
Regulation of mesangial cell apoptosis and proliferation by intracellular Ca2+ signals
9
Kidney International, Vol. 58 (2000), pp. 1876–1884 HORMONES – CYTOKINES – SIGNALING Regulation of mesangial cell apoptosis and proliferation by intracellular Ca 21 signals HILMI SALEH,EBERHARD SCHLATTER,DETLEF LANG,HANS-GERD PAUELS, and STEFAN HEIDENREICH Department of Medicine, University of Mu ¨ nster, Mu ¨ nster, Germany abolished thapsigargin-dependent, but not angiotensin II- or Regulation of mesangial cell apoptosis and proliferation by ATP-induced Ca 21 rises when administered during a 12-hour intracellular Ca 21 signals. preincubation. Background. In inflammatory glomerular diseases, prolifer- Conclusions. Our data suggest that a sustained increase of ation, as well as apoptosis of mesangial cells (MCs), has been [Ca 21 ] i may serve as a signal to trigger MC apoptosis. Growth shown histomorphologically. Both processes may regulate the factors such as PDGF can abolish apoptosis induced by eleva- cellular content of the mesangium by closely influencing each tions of [Ca 21 ] i by altering intracellular Ca 21 signaling. other. In the present study, we examined whether the cyto- plasmic free Ca 21 concentration [Ca 21 ] i is involved as a key second messenger in the regulation of proliferative and apo- ptotic events. In many forms of inflammatory or metabolic glomeru- Methods. Thapsigargin, an inhibitor of the endoplasmic lar diseases, mesangial cells (MCs) can change their phe- Ca 21 -Mg 21 -ATPase, was used as a test substance to investigate the role of [Ca 21 ] i in signaling MC apoptosis and growth in notype and become proliferative and matrix expanding. vitro. Apoptosis was determined by nuclear chromatin staining Paracrine or autocrine growth factors such as platelet- with Hoechst 33258, by a [ 3 H]-thymidine–based DNA fragmen- derived growth factor (PDGF) or insulin-like growth tation assay or by flow cytometry detecting binding of FITC- factor-1 [1, 2], vasoactive agonists, for example, angioten- conjugated annexin V. Proliferation was measured by [ 3 H]- sin II and endothelin [3, 4], or leukocyte-derived cyto- thymidine incorporation into acid-precipitable material and corroborated by cell counting. kines [5] have been shown to play a key role for the Results. Thapsigargin significantly induced apoptosis and in- induction of proliferation and remodeling of the mesan- hibited proliferation dose dependently in nanomolar concen- gium [6]. Apoptosis, or programmed cell death, is a trations without evoking necrotic damage when administered physiological process that is able to reduce surplus cell not longer than 12 hours. Significant apoptosis was measurable numbers during tissue development, differentiation, ma- after a six-hour treatment of MCs with thapsigargin. Determina- tion of [Ca 21 ] i by fura-2–dependent spectrofluorometry showed lignant transformation, or inflammation [7–9]. It is char- that thapsigargin was able to induce prolonged [Ca 21 ] i rises acterized by a fixed genetic program that is mostly trig- that could be prevented by preincubation with the intracellular gered or modified by signals from the environment. In Ca 21 chelator 1,2-bis(2-aminophenoxy)-ethane-N,N,N9, N9-tetra- glomerular diseases, apoptosis has been shown to occur acetic acid (BAPTA) acetomethyl ester (AM). BAPTA had in patients with proliferative glomerulonephritis [10] or no influence on MC viability but reversed thapsigargin-induced apoptosis to control levels. After thapsigargin treatment (100 in the experimental model of Thy 1.1 nephritis, where nmol/L, 12 hours), apoptotic MCs had a significantly higher glomerular hypercellularity is counterbalanced and re- [Ca 21 ] i of 251 6 25 nmol/L (N 5 41) as compared with MCs solved by apoptotic processes of MCs [11]. However, that were not or not yet apoptotic ([Ca 21 ] i of 116 6 20 nmol/L, the mechanisms of MC apoptosis induction in vivo or in N 5 26, P , 0,05). Platelet-derived growth factor (PDGF), a vitro and the involved second messenger pathways are well-characterized growth factor for MCs, reversed the effects of thapsigargin on proliferation and apoptosis in a similar fashion hardly understood. Previous studies have described that as BAPTA. PDGF acutely stimulated increases of [Ca 21 ] i but reactive oxidative intermediates, such as nitric oxide [12] or superoxide [13], or antibodies ligating the Fas receptor [14] or the Thy 1 epitope [15] can evoke programmed Key words: cell death, platelet-derived growth factor, thapsigargin, cell death of MCs. The concept that the cytosolic-free inflammatory glomerular disease, necrotic damage, metabolic glomeru- lar disease. calcium concentration [Ca 21 ] i may act as a key signal for apoptosis induction was first described for thymocytes Received for publication February 15, 1999 and lymphocytes, because a variety of proteases and and in revised form May 12, 2000 Accepted for publication May 16, 2000 endonucleases that are generally engaged as effector molecules in the apoptotic cascade are activated by rises 2000 by the International Society of Nephrology 1876
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Regulation of mesangial cell apoptosis and proliferation by intracellular Ca2+ signals

Kidney International, Vol. 58 (2000), pp. 1876–1884
HORMONES – CYTOKINES – SIGNALING
Regulation of mesangial cell apoptosis and proliferation byintracellular Ca21 signals
HILMI SALEH, EBERHARD SCHLATTER, DETLEF LANG, HANS-GERD PAUELS,and STEFAN HEIDENREICH
Department of Medicine, University of Munster, Munster, Germany
abolished thapsigargin-dependent, but not angiotensin II- orRegulation of mesangial cell apoptosis and proliferation byATP-induced Ca21 rises when administered during a 12-hourintracellular Ca21 signals.preincubation.Background. In inflammatory glomerular diseases, prolifer-
Conclusions. Our data suggest that a sustained increase ofation, as well as apoptosis of mesangial cells (MCs), has been[Ca21]i may serve as a signal to trigger MC apoptosis. Growthshown histomorphologically. Both processes may regulate thefactors such as PDGF can abolish apoptosis induced by eleva-cellular content of the mesangium by closely influencing eachtions of [Ca21]i by altering intracellular Ca21 signaling.other. In the present study, we examined whether the cyto-
plasmic free Ca21 concentration [Ca21]i is involved as a keysecond messenger in the regulation of proliferative and apo-ptotic events.
