
Production of bacteriocins by coagulase-negative staphylococci involved in bovine mastitis
11
Production of bacteriocins by coagulase-negative staphylococci involved in bovine mastitis Janaı ´na dos Santos Nascimento a , Patricia Carlin Fagundes a , Maria Aparecida Vasconcelos de Paiva Brito b , Ka ´tia Regina Netto dos Santos a , Maria do Carmo de Freire Bastos a, * a Instituto de Microbiologia Prof. Paulo de Go ´es, Departamento de Microbiologia Geral, Universidade do Brazil (UFRJ), CCS, Bloco I, sala I-1-059, Cidade Universita ´ria, 21941-590 Rio de Janeiro, RJ, Brazil b EMBRAPAGado de Leite, Juiz de Fora, MG, Brasil Received 8 June 2004; received in revised form 20 September 2004; accepted 10 October 2004 Abstract In the present study, 188 coagulase-negative Staphylococcus (CNS) strains were isolated from bovine mastitis cases from 56 different Brazilian dairy herds, located in the Southeast region of the country, and were tested for antimicrobial substance production. Twelve CNS strains (6.4%) exhibited antagonistic activity against a Corynebacterium fimi indicator strain. Most antimicrobial substances were sensitive to proteolytic enzymes suggesting that they might be bacteriocins (Bac). Amongst the CNS producers, six were identified as S. epidermidis, two as S. simulans, two as S. saprophyticus, one as S. hominis and one as S. arlettae. Plasmid profile analysis of these strains revealed the presence of at least one plasmid. The Bac + strains presented either no or few antibiotic resistance phenotypes. Three strains were shown to produce a bacteriocin either identical or similar to aureocin A70, a bacteriocin previously isolated from an S. aureus strain isolated from food. The remaining Bac + strains produce antimicrobial peptides that seem to be distinct from the best characterised staphylococcal bacteriocins described so far. Some of them were able to inhibit Listeria monocytogenes, an important food-borne pathogen, and several strains of Streptococcus agalactiae associated with bovine mastitis, suggesting a potential use of these bacteriocins either in the prevention or in the treatment of streptococcal mastitis. # 2004 Elsevier B.V. All rights reserved. Keywords: Coagulase-negative Staphylococcus spp.; Bacteriocin; Bovine mastitis; Cattle-bacteria 1. Introduction Bovine mastitis is an inflammation of the mammary glands usually due to a microbial infection that affects milk production and quality, being one of the most significant causes of economic loss to the dairy industry (Soltys and Quinn, 1999; Sordelli et al., 2000; Riffon et al., 2001; Bradley, 2002). Although several bacterial pathogens can cause mastitis, Staphylococcus aureus is the most important causative agent (followed by Streptococcus spp.), and www.elsevier.com/locate/vetmic Veterinary Microbiology 106 (2005) 61–71 * Corresponding author. Fax: +55 21 2560 8344. E-mail address: [email protected] (M.C.F. Bastos). 0378-1135/$ – see front matter # 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.vetmic.2004.10.014
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Production of bacteriocins by coagulase-negative staphylococci involved in bovine mastitis

www.elsevier.com/locate/vetmic
Veterinary Microbiology 106 (2005) 61–71
Production of bacteriocins by coagulase-negative
staphylococci involved in bovine mastitis
Janaına dos Santos Nascimento a, Patricia Carlin Fagundes a,Maria Aparecida Vasconcelos de Paiva Brito b, Katia Regina Netto dos Santos a,
Maria do Carmo de Freire Bastos a,*
a Instituto de Microbiologia Prof. Paulo de Goes, Departamento de Microbiologia Geral, Universidade do Brazil (UFRJ),
CCS, Bloco I, sala I-1-059, Cidade Universitaria, 21941-590 Rio de Janeiro, RJ, Brazilb EMBRAPA Gado de Leite, Juiz de Fora, MG, Brasil
Received 8 June 2004; received in revised form 20 September 2004; accepted 10 October 2004
Abstract
In the present study, 188 coagulase-negative Staphylococcus (CNS) strains were isolated from bovine mastitis cases from 56
different Brazilian dairy herds, located in the Southeast region of the country, and were tested for antimicrobial substance
production. Twelve CNS strains (6.4%) exhibited antagonistic activity against a Corynebacterium fimi indicator strain. Most
antimicrobial substances were sensitive to proteolytic enzymes suggesting that they might be bacteriocins (Bac). Amongst the
CNS producers, six were identified as S. epidermidis, two as S. simulans, two as S. saprophyticus, one as S. hominis and one as S.
