
Assinados os tratados argentino-brasilei ros - Coleção Digital ...
Upload
independentCategory
view
0download
0
Free Radical Biology & Medicine 45 (2008) 1403–1412
Contents lists available at ScienceDirect
Free Radical Biology & Medicine
j ourna l homepage: www.e lsev ie r.com/ locate / f reeradb iomed
Original Contribution
Rapid reactive oxygen species (ROS) generation induced by curcumin leads tocaspase-dependent and -independent apoptosis in L929 cells
Faisal Thayyullathil, Shahanas Chathoth, Abdulkader Hago, Mahendra Patel, Sehamuddin Galadari ⁎Cell Signaling Laboratory, Department of Biochemistry, Faculty of Medicine and Health Sciences, UAE University, P.O. Box 17666, Al Ain, United Arab Emirates
Abbreviations: Cur, curcumin; Cyt c, cytochrome c;MTT, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliuNAC, N-acetylcysteine; PBS, phosphate-buffered salinDCFH-DA, dichlorofluorescein diacetate; z-VAD-fmk, NAsp fluoromethylketone; AIF, apoptosis inducing fapolymerase; PAGE, polyacrylamide gel electrophoresanion channel; TBP, TATA binding protein; Cat, catalase;⁎ Corresponding author. Fax: +97137672033.
E-mail address: [email protected] (S. Galadar
0891-5849/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.freeradbiomed.2008.08.014
a b s t r a c t
a r t i c l e i n f oArticle history:
Evidence that curcumin ma Received 7 March 2008Revised 20 July 2008Accepted 5 August 2008Available online 16 August 2008Keywords:Reactive oxygen species (ROS)CurcuminCaspaseApoptosis inducing factor (AIF)p53Signaling
y have anticancer activities has renewed interest in its potential to prevent andtreat disease. In this study, we show that curcumin-mediated rapid generation of reactive oxygen species(ROS) leads to apoptosis by modulating different apoptotic pathways in mouse fibroblast L929 cells. We showfor the first time that curcumin-induced rapid ROS generation causes the release of apoptosis inducing factor(AIF) from the mitochondria to the cytosol and nucleus, hence, leading to caspase 3-independent apoptosis.However, our studies also show that curcumin induces the release of cytochrome c from mitochondria,causing activation of caspase 3, and concomitant PARP cleavage, which is the hallmark of caspase-dependentapoptosis. Furthermore, curcumin-induced ROS generation leads to the induction of the proapoptotic proteinp53 and its effector protein p21 and down-regulation of cell cycle regulatory proteins such as Rb and cyclinD1 and D3. Both glutathione (GSH) and N-acetylcysteine (NAC) pretreatment resulted in the completeinhibition of curcumin-induced ROS generation, AIF release from mitochondria, and caspase activation.Additionally, pretreatment of L929 cells with these antioxidants completely blocked the induction of p53-dependent p21 accumulation. In conclusion, our data show that in addition to caspase 3 activation,curcumin-induced rapid ROS generation leads to AIF release, and the activation of the caspase-independentapoptotic pathway.
© 2008 Elsevier Inc. All rights reserved.
Introduction
Curcumin, also known as diferuloymethane, is the major yellowpigment extracted from turmeric (Curcuma longa) and it is commonlyused as a flavoring agent in food. Curcumin has been reported topossess anti-inflammatory and antioxidant activities [1], and it hasbeen reported as a potent inhibitor of mutagenesis and carcinogenesis[2,3]. Several biological effects of curcumin have been reported. Theseinclude cell shrinkage, chromatin condensation, and oxidative DNAdamage [4]. Other reported signaling effects of curcumin include:inhibition of nitric oxide synthase, receptor tyrosine kinase, proteinkinase C, Akt, NFκB, and cyclin D1 and activation of p53 [5–9].Importantly, curcumin has been shown to induce apoptosis indifferent malignant cancer cell lines [10].
It is now well established that apoptosis or programmed celldeath is characterized by cell shrinkage, cytoplasmic, nuclear, and
ROS, reactive oxygen species;m bromide; GSH, glutathione;e; DMSO, dimethyl sulfoxide-benzoyloxycarbonyl-Val-Ala-ctor; PARP, poly(ADP-ribose)is; VDAC, voltage-dependentSOD, superoxide dismutase.
i).
l rights reserved.
chromatin condensation, membrane blebbing, protein fragmentationand DNA degradation, and finally breakdown of the cell into smallerunits or apoptotic bodies [11]. Many proteins are involved in thiscomplex process. Caspases, a family of cysteine-dependent aspartate-directed proteases, play a critical role in the initiation and executionof apoptosis [12]. Among this family of caspases, caspase 3, inparticular, is believed to be one of the most commonly involvedcaspases in the execution of apoptosis in various cell types [13]. Morerecently, caspase-independent apoptogenic proteins, such as apop-tosis inducing factor (AIF), endonuclease G (EndoG), and secondmitochondria-derived activator of caspase (smac) have been identi-fied and characterized [14–16].
Apoptosis inducing factor is a mitochondrial proteinwhose transloca-tion in the cytosol, as well as in the nucleus, mediates caspase-independent apoptosis in a number of model systems [17]. The humanfull-length AIF (613 amino acids) has no effect on apoptosis. The aminoterminal 102 amino acids of AIF, namely the mitochondrial localizationsequence, once removed, yield amature AIFmolecule. On the initiation ofapoptosis, the mature AIF releases into the cytosol and the nucleus,inducing chromatin condensation and large DNA fragmentation (50 kbp).During apoptosis, EndoG, like AIF, translocates to the nucleus where itcauses oligonucleosomal DNA fragmentation [16]. Subsequent studieshave demonstrated that EndoG catalyzes both high molecular DNAcleavage and oligonucleosomal DNA breakdown. Moreover, EndoGcooperateswith exonuclease andDnase1 to facilitateDNAprocessing [18].

1404 F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
Recently, it has been reported that curcumin-induced apoptosis isdue to the production of reactive oxygen species (ROS) [19]. These freeradicals are the by-products of normal cellular oxidative processes,and they are generated in the mitochondria and from other sources.They inflict serious damage to lipids, proteins, and DNA [20], and theyhave been suggested as regulating the process involved in theinitiation of apoptotic signaling [21]. Indeed, ROS have been demon-strated to perform certain functions in the early stages of apoptosis,and to induce the depolarization of the mitochondrial membrane,which eventually results in an increase in the level of otherproapoptotic molecules in the cytosol [22]. Although, ROS mayconstitute a direct cause of mitochondrial dysfunction, it remainscontroversial as to what is its role during apoptosis. Additionally, therole of ROS in Fas-mediated apoptosis has been demonstrated in avariety of cell types [23]. Taken together, the molecular eventsassociated with the role of ROS in the cytotoxic mechanisms have yetto be clarified. The tumor suppressor p53 is known to be an importantregulator of apoptotic cell death. It has been shown in several cell linesthat the growth arrest mediated by p53 is dependent on thetranscriptional activation of p21 [24], mitochondrial cytochrome crelease, and caspase activation [25].
To date, the mechanism of ROS-triggered cell death induced bycurcumin is poorly understood. In order to shed more light on themechanism of ROS-mediated apoptosis induced by curcumin, weinvestigated the effects of this compound on mouse fibroblast L929cells. Our study shows that in L929 cells curcumin induces a very rapidand significant ROS generation, which in turn leads to multipleapoptotic signal including both caspase-dependent and -independentpathways. Additionally, we have found that curcumin activates p53-dependent p21 accumulation and down-regulation of cell cycleregulatory proteins. Moreover, our study shows that antioxidantssuch as GSH and NAC inhibit curcumin-induced cell death, byinhibiting ROS generation.
Materials and methods
Reagents
Curcumin, glutathione, N-acetylcysteine, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT), proteinase K, RNase,glutathione assay chemicals, bovine liver catalase, super oxidedismutase, anti-rabbit IgG, and anti-mouse IgG were purchasedfrom Sigma Chemical Co. (St. Louis, MO). Oxidation-sensitive DCFH-DA (D-399) and -insensitive 5-(and -6)-carboxy-2′,7′-dichlorofluores-cein diacetate (C-369) were purchased fromMolecular Probes (Eugene,OR). RPMI 1640 GlutaMAX medium, fetal bovine serum (FBS), andtrypsin-EDTA were bought from GIBCO BRL (Grand Island, NY). Anti-p53, anti-p21, anti-BAX, anti-AIF, and anti-actin antibodies were fromSanta Cruz Biotechnology Inc. (Santa Cruz, CA). Anti-cytochrome cand anti-Rb antibodies were purchased from BD Pharmingen (SanDiego CA). Anti-PARP, anti-cyclin D1, anti-cyclin D3, and anti-pRbSer795 antibodies were purchased from Cell Signaling Technologies(Beverly, MA). Anti-VDAC (voltage-dependent anion channel 1) andanti-TBP (TATA binding protein) were purchased from Abcam (Cam-bridge, UK). Colorimetric CaspASE assay system and inhibitor ofcaspase 3 (z-VAD-fmk) were purchased from Promega (Madison, WI).
