
Microfoundations of Internal and External Absorptive Capacity Routines
Upload
uni-duesseldorfCategory
view
1download
0
THE ANATOMICAL RECORD 233:387-398 (1992)
Protein-Gold Transport in the Endocytic Complex of Trophotaenial Absorptive Cells in the Embryos of a Goodeid Teleost
JOACHIM F. SCHINDLER AND HARTMUT GREVEN Institut fur Anatomie, Uniuersitut Regensburg (J.F.S.), 0-8400 Regensburg, and Institut
fur Zoologie II, Uniuersitut Dusseldorf (H.G.), 04000 Dusseldorfi Germany
A B S T R A C T This paper reports adsorptive endocytosis of exogenous proteins by the trophotaenial absorptive cells (TACs) in the viviparous goodeid teleost, Ameca splendens. In vitro incubations were performed with gold conjugated to bovine serum albumin (Au-BSA), human transferrin (Au-HTf), fetuin (Au-Fet), and asialofetuin (Au-ASFet). Localization of gold label on the TAC surface was nearly exclusive to patches of an amorphous coat associated with part of the in- termicrovillous plasma membrane. On addition of excess BSA, HTf, Fet, or ASFet to incubation media containing, respectively, Au-BSA, Au-HTf, Au-Fet, or Au- ASFet, the density of gold particles adsorbed on the TAC surface decreased dras- tically. Moreover, attachment of the four protein-gold complexes to the same plasma membrane sites was suggested by reciprocal inhibitory effects. Further proteins such as hemoglobin, myoglobin, and cytochrome c were as well potent inhibitors of Au-BSA and Au-HTf binding and uptake.
Binding to TACs of native BSA or HTf was visualized by immunogold labeling. The interactions between proteins and binding sites required both the presence of Ca2+ and appropriate pH >6.6. Analyses of the concentration-dependent BSA and HTf binding curves, plotted from morphometric data, resulted in apparent disso- ciation constants, Kds, of approximately 5 x M, respectively.
Following binding at the TAC surface and internalization via clathrin-coated pits and vesicles the several ligands were routed along the lysosomal pathway with transit through the endosomal compartment. Prolonged incubation periods led to massive intracellular accumulation of tracer proteins. The effects of NH,C1 (10 mM) treatment on TACs included enormous cytoplasmic vacuolation, a reversible loss of protein binding sites on the Dlasma membrane, and a block in the transport
M and 4 x
- of protein-gold complexes to lysosomes.
In viviparous goodeid teleosts, matrotrophic rela- tionships include the maternal-fetal transfer of mem- brane impermeant molecules via an inner ovarian pla- cental association termed a trophotaenial placenta (Lombardi and Wourms, 1985a,b). Topographical rela- tionships in the gravid fish ovary have been illustrated previously (Lombardi and Wourms, 1985a; Schindler, 1990). In brief, an ovarian cavity accommodates the embryos during most of gestation until term. In this period, the placental association is composed of an in- ner ovarian epithelium and of embryonic extracorporal anal appendages, the so-called trophotaeniae. As there is no direct attachment of the fetal membranes to ma- ternal tissue, an embryotrophic liquid mediates in the transfer of nutrients. The occurrence of various pro- teins in the embryotrophic liquid has been recently demonstrated (Schindler et al., 1988). Trophotaeniae can thus provide a useful model for studies of molecular transport, and the bulk-phase endocytosis of horserad- ish peroxidase (HRP) as well as the transport of cat- ionized ferritin (CF) as an electrostatic adsorbate have been studied previously (Lombardi an Wourms, 1985b; Schindler and de Vries, 1987, 1988). As a new facet, present data demonstrate binding and internalization
0 1992 WILEY-LISS, INC
0 1992 Wiley-Liss, Inc.
of several protein-gold complexes and native proteins by a process resembling that observed in the receptor- mediated endocytosis of physiological ligands. Various probes of mammalian sources were employed since the homologous proteins from goodeid embryotrophic liq- uid are unavailable.
MATERIALS AND METHODS Animals
Gravid females of Ameca splendens (Miller and Fitzsimons, 1971) were anesthetized with tricaine methanesulfonate (MS 222, Sandoz). After decapita- tion, and then opening of the abdominal cavity, the ovary harboring embryos in late stages of gestation was excised. The embryos were then liberated into chilled preincubation buffer and the trophotaeniae sev-
Received May 10, 1991; accepted November 18, 1991. Address reprint requests to Dr. Joachim F. Schindler, Institut fur
Anatomie der Universitat Regensburg, Universitatsstrane 31, D-8400 Regensburg, Germany.

388 J.F. SCHINDLER AND H. GREVEN
ered. Most of the young fish could be raised up to en- sure continuation of the live stock.
Reagents and Buffers Bovine serum albumin (BSA), human diferric-trans-
ferrin (HTf), fetuin from fetal calf serum, type IV (Fet), asialofetuin (ASFet), asialomucin from bovine submax- illary gland (ASMuc), bovine hemoglobin (Hb), myoglo- bin of horse heart (Mb), cytochrome c, type IV from horse heart (Cyt.c), heparin, N-acetylneuraminic acid (NANA), N-acetyl-D-glucosamine, D( + )-galactose, D( + )-mannose, a-D( + )-fucose, goat anti-BSA IgG, and goat anti-human transferrin IgG were purchased from Sigma Chemical Co., St. Louis, MO, and rabbit anti- goat IgG coupled to 5 nm colloidal gold from Janssen Biotech. N.V. Olen, Belgium. All other chemicals were reagent grade.
Incubation medium, fish saline (FS), containing 113 mM NaC1, 5 mM KC1, 2 mM CaCl,, 1 mM MgSO,, 1 mM Na,HPO,, 1 mM NaHCO,, 1 mM sodium pyru- vate, 5 mM glucose, and 5 mM Hepes (pH 7.4) was prepared according to Wallace and Selman (1978). For studies on divalent cation dependency, FS was also pre- pared omitting Ca2 + , The extracellular medium was adjusted to pH 6.6 by substituting the buffer reagents in FS for succinic acid and bis Tris.
