitas vezes os sinais comportamentais precedem o
aparecimento de doenças ou queda de produção.
Observando conforto
É indiscutível que as vacas leiteiras em sistemas intensivos estão constantemente enviando
sinais que são sua "opinião" em relação ao ambiente em que vivem e como são tratadas. Estes
sinais devem ser analisados dentro de um todo e suas inter-relações estabelecidas para buscar-
se soluções factíveis. Hulsen (2011, 2013) desenvolveu manuais práticos que auxiliam no
treinamento de pessoal para entender um sistema sob o ponto de vista das vacas. São 3
perguntas básicas: 1) O que eu vejo? A resposta deve conter observações objetivas e precisas;
2) Como e por que isto acontece e qual sua causa? 3) O que isto significa, e o que devo fazer?
Resumindo, olhe, pense e faça.
Para continuar competitiva e sustentar-se no mercado, toda fazenda leiteira deve dar maior
importância aos desafios de saúde das vacas. As altas taxas anuais de descarte não apenas
representam queda de lucratividade do sistema, mas também colocam um questionamento
ético e moral sobre o direito de explorar a produção animal se isso tem um impacto negativo
tão grande na vida das vacas. Manutenção de saúde e aumento da expectativa de vida
produtiva das vacas só pode ser atingida com monitoramento constante de alterações de
conforto e bem-estar, que são os primeiros sinais da vaca que algo não está bem. Muito antes
do aparecimento de quadros clínicos. É muito triste ver proprietários que desenvolvem uma
espécie de "cegueira leiteira", oriunda do fato de ver diariamente animais sem conforto e sem
qualidade de vida e passar a considerar esta como uma situação normal.
Seguindo as orientações de Hulsen (2011): não olhe, apenas observe. Observe de todo até o
detalhe, de longe até bem próximo, do grupo até a vaca! Existe uniformidade de tamanho das
vacas? De higiene? De aspecto do pelo? De condição corporal? De escores de rúmen e
abdômen? Como os animais ocupam a área onde estão? Sua distribuição é homogênea? Das
vacas que não estão comendo ou bebendo, quantas estão deitadas nas camas? Menos que
85%? Por quê? Quantas vacas caminham com postura anormal? Isto é um problema? Qual a
causa? Estabeleça uma rotina de observação 2 ou 3 vezes ao dia, mantenha dados através do

260
tempo. Diferenças podem surgir entre categorias ou entre animais. Em relação à condição
corporal, quando mais que 10% das vacas estão muito magras ou muito gordas, uma revisão
no programa nutricional, escores de cocho e de locomoção deve ser feita. Escores de rúmen
e de abdômen indicam consumo nas últimas 24 e 48 h respectivamente.
Quantificando conforto-manejo-bem-estar
Uma série de diferentes escores relacionados à vaca e seu ambiente foram propostos, alguns
aceitos e utilizados no dia a dia da gestão de saúde de um rebanho. O que indicam os escores?
Eles são uma forma de expressão da vaca sobre elas próprias e o sistema onde vivem.
Classicamente, a filosofia produtivista prega que em se garantindo sanidade, nutrição e
genética, todas as necessidades do animal estarão satisfeitas. Esta cultura tem formado nas
últimas décadas exércitos de especialistas em suas áreas, mas completamente ignorantes
sobre o todo que envolve a vida de uma vaca leiteira. Infelizmente, vemos a assistência
técnica sendo feita com se existissem grupos de úteros que devem gestar, lotes de rúmen que
devem encher e fermentar, rebanhos de patas que devem andar e uma população de úberes
que devem produzir leite. O fato de que existe uma vaca ao redor destes e que estes são
apenas parte da vaca, acaba por ser negligenciado na tomada de decisões, principalmente nos
sistemas familiares sul-brasileiros que não contam com um técnico residente. A ilusão de que
apenas a implantação de tecnologias pontuais é a solução de todos os problemas tem levado
ao desequilíbrio e a falência de muitos sistemas. Um bom exemplo disto é a ideia de que
aumento de produção se faz através de melhoramento genético. Nos sistemas leiteiros
comerciais no sul do Brasil, tanto familiares como empresariais, existe de uma forma geral
um excesso de genética. Esta genética mais agressiva acaba por impor aos animais uma
exigência sobre nutrição, conforto térmico e de cama, capacidade de pessoal etc. que a nossa
realidade ambiental, agrícola e sociocultural não consegue prover. O resultado disto é uma
baixa expectativa de vida das vacas em relação a sua produção, e uma baixa fertilidade. É
fundamental entender que isto só se torna realidade através de muito sofrimento das vacas,
que acabam descartadas por 3 principais causas: mastite, infertilidade e problemas de casco.
Na filosofia evolucionista de produção animal devemos reconhecer o que não podemos
mudar como relevo, clima, capacidade produtiva máxima da terra e fatores financeiros de
mercado e de escolha da genética, recursos humanos, instalações, etc. que venham a criar um

261
sistema equilibrado e coerente com a interação dos fatores anteriores. O modelo gráfico
criado pelo autor para treinamento de pessoal é o de uma roda de carreta onde cada um dos
raios representam as subáreas: manejo, genética, sanidade, reprodução, nutrição, recursos
humanos e financeiros. No centro da roda está a vaca e sua satisfação com o sistema expressa
pelos escores. O tamanho da roda e sua capacidade de percorrer espaço na estrada representa
a produtividade do sistema. A queixa do produtor, ainda o principal motivo para buscar
assistência técnica, normalmente é o raio mais curto da roda. E a ideia, bastante arrogante da
ciência produtivista, é que o técnico deve "melhorar" este raio que está causando
desequilíbrio ao sistema. Analisando de uma forma holística, vemos que a queixa do
produtor, por exemplo, alto retorno ao cio nada mais é uma forma da vaca expressar seu
descontentamento com outra coisa. Ou seja, o problema está em outro raio que pode estar
mais longo, como genética, ou mais curto como conforto térmico. Na realidade ambos devem
existir, porque a qualidade de vida e desempenho da vaca é sempre resultado de uma
interação de todos os raios. Entendo que o técnico de sucesso hoje é aquele que tem como
meta equilibrar um sistema. Equilíbrio é atingido de uma forma mais viável economicamente
diminuindo raios longos, como genética, do que tentando aumentar raios curtos como
reprodução. Há que se ter a humildade de aceitar que as tecnologias conhecidas não servem
para todos os sistemas, e que todo erro de adoção tecnológica leva a queda de lucratividade
que é resultado da queda de qualidade de vida das vacas. É através dos escores que
quantificamos a qualidade e uniformidade de todos os raios. Os escores são o alfabeto da
linguagem da vaca, para entendê-la, não basta apenas conhecer os escores (as letras), mas
reconhecer suas inter-relações (as palavras) e definir soluções práticas para seu uso (as
frases).
Uma variedade de escores existe e outros podem ser criados para se quantificar a importância
de determinado problema ou resposta a uma mudança de manejo. A maioria dos escores tem
5 níveis, representando: muito baixo, baixo, médio, alto e muito alto. Em alguns se busca o
meio termo, como condição corporal, em outros como higiene o máximo, ou ainda o mínimo,
como em frequência respiratória. Neste capítulo vamos abordar 9 escores, utilizáveis em
todos os sistemas produtivos.

262
1) Escore de condição corporal
Em gado leiteiro o sistema mais utilizado é o de Edmonson (1989) que usa 5 níveis e estes
subdivididos em quartos. Não será discutida a avaliação dos escores, visto que existe vasta
literatura no tema, mas sim a necessidade do técnico em definir metas para cada rebanho, ou
ainda para cada vaca, visto a tremenda variabilidade de tamanho das vacas e tipo leiteiro nos
rebanhos sul-brasileiros. Como base deve-se considerar que animais que se aproximam do
"true-type" canadense, vacas grandes, angulosas, de alto volume, pico precoce e baixo sólido,
necessitam chegar ao parto com um escore entre 3 e 3,25 (diminuiu dos 3,75 de 20 anos
atrás). Como terá pouca reserva corporal para lactação, esta vaca necessitará de um altíssimo
aporte energético no início da lactação. A manutenção da exigência nutricional destas vacas
força as altas produtoras a um estado ruminal mais ácido com aparecimento de fezes tipo 2.
Esta exigência dificilmente será alcançada a pasto. Por outro lado, vacas de linhagem
Friesian, europeias, neozelandesas ou argentinas com maior musculosidade e deposição de
gordura, menor angulosidade, menor volume e alto teor de sólidos podem chegar ao parto
com uma maior reserva corporal para garantir a demanda para formação da gordura do leite.
Nestes animais, é possível exploração a pasto visando fermentação ruminal acética com
suplementação de concentrados, permitindo um escore fecal 3 durante o pico de lactação. O
escore de condição corporal deve ser aplicado pelo menos 5 vezes durante o ciclo produtivo:
na secagem, no pré-parto, no pico de lactação, no início da reprodução e no meio da lactação.
Reconhecer o "ponto de virada" de cada vaca é um segredo para o sucesso reprodutivo. Este
é o momento em que a vaca deixa de perder peso. Com o desencontro entre a demanda por
nutrientes e a baixa qualidade da dieta, a curva de perda de peso tem-se prolongado. Como
remédio, tem-se optado por um período de espera voluntário menor e início da reprodução
antes que a vaca perca muito peso. O resultado, além de não ser muito bom, acaba por exigir
a secagem de vacas com produção maior que 23 L, que representa seu pico de lucratividade.
O escore é a garantia de reservas para o desafio futuro, portanto deve ser avaliado dentro da
capacidade produtiva da vaca e da capacidade da fazenda em prover uma dieta que atenda
estas necessidades.

263
2) Escore de locomoção
Também descrito em 5 níveis que vão desde o caminhar normal (1) até séria claudicação (5).
O escore de locomoção é a opinião da vaca sobre a interação genética-ambiente-dieta. O
primeiro é parte dela, os demais são forçados a ela. A literatura sobre a avaliação dos escores
de locomoção é ampla (Sprechter et al., 1997). Em um primeiro momento, a vinda para a sala
de ordenha é quando se tem uma visão geral de conforto de locomoção. Olhando o grupo,
atente para as linhas de dorso. Que percentagem de vacas se desloca com o dorso arqueado?
Isto é compatível com a idade e nível de produção do grupo? Se existe um número pequeno
de vacas caminhando com desconforto (dorso arqueado), não há por que fazer análise
individual. Dedique sua hora técnica para outra coisa. É fundamental reconhecer que o escore
de locomoção é uma opinião real, mas tardia sobre o problema. Por exemplo, uma dieta com
baixa fibra efetiva, ou um ambiente com estresse térmico irão ambos diminuir tempo de
ruminação, produção de saliva e tamponamento ruminal. O desconforto no andar (escore 2),
pode levar de 10 a 20 dias para aparecer, causado pela laminite. A úlcera de sola (escore 3),
consequência da laminite leva de 20 a 60 dias para ser diagnosticada. Às vezes, o fator
estressante não existe mais, apenas o resultado. No exemplo, com ambas possíveis causas
haveria escore de consistência de fezes 2, e a causa (baixa FDN ou estresse térmico) seria
identificada pelo escore de composição de fezes ainda durante o desafio. Problemas podais
levam a queda de consumo: quando se monitora tempo de ruminação, nota-se uma grande
variabilidade na ruminação por longo tempo nestes animais. Como se a vaca comesse pouco
um dia porque tem dor, mas no próximo dia a fome é tanta que se obriga a comer com dor, e
o ciclo continua. Problemas de casco tem um impacto negativo enorme sobre a qualidade de
vida de vacas tanto confinadas como mantidas extensivamente.
3) Escore de casco
Utilizado como um exame complementar ao escore de locomoção (Hulsen, 2011). São 3
categorias que podem estar relacionadas aos escores de locomoção 1, 3, 4 e 5.
Resumidamente, escore 1 representa um casco (estojo córneo, sola, talão e espaço
interdigital) normal, escore 2 representa um único problema de grau moderado (úlcera de
sola) e escore 3 representa um problema de quadro grave ou uma combinação de alterações
(dermatite interdigital e necrose de sola).

264
4) Escore de pernas e de jarrete
O escore de pernas é uma medida do desvio de angulação dos membros posteriores em
relação à coluna. Representa uma medida do esforço da vaca em compensar o desconforto e
dor de apoio. Está divido em 3 níveis:
Escore 1: Quando as pinças apontam para a frente, paralelas à coluna com 0o de desvio até
um desvio lateral de até 17o.
Escore 2: Quando as pinças de uma ou ambas as pernas apontam entre 17o e 24o. Neste caso,
o ponto pivotante é o tarso. Este escore representa o alívio de peso sobre o dígito lateral,
colocando mais peso sobre o dígito medial. Pode ser uma tentativa de compensação de dor
solar na unha lateral, ou resultado de falta de casqueamento preventivo, visto que a taxa de
crescimento da unha lateral é maior que o da unha medial nos posteriores.
Escore 3: Quando o desvio lateral das pinças é maior que 24o, neste caso o ponto pivotante é
a articulação coxofemoral resultando em abdução de todo membro posterior.
O escore de pernas pode ser usado para avaliar a eficiência e frequência do programa de
casqueamento. Ainda dentro do escore de pernas, existe o escore de jarrete, também dividido
em 3 níveis que avaliam a condição da pele e anexos na região lateral do tarso. É usado para
avaliar qualidade das camas em sistemas de free-stall. Resumidamente, os graus são:
Grau 1: Pele e cobertura pilosa íntegra.
Grau 2: Pele engrossada, mas íntegra, perda de pelos, edema subcutâneo.
Grau 3: Áreas sem pelos, pele lesionada, extensa reação inflamatória subcutânea.
5) Escore de higiene
A quantidade de terra em sistemas abertos, ou de esterco seco ou fresco em ambos os
sistemas, aberto e fechado, aderidos ao corpo da vaca representa o nível de higiene oferecido
aos animais pelo proprietário. A higiene de úbere tem íntima relação com o aumento de
células somáticas nas amostras de tanque. A análise é feita em 3 regiões do corpo que são
ranqueadas em 4 níveis dependendo da quantidade de sujidades aderidas à pele e pelos. Às
regiões do úbere, coxas e abdômen, e pernas e cascos são atribuídos os graus 1 para região
limpa, até grau 4 para forte deposição de matéria orgânica aderia à pele. O escore de higiene
de úbere faz parte da gestão de higiene de ordenha, e serve para definir seus protocolos. O

265
escore de higiene de coxas e abdômen é uma avaliação da qualidade da cama em sistemas
fechados e da higiene dos locais de descanso em sistemas abertos. O escore de higiene de
pernas e cascos representa o grau de limpeza nos corredores, caminhos, praça de alimentação,
curral de espera, etc.
Como o escore avalia ambiente, deve-se fazer uma análise de grupo, não de animais
individuais. No entanto, é comum notar-se que os animais com coxas e abdômen mais sujos
ocupam uma posição inferior na ordem de dominância social do grupo, e lhes são permitidos
apenas os piores locais para deitar. Isto é indicativo que a qualidade do ambiente é marginal,
existe superlotação, ou ambos.
6) Escore de rúmen e abdômen
Assim como escore de condição corporal avalia reservas, ou deposição de gordura, os escores
de rúmen e abdômen avaliam consumo. Estes normalmente são paralelos aos níveis de
colesterol sérico por exemplo. O escore de rúmen representa consumo nas últimas 24 horas
e pode ser avaliado em qualquer hora do dia em sistemas fechados. Em sistemas abertos com
suplementação, após a alimentação da tarde é o melhor momento. A avaliação é feita
observando-se a aparência da fossa paralombar esquerda e pontuando-se da seguinte forma:
Escore 1: A fossa é côncava e profunda, as 3 linhas (última costela, apófises transversas
lombares e musculatura da coxa) são evidentes. Forçando-se com o punho no centro da fossa
existe a impressão de um vazio no abdômen. Uma vaca assim não comeu nada, ou pelo menos
muito pouco no último dia. Se o quadro for geral para o grupo, estão passando fome. Se
houver muita heterogeneidade, observar por competição e espaço de cocho. Se apenas um ou
poucos animais apresentarem o quadro, busque a causa primária que lhes está diminuindo
consumo.
Escore 2: A fossa ainda é côncava, mas apenas 2 das 3 linhas são evidentes. Em se fazendo
pressão, nota-se o rúmen semivazio. Este quadro é típico de animais recebendo dietas de alta
digestibilidade, alta proteína, baixa FDN e rápido tempo de passagem. Neste tipo de dieta, o
animal usa entre 50 e 60% da capacidade ruminal, rumina pouco, produz fezes grau 2 e está
em risco de desenvolver SARA (acidose ruminal subaguda). Este escore aparece
normalmente no pré-parto com a diminuição do consumo, e deve desaparecer até a terceira

266
semana de lactação em transições normais. A velocidade de evolução do escore 2 para 3
representa o desempenho da vaca naquela transição.
Escore 3: A fossa paralombar apresenta-se levemente côncava, apenas a linha formada pelas
apófises transversas lombares é visível. A pressão com o punho mostra resistência, indicando
um rúmen com conteúdo pastoso. Este representa o consumo de uma dieta equilibrada em
digestibilidade e fibra, entre 60 e 70% do volume ruminal sendo preenchido. Este quadro
normalmente resulta em fezes tipo 3, sendo o escore desejado desde os 15 até os 150 dias de
lactação para prevenir distúrbios digestivo-metabólicos.
Escore 4: A fossa paralombar apresenta-se plana, com volume abdominal visível. Apenas a
linha das apófises transversas aparece, mas pouco evidente. Neste caso, o volume ruminal
tem em torno de 85% de preenchimento, causado por uma dieta mais fibrosa, com menor
teor de concentrados e fermentação predominantemente acética. Este escore pode ser
buscado a partir da confirmação da prenhez ou meio da recuperação do peso perdido, ou seja,
a partir do dia 150 até a secagem e durante o período seco em animais que se visa manutenção
de peso. Deve passar para escore 3 quando iniciar a dieta pré-parto. Hoje entende-se que,
embora o meio e o fim da lactação com este escore, não represente o período mais produtivo
da vaca, acaba por ser o período mais lucrativo da lactação, pois tem-se uma vaca com dieta
mais barata, prenhe, ganhando peso e com raros problemas de saúde.
Escore 5: É aquele em que a fossa paralombar apresenta-se convexa e o rúmen tem conteúdo
semissólido (não confundir com timpanismo). O rúmen tem 100% de sua capacidade
preenchida, mas nunca por alto consumo, e sim por baixa digestibilidade do alimento que
tem alto tempo de passagem. Apesar de parecer bem alimentada, uma vaca assim acaba
ingerindo pouca quantidade de uma dieta de baixa qualidade. Este escore não é compatível
com produção leiteira.
O escore de abdômen representa o escore de rúmen de dois dias atrás, ou seja, o quanto a
vaca ingeriu nas últimas 72 horas. É classificado em 3 níveis observando a curvatura da
região baixa do ventre:
Escore 1: Linha do ventre bastante convexa, indicando consumo normal e satisfatório.
Escore 2: Linha do ventre entre reta e pouco convexa. Indica um animal com depressão de
consumo, e que deve ser pesquisada causa primária.

267
Escore 3: Linha de ventre de côncava, até reta. Indica um animal doente, que tem comido
pouco ou nada nos últimos 3 dias.
7) Escore de fezes
O aspecto, consistência e composição das fezes devem ser entendidos como a fase final do
processo digestivo e, portanto, a opinião da vaca sobre a dieta que lhe é oferecida. Existem
várias formas de classificação, tradicionalmente em relação à consistência:
Escore 1: Consistência líquida, diarreica. Pode ter como causa adaptação a uma nova dieta,
consumo exagerado de sal (após privação), enterites, acidose ruminal. A cor está sempre
relacionada ao tipo de dieta. Nunca pode ser considerado normal.
Escore 2: Consistência de iogurte. Aparece em dietas de alta digestibilidade, alta PB e NDT,
mas baixa FDN. Acaba sendo uma consequência do adensamento de dieta de altas produtoras
próximo ao pico de lactação. É o escore característico de rebanhos com SARA e,
frequentemente, acompanhado de escore de locomoção 2 e infertilidade. O escore 2 também
pode aparecer em sistemas semi-intensivos onde parte da dieta provém de pastejo no verão.
Nestes casos, as vacas comem concentrado e silagem, mas têm consumo de pasto e tempo de
ruminação diminuídos pelo estresse térmico. A composição da dieta parece certa, mas o que
a vaca realmente come e como processa não está certa. A forma prática de diferenciar entre
estes dois tipos de escore 2 é o exame de composição de fezes. Toma-se um copo (200 mL)
de material fecal fresca, coloca-se em uma peneira de cozinha e lava-se completamente com
água corrente. Se o material retido na peneira consistir de muco e grão inteiros ou quebrados,
trata-se de baixo consumo (não oferta) de fibra. Se na peneira restarem partículas de forragem
grandes (< 7 mm), indica falha na ruminação, característica de estresse térmico.
Escore 3: Este bolo fecal tem a forma de um pudim, mais espalhado que alto e com uma
depressão no centro. É característico de dietas com um equilíbrio entre fibra e concentrados,
com um ambiente ruminal de fermentação mista, propiônica e acética. Representa uma dieta
segura para prevenir queda de consumo e alterações digestórias. Busca-se escore fecal 3 no
início do último mês de gestação, depois a partir do dia em lactação (DEL) 15 até quando for
a opção de desadensar a dieta ao redor do dia 150. Normalmente acompanha escore de rúmen
3.

268
Escore 4: As fezes tem aspecto de bolo, mais alto que espalhado, com círculos marcados que
representam a passagem da ingesta pela válvula ileocecal. São típicos de pastos maduros, ou
dieta entre 10 e 12% de PB com elevado teor de fibra, ambiente ruminal com pH sempre
maior que 6,0. É a consistência ideal para vacas do meio para o fim da lactação e início do
período seco e normalmente acompanha o escore de rúmen 4.
Escore 5: São fezes com aspecto de biscoito, que lembram fezes de equinos ou terneiras que
pastam. Têm cor marrom escuro com muco por fora e tornam-se enegrecidas com o sol.
Escore típico de um animal doente e com baixíssimo consumo. Em nenhum momento pode
ser considerado normal em vacas adultas.
8) Escore de esfíncter (Escore de tetas)
Este escore classifica de forma quantitativa o impacto do sistema de ordenha sobre a saúde
do esfíncter do teto. Deve ser avaliado imediatamente após a retirada do conjunto de ordenha.
O escore é composto por 4 graus:
Escore 1: Não existe espessamento e o esfíncter está totalmente internalizado.
Escore2: Formação de um anel não rugoso, resultado de um prolapso inicial da musculatura
do esfíncter, mas sem queratinização.
Escore 3: Anel externo e bastante aparente no esfíncter, com alguma queratinização.
Escore 4: Esfíncter prolapsado, espessado na forma de um anel duro e com evidente
queratinização.
Sempre que houver mais de 20% de tetas com escores 3 e 4, ou quando os escores piorarem
em comparação com a última análise, deve-se fazer uma checagem completa no sistema de
ordenha: vácuo, pulsador, ciclo e borrachas. A queratinização de esfíncteres coloca a vaca
sempre em um grupo de risco mastite ambiental.
9) Escore de conforto térmico
Apesar de existirem curvas de ITU (índice temperatura umidade) onde seus pontos de corte
estão sendo permanentemente revistos, é indiscutível que a frequência respiratória da vaca
não recentemente alimentada representa sua opinião sobre quanto calor está sentindo.
Podemos classificar em 5 níveis, sendo os piores resultados obtidos antes da ordenha da tarde.

269
Escore 1: Dentro do intervalo fisiológico de 24 a 44 movimentos respiratórios por minuto.
Quase não se vê movimento de tórax.
Escore 2: De 44 a 70 movimentos por minuto, ofegante, mas com boca fechada e pouca
salivação. Movimentos de tórax facilmente vistos.
Escore 3: De 70 a 120 movimentos por minuto, ofegante, salivação com a boca por vezes
aberta, mas sem exposição da língua.
Escore 4: De 120 a 160 movimentos por minuto, pescoço estendido, cabeça elevada, muita
salivação com a boca aberta e a língua por vezes exposta.
Escore 5: Mais de 160 movimentos por minuto, inicia a exaustão, língua sempre exposta,
cabeça baixa, respiração abdominal.
O estresse térmico é o maior desafio ambiental na produção de leite durante o verão no sul
do Brasil. A combinação de dias e noites quentes com alta umidade impede que a vaca tenha
momentos no dia para esfriar. Esta situação é pior que a criação em clima desértico onde
apesar de atingir 48oC durante o dia, o ar é seco e durante a noite a temperatura chega aos
10oC.
O conjunto de escores de monitoramento aqui apresentado constitui uma ferramenta de fácil
aplicabilidade por qualquer pessoa que trabalhe na atividade leiteira. Para o técnico que pela
primeira vez veja um rebanho permite ter uma ideia bastante precisa sobre o dia a dia das
vacas antes de abordar o problema específico para o qual foi chamado. Quando a assessoria
é continuada, é a forma de acompanhar a evolução do resultado das alterações de manejo
implementadas. Ao contrário do que se pensa em um primeiro momento, este processo não
toma tempo, pois uma vez incorporado no modo de se ver uma vaca, torna-se automático
para o técnico cada vez que olha uma vaca, classificar seus escores.
Monitoramento de ruminação
É reconhecida desde muito tempo a alta correlação entre tempo de ruminação, saúde e
produção. Também se sabe que qualquer fator estressante acaba por diminuir o tempo ou a
distribuição diária da ruminação. Durante a ruminação, a vaca mantém os olhos semiabertos
em uma atitude que representa um alto estado de relaxamento. Isto representa para ela
segurança, conforto e ausência de dor. A redução na ruminação sempre foi um achado clínico
presente nas mais diversas doenças em bovinos, mesmo extra-digestórias. No entanto, a

270
quantificação do tempo diário de ruminação só era feita em pesquisa, observando-se animais
individuais em blocos de 10 minutos dentro de cada hora do dia. Foi em 2008, que a Empresa
Israelense SCR lançou no mercado uma coleira dotada de um microfone que gravava o ranger
de dentes durante a ruminação. O dispositivo então computava o tempo dedicado à ruminação
em blocos de 2 horas, e estes dados poderiam ser descarregados automaticamente para o
software assim que a vaca entrasse na ordenha. Como a coleira também possui um dispositivo
de nível, onde o deslocamento da bolha de ar também é detectado, a variação na atividade
diária do animal também era analisada. O objetivo inicial do sistema era produzir um detector
de cio mais confiável que os pedômetros, já em uso desde a década de 1970. A combinação
na flutuação das duas variáveis durante o ciclo elevou as taxas de detecção de estro de 62 a
68% com os pedômetros para 86% em sistemas fechados. Logo apareceram os primeiros
resultados da variabilidade em ruminação em diversos eventos como: parto, mastite,
problemas de casco, metrite, cetose, deslocamento de abomaso e é claro, cio.
Calamari et al. (2014) descreveram de forma inédita que a rapidez na recuperação nos níveis
de ruminação na primeira semana pós-parto está associada ao aparecimento ou não de
doenças de transição até o DEL 40. Resumidamente, o tempo de ruminação cai até 50% no
dia do parto, e retorna aos níveis pré-parto dentro de 5 dias. Vacas que atingem esta meta
terão um primeiro mês de lactação sem problemas. O brasileiro Ricardo Chebel foi o primeiro
a demonstrar que a variabilidade no tempo diário de ruminação na última semana de gestação
tem correlação elevada com eventos como cetose subclínica, retenção de placenta,
hipocalcemia subclínica e metrite. Estes resultados reforçam achados anteriores (Huzzey et
al., 2007) que demonstraram uma queda crescente no consumo voluntário pré-parto em vacas
que viriam a desenvolver metrite na segunda semana pós-parto. Também ficou claro que uma
queda em consumo voluntário é precedida por uma queda em ruminação nas últimas 12
horas, ou seja, não é o consumo que determina ruminação, mas a ruminação que leva ao
esvaziamento ruminal e permite consumo.
Entende-se que o período de stress que precede a instalação clínica de uma doença é
particularmente longo em bovinos. Essa tentativa de compensação à agressão pode iniciar 10
dias antes do aparecimento do quadro clínico. Infelizmente, os sinais de que uma vaca não
está bem são muito sutis, podendo aparecer como um leve aumento de temperatura à tarde,
posição das orelhas um pouco mais baixa, passada mais curta, e arrastar a pinça ao caminhar.

