
Regulation of constitutive and UVR-induced skin pigmentation by melanocortin 1 receptor isoforms
16
The FASEB Journal • FJ Express Full-Length Article Regulation of constitutive and UVR-induced skin pigmentation by melanocortin 1 receptor isoforms Francois Rouzaud,* ,†,1 Gertrude-E. Costin,* Yuji Yamaguchi,* Julio C. Valencia,* Werner F. Berens,* Kevin G. Chen,* Toshihiko Hoashi,* Markus Bo ¨hm, § Zalfa A. Abdel-Malek, ¶ and Vincent J. Hearing* *Laboratory of Cell Biology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland, USA; † Department of Medicinal Chemistry, University of Florida, Gainesville, Florida, USA; § Department of Dermatology and Ludwig Boltzmann Institute for Cell Biology and Immunobiology of the Skin, University of Mu ¨ nster, Munster, Germany; and ¶ Department of Dermatology, University of Cincinnati College of Medicine, Cincinnati, Ohio, USA ABSTRACT Melanin synthesized by epidermal mela- nocytes protects the skin against UVR-induced DNA damage and skin cancer. Exposure to UVR increases the synthesis of the photoprotective eumelanin on activation of MC1R, a melanoma susceptibility gene. We studied the expression of MC1R under UVR and -MSH stimulation in skin of different ethnic origins and in melanocytes of various pigmentary levels. This study identifies and characterizes a novel MC1R iso- form (MC1R350) generated by alternative splicing of the classically known MC1R (MC1R317). We demon- strate that the melanin content of melanocytes shows a significant positive correlation with MC1R317 levels but correlates inversely with the amount of MC1R350, suggesting that this latter isoform could act as a nega- tive regulator of melanin synthesis. We confirmed that hypothesis by showing that while MC1R317 signaling significantly increases the expression of MITF and tyrosinase, two key factors in the melanin synthesis pathway, MC1R350 dramatically hampers their expres- sion. In the skin, we show that UVR does not increase MC1R350 expression but does significantly increase MC1R317. Taken together, our results strongly suggest that MC1R350 acts as a negative regulator of skin pigmentation and demonstrate for the first time that MC1R isoform-specific expression is closely related to skin pigmentation and photoprotection.—Rouzaud, F., Costin, G-E., Yamaguchi, Y., Valencia, J. C., Berens, W. F., Chen, K. G., Hoashi, T., Bo ¨hm, M., Abdel-Malek, Z. A., Hearing, V. J. Regulation of constitutive and UVR-induced skin pigmentation by melanocortin 1 receptor isoforms. FASEB J. 20, E1338 –E1350 (2006) Key Words: splice variant melanocyte UVR Variations in human hair and skin colors are caused by differences in the distribution and type of melanins that are produced in melanocytes. Mela- nins exist in two distinct types: the brown/black eumelanins and the yellow/red pheomelanins. Indi- viduals with light skin and red hair have a predomi- nance of pheomelanin in their skin and hair and/or an impaired ability to produce eumelanin. Although human pigmentation shows a wide variety of pheno- types and is thought to be under the control of several genetic loci (1), recent studies have shown that the red hair phenotype can be associated pre- dominantly with variants of a single gene— exten- sion—which encodes the melanocortin 1 receptor (MC1R) (2–3). MC1R was the first member of the melanocortin receptors to be cloned (4 –5). The membrane-anchored 317 amino acid protein has 7 transmembrane spanning domains and plays a criti- cal role in the determination of skin and/or hair color in mammals. Common MC1R alleles in North- ern European populations, particularly R151C, R160W, and D294H, are associated with red hair and sun sensitivity (2–3, 6). These mutants all bind -MSH but are unable to activate adenylate cyclase in cultured cells (7– 8). Though knowledge of the down- stream signaling of MC1R is still incomplete, activa- tion of the receptor by either -MSH or ACTH increases the intracellular cAMP concentration, which in turn regulates the activity of a range of transcription factors (such as microphthalmia, or MITF) and melanogenic enzymes (such as tyrosi- nase) that modulate the amounts of eumelanin and pheomelanin (9 –10). MC1R is also a member of the G-protein-coupled receptor (GPCR) family. GPCRs act as recognition elements for a vast range of hormones, transmitters, and modulators (11). They have therefore been widely studied and are considered the most tractable class of proteins for drug design in the pharmaceutical industry (12). Many of the key early studies of GPCR have been reviewed recently (13) and include data from ligand binding studies, target size irradiation, and antibody (Ab) -induced receptor activation. However, in most 1 Correspondence: University of Florida, Department of Medicinal Chemistry, Rm. P5–26, 1600 SW Archer Rd., Gainesville, FL 32610, USA. E-mail: frouzaud@ufl.edu doi: 10.1096/fj.06-5922fje E1338 0892-6638/06/0020-1338 © FASEB
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Regulation of constitutive and UVR-induced skin pigmentation by melanocortin 1 receptor isoforms

The FASEB Journal • FJ Express Full-Length Article
Regulation of constitutive and UVR-induced skinpigmentation by melanocortin 1 receptor isoforms
Francois Rouzaud,*,†,1 Gertrude-E. Costin,* Yuji Yamaguchi,* Julio C. Valencia,*Werner F. Berens,* Kevin G. Chen,* Toshihiko Hoashi,* Markus Bohm,§
Zalfa A. Abdel-Malek,¶ and Vincent J. Hearing**Laboratory of Cell Biology, National Cancer Institute, National Institutes of Health, Bethesda,Maryland, USA; †Department of Medicinal Chemistry, University of Florida, Gainesville, Florida,USA; §Department of Dermatology and Ludwig Boltzmann Institute for Cell Biology andImmunobiology of the Skin, University of Munster, Munster, Germany; and ¶Department ofDermatology, University of Cincinnati College of Medicine, Cincinnati, Ohio, USA
ABSTRACT Melanin synthesized by epidermal mela-nocytes protects the skin against UVR-induced DNAdamage and skin cancer. Exposure to UVR increasesthe synthesis of the photoprotective eumelanin onactivation of MC1R, a melanoma susceptibility gene.We studied the expression of MC1R under UVR and�-MSH stimulation in skin of different ethnic originsand in melanocytes of various pigmentary levels. Thisstudy identifies and characterizes a novel MC1R iso-form (MC1R350) generated by alternative splicing ofthe classically known MC1R (MC1R317). We demon-strate that the melanin content of melanocytes shows asignificant positive correlation with MC1R317 levels butcorrelates inversely with the amount of MC1R350,suggesting that this latter isoform could act as a nega-tive regulator of melanin synthesis. We confirmed thathypothesis by showing that while MC1R317 signalingsignificantly increases the expression of MITF andtyrosinase, two key factors in the melanin synthesispathway, MC1R350 dramatically hampers their expres-sion. In the skin, we show that UVR does not increaseMC1R350 expression but does significantly increaseMC1R317. Taken together, our results strongly suggestthat MC1R350 acts as a negative regulator of skinpigmentation and demonstrate for the first time thatMC1R isoform-specific expression is closely related toskin pigmentation and photoprotection.—Rouzaud, F.,Costin, G-E., Yamaguchi, Y., Valencia, J. C., Berens,W. F., Chen, K. G., Hoashi, T., Bohm, M., Abdel-Malek,Z. A., Hearing, V. J. Regulation of constitutive andUVR-induced skin pigmentation by melanocortin 1receptor isoforms. FASEB J. 20, E1338–E1350 (2006)
Key Words: splice variant � melanocyte � UVR
Variations in human hair and skin colors arecaused by differences in the distribution and type ofmelanins that are produced in melanocytes. Mela-nins exist in two distinct types: the brown/blackeumelanins and the yellow/red pheomelanins. Indi-viduals with light skin and red hair have a predomi-
nance of pheomelanin in their skin and hair and/oran impaired ability to produce eumelanin. Althoughhuman pigmentation shows a wide variety of pheno-types and is thought to be under the control ofseveral genetic loci (1), recent studies have shownthat the red hair phenotype can be associated pre-dominantly with variants of a single gene— exten-sion—which encodes the melanocortin 1 receptor(MC1R) (2–3). MC1R was the first member of themelanocortin receptors to be cloned (4 –5). Themembrane-anchored 317 amino acid protein has 7transmembrane spanning domains and plays a criti-cal role in the determination of skin and/or haircolor in mammals. Common MC1R alleles in North-ern European populations, particularly R151C,R160W, and D294H, are associated with red hair andsun sensitivity (2–3, 6). These mutants all bind�-MSH but are unable to activate adenylate cyclase incultured cells (7– 8). Though knowledge of the down-stream signaling of MC1R is still incomplete, activa-tion of the receptor by either �-MSH or ACTHincreases the intracellular cAMP concentration,which in turn regulates the activity of a range oftranscription factors (such as microphthalmia, orMITF) and melanogenic enzymes (such as tyrosi-nase) that modulate the amounts of eumelanin andpheomelanin (9 –10).
MC1R is also a member of the G-protein-coupledreceptor (GPCR) family. GPCRs act as recognitionelements for a vast range of hormones, transmitters,and modulators (11). They have therefore been widelystudied and are considered the most tractable class ofproteins for drug design in the pharmaceutical industry(12). Many of the key early studies of GPCR have beenreviewed recently (13) and include data from ligandbinding studies, target size irradiation, and antibody(Ab) -induced receptor activation. However, in most
1 Correspondence: University of Florida, Department ofMedicinal Chemistry, Rm. P5–26, 1600 SW Archer Rd.,Gainesville, FL 32610, USA. E-mail: [email protected]
doi: 10.1096/fj.06-5922fje
E1338 0892-6638/06/0020-1338 © FASEB

cells and tissues, individual GPCRs are expressed at lowlevels; this is especially the case for human MC1R (14).Until recently, virtually every picture of GPCR structureand function had depicted the protein as a monomer.However, a growing number of biochemical, biophysi-cal, and functional studies suggest that GPCRs formfunctional, SDS-stable dimers (15). It was recentlyindicated that melanocortin receptors can formdimeric structures (16–18).
UV radiation (UVR) is among the most ubiquitousagents in the environment, and humans are inevitablyexposed to it. UVR not only initiates and promotes thetransformation of normal epidermal cells to cancercells via dysregulation of intracellular signaling path-ways and via its mutagenic effects on DNA, but also byaltering host immunity by reducing its capability forsurveillance against tumor or viral antigens (19–20).The significance of cutaneous pigmentation lies in thephotoprotective effect of melanin, particularly eumela-nin, against sun-induced carcinogenesis. Epidermalmelanocytes and keratinocytes respond to UVR byincreasing their expression of �-MSH and ACTH,which up-regulate the expression of MC1R and mediatethe pigmentary response to UVR, as proposed in stud-ies using mouse melanoma cells (21).
Constitutive skin pigmentation dramatically affectsthe incidence of skin cancer, and the photoprotec-tive function of eumelanin in the skin is highlysignificant (22–23). In the U.S., rates of basal andsquamous cell carcinomas are 50-fold higher in Cau-casians than in African Americans (24 –26), andAfrican Americans have a 13-fold lower incidence ofmelanoma than do Caucasians. Pigmentation pat-terns in the skin are determined not only by thelocation of melanocytes but also by the type andamount of melanins they produce. It has been shownthat melanin affords significant protection againstDNA damage in underlying skin cells (23), as it canabsorb UV very efficiently at most wavelengths. Levelsof melanin are �4-fold higher in African Americanskin than in Caucasian skin, and levels of DNAdamage were �7- to 8-fold lower in African Americanskin than in Caucasian skin (23).
