
Relationship of EP1-4 prostaglandin receptors with rat hypothalamic cell groups involved in...
13
Rapid Publication Relationship of EP 1-4 Prostaglandin Receptors With Rat Hypothalamic Cell Groups Involved in Lipopolysaccharide Fever Responses TAKAKAZU OKA, 1 KAE OKA, 1,2 THOMAS E. SCAMMELL, 1 CHARLOTTE LEE, 2 JOSEPH F. KELLY, 2 FRANCOIS NANTEL, 3 JOEL K. ELMQUIST, 1,2 AND CLIFFORD B. SAPER 1 * 1 Department of Neurology, Beth Israel Deaconess Medical Center and Program in Neuroscience, Harvard Medical School, Boston, Massachusetts 02115 2 Department of Medicine and Division of Endocrinology, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts 02115 3 Merck Frosst Center for Therapeutic Research, Dorval-Pointe-Claire, Quebec H9J 1R4, Canada ABSTRACT The action of prostaglandin E 2 (PGE 2 ) in the preoptic area is thought to play an important role in producing fever. Pharmacologic evidence suggests that, among the four subtypes of E-series prostaglandin (EP) receptors, i.e., EP 1 , EP 2 , EP 3 , and EP 4 , the EP 1 receptor mediates fever responses. In contrast, evidence from mice with EP receptor gene deletions indicates that the EP 3 receptor is required for the initial (,1 hour) fever after intravenous (i.v.) lipopolysac- charide (LPS). To investigate which subtypes of EP receptors mediate systemic infection-induced fever, we assessed the coexpression of Fos-like immunoreactivity (Fos-IR) and EP 1-4 receptor mRNA in nuclei in the rat hypothalamus that have been shown to be involved in fever responses. Two hours after the administration of i.v. LPS (5 mg/kg), Fos-IR was observed in the ventromedial preoptic nucleus, the median preoptic nucleus, and the paraventricular hypothalamic nucleus. In these nuclei, EP 4 receptor mRNA was strongly expressed and the Fos-IR intensely colocalized with EP 4 receptor mRNA. Strong EP 3 receptor mRNA expression was only seen within the median preoptic nucleus but Fos-IR showed little coexpression with EP 3 receptor mRNA. EP 2 receptor mRNA was not seen in the PGE 2 sensitive parts of the preoptic area. Although approx- imately half of the Fos-immunoreactive neurons also expressed EP 1 receptor mRNA, EP 1 mRNA expression was weak and its distribution was so diffuse in the preoptic area that it did not represent a specific relationship. In the paraventricular nucleus, EP 4 mRNA was found in most Fos-immunoreactive neurons and levels of EP 4 receptor expression increased after i.v. LPS. Our findings indicate that neurons expressing EP 4 receptor are activated during LPS-induced fever and suggest the involvement of EP 4 receptors in the production of fever. J. Comp. Neurol. 428: 20 –32, 2000. © 2000 Wiley-Liss, Inc. Indexing terms: prostaglandin E 2 ; ventromedial preoptic area; EP 1 receptor; EP 3 receptor; in situ hybridization Fever is an adaptive biologic response that improves survival during experimental bacteremia (Kluger et al., 1996). Prostaglandin E 2 (PGE 2 ), the predominant endog- enous isoform of PGE in the brain, is thought to be an essential mediator of fever (Blatteis and Sehic, 1997). Microinjections of PGE 1 , PGE 2 , or PGE receptor agonists into the brain in rats (Williams et al., 1977; Stitt, 1991; Grant sponsor: NIH; Grant numbers: NS33987, MH56537, and DK53301. *Correspondence to: Clifford B. Saper, Department of Neurology, Beth Israel Deaconess Medical Center and Program in Neuroscience, Harvard Medical School, Boston, MA 02115. E-mail: [email protected] Received 15 June 2000; Revised 7 August 2000; Accepted 8 August 2000 THE JOURNAL OF COMPARATIVE NEUROLOGY 428:20 –32 (2000) © 2000 WILEY-LISS, INC.
Transcript of Relationship of EP1-4 prostaglandin receptors with rat hypothalamic cell groups involved in...

Rapid Publication
Relationship of EP1-4 ProstaglandinReceptors With Rat Hypothalamic CellGroups Involved in Lipopolysaccharide
Fever Responses
TAKAKAZU OKA,1 KAE OKA,1,2 THOMAS E. SCAMMELL,1 CHARLOTTE LEE,2
JOSEPH F. KELLY,2 FRANCOIS NANTEL,3 JOEL K. ELMQUIST,1,2AND
CLIFFORD B. SAPER1*1Department of Neurology, Beth Israel Deaconess Medical Center and Program in
Neuroscience, Harvard Medical School, Boston, Massachusetts 021152Department of Medicine and Division of Endocrinology, Beth Israel Deaconess Medical
Center, Harvard Medical School, Boston, Massachusetts 021153Merck Frosst Center for Therapeutic Research, Dorval-Pointe-Claire,
Quebec H9J 1R4, Canada
ABSTRACTThe action of prostaglandin E2 (PGE2) in the preoptic area is thought to play an important
role in producing fever. Pharmacologic evidence suggests that, among the four subtypes ofE-series prostaglandin (EP) receptors, i.e., EP1, EP2, EP3, and EP4, the EP1 receptor mediatesfever responses. In contrast, evidence from mice with EP receptor gene deletions indicates thatthe EP3 receptor is required for the initial (,1 hour) fever after intravenous (i.v.) lipopolysac-charide (LPS). To investigate which subtypes of EP receptors mediate systemic infection-inducedfever, we assessed the coexpression of Fos-like immunoreactivity (Fos-IR) and EP1-4 receptormRNA in nuclei in the rat hypothalamus that have been shown to be involved in fever responses.Two hours after the administration of i.v. LPS (5 mg/kg), Fos-IR was observed in the ventromedialpreoptic nucleus, the median preoptic nucleus, and the paraventricular hypothalamic nucleus. Inthese nuclei, EP4 receptor mRNA was strongly expressed and the Fos-IR intensely colocalizedwith EP4 receptor mRNA. Strong EP3 receptor mRNA expression was only seen within themedian preoptic nucleus but Fos-IR showed little coexpression with EP3 receptor mRNA. EP2receptor mRNA was not seen in the PGE2 sensitive parts of the preoptic area. Although approx-imately half of the Fos-immunoreactive neurons also expressed EP1 receptor mRNA, EP1 mRNAexpression was weak and its distribution was so diffuse in the preoptic area that it did notrepresent a specific relationship. In the paraventricular nucleus, EP4 mRNA was found in mostFos-immunoreactive neurons and levels of EP4 receptor expression increased after i.v. LPS. Ourfindings indicate that neurons expressing EP4 receptor are activated during LPS-induced feverand suggest the involvement of EP4 receptors in the production of fever. J. Comp. Neurol. 428:20–32, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: prostaglandin E2; ventromedial preoptic area; EP1 receptor; EP3 receptor; in situ
hybridization
Fever is an adaptive biologic response that improvessurvival during experimental bacteremia (Kluger et al.,1996). Prostaglandin E2 (PGE2), the predominant endog-enous isoform of PGE in the brain, is thought to be anessential mediator of fever (Blatteis and Sehic, 1997).Microinjections of PGE1, PGE2, or PGE receptor agonistsinto the brain in rats (Williams et al., 1977; Stitt, 1991;
Grant sponsor: NIH; Grant numbers: NS33987, MH56537, and DK53301.*Correspondence to: Clifford B. Saper, Department of Neurology, Beth
Israel Deaconess Medical Center and Program in Neuroscience, HarvardMedical School, Boston, MA 02115. E-mail: [email protected]
Received 15 June 2000; Revised 7 August 2000; Accepted 8 August 2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 428:20–32 (2000)
© 2000 WILEY-LISS, INC.

Scammell et al., 1996; Oka et al., 1997) and other species(Feldberg and Saxena, 1971; Morimoto et al., 1988) haveshown that PGE acts on the region at the anteroventraltip of the third ventricle, including the organum vasculo-sum of the lamina terminalis (OVLT) and the adjacentpreoptic area (POA) to produce fever. PGE2 levels increasein this region during the febrile response to lipopolysac-charide (LPS) (Sirko et al., 1989; Van Dam et al., 1993;Sehic et al., 1996). In addition, blockade of PGE2 synthesisby microinjection of cyclooxygenase (COX) inhibitors intothe OVLT and adjacent preoptic region attenuates fever(Cranston and Rawlins, 1972; Morimoto et al., 1988;Scammell et al., 1998). However, the exact neuronal sub-strates of PGE action in producing fever remain unknown.
