
Slow relaxation in microcanonical warming of a Ising lattice
Upload
independentCategory
view
1download
0
Tmf
SRD
a
ARRA
KETPNEfR
1
n
nd[saicse
1h
Pharmacological Research 78 (2013) 18– 27
Contents lists available at ScienceDirect
Pharmacological Research
jo ur nal home p age: www.elsev ier .com/ locate /yphrs
RPV4 channel activation leads to endothelium-dependent relaxationediated by nitric oxide and endothelium-derived hyperpolarizing
actor in rat pulmonary artery
usanth V. Sukumaran, Thakur Uttam Singh ∗, Subhashree Parida, Ch.E. Narasimha Reddy,amasamy Thangamalai, Kannan Kandasamy, Vishakha Singh, Santosh Kumar Mishra
ivision of Pharmacology and Toxicology, Indian Veterinary Research Institute, Izatnagar 243122, Bareilly, Uttar Pradesh, India
r t i c l e i n f o
rticle history:eceived 21 May 2013eceived in revised form 27 August 2013ccepted 12 September 2013
eywords:ndotheliumRPV4 channelsulmonary arteryitric oxidendothelium-derived hyperpolarizingactorelaxation
a b s t r a c t
The purpose of the present study was to characterize TRPV4 channels in the rat pulmonary arteryand examine their role in endothelium-dependent relaxation. Tension, Real-Time polymerase chainreaction (Real-Time PCR) and Western blot experiments were conducted on left and right branchesof the main pulmonary artery from male Wistar rats. TRPV4 channel agonist GSK1016790A (GSK)caused concentration-related robust relaxation (Emax 88.6 ± 5.5%; pD2 8.7 ± 0.2) of the endothelium-intact pulmonary artery. Endothelium-denudation nearly abolished the relaxation (Emax 5.6 ± 1.3%)to GSK. TRPV4 channel selective antagonist HC067047 significantly attenuated GSK-induced relax-ation (Emax 56.2 ± 6.6% vs. control Emax 87.9 ± 3.3%) in endothelium-intact vessels, but had no effecton either ACh-induced endothelium-dependent or SNP-induced endothelium-independent relaxations.GSK-induced relaxations were markedly inhibited either in the presence of NO synthase inhibitor L-NAME (Emax 8.5 ± 2.7%) or sGC inhibitor ODQ (Emax 28.1 ± 5.9%). A significant portion (Emax 30.2 ± 4.4%)of endothelium-dependent relaxation still persisted in the combined presence of L-NAME and cyclooxy-genase inhibitor indomethacin. This EDHF-mediated relaxation was sensitive to inhibition by 60 mM K+
depolarizing solution or K+ channel blockers apamin (SKCa; KCa2.3) and TRAM-34 (IKCa; KCa3.1). GSK−10 −7
(10 − 10 M) caused either modest decrease or increase in the basal tone of endothelium-intactor denuded rings, respectively. We found a greater abundance (>1.5 fold) of TRPV4 mRNA and pro-tein expressions in endothelium-intact vs. denuded vessels, suggesting the presence of this channel inpulmonary endothelial and smooth muscle cells as well. The present study demonstrated that NO andEDHF significantly contributed to TRPV4 channel-mediated endothelium-dependent relaxation of the ratpulmonary artery.
. Introduction
Transient receptor potential vanilloid 4 (TRPV4) channels areonselective cation channels which have moderate selectivity for
Abbreviations: TRPV4, transient receptor potential vanilloid 4; L-NAME, NG-itro-l-arginine methyl ester; PE, phenylephrine; TRAM 34, 1-[(2-chlorophenyl)iphenylmethyl]-1H-pyrazole; GSK, GSK1016790A; ODQ, 1H-[1,2,4]oxadiazolo4,3-a]quinoxalin-1-one; DMSO, dimethyl sulphoxide; SNP, sodium nitroprus-ide; EDHF, endothelium-derived hyperpolarizing factor; ACh, acetylcholine; ATP,denosine triphosphate; 4�-PDD, 4�-phorbol-12,13-didecanoate; EET, epoxye-cosatrienoic acids; SKCa, small conductance potassium channel; IKCa, intermediateonductance potassium channel; MKHS, modified Krebs–Henseleit solution; sGC,oluble guanylyl cyclase; NO, nitric oxide; PCR, polymerase chain reaction; ECs,ndothelial cells; ANOVA, analysis of variance.∗ Corresponding author. Tel.: +91 581 2300291; fax: +91 581 2303284.
E-mail addresses: [email protected], [email protected] (T.U. Singh).
043-6618/$ – see front matter © 2013 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.phrs.2013.09.005
© 2013 Elsevier Ltd. All rights reserved.
Ca2+ (PCa/PNa = 6). These channels are present both in endothelialand vascular cells and are implicated in the regulation of vascu-lar tone [1,2]. It is well established that the interplay betweenendothelial cells (ECs) and the underlying vascular cells plays acritical role in determining vascular tone, regional blood flow andarterial pressure in different vascular beds. Vasoactive agonistslike acetylcholine (ACh), adenosine triphosphate (ATP), bradykininand mechanical stimuli, such as flow and shear stress causea rapid increase in intracellular Ca2+ leading to synthesis andrelease of endothelium-derived vasodilators like nitric oxide (NO),endothelium-derived hyperpolarizing factor (EDHF) and prostac-yclin. In several vascular beds, a rise in intracellular Ca2+ inECs activates the enzyme eNOS and cytosolic phospholipase A2
(that activates cyclooxygenase) for the synthesis of endothelium-derived vasodilators NO and prostacyclins, respectively. However,the factors/mechanisms mediating EDHF response in blood vesselsremain elusive. Nevertheless, accumulating evidence points to the
acolog
flc[aaim
liieamavtipTac4ibec
vmacFmebcm(ifdcrvnAncam
eeitoaretlhnw
S.V. Sukumaran et al. / Pharm
act that EDHF response is strictly dependent on activation of vascu-ar endothelial intermediate conductance (IKCa, KCa3.1) and smallonductance (SKCa, KCa2.3) Ca2+-sensitive potassium (K+) channels3]. Recent studies provide evidence that these channels in ECs arectivated by local Ca2+ signals/events (Ca2+ sparklets) [4,5]. Uponctivation of these potassium channels, the underlying hyperpolar-zing current from ECs then spreads to smooth muscle cells through
yoendothelial gap junctions leading to vasodilation [6].In ECs, the main mechanisms involved in the rise of intracellu-
ar calcium include Ca2+ release from intracellular stores and Ca2+
nflux through store-operated and Ca2+ permeable cation channelsn the plasma membrane [7]. In recent years, there is increasingvidence that TRPV4 channels provide a potential Ca2+ perme-ble mechanism in vascular ECs for the activation of vasodilatorechanisms [5,8]. A large number of physical and chemical stimuli
ctivate this channel to cause release of endothelium-derivedasodilators and regulate vascular tone. For example, vasodila-ion in response to shear stress in mouse mesenteric arteriesnvolved TRPV4 channels. Thus, both nitric oxide and EDHF com-onents of flow-mediated relaxations were markedly reduced inRPV4 (−/−) mice in comparison to wild type control [8]. Inddition to mechanical stimulus, chemical stimuli such as endo-annabinoids, arachidonic acid, acetylcholine and TRPV4-agonist�-phorbol-12,13-didecanoate (4�-PDD) have been reported to
nduce endothelium-dependent relaxation in peripheral and cere-ral blood vessels [9]. However, the role of TRPV4 channels inndothelium-dependent relaxation of pulmonary artery is notlear.