arlettae. Plasmid profile analysis of these strains revealed the presence of at least one plasmid. The Bac+ strains presented either
no or few antibiotic resistance phenotypes. Three strains were shown to produce a bacteriocin either identical or similar to
aureocin A70, a bacteriocin previously isolated from an S. aureus strain isolated from food. The remaining Bac+ strains produce
antimicrobial peptides that seem to be distinct from the best characterised staphylococcal bacteriocins described so far. Some of
them were able to inhibit Listeria monocytogenes, an important food-borne pathogen, and several strains of Streptococcus
agalactiae associated with bovine mastitis, suggesting a potential use of these bacteriocins either in the prevention or in the
treatment of streptococcal mastitis.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Coagulase-negative Staphylococcus spp.; Bacteriocin; Bovine mastitis; Cattle-bacteria
1. Introduction
Bovine mastitis is an inflammation of the
mammary glands usually due to a microbial infection
* Corresponding author. Fax: +55 21 2560 8344.
E-mail address: [email protected] (M.C.F. Bastos).
0378-1135/$ – see front matter # 2004 Elsevier B.V. All rights reserved
doi:10.1016/j.vetmic.2004.10.014
that affects milk production and quality, being one of
the most significant causes of economic loss to the
dairy industry (Soltys and Quinn, 1999; Sordelli et al.,
2000; Riffon et al., 2001; Bradley, 2002).
Although several bacterial pathogens can cause
mastitis, Staphylococcus aureus is the most important
causative agent (followed by Streptococcus spp.), and
.

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–7162
once it is established in the mammary glands of the
animal, it is very difficult to eradicate (Soltys and Quinn,
1999; Lammers et al., 2001). Coagulase-negative
staphylococci (CNS) have been considered a minor
pathogen of bovine mastitis. However, many studies
have recently shown the importance of CNS infection of
the bovine mammary glands (Soltys and Quinn, 1999;
Zhang and Maddox, 2000; Younis et al., 2003).
In the efforts to avoid mastitis, some vaccines that
can reduce the severity of this illness were generated.
However, these vaccines still do not control efficiently
the development of mastitis (Leitner et al., 2003). On
the other hand, it has been verified that the
indiscriminate treatment with antibiotics, without
either a technical prescription or identification tests
of the pathogen, can contribute to an increased
resistance of these micro-organisms, making the cure
of mastitis still more difficult (Gruet et al., 2001).
Moreover, it is important to stand out that these
medicines are set free in milk for some days, after their
administration, being able to cause problems for
consumers. Thus, the identification of alternative
methods for controlling this illness is essential. One of
these methods could be the use of bacteriocins.
Bacteriocins (Bac) are antimicrobial peptides or
proteins produced by bacteria with inhibitory activity
against other bacteria. These substances, specially
those produced by Gram-positive bacteria, present a
potential use either in food industries as biopreserva-
tives or in the prevention and treatment of some
infectious diseases, having medical and veterinary
applications (Jack et al., 1995; Oliveira et al., 1998b;
Sahl and Bierbaum, 1998).
Amongst the bacteriocin-producer staphylococcal
species, S. epidermidis is the most studied. Some of its
bacteriocins are very well characterised, such as Pep5,
epidermin, epicidin 280 and epilancin K7, which are
classified as lantibiotics. Pep5 is a 34-amino acid
peptide encoded by the structural gene pepA (Kaletta
et al., 1989). Epidermin is also a well-characterised
lantibiotic produced by S. epidermidis Tu3298. Its
structural gene, epiA, codes for a 22-amino acid
peptide (Augustin et al., 1992). Epicidin 280 is also a
30-amino acid lantibiotic, produced by S. epidermidis
BN280, which has 75% of homology with Pep5
(Heidrich et al., 1998). Epilancin K7, a 31-amino acid
lantibiotic produced by S. epidermidis K7, is encoded
by the gene elkA (van de Kamp et al., 1995).
