
Effect of anti-vascular endothelial growth factor antibody during early fetal development in rats
Upload
independentCategory
view
4download
0
Regression of Vessels in the Tunica Vasculosa LentisIs Initiated by Coordinated Endothelial Apoptosis:A Role for Vascular Endothelial Growth Factoras a Survival Factor for EndotheliumCHRISTOPHER A. MITCHELL,* WERNER RISAU, AND HANNES C.A. DREXLERMax-Planck Institut fur Physiologische und Klinische Forschung, W.G. Kerckhoff Institut, Bad Nauheim, Germany
ABSTRACT The development of the embry-onic lens is dependent on the formation andregression of the tunica vasculosa lentis (TVL),which is a transiently occurring capillary plexusthat surrounds the posterior part of the lens. Inthis study, by using the terminal deoxy-nucleo-tidyl transferase mediated nick end-labellingtechnique (TUNEL), electron microscopy, radioac-tive end-labelling of DNA extracted from TVL,and the Comet assay, we show that widespreadapoptosis of the endothelial cells that constitutethe TVL is occurring already at embryonic day17.5 (E17.5) of mouse development, much earlierthan was reported previously (Jack [1972a] Am.J. Ophthalmol. 74:261–272; Lang [1997] Cell DeathDiff. 4:12–20). In addition to apoptotic cell death,regression of this structure is associated withloss of capillary integrity, leakage of erythrocytesinto the vitreal compartment, and phagocytosisof the apoptotic endothelium by tissue macro-phages (hyalocytes).
In situ hybridization experiments with probesfor the flk-1 receptor and its high-affinity ligand,vascular endothelial growth factor (VEGF; Ter-man et al. [1992] Biochem. Biophys. Res. Com-mun. 187:1579–1586; Millauer et al. [1993] Cell72:835–846), revealed strong endothelial cell ex-pression for flk-1 in the eyes of E13.5–E17.5 em-bryos. VEGF mRNA was detected in lens epithe-lial cells located at the posterior pole of thedeveloping lens in E13.5 embryos, in close proxim-ity to the TVL capillaries. At later times (E14.5–E17.5), when the lens epithelial cells have differ-entiated into primary lens fiber cells, and a thicklenticular capsule is formed, the expression ofVEGF mRNA becomes restricted to the anteriorand equatorial portions of the lens. The physicalseparation of the VEGF-producing cells from theflk-1-expressing endothelium (due to the differen-tiation of the lens epithelial cells into lens fibercells and the formation of the lenticular capsule)may deprive the endothelium of an essentialsurvival factor and, thus, may constitute theprimary mechanism that is responsible for theinduction of endothelial cell apoptosis in this
model. Dev. Dyn. 1998;213:322–333.r 1998 Wiley-Liss, Inc.
Key words: embryonic lens; apoptosis; hyaloid;tunica vasculosa lentis; vascular en-dothelial growth factor
INTRODUCTION
The hyaloid vascular system (HVS) is a transientlyexisting network of capillaries that functions to nourishthe developing lens structure and may be involved inthe formation of the primary vascular vitreous of thedeveloping eye (Mann, 1964). In the mouse, the HVS isfirst recognized at embryonic day 10.5 (E10.5) and isessentially complete by E13.5. The cells that eventuallygive rise to the HVS are derived from two groups ofmesenchymal cells that invade either the gap betweenthe optic rim and the developing lens or through theoptic fissure. At the rim of the optic cup, one populationof primary mesenchymal cells differentiates into eitherelongated vascular cells or spherical hematopoieticprecursors. This wall of new capillaries is formed by onelayer of vascular precursor cells, which eventuallydifferentiate into the endothelium of the hyaloid ves-sels. Similarly, at the optic fissure, the hyaloid arteryforms in a segmental manner by the differentiation ofmesenchymal cells into either vascular and endothelialcells of the vessel wall or hematopoietic precursorslocated inside the developing vessels (Balazs et al.,1980).
In the rabbit, the first signs of HVS capillary regres-sion are observed as early as E21, when plateletthrombi adhere to the endothelial walls. At around thetime of birth, endothelial cells of the HVS containnumerous cytoplasmic glycogen and lipid droplets, andthe luminal surface of the endothelium shows promi-nent folding and microvillous projections (Jack, 1972a).In addition, cells with the ultrastructural appearancesof macrophages and that contain phagocytosed cellulardebris are observed to occlude the vessel lumen.
*Correspondence to: Dr. Christopher A. Mitchell, CRC Departmentof Clinical Oncology, City Hospital, Hucknall Road, Nottingham, NG51PB, United Kingdom. E-mail: [email protected]
Received 6 July 1998; Accepted 4 August 1998
DEVELOPMENTAL DYNAMICS 213:322–333 (1998)
r 1998 WILEY-LISS, INC.

Removal of cellular debris in the TVL is not associ-ated with the inflammatory response (Jack, 1972a) thataccompanies necrotic injury, supporting the proposalthat a developmentally programmed, apoptotic processis responsible for TVL regression. In fact, infiltration ofthe embryonic eye by inflammatory cells would have adeleterious effect on the development and tissue integ-rity of the eye. Retrospective analysis of early ultrastruc-tural studies of the hyaloid vasculature reveals severalexamples (Braekevelt and Hollenberg, 1970; Jack,1972a,b; Yamamoto and Fujimoto, 1984) of characteris-tically apoptotic endothelium in the TVL as well asvitreal hyalocytes with phagocytosed apoptotic bodies.Large-scale apoptosis is a characteristic of the develop-ment of a wide variety of embryonic and adult organsystems: One widely accepted hypothesis is that cells inthese organs default to an apoptotic program after thelevel of ‘‘survival factors’’ produced by neighboring cellsdrops below a critical threshold (Raff, 1992).
Recently, it has been shown that apoptosis in endothe-lial cells results in the cell surface expression of acti-vated tissue factor (Greeno et al., 1996), which is apotent procoagulant factor. Endothelial cell surfaceexpression of tissue factor results in the deposition offibrin, which, in turn, leads to the adhesion and activa-tion of platelets (Andree and Nemerson, 1995). There-fore, platelet adhesion to hyaloid vascular endothelium(HVE), which is the first ultrastructural evidence ofhyaloid regression in the rabbit (Jack, 1972a), may bemediated by the induction of apoptosis. The precisetrigger for the induction of apoptosis in the HVE isunknown; however, there are at least two potentialcontributing factors: The first is loss of contact with theextracellular matrix components that are necessary toensure the survival of endothelial cells (Meredith et al.,1993; Re et al., 1994; Drake et al., 1995; Stromblad andCheresh, 1996). The second possibility is that a survivalfactor, such as vascular endothelial growth factor VEGF(Araki et al., 1990; Alon et al., 1995; Benjamin et al.,1998), is down-regulated prior to the onset of endothe-lial apoptosis. Because the first regression phenomenaof the rabbit hyaloid system are observed at E21 (Jack,1972a), the event that initiates this regression mustoccur at or prior to E21 in the rabbit model. Therefore,we hypothesized that regression of the murine HVS isinitiated through programmed cell death (apoptosis) ofendothelial cells much earlier than was described previ-ously (Jack, 1972a; Lang and Bishop, 1993) and shortlyafter E13.5, when the TVL is fully formed. We used theterminal deoxy-nucleotidyl transferase mediated nickend-labelling technique (TUNEL), transmission elec-tron microscopy, radioactive end-labelling of DNA ex-tracted from TVL, and the comet assay to demonstratethat endothelial cells of the HVS show characteristics ofapoptosis as early as E17.5. We also showed thatendothelial cells of the TVL express flk-1 but that theyare separated progressively from the ligand (VEGF)beginning at E14.5–E17.5: From these data, we hypoth-esize that the physical separation of the source (includ-
ing the formation of the thick lens capsule during thistime) and the target of this juxtacrine survival signal inthe lens is the initiating event in the early, widespreadapoptosis of the TVL endothelium.
