
regulation of cerebrospinal fluid bicarbonate - NCBI
18
J. Phy8iol. (1977), 267, pp. 411-428 411 With 7 text-figura Prnted in Great Britain REGULATION OF CEREBROSPINAL FLUID BICARBONATE BY THE CAT CHOROID PLEXUS BY RUSSELL P. HUSTED AND DONAL J. REED From the Department of Pharmacology, University of Utah College of Medicine, Salt Lake City, Utah 84132, U.S.A. (Received 25 August 1976) SUMMARY 1. The regulation of cerebrospinal fluid (c.s.f.) bicarbonate concentra- tion was studied using the cat choroid plexus isolated in a chamber in sitU. 2. Decreases in plasma bicarbonate concentration caused relatively small changes in the c.s.f. bicarbonate concentration. 3. Alterations in c.s.f. bicarbonate concentration (c.s.f. HCO3- = 9 or 28 m-equiv/l.) were countered by changes in the bicarbonate concentra- tion of the fluid produced by the plexus or in the rate of bicarbonate transport which returned c.s.f. bicarbonate towards normal. 4. There was significant regulation of pH in the choroid plexus fluid during hypocapnia and hypercapnia. 5. Alterations of plasma acid-base status did not significantly alter the potential difference across the choroid plexus. However, the potential difference increased when c.s.f. bicarbonate was increased and decreased when c.s.f. bicarbonate was decreased. 6. The data indicate that the bicarbonate concentration in the c.s.f. is actively regulated by the choroid plexus during acid-base disturbances occurring either systemically or in the c.s.f. INTRODUCTION The pH of cerebrospinal fluid (c.s.f.) is subject to a greater degree of homoeostatic control than is plasma, particularly during metabolic acid-base disturbances (Fencl, Miller, & Pappenheimer, 1966; Hornbein & Pavlin, 1975; Pavlin & Hornbein, 1975a, b, c; Ponten & Siesj6, 1967). This relative stability of the composition of the c.s.f. is not only present in the adult animal but is also apparent in the neonatal rat (Johanson, Woodbury & Withrow, 1976) and the foetal and neonatal lamb (Hodson, Fenner, Brumley, Chernick & Avery, 1968). The mechanisms and sites
-
Upload
khangminh22 -
Category
Documents
-
view
13 -
download
0
Transcript of regulation of cerebrospinal fluid bicarbonate - NCBI

J. Phy8iol. (1977), 267, pp. 411-428 411With 7 text-figuraPrnted in Great Britain
REGULATION OF CEREBROSPINAL FLUID BICARBONATEBY THE CAT CHOROID PLEXUS
BY RUSSELL P. HUSTED AND DONAL J. REEDFrom the Department of Pharmacology, University of UtahCollege of Medicine, Salt Lake City, Utah 84132, U.S.A.
(Received 25 August 1976)
SUMMARY
1. The regulation of cerebrospinal fluid (c.s.f.) bicarbonate concentra-tion was studied using the cat choroid plexus isolated in a chamberin sitU.
2. Decreases in plasma bicarbonate concentration caused relativelysmall changes in the c.s.f. bicarbonate concentration.
3. Alterations in c.s.f. bicarbonate concentration (c.s.f. HCO3- = 9or 28 m-equiv/l.) were countered by changes in the bicarbonate concentra-tion of the fluid produced by the plexus or in the rate of bicarbonatetransport which returned c.s.f. bicarbonate towards normal.
4. There was significant regulation of pH in the choroid plexus fluidduring hypocapnia and hypercapnia.
5. Alterations of plasma acid-base status did not significantly alterthe potential difference across the choroid plexus. However, the potentialdifference increased when c.s.f. bicarbonate was increased and decreasedwhen c.s.f. bicarbonate was decreased.
6. The data indicate that the bicarbonate concentration in the c.s.f. isactively regulated by the choroid plexus during acid-base disturbancesoccurring either systemically or in the c.s.f.
INTRODUCTION
The pH of cerebrospinal fluid (c.s.f.) is subject to a greater degree ofhomoeostatic control than is plasma, particularly during metabolicacid-base disturbances (Fencl, Miller, & Pappenheimer, 1966; Hornbein &Pavlin, 1975; Pavlin & Hornbein, 1975a, b, c; Ponten & Siesj6, 1967).This relative stability of the composition of the c.s.f. is not only presentin the adult animal but is also apparent in the neonatal rat (Johanson,Woodbury & Withrow, 1976) and the foetal and neonatal lamb (Hodson,Fenner, Brumley, Chernick & Avery, 1968). The mechanisms and sites