Prostaglandin E type (EP) receptors are divided intofour subtypes: EP1, EP2, EP3, and EP4 receptors (Colemanet al., 1994; Ushikubi et al., 1995). Despite the establishedrole of PGE2 in producing fever, the EP receptor subtypesthat are crucial in the febrile response has remained con-troversial (Oka and Hori, 1994; Parrott and Vellucci, 1996;Oka et al., 1997, 1998; Ushikubi et al., 1998). Oka andHori administered a variety of drugs with agonist andantagonist properties for the four known EP receptors todissect their role in fever in rats (Oka and Hori, 1994).They found that intracerebroventricular (i.c.v.) injectionof 17-phenyl-v-trinor- PGE2 (an EP1 and EP3a receptoragonist), but not butaprost, M&B28767, or 11-deoxy-PGE1(EP2, EP3a, and EP4 receptor agonists, respectively), mim-icked PGE2 (i.c.v.) -induced fever and that SC19220 (anEP1 receptor antagonist, i.c.v.) blocked PGE2 (i.c.v.) fever.Similar pharmacologic study in pigs has suggested thatthe EP2 or EP3 receptor is necessary to produce fever(Parrott and Vellucci, 1996). On the other hand, Ushikubiand colleagues examined mice bearing genetic deletions ofthe EP1–4 receptors. They found that only EP3 knockoutanimals failed to show an early phase of fever (up to 1hour) after injection of LPS (into the tail vein) or PGE2(i.c.v.) (Ushikubi et al., 1998). These conflicting resultsmight be due to species differences. However, in the twopharmacologic studies, none of the drugs had actions con-fined entirely to a single receptor subtype except butap-
rost (Coleman et al., 1994; Boie et al., 1997), whereas inthe study of the knockout mice, the animals lacked thespecific EP receptors during development and throughoutthe body. Hence, it is unclear whether the responses re-corded were due to the same hypothalamic EP receptorsthat cause fever in intact animals.
All four EP receptor subtypes have been reported to bedistributed in the preoptic area of the hypothalamus(Sugimoto et al., 1994; Rage et al., 1997; Zhang andRivest, 1999; Nakamura et al., 1999). Recent reports haveindicated that there is up-regulation of EP4 receptor ex-pression in several areas of the brain, including thecorticotrophin-releasing hormone neurons of the paraven-tricular nucleus, after intravenous (i.v.) LPS orinterleukin-1b (Zhang and Rivest, 1999). In addition,paraventricular neurons that express Fos after i.c.v. PGE2also express EP4 receptors (Zhang and Rivest, 2000). How-ever, there has been no comprehensive analysis of thedistribution of the EP1–4 receptors of the PGE2-sensitiveanteromedial preoptic region. It is also known that neu-rons in the ventromedial preoptic nucleus (VMPO) and themedian preoptic nucleus (MnPO) show expression of theimmediate early gene product Fos in response to i.v. LPS(Sagar et al., 1995; Elmquist et al., 1996; Elmquist andSaper, 1996). However, the relationship of the EP receptorsubtypes with the neurons in the preoptic area that dem-onstrate LPS-induced Fos expression has not been ex-plored. Finally, we wanted to determine the relationshipof the EP4 gene induction with the Fos-expressing neuronsin the paraventricular nucleus (PVH) after i.v. LPS.Therefore, we mapped the distribution of mRNA for eachof the EP receptors in the hypothalamic regions that showchanges in Fos expression after i.v. LPS, to determine anychanges in EP gene expression, and the relationship of EPreceptor expression with neurons showing LPS-inducedFos.
MATERIALS AND METHODS
Animals
Adult, male, pathogen-free, Sprague-Dawley rats (250–350 g; Taconic, Germantown, NY) were used. Rats werehoused in a light- (12 hours on/off; lights on at 7:00 AM)and temperature- (22–23°C) controlled pathogen-free fa-cility with food and water available ad libitum. The pro-cedures used were approved by the Harvard MedicalSchool and Beth Israel Deaconess Medical Center Institu-tional Animal Care and Use Committees.
LPS administration and perfusion
Intravenous catheters were surgically implanted by us-ing aseptic techniques as described previously (Elmquistet al., 1996; Scammell et al., 1996). After surgery, ratswere housed individually in a sound-attenuated room andwere handled daily (5–7 days) to minimize stress duringthe actual injection procedures. On the experimental day,rats were injected with pyrogen-free saline (PFS) (Sigma,St. Louis, MO) or LPS from Salmonella typhimurium (5mg/kg in 0.15 ml) (Sigma; lot 23H4047) followed by a flushof PFS (0.10 ml) by means of i.v. injection catheter. LPSwas dissolved in PFS. Injections were given between 8:00AM and 9:00 AM to avoid circadian differences in baselineFos expression (Kononen et al., 1990). Two hours after i.v.administration of LPS or PFS, rats were deeply anesthe-
Abbreviations
3V third ventricleAC anterior commissureADP anterodorsal preoptic nucleusAVPV anteroventral periventricular nucleusAV3V anteroventral region of the third ventricledp dorsal parvicellular partl-OVLT lateral part of the organum vasculosum of the lamina ter-
minalisLPO lateral preoptic areaMnPO median preoptic nucleusm-OVLT medial part of the organum vasculosum of the lamina ter-
minalismp medial parvicellular partMPO/LPO border region between the medial preoptic area and the
lateral preoptic areaOC optic chiasmOVLT organum vasculosum of the lamina terminalispm posterior magnocellular partPS parastrial nucleusPVH paraventricular hypothalamic nucleusPVpo preoptic periventricular nucleusVMPO ventromedial preoptic nucleusVLPO ventrolateral preoptic nucleusvp ventral parvicellular part
21EP RECEPTORS ACTIVATED DURING LPS FEVER

tized with intraperitoneal (i.p.) chloral hydrate (7%, 350mg/kg) and perfused transcardially with diethyl pyrocar-bonate (DEPC)-treated 0.9% saline followed by 10% neu-tral buffered formalin (Sigma). The brains were removed,stored in the same fixative for 4 hours, submerged in 20%sucrose in DEPC-treated phosphate buffered saline (PBS).Six series of coronal sections were cut at 25 mm. Thesections were stored at 220°C in an antifreeze solution asdescribed previously (Elias et al., 1998) until in situ his-tochemical staining was initiated.
In situ hybridization histochemistry
Rat EP1, EP2, EP3, and EP4receptor cDNAs were gener-ated as described previously (Boie et al., 1997). Antisense EPreceptor 35S-labeled riboprobes were generated from thecDNA templates. The linearized plasmids were subject to invitro transcription with Sp6 (EP1, EP2, and EP3receptors) orT7 (EP4 receptor) polymerase according to the manufactur-er’s protocols (Promega, Madison, WI). The 35S-labeledprobes were diluted to 106 cpm/ml. The hybridization cock-tail was applied on the untreated, PFS-treated, or LPS-treated brain tissue sections, and the sections were incu-bated for 12–16 hours at 57°C. The following day, sectionswere incubated in 0.002% RNase A (Boehringer-Mannheim,Indianapolis, IN) for 30 minutes and rinsed in decreasingconcentrations of saline sodium citrate buffer containing 1mM dithiothreitol. Sections were placed in X-ray film cas-settes with BMR-2 film (Eastman-Kodak, Rochester, NY) for3–5 days. Slides were subsequently dipped in NTB2 photo-graphic emulsion (Eastman-Kodak), stored at 4°C for 15–21days, and developed with D-19 developer (Eastman-Kodak).Control hybridization experiments were done on adjacentsections by using each sense probe or tissue pretreatmentwith RNase A (200 mg/ml).
Dual-label in situ hybridizationhistochemistry/immunocytochemistry
The protocol used for combined in situ hybridizationhistochemistry and immunocytochemistry was a modifi-
cation of that described previously (Elias et al., 1999).PFS- or LPS-treated tissue sections were incubated in Fosrabbit primary antiserum (Ab-5, Oncogene Sciences, Cam-bridge, MA; 1:100,000) overnight followed by biotinylateddonkey anti-rabbit IgG (1:1,000; Vector, Burlingame, CA)for 60 minutes. Sections were then incubated in avidin-biotin complex (Vector Elite Kit; 1:500) and reacted with0.04% diaminobenzidine tetrahydrochloride (Sigma) and0.01% hydrogen peroxide dissolved in DEPC-treated PBS.Sections were mounted onto SuperFrost slides (Fisher,Pittsburgh, PA) and then processed for EP receptor in situhybridization as described above.
Data analysis
The relative grain densities of EP receptor mRNA sig-nals were assessed in comparison with those of the EP4receptor mRNA on the VMPO neurons on which the high-est density of silver grains were seen in the present studyand graded according to the scale of background level (2),weak but positive (1), moderate (11), strong (111), andvery strong (1111). The number of Fos-immunoreactivecells and double-labeled cells (Fos-immunoreactive neu-rons expressing EP receptors) were quantified at a mag-nification of 3100 or 3400 under brightfield illumination,and a grid was used to delineate the area of quantification.In the region surrounding the anteroventral tip of thethird ventricle (AV3V area), including portions of theOVLT and MnPO, a 600 3 600 mm grid was located so thatthe lower border abutted the third ventricle (3V) andextended 300 mm right and left of the midline over theOVLT (Fig. 1A). For the VMPO, a grid was located so thatthe medial edge abutted the 3V and extended 500 mmlaterally and 200 mm above and below the ventrolateralcorner of the 3V, at a level of just posterior to the OVLT(Fig. 1B) as described previously (Elmquist et al., 1996).For the MnPO, a 300 3 500 mm (horizontal 3 vertical)grid was set between the anterior commissure and the 3V(Fig. 1C). For the border region between the medial pre-optic area and the lateral preoptic area, a 300 3 700 mm
Fig. 1. The four boxed areas in which the number of Fos-immunoreactive cells and double-labeled cells (Fos-immunoreactiveneurons expressing EP receptors) were counted. Coronal sections ofthe rat brain are illustrated. Boxed areas represent the region sur-rounding the anteroventral tip of the third ventricle (AV3V), includ-ing the portions of the organum vasculosum of the lamina terminalis
(OVLT) and median preoptic nucleus (MnPO) (A), the ventromedialpreoptic nucleus (VMPO) (B), the MnPO (C), and the border regionbetween the medial preoptic area and the lateral preoptic area (MPO/LPO) (D). For other abbreviations, see list. Scale bar 5 500 mm in B(applies to A–D).