Since epidermal melanocytes respond to UVR byincreasing their expression of �-MSH, which up-regu-lates the expression of MC1R (27–28) and conse-quently enhances the response of melanocytes to mela-nocortins, we studied the expression of MC1R in skin ofdifferent ethnic origins and in primary normal humanmelanocytes (NHM) displaying a large range of pig-mentation phenotypes. We characterize for the firsttime MC1R350, a new human MC1R isoform differentfrom the one described by Tan et al. (29), whichcontains 350 amino acids and is generated by alterna-tive splicing of the more commonly known MC1R, herenamed MC1R317.
Its implication in human skin pigmentation is inves-tigated with respect to its influence on melanin produc-tion as well as tyrosinase and MITF expression.
MATERIALS AND METHODS
Cell culture
Primary NHM cultures established from neonatal foreskinswere maintained at 37°C with 5% CO2 in Medium 154(Cascade Biologics, Portland, OR, USA) supplemented withhuman melanocyte growth supplement containing bovinepituitary extract, FBS, bovine insulin, bovine transferrin,fibroblast growth factor, hydrocortisone, heparin, phorbol12-myristate 13-acetate (TPA) (Cascade Biologics), and 50ng/ml of gentamicin/amphothericin B (Biowhittaker, Walk-ersville, MD, USA). Donors were anonymous and no ethnicbackground information was available. We classified themaccording to visual pigmentation of the skin and to melanincontent, which correlates well with the extent of pigmenta-tion of the skin and derived melanocytes.
Cell growth assessment
Ten thousand cells were seeded in triplicate into 35 mmtissue culture dishes (BD Biosciences, San Jose, CA, USA) andharvested by trypsinization at several time points. Cells werecounted with a Coulter Particle Counter Z1 (BeckmanCoulter, Inc., Fullerton, CA, USA) and the doubling time wasdetermined.
Melanin content measurement
The melanin content was measured using a modification of apreviously reported method (30). Briefly, melanocytes werecultured until they became confluent. They were solubilizedin lysis buffer (1% Nonidet P-40; Calbiochem, San Diego, CA,USA; in PBS containing a protease inhibitor cocktail; Roche,Indianapolis, IN, USA) for 1 h on ice with occasional vortex-ing, and protein concentrations were measured with a bicin-coninic acid kit (Pierce, Rockford, IL, USA). Melanin pelletswere dissolved by incubation in 1 N NaOH at 37°C for 18 h.Aliquots of each sample were transferred to 96-well plates,quantitated by absorbance at 405 nm using an automaticmicroplate reader (Molecular Devices, Sunnyvale, CA, USA),and calibrated against a standard curve generated usingsynthetic melanin (Sigma, St. Louis, MO, USA).
Transient transfection of melanocytes
The Amaxa Nucleofectant system was used to transientlytransfect NHM according to the manufacturer’s instructions(Amaxa, Cologne, Germany). Two million cells were usedand seeded in 100 mm dishes directly after transfection.Assessment of transfection efficiency was performed by mea-suring the amount of plasmids present in the cells by semi-quantitative real-time RT-polymerase chain reaction (RT-PCR).
Electrophoresis and Western blot
NHM were harvested and solubilized in the lysis bufferdescribed above for 1 h on ice with occasional vortexing. Thesamples were centrifuged at 14,000 g for 30 min at 4°C andthe supernatants were recovered and kept at –70°C forfurther experiments. Protein concentrations of the sampleswere measured as noted above. For Western blot, sampleswere separated by electrophoresis under reducing conditions.Lysates were mixed 1:2 (v/v) with sample buffer (Bio-RadLaboratories, Hercules, CA, USA) containing 5% 2-mercap-toethanol and boiled for 10 min. Samples were separated by
E1339IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM

SDS/PAGE with 18% Tris-glycine gels (Invitrogen, Carlsbad,CA, USA) and transferred to polyvinylidene difluoride mem-branes (Millipore Corp., Bedford, MA, USA). Blots wereblocked in 5% nonfat milk in PBS for 1 h at room tempera-ture, then incubated with �PEP19 antiserum (1:300) toMC1R in 1% milk in PBS for 1 h at room temperature. The�PEP19 Ab was raised in our laboratory against residues266–279 of the murine Mc1r, which is situated in the thirdextracellular loop of the receptor (31). An anti-rabbit horse-radish peroxidase-linked whole Ab (Amersham Pharmacia,Piscataway, NJ, USA) was used as secondary Ab (dilution1:1,000). The immunoreactivity of the blots was detected byenhanced chemiluminescence Western blot detection kit(Amersham Pharmacia), according to the manufacturer’sinstructions.
[125I]�-MSH binding assays
Melanocytes were incubated for 2 h in the presence of 10 �Ci[125I](Lys11)(Nle4-D-Phe7)�-MSH (Sigma) for each 10 mmdish. Total proteins were extracted with the lysis bufferdescribed above and 40 �g of the total protein extract wasseparated by electrophoresis on Bis-Tris 4–12% gels (Invitro-gen) under native or reducing conditions. The dried gelswere exposed for 22 days in a storage phosphor screen, thenscanned with a STORM PhosphorImager (Molecular Dynam-ics, Sunnyvale, CA, USA) and analyzed with the Image Quantprogram.
5� and 3�-Rapid amplification of cDNA end (RACE)
Total cytoplasmic RNA was isolated from cultured melano-cytes using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA)according to the manufacturer’s recommendations. RACEreactions were performed with 200 ng RNA template usingthe SMART RACE cDNA amplification kit (Clontech, PaloAlto, CA). The gene-specific primers used for 5� RACE were1) 5�attctccaccagactcaccagccctagg3� and 2) 5�ggacacatacag-gcaccaaggctctgac3�, and for 3� RACE 3) 5�cagcaccccacctg-cagctgcatgttc3�, and 4) 5�agaacttcaacctcttcctcctcctc3�. Nestedpolymerase chain reaction (PCR) reactions were carried outin a 50 �l final volume with 50 ng cDNA template for the firstreaction from which 5 �l were used as a template for thesecond one, 0.5 mM dNTPs, 10 pmol of MC1R specificprimers, 100 pmol of Universal Primer Mix (or NestedUniversal Primer Mix), and 3 U of the Expand Long templateTaq polymerase mix (Roche, Indianapolis, IN, USA). After aninitial denaturation for 2 min at 94°C, 34 cycles at 94°C for30 s, 61°C for 30 s, and 68°C for 2 min were performed,followed by a final extension at 68°C for 5 min in a DNAThermal Cycler 9700 (Perkin Elmer, Boston, MA, USA).Fragments of interest were cloned into the pGEM®-T EasyVector (Promega, Madison, WI, USA).
Northern blot
Northern blot was performed following the NorthernMaxprocedure (Ambion, Austin, TX, USA). Briefly, 50 �g of totalRNA was denatured in formaldehyde buffer for 15 min at65°C before electrophoresis on a 1% agarose gel. Aftertransfer to a BrightStar-Plus membrane (Ambion), RNA wascross-linked using UV radiation (Stratalinker, Stratagene, LaJolla, CA, USA). A sequence-verified fragment of the humanMC1R cDNA was radiolabeled with deoxycytidine 5�[C32P]-triphosphate by random priming using the DECAprime IIlabeling kit (Ambion). Northern blots were hybridized for16 h at 42°C, then washed twice for 5 min in 2� saline-sodium
citrate (SSC), 0.1% SDS, and once for 15 min in 0.1� SSC,0.1% SDS at 42°C. Membranes were exposed on a MolecularDynamics phosphor screen for 10 days before analysis with aSTORM 860 PhosphorImager (Molecular Dynamics).
Sequencing analysis
Cloned fragments were analyzed by automatic sequencingusing the ABI PRISM Big Dye Terminator reaction kit (Ap-plied Biosystems, Foster City, CA, USA) and an automaticsequencer (ABI PRISM Cycle sequencing, Perkin Elmer,Boston, MA, USA) and sequencing files were analyzed withSequencher 4.0.5 software. Nucleotidic sequences were com-pared using the CLUSTALW (EMBL-European BioinformaticsInstitute, Cambridge, UK) and the basic local alignment searchtool (BLAST) (NCBI, Bethesda, MD, USA) programs. Receptorsecondary structure predictions were performed using the Pre-dict Protein program, accessible at: www.embl-heidelberg.de/predictprotein.
Immunohistochemistry for skin sections
Study subjects
1. This study involved nine healthy volunteer subjects whoseage ranged from 24 to 59 years old from three ethnic groups(African American, Asian, and Caucasian) and were selectedamong those used in an earlier study (23, 32). The biopsysites were all midback skin. This study was approved by theFDA Research Involving Human Subjects Committee, andinformed consent was obtained from each study subject.
2. Samples fixed with formaldehyde were sectioned at 3 �mand mounted on silane-coated glass slides. The expression ofMC1R was detected in paraffin-embedded sections by indirectimmunofluorescence using rabbit polyclonal antibody (pAb),�PEP20 raised in our laboratory against amino acids 319–338of MC1R350 according to our previously published protocol(33). Samples were deparaffinized twice with xylene for 5 minand dehydrated with graduated ethanol. Specimens werethen treated with cold methanol for 20 min and boiled with10 mM citrate buffer (pH 6.0) for 20 min. They weresubsequently incubated with 10% goat serum (Vector Labo-ratories, Burlingame, CA, USA) containing 2% BSA (Amer-sham Pharmacia) and 0.05% Tween20 for 30 min at 37°C,then with �PEP20 at 1:1,000 dilution with 2% goat serum at4°C overnight. The slides were incubated with Alexa Fluor(R)488 goat anti-rabbit IgG (H�L) (Molecular Probes, Eugene,OR, USA) at 37°C for 30 min at 1:500 dilution with 2% goatserum and covered by a drop of ProlongTM Antifade Kit(Molecular Probes) containing 1 �g/ml propidium iodide.The Alexa488 fluorescence was superimposed over pro-pidium iodide to show colocalization. Fluorescence was ana-lyzed and quantified using a LEICA DMRB/DMLD lasermicroscope (Leica, Wetzlar, Germany) and ScionImage com-puter software as previously reported (32). This system allowsone to eliminate background level fluorescence and to quan-tify fluorescence intensity from original images.
3. Deparaffinized skin sections were processed for MC1Rimmunostaining with N2–18, an Ab directed against theamino acids 2–18 of the human MC1R, as described beforewith slight modifications (34–35). In short, deparaffinizedsections were microwave-treated for epitope retrieval, fol-lowed by quenching with 0.33% hydrogen peroxide for 20min and blocking with 2% BSA for 25 min. Sections were thenincubated with the anti-MC1R Ab at 1 or 2 �g/ml for 45 min,followed by washing and Ab detection using the indirectimmunoperoxidase technique. For negative controls, sections
E1340 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal

were incubated with preimmune serum at the same concen-tration as the primary Ab.
