Relationship between coccolith Sr/Ca ratios and coccolithophore production and export in the Arabian...
20
Deep-Sea Research II 54 (2007) 581–600 Relationship between coccolith Sr/Ca ratios and coccolithophore production and export in the Arabian Sea and Sargasso Sea Heather M. Stoll a,b, , Patrizia Ziveri c,d , Nobumichi Shimizu e , Maureen Conte f , Susanna Theroux a a Department of Geoscience, Williams College, Williamstown, MA 01267, USA b Department of Geologia, University de Oviedo, Arias de Velasco, s/n 30005, Oviedo, Asturias, Spain c Department of Earth Science, Vrije University Amsterdam, De Boelelaan, 1085 1081HV Amsterdam, The Netherlands d ICTA, Autonomous University of Barcelona (UAB), Bellaterra, Spain e Department of Geology and Geophysics, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USA f Bermuda Biological Station for Research, Ferry Reach, St. George’s GE 01, Bermuda Accepted 15 January 2007 Available online 7 March 2007 Abstract Coccolithophore CaCO 3 production can account for 20–80% of biogenic carbonate exported from the photic zone, and coccoliths are a dominant biogenic carbonate in many deep-sea sediments. A new method for picking individual coccoliths from sediment traps and sediments for analysis using Secondary Ion Mass Spectrometry (ion probe) allows us to make precise Sr/Ca and Mg/Ca determinations on coccoliths from single species even in samples where material is limited. There are large biological effects in Sr/Ca partitioning in coccoliths that have been related to variations in coccolithophore productivity. In sediment traps from the Sargasso Sea at Bermuda and Arabian Sea in the Somali Basin, we can identify Sr/Ca variations in several species that are consistent with inferred seasonal variations in coccolithophore productivity in surface waters. In the Arabian Sea, coccolith Sr/Ca ratios in Calcidiscus leptoporus and Helicosphaera carteri are lowest during the nonproductive intermonsoon. They are highest during the upwelling of southwest monsoon and during the nutrient entrainment from strong winds of the northeast monsoon. These Sr/Ca variations match seasonal trends in coccolith export flux. Furthermore, Sr/Ca variations in C. leptoporus are larger, and this species also has the greater variation in export flux between southwest monsoon and intermonsoon seasons. At Bermuda, a 1996 fall bloom, driven by passage of a warm mode water eddy, induced a large increase in Sr/Ca of C. leptoporus coincident with an increase of C. leptoporus export. Over an annual series for 2004, highest Sr/Ca ratios of C. leptoporus in the summer months match the typical summer peak in surface standing stock of this species and the stimulation of its productivity by mesoscale cyclonic eddies and eddy–eddy interactions. High Sr/Ca did not coincide with the highest export of C. leptoporus coccoliths, likely because cyclonic eddies, unlike mode-water eddies, are dominated by small phytoplankton with low export efficiency. These data confirm a relationship between coccolith Sr/Ca ratios and productivity. Sr/Ca ratios of the smaller and more slowly sinking Emiliania huxleyi at Bermuda show very small (o4%) seasonal Sr/Ca variations. In contrast, nutrient- limited culture experiments for this species reveal significant (435%) increases in Sr/Ca with nitrate-stimulated growth ARTICLE IN PRESS www.elsevier.com/locate/dsr2 0967-0645/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.dsr2.2007.01.003 Corresponding author. Department of Geoscience, Williams College, Williamstown, MA 01267, USA. Tel.: +1 413 597 4541; fax: +1 413 597 4116. E-mail address: [email protected] (H.M. Stoll).
Transcript of Relationship between coccolith Sr/Ca ratios and coccolithophore production and export in the Arabian...

ARTICLE IN PRESS
0967-0645/$ - see
doi:10.1016/j.ds
�Correspondifax: +1413 597
E-mail addre
Deep-Sea Research II 54 (2007) 581–600
www.elsevier.com/locate/dsr2
Relationship between coccolith Sr/Ca ratios andcoccolithophore production and export in the
Arabian Sea and Sargasso Sea
Heather M. Stolla,b,�, Patrizia Ziveric,d, Nobumichi Shimizue,Maureen Contef, Susanna Therouxa
aDepartment of Geoscience, Williams College, Williamstown, MA 01267, USAbDepartment of Geologia, University de Oviedo, Arias de Velasco, s/n 30005, Oviedo, Asturias, Spain
cDepartment of Earth Science, Vrije University Amsterdam, De Boelelaan, 1085 1081HV Amsterdam, The NetherlandsdICTA, Autonomous University of Barcelona (UAB), Bellaterra, Spain
eDepartment of Geology and Geophysics, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USAfBermuda Biological Station for Research, Ferry Reach, St. George’s GE 01, Bermuda
Accepted 15 January 2007
Available online 7 March 2007
Abstract
Coccolithophore CaCO3 production can account for 20–80% of biogenic carbonate exported from the photic zone, and
coccoliths are a dominant biogenic carbonate in many deep-sea sediments. A new method for picking individual coccoliths
from sediment traps and sediments for analysis using Secondary Ion Mass Spectrometry (ion probe) allows us to make
precise Sr/Ca and Mg/Ca determinations on coccoliths from single species even in samples where material is limited. There
are large biological effects in Sr/Ca partitioning in coccoliths that have been related to variations in coccolithophore
productivity. In sediment traps from the Sargasso Sea at Bermuda and Arabian Sea in the Somali Basin, we can identify
Sr/Ca variations in several species that are consistent with inferred seasonal variations in coccolithophore productivity in
surface waters. In the Arabian Sea, coccolith Sr/Ca ratios in Calcidiscus leptoporus and Helicosphaera carteri are lowest
during the nonproductive intermonsoon. They are highest during the upwelling of southwest monsoon and during the
nutrient entrainment from strong winds of the northeast monsoon. These Sr/Ca variations match seasonal trends in
coccolith export flux. Furthermore, Sr/Ca variations in C. leptoporus are larger, and this species also has the greater
variation in export flux between southwest monsoon and intermonsoon seasons. At Bermuda, a 1996 fall bloom, driven by
passage of a warm mode water eddy, induced a large increase in Sr/Ca of C. leptoporus coincident with an increase of
C. leptoporus export. Over an annual series for 2004, highest Sr/Ca ratios of C. leptoporus in the summer months match the
typical summer peak in surface standing stock of this species and the stimulation of its productivity by mesoscale cyclonic
eddies and eddy–eddy interactions. High Sr/Ca did not coincide with the highest export of C. leptoporus coccoliths, likely
because cyclonic eddies, unlike mode-water eddies, are dominated by small phytoplankton with low export efficiency.
These data confirm a relationship between coccolith Sr/Ca ratios and productivity. Sr/Ca ratios of the smaller and more
slowly sinking Emiliania huxleyi at Bermuda show very small (o4%) seasonal Sr/Ca variations. In contrast, nutrient-
limited culture experiments for this species reveal significant (435%) increases in Sr/Ca with nitrate-stimulated growth
front matter r 2007 Elsevier Ltd. All rights reserved.
r2.2007.01.003
ng author. Department of Geoscience, Williams College, Williamstown, MA 01267, USA. Tel.: +1 413 597 4541;
4116.
ss: [email protected] (H.M. Stoll).

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600582
rates. The high dominance of E. huxleyi production during bloom events, coupled with suspension of small E. huxleyi
coccoliths below the photic zone with very long transport times, appears to attenuate significantly the amplitude of Sr/Ca
variations in trap material. Coccoliths from these traps possess low Mg/Ca ratios characteristic of low-Mg calcite, so that
higher Mg ratios obtained in some extant coccoliths in cultures are not representative of typical open-ocean coccoliths
recovered in sediment trap samples. A significant correlation of Mg/Ca with sea-surface temperature is observed only for
H. carteri coccoliths in the Arabian Sea, and not in C. leptoporus in either site.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Arabian Sea; Sargasso Sea; Coccoliths; Coccolithophores; Sr/Ca
1. Introduction
Despite their small overall contribution to photo-synthesis on a global scale, the productivity ofcoccolithophore algae may be disproportionatelyimportant to the marine carbon cycle. Coccolithsare believed to provide mineral ballast that con-tributes significantly to the sinking efficiency ofparticulate organic carbon to the deep ocean(Francois et al., 2002; Klaas and Archer, 2002).The rain of coccolith and foraminiferal carbonaterelative to that of organic carbon, known as the rainratio, also determines the pH, and hence carbonsequestration, in deep waters. Changes in the pastproduction and export of coccolithophore carbo-nate, or the ratio of carbonate to organic carbonproduction, may have contributed to past changesin atmospheric CO2 and climate. Consequently,there is great interest in reconstructing past varia-tions in the rain of coccoliths from surface waters.However, dissolution of coccoliths in water columnand sediments hampers reconstruction of coccolithrain rates from sediment accumulation rates.
Recently, the elemental chemistry of coccolitho-phore algae was proposed as an indicator ofcoccolithophore productivity and, potentially, ofcoccolith rain rates. An advantage is that thechemical ratios would not be sensitive to dissolutionof coccoliths or sediment focusing. Along core toptransects across upwelling zones, the Sr/Ca ratio ofthe coccolith fraction covaried with productivity(Stoll and Schrag, 2000; Stoll et al., 2002) and innutrient-limited batch cultures Sr/Ca covaried withcell growth and calcification rates (Rickaby et al.,2002). Different size fractions dominated by differ-ent coccolith species have different absolute Sr/Caratios, and different amplitudes of Sr/Ca variation.In core top transects, the amplitude of Sr/Cavariations was highest in large surface dwellingspecies of coccolithophores such as Calcidiscus
leptoporus and lowest in deep photic zone dwellers
such as Florisphaera profunda. The variable Sr/Caamplitudes may indicate the different productivityresponses of these different species to upwelling,since upwelling might be expected to impact mostsignificantly the surface dwelling species (Stoll et al.,2002).
Several questions remain regarding the nature ofthe relationship between coccolithophore produc-tion, coccolith export, and the elemental chemistryof coccoliths. It has been difficult to verify whetherchanges in Sr/Ca ratios correlate with coccolitho-phore production in surface waters and whetherchanges in Sr/Ca ratios of different species indicatethe degree to which the productivity of that speciesor niche is varying. Furthermore, since the Sr/Caratio of coccoliths is set during their production inthe photic zone, it is unclear how this ratio relates tococcolith export, which can be decoupled fromproductivity and can be sensitive to food webstructure. It has been difficult to evaluate thecontrols of variations in coccolith Sr/Ca and toinfer the importance of coccolithophore productiv-ity to the oceans’ biogeochemical cycles becausethere are very limited data on the productivity of thecoccolithophores, or individual species of cocco-lithophores, in the surface ocean.
Here we apply a new analytical technique,analysis of Sr/Ca and Mg/Ca ratios of individuallypicked coccoliths via Secondary Ion Mass Spectro-metry (SIMS), to coccoliths of several speciescollected in sediment traps in two contrastingoceanographic regimes, the oligotrophic gyre ofthe Sargasso Sea near Bermuda and the high-productivity seasonal upwelling zone of the ArabianSea. In both regions there are strong seasonalvariations in coccolithophore and total productionand export which are well characterized. Thisapproach allows us to compare the geochemistryof individual species of exported coccoliths withexport fluxes of those coccoliths and estimates oftheir production in the water column. We compare