Preparation of Protein-Gold Conjugates Gold sols of 5 nm (Au,) and 15 nm (Au,,) particle size
resulted from a tannic acid-sodium citrate reduction of a chloroauric acid solution (Muhlpfort, 1982). The op- timum amounts of BSA, HTf, Fet, ASFet, and ASMuc to stabilize Au, or Au,, were determined by adding increasing amounts of protein diluted in 5 mM NaCl until the colloid no longer flocculated after the addition of 10% NaCl solution (Horisberger and Rosset, 1977). To remove practically all unconjugated ligand, the col- loids were centrifuged four times and each time the pellets immediately resuspended in FS buffer contain- ing 0.5 mg/ml Carbowax 20-M. Stock solutions were adjusted to concentrations with OD515 0.1 and OD,,, 0.3 for Au,-conjugates and Au,,-conjugates, respec- tively.
lmmunogold Labeling After incubations with 0.1 mg/ml and 1 mg/ml BSA
or HTf for 1 hour at 25"C, trophotaeniae were fixed with 3% paraformaldehyde and 0.3% glutaraldehyde in 0.1 M cacodylate for 30 minutes, dehydrated in etha- nol, embedded in L R White resin, and polymerized at 4°C. Ultrathin sections on nickel grids were floated on drops of 10 mM NH,C1 in phosphate buffered saline (PBS) for 1 hour in order to block free aldehyde groups. Alternating incubations with antibodies and washing buffers were carried out in PBS supplemented with 0.5% gelatin. The sequence of incubations started with a 30-minute rinse, followed by exposure to either goat anti-BSA IgG (1:500) or goat anti-HTf IgG (1:500) for 30 minutes, continued by a wash interval of 30 min- utes, then exposure to auro anti-goat IgG (1:25) for 30 minutes, and a final 30-minute rinse. Postfixation was in 2% glutaraldehyde for 30 minutes.
Trophotaeniae exposed to 0.1 mg/ml BSA or HTf for 1 hour at 0°C were fixed as above and then the tissue immediately submitted to immunohistochemical treat-
ment. After postfixation with 2% glutaraldehyde, spec- imens were embedded for electron microscopy as usual. In similar experiments, Ca2+ deficient FS or buffer a t pH 6.6 was used.
Incubation Protocols Using Protein-Gold Sols Although there was no indication that age of the
embryos was critical to the results, only sibling com- parison was regarded in the same experiment. All me- dia were adjusted to pH 7.4. Unless otherwise indi- cated, incubation intervals were 1 hour and the temperature was maintained a t 25°C.
Pieces of trophotaeniae were incubated with stock solutions of Au,,-BSA, Au,-HTf, Au,,-HTf, Au,,-Fet, Au,,-ASFet, and Au,,-ASMuc at 0°C in the absence or presence of each 30 KM BSA, HTf, Fet, Mb, Hb, or
Protein-gold uptake was demonstrated by incubating trophotaeniae with stock solutions of Au,-HTf, Au,,- BSA, and Au15-Fet in the absence or presence of each 30 pM BSA, HTf, Fet, Mb, Hb, or Cyt.c. Similar proto- cols included the addition of 4 mg/ml heparin, 1 mg/ml NANA, N-acetylglucosamine, galactose, mannose, or fucose.
Time sequence analysis with diluted (1:15) Au,-HTf was at time intervals of 20 seconds, 1 minute, 3 min- utes, 10 minutes, 30 minutes, 1 hour, 2 hours, and 4 hours.
Double labeling experiments were performed with Au,-HTf and Au,,-HTf. Trophotaeniae were incubated with either tracer substance, chased in pure FS for 30 minutes or 1 hour, and then reincubated with the dif- ferent-sized gold probe.
Temperature-dependent uptake of Au,,-BSA was de- termined at O"C, 5"C, 1O"C, and 15°C.
The effects of an acidotropic agent were demon- strated by incubating a trophotaenia with Au,-HTf in the presence of 10 mM NH,Cl.
Incubation periods were terminated by immersing the trophotaeniae into fixative. For standard electron microscopy, the reagent contained 3% paraformalde-
Cyt.c.
Fig. 1 . Electron micrograph depicting a section of bilayered tropho- taenial epithelium with a subtending capillary (C). Several TACs cover a basal stratum of squamous epidermis-derived cells (BC), though another plane of section would probably demonstrate pseudo- stratification. A wide labyrinth of interstices is gaping between the two cell types bridged by interconnected processes from both sides. The organization of the TAC supranuclear cytoplasm gives rise to zonation in (I) an apical endocytic complex including the surface plasma membrane in contact with the ovary lumen (OL), and (11) an underlying mitochondria1 domain colocalizing large irregularly delin- eated dark bodies. x 3,800.
Fig. 2. Portion of a TAC highlighting the polymorphism of vacuolar components of the apical endocytic complex. Deep plasma membrane invaginations honeycomb the subsurface cytoplasm. Vacuoles identi- fied as endosomes (E) are characterized by electron lucent unilater- ally invaginated lumina (arrowhead). A small endosome is connected with two dense apical tubules (arrow). x 38,000.
Fig. 3. Portion of the apical surface o f a TAC after simultaneous fixation with paraformaldehyde, glutaraldehyde, and osmium tetrox- ide. An amorphous fuzzy coat (arrowhead) is associated with the ex- ternal leaflet of a clathrin-coated membrane invagination. The frag- ments of thick walled dense tubules (arrows) show a periodical, diagonally oriented stripy pattern. x 80,000.