Single and dual Ab labeling for confocalimmunofluorescence
NHM were seeded in 2-well Lab-Tek chamber slides (NalgeNunc Intl., Naperville, IL, USA). After three washes in PBS,the cells were fixed in 4% paraformaldehyde for 15 min at4°C. Then the cells were permeabilized with 0.01% Triton-X100 for 3 min at room temperature. For labeling with asingle MC1R Ab, melanocytes were blocked with 5% normalgoat serum for 1 h at room temperature. Melanocytes wereincubated with the MC1R antibodies �PEP20 (1:10 dilution)or SC-N19 (1:20 dilution) (Santa Cruz Biotechnology, SantaCruz, CA, USA) overnight at 4°C. After three washes in PBS,the polyclonal antibodies were reacted with goat anti-rabbitIgG labeled with Texas red (1:100) and the monoclonalantibodies were reacted with goat antimouse IgG labeled withfluorescein (1:100) (Vector), followed by nuclear counter-staining with 4�,6�-diam idino-2-phenylidole (DAPI) (Vector).Immunofluorescence signals were classified into three cate-gories according to whether they showed green, red, or yellowfluorescence. The latter was indicative of colocalization of thered and green fluorescence signals. All preparations wereexamined with a confocal microscope LSM 510 (Zeiss, Jena,Germany) equipped with HeNe, argon, krypton, and UV lasersources.
Fluorescence index (FI) measurements
The fluorescence index was obtained by measurement of thefluorescence signal from MC1R Ab in at least five melanocytesunder �-MSH-treated or untreated conditions. Briefly, origi-nal confocal microscopy images were obtained under samelaser gain intensity, size and magnification. These imageswere analyzed by using the software contained in the Zeiss 510LSM confocal microscope. All measurements were collectedto a database and are expressed as mean � sd.
Comparison of MC1R transcript levels
Comparative determination of MC1R transcript levels wereperformed by real-time semiquantitative RT-PCR using theSyBr Green detection on a Light Cycler (Roche) and specificprimers. Because the levels and the quality of mRNA may varyslightly according to the different types of treatment, wequantified the transcripts relative to the GAPDH housekeep-ing gene using the primers listed above by determining thedifference between the crossing point (Ct) of amplification ofthe target RNA and the GAPDH RNA (delta Ct GAPDH). TheCt is defined as the point when the amplification starts theexponential phase. Comparison of transcript levels then relieson differences between the delta Cts (delta-deltaCt, ddct). Weconsidered the MC1R transcript level in 964b� cells as areference, since they were the most highly pigmented. There-fore, the numbers of fold activation were calculated asfollows. Given the relation: nb fold � nb copies MC1R(target)/nb copies MC1R (reference), nb fold can be ex-pressed as: 10 (log(nb copies Mc1r (target) – log (nb copies Mc1r
(reference)).
RESULTS
Morphology and growth of NHM in culture
Eighteen different NHM cultures derived from humanneonatal foreskins obtained from individual donors
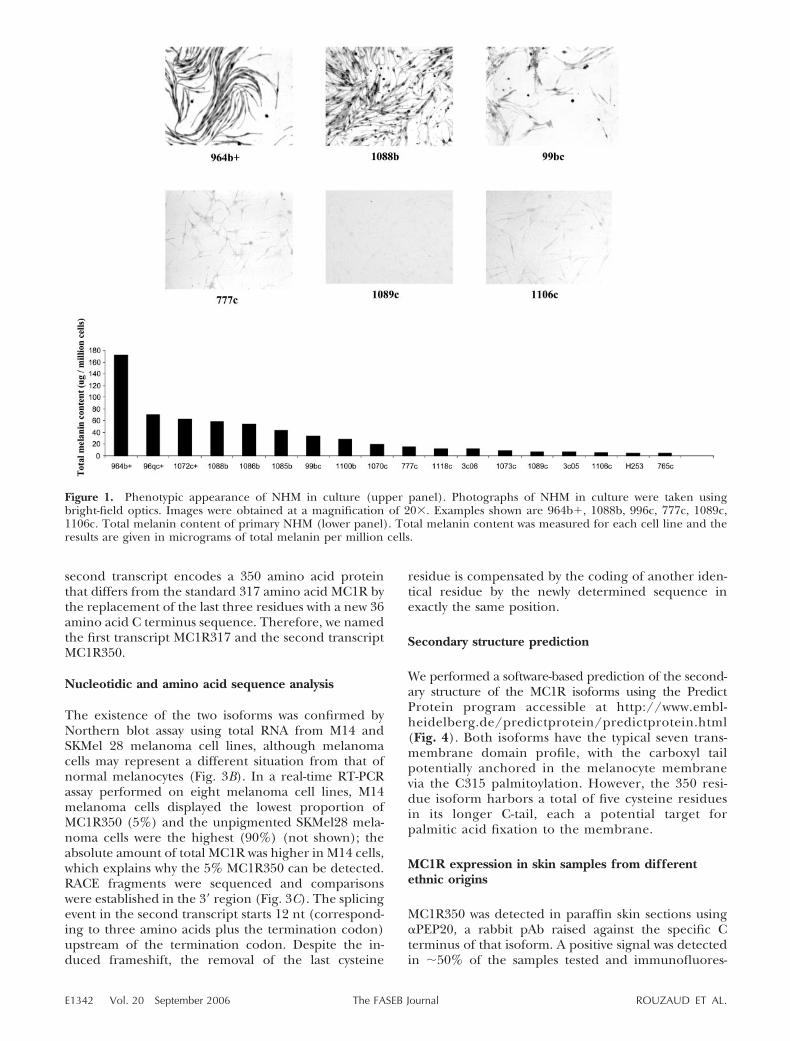
presenting different levels of skin pigmentation (do-nors were anonymous, therefore no information onethnicity was available) were established and main-tained under identical conditions. Microphotographswere taken using a bright-field filter (Fig. 1). Theintensity of pigmentation varied from one populationto another, spanning a large range of intensities fromvery dark to very lightly pigmented; measurements ofmelanin content were consistent with visible phenotype(Fig. 1).
Immunodetection of MC1R isoforms expressed bycultured melanocytes
Western blot experiments were performed to measureMC1R levels in the various melanocyte populations(Fig. 2). Immunodetection by �PEP19 revealed threepatterns of expression. Pattern 1 (shown in Fig. 2A) hada single band at 60 kDa whereas pattern 2 (Fig. 2B)revealed two sets of doublet bands, one at 30 and 36kDa and the other at �60 and 70 kDa. Pattern 3 (Fig.2C) showed only one band at 30 kDa and one at 60 kDa,and was the most common pattern.
The receptor complexes
A binding experiment was conducted to determine ifthe doublet of high MW bands (�60–70 kDa) revealedby Western blot correspond to a multi-MC1R complexstructure that could bind �-MSH. We used 125I-�MSHlabeling to detect the receptor, which revealed twodistinct signals at �30–35 kDa and 60–70 kDa andcorresponds to the sizes identified by immunoblotting(Fig. 2D). The intensity of the two signals is approxi-mately equivalent, suggesting that �-MSH binds withsimilar affinity to either the high or the low MW formsof the complexes. The same profile was obtained inboth native and reducing conditions, indicating thatthe bond involved in the complex formation is cova-lent. Our results suggest that what we are detecting at60–70 kDa corresponds to a dimeric MC1R, but ourdata cannot at this point exclude the possibility that itcould be the result of an interaction between MC1Rand another protein.
Characterization of untranslated regions (UTRs)
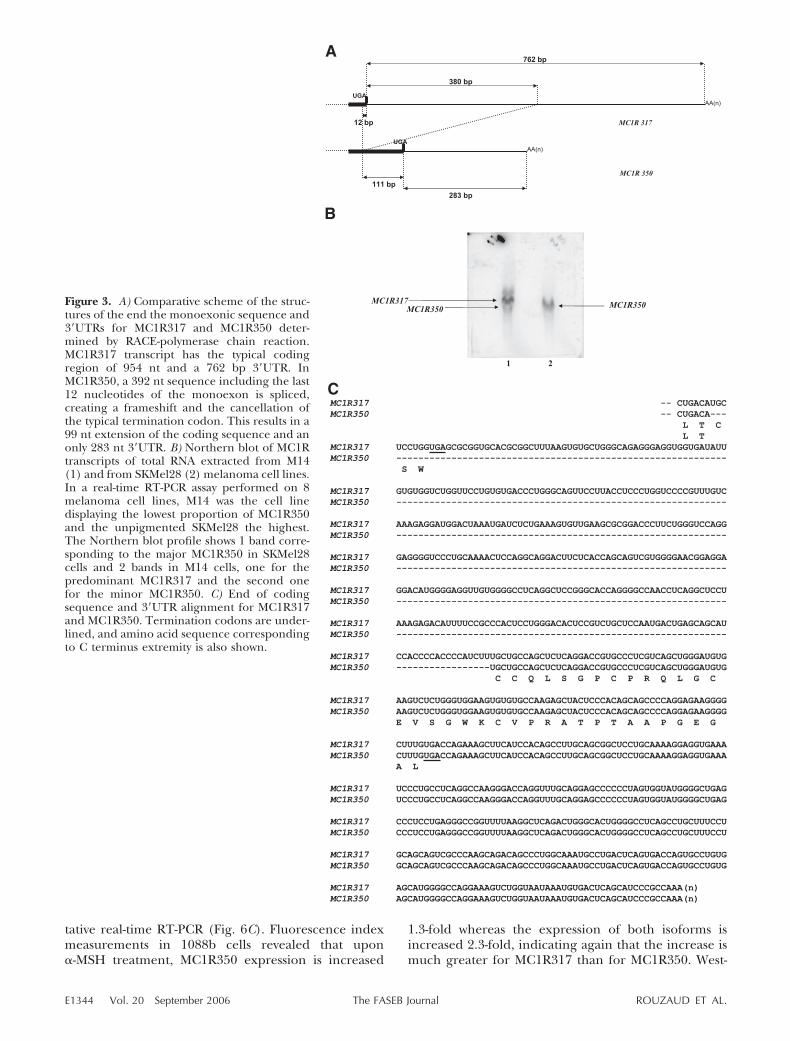
To characterize at the nucleotide level the isoformsdepicted in the Western blots, nested amplificationswere performed using two distinct adapter-ligatedcDNA libraries for each cell line. 5� RACE-PCR analysisrevealed only one DNA fragment corresponding to a5�untranslated region (5� UTR) of 547 bp. Similarinvestigations were performed to obtain the 3� UTRbut, surprisingly, different products were detected inthe 3� RACE-PCR experiments (Fig. 3A). The largestfragment corresponded to the typical 954 nt codingregion with a 762 nt 3� UTR. The smaller fragmentcorresponded to a 1053 nt coding region with a 283 nt3� UTR. The sequence translation indicates that the
E1341IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM
second transcript encodes a 350 amino acid proteinthat differs from the standard 317 amino acid MC1R bythe replacement of the last three residues with a new 36amino acid C terminus sequence. Therefore, we namedthe first transcript MC1R317 and the second transcriptMC1R350.
Nucleotidic and amino acid sequence analysis
The existence of the two isoforms was confirmed byNorthern blot assay using total RNA from M14 andSKMel 28 melanoma cell lines, although melanomacells may represent a different situation from that ofnormal melanocytes (Fig. 3B). In a real-time RT-PCRassay performed on eight melanoma cell lines, M14melanoma cells displayed the lowest proportion ofMC1R350 (5%) and the unpigmented SKMel28 mela-noma cells were the highest (90%) (not shown); theabsolute amount of total MC1R was higher in M14 cells,which explains why the 5% MC1R350 can be detected.RACE fragments were sequenced and comparisonswere established in the 3� region (Fig. 3C). The splicingevent in the second transcript starts 12 nt (correspond-ing to three amino acids plus the termination codon)upstream of the termination codon. Despite the in-duced frameshift, the removal of the last cysteine
residue is compensated by the coding of another iden-tical residue by the newly determined sequence inexactly the same position.