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 583
the sediment trap results with new data fromnitrate-limited chemostat cultures of Emiliania
huxleyi. These illustrate how coccolith chemistryvaries with growth rate and productivity of thisspecies, independent of export.
Our results show that Sr/Ca in C. leptoporus andHelicosphaera carteri is positively correlated withexport production of these species in the ArabianSea. In the Bermuda trap Sr/Ca of C. leptoporus
increases with C. leptoporus export during a fallmode water eddy event. However, increased Sr/Caratio during stimulation of C. leptoporus productionduring passage of summer cyclonic was notaccompanied by increased C. leptoporus export,likely because these eddies do not stimulate largerphytoplankton that enhance efficiency of export.This dataset provides the first direct comparisonsbetween coccolith export fluxes and coccolithchemistry.
2. Background on productivity and export in studied
trap sites
2.1. Arabian Sea off the coast of Somalia
A Technicap PPS-5 sediment trap array MST-9was moored in the Somali Basin at 10143.0680N and53134.4220E with a bottom depth of 4047m duringthe NIOP-JGOFS study in the Northwest IndianOcean. Traps were deployed at 1032m (MST9-E)and 3200m (MST9-G) from June 1992 to February1993. Details of sediment trap methods and designare given in (Brummer, 2002).
Monsoon-driven seasonal reversals in wind direc-tion in the Arabian Sea give rise to strong seasonalvariability in production. Strong summer southwestmonsoon (SWM) winds drive upwelling along thewestern boundary of the northeastern part of theIndian Ocean, resulting in a nearly 10 1C cooling ofsurface waters beginning in May and intensifyinguntil July. Upwelling of nutrients from a depth ofabout 200m increases surface concentrations ofnitrate (12 mmol) and phosphorus (1.1 mmol/kg),giving rise to a dramatic increase in productivityand surface chlorophyll, making this one of themost productive areas in the world ocean (datafrom World Ocean Atlas 2001). During the inter-monsoon period characterized by weak winds,typically from October–November and March–May, nutrients and productivity are low. Duringthe winter December–February, the strong winds ofthe NE monsoon entrain deep nutrients by vertical
mixing, thus stimulating a moderate enhancementin productivity.
Major seasonal shifts in phytoplankton commu-nity structure accompany the shifts in monsoonregimes and may be expected to influence exportefficiency. During the SWM, upwelling-influencedsites are dominated by diatoms (430% of primaryproduction) and Phaeocystis (Barlow et al., 1999).Small (o2 mm, non-coccolithophore) prymnesio-phytes comprise a lesser contribution to eukaryoticpopulation, and prokaryotic production (Barlowet al., 1999) accounts for o23% of primaryproduction. In contrast, during the intermonsoon,small prymnesiophytes (o2 mm) dominate thephytoplankton community, and diatoms andPhaeocystis contribute 7–21% of chlorophyll a
(Barlow et al., 1999). During the NEM, prokaryoteproduction dominates (450%) primary produc-tion, diatoms contribute an average of 8% ofprimary production and prymnesiophytes an aver-age of 11% (Brown et al., 2002). The highdominance of diatoms during the SWM leads tovery high export efficiency. Export ratios derivedfrom 234Th scavenging suggest that during the lateSWM, 17–28% of production is exported to deeptraps, whereas during the NEM and spring IM,5–10% of production is exported (Buesseler et al.,1998).
While coccolithophores are not a large propor-tion of the phytoplankton, they show strongseasonal variations in productivity. Measurementsof calcification by coccolithophores in the photiczone show three times higher community calcifica-tion rates and higher standing stocks of calciteduring the SWM compared to the October inter-monsoon (Balch et al., 2000). Fluxes of coccolitho-phores and coccoliths to the 1000-m trap increasedramatically during the upwelling of the SWMseason and during the wind-driven nutrient entrain-ment of the northeast monsoon season (Broerseet al., 2000). Fluxes of upper photic zone speciesE. huxleyi, Gephyrocapsa oceanica, Umbilicosphaera
sibogae, C. leptoporus, and to a lesser extentH. carteri, all increase significantly during theSWM. Fluxes of deep dwelling species F. profunda
and Gladiolithus flabellatus, which always inhabitdeep nutrient-rich waters, are much less variable.Comparable maxima in coccolith fluxes duringthe SWM and NEM were observed in sedimenttraps in the Northern Arabian Sea off the coast ofPakistan (Andruleit et al., 2000). The correspon-dence between seasonal trends in calcification rates

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600584
(Balch et al., 2000) and trap fluxes suggests thatelevated coccolith fluxes during the SWM aretracking higher coccolithophore production in sur-face waters and not merely enhanced exportefficiency of particles. This interpretation is furthersupported by the close correlation of coccolith andcoccosphere fluxes throughout high flux (SWM andNEM) and low flux (IM) periods, since the veryfragile coccospheres must result from recent pro-duction and not scavenging of material suspendedfor long periods in the water column. Fluxes ofcoccoliths to the deeper 3200-m sediment trap(Ziveri et al., 2000b) show greater short-termvariability during the SWM than those of the1000-m trap, but their fluxes track the overallpattern of minima in fluxes during the intermon-soon and maximum fluxes during the SWM. Theadditional variability probably results from variablelateral sediment movement of fine particles into thetrap due to strong currents. More rapidly sinkingplanktic foraminifera, which are unaffected bycurrents, show no high frequency variability, butrather single, clearly defined maxima in flux duringboth the SWM and NEM in the deep 3200-m trap(Conan et al., 2002). Since the 1000-m coccolithfluxes are more closely related to surface variationsin productivity and temperature, we use them forcomparison with coccolith chemistry in subsequentanalyses. Chemistry of the coccolith populations inthe 3200-m trap is unlikely to be affected by lateraltransport because over the integration period oftrap collection the temporal variations in oceano-graphic conditions and coccolithophore productiv-ity are regional in scope and affect the broad sourcearea of coccoliths.
2.2. Sargasso Sea at Bermuda
The Ocean Flux Program (OFP) sediment trapseries has sampled continuously since 1978. Thecurrent trap array is located at 311500N, 641100Wwith collection at 500, 1500, and 3200m. Trapsample processing has been previously summarized(Conte et al., 2001). In this study we presentanalyses for a 9-month interval from February toOctober of 2004 (1500-m trap), which captures themain succession from spring bloom to summerstratification. We also present analyses for a 2-month period from November 1996–January 1997(3200-m trap) that captures a high-productivityevent triggered by mixed-layer destratification dur-ing passage of a warm mesoscale feature (Conte
et al., 2003), also classified as a mode-water eddy(Sweeney et al., 2003).
The setting of the Bermuda sediment trap andhydrographic station has been summarized (Mi-chaels and Knap, 1996). Strong winter cooling inthe Sargasso Sea, accompanied by storm fronts,drives convective mixing down to 150–250m inFebruary or March. Entrainment of deeper nutri-ent-rich water stimulates increased surface nutrients(surface nitrate maximum of 0.3–0.71 mmol/kg;(Michaels and Knap, 1996) and a phytoplanktonspring bloom and increased productivity. Thisspring bloom typically coincides with highest massfluxes to the sediment traps.
In the summer, thermal stratification restrictsvertical nutrient entrainment. However, nitrogenfixation rates increase with summer eolian dustdeposition, which likely relieves iron limitation innitrogen-fixing diazotrophs (Orcutt et al., 2001).This fixed nitrogen fuels some additional produc-tion that draws down DIC in the summer (Orcuttet al., 2001). On subseasonal timescales, mesoscaleeddies and submesoscale features can enhancenutrient upwelling and mixing and biologicalproductivity (McGillicuddy et al., 1998; Conteet al., 2003; Sweeney et al., 2003; MourinoCarballado and McGillicuddy, 2006).
Compared to the Arabian Sea, prymnesiophytescomprise a more significant proportion of theeukaryotic phytoplankton community at the BATSsite. Prymnesiophytes, inferred to be largely cocco-lithophores, are most abundant during thespring bloom, when they comprise 44% of chlor-ophyll a, compared to 7% from diatoms and 17%from prokaryotes (Steinberg et al., 2001). Diatomblooms are rare (Steinberg et al., 2001) andclosely associated with the passage of mode-watereddies (Sweeney et al., 2003). During seasonsother than the spring bloom, prokaryotic pico-plankton often dominate (Steinberg et al., 2001). Onaverage, about 8% of primary production isexported to deep sediment traps (Steinberg et al.,2001).
Detailed study of the seasonal cycle of coccolitho-phore standing stocks for 1991–1993 in the watercolumn showed that the highest concentrations ofcoccolithophores occurred during the spring bloom.During the spring bloom, standing stocks weredominated by E. huxleyi (Haidar et al., 2000; Haidarand Thierstein, 2001). In the summer and fall,E. huxleyi abundances dropped and the coccolitho-phore population was dominated by F. profunda,

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 585
which dwelled in the deeper photic zone (100–150m),and Umbellosphera spp., which dwelled in the upperphotic zone (Fig. 1B). C. leptoporus abundancestypically peaked in the summer and/or fall, occa-sionally with a secondary peak during the springbloom (Haidar et al., 2000; Haidar and Thierstein,2001). H. carteri abundances peaked in the earlysummer through early fall period.
Our evaluation of the data on C. leptoporus
standing stocks for 1991–93 with the recent
Integrat
0
10
20
30
40
Dec-
90
Jan-9
1A
pr-
91
Jun-9
1A
ug-9
1O
ct-9
1D
ec-
91
Feb-9
2A
pr-
92
Jun
d
C. lept.
0
2000
4000
6000
8000
10000
Dec-9
0Jan-9
1A
pr-
91
Jun-9
1A
ug-9
1O
ct-
91
Dec-9
1F
eb-9
2A
pr-
92
J
0%
20%
40%
60%
80%
100%
Sep-9
1N
ov-
91
Dec-
91
Feb-9
2A
pr-
92
May-
92
Jul-92
Sep-9
2
10
6 s
ph
ere
s/m
2
10
6 s
ph
ere
s/m
2%
E.h
ux./(E
.hu
x+
F.
pro
f.)
(A)
(B)
(C)
Fig. 1. (A, B) Standing stock of coccospheres in the photic zone calculat
passage promoting nutrient upwelling (from Sweeney et al., 2003). Gra
2006). (C) Comparison of ratios of E. huxleyi/(E. huxleyi+F. profund
Adapted from Haidar et al. (2000). Due to long residence time of suspen
attenuated in the trap record.
compilation of dust accumulation (Sholkovitz andSedwick, 2006) shows no significant correspon-dence. However, our comparison for 1993, whenboth eddy passage data (Sweeney et al., 2003) andstanding stock data are available (Haidar et al.,2000; Haidar and Thierstein, 2001), shows a strongcorrespondence between peaks in standing stock ofC. leptoporus and passage of cold-core cycloniceddies, which promote upwelling of nutrient-richwater (Fig. 1A).
ed photic zone
-92
Aug-9
2O
ct-9
2D
ec-
92
Feb-9
3A
pr-
93
Jun-9
3A
ug-9
3O
ct-9
3D
ec-
93
Feb-9
4
ate
….SSA dataset available…
date
un-9
2A
ug-9
2O
ct-
92
Dec-9
2F
eb-9
3A
pr-
93
Jun-9
3A
ug-9
3O
ct-
93
Dec-9
3F
eb-9
4E. hux. F. prof.
Oct
-92
Dec-
92
Jan-9
3M
ar-
93
trap 3000m
photic zone
ed from Haidar and Thierstein (2001). Bars indicate cyclonic eddy
y circles denote dust Fe delivery (from Sholkovitz and Sedwick,
a) in photic zone and deep (3000m) sediment trap at Bermuda.
ded coccoliths, the seasonal cycle in the photic zone is significantly