In many forms of inflammatory or metabolic glomeru-Methods. Thapsigargin, an inhibitor of the endoplasmiclar diseases, mesangial cells (MCs) can change their phe-Ca21-Mg21-ATPase, was used as a test substance to investigate
the role of [Ca21]i in signaling MC apoptosis and growth in notype and become proliferative and matrix expanding.vitro. Apoptosis was determined by nuclear chromatin staining Paracrine or autocrine growth factors such as platelet-with Hoechst 33258, by a [3H]-thymidine–based DNA fragmen- derived growth factor (PDGF) or insulin-like growthtation assay or by flow cytometry detecting binding of FITC-
factor-1 [1, 2], vasoactive agonists, for example, angioten-conjugated annexin V. Proliferation was measured by [3H]-sin II and endothelin [3, 4], or leukocyte-derived cyto-thymidine incorporation into acid-precipitable material and
corroborated by cell counting. kines [5] have been shown to play a key role for theResults. Thapsigargin significantly induced apoptosis and in- induction of proliferation and remodeling of the mesan-
hibited proliferation dose dependently in nanomolar concen- gium [6]. Apoptosis, or programmed cell death, is atrations without evoking necrotic damage when administered
physiological process that is able to reduce surplus cellnot longer than 12 hours. Significant apoptosis was measurablenumbers during tissue development, differentiation, ma-after a six-hour treatment of MCs with thapsigargin. Determina-
tion of [Ca21]i by fura-2–dependent spectrofluorometry showed lignant transformation, or inflammation [7–9]. It is char-that thapsigargin was able to induce prolonged [Ca21]i rises acterized by a fixed genetic program that is mostly trig-that could be prevented by preincubation with the intracellular gered or modified by signals from the environment. InCa21 chelator 1,2-bis(2-aminophenoxy)-ethane-N,N,N9, N9-tetra-
glomerular diseases, apoptosis has been shown to occuracetic acid (BAPTA) acetomethyl ester (AM). BAPTA hadin patients with proliferative glomerulonephritis [10] orno influence on MC viability but reversed thapsigargin-induced
apoptosis to control levels. After thapsigargin treatment (100 in the experimental model of Thy 1.1 nephritis, wherenmol/L, 12 hours), apoptotic MCs had a significantly higher glomerular hypercellularity is counterbalanced and re-[Ca21]i of 251 6 25 nmol/L (N 5 41) as compared with MCs solved by apoptotic processes of MCs [11]. However,that were not or not yet apoptotic ([Ca21]i of 116 6 20 nmol/L,
the mechanisms of MC apoptosis induction in vivo or inN 5 26, P , 0,05). Platelet-derived growth factor (PDGF), avitro and the involved second messenger pathways arewell-characterized growth factor for MCs, reversed the effects
of thapsigargin on proliferation and apoptosis in a similar fashion hardly understood. Previous studies have described thatas BAPTA. PDGF acutely stimulated increases of [Ca21]i but reactive oxidative intermediates, such as nitric oxide [12]
or superoxide [13], or antibodies ligating the Fas receptor[14] or the Thy 1 epitope [15] can evoke programmedKey words: cell death, platelet-derived growth factor, thapsigargin,cell death of MCs. The concept that the cytosolic-freeinflammatory glomerular disease, necrotic damage, metabolic glomeru-
lar disease. calcium concentration [Ca21]i may act as a key signal forapoptosis induction was first described for thymocytesReceived for publication February 15, 1999and lymphocytes, because a variety of proteases andand in revised form May 12, 2000
Accepted for publication May 16, 2000 endonucleases that are generally engaged as effectormolecules in the apoptotic cascade are activated by rises 2000 by the International Society of Nephrology
1876

Saleh et al: MC apoptosis and Ca21 signaling 1877
in Ca21 [16, 17]. However, for other cell types such asneutrophilic granulocytes, elevations of [Ca21]i are fol-lowed by a rescue from apoptosis [18]. Thus, in the pres-ent study, the role of a rise of [Ca21]i as an activationsignal of MC apoptosis has been investigated. The selec-tive endoplasmic Ca21-ATPase inhibitor thapsigarginwas used as a test substance that induces a release ofCa21 from cytosolic stores and evokes Ca21 influx acrossthe plasma membrane [19, 20]. Hence, it mimics theaction of various vasoactive agonists or hormones on[Ca21]i. Furthermore, the effects of the growth factorPDGF on Ca21-dependent MC apoptosis and alterationsof [Ca21]i were studied.
METHODS
Reagents
Thapsigargin, angiotensin II, adenosine 59-triphos-phate (ATP), and 1,2-bis(2-aminophenoxy)-ethane-N,N,N9,N9-tetra-acetic acid-acetomethyl ester (BAPTA-AM) were obtained from Calbiochem (Bad Soden, Ger-many), PDGF-BB from Boehringer Mannheim (Mann-heim, Germany), [3H]-thymidine (specific activity 5.0 Ci/mmol) from Amersham-Buchler (Braunschweig, Ger-many), and FITC-labeled annexin V from BoehringerIngelheim Bioproducts (Heidelberg, Germany). Fura-2AM and all other reagents were purchased from Sigma(Deisenhofen, Germany).