RESULTS
In both wholemount samples (Fig. 1A–D) and sec-tions (data not shown), apoptotic endothelial cell nucleithat were stained by using the TUNEL technique wereevident from E17.5 to postnatal day 16 (P16; Fig. 1A–D;and data not shown). From E17.5 to P5, the structure ofthe TVL was maintained (Fig. 1A–D); that is, thehyaloid artery, the long vessels, and the interconnect-ing vessels of the TVL showed no macroscopic featuresof regression. After P5, there was a breakdown in thestructure of the TVL that began with the loss of thesmall interconnecting capillaries, followed by the longerbranches of the TVL, and, finally, the complete removalof the longer hyaloid vessels by P16 (data not shown).Intimately associated with the TVL were cells thatstained for the monocyte/macrophage marker F4/80(Fig. 1E,F). These cells were most abundant in theposterior portions of the TVL in embryonic (data notshown) and P1 mice (Fig. 1E,F), and they were directlyadherent to the capillaries of the TVL. Careful compari-son of TVL cells that were stained with F4/80 antibodyand those that were stained with TUNEL revealed thatthe vast majority of F4/80-positive hyalocytes wereextraluminal to the TVL capillaries (Fig. 1E,F), theywere particularly numerous at capillary branch points(Fig. 1F), and they were not detectable by using theTUNEL technique (Fig. 1B). In contrast, prior to P5,TUNEL-positive cells were located almost exclusivelywithin the capillary walls (compare Fig. 1A,B with Fig.1E,F). From P5 onward, cells that were clearly externalto the TVL capillaries were also TUNEL positive (Fig.1C): These cells are probably macrophages with in-gested apoptotic fragments containing TUNEL-labellednuclear material (Rotello et al., 1994).
DNA Fragmentation
DNA extracted from the cells of the TVL displayed aDNA fragmentation pattern that was characteristic ofcells undergoing apoptotic cell death (Fig. 2A, lanes 3and 4). This pattern was very similar to the patterndisplayed by DNA extracted from HL60 cells, whichwere rendered apoptotic by etoposide treatment andwhich served as positive controls (Fig. 2A, lane 1). DNAextracted from the sample in which lenses and TVLswere homogenized together also showed extensive DNAfragmentation (Fig. 2A, lane 2).
We performed the comet assay as an independentassay to determine whether cells comprising the TVLare in the process of degenerating. The vast majority ofcell nuclei from the TVL that we observed within theagarose gels showed characteristic comet tails (Fig. 2B),which indicate the presence of DNA strand breaks inthese cells and which correlate well with our results de-rived from the experiments using the TUNEL technique.
323APOPTOSIS IN ENDOTHELIUM OF THE TVL

Transmission Electron Microscopy
Ultrastructural evidence of endothelial apoptosis andof vessels that showed evidence of a breakdown of theirstructural integrity in the TVL was seen in samples oftissue from mice at both E15.5 (data not shown) andE17.5 (Fig. 3A–G). The most common feature observedat these times was condensation and marginalization ofheterochromatin to the borders of endothelial nuclei(Fig. 3A). In addition, the cytoplasm of endothelial cells
was electron dense and contained dilated mitochondria(Fig. 3A). On both the luminal surface and the ablumi-nal surface of the TVL endothelium, there were numer-ous membranous folds and microvillous projections(Fig. 3B), and the basal lamina often was separatedfrom the abluminal surface (Fig. 3E). Examples ofendothelial cell wall narrowing were found (Fig. 3C),and macrophages were observed in the process ofphagocytosing erythrocytes located within the vitreous
Fig. 1. Endothelial cell apoptosis as demonstrated by using immuno-histochemical terminal deoxy-nucleotidyl transferase mediated nick end-labelling (TUNEL) of eyes from embryonic day 17.5 (E17.5) to postnatalday 5 (P5) mice (A–D). In wholemount samples, extensive nuclear TUNELlabelling, which is indicative of endothelial apoptosis, is seen in thevessels of the tunica vasculosa lentis (TVL) as well as the hyaloid artery.Stars mark areas that were denuded of capillaries during the processingprocedure. A: E17.5, posterior view. Widespread apoptosis is evident inboth the hyaloid artery (arrowhead) and in TVL capillaries (small arrow).B: Enlarged view from the area indicated by the large arrow in A.TUNEL-positive cell nuclei are clearly identifiable within the contours of
the capillaries. C: P5, posterior view. There is also a clear difference in thenumber of small, connecting capillaries compared with E17.5 (cf. A).There are now several examples of cells external to the capillaries,probably hyalocytes, that are TUNEL positive (arrows). D: E17.5 controlTVL (TdT omitted). E,F: Wholemount immunohistochemical staining withF4/80 antibody of hyalocytes of the TVL in P1 mouse. E: Posterior view oflens with large numbers of macrophages on the long branches of thehyaloid artery and connecting vessels. Note that the F4/80-positive cellsare in a different location than the TUNEL-positive cells shown in B. F:Note that the F4/80-positive hyalocytes (arrows) are external to thecapillaries and commonly are located at vessel branching points.
324 MITCHELL ET AL.

(Fig. 3D), confirming a breakdown in capillary integ-rity. Endothelial cell nuclei with marginalized hetero-chromatin were also observed projecting perpendicu-larly into the vessel lumen (Fig. 3E) or with a clear zoneseparating them from the endothelial cytoplasm (Fig.3B). It was also noted that the majority of vessels thatwere examined had patent lumina (data not shown).The only cell type that was observed to be in mitosiswas located abluminal to endothelium and under theinvesting basal lamina (Fig. 3F), which is consistentwith the morphological description of a pericyte.
In the hyaloid artery, morphologically distinct, cuboi-dal endothelial cells also displayed condensed andmarginalized heterochromatin (Fig. 3G). Serial section-ing through the hyaloid artery of an E17.5 embryo eye(data not shown) revealed that many of these cuboidalendothelial cells were anuclear and exhibited extensivemembrane folding and microvillous projections, al-though they retained their junctional complexes (Fig.3G).