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600586
Despite the pronounced seasonal succession ofcoccolithophore species in the photic zone, theseasonal succession is significantly attenuated, andin some cases entirely eliminated, in the flux ofcoccoliths to deep (3000m) sediment traps over the1992 cycle (Haidar et al., 2000). The flux of F.
profunda to deep (3200m) sediment traps remainsnearly constant throughout the year despite themajor summer–fall peak in standing stock. The fluxof E. huxleyi coccoliths rises with the spring bloom,but remains elevated throughout the summer andearly fall despite low photic zone abundances.Consequently, the proportion of E. huxleyi/(E. huxleyi+F. profunda) coccoliths in the photiczone is 495% E. huxleyi during the spring bloom,and o10% E. huxleyi in early fall. However, theratio in deep sediment traps ranges only from 60%to 30% (Fig. 1C; (Haidar et al., 2000). A similardecoupling of surface deposition and deep trapfluxes has been observed for atmosphericallydelivered mineral dust particles at Bermuda (Jickellset al., 1998). Seasonal changes in the efficiency ofparticle export, likely in response to changingphytoplankton community structure, concomitantwith scavenging of coccoliths (and mineral dust)suspended for long periods at moderate depths,likely account for this seasonal decoupling (Haidaret al., 2000).
Transient flux variations due to mesoscale fea-tures are superimposed on the seasonal cycles at theBermuda Site. In late November 1996, the passageof a warm mesoscale feature enhanced nutriententrainment and mixed the water column down to180m, resulting in a fall high productivity event andincreased mass flux out of the photic zone (200mtrap) and into the deep (3200m) trap (Conte et al.,2003). Concentrations of pigment 190 hexanolox-yfucoxanthin derived from haptophytes (includingcoccolithophores) increased dramatically in theupper photic zone and slightly at the deep chlor-ophyll maximum, indicating significantly higherstanding stocks within the mesoscale feature. Fluxesof alkenones, derived from E. huxleyi, increased bya factor of 30 in the deep trap from pre-event topeak fluxes, confirming increased productivity ofcoccolithophores and E. huxleyi in particular. In thedeep trap, lipid analyses indicate that in the initialmass pulse much of the material is derived not fromsurface production, but from biological aggregationof refractory organic compounds and residualsilicates suspended in the water column. Thesubsequent mass pulse contains new labile phyto-
plankton-derived organic carbon (Conte et al.,2003).
3. Methods
3.1. Trap sample preparation and Sr/Ca
measurements
For samples from the OFP, a 5–7mg split of thedry o125-mm fraction was massed and transferredto a 50-mL centrifuge tube. Five millilitres of 0.45-mm filtered seawater with pH adjusted to 8.5 withNH4OH were used to resuspend the sediment.Sediment was oxidized using 2mL of sodiumhypochlorite and 2mL of 30% hydrogen peroxide(Bairbakhish et al., 2001). After oxidation sedimentwas recovered on 0.45-mm polycarbonate or cellu-lose nitrate filter under syringe or vacuum filtration,and the filter was rinsed three times with 10mL ofdistilled water with pH adjusted to 8.5 withNH4OH. Sediment was resuspended off the filterin 5mL of 2% NH4OH, and filtered through a 20-mm Nylon Spectramesh (Spectrumlabs) to obtainthe coccolith (o20 mm) fraction. All filtrationapparatus and mesh were acid washed, rinsed, andneutralized with 2% NH4OH between samples toeliminate cross contamination.
A wet split (3% aliquot) of the o20-mm sizefraction was filtered onto a 25-mm 0.4-mm poly-carbonate filter for determination of CaCO3
o20 mm. All wet splits were collected with aprecision pipette in a minimum of four aliquots(e.g., a 1000-mL split is taken as 4* 250 mL) toimprove precision by compensating for heterogene-ity in the sample suspension. The o20-mm poly-carbonate filter was placed in a 15-mL centrifugetube with 2mL of 0.5% HNO3. A 25% aliquot wetsplit of the 20–125 mm fraction was also acidified in0.5% HNO3. For both, Ca was analyzed usingflame atomic absorption spectrometry on a PerkinElmer AAnalyst 300 using matrix-match standards.
The smallest coccoliths were separated from theo20-mm fraction by repeated decanting (Stoll andZiveri, 2002) using a settling distance of 6 cm for280min. Light microscope analysis indicates thatthe carbonate fraction of the supernatant isdominated by coccoliths of E. huxleyi and F.
profunda in variable proportion. This small cocco-lith fraction was then dissolved in 0.1M acetic acid/ammonium acetate buffer and Sr/Ca ratios weremeasured via Inductively Coupled Argon PlasmaSpectrometry in axial mode (Thermo Elemental

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 587
model IRIS 1000 DUO at Middlebury College) withprecisiono1% (r.s.d).
A fraction enriched in larger coccoliths wasproduced by microfiltering to obtain a o3-mm and3–12-mm fraction (for 1996–1997 series) or for the2004 series by repeated decanting of the sedimentedfraction described above, using a settling time of40min at 3.5 cm. The sediment after this settling wasdominated (volumetrically) by coccoliths of C.
leptoporus, U. sibogae, H. carteri, and Coccolithus
pelagicus. An aliquot of the coarse coccolithfraction was suspended in ethanol, sonicated brieflyto break up aggregates, and then pipetted on a glassslide for picking of individual coccoliths for ionprobe analysis. For the 1996–1997 series, a split ofthis coarse fraction also was dissolved in 0.1Macetic acid/ammonium acetate buffer, and Sr/Caratios were measured via Inductively CoupledArgon Plasma Spectrometry (ICP), as was donefor the finest coccolith fraction. For MST-9, forseveral samples the o32-mm fraction, and micro-filtered fractions of medium (5–8 mm) and coarse(8–10 mm) coccoliths (processed as in (Broerse et al.,2000) were dissolved for Sr/Ca measurement viaICP. A split of the o32-mm fraction from MST9was suspended in ethanol and sonicated briefly, thenpipetted on a glass slide for picking of individualcoccoliths for ion probe analysis.
The method for ion probe analysis of individuallypicked coccoliths is detailed elsewhere (Stoll et al.,submitted). In summary, a tungsten needle mountedin a Narishige micromanipulator was used to pickup individual coccoliths of C. leptoporus and H.
carteri, and for OFP E. huxleyi under 400�magnification on a Nikon Diaphot 300 invertedmicroscope equipped with polarizer. For each
Fig. 2. SEM image of picked epoxy-mounted coccoliths of C. leptoporu
75 mm in the left and 50 mm on the right.
sample and species, a population of 12–20 pickednannofossils was placed in a well on a Tacky Dot(SPI) slide. Epoxy (Buehler Epoxide) was pouredinto an aluminum mount on the tacky dot slide;when hardened the Tacky Dot slide was removedfrom the epoxy using ethanol to dissolve the TackyDot polymer coating. Samples were further exposedby abrasion on an ethanol-wetted felt pad. Identi-fications were confirmed by interference figuresunder cross-polarized transmitted-light microscopy,by scanning electron microscopy, and by reflected-light microscopy (Fig. 2).
Sr/Ca and Mg/Ca ratios were measured on aCameca IMS3f Secondary Ion Mass Spectrometerat the Northeast National Ion Microprobe Facilityat Woods Hole. The analytical method used here isbased on previous studies of trace elements inmarine carbonates (Hart and Cohen, 1996). A 20-mm beam of negatively charged oxygen ions with acurrent of 2 nA was rastered across a 50-mm squarearea covering the population of picked nannofossils.An energy offset of �90V was used for suppressinginterferences of molecular ions. Data were collectedin three consecutive analyses of five cyclic scans ofthe magnetic field covering 24 Mg to 40 Ca to 88 Sr,with typical counting errors of 1% or better for Sr88. We used a standard reference calcite from theOka carbonatite complex, Quebec (OKA-C). Thestandard was run multiple times using the sameanalytical protocol as the samples at the onset ofeach daily session, and these values were used tocalibrate for differential ionization efficiency ofdifferent elements. The ion probe technique repro-duced to within 1% the Sr/Ca ratios of monospecificcultured H. carteri samples, which also had beenmeasured by ICP atomic emission spectroscopy
s (left) and H. carteri (right). Width of circular tacky dot area is

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600588
(Stoll et al., 2002). Analyses of replicate populationsof Toweius coccoliths from the same sample, pickedand analyzed several months apart, yield Sr/Caratios, which differ by 1%, respectively. Contribu-tions to mass 88 and mass 40 from epoxy resinrepresent o0.2% of counts for average samples andin no case exceed 1% of total sample counts. Thesedata suggest comparable analytical uncertainty forSr/Ca ratios via ion probe and ICP methods.
Mg/Ca ratios measured by the ion probe aresubject to greater uncertainty due to molecularinterferences on mass 24. Background counts ofmass 24 on epoxy resin may represent 10% ofcounts for average sample sizes, and can signifi-cantly affect the calculated Mg/Ca ratio when thecoccolith sample mass is small. We calculated themagnitude of this artifact for each analysis based onthe ratio of signal from epoxy to signal fromcoccoliths as inferred from measured Ca intensity.In this calculation we assumed the mass 24contribution from epoxy is the same when rasteringa population of coccoliths as when just rasteringepoxy alone. This is a reasonable approximationsince in cross section there is significant void spacein coccolith populations so their actual volumecontribution to rastered volume is small. Because ofthis assumption, the corrected Mg/Ca we calculatedrepresents a lower limit on the Mg/Ca of thecoccoliths. Subsequent Mg/Ca data are presentedshowing both the Mg/Ca measured and an error barextending to the lower limit representing Mg/Caratio corrected for epoxy contribution. Ratios arereported in mol% Mg as is conventional inbiomineralization studies (e.g., Stanley et al.,2005), which is equivalent to 10 times the Mg/Caratio expressed in mmolmol�1 units conventionalfor foraminiferal Mg/Ca analysis.
For determination of coccolith fluxes in the OFPtraps, following oxidation in bleach, splits of theo20-mm fraction were filtered onto polycarbonatefilters (for SEM examination) and cellulose filters(for light microscope examination). Coccoliths ofthe abundant species E. huxleyi and F. profunda
were counted using scanning electron micro-scopy, whereas minor species (o5% abundance)C. leptoporus were counted using transects inpolarized light microscopy. For counts of minorspecies C. leptoporus, sufficient fields of view wereexamined to attain total counts of 50–100 cocco-liths. Because archive trap samples from OFP aredried, it is not possible to calculate coccospherefluxes for the OFP series.
We attempted to apply the ion probe method toanalysis of monthly plankton samples collected oncellulose filters at Bermuda by Haidar and Thier-stein (2001). However, the density of coccosphereson filters was too sparse to allow for recovery ofsufficient coccoliths resuspended off the filter onto aglass slide for picking. In situ analysis of cocco-spheres on the filter was also not possible. Thesparse distribution and diversity of coccoliths on thefilter, and the limited magnification and resolutionof the optics on the ion probe, precluded focusingthe ion beam on discrete, identifiable coccospheres.
3.2. Culture methods
Three strains of E. huxleyi, AC284, AC320, andNAP4, which were isolated from the Mediterraneanand Atlantic (Ian Probert, ALGOBANK, Univer-sity of Caen, France), were grown in a nitrate-limited chemostat and nutrient-replete semi-contin-uous batch culture. The strains were pre-adapted tofull-growth media and ambient conditions for aminimum of six generations before being introducedinto the chemostat or batch culture experiments.Media was prepared from seawater collected at adepth of 60m off the coast of Woods Hole, MA byWHOI Environmental Systems Laboratory Sea-water was vacuum filtered using a Millipore Steri-cup Sterile Vacuum Filter Unit (pore size 0.22 mm)to eliminate particulates and possible bacterialcontamination. Media were enriched using therecipe for K/5 media of (Keller et al., 1987), forall components except nitrate and ammonium. Noammonium was added to the media. For nutrient-replete semi-continuous batch cultures, nitrate wasadded according to K/5 recipe. For nitrate-limitedchemostats, we evaluated the nitrate levels requiredso that media would support no more than200,000 cells/mL. Initial concentrations of nitratein the seawater were estimated using the WorldOcean Atlas (Conkright, 2002). Given initial nitrateof 5.8 mM, full K/5 nutrient replete media for semi-continuous culture had 176.8 mM NO3
� and nitrate-limited chemostat media had 12 176.8 mM NO3
�.The cultures were grown in 400-mL sterilized Pyrexbeakers in a laminar flow hood with 24 h illumina-tion intensity of 117 mEm�2 s�1 and constant tem-perature of 24 1C. Cells were suspended using asterilized, polycarbonate, magnetic floating stirbar.Beakers were capped with a sterilized cotton squarewrapped in cheesecloth and held in place with awatchglass to permit air exchange but preclude