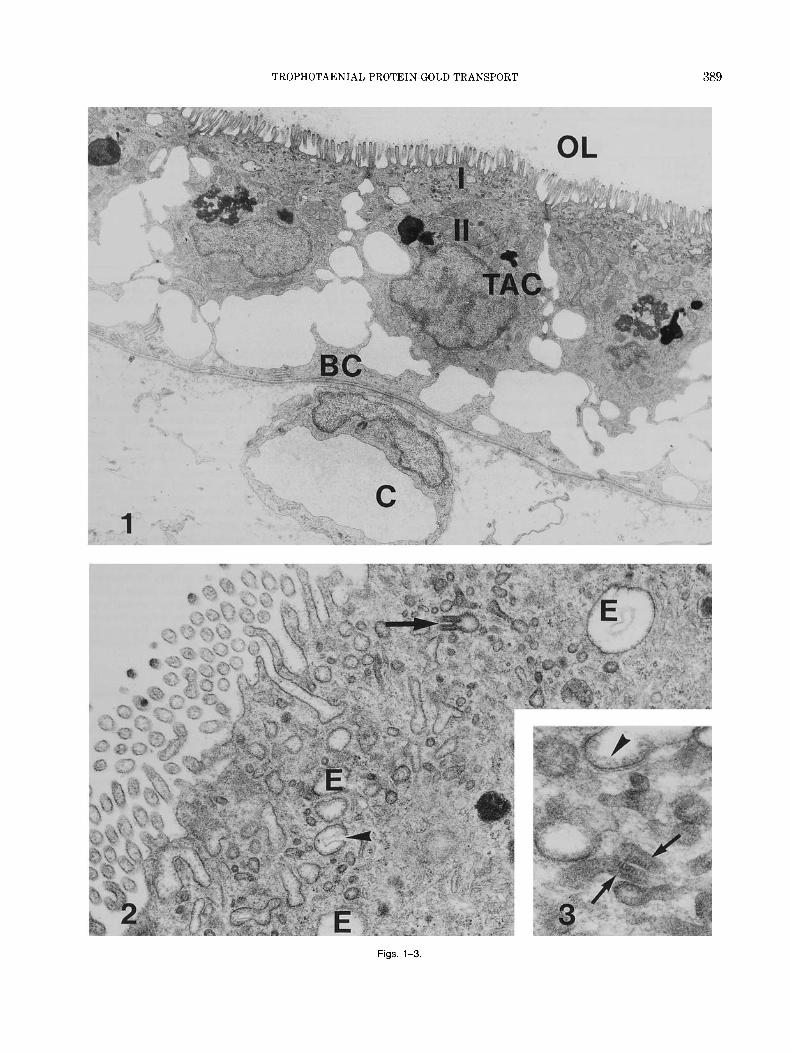
TROPHOTAENIAL PROTEIN-GOLD TRANSPORT 389
Figs. 1-3.

390 J.F. SCHINDLER AND H. GREVEN
TABLE 1. Comuetitive inhibition of urotein-gold binding to TACs'
Addition
Au,,-BSA (1) Au15-BSA (2)
AuK-HTF (2) Au,-HTf (1)
Au;,-HTf (1)
Au,,-Fet Au,,-ASFet Au, ,-ASMuc
Auls-HTf (2)
None + BSA + HTf 5.4 0.6 0.8 6.5 0.3 0.1
17.8 1.4 1.4 13.3 1.8 1.5
5.5 0.2 0.2 4.1 0.6 0.9 1.9 0.0 0.1 2.8 0.1 0.0
0.0 0.2 -
+ Fet + Mb + Hb + Cyts 1 .o 1.0 0.8 0.7 0.9 2.0 1.5 0.4 0.7 0.1 0.1 0.0
. .
0.3 0.4 0.4 0.9 1.8 1.5 1.9 1.2 1.3 0.1 - 0.2 0.6 - 0.6 - - -
- - -
- - -
'Prechilled trophotaeniae were incubated with stock solutions (1 ml) of the protein-gold conjugates tabulated in the absence or presence of 30 pM of unconjugated protein for 1 hour a t 0°C prior to fixation and embedding. Each figure represents the number of gold particlesipm plasma membrane (for morphometric details, see Materials and Methods section) averaging the analysis of approximately 500 pm of epithelial surface representing one individual. The experiments included embryos of two broods (1) and (2).
hyde and 1% glutaraldehyde in 0.1 M cacodylate sup- plemented with 2 mM CaC1,. Ultrathin sections were stained with uranyl acetate and lead citrate.
Morpho~efric Anaiyses
For determinations of the surface particle density of protein-gold complexes or immunogold label, the length of the apical plasma membrane was measured on low power electron micrographs. As microvilli were not a site of specific labeling, these were omitted in the evaluations of the effective surface by a morphomat (Digi-Plot WX 4671, Watanabe, Japan). The marker particles were counted while scrutinizing the respec- tive loci in the electron microscope a t high magnifica- tion. Only those particles that had contact with the amorphous coat of the trophotaenial absorptive cell (TAC) surface proper and in the plasma membrane in- vaginations were counted. To improve statistical rele- vance, three blocks of each preparation were micro- tomed, and only ultrathin sections exhibiting a silver interference color were analyzed. Care was taken that the epithelium was cut perpendicular to the basement lamina.
RESULTS Trophotaenial Morphology
The trophotaenial epithelium in goodeid teleosts showed a remarkably heterogeneous structure. Of its two cell types-TACs and epidermal cells-TACs were prevalent. On most of the trophotaenial surface, they formed a monolayer covering the basement lamina, or overlying a close stratum of epidermal cells (Fig. 1). Transitions between the two varieties produced pseu- dostratification. Epidermis, mostly two cell layers thick, showed on the surface in few randomly scattered patches.
In the present study, ultrastructural localization of events connected with protein transport in TACs was restricted to a zone of cytoplasm (1-3 (*.m thick) con- taining abundant membranous elements (Fig. 2). This major portion of the cell's vacuolar apparatus, termed the apical endocytic complex, was composed of surface plasma membrane invaginations, coated pits and ves- icles, endosomes, dark lysosomal bodies, and dense tu- bules. In addition, a good many unclassifiable profiles were also present on micrographs.