271
Todos eles são sinais que apenas são perceptíveis àquelas pessoas de observação aguçada e
que conhecem a fundo o comportamento de suas vacas. A queda de ruminação e aumento de
ócio, parecem ser os parâmetros mensuráveis automaticamente mais precoces a sofrer
alteração frente à tentativa de compensação de um desafio de saúde.
Em 2011, a startup brasileira Chipinside iniciou o desenvolvimento de uma coleira para
detecção de ruminação atividade e ócio através de um único sensor, um acelerômetro triaxial.
Em 2013, quando do lançamento do produto, outras duas empresas europeias lançavam
produtos similares. De uma forma geral, estes sistemas comparam o perfil de ruminação e a
atividade do dia em questão de uma determinada vaca com a sua média nos 5 ou 7 dias
anteriores. Cada vez que existe um período de queda de ruminação e aumento de atividade,
o sistema reconhece como cio. O indicativo é mostrado na tela do computador, mas hoje
também com acesso via celular. Apesar da importância amplamente reconhecida pelos
produtores e um preço competitivo, a coleira brasileira C-Tech não teve a penetração de
mercado inicialmente esperada. O sucesso no uso destes sistemas depende do
desenvolvimento de uma rotina diária de acompanhamento do comportamento das vacas
como forma de antever problemas futuros e instituir uma ação rápida. Um estudo europeu
mostrou que os produtores levavam em torno de um ano para familiarizar-se com os sistemas
e passar a fazer o melhor uso das informações fornecidas. O produtor de leite no Brasil
explora de uma forma bastante lenta a tecnologia de informação disponível para o
agronegócio. Um bom exemplo é a quantidade de produtores que já compraram um software
de gestão leiteira, mas o usam em parte ou esporadicamente. Parece que a rotina de inclusão
de dados, e o hábito de sentar-se ao computador serão bastante comuns na próxima geração
leiteira, mas não tanto na atual. Foi a análise desta realidade e a resposta de produtores em
um estudo piloto, que o equipamento deixou de ser vendido e passou a fazer parte de um
serviço de monitoramento. Foi então criado o "CowMed Assistant" onde, o produtor contrata
o serviço de acompanhamento que consiste na instalação do sistema e coleiras na propriedade
e os dados não só aparecem na fazenda, mas são enviados via internet e analisados na sede
da empresa no campus da UFSM. Desta forma, inicialmente por Whatsapp, agora iniciando
com um aplicativo próprio, o produtor recebe duas vezes por dia alertas de cio e de alterações
de saúde, e proximidade de parto. No grupo de cada propriedade participam produtor,
veterinário, inseminador, nutricionista, etc. Desta forma, informação sobre vacas em cio e

272
horário do início do cio são recebidas por todos simultaneamente. Quando são feitas
alterações na dieta, é informado o grau de adaptação de cada animal. A estratégia tem
mantido produtores em sua zona de conforto recebendo alertas sem a necessidade de analisar
gráficos. Por outro lado, tem facilitado a vida e qualificado a hora técnica dos clínicos que
acabam sendo chamados mais precocemente para o atendimento, aumentando a taxa de
sucesso dos tratamentos e a velocidade de recuperação. Correção de dietas e reconhecimento
de animais que não estão adaptados podem ser feitos antes que resulte em uma queda na
produção. E, por fim, a empresa recentemente lançou a nova coleira C-Tech plus que envia
dados para uma antena a cada hora, sem a necessidade de a vaca passar por um portal,
monitorando animais desde a entrada no pré-parto, e durante toda lactação.
Hoje, pode-se dizer que é impossível uma vaca desenvolver uma doença sem que antes exista
um aumento no tempo de ócio e diminuição na ruminação. Todo este corpo de informações
faz parte da construção de uma biblioteca de comportamentos que precedem doenças, assim
como perfis comportamentais no pré-parto que colocam o animal em um grupo de transição
de risco. No futuro, a produção leiteira com garantia de qualidade de vida para as vacas, não
será possível sem o uso de sistemas de monitoramento remoto. Nos próximos anos, uma série
de novos produtos apresentados na forma de tecnologias vestíveis e não invasivas já em
desenvolvimento chegarão ao mercado ao redor do mundo. O reconhecimento de variações
de temperatura corporal, de pH ruminal, do tempo do animal em pé e deitado, das variações
na deambulação será feito constantemente no animal livre, e de forma automática. Poderemos
chegar ao diagnóstico super-precoce, tão buscado na medicina humana. O veterinário terá a
oportunidade de trabalhar mais com a cabeça e de forma estratégica, visto que a necessidade
do trabalho braçal será diminuída. Ao mesmo tempo em que esta é uma oportunidade
fantástica de qualificar o trabalho daqueles que escolheram dedicar a vida "ao leite", também
abre uma série de necessidade em relação a outros conhecimentos que deverão ser adquiridos
ou aperfeiçoados por aqueles que buscam o sucesso na especialidade.
Referências
Albright JL, Arave CW (1997) The behaviour of cattle. CAB International, Oxford, UK.
Albright JL, Fulwider WK (2007) Dairy cattle behaviour, facilities, handling, transport, automation and well-
being. In Livestock Handling and Transport. 3rd edn. (Ed. T Grandin) pp. 109–133.CAB International, Oxford,
UK.

273
AssureWel (2010) Advancing Animal Welfare Assurance. Collaborative project of University of Bristol,
RSPCA and Soil Association of UK. <http://www.assurewel.org/dairycows>
Calamari, L., N. Soriani, G. Panella, F. Petrera, A. Minuti, and E. Trevisi. 2014. Rumination time around
calving: an early signal to detect cows at greater risk of disease. J. Dairy Sci. 97:1-13.
Chenoweth PJ, Landaeta-Hernandez AJ (1998) Maternal and reproductive behaviour of livestock. In: Genetics
and the behaviour of domestic animals. (Ed. T Grandin) pp. 145–165. Academic Press, Elsevier, US.
European Food Safety Authority (2009) Scientific opinion on the overall effects of farming systems on dairy
cow welfare and disease. The EFSA Journal 1143, 1–38.
Grandin T (Ed.) (1998) Genetics and the behaviour of domestic animals. Academic Press, Elsevier, US. Grandin
T (Ed.) (2007) Livestock Handling and Transport. 3rd ed.
Hemsworth PH, Coleman GJ, Barnett JL, Borg S, Dowling S (2002) The effects of cognitive behavioral
intervention on the attitude and behavior of stockpersons and the behavior and productivity of commercial dairy
cows. Journal of Animal Science 80, 68–78.
Hulsen J (2011) Cow Signals: A Practical Guide for Dairy Farm Management. Roodbont Publishers, Zutphen,
Netherlands.
Hulsen J (2013) Cow Signals Checkbook: Working on Health, Production and Welfare. Roodbont Publishers,
Zutphen, Netherlands.
Huzzey JM, Veira DM, Weary DM, and von Keyserlingk MA. 2007. Prepartum behavior and dry matter intake
identify dairy cows at risk for metritis. J. Dairy Sci. 90:3220-3233
International Dairy Federation (2008) Guide to Good Animal Welfare in: Dairy Production. Brussels, Belgium.
Mason G, Rushen J (2006) Stereotypic animal behaviour: Fundamentals and applications to welfare. 2nd ed.
Phillips C (2002) Cattle behaviour and welfare. 2nd ed. Blackwell Publishing, Oxford, UK.
Price EO (1998) Behavioural genetics and the process of animal domestication. In Genetics and the behaviour
of domestic animals. (Ed. T Grandin) pp. 31–65. Academic Press, Elsevier, US.
Sprechter DJ, Holstetler DE, Kaneene JB (1997) A lameness scoring system that uses posture and gait to predict
dairy cattle reproductive performance. Theriogenology 47, 1178–1187.
Von Keyserlingk MAG, Rushen J, de Passille AM, Weary DM (2009). Invited review: the welfare of dairy
cattle – key concepts and the role of science. Journal of Dairy Science 92, 4101–4111.
Welfare Quality (2009) Welfare quality assessment protocol for cattle. Welfare Quality Consortium, Lelystad,
Netherlands.
Whay HR, Main DCJ, Green LE, Webster AJF (2003) Assessment of the welfare of dairy cattle using
animal-based measurements: direct observations and investigations of farm records. The Veterinary Record
153(7), 197–202.

274
Pontos críticos da qualidade do leite
André Thaler Neto
Adriana Hauser
Marciel França
Veridiane Orsolin
A cadeia produtiva do leite passa atualmente por um período de busca por melhoria na
qualidade do leite in natura e consequentemente dos produtos finais destinados ao
consumidor. A necessidade de atendimento de parâmetros mínimos para os indicadores de
qualidade do leite se mostra em diversos pontos da cadeia, como na fazenda, onde a contagem
de células somáticas (CCS) pode ser usada como indicador de saúde da glândula mamária
das vacas, enquanto a composição e os atributos físicos do leite refletem principalmente
aspectos de alimentação, manejo e ambiência, enquanto a contagem bacteriana total reflete
principalmente aspectos de higiene e armazenamento do leite. Na indústria, o atendimento
desses requisitos permite melhoria na produção seja por um menor desafio tecnológico, maior
rendimento ou maior vida do produto final (Murphy et al., 2016).
Em geral, os diferentes indicadores de qualidade do leite se inter-relacionam em maior ou
menor magnitude, de modo que dificilmente podem ser solucionados de modo independente.
Assim sendo, alguns pontos são críticos em termos de atendimento aos critérios estabelecidos
pelas normas em vigor no país. Neste sentido, deve-se considerar que até a década de 1990
o Brasil não possuía uma normatização abrangente sobre atributos mínimos de qualidade em
nível de propriedade leiteira, tema que passou a ser fundamental para a cadeia produtiva
especialmente a partir do início do novo século.
Este novo panorama tem seu principal ponto de partida em 1996, quando o Ministério da
Agricultura, Pecuária e Abastecimento (MAPA) passou a trabalhar efetivamente no Plano
Nacional de Melhoria da Qualidade do Leite (PNMQL), com o objetivo de estabelecer
indicadores quantitativos de referência para a qualidade do leite cru e diminuir as perdas
qualitativas da matéria-prima decorrentes da temperatura inadequada de armazenamento e
de falhas durante o transporte até o estabelecimento beneficiador (Oliveira et al., 2000).
Como consequência ao PNMQL, em 2002, publicou-se a Instrução Normativa nº37/2002
(Brasil, 2002a), que instituiu a Rede Brasileira de Laboratórios de Controle da Qualidade do
Leite (RBQL), a qual foi incumbida de realizar análises laboratoriais oficiais e de cunho

275
fiscalizatório de amostras de leite cru das propriedades rurais e laticínios, dando o suporte
necessário às demandas geradas pela Instrução Normativa nº 51/2002, publicada no mesmo
ano, a qual estabeleceu os padrões de identidade e qualidade do leite cru refrigerado e o
Regulamento Técnico da Coleta e Transporte do Leite (Brasil, 2002b), estabelecendo os
requisitos físico-químicos, microbiológicos e de resíduos químicos do leite nas propriedades
rurais, os quais devem ser avaliados em análise compulsória mensal de uma amostra de leite,
a ser implantados a partir de 2005. A IN51/2002 estabeleceu limiares estanques para o leite
cru refrigerado, tais como no mínimo 3% de gordura, 8,4% de extrato seco desengordurado
e 2,9% de proteína, acidez titulável entre 14 e 18ºD, densidade entre 1,028 a 1,034 g/mL e
índice crioscópico variando entre -0,530ºH e -0,550ºH (Brasil, 2011). Além das IN do
MAPA, alguns parâmetros para indicadores de composição e de propriedades físicas do leite
também são regulamentados pelo Decreto 9.013/2017 (Brasil, 2017), o qual tem como
objetivos, além de estabelecer critérios mínimos de qualidade, identificar e evitar fraudes na
matéria-prima que chega às indústrias. Nesse documento, além dos critérios já mencionados,
estabelece-se um teor mínimo de lactose em 4,3%.
Por outro lado, os indicadores de qualidade microbiológica (contagem padrão em placas,
CPP, ou contagem bacteriana total, CBT) e de saúde da glândula mamária (contagem de
células somáticas, CCS) foram estabelecidos prazos para adequação dos produtores. Devido
ao fato de que parcela substancial dos produtores não conseguiram se adaptar aos critérios
mínimos para CBT e CCS nas datas limites estabelecidas, os prazos foram prorrogados em
dois momentos, pela Instrução Normativa nº 62/2011 (Brasil, 2011) e pela Instrução
Normativa nº 07/2016 (Brasil, 2016). Atualmente, as regiões Sul, Sudeste e Centro-oeste têm
como limites o valor de 3,0x105 UFC/mL para CPP e de 5,0x105 células/mL de leite para
CCS (Brasil, 2016). Deve-se considerar que estes valores fixos nos componentes sólidos e
nas propriedades físicas, com períodos de adaptação para CCS e CBT, na nossa opinião não
consideram as inter-relações entre estas variáveis.
As concentrações dos componentes sólidos do leite devem ser avaliadas no mínimo uma vez
ao mês em amostras de tanque de propriedades leiteiras, em equipamentos automatizados por
metodologia de infravermelho. A CBT e a CCS são analisadas através de técnicas de
citometria de fluxo, técnica em que é adicionado brometo de etídio ao leite, corando o DNA,
sendo o leite injetado num capilar acoplado a um sistema óptico que recebe um feixe de laser,

276
capaz de captar a fluorescência emitida. No caso da contagem bacteriana os equipamentos
automatizados estimam a contagem individual de bactérias (CIB), sendo que a partir de
amostras pareadas com análise de CPP em placas com meio de cultura e incubada a 36°C por
48 horas, são estimadas curvas de regressão da CPP em função da CIB, expressando o
resultado em UFC/mL (Brasil, 2011).
Os resultados destas análises mensais têm sido utilizados como referência em programas de
pagamento por qualidade, caracterização e fiscalização para identificação de não
conformidades causadas por adição de contaminantes, adulterantes ou leite obtido sob
condições não adequadas. Os programas de remuneração pela qualidade ainda são bastante
heterogêneos nas diferentes indústrias de laticínios do país. Estes programas são estratégias
fundamentais para a melhoria da qualidade do leite, sendo a falta de valorização da qualidade
do leite por parcela das indústrias de laticínios um ponto extremamente crítico para a
promoção da melhoria da qualidade do leite de forma sustentável em nível de propriedade
leiteira. Winck (2012) em um estudo sobre pagamento por qualidade em cooperativas do
Oeste de santa Catarina concluiu que o pagamento por qualidade impactou positivamente em
diversos aspectos sociais, econômicos e técnicos na cadeia produtiva, melhorando a
qualidade do leite e incentivando cooperativas e produtores a investirem na produção.
Objetiva-se neste texto apresentar os principais indicadores de qualidade do leite e mencionar
alguns aspectos que consideramos críticos para a melhoria da qualidade do leite e
atendimento dos critérios mínimos estabelecidos, nas condições atuais de produção do país,
considerando a heterogeneidade de sistemas de produção e níveis de adoção de tecnologia
no país.
Qualidade microbiológica do leite
Os principais microrganismos que contaminam o leite podem ser divididos segundo sua faixa
de temperatura de crescimento e multiplicação em psicrotróficos, mesófilos e termodúricos.
Os microrganismos mesófilos são o principal grupo identificado pela CBT, sendo capazes de
se multiplicar em temperaturas entre 5 e 50°C. Incluem em sua maioria bactérias Gram-
positivas dos gêneros Lactococcus, Streptococcus, Leuconostoc e Enterococcus, acidificando
o leite pela produção de ácido lático devido à fermentação da lactose (Santos e Fonseca,
2007). Este fato faz com que a elevada CBT muitas vezes esteja associada à redução nos

277
teores de lactose e, consequentemente do ESD. As bactérias termodúricas são bactérias
capazes de resistir a temperatura de pasteurização (63°C por 30 minutos, lenta; 71-75°C por
15-20 segundos, rápida), sendo que altas contagens podem estar relacionadas a falhas de
limpeza dos equipamentos de ordenha, rachaduras nas borrachas, biofilmes com depósitos
de minerais (pedras de leite) nas tubulações ou tetos muito sujos, podendo sobreviver por
anos por produzirem esporo em condições adversas (Bava et al., 2011), podendo causar
problemas na vida de prateleira do leite, como gelatinização do leite longa vida. São
exemplos de microrganismos termorresistentes: Micrococcus, Microbacterium,
Lactobacillus e Streptococcus. Gêneros como Clostridium e Bacillus são os formadores de
esporos (Santos e Fonseca, 2007). Geralmente os microrganismos mesófilos e termodúricos
apresentam baixa capacidade de se desenvolver sob condições de refrigeração na qual o leite
é armazenado, sendo consequência principalmente da contaminação inicial, quando o leite é
adequadamente armazenado.
Por outro lado, os microrganismos psicrotróficos apresentam capacidade de multiplicação a
temperaturas de refrigeração, menores que 7°C (mesmo sua temperatura ótima de
crescimento sendo maior) e podem ser responsáveis pela deterioração do leite cru refrigerado
e de seus derivados. Nesse grupo se incluem principalmente bactérias Gram-negativas dos
gêneros Pseudomonas, Achromobacter, Aeromonas e Alcaligenes, com habilidade de
produzir enzimas hidrolíticas termorresistentes as quais retêm de 30 a 100% de sua atividade
enzimática após tratamento térmico do leite, responsáveis pela redução no rendimento de
queijos, limitação da vida de prateleira, alteração de sabores, odores e aparência, bem como
perdas de proteína e gordura do leite (Murphy et al., 2016). Água não tratada, solo e vegetais
são as principais fontes, geralmente contaminando o leite em decorrência de falhas nos
procedimentos de limpeza e sanitização durante a produção. Temperatura de refrigeração do
leite inadequada ou período longo de estocagem do leite refrigerado (superior a 48 horas)
também estão associadas a altas contagens desses microrganismos (Sanvido, 2007). Apesar
de ainda não haver limites máximos de contaminação do leite por microrganismos
psicrotróficos, a baixa contagem destes constitui-se em um grande desafio no futuro, visando
melhorar a qualidade dos derivados lácteos brasileiros, visto que a refrigeração não é um
método eficiente de controle, dependendo principalmente da contaminação inicial, ou seja
das condições de higiene dos animais, dos equipamentos, do ambiente, incluindo a água, do

278
uso eficiente de detergentes e desinfetantes e das condições higiênicas do processo de
ordenha.
A análise dos dados de CBT do leite auxilia na avaliação dos procedimentos de limpeza e
desinfecção dos equipamentos de ordenha e tanques de refrigeração, higiene de ordenha,
ambiente que o animal está alojado, a temperatura e o tempo de armazenagem do leite, dentre
outros fatores. Altas contagens bacterianas podem impactar negativamente o rendimento
industrial, segurança do alimento, e na qualidade do leite e dos produtos lácteos. Apesar das
exigências atuais serem de valores menores que 300.000 UFC/mL, valores inferiores a
20.000 UFC/mL são esperados quando boas práticas de higiene são adotadas (Ribeiro Neto
et al., 2012). Além de aspectos higiênicos e de conservação, fatores ambientais também
podem afetar a CBT. Bueno et al. (2008) constataram que a CBT foi significativamente
influenciada pela umidade ambiente e precipitação pluviométrica, sendo significativamente
maior no período das chuvas, com maior contaminação ambiental, acúmulo de lama e maior
ocorrência de tetos sujos no momento da ordenha. A qualidade da água utilizada nas
atividades de ordenha pode constituir expressiva fonte de bactérias contaminantes do leite,
causando a elevação da CBT (Felipus, 2017; João et al., 2011). Entretanto, os diversos fatores
envolvidos atuam de forma conjunta, sendo difícil dimensionar seu efeito isoladamente. Por
exemplo, em um trabalho com cloração de água, observamos que nos períodos em que a água
utilizada na sala de ordenha em propriedades leiteiras da região serrana de Santa Catarina era
clorada, havia uma melhoria altamente significativa na qualidade da água, tanto em termos
de bactérias psicrotróficas como mesófilas, assim como de coliformes, em relação aos
períodos em que não havia cloradores nas mesmas propriedades. Entretanto, esta melhoria
não se refletiu em melhoria na qualidade do leite, provavelmente pela interação com outros
fatores, como uso adequado de detergentes e desinfetantes (Silva et al., 2017).
A redução da CBT é uma das mais importantes tarefas visando oferecer ao consumidor um
produto higiênico e seguro. A maioria das ações para redução da CBT estão ligadas ao
manejo e higiene na propriedade, por isso se torna fundamental o treinamento das pessoas
envolvidas, pois boas práticas de ordenha são ações simples, eficientes, mas que exigem
mudanças e adaptações na rotina do produtor. Além da limpeza dos animais que chegam para
serem ordenhados, do correto pré-dipping e da secagem dos tetos, a higiene pessoal dos
ordenhadores também é importante. As propriedades que apresentam infraestrutura mais

279
adequada para a produção, maior adoção das práticas recomendadas de manejo de ordenha
produzem leite com melhor qualidade (Werncke et al., 2016). É importante manter os
equipamentos de ordenha em bom estado de conservação, ter cuidados com procedimentos
de limpeza e sanitização, detergentes, concentração, frequência de limpeza, temperatura da
água (Bava et al., 2011). Em um trabalho no Vale do Itajaí (SC) observamos que as condições
de higiene da sala do leite e dos resfriadores também esteve altamente relacionado à
qualidade microbiológica do leite coletado pelos caminhões nas propriedades leiteiras
(Felipus, 2017). Como as ações de higiene para reduzir a CBT são tecnicamente simples em
sua maioria, as propriedades leiteiras mais profissionalizadas têm obtido resultados bastante
expressivos. Entretanto, quando analisamos grandes bancos de dados de indústrias de
laticínios e propriedades leiteiras, observamos na nossa prática que ainda estamos longe de
atingir as metas estabelecidas em termos globais. Isto se deve a diversos fatores, mas a cultura
geral da nossa população deve ser mencionada, de modo que muitas vezes práticas de baixa
qualidade higiênica do leite podem estar associadas às práticas de higiene pessoal dos
produtores rurais, sendo necessários programas voltados à saúde no campo e não somente
aos produtos comercializados.
Outro desafio é o armazenamento do leite na propriedade, o qual pode ser realizado por meio
de tanques de imersão ou expansão. O tanque de imersão deve ter capacidade para resfriar
ao leite a 7°C em no máximo três horas após a ordenha e os resfriadores por expansão direta
para 4ºC em até três horas após a ordenha, independentemente de sua capacidade (Brasil,
2011), estando a manutenção dessa temperatura diretamente ligada com a multiplicação de
microrganismo no leite e a CBT. Em um trabalho avaliando o ciclo de armazenamento do
leite em propriedades leiteiras com coleta de leite a cada 48 horas observamos que em
amostras de leite com contaminação inicial aceitável (CBT de aproximadamente 100.000
UFC/mL), quando respeitadas as especificações quanto ao volume de leite a ser resfriado, os
tanques de expansão permitem a manutenção da qualidade microbiológica do leite em termos
de CBT e contagem de microrganismos psicrotróficos, ao longo do tempo (Reche et al.,
2015).
Assim como na produção e no armazenamento, o transporte do leite até as indústrias de
laticínios constitui um ponto crítico a ser considerado. Em um trabalho realizado em uma
indústria de laticínio verificamos que a implementação de práticas que reduzam a

280
contaminação e perda de qualidade da matéria prima durante o transporte, manutenção do
leite em baixas temperaturas e redução do tempo decorrido nas rotas de coleta da matéria
prima e o seu descarregamento no estabelecimento processador apresentam importante
relação com a contagem de microrganismos psicrotróficos e a CBT (Felipus, 2017), sendo
que o transporte do leite cru a granel piorou consideravelmente a qualidade microbiológica
do leite, com valores muito superiores aos esperados pela média ponderada a partir do volume
nas propriedades de leite, porém, não demonstrou influência nos resultados físico-químicos
e de composição do leite recebido na indústria (Tabela 1). Isto demonstra que se o transporte
do leite não for feito de modo muito criterioso, a indústria que remunera o produtor por
qualidade estará pagando por um produto de determinada qualidade, a partir das análises de
amostras dos tanques nas propriedades e recebendo um produto muito inferior na sua
plataforma de descarregamento.
Tabela 1. Médias dos indicadores de qualidade do leite para os resultados ponderados das
propriedades rurais e nos tanques isotérmicos dos caminhões de transporte (Felipus, 2017)
Saúde da glândula mamária
A CCS, como indicador de saúde da glândula mamária, possui papel central na qualidade do
leite. Este fato advém não só da indicação da saúde da glândula mamária, bem como da
relação da CCS com diversos outros indicadores de qualidade do leite, em especial os teores
de lactose e, consequentemente, do extrato seco desengordurado (ESD), do teor de cloretos
do leite e de atributos físicos do leite, como a acidez titulável.

281
Para o produtor a elevada CCS é um importante indicador de perdas econômicas, visto que a
mastite subclínica reduz consideravelmente a produção de leite. Na Suécia, Hagnestam-
Nielsen et al. (2009), observaram que vacas com 500.000 células/mL já apresentavam perdas
na produção de leite, variando de 0,7 a 2,0 kg (3 a 9%) em vacas primíparas e de 1,1 a 3,7 kg
(4 a 18%) em multíparas, dependendo do estágio de lactação. Além destas perdas o aumento
de quadros clínicos de mastite, o aumento da taxa de descarte, com consequentes custos de
reposição, a diminuição do valor de mercado dos animais, o aumento do risco de
contaminação do leite com antibióticos são alguns dos demais fatores de perdas relacionadas
à mastite na propriedade.
Para a indústria o leite com elevada CCS representa prejuízos consideráveis, com destaque
para a redução do rendimento na fabricação do queijo, visto que durante a mastite, a síntese
de caseína é diminuída (Mazal et al., 2007), o que pode ser explicado possivelmente pela
redução na capacidade de síntese e secreção da caseína pelo epitélio glandular que está
lesionado pelas toxinas bacterianas (Oliver e Calvinho, 1995). Apesar dos níveis de proteína
total serem mantidos, isto ocorre às custas do aumento dos teores de nitrogênio não-
caseinoso, devido ao aumento da concentração de proteínas séricas no leite (Auldist, 1996),
diminuindo consequentemente o rendimento na fabricação do queijo. Em um experimento
com produção de queijo colonial com leite pasteurizado em escala industrial, observamos
uma piora de 5,3% no rendimento do queijo quando comparamos partidas de queijo feitas a
partir de leite com mais de 500.000 células/mL (652.000 células/mL) em relação àqueles
abaixo deste limiar (média de 373.000 células/mL), com consequente aumento no teor de
caseína no soro do queijo (Mattiello, 2015).
Outro problema decorrente da elevada CCS é a redução dos teores de lactose em função do
aumento da CCS, demonstrada em um trabalho que realizamos com dados de controle leiteiro
em Santa Catarina (Alessio et al., 2016). A redução do teor de lactose em função da mastite
subclínica, indicada pela elevada CCS, deve-se a fatores tais como mudanças na homeostase
da glândula mamária (Pessoa et al., 2012) e aumento na concentração plasmática de lactose
(Ben Chedly et al., 2009). A redução nos teores de lactose vai se refletir no ESD, podendo
levar a não conformidades no leite ao chegar à plataforma de recepção das indústrias,
resultando em prejuízos para toda a cadeia produtiva do leite.