Our laboratory has been investigating bacteriocin
production by Staphylococcus strains, which is
generally associated to plasmids. Amongst the
bacteriocins produced by S. aureus and studied by
our group, aureocins A70 and A53 are the best
characterised. Aureocin A70 is a multi-peptide, non-
lantibiotic bacteriocin produced by S. aureus A70
which is encoded by an 8.0 kb-plasmid named pRJ6
(Giambiagi-deMarval et al., 1990). Aureocin A70 was
the first bacteriocin described which is composed of
four related small cationic peptides, which are
encoded by the aurABCD operon (Netz et al.,
2001). Aureocin A53 is a 51-amino acid peptide
produced by S. aureus A53. It is encoded on a 10.4 kb
plasmid, pRJ9 (Netz et al., 2002).
As well as other antimicrobial substances, bacter-
iocins produced by staphylococci (staphylococcins)
could be employed, in their purified form, on the
control of bovine mastitis. Therefore, in the present
study, in an attempt to detect new bacteriocins with
potential medical and veterinary applications, we
extended our studies on bacteriocin production to CNS
strains, which were associated with bovine mastitis in
different Brazilian dairy herds.
2. Materials and methods
2.1. Bacterial strains and culture conditions
One hundred and eighty-eight coagulase-negative
Staphylococcus strains involved in bovine mastitis
were isolated from 56 different Brazilian dairy herds
located in the Southeast region of the country, and
used in the screening for bacteriocin production.
Seventy-four Streptococcus agalactiae strains were
also isolated from bovine mastitis cases and used as
indicators. These strains were isolated according to
procedures recommended by Harmon et al. (1990).
S. aureus and S. epidermidis strains from previous
studies (Table 1) were used either as producers or as
indicators in the inhibition assays. Staphylococcus
strains were grown in either TSB (Difco) or BHI
(Difco), at 37 8C for 18 h. TSB was used to grow the
strains for DNA isolation and BHI was used in all
bacteriocin assays. The bacteria used as indicators
were grown in BHI medium, except for lactic acid
bacteria, that were cultivated in MRS medium (BBL)

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–71 63
Table 1
Staphylococcus strains previously described and used in this study
Strain Relevant features Reference/source
Staphylococcus aureus
A53 Bac+ (aureocin A53), Imm+, plasmid: pRJ9 (10.4 kb) Giambiagi-deMarval et al. (1990)
A70 Bac+ (aureocin A70), Imm+, plasmid: pRJ6 (8.0 kb) Giambiagi-deMarval et al. (1990)
A70 Bac� Strain A70 cured of pRJ6 Giambiagi-deMarval et al. (1990)
Staphylococcus epidermidis
Tu 3298 Bac+ (epidermin) Augustin et al. (1992)
5 Bac+ (Pep5) Kaletta et al. (1989)
5 (cured) Bac� Pep5� Bierbaum et al. (1994)
BN280 Bac+ (epicidin 280) Heidrich et al. (1998)
K7 Bac+ (epilancin K7) van de Kamp et al. (1995)
Bac, bacteriocin; Imm, immunity.
and incubated at 28 8C for 48 h under microaerophilic
atmosphere. Bacteria were stored in their appropriate
culture medium with 40% glycerol (w/v) at �20 8Cuntil needed. When necessary, the media were
supplemented with agar at 1.5% (w/v) or 0.6% (w/v).
2.2. Assay for antimicrobial substance production
This assay was done as described previously by
Giambiagi-deMarval et al. (1990). C. fimi NCTC 7547
was used as the indicator strain for production of
antimicrobial substances.
2.3. Identification of the strains
Only the CNS strains that exhibited antimicrobial
substance production were identified, using conven-
tional biochemical tests (Kloos and Schleifer, 1986)
or, when necessary, using a commercial kit for
identification (API Staph, BioMerieux). S. agalactiae
strains were identified on the basis of Gram staining,
presence of hemolysis, negative results on tests for the
presence of catalase and esculin hydrolysis, and
positive results on tests for CAMP and hydrolysis of
sodium hypurate.