Part of the extracellular plasma membrane on the TAC surface proper was covered by a thick amorphous coat, which was regularly more dense in the membrane invaginations showing a cytoplasmic clathrin coat (Fig. 3). Consequently, the luminal face of internalized vesicles and endosomes was likewise covered by the amorphous coat. Sorting of membrane in the endo- somes was frequently indicated by segments of smooth membrane delimiting unilateral invaginations of the vacuoles (Fig. 2; cf. Schindler and de Vries, 1988). Lysosomes were identified by having electron opaque contents, a positive reaction in response to acid phos- phatase histochemistry (Greven and Robenek, 19861, and a smooth inner and outer surface. After simul- taneous fixation with paraformaldehyde, glutaralde- hyde, and osmium tetroxide, the dense tubules re- vealed a periodical substructure (Fig. 31, similar to the one originally demonstrated in a variety of absorptive epithelia by Hatae et al. (19861. Such dense tubules, which had lengths up to 1.5 pm, were occasionally seen very close to the apical plasma membrane but were never seen connected to the cell surface. Fusions with endosomes, on the other hand, occurred fairly regu- larly.
Binding of Protein-Gold Conjugates to TACs is Inhibited by Various Native Proteins
Protein-gold conjugates of BSA, HTf, Fet, and ASFet bound to the TAC surface a t 0°C. Label was nearly exclusively localized a t the amorphous coat present on part of the intermicrovillous surface membrane and in coated invaginations. To assay whether the binding was competitively inhibitable, different batches of each protein-gold complex were also used in incubation me- dia supplemented with 30 pM of the respective uncon- jugated protein. Distinctly lower values determined in these experiments suggested that high percentages of the membrane associated gold in fact represented in- hibitable, receptor-like binding (Table 1). The question arose, as to whether different proteins competed for the same adhesion sites. Therefore, inhibition experiments included the addition of various proteins such as BSA, HTf, Fet, Hb, Mb, or Cyt.c to the sols of a definite gold complex. The data of morphometric analyses obtained from preparations with two sizes of gold beads indicate

TROPHOTAENIAL PROTEIN-GOLD TRANSPORT 391
Figs. 4-6. Time course of Au,-HTf traficking. Trophotaeniae were
Fig. 4. Binding of gold beads a t the amorphous extracellular coat
Fig. 5. Occasional marker beads (arrow) reached the lysosomal com- partment (L) within 10 minutes of incubation. Several variously ori- ented dense tubules remained unlabeled. x 40,000.
Fig. 6. Labeling ofendosomes (El and lysosomes (L) after 30 minutes of incubation. Cross-sections of dense tubules (arrowheads). x 40,000.
incubated with Au,-HTf (dilution 1:15) a t 25°C.
after 20 seconds of incubation. E, endosome. x 48,000.
the existence of protein binding sites on TACs that accept a variety of molecules (Table 1).
fucose interfere with binding and uptake of the protein- gold complexes.
Distribution of Protein-Gold Conjugates in the Endocytic Complex of TACs
Time sequence analysis of the Au,-HTf trafficking in TACs demonstrated clustering in coated pits after ap- proximately 20 seconds, entry via coated vesicles after approximately 1 minute, delivery to endosomes after approximately 3 minutes, and the appearance of occa- sional marker beads in the lysosomal compartment within 10 minutes of internalization (Figs. 4-6). Pro- longed exposure to protein saline over periods of 1 hour, 2 hours, and 4 hours, resulted in massive accu- mulation of gold label in the lysosomal compartment, and the trophotaeniae increasingly took a pinkish hue. At no time were the dense tubules labeled suggesting that this organelle was not a vehicle in the lysosomal transport chain.
The capacity of TACs to ingest Au,-HTf (Fig. 71, Au,,-HTf, Au,,-BSA (Fig. 81, Au,,-Fet (Fig. 91, or Au,,-ASMuc differed considerably. Dense labeling pat- terns were obtained with the gold conjugates of HTf and BSA. The Au,,-Fet complex acted as a relatively weak ligand, and Au,,-ASMuc failed to bind at the TAC surface. In the presence of native proteins such as HTf, BSA, Fet, Hb, Mb, or Cyt.c labeling distinctly fell off for all the gold probes (Fig. 10). Neither were the strongly anionic molecules heparin or NANA endocytic inhibitors, nor did the electrically neutral monocarbo- hydrates N-acetylglucosamine, galactose, mannose, or
Double Labeling with Au,-HTf and Au,,-HTf Regular lysosomal colocalization of Au,-HTf and
Au,,-HTf in TACs consecutively exposed to either ligand for 1 hour with an intermediate 30-minute in- terval in pure buffer was evident (Fig. 11). This dis- tinctly decreased as the interval between probes was increased to 1 hour. Inverse distributive patterns re- sulted from pulsing in the reverse order. The failure of the chaser to reach the same lysosomes with the chase given too long a start suggests that the terminals were only transitorily accessible to endocytized material. This was also demonstrated by unlabeled lysosomes, which were regularly found even after prolonged incu- bation periods with protein-gold complexes (Fig. 12). Several bands of serial sections were investigated to rule out the possibility that lysosomes were excentri- cally labeled.
Temperature-Dependent Uptake of Au , ,-6SA As receptor-mediated endocytosis in the mammalian
cell is temperature-dependent in a characteristic way (Weigel and Oka, 1981), exposure of trophotaeniae to Au,,-BSA a t O"C, 5"C, 1O"C, and 15°C was meant to demonstrate analogous effects in TACs. At 0°C endocy- tosis was negligible, and a gradual increase in uptake was observed between 5°C and 10°C. The ligand appar- ently failed to reach the lysosomal compartment. This blocking disappeared at 15°C. In eukaryotic cells, lyso-

392 J.F. SCHINDLER AND H. GREVEN
Figs. 7-1 1

TROPHOTAENIAL PROTEIN-GOLD TRANSPORT 393
Fig. 12. Distribution of Au -HTf (diluted 1:15) in a TAC after 2 hours of continuous uptake. Apparently void lysosomes (L) would indicate that they were no longer accessible to incoming endocytotic traffic. E, endosome. X 54,000.