282
Não iremos abordar neste texto todos os aspectos relacionados à prevenção da mastite
subclínica e a consequente redução de células somáticas, por tratar-se de tema complexo que
demanda abordagem separada. Em termos de pontos críticos para a redução devemos
considerar que eles podem ser similares à boa parte da temática abordada para CBT, quando
os pontos críticos da propriedade leiteira se encontram na higiene da ordenha. Entretanto,
muitas propriedades sem problemas com CBT apresentam resultados muito ruins para CCS
por estarem relacionados a fatores ligados aos animais, aos equipamentos de ordenha ou a
técnicas específicas relacionadas à prevenção e/ou tratamento das mastites. Podemos ilustrar
este aspecto multifatorial com o resultado de uma pesquisa que realizamos em propriedades
com controle leiteiro em Santa Catarina (Cardoso et al., 2015), onde foram identificados
como principais fatores relacionados ao aparecimento de novas infecções intramamárias
fatores ligados aos animais (ordem de parto e a profundidade de úbere), ao ambiente (escore
de limpeza dos úberes), ao animal e ao sistema de ordenha (presença de hiperqueratose em
ponta de teta e adoção de linha de ordenha), enquanto para a cronificação da mastite
subclínica o estágio de lactação, a profundidade do úbere e a manutenção dos equipamentos
de ordenha foram fundamentais. A redução da CCS continua sendo um desafio à pecuária
leiteira em quase todo o mundo e é um ponto crítico para a melhoria da qualidade do leite no
seu sentido mais amplo.
Composição do leite
Teor de gordura
O teor de gordura ainda não representa um grande desafio para a cadeia produtiva como um
todo, visto que parcela considerável das propriedades de leite do país ainda não possuem
animais com elevada produção de leite e, consequentemente com teores de gordura
relativamente altos, de modo que o pool de leite recebido pelas indústrias apresenta teores
relativamente elevados de gordura. Além disto, o fato de que os teores de gordura são
manipuláveis pelas dietas e pelo manejo alimentar adotado pelos animais, os produtores
podem adotar estratégias de produção voltadas ao mercado, de acordo com as tabelas de
remuneração por sólidos do leite adotados pelas indústrias.

283
Por outro lado, nas regiões em que são utilizados sistemas de produção com elevada produção
por vaca, muitas vezes associada à elevada suplementação com carboidratos rapidamente
fermentáveis, a manutenção de adequados teores de gordura no leite torna-se um dos grandes
desafios da produção eficiente de leite.
Por entendermos o teor de gordura como um dos principais indicadores de saúde ruminal de
vacas leiteiras e, consequentemente um tema central na nutrição de vacas leiteiras, não
abordaremos este tema neste texto, visto que sua complexidade exige uma abordagem
profunda. Mas, pelo menos dois aspectos não nutricionais fundamentais para o teor de
gordura precisam ser considerados. O melhoramento genético para produção de sólidos do
leite é fundamental para garantir bons teores de sólidos do leite em longo prazo e uma
adequada ambiência, especialmente em termos de redução do estresse térmico e,
consequentemente à capacidade de consumo de alimentos pelas vacas leiteiras. Sem haver
reocupação com estes fatores dificilmente os resultados dos programas de alimentação de
vacas leiteiras terão bons resultados em termos de teores de gordura no leite.
Extrato seco desengordurado (ESD)
O ESD é composto por todos os componentes do leite, excetuando-se a gordura, sendo a
lactose e a proteína seus principais componentes. A lactose é importante para as propriedades
coligativas do leite: pressão osmótica, abaixamento do ponto crioscópico e elevação do ponto
de ebulição, sendo responsável por cerca de 50% na pressão osmótica do leite. Variações nos
teores de lactose estão associadas com variações recíprocas de constituintes solúveis em
água, especialmente sódio e cloretos (Varnam e Sutherland, 2001). Em situações de
inflamação da glândula mamária, ocorrem lesões nas junções das células secretoras de leite,
acarretando em alteração na permeabilidade. Essa alteração permite a passagem de lactose
para a corrente sanguínea e de íons da corrente sanguínea para o leite, especialmente o
cloreto, gerando alterações na composição do leite. Outro fator que reduz o teor de lactose é
o déficit energético. Em um experimento em Santa Catarina observamos reduções
importantes nos teores de lactose no outono podendo ser atribuído à deficiência nutricional
que ocorre nesta época do ano, especialmente devido à deficiência quali-quantitativa de
volumosos, com aumento na primavera, época com maior disponibilidade de pastagens, em
quantidade e qualidade no sul do Brasil, sobretudo de aveia (Avena spp.) e azevém (Lolium

284
multiflorum) (Alessio et al., 2016). Neste mesmo estudo foram identificadas reduções severas
nos teores de lactose em função do aumento da CCS. De modo diferente dos teores de
gordura, pouca variação nos teores de lactose é observada em função da raça ou estágio de
lactação. Outro fator que afeta consideravelmente os teores de lactose é o estresse térmico.
Wheelock et al. (2010) observaram redução de 200 a 400 g diárias na produção de lactose
em vacas leiteiras sob estresse térmico, sendo a glicose consumida em maior velocidade nas
vacas sob estresse térmico. Há um aumento da glicólise e da respiração anaeróbia para manter
o equilíbrio energético durante situações de estresse térmico (Tian et al., 2015), sendo que a
diminuição da glicemia pode explicar a redução do teor de lactose (Schwartz et al., 2009).
A proteína do leite sofre influência racial e do nível de produção de leite, porém menos
marcante que a gordura (CSIRO, 2007), sendo o melhoramento genético fundamental para
manutenção de bons teores de proteína a longo prazo. Outro fator determinante no teor de
proteína do leite é a dieta, sendo que vacas alimentadas com dietas ricas em carboidratos não
fibrosos produzem leite com mais proteína (Ishler et al., 1996). O aumento na CCS não
determina grandes alterações no teor total de proteína do leite (Tomazi et al., 2015), devido
à redução na síntese de caseína, acompanhada por aumento na passagem de proteínas
plasmáticas para o leite (Urech et al., 1999), trazendo prejuízos na produção de derivados,
como queijos (Mattiello, 2015; Urech et al., 1999), porém não é significativa na redução do
ESD, especialmente a ponto de gerar inconformidade. O estresse térmico também pode
determinar redução no teor de proteína do leite, visto que o catabolismo muscular aumenta
durante o estresse térmico, ocorrendo maior concentração no nitrogênio ureico plasmático,
favorecendo uma maior redistribuição do nitrogênio proteico para ureia (Wheelock et al.,
2010) e diminuindo a habilidade da síntese proteica das células mamárias, reduzindo-se o
teor de caseína (Bernabucci et al., 2002). Além disto, também ocorre redução no consumo
de alimento pelas vacas leiteiras em estresse térmico (Rhoads et al., 2009), podendo reduzir
os teores de lactose e proteína.
Como resposta às variações nos teores de lactose e proteínas observa-se considerável
variação no teor de ESD. Em um estudo no sul do Rio Grande do Sul, foram encontradas
inconformidades nos meses de fevereiro, março e abril, com ESD abaixo dos 8,4%
preconizados. Essa variação foi explicada por teores de proteína menores, porém
principalmente por uma queda no teor de lactose, o que pode ser explicado pela queda na

285
qualidade da alimentação durante o outono (Gonzalez et al., 2004). Em um estudo com mais
de cem mil amostras de tanques de propriedades leiteiras em 2015 (em andamento) também
observamos baixos teores de ESD no outono, com elevado percentual de amostras abaixo de
8,4% (não conformes), além do verão (Figura 1).
Durante o outono, os baixos teores de ESD e, consequente aumento de não conformidade,
devem-se principalmente à redução no teor de lactose, sendo parcialmente compensados pelo
aumento nos teores de proteína, o que pode estar relacionado à menor produção de
leite/vaca/dia nas condições de menor oferta alimentar. No verão a diminuição do ESD deve-
se aos teores menores de proteína e lactose, o que pode ser atribuído apo estresse térmico.
Neste estudo também se observa que os demais fatores que deprimem os teores de ESD são
a elevada CCS, pelo seu impacto sobre o teor de lactose e pelo tamanho das propriedades,
sendo que os produtores que comercializam pequenas quantidades de leite são mais
vulneráveis a apresentar não conformidades, especialmente no outono, visto que estas
propriedades apresentam menor capacidade de manter uma condição adequada de
alimentação ao longo de todo o ano, dados entre outras peculiaridades, pela menor
capacidade de conservação de forragem e pelo nível de tecnificação predominantemente
menor.
Figura 1. Percentagem de não conformidade para extrato seco desengordurado (ESD) e
médias de proteína, lactose e ESD

286
Gabbi et al. (2013) analisando dados de rebanhos leiteiros dos estados do Rio Grande do Sul
e de Santa Catarina, observaram que a maior diferenciação entre as propriedades leiteiras se
dá pelas estratégias de alimentação das vacas leiteiras. Estes resultados demonstram que a
redução das não conformidades em ESD passa pela melhoria da estrutura das propriedades,
da alimentação das vacas leiteiras e da saúde da glândula mamária.
Propriedades físicas do leite
Uma série de alterações nas propriedades físicas do leite tem causado prejuízos a toda a
cadeia produtiva do leite, seja pelas dificuldades que impõem ao processamento industrial do
leite, em especial ao tratamento térmico, assim como a dificuldade em distinguir as situações
de fraudes. Salienta-se que a maioria destas alterações são mais frequentes no verão, sob
condições de estresse térmico. Vamos descrever algumas destas alterações que podem ser
encontradas em condições reais de produção.
Leite instável não ácido (LINA)
O leite que precipita ao teste do álcool e não apresenta acidez titulável acima de 18° Dornic
é denominado leite instável não ácido (LINA) (Fischer et al., 2012). Como há perda da
estabilidade da caseína, ocorrem perdas no processo térmico, do leite, especialmente no
processo UHT (ultra high temperature) (Marques et al., 2007). A prova do álcool é realizada
na propriedade antes do carregamento do leite, como teste de rejeição ou aceitação do leite
pela indústria, sendo pouco seletivo e específico. A prova avalia a estabilidade das proteínas
lácteas submetidas a desidratação provocada pelo álcool, sendo usada para estimar a
estabilidade do leite quando submetido ao tratamento térmico (Marques et al., 2007; Horne,
2016). A adição de álcool ao leite modifica a constante dielétrica do meio e modifica o
equilíbrio que envolve a coagulação das proteínas do leite (Horne, 2016). A coagulação do
leite induzida pelo calor resulta da redução da estabilidade coloidal das micelas de caseína,
as quais são associações hidratadas de coloides estabilizados por eletrólitos. O núcleo das
micelas consiste em uma matriz de proteína amorfa e nanoclusters amorfos de fosfato de
cálcio, também chamado de fosfato de cálcio coloidal (CCP) e água, enquanto a kappa-
caseína localiza-se na superfície das micelas. Para manter a estabilidade micelar, interações
entre fosfato de cálcio e proteínas são necessárias, na forma de interações hidrofóbicas e

287
eletrostáticas, ligações de dissulfeto, ligações de hidrogênio e interações de van de Waals.
Por outro lado, a estabilidade coloidal das micelas de caseína é governada principalmente
pela extremidade C-terminal da κ-caseína, que se projeta da superfície da micela, sendo
predominantemente de natureza estérica. (Huppertz, 2016). Muitos processos, incluindo
tratamento UHT, esterilização e produção de leite em pó, em que a formação de géis ou a
sedimentação devem ser evitados, são favorecidos pelo leite com alta estabilidade. Em outros
processos, como produção de queijos e iogurte, em que a coagulação e a formação de géis
são necessárias, uma menor estabilidade do leite pode ser até benéfica, já que pode suportar
calor suave (Tsioulpas et al., 2007).
A estabilidade depende de diversos outros fatores, tais como o pH. Em seu pH natural, o
leite é estável ao tratamento térmico e suporta temperaturas de até 140°C por períodos
maiores que 15 minutos, sem sinais visíveis de instabilidade. No entanto, mesmo pequenas
mudanças no pH, com variações de 0,1 unidade de pH, podem resultar em mudanças na
estabilidade do leite ao calor (Huppertz, 2016). Alterações relativamente pequenas nas
concentrações de sais de leite, especialmente de cálcio, fósforo inorgânico e citrato, também
podem ter efeitos importantes sobre as características tecnológicas do leite, podendo estar
relacionado à alimentação das vacas (Fox e Sweeney, 1998). Fagnani et al. (2014) no Paraná
observaram que quando comparado ao leite normal, o leite LINA apresentou maior
concentração de potássio (43,42 versus 38,50 mmol/L; P< 0,05), menor concentração de
cálcio total (90,87 versus 104,54 mmol/L; P< 0,05) e menor concentração de fosfatos (58,48
versus 73,67 mg/dL; P< 0,05).
O balanço ácido-básico é fundamental para a manutenção do pH sanguíneo e a saúde
animal. Em vacas de leite, alterações no equilíbrio ácido-básico podem ser acompanhados de
distúrbios eletrolíticos e consequentemente, podem influenciar na composição e no balanço
iônico do leite, além da produção de leite (Marques, 2011; Fischer et al., 2012). Fagnani et
al. (2014), ao pesquisarem a relação entre distúrbios metabólicos e leite LINA, encontraram
que a alcalose metabólica compensada foi o distúrbio metabólico mais prevalente associado
ao LINA. Werncke (2017) observou redução da estabilidade do leite ao teste do álcool pela
indução de acidose em vacas leiteiras.
O melhoramento genético para desempenho produtivo aumenta as exigências nutricionais
das vacas leiteiras e a susceptibilidade destas a fatores estressantes, com destaque para o

288
estresse térmico, em função do incremento calórico a partir da maior ingestão de alimentos.
Deste modo, as deficiências nutricionais e o estresse térmico, de modo isolado ou em
conjunto, exercem papel preponderante no desencadeamento de casos de LINA em condições
de campo. Diversos trabalhos têm demonstrado o aumento do aparecimento de quadros de
LINA em épocas de menor oferta de alimentos, em especial o outono, assim como em
propriedades menos estruturadas (Werncke et al., 2016), sob condições de restrição alimentar
(Gabbi et al., 2016). A restrição, provocada pela redução de 40 a 50% da quantidade de
alimento oferecida, por um determinado período de tempo, reduz a produção de leite e
aumentou a frequência da ocorrência do LINA (Zanela et al., 2006) e produz lesões nas
junções fortes entre as células secretoras de leite nos alvéolos mamários, indicadas pelo
aumento na concentração de lactose no plasma sanguíneo (Stumpf, 2013). Schmidt (2014),
em um experimento de indução de leite LINA pela restrição de 50% das exigências de
proteína e energia de vacas de raça Holandesa e Holandesa x Jersey em Lages (SC), observou
uma recuperação mais rápida do quadro de instabilidade do leite ao teste do álcool, no
tratamento em que as vacas receberam uma dieta atendendo 100% das exigências de energia
e proteína. Observou também aumento na produção de leite e na concentração de lactose.
Houve alguma recuperação nos quadros de LINA com suplementação energética, porém o
agravamento do quadro de LINA, juntamente com severa redução na produção de leite e
condição corporal das vacas quando era realizada suplementação proteica, porém com déficit
energético (Figura 2).
Os relatos de quadros de LINA durante o verão são bastante frequentes, sendo que Abreu
(2008) demonstrou este problema experimentalmente. Enquanto a restrição de sombra em
vacas em pastejo sob condições de estresse térmico intenso em Itapiranga (SC) provocou
uma redução severa na resistência do leite ao teste do álcool, em um experimento similar sob
condições de estresse moderado, o mesmo autor não observou redução na resistência ao teste
do álcool.

289
Figura 2. Concentração média de álcool em que ocorreu a coagulação do leite em função dietas
visando suprir as exigências de energia (100E+50P), proteína (50E+100P) ou ambas
(100E+100P). Fonte: Schmidt (2014)
Baixa acidez titulável
A acidez titulável serve como um indicador de leite ácido, sendo que a IN 62 estabelece como
requisitos de acidez titulável do leite cru refrigerado 0,14 a 0,18 g de ácido láctico por 100
mL ou 14 a 18oD (Brasil, 2011a). O leite recém ordenhado apresenta-se ligeiramente ácido,
denominada acidez natural, tendo como origem seus componentes normais, como albumina,
citratos, dióxido de carbono, caseínas e fosfatos (Brasil, 2013). A lactose pode ser fermentada
por ação de microrganismos com formação de ácidos orgânicos, em especial o ácido láctico,
resultando na chamada acidez adquirida, a qual em conjunto com a acidez natural, forma a
acidez real do leite. Quando o leite é obtido em condições inadequadas de higiene e
refrigeração deficiente, a acidez aumenta, podendo tornar o leite impróprio para o consumo
humano (Brasil, 2013). A variação do teor de proteína e das frações proteicas também afeta
a acidez titulável do leite cru (Töpel, 2016), sendo que o leite produzido por vacas sob
estresse térmico apresenta menores teores de proteína e lactose, além de um pH mais elevado
e menor acidez titulável. No estresse térmico também ocorre uma alteração nas frações
proteicas do leite e o teor de αs e β caseína diminui, proteínas ricas em grupos fosfato, sendo

290
os constituintes ácidos das micelas de caseína. Assim, seu menor teor no leite pode explicar
o menor conteúdo de fósforo no leite, com maior pH do leite e menor acidez titulável,
comumente registrada durante os meses de verão. A fosforilação das αs e β-caseínas necessita
da presença do fosfato de ATP, sendo que essa fosforilação é significativamente prejudicada
em condições de déficit de energia, de modo que se supõe que o menor teor de αs e β-caseína
no leite de verão podem ser pelo menos parcialmente devido à redução do aporte de energia
e proteína via alimentação (Bernabucci et al., 2013).
Trabalhos em desenvolvimento pelo nosso grupo de pesquisa demonstram o grande aumento
no percentual de propriedades leiteiras com acidez titulável abaixo de 14oD no verão,
inclusive em amostras conjuntas de tanques de caminhão, resultando em considerável
preocupação para as indústrias de laticínios devido às inconformidades em relação aos
regulamentos de qualidade do leite. Os resultados parciais destes trabalhos também
demonstram o potencial de redução da acidez titulável e do teor de lactose pelo aumento da
CCS, tanto em amostras de tanque de resfriamento (Arruda Júnior et al., 2016), como em
amostras individuais de vacas (Arruda Júnior et al., 2015).
Alterações no índice crioscópico
O índice crioscópico (IC), medida do ponto de congelamento do leite é uma prova utilizada
principalmente para detectar fraude por adição de água ao leite devendo estar entre -0,530oH
a -0,550oH, o que corresponde em graus Celsius a -0,512oC a -0,531oC (Brasil, 2011a), sendo
que valores mais altos (próximos de zero) podem indicar adição de água e valores mais baixos
(mais afastados de zero) podem indicar maior quantidade de solutos no leite. Porém, em
algumas situações, o leite apresenta distúrbios nesses valores, sem que haja fraude proposital,
podendo resultar na aplicação de penalidades ao produtor.
A lactose e íons dissolvidos respondem por cerca de 80% da diminuição do índice
crioscópico e o nitrogênio não proteico, principalmente ureia, ácidos graxos de cadeia curta
e CO2 são responsáveis por cerca de 20% (Töpel, 2016), sendo que estes componentes não
são independentes entre si, podendo a variação de um componente ser compensada por outros
componentes. A influência da composição é responsável também pela variação do IC ao
longo da lactação, em que animais com lactação mais avançada tem menor chance de ter um
IC alto, similar ao encontrado em casos de fraude por adição de água (Botaro e Santos, 2008;

291
Santos, 2012). Essa variação ao longo da lactação pode gerar ocasiões de inconformidade em
rebanhos que apresentam parições estacionais (Henno et al., 2008). Em um estudo no Canadá
foram observadas variações estacionais no ponto de congelamento do leite com maior IC no
verão e valores menores no inverno (Schukken et al., 1992). Esse comportamento também
foi relatado na Europa, em rebanhos recebendo dieta balanceada associada a pastagem. Este
fato pode estar relacionado ao maior consumo de água pelos animais (Henno et al., 2008),
uma vez que a osmolaridade do leite e do sangue estão intimamente relacionadas (Shipe,
1959). Pagno et al. (2013) observaram aumento do IC em vacas com restrição hídrica na
pastagem e que recebiam água antes da ordenha, em Castro (PR), sendo que após a supressão
da oferta de água antes da ordenha o IC retornou a valores normais. A alimentação das vacas
também tem impacto sobre o IC, porém apenas em casos extremos (fome) há possibilidade
de este fator, isoladamente, elevar o IC acima do limite regulamentado (Bowman et al.,
2005). Porém, dietas desequilibradas, com baixa energia, deficiência de minerais ou excesso
de proteína podem levar a um aumento do índice crioscópico (Töpel, 2016; Bowman et al.,
2005).
Outras causas de aumento no IC não relacionados a fraude intencional podem ocorrer. Um
deles é a água residual de lavagem automática em sistemas de ordenha, condução e
resfriamento de leite, assim como a condensação de água nas tubulações de condução do
leite, onde a água se acumula e mistura ao leite. Por outro lado, o aumento da acidez do leite
pela fermentação da lactose em ácido láctico, causa diminuição do índice crioscópico do
leite, assim como vacas privadas de água podem produzir leite com baixo IC, devido à
desidratação (Botaro e Santos, 2008; Santos, 2012; Shipe, 1959).
Aumento nos teores de cloretos no leite
O teste de cloretos no leite é feito com o objetivo de identificar fraudes, visto que a adição
de cloretos pode ser utilizada com a finalidade de “mascarar” uma adulteração no leite pela
adição de água, permitindo a correção da densidade e a crioscopia do leite. Esta análise,
quando empregada paralelamente às outras provas de controle de qualidade, constitui uma
ferramenta importante para confirmar suspeitas de fraudes por adição de água e sal ao leite
(Brasil, 2014).

292
A pesquisa de cloretos em leite fluído fundamenta-se na reação do nitrato de prata com
cloretos em presença de cromato de potássio como indicador. O teste é considerado positivo
se a solução resultante apresentar coloração amarela e é considerado negativo com
aparecimento de precipitado marrom. Devido a coloração natural do leite o precipitado
marrom é visualizado como alaranjado (Brasil, 2014). Um resultado positivo não significa
necessariamente fraude por adição de cloretos à amostra. Significa que a concentração de
cloretos na amostra é superior à quantidade encontrada em animais sadios (0,08 a 0,1%).
Alguns fatores podem influenciar no teor de cloretos presente no leite, tais como diferenças
individuais, alimentação, estado de hidratação, raça, espécie, número de lactações, variações
diurnas, estágio de lactação, sazonalidade, além de alterações patológicas. O teor pode
aumentar com o avanço do período de lactação ou na fase precoce da lactação, ou em
condições de transtornos da secreção, especialmente nos casos de mastite. A não
consideração desses fatores pode resultar em testes com resultados falsos positivos, com
graves consequências econômicas, representadas principalmente pelo descarte do leite
(Brasil, 2014)
Variações na concentração de cloretos no leite em função do estágio da lactação foram
observadas por Raimondo et al. (2009) ao avaliarem leite de vacas Jersey a partir de 12 horas
após o parto até 30 dias de lactação, encontrando concentrações máximas de cloretos no
período colostral (154,1 ± 29,3 mg/dL) em amostras coletadas até 12 horas após o parto. A
partir do segundo dia de lactação, houve uma diminuição significativa de cloretos que se
estendeu até 30 dias de lactação. Estes autores consideram valores de referência para cloretos
em leite de vacas Jersey, valores entre 137,7 e 173,4 mg/dL.
Alterações na permeabilidade vascular da glândula mamária podem afetar a concentração de
determinados íons no leite e os mais afetados são sódio, potássio, cálcio e cloro. Infecções
bacterianas na glândula mamária causam destruição do epitélio secretor, abertura das tight
junctions entre as células e aumento da permeabilidade capilar (Kitchen, 1981). Ocorre um
aumento de sódio e potássio no leite em casos de lesões. Por outro lado, a concentração de
potássio e cálcio diminui no leite pois estes, devido às mudanças de permeabilidade, escapam
para o sangue (Ogola et al., 2007). Essas alterações também se refletem em um aumento no
pH e na condutividade elétrica (CE) do leite pelo aumento da concentração de sódio e cloretos
no leite. A CE também é usada como um indicador da ocorrência de mastite, especialmente

293
para diagnóstico em sistemas de ordenha (Córdova, 2016; Della Libera et al., 2001). O teor
de cloretos também pode aumentar quando a temperatura ambiental aumenta, sendo que
aumentos de 30 a 100% na concentração de cloretos podem ocorrer quando vacas são
mantidas em temperaturas variando entre 29 a 32oC. A concentração de cloretos aumenta
para compensar a diminuição de lactose, sendo a variação na proporção cloretos/lactose um
mecanismo fisiológico para manter a pressão osmótica das células da glândula mamária
(Bernabucci e Calamari, 1998). Assim sendo, a reação de amostras de leite positivas para
cloreto, especialmente no verão, tem sido uma preocupação em diversas indústrias de
laticínio.
Conclusões
Os diversos indicadores de qualidade do leite estão inter-relacionados, de modo que
dificilmente serão obtidos resultados positivos para um indicador sem considerar o conjunto
de indicadores. Dentre os pontos críticos da qualidade do leite destaca-se a contagem de
células somáticas, a qual, indica, de forma isolada, a saúde da glândula mamária, com
elevados impactos na rentabilidade da propriedade leiteira e no rendimento industrial. Além
disso, elevadas contagens de células somáticas podem determinar reduções consideráveis nos
teores de lactose e caseína, com consequências sobre o ESD, além de aumentar a
probabilidade de ocorrência de não conformidades por baixa acidez titulável ou provas
positivas para cloretos.
Outro ponto crítico é a alimentação animal que pode afetar os teores de praticamente todos
os componentes do leite, aumentar consideravelmente o risco de ocorrência de LINA além
de alterações em outros aspectos físicos do leite. Merece destaque também o estresse térmico,
visto estar relacionado de forma isolado ou em conjunto com outros fatores tais como erros
de alimentação animal, com as mais importantes alterações nas propriedades físicas do leite.
Assim sendo, não podemos esperar resultados excelentes na qualidade do leite sem melhoria
na ambiência das vacas.
Referências
Abreu, A. S. Leite instável não ácido e propriedades físico-químicas do leite de vacas Jersey. 2008. 97 f.
Dissertação (Mestrado em Zootecnia) Universidade Federal de Santa Maria, 2008.

294
Alessio, D. R. M. et al. Multivariate analysis of lactose content in milk of Holstein and Jersey cows. Semina:
Ciências Agrárias, v. 37, n. 4, p. 2641–2652, 2016.
Arruda Junior, L. C.; Alessio, D. R. M.; Abreu, A.S.; Werncke, D.; Reche, N. L. M.; Voges, J. G.; Fischer, V.;
Thaler Neto, A. Baixa acidez titulável em leite de tanques de resfriamento. In: Congresso Brasileiro de
Qualidade do Leite, 6, 2015, Curitiba - PR. Anais..., Curitiba: CBQL, 2015.
Arruda Júnior, L. C.; Alessio, D. R. M.; Schmidt, F. A.; Pelizza, A. et al. Low titratable acidity in individual
milk samples of cows. In: Reunião Anual da Sociedade Brasileira de Zootecnia: Produção Animal Para as
Novas Gerações, 53, 2016, Gramado - RS. Anais...Porto Alegre: UFRGS, 2016.
Auldist, M J et al. Effects of somatic cell count and stage of lactation on raw milk composition and the yield
and quality of Cheddar cheese. Journal of Dairy Research, v. 63, n. 2, p. 269–80, 1996.
Bava, L.; Zucali, M.; Sandrucci, A.; Brasca, M.; Vanoni, L.; Zanini, L.; Tamburini, A. Effect of cleaning
procedure and hygienic condition of milking equipment on bacterial count of bulk tank milk. Journal of Dairy
Research, v. 78, p. 211–219, 2011.
Ben Chedly, H.; Lacasse, P.; Marnet, P. G.; Wiart-Letort, S.; Finot, L.; Boutinaud, M. Cell junction disruption
after 36 h milk accumulation was associated with changes in mammary secretory tissue activity and dynamics
in lactating dairy goats. Journal of Physiology and Pharmacology, v. 60, n. 3, p. 105-111, 2009.
Bernabucci, U.; Basiricó, L.; Morera, P. Impact of hot environment on colostrum and milk composition.
Cellular and Molecular Biology, v. 59, n. 1, p. 67–83, 2013.
Bernabucci, U.; Calamari, L. Effects of heat stress on bovine milk yield and composition. Zootecnica e
Nutrizione Animale, v. 24, p. 247–257, 1998.
Bernabucci, U.; Lacetera, N.; Ronchi, B.; Nardone, A. Effects of the hot season on milk protein fractions in
Holstein cows. Anim. Res. 51: 25-33. 2002.
Botaro, B.; Santos, M. V. Entendendo a variação da crioscopia do leite - Radar Técnico - Qualidade do Leite -
MilkPoint. Disponível em: <https://www.milkpoint.com.br/radar-tecnico/qualidade-do-leite/entendendo-a-
variacao-da-crioscopia-do-leite-46948n.aspx>. Acesso em: 24 ago. 2017.
Bowman, M; Gill, R.; Reynolds, C. Significance of Nutritional Effects on the Freezing Point of Milk. Ontario
Agri-Business Association Nutrition Committee; 2005.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa nº 37, de 18 de abril de 2002.
Diário Oficial da República Federativa do Brasil, Brasília, p. 8, 24 de abril. 2002a. Seção 1.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa nº 51, de 18 de setembro de
2002. Diário Oficial da República Federativa do Brasil, Brasília, p. 13, 20 de set. 2002b. Seção 1.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa nº 62, de 29 de dezembro de
2011. Diário Oficial da República Federativa do Brasil, 31 dez. 2011a. Seção 1, p. 6.
Brasil. Método de Ensaio - Depressão do Ponto de Congelamento do Leite Fluido. v. 10-01–01, p. 1–7, 2011b.
Brasil. Determinação de acidez titulável em leite fluido. p. 4–7, 2013.
Brasil. Pesquisa de cloretos em Leite Fluido por Colorimetria. p. 1–4, 2014.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa nº 7, de 03 de maio de 2016.
Diário Oficial da República Federativa do Brasil, Brasília, n. 84, p. 18, 03 de mai. 2016. Seção 1.
Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Decreto Nº 9.013, DE 29 de março de 2017. Diário
Oficial da República Federativa do Brasil, Brasília, 2017.
Bueno, V. F. F.; Mesquita, A. J.; Oliveira, A. N.; Nicolau, E. S.; Neves, R. B. S. Contagem bacteriana total do
leite: relação com a composição centesimal e período do ano no Estado de Goiás. Revista Brasileira de Ciência
Veterinária, v. 15, n. 1, p. 40–44, 2008.
Cardozo, L.L. et al. Risk factors for the occurrence of new and chronic cases of subclinical mastitis in dairy
herds in southern Brazil. Journal of Dairy Science, v. 98, n. 11, p. 7675–7685, 2015.