2.4. Effects of proteolytic enzymes and of 0.2N
NaOH on AMS activity
The effects of trypsin (Sigma), proteinase K
(Boehinger Mannheim), protease XXIII (Sigma)
and 0.2N NaOH on AMS activity were determined
by the method described by Giambiagi-deMarval et al.
(1990). Lack of inhibition zones when C. fimi
NCTC7547 was used as the indicator strain indicated
that the antimicrobial compound had either a
proteinaceous or an acidic nature.
2.5. Antibiotic resistance
Resistance patterns were determined by disc
diffusion on Mueller-Hinton agar (Oxoid), according
to the National Committee for Clinical Laboratory
Standards (NCCLS, 2002). The following antibiotics
(Sensifar) were used: ampicillin (10 mg), cephalothin
(30 mg), ciprofloxacin (5 mg), clindamycin (2 mg),
chloramphenicol (30 mg), erythromycin (15 mg),
gentamicin (10 mg), imipenen (10 mg), mupirocin
(5 mg), oxacillin (1 mg), penicillin (10 U), rifampin
(5 mg), tetracycline (30 mg) and vancomycin (30 mg).
The diameters of the inhibition zones were interpreted
according to the NCCLS guidelines after 24 h of
incubation at 37 8C. Strains which showed resistance
to b-lactamic drugs were also tested for the presence
of the methicillin-resistance gene (mecA) by PCR.
2.6. Isolation of plasmid DNA
Whole-cell lysates were prepared as described by
Giambiagi-deMarval et al. (1990).
2.7. DNA–DNA hybridisation assays
Southern blots and hybridisations were performed
as described by Bastos and Murphy (1988). The 5.2-kb
HindIII-A fragment of pRJ6 (encompassing all genes

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–7164
required for aureocin A70 production) was used as a
probe. 32P-labelled probes were prepared by the
Random-Primers DNA Labelling System (Invitrogen)
following the manufacturer’s recommendations.
2.8. PCR amplification
Total DNA was extracted by boiling, as described
previously by Nunes et al. (1999). The primers used
for amplification of the aurABCD operon and the
genes aucA, eciA and elkA (the structural genes of
aureocin A70, aureocin A53, epicidin 280 and
epilancin K7, respectively) as well as for amplification
of the methicillin-resistance gene (mecA) and genes
orfA and orfB (presumably involved in the immunity
to aureocin A70) are listed in Table 2. Each PCR
reaction contained: 1� PCR reaction buffer (Invitro-
gen), 2.5 mM concentration of each deoxyribonucleo-
side triphosphate, 2.5 U of Taq-Polymerase (Invi-
trogen) and 50 pmol of each primer. The amplification
was done in a Programmable Thermal Controller
(PTC-100TM, MJ Research, USA) and the cycles were
performed as described by Nunes et al. (1999). A 100-
bp DNA ladder (Invitrogen) was used as the molecular
size marker.
2.9. Determination of the inhibitory spectrum
of each AMS+ strain
To determine the inhibitory spectrum of each
AMS, several strains from different species of Gram-
positive bacteria were tested as indicators. The
Table 2
Primers used in this work
Gene(s) to be amplified Relevant features Primers
aurABCD Aureocin A70 structural genes Forward: P
reverse: P
aucA Aureocin A53 structural gene Forward: A
reverse: A
orfA and orfB Immunity genes to aureocin A70 Forward: R
reverse: R
elkA Epilancin K7 structural gene Forward: K
reverse: K
eciA Epicidin 280 structural gene Forward: E
reverse: E
mecA Methicillin-resistance gene Forward: M
reverse: M
inhibitory action of the AMS produced by the
CNS strains against S. agalactiae involved in
bovine mastitis was also tested using 74 indicator
strains.
2.10. Agarose gel electrophoresis
Agarose gel electrophoreses were performed as
described by Sambrook et al. (1989). Products
amplified by PCR were analysed in 1.4% (w/v)
agarose gels and the plasmid profiles were determined
in 0.7% (w/v) agarose gels. The gels were stained in an
aqueous solution containing ethidium bromide
(0.5 mg/ml) and visualised on a UV transiluminator.