Cell culture and curcumin treatment
L929 mouse fibroblast cells (ATCC) were grown in RPMI 1640containing GlutaMAX medium supplemented with 10% (V/V) heat-inactivated fetal bovine serum without antibiotics in an incubatorcontaining a humidified atmosphere of 95% air and 5%CO2 at 37°C.Curcumin was dissolved in dimethyl sulfoxide (DMSO) at a concen-tration of 20 mM and was stored in a dark colored bottle at -20°C. Thestock was diluted to the required concentration with DMSO when
needed. Prior to curcumin treatment cells were grown to about 80%confluence, and then exposed to curcumin at different concentrations(0–100 μM) and for a different period of time (0–24 h). Cells grown in amedium containing an equivalent amount of DMSOwithout curcuminserved as control.
Cell viability and cytotoxicity assay
Cell viability following treatment with different concentrations ofcurcumin was determined by trypan blue dye exclusion test. Aftertreatment with curcumin, floating L929 cells in the medium of eachplatewere transferred to centrifuge tubes. Adherent cellswere detachedwith trypsin and added to the corresponding floating cells beforecentrifugation. The pellet were resuspended and the cells were stainedwith 0.4% trypan blue, and the numbers of trypan blue positive andnegative cells were counted on a hemocytometer by light microscopy.
Cytotoxicity assays were carried out as described previously withslightmodifications [10]. Cells were grown in 96-well microtiter plates(10,000 cells/well) and they were incubated for 24 h with or withoutdifferent concentrations of curcumin (0–100 μM). At the required timepoint 100 μl media were removed and 25 μl of MTT (5 mg/ml) wasadded to eachwell. The plates were incubated for a further 4 h at 37°C.After incubation the plateswere centrifuged at 1500 rpm for 5min andthemediawere removed from all thewells. The formazan crystals thatform were then solubilized in a 200 μl of DMSO. The colored solutionwas quantified at 570 nm by using Anthos Labtec HT2 96-well platereader. The cytotoxicity was expressed as percentage over control.
DNA fragmentation analysis
Apoptotic DNA fragments were isolated from the apoptotic cells asfollows: L929 cells were grown at a density of 2× 106 cells per plateand incubated with various concentrations of curcumin. Cells werethen washed with PBS and incubated with 200 μl of lysis buffer(50 mM Tris-HCl (pH 7.5), 3% nonionic detergent IGPAL CA-630[(octylphenoxy) polyethoxyethanol], and 20 mM EDTA) for 10 min.The sampleswere centrifuged at 1000 g for 5min in order to collect thesupernatant which contain apoptotic DNA fragment. SDS (50 μl, 5%)was added and the supernatants were incubated with 0.4 μg/mlRNase at 56°C for 2 h to remove the cellular RNA. Proteinase K(1.5 μg/ml) was then added to the supernatant at 56°C and it wasfurther incubated for 2 h to remove the proteins. The DNA was thenprecipitated with 0.1 vol of 3 M sodium acetate and 2.5 vol of ice-coldabsolute ethanol. After centrifugation, the DNA pellet was washedwith 70% ethanol and then air-dried. The dried pellet wereresuspended in 20 μl TE buffer (10 mM Tris-HCl, pH 7.5, and0.1 mM EDTA) and incubated at 65°C for 5 min. Finally theresuspended DNA was subjected to electrophoresis on a 2% agarosegel at a constant voltage of 40 V for 3–4 h.
Measurement of PARP cleavage
To determine the activation of caspase 3 activity we examined thecleavage of poly(ADP-ribose) polymerase (PARP). Thirty micrograms ofwhole cell extract was resolved on 8% polyacrylamide gel, transferred tonitrocellulose membrane, blocked with 5% nonfat milk protein, probedwith PARP antibodies (1:2000), and detected by using enhancedchemiluminescence reagent (Pierce Biotechnology (Rockford, IL).
Enzymatic caspase 3 assay
The enzymatic assay of caspase induced by curcuminwas measuredby using the manufacture's protocol (Promega (Madison, WI)). Briefly,cells were lysed in a lysis buffer by freeze and thawing. The lysed cellswere centrifuged at 14,000 rpm for 15 min. Fifty micrograms of proteinwas incubated with 30 μl of caspase assay buffer and 2 μl of caspase 3
1405F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
(DEVD-pNA) colorimetric substrate at 37°C for 4 h. The optical densityof the reaction mixture was quantitated spectrophotometrically at awavelength of 405 nm by using an Athos Labtec HT2 96-well platereader.
Measurement of intracellular ROS levels
Generation of ROS was measured by the oxidative-sensitivefluorescent probe DCFH-DA. DCFH-DA is a nonpolar compound thatreadily diffuses into cells, where it is cleaved by intracellular esterasesto form DCFH and thereby is trapped inside the cells. DCFH is oxidizedto the highly fluorescent 2,7-dichlorofluorescein (DCF) following ROSgeneration. After treatment of cells with curcumin, attached cells werewashed twicewithHank's buffer and then loaded for 60minwith 50 μgof DCFH-DA in 2ml of culturemedia. The cellswere againwashed three
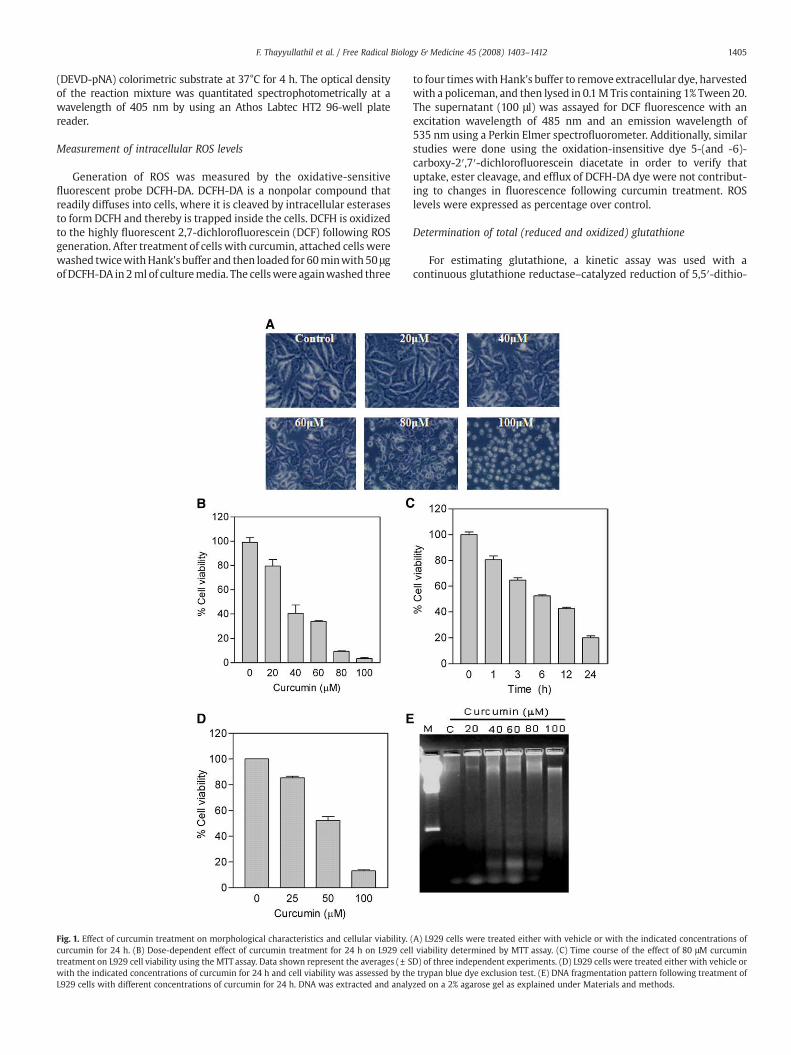
Fig. 1. Effect of curcumin treatment on morphological characteristics and cellular viability.curcumin for 24 h. (B) Dose-dependent effect of curcumin treatment for 24 h on L929 celtreatment on L929 cell viability using the MTT assay. Data shown represent the averages (± Swith the indicated concentrations of curcumin for 24 h and cell viability was assessed by thL929 cells with different concentrations of curcumin for 24 h. DNA was extracted and analy
to four timeswith Hank's buffer to remove extracellular dye, harvestedwith a policeman, and then lysed in 0.1M Tris containing 1% Tween 20.The supernatant (100 μl) was assayed for DCF fluorescence with anexcitation wavelength of 485 nm and an emission wavelength of535 nm using a Perkin Elmer spectrofluorometer. Additionally, similarstudies were done using the oxidation-insensitive dye 5-(and -6)-carboxy-2′,7′-dichlorofluorescein diacetate in order to verify thatuptake, ester cleavage, and efflux of DCFH-DA dye were not contribut-ing to changes in fluorescence following curcumin treatment. ROSlevels were expressed as percentage over control.