R. F. HUSTED AND D. J. REED
of regulation of c.s.f. acid-base status still remain the subject of con-siderable controversy (see Siesj6, 1972).
It has been suggested that the concentration of bicarbonate andhydrogen ion in the c.s.f. is determined passively by the concentration andpotential gradients that exist between c.s.f. and plasma (Forster, Dempsy&; Chosy, 1975; Hornbein & Pavlin, 1975; Kjallquist & Siesjd, 1968;Pavlin & Hornbein, 1975 a, b, c). The production of organic buffer sub-stances (principally lactate and ammonia) by the brain has been putforth as a factor contributing to the stability of c.s.f. pH (Hasan &Kazemi, 1976; Kazemi, Shore, Shih & Shannon, 1973). Active transportof, hydrogen or bicarbonate ions or both, across the blood-brain andblood-c.s.f. barriers has also been proposed as a mechanism contributingto the control of c.s.f. acid-base status (Pappenheimer, 1970; Severinghaus,Mitchell, Richardson & Singer, 1963).The in 8itU isolated choroid plexus preparation developed by Miner
& Reed (1972) provides a unique system in which to study the role of thechoroid plexus in the regulation of c.s.f. composition. In a previous studyit was determined that the choroid plexus actively transported potassiumduring alterations of c.s.f. and plasma potassium concentrations (Husted &Reed, 1976). The present study was designed to study the role of thechoroid plexus in the regulation of c.s.f. bicarbonate concentration duringalterations in cerebrospinal fluid and plasma acid-base status.
METHODS
Experimental. Adult cats of either sex, body weight 2 7-6 kg, were anaesthetizedwith pentobarbitone sodium, 32 mg/kg, i.P. A tracheotomy was performed and theanimal was maintained on artificial respiration throughout the experiment. Thefemoral artery was cannulated for monitoring blood pressure and for collectingblood samples. The femoral vein was cannulated to permit intravenous administra-tion ofsupplemental anaesthetic. Body temperature was maintainednear normalwithan electric heating pad. The choroid plexus was isolated in a chamber in situ asdescribed in detail by Miner & Reed (1972). In brief, the method requires an extensiveunilateral craniotomy, removal of the cerebral cortex overlying the left lateralventricle and exposure of the plexus within the lateral ventricle. An appropriatesegment of the plexus is then separated from its underlying connective tissueattachments and isolated within a chamber with the blood and nerve supply intact.
Collection of chamber fluid. After completion of the isolation procedure all thefluid in the chamber was removed. Fluid secreted by the choroid plexus was allowedto collect in the chamber under a layer of liquid petrolatum that was added toeliminate fluid evaporation. In some experiments 30 or 50 p1. artificial c.s.f. (de-scribed below) was added to the chamber at the start of the collection period. At theend of the collection period all the fluid beneath the liquid petrolatum (designatedchamber fluid) was removed from the chamber and the collection procedure repeatedas long as the preparation was stable, i.e. adequate blood pressure, plexus bloodflow and fluid secretion rate. Collection periods were either 15 or 30 min depending
412

C.S.F. ACID-BASE REGULATION 413
upon the rate of fluid secretion by the plexus and the volume of the chamber thatwas used. The chamber fluid includes the fluid that was secreted by the choroidplexus and the artificial c.s.f. added at tl'e start of the collection period, if any. Anarterial blood sample was obtained simultaneously with the c.s.f. sample.
Arti~ficial cerebroepinal fluids. The artificial c.s.f. described by Merlis (1940) wasused as the normal-bicarbonate c.s.f. The low-HCO3 and high-HCO3 c.s.f.s wereprepared by interchanging NaCl and NaHCO3 in the Merlis artificial c.s.f. such thatthe final bicarbonate concentrations were approximately 0-5 and 1-5 times thenormal concentration, respectively.
Reapiration. Normal arterial Poo, (29.6 + 0-5 mmHg, measured in sixty-six cats assoon as possible after anaesthesia) was maintained by artificial respiration withroom air at 24 breaths/min and a tidal volume of 40 ml. Low Pco, was achieved byincreasing tidal volume to 80 ml. room air. Hypercapnia was produced by hyper-ventilation (80 ml. tidal volume) with a mixture of either 5% CO2, 20% 0°2, 75% N2or 10% COV, 20% 02, 70% N2.
Potential measurements. Micro-electrodes were pulled and filled by boiling in asolution containing 3M-KCl and 0 01 M-HCl. Acidification of the KCl has been foundto abolish tip potentials (Williams, Withrow & Woodbury, 1971). The micro-electrode was placed into the fluid in the chamber. The indifferent electrode was ahypodermic needle placed in the platysma muscle of the lower neck.
Analyses. Plasma pH and PCO2 were measured with an Instrumentation Labora-tory blood gas apparatus (model 213). The total CO2 of 25 #tl. chamber fluid wasdetermined by gas chromatography (Fisher Clinical Gas Partitioner). A 20 u1.sample (chamber fluid or plasma) was diluted 1: 200 with Li2SO4 solution for deter-mination of sodium, potassium and chloride. Sodium and potassium were deter-mined with an Instrumentation Laboratory, model 143, internal standard flamephotometer. Chloride was determined with a Buchler-Cotlove chloridometer.Protein concentrations were determined by the method of Pesce & Strande (1973) on20 ju. samples of chamber fluid and a comparable volume of a 1:20 dilution ofplasma.
Calculation. Plasma bicarbonate concentration was determined from plasma pHand PCo2 by use of the Henderson-Hasselbalch equation. The PCO2 of c.s.f. wascalculated from the arterial value as described by Ponten & Siesj6 (1966). The pHand bicarbonate concentration of c.s.f. were calculated from the c.s.f. total CO2 andPC02 using values of S (CO2 solubility) and pK' determined by Mitchell, Herbert &Carman (1965). Concentrations of protein and electrolytes in the newly formed c.s.f.were calculated by subtracting the amount of the substance initially added to thechamber in the artificial c.s.f. from the total amount in the chamber fluid anddividing by the volume of new c.s.f. secreted during the collection period. Thus it isassumed that any change in chamber fluid concentration was due to the addition ofthe measured volume of new fluid of the calculated composition. This assumptionmay not be absolutely correct but it is not important for the purposes of thispresentation whether the change in the quantity of the substance in the chamber isexpressed as a flux or as a change in concentration in the new fluid. The formulathat was used is:
new c.s.f. (X) - (X)6F VC, - (X)AVAVCF A
where (X) is the concentration of the substance in the fluid, V is the fluid volumeand the subscripts CF and A refer to chamber fluid and added artificial c.s.f.,respectively. The regulation of c.s.f. and plasma pH during respiratory acid-basechanges was quantitated as described by Siesj6 (1971).