295
Córdova, H.A. Produção e qualidade do leite, frequência de ordenha, profundidade de úbere e comportamento
de vacas da raça holandesa em ordenha robótica. 2016. 142 f. Tese (Doutorado em Ciência Animal) –
Universidade do Estado de Santa Catarina. Centro de Ciências Agroveterinárias, Lages, SC, 2016.
CSIRO. Nutrient requirements of domesticated ruminants. CSIRO Publishing, 2007.
Della Libera, A. M. M. P. et al. Características físico-químicas e microbiológicas do leite de vacas sem
alterações ao exame físico da glândula mamária e com alta contagem de células somáticas. Rev. Bras. Saúde
Prod. Anim., v. 1, n. 2, p. 42–47, 2001.
Fagnani, R.; Beloti, V.; Battaglini, A. P. P. Acid-base balance of dairy cows and its relationship with alcoholic
stability and mineral composition of milk. Pesquisa Veterinária Brasileira, v. 34, n. 5, p. 398–402, 2014.
Felipus, N. C. Impacto do transporte a granel na qualidade microbiológica e físico-química e na composição do
leite cru refrigerado em indústria de laticínios. 2017. 93p. Dissertação (Mestrado em Ciência Animal – Área:
Produção Animal) – Universidade do Estado de Santa Catarina. Programa de Pós-graduação em Ciência
Animal, Lages, 2017.
Fischer, V.; Ribeiro, M. E. R.; Zanela, M. B.; et al. Leite instável não ácido: Um problema solucionável? Revista
Brasileira de Saúde e Produção Animal, v. 13, n. 3, p. 838–849, 2012.
Fox, P. F.; Sweeney, P. L. H. Dairy Chemistry and Biochemistry. London: Thomson Science; 1998; 478p.
Gabbi, A.M. et al. Typology and physical-chemical characterization of bovine milk produced with different
productions strategies. Agr. Syst.., v.121, p. 130-134, 2013.
Gabbi, A.M. et al. Milk traits of lactating cows submitted to feed restriction. Tropical Animal Health and
Production, v. 48, n. 1, p. 37-43, 2016.
Gonzalez, H. et al. Avaliação da qualidade do leite na bacia leiteira de Pelotas, RS. Efeito dos meses do ano.
Revista Brasileira de Zootecnia, v. 33, n. 6, p. 1531–1543, 2004.
Hagnestam-Nielsen, C.; Emanuelson, U.; Berglund, B.; Strandberg, E. Relationship between somatic cell count
and milk yield in different stages of lactation. Journal of Dairy Science, v.92, n.7, p.3124-33, 2009
Henno, M.; Ots, M.; Jõudu, I.; Kaart, T.; Kärt, O. Factors affecting the freezing point stability of milk from
individual cows. International Dairy Journal, v. 18, n. 2, p. 210–215, 2008.
Horne, D. S. Ethanol Stability and Milk Composition. Advanced Dairy Chemistry, v. 1B, p. 225–246, 2016.
Huppertz, T. Heat Stability of Milk. In: S.S.B. Media (Ed.); Advanced Dairy Chemistry. p. 179–196, 2016.
New York.
Ishler, V.; Heinrichs, J.; Varga, G. From feed to milk: Understanding rumen function. Pennsylvania State
University, 1996. Disponível em: http://extension.psu.edu/animals/dairy/nutrition/nutrition-and-
feeding/rumen-function/from-feed-to-milk-understanding-rumen-function
João, J. H.; Lima-Rosa, C. A. V.; Neto, A. T.; Picinin, L. C. A.; Fuck, J. J.; Marin, G. Qualidade da água
utilizada na ordenha de propriedades leiteiras do Meio Oeste Catarinense, Brasil. Revista de Ciências
Agroveterinárias, v. 10, n. 1, p. 9–15, 2011.
Kitchen, B. Y. B. J. Review of the progress of Dairy Science: Bovine mastitis: milk compositional changes and
related diagnostic tests. J. Dairy Sci., v. 48, p. 167–188, 1981.
Marques, L. T. et al. Produção leiteira, composição do leite e perfil bioquímico sanguíneo de vacas lactantes
sob suplementação com sal aniônico. Revista Brasileira de Zootecnia, v. 40, n. 5, p. 1088-1094, 2011.
Marques, L. T.; Zanela, M. B.; Ribeiro, M. E. R.; Stumpf Júnior, W.; Fischer, V. Ocorrência do leite instável
ao álcool 76% e não ácido (LINA) e efeito sobre os aspectos físico-químicos do leite. Revista Brasileira de
Agrociencia, v. 13, n. 1, p. 91–97, 2007.
Mattiello, C. A. Avaliação do rendimento industrial, atributos microbiológicos e físico-químicos de queijo
colonial produzido a partir de leite com dois diferentes níveis de células somáticas. 2015. 106 f. Dissertação
(Mestrado em Ciência Animal) Universidade do Estado de Santa Catarina. Centro de Ciências Agroveterinárias,
Lages, SC, 2015.

296
Mazal, G. et al. Effect of somatic cell count on prato cheese composition. Journal of Dairy Science, v. 90, n. 2,
p. 630–636, fev. 2007.
Murphy, S. C. et al. Influence of raw milk quality on processed dairy products: How do raw milk quality test
results relate to product quality and yield? Journal of Dairy Science, v. 99, n. 12, p. 10128–10149, 2016.
Ogola, H.; Shitandi, A.; Nanua, J. Effect of mastitis on raw milk compositional quality. Journal of Veterinary
Science, v. 8, n. 3, p. 237–242, 2007.
Oliver, S P; Calvinho, L F. Influence of inflammation on mammary gland metabolism and milk composition.
Journal of Animal Science, v. 73, suppl. 2, p. 18, 1995.
Oliveira, L.C.; Gomes, M.F.; Velloso, C.R.V. Modernização da legislação sanitária federal sobre leite e
derivados. In: Castro, M.C.D.; Portugal, J.A.B. Perspectivas a Avanços em Laticínios. Juiz de Fora. EPAMIG.
Centro Tecnológico da Zona da Mata, ILCT,2000. 278 p.
Pagno, K. C. A.; Jesuz, E. R.; Carneiro M. G. B.; Pagno, J. T. Análise do índice crioscópico do leite de vacas
expostas à mudança no manejo hídrico. In: Congresso Brasileiro de Qualidade do Leite, 5, 2013, Águas de
Lindóia. Anais... Botucatu: Faculdade de Medicina Veterinária e Zootecnia, 2013.
Pessoa, R. B.; Blagitz, M. G.; Batista, C. F.; Santos, B. P. et al. Avaliação da apoptose de leucócitos polimorfo
nucleares CH138+ em leite bovino de alta e baixa contagem de células somáticas dados preliminares. Arq. Bras.
Med. Vet. Zootec, Belo Horizonte, v. 64, n. 3, p. 533-539, 2012.
Raimondo, R. F. S.; Birgel, D. B.; Saut, J. P. E.; Komninou, E. R.; Birgel Junior, E. H. Teores de cloretos,
lactose e índice cloretos/lactose na secreção láctea de bovinos da raça Jersey no primeiro mês da lactação.
Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 61, n. 1, p. 259–265, 2009.
Reche, N. L. M.; Neto, A. T.; D`Ovideo, L.; Felipus, N. C.; Pereira, L. C.; Cardozo, L. L.; Lorenzetti, R. G.;
Picinin, L. C. A. Multiplicação microbiana no leite cru armazenado em tanques de expansão direta. Ciência
Rural, p. 828–834, 2015.
Rhoads, M. L., Rhoads, R. P., Van Baale, M. J., Collier, R. J. et al. Effects of heat stress and plane of nutrition
on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci.
2009; 92:1986–1997.
Ribeiro Neto, A. C.; Barbosa, S. B. P.; Jatobá, R. B.; Silva, A. M.; Silva, C. X.; Silva, M. J. A.; Santoro, K. R.
Qualidade do leite cru refrigerado sob inspeção federal na região Nordeste. Arquivo Brasileiro de Medicina
Veterinária e Zootecnia, v. 64, n. 5, p. 1343-1351, 2012
Santos, M. V. Fatores que podem alterar a crioscopia do leite - MilkPoint. Disponível em:
<https://www.milkpoint.com.br/mypoint/6239/p_fatores_que_podem_alterar_a_crioscopia_do_leite_4319.asp
x>. Acesso em: 24 ago. 2017.
Santos, M. V.; Fonseca, L. F. L. Estratégias para controle de mastite e melhoria da qualidade do leite. São
Paulo: Manole, 2007. 314 p.
Sanvido, V. Efeito do tempo de armazenamento do leite cru e da temperatura de estocagem do leite pasteurizado
sobre sua vida de prateleira. 2007. 78 f. Universidade Estadual de Campinas, 2007.
Shipe, W. F. The freezing point of milk. A review. Journal of Dairy Science, v. 42, n. 11, p. 1745–1762, 1959.
Schmidt, F. A. Efeito do suprimento das exigências de energia e/ou proteína na recuperação da instabilidade do
leite ao teste do álcool, 2014. 78p. Dissertação (Mestrado em Ciência Animal) – Centro de Ciências
Agroveterinárias, Universidade do Estado de Santa Catarina, Lages, 2014.
Schukken, Y. H.; Fulton, C. D.; Leslie, K. E. N. E. Freezing point of bulk milk in Ontario - An observational
study. Journal of Food Protection, v. 55, n. 12, p. 995–998, 1992.
Shwartz, G.; Rhoads, M. L.; Van Baale, M. J.; Rhoads, R. P.; Baumgard, L. H. Effects of a supplemental yeast
culture on heat-stressed lactating Holstein cows. J. Dairy Sci. v.92, n. 3, p. 935-942, 2009.
Silva, C. G.; Alessio, D.R.M.; Knob, D. A.; D'Ovidio, L.; Thaler Neto, A. Influência da sanificação da água e
das práticas de ordenha na qualidade do leite. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 70,
p. 615-622, 2018.

297
Stumpf, M. T. et al. Severe feed restriction increases permeability of mammary gland cell tight junctions and
reduces ethanol stability of milk. Animal, v. 7, n. 7, p. 1137-1142, 2013.
Sundekilde, U. K.; Frederiksen, P. D.; Clausen, M. R.; Larsen, L. B.; Bertram, H. C. Relationship between the
metabolite profile and technological properties of bovine milk from two dairy breeds elucidated by NMR-based
metabolomics. Journal of Agricultural and Food Chemistry, v. 59, n. 13, p. 7360–7367, 2011.
Tian, H.; Wang, W. Y.; Zheng, N.; Cheng, J. B. et al. Identification of diagnostic biomarkers and metabolic
pathway shifts of heat-stressed lactating dairy cows. J. Proteomics. 2015; 125: 17-28.
Tomazi, T. et al. Bovine subclinical intramammary infection caused by coagulase-negative staphylococci
increases somatic cell count but has no effect on milk yield or composition. Journal of dairy science, v. 98, n.
5, p. 3071–8, 2015.
Töpel, A. Chemie und Physik der Milch. Naturstoff – Rohstoff – Lebensmittel. Hamburg: Behr's Verlag; 2015;
758p.
Tsioulpas, A.; Lewis, M. J.; Grandison, A. S. Effect of minerals on casein micelle stability of cows’ milk.
Journal of Dairy Research, v. 74, n. 2, p. 167–173, 2007.
Urech, E.; Puhan, Z.; Schällibaum, M. Changes in milk protein fraction as affected by subclinical mastitis.
Journal of Dairy Science, v. 82, n. 11, p. 2402–2411, 1999.
Varnam, A.; Sutherland, J. P. Milk and milk products: technology, chemistry and microbiology. v. 1.
Gaithersburg, Maryland: Aspen Publication; 2001; 453p.
Zanela, M. B.; Fischer, V.; Ribeiro, M. E. R.; et al. Leite instável não-ácido e composição do leite de vacas
Jersey sob restrição alimentar. Pesquisa Agropecuária Brasileira, v. 41, n. 5, p. 835–840, 2006.
Werncke, D. et al. Qualidade do leite perfil das propriedades leiteiras no sul de Santa Catarina: abordagem
multivariada. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 68, n. 2, p. 506–516, 2016.
Wheelock, J. B; Rhoads, R. P.; Van Baale, M. J.; Sandders, S. R. et al. Effects of heat stress on energetic
metabolism in lactating Holstein cows. J. Dairy Sci. v.93, n. 2, p.644-655, 2010.
Winck, C. A. Impactos do pagamento pela qualidade na cadeia produtiva do leite na região Oeste de Santa
Catarina. 119 f., 2012. Tese (Doutorado em Agronegócios), Programa de Pós-Graduação em Agronegócios -
Centro de Estudos e Pesquisas em Agronegócios da Universidade Federal do Rio Grande do Sul (UFRGS),
2012.

298
Relação entre metabolismo e reprodução em vacas leiteiras
Álan Maia Borges
Telma da Mata Martins
Ana Carolina Leite
Philipe Pimenta Nunes
Nos últimos anos, verifica-se elevado crescimento da produtividade e do tamanho dos
rebanhos leiteiros, visando atender a crescente demanda mundial de leite e seus derivados.
Em paralelo, nota-se redução da eficiência reprodutiva, o que compromete o desempenho
atual dos laticínios, uma vez que a produção de leite depende da capacidade da vaca se tornar
gestante. As adaptações metabólicas para a alta produção de leite e a elevada ocorrência de
doenças puerperais estão diretamente relacionadas com a redução da fertilidade (LUCY,
2001). O objetivo dessa revisão é descrever os mecanismos pelos quais o metabolismo pode
estar relacionado com problemas reprodutivos de vacas de alta produção.
Alterações metabólicas e reprodutivas em vacas leiteiras
Durante as últimas semanas que antecedem o parto pode-se verificar alterações metabólicas
e hormonais que visam adaptar o organismo para o início da lactação. À medida que o parto
se aproxima as concentrações de glicocorticoides, somatotropina e prolactina e estrógeno
aumentam e, por outro lado, a concentração de progesterona diminui (Figura 1; BELL, 1995).
Este período de periparto das vacas leiteiras é caracterizado por diminuição da ingestão de
alimentos e aumento súbito na demanda energética, imposta pelo início de lactação. Vacas
de alta produção recém-paridas, geralmente, apresentam redução mais acentuada da ingestão
de alimentos e depressão do sistema imunológico, tornando-se altamente suscetíveis ao
estabelecimento de doenças após o parto (Butler, 2006; Sordillo, 2009). A adoção de
programas de controle, a identificação precoce do animal doente e a adoção de cuidados para
promover a sua recuperação são primordiais para minimizar as consequências na reprodução
e na produção (Van Saun, 2000; Smith e Risco, 2005).
A maior produtividade dos rebanhos leiteiros tem aumentado significativamente a demanda
de nutrientes para a síntese de leite. Quando o animal não apresenta aumento da ingestão de
alimentos para atender essa demanda, é necessário mobilizar reservas corporais (Grummer,
1995). Esse mecanismo compensatório pode gerar distúrbios metabólicos devido à redução

299
drástica da ingestão de alimentos ocorrida no periparto de vacas de alta produção (Figura 2;
Bertics et al., 1992).
Figura 1. Mudanças na concentração sérica de hormônios envolvidos na
homeorrese de vacas leiteiras no periparto (adaptado de Bell, 1995)
Figura 2. Consumo de matéria seca (CMS) no periparto de vacas leiteiras
(adaptado de Bertics et al., 1992)

300
Muitas evidências confirmam que os hormônios metabólicos, como a somatotropina (GH), a
insulina, os fatores de crescimento semelhantes à insulina (IGFs I e II), a leptina, dentre
outros, exercem importantes papéis no controle da função reprodutiva dos mamíferos, agindo
como mediadores ou sinalizadores dos efeitos da ingestão de alimentos e do balanço
energético sobre a fertilidade de bovinos (GIL, 2003). Seguem abaixo algumas considerações
sobre a influência de alterações metabólicas na reprodução das vacas leiteiras.
O aumento da produção de leite está relacionado com a elevação da concentração de GH,
hormônio catabólico. Por meio dos seus efeitos lipolíticos e diabetogênicos, o GH favorece
a disponibilidade de precursores para a síntese do leite, pois aumenta a mobilização de ácidos
graxos do tecido adiposo e reduz a utilização de glicose pelos tecidos periféricos ao diminuir
a sensibilidade dos mesmos à ação da insulina (Peel e Bauman, 1987; Bell, 1995). A redução
da sensibilidade pancreática à glicose implica queda na secreção de insulina (Riis, 1983).
Quando a mobilização de tecido adiposo é elevada, a taxa de lipólise pode sobrepassar a de
lipogênese, liberando maior quantidade de ácidos graxos não esterificados (AGNE) para a
circulação. A concentração plasmática de AGNE começa a aumentar na última semana da
gestação, podendo ser utilizado como fonte de energia pela glândula mamária e outros
órgãos. No fígado, os AGNE podem ser oxidados para produção de energia, podem originar
corpos cetônicos (principalmente beta-hidroxibutirato, -OHB), ou podem ser esterificados
com gliceróis e exportados como triglicerídeos. O fígado dos ruminantes possui capacidade
limitada para exportar triglicerídeos, sendo que a maior mobilização em relação à baixa
exportação leva ao acúmulo hepático de gordura, predispondo o animal a doenças
metabólicas (Bell e Bauman, 1996; Head e Gulay, 2001). Vacas obesas tendem a apresentar
redução mais acentuada na ingestão de alimentos e maior mobilização de tecido adiposo no
pós-parto, portanto, são mais suscetíveis às ocorrências de esteatose hepática e cetose (Reid
et al., 1986).
A queda na ingestão de matéria seca com consequente redução das concentrações séricas de
glicose e insulina, aliados à intensa mobilização de AGNE no período pós-parto em vacas de
alta produção, são responsáveis pela conformação de quadro de estresse metabólico,
resultando em processo inflamatório sistêmico denominado inflamação metabólica (Leblanc,
2014). A mobilização de AGNE do tecido adiposo, por meio da ação do GH, também é
acompanhada pela liberação das citocinas pró-inflamatórias, de fator de necrose tumoral α

301
(TNFα) e interleucina 6 (IL-6) (Ingvartsen e Moyes, 2013). Além disso, os próprios AGNE
são capazes de se ligarem a receptores celulares de reconhecimento de padrões microbianos,
principalmente ao receptor tipo-toll 4 (TLR4), distribuídos em diversas células e tecidos do
organismo tais como fígado, hipotálamo, ovários, útero, dentre outros. Ao se ligar a tais
receptores, os AGNE deflagram resposta inflamatória por meio da indução da síntese de
citocinas como TNFα, IL-1β, IL-6 (Hotamisligil e Erbay, 2008). TNFα e IL-1β são
responsáveis por bloquear a via de sinalização intracelular da insulina, bloqueando assim sua
ação sobre as células do organismo e exacerbando o quadro de resistência periférica a esse
hormônio. IL-6 e IL-1β são responsáveis por induzir a produção de proteínas de fase aguda
pelo fígado, amplificando o quadro sistêmico de inflamação metabólica (Bertoni et al., 2006;
Trevisi et al., 2012). O estresse metabólico, associado ao desenvolvimento de resistência
periférica à insulina, é um fator que contribuem para a redução da imunidade no período de
transição de vacas de alta produção, predispondo-as à diversas afecções no período pós-parto,
prolongando a duração do período do parto à nova concepção (Leblanc, 2014).
As alterações metabólicas decorrentes da intensa mobilização de reservas corporais e o
prolongamento do período de balanço energético negativo podem atrasar a ocorrência da
primeira ovulação após o parto. A influência do balanço energético negativo no retorno da
atividade ovariana luteal cíclica é devida ao atraso no aumento da frequência e amplitude dos
pulsos de gonadotropinas (FSH e LH) na presença de altas concentrações de GH, AGNE e
corpos cetônicos, e baixas concentrações de insulina, glicose e IGF-I (Figura 3; Bell e
Bauman, 1996). O GH regula a síntese e a secreção hepática de IGF-I (Gong, 2002). Apesar
da elevada concentração de GH circulante no início da lactação, o fígado encontra-se
refratário à ação desse hormônio, implicando redução da concentração plasmática de IGF-I
(Figura 4; Spicer et al., 1990). O desacoplamento do eixo somatotrópico ocorrido durante o
período de balanço energético negativo está relacionado com a condição de hipoinsulinemia
(Spicer et al., 1990; Lucy, 2003).
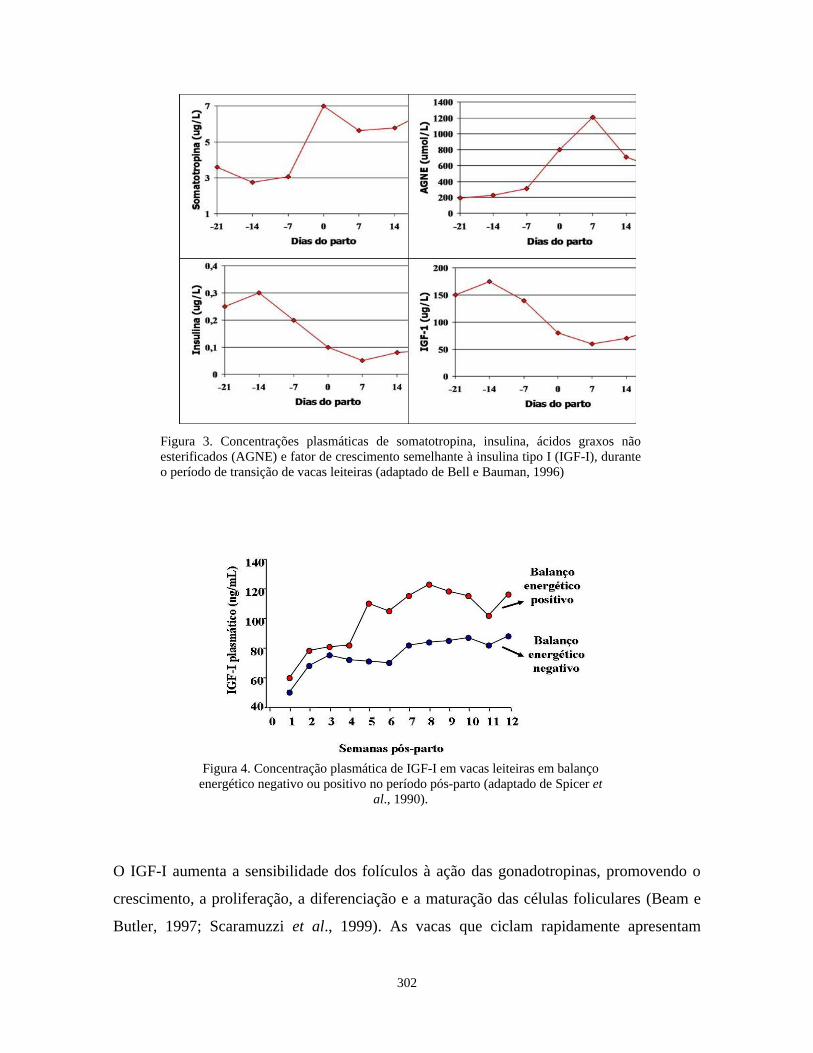
302
Figura 3. Concentrações plasmáticas de somatotropina, insulina, ácidos graxos não
esterificados (AGNE) e fator de crescimento semelhante à insulina tipo I (IGF-I), durante
o período de transição de vacas leiteiras (adaptado de Bell e Bauman, 1996)
Figura 4. Concentração plasmática de IGF-I em vacas leiteiras em balanço
energético negativo ou positivo no período pós-parto (adaptado de Spicer et
al., 1990).
O IGF-I aumenta a sensibilidade dos folículos à ação das gonadotropinas, promovendo o
crescimento, a proliferação, a diferenciação e a maturação das células foliculares (Beam e
Butler, 1997; Scaramuzzi et al., 1999). As vacas que ciclam rapidamente apresentam

303
aumento precoce da concentração plasmática de IGF-I, enquanto as vacas em anestro
prolongado não exibem aumento da concentração de IGF-I até a sétima semana após o parto
(Figura 5; Beam e Butler, 1997).
Figura 5. Concentração plasmática média de IGF-I em vacas leiteiras
ovulando ou não no período pós-parto (adaptado de Beam e Butler, 1997).
De maneira semelhante ao IGF-I, a insulina exerce importante papel na resposta ovariana à
ação das gonadotropinas, controlando o desenvolvimento folicular. A insulina atua por meio
da ligação a receptores específicos localizados nas células da teca e da granulosa. Dentre
outros efeitos metabólicos, o estímulo do transporte de glicose para o interior das células é
um dos mais importantes, pois esse metabólito é a principal fonte energética para os folículos
(Rabiee et al., 1997). O período de balanço energético negativo é caracterizado pela redução
na concentração circulante de insulina (Sinclair et al., 2002). A baixa concentração
plasmática de insulina pode implicar redução na produção de androstenediona e estradiol,
comprometendo a capacidade dos folículos adquirirem receptores de LH e impedindo a
ocorrência da ovulação (Stewart et al., 1995). Além de agir diretamente nos ovários, a
insulina influencia na liberação do LH (Monget e Martin, 1997). Vacas leiteiras alimentadas
com dietas que promovem aumento da concentração de insulina no pós-parto apresentam
menor período de serviço (Gong, 2002).
Mudanças nutricionais agudas e crônicas e redução da condição corporal implicam em
diminuição da síntese e secreção de leptina, hormônio peptídico produzido pelos adipócitos,

304
que age diretamente sobre o hipotálamo, regulando a ingestão de alimentos, o balanço
energético, a termogênese e a fertilidade. Em vacas leiteiras, a concentração de leptina
encontra-se positivamente correlacionada com as concentrações de insulina e glicose, e
negativamente correlacionada com as concentrações de GH e AGNE. Durante o período de
balanço energético negativo, ocorre diminuição na síntese e na secreção de leptina, o que
contribui para a inibição da secreção de GnRH. Após a recuperação da condição corporal, o
aumento da concentração de leptina circulante estimula a secreção de GnRH e auxilia no
retorno da atividade ovariana luteal cíclica (Williams et al., 1999; Williams, 2001).
Relação dos distúrbios metabólicos e reprodutivos em vacas leiteiras de alta produção
A partir da década de 1990, os estudos sobre a redução da fertilidade das vacas leiteiras de
alta produção começaram a relacionar as alterações ocorridas no desenvolvimento folicular
durante o período de balanço energético negativo, com a menor qualidade dos oócitos e
embriões, e, consequentemente, com a diminuição das taxas de concepção (Britt, 1992;
O’Callanghan e Boland, 1999). A menor qualidade dos oócitos e embriões é resultante das
condições adversas às quais os folículos foram expostos durante o balanço energético
negativo, já que a foliculogênese pode durar até 60 dias. A produção de progesterona pelo
corpo lúteo formado a partir do folículo ovulatório exposto a condições adversas também se
encontra comprometida (Figura 6; Britt, 1992).
O período de balanço energético negativo é caracterizado pelos baixos níveis de glicose,
insulina e IGF-I e pela menor frequência dos pulsos de LH. Em conjunto, estes fatores
limitam a produção de estrógeno pelo folículo dominante, diminuindo a qualidade dos
oócitos, a capacidade de desenvolvimento do embrião e a concentração plasmática de
progesterona (Butler, 2003). Os estudos de Sartori (2002) e Sartori et al. (2002)
demonstraram que os embriões de novilhas e vacas secas são de melhor qualidade que os
embriões de vacas em lactação, o que pode justificar, pelo menos em parte, a subfertilidade,
as menores taxas de concepção e a alta mortalidade embrionária precoce em animais
lactantes.