The presence of TRPV4 channels has also been demonstrated inascular smooth muscle cells of rat cerebral artery [10], smoothuscle cells of rat pulmonary arteries [11,12], and rat and mouse
ortic smooth muscle cells [13]. However depending on the vas-ular beds, TRPV4 channels regulate the vascular tone differently.or instance, activation of TRPV4 channels in vascular smoothuscle cells of rat cerebral and mouse mesenteric arteries with
ndogenous ligand epoxyeicosatrienoic acids (EET) leads to mem-rane hyperpolarization and vasodilation involving BKCa (KCa1.1)hannels [14]. According to a recently published work on rat cre-aster arterioles (myogenically active) and mesenteric arteries
myogenically not active), it has been demonstrated that decreasingntraluminal pressure activated TRPV4 channels to increase therequency of endothelial cell Ca2+ events leading to endothelium-ependent vasodilation [5]. Previous studies provide evidence thatonstant release of endothelial vasodilators maintains low arte-ial pressure in the pulmonary vascular bed [15]. Therefore, it isery much relevant to understand the contribution of TRPV4 chan-els to endothelium-dependent vasodilation in pulmonary arteries.ccording to a recent study, however, activation of TRPV4 chan-els with specific agonist 4�-PDD increased the intracellular Ca2+
oncentration through ryanodine receptor (RyR2) in rat pulmonaryrtery smooth muscle cells, and caused a modest increase in pul-onary artery tone [16].TRPV4 channels have been identified in the microvascular
ndothelium of mouse lungs. Activation of these channels increasesndothelial permeability [17]. TRPV4 channels have also beendentified in human pulmonary artery endothelial cells [18], butheir role in the control of vascular tone is not clear. We andthers have previously demonstrated that NO and EDHF playn important role in endothelium-dependent relaxation of theat main pulmonary artery [19–21]. Therefore, using rat as anxperimental model, the first objective of the present study waso characterize the presence of TRPV4 channels in the vascu-
ar endothelium in this vessel. To achieve this objective, weave used selective agonist (GSK1016790A) [22] and antago-ist (HC067047) [23] of TRPV4 channels in this study. Further,e determined the relative expression of TRPV4 channel mRNAical Research 78 (2013) 18– 27 19
and protein in endothelium-intact and denuded vessels. The sec-ond objective was to determine the role of these channels inendothelium-dependent mechanisms regulating the tone of thisvessel. Appropriate pharmacological tools have been employedto delineate endothelium-dependent pathways activated throughTRPV4 channels. These included nitric oxide synthase inhibitor NG-nitro-l-arginine methyl ester (L-NAME), cyclooxygenase inhibitorindomethacin, small conductance potassium channel (SKCa) chan-nel inhibitor apamin and intermediate conductance potassiumchannel (IKCa) inhibitor TRAM-34.
2. Materials and methods
2.1. Animals
Healthy adult male Wistar rats (150–200 g) were procuredfrom the Laboratory Animal Resource Section, Indian VeterinaryResearch Institute, Izatnagar, U.P., India. Animals were kept foracclimatization for a period of seven days before conduction ofexperiments. All protocols, employed in this study, were approvedby the Institutional Animal Ethics Committee, Indian VeterinaryResearch Institute, Izatnagar.
2.2. Tension recording
The animals were bled to death by excising the abdominal aortaunder urethane (1.2 g/kg body weight intraperitoneally) anesthe-sia. Heart and lungs en bloc were removed and transferred to icecold Modified Krebs–Henseleit solution (MKHS) of the followingcomposition (in mM): NaCl 118, KCl 4.7, CaCl2 2.5, MgSO4 1.2,NaHCO3 11.9, KH2PO4 1.2 and d-glucose 11.1 (pH 7.4). Both the leftand right branches of the main pulmonary artery were dissectedcarefully and cleaned of adhering connective tissues under a dis-secting microscope. The arterial segments were mounted on twostainless steel hooks and suspended in 10 ml organ baths containingMKHS, maintained at 37 ◦C, and aerated continuously with medi-cal gas (21% O2 + 5% CO2 + 71% N2) mixture. A passive tension of1.0 g [24] was applied during the equilibration period of 90 min andthe bath solution was changed every 15 min. Tension was recordedusing a high sensitivity isometric force transducer and stored in acomputer using Chart version 6.1.3 software program (Powerlab,AD Instruments, Bella Vista, NSW, Australia) for further analysis.
After the equilibration period, tissue viability was examined byrecording the contraction to high K+ (80 mM)-depolarizing solu-tion. KCl 80 mM was prepared as normal MKHS but with equimolarsubstitution of NaCl with KCl. Cumulative concentration–responsecurves to relaxants were elicited in arterial segments pre-contracted with a sub-maximal concentration of phenylephrine(0.1–3 �M) to achieve near identical pre-contraction levels.Endothelial integrity was examined by adding 1 �M of ACh at theplateau of phenylephrine contraction. The arterial rings were con-sidered endothelium-intact, if 1 �M ACh caused >80% reversal ofphenylephrine-induced contraction. In the endothelium-denudedpreparations, relaxation response to 1 �M of ACh was either absentor it was less than 10% of the maximal reversal of 1 �M ofphenylephrine-induced contraction. Endothelial denudation wasachieved by gently passing a wet cotton swab through the lumenof the arterial ring. Endothelium removal, pretreatment with L-NAME or ODQ sensitized the pre-contractions to phenylephrinebefore eliciting relaxation responses to different vasodilators. Onthe other hand, indomethacin attenuated contractile responses
to phenylephrine. Accordingly, an appropriate concentration ofphenylephrine, as stated above, was used to achieve matching con-traction level of the arterial segments before eliciting relaxationsto vasodilators.
2 acolog
2
2a
rapr
2S
aetoacectSa
2po
ecp1apyTotbcrf
2E
rlpccipSip
2cp
e-
0 S.V. Sukumaran et al. / Pharm
.3. Experimental protocols
.3.1. Role of endothelium in GSK1016790A-induced pulmonaryrtery relaxation
To elucidate the role of vascular endothelium in GSK-mediatedelaxation, a concentration–response curve to TRPV4 channelgonist GSK (10−10 − 10−7 M) was elicited in phenylephrinere-contracted endothelium-intact/endothelium-denuded arterialings of the rat pulmonary artery.
.3.2. Role of TRPV4 channel on GSK1016790A, ACh andNP-induced relaxation in pulmonary artery
To confirm the role of TRPV4 channel in GSK-mediated relax-tion, a concentration–response curve to GSK (10−10–10−7 M) waslicited in arterial rings pretreated for 30 min with 1 �M of a selec-ive TRPV4 channel antagonist HC067047. To examine the rolef TRPV4 channels in mediating endothelium-dependent relax-tion to muscarinic receptor agonist ACh, concentration–responseurve to ACh (10−9–10−4.5 M) was elicited either in the pres-nce or absence of 1 �M of HC067047. Specificity of TRPV4hannel antagonism by HC067047 was determined by elici-ing relaxation response to endothelium-independent vasodilatorNP (10−9–10−5 M) either in the presence or absence of thentagonist.
.3.3. Elucidation of the role of NO, prostacyclin and EDHFathways in TRPV4-mediated GSK1016790A-induced relaxationf rat pulmonary artery
In order to examine the role of NO-sGC pathway in GSK-inducedndothelium-dependent relaxation, a concentration–responseurve to GSK (10−10–10−7 M) was elicited in phenylephrinere-contracted pulmonary arterial rings pretreated with either00 �M L-NAME or 10 �M ODQ for 30 min. Indomethacin is
non selective COX inhibitor which inhibits synthesis ofrostanoids. To evaluate the contribution of endothelial prostac-clin to GSK-induced relaxation, the relaxant responses to theRPV4 agonist was elicited either in the presence or absencef 10 �M indomethacin (30 min pre-incubation). The contribu-ion of EDHF pathway to GSK-induced relaxation was evaluatedy eliciting concentration-dependent relaxation to the TRPV4hannel agonist on phenylephrine-contracted pulmonary arteryings pre-treated with 100 �M L-NAME plus 10 �M indomethacinor 30 min.