Determination of total (reduced and oxidized) glutathione
For estimating glutathione, a kinetic assay was used with acontinuous glutathione reductase–catalyzed reduction of 5,5′-dithio-
(A) L929 cells were treated either with vehicle or with the indicated concentrations ofl viability determined by MTT assay. (C) Time course of the effect of 80 μM curcuminD) of three independent experiments. (D) L929 cells were treated either with vehicle ore trypan blue dye exclusion test. (E) DNA fragmentation pattern following treatment ofzed on a 2% agarose gel as explained under Materials and methods.
1406 F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
bis(2-nitrobenzoic acid) (DTNB) to the chromophore, whose rate wasmonitored spectrophotometrically at 412 nm. Cells were lysed in400 μl of 5% sulfosalicylic acid by freeze-thawing. After centrifugation(10,000 g for 5 min), the supernatant was transferred to anothertube. Then, 10 μl of aliquot was mixed with 150 μl potassiumphosphate buffer, pH 7.0, containing 5 mM EDTA, 1.5 mg/ml DTNB,and 6 U/ml glutathione reductase. Finally, 50 μl of (0.16 mg/ml)NADPH in potassium phosphate buffer was also added. Theabsorbance was read at 412 nm and quantified using Bio-Radmicroplate reader.
Western blotting
Cells were washed twice with phosphate buffered saline (PBS)and lysed in a RIPA lysis buffer [50 mM Tris HCl (pH 7.4), 1% NP-40,40 mM NaF, 10 mM NaCl, 10 mM Na3VO4, 1 mM phenylmethylsufonylfluoride (PMSF), 10 mM dithiothreitol (DTT), and EDTA-free Roshprotease inhibitor tablets per 20 ml buffer]. The cell lysates werecentrifuged at 14,000 rpm for 15 min. Total protein, determined bythe Bio-Rad protein assay, was mixed with 6X loading buffer andboiled at 100°C for 3 min. Samples at 40 μg/lane were resolved bySDS-PAGE and the separated proteins were transferred ontonitrocellulose membranes by the wet transfer method using a Bio-Rad electrotransfer apparatus. Following transfer the blots wereblocked with 5% nonfat milk in Tris buffer saline containing 0.1%Tween 20 and then incubated with primary antibodies followed bysecondary antibodies. Proteins were visualized by using theenhanced chemiluminescence system. Western blot data presentedare representative of those obtained in at least three separateexperiments.
Measurement of cytochrome c release
Release of cytochrome c from the mitochondria to cytosol wasmeasured by Western blot analysis as previously described [26].Briefly cells were washed once with ice-cold PBS and gently lysed for30 s in 100 μl ice cold lysis buffer (250 mM sucrose, 1 mM EDTA, 0.05%digitonin, 25 mM Tris, pH 6.8, 1 mM dithiothreitol, 1 μg/ml leupeptin,1 μg/ml pepstatin, 1 μg/ml aprotinin, 1 mM benzamidine, and 0.1 mMphenylmethylsulfonyl fluoride). The lysates were centrifuged at12,000 g at 4°C for 5 min in order to obtain the supernatant (cytosolicfraction) and the pellet (fraction that contains the mitochondria). Fiftymicrograms of each supernatant or the pellet proteins was subjectedto electrophoresis on a 15% SDS-polyacrylamide gel and analyzed byWestern blot using anti-cytochrome c antibody.
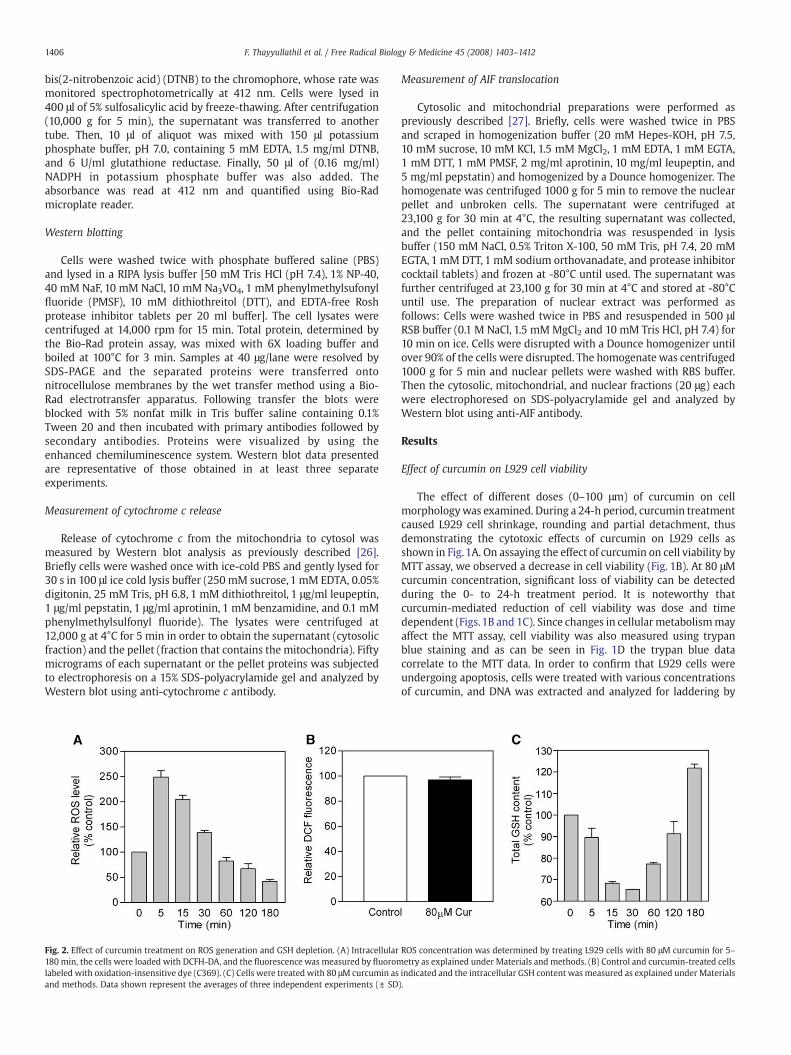
Fig. 2. Effect of curcumin treatment on ROS generation and GSH depletion. (A) Intracellular180 min, the cells were loaded with DCFH-DA, and the fluorescence was measured by fluoromlabeled with oxidation-insensitive dye (C369). (C) Cells were treated with 80 μM curcumin asand methods. Data shown represent the averages of three independent experiments (± SD
Measurement of AIF translocation
Cytosolic and mitochondrial preparations were performed aspreviously described [27]. Briefly, cells were washed twice in PBSand scraped in homogenization buffer (20 mM Hepes-KOH, pH 7.5,10 mM sucrose, 10 mM KCl, 1.5 mM MgCl2, 1 mM EDTA, 1 mM EGTA,1 mM DTT, 1 mM PMSF, 2 mg/ml aprotinin, 10 mg/ml leupeptin, and5 mg/ml pepstatin) and homogenized by a Dounce homogenizer. Thehomogenate was centrifuged 1000 g for 5 min to remove the nuclearpellet and unbroken cells. The supernatant were centrifuged at23,100 g for 30 min at 4°C, the resulting supernatant was collected,and the pellet containing mitochondria was resuspended in lysisbuffer (150 mM NaCl, 0.5% Triton X-100, 50 mM Tris, pH 7.4, 20 mMEGTA, 1 mM DTT, 1 mM sodium orthovanadate, and protease inhibitorcocktail tablets) and frozen at -80°C until used. The supernatant wasfurther centrifuged at 23,100 g for 30 min at 4°C and stored at -80°Cuntil use. The preparation of nuclear extract was performed asfollows: Cells were washed twice in PBS and resuspended in 500 μlRSB buffer (0.1 M NaCl, 1.5 mM MgCl2 and 10 mM Tris HCl, pH 7.4) for10 min on ice. Cells were disrupted with a Dounce homogenizer untilover 90% of the cells were disrupted. The homogenate was centrifuged1000 g for 5 min and nuclear pellets were washed with RBS buffer.Then the cytosolic, mitochondrial, and nuclear fractions (20 μg) eachwere electrophoresed on SDS-polyacrylamide gel and analyzed byWestern blot using anti-AIF antibody.