305
Figura 6. Os folículos ovulatórios no pós-parto foram numerados de 1 a 6 e cada um
originou um corpo lúteo. O primeiro e o segundo folículos ovulatórios começaram seu
desenvolvimento durante a lactação anterior ou no período seco, quando as condições
metabólicas eram desejáveis. Ao contrário, o terceiro, o quarto e o quinto folículos
desenvolveram durante o período de balanço energético negativo e, consequentemente,
foram expostos aos efeitos prejudiciais das alterações metabólicas ocorridas no pós-
parto (adaptado de Britt, 1992).
Existem correlações significativas entre a constituição plasmática e o fluido intrafolicular,
quanto à presença de glicose, AGNE, -OHB, dentre outros. As alterações metabólicas
ocorridas durante o período de balanço energético negativo modificam a composição do
fluido intrafolicular, afetando a qualidade das células da granulosa. As células da granulosa
são de extrema importância para a esteroidogênese e o controle dos processos reprodutivos.
As alterações metabólicas também são prejudiciais para as células do cumulus oophorus e
para o oócito (Leroy, 2005, Vanholder et al., 2005). A menor qualidade dos oócitos e o
comprometimento ao desenvolvimento embrionário podem ser resultados de alterações no
desenvolvimento folicular, no ambiente uterino e nas tubas. Os embriões de vacas leiteiras
de alta produção contêm 45% a mais de lipídeos que os de vacas secas. O maior conteúdo
lipídico está associado com a alta sensibilidade embrionária ao estresse oxidativo, menor
função mitocondrial e menor capacidade de sobreviver após o congelamento (Figura 7;
Leroy, 2005).

306
Figura 7. Esquema dos possíveis mecanismos pelas quais a qualidade do embrião pode
ser prejudicada em vacas leiteiras de alta produção (adaptado de Leroy, 2005)
NEFA = ácidos graxos não esterificados; β-OHB = beta-hidroxibutirato
Nos últimos anos, outros pesquisadores também têm buscado estudar o impacto dessas
alterações metabólicas relacionadas ao balanço energético negativo e a fertilidade de vacas
de alta produção, deixando de se concentrarem apenas no ambiente ovariano e/ou uterino e
tentando investigar outras porções da genitália tubular feminina. Assim, as concentrações de
AGNE no sangue são conhecidas por alterarem o desenvolvimento folicular e, atualmente,
verifica-se que também podem, muito provavelmente, modificar o microambiente das tubas
uterinas. O oócito no final do período de maturação dentro do folículo pré-ovulatório e o
embrião nos primeiros dias de clivagem celular são bastante sensíveis às mudanças
metabólicas e, por isso o estresse metabólico pode comprometer a fertilidade pela atuação
nos microambientes foliculares e tubáricas. A exposição a AGNE reduz a proliferação
celular, a migração celular e a capacidade de ligação ao espermatozóide, porém, as células
da tuba uterina parecem desempenhar papel ativo na regulação das concentrações luminais
de AGNE por meio da maior permeabilidade, acúmulo de lipídeos intracelulares e no
metabolismo de ácidos graxos, o que poderia minimizar os efeitos negativos dos AGNE sobre
o embrião (Jordens et al., 2017; Leroy, 2017). As mudanças ocorridas na fisiologia
reprodutiva das vacas leiteiras de alta produção também podem estar relacionadas com a
maior intensidade do metabolismo dos hormônios esteroides no fígado (Wiltbank et al.,
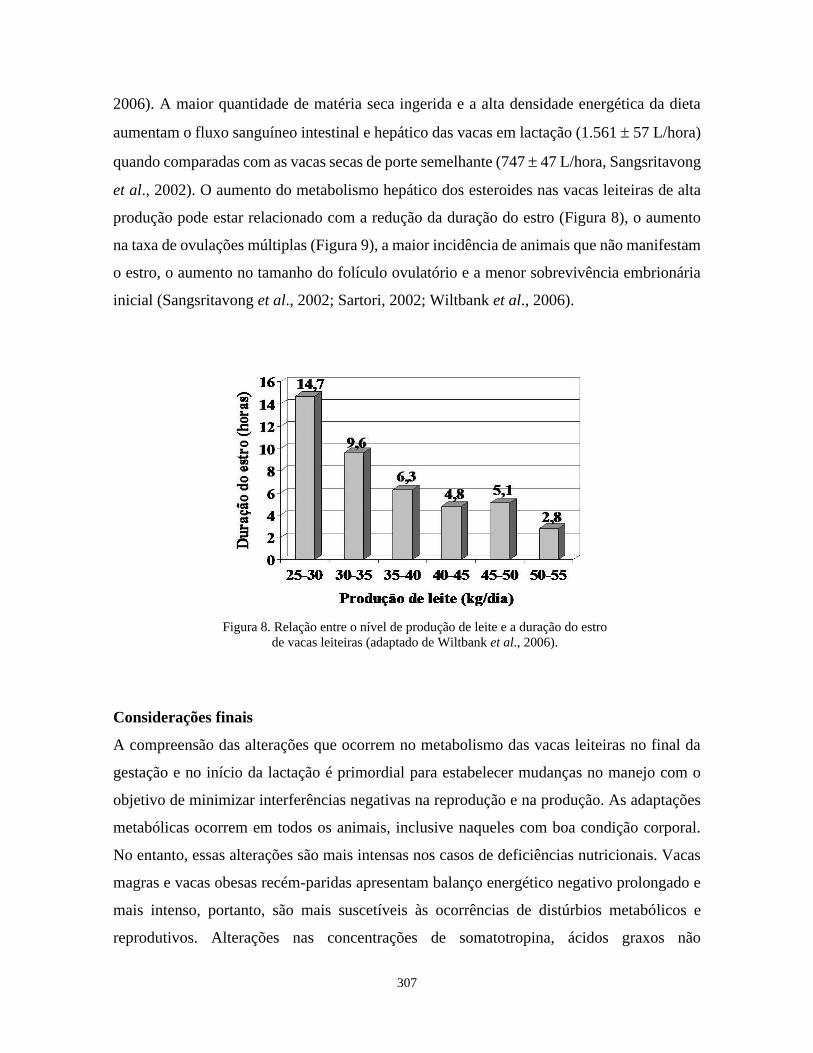
307
2006). A maior quantidade de matéria seca ingerida e a alta densidade energética da dieta
aumentam o fluxo sanguíneo intestinal e hepático das vacas em lactação (1.561 57 L/hora)
quando comparadas com as vacas secas de porte semelhante (747 47 L/hora, Sangsritavong
et al., 2002). O aumento do metabolismo hepático dos esteroides nas vacas leiteiras de alta
produção pode estar relacionado com a redução da duração do estro (Figura 8), o aumento
na taxa de ovulações múltiplas (Figura 9), a maior incidência de animais que não manifestam
o estro, o aumento no tamanho do folículo ovulatório e a menor sobrevivência embrionária
inicial (Sangsritavong et al., 2002; Sartori, 2002; Wiltbank et al., 2006).
Figura 8. Relação entre o nível de produção de leite e a duração do estro
de vacas leiteiras (adaptado de Wiltbank et al., 2006).
Considerações finais
A compreensão das alterações que ocorrem no metabolismo das vacas leiteiras no final da
gestação e no início da lactação é primordial para estabelecer mudanças no manejo com o
objetivo de minimizar interferências negativas na reprodução e na produção. As adaptações
metabólicas ocorrem em todos os animais, inclusive naqueles com boa condição corporal.
No entanto, essas alterações são mais intensas nos casos de deficiências nutricionais. Vacas
magras e vacas obesas recém-paridas apresentam balanço energético negativo prolongado e
mais intenso, portanto, são mais suscetíveis às ocorrências de distúrbios metabólicos e
reprodutivos. Alterações nas concentrações de somatotropina, ácidos graxos não

308
esterificados, insulina, glicose, fator de crescimento semelhante à insulina tipo I, leptina,
dentre outros hormônios e metabólitos, estão relacionadas com as ocorrências de distúrbios
reprodutivos, principalmente em vacas de alta produção, devido às interferências no eixo
hipotalâmico-hipofisário-ovariano e no eixo somatotrópico. A redução da fertilidade desses
animais é decorrente das influências na qualidade dos folículos, dos oócitos, do corpo lúteo
e do ambiente uterino, que resultam em baixas taxas de concepção e menor sobrevivência
embrionária. É necessário realizar mais estudos com vacas mestiças leiteiras, pois esses
animais apresentam particularidades ainda não esclarecidas em relação às ocorrências de
distúrbios metabólicos e reprodutivos.
Figura 9. Relação entre o nível de produção de leite e a incidência de
ovulações múltiplas em vacas leiteiras (adaptado de Wiltbank et al., 2006).
Referências
Artunduaga, M. A. T. Efeito de dietas com fontes lipídicas e gliconeogênicas no período de transição de
primíparas leiteiras sobre: perfil metabólico, produção de leite e reprodução. 2010. 96p. Tese (Doutorado em
Zootecnia) - Escola de Veterinária da UFMG, Belo Horizonte, MG.
Beam, S. W.; Butler, W. R. Energy balance and ovarian follicle development prior to first ovulation postpartum
in dairy cows receiving three levels of dietary fat. Biol. Reprod., v. 56, p. 133-142, 1997.
Bell, A. W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation.
J. Anim. Sci., v.73, p. 2804-2819, 1995.
Bell, A. W.; Bauman, D. E. The transition cow: actualized homeorhesis. In: Cornell Nutrition Conference for
Feed Manufacturers, 1996, Nova York. Anais…. Nova York, p. 150-157, 1996.
Bertics, S. J.; Grummer, R. R.; Cadorniga-Valino, C. et al. Effect of prepartum dry matter intake on liver
triglyceride concentration and early lactation. J. Dairy Sci., v. 75, p. 1914-1922, 1992.

309
Bertoni, G.; Trevisi, E.; Ferrari, A. R. et al. The dairy cow performances can be affected by inflammations
occurring around calving. In: 57th EAAP Meeting, 2006, Antalya, Turquia. P. 325, 2006.
Britt, J. H. Reproductive performance as affected by nutrition and environment. In: III Florida Ruminant
Nutrition Symposium, 1992, Orlando, Anais… Orlando, p. 93-101, 1992.
Butler, W. R. Efeito do balanço energético negativo na fertilidade de vacas leiteiras. In: X Curso Novos
Enfoques na Produção e Reprodução de Bovinos, 2006, Uberlândia. Anais...Uberlândia, 2006.
Butler, W. R. Energy balance relationships with follicular development, ovulation and fertility in postpartum
dairy cows. Liv. Prod. Sci., v.83, p. 211-218, 2003.
Butler, W. R. Nutrition, negative energy balance and fertility in the postpartum dairy cow. Cattle Pract., v. 13,
p. 13-18, 2005.
Gil, C. V. Effect of nutrition on follicle development and ovulation rate in the ewe. 2003. 56p. Tese (Doutorado
em Medicina Veterinária) – Faculdade de Medicina Veterinária, Uppsala.
Gong, J. G. Influence of metabolic hormones and nutrition on ovarian follicle development in cattle: practical
implications. Dom. Anim. Endocrinol., v. 23, p. 229-241, 2002.
Grimard, B.; Humblot, P.; Ponter, A. A. et al. Influence of postpartum energy restriction on energy status,
plasma LH and estradiol secretion and follicular development in suckled beef cows. J. Reprod. Fert., v. 104, p.
173-179, 1995.
Grummer, R. R. Impact of changes in organic nutrients metabolism on feeding the transition cow. J. Anim. Sci.,
v. 73, p. 2820-2833, 1995.
Head, H. H.; Gulay, M. S. Recentes avanços na nutrição de vacas no período de transição. In: II Simpósio
Internacional de Bovinocultura de Leite: Novos Conceitos Em Nutrição, 2001, Lavras. Anais... Lavras, p. 121-
137, 2001.
Hotamisligil, G. S.; Erbay, E. Nutrient sensing and inflammation in metabolic diseases. Nat. Rev. Immunol., v.
8, p. 923-934, 2008.
Ingvartsen, K. L.; Moyes, K. M. Nutrition, immune function and health of dairy cattle. Animal, v. 7, s. 1, p.
112-122, 2013.
Jordaens L, Van Hoeck, V.; Maillo, V. et al., Maternal metabolic stress may affect oviduct gatekeeper function.
Reproduction. doi: 10.1530/REP-16-0569, 2017
Leblanc, S. J. Reproductive tract inflammatory disease in postpartum dairy cows. Animal, v. 8, s. 1, p. 54-63,
2014.
Leroy, J. L.; Bie, J.; Jordaens, L. et al. Negative energy balance and metabolic stress in relation to oocyte and
embryo quality: na update on possible pathways reducing fertility in dairy cows. Proceedings of the 31th Annual
Meeting of the Brazilian Embryo Technology Society (SBTE). Anim. Reprod., v.14, n.3, p.497-506, 2017.
Leroy, J.L.M.R. Metabolic changes in high producing dairy cows and the consequences on oocyte and embryo
quality. 2005. 252 p. Tese (Ph.D. em Reprodução, Obstetrícia e Saúde de Rebanho) – Escola de Veterinária da
Universidade de Ghent, Ghent.
Lucy, M. C. Physiological mechanisms leading to reproductive decline in dairy cattle. In: II Biannual W. E.
Petersen Symposium – Reproductive loss in dairy cows: is the trend reversible? 2003, Minnesota. Anais…
Minnesota, p. 3-12, 2003.
Lucy, M. C. Reproductive loss in high-producing dairy cattle: where will it end? J. Dairy Sci., v. 84, p. 1277-
1293, 2001.
Monget, P.; Martin, G. B. Involvement of insulin-like growth factors in the interactions between nutrition and
reproduction in female mammals. Hum. Reprod., v. 12 (Supl. 1), p. 33-52, 1997.
O’Callaghan, D.; Boland, M. P. Nutritional effects on ovulation. Anim. Sci., v. 68, p. 299-314, 1999.
Peel, C. J.; Bauman, D. E. Somatotropin and lactation. J. Dairy Sci., v. 70: p. 474-386, 1987.

310
Rabiee, A. R.; Lean, I. J.; Gooden, J. M. et al. Short-term studies of ovarian metabolism in the ewe. Anim.
Reprod. Sci., v. 47, p. 43-58, 1997.
Reid, I. M.; Roberts, C. J.; Treacher, R.J. et al. Effect of body condition at calving on tissue mobilization,
development of fatty liver and blood chemistry of dairy cows. Anim. Prod., v. 43, p. 7-15, 1986.
Riis, P. M. Adaptation of metabolism to various conditions: nutritional and other environmental conditions. In:
Dynamic Biochemistry of Animal Production, Copenhagen, Elsevier, p. 319-357, 1983.
Sangsritavong, S.; Combs, D.K.; Sartori, R.; Wiltbank, M.C. High feed intake increases blood flow and
metabolism of progesterone and estradiol-17ß in dairy cattle. J. Dairy Sci., v. 85, p. 2831–2842, 2002.
Sartori, R. Ovarian function, circulating steroids, and early embryonic development in dairy cattle. 2002. 171p.
Tese (Ph.D. Dairy Science) – University of Wisconsin, Madison.
Sartori, R., Sartor-Bergfelt, R.; Mertens, S. A. et al. Fertilization and early embryonic development in heifers
and lactating cows in summer and lactating and dry cows in winter. J. Dairy Sci., v. 85, p. 2803-2812, 2002.
Scaramuzzi, R. J.; Murray, J. F.; Downing, J. A. et al. The effects of exogenous growth hormone on follicular
steroid secretion and ovulation rate in sheep. Dom. Anim. Endocrinol., v. 17, p. 269-277, 1999.
Sinclair, K. D.; Revilla, R.; Roche, J. F. et al. Ovulation of the first dominant follicle arising after day 21
postpartum in suckling beef cows. J. Anim. Sci., v. 75, p. 115-126, 2002.
Smith, B. I.; Risco, C. A. Management of periparturient disorders in dairy cattle. Vet. Clin. Food. Anim., v. 21,
p. 503-521, 2005.
Sordillo, L. M.; Contreras, G. A.; Aitken, S. L. Metabolic factors affecting the inflammatory response of
periparturient dairy cows. Anim. Health Res. Rev., v. 10, p. 53-63, 2009.
Spicer, L. J.; Tucker, W. B.; Adams, G. D. Insulin-like growth factor-I in dairy cows: relationships among
energy balance, body condition, ovarian activity, and estrous behavior. J. Dairy Sci., v. 73, p. 929-937, 1990.
Stevenson, J. S. Relationships among luteinizing hormone, estradiol, progesterone, glucocorticoids, milk yield,
body weight and postpartum ovarian activity in Holstein cows. J. Anim. Sci., v. 48, p. 570-577, 2006.
Stewart, R. E.; Spicer, L. J.; Hamilton, T. D. et al. Effects of insulin-like growth factor I and insulin on
proliferation and on basal and luteinizing hormone-induced steroidogenesis of bovine thecal cells: involvement
of glucose and receptors for insulin-like growth factor I and luteinizing hormone. J. Anim. Sci., v. 73, p. 3719-
3731, 1995.
Trevisi, E.; Amadori, M.; Cogrossi, S. et al. Metabolic stress and inflammatory response in high-yielding,
periparturient dairy cows. Res. Vet. Sci., v. 93, p. 695-704, 2012.
Van Saun, R. J. Transition cow nutrition and management: the key to herd reproductive performance. Penn
State University, 2000: Disponível em: http://vbs.psu.edu/ext/resources/pdf/transition-cow-health-and-
nutrition.
Vanholder, T.; Leroy, J. L. M. R.; Van Soom, A. et al. Effect of non-esterified fatty acids on bovine granulosa
cell steroidogenesis and proliferation in vitro. Anim. Reprod. Sci., v. 87, p. 33-44, 2005.
Williams, G. L. Implicações da amamentação e manejo da cria na eficiência reprodutiva futura de vacas de
corte. In: V Curso Novos Enfoques na Produção e Reprodução de Bovinos, 2001, Uberlândia. Anais...,
Uberlândia, p.65-73, 2001.
Williams, L. M.; Adam, C. L.; Mercer, J. G. et al. Leptin receptor and neuropeptide Y gene expression in the
sheep brain. J. Neuroendocrinol., v. 11, p.165-169, 1999.
Wiltbank, M.; Lopez, H.; Sartori, R. et al. Changes in reproductive physiology of lactating dairy cows due to
elevated steroid metabolism. Theriogenology, v. 65, p. 17-29, 2006.

311
Prevenção das acidoses ruminais em rebanhos leiteiros: novos
conceitos
Enrico Lippi Ortolani
A produtividade leiteira teve um incrível aumento graças aos avanços em genética,
nutrição, manejo e controle de enfermidades. Isso foi acompanhado por aumento na
ingestão de matéria seca (MS), com destaque à energia, na forma de carboidratos
solúveis. Essa mudança favoreceu o surgimento de quadros de acidose ruminal devido à
ingestão excessiva ou anormal de carboidratos solúveis.
Para entendermos melhor essa enfermidade é necessário revisar brevemente conceitos de
pH, acidez e os mecanismos de controle de pH ruminal. O pH é uma escala logarítmica
(0 a 14) que mede o grau de acidez (concentração de íons H+) e da alcalinidade
(concentração de íons OH-) dos compostos. Se considera pH 7,0 como neutro e os valores
inferiores e superiores ácidos ou alcalinos, respectivamente. Por ser logarítmico a cada
grau de pH que diminui ou aumenta ocorre uma alteração de concentração de íons H+ ou
de OH- na ordem de 10 vezes. Assim, se o pH diminui de 7,0 para 4,0 existe um
incremento de 1.000 vezes na concentração de H+. A força de um ácido se mede pelo seu
pK, ou seja, o pH ao qual a metade do H+ está associado ou não ao ácido. Quanto menor
o pK mais forte será o ácido.
O pH ruminal depende do balanço de produção de ácidos e de bases em seu meio. Os
principais ácidos graxos de cadeia curta (AGCC), antigamente denominados de ácidos
graxos voláteis, produzidos no rúmen são o acético (pK 5,0), o propiônico (pK 4,8) e o
butírico (pK 4,7). A principal base é o bicarbonato proveniente da saliva deglutida e em
menor grau a amônia (NH3), produzida a partir da hidrólise ruminal de proteína ou ureia.
A absorção dos ácidos ocorre pelas papilas da parede do rúmen e do omaso. Ácidos
associados com íon H+ são mais absorvidos que não-associados. O pH normal do fluido
do rúmen está entre 5,9 a 7,1 com produção total de ácidos não superior a 90 mmol/L de
conteúdo (Ortolani et al., 1982).

312
Tipos de acidose ruminal e suas patogenias
Existem dois tipos de acidoses ruminais: a mais branda e a mais intensa. A primeira é
gerada por aumento anormal de AGCC, em especial de propiônico e butírico, denominada
de acidose ruminal por AGCC (ARAGCC) produzindo um pH ruminal entre 5,2 a 5,8
por no mínimo 5 h de duração e com produção de ácidos superior a 90 mmol/L. A segunda
forma decorre da produção anormal de ácido láctico (pK 3,7) provocando a acidose
láctica ruminal (ALR), gerando um pH ruminal inferior a 5,2 com qualquer duração e
com produção total de ácidos superior a 200 mmol/L.
A produção de excesso de ácidos está intimamente relacionada com o tipo e a quantidade
de concentrado ingerido. A geração de ácidos ruminais é maior quanto menor for a cadeia
dos carboidratos solúveis na dieta. Assim, os monossacarídeos (por exemplo, glicose e
frutose presentes nas frutas) são mais acidogênicos que os dissacarídeos (lactose -no soro
lácteo- e sacarose -presente no melaço-) e estes mais acidogênicos que os polissacarídeos
(principalmente o amido). A ordem de disponibilidade do amido para a fermentação é
decrescente nos seguintes grãos: trigo, aveia, arroz, milho e sorgo. A mesma ordem ocorre
por processamento dos grãos de milho: floculação, silagem de milho úmido, fubá, quirera
e grãos inteiros (Owens et al., 1999; Krauze & Oetzel, 2006).
Outro ponto importante no controle da acidose é a salivação. O pH da saliva é alcalino
pela presença de bicarbonato, fundamental para o tamponamento ruminal. As glândulas
salivares recebem influência positiva do nervo vago, o qual é muito estimulado pelo atrito
das fibras dietéticas na parede ruminal como se fossem uma “escova de dente”. Quanto
maior o tamanho das fibras mais abundante será a salivação. Assim, uma partícula da
forragem de 1 cm tem um efeito 80x no estímulo da salivação, o caroço de algodão 70x,
a polpa cítrica 30x, o grão inteiro de milho 5,4x, quirera 2,4x, fubá 1,8x e milho floculado
1x.
A fermentação dos alimentos é feita pelos microrganismos ruminais. Na alimentação rica
em forragens predominam as bactérias Gram negativas (celulolíticas). Quanto mais
aumentam os concentrados energéticos, mais proliferam bactérias Gram positivas
(amilolíticas). Na ARAGCC se multiplicam outras bactérias Gram positivas, inicialmente
o Streptoccocus bovis (pH 5,1 a 5,8) que produz ácido propiônico e butírico e uma
pequena quantidade de láctico, e depois o Lactobaccilus spp. (menor que pH 5,2) que

313
origina apenas ácido láctico. São fundamentais no controle da acidose o estímulo ao
crescimento de bactérias lactilíticas (Megasphera elsdeni, Selenomonas ruminantium)
que transformam ácido láctico em propiônico, menos forte. Essas bactérias têm uma ação
maior no pH 5,8 porém morrem em pH inferior a 5,3. Suas populações crescem
lentamente, assim a adaptação às dietas ricas em concentrados energéticos deve ser
gradual (Owens et al., 1998).
A quantidade de concentrados para provocar acidose é muito variável. É possível gerar
ARAGCC em vacas sem prévia adaptação quando de repente se oferece mais de 35% de
matéria seca de concentrados ou ALR com mais de 50%. Quanto mais pesadas forem as
vacas, menores serão as quantidades de concentrados ingeridas para causar acidose
(Ortolani, 1995).
A morbidade da ARAGCC é muito variável no rebanho, sendo mais frequente ao redor
de 80 a 140 dias de lactação, momento em que a ingestão de matéria seca é máxima.
Porém, em primíparas, a morbidade é maior no terço médio da lactação. Essa situação
provavelmente ocorra, pois, as novilhas parecem ter um menor controle na ingestão de
concentrados energéticos e quando são mantidas em conjunto com multíparas, as
primeiras reduzem o número de visitas ao cocho, o que as faz comer em maior quantidade
em cada visita. Algumas vacas multíparas têm um risco maior de apresentarem acidose
que outras. As mais resistentes controlam mais a ingestão de concentrados após um
quadro prévio de acidose que as susceptíveis. Nas susceptíveis desenvolvem-se mais as
rumenites promovendo menor absorção de AGCC, tornando as recorrências do quadro
mais frequente, de maior intensidade e com recuperação mais lenta (Krauze & Oetzel,
2006; Beauchemin et al., 2014).
Na ALR a alta produção de ácidos e a grande dissociação destes poderá aumentar a
osmolaridade ruminal acima da sanguínea (300 mOsm/L) promovendo a migração de
fluidos e eletrólitos para o rúmen, gerando assim desidratação. A diarreia é discreta na
ARAGCC e exuberante na ALR, ambas com duração de até três dias. O pH das fezes
diminui de 6,0-7,0 para menos de 6,0 na ALR. A absorção do ácido láctico pode gerar
um estado variado de acidose metabólica acompanhado de acidúria temporária. A
laminite asséptica ocorre em muitas vacas dentro de dois a três dias na ALR, ou semanas
e meses depois de episódios de ARAGCC, causando claudicação e presença de linhas de

314
estresse nos cascos (Sousa, 2017). Algumas vacas podem contrair quadros pneumônicos
e polioencefalomalacia depois da acidose ruminal (Owens et al., 1998; Ortolani et al.,
2010; Danscher et al., 2015).
Devido às dificuldades de evidenciação de sintomas clínicos na ARAGCC têm sido
estudadas alterações no comportamento das vacas. Assim, Ortolani (2018) verificou que
existe uma diminuição em 63% na ingestão de matéria seca no primeiro dia da
enfermidade, sendo restabelecido apenas no 3º dia posterior. O tempo devotado às
atividades básicas também se alteram com diminuição de cerca 50% na ruminação, 35 %
na ingestão de alimentos, e aumento de 35% no tempo de ócio e de posição em decúbito
esternal.
Diagnóstico da acidose ruminal
O diagnóstico é feito por meio do exame físico e laboratorial. A enfermidade provoca
depressão no estado geral, diminuições do apetite, da produção leiteira, da frequência
ruminal e da ruminação, causa diarreia e desidratação entre outros sintomas supracitados.
Na ALR ocorre uma diminuição no pH fecal e da urina. No exame do fluido ruminal se
encontram alterações de cor (mais amarelado), odor (mais ácido) e diminuição do pH
entre outras mudanças.
Manejo preventivo da acidose ruminal
A prevenção é a chave do manejo da acidose ruminal. A primeira medida é a identificação
contínua da presença dos casos clínicos. Outros indicadores indiretos devem ser tomados
em conta na adoção de medidas preventivas. Um deles é o índice de ruminação do
rebanho que deve ser superior a 40% em vacas que não estejam comendo, se deslocando
ou dormindo. O índice de laminite deve ser inferior a 10%. A acidose interfere
negativamente na produção de gordura no leite. Vacas Holandesas e Jersey devem ter um
porcentual de gordura no leite superior a 3,4% e 4,3% e uma relação gordura: proteína
(G:P) maior que 1,1 e 1,2, respectivamente. Valores de G:P inferiores a 1,0 das vacas são
fortes indicadores de alta frequência de ARAGCC no rebanho (Enemark, 2008; Krauze
& Oetzel, 2008).