22 T. OKA ET AL.

(horizontal 3 vertical) grid was set so that the medialborder was located just above the lateral edge of the opticchiasm and the lower border was abutted on the brainsurface (Fig. 1D). Double-labeled cells were those thatexhibited a clearly brown nucleus (Fos-IR) and a cluster ofsilver grains (EP receptor mRNA) over the cell body de-lineating a region with hybridization levels at least 33above background.
The mean silver grain density over the parvicellular andmagnocellular PVH were assessed as percentage optical den-sity. Briefly, digital darkfield photomicrographs were ac-quired in Adobe Photoshop (Adobe Systems, Mountain View,CA). The gray-scale was inverted and adjusted for maximalcontrast (background around silver grains 5 0, silver grains5255). Images were then imported into NIH Image 1.62, andthe mean density (corresponding to percent of field occupiedby silver grains) was calculated over the third ventricle(background labeling). This density was then subtractedfrom the density measured over the parvicellular and mag-nocellular PVH. The result was rescaled (0 5 backgroundlabeling density over third ventricle, 100 5 pure black field)and presented as percentage optical density 6 SEM. Stu-dent’s t-test (in case of equal variance) or Welch’s t-test (incase of unequal variance) were used to compare the numberof Fos-immunoreactive cells between the PFS-treatedgroups and the LPS-treated ones.
Production of photomicrographs
Photomicrographs were produced by capturing imageswith a digital camera (Kodak DCS 460; Eastman-Kodak)mounted directly on the microscope (Zeiss Axioplan), anddownloaded to an Apple Macintosh Power PC computer(Apple Computers, Cupertino, CA). Image-editing soft-ware (Adobe Photoshop) was used to combine photomicro-graphs into plates. Only the brightness, sharpness, andcontrast were adjusted. Figures were printed on a dye-sublimation printer (Kodak XLS8600PS; Eastman-Kodak). Cytoarchitectonic illustrations were made by us-ing a camera lucida and importing images into Canvas 5(Deneba Systems, Miami, FL).
RESULTS
EP receptor distribution within the PGE2-sensitive anteromedial preoptic region
Among the four subtypes of EP receptors examined, EP4receptor mRNA was the most strongly expressed within thePGE2-sensitive part of the preoptic area. In contrast to thedistinct localization of EP4and EP3 receptor mRNAs, expres-sion of EP1 receptor mRNA was weak and diffuse through-out this region and no EP2 receptor mRNA was detected(Table 1). For EP3 and EP4 receptor mRNA, silver grainswere condensed in clusters primarily over neuronal cell bod-ies and did not represent a vascular pattern, although we didnot attempt to identify the types of labeled cells.
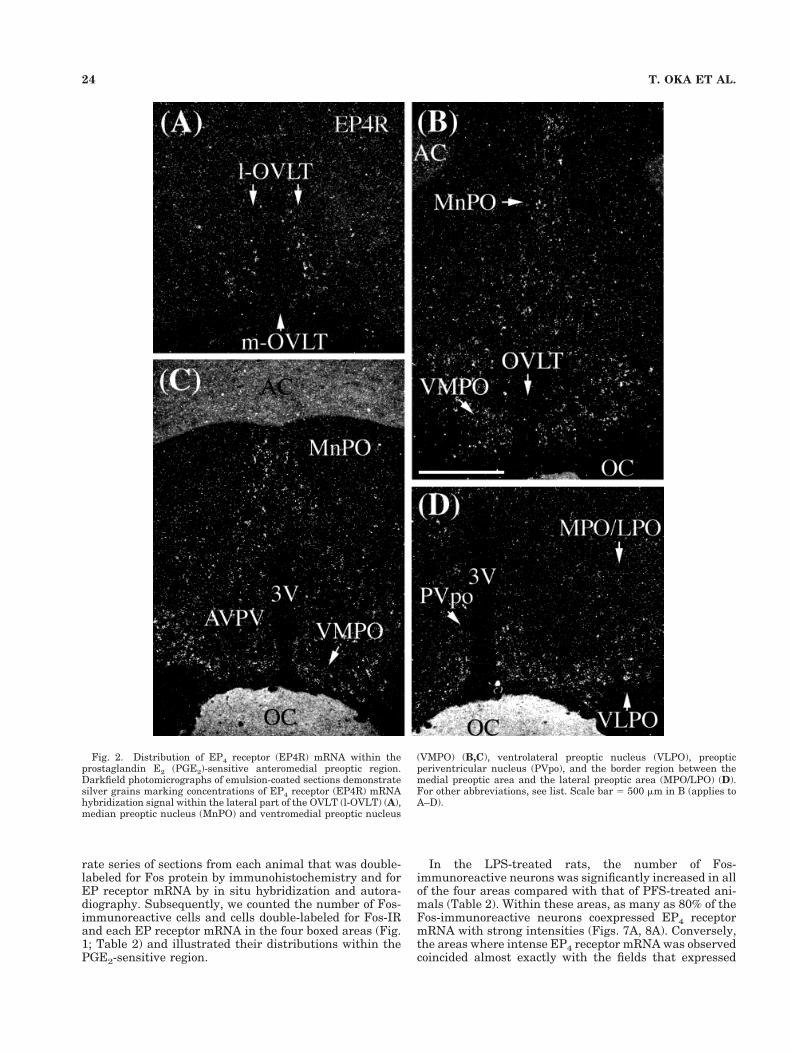
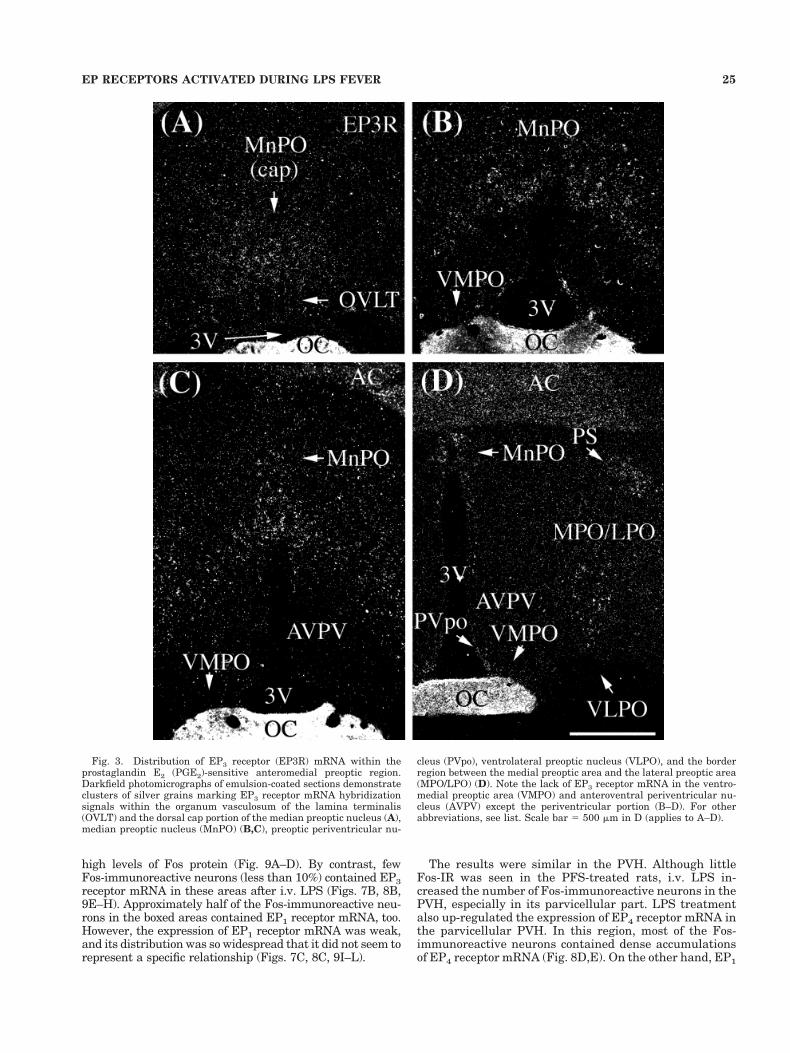
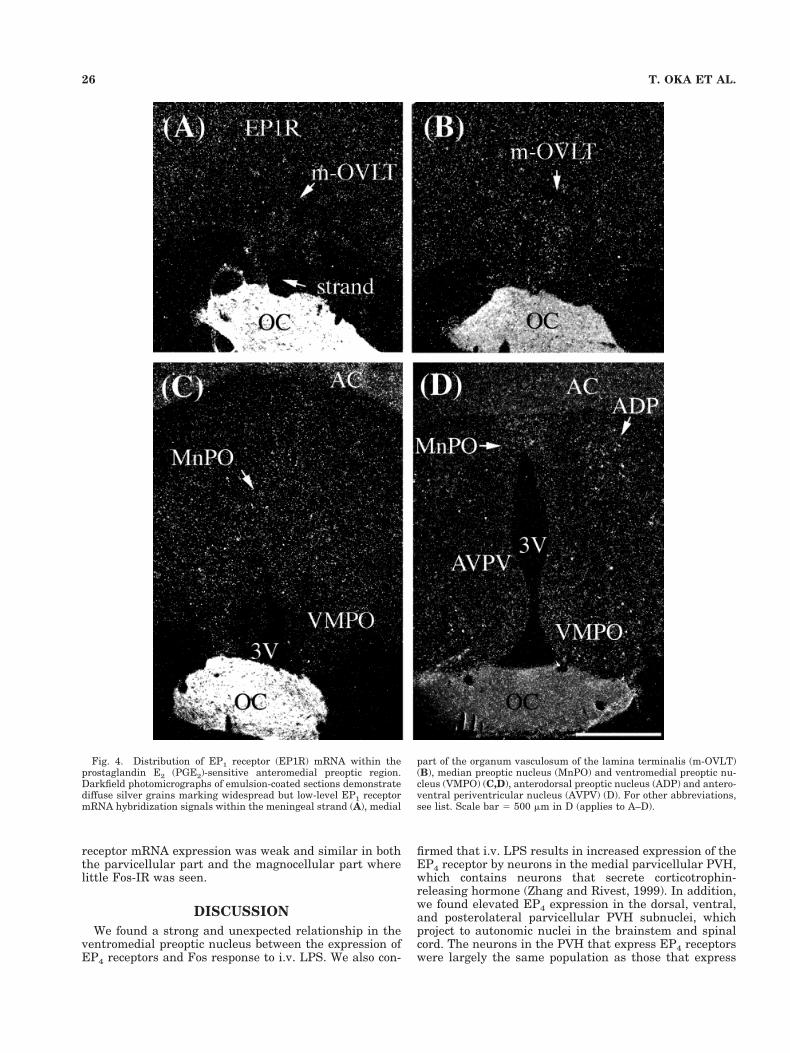
In the OVLT, EP4 receptor mRNA was strongly ex-pressed in the lateral portion of the nucleus (Fig. 2A). Incontrast, EP3 receptor mRNA signal was strong within theportion of the MnPO forming a dorsal cap over the OVLTbut weak within the OVLT itself (Fig. 3A). EP1 receptormRNA was moderately expressed within the meningealstrand just rostral to the OVLT (Fig. 4A) and the medialportion of the OVLT (Fig. 4B).