.3.4. Effect of SKCa and IKCa channel blockers or 60 mM K+ onDHF response elicited by GSK1016790A in rat pulmonary artery
In order to first confirm that EDHF contributes to the relaxantesponse elicited by GSK through vascular smooth muscle hyperpo-arization, the arterial rings were incubated with100 �M L-NAMElus 10 �M indomethacin for 30 min and a concentration–responseurve to GSK (10−10–10−7 M) was developed in 60 mM K+ pre-ontracted rings. To elucidate the role of SKCa and IKCa channelsn EDHF-mediated relaxation response to GSK (10−10–10−7 M), theulmonary arterial rings were incubated with specific blockers ofKCa (apamin) plus IKCa channels (TRAM 34) for 30 min before elic-ting the concentration–response curve to GSK on phenylephrinere-contracted tissues.
.3.5. Relative mRNA and protein expression study of TRPV4hannels in endothelium-intact and endothelium-denuded
ulmonary arteryWe employed Real-Time PCR technique to examine relativexpression of TRPV4 channels mRNA in endothelium-intact anddenuded pulmonary arteries.
ical Research 78 (2013) 18– 27
2.4. mRNA expression study
2.4.1. Total RNA isolationPulmonary artery was isolated in 0.1% diethyl pyrocarbonate-
treated and autoclaved phosphate buffer saline (PBS). Tissues werestored in RNA later solution until use. During use, the tissues weresnap frozen in liquid nitrogen and processed for RNA isolation.Total RNA was isolated with RNA isolation kit (IBI Scientific, USA)as per the manufacturer’s instructions. The purity of the RNA waschecked by A260/A280 ratio and A230/A260 ratio in a nanodropand quantified as follows: 1OD = 40 �g/ml.
2.4.2. Quantitative Real-Time PCR reactionReal-Time PCR was conducted using SYBR Green I master
mix (KAPATM SYBR Fast qPCR Master Mix (2X) Universal, KAPABiosystems, USA). Each sample was run in duplicate in 20 �lreaction mixture. The 20 �l reaction mixture consisted of 10 �lSYBR Green master mix, 1.0 �l from 5 pmoles/�l stock solu-tion of each of gene specific forward and reverse primers, 1 �lof cDNA and volume was made up to 20 �l with nuclease-freewater. The Real-Time PCR reaction started with initial incuba-tion at 95 ◦C for 10 min followed by 35 cycles of amplificationwith denaturation at 95 ◦C for 35 s, annealing (temperature var-ied depending on primer) for 30 s and extension at 72 ◦C for30 s each. The optimum annealing temperatures were deter-mined by PCR for the respective gene using the specific primersas follows, for the TRPV4 gene (Acc. No. NM 023970) wereF 5′-AGAAAGCGCCCATGGATT-3′, R 5′-TCTGTGGCTGCTTCTCTACG-3′ (102 bp) [25] at 60 ◦C and for the GAPDH gene (Acc.No. GU 214026.1) were F5′-AACTTTGGCATTGTGGAAGG-3′, R5′-ACACATTGGGGGTAGGAACA-3′ (223 bp) at 56 ◦C, respectively. Toassess the specificity of the amplified product, dissociation curvewas generated at temperature of 55 ◦C through 95 ◦C. The resultwas expressed as threshold cycle values (CT). This value is the cyclenumber when the fluorescence of the reporter dye is appreciablyhigher than the background fluorescence. The threshold, automat-ically adjusted by the instrument, was used for the generation ofCT values.
2.5. Protein expression study by Western blot
Endothelium-intact and endothelium-denuded pulmonaryarteries were frozen in liquid nitrogen. The frozen pulmonaryarteries were crushed and homogenized with ice-cold lysis buffer(100 mM Tris–HCl, pH 7.5; 100 mM EDTA, pH 7.5; 100 mM PMSF;1% Triton X-100; 10 �l of 100× protease inhibitor cocktail) usinghomogenizer. The homogenate was centrifuged at 4 ◦C with1000 × g for 5 min, the supernatant [25] was collected, and Brad-ford protein assay kit was used for the measurement of protein inthe samples. Aliquots of the samples (40 �g of protein) was dilutedin loading buffer and denatured for 3 min at 100 ◦C in boiling waterbath. Samples were separated by gradient sodium dodecyl sul-fate (SDS)–polyacrylamide gel electrophoresis (SDS/PAGE) usinga GeiNeiTM, High power, DC power supply system. Proteinswere electrophoretically transferred onto polyvinylidene difluo-ride (PVDF) membrane using GeNeiTM, Electro Transfer MidiDual-08-04 system (2 h at 100 V in 25 mM Tris, 192 mM glycineand 20% methanol). After transfer, the membrane was washed fourtimes for 20 min duration with phosphate buffered saline–Tween-20 (PBS-T containing PBS, pH 7.4 and 0.05% Tween-20, v/v) andblocked for 2 h at room temperature in blocking buffer containing5% skimmed milk powder in PBS (w/v). Blots were then incubated
for overnight at 4 ◦C with primary antibody in PBS against TRPV4channel (NB100-74960, Novus Biologicals, USA) at 1:500 dilutionand GAPDH (SC-25778, Santa Cruz Biotechnology, USA) at 1:200dilutions in PBS-T. The primary antibody was removed and blot was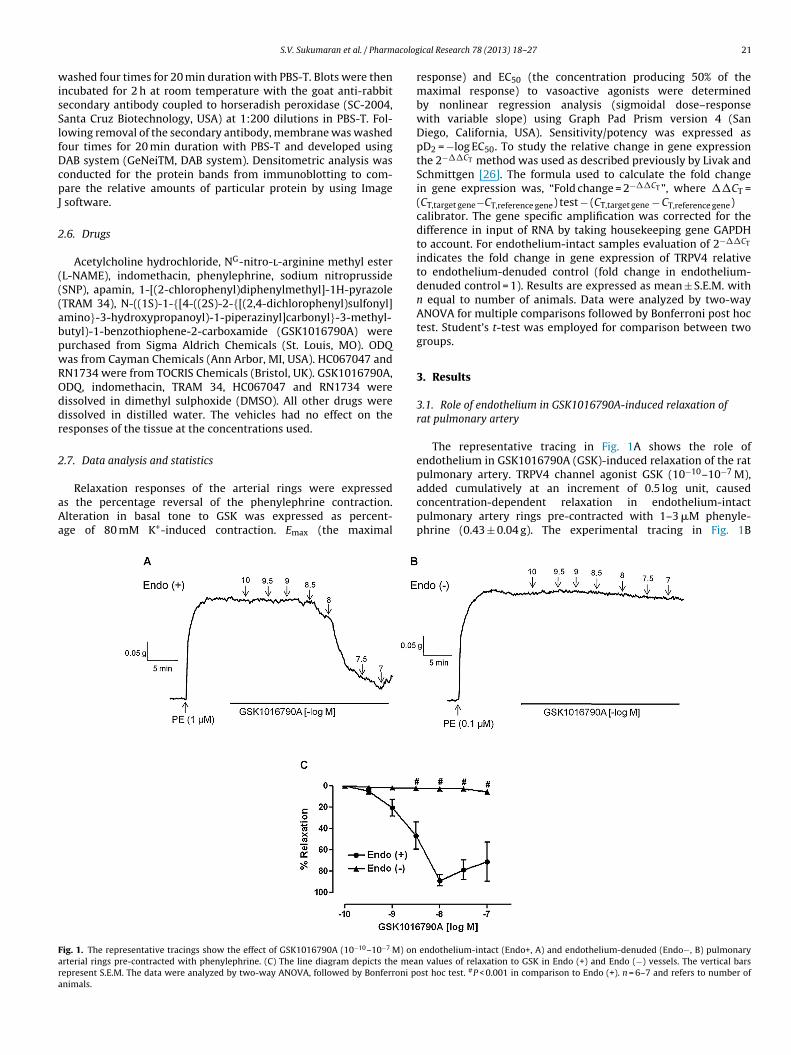
acolog
wisSlfDcpJ
2
(((abpwROddr
2
aAa
Fara
S.V. Sukumaran et al. / Pharm
ashed four times for 20 min duration with PBS-T. Blots were thenncubated for 2 h at room temperature with the goat anti-rabbitecondary antibody coupled to horseradish peroxidase (SC-2004,anta Cruz Biotechnology, USA) at 1:200 dilutions in PBS-T. Fol-owing removal of the secondary antibody, membrane was washedour times for 20 min duration with PBS-T and developed usingAB system (GeNeiTM, DAB system). Densitometric analysis wasonducted for the protein bands from immunoblotting to com-are the relative amounts of particular protein by using Image
software.