Fig. 7. Binding and uptake of Au,-HTf by a TAC after a 1-hour incubation period. Gold beads adhere to restricted areas of the inter- microvillous plasma membrane associated with a fuzzy coat. An en- dosome (El apparently in the process of fusing with several vesicles is moderately labeled. Intracellular accumulation of tracer protein leads to densely labeled lysosomes (L). x 57,000.
E, Endosome; L, Lysosome. x 38,000.
bation for 1 hour. E, Endosome; L, Lysosome. x 35,000.
Fig. 8. Apical portion of a TAC incubated with Au,,-BSA for 1 hour.
Fig. 9. Micrograph depicting Au,,-Fet uptake by a TAC after incu-
Fig. 10. Incubation with Au,-HTf in the presence of 30 pM uncon- jugated HTf for 1 hour at 25°C. A lysosome (L) is labeled with only few gold beads. x 37,000.
Fig. 1 1 . Micrograph depicting double labeling with Au,,-HTf and Au,-HTf in a TAC consecutively incubated with the ligands for 1 hour with a n intermediate 30-minute chase interval in pure buffer between probes. Colocalization of both gold sizes occurs in a lysosome (L). No counterstain. x 54,000.
soma1 sequestration of external proteins seems to be a universal process involving two temperature-sensitive rate-limiting steps. In the cells of homeothermic ani- mals the budding off of micropinocytic vesicles is initi- ated at temperatures higher than 10°C. The intracel- lular fusion events for conveyance to lysosomes require the temperature rise to approximately 20°C (Dunn et al., 1980; Weigel and Oka, 1981; Sullivan et al., 1987). In the poikilothermal goodeids, the corresponding val- ues were each lower by about 10°C.
Effects of NH,CI In receptor-mediated endocytosis, drugs that raise
vacuolar pH may disrupt both lysosomal degradation and receptor recycling (Dean et al., 1984). To deter- mine the effects of a pH elevator on the fine structure and protein-gold transport in TACs, trophotaeniae

394 J.F. SCHINDLER AND H. GREVEN
Fig. 13. Endosomal (E) vacuolation in a TAC incubated with Au,-HTf in the presence of 10 mM NH,Cl.
Fig. 14. Lysosomal (L) swelling in a TAC induced by 10 mM NH,CI. x 16,000.
x 36,000.
were incubated for 1 hour with Au,-HTf in the pres- ence of 10 mM NH,C1 a t 25°C. Most conspicuous, the cytoplasm showed tremendous vacuolation (Fig. 13), a phenomenon known as osmotic swelling induced by various weakly basic compounds (Ohkuma and Poole, 1981). In many TACs, a single huge endosome formed in the apical cytoplasm. Swelling of the lysosomes was less pronounced resulting in vacuoles with an electron lucent halo surrounding a core of dark stained material (Fig. 14). The endosomes contained gold beads mostly aggregated a t the peripheral rim. Lysosomal transport was apparently totally blocked as this organelle re- mained virtually unlabeled.
In response to NH,C1 (10 mM) treatment of tropho- taeniae for 30 minutes a t 25"C, and then exposure to protein-gold sols in the continued presence of drug for 1 hour a t O'C, gold labeling of the apical surface mem- brane was reduced by approximately 40% of control as determined morphometrically. The effect was revers- ible in the absence of ammonia within 20 minutes. No depletion of surface binding sites was apparently no- ticeable on TACs analogously preincubated a t 0°C.
Localization of Native BSA and HTf by lmmunogold Labeling
After exposing trophotaeniae to native BSA or HTf for 1 hour at 0°C followed by short fixation intervals and the incubations with anti-BSA IgG or anti-HTf IgG, respectively, surface immunomarking of whole specimens was accomplished with 5 nm gold conju- gated secondary antibodies. In electron microscopy, gold beads were mainly found adherent to the amor- phous extracellular coat on the TAC intermicrovillous plasma membrane (Fig. 15). In response to the omis- sion of Ca2+ from, or the addition of EDTA to incuba- tion media, environmental pH 6.6, or excess nonimmu- nolocalized protein, the density of marker beads was significantly reduced (Table 2).
In parallel, material incubated with BSA or HTf at concentrations of 0.1 and 1 mg/ml for 1 hour at 25°C was embedded in L R White resin, and examined by on-grid immunomarking of ultrathin sections. The an- tigenicity of both BSA and HTf was well retained, al- beit a t the expense of structural preservation. Most

TROPHOTAENIAL PROTEIN-GOLD TRANSPORT 395
Fig. 15. Surface immunomarking of a TAC after incubation with 1 mglml HTf for 30 minutes at 0°C. The 5 nm gold conjugated secondary antibody is localized to the fuzzy extracellular coat mainly present in two coated pits in the process of being formed (arrows). x 72,000.
Fig. 17. Portion of a TAC embedded in L R White resin after incu- bation with 0.1 mg/ml BSA for 1 hour. Immunogold marking demon- strates protein uptake in the endosomal (E) and lysosomal (L) com- partment. Labeling intensities appear relatively poor at the low concentration of antigen. x 47,500.
Fig. 16. Portion of a TAC embedded in L R White resin after incu- bation with 1 mgiml HTf for 1 hour. Immunogold labeling is intense on endosomes (E) and even more so on dark lysosomes (L). x 26,000.
membranous outlines of cell organelles were not por- trayed yet the endosomal and lysosomal compartments defined themselves against the coarsely precipitated cytoplasmic matrix. The pattern of both gold anti-BSA and gold anti-HTf labeling (Figs. 16,17) was much the same as the one previously seen after incubations with several protein-gold conjugates. Minor differences were mostly due to less dense distribution of antibody bind- ing sites on dark lysosomes after immunomarking. This may be a result of protein degradation that does not, however, affect labeling intensities after ingestion of protein-gold complexes. Only negative results were produced in the controls.