315
A segunda medida é a adaptação gradual aos concentrados, principalmente no período do
pré-parto. Essa providência tem dois motivos: aumentar adequadamente a população de
microrganismos lactilíticos e promover o crescimento das papilas ruminais. Vacas que
vão receber na dieta pós-parto até 50% a 60% MS de concentrados devem ter incluídos
gradualmente na ração 3 a 4 kg desses alimentos no último mês de gestação (Krauze &
Oetzel, 2008).
A terceira medida é evitar que as vacas fiquem demasiado tempo sem receber alimento
no decorrer do dia. Vacas famintas, por exemplo por 18 horas, comem cerca de 20% a
mais da mesma dieta quando realimentadas provocando uma queda drástica no pH
ruminal, por muitas horas. Assim deve-se distribuir a dieta e acompanhar sua sobra no
comedouro por no mínimo duas vezes ao dia. Sobras de alimento inferior a 5% é
indicativo que as vacas estão famintas. A sobra de alimentos ideal antes de uma nova
oferta deve ser entre 5% a 10 % do oferecido (Beuachemin & Penner, 2014).
A quarta medida é estimular a salivação. Como já mencionado, o tamanho da partícula
da forragem estimula mais ou menos a salivação. Para o tanto mensure o tamanho da
partícula de forragem oferecida por meio de peneiras sequenciais (Penn State Separator)
com malhas de diferentes diâmetros (longa > 1,9 cm; média 0,8 a 1,9 cm e curta < 0,8
cm). Porém, é um erro pensar que o excesso de fibra longa é benéfico para a vaca. Em
dietas com alto concentrado, o excesso de fibra longa favorecerá o surgimento de acidose.
Isso se deve à eleição do consumo de fibras curtas em detrimento das longas, deixando
estas no comedouro. Para evitar isso, algumas medidas são propostas. Estudo recente
indica que, em dietas muito energéticas, a melhor distribuição de fibra é algo como 8%
de longa, 40% de média e 52 % de curta. Para garantir um bom consumo de fibra longa
podem-se adicionar na dieta “colantes” como a glicerina (10% MS) ou melaço de cana
ou soja líquido que aderem todas as fibras aumentando seu consumo como um todo. É
fundamental também, para evitar a seleção de alimentos pela vaca, o oferecimento de
uma dieta completa totalmente misturada (TMR), distribuída por um vagão forrageiro.
Manter sempre ajustado o triturador de forragem para não fornecer volumosos picados
em demasia. Não deixar o feno ser misturado por muito tempo no vagão, pois isso
aumenta sua fragmentação. Oferecer, sempre que possível, forragem fresca, pois o
consumo de fibra longa aumenta (Krauze & Oetzel, 2008; Beauchemin & Penner, 2014).

316
No caso de todas estas medidas não funcionarem, principalmente em dietas muito ricas
em concentrados com alto risco de acidose, devem se adicionar tampões na dieta. Os
melhores resultados na prevenção são obtidos com bicarbonato de sódio (1,25% MS).
A quinta medida é a regularização do apetite e o acesso ao comedouro. Como já
mencionado, a dieta deve ser oferecida duas vezes ao dia, o que regulariza o pH e aumenta
o índice de ruminação. Deve-se evitar a superpopulação de vacas para não congestionar
o comedouro, sendo o ideal um espaço de 80 a 100 cm/vaca em lactação, principalmente
quando se misturam primíparas e multíparas. É essencial manter o bem-estar dos animais,
em especial no tocante ao estresse térmico.
Finalmente, sugere-se empregar aditivos para prevenir a acidose, os quais podem ser
naturais ou artificiais. O principal aditivo natural é a base de probiótico contendo
leveduras secas (Sacharomyces cerevisae 4 g/vaca/dia) que tem um papel preventivo
somente em ARAGCC, ajudando manter o pH ruminal acima de 5,6 e por aumentar as
bactérias lactilíticas. Os aditivos artificiais são a base de ionóforos e certos antibióticos.
Os ionóforos (monensina, lasalocida e salinomicina) são antimicrobianos que inibem o
crescimento de bactérias ruminais Gram positivas, em especial Streptoccocus bovis,
atuando no fluxo de cátions monovalentes ou divalentes interferindo no metabolismo
energético. Os antibióticos mais empregados são a virginiamicina e a tilosina com ação
bacteriostática sobre as bactérias Gram positivas produtoras de ácido láctico.
Recentemente um experimento conduzido pela nossa equipe demonstrou que a adição de
monensina (30 ppm) e virginiamicina (25 ppm) mitiga quadros de ALR muito intensos.
Referências
Beauchemin, K.A.; Penner, G. New developments in understanding ruminal acidosis in dairy cows.
3/February/2014. Article.extension.org/pages/26022, consultado 20/mayo/2016 em: <
https://articles.extension.org/pages/26022/new-developments-in-understanding-ruminal-acidosis-in-dairy-
cows>.
Danscher, A.M.; Shucong, L.; Andersen, P. H.; Khafipour, E. Kristensen, N.B.; Plaizier, J.C. Indicators of
subacute ruminal acidosis (SARA) in Danish Holstein cows. Acta Vet. Scand. v.57, p. 39-46, 2015.
Enemark, J.M.D. The monitoring prevention and treatment of subacute ruminal acidosis (SARA), a review.
The Vet. J. v.6, p. 32-43, 2008.
Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute rumen acidosis in dairy herds: a
review. Anim. Feed Sci. Technol. v.126, p. 215-236, 2006.
Oliveira, F.L.C., Sousa, R.S., Santos, J.A. et al. Use of virginiacymin and monensin to mitigate rumen
lactic acidosis in beef cattle. Proceedings of the Congress of the World Association for Buiatrics, 29.
2016, Dublin, Ireland. Anais... Dublin: 2016.

317
Ortolani, E.L.; Souza, R.; Benesi, F.J. The pH of the bovine ruminal fluid as influenced by species and diet.
Arq. Bras. Med. Vet. Zoot. v.34, n.2, 23-32, 1982.
Ortolani, E.L. Induction of lactic acidosis in cattle with sucrose: relationship between dose, rumen fluid pH
and animal size. Vet. Hum. Toxicol. v.37, n.5, p. 462-464, 1995.
Ortolani, E.L.; Maruta, C.A.; Minervino, A.H.H. Quadro clínico de zebuínos e taurinos submetidos à
acidose láctica ruminal aguda. Braz. J. Vet. Res. Anim. Sci. v.47, p. 253-261, 2010.
Owens, F.N. Secrist, D.S.; Hill,W.J.; Gill, D.R. Acidosis in cattle: a review. J. Ani. Sci., v.76, p.275-
286,1998.
Sousa, R.J. Avaliação de anti-inflamatórios não esteroidais no tratamento da laminite asséptica aguda
decorrente de acidose ruminal por oligofrutose em bovinos. Tese doutoramento, Faculdade de Medicina
Veterinária e Zootecnia, Universidade de São Paulo, 114 p., 2017.

318
Hipocalcemia puerperal bovina
Luis Albornoz
J. P. Albornoz
M. Morales
L. E. Fidalgo
Transtornos metabólicos
A necessidade de satisfazer os requerimentos de alimentos de origem animal da
população e de aumentar a rentabilidade das empresas pecuárias, tem motivado a seleção
das espécies animais para obter o máximo de otimização. Entre estas, o bovino ocupa um
lugar relevante e para aumentar sua capacidade produtiva têm sido utilizados diversos
procedimentos tais como seleção genética, novos sistemas de alimentação,
procedimentos de manejo e utilização de biotecnologias (Contreras, 1998). Com estas
medidas têm se conseguido maiores produções, porém com aumento do risco de sofrer
alterações da saúde e, agravado pela escassez de alimentos e problemas de manejo, a
saúde se compromete ainda mais em rebanhos que tenham níveis produtivos
relativamente baixos (Contreras, 1998).
Quando aumenta a produção por vaca, se trabalha com indivíduos selecionados, cuja
adequação orgânica permite ter bom rendimento, porém são animais mais suscetíveis de
sofrer enfermidades provocadas por uma alteração do metabolismo devido à sobrecarga da
atividade metabólica que exigem os maiores níveis produtivos. Nessas vacas, o metabolismo
energético, proteico e mineral frequentemente está alterado (Corbellini, 2000; Overton et al.,
2017). Esse aumento da produção induz una maior frequência e intensidade de apresentação
das enfermidades metabólicas o da produção, provocadas por um desequilíbrio entre os
nutrientes que ingressam no organismo, seu metabolismo e os egressos através das fezes,
urina, leite, feto, etc. Lamentavelmente, a maioria destas enfermidades são de difícil
percepção limitando a produção de forma persistente e diminuindo a rentabilidade (Payne,
1981; Overton et al., 2017).
Tanto uma deficiente como uma excessiva alimentação, seja de minerais, energia ou
proteína pode causar problemas, e muitas vezes esses desequilíbrios têm efeitos aditivos
ou se potencializam criando um transtorno maior do que considerado isoladamente. A

319
fertilidade e os problemas metabólicos estão relacionados e não podem ser separados
(Lotthamer, 1992). Caso típico destas situações são os eventos de crescimento, gestação
e lactação. Este efeito distribuidor da energia é conhecido como homeorresis (Baumam
& Currie, 1980), do vocábulo grego que significa "fluxo uniforme", ou como teleorresis
("fluxo dirigido"). Isto implica conseguir a uniformidade do fluxo de nutrientes para
sustentar um estado fisiológico, como por exemplo, a lactação. Para a manutenção da
gestação e o preparo da lactação são necessárias adaptações metabólicas nos tecidos de
uma forma coordenada (Bauman & Currie, 1980; Van Saun, 1997). Os tecidos primários
de adaptação são homeorréticos e, neste grupo, estão os tecidos adiposo,
musculoesquelético e hepático. As mudanças metabólicas associadas com a transição
desde a gestação até a lactação ocorrem em todos os animais, incluídos aqueles bem
alimentados. Entretanto, estas mudanças metabólicas podem ser exageradas quando o
fornecimento dietético de energia, proteínas ou minerais são insuficientes, o que leva à
manifestação de transtornos metabólicos (Van Saun, 1997).
As perdas econômicas associadas às enfermidades do periparto têm elevado custo,
representado pela perda de produção de leite, diminuição da eficiência reprodutiva,
despesas veterinárias, aumento de mão de obra, tratamentos, substituição de animais, etc.
(Corbellini, 2000; Van Saun, 2010). Em torno de 50% de todas as lactações são afetadas
por pelo menos uma enfermidade no periparto, o que sugere perdas econômicas
importantes (Corbellini, 2000; Dyk et al., 1995). Estudos epidemiológicos mostram que
doenças durante o período de transição não são eventos produzidos independentemente,
mas são um complexo de transtornos relacionados entre si (Van Saun, 1997). Por
exemplo, se uma vaca sofrer hipocalcemia puerperal (HP) tem 4 vezes mais probabilidade
de ter retenção de placenta e 2 a 3 vezes más de ter deslocamento de abomaso. A retenção
de placenta aumenta 16,4 vezes a probabilidade de sofrer cetose, e as vacas com
deslocamento de abomaso são 1,8 vezes mais propensas de sofrer HP, retenção de
placenta ou cetose (Curtis et al., 1983). É, portanto, evidente que se uma vaca está sadia
durante o período de transição, cabe esperar um período de lactação produtivo e com
grandes expectativas de uma boa eficiência reprodutiva (Van Saun, 1997).
Dentre os distúrbios metabólicos que podem sofrer as vacas no período do periparto,
existem os desequilíbrios minerais, mas também estão os desequilíbrios energéticos e

320
proteicos. Por exemplo, a síndrome de mobilização adiposa é a principal consequência
de uma pobre adaptação do animal ao balanço energético negativo (BEN) e causa de
outras patologias do periparto (cetose, deslocamento de abomaso, retenção de placenta,
mastite, redução da produção, problemas reprodutivos e aumento da susceptibilidade a
doenças infecciosas (Fernández, 2009; Roberts et al., 2012; Santschi et al., 2011).
Hipocalcemia puerperal
O final do período de gestação e início da lactação representam situações fisiológicas de
estresse e de mudanças dramáticas na demanda de nutrientes, que requer uma perfeita
coordenação do metabolismo para satisfazer o aumento significativo dos requerimentos
logo após o parto. O metabolismo dos minerais não escapa a essas grandes mudanças,
especialmente o do cálcio. Todas as vacas sofrem diminuição do cálcio do sangue desde
um dia antes do parto até dois ou três dias após o parto (Albornoz, 2006), esperando que
os mecanismos homeostáticos que regulam o metabolismo do cálcio se adaptem à grande
demanda deste mineral (Holmes, 2003).
A HP é um distúrbio metabólico que ocorre no periparto especialmente em vacas leiteiras
de alta produção. O transtorno se caracteriza por um quadro clínico que inclui inapetência,
tetania, paralisia flácida, inibição da micção e da defecação, decúbito, coma e
eventualmente morte. Do ponto de vista bioquímico nota-se uma rápida diminuição da
concentração de cálcio y fósforo no sangue, relacionada com a formação de colostro
(Albornoz, 2006; Contreras, 2002b; Horst et al.,1997; Horst et al., 1998; Houe et al.,
2001). Os níveis sanguíneos de referência de cálcio em vacas leiteiras variam entre 8,6 e
9,6 mg/dL (Blood & Radostitis 1992; Goff, 2008). Quando o cálcio plasmático está
abaixo desses limites, funções como a neurotransmissão, contração muscular e regulação
hormonal podem ser afetadas (Blood & Radostits, 1992; Corbellini, 2000; Radostits,
2001). A forma clínica do transtorno (paresia puerperal hipocalcêmica) apresenta-se
principalmente em granjas leiteiras intensivas. A incidência média pode estimar-se entre
5 a 10%. É observada mais frequentemente em vacas de alta produção e de mais idade
(Radostits, 2001; Stöber, 2005).

321
O cálcio no organismo
O cálcio é um cátion multivalente muito importante no organismo. Este mineral mantém
a integridade da estrutura de ossos e dentes, sendo fundamental para controlar uma grande
quantidade de processos bioquímicos. No organismo, aproximadamente 98% do cálcio
encontra-se nos ossos, 1% nas células e 0,3% no líquido extracelular. Do total de cálcio
no plasma, 55% encontra-se em forma ionizada como Ca2+ ativo, 35% unido a proteínas,
principalmente albumina e 10% constitui complexos em formas não iónicas como o
bicarbonato de cálcio. O equilíbrio entre o Ca ionizado e o Ca unido a proteínas depende
do pH sanguíneo. A alcalose diminui a concentração de Ca total, enquanto a acidose tem
o efeito contrário (Holmes, 2003; Rosol et al., 2000; Stöber, 2005).
Fisiopatologia da hipocalcemia
No periparto, o cálcio sofre uma rápida depleção passando do plasma à glândula mamária
sem dar tempo de uma compensação por parte dos mecanismos endócrinos (Alonso,
1997; Dhiman & Sasidharan, 1998; Horst et al., 1998). Durante o parto ou logo após, a
hipocalcemia é inevitável e caracterizada por concentrações de Ca sanguíneo < 8 mg/dL
(Risco, 2001a; Risco, 2001b). O feto necessita 5,3 g de Ca por dia, enquanto são
requeridos entre 13 e 18 g para a secreção de colostro (Alonso, 1997; Corbellini, 2000).
Ao redor do 1º dia pós-parto as vacas que produzem em torno de 10 L de colostro utilizam
como mínimo 23 g de Ca, aproximadamente 6 a 9 vezes mais do Ca contido no líquido
extracelular. Portanto, as necessidades totais de Ca aumentam radicalmente após o parto
passando de 10 a 15 g/dia para 30 a 50 g/dia (Alonso, 1997; Corbellini, 1998; Horst et
al., 1998; Risco, 2001b). Durante o período seco, quando os requerimentos de cálcio são
mínimos (a drenagem fetal é de 10 a 12 g/dia), os mecanismos homeostáticos encontram-
se relativamente inativos (Horst, 1997). No momento do parto, as necessidades de Ca
crescem subitamente e quase todas as vacas sofrem um momentâneo desequilíbrio na
regulação do Ca sanguíneo, sem chegar a ser uma verdadeira deficiência de Ca
(Corbellini, 2000; De Garis & Lean, 2009).
Os mecanismos de homeostase que regulam a calcemia interveem rapidamente
restituindo o Ca por três vias (Holmes, 2003): (a) absorção intestinal de cálcio, (b)
liberação de cálcio dos ossos para o sangue e (c) reabsorção cálcio em nível renal. O

322
atraso no início dos mecanismos de homeostase é a causa mais importante da HP (Alonso,
1997). Existem três fatores que afetam a homeostase do Ca e as variações de um ou mais
deles parecem ter importância decisiva no desencadeamento do transtorno: (1) Perda
excessiva de Ca para o colostro (Alonso, 1997): um alto potencial genético e boa
alimentação energética e proteica no pré-parto, principalmente quando há ganho da
condição corporal de 20 a 30 dias antes do parto, predispõem ao transtorno porque a
secreção inicial de colostro é abundante (Corbellini 2000; Roche, 2003). (2) Diminuição
na absorção de Ca no intestino: na absorção intestinal de Ca participam dois mecanismos
de transporte -ativo e passivo- através das células epiteliais do intestino delgado. O
transporte ativo, que é o mais importante, realiza-se com mediação do hormônio da
paratireoide (PTH) e da vitamina D3, enquanto transporte passivo ocorre por diferença de
gradientes de concentração (Alonso, 1997). (3) A mobilização de Ca a partir dos
depósitos de Ca ósseo pode não ser suficientemente rápida para manter a calcemia
(Alonso, 1997): durante períodos de baixa ingesta de Ca, o osso é a maior fonte deste
mineral (Horst et al.,1997). O Ca existe no osso em dois estados, uma pequena quantidade
está facilmente disponível em solução ao redor das células trabeculares ósseas e os
canalículos ósseos (Horst et al.,1997). O Ca solúvel que se encontra nos fluidos do osso
é separado dos fluidos extracelulares pelas células ósseas sinciciais sob estimulação do
PTH. Estas células rapidamente transferem os fluidos cálcicos do osso à corrente
sanguínea (Goff et al., 2004; Horst et al.,1997). A quantidade de Ca proveniente dos
fluidos ósseos em vacas leiteiras adultas alimentadas com dietas de alto conteúdo de
forragens pode ser estimado entre 6 a 10 g de Ca por dia (Goff & Horst, 1997).
Regulação endócrina da calcemia
Vários hormônios estão envolvidos no metabolismo do cálcio e do fósforo. Dois desses
hormônios, o PTH e a calcitonina (CT), têm efeito na atividade do Ca extracelular e o
osso (Goff et al., 2008; De Garis & Lean, 2009; Holmes, 2004). A glândula paratireoide
secreta PTH em resposta a uma queda da calcemia ou a um aumento da fosfatemia e tem
uma ação hipercalcemiante (Goff et al., 2008; Thiede, 1994). A CT é secretada em
resposta a uma hipercalcemia (Payne, 1981; Underwood, 1983). Um 3º hormônio, o 1,25-
dihidroxi-colecalciferol (DHCC) é derivado da vitamina D e sintetizado em pele, fígado

323
e rim. Esses três hormônios atuam juntos para manter constante o nível de cálcio e fósforo
no líquido extracelular e regular o metabolismo ósseo (Holmes, 2004; Horst et al., 1997).
Outros hormônios como os estrógenos, andrógenos, hormônio do crescimento e cortisol
também afetam o metabolismo do cálcio (Holmes, 2004).
PTH
O PTH é secretado em resposta a hipocalcemia e tem como efeito aumentar a
concentração do Ca mediante um efeito lento, porém prolongado (Payne, 1981). Para
regular a calcemia o PTH tem cinco ações distintas (Holmes 2004): (a) aumenta a
liberação de Ca dos ossos ao plasma; (b) aumenta a reabsorção de Ca dos túbulos renais;
(c) aumenta a atividade da enzima renal 1-α-hidroxilase; (d) diminui a reabsorção de
fósforo inorgânico nos túbulos renais; (e) aumenta a absorção intestinal de Ca, de forma
indireta mediada pela vitamina D3. Diante de qualquer diminuição de Ca sanguíneo, o
PTH é secretado atuando em poucos minutos aumentando a reabsorção renal no filtrado
glomerular. Se as necessidades de Ca são pequenas, a calcemia retorna à normalidade e
a secreção de PTH volta aos valores basais. Entretanto, se as necessidades de Ca são
grandes, a secreção de PTH estimula também a liberação de Ca do sistema ósseo (Horst
et al., 1997; Oetzel, 2002; Underwood 1983).
Calcitonina
A calcitonina atua, principalmente, em dois órgãos: osso e rim, induzindo um aumento
da saída de cálcio e fósforo desde o líquido extracelular e diminuindo a taxa de entrada
de estes íons ao líquido extracelular ou ambas simultaneamente. A magnitude da
diminuição plasmática de Ca é diretamente proporcional à taxa basal de recâmbio ósseo.
Portanto, os animais jovens em fase de crescimento são mais afetados pela CT, enquanto
os animais adultos com esqueletos mais estáveis respondem minimamente à CT. A ação
hipocalcemiante da CT se deve à inibição da osteólise osteocítica e da liberação óssea
osteoclástica, especialmente quando se encontram estimuladas pelo PTH. Apesar de ter
um efeito menos importante do que nos ossos, a CT reduz a reabsorção renal de cálcio e
fósforo, causando aumento na liberação renal destes íons que levam a hipocalcemia e

324
hipofosfatemia (Venjakob et al., 2017). A CT também atua para reduzir a reabsorção de
sódio, magnésio e potássio no túbulo proximal do rim (Holmes, 2004).
Vitamina D₃
A vitamina D₃ atua como um hormônio. Embora possa ser sintetizada no organismo,
também é uma vitamina uma vez que, se não for sintetizada em quantidades suficientes,
deve ser ingerida (Holmes, 2004). Atua sinergicamente com PTH aumentando o Ca
sanguíneo por estimular a liberação óssea e aumentar a reabsorção renal de Ca (Horst et
al.,1997). A vitamina D₃ (DHCC) é importante por estimular o transporte do cálcio
ingerido na dieta através do epitélio intestinal (Horst et al.,1997). O Ca pode ser absorvido
através do intestino por mecanismos dependente e independente do DHCC. No caso do
mecanismo independente, a absorção de Ca é primariamente por difusão passiva (Oetzel,
2002) enquanto no mecanismo dependente de DHCC a absorção ocorre por transporte
ativo através das células do epitélio intestinal, o que ocorre quando a dieta é baixa ou
quando a demanda é alta em Ca (Horst et al.,1997; Hove & Hilde, 1984; Oetzel, 2002).
Este processo requer DHCC, o qual estimula a síntese de uma proteína que transporta o
Ca através das células do epitélio intestinal (Horst et al.,1997).
Nas vacas leiteiras de alta produção, as demandas de Ca antes do parto estão em seu
menor nível. As demandas de Ca pelo feto são relativamente baixas, similares às
quantidades que se necessitam para enfrentar a última etapa da lactação. Assim, os
mecanismos de liberação óssea permanecem imóveis e a absorção de Ca intestinal se
encontra em sua forma passiva nesse momento (Oetzel, 2002). As dietas típicas usadas
para alimentar as vacas nessa etapa da vida reprodutiva excedem os requerimentos de Ca,
de modo que as demandas de Ca para a manutenção dos tecidos corporais e o
desenvolvimento do feto podem ser cobertas totalmente sem a absorção dependente da
vitamina D (Oetzel, 2002). Durante os primeiros dias de lactação, a homeostase do Ca é
restaurada pelo PTH, que reduz as perdas urinárias de Ca, estimula a liberação de Ca dos
ossos e aumenta a síntese de DHCC, melhorando o transporte intestinal ativo de Ca
(Oetzel, 2002).
A capacidade de adaptação à hipocalcemia é influenciada por numerosos fatores. As
restrições de Ca melhoram a capacidade para responder à hipocalcemia, mas têm

325
relativamente efeitos pequenos na incidência de hipocalcemia clínica (Oetzel, 2002). O
risco mais importante na apresentação de hipocalcemia é o equilíbrio ácido-básico do
animal no momento do parto. Existe atualmente uma tendência a considerar a
hipocalcemia como uma “intoxicação nutricional subclínica” causada por excesso de
potássio (K), principal indutor de alcalose metabólica (Corbellini, 2000).
O Ca é requerido para o normal funcionamento de uma extensa variedade de tecidos e
processos fisiológicos. É necessário para a formação óssea, para a contração muscular, a
transmissão nervosa, a coagulação do sangue e na regulação de alguns hormônios (Goff
et al., 2008; Horst et al.,1994; Radostits, 2001). Em geral, os vertebrados mantêm a
calcemia com eficiência, com exceção das vacas parturientes adultas leiteiras, as quais
desenvolvem hipocalcemia (Horst et al., 1994). Os processos de absorção intestinal
(Hove & Hilde, 1984) e de liberação óssea de Ca estão sob a influência da regulação
endócrina do Ca, o PTH e o DHCC. Algumas desordens endócrinas resultantes de
deficiência ou excesso de PTH ou vitamina D foram hipotetizados como defeitos
primários em vacas com HP, porém essas hipóteses foram rejeitadas quando
pesquisadores acharam níveis elevados de PTH e DHCC no sangue de animais que
sofriam paresia puerperal hipocalcêmica (Corbellini, 1998; Horst et al., 1994). As lesões
celulares envolvidas ainda permanecem sem serem identificadas (Horst et al., 1994), mas
a etiologia da paresia puerperal hipocalcêmica ainda é motivo de estudo, pois não se
conhece a forma detalhada de todos os mecanismos metabólicos envolvidos. Uma falta
de resposta do tecido ósseo à ação mobilizadora de Ca e P por parte do PTH e o DHCC,
devido a um estado de alcalose metabólica, parece ser a causa mais provável (Contreras,
2002b; Corbellini, 1998; Corbellini, 2000; Goff et al., 2008).
Fatores que influem na homeostase de cálcio
Alcalose metabólica
A alcalose metabólica predispõe as vacas à HP clínica e subclínica. Nesse caso, a
conformação dos receptores do PTH sofre alteração fazendo menos sensíveis os tecidos
ósseo e renal ao PTH. A alcalose metabólica decorre, em grande medida, de uma dieta