Animals and culture of glomerular mesangial cellsFig. 1. Dose-dependent effects of thapsigargin on mesangial cell (MC)
Mesangial cells, derived from 6- to 10-week-old male apoptosis (A) and proliferation (B). MCs were synchronized and limitedin growth by reducing serum concentration of the culture mediumSD rats, were isolated, cultured, and characterized as(CM), as described in the Methods section, before being treated withdescribed previously [21]. MC culture medium (CM) thapsigargin for 12 hours. Apoptosis was determined by a [3H]-thymi-
consisted of RPMI 1640 supplemented with 1 mmol/L dine–based DNA fragmentation assay. Means 6 SEM are given takingcontrol as 100%. At least six independent experiments were done, withl-glutamine, 50 U/mL penicillin, and 50 mg/mL strepto-the total number of cultures indicated within the graph bars. *P , 0.05mycin (Seromed, Berlin, Germany). For the first two compared with control.
passages, CM also contained 5 mg/mL bovine insulin, 5mg/mL transferrin, and 5 ng/mL selenite. CM was sup-
cubated in CM with 0.5% FCS for 12 hours in order toplemented with 0.5 to 10% fetal calf serum (FCS). Forenable processing of apoptosis. CM was removed care-experiments, MCs between the third and fifth passagesfully and MCs solubilized in 1 mL hypotonic lysis bufferwere exclusively used.[0.5% Triton X-100, 5 mmol/L Tris, 20 mmol/L ethylene-
Quantitation of apoptosis by DNA diaminetetraacetic acid (EDTA), pH 8] for 30 minutesfragmentation assay on ice. Cell lysates were centrifuged at 13,000 3 g for
20 minutes to separate intact chromatin from fragmentedQuantitation of DNA fragmentation was performedDNA. Supernatants containing cleaved DNA and theafter labeling MC with [3H]-thymidine according to apellets that were dissolved in 0.5 N NaOH were sepa-method described by Higuchi and Aggarwal [22].rately transferred for scintillation counting. For the totalMesangial cells subcultured in 24-well dishes at a den-counts, MCs were lyzed by the addition of 20 mL of 20%sity of 3 3 104 cells/mL in CM supplemented with 10%sodium dodecyl sulfate (SDS) without performing high-FCS were synchronized and slowed down in growth byspeed centrifugation. The percentage of cleaved DNAreducing serum in CM to 5% FCS at day 2, to 2.5% FCSwas calculated according to the following formula:at day 3, and to 0.5% at day 4. Cells were then incubated
with 3 mCi/mL [3H]-thymidine for 20 hours, washed ex- DNA fragmentationtensively, and treated with the test substances for 1 to20 hours. Afterward, MCs were again washed and postin- 5 cpm of test sample (supernatant) 3 100/total cpm

Saleh et al: MC apoptosis and Ca21 signaling1878
Table 1. Determination of cell counts, apoptosis and necrosis of mesangial cells
Apoptocic cells Necrotic cells(Hoechst 332581) (propidium iodide1)
Designation/ Intact cell countstreatment of cells N 3 104/mL %
Control6 hours 4.360.3 262 ,112 hours 4.460.3 362 ,120 hours 4.160.4 564 ,1
Thapsigargin 100 nmol/L6 hours 4.060.1a 762a ,112 hours 3.660.3a 1664a 46320 hours 2.260.5a 3067a 1466a
BAPTA 30 minutes→ Thapsigargin 12 hours 4.360.4b 563b 362PDGF 1.25 nmol/L 12 hours 8.961.0a ,1 ,1
Data are means 6 SEM. Intact mesangial cell counts were determined by counting cells microscopically. Quantitation of apoptosis has been performed byfluorescence microscopy using Hoechst 33258 staining, or necrosis by propidium iodide staining detected by flow cytometry. Three independent cultures have beenanalyzed.
aP , 0.05 as compared with control for the indicated time pointsbP , 0.05 as compared with 12-hour thapsigargin treatment without BAPTA preincubation
Baseline DNA fragmentation was scattered between 80% ethanol for 5 minutes. The nuclear dye Hoechst33258 was added at a concentration of 5 mmol/mL for2 and 15% when MCs were cultured in medium alone.
The percentage of increases or decreases of apoptosis 20 minutes at 48C. Thereafter, slides were rinsed threetimes in PBS and embedded in 5% glycerol for analysisby the tested substances in relationship to basal apoptosis
(% of control) are depicted in the figures. by immunofluorescence microscopy. The percentage ofapoptotic cells was determined by counting nuclei withCell necrosis was determined by trypan blue exclusion
[10% trypan blue in phosphate-buffered saline (PBS)] condensed or fragmented chromatin after Hoechst 33258staining from 100 MCs for each experiment.or by low propidium iodide (PI) staining using flow cy-
tometry (discussed later in this article).Proliferation assay
Detection of apoptosis by annexin V binding Proliferation of subconfluent MCs was quantitated bydetermination of the incorporation of [3H]-thymidine.Mesangial cell binding of FITC-labeled annexin V in-
dicating early stages of apoptosis was measured by flow MCs (4 3 104/mL) that were growth limited as describedpreviously were washed twice with PBS and treated withcytometry. MCs cultured and treated as outlined pre-
viously in this article were trypsinized from culture flasks, test substances in CM with 0.5% FCS for 12 hours. Forthe last six hours of the incubation period, MCs werewashed extensively in ice-cold PBS, and incubated in
PBS containing 2% bovine serum albumin (BSA) and incubated with 3 mCi/mL [3H]-thymidine, and werewashed and solubilized in 1 mL 0.1% SDS. Precipitation0.01% NaN3 on ice to block unspecific binding of fluoro-
chromes. Unfixed cells were then labeled with annexin on ice was performed with 200 mL of 20% trichloroaceticacid (TCA) overnight. Acid-precipitable material wasV-FITC (2.5 mg/mL in PBS containing 1.8 mmol/L CaCl2)
and PI (5 mg/mL) for 15 minutes on ice before washing pelleted by centrifugation, dissolved in 0.5 N NaOH, andtransferred for liquid scintillation counting. Further-again in saline. MCs that exhibited low staining of nu-
clear chromatin by PI and high binding of annexin V more, MC growth was analyzed by counting cells micro-scopically.were identified as apoptotic cells; high annexin V binding
and PI staining indicated necrotic damage. Cells wereMicrospectrofluorometric measurement of [Ca21]ianalyzed on a FACScan flow cytometer (Becton Dickin-
son, Mountain View, CA, USA) for a total of 10,000 Mesangial cells (5 3 103/mL) were cultured on 35 mmglass cover slips and were slowed down in growth asevents.described previously in this article. MCs were incubated
Visualization of chromatin fragmentation with 1 mmol/L of the [Ca21]i-sensitive dye fura-2 AMdissolved with surfactant Pluronic F-127 (0.1 g/L) for 30Mesangial cells (2 3 103/mL) were seeded out to four-
well glass chamber slides (Nunc, Wiesbaden, Germany) minutes in Ham’s F-12 medium (GIBCO BRL, Eg-genstein, Germany) at 378C as described previously [23].and synchronized as described previously in this article.