Concentration-Dependent Binding of BSA and HTf
Saturation of BSA and HTf binding to TACs was demonstrated by morphometric analyses of particle densities on ultrathin sections of trophotaeniae incu- bated with the appropriate concentrations of protein a t 0°C and surface immunogold labeling of whole speci- mens. The apparent dissociation constants, K,s, calcu- lated from a Scatchard (1948) transformation (Fig. 18) of these data were 5 x lop7 M and 4 x lop7 M for BSA and HTf, respectively. These values are in good agree- ment with those obtained in experiments with radioio- dinated proteins (data presented elsewhere). The use of

396 J.F. SCHINDLER AND H. GREVEN
TABLE 2. Inhibition of HTf and BSA binding'
Density of Immunogold Label on the TAC Surface
Variation HTf (%) BSA (%)
None 100 t 17 100 2 6 -Ca2+ 18 ? 4 23 t 10 +EDTA (5 mM) 20 ? 6 17 t 2
pH 6.6 8 t 2 14 2 5 +BSA (20 mg/ml) 27 t 8 - + HTf (20 mg/ml) - 33 t 3
'After a 30-minute rinse in FS at 25"C, trophotaeniae were incubated with 0.1 mg/ml HTf or BSA for 1 hour at 0°C under the conditions tabulated. Fixation, embedding, immunocytochemistry of whole spec- imens, and morphometric analyses are described in the Materials and Methods section. The density of immunogold label on the TAC surface of a given sample was determined by analysis of approximately 500 pm of plasma membrane. Results are expressed as mean 2S.D. of three determinations representing different animals.
native proteins did not permit the correction for non- specific antigen binding but by localizing granular la- bel it was guaranteed that only molecules adherent to the amorphous surface coat were regarded.
DISCUSSION A Nonselective Protein Binding System
Several experiments in this study demonstrate the binding to TAGS of exogenous proteins that would nor- mally bind to cells only that express the corresponding highly specific receptors. Most remarkably, reciprocal competitive inhibition suggests a common type of bind- ing sites for all the ligands. This phenomenon could be alternatively explained by a steric factor that prevents a ligand from attaching to its own specific receptor af- ter the occupancy of a different binding site by another ligand. On the other hand, such a steric factor becomes the more unrealistic the greater the number of mole- cules involved.
Binding sites on cells that would be expected to nor- mally accept a great variety of ligands are the plasma membrane lectins. Such carbohydrate specific systems exist in, e.g., the asialoglycoprotein receptor and sim- ilar glycoreceptor systems of hepatocytes (Ashwell and Harford, 1982; Breitfeld et al., 1985), or the mannose 6-phosphate receptor for lysosomal enzymes (von Fig- ura and Hasilik, 1986). In the present system, however, lectin-like interactions can be definitely ruled out, since BSA and some of the inhibitor compounds used are nonglycosylated.
Nor does surface binding to TAGS of various protein ligands apparently result from overall electrostatic ad- hesion, since molecules are adsorbed with either sur- plus negative or surplus positive charges. The presence of strongly negative agents in the medium is without effects and cationized substances tend to aggregate a t the microvilli tips (data not presented). On the other hand, pH dependence of attachment would indicate the involvement of ionic bonds.
It follows from this that protein binding to TACs is quite unlike carbohydrate specific or purely electro- static interactions. As a consequence, present results suggest a relatively nonselective protein binding sys- tem, with docking mechanisms obviously not requiring a high degree of complementarity between ligand and
binding site. The interactions are associated with the protein structure of ligands, competitively inhibitable, saturable, and pH- and Ca2+-dependent. Displacement of gold label on the TAC surface by the native proteins indicates specific binding regardless of how many other proteins will do the very same thing. Such promiscuity demonstrates a t best the ubiquity of recognition deter- minants.
Are the proteins in this study to be called opportu- nistic ligands? We do not think so. Such classification would imply that the proteins can avail themselves of nonspecific adhesion sites or specific receptor systems similar to some enveloped viruses (Helenius and Marsh, 1982). On the one hand, the phenomenon of massive nonselective protein binding should be more ubiquitous then, and, on the other hand, widely distrib- uted physiological molecules are not likely to adapt to the requirements of some receptors on some absorptive cells. The reverse would seem to be more realistic. Ex- isting membrane constituents could possibly adopt properties of a n endocytic receptor. In the present case, virtually indiscriminate binding of proteins would be suggestive of proteinases involved. Yet, we want to hold this conjecture in abeyance, pending further ex- perimental data.
Endocytotic Uptake: Ligand and Binding Site Pathway Binding of protein ligands to TACs is rapidly fol-
lowed by endocytotic uptake. The use of diluted pro- tein-gold sols would exclude more than negligible fluid-phase uptake by implication. In adsorptive en- docytosis, protein-gold conjugates can trace intracellu- lar pathways by time sequence analysis, as the ligands remain coupled to colloidal gold after internalization (Woods et al., 1989). A major disadvantage is, however, that considerable batch-to-batch variation of unspecific binding occurs, and, moreover, the complexes behave as quasi-polymeric ligands. In this study, the tracer experiments with protein-gold complexes produced re- sults similar to the immunohistochemical data ob- tained with native proteins.
Collectively, the data gained from two approaches reveal a consistent picture of the ligand pathway irre- spective of the tracer applied. Upon entry via coated vesicles, rather swift transit through the endosomal compartment is followed by segregation to lysosomes. Tracer molecules did not enter dense apical tubules to more than a negligible extent, this being consistent with former results assigning that organelle a role in membrane recycling rather than providing a port for the importation of macromolecules (Schindler and de Vries, 1988).
While the ligand itinerary thus clearly indicates a typical lysosomal pathway, the pathway of binding sites still escapes exact definition. Only circumstantial evidence is available suggesting multiple rounds of in- ternalization rather than one-way trafficking with subsequent lysosomal degradation. Protein-gold conju- gates accumulate in lysosomes after prolonged periods of incubation. Such virtual amassment can be ad- vanced as indirect evidence for a shuttle service. Our results show that the continuous endocytotic uptake over 4 hours does not influence the capacity of TACs to internalize proteins and depletion of cell surface bind- ing sites was apparently not noticeable. On the other

TROPHOTAENIAL PROTEIN-GOLD TRANSPORT 397 ...-I m 1 0
0 Bound
0 1 2 10 20
BSA (PM)
Fig. 18. Saturation binding of BSA and HTf to TACs at 0°C. Em- bryos were incubated for 30 minutes a t 25°C in FS t o deplete surface binding sites from protein. Thereafter, pieces of trophotaeniae were exposed to appropriate concentrations of BSA (A) or HTf (B) for 1 hour at 0°C. Fixation, surface immunogold labeling, embedding, and mor- phometric analyses were as described in the Materials and Methods section. Each figure represents the average of the particle density on
hand, a reversible weak base-induced loss of surface binding sites was demonstrated. Such effect has been generally connected with receptor recycling (Dean et al., 1984). Probably, ligands dissociate from membrane in the acidified endosomal compartment as inferred from the sensitivity to environmental pH of protein binding on the surface plasma membrane.