Secondary structure prediction
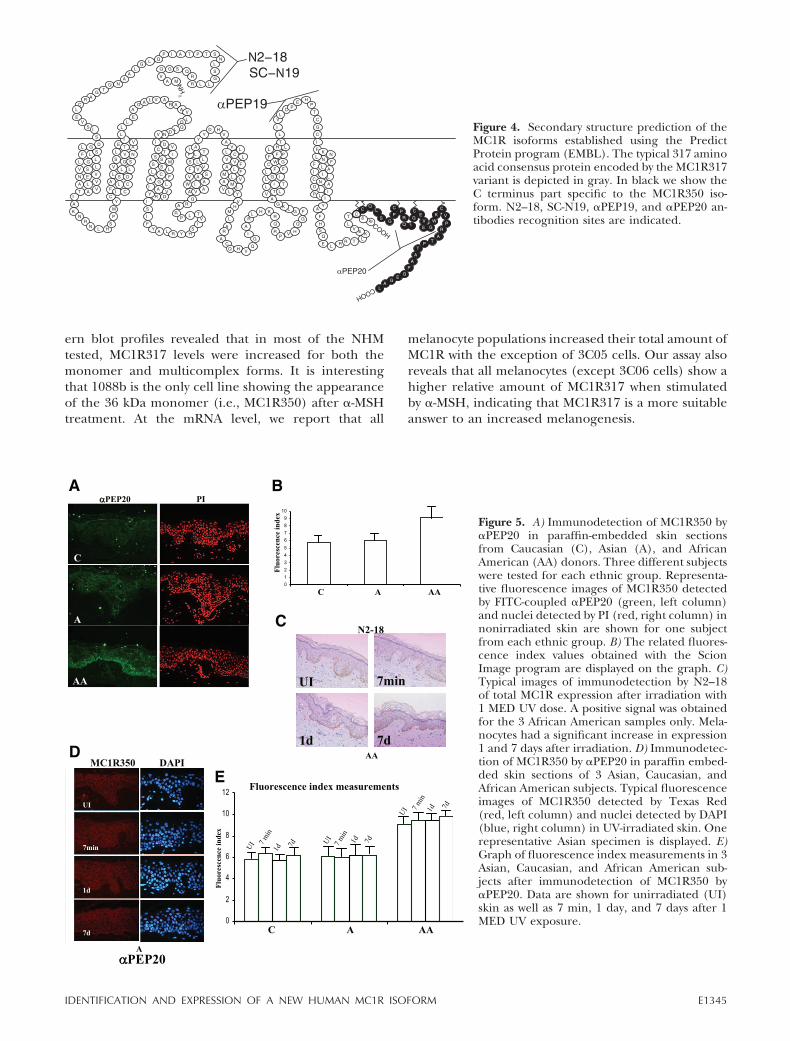
We performed a software-based prediction of the second-ary structure of the MC1R isoforms using the PredictProtein program accessible at http://www.embl-heidelberg.de/predictprotein/predictprotein.html(Fig. 4). Both isoforms have the typical seven trans-membrane domain profile, with the carboxyl tailpotentially anchored in the melanocyte membranevia the C315 palmitoylation. However, the 350 resi-due isoform harbors a total of five cysteine residuesin its longer C-tail, each a potential target forpalmitic acid fixation to the membrane.
MC1R expression in skin samples from differentethnic origins
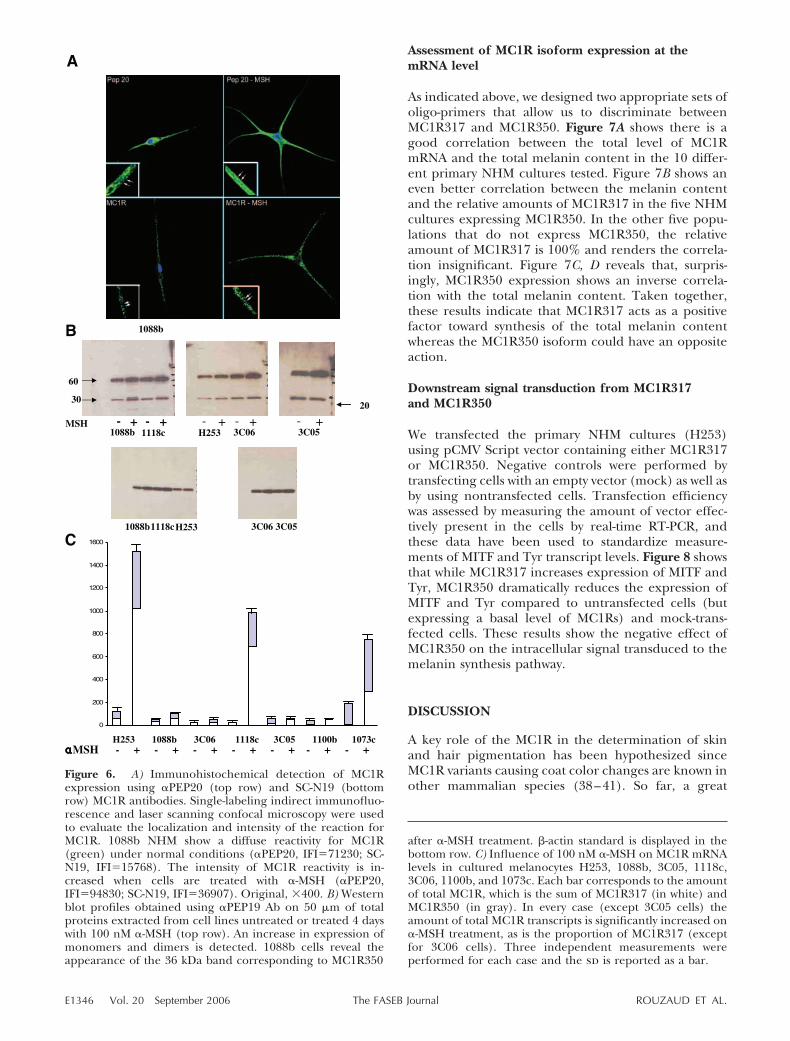
MC1R350 was detected in paraffin skin sections using�PEP20, a rabbit pAb raised against the specific Cterminus of that isoform. A positive signal was detectedin �50% of the samples tested and immunofluores-
Figure 1. Phenotypic appearance of NHM in culture (upper panel). Photographs of NHM in culture were taken usingbright-field optics. Images were obtained at a magnification of 20�. Examples shown are 964b�, 1088b, 996c, 777c, 1089c,1106c. Total melanin content of primary NHM (lower panel). Total melanin content was measured for each cell line and theresults are given in micrograms of total melanin per million cells.
E1342 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal

cence measurements indicated similar low levels ofMC1R350 expression in Caucasian and in Asian skin(Fig. 5A, B) and a significant 25% higher level inAfrican American skin. Due to the structural character-istics of the two isoforms, it is not possible to design anAb that would recognize MC1R317 only. Therefore, anyAb that recognizes MC1R317 will also detect de factoMC1R350, revealing the cumulative expression of thetwo isoforms. We therefore used N2–18 Ab (34–35)and were able to detect the MC1Rs in African Americansamples only (Fig. 5C). The N2–18 appears not to besensitive enough to detect MC1R in Caucasian andAsian skin, where its level of expression is lower.
Influence of 1 minimal erythemal dose (MED) UVRon the MC1Rs expression in skin
�PEP20 and N2–18 were used to monitor respectivelyMC1R350 and total MC1R expression 7 min, 1 day, and
7 days after irradiation with 1MED of UV. MC1R350expression did not significantly vary after UV irradia-tion in any of the ethnic groups examined (Fig. 5D, E).At 1 day and 7 days after irradiation, a significantincrease was detected by staining with N2–18 whichtargets the total MC1R corresponding to the sum ofMC1R350 and MC1R317 (Fig. 5C). Therefore, we at-tribute the increase detected by N2–18 to MC1R317.
Influence of �-MSH on MC1R expressionin cultured NHM
It has been known for a long time that �-MSH is aprincipal mediator of the melanogenic effects of UVR(28, 36–37, 31). Therefore, we treated cultured pri-mary NHM for 4 days with 100 nM �-MSH and assessedthe changes in MC1R expression by confocal immuno-fluorescence (Fig. 6A) and Western blot (Fig. 6B) usingSC-N19 and �PEP19 antibodies as well as semiquanti-
Figure 2. Several isoforms of MC1R expressed by melanocytes are observed in different human cell lines. Western blot profilesobtained using �PEP19 Ab on 50 �g of total proteins extracted from cell lines are shown. A) Detection using �PEP19 Ab ofMC1R proteins reveal a single band at 60 kDa. B) Immunodetection revealed two sets of two bands each: one at 30 kDa and 36kDa, and a second one with a greater intensity at �60 kDa and 70 kDa. C) The most widely represented with 1 band at 30 kDaand a second one at 60 kDa. D) [125I] �-MSH binding assay. 964b� melanocytes were incubated with 10 �Ci of[125I](Lys11)(Nle4-D-Phe7)�-MSH for 2 h, then washed as described in Materials and Methods. �-MSH that remained bound toMC1R allowed the detection of 2 pools of signals in native and reducing conditions. The lower signal corresponds to a molecularsize of �30 to 35 kDa and the higher signal gives a size of �60 to 70 kDa. There is no detectable difference between theintensities of the signals in either native or reducing environments. The corresponding Western blot profile is displayed for bandsize comparison.
E1343IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM
tative real-time RT-PCR (Fig. 6C). Fluorescence indexmeasurements in 1088b cells revealed that upon�-MSH treatment, MC1R350 expression is increased
1.3-fold whereas the expression of both isoforms isincreased 2.3-fold, indicating again that the increase ismuch greater for MC1R317 than for MC1R350. West-
Figure 3. A) Comparative scheme of the struc-tures of the end the monoexonic sequence and3�UTRs for MC1R317 and MC1R350 deter-mined by RACE-polymerase chain reaction.MC1R317 transcript has the typical codingregion of 954 nt and a 762 bp 3�UTR. InMC1R350, a 392 nt sequence including the last12 nucleotides of the monoexon is spliced,creating a frameshift and the cancellation ofthe typical termination codon. This results in a99 nt extension of the coding sequence and anonly 283 nt 3�UTR. B) Northern blot of MC1Rtranscripts of total RNA extracted from M14(1) and from SKMel28 (2) melanoma cell lines.In a real-time RT-PCR assay performed on 8melanoma cell lines, M14 was the cell linedisplaying the lowest proportion of MC1R350and the unpigmented SKMel28 the highest.The Northern blot profile shows 1 band corre-sponding to the major MC1R350 in SKMel28cells and 2 bands in M14 cells, one for thepredominant MC1R317 and the second onefor the minor MC1R350. C) End of codingsequence and 3�UTR alignment for MC1R317and MC1R350. Termination codons are under-lined, and amino acid sequence correspondingto C terminus extremity is also shown.
E1344 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal
ern blot profiles revealed that in most of the NHMtested, MC1R317 levels were increased for both themonomer and multicomplex forms. It is interestingthat 1088b is the only cell line showing the appearanceof the 36 kDa monomer (i.e., MC1R350) after �-MSHtreatment. At the mRNA level, we report that all
melanocyte populations increased their total amount ofMC1R with the exception of 3C05 cells. Our assay alsoreveals that all melanocytes (except 3C06 cells) show ahigher relative amount of MC1R317 when stimulatedby �-MSH, indicating that MC1R317 is a more suitableanswer to an increased melanogenesis.