ARTICLE IN PRESS
E. huxleyi
2.0
2.5
3.0
3.5
0 0.25 0.5 0.75 1
growth rate
Sr/
Ca (
mm
ol/m
ol)
full nutrient (K/5)
med. chemostat
slow chemostat
Fig. 3. Sr/Ca ratios of E. huxleyi grown in culture vs. growth rate
(day�1). Different experimental conditions (slow chemostat,
medium chemostat, and full-nutrient batch) are illustrated by
different symbols. Within each experimental condition, a point is
shown for each of the three strains (except for the medium
chemostat in which only two strains were successfully cultured).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 589
bacterial contamination. Cellular growth rates(day�1) were calculated from cell density checkedby daily cell counts on a Coulter Counter. In thecase of chemostat experiments these counts wereadjusted for culture dilution rate. For chemostatcultures, an Ismatec IP High-precision Multichan-nel Peristaltic Pump with Pharmed santoprene gas-tight tubing was run continuously to cycle newmedia into the culture vessel and pump outequivalent volume of cell-bearing culture. The pumpwas used to dilute the culture volume of 390mL atrates of 130 or 260mLday�1 in the slow andmedium growth rate experiments, respectively. Thisset of experimental conditions resulted in slowestgrowth rates for the slow chemostat, modest growthrates for the medium chemostat, and maximumgrowth rates in the nutrient-replete batch culture.After the chemostat had run for 7–14 days, or thebatch cultures had reached cell densities of 150,000cells/mL, 100–200mL of each culture were har-vested by centrifugation. The pellet was rinsed fivetimes in distilled water adjusted to pH 8.5 withNH4OH; then a split was dissolved in 0.1M aceticacid/ammonium acetate buffer and Sr/Ca ratioswere measured via ICP in axial mode (ThermoElemental model IRIS 1000 DUO at MiddleburyCollege) with precisiono1% (r.s.d).
4. Results and discussion
4.1. Cultured E. huxleyi
Sr/Ca ratios of cultured E. huxleyi range from2.33 to 3.33mmolmol�1, with lowest Sr/Ca ratiosobserved in the slowest growing slow chemostatcultures, and highest Sr/Ca ratios in the fastestgrowing chemostat and nutrient-replete batch cul-tures (Fig. 3). These data are supportive of apositive relationship between coccolith Sr/Ca ratiosand nutrient-limited growth rate in E. huxleyi assuggested by Rickaby et al. (2002). Sr/Ca ratios ofE. huxleyi in these culture experiments are compar-able to those measured from sediment traps atBermuda (2.6–2.8mmolmol�1). If the averagerelationship between Sr/Ca and growth rate iscomparable in strains at Bermuda and thosecultured, it would suggest that Bermuda E. huxleyi
cells were produced at 0.1–0.35 day�1. This range iscomparable to inferred growth rates of alkenoneproducers in high-productivity settings: Bidigareet al. (1997) calculated average growth rates ofalkenone producers in the Equatorial Pacific upwel-
ling zone and reported maximum mcell of 0.4 andminimum rates of 0.25. Average growth rates ofprymnesiophytes at BATS range from 0.2 to0.6 day�1 (Goericke, 1998).
4.2. Arabian Sea Sr/Ca records
Sr/Ca ratios of C. leptoporus track the broadseasonal productivity shifts and changes in cocco-lithophore flux in the Arabian Sea, with ratios thatare significantly lower during the fall intermonsoonperiod than during either the SWM or NEM(Fig. 4). The maximum Sr/Ca ratios in C. leptoporus
(July 19–26) precede the maximum export ofC. leptoporus (September 13–27) by nearly 2months. We believe that the high Sr/Ca C.
leptoporus arriving at the trap in mid July wereproduced during high productivity of the upwellingSWM season because they lag by over a month theonset of upwelling indicated by cold sea-surfacetemperatures (June 15) and lag the arrival of thefirst foraminifera with d18O signatures reflectingcold upwelled waters in the deep trap (June 21–July5; Conan et al., 2002). Sinking rates of at least85mday�1 would be required for peak Sr/Ca toreflect the onset of upwelling, and these rates arecomparable to bulk settling velocity of 150m/dinferred from the lag between SST and light d15Nvalues in trap organic carbon (Knappertsbusch andBrummer, 1995; Brummer, 2002) as well as settling
ARTICLE IN PRESS
0
2
4
6
8
10
12
Jun-9
2
Jul-92
Aug-9
2
Sep-9
2
Oct
-92
Nov-
92
Dec-
92
Jan-9
3
Feb-9
3
10
3sh
ells/m
2/d
0E+00
1E+06
2E+06
3E+06
4E+06
5E+06
6E+06
sp
here
s/m
2/d
SWM IM NEM(A)
(B)
(C)
(D)
0
0.2
0.4
0.6
0.8
1
1.2
Jun-9
2
Jul-92
Aug-9
2
Sep-9
2
Oct
-92
Nov-
92
Dec-
92
Jan-9
3
Feb-9
3
mg
/m2/d
C. leptoporus
H. carteri
2.0
2.5
3.0
3.5
Jun-9
2
Jul-92
Aug-9
2
Sep-9
2
Oct
-92
Nov-
92
Dec-
92
Jan-9
3
Feb-9
3
Sr/
Ca o
fC. le
pt.
2.5
3.0
3.5
4.0
Sr/
Ca o
f H
.cart
eri
H. carteri
C. leptoporus
20
24
28
Jun-9
2
Jul-92
Aug-9
2
Sep-9
2
Oct
-92
Nov-
92
Dec-
92
Jan-9
3
Feb-9
3
SS
T
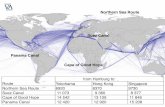
Fig. 4. Seasonal cycle in Arabian Sea at MST9, 1992–93. (A) Total coccosphere fluxes from MST9 trap (Broerse et al., 2000) (open
diamond symbols and heavy line showing 3 point moving average), and total fluxes of planktic foraminifera (trianges, from Conan et al.,
2002). (B) Ion probe Sr/Ca (mmolmol�1) measurements on C. leptoporus (filled circle, left axis) and H. carteri (open square, right axis). (C)
Mass fluxes of C. leptoporus and H. carteri coccoliths in the MST9 1000m trap (from Broerse et al., 2000). (D) Onset of upwelling is
indicated by the abrupt drop in sea-surface temperature as recorded by satellite and ship-based measurements (from Broerse et al., 2000).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600590
velocities of 143–243mday�1 calculated for cocco-liths during high productivity North Atlanticblooms (Ziveri et al., 2000a). The 1995 JGOFScruise sampling showed that there is a significant(1–2 month) lag between the onset of increasedprimary production triggered by SWM, and theincrease in export from the surface ocean, which islikely controlled by the onset and decline of thediatom bloom (Buesseler et al., 1998). Sr/Ca ratiosof H. carteri also track the broad productivitydecrease from the peak at the SWM to minimaduring the fall intermonsoon (Fig. 4). As was thecase for C. leptoporus, the maximum Sr/Ca ratiosprecede maximum export of H. carteri, probably forsimilar reasons.
Sr/Ca ratios of H. carteri are higher than those ofC. leptoporus, but the magnitude of Sr/Ca variationbetween SWM and IM is much larger in C.
leptoporus (50%) than that of H. carteri (nearly30%). The amplitude of variation in mass fluxbetween the intermonsoon and SWM is nearlydouble in C. leptoporus (factor of 10 flux increase)compared to H. carteri (factor of 5 flux increase).This suggests that the amplitude of Sr/Ca variation
among these two species reflects the degree to whichthe physiology or niche of that species has beenstimulated to increased productivity. The data areconsistent with that observed over spatial transectsin upwelling regions where C. leptoporus, a surface-dwelling species most stimulated by upwelling,shows much larger amplitude Sr/Ca increases thandeep-dwelling F. profunda, which is not stimulatedby upwelling (Stoll et al., 2002).
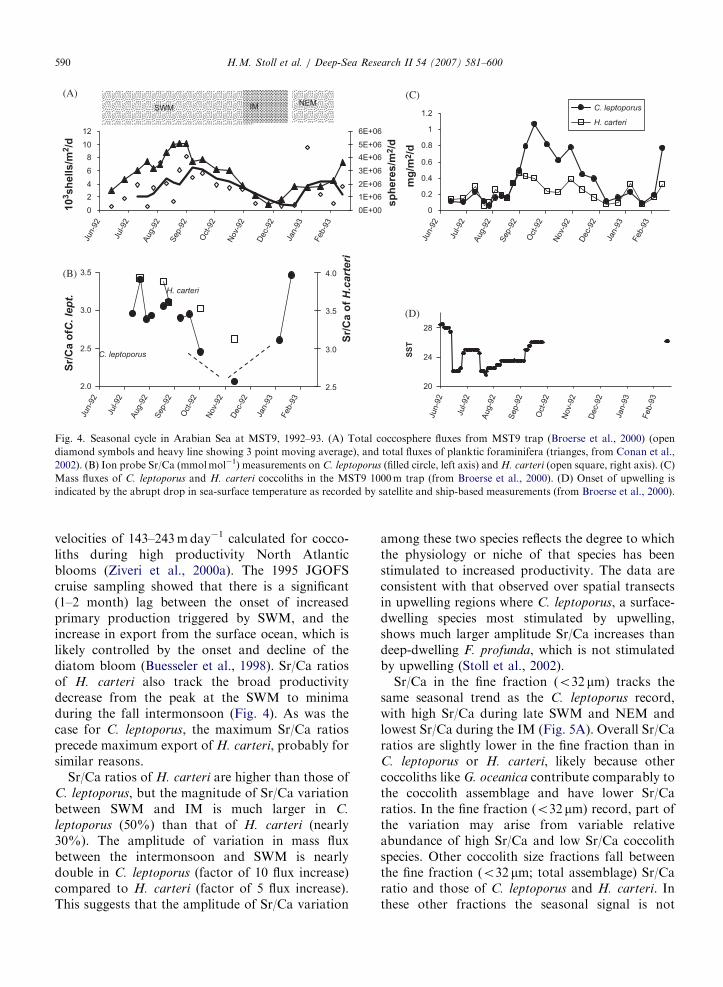
Sr/Ca in the fine fraction (o32 mm) tracks thesame seasonal trend as the C. leptoporus record,with high Sr/Ca during late SWM and NEM andlowest Sr/Ca during the IM (Fig. 5A). Overall Sr/Caratios are slightly lower in the fine fraction than inC. leptoporus or H. carteri, likely because othercoccoliths like G. oceanica contribute comparably tothe coccolith assemblage and have lower Sr/Caratios. In the fine fraction (o32 mm) record, part ofthe variation may arise from variable relativeabundance of high Sr/Ca and low Sr/Ca coccolithspecies. Other coccolith size fractions fall betweenthe fine fraction (o32 mm; total assemblage) Sr/Caratio and those of C. leptoporus and H. carteri. Inthese other fractions the seasonal signal is not
ARTICLE IN PRESS
MST9G Arabian Sea
2.0
2.5
3.0
3.5
4.0
May-9
2
Jul-92
Sep-9
2
Oct-
92
Dec-9
2
Jan-9
3
Mar-
93
Sr/
Ca (
mm
ol/m
ol)
C. lept.(ion probe)
H. cart.(ion probe)
<32 um (ICP)
med. Coccos (ICP)
coarse coccos (ICP)
SWM NEM
2
2.5
3
3.5
4
4.5
Nov-96 Dec-96 Jan-97 Feb-97
Sr/
Ca (
mm
ol/m
ol)
C. lept (ion
probe)
E. hux (ion
probe)
H. carteri
(ion probe)
fine
coccoliths
(ICP)
coarse
coccos
(ICP)
onset of mixing
(A)
(B)
Fig. 5. Detailed Sr/Ca data on all measured fractions from (A) MST9 trap in Arabian Sea, (B) OFP trap in Bermuda. In (A), upwelling
period during southwest monsoon (SWM) indicated by depressed sea-surface temperatures and period of enhanced winds during northeast
monsoon (NEM) are highlighted. In (B), arrow denotes the onset of the mesoscale mixing event as defined by homogenization of water
temperatures between surface and 54m depth (Conte et al., 2003).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 591
resolved due to lack of sample coverage during theIM.
4.3. Bermuda Sr/Ca records
4.3.1. Fall 1996 mesoscale mixing event
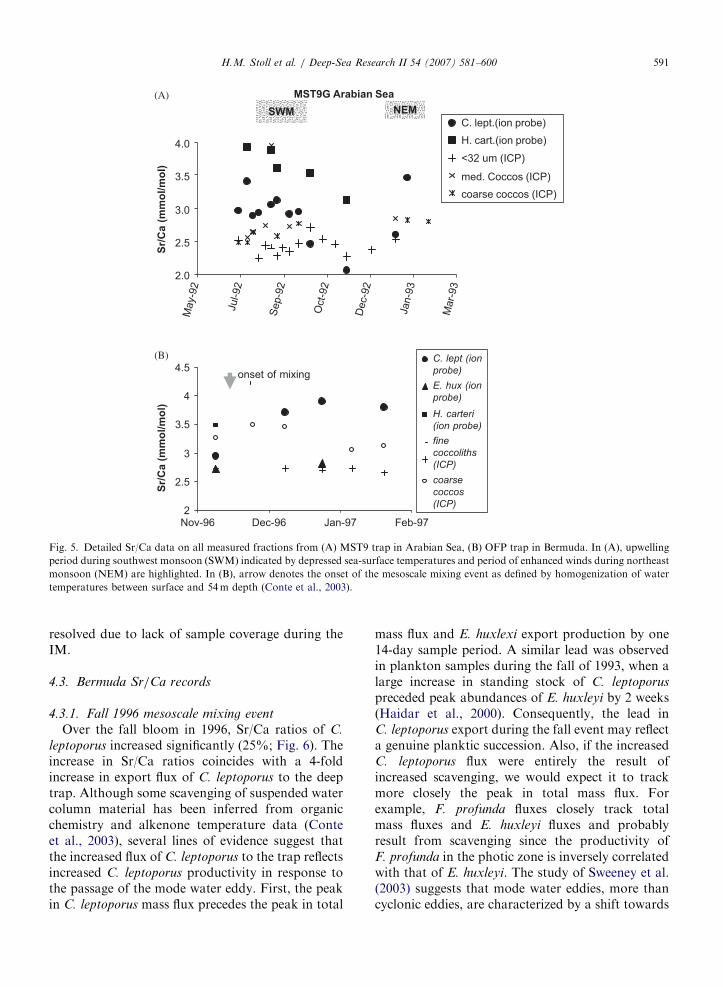
Over the fall bloom in 1996, Sr/Ca ratios of C.
leptoporus increased significantly (25%; Fig. 6). Theincrease in Sr/Ca ratios coincides with a 4-foldincrease in export flux of C. leptoporus to the deeptrap. Although some scavenging of suspended watercolumn material has been inferred from organicchemistry and alkenone temperature data (Conteet al., 2003), several lines of evidence suggest thatthe increased flux of C. leptoporus to the trap reflectsincreased C. leptoporus productivity in response tothe passage of the mode water eddy. First, the peakin C. leptoporus mass flux precedes the peak in total
mass flux and E. huxlexi export production by one14-day sample period. A similar lead was observedin plankton samples during the fall of 1993, when alarge increase in standing stock of C. leptoporus
preceded peak abundances of E. huxleyi by 2 weeks(Haidar et al., 2000). Consequently, the lead inC. leptoporus export during the fall event may reflecta genuine planktic succession. Also, if the increasedC. leptoporus flux were entirely the result ofincreased scavenging, we would expect it to trackmore closely the peak in total mass flux. Forexample, F. profunda fluxes closely track totalmass fluxes and E. huxleyi fluxes and probablyresult from scavenging since the productivity ofF. profunda in the photic zone is inversely correlatedwith that of E. huxleyi. The study of Sweeney et al.(2003) suggests that mode water eddies, more thancyclonic eddies, are characterized by a shift towards