VEGF and flk-1 mRNA ExpressionDuring Lens Development
In E13.5 eyes, there was strong labelling for VEGFmRNA over the primary lens epithelial cells at theposterior pole of the lens (Fig. 4A,B), with less intensesignal intensity in the superficial layers of the retina.From E14.5 to E17.5, labelling for VEGF becamerestricted to transitional lens epithelial cells at theequator and fiber cells deep within the structure of thelens, with the consequence that expression of VEGFmRNA became located more anteriorly within the lensstructure (Fig. 4C–E).
Staining for flk-1 in sections of eyes from E13.5–E17.5 embryos was found in cells adjacent to the lensstructure and also on the vitreal side of the retina (Fig.4F–J). Extensive staining for flk-1 was seen throughoutthe rest of the embryo (data not shown), and the patternof staining was consistent with that seen in previousstudies for flk-1 (Millauer et al., 1993; Yamaguchi et al.,1993) or by using the endothelial cell markers endoglin(Ge and Butcher, 1994), PECAM-1 (Vecchi et al., 1994),and MECA 32 (Hallmann et al., 1995; data not shown).In E13.5 embryo eyes, flk-1-expressing cells were lo-cated in three distinct areas, in a round patch of cells inthe center of the vitreal space and in two crescentricpatches located at the anterior and posterior poles ofthe eye anlage (Fig. 4G). In E14.5–E17.5 eyes, flk-1staining was prominent in cells that were juxtaposed tothe lens and adjacent to the vitreal surface of theretina, consistent with staining of the TVL (Fig. 4H–J).In addition, a thick lenticular capsule began to formaround the lens by E14.5. Because endothelial cells ofthe TVL express flk-1 but are progressively separatedfrom the ligand (VEGF) beginning at E14.5–E17.5, wehypothesize that the physical separation of the source(including the formation of the thick lens capsuleduring this time) and target of this juxtacrine survivalsignal in the lens may be the initiating event in theearly, widespread apoptosis of the TVL endothelium.
DISCUSSION
In this study, we have shown that endothelial cells,which are the major cellular constituent of the TVL,undergo apoptotic cell death, as revealed by detection offragmented DNA (TUNEL, end labelling of fragmented
Fig. 2. A: DNA fragmentation by radioactive labelling with Klenowpolymerase and agarose gelelectrophoresis. DNA was extracted frometoposide-treated HL60 cell (lane 1), from homogenized TVLs and lenses(lane 2), and from TVLs that were not homogenized (lanes 3 and 4).Exposure times were 4 hr for lane 1 and 42 hr for lanes 2–4. B: Detection
of DNA strand breaks in TVL cells by using the comet assay (3400). DNAwithin cell nuclei and comet tails was stained with ethidium bromide. Theincreased signal intensity in lane 2 is most likely due to the additionalextraction of apoptotic DNA fragments from the nuclei of lens fiber cells,which also apoptose during the process of lens differentiation.
325APOPTOSIS IN ENDOTHELIUM OF THE TVL
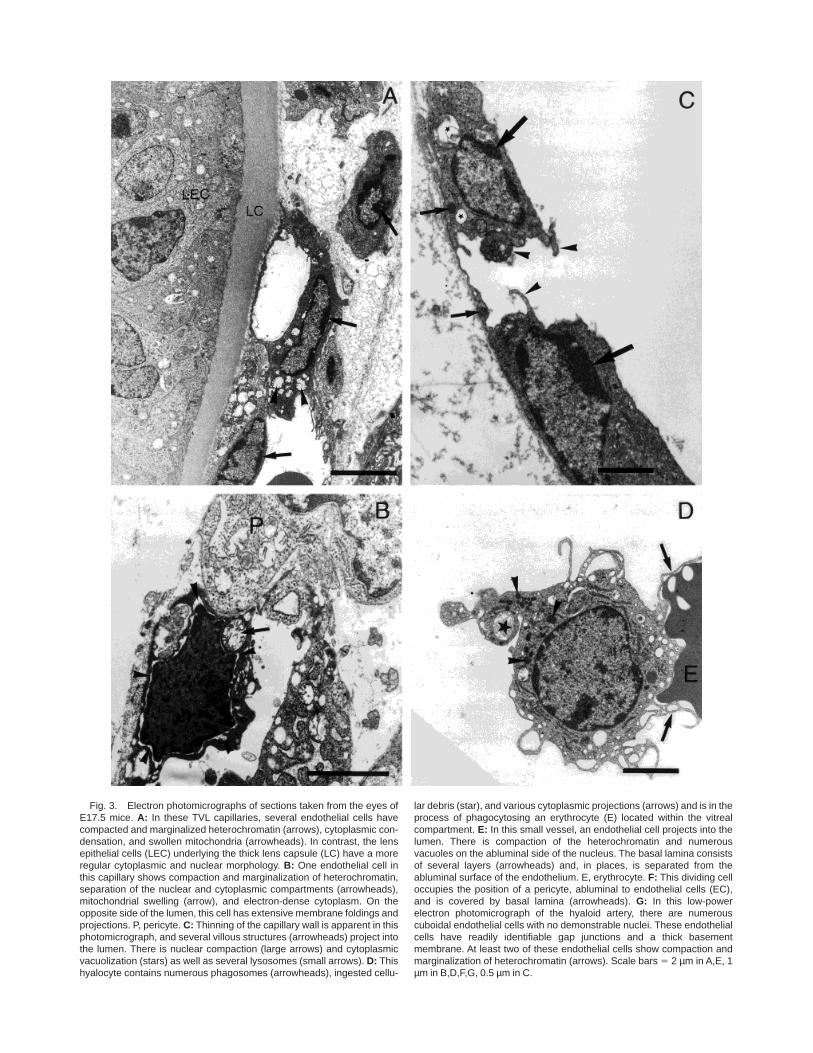
Fig. 3. Electron photomicrographs of sections taken from the eyes ofE17.5 mice. A: In these TVL capillaries, several endothelial cells havecompacted and marginalized heterochromatin (arrows), cytoplasmic con-densation, and swollen mitochondria (arrowheads). In contrast, the lensepithelial cells (LEC) underlying the thick lens capsule (LC) have a moreregular cytoplasmic and nuclear morphology. B: One endothelial cell inthis capillary shows compaction and marginalization of heterochromatin,separation of the nuclear and cytoplasmic compartments (arrowheads),mitochondrial swelling (arrow), and electron-dense cytoplasm. On theopposite side of the lumen, this cell has extensive membrane foldings andprojections. P, pericyte. C: Thinning of the capillary wall is apparent in thisphotomicrograph, and several villous structures (arrowheads) project intothe lumen. There is nuclear compaction (large arrows) and cytoplasmicvacuolization (stars) as well as several lysosomes (small arrows). D: Thishyalocyte contains numerous phagosomes (arrowheads), ingested cellu-
lar debris (star), and various cytoplasmic projections (arrows) and is in theprocess of phagocytosing an erythrocyte (E) located within the vitrealcompartment. E: In this small vessel, an endothelial cell projects into thelumen. There is compaction of the heterochromatin and numerousvacuoles on the abluminal side of the nucleus. The basal lamina consistsof several layers (arrowheads) and, in places, is separated from theabluminal surface of the endothelium. E, erythrocyte. F: This dividing celloccupies the position of a pericyte, abluminal to endothelial cells (EC),and is covered by basal lamina (arrowheads). G: In this low-powerelectron photomicrograph of the hyaloid artery, there are numerouscuboidal endothelial cells with no demonstrable nuclei. These endothelialcells have readily identifiable gap junctions and a thick basementmembrane. At least two of these endothelial cells show compaction andmarginalization of heterochromatin (arrows). Scale bars 5 2 µm in A,E, 1µm in B,D,F,G, 0.5 µm in C.