Results
Effect of curcumin on L929 cell viability
The effect of different doses (0–100 μm) of curcumin on cellmorphologywas examined. During a 24-h period, curcumin treatmentcaused L929 cell shrinkage, rounding and partial detachment, thusdemonstrating the cytotoxic effects of curcumin on L929 cells asshown in Fig.1A. On assaying the effect of curcumin on cell viability byMTT assay, we observed a decrease in cell viability (Fig. 1B). At 80 μMcurcumin concentration, significant loss of viability can be detectedduring the 0- to 24-h treatment period. It is noteworthy thatcurcumin-mediated reduction of cell viability was dose and timedependent (Figs.1B and 1C). Since changes in cellularmetabolismmayaffect the MTT assay, cell viability was also measured using trypanblue staining and as can be seen in Fig. 1D the trypan blue datacorrelate to the MTT data. In order to confirm that L929 cells wereundergoing apoptosis, cells were treated with various concentrationsof curcumin, and DNA was extracted and analyzed for laddering by
ROS concentration was determined by treating L929 cells with 80 μM curcumin for 5–etry as explained under Materials and methods. (B) Control and curcumin-treated cellsindicated and the intracellular GSH content was measured as explained under Materials).
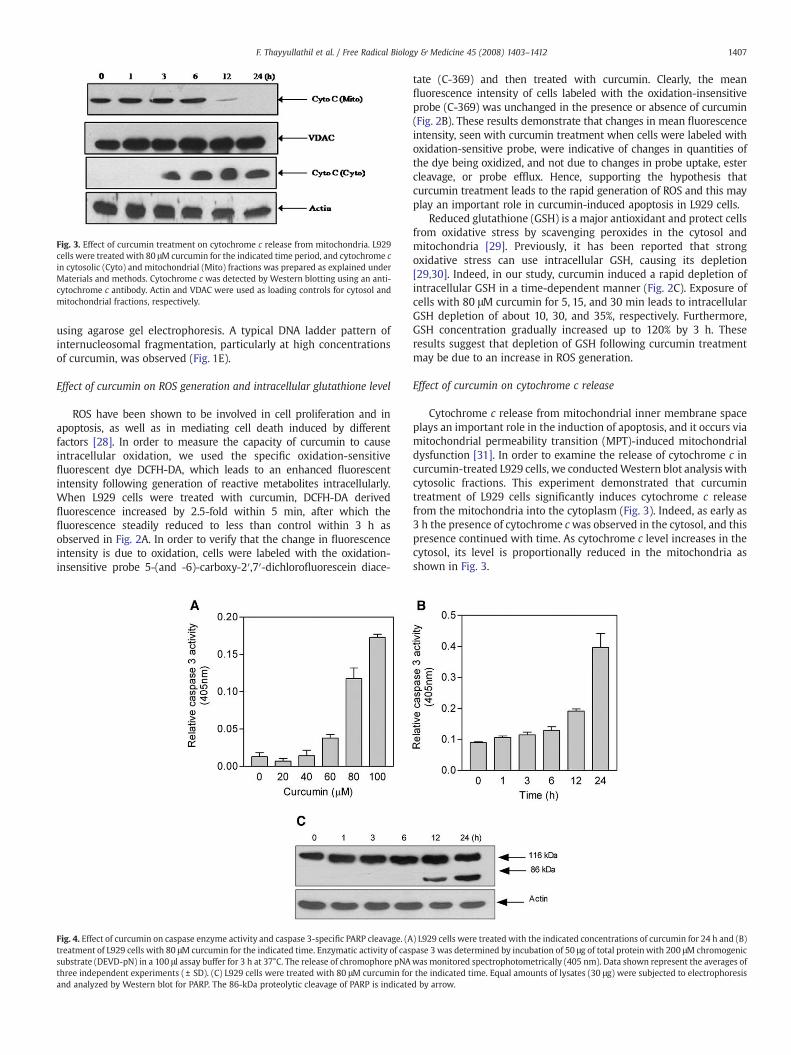
Fig. 3. Effect of curcumin treatment on cytochrome c release from mitochondria. L929cells were treated with 80 μM curcumin for the indicated time period, and cytochrome cin cytosolic (Cyto) and mitochondrial (Mito) fractions was prepared as explained underMaterials and methods. Cytochrome c was detected by Western blotting using an anti-cytochrome c antibody. Actin and VDAC were used as loading controls for cytosol andmitochondrial fractions, respectively.
1407F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
using agarose gel electrophoresis. A typical DNA ladder pattern ofinternucleosomal fragmentation, particularly at high concentrationsof curcumin, was observed (Fig. 1E).
Effect of curcumin on ROS generation and intracellular glutathione level
ROS have been shown to be involved in cell proliferation and inapoptosis, as well as in mediating cell death induced by differentfactors [28]. In order to measure the capacity of curcumin to causeintracellular oxidation, we used the specific oxidation-sensitivefluorescent dye DCFH-DA, which leads to an enhanced fluorescentintensity following generation of reactive metabolites intracellularly.When L929 cells were treated with curcumin, DCFH-DA derivedfluorescence increased by 2.5-fold within 5 min, after which thefluorescence steadily reduced to less than control within 3 h asobserved in Fig. 2A. In order to verify that the change in fluorescenceintensity is due to oxidation, cells were labeled with the oxidation-insensitive probe 5-(and -6)-carboxy-2′,7′-dichlorofluorescein diace-
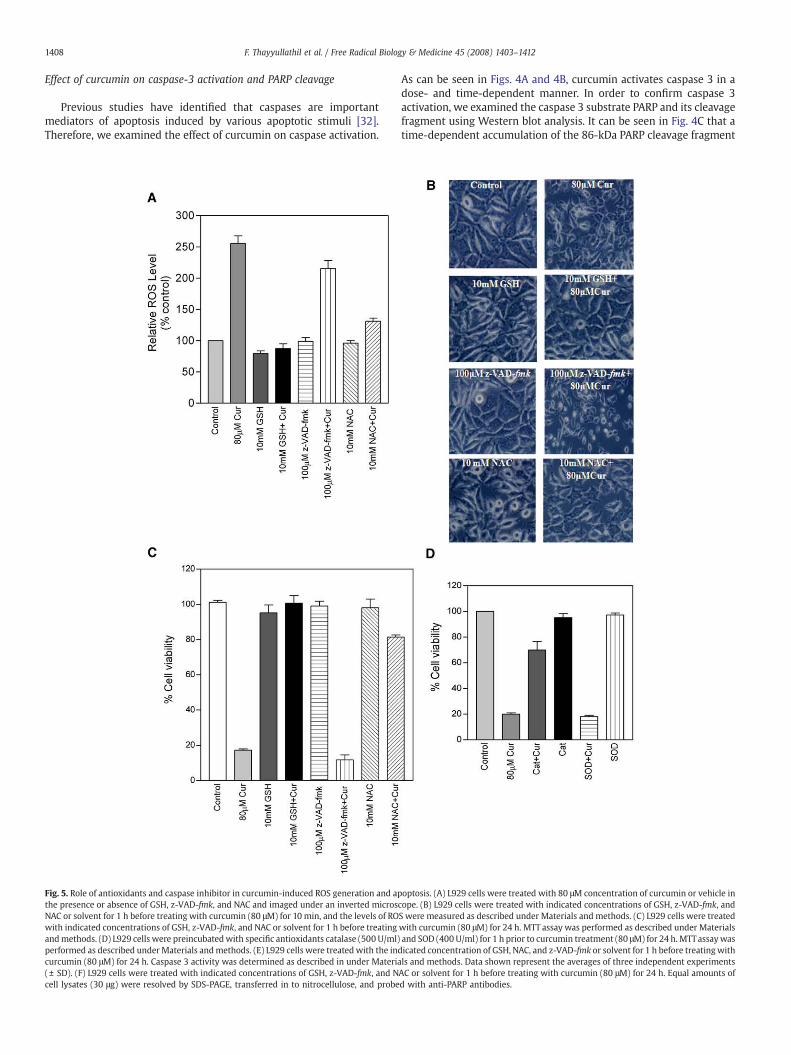
Fig. 4. Effect of curcumin on caspase enzyme activity and caspase 3-specific PARP cleavage. (Atreatment of L929 cells with 80 μM curcumin for the indicated time. Enzymatic activity of cassubstrate (DEVD-pN) in a 100 μl assay buffer for 3 h at 37°C. The release of chromophore pNAthree independent experiments (± SD). (C) L929 cells were treated with 80 μM curcumin forand analyzed by Western blot for PARP. The 86-kDa proteolytic cleavage of PARP is indicate
tate (C-369) and then treated with curcumin. Clearly, the meanfluorescence intensity of cells labeled with the oxidation-insensitiveprobe (C-369) was unchanged in the presence or absence of curcumin(Fig. 2B). These results demonstrate that changes in mean fluorescenceintensity, seen with curcumin treatment when cells were labeled withoxidation-sensitive probe, were indicative of changes in quantities ofthe dye being oxidized, and not due to changes in probe uptake, estercleavage, or probe efflux. Hence, supporting the hypothesis thatcurcumin treatment leads to the rapid generation of ROS and this mayplay an important role in curcumin-induced apoptosis in L929 cells.