ARTICLE IN PRESS
2
2.5
3
3.5
4
4.5
Nov-96 Dec-96 Jan-97 Feb-97
Sr/
Ca (
mm
ol/m
ol)
C. lept (ion
probe)
E. hux (ion
probe)
fine
coccoliths
(ICP)
onset of mixing
0.E+00
1.E+09
2.E+09
3.E+09
4.E+09
5.E+09
Nov-96 Dec-96 Jan-97 Feb-97
lth
s/m
2/d
(Eh
ux+
F. p
rof)
i
0.E+00
2.E+06
4.E+06
6.E+06
8.E+06
1.E+07
lith
s/m
2/d
(C. le
pt)
F. profunda
E. huxleyi
C. leptoporus
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Nov-96 Dec-96 Jan-97 Feb-97
Eh
ux/(
Eh
ux+
F. p
rof)
0
20
40
60
80
mg
/m2/d
ay f
lux
E.hux/(Ehux.+F. prof.)
trap flux
(A)
(B)
(C)
Fig. 6. Fall 1996 mode-water eddy event in Sargasso Sea at OFP. (A) Sr/Ca ratios of coccoliths, (B) export production, and (C) total trap
mass flux and proportion of E. huxleyi/(E.huxleyi+F. profunda). In (A), arrow denotes the onset of the mesoscale mixing event as defined
by homogenization of water temperatures between surface and 54m depth (Conte et al., 2003).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600592
larger phytoplankton like diatoms, which facilitatemore efficient export from the photic zone to thetraps. Indeed, pigment data from BATS verifyincreased diatom production during the fall 1996event (Conte et al., 2003).
During the fall bloom the Sr/Ca ratios of the E.
huxleyi and F. profunda fraction shows only a veryslight increase in Sr/Ca ratios, which are near thelevel of analytical precision. While the variablecontribution of E. huxleyi and F. profunda to thisfraction may partially obscure geochemical changesoccurring in either species, ion probe analyses
confirm that Sr/Ca ratios of pure E. huxleyi duringthe bloom are only 3.5% higher than those justbefore the fall bloom, barely resolvable fromanalytical uncertainty. The 50-fold increase inE. huxleyi flux to the deep trap, 30-fold increase inalkenone flux to the trap (Conte et al., 2003), anddramatic increase in the E. huxleyi/F. profunda
ratios in trap samples suggest that E. huxleyi
production did increase dramatically during themesoscale mode water event. Since the culture datahere and elsewhere show that E. huxleyi produced atdifferent nitrate levels and different growth rates

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 593
exhibit a significant range (at least 35%) in Sr/Caratios, the lack of contrast in Sr/Ca ratios betweenthe coccoliths collected in the trap just before theonset of the fall bloom and during the bloomsuggests that coccoliths collected at both timeintervals were produced under similar physiologicalconditions (Fig. 3; Rickaby et al., 2002). Conse-quently, we infer that coccoliths of E. huxleyi
arriving at the trap just before the fall bloom aredominated by suspended coccoliths produced dur-ing the previous spring bloom. Alkenone tempera-tures in deep trap sediments from early fall 1996suggest that alkenones settling to the deep trap inlate fall were likely derived from the spring bloom(Conte et al., 2003). Data from 1991 to 1992indicate that over 90% of the annual standing stockof E. huxleyi in the photic zone occurs inJanuary–May (Haidar et al., 2000; Haidar andThierstein, 2001). If the growth rate of E. huxleyi
were constant throughout the year, then more than90% of E. huxleyi coccoliths would be producedduring the spring bloom. This percentage wouldbe even greater if growth rates of E. huxleyi
were higher during the spring bloom than duringthe summer and fall. The long residence time ofE. huxleyi coccoliths in the water column, inferredfrom the constancy of the sediment trap E. huxleyi /F. profunda ratio (Fig. 1C; Haidar et al., 2000;Haidar and Thierstein, 2001), could be sufficient toattenuate the range of Sr/Ca variations encounteredin sediment trap samples. For example, if coccolithsproduced during E. huxleyi’s high-productivityseason had Sr/Ca ratios 30% higher than thoseproduced during E. huxleyi’s lower-productivityseason, but E. huxleyi coccoliths arriving at thetrap always contained a maximum of 100% andminimum of 85% coccoliths derived from the high-productivity season, then the contrast in Sr/Cawould be reduced from 30% to less than 4%.
Sr/Ca ratios from the coarse coccolith fractionfall mostly between those of small coccoliths and C.
leptoporus and do not parallel the Sr/Ca trends of C.
leptoporus, either because other dominant contribu-tors to the coarse coccoliths like H. carteri have adifferent Sr/Ca trend or because changes in therelative contribution of coccoliths with differentabsolute Sr/Ca ratios obscures the trend (Fig. 5B).
4.3.2. 2004 seasonal cycle
Sr/Ca ratios of C. leptoporus rise slightly in midsummer and then significantly in late summer andremain high through the fall (Fig. 7). The highest
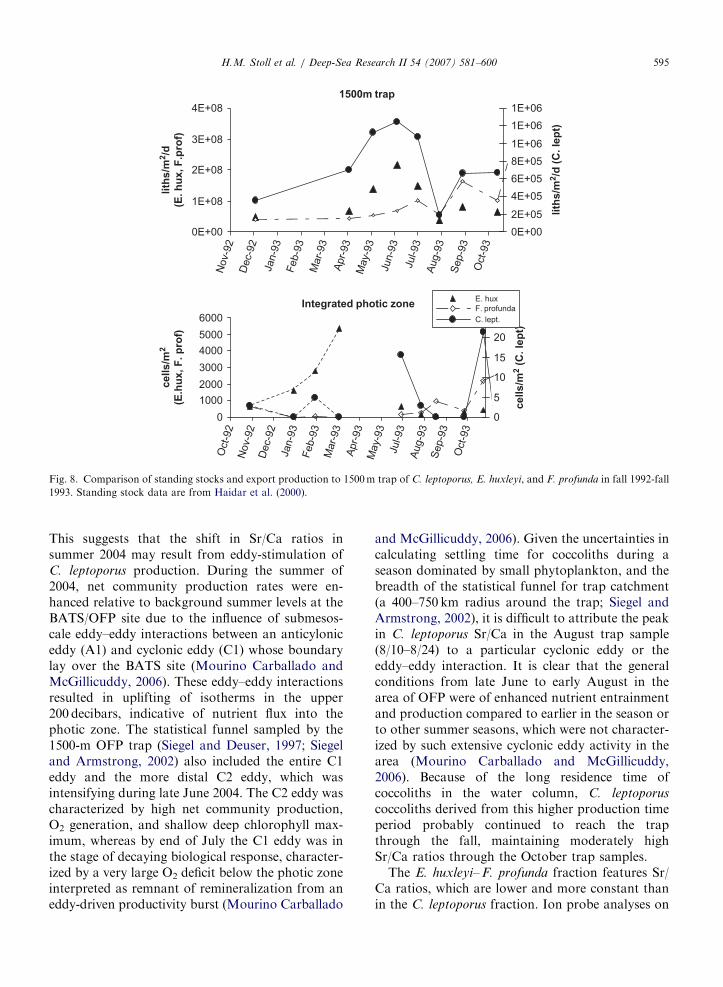
ratios in the summer of 2004 are comparable tothose observed at the peak of the fall 1996productivity event, whereas the ratios in spring2004 are comparable to those observed prior toonset of the fall 1996 event. On the annualtimescale, the peak in C. leptoporus Sr/Ca ratios inlate summer does not correspond with maximumexport fluxes of C. leptoporus, which instead occurduring the spring bloom. One explanation is that thehigh C. leptoporus flux during the spring bloom doesnot represent new production of C. leptoporus, butrather, enhanced export efficiency and perhapsscavenging of suspended C. leptoporus coccoliths.For example, during the spring bloom in 1993,integrated photic zone standing stocks of C. lepto-
porus increased only slightly (o50% relative to latefall 1992 levels), much less than during summer andfall events in 1993 when standing stock increased 6and 7 times over fall 1992 levels (Haidar et al., 2000;Haidar and Thierstein, 2001). However, the flux ofC. leptoporus to the 1500-m trap tripled during thespring bloom in 1993 (Fig. 8). Conversely, the majorsummer peak in C. leptoporus standing stock in1993 corresponds to lower rates of C. leptoporus
export. This may reflect a dramatic increase inexport efficiency and/or scavenging during thespring bloom when large phytoplankton like dia-toms are present, contrasting with low exportefficiency in the summer when cyclonic eddiesincrease production of small Synechococcus (Swee-ney et al., 2003). Alternatively, the high flux duringthe 1993 spring bloom could reflect a very transientincrease in C. leptoporus standing stock, which wasnot captured by the resolution of plankton sam-pling. However, this latter explanation seemsunlikely given the long duration of elevatedC. leptoporus fluxes and their close correspondencewith elevated total mass fluxes, productivity, andfluxes of E. huxleyi. Furthermore, higher exportefficiency and strong scavenging during the springbloom is also implicated in the higher fluxes ofF. profunda in the traps in spring of 1993, and moregenerally in the peak flux of dust to deep sedimenttraps during the spring bloom despite delivery of80% of dust in summer months (Jickells et al.,1998). These data suggest that export fluxes ofC. leptoporus in the OFP traps may not be a goodindicator of C. leptoporus production in surfacewaters, especially in the cases of productionstimulated by cyclonic eddies.
The high late summer and fall Sr/Ca ratios ofC. leptoporus are consistent with higher summer and