R. F. HUSTED AND D. J. REED
RESULTS
The wet weight of the portion of choroid plexus in the chamber variedfrom 2 to 6 mg while the rate of fluid accumulation in the chamberranged from 0 9 to 2-4 1d. min-. Thus the rate of fluid secretion by thechoroid plexus was about 0-4 ul. min' . mg-' as has been previouslyreported (Husted & Reed, 1976; Miner & Reed, 1972). The rate of c.s.f.formation was unaffected by changes in Pco, in the range from 10-7 to87-3 mmHg.The protein content of the new c.s.f. during normocapnia was similar
to that reported previously using this technique (Husted & Reed, 1976;Miner & Reed, 1972). The protein concentration of the c.s.f. remainedreasonably constant throughout any one experiment, although the firstchamber fluid sample often had a slightly higher protein content thansubsequent samples. Hyperventilation and moderate hypercapnia (5 %C02) caused no change in the protein content of the formed fluid. The moresevere hypercapnia due to inhalation of 10% CO2 resulted in increases inprotein content to values that approached plasma protein levels and, onoccasion, caused frank bleeding into the chamber. Expansion of the brainvolume also occurred during CO2 inhalation and with 10% C02 the expan-sion was of a magnitude sufficient to displace the chamber and cause lossof the fluid that had accumulated.
Technical problems including the marked brain expansion and highprotein levels of the samples during severe hypercapnia caused us toabandon further studies with 10% C02 after only a few c.s.f. samples hadbeen obtained.
There was not a significant correlation of concentrations of Na, K, orCl in the chamber fluid with the concentration of these electrolytes inplasma within the ranges that were observed. However, no attempt wasmade in this study to alter the concentration of any of these ions in eitherfluid.The relationship of chamber fluid bicarbonate concentration to plasma
bicarbonate concentration is shown in Fig. 1. Each point on the graphrepresents the values for one chamber fluid sample and its correspondingplasma sample. The filled symbols represent samples taken when noartificial c.s.f. was added to the chamber and open symbols representsamples taken either 15 or 30 min after addition of the normal-HCO3 c.s.f.The range of plasma bicarbonate concentrations observed was due tovariations in this parameter between animals at the start of the experi-ment and to progressive metabolic acidosis in some animals during thecourse of the experiment. The continuous line was calculated by linearregression analysis of all the data points and has a slope of 0 129 + 0-16I
414
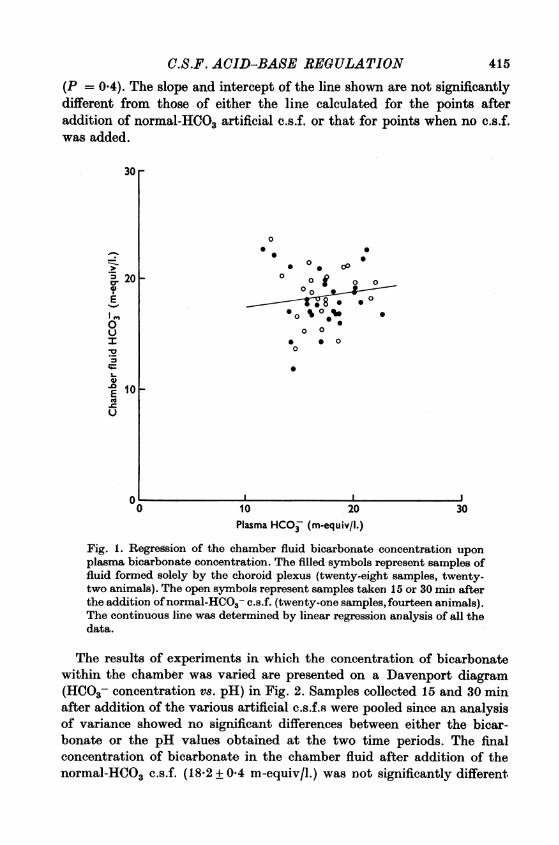
C.S.F. ACID-BASE REGULATION 415(P = 0.4). The slope and intercept of the line shown are not significantlydifferent from those of either the line calculated for the points afteraddition of normal-HCO3 artificial c.s.f. or that for points when no c.s.f.was added.
30
0
0> co~~~~~~
.r 20 _ ° O°0,E --8 E E °
O 0 000* SO0
._X
- 10
U
00 10 20 30
Plasma HCOj (m-equiv/l.)
Fig. 1. Regression of the chamber fluid bicarbonate concentration uponplasma bicarbonate concentration. The filled symbols represent samples offluid formed solely by the choroid plexus (twenty-eight samples, twenty-two animals). The open symbols represent samples taken 15 or 30 min afterthe addition ofnormal-HCO3- c.s.f. (twenty-one samples, fourteen animals).The continuous line was determined by linear regression analysis of all thedata.
The results of experiments in which the concentration of bicarbonatewithin the chamber was varied are presented on a Davenport diagram(HCO3- concentration vs. pH) in Fig. 2. Samples collected 15 and 30 minafter addition of the various artificial c.s.f.s were pooled since an analysisof variance showed no significant differences between either the bicar-bonate or the pH values obtained at the two time periods. The finalconcentration of bicarbonate in the chamber fluid after addition of thenormal-HCO3 c.s.f. (18-2 + 0-4 m-equiv/l.) was not significantly different.

R. F. HUSTED AND D. J. REEDfrom that of the added c.s.f. (18 m-equiv/1.) or the c.s.f. formed when noartificial c.s.f. was added to the chamber (18.4 + 0-5 m-equiv/l.). Afteraddition of the low-HCO3 c.s.f. the concentration of bicarbonate in thechamber increased significantly (P < 0.01) from 9 m-equiv/l. to a mean of14-8 + 0-4 m-equiv/l. The bicarbonate concentration of the fluid in thechamber decreased (P < 0.01) from 28 m-equiv/l. to 23-7 + 0-9 m-equiv/l.after the high-HCO3 c.s.f. was added.
35
30
25
1-1
> 20Cr
E15
0UI
10
5
7-0
PcO240
7-2 7-4 7-6pH
Fig. 2. Davenport diagram of the chamber fluid samples obtained afteraddition ofnormal-HCOG- c.s.f. (U, thirty-four samples, seventeen animals),low-HCO3- c.s.f. (*, seventeen samples, twelve animals), high-HCOG- c.s.f.(A, thirteen samples, nine animals) or no c.s.f. (@, thirty-eight samples,twenty-four animals). The open symbols represent the values for the low-HCO3- (C) and high-HCO3- (A) artificial c.s.f.s that were added to thechamber. The bars indicate one S.E. of mean.
Attempts to measure the intracellular potential of the choroid plexuscells were unsuccessful primarily due to the movements of the plexuswith respiration and the vascular pulse. Such movement prevented thetip of the electrode from remaining in a cell for sufficient time to allowan accurate measurement of the potential difference. During several ofthese attempts plexus capillaries were entered and the potential betweenchamber fluid and the capillaries ranged from 2 to 5 mV (chamber fluid
416