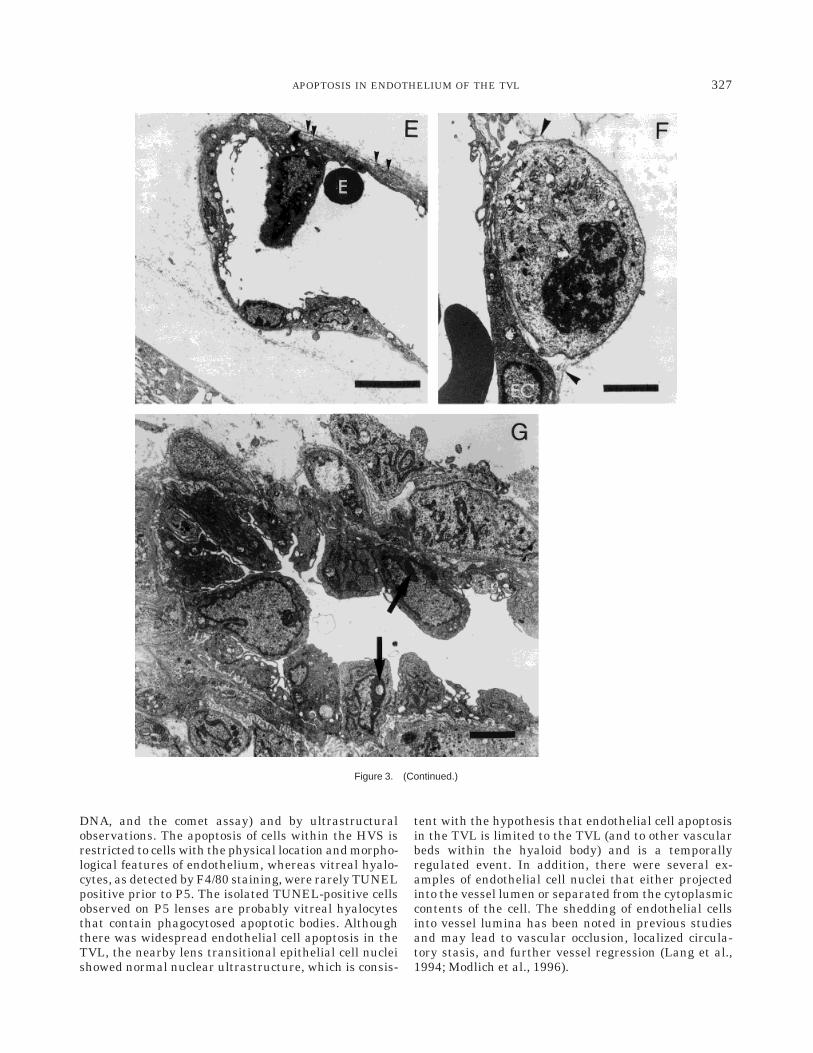
DNA, and the comet assay) and by ultrastructuralobservations. The apoptosis of cells within the HVS isrestricted to cells with the physical location and morpho-logical features of endothelium, whereas vitreal hyalo-cytes, as detected by F4/80 staining, were rarely TUNELpositive prior to P5. The isolated TUNEL-positive cellsobserved on P5 lenses are probably vitreal hyalocytesthat contain phagocytosed apoptotic bodies. Althoughthere was widespread endothelial cell apoptosis in theTVL, the nearby lens transitional epithelial cell nucleishowed normal nuclear ultrastructure, which is consis-
tent with the hypothesis that endothelial cell apoptosisin the TVL is limited to the TVL (and to other vascularbeds within the hyaloid body) and is a temporallyregulated event. In addition, there were several ex-amples of endothelial cell nuclei that either projectedinto the vessel lumen or separated from the cytoplasmiccontents of the cell. The shedding of endothelial cellsinto vessel lumina has been noted in previous studiesand may lead to vascular occlusion, localized circula-tory stasis, and further vessel regression (Lang et al.,1994; Modlich et al., 1996).
Figure 3. (Continued.)
327APOPTOSIS IN ENDOTHELIUM OF THE TVL

After the early and widespread induction of apoptosisof the TVL, regression and removal of the TVL may becompleted by two main processes: The radial growth ofthe lens itself that, without a concomitant growth of theTVL, causes a stretching and breaking of the capillariesconnecting the longitudinal vessels (Latker and Ku-wabara, 1981), and the detachment and phagocytosis ofapoptotic endothelial cells and extravasated vascularcomponents by vitreal hyalocytes (Balazs et al., 1980;Yamamoto and Fujimoto, 1984).
It has been proposed previously that the regression ofthe TVL undergoes a coordinated and species-specificpattern of regression and that the timing of the initia-tion of regression events in mice occurs around P5–P6(Balazs et al., 1980; Lang, 1997). In contrast, we haveobserved regression events, including widespread apop-tosis of endothelial cells, loss of endothelial basementmembrane integrity, and vitreal hyalocytes with in-gested cellular debris, in the eyes of E17.5 mice.
Whether the hyalocytes of the vitreous are directlyresponsible for eliciting the cell death program in TVLendothelium or function purely as phagocytic cellswithin the vitreous is unclear. To target hyalocytes inthe primary vitreous, Lang and coworkers (Lang andBishop, 1993) used a diphtheria toxin transgene underthe control of a granulocyte macrophage-colony stimu-lating factor (GM-CSF) promoter. When the developingeyes of these mice were examined, there was somepersistence of the hyaloid system, particularly in thelong vessels of the TVL and in the hyaloid artery. Thispersistence of the hyaloid system was attributed to theelimination of vitreal hyalocytes, and the authors con-cluded that the hyalocytes were directly responsible forthe endothelial cell death in this system (Lang andBishop, 1993; Lang et al., 1994). However, physicalpersistence of a nonfunctional hyaloid vasculature wouldalso be observed if the cell type responsible for theremoval of the apoptotic cells (the hyalocyte) waseliminated from the system (Lang and Bishop, 1993).