After treatment with the test substances, cells were fixed The measurements were performed in a constantly per-fused (10 mL/min) chamber, where the cover slips werein 4% paraformaldehyde in PBS for 10 minutes on ice,
rinsed twice in PBS, and permeabilized with ice cold fixed. MCs were perfused with buffer containing 110

Saleh et al: MC apoptosis and Ca21 signaling 1879
Fig. 3. Effect of thapsigargin on [Ca21]i of untreated MC (2BAPTA)or MC treated with BAPTA-AM (1BAPTA, 10 mmol/L) for 30 minutesand effect of external Ca21 removal on thapsigargin-induced [Ca21]i.Representative tracings of fura-2 fluorescence ratios out of six indepen-Fig. 2. Representative immunofluorescence micrographs of mesangialdent experiments are given. External Ca21 depletion was achieved bycells (MCs) treated without (A) or with thapsigargin (B, 100 nmol/Lsuperfusing cells in Ca21-free solution supplemented with 5 mmol/Lfor 12 h). Nuclear staining was performed with the Hoechst 33258 dye,EGTA.which gave a homogeneous chromatin architecture for control cells but
intensive nuclear condensation and fragmentation after thapsigargintreatment. Representative images out of six experiments are shown. Fig. 4. Induction of apoptosis by thapsigargin (TG; 100 nmol/L, 12 h)
of MCs pretreated without or with BAPTA-AM (10 mmol/L, 30 min).BAPTA-AM alone did not alter basal apoptosis significantly, but re-versed thapsigargin-induced apoptosis to control levels. Means 6 SEMare given with the numbers of cultures indicated within the graph bars.
mmol/L NaCl, 25 mmol/L NaHCO3, 10 mmol/L HEPES, *P , 0.05 as compared with control; #P , 0.05 as compared with TG.3.6 mmol/L KCl, 5 mmol/L glucose, 1 mmol/L MgCl2,and 1.3 mmol/L Ca-gluconate for 20 minutes at 378Cbefore the experiments were started. MCs were exited
gisches Institut, Universitat Freiburg, Freiburg, Ger-at wavelengths of 340, 360, and 380 nm with a xenon-many). At the end of each experiment, calibration ofquartz lamp (XBO 75 W; Zeiss, Jena, Germany) using[Ca21]i was performed by incubation of cells with iono-a filter wheel rotating at 10 Hz. Fluorescence was mea-mycin (1 mmol/L) in the presence (1.3 mmol/L) and ab-sured by a photon-counting tube (Hamamatsu H 3460-sence of extracellular Ca21 [buffered with 5 mmol/L eg-04, Herrsching, Germany) at 500 to 530 nm. Ten datatazic acid (EGTA)]. Maximum and minimum values ofpoints were averaged, resulting in a time resolution ofthe 340/380 nm excitation fluorescence ratio were used1 Hz. Fluorescence signals were taken from five MCsfor all calibrations to estimate the [Ca21]i.using an adjustable diaphragm. Autofluorescence was
To determine whether thapsigargin-induced MC apo-measured before loading cells with fura-2 and subtractedptosis was a direct consequence of [Ca21]i elevation, MCsfrom the original data for each experiment. The experi-stained by annexin V-FITC (2.5 mg/mL for 15 minutes)ments were analyzed and controlled with a PC-486 com-
puter system and specific software (U. Frobe, Physiolo- and loaded with fura-2 AM were aimed visually by spec-

Saleh et al: MC apoptosis and Ca21 signaling1880
Fig. 5. Induction of MC apoptosis by thapsigargin (100 nmol/L, 12 hours) and reversal of apoptosis by pretreatment of cells with BAPTA-AM(BAPTA, 10 mmol/L, 30 minutes) indicated by flow cytometry. In this multiparameter flow cytometry, low nuclear staining by PI excludedsignificant necrotic cell damage (#8%). Apoptosis was determined by binding of FITC-labeled annexin-V. Percentages of the cells within the fourquadrants are depicted with the number in the lower right quadrant indicating apoptotic cells. A representative experiment out of three is shown.