C.S.F. ACID-BASE REGULATION 417positive). There was not a significant effect of changes in plasma acid-basestatus on the potential difference measured between the c.s.f. in thechamber and the reference electrode. Since the absolute potential differencewas not measured in most experiments, the relationship of changes in
3 A
2 _
> AE A
0
0-23-1
i -2 /
-4-6 -4 -2 o 2 4 6
A chamber fluid HCO3 (m-equiv/1.)
Fig. 3. The correlation of the changes in bicarbonate concentration andpotential difference from their control values. Symbols as in Fig. 2. Thepoints are the mean of one to three determinations in each of five animals.The continuous line was calculated by linear regression analysis.
chamber fluid bicarbonate concentration to changes in potential differencefrom their respective control values is shown in Fig. 3. The mean changesin potential difference were - 1-9 +0O5 mV after addition of the low-HCO3 c.s.f. to the chamber and + P8+0 4 mV after the high-H003. Inthe experiments in which the potential difference was measured thecalculated concentration of bicarbonate in the new c.s.f. was 16*7 + 1*7 m-equiv/l. during the control periods. After addition of low-HCO3 c.s.f. thenew c.s.f. bicarbonate concentration increased to 23-4 + 1-8 m-equiv/l. Theconcentration of bicarbonate in the new c.s.f. was only 2-4 + 4-2 m-equiv/l.after the high-HCO3 c.s.f. was added to the chamber.
Figs. 4, 5 and 6 show the effects of hyperventilation (mean PCO2 =13*9 + 04 mmHg) and hypercapnia (mean PCO2 = 39*5 + 0 7 or 68-7 +7 0 mmHg) on c.s.f. and blood acid-base status. Normal-HCO3 c.s.f. wasadded to the chamber during all collection periods. Plasma Po2 was
R. F. HUSTED AND D. J. REEDgreater than 80 mmHg during all experimental periods. The % pHregulation (Siesjo, 1971) is a convenient method to quantitate the responseof the system to respiratory acid-base disturbances. It is defined as thedifference between the maximum change in pH and the observed change
100r
80 F
C.2
00
IC.
0
60 F
401-
20 F
0
II
0
41
2Time (hr)
I0
S
-443I
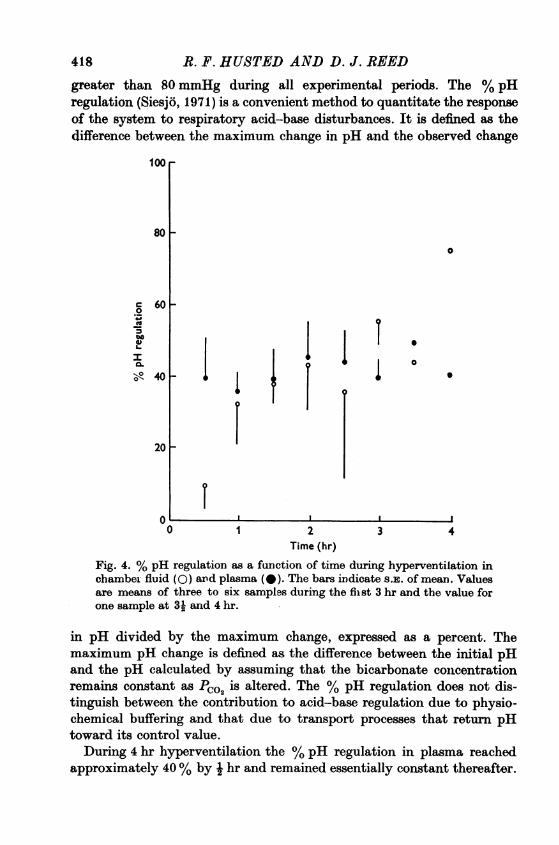
Fig. 4. % pH regulation as a function of time during hyperventilation inchamber fluid (0) and plasma (-). The bars indicate S.E. of mean. Valuesare means of three to six samples during the first 3 hr and the value forone sample at 3k and 4 hr.
in pH divided by the maximum change, expressed as a percent. Themaximum pH change is defined as the difference between the initial pHand the pH calculated by assuming that the bicarbonate concentrationremains constant as Pco2 is altered. The % pH regulation does not dis-tinguish between the contribution to acid-base regulation due to physio-chemical buffering and that due to transport processes that return pHtoward its control value.
During 4 hr hyperventilation the % pH regulation in plasma reachedapproximately 40% by i hr and remained essentially constant thereafter.
I I I
418

C.S.F. ACID-BASE REGULATION
On the other hand the % pH regulation in chamber fluid did not reach40% during the first hour of hyperventilation but appeared to increasethroughout the 4 hr period of study (Fig. 4).
Fig. 5 shows similar results that were obtained during hypercapnia.
100
80
0*Z; 60 -
goo~40¶
20 2
0 1 2 3 4Time (hr)
Fig. 5. % pH regulation as a function of time during inhalation of 5%(A, A) or 10% (O, *), CO2 in chamber fluid (A, E-I) and plasma (A, U).Chamber fluid values are the means oftwo samples and plasma the means offour samples during 5% C02. Values during 10% C02 are for single samples.
The % pH regulation in plasma during moderate hypercapnia wasapproximately 40 % from 1 to 4 hr although there may have been aslight increase with time. With 5 % C02 the % pH regulation in chamberfluid lagged behind that of plasma during the early time periods but hadreached almost complete regulation of chamber fluid pH by 4 hr. The % pHregulation of chamber fluid was similar during 10% C02 inhalation.However the regulation of plasma pH was much less during 10% C02than during 5% C02.
419
z6 PHY 267