EP4 receptor mRNA was also strongly expressed in theMnPO (Fig. 2B), VMPO (Fig. 2C), ventrolateral preoptic
nucleus, and preoptic periventricular nucleus (Fig. 2D).Its expression was moderate in the border region betweenthe medial preoptic area and the lateral preoptic area (Fig.2D), weak in the anteroventral periventricular nucleus(Fig. 2C), and absent in the lateral preoptic area. EP3receptor mRNA was expressed strongly within the MnPO(Fig. 3B,C) and moderately within the ventrolateral pre-optic nucleus, preoptic periventricular nucleus, and bor-der region between the medial preoptic area and the lat-eral preoptic area (Fig. 3D). EP3 receptor mRNA was notfound in the VMPO or anteroventral periventricular nu-cleus except in their periventricular portions (Fig. 3B–D).EP1 receptor mRNA was diffusely distributed throughoutthe preoptic area. Among these regions, it was moderatelyexpressed in the MnPO, VMPO (Fig. 4C,D), anteroventralperiventricular nucleus, anterodorsal preoptic nucleus,and the preoptic periventricular nucleus (Fig. 4D).
Distribution and density of each EP receptor within thePGE2-sensitive anteromedial preoptic region was similarin the untreated, PFS-treated, and LPS-treated rats. Hy-bridization with sense strand cRNA probes or hybridiza-tion with antisense probe after RNase treatment did notresult in any specific signal.
EP receptor distribution within the PVH
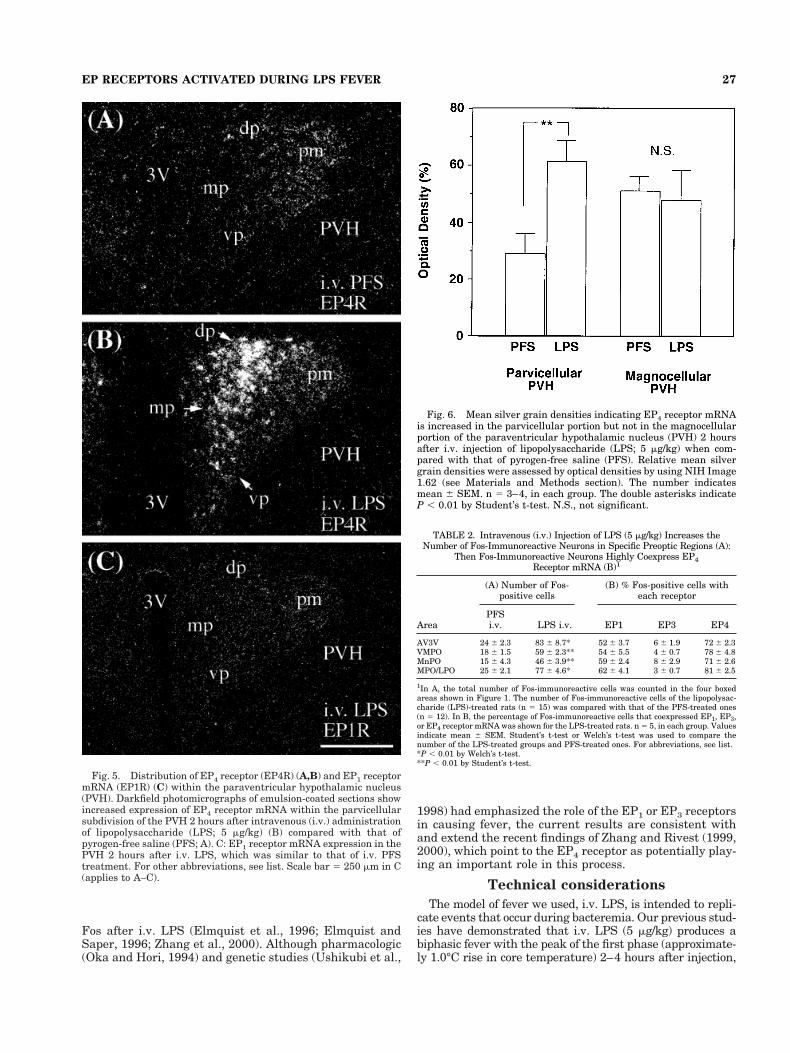
EP4 and EP1 receptor mRNAs were observed withinboth the parvicellular and magnocellular portions of thePVH (Fig. 5). Neither EP3 nor EP2 receptor mRNA wasfound in the PVH. EP4 receptor mRNA was more stronglyexpressed in the magnocellular portion in the untreatedand PFS-treated rats (Fig. 5A). However, in the LPS-treated rats, EP4 receptor mRNA expression was greaterover the parvicellular subnuclei, including the dorsal, ven-tral, and posterolateral subnuclei, which project to theautonomic targets, as well as the medial subnucleus,which contains corticotrophin-releasing hormone neurons(Figs. 5B, 6). EP1 receptor mRNA expression was weak(Fig. 5C) and unaffected by LPS treatment.
Colocalization of Fos-IR with EP receptormRNA in the PGE2-sensitive anteromedial
preoptic region and the PVH
To investigate which EP receptor-containing neuronsare activated during i.v. LPS fever, we prepared a sepa-
TABLE 1. Distribution of EP Receptor mRNAs in the Preoptic Area1
Brain regionEP1R
mRNAEP3R
mRNAEP4RmRNA
Meningeal strand 1/11 2 2Organum vasculosum of the lamina terminalis 11 1 111Median preoptic nucleus 11 111 1111Ventromedial preoptic nucleus 11 2/1 1111Anterodorsal preoptic nucleus 11 2 2Anteroventral periventricular nucleus 1/11 2 1Preoptic periventricular nucleus 11 11 111Ventrolateral preoptic nucleus 1 1/11 111Medial preoptic nucleus 1 2/1 1Border region between the medial preoptic area
and the lateral preoptic area1 11 11/111
Lateral preoptic area 1 2 2Ependymal layer of the third ventricle 2 11 2
1Qualitative analysis of hybridization signal for EP1, EP3, and EP4 receptor (EP1R,EP3R, and EP4R, respectively) mRNAs in the preoptic area of nontreated rats. In situhybridization histochemistry was accomplished by using specific 35S-labeled antisenseriboprobes complementary to the rat EP1, EP3, or EP4 receptors mRNA. Relative graindensities: 1111, very strong; 111, strong; 11, moderate; 1, weak but positive; 2,background level.
23EP RECEPTORS ACTIVATED DURING LPS FEVER
rate series of sections from each animal that was double-labeled for Fos protein by immunohistochemistry and forEP receptor mRNA by in situ hybridization and autora-diography. Subsequently, we counted the number of Fos-immunoreactive cells and cells double-labeled for Fos-IRand each EP receptor mRNA in the four boxed areas (Fig.1; Table 2) and illustrated their distributions within thePGE2-sensitive region.