Figure 4. Secondary structure prediction of theMC1R isoforms established using the PredictProtein program (EMBL). The typical 317 aminoacid consensus protein encoded by the MC1R317variant is depicted in gray. In black we show theC terminus part specific to the MC1R350 iso-form. N2–18, SC-N19, �PEP19, and �PEP20 an-tibodies recognition sites are indicated.
Figure 5. A) Immunodetection of MC1R350 by�PEP20 in paraffin-embedded skin sectionsfrom Caucasian (C), Asian (A), and AfricanAmerican (AA) donors. Three different subjectswere tested for each ethnic group. Representa-tive fluorescence images of MC1R350 detectedby FITC-coupled �PEP20 (green, left column)and nuclei detected by PI (red, right column) innonirradiated skin are shown for one subjectfrom each ethnic group. B) The related fluores-cence index values obtained with the ScionImage program are displayed on the graph. C)Typical images of immunodetection by N2–18of total MC1R expression after irradiation with1 MED UV dose. A positive signal was obtainedfor the 3 African American samples only. Mela-nocytes had a significant increase in expression1 and 7 days after irradiation. D) Immunodetec-tion of MC1R350 by �PEP20 in paraffin embed-ded skin sections of 3 Asian, Caucasian, andAfrican American subjects. Typical fluorescenceimages of MC1R350 detected by Texas Red(red, left column) and nuclei detected by DAPI(blue, right column) in UV-irradiated skin. Onerepresentative Asian specimen is displayed. E)Graph of fluorescence index measurements in 3Asian, Caucasian, and African American sub-jects after immunodetection of MC1R350 by�PEP20. Data are shown for unirradiated (UI)skin as well as 7 min, 1 day, and 7 days after 1MED UV exposure.
E1345IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM
Assessment of MC1R isoform expression at themRNA level
As indicated above, we designed two appropriate sets ofoligo-primers that allow us to discriminate betweenMC1R317 and MC1R350. Figure 7A shows there is agood correlation between the total level of MC1RmRNA and the total melanin content in the 10 differ-ent primary NHM cultures tested. Figure 7B shows aneven better correlation between the melanin contentand the relative amounts of MC1R317 in the five NHMcultures expressing MC1R350. In the other five popu-lations that do not express MC1R350, the relativeamount of MC1R317 is 100% and renders the correla-tion insignificant. Figure 7C, D reveals that, surpris-ingly, MC1R350 expression shows an inverse correla-tion with the total melanin content. Taken together,these results indicate that MC1R317 acts as a positivefactor toward synthesis of the total melanin contentwhereas the MC1R350 isoform could have an oppositeaction.
Downstream signal transduction from MC1R317and MC1R350
We transfected the primary NHM cultures (H253)using pCMV Script vector containing either MC1R317or MC1R350. Negative controls were performed bytransfecting cells with an empty vector (mock) as well asby using nontransfected cells. Transfection efficiencywas assessed by measuring the amount of vector effec-tively present in the cells by real-time RT-PCR, andthese data have been used to standardize measure-ments of MITF and Tyr transcript levels. Figure 8 showsthat while MC1R317 increases expression of MITF andTyr, MC1R350 dramatically reduces the expression ofMITF and Tyr compared to untransfected cells (butexpressing a basal level of MC1Rs) and mock-trans-fected cells. These results show the negative effect ofMC1R350 on the intracellular signal transduced to themelanin synthesis pathway.
DISCUSSION
A key role of the MC1R in the determination of skinand hair pigmentation has been hypothesized sinceMC1R variants causing coat color changes are known inother mammalian species (38–41). So far, a great
Figure 6. A) Immunohistochemical detection of MC1Rexpression using �PEP20 (top row) and SC-N19 (bottomrow) MC1R antibodies. Single-labeling indirect immunofluo-rescence and laser scanning confocal microscopy were usedto evaluate the localization and intensity of the reaction forMC1R. 1088b NHM show a diffuse reactivity for MC1R(green) under normal conditions (�PEP20, IFI�71230; SC-N19, IFI�15768). The intensity of MC1R reactivity is in-creased when cells are treated with �-MSH (�PEP20,IFI�94830; SC-N19, IFI�36907). Original, �400. B) Westernblot profiles obtained using �PEP19 Ab on 50 �m of totalproteins extracted from cell lines untreated or treated 4 dayswith 100 nM �-MSH (top row). An increase in expression ofmonomers and dimers is detected. 1088b cells reveal theappearance of the 36 kDa band corresponding to MC1R350
after �-MSH treatment. -actin standard is displayed in thebottom row. C) Influence of 100 nM �-MSH on MC1R mRNAlevels in cultured melanocytes H253, 1088b, 3C05, 1118c,3C06, 1100b, and 1073c. Each bar corresponds to the amountof total MC1R, which is the sum of MC1R317 (in white) andMC1R350 (in gray). In every case (except 3C05 cells) theamount of total MC1R transcripts is significantly increased on�-MSH treatment, as is the proportion of MC1R317 (exceptfor 3C06 cells). Three independent measurements wereperformed for each case and the sd is reported as a bar.
E1346 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal
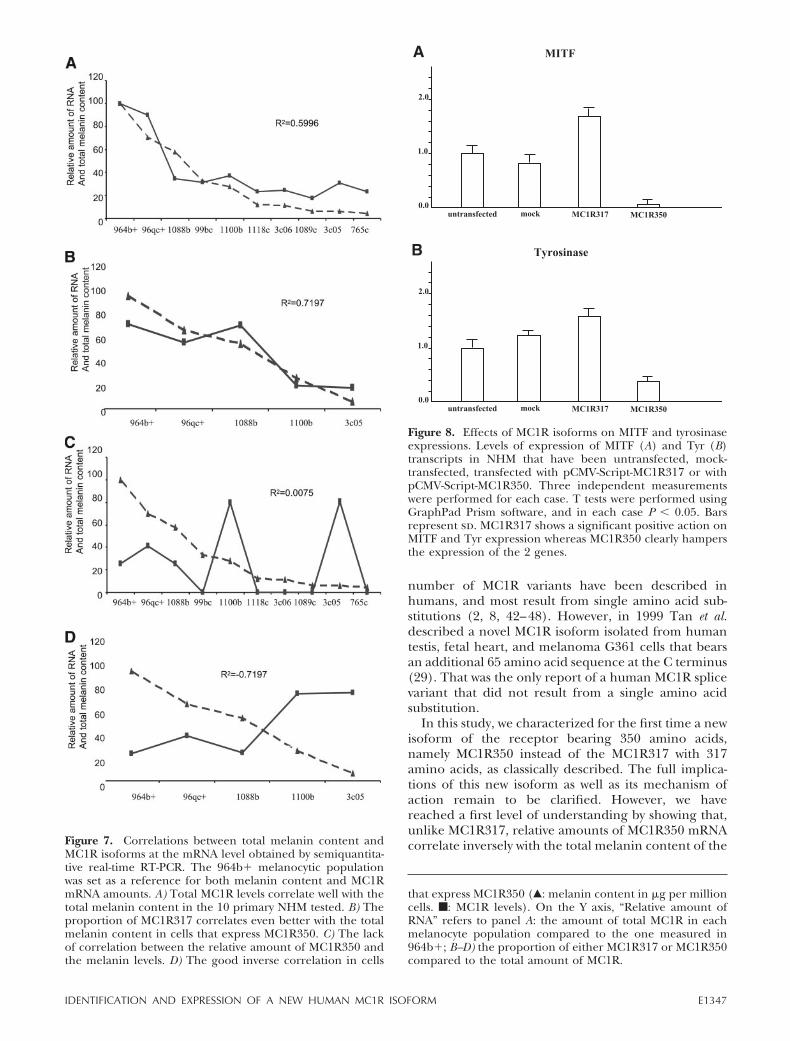
number of MC1R variants have been described inhumans, and most result from single amino acid sub-stitutions (2, 8, 42–48). However, in 1999 Tan et al.described a novel MC1R isoform isolated from humantestis, fetal heart, and melanoma G361 cells that bearsan additional 65 amino acid sequence at the C terminus(29). That was the only report of a human MC1R splicevariant that did not result from a single amino acidsubstitution.
In this study, we characterized for the first time a newisoform of the receptor bearing 350 amino acids,namely MC1R350 instead of the MC1R317 with 317amino acids, as classically described. The full implica-tions of this new isoform as well as its mechanism ofaction remain to be clarified. However, we havereached a first level of understanding by showing that,unlike MC1R317, relative amounts of MC1R350 mRNAcorrelate inversely with the total melanin content of theFigure 7. Correlations between total melanin content and
MC1R isoforms at the mRNA level obtained by semiquantita-tive real-time RT-PCR. The 964b� melanocytic populationwas set as a reference for both melanin content and MC1RmRNA amounts. A) Total MC1R levels correlate well with thetotal melanin content in the 10 primary NHM tested. B) Theproportion of MC1R317 correlates even better with the totalmelanin content in cells that express MC1R350. C) The lackof correlation between the relative amount of MC1R350 andthe melanin levels. D) The good inverse correlation in cells
Figure 8. Effects of MC1R isoforms on MITF and tyrosinaseexpressions. Levels of expression of MITF (A) and Tyr (B)transcripts in NHM that have been untransfected, mock-transfected, transfected with pCMV-Script-MC1R317 or withpCMV-Script-MC1R350. Three independent measurementswere performed for each case. T tests were performed usingGraphPad Prism software, and in each case P 0.05. Barsrepresent sd. MC1R317 shows a significant positive action onMITF and Tyr expression whereas MC1R350 clearly hampersthe expression of the 2 genes.
that express MC1R350 (Œ: melanin content in �g per millioncells. f: MC1R levels). On the Y axis, “Relative amount ofRNA” refers to panel A: the amount of total MC1R in eachmelanocyte population compared to the one measured in964b�; B–D) the proportion of either MC1R317 or MC1R350compared to the total amount of MC1R.
E1347IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM

NHM tested. This shows that MC1R350 is not positivelyinvolved in melanin synthesis but is perhaps a negativeregulator. We have further demonstrated the negativeimpact of MC1R350 expression on MITF and tyrosi-nase, two key players of melanogenesis. When primaryNHM are treated with �-MSH, most of the time theyincrease their level of MC1R350 amount; in contrast,however, in the majority of the cases they also decreasetheir proportion of this isoform (Fig. 6C), as largeincreases in MC1R317 appear to be a more suitableanswer to stimulation of the melanogenic pathway formelanocytes. Therefore, one can expect that the mech-anism revealed for NHM also applies in the skin. Sinceour immunohistochemical studies have shown thatAfrican American skins have higher absolute levels ofMC1R350 compared to Asian and Caucasian skins (Fig.5), similar to the way that �-MSH-treated NHM increasetheir levels of MC1R350, we expect that darker skinshave a greater relative amount of MC1R317. Indeed,UVR exposure does not increase MC1R350 expressionbut does significantly increase that of MC1R317, there-fore increasing its relative amount as well as melaninproduction. In the model we propose, levels of melaninproduction appear to be regulated by the balancebetween the two isoforms of MC1R, and the relativeamounts of MC1R317 and MC1R350 act like a rheostatto control the melanin production via MITF and tyrosi-nase. However, regulation of the balance itself will beinteresting to investigate since MC1R350 is generatedfrom MC1R317 by alternative splicing, and thus thenonspliced RNA could be considered an immaturetranscript. On the other hand, from the protein pointof view we cannot rule out the possibility that MC1R350might get cleaved to the shorter form by a protease,even though the C terminus would be slightly modified,in which case MC1R350 could be considered a prore-ceptor.