ARTICLE IN PRESS
2.4
2.6
2.8
3.0
3.2
3.4
3.6
3.8
Feb-
04
Mar-
04
Apr-
04
May-
04
May-
04
Jun-04 Jul-04 Aug-
04
Sep-
04
Oct-04 Nov-
04
Sr/
Ca (
mm
ol/m
ol)
C. lept (ion probe)
E. hux (ion probe)
H. carteri (ion probe)
Small coccos (ICP)
eddy activity
0E+00
1E+09
2E+09
3E+09
4E+09
5E+09
Feb-04 Mar-04 Apr-04 May-04 May-04 Jun-04 Jul-04 Aug-04 Sep-04 Oct-04 Nov-04
lith
s/m
2/d
ay(E
. h
ux, F
. p
rof.
)
0E+00
1E+07
2E+07
3E+07
4E+07
5E+07
6E+07
lith
s/m
2/d
ay (
C. le
pt)
0
20
40
60
80
100
Feb-
04
Mar-
04
Apr-
04
May-
04
May-
04
Jun-
04
Jul-04 Aug-
04
Sep-
04
Oct-
04
Nov-
04
julian day midpoint
mg
/m2/d
0%
20%
40%
60%
80%
100%
%E
hu
x/(
Eh
ux+
Fp
)
trap flux
trap ratio
C. lept.
Ehux
F. profunda
(A)
(B)
(C)
Fig. 7. Seasonal cycle in Sargasso Sea at OFP in 2004. (A) Sr/Ca ratios of coccoliths, (B) export production of coccoliths, and (C) total
trap mass flux and proportion of E. huxleyi/(E. huxleyi+F. profunda). Box denotes timing of cyclonic eddies and eddy–eddy interactions
within the area sampled by the 1500m OFP trap, as identified by Mourino and McGillicuddy (in review).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600594
fall productivity of this species in surface waters,likely coincident with eddy-enhancement of produc-tion (Fig. 1A; Haidar et al., 2000; Haidar andThierstein, 2001). In detail, for the 1993 dataset of
photic zone standing stock, both summer and fallpeaks in C. leptoporus standing stock coincide withcyclonic eddy events that promote upwelling ofnutrient-rich water into the photic zone (Fig. 1A).
ARTICLE IN PRESS
1500m trap
0E+00
1E+08
2E+08
3E+08
4E+08
Nov-9
2
Dec-9
2
Jan-9
3
Feb-9
3
Mar-
93
Apr-
93
May-9
3
Jun-9
3
Jul-93
Aug-9
3
Sep-9
3
Oct-
93
lith
s/m
2/d
(E. h
ux, F
.pro
f)
0E+00
2E+05
4E+05
6E+05
8E+05
1E+06
1E+06
1E+06
lith
s/m
2/d
(C
. le
pt)
Integrated photic zone
0
1000
2000
3000
4000
5000
6000
Oct-
92
Nov-9
2D
ec-9
2
Jan-9
3
Feb-9
3
Mar-
93
Apr-
93
May-9
3
Jul-93
Aug-9
3S
ep-9
3
Oct-
93
cells/m
2
(E.h
ux, F
. p
rof)
10
15
20
25
cells/m
2 (
C. le
pt)
F. profunda
C. lept.
E. hux
0
5
Fig. 8. Comparison of standing stocks and export production to 1500m trap of C. leptoporus, E. huxleyi, and F. profunda in fall 1992-fall
1993. Standing stock data are from Haidar et al. (2000).
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 595
This suggests that the shift in Sr/Ca ratios insummer 2004 may result from eddy-stimulation ofC. leptoporus production. During the summer of2004, net community production rates were en-hanced relative to background summer levels at theBATS/OFP site due to the influence of submesos-cale eddy–eddy interactions between an anticyloniceddy (A1) and cyclonic eddy (C1) whose boundarylay over the BATS site (Mourino Carballado andMcGillicuddy, 2006). These eddy–eddy interactionsresulted in uplifting of isotherms in the upper200 decibars, indicative of nutrient flux into thephotic zone. The statistical funnel sampled by the1500-m OFP trap (Siegel and Deuser, 1997; Siegeland Armstrong, 2002) also included the entire C1eddy and the more distal C2 eddy, which wasintensifying during late June 2004. The C2 eddy wascharacterized by high net community production,O2 generation, and shallow deep chlorophyll max-imum, whereas by end of July the C1 eddy was inthe stage of decaying biological response, character-ized by a very large O2 deficit below the photic zoneinterpreted as remnant of remineralization from aneddy-driven productivity burst (Mourino Carballado
and McGillicuddy, 2006). Given the uncertainties incalculating settling time for coccoliths during aseason dominated by small phytoplankton, and thebreadth of the statistical funnel for trap catchment(a 400–750 km radius around the trap; Siegel andArmstrong, 2002), it is difficult to attribute the peakin C. leptoporus Sr/Ca in the August trap sample(8/10–8/24) to a particular cyclonic eddy or theeddy–eddy interaction. It is clear that the generalconditions from late June to early August in thearea of OFP were of enhanced nutrient entrainmentand production compared to earlier in the season orto other summer seasons, which were not character-ized by such extensive cyclonic eddy activity in thearea (Mourino Carballado and McGillicuddy,2006). Because of the long residence time ofcoccoliths in the water column, C. leptoporus
coccoliths derived from this higher production timeperiod probably continued to reach the trapthrough the fall, maintaining moderately highSr/Ca ratios through the October trap samples.
The E. huxleyi– F. profunda fraction features Sr/Ca ratios, which are lower and more constant thanin the C. leptoporus fraction. Ion probe analyses on