Because TVL regression is regulated temporally andis initiated through a coordinated, widespread endothe-lial apoptosis, and because the pattern of events ap-pears to be consistent regardless of the species in-volved, one of the major questions is what event(s)initiates the coordinated apoptosis observed in TVLendothelium at E17.5? Because the early growth of thelens is dependent on the formation and regression ofthe TVL (Mann, 1964), then, from a teleological view-point, the growth and regression of the TVL should bemediated through signals from the maturing lens an-lage. Several lines of evidence are consistent with thishypothesis. Transgenic animals in which the differentia-tion of lens epithelial cells is disturbed, such as thoseexpressing fibroblast growth factor 1 (FGF1) or diphthe-ria toxin, under the control of the lens epithelialcell-specific aA-crystallin promoter (Chepelinsky et al.,1985) demonstrate a microphthalmia in addition to aregional persistence of the HVS (Breitman et al., 1987;Key et al., 1992; Robinson et al., 1995). In the earliest
stages of lens development, when the primary lens fibercells begin to differentiate (around E13.5 in mice), thesecells show strong mRNA expression for FGF1 andFGF2 (de Iongh and McAvoy, 1992), platelet-derivedgrowth factor (PDGF; Mudhar et al., 1993), VEGF (thisstudy), and polypeptide growth factors, which are mito-genic, chemoattractive, or survival factors for endothe-lium or pericytes. Further differentiation of the pri-mary and secondary lens fiber cells and formation of thethick lenticular capsule, which begins at E13.5–E15.5,are associated with a physical separation of thesegrowth and survival factors from the adjacent TVLand/or a down-regulation of the mRNA for PDGF, FGF1and FGF2, and VEGF (de Iongh and McAvoy, 1992;Mudhar et al., 1993; this study) and, thus, may be themechanism for the widespread apoptosis demonstratedin this study. The coordinated induction of apoptosis inmurine TVL endothelium occurs at around E15.5 (notshown); it is precisely at this time that expression ofVEGF becomes restricted (from E15.5 to E17.5) to lensfiber cells deep within the structure of the lens andtransitional epithelial cells at the lens equator. Becausethe expression of flk-1, a high affinity receptor formVEGF (Terman et al., 1992; Millauer et al., 1993), isexpressed at this time on endothelial cells of thejuxtaposed TVL, we hypothesized that endothelial cellsof the TVL, which have lost a critical juxtacrine sur-vival signal, undergo apoptosis. The mechanism oforgan regression that occurs by the loss of a juxtacrine/paracrine survival signal has been described for severalother systems and cytokines (Raff, 1992), includingerythropoietin, which rescues erythroid progenitor cells(Koury, 1992); colony stimulating factors, which sup-press apoptosis of hemopoietic precursor cells (Williamset al., 1990); epidermal growth factor, which reducesthe apoptosis that occurs during kidney development(Koseki et al., 1992; Coles et al., 1993); nerve growthfactor (NGF), which increases the survival of embryonicsympathetic neurons (Korsching and Thoenen, 1983;
Fig. 4. Darkfield photomicrographs of in situ autoradiographs thatwere generated with probes to vascular endothelial growth factor (VEGF;A–E) and flk-1 (F–J). A: E13.5 sense control. B: E13.5 VEGF antisense.Note the strong expression of VEGF over the lens epithelial cells at theposterior pole of the lens (arrowhead). C: E14.5 VEGF. The signal forVEGF is restricted to primary lens fiber cells (arrowheads) and transitionallens epithelium at the lens equator (arrows). D: E15.5 VEGF antisense.The signal for VEGF mRNA is located deep within the lens structure(arrowhead). E: E17.5 VEGF antisense. The signal for VEGF is limited tothe lens fiber cells in the anterior one-half of the lens (arrowhead) and to alayer on the superficial surface of the retina (arrows) that probably iscomprised of astrocytes migrating along the surface of the retina, asdescribed by Stone et al. (1996). F: E13.5 flk-1 sense control. G: E13.5flk-1 antisense. Note that, at this stage, the capillaries of the TVL aredensely packed, so that the patch of labelling in the central portion of theeye (arrow) corresponds to the hyaloid vessels. In addition, there arebands of flk-1-expressing cells at the posterior (arrowheads) and anteriorof the eye anlage. H: E14.5 flk-1 antisense. Discrete points of labellingcan be seen next to the retina (arrowheads), with strong expression in thehyaloid artery (arrow) and surrounding the lens (L). I: E15.5 flk-1antisense. J: E17.5 flk-1 antisense. Scale bars 5 100 µm. in A–D and F–I,200 µm in E,J.
328 MITCHELL ET AL.

Figure 4.
329APOPTOSIS IN ENDOTHELIUM OF THE TVL

Cowan et al., 1984; Levi-Montalcini, 1987; Oppenheimet al., 1988; Purves et al., 1988; Barde, 1989); andinsulin-like growth factor II (IGF-II), which inhibitsprogrammed cell death in dystrophic myofibers (Smithet al., 1995). The proposal that VEGF can act as asurvival factor for the transient growth of the TVL bygiving trophic support to embryonic capillaries is alsoconsistent with the data from the study by Alon et al.(1995), in which intraocular injection of VEGF rescuedretinal endothelium from apoptotic cell death that wasinduced experimentally by subjecting neonatal rats toconditions of hyperoxia-normoxia (Alon et al., 1995;Benjamin et al., 1998). Experiments with transgenicanimals are in progress to determine whether in-creased expression of three isoforms of VEGF in lensepithelial cells (using the aA-crystallin promoter; Chepe-linsky et al., 1985) can alter the pattern or extent ofendothelial cell apoptosis during development and re-gression of the murine TVL.
EXPERIMENTAL PROCEDURESAnimals
Whole eyes were collected from C57Bl mice aged fromE15.5 to P16. Briefly, female C57Bl mice were matedovernight, and, the following morning, the presence of avaginal plug was designated as E0.5. After pregnantmice were killed, the embryos were collected by cesar-ean section. Embryos (E15.5–E19.5) or neonatal mice(P0–P16) were killed, and their eyes were enucleated.The whole eyes were immediately immersed in either asolution of 4% paraformaldehyde in phosphate-bufferedsaline (PBS), pH 7.2, or 4% paraformaldehyde/2.5 %glutaraldehyde, pH 7.2, overnight. Whole embryos youn-ger than E15.5 were immersed in fixation solution orwere processed for frozen sectioning, because it was notpossible to isolate the lens successfully with the atten-dant TVL from the eyes of these mice.
Wholemount Immunohistochemistry
The sclera, cornea, and pupil were removed from thefixed eye with the aid of a dissecting microscope (Zeiss,Jena, Germany), and the lens with the TVL stillattached was carefully dissected free of the retinal cup.Both the lens and the retinal cup were placed in a singlewell of a flat-bottom, 96-well plate (Becton Dickinson;Lincoln Park, NJ) and washed three times in PBS, pH7.2, for 5 min each.