One way to control skin pigmentation was elucidatedat the beginning of the 1990s with the molecularcharacterization of the MC1R and its antagonist agoutisignal protein (ASP) (4, 49), and we now know thatMC1R and ASP are involved in the qualitative andquantitative regulation of mammalian pigmentation(50–51). ASP is produced in hair follicles and acts onfollicular melanocytes to inhibit eumelanin synthesis(52–53). Recently, the 8818G allele of the ASP gene hasbeen associated with a darker skin color in AfricanAmericans (54). Here, we report for the first time thatMC1R isoform-specific expression is closely linked todarker skin pigmentation and photoprotection.
The secondary structure prediction indicates that theC-tail of MC1R350 bears not only one (as doesMC1R317), but five, cysteine residues that could bepalmitoylated in order to further anchor the tail of thereceptor in the melanocyte membrane, which mightsignificantly increase its stability, localization, or func-tion. The carboxyl-terminal domain for all knownMC1R proteins is strongly conserved, particularly thecysteine in position 315. In domestic dogs, a pheomela-nic hair pattern is induced by the nonfunctional
R306ter mutation at the Mc1r locus (40, 55), whichresults from an early termination codon and the loss ofthe last 12 residues of the receptor, including the C315.By analogy to other G-protein-coupled receptors, theMC1R cytoplasmic tail is not expected to directly par-ticipate in signaling, but might impair coupling indi-rectly if C315 is palmitoylated and participates in recep-tor localization or trafficking (56–59). Biochemical andpharmacological studies may help to resolve the influ-ence of this longer C terminus on MC1R function andassess whether or not this sequence is responsible forthe negative effect of MC1R350 on the melanogenicpathway.
The Western blot profiles using �PEP19 also revealeda band (or a doublet) with a MW �65–70 kDa, i.e.,twice the size expected for the MC1R. Using 125I-�MSHlabeling, we detected two distinct binding signals, eachcorresponding to the sizes seen by immunoblotting.The sum of these results clearly shows that MC1R canbe involved in a multiproteic structure that is able tobind �-MSH with approximately the same affinity as thesingle receptor. Nevertheless, further studies areneeded to establish precisely the nature of the multi-proteic complex and its properties. We show that bothstructures are stable under reducing conditions, whichstrongly suggests they result from covalent bonds be-tween the entities and not from disulfide bonds be-tween the cysteine residues.
One of the most pressing and tantalizing issues raisedby these results is the significance of the MC1R isoformsand their potential dimerization for the pharmacologyof MC1R function, and whether this will result in theidentification and development of novel ligands thatinteract selectively with such receptors. It was recentlyreported that MC1R forms dimeric structures (18), andthat might be what we are detecting here, in which casefurther studies will be necessary to establish whetherMC1R can form heterodimers in an MC1R317-MC1R350 complex. Does MC1R350, as a negative reg-ulator of pigmentation, offer novel possibilities in termsof ways to inhibit melanogenesis? Melanocortin peptidestructure-activity relationship studies are providing keyinsights regarding the design and synthesis of melano-cortin selective agonists and/or antagonists and tre-mendous efforts have been recently put into the designand development of ligands for the melanocortin re-ceptors with properties not present in the endogenouspeptides, such as improved potency, receptor selectiv-ity, and bioavailability (60).
MC1R350 has been detected in �50% of the celllines studied, but so far we have not identified any cellline that expresses exclusively MC1R350. A largerscreening process hopefully will identify melanocytesproducing only MC1R350, which would be valuable asan in vivo model to further study the pharmacology andthe functionality of this isoform.
This research was supported by the Intramural ResearchProgram of the NIH, National Cancer Institute, Center forCancer Research, and also in part by NIH grant R01
E1348 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal

ES009110 (Z. Abdel-Malek). The authors wish to thank Dr.Janusz Z. Beer and Barbara Z. Zmudska of the FDA forproviding some of the specimens used in this study, and Pr.J. L. Garcia-Borron from the University of Murcia, Spain forinsightful discussions.
REFERENCES
1. Harrison, G. A. (1973) Differences in human pigmentation:measurement, geographic variation, and causes. J. Invest. Derma-tol. 60, 418–426
2. Valverde, P., Healy, E., Jackson, I., Rees, J. L., and Thody, A. J.(1995) Variants of the melanocyte-stimulating hormone recep-tor gene are associated with red hair and fair skin in humans.Nat. Genet. 11, 328–330
3. Box, N. F., Wyeth, J. R., O’Gorman, L. E., Martin, N. G., andSturm, R. A. (1997) Characterization of melanocyte stimulatinghormone receptor variant alleles in twins with red hair. Hum.Mol. Genet. 1891–1897
4. Mountjoy, K. G., Robbins, L. S., Mortrud, M. T., and Cone, R. D.(1992) The cloning of a family of genes that encode themelanocortin receptors. Science 257, 1248–1251
5. Chhajlani, V., and Wikberg, J. E. (1992) Molecular cloning andexpression of the human melanocyte stimulating hormonereceptor cDNA. FEBS Lett. 309, 417–420
6. Smith, R., Healy, E., Siddiqui, S., Flanagan, N., Steijlen, P. M.,Rosdahl, I., Jacques, J. P., Rogers, S., Turner, R., Jackson, I. J., etal. (1998) Melanocortin 1 receptor variants in an Irish popula-tion. J. Invest. Dermatol. 111, 119–122
7. Frandberg, P. A., Doufexis, M., Kapas, S., and Chhajlani, V.(1998) Human pigmentation phenotype: a point mutationgenerates nonfunctional MSH receptor. Biochem. Biophys. Res.Commun. 245, 490–492
8. Schioth, H. B., Phillips, S. R., Rudzish, R., Birch-Machin, M. A.,Wikberg, J. E., and Rees, J. L. (1999) Loss of function mutationsof the human melanocortin 1 receptor are common and areassociated with red hair. Biochem. Biophys. Res. Commun. 260,488–491
9. Suzuki, I., Cone, R. D., Im, S., Nordlund, J., and Abdel-Malek,Z. A. (1996) Binding of melanotropic hormones to the melano-cortin receptor MC1R on human melanocytes stimulates prolif-eration and melanogenesis. Endocrinology 137, 1627–1633
10. Abdel-Malek, Z., Swope, V. B., Suzuki, I., Akcali, C., Harriger,M. D., Boyce S. T., Urabe K., and Hearing, V. J. (1995)Mitogenic and melanogenic stimulation of normal humanmelanocytes by melanotropic peptides. Proc. Natl. Acad. Sci.U. S. A. 92, 1789–1793
11. Bockaert, J., and Pin, J. P. (1999) Molecular tinkering of Gprotein-coupled receptors: an evolutionary success. EMBO J. 18,1723–1729
12. Wilson, S., Bergsma, D. J., Chambers, J. K., Muir, A. I., Fantom,K. G., Ellis, C., Murdock, P. R., Herrity, N. C., and Stadel, J. M.(1998) Orphan G-protein-coupled receptors: the next genera-tion of drug targets? Br. J. Pharmacol. 125, 1387–1392
13. Salahpour, A., Angers, S., and Bouvier, M. (2000) Functionalsignificance of oligomerization of G-protein-coupled receptors.Trends Endocrinol. Metab. 11, 163–168
14. Donatien, P. D., Hunt, G., Pieron, C., Lunec, J., Taieb, A., andThody, A. J. (1992) The expression of functional MSH receptorson cultured human melanocytes. Arch. Dermatol. Res. 284, 424–426
15. Hebert, T. E., and Bouvier, M. (1998) Structural and functionalaspects of G protein-coupled receptor oligomerization. Biochem.Cell Biol. 76, 1–11
16. Milligan, G. (2004) G protein-coupled receptor dimerization:function and ligand pharmacology. Mol. Pharmacol. 66, 1–7
17. Mandrika, I., Petrovska, R., and Wikberg, J. (2005) Melanocor-tin receptors form constitutive homo- and heterodimers. Bio-chem. Biophys. Res. Commun. 326, 349–354
18. Sanchez-Laorden, B. L., Sanchez-Mas, J., Martinez-Alonso, E.,Martinez-Menarguez, J. A., Garcia-Borron, J. C., and Jimenez-Cervantes, C. (2006) Dimerization of the human melanocortin1 receptor: functional consequences and dominant-negativeeffects. J. Invest. Dermatol. 126, 172–181
19. Imoto, K., Kobayashi, N., Katsumi, S., Nishiwaki, Y., Iwamoto,T. A., Yamamoto, A., Yamashina, Y., Shirai, T., Miyagawa, S.,Dohi, Y., et al. (2002) The total amount of DNA damagedetermines ultraviolet-radiation-induced cytotoxicity after uni-formor localized irradiation of human cells. J. Invest. Dermatol.119, 1177–1182
20. Rouzaud, F., Kadekaro, A. L., Abdel-Malek, Z. A., and Hearing,V. J. (2005) MC1R and the response of melanocytes to ultravi-olet radiation. Mutat. Res. 571, 133–152
21. Chakraborty, A. K., Funasaka, Y., Slominski, A., Bolognia, J.,Sodi, S, Ichihashi, M., and Pawelek, J. M. (1999) UV light andMSH receptors. Ann. N. Y. Acad. Sci. 885, 100–116
22. Tadokoro, T., Kobayashi, N., Zmudzka, B. Z., Ito, S., Waka-matsu, K., Yamaguchi, Y., Korossy, K. S., Miller, S. A., Beer, J. Z.,and Hearing, V. J. (2003) UV-induced DNA damage andmelanin content in human skin differing in racial/ethnicorigin. FASEB J. 17, 1177–1179
23. Halder, R. M., and Bridgeman-Shah, S. (1995) Skin cancer inAfrican Americans. Cancer 75, 667–673
24. Preston, D. S., and Stern, R. S. (1992) Nonmelanoma cancers ofthe skin. N. Engl. J. Med. 327, 1649–1662
25. Kricker, A, Armstrong, B. K., and English, D. R. (1994) Sunexposure and non-melanocytic skin cancer. Cancer Causes Control5, 367–392
26. Scott, M. C., Suzuki, I., and Abdel-Malek, Z. A. (2002) Regula-tion of the human melanocortin 1 receptor expression inepidermal melanocytes by paracrine and endocrine factors andby ultraviolet radiation. Pigment Cell Res. 15, 433–439
27. Tan, C. P., McKee, K. K., Weinberg, D. H., MacNeil, T., Palyha,O. C., Feighner, S. D., Hreniuk, D. L., Van Der Ploeg, L. H.,MacNeil, D. J., and Howard, A. D. (1999) Molecular analysis ofa new splice variant of the human melanocortin-1 receptor.FEBS Lett. 451, 137–141
28. Siegrist, W., and Eberle, A. N. (1986) In situ melanin assay forMSH using mouse B16 melanoma cells in culture. Anal. Biochem.159, 191–197
29. Rouzaud, F., Annereau, J. P., Valencia, J. C., Costin, G. E., andHearing, V. J. (2003) Regulation of melanocortin 1 receptorexpression at the mRNA and protein levels by its natural agonistand antagonist. FASEB J. 17, 2154–2156