Reduced glutathione (GSH) is a major antioxidant and protect cellsfrom oxidative stress by scavenging peroxides in the cytosol andmitochondria [29]. Previously, it has been reported that strongoxidative stress can use intracellular GSH, causing its depletion[29,30]. Indeed, in our study, curcumin induced a rapid depletion ofintracellular GSH in a time-dependent manner (Fig. 2C). Exposure ofcells with 80 μM curcumin for 5, 15, and 30 min leads to intracellularGSH depletion of about 10, 30, and 35%, respectively. Furthermore,GSH concentration gradually increased up to 120% by 3 h. Theseresults suggest that depletion of GSH following curcumin treatmentmay be due to an increase in ROS generation.
Effect of curcumin on cytochrome c release
Cytochrome c release from mitochondrial inner membrane spaceplays an important role in the induction of apoptosis, and it occurs viamitochondrial permeability transition (MPT)-induced mitochondrialdysfunction [31]. In order to examine the release of cytochrome c incurcumin-treated L929 cells, we conductedWestern blot analysis withcytosolic fractions. This experiment demonstrated that curcumintreatment of L929 cells significantly induces cytochrome c releasefrom the mitochondria into the cytoplasm (Fig. 3). Indeed, as early as3 h the presence of cytochrome cwas observed in the cytosol, and thispresence continued with time. As cytochrome c level increases in thecytosol, its level is proportionally reduced in the mitochondria asshown in Fig. 3.
) L929 cells were treated with the indicated concentrations of curcumin for 24 h and (B)pase 3 was determined by incubation of 50 μg of total proteinwith 200 μM chromogenicwas monitored spectrophotometrically (405 nm). Data shown represent the averages ofthe indicated time. Equal amounts of lysates (30 μg) were subjected to electrophoresisd by arrow.
1408 F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
Effect of curcumin on caspase-3 activation and PARP cleavage
Previous studies have identified that caspases are importantmediators of apoptosis induced by various apoptotic stimuli [32].Therefore, we examined the effect of curcumin on caspase activation.
Fig. 5. Role of antioxidants and caspase inhibitor in curcumin-induced ROS generation and apthe presence or absence of GSH, z-VAD-fmk, and NAC and imaged under an inverted microsNAC or solvent for 1 h before treating with curcumin (80 μM) for 10 min, and the levels of ROwith indicated concentrations of GSH, z-VAD-fmk, and NAC or solvent for 1 h before treatingandmethods. (D) L929 cells were preincubatedwith specific antioxidants catalase (500 U/ml)performed as described underMaterials andmethods. (E) L929 cells were treated with the indcurcumin (80 μM) for 24 h. Caspase 3 activity was determined as described in under Materia(± SD). (F) L929 cells were treated with indicated concentrations of GSH, z-VAD-fmk, and Ncell lysates (30 μg) were resolved by SDS-PAGE, transferred in to nitrocellulose, and probe
As can be seen in Figs. 4A and 4B, curcumin activates caspase 3 in adose- and time-dependent manner. In order to confirm caspase 3activation, we examined the caspase 3 substrate PARP and its cleavagefragment using Western blot analysis. It can be seen in Fig. 4C that atime-dependent accumulation of the 86-kDa PARP cleavage fragment
optosis. (A) L929 cells were treated with 80 μM concentration of curcumin or vehicle incope. (B) L929 cells were treated with indicated concentrations of GSH, z-VAD-fmk, andS were measured as described under Materials and methods. (C) L929 cells were treatedwith curcumin (80 μM) for 24 h. MTT assay was performed as described under Materialsand SOD (400 U/ml) for 1 h prior to curcumin treatment (80 μM) for 24 h. MTTassaywasicated concentration of GSH, NAC, and z-VAD-fmk or solvent for 1 h before treating withls and methods. Data shown represent the averages of three independent experimentsAC or solvent for 1 h before treating with curcumin (80 μM) for 24 h. Equal amounts ofd with anti-PARP antibodies.
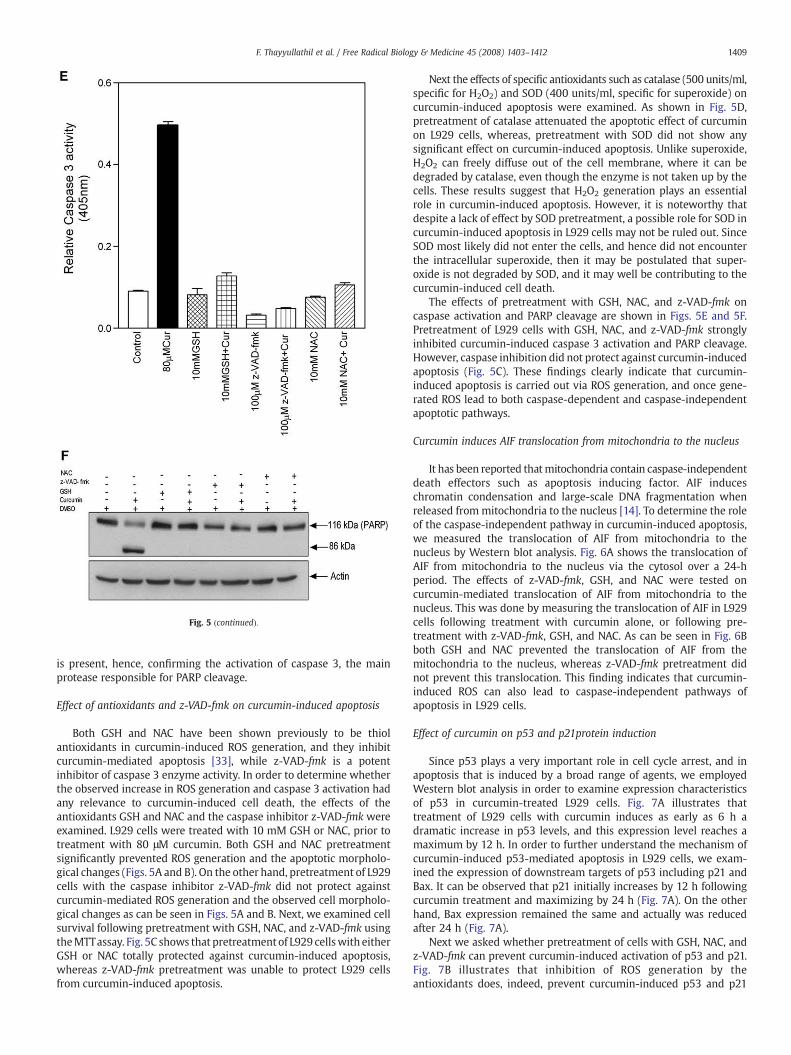
Fig. 5 (continued).
1409F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
is present, hence, confirming the activation of caspase 3, the mainprotease responsible for PARP cleavage.
Effect of antioxidants and z-VAD-fmk on curcumin-induced apoptosis
Both GSH and NAC have been shown previously to be thiolantioxidants in curcumin-induced ROS generation, and they inhibitcurcumin-mediated apoptosis [33], while z-VAD-fmk is a potentinhibitor of caspase 3 enzyme activity. In order to determine whetherthe observed increase in ROS generation and caspase 3 activation hadany relevance to curcumin-induced cell death, the effects of theantioxidants GSH and NAC and the caspase inhibitor z-VAD-fmk wereexamined. L929 cells were treated with 10 mM GSH or NAC, prior totreatment with 80 μM curcumin. Both GSH and NAC pretreatmentsignificantly prevented ROS generation and the apoptotic morpholo-gical changes (Figs. 5A and B). On the other hand, pretreatment of L929cells with the caspase inhibitor z-VAD-fmk did not protect againstcurcumin-mediated ROS generation and the observed cell morpholo-gical changes as can be seen in Figs. 5A and B. Next, we examined cellsurvival following pretreatment with GSH, NAC, and z-VAD-fmk usingtheMTTassay. Fig. 5C shows that pretreatment of L929 cellswith eitherGSH or NAC totally protected against curcumin-induced apoptosis,whereas z-VAD-fmk pretreatment was unable to protect L929 cellsfrom curcumin-induced apoptosis.