In the LPS-treated rats, the number of Fos-immunoreactive neurons was significantly increased in allof the four areas compared with that of PFS-treated ani-mals (Table 2). Within these areas, as many as 80% of theFos-immunoreactive neurons coexpressed EP4 receptormRNA with strong intensities (Figs. 7A, 8A). Conversely,the areas where intense EP4 receptor mRNA was observedcoincided almost exactly with the fields that expressed
Fig. 2. Distribution of EP4 receptor (EP4R) mRNA within theprostaglandin E2 (PGE2)-sensitive anteromedial preoptic region.Darkfield photomicrographs of emulsion-coated sections demonstratesilver grains marking concentrations of EP4 receptor (EP4R) mRNAhybridization signal within the lateral part of the OVLT (l-OVLT) (A),median preoptic nucleus (MnPO) and ventromedial preoptic nucleus
(VMPO) (B,C), ventrolateral preoptic nucleus (VLPO), preopticperiventricular nucleus (PVpo), and the border region between themedial preoptic area and the lateral preoptic area (MPO/LPO) (D).For other abbreviations, see list. Scale bar 5 500 mm in B (applies toA–D).
24 T. OKA ET AL.
high levels of Fos protein (Fig. 9A–D). By contrast, fewFos-immunoreactive neurons (less than 10%) contained EP3receptor mRNA in these areas after i.v. LPS (Figs. 7B, 8B,9E–H). Approximately half of the Fos-immunoreactive neu-rons in the boxed areas contained EP1 receptor mRNA, too.However, the expression of EP1 receptor mRNA was weak,and its distribution was so widespread that it did not seem torepresent a specific relationship (Figs. 7C, 8C, 9I–L).
The results were similar in the PVH. Although littleFos-IR was seen in the PFS-treated rats, i.v. LPS in-creased the number of Fos-immunoreactive neurons in thePVH, especially in its parvicellular part. LPS treatmentalso up-regulated the expression of EP4 receptor mRNA inthe parvicellular PVH. In this region, most of the Fos-immunoreactive neurons contained dense accumulationsof EP4 receptor mRNA (Fig. 8D,E). On the other hand, EP1
Fig. 3. Distribution of EP3 receptor (EP3R) mRNA within theprostaglandin E2 (PGE2)-sensitive anteromedial preoptic region.Darkfield photomicrographs of emulsion-coated sections demonstrateclusters of silver grains marking EP3 receptor mRNA hybridizationsignals within the organum vasculosum of the lamina terminalis(OVLT) and the dorsal cap portion of the median preoptic nucleus (A),median preoptic nucleus (MnPO) (B,C), preoptic periventricular nu-
cleus (PVpo), ventrolateral preoptic nucleus (VLPO), and the borderregion between the medial preoptic area and the lateral preoptic area(MPO/LPO) (D). Note the lack of EP3 receptor mRNA in the ventro-medial preoptic area (VMPO) and anteroventral periventricular nu-cleus (AVPV) except the periventricular portion (B–D). For otherabbreviations, see list. Scale bar 5 500 mm in D (applies to A–D).
25EP RECEPTORS ACTIVATED DURING LPS FEVER
receptor mRNA expression was weak and similar in boththe parvicellular part and the magnocellular part wherelittle Fos-IR was seen.
DISCUSSION
We found a strong and unexpected relationship in theventromedial preoptic nucleus between the expression ofEP4 receptors and Fos response to i.v. LPS. We also con-
firmed that i.v. LPS results in increased expression of theEP4 receptor by neurons in the medial parvicellular PVH,which contains neurons that secrete corticotrophin-releasing hormone (Zhang and Rivest, 1999). In addition,we found elevated EP4 expression in the dorsal, ventral,and posterolateral parvicellular PVH subnuclei, whichproject to autonomic nuclei in the brainstem and spinalcord. The neurons in the PVH that express EP4 receptorswere largely the same population as those that express
Fig. 4. Distribution of EP1 receptor (EP1R) mRNA within theprostaglandin E2 (PGE2)-sensitive anteromedial preoptic region.Darkfield photomicrographs of emulsion-coated sections demonstratediffuse silver grains marking widespread but low-level EP1 receptormRNA hybridization signals within the meningeal strand (A), medial
part of the organum vasculosum of the lamina terminalis (m-OVLT)(B), median preoptic nucleus (MnPO) and ventromedial preoptic nu-cleus (VMPO) (C,D), anterodorsal preoptic nucleus (ADP) and antero-ventral periventricular nucleus (AVPV) (D). For other abbreviations,see list. Scale bar 5 500 mm in D (applies to A–D).
26 T. OKA ET AL.
Fos after i.v. LPS (Elmquist et al., 1996; Elmquist andSaper, 1996; Zhang et al., 2000). Although pharmacologic(Oka and Hori, 1994) and genetic studies (Ushikubi et al.,
1998) had emphasized the role of the EP1 or EP3 receptorsin causing fever, the current results are consistent withand extend the recent findings of Zhang and Rivest (1999,2000), which point to the EP4 receptor as potentially play-ing an important role in this process.
Technical considerations
The model of fever we used, i.v. LPS, is intended to repli-cate events that occur during bacteremia. Our previous stud-ies have demonstrated that i.v. LPS (5 mg/kg) produces abiphasic fever with the peak of the first phase (approximate-ly 1.0°C rise in core temperature) 2–4 hours after injection,
Fig. 5. Distribution of EP4 receptor (EP4R) (A,B) and EP1 receptormRNA (EP1R) (C) within the paraventricular hypothalamic nucleus(PVH). Darkfield photomicrographs of emulsion-coated sections showincreased expression of EP4 receptor mRNA within the parvicellularsubdivision of the PVH 2 hours after intravenous (i.v.) administrationof lipopolysaccharide (LPS; 5 mg/kg) (B) compared with that ofpyrogen-free saline (PFS; A). C: EP1 receptor mRNA expression in thePVH 2 hours after i.v. LPS, which was similar to that of i.v. PFStreatment. For other abbreviations, see list. Scale bar 5 250 mm in C(applies to A–C).
Fig. 6. Mean silver grain densities indicating EP4 receptor mRNAis increased in the parvicellular portion but not in the magnocellularportion of the paraventricular hypothalamic nucleus (PVH) 2 hoursafter i.v. injection of lipopolysaccharide (LPS; 5 mg/kg) when com-pared with that of pyrogen-free saline (PFS). Relative mean silvergrain densities were assessed by optical densities by using NIH Image1.62 (see Materials and Methods section). The number indicatesmean 6 SEM. n 5 3–4, in each group. The double asterisks indicateP , 0.01 by Student’s t-test. N.S., not significant.
TABLE 2. Intravenous (i.v.) Injection of LPS (5 mg/kg) Increases theNumber of Fos-Immunoreactive Neurons in Specific Preoptic Regions (A):
Then Fos-Immunoreactive Neurons Highly Coexpress EP4Receptor mRNA (B)1
Area
(A) Number of Fos-positive cells
(B) % Fos-positive cells witheach receptor
PFSi.v. LPS i.v. EP1 EP3 EP4
AV3V 24 6 2.3 83 6 8.7* 52 6 3.7 6 6 1.9 72 6 2.3VMPO 18 6 1.5 59 6 2.3** 54 6 5.5 4 6 0.7 78 6 4.8MnPO 15 6 4.3 46 6 3.9** 59 6 2.4 8 6 2.9 71 6 2.6MPO/LPO 25 6 2.1 77 6 4.6* 62 6 4.1 3 6 0.7 81 6 2.5
1In A, the total number of Fos-immunoreactive cells was counted in the four boxedareas shown in Figure 1. The number of Fos-immunoreactive cells of the lipopolysac-charide (LPS)-treated rats (n 5 15) was compared with that of the PFS-treated ones(n 5 12). In B, the percentage of Fos-immunoreactive cells that coexpressed EP1, EP3,or EP4 receptor mRNA was shown for the LPS-treated rats. n 5 5, in each group. Valuesindicate mean 6 SEM. Student’s t-test or Welch’s t-test was used to compare thenumber of the LPS-treated groups and PFS-treated ones. For abbreviations, see list.*P , 0.01 by Welch’s t-test.**P , 0.01 by Student’s t-test.
27EP RECEPTORS ACTIVATED DURING LPS FEVER
followed by a long-lasting second phase (which starts 4 hoursafter injection and peaks approximately 6 hours after injec-tion, approximately 1.5°C rise) (Elmquist et al., 1996; Scam-mell et al., 1998). Therefore, in the present study, we chosethe 2-hour time point, at which there is a reliable fever ofapproximately 1°C to investigate the coexpression of Fos-IRand EP receptor mRNA. We did not investigate the timecourse of changes in EP receptor mRNA expression, al-though this would clearly be of interest and will be thesubject of later studies.
This model is critically dependent on the effects ofPGs in the anteromedial preoptic area, especially theVMPO, MnPO, and OVLT (Scammell et al., 1998). Thisbrain region binds PGE2 in both rats (Matsumura et al.,1990, 1992) and monkeys (Watanabe et al., 1988). How-ever, the effects of different neurons in this region thatbind PGE2 on different physiologic responses has not
been explored previously. Hence, whereas we discussour findings in terms of the physiology of fever re-sponses, the changes we have observed may underlieother aspects of the central nervous system response toimmune stimuli as well.