30. Tadokoro, T., Yamaguchi, Y., Batzer, J., Coelho, S. G., Zmudzka,B. Z., Miller, S. A., Wolber, R., Beer, J. Z., and Hearing, V. J.(2005) Mechanisms of skin tanning in different racial/ethnicgroups in response to ultraviolet radiation. J. Invest. Dermatol.124, 1326–1332
31. Jimenez, M., Maloy, W. L., and Hearing, V. J. (1989) Specificidentification of an authentic clone for mammalian tyrosinase.J. Biol. Chem. 264, 3397–3403
32. Bohm, M., Metze, D., Schulte, U., Becher, E., Luger, T. A., andBrzoska, T. (1999) Detection of melanocortin-1 immunoreactivityin human skin cells in culture and in situ. Exp. Dermatol. 8, 453–461
33. Bohm, M., Brzoska, T., Schulte, U., Schiller, M., Kubitscheck,U., and Luget, T. A. (1999) Characterization of a polyclonalantibody against the human melanocortin-1 receptor. Ann.N. Y. Acad. Sci. 885, 372–382
34. Pawelek, J. M., Chakraborty, A. K., Osber, M. P., Orlow, S. J.,Min, K. K., Rosenzweig, K. E., and Bolognia, J. L. (1992)Molecular cascades in UV-induced melanogenesis: a central rolefor melanotropins? Pigment Cell Res. 5, 348–356
35. Im, S., Moro, O., Peng, F., Medrano, E. E., Cornelius, J.,Babcock, G., Nordlund, J. J., and Abdel-Malek, Z. A. (1998)Activation of the cyclic AMP pathway by alpha-melanotropinmediates the response of human melanocytes to ultraviolet Bradiation. Cancer Res. 58, 47–54
36. Robbins, L. S., Nadeau, J. H., Johnson, K. R., Kelly, M. A.,Roselli-Rehfuss, L., Baack, E., Mountjoy, K. G., and Cone, R. D.(1993) Pigmentation phenotypes of variant extension locusalleles result from point mutations that alter MSH receptorfunction. Cell 72, 827–834
37. Klungland, H., Vage, D. I., Gomez-Raya, L., Adalsteinsson, S.,and Lien, S. (1995) The role of melanocyte-stimulating hor-mone (MSH) receptor in bovine coat color determination.Mamm. Genome 6, 636–639
38. Newton, J. M., Wilkie, A. L., He, L., Jordan, S.A., Metallinos,D. L., Holmes, N. G., Jackson, I. J., and Barsh, G. S. (2000)Melanocortin 1 receptor variation in the domestic dog. Mamm.Genome 11, 24–30
E1349IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM

39. Rouzaud, F., Martin, J., Gallet, P. F., Delourme, D., Goulemot-Leger, V., Amigues, Y., Menissier, F., Leveziel, H., Julien, R., andOulmouden, A. (2000) A first genotyping assay of French cattlebreeds based on a new allele of the extension gene encoding themelanocortin-1 receptor (Mc1r). Genet. Sel. Evol. 32, 511–520
40. Rana, B. K., Hewett-Emmett, D., Jin L., Chang, B. H., Sam-buughin, N., Lin, M., Watkins, S., Bamshad, M., Jorde, L. B., andRamsay, M. (1999) High polymorphism at the human melano-cortin 1 receptor locus. Genetics 151, 1547–1557
41. Jimenez-Cervantes, C., Germer, S., Gonzalez, P., Sanchez, J.,Sanchez, C. O., and Garcia-Borron, J. C. (2001) Thr40 andMet122 are new partial loss-of-function natural mutations of thehuman melanocortin 1 receptor. FEBS Lett. 508, 44–48
42. Jimenez-Cervantes, C., Olivares, C., Gonzalez, P., Morandini, R.,Ghanem, G., and Garcia-Borron, J. C. (2001) The Pro162variant is a loss-of-function mutation of the human melanocor-tin 1 receptor gene. J. Invest. Dermatol. 117, 156–158
43. Box, N. F., Duffy, D. L., Irving, R. E., Russell, A., Chen, W.,Griffyths, L. R., Parsons, P. G., Green, A. C., and Sturm, R. A.(2001) Melanocortin-1 receptor genotype is a risk factor forbasal and squamous cell carcinoma. J. Invest. Dermatol. 116,224–229
44. Sturm, R. A., Teasdale, R. D ., and Box, N. F. (2001) Humanpigmentation genes: identification, structure and consequencesof polymorphic variation. Gene 277, 49–62
45. Scott, M. C., Wakamatsu, K., Ito, S., Kadekaro, A. L., Kobayashi,N., Groden, J., Kavanagh, R., Takakuwa, T., Virador, V., Hear-ing, V. J., et al. (2002) Human melanocortin 1 receptor variants,receptor function and melanocyte response to UV radiation.J. Cell Sci. 115, 2349–2355
46. John, P. R., and Ramsay, M. (2002) Four novel variants in MC1Rin red-haired South African individuals of European descent:S83P, Y152X, A171D, P256S. Hum. Mutat. 19, 461–462
47. Lu, D., Willard, D., Patel, I. R., Kadwell, S., Overton, L., Kost, T.,Luther, M., Chen, W., Woychik, R. P., and Wilkison, W. O.(1994) Agouti protein is an antagonist of the melanocytestimulating- hormone receptor. Nature 371, 799–802
48. Little, C. C. (1957) The Inheritance of Coat Color in Dogs, Howell,New York
49. Searle, A. G. (1968) Comparative Genetics of Coat Colour inMammals. Academic Press, New York
50. Silvers, W. K., and Russel, E. S. (1955) An experimental ap-proach to action of genes at the agouti locus in mouse. J. Exp.Zool. 130, 199–220
51. Silvers, W. K. (1979) The Coat Colors of Mice: A Model forMammalian Gene Action and Interaction. Springer-Verlag, Basel,Switzerland
52. Bonilla, C., Boxill, L. A., Donald, S. A., Williams, T., Sylvester,N., Parra, E. J., Dios, S., Norton, H. L., Shriver, M. D., andKittles, R. A. (2005) The 8818G allele of the agouti signalingprotein (ASIP) gene is ancestral and is associated with darkerskin color in African Americans. Hum. Genet. 116, 402–406
53. Everts, R. E., Rothuizen, J., and van Oost, B. A. (2000) Identifi-cation of a premature stop codon in the melanocyte-stimulatinghormone receptor gene (MC1R) in Labrador and Goldenretrievers with yellow coat colour. Anim. Genet. 31, 194–199
54. Eason, M. G., Jacinto, M. T., Theiss, C. T., and Liggett, S. B.(1994) The palmitoylated cystein of the cytoplasmic tail of alpha2A-adrenergic receptors confers subtype-specific agonist-pro-moted downregulation. Proc. Natl. Acad. Sci. U. S. A. 91, 11178–11182
55. Kawate, N., and Menon, K. M. (1994) Palmitoylation of lutein-izing hormone/human choriogonadotropin receptors in trans-fected cells. Abolition of palmitoylation by mutation of Cys-621and Cys-622 residues in the cytoplasmic tail increases ligand-induced internalization of the receptor. J. Biol. Chem. 269,30651–30658
56. Schulein, R., Liebenhoff, U., Muller, H., Birnbaumer, M., andRosenthal, W. (1996) Properties of the human arginine vaso-pressin V2 receptor after site-directed mutagenesis of its puta-tive palmitoylation site. Biochem. J. 313, 611–616
57. Tanaka, K., Nagayama, Y., Nishihara, E., Namba, H., Yamashita,S., and Niwa, M. (1998) Palmitoylation of human thyrotropinreceptor: slower intracellular trafficking of the palmitoylation-defective mutant. Endocrinology 139, 803–806
58. Irani, B. G., Holder, J. R., Todorovic, A., Wilczynski, A. M.,Joseph, C. G., Wilson, K. R., and Haskell-Luevano, C. (2004)Progress in the development of melanocortin receptor selectiveligands. Curr. Pharm. Des. 10, 3443–3479
Received for publication March 3, 2006.Accepted for publication April 10, 2006.
E1350 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal

The FASEB Journal • FJ Express Summary
Regulation of constitutive and UVR-induced skinpigmentation by melanocortin 1- receptor isoforms
Francois Rouzaud,*,†,1 Gertrude-E. Costin,* Yuji Yamaguchi,* Julio C. Valencia,*Werner F. Berens,* Kevin G. Chen,* Toshihiko Hoashi,* Markus Bohm,§
Zalfa A. Abdel-Malek,¶ and Vincent J. Hearing**Laboratory of Cell Biology, National Cancer Institute, National Institutes of Health, Bethesda,Maryland, USA; †Department of Medicinal Chemistry, University of Florida, Gainesville, Florida,USA; §Department of Dermatology and Ludwig Boltzmann Institute for Cell Biology andImmunobiology of the Skin, University of Munster, Munster, Germany; and ¶Department ofDermatology, University of Cincinnati College of Medicine, Cincinnati, Ohio, USA
To read the full text of this article, go to http://www.fasebj.org/cgi/doi/10.1096/fj.05-5922fje
UV radiation (UVR) is among the most ubiquitousagents in the environment, and humans are inevitablyexposed to it. Constitutive skin pigmentation dramati-cally affects the incidence of sun-induced skin cancer,and the photoprotective function of melanin in theskin is highly significant. Since epidermal melanocytesrespond to UVR by increasing their expression of�-MSH which up-regulates the expression of MC1R andconsequently enhances the response of melanocytes tomelanocortins, we studied the expression of MC1R inskin of different ethnic origins and in primary normalhuman melanocytes (NHM) displaying a large range ofpigmentation phenotypes.
PRINCIPAL FINDINGS
1. Detection and characterization of MC1R isoformsexpressed by NHM
Western blot was used to detect MC1R expression in 15NHM cultures differing in their melanin content. Im-munodetection by �PEP19, a MC1R-specific antibody(Ab), revealed three patterns of expression. Pattern 1had a single band at 60 kDa whereas pattern 2 revealedtwo sets of doublet bands: one at 30 and 36 kDa and theother at �60 and 70 kDa. Pattern 3 showed only oneband at 30 kDa and one at 60 kDa, and was the mostcommon pattern. An 125I-�MSH labeling study helpeddetermine that all those bands can bind �-MSH andthat the 60 and 70 kDa bands represent a multi-MC1Rcomplex covalent structure. To characterize at thenucleotide level the isoforms depicted in the Westernblots, RACE-polymerase chain reaction (PCR) amplifi-cations were performed using two distinct adapter-ligated cDNA libraries for each NHM culture. 5� RACE-PCR analysis revealed only one 5�untranslated region(5�UTR) of 547 bp identical for all NHM. The 3�RACE-PCR investigations revealed the typical 954 nt
monoexon with a 762 nt 3�UTR, as well as a smallerfragment corresponding to a 1053 nt coding regionwith a 283 nt 3�UTR. The sequence translation indi-cates that the second transcript encodes a 350 aminoacid protein, which differs from the typical 317 aminoacid MC1R by the replacement of the last three resi-dues with a new 36 amino acid C terminus sequence.Therefore, we named the first transcript MC1R317 andthe second transcript MC1R350. A software-based pre-diction of the secondary structure of the MC1R iso-forms indicated that both isoforms have the typicalseven transmembrane domain profile, with the car-boxyl tail potentially anchored in the melanocyte mem-brane via the C315 palmitoylation. However, MC1R350harbors a total of five cysteines in its longer C-tail, eachof them being a potential target for palmitic acidfixation to the membrane.