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600596
two pure E. huxleyi populations, one from thespring bloom, and one from late fall, show valuesthat differ by only 4%. This is close to the limit ofanalytical precision. As was the case for the fall1996 samples, the large majority of E. huxleyi
coccoliths reaching the trap in summer and late fallprobably derive from the spring bloom. The longresidence time of spring-bloom-derived material inthe water column is consistent with the elevatedratios of E. huxleyi/F. profunda observed in the trapin the late summer and fall when data of (Haidaret al., 2000) indicate that photic zone productionis dominated almost exclusively by F. profunda
(Figs. 7, 1C).
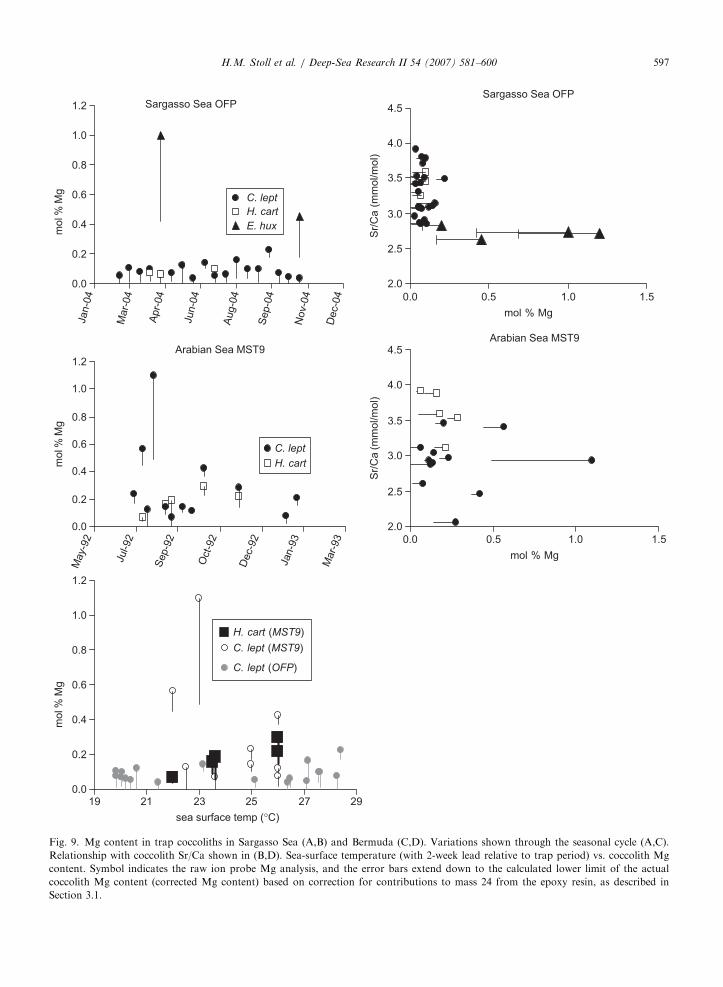
4.4. Mg content of coccoliths
Coccoliths of C. leptoporus and H. carteri fromBermuda show very low Mg content (o0.2mol%;Fig. 9A). Mg content shows no correlation with Sr/Ca ratios (Fig. 9B) and, in the case of the fullseasonal record from C. leptoporus, shows norelationship with seasonal variations in C. lepto-
porus export production and no significant correla-tion to sea-surface temperature (r2 ¼ 0.05; Fig. 9E).Low Mg calcite with slightly higher Mg content of0.2–0.4mol% (corrected values as described inSection 3.1) is observed in the two samples ofE. huxleyi from the Bermuda trap, with the highestvalue observed during the spring bloom. Mgcontents of E. huxleyi measured from the fall 1996event are comparable (not plotted), with Mgcontent 0.4mol% in the pre-productivity eventsample and 0.1% at the peak of the E. huxleyi
export pulse. These data suggest the potential forgreater variability in Mg content of E. huxleyi,which in part reflects greater analytical uncertaintydue to the low mass of E. huxleyi coccoliths and thegreater proportional contribution of mass 24 fromepoxy. No consistent relationship is observedbetween Mg content and export production orresidence time of coccoliths in the water column.
Mg contents for C. leptoporus and H. carteri
coccoliths from Arabian Sea traps are slightlyhigher and more variable (Fig. 9C), perhaps becausethese samples were not subject to oxidative or ionexchange cleaning prior to analysis. Still, only twosamples had corrected mol%Mg greater than 0.4%.Corrected Mg contents for C. leptoporus were notsignificantly correlated to sea-surface temperature(r2 ¼ 0.14), while those for H. carteri showed astrong positive correlation temperature (r2 ¼ 0.8;
Fig. 9E). As was the case for Bermuda traps, Mgcontent shows no correlation with Sr/Ca ratios orcoccolith export flux (Fig. 9D).
Mg content also was measured in C. pelagicus
isolated from the NABE48 sediment trap (48N21W) collected in 1989. Samples from bloom andnon-bloom collections had low Mg contents of 0.5and 0.4mol% Mg, respectively.
While the Mg/Ca ratio in many biogenic carbo-nates is positively correlated with sea-surfacetemperature, only the H. carteri samples from theArabian Sea traps show a consistent correlationbetween coccolith Mg/Ca ratios and sea-surfacetemperature. In C. leptoporus, tight cellular controlover calcification and the chemistry of coccolithsmay preclude the expression of such a relationship.Alternatively, Mg present in other phases likecoccolith-bound organic fractions obscures a tem-perature dependence of Mg content in the calciteitself.
The low Mg content of coccoliths recovered fromsediment traps are consistent with previous studies(Siesser, 1977) and culture data of these and otherspecies (Stoll et al., 2001) which indicate very lowMg in coccolith calcite. The data contrast withresults from Stanley et al. (2005) indicating that twococcolithophore species, Pleurochrisis carterae andOchrosphaera neopolitana, secrete high-Mg calcite44mol% when cultured in modern seawater.Stanley et al. (2005) suggested that the high Mg/Ca ratio of seawater in the late Cenozoic, comparedwith lower seawater Mg/Ca inferred for LateCretaceous and Earliest Cenozoic, promotes highMg/Ca in coccoliths. They suggest that high Mg/Cacurtails growth rates for most extant coccolitho-phore species, and that coccolithophores were moresuccessful during the late Cretaceous and earlyCenozoic due to lower seawater Mg/Ca ratios. Ourdata suggest that the dominant modern coccolitho-phores responsible for most coccolith carbonateproduction in the modern ocean (e.g., C. leptoporus,C. pelagicus, H. carteri, E. huxleyi, and G. oceanica;Ziveri et al., 2007) are able to maintain very low Mgcalcite despite the high modern seawater Mg/Caratio. These dominant calcifiers of the modernCoccolithaceae (C. pelagicus)+Calcidiscaceae (C.
leptoporus), and Noelaerhabdaceae (E. huxleyi andGephyrocapsa) families, are not new lineages, butrather descendents from lineages of Coccolithaceaeand Prinsiaceae (Bown et al., 2004), respectively,which were the dominant coccoliths in earlyPaleocene European chalk sequences (Bown,
ARTICLE IN PRESS
Sargasso Sea OFP
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Jan-0
4
Mar-
04
Apr-
04
Jun-0
4
Aug-0
4
Sep-0
4
Nov-0
4
Dec-0
4
mol %
Mg
C. lept
H. cart
E. hux
Arabian Sea MST9
0.0
0.2
0.4
0.6
0.8
1.0
1.2
May-
92
Jul-92
Sep-9
2
Oct
-92
Dec-
92
Jan-9
3
Mar-
93
mol %
Mg
C. lept
H. cart
Sargasso Sea OFP
2.0
2.5
3.0
3.5
4.0
4.5
0.0 0.5 1.0 1.5
mol % Mg
Sr/
Ca (
mm
ol/m
ol)
Arabian Sea MST9
2.0
2.5
3.0
3.5
4.0
4.5
0.0 0.5 1.0 1.5
mol % Mg
Sr/
Ca (
mm
ol/m
ol)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
19 21 23 25 27 29
sea surface temp (°C)
mol %
Mg
H. cart (MST9)
C. lept (MST9)
C. lept (OFP)
Fig. 9. Mg content in trap coccoliths in Sargasso Sea (A,B) and Bermuda (C,D). Variations shown through the seasonal cycle (A,C).
Relationship with coccolith Sr/Ca shown in (B,D). Sea-surface temperature (with 2-week lead relative to trap period) vs. coccolith Mg
content. Symbol indicates the raw ion probe Mg analysis, and the error bars extend down to the calculated lower limit of the actual
coccolith Mg content (corrected Mg content) based on correction for contributions to mass 24 from the epoxy resin, as described in
Section 3.1.
H.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 597

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600598
2005). The dominant modern calcifiers of the openocean appear to have evolved close cellular regula-tion of Ca uptake that very effectively excludes Mgfrom the calcifying vesicle.
5. Conclusions and implications
5.1. Coccolith Sr/Ca and coccolithophore production
Our data suggest that in both the upwelling-dominated system of the Arabian Sea and theoligotropic gyre of the Sargasso Sea, stimulus incoccolithophore production in the photic zone leadsto higher Sr/Ca ratios in coccoliths captured insediment traps. This is true for monsoon-stimulatedproduction of H. carteri and C. leptoporus in theArabian Sea caused by summer upwelling and wind-driven winter convective mixing, and is true formesoscale eddy-driven nutrient enhancement of C.
leptoporus production in the Sargasso Sea.
5.2. Coccolith Sr/Ca and coccolith export
In the Arabian Sea, this correlation betweencoccolithophore production and coccolith Sr/Caalso extends to coccolith and coccosphere export,whereas in Bermuda, the export production of C.
leptoporus (and other coccoliths) is frequentlydecoupled from their production. To a large degreethis may reflect a very high efficiency of coccolithsinking during the highest productivity period in theArabian Sea, and lower residence time of coccolithsin the water column, at least for the larger coccolithsof H. carteri and C. leptoporus examined in thissetting. In the Arabian Sea, especially during theSWM, diatoms and large phytoplankton comprise amuch larger portion of the phytoplankton commu-nity. Diatoms and other large phytoplankton areresponsible for a disproportionately large share ofcarbon export because their large size causes highersinking rates and shorter food webs. In addition,some varieties make flocculent masses that increasesinking rate (Buesseler, 1998).
The closer coupling of export flux and coccolithSr/Ca and production in the Arabian Sea site mayin part reflect the seasonal synchroneity in the max-imum production of dominant upper photic zonecoccoliths (including C. leptoporus and H. carteri)and overall production including large diatoms, allstimulated by the upwelling of the SWM. None-theless, coccolith export production also closelyfollows the second peak in coccolith Sr/Ca during
the northeast monsoon, when diatoms are not sucha significant portion of the phytoplankton andexport efficiency is expected to be lower. Thissuggests that another, as yet unidentified, factor isin part responsible for the close coupling of exportand production in the Arabian Sea.
Given that the elemental chemistry of individualcoccoliths is set in surface waters and is unaffectedby the timing or efficiency of export, the Sr/Cachemistry of coccoliths recovered in the traps mayserve as an effective marker for processes of exportand productivity. Because of the greater resistanceof coccoliths to degradation compared to organiccomponents, the chemistry of coccoliths in sedimenttraps may yield more salient signals of productionpulses than other indicators. For example, in theBermuda region Sr/Ca of C. leptoporus may serve asan indicator of export of material derived from eddystimulation of productivity. Because of mixing ofcoccoliths from different production regimes in trapsamples and the long residence time of suspendedcoccoliths in the water column in some settings, thefull amplitude of Sr/Ca variation among high- andlow-productivity seasons may be underestimated byseasonal trap time series. If it becomes possible toobtain high-volume filter samples of coccoliths fromthe photic zone to constrain end member coccolithchemistry, then it should be possible to evaluate theextent of mixing and selective preservation ofcoccoliths from different seasons in the sedimenttrap material. This would provide further informa-tion on the processes and timing of coccolith exportrelevant to models of ocean biogeochemical cycling.
5.3. Magnitude of coccolith Sr/Ca variations for a
given species over seasonal cycles
The different magnitudes of coccolith Sr/Cavariation observed in different species over seasonalproductivity cycles reflect several processes. Thereappear to be genuine differences in the seasonalamplitude of Sr/Ca change arising from differentdegrees of productivity stimulation in differentniches. For example, in the Arabian Sea setting,the different magnitudes of Sr/Ca change amongspecies are proportional to the degree of change inexport production of the different species. Inaddition, it is possible that coccolithophores ofdifferent physiology have different slopes of re-sponse of coccolith Sr/Ca to a given degree ofproductivity stimulation. According to one mechan-ism for coccolith Sr/Ca regulation, at lower nutrient