Next the effects of specific antioxidants such as catalase (500 units/ml,specific for H2O2) and SOD (400 units/ml, specific for superoxide) oncurcumin-induced apoptosis were examined. As shown in Fig. 5D,pretreatment of catalase attenuated the apoptotic effect of curcuminon L929 cells, whereas, pretreatment with SOD did not show anysignificant effect on curcumin-induced apoptosis. Unlike superoxide,H2O2 can freely diffuse out of the cell membrane, where it can bedegraded by catalase, even though the enzyme is not taken up by thecells. These results suggest that H2O2 generation plays an essentialrole in curcumin-induced apoptosis. However, it is noteworthy thatdespite a lack of effect by SOD pretreatment, a possible role for SOD incurcumin-induced apoptosis in L929 cells may not be ruled out. SinceSOD most likely did not enter the cells, and hence did not encounterthe intracellular superoxide, then it may be postulated that super-oxide is not degraded by SOD, and it may well be contributing to thecurcumin-induced cell death.
The effects of pretreatment with GSH, NAC, and z-VAD-fmk oncaspase activation and PARP cleavage are shown in Figs. 5E and 5F.Pretreatment of L929 cells with GSH, NAC, and z-VAD-fmk stronglyinhibited curcumin-induced caspase 3 activation and PARP cleavage.However, caspase inhibition did not protect against curcumin-inducedapoptosis (Fig. 5C). These findings clearly indicate that curcumin-induced apoptosis is carried out via ROS generation, and once gene-rated ROS lead to both caspase-dependent and caspase-independentapoptotic pathways.
Curcumin induces AIF translocation from mitochondria to the nucleus
It has been reported thatmitochondria contain caspase-independentdeath effectors such as apoptosis inducing factor. AIF induceschromatin condensation and large-scale DNA fragmentation whenreleased from mitochondria to the nucleus [14]. To determine the roleof the caspase-independent pathway in curcumin-induced apoptosis,we measured the translocation of AIF from mitochondria to thenucleus by Western blot analysis. Fig. 6A shows the translocation ofAIF from mitochondria to the nucleus via the cytosol over a 24-hperiod. The effects of z-VAD-fmk, GSH, and NAC were tested oncurcumin-mediated translocation of AIF from mitochondria to thenucleus. This was done by measuring the translocation of AIF in L929cells following treatment with curcumin alone, or following pre-treatment with z-VAD-fmk, GSH, and NAC. As can be seen in Fig. 6Bboth GSH and NAC prevented the translocation of AIF from themitochondria to the nucleus, whereas z-VAD-fmk pretreatment didnot prevent this translocation. This finding indicates that curcumin-induced ROS can also lead to caspase-independent pathways ofapoptosis in L929 cells.
Effect of curcumin on p53 and p21protein induction
Since p53 plays a very important role in cell cycle arrest, and inapoptosis that is induced by a broad range of agents, we employedWestern blot analysis in order to examine expression characteristicsof p53 in curcumin-treated L929 cells. Fig. 7A illustrates thattreatment of L929 cells with curcumin induces as early as 6 h adramatic increase in p53 levels, and this expression level reaches amaximum by 12 h. In order to further understand the mechanism ofcurcumin-induced p53-mediated apoptosis in L929 cells, we exam-ined the expression of downstream targets of p53 including p21 andBax. It can be observed that p21 initially increases by 12 h followingcurcumin treatment and maximizing by 24 h (Fig. 7A). On the otherhand, Bax expression remained the same and actually was reducedafter 24 h (Fig. 7A).
Next we asked whether pretreatment of cells with GSH, NAC, andz-VAD-fmk can prevent curcumin-induced activation of p53 and p21.Fig. 7B illustrates that inhibition of ROS generation by theantioxidants does, indeed, prevent curcumin-induced p53 and p21
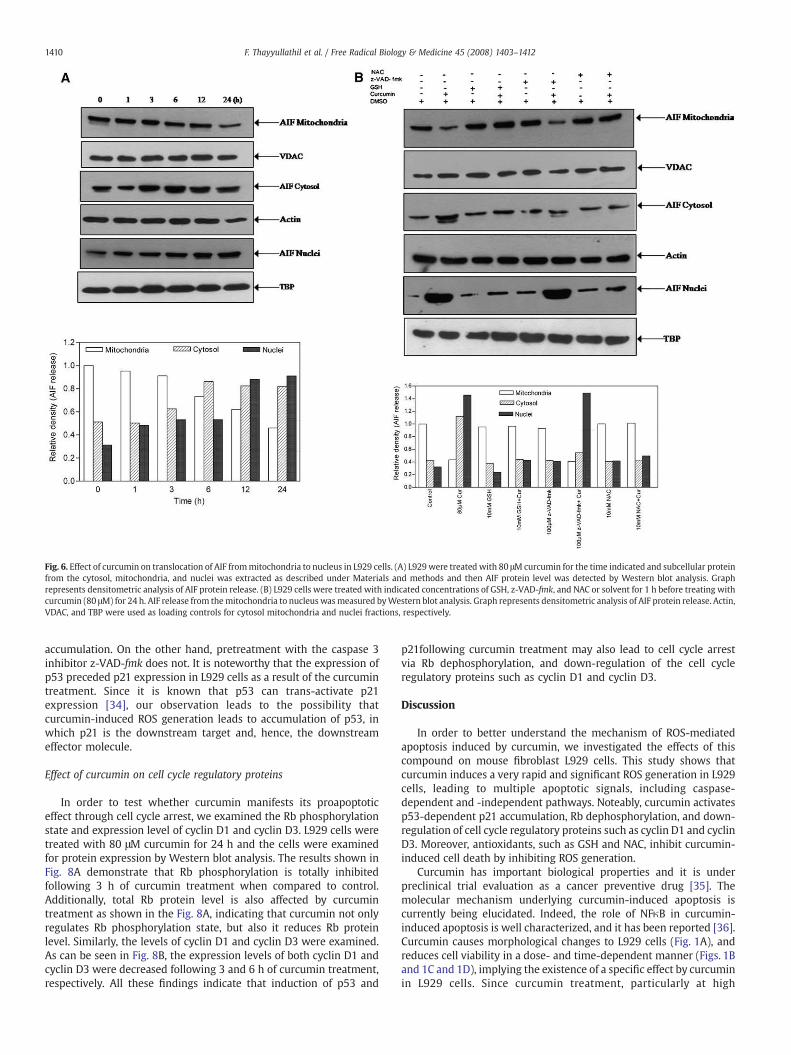
Fig. 6. Effect of curcumin on translocation of AIF frommitochondria to nucleus in L929 cells. (A) L929were treated with 80 μM curcumin for the time indicated and subcellular proteinfrom the cytosol, mitochondria, and nuclei was extracted as described under Materials and methods and then AIF protein level was detected by Western blot analysis. Graphrepresents densitometric analysis of AIF protein release. (B) L929 cells were treated with indicated concentrations of GSH, z-VAD-fmk, and NAC or solvent for 1 h before treating withcurcumin (80 μM) for 24 h. AIF release from themitochondria to nucleus wasmeasured byWestern blot analysis. Graph represents densitometric analysis of AIF protein release. Actin,VDAC, and TBP were used as loading controls for cytosol mitochondria and nuclei fractions, respectively.
1410 F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
accumulation. On the other hand, pretreatment with the caspase 3inhibitor z-VAD-fmk does not. It is noteworthy that the expression ofp53 preceded p21 expression in L929 cells as a result of the curcumintreatment. Since it is known that p53 can trans-activate p21expression [34], our observation leads to the possibility thatcurcumin-induced ROS generation leads to accumulation of p53, inwhich p21 is the downstream target and, hence, the downstreameffector molecule.
Effect of curcumin on cell cycle regulatory proteins
In order to test whether curcumin manifests its proapoptoticeffect through cell cycle arrest, we examined the Rb phosphorylationstate and expression level of cyclin D1 and cyclin D3. L929 cells weretreated with 80 μM curcumin for 24 h and the cells were examinedfor protein expression by Western blot analysis. The results shown inFig. 8A demonstrate that Rb phosphorylation is totally inhibitedfollowing 3 h of curcumin treatment when compared to control.Additionally, total Rb protein level is also affected by curcumintreatment as shown in the Fig. 8A, indicating that curcumin not onlyregulates Rb phosphorylation state, but also it reduces Rb proteinlevel. Similarly, the levels of cyclin D1 and cyclin D3 were examined.As can be seen in Fig. 8B, the expression levels of both cyclin D1 andcyclin D3 were decreased following 3 and 6 h of curcumin treatment,respectively. All these findings indicate that induction of p53 and
p21following curcumin treatment may also lead to cell cycle arrestvia Rb dephosphorylation, and down-regulation of the cell cycleregulatory proteins such as cyclin D1 and cyclin D3.
Discussion
In order to better understand the mechanism of ROS-mediatedapoptosis induced by curcumin, we investigated the effects of thiscompound on mouse fibroblast L929 cells. This study shows thatcurcumin induces a very rapid and significant ROS generation in L929cells, leading to multiple apoptotic signals, including caspase-dependent and -independent pathways. Noteably, curcumin activatesp53-dependent p21 accumulation, Rb dephosphorylation, and down-regulation of cell cycle regulatory proteins such as cyclin D1 and cyclinD3. Moreover, antioxidants, such as GSH and NAC, inhibit curcumin-induced cell death by inhibiting ROS generation.