trofluorescence microscopy for [Ca21]i measurements. creased apoptosis by 138 6 30% above medium control.Time kinetic studies revealed that a significant increaseAnnexin V-FITC–positive apoptotic cells and annexin
V-FITC–negative intact cells were analyzed separately of MC apoptosis was demonstrable at minimum incuba-tion times of six hours (Table 1). At incubation timesfor [Ca21]i.above 12 hours, necrosis rates above 10% were found
Statistics (12 6 3% necrotic MC after 20 hours of treatment withthapsigargin; Table 1).Data are presented as mean values 6 SEM. The num-
ber of experiments (N) is depicted within the figure bars. In the following experiments, thapsigargin was admin-istered at a concentration of 100 nmol/L for 12 hours ifOne-way analysis of variance (ANOVA) test (Microcal
Origin 5.0 Software, Northhampton, MA, USA) was not otherwise indicated. Furthermore, thapsigargin sig-nificantly reduced MC proliferation at concentrationsused for statistical analysis. P values of less than 0.05
were considered significant. For statistics and depiction and incubation times that were chosen for apoptosisstudies (Fig. 1B). Experiments with flow cytometry de-of data in the figures, changes of the level of apoptosis
or proliferation of MC by the agonists are expressed tecting phosphatidylserine exposure of apoptotic cellsby annexin V staining showed that under control condi-in percentage taking control medium as 100% because
baseline apoptosis scattered between 2 and 15%. tions, 3 6 2% of MC bound annexin V, whereas aftertreatment with thapsigargin (100 nmol/L for 12 hours),20 6 4% of cells exerted an increased annexin V staining,
RESULTSindicative for apoptosis (N 5 12, P , 0.05). Representa-
Effects of thapsigargin on MC apoptosis tive flow cytometry tracings after labeling MCs withand proliferation annexin V-FITC for apoptosis detection and PI for ne-
crosis detection are given in Figure 5 (upper panel).As shown in Figure 1A, the Ca21-ATPase inhibitorthapsigargin induced apoptosis in nanomolar concentra- Staining MCs with the nuclear dye Hoechst 33258 as
a further method to confirm apoptosis in later stagestions dose dependently when MCs were treated for 12hours. A thapsigargin concentration of 100 nmol/L in- revealed significant chromatin condensation and frag-
Saleh et al: MC apoptosis and Ca21 signaling 1881
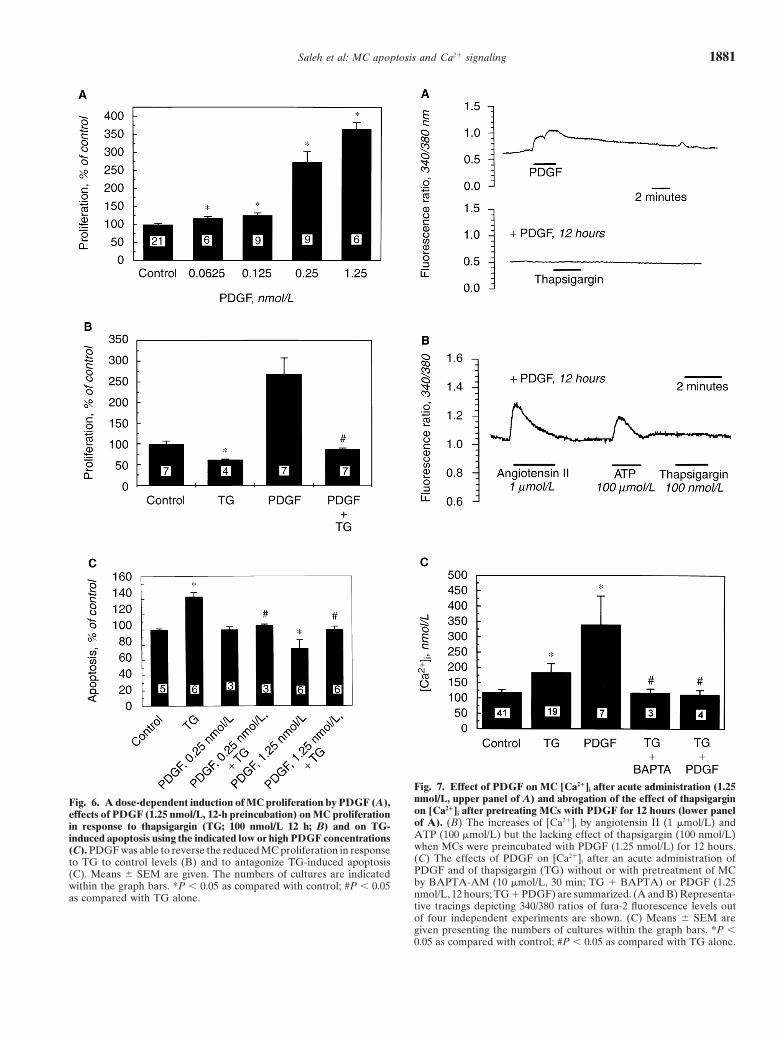
Fig. 7. Effect of PDGF on MC [Ca21]i after acute administration (1.25nmol/L, upper panel of A) and abrogation of the effect of thapsigarginFig. 6. A dose-dependent induction of MC proliferation by PDGF (A),on [Ca21]i after pretreating MCs with PDGF for 12 hours (lower paneleffects of PDGF (1.25 nmol/L, 12-h preincubation) on MC proliferationof A). (B) The increases of [Ca21]i by angiotensin II (1 mmol/L) andin response to thapsigargin (TG; 100 nmol/L 12 h; B) and on TG-ATP (100 mmol/L) but the lacking effect of thapsigargin (100 nmol/L)induced apoptosis using the indicated low or high PDGF concentrationswhen MCs were preincubated with PDGF (1.25 nmol/L) for 12 hours.(C). PDGF was able to reverse the reduced MC proliferation in response(C) The effects of PDGF on [Ca21]i after an acute administration ofto TG to control levels (B) and to antagonize TG-induced apoptosisPDGF and of thapsigargin (TG) without or with pretreatment of MC(C). Means 6 SEM are given. The numbers of cultures are indicatedby BAPTA-AM (10 mmol/L, 30 min; TG 1 BAPTA) or PDGF (1.25within the graph bars. *P , 0.05 as compared with control; #P , 0.05nmol/L, 12 hours; TG 1 PDGF) are summarized. (A and B) Representa-as compared with TG alone.tive tracings depicting 340/380 ratios of fura-2 fluorescence levels outof four independent experiments are shown. (C) Means 6 SEM aregiven presenting the numbers of cultures within the graph bars. *P ,0.05 as compared with control; #P , 0.05 as compared with TG alone.