.6. Drugs
Acetylcholine hydrochloride, NG-nitro-l-arginine methyl esterL-NAME), indomethacin, phenylephrine, sodium nitroprussideSNP), apamin, 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazoleTRAM 34), N-((1S)-1-{[4-((2S)-2-{[(2,4-dichlorophenyl)sulfonyl]mino}-3-hydroxypropanoyl)-1-piperazinyl]carbonyl}-3-methyl-utyl)-1-benzothiophene-2-carboxamide (GSK1016790A) wereurchased from Sigma Aldrich Chemicals (St. Louis, MO). ODQas from Cayman Chemicals (Ann Arbor, MI, USA). HC067047 andN1734 were from TOCRIS Chemicals (Bristol, UK). GSK1016790A,DQ, indomethacin, TRAM 34, HC067047 and RN1734 wereissolved in dimethyl sulphoxide (DMSO). All other drugs wereissolved in distilled water. The vehicles had no effect on theesponses of the tissue at the concentrations used.
.7. Data analysis and statistics
Relaxation responses of the arterial rings were expresseds the percentage reversal of the phenylephrine contraction.lteration in basal tone to GSK was expressed as percent-ge of 80 mM K+-induced contraction. Emax (the maximal
ig. 1. The representative tracings show the effect of GSK1016790A (10−10–10−7 M) onrterial rings pre-contracted with phenylephrine. (C) The line diagram depicts the meaepresent S.E.M. The data were analyzed by two-way ANOVA, followed by Bonferroni ponimals.
ical Research 78 (2013) 18– 27 21
response) and EC50 (the concentration producing 50% of themaximal response) to vasoactive agonists were determinedby nonlinear regression analysis (sigmoidal dose–responsewith variable slope) using Graph Pad Prism version 4 (SanDiego, California, USA). Sensitivity/potency was expressed aspD2 = −log EC50. To study the relative change in gene expressionthe 2−��CT method was used as described previously by Livak andSchmittgen [26]. The formula used to calculate the fold changein gene expression was, “Fold change = 2−��CT ”, where ��CT =(CT,target gene−CT,reference gene) test − (CT,target gene − CT,reference gene)calibrator. The gene specific amplification was corrected for thedifference in input of RNA by taking housekeeping gene GAPDHto account. For endothelium-intact samples evaluation of 2−��CT
indicates the fold change in gene expression of TRPV4 relativeto endothelium-denuded control (fold change in endothelium-denuded control = 1). Results are expressed as mean ± S.E.M. withn equal to number of animals. Data were analyzed by two-wayANOVA for multiple comparisons followed by Bonferroni post hoctest. Student’s t-test was employed for comparison between twogroups.
3. Results
3.1. Role of endothelium in GSK1016790A-induced relaxation ofrat pulmonary artery
The representative tracing in Fig. 1A shows the role ofendothelium in GSK1016790A (GSK)-induced relaxation of the ratpulmonary artery. TRPV4 channel agonist GSK (10−10–10−7 M),
added cumulatively at an increment of 0.5 log unit, causedconcentration-dependent relaxation in endothelium-intactpulmonary artery rings pre-contracted with 1–3 �M phenyle-phrine (0.43 ± 0.04 g). The experimental tracing in Fig. 1Bendothelium-intact (Endo+, A) and endothelium-denuded (Endo−, B) pulmonaryn values of relaxation to GSK in Endo (+) and Endo (−) vessels. The vertical barsst hoc test. #P < 0.001 in comparison to Endo (+). n = 6–7 and refers to number of

2 acolog
iidaerIwmwc
3Gp
eHptpnHua
itpwch99
FaeA
2 S.V. Sukumaran et al. / Pharm
llustrates that GSK-induced relaxation was nearly abolishedn endothelium-denuded vessels. The line diagram in Fig. 1Cepicts concentration–response curves of GSK-induced relax-tion in endothelium-intact and -denuded arterial rings. Inndothelium-intact vessels, GSK elicited concentration-dependentelaxation with Emax of 88.6 ± 5.5% and pD2 of 8.7 ± 0.2 (n = 6).n endothelium-denuded vessels, the precontraction level
ith 0.1 �M phenylephrine was 0.43 ± 0.04 g, and the maxi-al relaxation produced by GSK was 5.6 ± 1.3% (n = 7), whichas significantly (P < 0.001) different from endothelium-intact
ontrol.
.2. Effect of HC067047, a selective blocker of TRPV4 channel, onSK1016790A, acetylcholine and SNP-induced relaxations ofulmonary artery rings
Fig. 2A shows that 1 �M HC067047 attenuated GSK-inducedndothelium-dependent relaxations. Incubation of tissues withC067047 for 30 min as such had no effect on basal tone of the ratulmonary artery. It had no significant effect on the precontrac-ion tone (0.39 ± 0.03 g vs. control, 0.37 ± 0.02 g) elicited by 1 �Mhenylephrine. The maximal relaxation to GSK, however, was sig-ificantly (P < 0.05) reduced (56.2 ± 6.6%; n = 7) in the presence ofC067047 in comparison to control (87.9 ± 3.3%, n = 6). The pD2 val-es of GSK in the presence and absence of HC067047 were 7.2 ± 0.2nd 8.5 ± 0.2, respectively.
HC067047 was used to elucidate the role of TRPV4 channeln mediating ACh-induced endothelium-dependent relaxations inhe rat pulmonary artery. In the presence of HC067047, 1 �Mhenylephrine-induced precontraction tone of the arterial ringsas 0.32 ± 0.03 g, which was not significantly different from the
ontrols (0.30 ± 0.03 g). As shown in Fig. 2B, 1 �M HC067047ad no effect on ACh-induced relaxations (pD2, 6.6 ± 0.01; Emax,6.0 ± 2.9%; n = 7) in comparison to control (pD2, 6.5 ± 0.1; Emax,7.3 ± 3.5%; n = 6). In order to further confirm that TRPV4 channels
ig. 2. Effect of a selective TRPV4 antagonist HC067047 (1 �M) on GSK1016790A (10−
tions in endothelium-intact pulmonary arterial rings. Effect of RN1734 (another selectivndothelium-intact pulmonary arterial rings. Line diagrams depict the mean value of GSKNOVA, followed by Bonferroni post hoc test. *P < 0.05 and #P < 0.001 in comparison to co
ical Research 78 (2013) 18– 27
are not involved in ACh-mediated endothelium-dependent relax-ation of the rat pulmonary artery, we did some experiments withanother selective blocker of TRPV4 channels, RN1734 [27]. Incuba-tion of the tissues with 20 �M RN1734 for 30 min had no significanteffect on the basal tone. In the presence of RN1734, 1 �M phenyle-phrine caused a precontraction tone of 0.27 ± 0.02 g, which was notsignificantly different from the controls (0.26 ± 0.02 g). Fig. 2C illus-trates that RN1734 had no significant effect on the concentration-dependent relaxation to ACh. Thus, the pD2 and Emax of ACh in thepresence of RN1734 were 7.2 ± 0.1 and 101.6 ± 1.8%, respectively(n = 6) vs. control (pD2 7.1 ± 0.1; Emax 97.9 ± 1.8%; n = 6).