Curcumin has important biological properties and it is underpreclinical trial evaluation as a cancer preventive drug [35]. Themolecular mechanism underlying curcumin-induced apoptosis iscurrently being elucidated. Indeed, the role of NFκB in curcumin-induced apoptosis is well characterized, and it has been reported [36].Curcumin causes morphological changes to L929 cells (Fig. 1A), andreduces cell viability in a dose- and time-dependent manner (Figs. 1Band 1C and 1D), implying the existence of a specific effect by curcuminin L929 cells. Since curcumin treatment, particularly at high

Fig. 7. Effect of curcumin treatment on p53, p21, and Bax expression in L929 cells. (A)L929 cells were treated with 80 μM curcumin for the indicated time and proteinexpression level was measured by Western blot analysis. (B) Effect of GSH, z-VAD-fmk,and NAC on expression level of p53 and p21 proteins. L929 cells were incubated withGSH, z-VAD-fmk, and NAC or solvent for 1 h before treatment with curcumin. Equalamounts of cell lysates (40 μg) were subjected to electrophoresis and analyzed byWestern blotting performed as indicated under Materials and methods. Western blotdata presented are representative of those obtained in at least three separateexperiments.
Fig. 8. Effect of curcumin on cell cycle regulatory proteins. (A) L929 cells were treatedwith 80 μM curcumin for the indicated time. The phosphorylated Rb and total Rbprotein were detected with specific antibodies. (B) L929 cells were treated with 80 μMcurcumin for different time periods and expression levels of cyclin D1 and cyclin D3proteins were measured by Western blot analysis. Western blot data presented arerepresentative of at least three separate experiments.
1411F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
concentrations, causes a typical DNA ladder pattern of internucleoso-mal fragmentation (Fig. 1E) this specific effect of curcumin on L929cells is leading these cells to undergo apoptosis.
At apoptosis-inducing concentrations, curcumin induces ROSformationwithin 5min of treatment (Fig. 2A), and this ROS generationis real (the dye DCFH-DA being oxidized), and not due to changes inprobe uptake, ester cleavage, or probe efflux (Fig. 2B). ROS includesfree radicals such as superoxide (O2 ), hydroxy radical (OH-), andnonradical derivatives of oxygen such as H2O2. This free radicalgeneration has been shown to accelerate cell death by damagingcellular components, including DNA, proteins, and lipid membranes[37]. Additionally, it is observed that curcumin rapidly depletedintracellular GSH, which potentiates the elevation of ROS such asH2O2. Thus, our data support the fact that oxidative stress plays a roleas a common mediator of apoptosis.
Caspases play critical role in the initiation and execution ofapoptosis which is activated by cytochrome c [12,13]. Curcumininduces a dramatic mitochondrial cytochrome c release in L929 cells(Fig. 3), a critical step in the formation of the apoptosome, whichultimately cleaves procaspase 3 to form the active caspase 3. In thisstudy, curcumin induces the activation of caspase 3 in a dose- andtime-dependent manner (Figs. 4A and B). Based on these findings it isconceivable that ROS generation leads to the release of cytochrome c,caspase 3 activation, and PARP cleavage during the curcumin-inducedcaspase-dependent apoptotic process. Moreover, the rapid ROSgeneration following curcumin treatment and the resultant effectsare all inhibited by pretreatment of L929 cells with the antioxidantsGSH and NAC (Fig. 5), a point which underscores the role of ROS incurcumin mechanism of action.
Apoptosis-inducing factor is a mitochondrial flavoprotein, whichnormally resides in the inner mitochondrial membrane, and it
possesses an NADH oxidase activity [38]. It has been proposed thatAIF acts as a scavenger of ROS, particularly peroxides [14].Translocation of AIF from the mitochondria to the cytosol as wellas to the nucleus exerts caspase-independent apoptosis in anumber of model systems [39,40]. Importantly, various proapopto-tic effects of AIF are not inhibited by pharmacological caspaseinhibitors such as z-VAD-fmk or boc-aspartyl(Ome)-fluoromethylk-etone (BAF), indicating that AIF can trigger nuclear apoptosis in acaspase-independent manner [41]. In this study, curcumin inducesAIF translocation from mitochondria to the cytosol and the nucleus(Fig. 6A). Pretreatment with GSH and NAC prevents this transloca-tion of AIF and apoptosis, whereas z-VAD-fmk pretreatment doesnot (Fig. 6B). This provides obvious support for the contention thatcurcumin-induced ROS is also able to induce caspase-independentapoptosis.
The tumor suppressor protein p53 plays a key role in theinduction of apoptosis, and cell cycle arrest in response to a varietyof genotoxic stresses, but the underlying mechanism for theprogression of p53-dependent apoptosis is largely unknown.However, some reports suggest a role for ROS as a potential mediatorof p53-dependent apoptosis [42]. In this study, treatment of L929cells with curcumin results in marked up-regulation of p53 and p21protein (Fig. 7A), Rb protein dephosphorylation, and a down-regulation of cyclin D1 and cyclin D3 (Figs. 8A and 8B). Furthermore,pretreatment with antioxidants, such as GSH and NAC, prevents theactivation of p53 and p21, as well as protecting L929 cells fromundergoing apoptosis. However, pretreatment with the caspase 3inhibitor is ineffective (Fig. 7B). Therefore, it is likely that curcumin-induced ROS generation is associated with cell cycle arrest throughinduction of p53, reduction of cyclin D1 and D3, and depho-sphorylation of Rb in L929 cells. Moreover, p21 can be up regulatedin both a p53-dependent and a p53-independent manner [43]. Sincetreatment of L929 cells with curcumin results in concomitant up-regulation of p21 and p53 (Fig. 7B), it is likely that p21 induction incurcumin-treated L929 cells might be due to the up-regulation ofp53 protein.

1412 F. Thayyullathil et al. / Free Radical Biology & Medicine 45 (2008) 1403–1412
Concluding remarks
In conclusion, we have demonstrated that curcumin-inducedapoptosis in L929 cells involves at least three different signalingpathways, both caspase-dependent and caspase-independent, withROS being a prime initiating signaling candidate. A molecularunderstanding of the mechanism of action of curcumin is aprerequisite in establishing its therapeutic relevance. We feel thatour study has contributed to this understanding. However, thesignificance of activation of multiapoptotic signaling by curcuminrequires more in-depth investigations.
Acknowledgments
This work was financially supported by grants from The Terry FoxFoundation for Cancer Research (UAE-05-98), The Sheikh HamdanAward for Medical Sciences, and grants from the Faculty of Medicineand Health Sciences, United Arab Emirates University. We thank Prof.Yusuf A. Hannun (Department of Biochemistry and Molecular Biology,MUSC, USA) for reviewing this manuscript.
References
[1] Anto, R. J.; Kuttan, G.; Dinesh Babu, K.; Rajasekharan, K. N.; Kuttan, R. Anti-inflammatory activity of natural and synthetic curcuminoids. Pharm. Pharmacol.Commun. 4:103–106; 1998.
[2] Azuine, M. A.; Kayal, J. J.; Bhide, S. V. , Protective role of aqueous turmeric extractagainst mutagenicity of direct-acting carcinogens as well as benzo[a]pyrene-induced genotoxicity and carcinogenicity. J. Cancer Res. Clin. Oncol. 118:447–452;1992.
[3] Rao, C. V.; Rivenson, A.; Simi, B.; Reddy, B. S. Chemoprevention of coloncarcinogenesis by dietary curcumin, a naturally occurring plant phenoliccompound. Cancer Res. 55:259–266; 1995.
[4] Sreejayan, N.; Rao, M. N. A. Curcuminoids as potent inhibitors of lipidperoxidation. J. Pharm. Pharmacol. 46:1013–1016; 1994.
[5] Pan, M. H.; Lin-Shiau, S. Y.; Lin, J. K. Comparative studies on the suppression ofnitric oxide synthase by curcumin and its hydrogenated metabolites throughdown-regulation of IκB kinase and NFκB activation in macrophages. Biochem.Pharmacol. 60:1665–1676; 2000.
[6] Singh, S.; Aggarwal, B. B. Activation of transcription tactor NFκB is suppressed bycurcumin (diferuloylmethane). J. Biol. Chem. 270:24995–25000; 1995.
[7] Aggarwal, S.; Ichikawa, H.; Takada, Y.; Sandur, S. K.; Shishodia, S.; Aggarwal, B. B.Curcumin (diferuloylmethane) down-regulates expression of cell proliferationand antiapoptotic and metastatic gene products through suppression of IκBαkinase and Akt activation. Mol. Pharmacol. 69:195–206; 2006.