Saleh et al: MC apoptosis and Ca21 signaling1882
mentation only in MCs treated with thapsigargin for at consecutively exposing cells to thapsigargin (100 nmol/L)for further 12 hours normalized MC growth to basalleast six hours (Table 1). Figure 2 provides typical images
of intact or late apoptotic MCs treated for 12 hours with levels and reversed the reduction of proliferation in-duced by thapsigargin (Fig. 6B). At lower PDGF concen-100 nmol/L thapsigargin after Hoechst staining.trations of 0.25 nmol/L, this treatment had no influence
Effects of thapsigargin on [Ca21]i and abrogation of on basal MC apoptosis, but reversed the thapsigargin-apoptosis by Ca21 buffering induced increase of apoptosis back to baseline. At higher
PDGF concentrations (1.25 nmol/L), PDGF per se re-Figure 3 shows that thapsigargin (100 nmol/L) mark-edly elevated [Ca21]i of MCs as presented by the 340/ duced the basal MC DNA fragmentation rate (Fig. 6C).380 fluorescence ratio. This increase of [Ca21]i was com-
Effects of PDGF on [Ca21]i and thapsigargin-inducedpletely abrogated when MCs were preincubated with the[Ca21]i increasesintracellular Ca21-chelator BAPTA-AM (10 mmol/L) for
30 minutes. These data are summarized in Figure 7C. Treatment of MCs with PDGF led to an acute increaseof [Ca21]i, as shown by the 340/380 fluorescence ratios.Removing Ca21 from extracellular medium by addition
of EGTA (1 to 5 mmol/L) and culturing MCs in nomi- However, preincubation with PDGF for 12 hours andthen exposing cells to thapsigargin abolished the rise ofnally Ca21-free medium highly diminished the adherence
of MCs so that MC apoptosis was not measurable under [Ca21]i completely. Figure 7A gives original tracings ofrepresentative experiments. Figure 7C summarizes thethese conditions. However, detecting [Ca21]i of MCs
after removal of Ca21 in the presence of EGTA (5 acute effects of thapsigargin (100 nmol/L) and PDGF(1.25 nmol/L) on [Ca21]i of MCs and compares themmmol/L) gave thapsigargin-induced [Ca21]i elevations
that were similar as for cells left in the presence of Ca21 with the lack of effects of thapsigargin when MCs werepreincubated with BAPTA-AM for 30 minutes or PDGF(D[Ca21]i 121 6 34 nmol/L in Ca21-free solution, N 5 6,
vs. 78 6 36 nmol/L, N 5 19; lower panel of Fig. 3). for 12 hours. Interestingly, angiotensin II and ATP, bothApplication of BAPTA-AM for 30 minutes did not well-known agonists leading to [Ca21]i rises of MCs,
alter the baseline DNA fragmentation of MCs, but re- were, in contrast to thapsigargin, still able to evoke Ca21
versed the thapsigargin-induced increase of apoptosis peaks after a 12-hour preincubation with PDGF. A rep-back to control levels. This was shown by the [3H]-thymi- resentative tracing is shown in Figure 7B. Angiotensindine–based DNA fragmentation assay (Fig. 4), flow cy- II (1 mmol/L) elevated [Ca21]i from 143 6 18 to 396 6tometry detecting annexin V binding (Fig. 5), as well as 38 nmol/L (N 5 5) after preincubation of MCs withby Hoechst 33258 staining (Table 1). These data suggest PDGF (1.25 nmol/L), which was similar to MCs withoutthat thapsigargin evoked apoptosis of MCs by leading PDGF pretreatment (data not shown). Simultaneous ad-to an increase of [Ca21]i consecutive to a release from ministration of PDGF and thapsigargin to MCs inducedinternal cytoplasmic stores and inducing secondary Ca21- a significant rise of [Ca21]i without exerting additive ordependent mechanisms responsible for DNA degrada- potentiating effects (data not shown).tion.
DISCUSSION[Ca21]i of MCs treated with thapsigargin in intact andapoptotic cells The endoplasmic reticular Ca21-ATPase inhibitor
thapsigargin was used in the present study to investigateSince not all MCs treated with thapsigargin showedwhether an increase of [Ca21]i can serve as a signal tofeatures of apoptosis, we examined whether differencesinitiate MC apoptosis. Many previous studies haveof basal [Ca21]i were measurable between intact (annexinshown that a rise of [Ca21]i by thapsigargin involves anV negative) and apoptotic (annexin V positive) cells.initial release of the cation from microsomal, nonmito-Intact thapsigargin-treated MCs had a significantly lowerchondrial Ca21 pools, which induces and is followed by[Ca21]i of 116 6 20 nmol/L (N 5 26) as compared witha capacitative Ca21 influx from the extracellular spaceapoptotic thapsigargin-treated cells, which had a [Ca21]i
[19, 20]. Our results confirmed that thapsigargin elevatedof 251 6 25 nmol/L (N 5 41, P , 0.05).[Ca21]i of MCs also in nominally Ca21-free medium. In
Effects of PDGF on proliferation and apoptosis and contrast to Ca21 ionophores, which permeabilize cellcounter-regulation of the action of thapsigargin membranes and organelle membranes unspecifically,
thapsigargin is a suitable and specific test substance toThe well-characterized growth factor PDGF dose de-examine the significance of Ca21-signaling in apoptosispendently induced proliferation of MCs as determined[24, 25]. With respect to increasing [Ca21]i, it mimics theby thymidine incorporation at nanomolar concentrationsaction of various natural mediators such as vasopres-when cells were treated for 12 hours (Fig. 6A). Cellsor agonists (for example, endothelin, angiotensin II),counting corroborated MC growth by PDGF (Table 1).
Preincubation with PDGF (1.25 nmol/L) for 12 hours and growth factors (for example, PDGF, basic fibroblast

Saleh et al: MC apoptosis and Ca21 signaling 1883
growth factor) or cytokines (for example, interleukin-2) previous studies [31, 32]. In comparison to thapsigargin,[Ca21]i increases after PDGF exposure were similar with[19, 23, 26].