Samples to be processed for wholemount immunohis-tochemical staining were incubated in 0.1 M glycine inPBS (1 hr at room temperature; RT), dehydratedthrough a graded series of methanol (25%, 50%, 75%,and 2 changes of 100%; 15 min each at RT), treated with5% H2O2 in methanol (5 hr), and washed with twochanges of 100% methanol (15 min each). Samples wererehydrated through descending grades of methanol(75%, 50%, 25%, and 2 changes of PBS; 15 min each atRT). At this point, samples that were to be used forTUNEL staining were processed with the in situ cell-death detection kit (Boehringer-Mannheim, Mann-
heim, Germany), according to the manufacturer’s in-structions.
For immunohistochemical detection of antigens,samples were incubated in a blocking buffer containing10% preimmune serum (depending on the species inwhich the secondary antibody was raised), 1% bovineserum albumin, and 0.05% sodium azide in PBS, pH7.2. Samples were washed subsequently in PBS (threetimes at RT) before the tissue was permeabilized byimmersion in a solution of 3% instant skim milk powderand 0.1% Triton X-100 in PBS (PBSMT; twice for 1 hr atRT). Samples were then incubated in the primaryantibody (diluted 1:100 in PBSMT; 48°C overnight),extensively washed with PBSMT (five times for 1 hreach at RT), and incubated in the horseradish peroxi-dase-coupled secondary antibody (diluted 1:200 in PB-SMT; 48°C overnight). Samples were then washedextensively with PBSMT (five times for 1 hr each) andin two changes of 0.1% Triton X-100 in PBS (PBT) for 10min each. Development consisted of incubation ofsamples in a solution of 3,38,4,48-tetraaminobiphenyl(Sigma, St. Louis, MO; one tablet in 12 ml of PBT plus1:1000 H2O2) for 5–10 min at RT. The samples werethen rinsed in PBT (three times for 5 min each) andPBS (three times for 5 min each), equilibrated in 50%glycerol, and transferred into 70% glycerol, and thespecimens were viewed under a dissecting microscope.The antibodies used in this study were rat antimouseimmunoglobulin G (IgG) F4/80 (Austyn and Gordon,1981; Hume et al., 1983; Hume et al., 1984), endoglin(Ge and Butcher, 1994), and platelet/endothelial celladhesion molecule I (PECAM; Vecchi et al., 1994).
Detection of DNA Laddering by End LabellingWith Klenow Polymerase
Fresh 17.5E eyes were collected in ice-cold PBS priorto dissection. From each eye, the lens with the adherentTVL was dissected free (taking care not to damage thelens capsule) and incubated in lysis buffer (1% TritonX-100; 10 mM Tris HCl, pH 7.5; 10 mM EDTA) for 20min to extract the fragmented DNA. The unfragmentedDNA remained associated with the cell nuclei and waspelleted together with the cellular debris and the lenscapsule during the subsequent centrifugation at313,000 rpm on a bench-top microcentrifuge. Thesupernatants were extracted once with phenol and oncewith phenol/chloroform/isoamylalcohol (25:24:1), andDNA was precipitated by the addition of 0.1 volume 3 Msodium acetate and 2.5 volume ethanol. The DNAprecipitates were washed with 75% ethanol, air dried,and resuspended in 50 µl TE (10 mM Tris, 1 mMEDTA).
In one sample, the lenses with accompanying TVLs(n 5 6) were disrupted by vigorous pipetting, whereas,in the other two samples (n 5 6 each), care was taken toleave the lenses intact to extract DNA only from thecells within the TVL. To obtain a reference samplecontaining apoptotic DNA fragments, HL60 cells (1 3
330 MITCHELL ET AL.

106/ml) were treated with 10 µM etoposide for 18 hr andextracted in an analogous manner.
The labelling procedure was carried out according tothe protocol of Rosl (1992) with a few minor modifica-tions. Briefly, the samples were incubated with 2.5 UKlenow polymerase and 0.5 mCi of 32P-dCTP in thepresence of 10 mM Tris/HCL, pH 7.5, and 5 mM MgCl2.The reaction was incubated at RT for 20 min and wasterminated by the addition of 10 mM EDTA. After theaddition of 50 µg tRNA, two consecutive precipitationcycles with ammonium acetate (2.5 M final concentra-tion)/isopropanol were performed to remove unincorpo-rated label, and the pellet was resuspended in 10 mMTris/HCl, pH 7.5, and 1 mM EDTA. Aliquots wereloaded onto a 2% agarose gel and separated by electro-phoresis at 100 V for 2 hr. After the drying of the gel on3 MM Whatman paper (Whatman, Hillsboro, OR), thegel was scanned on a phosphoimager (Fujifilm BAS-2500; Fuji, Tokyo, Japan).
Detection of Fragmented DNA in the TVLby Using the Comet Assay
Comet assays were performed as described previ-ously with minor modifications (Olive et al., 1996).Briefly, lens capsules with their attached TVL (n 5 2)were separated from the lens substance by microdissec-tion and dispersed by gentle pipetting in 200 µl ofice-cold PBS. From this cell suspension, 10 µl wereadded to 150 µl of 1% low-gelling-temperature agrose(Life Technologies, Gaithersburg, MD) dissolved indouble-distilled water and preheated to 40°C. A total of70 µl of this mixture was pipetted onto a precut piece ofGelfix electrophoresis support (Serva, Heidelberg, Ger-many) and immediately covered with a glass coverslipto obtain an even spreading of the gel. After removal ofthe coverslip, the slides were immersed in an alkalinelysis solution containing 1 M NaCl, 0.03 M NaOH, and2% sarcosyl for 1 hr, followed by a 1-hr wash in tworinses of 0.03 M NaOH plus 2 mM EDTA beforeelectrophoresis in fresh rinse solution at 40 V for 7 min.The slides were then rinsed with distilled water andstained in a solution of 2.5 µg/ml propidium iodide for20 min. Comets were viewed with the aid of a Leitzepifluorescence microscope (Leitz, Wetzlar, Germany).
In Situ Hybridization
The in situ hybridization technique that was used isidentical to the technique described previously (Breieret al., 1992). Mouse embryos or fixed eyes with thecornea and sclera dissected away were embedded inTissue Tek medium (Miles Scientific, Elkhart, IN).Sections were cut in a cryostat at 220°C, dried onsilane-subbed slides (Sigma), and fixed in 4% parafor-maldehyde. Sections were digested with pronase (40µg/ml) for 10 min at RT. The digestion was stopped in0.2% glycine. Slides were rinsed with PBS, fixed in 4%paraformaldehyde, and rinsed again in PBS. Sectionswere acetylated in acetic anhydride diluted 1:400 with0.1 mM triethanolamine for 10 min and rinsed with
PBS. Prehybridization was performed in buffer contain-ing 50% formamide; 10% dextran sulphate; 10 mMTris-HCl, pH 7.5; 10 mM sodium phosphate, pH 6.8; 2 3standard saline citrate (SSC); 5 mM EDTA; 150 µg/mlyeast tRNA; 0.1 mM UTP, 0.1 mM ADPbS, 1 mMATPgS; 10 mM dithiothreitol (DTT); and 2 mM 2-mer-captoethanol for 1 hr at 48°C. Sections were dehydratedin graded ethanol and stored at 270°C.