326
com predomínio catiônico (K+, Na+, Ca2+ e Mg2+) sobre os ânions (cloretos, fosfatos e
sulfatos) no sangue (Goff, 2008).
Hipomagnesemia
A hipomagnesemia provoca uma diminuição na capacidade de mobilização de Ca em
resposta a um estímulo hipocalcêmico (Bednarek et al., 2000). Isto explica a maior
suscetibilidade à hipocalcemia nas vacas hipomagnesêmicas do que nas
normomagnesêmicas, o que se conhece como hipocalcemia Mg-dependente. Nesse caso,
ocorreria uma menor produção de PTH, reduzindo o tecido sensível ao hormônio e uma
menor resposta aos tecidos “alvo” para a mobilização de Ca (Corbellini, 1998; Goff et
al., 2004; Goff, 2008; Mulligan & Doherty, 2008; Roche, 2003) ou também uma
interferência do metabolismo da vitamina D, pois o Mg é necessário para a hidroxilação
no fígado do 25-OH-colecalciferol, intermediário na síntese de DHCC (Contreras,
2002b). O excesso de K (> 2% MS) altera o metabolismo do Mg, interferindo sua
absorção em nível ruminal, muito embora na porção pós-ruminal haja uma compensação
na absorção de Mg. Um aumento da concentração de K na dieta entre 2,6 a 4,3% reduz a
absorção de Mg em aproximadamente 82% (Roche, 2003). Também o excesso de amônio
ruminal, produzido por excesso de proteína degradável ou deficiência de energia na dieta,
provocam interferência na absorção de Mg (Contreras, 2002a). Foi observado que 70%
das vacas em sistemas de pastoreio eram hipomagnesêmicas no dia do parto (Roche,
2003). Albornoz (2006) observou uma tendência a incremento de Mg no dia do parto,
embora os valores possam ser considerados hipomagnesêmicos. As vacas nesse estudo
mostraram hipomagnesemia com valores que se elevaram no dia do parto (1,78 ± 0,26
mg/dL; n= 35) aproximando-se aos limites inferiores de referência (1,8 mg/dL).
Venjakob et al. (2017) encontraram uma correlação negativa entre Ca e Mg indicando a
importância de estudos mais aprofundados sobre a regulação fisiológica desses minerais
ao redor do parto.
Desenvolvimento da hipocalcemia
As concentrações sanguíneas de cálcio e fósforo no bovino são de 8,0-10,6 mg/dL e 3,2-
7,1 mg/dL, respectivamente. Em vacas sadias no momento do parto, as concentrações de

327
cálcio e fósforo diminuem levemente (Albornoz, 2006) e depois se restabelecem, o que
se conhece como hipocalcemia fisiológica. Porém, quando o descenso é intenso, por
insuficiente capacidade de mobilização de Ca, se apresenta o transtorno metabólico
conhecido como paresia puerperal hipocalcêmica ou febre do leite). As causas de uma
mobilização insuficiente de cálcio são: (1) Síntese ou secreção insuficiente de PTH,
hormônio responsável pela extração de Ca e P do osso para aumentar suas concentrações
no sangue. (2) Síntese insuficiente do precursor da vitamina D, a qual promove a absorção
de Ca no intestino. (3) Insuficiente resposta dos tecidos (rim, osso, intestino) aos
estímulos de PTH e/ou DHCC. (4) Diminuição do apetite por doenças concomitantes. (5)
Hipomagnesemia, que altera a secreção de PTH e o metabolismo da vitamina D. (6)
Estresse, que favorece a secreção da calcitonina, hormônio hipocalcemiante.
Fatores predisponentes da hipocalcemia
Raça
Inúmeras pesquisas sugerem que algumas raças de vacas leiteiras são mais suscetíveis à
HP, tais como Channel Island, Swedish Red and White e Jersey (Horst et al., 1997; Lean
et al., 2006; Roche, 2003; Lean et al., 2006). A exata razão desta maior suscetibilidade
não está clara, mas tem sido demonstrado que os receptores intestinais para DHCC são
menores em vacas Jersey que em Holandesas ajustados por idade (Horst et al., 1997). O
baixo número de receptores pode resultar de uma perda de tecido sensível ao DHCC
(Horst et al.., 1997; Houe et al., 2001).
Idade
À medida que aumenta a idade das vacas, também aumenta a incidência de HP (Alonso,
1997; Horst et al., 1997; Houe et al., 2001), sendo muito rara a ocorrência em vacas de
primeira lactação e aumentando a partir da terceira lactação (Horst et al., 1997). Inúmeros
fatores contribuem para que a idade avançada seja um importante fator predisponente da
HP, tais como a maior produção de leite das vacas adultas, as quais têm maior demanda
de Ca, a queda na capacidade de mobilizar Ca ósseo, a diminuição do transporte ativo de

328
Ca no intestino e a menor produção de DHCC. Coletivamente todos esses problemas
produzem una falta de resposta às necessidades agudas de Ca (Horst et al., 1997).
Dieta e condição corporal
Quando as vacas são alimentadas com níveis baixos de Ca ou quando se ajusta a relação
Ca/P para 2:1 tem sido notada uma menor incidência de HP (Horst et al., 1997).
Um alto score na condição corporal aumenta os riscos de HP (Houe et al., 2001). A
condição corporal pode ser medida em uma escala que vai de 1 a 5 com frações de 0,25
na qual score 1 significa um animal extremamente magro e score 5 um animal
extremamente obeso (Ferguson et al., 1994; Houe et al., 2001). Foi observado que vacas
com condição corporal maior que 4 tinham um risco de HP maior (odds ratio de 4,3)
(Houe et al., 2001).
Nível de produção
A incidência de HP está associada positivamente com o nível de produção de leite. Foi
observado um aumento de 0,05% na incidência de HP por cada kg de produção de gordura
produzida na lactação anterior. Outro estudo demonstrou que o risco para HP teve um
OR de 1,5 para vacas que produziram 7 kg de leite corrigidos por gordura comparadas
com vacas que produziram 4 kg de leite nas mesmas condições (Houe et al., 2001).
Distúrbios relacionados com a hipocalcemia
Pela ação coordenada de órgãos como fígado, rim, intestino delgado e osso e através da
ação homeostática dos hormônios calciotrópicos, aumenta a capacidade intestinal de
absorção intestinal de Ca, sua mobilização (ressorção) óssea e sua reabsorção renal de
Ca. Uma falha nesses mecanismos geralmente causada por erros de manejo no pré-parto,
pode levar não somente a uma alta incidência de HP clínica, mas também HP subclínica,
responsável por transtornos como partos demorados e retenção de placenta (Corbellini,
1997). A paresia puerperal ou febre do leite é a manifestação clínica da HP, sendo um
dos distúrbios metabólicos mais comuns do periparto na vaca leiteira multípara (Risco,
2001b). Embora não esteja totalmente esclarecida a relação entre hipocalcemia e paralisia
muscular flácida, os íons de Ca têm importância na condução do estímulo nervoso e nas

329
células musculares, bem como na transmissão do estímulo da placa motora terminal e na
contração muscular (Stöber, 2005). Os sinais clínicos ocorrem por mudanças no tono
neuromuscular, paralisia flácida e, eventualmente, coma. A maioria dos casos não
tratados morrem entre 1 a 2 dias (Houe et al., 2001; Risco, 2001b). Foi observado que a
paresia ou paralisia esteve associada com níveis de Ca plasmático inferiores a 4,0 ou 5,0
mg/dL (Oetzel, 1996; Risco, 2001a). Nem todas as vacas que sofrem HP desenvolvem os
sinais clínicos.
A HP causa um aumento significativo na incidência de outras enfermidades, tais como
mastite, retenção de placenta, deslocamento de abomaso, distocias e cetose, as quais
evidentemente agravam o BEN e atentam contra a vida produtiva da vaca (Corbellini,
2000). Além do mais, reduzem a capacidade das células do sistema imunitário em
resposta a estímulos contribuindo ao aumento de doenças infecciosas como mastite e
metrite (Reinhardt, 2011). A diminuição de Ca no sangue reduz a contratilidade muscular,
começando pela musculatura lisa (Corbellini, 2000), diminuindo consequentemente a
motilidade do rúmen e do abomaso, aumentando a incidência de deslocamento de
abomaso e reduzindo o consumo. A redução da contratilidade muscular também impede
o eficiente fechamento do mamilo, o que leva a aumentar a frequência de mastite
(Reinhardt, 2011).
Foram encontradas associações significativas calculadas pelo índice de OR (odd ratio)
entre HP e distocia de 6,5; retenção de placenta de 3,2; deslocamento de abomaso à
esquerda de 3,4; e metrite e cetose clínicas de 1,7 (Curtis et al., 1983; Oetzel, 1996; Risco,
2001a). Também se encontraram associações significativas entre HP e prolapso uterino,
cios silenciosos, cistos ovarianos e infertilidade (Houe et al., 2001; Risco, 2001a).
A produção de leite também se vê afetada. Assim, se encontrou que vacas com HP tiveram
uma redução na produção de leite entre 1,1 a 2,9 kg/dia (Houe et al., 2001), considerando
apenas o efeito direto, pois se fossem considerados os efeitos indiretos, a perda de leite
seria maior, por exemplo, efeitos por infecção do trato reprodutivo e alongamento do
período parto-concepção. O peso corporal também é afetado notando perdas de peso
consideráveis (Houe et al., 2001)

330
Prevenção da hipocalcemia
Muitos princípios de controle da HP têm sido descritos pela literatura nos últimos 50
anos, mas somente alguns têm sido aplicados a rebanhos comerciais (Thilsing et al.,
2002). Dentro deles podem ser mencionados: (a) Restrição da ingesta de Ca durante as
últimas semanas de gestação ou diminuição relativa por incremento do P (Contreras,
2002b; Thilsing et al., 2002). (b) Suplementação com sais aniônicos de forma a acidificar
a dieta durante as últimas semanas de gestação (Contreras, 2002b; Dishington, 1975;
Goff, 2008; Thilsing et al., 2002;). (c) Suplementação de Ca via oral ao redor do parto
(Bostedt & Bless, 1993; Dhiman & Sasidharan, 1998; Goff, 2008; Jonsson & Pehrson,
1970; Queen, 1993). (d) Combinação de tratamentos com dietas acidogênicas durante o
pré-parto e administração de CaCl2 pós-parto, obtendo maiores níveis séricos de Ca logo
após o parto (Afshar et al., 2018). (e) Administração de vitamina D ou metabolitos
análogos 2 a 5 dias antes do parto, tratamento que aumenta efetivamente a absorção de
Ca no intestino. Este tratamento não tem efeito se a vaca pare antes. Como nem sempre
é possível predizer o momento exato do parto, se deve desaconselhar a repetição do
tratamento pois implica o perigo de calcinose por hipervitaminose D₃ (Bostedt & Bless,
1993; Goff, 1989; Goff, 2008; Stöber, 2005).
Existem outras medidas de controle menos específicas e comuns para a prevenção da HP
que incluem algumas das seguintes práticas de manejo (Thilsing et al., 2002; Roche,
2003; Goff et al., 1989): controlar os níveis de Mg, K e Na durante o pré-parto, controlar
a condição corporal, controlar a ingesta de glicídeos durante o pré-parto, encurtar o
período seco, ordenhar durante o pré-parto, reduzir a ordenha no início da lactação e
administrar PTH.
Ingesta de magnésio
Rebanhos leiteiros com alta incidência de HP com frequência têm concentrações de Mg
sanguíneo abaixo do valor de referência durante o periparto (Venjakob 2017; De Garis &
Lean, 2009; Contreras 2002a; Bednarek et al., 2000; Wang et al., 1994). Existe
concorrência entre o Mg e o Ca para sua absorção no intestino e sua reabsorção nos
túbulos renais (Contreras, 2002a; Wang et al., 1994). Mais importante é que deficiência
de Mg pode diminuir a mobilização de Ca ósseo (Corbellini, 1998; Roche, 2003). Foi

331
observado que quando se alimentam vacas com 71 g de Mg/dia durante o período seco
existe maior mobilização de Ca ósseo que quando se alimentam com 17 g de Mg/dia
(Wang et al., 1994). À medida que se aprofunda no conhecimento da etiopatogenia da
HP, se desenvolvem métodos preventivos, geralmente encaminhados a aumentar de
forma mais rápida a disponibilidade de Ca sanguíneo, seja pelo incremento na taxa de
absorção intestinal ou seja pela mobilização óssea (Alonso, 1997).
Medidas profiláticas
Nas medidas preventivas tem que distinguir entre as farmacológicas (destinadas ao
indivíduo) e a adaptação à alimentação das vacas durante o pré-parto, a qual é efetiva
depois de um tempo prolongado, mas tem a vantagem de atuar sobre todas as vacas em
risco de sofrer HP (Stöber, 2005).
Algumas medidas profilácticas que podem ser mencionadas incluem as seguintes:
- Dietas com baixo conteúdo de Ca: As recomendações dietárias prévias ao parto têm por
objetivo preparar a vaca para que a homeostase do Ca funcione de forma ótima no início
da lactação. Com baixo fornecimento de Ca no pré-parto favorece-se a absorção ativa
intestinal e rápida liberação óssea (Contreras, 2002b; Goff & Horst, 1997; Goff et al.,
2004). Para isto, é necessário que a vaca consuma menos de 20 g de Ca/dia, o que com a
alimentação habitual é difícil (Stöber, 2005). A ingesta de Ca durante o pré-parto pode
ser restrita substituindo em parte ou totalmente a forragem de leguminosas na dieta por
forragem seca de gramíneas, silagem de milho e/ou concentrados, porém é difícil atingir
menos de 50 g/dia, o que não resulta efetivo para prevenir a HP (Oetzel, 1993).
- Dietas com baixa relação Ca/P: Em alguns países é comum a administração parenteral
de fósforo entre 60 a 45 dias antes do parto. A hipótese é que a HP pode ser prevenida
com a suplementação de fósforo e rações pobres em Ca no final da gestação (Manston,
1967). Isto estimularia a atividade da glândula paratireoide no período seco e prepararia
as vacas para a demanda de Ca após o parto. Inversamente, dietas ricas em Ca (entre 100
a 125 g/dia) no mesmo período, aumentam a incidência de HP, provavelmente por
diminuir a atividade da paratireoide. Deve enfatizar-se a importância da relação Ca/P
como preventivo do transtorno, menos do que as quantidades absolutas de cada mineral
separadamente (Rajaratne et al., 1994; Alonso, 1997). Contreras et al. (1996) e Roche et

332
al. (2013) estudaram perfis metabólicos em sistemas de pastoreio, encontrando que a
relação Ca/P é menor em vacas gestantes do que em vacas em lactação. Albornoz (2014)
observa que a relação Ca/P é variável conforme a estação do ano encontrando que, no
outono, a relação é menor em vacas gestantes (1,66 ± 0,31) do que em lactantes (1,92 ±
0,44) enquanto na primavera, a relação é maior em vacas gestantes (1,89 ± 0,45) do que
em lactantes (1,75 ± 0,42). Alguns autores opinam que a hipofosfatemia é a maior causa
de recidivas e pobres respostas na rotina de terapia para a HP clínica (Kojouri &
Karimzadeh, 2002). Recomenda-se para rebanhos leiteiros 0,42% de P na matéria seca
(MS) ingerida e uma relação Ca/P de 1,4 (Call et al., 1987; Gerloff et al., 1996). Também
é recomendado o fornecimento de 0,25% de Mg na MS ingerida (Albornoz, 2006).
- Uso do balanço iônico: Os organismos vivos devem manter a eletroneutralidade do
plasma, ou seja, que a soma dos cátions (carregados positivamente) devem equilibrar-se
com a de ânions (carregados negativamente). Este é o conceito básico do chamado
balanço cátion-ânion da dieta (BCAD), balanço eletrolítico da dieta, diferença cátion-
ânion da dieta (DCAD) ou diferença iônica (García, 1997). Os cátions Na+ e K+ e os
ânions Cl- e S= são os principais a levar em consideração ao avaliar o BCAD (García
1997; Goff et al., 2004). Uma dieta predominante em ânions é considerada acidogênica,
enquanto uma dieta predominante em cátiones é alcalogênica. Em uma dieta rica em Cl
e S, a redução do BCAD pode causar as seguintes mudanças fisiológicas: aumento na
concentração de H+ no sangue, diminuição na concentração de bicarbonato sanguíneo,
descenso do pH sanguíneo, redução do bicarbonato urinário, redução do pH urinário e
redução da ingesta de matéria seca (Charbonneau et al., 2006; Oetzel, 2000). Esses efeitos
são importantes na vaca seca, pois permitem a manipulação fisiológica da vaca através
de modificações dietéticas. O descenso do pH sanguíneo mobiliza Ca ósseo para
neutralizar a acidez (sai como carbonato de Ca), ao tempo que incrementa a absorção
intestinal de Ca, reduzindo a incidência do transtorno (García 1997; Oetzel 2002).
Trabalhos sugerem que o uso do BCAD resulta em: (a) diminuição dos casos clínicos de
HP (Chan et al., 2005; Charbonneau et al., 2006; Oetzel, 1993; Oetzel, 2002); (b)
incremento na produção de leite (Oetzel, 1993; Risco, 2001a); e (c) melhora no
desempenho reprodutivo (Oetzel, 1993a; Risco, 2001a). Outros trabalhos sugerem que
um BCAD menos positivo reduziria os níveis de K, o que aumentaria a absorção de Mg,

333
melhorando a capacidade das vacas para manter a homeostase do Ca (Roche, 2003). A
acidificação artificial da ração só é possível de forma limitada porque sua palatabilidade
diminui se o BCAD reduz mais de 300 mEq/kg MS de alimento. Deve evitar-se uma
diminuição da ingesta nesta fase crítica porque aumenta o risco de aumentar o BEN
(Stöber, 2005).
- Administração de géis de cálcio: Tem por objetivo normalizar a calcemia ao parto e
reduzir a concentração de ácidos graxos não esterificados e de beta-hidroxibutirato no
pós-parto (Corbellini, 2000). O cloreto de Ca (ClCa2) fornecido vários dias antes do parto
e por 1 a 2 dias após o parto deu resultados efetivos para diminuir a incidência de HP
clínica (Oetzel, 1993; Horst et al., 1997; Dhiman & Sasidharan, 1998; Stöber, 2005).
Também é usada uma pasta comercial de ClCa2 (54 g de Ca) fornecida antes do parto e
12 e 24 horas após o parto (Goff & Horst, 1994; Hernández, 1999). A incidência de HP
clínica e deslocamento de abomaso diminuíram significativamente (Oetzel, 1993). A
solução de ClCa2 e preparações de gel oferecem um produto muito solúvel com 36% de
Ca rapidamente absorbido (Horst et al.,1997). Entretanto, soluções e géis de ClCa2
possuem desvantagens, pois podem ser cáusticas e causar ulceração em boca, esófago e
mucosa digestiva (Goff & Horst, 1994; Goff et al., 2004; Oetzel, 1993; Thilsing et al.,
2002). O ClCa2 reduz o pH sanguíneo, o que pode ser benéfico no período do periparto
da vaca porque há um efeito aditivo de ânions na dieta pré-parto e, consequentemente,
preventivo da hipocalcemia, aumentando a sensibilidade dos tecidos ao PTH (Horst et
al., 1997). Contudo, doses orais excessivas de ClCa2 podem induzir acidose metabólica e
causar inapetência em um momento em que a ingesta de alimento já está comprometida
(Goff & Horst, 1993; Goff & Horst, 1994). Uma alternativa aos sais de Ca, é o propionato
de Ca, que é fornecido em um preparado de pasta espesso e pode aumentar o Ca
sanguíneo. Os efeitos de aumento da calcemia não são tão rápidos como com o ClCa2
mas a atividade do propionato de Ca é mais prolongada (Goff & Horst, 1994), não tem
efeito acidificante do pH sanguíneo (Goff & Horst, 1994) e serve como precursor
gliconeogénico diminuindo o BEN (Alonso, 1997; Corbellini, 2000; Goff, 2004). A
desvantagem é que tem apenas 21% de Ca, pelo que requer o fornecimento de grande
volume de preparado oral.

334
Perdas econômicas causadas pela hipocalcemia
Em países como Argentina e Chile, estima-se uma prevalência de HP em vacas de alta
produção entre 6 a 16% (Corbellini, 2000; Contreras, 2002b). Na Suécia, estima-se um
risco de incidência de 5 a 10% (Houe et al., 2001) enquanto nos EUA consideram-se
valores entre 5 a 9% do total de vacas leiteiras (Goff et al., 1989; Mulligan & Doherty,
2008; Wang et al., 1994). Estudos norte-americanos (Horst et al., 1997) concluem que a
HP é um transtorno que causa graves perdas econômicas fundamentalmente por causa de
custos de tratamento, mortes (15 a 30% dos animais são refratários a uma ou duas
aplicações parenterais de borogliconato de Ca) e complicações secundárias, como atonia
ruminal, perda de apetite, mastite clínica, retenção de placenta, metrite, degeneração e
necrose de células musculares (principalmente membros posteriores), e pneumonia por
aspiração (Corbellini, 1998), podendo reduzir a vida produtiva da vaca em 3,4 anos
(Alonso, 1997; Horst et al., 1997).
O custo médio por casos de HP foi estimado em U$334/vaca (Corbellini, 1998; Goff et
al., 2004; Horst et al., 1997). Este valor está baseado nos custos diretos associados com
tratamento de casos clínicos e com perda de produção. A presença de HP aumenta a
incidência de cetose, mastite, metrite, partos distócicos, deslocamento de abomaso,
prolapso uterino e retenção de placenta (Hernández et al., 1999; Horst et al., 1997; Risco,
1984). Pode-se afirmar que vacas afetadas de HP têm entre 3 a 9 vezes mais
possibilidades de sofrer outros distúrbios no periparto (Wang et al., 1994). O custo direto
por tratamentos da HP clínica em EUA tem sido estimado em U$15 milhões, enquanto
problemas secundários causados pela HP podem incrementar o custo anual da doença em
mais de U$120 milhões (Goff et al., 1989; Wang et al., 1994). A prevenção desta
enfermidade pode sem dúvida aumentar de grande maneira a rentabilidade da produção
leiteira (Wang et al., 1994).
Referências
Afshar S., Rasooli A., Nouri M., Shahryari A., Khosravi M., Constable P. (2018). Effect of postparturient
oral calcium administration on serum total calcium concentration in Holstein cows fed diets of different
dietary cation-anion difference in late gestation. Research in Veterinary Science, 117: 118-124.
Albornoz L. (2006). Hipocalcemia puerperal: variaciones de minerales en el periparto y evaluación de
tratamientos. Tesis de Maestría en Salud Animal. Facultad de Veterinaria, Universidad de La República,
Uruguay.

335
Albornoz L. (2014). Estudio comparativo del balance energético negativo peripartal en vacas lecheras en
diferentes sistemas de producción en Uruguay y España. Tesis de Doctorado. Facultad de Veterinaria,
Universidade de Santiago de Compostela, Lugo, España.
Alonso A.J. (1997). Profilaxis de la paresia puerperal hipocalcémica bovina. Med Vet 14: 610-614.
Bauman D. E., Currie W. B. (1980). Partitioning of nutrients during pregnancy and lactation: a review of
mechanisms involving homeostasis and homeorhesis. J Dairy Sci 63: 1514-1529.
Bednarek D., Kondracki M., Gzeda M. (2000). Magnesium and calcium in prophylaxis of parturient paresis
in dairy cows. Abstracts. Medycyna Weterynaryjna 56: 367-371.
Blood D.C, Radostits O.M. (1992). Medicina Veterinaria. Séptima edición. McGraw-Hill. 1059 p.
Bostedt H., Bless S. (1993). Investigations of the effect of various prophylactic measures on the incidence
of bovine milk fever. Abstracts. Tierarztliche Umschau 48: 424-431.
Call J.W., Butcher J.E., Shupe J.L, Lamb R.C., Boman R.L., Olson A.E. (1987). Clinical effects of low
dietary phosphorus concentrations in feed given to lactating dairy cows. Am J Vet Res 1: 133-136.
Chan P.S., West J.W., Bernard J.K., Fernandez J.M. (2005). Effects of dietary cation-anion difference on
intake, milk yield, and blood components of the early lactation cow. J Dairy Sci 88: 4384-4392.
Charbonneau E., Pellerin D., Oetzel G. (2006). Impact of lowering dietary cation-anion difference in
nonlactating dairy cows: a meta-analysis. J Dairy Sci 89: 537-548.
Contreras, P. (1998). Síndrome de movilización grasa en vacas lecheras al inicio de la lactancia y sus
efectos en salud y producción de los rebaños. Arch Med Vet 30:17-27.
Contreras P. (2002a). Hipomagnesemia: efectos y procedimientos de prevención en los rebaños. VII
Congreso de la Sociedad Española de Medicina Interna Veterinaria. León: Universidad de León: 20-29.
Contreras P. (2002b). Paresia puerperal hipocalcémica. VII Congreso de la Sociedad Española de Medicina
Interna Veterinaria. León. Universidad de León: 30-34.
Corbellini C. (1997). Nutrición y fertilidad en vacas lecheras influencia del plano energético proteico y
formas de monitoreo. Primeras Jornadas de Reproducción Bovina. Villa María, Argentina, p. 21-27.
Corbellini C. (1998). Anais do Seminário Internacional sobre Deficiências Minerais em Ruminantes.
UFRGS, Porto Alegre, RS, Brasil.
Corbellini C. (2000). Influencia de la nutrición en las enfermedades de la producción de las vacas lecheras
en transición. XXI Congreso Mundial de Buiatría, Punta del Este, Uruguay, 689: 16.
Curtis, C.R., Erb, H.N., Sniffen, C.J., Smith, R.D., Powers, P.A., Smith, M.C., White, M.E., Hillman, R.B.,
Pearson, E.J. (1983). Association of parturient hypocalcemia with eight periparturient disorders in Holstein
cows. JAVMA 183: 559-561.
De Garis P.J., Lean I.J. (2009) Milk fever in dairy cows: A review of pathophysiology and control
principles. The Veterinary Journal 176: 58–69.
Dhiman T.R., Sasidharan V. (1998). Effectiveness of calcium chloride in increasing blood calcium
concentrations of periparturient dairy cows. J Anim Sci 77: 1597-1605.
Dishington I.W. (1975). Prevention of milk fever (hypocalcemic paresis puerperalis) by dietary salt
supplements. Acta Vet Scand 16: 503-512.
Dyk, P.B., Emery, R.S., Liesman, J.L., Bucholtz, H.F., VandeHaar, M.J. (1995). Prepartum non esterified
fatty acids in plasma are higher in cows developing periparturient health problems. J Dairy Sci 78 (Suppl.
1): 264, Abstract P337.
Ferguson J.D., Galligan D.T., Thomsen N. (1994). Principal descriptors of body condition score in Holstein
cows. J Dairy Sci 77: 2695.

336
Fernández G. (2009). El periodo de transición en la vaca lechera. Sistema de Revisiones en Investigación
Veterinaria de San Marcos. Seminario Avanzado de Investigación-Cajamarca, Universidad Nacional de
Cajamarca, Perú, p. 1-5.
García A. (1997). El empleo del balance iónico para prevenir la hipocalcemia. Prácticas Veterinarias,
Uruguay. Año 3: 78.
Gerloff, B., Swenson J. (1996). Acute recumbency and marginal phosphorus deficiency in dairy cattle.
JAVMA 5: 716-719.
Goff J.P., Kehrli M.E., Horst R.L. (1989) Periparturient hypocalcemia in cows: prevention using
intramuscular parathyroid hormone. J Dairy Sci 72: 1182-1187.
Goff J.P., Horst R.L. (1993). Oral administration of calcium salts for treatment of hypocalcemia in cattle.
J Dairy Sci 76: 101-108.
Goff J.P., Horst R.L. (1994). Calcium salts for treating hypocalcemia: carrier effects, acid base balance,
and oral versus rectal administration. J Dairy Sci 77: 1451-1456.
Goff J.P., Jardon P.W., Wedom J., Horst R.L. (1995). Field trials of an oral calcium propionate paste as an
aid in preventing hypocalcemia and ketosis. J Dairy Sci 78 Suppl 1,105: 186.
Goff J.P., Horst R.L. (1997). Effects of the addition of potassium or sodium, but not calcium, to prepartum
rations on milk fever in dairy cows. J Dairy Sci 80: 176-186.
Goff J.P., Ruiz R., Horst L. (2004). Relative acidifying activity of anionic salts commonly used to prevent
milk fever. J Dairy Sci. 87: 1245-1255
Goff J.P. (2008). The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in
dairy cows. The Veterinary Journal 176: 50–57
Hernandez J., Risco C., Elliot J. (1999). Effect of oral administration of a calcium chloride gel on blood
mineral concentrations, parturient disorders, reproductive performance, and milk production of dairy cows
with retained fetal membranes. JAVMA 215: 72-76.
Holmes C.T. (2003). Trastornos del metabolismo del calcio en vacas lecheras y su prevención.
Residenciado de la Pontificia Universidad Católica de Chile. Facultad de Agronomía e Ingeniería Forestal.
Departamento de Zootecnia.
Horst R.L., Goff J.P., Reinhardt T.A. (1994). Symposium: Calcium metabolism and utilization. J Dairy Sci
77: 1936-1951.
Horst R.L., Goff J.P., Reinhardt T.A. (1997). Strategies for preventing milk fever in dairy cattle. J Dairy
Sci 80:1269-1280.
Horst R.L., Kimura K., Goff J.P. (1998). Effect of mastectomy on plasma calcium and vitamin A and E
metabolism in the periparturient dairy cows. National Animal Disease Center, 10th International Conference
on Production Diseases in Farm Animals A8: 34.
Houe H., Ostergaard S., Thilsing Hansen T., Jorgensen R.J., Larsen T., Sorensen J.T., Agger J.F., Blom
J.Y. (2001). Milk fever and subclinical hypocalcaemia. An evaluation of parameters on incidence risk,
diagnosis, risk factors and biological effects as input for a decision support system for disease control. Acta
Vet Scand 42: 1-29.
Hove K., Hilde B.L. (1984). Plasma calcium in the lactating cow: dependence on continuous intestinal
absorption. Can J Anim Sci 64 (Suppl.): 227-228.
Jonsson G., Pehrson B. (1970). Trials with prophylactic treatment of parturient paresis. Vet. Rec. 87: 575-
583.
Kojouri G.A., Karimzadeh J. (2002). A study of serum calcium, phosphorus and magnesium concentrations
in cattle affected with milk fever in Shahrekord district. Abstracts. Journal Faculty Veterinary Medicine,
University of Tehran 57: 69-71.