Our data show that treatment with thapsigargin re- respect to height and duration, so we can widely excludethat the pattern of Ca21 peaking determines whether asulted in apoptosis of MCs, which were in a synchronized
and growth-limited state. MCs kept in a proliferative proliferative or an apoptotic pathway will be initiated.PDGF induced MC proliferation and using higher con-state by serum supplementation (.1% FCS in medium)
were resistant to the apoptosis-inducing effects of thapsi- centrations (1.25 nmol/L) even suppressed basal apopto-sis. When MCs were pretreated with PDGF for 12 hoursgargin. We detected features of apoptosis by three differ-
ent techniques; this study shows that thapsigargin and were consequently exposed to thapsigargin, Ca21-triggered apoptosis disappeared. Most interestingly, pre-induced nuclear chromatin condensation and fragmen-
tation by Hoechst 33258 nuclear staining as well as treatment with the growth factor exclusively abolishedthapsigargin-induced elevations of [Ca21]i but not angio-plasma membrane alterations indicative for apoptosis
using annexin V-based flow cytometry. An increase of tensin II- or ATP-induced Ca21 fluxes. These data indi-cate that increases of [Ca21]i per se are not sufficient to[Ca21]i was an essential trigger to activate apoptotic pro-
cesses of MCs because intracytoplasmic chelation of Ca21 induce apoptosis or proliferation, although during bothcellular processes, Ca21 signaling occurs. Depletion ofby BAPTA-AM completely antagonized the effects of
thapsigargin and reversed apoptosis to basal levels. Fur- cytosolic Ca21 stores by growth factors could be excludedsince agonists were still able to induce [Ca21]i peaks afterthermore, MCs treated with thapsigargin showed sig-
nificantly higher [Ca21]i when they were identified as PDGF pretreatment.Growth factors induce their effects via cell membraneapoptotic cells by annexin V binding as compared with
MCs that were still intact and negative for annexin V receptors, which are coupled to protein tyrosin kinases.During the downstream events, phospholipase C-g andbinding. When MCs were treated with thapsigargin for
more than 12 hours, progression of cells into necrotic phosphatidylinositol 3 kinase are involved, which gener-ate rises of [Ca21]i, mobilization of transcription factors,stages took place, but the percentage of apoptotic MCs
never exceeded one third (Table 1). MCs that peaked and nuclear expression of protooncogenes [32–34]. Bythe activation of these cascades, stimulation of Ca21-with [Ca21]i were identified as senescent cells, but it is not
clear by which mechanisms the surviving cells quenched dependent endonucleases, phospholipase A, or prote-[Ca21]i elevations and were rescued from apoptosis. ases engaged in apoptotic pathways [35] must be blockedProbably, MCs that locally produce high levels of auto- by yet unknown mechanisms. Our results suggest thatcrine growth factors such as PDGF are protected against growth factors can alter Ca21 signaling and cell survivalapoptosis safely. depending on the duration of time they are present at
Thymocytes and lymphocytes are cells in which apo- sufficient concentrations.ptosis in response to a rise of [Ca21]i could be first demon- In conclusion, our study shows that Ca21 signals arestrated [27, 28]. This was similarly achieved by Ca21 involved in MC proliferation as well as in apoptosis andionophores, thapsigargin, and dexamethasone [29]. Since might serve as a regulatory link. Mediators such as PDGFall of these substances increased [Ca21]i, it was suggested support not only cell growth, but rescue MCs from apo-that Ca21 is a second messenger signal generally engaged ptosis as survival factors by altering Ca21 homeostasis.as a trigger of apoptosis [17, 20]. In nonlymphoid bloodcells such as neutrophils or monocytes, however, eleva- ACKNOWLEDGMENTtions of [Ca21]i retarded and antagonized apoptotic pro- This study was supported by the IZKF Universitat Munster, grantcesses [18, 30], so that [Ca21]i peaks are not able to B5, Munster, Germany.initiate apoptotic pathways per se. It is striking that inlymphocytes or lymphoid cell lines, all exerting a high REFERENCESproliferative capacity, increases of [Ca21]i trigger apopto- 1. Schulz PJ, DiCorleto PE, Silver BJ, Abboud HE: Mesangialsis, whereas in neutrophils or monocytes, cells that are cells express PDGF mRNA and proliferate in response to PDGF.
Am J Physiol 255:F674–F679, 1988terminally differentiated and not or hardly not able to2. Doi T, Striker LJ, Elliot SJ, Conti FG, Striker GE: Insulin-likeproliferate, a rescue from apoptosis takes place [17]. MCs
growth factor-1 is a progression factor for human mesangial cells.that divide in culture and also significantly proliferate in Am J Pathol 134:395–404, 1989
3. Wolthuis A, Boes A, Rodemann HP, Grond J: Vasoactive agentsvivo during different glomerular diseases fit into thisaffect growth and protein synthesis of cultured rat mesangial cells.scheme by initiating apoptosis after elevation of [Ca21]i. Kidney Int 41:124–131, 1992
This linkage between proliferation of cells and sensi- 4. Simonson MS, Wann S, Mene P, Dubyak GR, Kester M, Naka-zato Y, Sedor JR, Dunn MJ: Endothelin stimulates phospholipasetivity to Ca21 signal-dependent apoptosis prompted usC, Na1/H1 exchange, c-fos expression and mitogenesis in rat mes-to test the effects of the growth factor PDGF on thapsi-angial cells. J Clin Invest 83:708–712, 1989
gargin-induced MC apoptosis. PDGF acutely caused a 5. Sterzel RB, Schulze-Lohoff E, Marx M: Cytokines and mesan-gial cells. Kidney Int 43(Suppl 39):S26–S31, 1993marked increase of [Ca21]i as it has been shown in several

Saleh et al: MC apoptosis and Ca21 signaling1884
6. Floege J, Eng E, Young BA, Johnson RJ: Factors involved in TNF receptor in TNF-induced cytotoxicity, DNA fragmentation,and differentiation. J Immunol 152:4017–4025, 1994the regulation of mesangial cell proliferation in vitro and in vivo.