To illustrate that the TRPV4 channel blocker specifically antag-onized the activation of endothelial TRPV4 channels by GSK inthe rat pulmonary artery, the effect of HC067047 was assessedon relaxations induced by endothelium-independent vasodilatorsodium nitroprusside (SNP). Fig. 2D illustrates that 1 �M HC067047had no effect on SNP-induced relaxations (pD2, 7.4 ± 0.1 and Emax,107.6 ± 2.3%; n = 6 vs. control; pD2, 7.3 ± 0.1 and Emax, 112.6 ± 2.1%;n = 7).
3.3. Role of nitric oxide, prostacyclin and EDHF inGSK1016790A-mediated relaxation of pulmonary artery
In order to elucidate the contribution of NO to GSK-inducedendothelium-dependent relaxation in the rat pulmonary artery,relaxation to the TRPV4 agonist was elicited in the pres-ence and absence of L-NAME. Pulmonary arterial rings wereincubated with 100 �M L-NAME for 30 min before elicitingthe concentration–response curve to GSK in phenylephrine-precontracted tissues. L-NAME significantly increased (26.5 ± 7.5%of 80 mM KCl, n = 10) the basal tone of the pulmonary artery. In
the presence of 100 �M L-NAME, 0.1 �M phenylephrine-inducedpre-contraction tone was 0.37 ± 0.03 g, which was not significantlydifferent from control (0.37 ± 0.04 g) elicited with 1–3 �M phenyle-phrine. As shown in Fig. 3A, the maximal relaxation elicited by GSK10–10−7 M; A), ACh (10−9–10−4.5 M; B) and SNP (10−9–10−5 M; D)-induced relax-e antagonist of TRPV4 channel) on ACh (10−9 − 10−4.5 M; C)-induced relaxations in
relaxation. The vertical bars represent S.E.M. The data were analyzed by two-wayntrol. n = 6–7.
S.V. Sukumaran et al. / Pharmacological Research 78 (2013) 18– 27 23
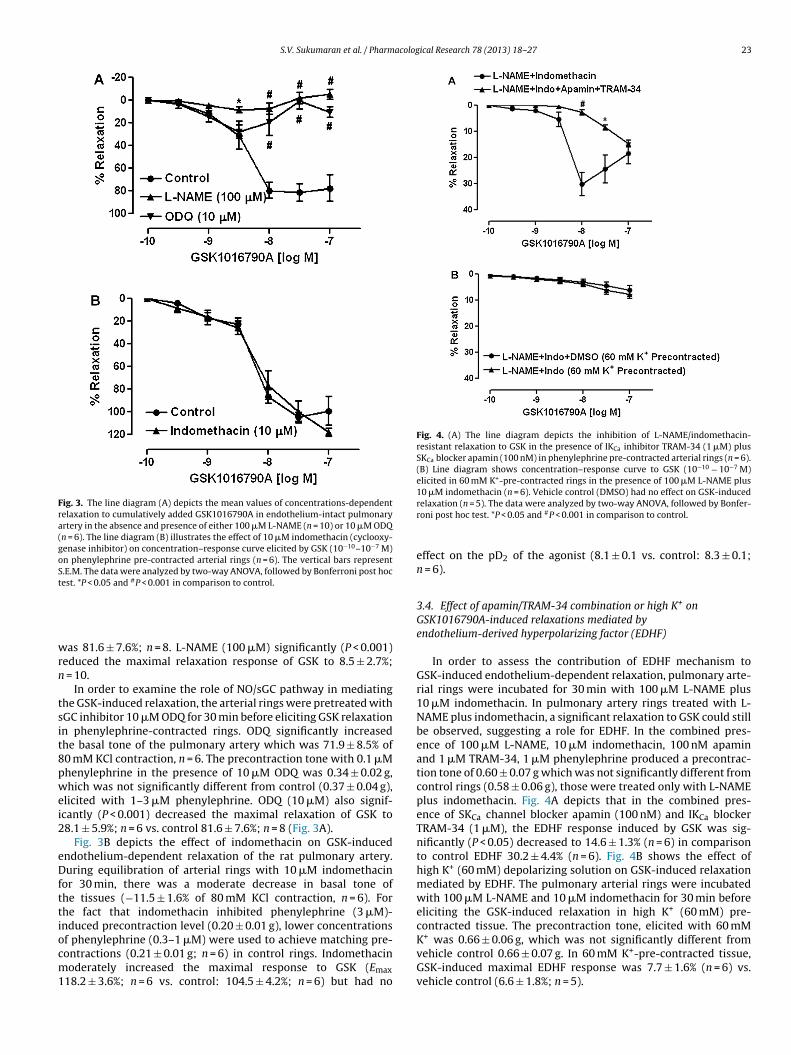
Fig. 3. The line diagram (A) depicts the mean values of concentrations-dependentrelaxation to cumulatively added GSK1016790A in endothelium-intact pulmonaryartery in the absence and presence of either 100 �M L-NAME (n = 10) or 10 �M ODQ(n = 6). The line diagram (B) illustrates the effect of 10 �M indomethacin (cyclooxy-genase inhibitor) on concentration–response curve elicited by GSK (10−10–10−7 M)on phenylephrine pre-contracted arterial rings (n = 6). The vertical bars representS.E.M. The data were analyzed by two-way ANOVA, followed by Bonferroni post hoctest. *P < 0.05 and #P < 0.001 in comparison to control.
wrn
tsit8pwei2
eDfttiocm1
Fig. 4. (A) The line diagram depicts the inhibition of L-NAME/indomethacin-resistant relaxation to GSK in the presence of IKCa inhibitor TRAM-34 (1 �M) plusSKCa blocker apamin (100 nM) in phenylephrine pre-contracted arterial rings (n = 6).(B) Line diagram shows concentration–response curve to GSK (10−10 − 10−7 M)elicited in 60 mM K+-pre-contracted rings in the presence of 100 �M L-NAME plus10 �M indomethacin (n = 6). Vehicle control (DMSO) had no effect on GSK-induced
contracted tissue. The precontraction tone, elicited with 60 mMK+ was 0.66 ± 0.06 g, which was not significantly different fromvehicle control 0.66 ± 0.07 g. In 60 mM K+-pre-contracted tissue,
as 81.6 ± 7.6%; n = 8. L-NAME (100 �M) significantly (P < 0.001)educed the maximal relaxation response of GSK to 8.5 ± 2.7%;
= 10.In order to examine the role of NO/sGC pathway in mediating
he GSK-induced relaxation, the arterial rings were pretreated withGC inhibitor 10 �M ODQ for 30 min before eliciting GSK relaxationn phenylephrine-contracted rings. ODQ significantly increasedhe basal tone of the pulmonary artery which was 71.9 ± 8.5% of0 mM KCl contraction, n = 6. The precontraction tone with 0.1 �Mhenylephrine in the presence of 10 �M ODQ was 0.34 ± 0.02 g,hich was not significantly different from control (0.37 ± 0.04 g),
licited with 1–3 �M phenylephrine. ODQ (10 �M) also signif-cantly (P < 0.001) decreased the maximal relaxation of GSK to8.1 ± 5.9%; n = 6 vs. control 81.6 ± 7.6%; n = 8 (Fig. 3A).
Fig. 3B depicts the effect of indomethacin on GSK-inducedndothelium-dependent relaxation of the rat pulmonary artery.uring equilibration of arterial rings with 10 �M indomethacin
or 30 min, there was a moderate decrease in basal tone ofhe tissues (−11.5 ± 1.6% of 80 mM KCl contraction, n = 6). Forhe fact that indomethacin inhibited phenylephrine (3 �M)-nduced precontraction level (0.20 ± 0.01 g), lower concentrationsf phenylephrine (0.3–1 �M) were used to achieve matching pre-ontractions (0.21 ± 0.01 g; n = 6) in control rings. Indomethacin
oderately increased the maximal response to GSK (Emax18.2 ± 3.6%; n = 6 vs. control: 104.5 ± 4.2%; n = 6) but had no
relaxation (n = 5). The data were analyzed by two-way ANOVA, followed by Bonfer-roni post hoc test. *P < 0.05 and #P < 0.001 in comparison to control.
effect on the pD2 of the agonist (8.1 ± 0.1 vs. control: 8.3 ± 0.1;n = 6).