337
Lean I.J., De Garis P.J., McNeil D.M., Block E. (2006). Hypocalcemia in dairy cows: meta-analysis and
dietary cation anion difference theory revisited. J Dairy Sci 89: 669-684.
Lotthamer K.H. (1992). Influencia de algunos factores nutricionales sobre metabolitos, enzimas y
minerales en el suero sanguíneo y leche en vacas lecheras. XX Jornadas Uruguayas de Buiatría, VII
Congreso Latinoamericano.
Manston B. (1967). The influence of dietary calcium and phosphorus concentration on their absorption in
the cow. J Agric Sci 68: 263-268.
Mulligan F.J., Doherty M.L. (2008). Production diseases of the transition cow. The Veterinary Journal 176:
3-9.
Oetzel G. (1993). Use of anionic salts for prevention of milk fever in dairy cattle. The Compendium 15:
1138-1146.
Oetzel G. (1996). Effect of calcium chloride gel treatment in dairy cows on incidence of peripartum
diseases. JAVMA 209: 958-961.
Oetzel G. (2000). Management of dry cows for the prevention of milk fever and other mineral disorders.
Veterinary Clinics Food Animal 16: 369-386.
Oetzel G. (2002). The dietary cation-anion difference concept in dairy cattle nutrition: possibilities and
pitfalls. XXII World Buiatrics Congress Hannover Alemania: 198-208.
Overton T.R., McArt J.A., Nydam D. (2017). A 100-year review: metabolic health indicators and
management of dairy cattle Journal of Dairy Science 100: 10398-10417.
Payne J.M. (1981). Enfermedades metabólicas de los animales zootécnicos. Ed. Acribia Zaragoza, España.
Queen G. (1993). Effects of oral administration of a calcium-containing gel on serum calcium concentration
in postparturient dairy cows. JAVMA 202: 607-609.
Radostits O.M., Gay C.C., Blood D.C., Hinchcliff K.W. (2001) Medicina Veterinaria. Tratado de las
enfermedades del ganado bovino, ovino, porcino, caprino y equino. 9ª edición. v. II, 1687-1698.
Rajaratne A.A., Scott D., Buchan W. (1994). Effects of a change in phosphorus requirement on phosphorus
kinetics in the sheep. Res Vet Sci 56: 262-264.
Reinhardt T.A., Lippolis J.D., McCluskey B.J., Goff J.P., Horst R.L. (2011). Prevalence of subclinical
hypocalcemia in dairy herds. The Veterinary Journal 188: 122–124.
Risco C., Reynolds J., Hird D. (1984). Uterine prolapse and hypocalcemia in dairy cows. JAVMA 185:
1510-1513.
Risco C. (2001a). Management of the postpartum dairy cow to maximize pregnancy rate. 34th Annual
Conference of the American Association of Bovine Practicioners, Vancouver.
Risco C. (2001b). Nutritional management of dry cows and calving related disorders. 34th Annual
Conference of the American Association of Bovine Practitioners, Vancouver.
Roberts T., Chapinal N., LeBlanc S.J., Kelton D.F., Dubuc J., Duffield T.F. (2012). Metabolic parameters
in transition cows as indicators for early-lactation culling risk. J. Dairy Sci. 95: 3057-3063.
Roche J.R. (2003). Hypocalcaemia and DCAD for the pasture-based transition cow. A review. Acta Vet
Scand Suppl. 97: 65-74.
Roche J.R., MacDonald K.A., Schütz K.E., Matthews L. R., Verkerk G. A., Meier S., Loor J.J., Rogers
A.R., McGowan J., Morgan S.R., Taukiri S., Webster J.R. (2013). Calving body condition score affects
indicators of health in grazing dairy cows. J Dairy Sci 96: 5811-25.
Rosol T.J., Chew, D.J., Nagode, L.A., Schenck, P. (2000). Disorders of calcium. hipercalcemia and
hipocalcemia. In: Dibartola, S.P. Fluid therapy in small animal practice, Ed. Saunders, Philadelphia, pp.
108-162.

338
Santschi D.E., Lefebvre, D.M., Cue, R.I., Girard, C.L. (2011). Incidence of metabolic disorders and
reproductive performance following a short (35-d) or conventional (60-d) dry period management in
commercial Holstein herds. J Dairy Sci 94: 3322-3330.
Stöber M. (2005). Medicina interna y cirugía del bovino. Paresia puerperal hipocalcémica. Editorial
Intermédica 4ª Ed. pp 1125-1132.
Thiede M.A. (1994). Parathyroid hormone related protein: a regulated calcium mobilizing product of the
mammary gland. J Dairy Sci 77: 1952-1963.
Thilsing H.T., Jorgensen R., Ostergaard, J. (2002). Milk fever control principles: a review. Acta Vet Scand
43: 1-19.
Underwood E.J. (1983). Los minerales en la nutrición del ganado. Editorial Acribia, Zaragoza, España.
Van Saun, R.J. (1997). Prepartum nutrition: the key to diagnosis and management of periparturient
disease. http://vbs.psu.edu/extension-
1/copy_of_resources/pdf/transitioncowhealthandnutrition/prepartum%20nutr.pdf/view. Acessado em:
31/05/2014.
Van Saun R.J. (2010). Indicators of dairy cow transition risks metabolic profiles revisited. XXVI Congreso
Mundial de Buiatría. Santiago, Chile. pp 65-77.
Venjakob P.L., Borchardt S. (2017). Hypocalcemia-cow-level prevalence and preventive strategies in
German dairy herds. Journal of Dairy Science 100: 9258-9266.
Wang C., Risco C., Donovan G., Merrit A., Beede D., Velez J. (1994). Recent advances in prevention of
parturient paresis in dairy cows. The Compendium 10: 1373-1377.

339
Impacto da nutrição na composição e qualidade do leite
Leopoldo Braz Los
O leite tem uma participação indispensável a alimentação humana, principalmente nos
primeiros anos de vida, fornecendo um balanço perfeito de macro e micronutrientes, como
proteínas de alta qualidade, carboidratos, gordura e sais minerais indispensáveis para o
crescimento, desenvolvimento e manutenção da saúde. A qualidade do leite como matéria
prima é um dos maiores requisitos para a consolidação da indústria de lácteos. O controle de
qualidade nas últimas décadas vem restringindo as adulterações do leite in natura como a
determinação de acidez, índice crioscópico e densidade. Segundo Chapaval (1999), um leite
de boa qualidade tem as seguintes características:
• Livre de microrganismos
• Baixa contagem de células somáticas
• Livre de sedimentos e materiais estranhos
• Sabor levemente adocicado e levemente aromático
• Livre de odores e aromas estranhos
• Estar de acordo com os padrões legais para os componentes.
Nos últimos anos os laticínios em vários países têm aplicado programas de melhorias de
qualidade nas fazendas, com medidas que asseguram a segurança da utilização do leite para
produtos lácteos. Essas práticas de produção devem assegurar que o leite seja produzido de
animais saudáveis, livre de resíduos e com sustentabilidade. Os pilares de um programa de
boas práticas de produção são: saúde animal, higiene de ordenha, alimentação animal, bem-
estar animal e ambiente.
Algumas indústrias de leite no Brasil já têm feito um sistema de pagamento pelo leite de
acordo com a qualidade, tanto em contagem de células somáticas e qualidade microbiológica,
quanto em sólidos do leite, como gordura e proteína que impactam diretamente no
rendimento industrial para a produção dos derivados do leite. Com isso a qualidade do leite,

340
não só o volume de leite tem sido o foco de trabalho pelo produtor porque influencia
diretamente na rentabilidade da atividade.
A quantidade e qualidade do leite produzido pela vaca depende de vários fatores como a
nutrição, raça, estágio de lactação, condição ambiental, saúde, técnica e frequência de
ordenha e seleção genética.
Os fatores nutricionais são os que podem ser controlados mais diretamente e que tem
respostas de prazo mais curto, mas que demandam de um conhecimento mais aprofundado
em manejo dos animais, metabolismo, fermentação ruminal e secreção do leite.
O programa nutricional adequado, o qual possui uma formulação e um manejo alimentar
adequado, proporciona além de um alto volume de leite produzido, um leite com qualidade
com altos níveis de sólidos de acordo com cada raça, os animais saudáveis sem problemas
metabólicos, permitindo que a vaca reproduza e persista no rebanho e tenha alta eficiência
de conversão dos alimentos em leite o que torna a atividade rentável ao produtor de leite.
Os principais componentes do leite são a gordura, a proteína e a lactose, que são sintetizadas
por células que estão nos alvéolos da glândula mamárias através de nutrientes que são
transportados pelo sangue.
Gordura do leite
A gordura do leite é componente que mais sofre variação, podendo variar de 2 a 3 pontos
percentuais. Em geral, é o componente com maior facilidade de manipulação pelo
nutricionista, mas multifatorial, onde a composição da dieta, manejo alimentar, conforto
animal, raças e caraterísticas individuais tem impacto relevante no conteúdo.
Os ácidos graxos de cadeia curta são produzidos no úbere a partir de ácidos graxos voláteis
de cadeia curta (VFA), principalmente acetato e butirato, produzidos a partir da fermentação
da fibra no rúmen. Além disso, alguns ácidos graxos de cadeia curta no leite são feitos de
beta-hidroxibutirato circulando na corrente sanguínea. Ácidos graxos de cadeia longa não
são produzidos no úbere, mas vêm dos ácidos graxos da dieta, dos microrganismos do rúmen
e da gordura corporal da vaca. Os ácidos graxos de cadeia mais longa no leite são extraídos
diretamente dos ácidos graxos circulantes no sangue. Os ácidos graxos de cadeia curta e
cadeia longa são combinados (cerca de 50/50) para formar a gordura do leite.

341
No rúmen cerca de 60 a 90% das ligações insaturadas de ácidos graxos das gorduras que
entram no rúmen via alimentação os microrganismos ruminais conseguem saturar. Ácidos
graxos totalmente saturados ou ácidos graxos monoinsaturados com configuração trans
(átomos de hidrogênio em ambos os lados da ligação dupla e não no mesmo lado da ligação
dupla, como na configuração cis) podem escapar do rúmen. Quantidades aumentadas desses
ácidos graxos trans no intestino estão correlacionadas com a baixa síndrome da gordura do
leite. A síntese mamária de gordura a partir de AGV de cadeia curta é inibida pelos ácidos
graxos trans que chegam à glândula mamária.
A quantidade de ácidos graxos trans que chegam ao intestino da vaca é influenciada por
vários fatores. Quando as vacas são alimentadas com dietas contendo grandes quantidades
de ácidos graxos insaturados disponíveis no rúmen, mais ácidos graxos trans escapam do
rúmen e chegam ao intestino. Vacas com acidose ruminal aumentam também a passagem de
ácidos graxos trans do rúmen. A diminuição na relação ruminal acetato: propionato
observada com a acidose ruminal é um sinal de alteração na fermentação ruminal, que
também aumenta a passagem ruminal de ácidos graxos trans e diminui a concentração de
gordura no leite.
A gordura do leite é totalmente dependente dos ácidos graxos de cadeia curta, ao redor da
metade da gordura do leite é feita a partir de ácidos graxos de cadeia curta, especificamente
acetato e butirato. Estes são feitos no rúmen a partir da fermentação das frações de fibra da
dieta. Uma boa fermentação da fibra se faz com alimentação forrageiras de alta
digestibilidade e subprodutos, controlando o pH do rúmen, controlando os níveis de gordura
disponível na dieta (< 5%) e fornecendo quantidades adequadas de nitrogênio e aminoácidos
no rúmen. Quando as gorduras alimentares são suplementadas para atender às necessidades
gerais de energia da vaca, sem extrapolar os limites, aumentam a síntese da gordura do leite,
aumentam a produção de leite, sem causar alterações no teor de gordura do leite.
Ácidos graxos insaturados disponíveis no rúmen vêm de fontes vegetais. Gorduras em
sementes inteiras, como soja integral e sementes de algodão inteiras, estão lentamente
disponíveis no rúmen. Esses ácidos graxos insaturados lentamente disponíveis geralmente
serão completamente alterados para ácidos graxos saturados antes de deixarem o rúmen. Mas
se o ambiente ruminal estiver comprometido ou se grandes quantidades de sementes forem
alimentadas, as gorduras podem sair do rúmen como ácidos graxos trans parcialmente

342
saturados que podem levar à depressão da gordura do leite. Se alimentar com óleos vegetais,
pode se esperar que alguns deles deixem o rúmen como ácidos graxos trans e potencialmente
causem impacto na síntese da gordura do leite.
Proteína do leite
A proteína do leite é um componente que sofre menor variação quando comparada à gordura,
também é multifatorial, e de difícil manipulação pelo nutricionista. A proteína do leite na
glândula mamária é sintetizada principalmente a partir de aminoácidos no sangue e são os
precursores primários usados para sintetizar a proteína do leite. Do nitrogênio total do leite,
aproximadamente 76% corresponde a caseína, 18% a proteínas do soro do leite e 6% de
nitrogênio não proteico. A produção de proteína é geralmente limitada pelo aminoácido que
tem o menor suprimento em relação à exigência da vaca. Esse aminoácido é chamado de
“primeiro aminoácido limitante”. É o elo perdido da cadeia de proteínas e, quando é usado,
a produção de proteínas é interrompida. A vaca recebe aminoácidos no intestino de duas
fontes primárias. Os microrganismos do rúmen fornecem 50 a 75% dos aminoácidos. Por
isso a energia contida na dieta pode limitar a síntese de proteína do leite, já que limita a
síntese de proteína microbiana, e a proteína não degradável no rúmen (proteína bypass)
fornece o restante. A eficiência da conversão do nitrogênio da dieta em proteína do leite pela
vaca é bastante baixa (25 a 30%).
A vaca usa muitos aminoácidos para a manutenção de seus sistemas vitais, como o
funcionamento do intestino, fígado e outros tecidos. Isso faz com que a proteína do leite seja
difícil de mudar nutricionalmente. Fazer o balanceamento de aminoácidos, em vez de
simplesmente utilizar as quantidades de aminoácidos individuais disponíveis para a produção
de proteínas na glândula mamária, é mais efetivo para o aumento da proteína do leite a
produção de proteína do leite.
O entendimento do uso de aminoácidos pelo nutricionista permite que se trabalhe com
precisão no equilíbrio das dietas para a melhora de proteína do leite. São necessárias mais
pesquisas para ajudar os nutricionistas a prever quanto de cada aminoácido será produzido
pelos microrganismos do rúmen, quanto que cada aminoácido passa pelo rúmen e é absorvido
no intestino delgado, e análises laboratoriais acessíveis para determinar o conteúdo de
aminoácido de cada ingrediente da dieta.

343
Estratégias para aumentar a proteína do leite
A fermentação em ruminantes é resultado de um processo evolutivo de características
anatômicas e simbióticas a qual permite que se utilize carboidratos estruturais como fontes
de energia e compostos nitrogenados não proteicos como fonte de proteína. O resultado da
fermentação são os ácidos graxos de cadeia curta, proteína microbiana, metano, dióxido de
carbono, vitaminas do complexo B e K, etc.
Maximizar a produção de proteína microbiana é fundamental numa estratégia de aumento de
proteína, o perfil de aminoácidos dos microrganismos é muito semelhante ao da proteína do
leite, fazendo com que sua proteína tenha alto potencial de conversão em proteína do leite.
Com isso o fornecimento de forragens de alta digestibilidade, maximizando o consumo de
matéria seca, evitando a acidose subclínica, fornecendo quantidades adequadas de proteína
solúvel e degradável, e sincronizando os carboidratos e proteínas disponíveis no rúmen de
hora em hora no rúmen é a principal estratégia de aumento de proteína no leite.
O pool de aminoácidos da proteína que é não degradável no rúmen também afetará a
produção de proteína do leite, mas este tem dependência de ingredientes que são utilizados
na dieta, já que cada um tem uma composição de aminoácidos específica e uma quantidade
de proteína que não é degradada no rúmen. O milho e seus subprodutos, tais como grãos de
destilaria e farelo de glúten de milho, são conhecidos por serem baixos em lisina. A soja é
conhecida por ser baixa em metionina. Há também aminoácidos de bypass (protegidos da
degradação ruminal) individuais que agora estão sendo incorporados nonas dietas. Os
aminoácidos que passam da degradação ruminal, devem fornecer um perfil de aminoácidos
que complemente a proteína microbiana produzida no rúmen. O objetivo é combinar as duas
fontes de aminoácidos para fazer um suprimento de aminoácidos intestinais semelhante ao
necessário para a produção de proteína do leite. Nas dietas características do Brasil, a
metionina é o aminoácido mais limitante devido ao perfil de concentrado utilizado para a
suplementação de proteína na dieta, o farelo de soja. Muitas vezes quando se trabalha com
outra fonte de concentrado para fazer o equilíbrio de aminoácidos, ao exemplo da utilização
de subprodutos de milho para suprir a deficiência de metionina na dietas com farelo de soja,
a redução de farelo de soja para a entrada destes na dieta causa uma deficiência de outro
aminoácido como neste caso a lisina, sendo desta forma na maioria das vezes necessário a

344
correção com uma fonte de aminoácido bypass específico para o aminoácido limitante, para
não causar a deficiência. A maioria dos estudos com aminoácidos suplementares de bypass
aumenta o rendimento e proteína do leite.
O conteúdo energético da dieta energia é necessária para manter a produção e proteína do
leite. No início da lactação, o aumento da energia estimula a produção de leite e quantidade
proteína do leite produzida, com pouco efeito sobre a porcentagem de proteína no leite. Mais
tarde na lactação, a energia aumenta a concentração de proteína no leite até certo ponto. Parte
dessa resposta na proteína do leite pode ser devida à glicose e acetato extra disponíveis no
úbere, mas a energia adicionada pode causar mais um aumento na síntese de proteína
microbiana que aumenta a oferta de aminoácidos no úbere.
A alimentação de mais carboidratos disponíveis no rúmen pode aumentar a produção de
proteína do leite. A combinação da fermentação da energia ruminal combinada com as
frações proteicas também pode ser eficaz na melhoria da eficiência do nitrogênio. Existem
diferenças substanciais entre as fontes de amido e dentro dos grãos, devido ao processamento
nas taxas de liberação de energia no rúmen. Como exemplo, a digestibilidade ruminal do
amido diminui de 70% com o milho moído para 54% com o milho moído grosseiramente.
Além disso, tem sido demonstrado que a substituição de alguns dos amidos da dieta por
açúcares de fermentação rápida pode melhorar a captura ruminal de N para a produção de
proteína microbiana influenciando no teor de proteína do leite
A gordura na dieta em excesso diminui a produção de proteína do leite, mas a razão para isso
ainda não está clara. A substituição de gorduras por carboidratos no rúmen pode deprimir a
síntese de proteína microbiana e, assim, diminuir a quantidade de aminoácidos disponíveis
para a síntese. A gordura também pode inibir o crescimento de alguns microrganismos
diretamente. Alguns nutricionistas recomendam a adição de 1% a mais de proteína não
degradável no rúmen para cada 3% de gordura adicionada em uma dieta.
Ingestão de matéria seca
Promover o adequado consumo de matéria está relacionada à minimização do balanço
proteico e energético negativo durante o início da lactação, o qual aumenta a proporção de
gordura no leite. À medida que as vacas leiteiras passam para um balanço energético positivo,

345
o peso corporal é recuperado, a perda da condição corporal é minimizada e as vacas produzem
leite com composição normal de gordura e proteína.
As vacas leiteiras de alta produção devem ingerir de 3,6 a 4,0% do seu peso corporal
diariamente como matéria seca. Se um rebanho leiteiro está consumindo menos matéria seca
do que 3,6 a 4,0% do peso corporal, a produção de gordura do leite e componentes proteicos
pode ser limitada. O aumento da frequência de alimentação aumenta a gordura do leite e o
componente proteico, especialmente com dietas ricas em fibras e com alto teor de grãos, uma
maior resposta é observada em dietas com menos de 45% de forragem e quando o
concentrado é alimentado separadamente, como na alimentação de sala de ordenha. Quando
as dietas são alimentadas em dietas totais, a frequência de alimentação não é tão importante,
desde que a ração permaneça palatável e seja alimentada pelo menos uma vez ao dia.
Gerenciamento de alimentação
Qualquer manejo em que as vacas comam anormalmente ou limite a ingestão de alimentos,
pode afetar os componentes do leite. Os exemplos incluem: falta de espaço nos cochos, lote
de novilhas com vacas mais velhas ou acima da capacidade total das camas. As dietas que
favorecem a seleção de alimentos e alimentação com pouca frequência em um sistema
convencional (não dieta total misturada TMR) tem potencial de alterar a qualidade do leite,
assim como a alimentação de dieta total se não tiver o tamanho adequado de partículas, se
não tiver suficientes diminui a ruminação e com isso reduz o tamponamento via saliva e baixa
o pH ruminal diminuindo atividade de microrganismos que degradam a fibra dietética, e se
tiver partículas com tamanho maior que o recomendado podem favorecer a seleção de
ingredientes e o animais consumam mais concentrado que volumoso e favorecer a alteração
nos componentes do leite pela falta de ruminação e queda do pH ruminal. Alimentando-as
com ingredientes proteicos antes do fornecimento de enérgicos e alimentando os
concentrados antes da forragem em sistemas não TMR, todas essas práticas de manejo da
alimentação alteram os componentes do leite de vacas leiteiras. Outras condições que podem
alterar é a limitação no número de refeições por dia (uma ou duas refeições por dia versus 10
a 15) ou permitir que as vacas comam refeições de alto concentrado em parte do tempo e
altas refeições de forragem no resto do dia. Deve se assegurar de que a dieta fresca esteja
disponível 20 horas por dia, e que a ração estragada seja removida dos cochos. Conforto,

346
sombra ou o resfriamento devem ser fornecidos durante os dias em estresse térmico para
ajudar a manter a ingestão normal e os padrões normais das refeições.
A dieta provavelmente é o fator que mais influência nos produtos finas da fermentação, no
número e proporção relativa das diferentes espécies ruminais, fazer as mudanças abruptas de
dietas fazem com que os microrganismos alterem seu balanço de fermentação, com isso abre
porta para microrganismos facultativos oportunistas dominarem a fermentação e como
exemplo através da produção de diferentes ácidos o abaixamento do pH ruminal causando
distúrbios metabólicos. Fazer a mudança gradualmente para permitir que o tempo dos
microrganismos do rúmen se adaptem e não ocorra distúrbios metabólicos e não diminua
produção de proteína microbiana no rúmen que pode reduzir a proteína do leite, e também
menor população microbiana para digerir a dieta, limita a produção de ácidos graxos de
cadeia curta no rúmen, diminuindo a produção de leite.
Um dos problemas mais comuns na mudança abrupta de dieta, é a acidose que ocorre quando
se muda de uma dieta rica em fibras para alta concentrado que é rica em carboidratos
fermentáveis (amido e açúcares). Grandes quantidades de amido e açúcar estimulam as
bactérias que produzem ácido láctico. Nesse caso, as bactérias que normalmente usam ácido
láctico não conseguem acompanhar a produção. A quantidade de acidez no rúmen é medida
por leituras de pH. Quando o pH do rúmen cai, a digestão de fibras é deprimida porque o
grupo de microrganismos que degradam a fibra são sensíveis a pH. Como os produtos da
digestão de fibras são usados para a síntese da gordura do leite, uma queda no teste da gordura
do leite é um indicio de acidose. Além disso, o acúmulo de ácido provoca um influxo de água
dos tecidos para o intestino e, portanto, um sinal comum de acidose é a diarreia. Se o pH do
rúmen diminuir e cair abaixo de 5,5 muitas outras bactérias ruminais também começam a ser
afetadas. À medida que o ácido láctico se acumula, é absorvido e diminui o pH do sangue.
Altos níveis de ácido também podem causar úlceras no rúmen resultando em infiltração de
bactérias no sangue que podem causar abscessos no fígado. A acidose subaguda também é
caracterizada por ingestão de cíclica porque os animais comem menos durante períodos de
sofrimento; então, se o rúmen se adaptar, seu apetite retorna.
Outros problemas comuns de acidose ocorrem em dietas em que a ingestão de fibra efetiva é
muito baixa, dietas formuladas com muito concentrado e pouca fibra efetiva, tamanho de
partícula abaixo do recomendado e dietas com problema de manejo alimentar onde os

347
animais conseguem selecionar os ingredientes da dieta. Quando os animais não ruminam
normalmente, a falta de saliva, que contém um tampão natural, contribui para o baixo pH
ruminal. O tamanho adequado da partícula na dieta parece necessário para evitar a da gordura
do leite baixa. As vacas necessitam de fibra e forragem para estimular a atividade
mastigatória e a produção de saliva, sendo ambas necessárias para manter um pH ruminal
adequado e um rúmen sadio.
O uso de aditivos como o tamponante bicarbonato de sódio minimizam a queda de pH quando
é utilizado dietas com grandes quantidades de concentrados rapidamente fermentáveis que
produz excesso ácidos e os sistemas tamponantes, principalmente o fluxo salivar não são
suficientes. Isso permite manter ativas as bactérias que digerem a fibra não reduzindo assim
a quantidade de ácido acético produzido que faz com que não reduza a porcentagem de
gordura.
Recomendações
− Controle de carboidratos fermentáveis; amido< 28% (%dieta total)
− Relação concentrado:forragem< 55%
− Adaptação na mudança de dieta alta forragem para alta concentração
− Evitar teor de fibra na dieta; <30% FDN e <19% forragem NDF (% dieta total)
− Tempo de mistura de dieta total em excesso, redução excessiva de tamanho de partícula;
− Seleção de alimentos pela vaca; avaliar tamanho de partícula do fornecido: sobra;
− Uso de subprodutos fibrosos em substituição aos grãos;
− Aumento na frequência de alimentação;
− Uso de dieta total mistura;
− Utilização de tamponantes na dieta;
− Baixo teor de gordura insaturada na dieta;
Observar no rebanho sintomas de acidose:
• Teste de baixo teor de gordura no leite; <3,0 a 3,3%;
• Proteína baixa do leite;
• Consumo irregular;
• Diarreia;

348
• Baixa ruminação (<50% das vacas deitadas não mastigando sua ruminação);
• Oscilação na produção de leite;
• Fezes inconsistentes no mesmo grupo de alimentação;
• Fezes espumosas, contém bolhas de gás;
• Aparência de mucina / fibrina nas fezes;
• Aumento do tamanho de partícula de fibra (> 1,0 cm) nas fezes;
• Aparência de grão não digerido, moído (≤ 0,4 cm) nas fezes;
• Eficiência de alimentação reduzida;
Conclusão
O planejamento nutricional de um rebanho pode otimizar a produção tanto de leite, mas como
componentes, mas o foco deve ser rotineiramente na produção de forragens, manejo
alimentar e nutrição. O aumento da produção de sólidos do leite são um indicado de melhor
saúde para os animais, além de aumentar o faturamento ao produtor com a venda do leite.
Referência
CHAPAVAL, L. Detecção de enterotoxinas produzidas por Staphylococcus aureus no leite bovino por
eletroforese capilar e identificação dos isolados enterotoxigênicos via PCR. 1999. 25f. Tese doutorado –
Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Piracicaba, 1999.