[8] Mukhopadhyay, A.; Banerjee, S.; Joe Stafford, L.; Xia, C.; Liu, M.; Aggarwal, B. B.Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma proteinphosphorylation. Oncogene 21:8852–8861; 2002.
[9] Jiang, M. C.; Yang-Yen, H. F.; Lin, J. K.; Yen, J. J. Differential regulation of p53, c-myc,bcl-2 and bax protein expression during apoptosis induced by widely divergentstimuli in human hepatoblastoma cells. Oncogene 13:609–616; 1996.
[10] Anto, R. J.; Mukhopadhyay, A.; Denning, K.; Aggarwal, B. B. Curcumin (difer-uloylmethane) induces apoptosis through activation of caspase-8, BID cleavageand cytochrome c release: its suppression by ectopic expression of Bcl-2 andBcl-xl. Carcinogenesis 23:143–150; 2002.
[11] Thompson, C. B. Apoptosis in the pathogenesis and treatment of disease. Science267:1456–1462; 1995.
[12] Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathwaysof caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol. 15:269–290;1999.
[13] Izban, K. F.; Wrone-Smith, T.; Hsi, E. D.; Schnitzer, B.; Quevedo, M.; Alkan, S.Characterization of the interleukin-1β-converting enzyme/Ced-3-family protease,caspase 3/CPP32, in Hodgkin's disease: lack of caspase 3 expression in nodularlymphocyte predominance Hodgkin's. disease. Am. J. Pathol.154:1439–1447; 1999.
[14] Susin, S. A.; Lorenzo, H. K.; Zamzami, N.; Marzo, I.; Snow, B. E.; Brothers, G. M.;et al. Molecular characterization of mitochondrial apoptosis-inducing factor.Nature 397:441–446; 1999.
[15] Du, J.; Suzuki, H.; Nagase, F.; Akhand, A. A.; Yokoyama, T.; Miyata, T.; Kurokawa, K.;
Nakashima, I. Methylglyoxal induces apoptosis in Jurkat leukemia T cells byactivating c-Jun N-terminal kinase. J. Cell. Biochem. 77:333–344; 2000.
[16] Li, L. Y.; Luo; Wang, X. Endonuclease G is an apoptotic DNase when released frommitochondria. Nature 412:95–99; 2001.
[17] Susin, S. A.; Lorenzo, H. K.; Zamzami, N.; Marzo, I.; Brenner, C.; Larochette, N.; et al.Mitochondrial release of caspase-2 and -9 during the apoptotic process. J. Exp.Med. 189:381–394; 1999.
[18] Widalk, P.; Li, L. Y.; Wang, X.; Garrard, W. T. Action of recombinant humanapoptotic endonuclease G on nacked DNA and chromatin substrate: cooperationwith exonuclease and DNase 1. J. Biol. Chem. 276:48404–48409; 2001.
[19] Bhaumik, S.; Anjum, R.; Rangaraj, N.; Pardhasaradhi, B. V. V.; Khar, A. Curcuminmediated apoptosis in AK-5 tumor cells involves the production of reactive oxygenintermediates. FEBS Lett. 456:311–314; 1999.
[20] Orrenius, S. Mechanisms of oxidative cell damage. In: Pol, G., Albano, E., Dianzani,M.U. (Eds.), Free radicals: from basic science to medicine. Birkhauser Verlag, Basel,pp. 47–64; 1993.
[21] Vaux, D. L.; Korsmeyer, S. J. Cell death in development. Cell 96:245–254; 1999.[22] Shih, C. M.; Ko, W. C.; Wu, J. S.; Wei, Y. H.; Wang, L. F.; Chang, E. E.; et al. Mediating
caspase-independent apoptosis by cadmium through mitochondria-ROS pathwayin MRC-5 fibroblasts. J. Cell. Biochem. 91:383–397; 2004.
[23] Bauer, M. K.; Vogt, M.; Los, M.; Siegel, J.; Wesselborg, S.; Schulze-Osthoff, K. Role ofreactive oxygen intermediates in activation-induced CD95 (APO-1/Fas) ligandexpression. J Biol. Chem. 273:8048–8055; 1998.
[24] Levine, A. J. p53, the cellular gatekeeper for growth and division. Cell 88:323–331;1997.
[25] Henkels, K. M.; Turchi, J. J. Cisplatin-induced apoptosis proceeds by caspase 3-dependent and -independent pathways in cisplatin-resistant and -sensitivehuman ovarian cancer cell lines. Cancer Res. 59:3077–3083; 1999.
[26] Pique, M.; Barragan, M.; Dalmau, M.; Bellosillo, B.; Pons, G.; Gil, J. Aspirin inducesapoptosis through mitochondrial cytochrome c release. FEBS Lett. 480:193–196;2000.
[27] Liu, X.; Kim, C. N.; Yang, J.; Jemmerson, R.; Wang, X. Iduction of apoptotic programin cell free extract: requirement for dATP and cytochrome c. Cell 86:147–157;1996.
[28] Buttke, T. M.; Sandstrom, P. A. Oxidative stress as amediator of apoptosis. Immunol.Today 15:7–10; 1994.
[29] Voehringer, D. W. BCl-2 and glutathione: alterations in cellular redox state thatregulate apoptosis sensitivity. Free Radic. Biol. Med. 27:945–950; 1999.
[30] Armstrong, J. S.; Steinauer, K. K.; Hornung, B.; Irish, J. M.; Lecane, P.; Birrell, G. W.;Peehl, D. M.; Knox, S. J. Role of glutathione depletion and reactive oxygen speciesgenerations in apoptotic signaling in a human B lymphoma cell line. Cell DeathDiffer. 9:252–263; 2002.
[31] Green, D. R.; Reed, J. C. Mitochondria and apoptosis. Science 281:1309–1312; 1998.[32] Desagher, S.; Osen-Sand, A.; Nichols, A.; Eskes, R.; Montessuit, S.; Lauper, S.; et al.
Bid-induced conformational change of Bax is responsible for mitochondrialcytochrome c release during apoptosis. J. Cell Biol. 144:891–901; 1999.
[33] Jung, E. M.; Lim, J. H.; Lee, T. J.; Park, J. W.; Choi, K. S.; Kwon, T. K. Curcuminsensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through reactive oxygen species-mediated upregulation ofdeath receptor 5 (DR5). Carcinogenesis 26:1905–1913; 2005.
[34] Choudhuri, T.; Pal, S.; Das, T.; Sa, G. Curcumin selectively induces apoptosis inderegulated cyclin D1-expressed cells at G2 phase of cell cycle in a p53-dependentmanner. J. Biol. Chem. 280:20059–20068; 2005.
[35] Kawamori, T.; Lubet, R.; Steele, V. E.; Kelloff, G. J.; Kaskey, R. B.; et al.Chemopreventive effect of curcumin, a naturally occurring anti-inflammatoryagent, during the promotion/progression stages of colon cancer. Cancer Res.59:597–601; 1999.
[36] Anto, R. J.; Maliekal, T. T.; Karunagaran, D. L-929 Cells harboring ectopicallyexpressed RelA resist curcumin-induced apoptosis. J. Biol. Chem. 275:15601–15604;2000.
[37] Halliwell, B.; Gutteridge, J. M. C. Free radicals in biology and medicine, second ed.Clarendon, Oxford; 1989.
[38] Punj, V.; Chakrabarty, A. M. Redox proteins in mammalian cell death: anevolutionarily conserved function in mitochondria and prokaryotes. Cell. Micro-biol. 5:225–231; 2003.
[39] Cande, C.; Cecconi, F.; Dessen, P.; Kroemer, G. Apoptosis-inducing factor (AIF): key tothe conserved caspase-independent pathways of cell death? Cell Sci. 115:4727–4734; 2002.
[40] Cregan, S. P.; Dawson, V. L.; Slack, R. S. Role of AIF in caspase-dependent andcaspase-independent cell death. Oncogene 23:2785–2796; 2004.
[41] Cande, C.; Vahsen, N.; Garrido, C.; Kroemer, G. Apoptosis-inducing factor (AIF):caspase-independent after all. Cell Death Differ. 11:591–595; 2004.
[42] Johnson, T. M.; Yu, Z. X.; Ferrans, V. J.; Lowenstein, R. A.; Finkel, T. Reactive oxygenspecies are downstream mediators of p53-dependent apoptosis. Proc. Natl. Acad.Sci. USA 93:11848–11852; 1996.
[43] Gartel, A. L.; Serfas, M. S.; Tyner, A. L. p21-negative regulator of the cell cycle. Proc.Soc. Exp. Biol. Med. 213:138–149; 1996.
Copyright © 2022 FDOKUMEN




