R. F. HUSTED AND D. J. REED
The changes in bicarbonate concentration that occurred during altera-tion of plasma Pco, are shown in Fig. 6. In both hyperventilation andmoderate hypercapnia the changes in chamber fluid bicarbonate laggedbehind those of plasma but the chamber fluid and plasma changes in
6
4
Ia
0 Ioa
2 -
0V
"
I-.t
a
4 U1
-2k
-41-
-8
Ir
I
II
1t
Er IT
I I
0
8
I I I
0 1 2
Time (hr)
3 4
Fig. 6. Changes in bicarbonate concentration as a fiction of time duringhyperventilation (O. 0) 5%5 CO2 inhalation (A, A), or 100 0C2 inhala-tion (O.' U). Symbols and number of samples as in Figs. 4 and 5.
bicarbonate were similar during the last 2 hr of either disturbance. Theincrease in plasma bicarbonate was similar in both moderate and severehypercapnia. However, the increase in chamber fluid bicarbonate with10% 002 was much greater than that during 5 % C02 and greater thanthat occurring in plasma during either 5 or 10% C02 inhalation. It must be
o |420
-6 ~

C.S.F. ACID-BASE REGULATION 421mentioned that the chamber fluid samples obtained during 10%O02had amuch higher protein concentration than was normally accepted. Anincrease in chamber fluid protein concentration of the magnitude seen insevere hypercapnia was commonly found to be correlated with a decreasein the measured bicarbonate concentration. Therefore, it is possiblethat the measured change in chamber fluid bicarbonate with severe hyper-capnia underestimated the actual change that occurred.
150 F
.
40 0* .0.oa 00 00
%0 @00 00
0 0 0 4a0 0.o 0
130 I-
0 -o0
0
0
0
0
0
0o
0
0
0
0
0
a
0
@00 a
0
0
120 1-
0 0
0
1100 10 20 30
Chamber fluid HCO5 (m-equiv/l.)
Fig. 7. Chamber fluid chloride concentration as a function of chamber fluidbicarbonate concentration (114 samples, twenty-nine animals). The filledsymbols represent samples taken during hyperventilation. The open
symbols represent all other samples regardless of added c.s.f. and plasmacomposition. The continuous line has a slope of -1 and is intended merely as
a reference.16-2
140
0~:-
._
4,.0E
';-cU5
_L_a a I

R. F. HUSTED AND D. J. REED
Chamber fluid chloride is shown as a function of chamber fluid bi-carbonate in Fig. 7. All chamber fluid samples where both chloride andbicarbonate were measured are shown regardless of the composition ofthe added c.s.f. The filled symbols represent values obtained duringhyperventilation. The slope of the line shown is -1 and most of thevariability about the line is due to differences in the sum of chloride andbicarbonate in the chamber fluid of different animals. The slopes of thechloride-bicarbonate relationship for individual animals were calculatedand the mean slope was not significantly different from -1. This indicatesthat the decreases in bicarbonate were probably not due to altered con-centrations of glycolytic intermediates.
DISCUSSION
The primary objective of this study was to evaluate the role of thechoroid plexus in acid-base homoeostasis of the c.s.f. This was done bydetermining the effects of changing systemic or c.s.f. acid-base status onthe composition of fluid formed by the choroid plexus. The first questionthat had to be addressed was that of the concentration of bicarbonate inthe newly formed c.s.f. Calculation of bicarbonate based on charge balance(i.e. Na +K - Cl) had indicated that the plexus fluid bicarbonate con-centration was 5-7 m-equiv/l. greater than that of large cavity c.s.f.(Ames, Sakanoue & Endo, 1964; Miner & Reed, 1972). Analysis of data onthe kinetics of labelled bicarbonate movement from blood to c.s.f. ledVogh & Maren (1975) to speculate that the concentration of bicarbonate innewly formed c.s.f. might be as high as 72 m-equiv/l. The concentration ofbicarbonate was thought to be reduced subsequently by addition ofacidic metabolites from the brain tissue. In this study the concentrationof bicarbonate was determined in fluid formed by the choroid plexusisolated in a chamber in situ and therefore the c.s.f. formed had nocontact with non-choroidal tissue. The concentration of bicarbonatein the fluid collected from the chamber was approximately 18x5 m-equiv/l.when plasma acid-base status was normal for animals at this altitude(1350 m).In the present study hypercapnia had no effect on the rate of c.s.f.
production. In previous studies hypercapnia has been found to increase(Ames, Higashi & Nesbett, 1965), decrease (Evans, Reynolds, Reynolds &Saunders, 1976) or have no effect (Hochwald, Malhan & Brown, 1973) onc.s.f. formation. Cutler & Barlow (1966) and Lending, Slobody & Mestern(1961) found an increase in protein entry into brain and c.s.f. duringhypercapnia. The entry of sucrose into the c.s.f. was also increased byhypercapnia (Evans et al. 1976). Hochwald et al. (1973) found that c.s.f.
422