Single-stranded [35S]RNA probes were generated byin vitro transcription of pGEM-mflk-1.ecl1 and pBS(KS)-mVEGF164 clones. These clones contained 2.6 kb and563 base pairs of the coding region of the mflk-1 ormVEGF cDNA, respectively. The RNA probes weregenerated by using 100 mCi (35S)UTP and T3, T7, orSP6 RNA polymerases essentially as described by themanufacturer (Stratagene, La Jolla, CA). The flk-1probe was subjected to alkaline hydrolysis with 2 MNaOH at 48°C for 13 min. RNA probes were purified byethanol precipitation in the presence of ammoniumacetate, dissolved in 50% formamide and 10 mM DTT,and stored at 220°C. Hybridization was performed inprehybridization buffer supplemented with 5 3 104
counts/min/ml 35S-labelled RNA probe. Sections werehybridized overnight in a (50% formamide) humidchamber at 48°C. Depending on the size of the section,approximately 20 µl of buffer/section were used. Slideswere washed in 2 3 SSC, 50% formamide for 2 hr at45–48°C followed by RNAse digestion (20 µg/ml) for 15min. Slides were washed in 2 3 SSC, 50% formamideovernight and dried at RT. After dehydration in gradedethanol, slides were coated with Kodak NTB-2 emul-sion (Eastman-Kodak, Rochester, NY) diluted 1:1 withwater and exposed for 10–14 days. Slides were devel-oped and stained with toluidine blue (0.2% in 0.2 Msodium acetate, pH 4.3).
Electron Microscopy
The whole eyes were fixed overnight in a 4% glutaral-dehyde/1% sucrose solution in PBS, pH 7.2, and subse-quently washed in PBS three times. Samples werepostfixed in 1% OsO4, dehydrated in graded concentra-tions of ethanol, infiltrated with araldite, and embed-ded in flat rubber molds. Critical orientation of speci-mens was carried out under a dissecting microscope.Semithin sections were prepared from each block,stained with 0.1% toluidine blue in 5% borax, andexamined under a light microscope. The blocks weretrimmed to 1 mm2, including the area of interest, andultrathin sections (50 nm) were mounted on 200-mesh,thin-bar copper grids, stained with lead citrate, coatedwith carbon, and examined in a Phillips CM10 transmis-sion electron microscope (Phillips, Holland) at an accel-erating voltage of 80 kV.
ACKNOWLEDGMENTS
The authors thank Dr. Eli Keshet for useful discus-sions, Dr. Georg Breier for providing probes for use withthe in situ hybridization technique, Dr. Matthias Claussfor critical review of the paper, and Prof. Dr. Jutta
331APOPTOSIS IN ENDOTHELIUM OF THE TVL

Schaper, Dr. Sawa Kostin, and Fr. Annemarie Mobsfrom the Department of Experimental Cardiology,Kerckhoff Institut, Bad Nauheim for assistance withelectron microscopy. We are also grateful to Fr. HiltrudHolzinger for assistance with the photomicrographs.
REFERENCESAlon T, Hemo I, Itin A, Peer J, Stone J, Keshet E. Vascular endothelial
growth factor acts as a survival factor for newly formed retinalvessels and has implications for retinopathy of prematurity. NatureMed. 1995;1:1024–1028.
Andree HAM, Nemerson Y. Tissue factor: Regulation of activity by flowand phospholipid surfaces. Blood Coagul. Fibrin. 1995;6:189–197.
Araki S, Simada Y, Kaji K, Hayashi, H. Role of protein kinase C in theinhibition by fibroblast growth factor of apoptosis in serum depletedendothelial cells. Biochem. Biophys. Res. Commun. 1990;172:1081–1085.
Austyn JM, Gordon S. F4/80, a monoclonal antibody directed specifi-cally against the mouse macrophage. Eur. J. Immunol. 1981;11:805–815.
Balazs EA, Toth LZ, Ozanics V. Cytological studies on the developingvitreous as related to the hyaloid vessel system. Alb. Von Graefe’sArch. Klin. Exp. Ophthalmol. 1980;213:71–85.
Barde YA. Trophic factors and neuronal survival. Neuron 1998;2:1525–1534.
Benjamin L, Heino I, Keshet E. A plasticity window for blood vesselremodelling is defined by pericyte coverage of the preformed endothe-lial network and is regulated by PDGF B and VEGF. Development1998;125:1591–1598.
Braekevelt CR, Hollenberg MJ. Comparative electron microscopicstudy of development of hyaloid and retinal capillaries in albinorats. Am. J. Ophthalmol. 1970;69:1032–1046.
Breier G, Albrecht U, Sterrer S, Risau W. Expression of vascularendothelial growth factor during embryonic angiogenesis and endo-thelial cell differentiation. Development 1992;114:521–532.
Breitman ML, Clapoff S, Rossant J, Tsui LC, Glode LM, Maxwell IH,Bernstein A. Genetic ablation: Targeted expression of a toxin genecauses microphthalmia in transgenic mice. Science 1987;238:1563–1565.
Chepelinsky AB, King CR, Zelenka PS, Piatigorsky J. Lens-specificexpression of the chloramphenicol acetyltransferase gene promotedby 58 flanking sequences of the murine alpha A-crystallin gene inexplanted chicken lens epithelia. Proc. Natl. Acad. Sci. USA 1985;82:2334–2338.
Coles HSR, Burne JF, Raff MC. Large-scale normal cell death in thedeveloping rat kidney and its reduction by epidermal growth factor.Development 1993;118:777–784.
Cowan WM, Fawcett JW, O’Leary DD, Stanfield BB. Regressive eventsin neurogenesis. Science 1984;225:1258–1265.
de Iongh R, McAvoy JW. Distribution of acidic and basic fibroblastgrowth factors (FGF) in the foetal rat eye: Implications for lensdevelopment. Growth Factors 1992;6:159–177.
Drake CJ, Cheresh DA, Little CD. An antagonist of integrin alpha vbeta 3 prevents maturation of blood vessels during embryonicneovascularization. J. Cell Sci. 1995;108:2655–2661.
Ge AZ, Butcher EC. Cloning and expression of a cDNA encoding mouseendoglin, an endothelial cell TGF-beta ligand. Gene 1994;138:201–206.
Greeno EW, Bach RR, Moldow CF. Apoptosis is associated withincreased cell surface tissue factor procoagulant activity. Lab.Invest. 1996;75:281–289.
Hallmann R, Mayer DN, Berg EL, Broermann R, Butcher EC. Novelmouse endothelial cell surface marker is suppressed during differen-tiation of the blood brain barrier. Dev. Dyn. 1995;202:325–332.