3.4. Effect of apamin/TRAM-34 combination or high K+ onGSK1016790A-induced relaxations mediated byendothelium-derived hyperpolarizing factor (EDHF)
In order to assess the contribution of EDHF mechanism toGSK-induced endothelium-dependent relaxation, pulmonary arte-rial rings were incubated for 30 min with 100 �M L-NAME plus10 �M indomethacin. In pulmonary artery rings treated with L-NAME plus indomethacin, a significant relaxation to GSK could stillbe observed, suggesting a role for EDHF. In the combined pres-ence of 100 �M L-NAME, 10 �M indomethacin, 100 nM apaminand 1 �M TRAM-34, 1 �M phenylephrine produced a precontrac-tion tone of 0.60 ± 0.07 g which was not significantly different fromcontrol rings (0.58 ± 0.06 g), those were treated only with L-NAMEplus indomethacin. Fig. 4A depicts that in the combined pres-ence of SKCa channel blocker apamin (100 nM) and IKCa blockerTRAM-34 (1 �M), the EDHF response induced by GSK was sig-nificantly (P < 0.05) decreased to 14.6 ± 1.3% (n = 6) in comparisonto control EDHF 30.2 ± 4.4% (n = 6). Fig. 4B shows the effect ofhigh K+ (60 mM) depolarizing solution on GSK-induced relaxationmediated by EDHF. The pulmonary arterial rings were incubatedwith 100 �M L-NAME and 10 �M indomethacin for 30 min beforeeliciting the GSK-induced relaxation in high K+ (60 mM) pre-
GSK-induced maximal EDHF response was 7.7 ± 1.6% (n = 6) vs.vehicle control (6.6 ± 1.8%; n = 5).

24 S.V. Sukumaran et al. / Pharmacological Research 78 (2013) 18– 27
Fig. 5. Effect of GSK1016790A (GSK) on basal tone of rat pulmonary artery. The representative experimental tracing shows concentration-dependent decrease in basal tone tocumulatively added GSK (10−10–10−7 M) in endothelium-intact (A; n = 7) and concentration-dependent increase in basal tone to cumulatively added GSK (10−10–10−7 M) ine ean vai e data
3ea
a
FeGd
ndothelium-denuded (B; n = 7) pulmonary artery. (C) The line diagram depicts the mntact and -denuded arterial rings, respectively. The vertical bars represent S.E.M. Thnd # P < 0.001 in comparison to Endo (+).
.5. Effect of GSK1016790A on the basal tone ofndothelium-intact and endothelium-denuded rat pulmonary
rteryEffect of GSK on basal tone was determined by exposure of therterial rings to different concentrations of GSK (10−10–10−7 M)
ig. 6. Relative mRNA and protein expression study of TRPV4 channels in endothelium-dxpression (n = 4) and (B) protein expression of TRPV4 channel in endothelium-denuded
APDH and TRPV4 channel in endothelium-intact and -denuded rat pulmonary artery. Tenuded mRNA and protein expression.
lues of concentration-dependent relaxation and contraction to GSK in endothelium-a were analyzed by two-way ANOVA followed by Bonferroni post hoc test. *P < 0.05
in endothelium-intact as well as in endothelium-denuded rings.Raw tracings in Fig. 5A (Endo+) and B (Endo−) illustrate the
relaxant and contractile responses to GSK, respectively. Fig. 5Cshows concentration–response curves to GSK (10−10–10−7 M)added cumulatively at an increment of 0.5 log unit in endothelium-intact and -denuded arterial rings. The maximum relaxation to GSKenuded and -intact pulmonary artery. The bar diagram (A) depicts relative mRNAand endothelium-intact rat pulmonary artery (n = 4). (C) Representative blots showhe data were analyzed by Student’s t-test. *P < 0.05 as compared to endothelium-

S.V. Sukumaran et al. / Pharmacological Research 78 (2013) 18– 27 25
el ago
wcc
3ea
iarise
iaiieni
4
crwtcttfddieovi
arpot
Fig. 7. Schematic diagram depicts the pathways involved in TRPV4 chann
as 4.8 ± 2.2% (n = 7) in endothelium-intact vessel. The maximumontraction to GSK was 11.3 ± 4.9% (n = 7) of 80 mM K+-inducedontraction in endothelium-denuded rings.
.6. Relative expression of mRNA and protein of TRPV4 channel inndothelium-intact and endothelium-denuded rat pulmonaryrtery
Real-Time PCR study was performed to assess the fold changen the expression of TRPV4 channel in endothelium-denudednd endothelium-intact rat pulmonary artery. Fig. 6A depicts theelative expression of TRPV4 channel in this artery. In endothelium-ntact pulmonary artery, TRPV4 channel mRNA expression wasignificantly 1.9 ± 0.2 fold (P < 0.05; n = 4) greater in comparison tondothelium-denuded artery.
Fig. 6B and C depict the TRPV4 channel protein expressionn the endothelium-denuded and endothelium-intact pulmonaryrtery. The relative unit of protein expression (TRPV4/GAPDH)n endothelium-denuded artery was set to 1.0. The alterationn the protein expression in endothelium-intact artery has beenxpressed accordingly. TRPV4 channel protein expression was sig-ificantly (P < 0.05) higher in endothelium-intact (1.7 ± 0.2; n = 4)
n comparison to endothelium-denuded artery.
. Discussion
The major findings of the present study are as follows: (1) TRPV4hannel agonist GSK1016790A caused endothelium-dependentelaxation (>80%) of the rat pulmonary artery rings pre-contractedith phenylephrine; (2) endothelium denudation nearly abolished
he relaxation response to GSK in this vessel; (3) selectiveTRPV4hannel antagonist HC067047 attenuated the relaxation responseo GSK; (4) both nitric oxide and EDHF, but not prostacyclin con-ributed to GSK-induced relaxation; (5) we found no evidenceor the role of TRPV4 channels in ACh-induced endothelium-ependent relaxation in this vessel; (6) GSK caused modestecrease (∼5%) in the pulmonary artery basal tone in endothelium-
ntact rings, while it marginally increased the basal tone (∼11%) inndothelium-denuded vessels; (7) there was a greater abundancef the TRPV4 mRNA and protein expressions in endothelium-intacts. endothelium-denuded arteries. The implications of these find-ngs are discussed.
Our tension recording studies revealed that pharmacologicalctivation of TRPV4 channels with the selective agonist GSK caused
obust relaxation of endothelium-intact rat pulmonary artery ringsre-contracted with phenylephrine. Further, TRPV4 channel antag-nist HC067047 significantly attenuated the vasodilator responseo GSK. The selectivity of TRPV4 channel antagonism by HC067047nist GSK1016790A-induced relaxation of rat isolated pulmonary artery.
was confirmed by the observation that the antagonist did not mod-ify the endothelium-independent relaxation produced by sodiumnitroprusside in the pulmonary artery. These findings suggestthat TRPV4 channels are present on the endothelial cells of therat pulmonary artery and their activation leads to endothelium-dependent relaxation. The presence of these channels in vascularendothelial as well as smooth muscle cells is further confirmedby demonstrating a relatively greater expression of TRPV4 chan-nel mRNA as well as protein in endothelium-intact in comparisonto denuded vessels. According to a previous study, the presenceof TRPV4 channels mRNA and protein has been shown in therat pulmonary artery smooth muscle cells only [25]. The involve-ment of TRPV4 channels in endothelium-dependent relaxation ofrat pulmonary artery is consistent with previous reports show-ing that the activation of TRPV4 channel by its agonists causedendothelium-dependent vasodilation in rat carotid artery [28],mouse and rat aorta [29]. Acetylcholine has been reported tobe an endogenous agonist for TRPV4 channels in mouse mesen-teric and carotid arteries [30]. These authors have demonstratedthat in the TRPV4 knockout mouse model, acetylcholine-inducedendothelium-dependent vasodilation is reduced both in vivo andin vitro. In the present study, we observed that TRPV4 channelantagonist HC067047 did not modify ACh-induced endothelium-dependent relaxation in the rat pulmonary artery. It is furtherconfirmed by the finding that another TRPV4 selective blockerRN1734 had no significant effect on concentration-dependentendothelium-dependent relaxation to ACh in the rat pulmonaryartery. The reasons for this discrepancy in the activation of TRPV4channels by ACh in mouse mesenteric/carotid [30] vs. rat pul-monary artery (present study) remain to be determined, but couldbe due to differences in species and vascular beds. Such differencesin the activation of TRPV4 channels by endogenous ligands in differ-ent blood vessels/species have been reported recently. For example,arachidonic acid metabolite 11,12-EET activated TRPV4 channelsin the mouse mesenteric artery [4], but not in the human coronaryarterioles [27].