C.S.F. ACID-BASE REGULATIONprotein concentration increased in normal animals during hypercapniawhile in hydrocephalic animals there was no change in c.s.f. protein. Thedifference between normal and hydrocephalic cats was felt to be due to thedestruction of the choroid plexus during hydrocephalus. In the currentstudy the protein concentration of the plexus fluid increased during severehypercapnia to a much greater extent than was reported for bulk c.s.f. inthe studies cited above. Thus the choroid plexus may be the site ofincreased entry of protein into the c.s.f. in hypercapnia.When changes occurred in plasma bicarbonate concentration without
changes in plasma Pco, the change in the concentration of bicarbonatein the plexus fluid was very small with the best estimate from Fig. 1being 13% of the plasma change. Fencl et al. (1966) and Pont6n & Siesj6(1967) found that the change in the c.s.f. bicarbonate was 35% of thechange in plasma bicarbonate during metabolic acid-base disturbances.Pavlin & Hornbein (1975a, b) found that during metabolic acidosis ormetabolic alkalosis the change in c.s.f. bicarbonate was about 40 % aslarge as the corresponding change in plasma. The better regulation ofbicarbonate concentration in the freshly secreted fluid indicates that thecontrol of bicarbonate may be more efficient at the blood-c.s.f. barrier(choroid plexus) than at the blood-brain barrier (capillary-glial complex).It should also be noted that in the intact system, changes in the c.s.f.-blood potential difference occur that can account for most of the regulationof c.s.f. bicarbonate during metabolic acidosis and alkalosis (Pavlin &Hornbein, 1975 a, b). However, there are no comparable changes in thepotential difference in the isolated choroid plexus system. Thus, activetransport of bicarbonate may be involved in the maintenance of c.s.f.composition at the choroid plexus when plasma bicarbonate concentrationis altered.
Previous studies in other systems have shown no effect of c.s.f. acid-base status on the potential between c.s.f. and blood (Held, Fencl &Pappenheimer, 1964; Welch & Sadler, 1965). However, in the presentstudies alterations in bicarbonate concentration on the c.s.f. side of theisolated choroid plexus caused changes in the potential difference thatopposed correction of the acid-base disturbance. Despite the increasein the potential difference that occurred when high-HCO3 c.s.f. was addedto the chamber the choroid plexus decreased the concentration of bicar-bonate in the chamber and in some cases the total bicarbonate in thechamber was less at the end of the collection period than at the beginningwhile the rate of c.s.f. formation was unaltered from control. Constantc.s.f. formation with more rapid accumulation of bicarbonate was notedafter addition of the low-HCO3 c.s.f. even though the potential differencewas less than during control periods. It appears, therefore, that active
423

R. F. HUSTED AND D. J. REEDtransport of hydrogen or bicarbonate can occur in both directions acrossthe choroid plexus in response to alterations of c.s.f. bicarbonate duringa normal plasma acid-base state.A comparison of the time course of changes in plasma and c.s.f. bi-
carbonate concentration during altered ventilation reveals certain con-sistent differences. Plasma bicarbonate concentration decreased duringthe first 15 min (Nilsson & Busto, 1973; Ponten, 1966) but then remainedconstant during the remainder of 4-6 hr of hyperventilation (Arieff,Kerian, Massry & DeLima, 1976; Kazemi, Shannon & Carvallo-Gil,1967; Wichser & Kazemi, 1975). Ponten (1966) found no decrease inc.s.f. bicarbonate during the first 15 min of moderate hyperventilationbut c.s.f. bicarbonate did decrease within 30 min. Cerebrospinal fluidbicarbonate continued to decrease for at least 6 hr (Kazemi et al. 1967).The pattern of rapid changes in plasma bicarbonate concentration andthe delayed but quantitatively larger changes in c.s.f. bicarbonate alsooccur during hypercapnia (Arieff et al. 1976; Kazemi et al. 1967; Pont6n,1966; Wichser & Kazemi, 1975). In the present studies the time courseof changes in plasma bicarbonate was similar to that discussed above.The changes in chamber fluid bicarbonate were delayed in the hypocapnicand moderately hypercapnic periods and did not significantly exceedthose of plasma. Despite similar changes in bicarbonate concentration,the chamber fluid pH returned closer to its control value than did that ofplasma as indicated by the higher % pH regulation in chamber fluidduring the later time periods. Severe hypercapnia caused a rise in plasmabicarbonate similar to that seen in moderate hypercapnia both in time andquantity. However, chamber fluid bicarbonate increased more rapidlywith the higher Pco2. It is interesting to note that the % pH regulation inchamber fluid was similar during both moderate and severe hypercapniawhile plasma pH regulation decreased with increasing Pco2.The greater change in c.s.f. bicarbonate than in plasma bicarbonate
has been explained by the changes in potential difference that occurduring alterations of ventilation. In most experimental animals hyper-capnia leads to an increase in the c.s.f.-blood potential (Held et al. 1964;Kjallquist, 1970; Kjallquist & Siesjo, 1967). However, in the cat hyper-capnia causes a negative shift in the potential measured between bulkc.s.f. in the ventricles or cisterna magna and extrameningeal tissue(Mottschall & Loescheke, 1963; Pannier, Weyne & Leusen, 1971; Woody,Marshall, Besson, Thompson, Aleonard & Albe-Fessard, 1970). Therefore,in the cat the ratio of bicarbonate concentration in c.s.f. to plasmabicarbonate should decrease during hypercapnia if the change in c.s.f.bicarbonate is controlled passively by the changes in potential differenceand plasma bicarbonate. Pannier et al. (1971) found that the c.s.f.
424

C.S.F. ACID-BASE REGULATIONplasma bicarbonate ratio actually increased during hypercapnia in the cat.In the present study the potential difference across the choroid plexus wasnot altered by hyperventilation or hypercapnia.The similarity of the changes in c.s.f. and plasma bicarbonate con-
centrations during hypocapnia and moderate hypercapnia might suggestthat passive processes are controlling c.s.f. pH when ventilatory status isaltered. However, it is known that as the severity of a respiratory acid-base disturbance increases the disparity between plasma and c.s.f.acid-base regulation becomes more apparent (Lee, Chu, Posner & Plum,1969; Shibata, Saitoh, Takahashi & Okubo, 1976). The same trend is alsoapparent in this study if the results of 5 and 10% CO2 inhalation arecompared. With 10% CO2 the increase in chamber fluid HCO3 was muchgreater than that of the plasma. Thus, active transport may be involvedin the regulation of c.s.f. bicarbonate by the choroid plexus during respira-tory acid-base disturbances and becomes more apparent as the insultbecomes more severe.The potentials measured across the isolated choroid plexus do not
behave like the potentials measured between bulk c.s.f. and blood. Thus,it is likely that the potentials measured between bulk c.s.f. and plasmaare due to processes occurring at the blood-brain barrier. The potentialdifference between bulk c.s.f. and plasma can be altered by varying plasmapotassium concentration without changes in plasma acid-base status andyet c.s.f. bicarbonate concentration will be maintained at control levels(Bledsoe & Mines, 1975; Cameron, Caronna & Miller, 1973). Thus, theregulation of c.s.f. bicarbonate is not controlled significantly by changesin potential difference and may occur primarily across membranes of thechoroid plexus where the potential difference is unaffected by plasmaacid-base status.Acute respiratory acid-base disturbances are buffered in c.s.f., at least
partially, by increases in lactate concentration during hyperventilation(Van Vaerenbergh, Demeester & Leusen, 1965; Severinghaus et al. 1963;Kjallquist, Nardini & Siesjd, 1969) and by increases in ammonia concentra-tion during hypercapnia (Hasan & Kazemi, 1976; Wichser & Kazemi,1975). These organic buffers are probably released from the brain into thec.s.f. However, during prolonged acid-base disturbances the changes inc.s.f. bicarbonate are correlated with opposite changes in c.s.f. chlorideconcentration (Severinghaus et al. 1963). The concentration of bicarbonatein fluid formed by the plexus was also inversely related to chlorideconcentration and thus there was probably no change in lactate orammonia in the newly formed c.s.f. Therefore, the regulation of c.s.f.bicarbonate during respiratory disturbances of acid-base equilibriummay be due to acute changes in both plasma bicarbonate and brain buffer
425