It is unlikely that the EP receptor probes we used lackedsufficient selectivity. The cRNAs were raised against uniqueparts of the EP receptor mRNAs, and each produced a spe-cific pattern of staining in the brain. Our pattern of localiza-tion for the EP1, EP3, and EP4 receptors is consistent withprevious reports (Sugimoto et al., 1994; Rage et al., 1997;Nakamura et al., 1999; Zhang and Rivest, 1999, 2000). How-ever, we provide more detailed mapping of these receptorsubtypes in the anteromedial preoptic area, and the relation-ship of that labeling with LPS-induced Fos. Although wefound EP2 receptor mRNA in the bed nucleus of the striaterminalis as had been reported, we could not identify obvi-
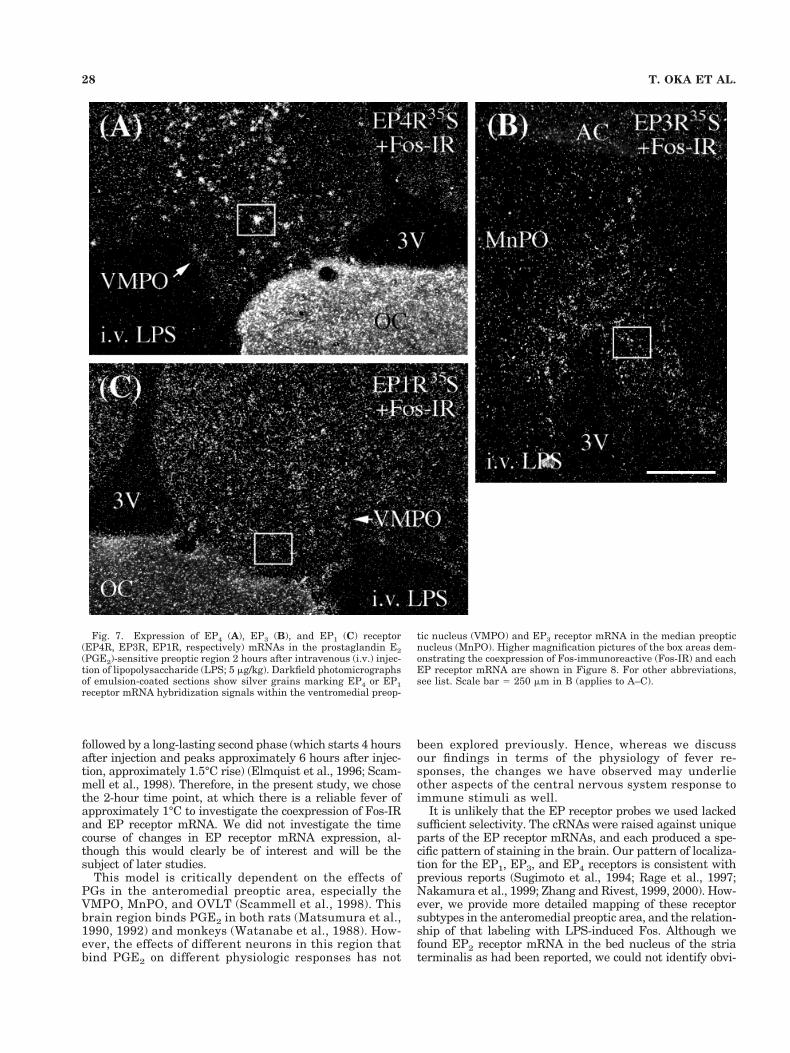
Fig. 7. Expression of EP4 (A), EP3 (B), and EP1 (C) receptor(EP4R, EP3R, EP1R, respectively) mRNAs in the prostaglandin E2(PGE2)-sensitive preoptic region 2 hours after intravenous (i.v.) injec-tion of lipopolysaccharide (LPS; 5 mg/kg). Darkfield photomicrographsof emulsion-coated sections show silver grains marking EP4 or EP1receptor mRNA hybridization signals within the ventromedial preop-
tic nucleus (VMPO) and EP3 receptor mRNA in the median preopticnucleus (MnPO). Higher magnification pictures of the box areas dem-onstrating the coexpression of Fos-immunoreactive (Fos-IR) and eachEP receptor mRNA are shown in Figure 8. For other abbreviations,see list. Scale bar 5 250 mm in B (applies to A–C).
28 T. OKA ET AL.
ous expression within the PGE2-sensitive regions, whereweak EP2 expression had been reported (Zhang and Rivest,1999). The present results for the EP1 receptor also differfrom those of Batshake, who reported hybridization only inthe PVH and supraoptic nucleus in mice (Batshake et al.,1995). However, they did not undertake a detailed neuro-anatomic survey in mice, and there may be species differ-ences in expression patterns.
It is always possible that our probes were not signifi-cantly sensitive to pick up all of the EP receptor-positiveneurons. However, on both our X-ray film autoradiographsand emulsion coated slides, the patterns were similar. Wefound the same pattern in rats that were untreated, and inones that received i.v. PFS or LPS 2 hours before death.Hence, the pattern of EP receptor hybridization does notseem to change with physiologic manipulation, althoughEP4 receptor mRNA was up-regulated in the parvicellularportion of the PVH by i.v. LPS.
Distinctive distribution and activationpattern of EP receptors within the PGE2-
sensitive anteromedial preoptic region
Our study identified distinctive distributions for theEP1, EP3, and EP4 receptor mRNAs within the PGE2-sensitive anteromedial preoptic region. Among these, onlythe EP4 receptor mRNA was strongly expressed through-out the PGE2-sensitive region, including the OVLT, theVMPO, and the MnPO. Most Fos-immunoreactive neu-rons in this region after i.v. LPS also contained EP4 re-ceptor mRNA. In contrast, little EP3 receptor mRNAwas found within the OVLT and VMPO. Furthermore,despite strong EP3 receptor mRNA expression withinthe MnPO, few Fos-immunoreactive neurons containedEP3 receptor mRNA. EP1 receptor mRNA was foundthroughout the PGE2-sensitive region. However, its ex-pression was weak, and, although approximately half ofthe Fos-immunoreactive cells contained EP1 receptor
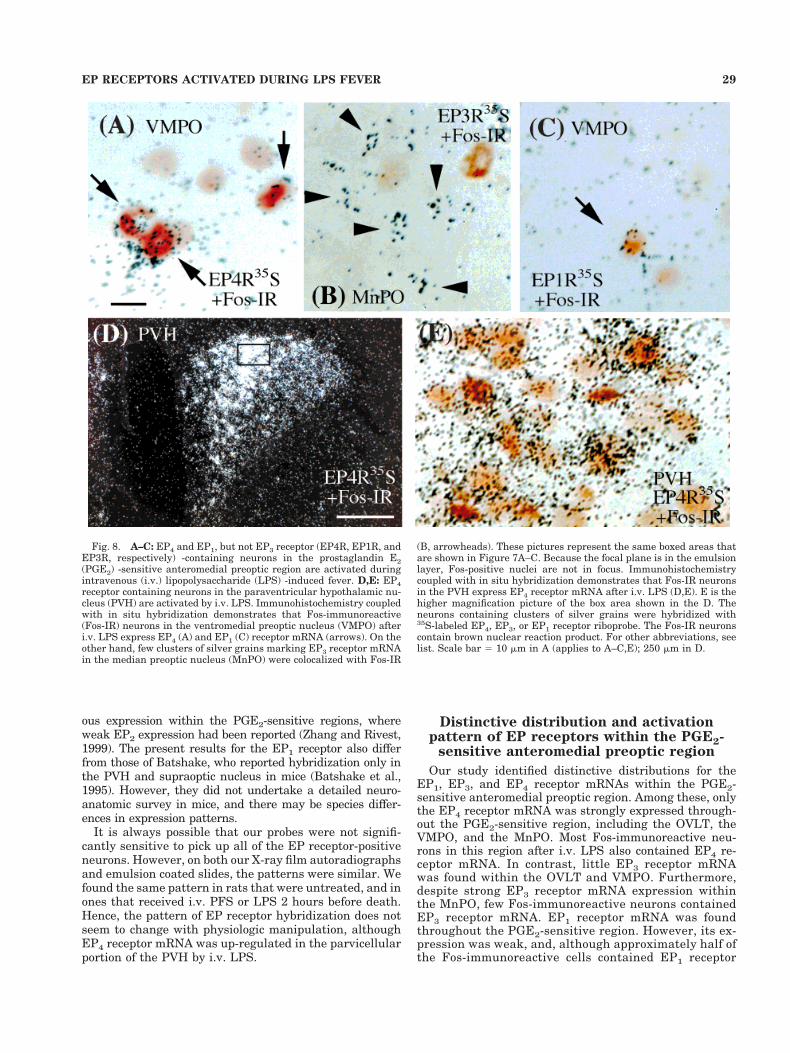
Fig. 8. A–C: EP4 and EP1, but not EP3 receptor (EP4R, EP1R, andEP3R, respectively) -containing neurons in the prostaglandin E2(PGE2) -sensitive anteromedial preoptic region are activated duringintravenous (i.v.) lipopolysaccharide (LPS) -induced fever. D,E: EP4receptor containing neurons in the paraventricular hypothalamic nu-cleus (PVH) are activated by i.v. LPS. Immunohistochemistry coupledwith in situ hybridization demonstrates that Fos-immunoreactive(Fos-IR) neurons in the ventromedial preoptic nucleus (VMPO) afteri.v. LPS express EP4 (A) and EP1 (C) receptor mRNA (arrows). On theother hand, few clusters of silver grains marking EP3 receptor mRNAin the median preoptic nucleus (MnPO) were colocalized with Fos-IR
(B, arrowheads). These pictures represent the same boxed areas thatare shown in Figure 7A–C. Because the focal plane is in the emulsionlayer, Fos-positive nuclei are not in focus. Immunohistochemistrycoupled with in situ hybridization demonstrates that Fos-IR neuronsin the PVH express EP4 receptor mRNA after i.v. LPS (D,E). E is thehigher magnification picture of the box area shown in the D. Theneurons containing clusters of silver grains were hybridized with35S-labeled EP4, EP3, or EP1 receptor riboprobe. The Fos-IR neuronscontain brown nuclear reaction product. For other abbreviations, seelist. Scale bar 5 10 mm in A (applies to A–C,E); 250 mm in D.