Trophotaenial Protein Endocytosis: Present Vs. Previous Results
Endocytosis by goodeid trophotaeniae of conven- tional tracer substances, such as HRP, native ferritin (NF), or CF has been previously demonstrated (Lom- bardi and Wourms, 1985b; Grolje Wichtrup and Greven, 1985; Schindler and de Vries, 1987, 1988, 1990). Yet one must take into consideration the fact that protocols using high concentrations of content markers can produce rather trivial results demonstrat- ing no more than the ability of many eukaryotic cells to continuously internalize portions of their plasma mem- brane via micropinocytosis. Cationized adsorbates sim- ply verify the presence of negative charges in the pi- nocytic vesicle membrane.
Nonetheless, HRP and NF should bind to a nonselec- tive protein binding system irrespective of dosage. Lombardi and Wourms (1985b) reported the detection of HRP reaction product within tubular surface invagi- nations on the TACs in A. splendens. At variance with this finding, Schindler and de Vries (1988) stated the complete absence of surface label on the TACs in Xe- noophorus captiuus, and yet there was indirect evi- dence of HRP uptake other than by a fluid-phase mech- anism. A rigorous reinvestigation revealed the discrepancies in surface labeling to be obviously due to the use of different types of HRP (data not presented). The glycoprotein, HRP, seems to be a poor ligand for the protein binding system, similar to Fet. Unlike HRP, NF is nonglycosylated. It appreciably binds to the TAC surface (Grolje Wichtrup and Greven, 1985). Further investigation has shown that NF competi-
15
10
5
0 0 1 2 10 20
HTf (PM)
approximately 500 pm of TAC plasma membrane. Nonspecific immu- nolabeling defined as the values determined in controls by performing the immunomarking procedure on specimens incubated in the ab- sence of antigen, regularly less than 10% of the total binding, was subtracted from the data. Inserts: Scatchard analysis of the binding data. Ordinate: particles X pm-l x liter x pM-’. Abscissa: particles x pm-l.
tively inhibits the binding of protein-gold conjugates and vice versa (data not included in this study).
There is agreement about the channelling along the degradative pathway of HRP in TACs, but there is dis- agreement about the transport mechanism. The scheme drawn by Lombardi and Wourms (1985b) de- scribes the passage of HRP via endocytotic vesicles into an apical canalicular system and from there the deliv- ery to small collecting vesicles. Schindler and de Vries (1988) describe the uptake via clathrin-coated vesicles and direct transport to endosomes after shedding of the coat. Dense apical tubules (probably identical with the apical canalicular system) were mostly unlabeled or only faintly HRP-reactive. As outlined before, present results suggest the lysosomal pathway in TACs not to include transfer through dense apical tubules.
Biological Implications At first sight, virtually random acceptance of protein
ligands does not make much sense, and thus the effect might be rashly dismissed as some sort of nonspecific endocytosis. On closer examination, however, it comes to be realized that a “scavenger pathway” for the clear- ance of physiological macromolecules from a body fluid neither needs high selectivity nor extreme affinity. As- suming that, similar to blood serum, embryotrophe predominantly contains albumin, this would be a main natural ligand and could at the same time give other molecules a pickaback. A special ligand that finds its very own specific receptor and has a high-affinity bind- ing could still evade the lysosomal pathway.
Similar mechanisms might well be operative in other absorptive epithelia, such as, e.g., the kidney proximal tubule (Maunsbach, 1966a,b), the ra t visceral yolk sac (Williams et al., 1975; Moore et al., 1977; Dunton et al., 1988), the rete testis (Morales and Hermo, 19861, or the ductuli efferentes (Hermo and Morales, 1984). The quo- tations refer to reports of adsorptive 1251-BSA, albu- min-gold, or NF endocytosis lacking recognition of specific receptors. An adsorptive “scavenger pathway”

398 J.F. SCHINDLER AND H. GREVEN
may not always stand out against an expectable “non- specific background.” Our own experience shows that the rate of protein uptake by the TACs of A. splendens is unparalleled indeed-even among other goodeid spe- cies.
ACKNOWLEDGMENTS The skillful technical assistance of Mrs. R. Kromas is
gratefully acknowledged. This study was supported by the Deutsche Forschungsgemeinschaft (Schi 268/1-2).
LITERATURE CITED Ashwell, G., and J . Harford 1982 Carbohydrate-specific receptors of
the liver. Annu. Rev. Biochem., 51.531-554. Breitfeld, P.P., C.F. Simmons Jr., G.J.A.M. Strous, H.J. Geuze, and
A.L. Schwartz 1985 Cell biology of the asialoglycoprotein recep- tor system: a model of receptor-mediated endocytosis. Int. Rev. Cytol., 97t47-95.
Dean, R.T., W. Jessup, and C.R. Roberts 1984 Effects of exogenous amines on mammalian cells, with particular reference to mem- brane flow. Biochem. J., 21 7t27-40.
Dunn, W.A., A.L. Hubbard, and N.N. Aronson J r . 1980 Low temper- ature selectively inhibits fusion between pinocytic vesicles and lysosomes during heterophagy of ‘251-asialofetuin by the perfused rat liver. J. Biol. Chem., 2555971-5978.