R. F. HUSTED AND D. J. REEDconcentrations. As newly formed c.s.f. with the proper concentration ofbicarbonate replaces that in the ventricles the choroid plexus maybecome the major factor in homoeostatic control of c.s.f. pH.
This work was supported by U.S.P.H.S. grants N3 12869, GM 00153 andDE 00252.
REFERENCES
AMEs, A. III, HIGASHI, K. & NESBETT, F. B. (1965). Effects of PcR, acetazolamideand ouabain on volume and composition of choroid-plexus fluid. J. Phyaiol. 181,516-524.
AMES, A. III, SAKANOUE, M. & ENDo, S. (1964). Na, K, Ca, Mg, and Cl concentrationsin choroid plexus fluid and cisternal fluid compared with plasma ultraffiltrate.J. Neurophysiol. 27, 672-681.
ARIEF, A. I., KERIN, A., MASSRY, S. G. & DELIMA, J. (1976). Intracellular pH ofbrain: alterations in acute respiratory acidosis and alkalosis. Am. J. Phyaiol. 230,804-812.
BLEDSOE, S. W. & MnEs, A. H. (1975). Effect of plasma (K+) on the DC potentialand on ion distributions between CSF and blood. J. apple. Phyaiol. 39, 1012-1016.
CAMERON, I. R., CARONNA, J. & MILLER, R. (1973). The effect of acute hyperkalemiaon the c.s.f.-blood potential difference and the control of c.s.f. pH. J. Phyaiol.232, 102-103P.
CUTLER, R. W. P. & BARLOW, C. F. (1966). The effect of hypercapnia on brainpermeability to protein. Arch8 Neurol., Chicago 14, 54-63.
EVANs, C. A. N., REYNOLDS, J. M., REYNOLDS, M. L. & SAUNDERS, N. R. (1976).The effect of hypercapnia on a blood-blain barrier mechanism in foetal and new-borr sheep. J. Phyaiol. 255, 701-714.
FENCL, V., MILLER, T. B. & PAPPENHEIMER, J. R. (1966). Studies on the respiratoryresponse to disturbances of acid-base balance, with deductions concering theionic composition of cerebral interstitial fluid. Am. J. Physiol. 210, 459-472.
FORSTER, H. V., DEMPSEY, J. A. & CHOSY, L. W. (1975). Incomplete compensationof CSF (H+) in man during acclimatization to high altitude (4,300 m). J. apple.Phyaiol. 38, 1067-1072.
HASAN, F. M. & KAZEMI, H. (1976). Dual contribution theory of regulation of CSFHCO- in respiratory acidosis. J. apple. Phyaiol 40, 559-567.
HELD, D., FENCL, V. & PAPPENHEIMER, J. R. (1964). Electrical potential of cere-brospinal fluid. J. Neurophyaiol. 27, 942-959.
HocHwALD, G. M., MALHAN, C. & BROWN, J. (1973). Effect of hypercapnia onCSF turnover and blood-CSF barrier to protein. Archa Neurol., Chicago 28, 150-155.
HODSON, W. A., FENNER, A., BRuIMLEY, G., CHERNICK, V. & AVERY, M. E. (1968).Cerebrospinal fluid and blood acid-base relationships iD fetal and neonatal lambsand pregnant ewes. Reap. Phyaiol. 4, 322-332.
HORNBEIN, T. F. & PAVLIN, E. G. (1975). Distribution of H+ and HCO- betweenCSF and blood during respiratory alkalosis in dogs. Am. J. Phyaiol. 228, 1149-1154.
HuSTED, R. F. & REED, D. J. (1976). Regulation of cerebrospinal fluid potassium bythe cat choloid plexus. J. Physiol. 259, 213-221.
JOHANSON, C. E., WOODBURY, D. M. & WITHROW, C. D. (1976). Distribution ofbicarbonate between blood and cerebrospinal fluid in the neonatal rat in metabolicacidosis and alkalosis. Life Sci. Oxford 19, 691-700.
KAZEMI, H., SHANNON, D. C. & CARVALLO-GIL, E. (1967). Brain CO2 bufferingcapacity in respiratory acidosis and alkalosis. J. apple. Phyaiol. 22, 241-246.
426