3. Results
3.1. Detection and identification of the AMS+ strains
One hundred and eighty-eight coagulase-negative
Staphylococcus (CNS) strains were isolated from
bovine mastitis cases from 56 different Brazilian dairy
herds, located in the Southeast region of the country,
and were tested for AMS production. Twelve CNS
strains (6.4%), belonging to six different herds,
exhibited antimicrobial substance production against
C. fimi (inhibition zones between 17 and 38 mm).
Among the CNS producers, six were identified as S.
epidermidis, two as S. simulans, two as S. saprophy-
ticus, one as S. hominis and one as S. arlettae
(Table 3).
Size (bp)
of the fragment
to be amplified
4B, 50-CCTTATAACTTCGAATGCT-30;5, 50-AATTATTAACAAGAGAAA-30
525
UC1, 50-GAAGTTGTGAAAACTATTA-30;UC2, 50-CATAAAACAAAGAGCCAAAGT-30
322
J61, 50-GACGAGGGTATTGCATA-30;J62, 50-CTAAGCATTCGATAAGG-30
722
71, 50-ATGAATAACTCATTATTC-30;72, 50-ATGGAA AACAAAAAAG-30
171
pin1, 50-CAGGAG GGATATATTATGG-30;pin2, 50-CAATCACTACTATTGACAATCAC-30
165
RS1, 50-TAGAAATGA CTG AACGTCCG-30;RS2, 50-TTGCGATCAATGTTACCG TAG-30
154

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–71 65
Table 3
Characteristics of the bacteriocin-producer CNS strains
Strains Identification Sensitivity of the Bac to proteolytic enzymes Resistance profile Plasmid formsa
Protease XXIII Proteinase K Trypsin
A70b S. aureus S S S � 8.0 (pRJ6)
494 S. epidermidis R S S Emc >25; 15; 6.7; 5.5; 3.7; 3.2; 2.2
2166 S. epidermidis R S S � 12; 6.2
2167 S. epidermidis S R R Emc 12; 6.2
2457 S. simulans S R S Emc >27
3024 S. epidermidis S S S Ap, Ce, Gm, Ox, Pc, Tc 15; 8.1; 4.5
3154 S. saprophyticus R S S Emc 15; 8
3299 S. simulans R S S � >25; 3.5
3414 S. epidermidis R S S � >25
3419 S. hominis R R R � 4
3528 S. arlettae S S S Ap, Pc 15; 8
3576 S. saprophyticus S S S Ap, Pc 15; 8
3577 S. epidermidis S R R � >25 (2)d; 2.5; 1.5
R, resistant; S, sensitive; (�) sensitive to all the antibiotics tested; Ap, ampicillin; Ce, cephalothin; Em, erythromycin; Gm, gentamicin; Ox,
oxacillin; Pc, penicillin; Tc, tetracycline.a Size in kb.b Strain A70 was included in these experiments as a positive control.c Intermediate resistance.d The number between parentheses indicates the presence of two plasmid bands with size larger than 25 kb.
3.2. Effects of proteolytic enzymes on AMS activity
The results are also presented in Table 3. The AMS
were resistant to 0.2N NaOH, discarding the
possibility that the inhibition exhibited was due to
organic acids produced by the producer strain during
its metabolism. Except for the AMS produced by the
strain 3419, the remaining AMS were sensitive to at
least one proteolytic enzyme tested, indicating that
these AMS present a biological active proteinaceus
component in their structure, the main characteristic
of a typical bacteriocin. Therefore, from hereafter, the
AMS+ strains will be considered Bac+. Bacteriocins
produced by strains 3154, 3528 and 3576, as well as
aureocin A70, showed a high sensitivity to proteolytic
enzymes, being sensitive to all enzymes tested.
3.3. Antibiotic resistance profile
The Bac+ strains did not exhibit a great number of
resistance markers, except for strain 3024, that showed
resistance to six antibiotics (Table 3). The remaining
strains were resistant to either two (ampicillin and
penicillin) or none of the antibiotics tested. Strains
494, 2167, 2457 and 3154 presented intermediate
resistance to erythromycin. Strains that showed
resistance to one or more b-lactamic drugs were also
tested for the presence of the mecA gene, which
encodes methicillin resistance. Among the strains
tested, only strain 3024 showed amplification of the
mecA gene (data not shown).