23. Schlatter E, Ankorina I, Haxelmans S, Kleta R: Effects ofKidney Int 43(Suppl 39):S47–S54, 1993diadenosine polyphosphates, ATP and angiotensin II on cytosolic7. Wyllie AH, Kerr JFR, Currie AR: Cell death: The significanceCa21-activity and contraction of rat mesangial cells. Pflugers Archof apoptosis. Int Rev Cytol 68:251–306, 1980430:721–728, 19958. Ellis RE, Yuan JY, Horvitz HR: Mechanisms and functions of
24. Tsukamoto A, Kaneko Y: Thapsigargin, a Ca21-ATPase inhibitor,cell death. Annu Rev Cell Biol 7:663–698, 1991depletes the intracellular Ca21 pool and induces apoptosis in human9. Cohen JJ: Apoptosis. Immunol Today 14:126–130, 1993hepatoma cells. Cell Biol Int 17:969–970, 199310. Harris DJ: Cell death in the diseased glomerulus. Histopathology
25. Li WW, Alexandre S, Cao X, Lee AS: Transactivation of the12:679–683, 1988grp78 promotor by Ca21 depletion: A comparative analysis with11. Baker AJ, Mooney A, Hughes J, Lombardi D, Johnson RJ, SavillA23187 and the endoplasmic reticulum Ca21-ATPase inhibitorJ: Mesangial cell apoptosis: The major mechanism for resolutionthapsigargin. J Biol Chem 268:12003–12009, 1993of glomerular hypercellularity in experimental mesangial prolifera-
26. Wong WL, Brostrom MA, Kuznetsov G, Gmitter-Yellen D,tive nephritis. J Clin Invest 94:2105–2116, 1994Brostrom CO: Inhibition of protein synthesis and early protein12. Muhl H, Sandau K, Brune B, Briner VA, Pfeilschifter J: Nitricprocessing by thapsigargin in cultured cells. Biochem J 289:71–79,oxide donors induce apoptosis in glomerular mesangial cells, epi-1993thelial cells and endothelial cells. Eur J Pharmacol 317:137–149,
27. Kaiser N, Edelman IS: Calcium dependence of glucocorticoid-1996induced lymphocytolysis. Proc Natl Acad Sci USA 74:638–642, 197713. Sandau K, Pfeilschifter J, Brune B: The balance between nitric
28. Wyllie AH: Glucocorticoid-induced thymocyte apoptosis is asso-oxide and superoxide determines apoptotic and necrotic death ofciated with endogenous endonuclease activation. Nature 284:555–rat mesangial cells. J Immunol 158:4938–4946, 1997556, 198014. Gonzales-Cuadrado S, Lopez-Armada MJ, Gomez-Guerrero 29. Kaiser N, Edelman IS: Further studies on the role of calcium inC, Subira D, Garcia-Sahuquillo A, Ortiz-Gonzalez A, Neilson glucocorticoid-induced lymphocytolysis. Endocrinology 103:936–EG, Egido J, Ortiz A: Anti-Fas antibodies induce cytolysis and 942, 1978
apoptosis in cultured human mesangial cells. Kidney Int 49:1064– 30. Macintyre EA, Roberts PJ, Jones M, Van Der Schoot CE, Fava-1070, 1996 laro EJ, Tidman N, Linch DC: Activation of human monocytes
15. Sato T, van Dixhoorn MAG, Schroeijers WEM, Huizinga TWJ, occurs on cross-linking monocytic antigens to an Fc receptor. JReutelingsperger CPM, van Es LA, Daha MR: Apoptosis of Immunol 142:2377–2383, 1989cultured rat glomerular mesangial cells induced by IgG2a mono- 31. Heidenreich S, Tepel M, Lang D, Rahn KH, Zidek W: Differen-clonal anti-Thy-1 antibodies. Kidney Int 49:403–412, 1996 tial effects of insulin-like growth factor 1 and platelet-derived
16. Martin SJ, Green DR: Protease activation during apoptosis: growth factor on growth response, matrix formation, and cytosolicDeath by a thousand cuts? Cell 82:349–352, 1995 free calcium of glomerular mesangial cells of spontaneously hyper-
17. McConkey DJ, Orrenius S: The role of calcium in the regulation tensive and normotensive rats. Nephron 68:481–488, 1994of apoptosis. J Leukoc Biol 59:775–783, 1996 32. Whiteside C, Munk S, Zhou X, Miralem T, Templeton DM:
18. Whyte MKB, Hardwick SJ, Meager LC, Savill JS, Haslett C: Chelation of intracellular calcium prevents mesangial cell prolifera-Transient elevations of cytosolic free calcium retard subsequent tive responsiveness. J Am Soc Nephrol 9:14–25, 1998apoptosis in neutrophils in vitro. J Clin Invest 92:446–455, 1993 33. Valius M, Kazlauskas A: Phospholipase C-g1 and phosphatidyl-
19. Clapham DE: Calcium signaling. Cell 273:259–268, 1995 inositol 3 kinase are the downstream mediators of the PDGF recep-20. Jiang S, Chow SC, Nicotera P, Orrenius S: Intracellular Ca21 tor’s mitogenic signal. Cell 73:321–334, 1993
signals activate apoptosis in thymocytes: Studies using the Ca21- 34. Timmerman LA, Clipstone NA, Ho SN, Northrop JP, CrabtreeATPase inhibitor thapsigargin. Exp Cell Res 212:84–92, 1994 GR: Rapid shuttling of NF-AT in discrimination of Ca21 signals
21. Heidenreich S, Tepel M, Schluter H, Harrach B, Zidek W: and immunosuppression. Nature 383:837–840, 1996Regulation of rat mesangial cell growth by diadenosine phosphates. 35. Nicotera P, Zhivotovsky B, Orrenius S: Nuclear calcium trans-J Clin Invest 95:2862–2867, 1995 port and the role of calcium in apoptosis. Cell Calcium 16:279–288,
199422. Higuchi M, Aggarwal BB: Differential roles of two types of the




![Docking of LDCVs Is Modulated by Lower Intracellular [Ca2+] than Priming](https://static.fdokumen.com/doc/165x107/6344ad896cfb3d40640952c4/docking-of-ldcvs-is-modulated-by-lower-intracellular-ca2-than-priming.jpg)






![Near-Membrane [Ca2+] Transients Resolved Using the Ca2+ Indicator FFP18](https://static.fdokumen.com/doc/165x107/631286873ed465f0570a4533/near-membrane-ca2-transients-resolved-using-the-ca2-indicator-ffp18.jpg)