C.S.F. ACID-BASE REGULATIONKAZEMI, H., SHORE, N. S., SHIH, V. E. & SHANNON, D. C. (1973). Brain organic
buffers in respiratory acidosis and alkalosis. J. apple. Physiol. 34, 478-482.KJALLQUIST, A. (1970). The CSFfblood potential in sustained acid-base changes in
the rat. With calculations of electrochemical potential differences for H+ andHCO3-. Acta physiol. scand. 78, 85-93.
KJALLQUIST, A. & SIESJO, B. K. (1967). The CSF/blood potential in sustainedacidosis in the rat. Acta physiol 8cand. 71, 255-256.
KJALLQUIST, A. & SIEsJo, B. K. (1968). Regulation of CSF pH-Influence of theCSF/plasma potential. Scand. J. clin. Lab. Invest. suppl. 102, I: C.
KJALLQUIST, A., NARDINI, M. & SIESJO, B. K. (1969). The regulation of extra- andintracellular acid-base parameters in the rat brain during hyper- and hypocapnia.Acta physiol. 8cand. 76, 485-494.
LEE, J. E., CHU, F., POSNER, J. B. & PLUM, F. (1969). Buffering capacity of cerebro-spinal fluid in acute respiratory acidosis in dogs. Am. J. Physiol. 217, 1035-1038.
LENDING, M., SLOBODY, L. B. & MESTERN, J. (1961). Effect of hyperoxia, hyper-capnia, and hypoxia on blood-cerebrospinal fluid barrier. Am. J. Physiol. 200,959-962.
MERLIS, J. K. (1940). The effect of changes in the calcium content of cerebro-spinal fluid on spinal reflex activity in the dog. Am. J. Physiol. 131, 67-72.
MINER, L. C. & REED, D. J. (1972). Composition of fluid obtained from choroidplexus tissue isolated in a chamber in sitU. J. Physiol. 227, 127-139.
MITCHELL, R. A., HERBERT, D. A. & CARMAN, C. T. (1965). Acid base constants andtemperature coefficients for cerebrospinal fluid. J. apple. Physiol. 20, 27-30.
MOTTSCHALL, H. -J. & LOESOHEKE, H. H. (1963). Messungen des transmeningealenpotentials der katze bei aenderungen des C02-drucks und der H+-ionenkonzentra-tion im blut. Pflugerm Arch. ges. Physiol. 277, 662-670.
NILSSON, L. & BUSTO, R. (1973). Controlled hyperventilation and its effects onbrain energy and acid base parameters. Acta anaesth. scand. 17, 243-252.
PANNIER, J. L., WEYNE, J. & LEUSEN, I. (1971). The CSF/blood potential and theregulation of the bicarbonate concentration of CSF during acidosis in the cat.Life Sci. Oxford 10, 287-300.
PAPPENHEIMER, J. R. (1970). Transport of HCO3- between brain and blood. InCapillary Permeability, ed. CRONE, C. & LASSEN, N. A., pp. 454-458. Copenhagen:Munksgaard; New York: Academic Press.
PAVLIN, E. G. & HORNBEIN, T. F. (1975a). Distribution of H+ and HCO3- betweenCSF and blood during metabolic acidosis in dogs. Am. J. Physiol. 228, 1134-1140.
PAVLIN, E. G. & HORNBEIN, T. F. (1975b). Distribution of H+ and HCO3- betweenCSF and blood during metabolic alkalosis in dogs. Am.J. Physiol. 228, 1141-1144.
PAVLIN, E. G. & HORNBEIN, T. F. (1975c). DistIibution of H+ and HCO3- betweenCSF and blood during respiratory acidosis in dogs. Am. J. Physiol. 228, 1145-1148.
PESCE, M. A. & STRANDE, C. S. (1973). A new micromethod for determination ofprotein in cerebrospinal fluid and urine. Clin. Chem. 19, 1265-1267.
PONTEiN, U. (1966). Consecutive acid-base changes in blood, brain tissue and cerebro-spinal fluid during respiratory acidosis and baseosis. Acta neurol. 8cand. 42,455-471.
PONTEN, U. & SIESJO, B. K. (1966). Gradients of CO2 tension in the brain. Actaphysiol. 8cand. 67, 129-140.
PONTE'N, U. & SIESJO, B. K. (1967). Acid- base relations in arterial blood and cerebro-spiral fluid of the unanesthetized rat. Acta physiol. scand. 71, 89-95.
SEVERINGHAUS, J. W., MITCHELL, R. A., RICHARDSON, B. W. & SINGER, M. M.(1963). Respiratory control at high altitude suggesting active transport regulationof CSF pH. J. appl. Physiol. 18, 1155-1166.
427

428 R. F. HUSTED AND D. J. REEDSHIBATA, H., SAITOH, Y., TAKAHASHI, H. & OKUBO, T. (1976). The apparent buffer
value of cerebrospinal fluid in acute hypercapnia. Bull. Eur. Physiopath. Reap.12, 297-310.
SrEsj6, B. K. (1971). Quantification ofpH regulation in hypercapnia and hypocapnia.Scand. J. clin. Lab. Invest. 28, 113-119.
SIESJo, B. K. (1972).-The regulation of cerebrospinal fluid pH. Kidney Int. 1, 360-374.
VAN VAERENBERGH, P. J. J., DEMEESTER, G. & LEUSEN, I. (1965). Lactate incerebrospinal fluid during hyperventilation. Arch8 int. Physiol. 73, 738-747.
VoGH, B. P. & MAREN, T. H. (1975). Sodium, chloride, and bicarbonate movementfrom plasma to cerebrospinal fluid in cats. Am. J. Physiol. 228, 673- 683.
WELCH, K. & SADLER, K. (1965). Electrical potentials of choroid plexus of the rabbit.J. Neurosurg. 22, 344-349.
WICHsER, J. & KAZEMI, H. (1975). CSF bicarbonate regulation in respiratoryacidosis and alkalosis. J. apple. Physiol. 38, 504-511.
WILLIAs, J. A., WInrEow, C. D. & WOODBuIRY, D. M. (1971). Effects of ouabainand diphenylhydantoin on transmembrane potentials, intracellular electrolytes,and cell pH of rat muscle and liver in vivo. J. Physiol. 212, 101-115.
WOODY, C. D., MARSHALL, W. H., BEssON, J. M., THOMPSON, H. K., ALEONARD,P. & ALBE-FESSARD, D. (1970). Brain potential shift with respiratory acidosis inthe cat and monkey. Am. J. Physiol. 218, 275-283.




