Dunton, A,, M.K. Pratten, and F. Beck 1988 Uptake and digestion of 1251-labelled bovine serum albumin by rat visceral yolk sac cul- tured in vitro as a closed vesicle. Placenta, 9r303-311.
von Figura, K., and A. Hasilik 1986 Lysosomal enzymes and their receptors. Annu. Rev. Biochem., 55.167-193.
Greven, H., and H. Robenek 1986 Die Aufnahme von Protein-Gold- Komplexen durch die Trophotanien von Amecu splendens Miller and Fitzsimons (Teleostei, Cyprinodontriformes). Verh. Dtsch. Zool. Ges., 79r387 (Abstract).
GroRe Wichtrup, L., and H. Greven 1985 Uptake of ferritin by the trophotaeniae of the goodeid fish, Amecu splendens Miller and Fitzsimons 1971 (Teleostei, Cyprinodontiformes). Cytobios, 42: 33-40.
Hatae, T, M. Fujita, and H. Sagara 1986 Helical structures in the apical tubules of several absorbing epithelia. Kidney proximal tubule, visceral yolk sac and ductuli efferentes. Cell Tissue Res., 244t39-46.
Helenius, A., and M. Marsh 1982 Endocytosis of enveloped animal viruses. Ciba Found. Symp., 92t59-76.
Hermo, L., and C. Morales 1984 Endocytosis in nonciliated epithelial cells ofthe ductuli efferentes in the rat. Am. J . Anat., 171:59-74.
Horisberger, M., and J. Rosset 1977 Colloidal gold, a useful marker for transmission and scanning electron microscopy. J . Histochem. Cytochem., 25295 -305.
Lombardi, J . , and J.P. Wourms 1985a The trophotaenial placenta of a viviparous fish. 11. Ultrastructure of trophotaeniae, the embry- onic component. J . Morphol., 184:293-309.
Lombardi, J., and J.P. Wourms 198513 The trophotaenial placenta of a viviparous fish. 111. Protein uptake by trophotaeniae, the embry- onic component. J . Exp. Zool., 236:165-179.
Maunsbach, A.B. 1966a Absorption of P5-labeled homologous albu-
min by rat kidney proximal tubule cells. A study of microperfused single proximal tubules by electron microscopic autoradiography and histochemistry. J . Ultrastruct. Res., 15t197-241.
Maunsbach, A.B. 1966b Absorption of ferritin by rat kidney proximal tubule cells. Electron microscopic observations on the initial up- take phase in cells of microperfused single proximal tubules. J . Ultrastruct. Res., 16:l-12.
Miller, R.R., and J.M. Fitzsimons 1971 Amecu splendens, a new genus and species of goodeid fish from Western Mexico, with remarks on the classification of the Goodeidae. Copeia, 1971r1-13.
Moore, A.T., K.E. Williams, and J.B. Lloyd 1977 The effect of chem- ical treatment of albumin and orosomucoid on rate of clearance from the rat bloodstream and rate of pinocytic capture by rat yolk sac cultured in vitro. Biochem. J., 164:607-616.
Morales, C., and L. Hermo 1986 lntracellular pathways of endocy- tosed transferrin and non-specific tracers in epithelial cells lining the rete testis of the rat. Cell Tissue Res., 245:323-330.
Muhlpfort, H. 1982 The preparation of colloidal gold particles using tannic acid as an additional reducing agent. Experientia (Basel), 38:1127-1128.
Ohkuma, S., and B. Poole 1981 Cytoplasmic vacuolation of mouse peritoneal macrophages and the uptake into lysosomes of weakly basic substances. J. Cell Biol., 9Ot656-664.
Scatchard, G. 1948 The attraction of proteins for small molecules and ions. Ann. N.Y. Acad. Sci., 51t660-672.
Schindler, J.F. 1990 Retrograde trafficking of tracer protein by the inner ovarian epithelium in gravid goodeid teleosts. Anat. Rec., 226t177-186.
Schindler, J.F., and U. de Vries 1987 Protein uptake and transport by trophotaenial absorptive cells in two species of goodeid embryos. J . Exp. Zool., 241t17-29.
Schindler, J.F., and U. de Vries 1988 Maternal-embryonic relation- ships in the goodeid teleost, Xenoophorus cuptiuus. The vacuolar apparatus in trophotaenial absorptive cells and its role in mac- romolecular transport. Cell Tissue Res., 253r115-128.
Schindler, J.F., and U. de Vries 1990 Effects of ammonia, chloroquine, and monensin on the vacuolar apparatus of an absorptive epithe- lium. Cell Tissue Res., 259r283-292.
Schindler, J.F., R. Kujat, and U. de Vries 1988 Maternal-embryonic relationships in the goodeid teleost, Xenoophorus cuptiuus. The internal ovarian epithelium and the embryotrophic liquid. Cell Tissue Res., 254r177-182.
Sullivan, P.C., A.L. Ferris, and B. Storrie 1987 Effects of temperatur.;, pH elevators, and energy production inhibitors on horseradish peroxidase transport through endocytic vesicles. J. Cell. Physiol., 131 58-63,
Wallace, R.A., and K. Selman 1978 Oogenesis in Fundulus heterocli- tus. I. Preliminary observations on oocyte maturation in vivo and in vitro. Dev. Biol., 62t354-369.
Weigel, P.H., and J.A. Oka 1981 Temperature dependence of endocy- tosis mediated by the asialoglycoprotein receptor in isolated rat hepatocytes. Evidence for two potentially rate-limiting steps. J. Biol. Chem., 256:2615-2617.
Williams, K.E., E.M. Kidston, F. Beck, and J.B. Lloyd 1975 Quanti- tative studies of pinocytosis. 11. Kinetics of protein uptake and digestion by rat yolk sac cultured in vitro. J . Cell Biol., 64r123- 134.
Woods, J.W., J. Goodhouse, and M.G. Farquhar 1989 Transferrin re- ceptors and cation-independent mannose-6-phosphate receptors deliver their ligands to two distinct subpopulations of multive- sicular endosomes. Eur. J . Cell Biol., 5Or132-143.
Copyright © 2022 FDOKUMEN




