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600 599
availability coccolithophores extrude more extra-cellular polysaccharides, which bind Sr preferen-tially to Ca, depressing the Sr/Ca ratio of free ionsin the near-cell region and causing a lower Sr/Caratio in calcifying vescicle and coccoliths duringnutrient limitation (Langer et al., 2006). Given thatcell size and other factors unique to a given speciesmay regulate the degree of polysaccharide extrusionin response to nutrients, it is possible that differentspecies may have different amplitudes of Sr/Cachange in response to a given productivity stimulus.
The expression of these types of primary changesin coccolith Sr/Ca in the sediment trap record alsoappears to depend on the efficiency of settling andon the degree of seasonality in production by agiven species. Species with very narrow ecologicaltolerance, or a very high degree of proliferation in avery narrow set of conditions, may not record arange of conditions but rather record only thoseduring which that particular species grows orproliferates. For example, in both Arabian Seaand Bermuda production of C. leptoporus seems tooccur over broader range of conditions and so itproduces coccoliths during both less and morefavorable conditions exhibiting a large range ofSr/Ca. In contrast, at least in oceanic settings,E. huxleyi seems to produce prolifically, but only atits most favorable conditions. In addition, part ofthe differences in the response of C. leptoporus vs.E. huxleyi at Bermuda may reflect the tenfolddifference in size and settling rate. Individual smallE. huxleyi and F. profunda coccoliths can remainsuspended 10 times longer than larger coccolithslike C. leptoporus. Consequently, the smaller cocco-liths are more likely to have long residence time inwater column and more attenuated geochemicalsignal due to mixing of populations produced indifferent seasons. This latter aspect is likely to affectthe magnitude of the Sr/Ca signal over seasonaltimescales. However, for reconstruction of pastcoccolithophore productivity from coccolith Sr/Caratios, seasonal and longer-scale mixing will beinherent in sediment records from most parts of theocean and Sr/Ca shifts in downcore records willreflect changes in mean conditions.
Acknowledgments
We thank Tyler Auer and Nora Matell fortechnical assistance picking coccoliths, and J.C.Weber for assistance in collecting OFP data andsamples. Research supported by NSF OCE-0424474
to H.M. Stoll and a fellowship to H. Stoll from theSpanish Ministry of Education co-funded by theEuropean Social Fund. Sediment trap samples fromthe Arabian Sea are part of the Netherlands IndianOcean Programme and were provided by Gert-JanBrummer and supported by NWO 854.00.005.Sediment trap samples from Bermuda are collectedthrough the Ocean Flux Program which is sup-ported by NSF OCE-0509602 and OCE-0325627.
References
Andruleit, H.A., von Rad, U., Bruns, A., Ittekkot, V., 2000.
Coccolithophore fluxes from sediment traps in the north-
eastern Arabian Sea off Pakistan. Marine Micropaleontology
38, 285–308.
Bairbakhish, A.N., Bollmann, J., Sprengel, C., Thierstein, H.R.,
2001. Disintegration of aggregates and coccospheres in
sediment trap samples. Marine Micropaleontology, 219–223.
Balch, W.M., Drapeau, D., Fritz, J., 2000. Monsoonal forcing of
calcification in the Arabian Sea. Deep-Sea Research II 47,
1301–1337.
Barlow, R.G., Mantoura, R.F.C., Cummings, D.G., 1999. Mon-
soonal influence on the distribution of phytoplankton pigments
in the Arabian Sea. Deep-Sea Research II 46 (3–4), 677–699.
Bidigare, R.R., Fluegge, A., et al., 1997. Consistent fractionation of
C-13 in nature and in the laboratory: growth-rate effects in some
haptophyte algae. Global Biogeochemical Cycles 11, 279–292.
Bown, P., 2005. Calcareous nannoplankton evolution: a tale of
two oceans. Micropaleontology 51 (4), 299–308.
Bown, P., Lees, J., Young, J., 2004. Calcareous nannoplankton
evolution and diversity through time. In: Thierstein, H.,
Young, J. (Eds.), Coccolithophores: from Molecular Pro-
cesses to Global Impact. Springer, Berlin, pp. 482–508.
Broerse, A.T.C., Brummer, G.J.A., Van Hinte, J.E., 2000.
Coccolithophore export production in response to monsoonal
upwelling off Somalia (northwestern Indian Ocean). Deep-
Sea Research II 47 (9–11), 2179–2205.
Brown, S.L., Landry, M.R., Christensen, S., Garrison, D.,
Gowing, M.M., Bidigare, R.R., Campbell, L., 2002. Micro-
bial community dynamics and taxon-specific phytoplankton
production in the Arabian Sea during the 1995 monsoon
seasons. Deep-Sea Research II 49 (12), 2345–2376.
Brummer, G.J., 2002. Monsoon-driven export fluxes and early
diagenesis of particulate nitrogen and its delta 15N across the
Somalia margin. Geological Society Special Publications 195,
353–370.
Buesseler, K., Ball, L., Andrews, J., Benitez-Nelson, C., Bela-
stock, R., Chai, F., Chao, Y., 1998. Upper ocean export of
particulate organic carbon in the Arabian Sea derived from
thorium-234. Deep-Sea Research Part II-Topical Studies in
Oceanography 45 (10–11), 2461–2487.
Buesseler, K.O., 1998. The decoupling of production and
particulate export in the surface ocean. Global Biogeochem-
ical Cycles 12 (2), 297–310.
Conan, S.M.H., Ivanova, E.M., Brummer, G.J.A., 2002.
Quantifying carbonate dissolution and calibration of for-
aminiferal dissolution indices in the Somali Basin. Marine
Geology 182, 325–349.

ARTICLE IN PRESSH.M. Stoll et al. / Deep-Sea Research II 54 (2007) 581–600600
Conkright, M.E.E.A., 2002. World Ocean Atlas 1001; Objective
Analyses, Data Statistics, and Figures, CD-ROM Documenta-
tion. National Oceanographic Data Center, Silver Spring, MD.
Conte, M.H., Dickey, T.D., Weber, J.C., Johnson, R.J., Knap,
A.H., 2003. Transient physical forcing of pulsed export of
bioreactive material to the deep Sargasso Sea. Deep-Sea
Research I 50, 1157–1187.
Conte, M.H., Ralph, N., Ross, E.H., 2001. Seasonal and
interannual variability in deep ocean particle fluxes at the
Oceanic Flux Program (OFP)/Bermuda Atlantic Time Series
(BATS) site in the western Sargasso Sea near Bermuda. Deep-
Sea Research II 48, 1471–1505.
Francois, R., Honjo, S., Krishfield, R., Manganini, S., 2002. Factors
controlling the flux of organic carbon to the bathypelagic zone
of the ocean. Global Biogeochemical Cycles 16.
Goericke, R., 1998. Response of phytoplankton community
structure and taxon-specific growth rates to seasonally
varying physical forcing in the Sargasso Sea off Bermuda.
Limnology and Oceanography 43 (5), 921–935.
Haidar, A.T., Thierstein, H.R., 2001. Coccolithophore dynamics
off Bermuda (N. Atlantic). Deep-Sea Research Part II-
Topical Studies in Oceanography 48, 1925–1956.
Haidar, A.T., Thierstein, H.R., Deuser, W.G., 2000. Calcareous
phytoplankton standing stocks, fluxes and accumulation in
Holocene sediments off Bermuda (N. Atlantic). Deep-Sea
Research II 47, 1907–1938.
Hart, S.R., Cohen, A.L., 1996. An ion probe study of annual
cycles of Sr/Ca and other trace elements in corals. Geochimica
Et Cosmochimica Acta 60, 3075–3084.
Jickells, T.D., Dorling, S., Deuser, W.G., Church, T.M.,
Arimoto, R., Prospero, J.M., 1998. Air-borne dust fluxes to
a deep water sediment trap in the Sargasso Sea. Global
Biogeochemical Cycles 12, 311–320.
Keller, M.D., Selvin, R.C., Claus, W., Guilland, R.R.L., 1987.
Media for the culture of oceanic ultraphytoplankton. Journal
of Phycology 23, 633–638.
Klaas, C., Archer, D. E., 2002. Association of sinking organic
matter with various types of mineral ballast in the deep sea:
implications for the rain ratio. Global Biogeochemical Cycles 16.
Knappertsbusch, M., Brummer, G.J.A., 1995. A Sediment Trap
Investigation of Sinking Coccolithophorids in the North-
Atlantic. Deep-Sea Research I 42, 1083–1109.
Langer, G., Nehrke, G., et al., 2006. Coccolith strontium to
calcium ratios in Emiliania huxleyi: the dependence on
seawater strontium and calcium concentrations. Limnology
and Oceanography 51, 310–320.
McGillicuddy, D.J., Robinson, A.R., et al., 1998. Influence of
mesoscale eddies on new production in the Sargasso Sea.
Nature 394, 263–266.
Michaels, A.F., Knap, A.H., 1996. Overview of the US JGOFS
Bermuda Atlantic Time-series Study and the Hydrostation S
program. Deep-Sea Research II 43, 157–198.
Mourino Carballado, B., McGillicuddy, D.J., 2006. Mesoscale
variability in the metabolic balance of the Sargasso.
Limnology and Oceanography 51 (6), 2675–2689.
Orcutt, K.M., Lipschultz, F., Gundersen, K., Arimoto, R.,
Michaels, A.F., Knap, A.H., Gallon, J.R., 2001. A seasonal
study of the significance of N-2 fixation by Trichodesmium
spp. at the Bermuda Atlantic Time-series Study (BATS) site.
Deep-Sea Research II 48, 1583–1608.
Rickaby, R.E.M., Schrag, D.P., Zondervan, I., Riebesell, U.,
2002. Growth rate dependence of Sr incorporation during
calcification of Emiliania huxleyi. Global Biogeochemical
Cycles 16 (1).
Sholkovitz, E.R., Sedwick, P.N., 2006. Open-ocean deployment
of a buoy-mounted aerosol sampler on the Bermuda Testbed
Mooring: Aerosol iron and sea salt over the Sargasso Sea.
Deep-Sea Research I 53 (3), 547–560.
Siegel, D.A., Armstrong, R.A., 2002. Trajectories of sinking
particles in the Sargasso Sea: modeling of statistical funnels
above deep-ocean sediment traps (vol. 44, pp. 1519, 1997).
Deep-Sea Research I 49, 1115–1116.
Siegel, D.A., Deuser, W.G., 1997. Trajectories of sinking particles
in the Sargasso Sea: modeling of statistical funnels above deep-
ocean sediment traps. Deep-Sea Research I 44, 1519–1541.
Siesser, W.G., 1977. Chemical Composition of Calcareous
Nannofossils. South African Journal of Science 73 (9), 283.
Stanley, S.M., Ries, J.B., Hardie, L.A., 2005. Seawater chemistry,
coccolithophore population growth, and the origin of
Cretaceous chalk. Geology 33 (7), 593–596.
Steinberg, D.K., Carlson, C.A., Bates, N.R., Johnson, R.J.,
Michaels, A.F., Knap, A.H., 2001. Overview of the US
JGOFS Bermuda Atlantic Time-series Study (BATS): a
decade-scale look at ocean biology and biogeochemistry.
Deep-Sea Research II 48 (8–9), 1405–1447.
Stoll, H.M., Encinar, J.D., Alonso, J.I.G., Rosenthal, Y.,
Probert, I., Klaas, C., 2001. A first look at paleotemperature
prospects from Mg in coccolith carbonate: cleaning techni-
ques and culture measurements. Geochemistry Geophysics
Geosystems 2.
Stoll, H.M., Schrag, D.P., 2000. Coccolith Sr/Ca as a new
indicator of coccolithophorid calcification and growth rate.
Geochemistry Geophysics Geosystems 1, 1–24.
Stoll, H.M., Shimizu, N., Arevalos, A., Matell, N., Banasiak, A.,
Zeren, S., submitted. Insights on coccolith chemistry from a
new ion probe method for analysis of individually picked
coccoliths. Geochemistry Geophysics Geosystems.
Stoll, H.M., Ziveri, P., 2002. Separation of monospecific and
restricted coccolith assemblages from sediments using differ-
ential settling velocity. Marine Micropaleontology 46 (1–2),
209–221.
Stoll, H.M., Ziveri, P., Geisen, M., Probert, I., Young, J.R., 2002.
Potential and limitations of Sr/Ca ratios in coccolith
carbonate: new perspectives from cultures and monospecific
samples from sediments. Philosophical Transactions of the
Royal Society of London Series a-Mathematical Physical and
Engineering Sciences 360 (1793), 719–747.
Sweeney, E.N., McGillicuddy, D.J., Buesseler, K.O., 2003.
Biogeochemical impacts due to mesoscale eddy activity in
the Sargasso Sea as measured at the Bermuda Atlantic Time-
series Study (BATS). Deep-Sea Research Part II-Topical
Studies in Oceanography 50, 3017–3039.
Ziveri, P., Bernardi, B., Baumann, K.-H., Stoll, H.M., Mortyn,
P.G., 2007. Sinking of coccolith carbonate and potential
contribution to organic carbon ballasting in the deep ocean.
Deep-Sea Research II, in press [doi:10.1016/j.dsr2.2007.01.006].
Ziveri, P., Broerse, A.T.C., van Hinte, J.E., Westbroek, P.,
Honjo, S., 2000a. The fate of coccoliths at 48 degrees N 21
degrees W, northeastern Atlantic. Deep-Sea Research Part II-
Topical Studies in Oceanography 47 (9–11), 1853–1875.
Ziveri, P., Giunta, S., Broerse, A., Ganssen, G., 2000b.
Coccolithophorid response to an upwelling hydrographic
system: example from offshore Somalia. Eighth International
Nannoplankton Association Conference, Bremen, Germany.