2. MC1R expression and UV response in skinfrom different ethnic origins
MC1R350 was detected in paraffin skin sections using�PEP20, a new rabbit polyclonal antibody (pAb) weraised against the specific C terminus of that isoform.Immunofluorescence measurements indicated similarlow levels of MC1R350 expression in Caucasian andAsian skin (Fig. 1A, B), and a significant 25% higherlevel in African American skin. Due to the structuralcharacteristics of the two isoforms, it is not possible todesign an Ab that would recognize MC1R317 only.Therefore, any Ab that recognizes MC1R317 will alsodetect de facto MC1R350, revealing the cumulativeexpression of the two isoforms. Therefore, �PEP20 andanother previously reported MC1R Ab (N2–18) were
1 Correspondence: University of Florida Department ofMedicinal Chemistry, Rm. P5–26, 1600 SW Archer Rd.,Gainesville, FL 32610, USA. [email protected]
doi: 10.1096/fj.06-5922fje
19270892-6638/06/0020-1927 © FASEB

used to monitor respectively MC1R350 and total MC1Rexpression 7 min, 1 day, and 7 days after irradiation,with 1 minimal erythemal dose (MED) of UVR. At 1 dayand 7 days after irradiation, a significant increase wasdetected by staining with N2–18, which targets the totalMC1R corresponding to the sum of MC1R350 andMC1R317 (Fig. 1C). MC1R350 expression did notsignificantly vary after UV irradiation in any of theethnic groups examined (Fig. 1D, E). Therefore, weattribute the increase detected by N2–18 to MC1R317.
3. Influence of �MSH on MC1R expressionin cultured NHM
It has been known for a long time that �MSH canmimic the melanogenic effects of UVR. Confocal im-munofluorescence and Western blot using SC-N19 (acommercially available MC1R Ab) and �PEP19 as wellas semiquantitative real-time RT-PCR were used toassess changes in MC1R isoforms expression in NHMon �-MSH treatment. Fluorescence index measure-ments indicated that the increase in expression is muchgreater for MC1R317 than for MC1R350. Western blotprofiles revealed that in most of the NHM tested,MC1R317 levels were increased for both the monomerand the multicomplex forms. At the mRNA level, all
NHM cultures tested but one increased their totalamount of MC1R and all NHM cultures, but oneshowed a higher relative amount of MC1R317 whenstimulated by �-MSH, indicating that MC1R317 is amore suitable answer to an increased melanogenesis.
4. Effects of MC1R isoforms on the melaninbiosynthesis pathway
Our real-time RT-PCR study shows a good correlationbetween the total level of MC1R mRNA and the totalmelanin content in the 10 different NHM culturestested, and that correlation is even better if we considerMC1R317 only. However, MC1R350 expression showsan inverse correlation with the total melanin content,suggesting that MC1R317 acts as a positive factor to-ward the total melanin content synthesis whereas theMC1R350 isoform could have an opposite action. Tran-sient transfections of NHM with MC1R317 or MC1R350show that while MC1R317 increases expression of MITFand tyrosinase, MC1R350 dramatically reduces them(Fig. 2). These results show the negative impact ofMC1R350 on the intracellular signal transduced to themelanin synthesis pathway.
Figure 1. A) Immunodetection of MC1R350 by�PEP20 in paraffin-embedded skin sectionsfrom Caucasian (C), Asian (A), and AfricanAmerican (AA) donors. Three different subjectswere tested for each ethnic group. Representa-tive fluorescence images of MC1R350 detectedby FITC-coupled �PEP20 (green, left column)and nuclei detected by PI (red, right column) innonirradiated skin are shown for one subjectfrom each ethnic group. B) The related fluores-cence index values obtained with the ScionImage program are displayed on the graph inpanel C. Typical images of immunodetection byN2–18 of total MC1R expression after irradia-tion with 1 MED UV dose. A positive signal wasobtained for the three African American sam-ples only. Melanocytes had a significant increasein expression 1 and 7 days after irradiation. D)Immunodetection of MC1R350 by �PEP20 inparaffin-embedded skin sections of three Asian,Caucasian, and African American subjects. Typ-ical fluorescence images of MC1R350 detectedby Texas Red (red, left column) and nucleidetected by 4�,6�-diam idino-2-phenylidole(blue, right column) in UV-irradiated skin. Onerepresentative Asian specimen is displayed. E)Graph of fluorescence index measurements in 3Asian, Caucasian, and African American sub-jects after immunodetection of MC1R350 by�PEP20. Data are shown for unirradiated (UI)skin as well as 7 min, 1 day, and 7 days after 1MED UV exposure.
1928 Vol. 20 September 2006 ROUZAUD ET AL.The FASEB Journal
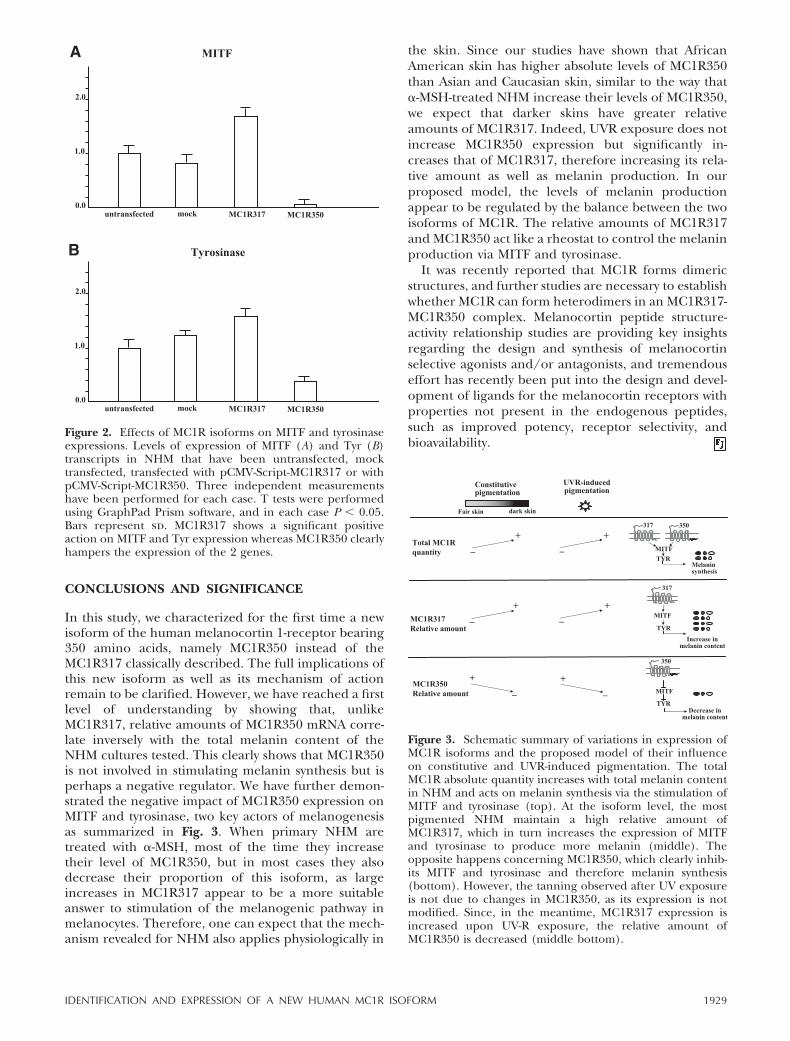
CONCLUSIONS AND SIGNIFICANCE
In this study, we characterized for the first time a newisoform of the human melanocortin 1-receptor bearing350 amino acids, namely MC1R350 instead of theMC1R317 classically described. The full implications ofthis new isoform as well as its mechanism of actionremain to be clarified. However, we have reached a firstlevel of understanding by showing that, unlikeMC1R317, relative amounts of MC1R350 mRNA corre-late inversely with the total melanin content of theNHM cultures tested. This clearly shows that MC1R350is not involved in stimulating melanin synthesis but isperhaps a negative regulator. We have further demon-strated the negative impact of MC1R350 expression onMITF and tyrosinase, two key actors of melanogenesisas summarized in Fig. 3. When primary NHM aretreated with �-MSH, most of the time they increasetheir level of MC1R350, but in most cases they alsodecrease their proportion of this isoform, as largeincreases in MC1R317 appear to be a more suitableanswer to stimulation of the melanogenic pathway inmelanocytes. Therefore, one can expect that the mech-anism revealed for NHM also applies physiologically in
the skin. Since our studies have shown that AfricanAmerican skin has higher absolute levels of MC1R350than Asian and Caucasian skin, similar to the way that�-MSH-treated NHM increase their levels of MC1R350,we expect that darker skins have greater relativeamounts of MC1R317. Indeed, UVR exposure does notincrease MC1R350 expression but significantly in-creases that of MC1R317, therefore increasing its rela-tive amount as well as melanin production. In ourproposed model, the levels of melanin productionappear to be regulated by the balance between the twoisoforms of MC1R. The relative amounts of MC1R317and MC1R350 act like a rheostat to control the melaninproduction via MITF and tyrosinase.
It was recently reported that MC1R forms dimericstructures, and further studies are necessary to establishwhether MC1R can form heterodimers in an MC1R317-MC1R350 complex. Melanocortin peptide structure-activity relationship studies are providing key insightsregarding the design and synthesis of melanocortinselective agonists and/or antagonists, and tremendouseffort has recently been put into the design and devel-opment of ligands for the melanocortin receptors withproperties not present in the endogenous peptides,such as improved potency, receptor selectivity, andbioavailability.
Figure 2. Effects of MC1R isoforms on MITF and tyrosinaseexpressions. Levels of expression of MITF (A) and Tyr (B)transcripts in NHM that have been untransfected, mocktransfected, transfected with pCMV-Script-MC1R317 or withpCMV-Script-MC1R350. Three independent measurementshave been performed for each case. T tests were performedusing GraphPad Prism software, and in each case P � 0.05.Bars represent sd. MC1R317 shows a significant positiveaction on MITF and Tyr expression whereas MC1R350 clearlyhampers the expression of the 2 genes.
Figure 3. Schematic summary of variations in expression ofMC1R isoforms and the proposed model of their influenceon constitutive and UVR-induced pigmentation. The totalMC1R absolute quantity increases with total melanin contentin NHM and acts on melanin synthesis via the stimulation ofMITF and tyrosinase (top). At the isoform level, the mostpigmented NHM maintain a high relative amount ofMC1R317, which in turn increases the expression of MITFand tyrosinase to produce more melanin (middle). Theopposite happens concerning MC1R350, which clearly inhib-its MITF and tyrosinase and therefore melanin synthesis(bottom). However, the tanning observed after UV exposureis not due to changes in MC1R350, as its expression is notmodified. Since, in the meantime, MC1R317 expression isincreased upon UV-R exposure, the relative amount ofMC1R350 is decreased (middle bottom).
1929IDENTIFICATION AND EXPRESSION OF A NEW HUMAN MC1R ISOFORM





![EVcc`c Wc`^ 2W hZ]] SV UVR]e+ 45D - Daily Pioneer](https://static.fdokumen.com/doc/165x107/632551385c2c3bbfa803227c/evccc-wc-2w-hz-sv-uvre-45d-daily-pioneer.jpg)