In pulmonary artery with active endothelium, inhibition ofNO synthase by L-NAME or inhibition of sGC by ODQ markedlyattenuated the vasodilator response to GSK, thereby suggestinga significant contribution of NO/sGC pathway in TRPV4 channel-mediated endothelium-dependent relaxation of the rat pulmonaryartery. Our finding is consistent with earlier studies showing TRPV4channel-linked elevation in intracellular Ca2+ and NO release inmouse mesenteric [30] and rat carotid [28] arteries. According to a
recent report, there is evidence for the role of endogenous pros-tacyclin in endothelium-dependent relaxation caused by TRPV1agonist, anandamide in the rat small pulmonary artery [31]. Weobserved that cyclooxygenase inhibitor indomethacin, however,
2 acolog
hro
rwtriiuTtEawtGmslid
ldwowwpf3padhetTStcenwtntltrsasr
cetcpimrt
[
[
[
[
[
[
[
[
[
6 S.V. Sukumaran et al. / Pharm
ad no inhibitory effect on GSK-induced endothelium-dependentelaxation in the rat main pulmonary artery, suggesting little rolef prostacyclin.
One of the intriguing observations is that the relaxationesponse to GSK in the presence of L-NAME was ∼10%, whichas significantly potentiated (∼30%) when the tissues were
reated with L-NAME plus indomethacin to elicit EDHF-mediatedelaxation. Thus, the augmentation of the EDHF response byndomthacin indicates that GSK in the absence of the COX inhibitors releasing some endothelial contractile prostanoid that is atten-ating the EDHF-mediated relaxation in the rat pulmonary artery.his hypothesis is supported by a previous report, which showedhat endothelial thromboxane A2 inhibited SKCa component ofDHF response after NO synthase block in the rat middle cerebralrtery [32]. We also demonstrated that similar to indomethacinhich inhibits thromboxane A2 synthesis, thromboxane A2 recep-
or antagonist ICI-192,605 potentiated the relaxant response toSK in the presence of L-NAME. This was confirmed as an EDHF-ediated relaxation by the observation that it was abolished in tis-
ues pre-contracted with 60 mM K+ depolarizing solution (unpub-ished observations). We and others have previously shown thatn addition to NO, EDHF significantly contributes to endothelium-ependent relaxation in the rat pulmonary artery [19,21].
Depolarization of arterial rings in vitro by increasing extracellu-ar potassium prevents the relaxation response to EDHF because theepolarizing solution creates a K+ gradient that does not favor out-ard movement of K+ and hyperpolarization [33]. Accordingly, we
bserved that the EDHF response to GSK in the rat pulmonary arteryas nearly abolished when the arterial rings were pre-contractedith high K+ (60 mM), suggesting the role of K+ channels and hyper-olarization mechanisms mediating the EDHF response. This wasurther substantiated by the observation that apamin plus TRAM-4 markedly attenuated GSK-induced EDHF responses in the ratulmonary artery. The role of IKCa and SKCa channels in medi-ting EDHF-induced relaxation of rat pulmonary artery has beenemonstrated earlier [21,34]. In the mouse mesenteric artery, itas been reported that TRPV4 channels activated by GSK and AChlicited EDHF-mediated vasodilation [4]. These authors showedhat local Ca2+ signals (sparklets) represent Ca2+ influx throughRPV4 channels with subsequent activation of endothelial IKCa andKCa channels leading to hyperpolarization and vasodilation. Fur-her, Bagher and co-workers [5] recently demonstrated that TRPV4hannels were involved in increasing the frequency of EC Ca2+
vents, the consequence of which was the activation of IKCa chan-els and vasodilation of the rat cremaster arterioles. In agreementith these findings in peripheral blood vessels, the observations of
he present investigation suggest that the endothelial TRPV4 chan-els significantly contribute to the EDHF-mediated relaxation ofhe rat pulmonary artery. The mechanisms involved in endothe-ial hyperpolarization and arterial dilation have been explained inerms of electrotonic spread of endothelial hyperpolarizing cur-ent to smooth muscle cells through gap junctions resulting inmooth muscle hyperpolarization and vasodilation [6]. This is inccordance with the role of myoendothelial gap junctions in thepread of endothelial hyperpolarization (EDHF) to vascular cells ofat pulmonary artery as reported earlier [35].
In the rat pulmonary artery with intact endothelium, GSKaused a moderate decrease (∼5%) in the basal tone. However, inndothelium-denuded arterial rings, GSK produced an increase inhe basal tone, which was about ∼11% of K+ contraction. This isonsistent with the involvement of endothelial TRPV4 channelsroducing inhibitory basal tone on this vessel. Regarding GSK-
nduced increase in basal tone in endothelium-denuded rings, itay involve Ca2+ influx through vascular cell TRPV4 channels
esulting in increased intracellular Ca2+ and smooth muscle con-raction. This hypothesis is supported by an earlier study, which
[
ical Research 78 (2013) 18– 27
demonstrated that TRPV4 channel activation with the agonist4�-PDD increased intracellular Ca2+ in pulmonary artery smoothmuscle cells resulting in modest rise (<10% of K+-contraction)in arterial basal tone [16]. Collectively these results indicatethat TRPV4 signaling in vascular endothelium and smooth mus-cle cells of pulmonary artery have opposing actions; while theformer is responsible for vasodilation, the latter mediates arte-rial constriction. Fig. 7 summarizes the pathways involved in theendothelium-dependent relaxation of the rat pulmonary artery,resulting from the activation of TRPV4 channels.
5. Conclusion
The results of the present study suggest that endothelial TRPV4channels are present in the endothelium of the rat pulmonaryartery and their activation causes endothelium-dependent relax-ation involving NO and EDHF. However, it remains to be seenwhether any endogenous agonist or blood flow in pulmonary cir-culation would activate these channels to regulate the pulmonaryarterial tone in health and diseases.
References
[1] Watanabe H, Vriens J, Janssens A, Wondergem R, Droogmans G, Nilius B.Modulation of TRPV4 gating by intra- and extracellular Ca2+. Cell Calcium2003;33:489–95.
[2] Earley S. Endothelium-dependent cerebral artery dilation mediated by tran-sient receptor potential and Ca2+-activated K+channels. J Cardiovasc Pharmacol2011;57:148–53.
[3] Garland CJ, Hiley CR, Dora KA. EDHF: spreading the influence of the endothe-lium. Br J Pharmacol 2011;164:839–52.
[4] Sonkusare SK, Bonev AD, Ledoux J, Liedtke W, Kotlikoff MI, Heppner TJ, et al.Elementary Ca2+ signals through endothelial TRPV4 channels regulate vascularfunction. Science 2012;336:597–601.
[5] Bagher P, Beleznai T, Kansui Y, Mitchell R, Garland CJ, Dora KA. Low intravas-cular pressure activates endothelial cell TRPV4 channels, local Ca2+ events, andIKCa channels, reducing arteriolar tone. Proc Natl Acad Sci 2012;109:18174–9.