3.4. Plasmid profiles
All strains showed at least one plasmid form
(Table 3). Strains 3154 (S. saprophyticus), 3528 (S.
arlettae) and 3576 (S. saprophyticus) exhibited an
identical plasmid profile, and, interestingly, they carry
a plasmid with a size similar to that of pRJ6 (8.0 kb), a
bacteriocinogenic plasmid previously characterised
by our group and that encodes aureocin A70. These
three strains were isolated from the same herd. Strains
2166 and 2167 (both S. epidermidis and isolated from
the same herd), also presented an identical plasmid
profile.
3.5. DNA–DNA hybridisation assays
Since strains 3154, 3528 and 3576 carry a plasmid
with a size similar to that of pRJ6, the homology
between these plasmids and pRJ6 was investigated. A
strong signal was detected to the 8.0-kb plasmids
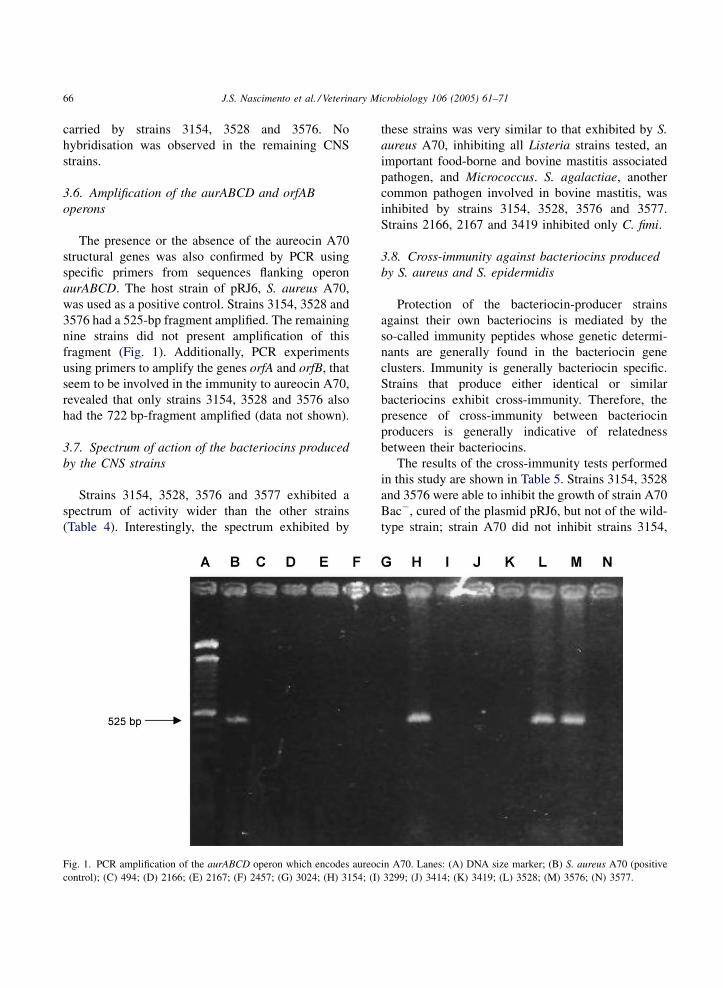
J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–7166
carried by strains 3154, 3528 and 3576. No
hybridisation was observed in the remaining CNS
strains.
3.6. Amplification of the aurABCD and orfAB
operons
The presence or the absence of the aureocin A70
structural genes was also confirmed by PCR using
specific primers from sequences flanking operon
aurABCD. The host strain of pRJ6, S. aureus A70,
was used as a positive control. Strains 3154, 3528 and
3576 had a 525-bp fragment amplified. The remaining
nine strains did not present amplification of this
fragment (Fig. 1). Additionally, PCR experiments
using primers to amplify the genes orfA and orfB, that
seem to be involved in the immunity to aureocin A70,
revealed that only strains 3154, 3528 and 3576 also
had the 722 bp-fragment amplified (data not shown).