Hume DA, Perry VH, Gordon S. Immunohistochemical localisation ofa macrophage-specific antigen in developing mouse retina: Phagocy-tosis of dying neurons and differentiation of microglial cells to form aregular array in the plexiform layers. J. Cell Biol. 1983;97:253–257.
Hume DA, Perry VH, Gordon S. The mononuclear phagocyte system ofthe mouse defined by immunohistochemical localisation of antigen
F4/80: Macrophages associated with epithelia. Anat. Rec. 1984;210:503–512.
Jack R. Regression of the hyaloid vascular system. An ultrastructuralanalysis. Am. J. Ophthalmol. 1972a;74:261–272.
Jack RL. Ultrastructure of the hyaloid vascular system.Arch. Ophthal-mol. 1972b;87:555–567.
Key B, Liu L, Potter SS, Kaur S, Akeson R. Lens structures existtransiently in development of transgenic mice carrying an alpha-crystallin-diphtheria toxin hybrid gene. Exp. Eye Res. 1992;55:357–367.
Korsching S, Thoenen H. Nerve growth factor in sympathetic gangliaand corresponding target organs of the rat: Correlation with densityof sympathetic innervation. Proc. Natl.Acad. Sci. USA1983;80:3513–3516.
Koseki C, Herzlinger D, Alawqati Q. Apoptosis in metanephric develop-ment. J. Cell Biol. 1992;119:1327–1333.
Koury MJ. Programmed cell death (apoptosis) in hematopoiesis. Exp.Hematol. 1992;20:391–394.
Lang R. Apoptosis in mammalian eye development: lens morphogen-esis, vascular regression and immune privilege. Cell Death Diff.1997;4:12–20.
Lang RA, Bishop JM. Macrophages are required for cell death andtissue remodelling in the developing mouse eye. Cell 1993;74:453–462.
Lang R, Lustig M, Francois F, Sellinger M, Plesken H. Apoptosisduring macrophage-dependent ocular tissue remodelling. Develop-ment 1994;120:3395–3403.
Latker CH, Kuwabara T. Regression of the tunica vasculosa lentis inthe postnatal rat. Invest. Ophthalmol. Vis. Sci. 1981;21:689–699.
Levi-Montalcini R. The nerve growth factor 35 years later. Science1987;237:1154–1162.
Mann IC. ‘‘The Development of the Human Eye.’’ New York: Grune andStratton, Inc., 1964.
Meredith JEJ, Fazeli B, Schwartz MA. The extracellular matrix as acell survival factor. Mol. Biol. Cell 1993;4:953–961.
Millauer B, Wizigmann-Voos S, Schnurch H, Martinez R, Moller NP,Risau W, Ullrich A. High affinity VEGF binding and developmentalexpression suggest Flk-1 as a major regulator of vasculogenesis andangiogenesis. Cell 1993;72:835–846.
Modlich U, Kaup FJ, Augustin HG. Cyclic angiogenesis and bloodvessel regression in the ovary—Blood vessel regression duringluteolysis involves endothelial cell detachment and vessel occlusion.Lab. Invest. 1996;74:771–780.
Mudhar HS, Pollock RA, Wang C, Stiles CD, Richardson WD. PDGFand its receptors in the developing rodent retina and optic nerve.Development 1993;118:539–552.
Olive PL, Viske CM, Banath JP. Use of the comet assay to identify cellssensitive to tirapamazine in multicell spheroids and tumors in mice.Cancer Res. 1996;56:4460–4463.
Oppenheim RW, Haverkamp LJ, Prevette D, McManaman JL, AppelSH. Reduction of naturally occurring motoneuron death in vivo by atarget-derived neurotrophic factor. Science 1988;240:919–22.
Purves D, Snider WD, Voyvodic JT. Trophic regulation of nerve cellmorphology and innervation in the autonomic nervous system.Nature 1988;336:123–128.
Raff MC. Social controls on cell survival and cell death. Nature1992;356:397–400.
Re F, Zanetti A, Sironi M, Polentarutti N, Lanfrancone L, Dejana E,Colotta F. Inhibition of anchorage-dependent cell spreading triggersapoptosis in cultured human endothelial cells. J. Cell Biol. 1994;127:537–546.
Robinson ML, Overbeek PA, Verran DJ, Grizzle WE, Stockard CR,Friesel R, Maciag T, Thompson JA. Extracellular FGF-1 acts as alens differentiation factor in transgenic mice. Development 1995;121:505–514.
Rosl F. A simple and rapid method for detection of apoptosis in humancells. Nucleic Acids Res. 1992;20:52–43.
Rotello RJ, Fernandez PA, Yuan J. Anti-apogens and anti-engulfens:monoclonal antibodies reveal specific antigens on apoptotic andengulfment cells during chicken embryonic development. Develop-ment 1994;120:1421–1431.
332 MITCHELL ET AL.

Smith J, Fowkes G, Schofield PN. Programmed cell death in dystro-phic (Mdx) muscle is inhibited by Igf-II. Cell Death Diff. 1995;2:243–251.
Stone J, Chan-Ling T, Pe’er J, Itin A, Gnessin H, Keshet E. Roles ofvascular endothelial growth factor and astrocyte degeneration inthe genesis of retinopathy of prematurity. Invest. Ophthalmol. Vis.Sci. 1996;37:290–299.
Stromblad S, Cheresh DA. Integrins, angiogenesis and vascular cellsurvival. Chem. Biol. 1996;3:881–885.
Terman BI, Dougher-Vermazen M, Carrion ME, Dimitrov D, Armel-lino DC, Gospodarowicz D, Bohlen P. Identification of the KDRtyrosine kinase as a receptor for vascular endothelial cell growthfactor. Biochem. Biophys. Res. Commun. 1992;187:1579–1586.
Vecchi A, Garlanda C, Lampugnani M G, Resnati M, Matteucci C,Stoppacciaro A, Schnurch H, Risau W, Ruco L, Mantovani A, DejanaE. Monoclonal antibodies specific for endothelial cells of mouse bloodvessels. Their application in the identification of adult and embry-onic endothelium. Eur. J. Cell Biol. 1994;63:247–254.
Williams GT, Smith CA, Spooncer E, Dexter TM, Taylor DR. Haemopoi-etic colony stimulating factors promote cell survival by suppressingapoptosis. Nature 1990;343:76–79.
Yamaguchi TP, Dumont DJ, Conlon RA, Breitman ML, Rossant J.Flk-1, an flt-related receptor tyrosine kinase is an early marker forendothelial cell precursors. Development 1993;118:489–498.
Yamamoto K, Fujimoto S. Phagocytic activity of hyalocytes in postna-tal hyaloid artery. Jpn. J. Ophthalmol. 1984;28:189–195.
333APOPTOSIS IN ENDOTHELIUM OF THE TVL
Copyright © 2022 FDOKUMEN




