29EP RECEPTORS ACTIVATED DURING LPS FEVER

mRNA, this did not seem to represent a special rela-tionship.
This association with Fos expression might reflect thatstimulation of EP4 receptors increases cAMP, which is aneffective stimulus for Fos expression, whereas the EP1receptor increases intracellular calcium and EP3 receptor
decreases or increases cAMP depending on its isoform(Ushikubi et al., 1995). As the c-fos gene is activated byeither increased intracellular calcium or cAMP (Shengand Greenberg, 1990), it is understandable that activationof the EP3a receptor isoform, which causes reduced intra-cellular cAMP (Boie et al., 1997), might not produce neu-
Fig. 9. Distribution of activated EP receptor containing neuronswithin the prostaglandin E2 (PGE2)-sensitive anteromedial preopticregion 2 hours after intravenous (i.v.) administration of lipopolysac-charide (LPS). Coronal sections of the rat brain are illustrated.Squares represent Fos-immunoreactive (Fos-IR) neurons expressingEP4, EP3, or EP1 receptor mRNA. Small dots represent Fos-immunoreactive neurons. Gray areas represent the regions where EP
receptor mRNA is predominantly expressed with darker areas repre-senting a greater density of silver grains. A–D: Fos-like IR and itscoexpression with EP4 receptor mRNA (EP4R1Fos-IR). E–H: Fos-like-IR and its coexpression with EP3 receptor mRNA (EP3R1Fos-IR). I–L: Fos-like-IR and its coexpression with EP1 receptor mRNA(EP1R1Fos-IR). Scale bar 5 500 mm in B (applies to A–L).
30 T. OKA ET AL.

ronal Fos expression, even though it may mediate cellularresponses. The EP3 receptor probe that we used does notdiscriminate between isoforms. Thus, Fos immunohisto-chemistry might fail to implicate a role of some EP3 re-ceptor isoforms in the fever response if these receptorsdecreased intracellular cAMP. Nonetheless, the absence ofEP3 receptor mRNA in the VMPO, one of the most PGE2-sensitive regions, raises the question whether the EP3receptor can be solely responsible for fever in rats. Rather,the distinctive distribution of each EP receptor mRNA inthis region, i.e., EP1 receptor mRNA in the medial OVLT,EP4 receptor mRNA in the lateral OVLT and VMPO, andthe EP3 receptor mRNA in the MnPO, might suggest aninvolvement of each EP receptor in different componentsor phases of the fever response.
The present study demonstrated that EP3 and EP4 re-ceptor mRNA expression is also evident in the borderregion between the medial preoptic area and the lateralpreoptic area, extending down to the ventrolateral preop-tic nucleus. The ventrolateral preoptic nucleus has beenimplicated in the regulation of sleep (Sherin et al., 1996).Thus, EP receptors in the border region between the me-dial preoptic area and the lateral preoptic area may beinvolved in LPS-induced changes in vigilance as well.PGE2-sensitive sleep-related neurons have been demon-strated in this area (Koyama and Hayaishi, 1994) and thenumber of Fos-immunoreactive neurons in the ventrolat-eral preoptic nucleus is increased during sleep (Sherin etal., 1996).
From the present study, we cannot specify the subcel-lular sites expressing EP receptors. Although the EP3receptor is thought to cause presynaptic inhibition in thecortex of mice (Exner and Schlicker, 1995), the site of EPreceptors involved in fever response may be postsynapticas PGE2-induced firing rate changes of the thermosensi-tive neurons have been observed even in Ca21 free me-dium, which blocks synaptic activity (Matsuda et al.,1992).
Role of the PVH in fever
As previously described by Zhang and Rivest (1999),EP4 receptor mRNA was also up-regulated in the parvi-cellular PVH after i.v. LPS. Although they emphasizedup-regulation of EP4 receptors on corticotrophin-releasinghormone neurons in the medial parvicellular PVH, wefound EP4 expression up-regulated in the dorsal, ventral,and posterolateral parvicellular subnuclei as well. TheEP4 receptor expression was colocalized with Fos expres-sion in each of these cell groups. We previously showedthat Fos-positive neurons in the dorsal parvicellular PVHafter i.v. LPS project to the sympathetic preganglioniccolumn in the spinal cord (Zhang et al., 2000). Hence, theEP4 activation in the PVH most likely includes the auto-nomic as well as endocrine components of the nucleus.
The PVH is approximately 2 mm away from the mostsensitive preoptic sites at which PGE2 injection causesfever. Hence, finding EP receptors in the PVH, expressedspecifically by neurons that are activated during the feverresponse, is remarkable. One interpretation is that theseneurons can be activated by several routes, including dif-fusion of PGE2 from the POA. Alternatively, PGE2 may bemade by perivascular microglial cells or by endothelialcells lining small blood vessels in the PVH region duringfever (Van Dam et al., 1993; Cao et al., 1995; Elmquist etal., 1997; Lacroix and Rivest, 1998), or by PVH neurons,
which themselves express COX (Breder et al., 1995). Al-though neuronal expression of COX is not increased dur-ing fever, there is baseline expression by neurons both inthe VMPO and PVH (Breder et al., 1995). Hence, thepresence of EP4 receptors at these sites may indicate thepresence of another, previously unrecognized, paracrinepathway for activating the neurons that participate in thefever response.
The parvicellular PVH as well as the anteromedial pre-optic region may be necessary for producing fever. Lesionsof the parvicellular PVH block fever response to i.p. LPS(Horn et al., 1994; Caldeira et al., 1998) or i.v. LPS (Lu etal., submitted for publication) and neurons of the dorsalparvicellular PVH that project to the spinal cord (Zhang etal., 2000) are activated by i.v. LPS. The microinjection ofPGE2 into the PVH is reported to increase core tempera-ture in rats (Bhatnagar et al., 1993), although its effect isless remarkable than that of PGE2 injection into the an-teromedial preoptic region. On the other hand, the micro-injection of 17-phenyl-v-trinor-PGE2 (an EP1 and EP3a
receptor agonist) into the PVH did not increase core tem-perature (Oka et al., 1997). We could not find EP2 or EP3receptor mRNA in the parvicellular PVH in the presentstudy. Therefore, taking the above findings into account, itmight be possible that i.v. LPS produces fever, at least inpart, by the direct action of PGE2 on the EP4 receptorcontaining neurons in the parvicellular PVH.
In conclusion, the present findings indicate that neu-rons expressing EP4 receptor are activated during LPS-induced fever and suggest the involvement of EP4 recep-tors in the production of fever, at least beyond the firsthour after i.v. LPS injection (Ushikubi et al., 1998). It willbe important to test animals with EP4 gene deletions forthe later stages of fever disruption. However, the develop-ment of site-selective EP4 receptor genetic deletions, ordrugs that are specific for the EP4 receptor, may be nec-essary to test the importance of hypothalamic EP4 recep-tors specifically.
LITERATURE CITED
Batshake B, Nilsson C, Sundelin J. 1995. Molecular characterization of themouse prostanoid EP1 receptor gene. Eur J Biochem 231:809–814.
Bhatnagar S, Meaney MJ, Amir S. 1993. The effects of prostaglandin E2injected into the paraventricular nucleus of the hypothalamus onbrown adipose tissue thermogenesis in spontaneously hypertensiverats. Brain Res 613:285–287.
Blatteis CM, Sehic E. 1997. Prostaglandin E2: A putative fever mediator.In: Mackowiak PA, editor. Fever basic mechanisms and management.Philadelphia: Lippincott–Raven. p 117–145.
Boie Y, Stocco R, Sawyer N, Slipetz DM, Ungrin MD, Neuschafer-Rube F,Puschel GP, Metters KM, Abramovitz M. 1997. Molecular cloning andcharacterization of the four rat prostaglandin E2 prostanoid receptorsubtypes. Eur J Pharmacol 340:227–241.
Breder CD, Dewitt D, Kraig RP. 1995. Characterization of inducible cyclo-oxygenase in rat brain. J Comp Neurol 355:296–315.
Caldeira JC, Franci CR, Pela IR. 1998. Bilateral lesion of hypothalamicparaventricular nucleus abolishes fever induced by endotoxin and bra-dykinin in rats. Ann N Y Acad Sci 856:294–297.
Cao C, Matsumura K, Yamagata K, Watanabe Y. 1995. Induction bylipopolysaccharide of cyclooxygenase-2 mRNA in rat brain; its possiblerole in the febrile response. Brain Res 697:187–196.