[6] Edwards G, Félétou M, Weston AH. Endothelium-derived hyperpolarising fac-tors and associated pathways: a synopsis. Pflugers Archiv 2010;459:863–79.
[7] Nilius B, Droogmans G. Ion channels and their functional role in vascularendothelium. Physiol Rev 2001;81:1415–59.
[8] Mendoza SA, Fang J, Gutterman DD, Wilcox DA, Bubolz AH, Li R, et al. TRPV4-mediated endothelial Ca2+ influx and vasodilation in response to shear stress.Am J Physiol: Heart Circ Physiol 2010;298:H466–76.
[9] Filosa JA, Yao X, Rath G. TRPV4 and the regulation of vascular tone. J CardiovascPharmacol 2013;61:113–9.
10] Marrelli SP, O’Neil RG, Brown RC, Bryan Jr RM. PLA2 and TRPV4 channels reg-ulate endothelial calcium in cerebral arteries. Am J Physiol: Heart Circ Physiol2007;292:H1390–7.
11] Martin E, Dahan D, Cardouat G, Gillibert-Duplantier J, Marthan R, Savineau JP,et al. Involvement of TRPV1 and TRPV4 channels in migration of rat pulmonaryarterial smooth muscle cells. Pflugers Archiv 2012;464:261–72.
12] Yang XR, Lin AH, Hughes JM, Flavahan NA, Cao YN, Liedtke W, et al. Up regulationof osmomechano sensitive TRPV4 channel facilitates chronic hypoxia-inducedmyogenic tone and pulmonary hypertension. Am J Physiol: Lung Cell MolecularPhysiol 2012;302:L555–68.
13] Watanabe H, Murakami M, Ohba T, Takahashi Y, Ito H. TRP channel and cardio-vascular disease. Pharmacol Ther 2008;118:337–51.
14] Earley S, Heppner TJ, Nelson MT, Brayden JE. TRPV4 forms a novel Ca2+
signaling complex with ryanodine receptors and BKCa channels. Circ Res2005;97:1270–9.
15] Cooper CJ, Landzberg MJ, Anderson TJ, Charbonneau F, Creager MA, Ganz P, et al.Role of nitric oxide in the local regulation of pulmonary vascular resistance inhumans. Circulation 1996;93:266–71.
16] Dahan D, Ducret T, Quignard JF, Marthan R, Savineau JP, Estève E. Implicationof the ryanodine receptor in TRPV4-induced calcium response in pulmonaryarterial smooth muscle cells from normoxic and chronically hypoxic rats. Am JPhysiol: Lung Cell Molecular Physiol 2012;303:L824–33.
17] Alvarez DF, King JA, Weber D, Addison E, Liedtke W, Townsley MI. Transientreceptor potential vanilloid 4-mediated disruption of the alveolar septal bar-rier: a novel mechanism of acute lung injury. Circ Res 2006;99:988–95.
18] Fantozzi I, Zhang S, Platoshyn O, Remillard CV, Cowling RT, Yuan JX. Hypoxiaincreases AP-1 binding activity by enhancing capacitative Ca2+ entry in human
pulmonary artery endothelial cells. Am J Physiol: Lung Cell Molecular Physiol2003;285:L1233–45.19] Chen G, Suzuki H, Weston AH. Acetylcholine releases endothelium-derivedhyperpolarizing factor and EDRF from rat blood vessels. Br J Pharmacol1988;95:1165–74.

acolog
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S.V. Sukumaran et al. / Pharm
20] Karamsetty MR, Nakashima JM, Ou L, Klinger JR, Hill NS. EDHF contributes tostrain-related differences in pulmonary arterial relaxation in rats. Am J Physiol:Lung Cell Molecular Physiol 2001;280:L458–64.
21] Subramani J, Leo MDM, Kathirvel K, Arunadevi R, Singh TU, Prakash VR, et al.Essential role of nitric oxide in sepsis-induced impairment of endothelium-derived hyperpolarizing factor-mediated relaxation in rat pulmonary artery.Eur J Pharmacol 2010;630:84–91.
22] Jin M, Wu Z, Chen L, Jaimes J, Collins D, Walters ET, et al. Determinants of TRPV4activity following selective activation by small molecule agonist GSK1016790A.PLoS ONE 2011;6:e16713.
23] Everaerts W, Zhend X, Ghosh D, Vriens J, Gevaert T, Gilbert JP, et al. Inhibitionof the cation channel TRPV4 improves bladder function in mice and rats withcyclophosphamide-induced cystitis. Proc Natl Acad Sci 2010;107:19084–9.
24] Mishra SK, Abbot SE, Choudhury Z, Cheng M, Khatab N, Maycock NJR,et al. Endothelium-dependent relaxation of rat aorta and main pulmonaryartery by the phytoestrogens genistein and daidzein. Cardiovasc Res 2000;46:539–46.
25] Yang XR, Lin MJ, McIntosh LS, Sham JSK. Functional expression of transientreceptor potential melastatin- and vanilloid related channels in pulmonaryarterial and aortic smooth muscle. Am J Physiol: Lung Cell Molecular Physiol2006;290:L1267–76.
26] Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingrealtime quantitative PCR and the 2−��CT Methods. Methods 2001;25:
402–8.27] Zheng X, Zinkevich NS, Gebremedhin D, Gauthier KM, Nishijima Y, Fang J, et al.Arachidonic acid-induced dilation in human coronary arterioles: convergenceof signaling mechanisms on endothelial TRPV4-mediated Ca2+ entry. J Am HeartAssoc 2013;2:e000080.
[
ical Research 78 (2013) 18– 27 27
28] Köhler R, Heyken WT, Heinau P, Schubert R, Si H, Kacik M, et al. Evidence fora functional role of endothelial transient receptor potential V4 in shear stress-induced vasodilatation. Arterioscler Thromb Vasc Biol 2006;26:1495–502.
29] Willette RN, Bao W, Nerurkar S, Yue TL, Doe CP, Stankus G, et al. Systemic acti-vation of the transient receptor potential vanilloid subtype 4 channel causesendothelial failure and circulatory collapse: Part 2. J Pharmacol Exp Ther2008;326:443–52.
30] Zhang DX, Mendoza SA, Bubolz AH, Mizuno A, Ge ZD, Li R, et al. Tran-sient receptor potential vanilloid type 4-deficient mice exhibit impairedendothelium-dependent relaxation induced by acetylcholine in vitro and invivo. Hypertension 2009;53:532–8.
31] Baranowska-Kuczko M, MacLean MR, Kozłowska H, Malinowska B.Endothelium-dependent mechanisms of the vasodilatory effect of theendocannabinoid, anandamide, in the rat pulmonary artery. Pharmacol Res2012;66:251–9.
32] McNeish AJ, Garland CJ. Thromboxane A2 inhibition of SKCa after NO synthaseblock in rat middle cerebral artery. Br J Pharmacol 2007;151:441–9.
33] Gerber RT, Anwar MA, Poston L. Enhanced acetylcholine induced relax-ation in small mesenteric arteries from pregnant rats: an important rolefor endothelium-derived hyperpolarizing factor (EDHF). Br J Pharmacol1998;125:455–60.
34] Kroigaard C, Dalsgaard T, Simonsen U. Mechanisms underlying epithelium-dependent relaxation in rat bronchioles: analogy to EDHF-type relaxation
in rat pulmonary arteries. Am J Physiol: Lung Cell Molecular Physiol2010;298:L531–42.35] Morio Y, Carter EP, Oka M, McMurtry IF. EDHF-mediated vasodilation involvesdifferent mechanisms in normotensive and hypertensive rat lungs. Am JPhysiol: Heart Circ Physiol 2003;284:H1762–70.
Copyright © 2022 FDOKUMEN




