Coleman RA, Smith WL, Narumiya S. 1994. International Union of Phar-macology classification of prostanoid receptors: properties, distribu-tion, and structure of the receptors and their subtypes. Pharmacol Rev46:205–229.
Cranston WI, Rawlins MD. 1972. Effects of intracerebral micro-injection ofsodium salicylate on temperature regulation in the rabbit. J Physiol(Lond) 222:257–266.
31EP RECEPTORS ACTIVATED DURING LPS FEVER

Elias CF, Aschkenasi C, Lee C, Kelly J, Ahima RS, Bjorbaek C, Flier JS,Saper CB, Elmquist JK. 1999. Leptin differentially regulates NPY andPOMC neurons projecting to the lateral hypothalamic area. Neuron23:775–786.
Elias CF, Saper CB, Maratos-Flier E, Tritos NA, Lee C, Kelly J, Tatro JB,Hoffman GE, Ollmann MM, Barsh GS, Sakurai T, Yanagisawa M,Elmquist JK. 1998. Chemically defined projections linking the medio-basal hypothalamus and the lateral hypothalamic area. J Comp Neurol402:442–459.
Elmquist JK, Saper CB. 1996. Activation of neurons projecting to theparaventricular hypothalamic nucleus by intravenous lipopolysaccha-ride. J Comp Neurol 374:315–331.
Elmquist JK, Scammell TE, Jacobson CD, Saper CB. 1996. Distribution ofFos-like immunoreactivity in the rat brain following intravenous lipo-polysaccharide administration. J Comp Neurol 371:85–103.
Elmquist JK, Breder CD, Sherin JE, Scammell TE, Hickey WF, Dewitt D,Saper CB. 1997. Intravenous lipopolysaccharide induces cyclooxygen-ase 2-like immunoreactivity in rat brain perivascular microglia andmeningeal macrophages. J Comp Neurol 381:119–129.
Exner HJ, Schlicker E. 1995. Prostanoid receptors of the EP3 subtypemediate the inhibitory effect of prostaglandin E2 on noradrenalinerelease in the mouse brain cortex. Naunyn Schmiedebergs Arch Phar-macol 351:46–52.
Feldberg W, Saxena PN. 1971. Further studies on prostaglandin E 1 feverin cats. J Physiol (Lond) 219:739–745.
Horn T, Wilkinson MF, Landgraf R, Pittman QJ. 1994. Reduced febrileresponses to pyrogens after lesions of the hypothalamic paraventricu-lar nucleus. Am J Physiol 267:R323–R328.
Kluger MJ, Kozak W, Conn CA, Leon LR, Soszynski D. 1996. The adaptivevalue of fever. Infect Dis Clin North Am 10:1–20.
Kononen J, Koistinaho J, Alho H. 1990. Circadian rhythm in c-fos-likeimmunoreactivity in the rat brain. Neurosci Lett 120:105–108.
Koyama Y, Hayaishi O. 1994. Modulation by prostaglandins of activity ofsleep-related neurons in the preoptic/anterior hypothalamic areas inrats. Brain Res Bull 33:367–372.
Lacroix S, Rivest S. 1998. Effect of acute systemic inflammatory responseand cytokines on the transcription of the genes encoding cyclooxygen-ase enzymes (COX-1 and COX-2) in the rat brain. J Neurochem 70:452–466.
Matsuda T, Hori T, Nakashima T. 1992. Thermal and PGE2 sensitivity ofthe organum vasculosum lamina terminalis region and preoptic area inrat brain slices. J Physiol (Lond) 454:197–212.
Matsumura K, Watanabe Y, Onoe H, Watanabe Y, Hayaishi O. 1990. Highdensity of prostaglandin E2 binding sites in the anterior wall of the 3rdventricle: a possible site of its hyperthermic action. Brain Res 533:147–151.
Matsumura K, Watanabe Y, Imai-Matsumura K, Connolly M, Koyama Y,Onoe H, Watanabe Y. 1992. Mapping of prostaglandin E2 binding sitesin rat brain using quantitative autoradiography. Brain Res 581:292–298.
Morimoto A, Murakami N, Nakamori T, Watanabe T. 1988. Multiplecontrol of fever production in the central nervous system of rabbits.J Physiol (Lond) 397:269–280.
Nakamura K, Kaneko T, Yamashita Y, Hasegawa H, Katoh H, Ichikawa A,Negishi M. 1999. Immunocytochemical localization of prostaglandinEP3 receptor in the rat hypothalamus. Neurosci Lett 260:117–120.
Oka K, Oka T, Hori T. 1997. Prostaglandin E2 may induce hyperthermiathrough EP1 receptor in the anterior wall of the third ventricle andneighboring preoptic regions. Brain Res 767:92–99.
Oka K, Oka T, Hori T. 1998. PGE2 receptor subtype EP1 antagonist mayinhibit central interleukin-1beta-induced fever in rats. Am J Physiol275:R1762–R1765.
Oka T, Hori T. 1994. EP1-receptor mediation of prostaglandin E2-inducedhyperthermia in rats. Am J Physiol 267:R289–R294.
Parrott RF, Vellucci SV. 1996. Effects of centrally administered prosta-glandin EP receptor agonists on febrile and adrenocortical responses inthe prepubertal pig. Brain Res Bull 41:97–103.
Rage F, Lee BJ, Ma YJ, Ojeda SR. 1997. Estradiol enhances prostaglandinE2 receptor gene expression in luteinizing hormone-releasing hormone(LHRH) neurons and facilitates the LHRH response to PGE2 by acti-vating a glia-to-neuron signaling pathway. J Neurosci 17:9145–9156.
Sagar SM, Price KJ, Kasting NW, Sharp FR. 1995. Anatomic patterns ofFos immunostaining in rat brain following systemic endotoxin admin-istration. Brain Res Bull 36:381–392.
Scammell TE, Elmquist JK, Griffin JD, Saper CB. 1996. Ventromedialpreoptic prostaglandin E2 activates fever-producing autonomic path-ways. J Neurosci 16:6246–6254.
Scammell TE, Griffin JD, Elmquist JK, Saper CB. 1998. Microinjection ofa cyclooxygenase inhibitor into the anteroventral preoptic region at-tenuates LPS fever. Am J Physiol 274:R783–789.
Sehic E, Szekely M, Ungar AL, Oladehin A, Blatteis CM. 1996. Hypotha-lamic prostaglandin E2 during lipopolysaccharide-induced fever inguinea pigs. Brain Res Bull 39:391–399.
Sheng M, Greenberg ME. 1990. The regulation and function of c-fos andother immediate early genes in the nervous system. Neuron 4:477–485.
Sherin JE, Shiromani PJ, McCarley RW, Saper CB. 1996. Activation ofventrolateral preoptic neurons during sleep. Science 271:216–219.
Sirko S, Bishai I, Coceani F. 1989. Prostaglandin formation in the hypo-thalamus in vivo: effect of pyrogens. Am J Physiol 256:R616–R624.
Stitt JT. 1991. Differential sensitivity in the sites of fever production byprostaglandin E1 within the hypothalamus of the rat. J Physiol (Lond)432:99–110.
Sugimoto Y, Shigemoto R, Namba T, Negishi M, Mizuno N, Narumiya S,Ichikawa A. 1994. Distribution of the messenger RNA for the prosta-glandin E receptor subtype EP3 in the mouse nervous system. Neuro-science 62:919–928.
Ushikubi F, Hirata M, Narumiya S. 1995. Molecular biology of prostanoidreceptors; an overview. J Lipid Mediat Cell Signal 12:343–359.
Ushikubi F, Segi E, Sugimoto Y, Murata T, Matsuoka T, Kobayashi T,Hizaki H, Tuboi K, Katsuyama M, Ichikawa A, Tanaka T, Yoshida N,Narumiya S. 1998. Impaired febrile response in mice lacking the pros-taglandin E receptor subtype EP3. Nature 395:281–284.
Van Dam AM, Brouns M, Man AHW, Berkenbosch F. 1993. Immunocyto-chemical detection of prostaglandin E2 in microvasculature and inneurons of rat brain after administration of bacterial endotoxin. BrainRes 613:331–336.
Watanabe Y, Watanabe Y, Hayaishi O. 1988. Quantitative autoradio-graphic localization of prostaglandin E2 binding sites in monkey dien-cephalon. J Neurosci 8:2003–2010.
Williams JW, Rudy TA, Yaksh TL, Viswanathan CT. 1977. An extensiveexploration of the rat brain for sites mediating prostaglandin-inducedhyperthermia. Brain Res 120:251–262.
Zhang J, Rivest S. 1999. Distribution, regulation and colocalization of thegenes encoding the EP2- and EP4-PGE2 receptors in the rat brain andneuronal responses to systemic inflammation. Eur J Neurosci 11:2651–2668.
Zhang J, Rivest S. 2000. A functional analysis of EP4 receptor-expressingneurons in mediating the actions of prostaglandin E2 within specificnuclei of the brain in response to circulating interluekin-1b. J Neuro-chem 74:2134–2145.
Zhang Y-H, Lu J, Elmquist JK, Saper CB. 2000. Lipopolysaccharide acti-vates specific populations of hypothalamic and brainstem neurons thatproject to the spinal cord. J Neurosci 20:6578–6586.
32 T. OKA ET AL.




