3.7. Spectrum of action of the bacteriocins produced
by the CNS strains
Strains 3154, 3528, 3576 and 3577 exhibited a
spectrum of activity wider than the other strains
(Table 4). Interestingly, the spectrum exhibited by
Fig. 1. PCR amplification of the aurABCD operon which encodes aureoc
control); (C) 494; (D) 2166; (E) 2167; (F) 2457; (G) 3024; (H) 3154; (I)
these strains was very similar to that exhibited by S.
aureus A70, inhibiting all Listeria strains tested, an
important food-borne and bovine mastitis associated
pathogen, and Micrococcus. S. agalactiae, another
common pathogen involved in bovine mastitis, was
inhibited by strains 3154, 3528, 3576 and 3577.
Strains 2166, 2167 and 3419 inhibited only C. fimi.
3.8. Cross-immunity against bacteriocins produced
by S. aureus and S. epidermidis
Protection of the bacteriocin-producer strains
against their own bacteriocins is mediated by the
so-called immunity peptides whose genetic determi-
nants are generally found in the bacteriocin gene
clusters. Immunity is generally bacteriocin specific.
Strains that produce either identical or similar
bacteriocins exhibit cross-immunity. Therefore, the
presence of cross-immunity between bacteriocin
producers is generally indicative of relatedness
between their bacteriocins.
The results of the cross-immunity tests performed
in this study are shown in Table 5. Strains 3154, 3528
and 3576 were able to inhibit the growth of strain A70
Bac�, cured of the plasmid pRJ6, but not of the wild-
type strain; strain A70 did not inhibit strains 3154,
in A70. Lanes: (A) DNA size marker; (B) S. aureus A70 (positive
3299; (J) 3414; (K) 3419; (L) 3528; (M) 3576; (N) 3577.

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–71 67
Table 4
Spectrum of activity of the bacteriocins produced by coagulase-negative Staphylococcus strains
Indicator strains Producer strains
A70 494 2166 2167 2457 3024 3154 3299 3414 3419 3528 3576 3577
Bacillus megaterium F4 + � � � � � � � � � � � �Corynebacterium fimi NCTC 7547 + + + + + + + + + + + + +
Enterococcus faecium E86 NT � � � � � � � � � � � +
Lactobacillus casei t � � � � � � � � � � � �Lactobacillus casei ATCC 393 t � � � � � � � � � � � �Lactococcus lactis ATCC 11454 t � � � � � � � � � � � �Leuconostoc lactis ATCC 19256 t � � � � � � � � � � � �Listeria monocytogenes 11LM + � � � � � + � � � + + +
Listeria monocytogenes L1/2A + + � � � � + � � � + + +
Listeria monocytogenes 7848 + � � � � � + � � � + + +
Listeria innocua 397 + � � � � � + � � � + + +
Micrococcus spp. + � � � + � + + � � + + +
Paenibacillus polymyxa SCE-2 + � � � � � � � � � � � �Pediococcus pentosaceus ATCC 43200 � � � � � + � � + � � + �Streptococccus pyogenes � � � � � � � + � � � � �Streptococcus agalactie � � � � � � + � � � + + +
(+) Inhibition; (�) no inhibition; NT, not tested; t, turbid zone of inhibition. Only the indicator strains that were inhibited by at least one of the
producer strains are shown in this table. Enterococcus faecalis FA2-2, E. faecalis OG1X, Lactobacillus brevis ATCC 14869 and Leuconostoc
mesenteroides ATCC 8293 were also tested, however, they were not inhibited by any of the producer strains.
3528 and 3576 suggesting the presence of cross-
immunity among the bacteriocins produced by these
four strains. Strain 3577 was also able to inhibit strain
A70 Bac� but not A70. However, strain A70 inhibited
strain 3577, suggesting that the bacteriocins produced
Table 5
Immunity/resistance to bacteriocins among bacteriocinogenic CNS and S
Indicator strains Producer strains
494 2166 2167 2457 3024
S. aureus A70 � � � � �S. aureus A70 Bac� � � � � �S. aureus A53 � � � � �S. epidermidis Tu 3298 � � � � �S. epidermidis 5 � � � � �S. epidermidis BN280 � � � � �S. epidermidis K7 � � � � �
Producer strains Indicator strains
494 2166 2167 2457
S. aureus A70 (aureocin A70) � � + +
S. aureus A53 (aureocin A53) � � � +
S. epidermidis Tu 3298 (epidermin) + + + +
S. epidermidis 5 (Pep5) + + + +
S. epidermidis BN280 (epicidin 280) � � � �S. epidermidis K7 (epilancin K7) � � � �(+) Inhibition; (�) no inhibition.
by both strains are different. Strain 3299 inhibited only
strain A53, indicating that its bacteriocin is distinct
from aureocin A53. The producer strains of Pep5 and
epidermin were able to inhibit all the 12 CNS strains
associated with bovine mastitis, suggesting that none
. aureus strains
3154 3299 3414 3419 3528 3576 3577
� � � � � � �+ � � � + + +
+ + � � + + �� � � � � � �� � � � � � �� � � � � � �� � � � � � �
3024 3154 3299 3414 3419 3528 3576 3577
� � + � � � � +
� + � � � + + �+ + + + + + + +
+ + + + + + + +
� � � � � � � �� � � � � � � �

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–7168
Fig. 2. Inhibition of Streptococcus agalactiae strains involved in bovine mastitis by Bac+ coagulase-negative Staphylococcus strains. Seventy-
four S. agalactiae strains involved in bovine mastitis were tested as indicators. (&) Number of inhibited strains and (&) percentage of inhibited
strains.
SCN bacteriocin is related to Pep5 and epidermin. The
producers of epicidin 280 and epilancin K7, however,
were neither able to inhibit nor to be inhibited by any
of the CNS strains. In such cases, the absence of
inhibition could result from either bacteriocin
immunity or resistance (i.e, inability of the bacteriocin
to act on the cells).
3.9. Amplification of the structural genes for
aureocin A53, epilancin K7 and epicidin 280
To discard the possibility that the CNS bacteriocins
are related to aureocin A53, epilancin K7 and epicidin
280, PCR experiments for the amplification of the
structural genes of these staphylococcins were
performed and none of the 12 CNS strains involved
in bovine mastitis showed amplification of the
expected fragments (data not shown).
3.10. Inhibition of Streptococcus agalactiae strains
involved in bovine mastitis
Most of the S. agalactiae strains used as indicators
(78.4%) were inhibited by strain 3299 (Fig. 2). Strains
2166, 2167, 3154, 3419 and 3577 were able to inhibit
at least 40% of the indicators strains. The percentage
of inhibition observed with the remaining strains lied
between 10 and 38%.
4. Discussion
Previous studies have shown the production of
AMS by S. aureus strains from different sources.
Giambiagi-deMarval et al. (1990) found 13 (9.5%)
AMS producers amongst 137 strains isolated from
foods and not hospitalised patients. From 1990 to
1992, 163 S. aureus strains were isolated from
different clinical specimens of human origin, in four
hospitals from Rio de Janeiro. Fifty-three strains
(32.5%) were shown to produce AMS, but only three
(1.8%) presented a high antagonistic activity against
C. fimi (Gamon et al., 1999). Oliveira et al. (1998a),
studying 46 S. aureus strains isolated from apparently
healthy cattle, identified only four (8.7%) bacteriocin-
producer strains. Recently, Nascimento et al. (2002)
studied 50 strains of S. aureus isolated from mastitic
cows and verified that 24% of these strains showed a
high inhibitory activity against C. fimi. Therefore, a
higher incidence of AMS+ S. aureus strains was found
amongst the isolates involved in bovine mastitis. In the
present work, however, amongst 188 CNS, we found
only 12 (6.4%) producer strains. AMS produced by
most strains were sensitive to proteolytic enzymes
suggesting they might be bacteriocins. The only
exception was strain 3419. However, bacteriocins
resistant to proteolytic digestion has already been
reported in the literature (Netz et al., 2002). Although
bacteriocin production is not a characteristic found in
most Staphylococcus strains, it may confer a
competitive advantage to the producer strains for
nutrition in the surroundings, helping in the occupa-
tion of determined ecological niches.
In this study, most of the Bac+ CNS strains were
identified as S. epidermidis. The remaining strains
were S. simulans, S. saprophyticus, S. hominis and S.
arlettae. Bacteriocin production has already been
described in S. epidermidis (Sahl and Bierbaum,

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–71 69
1998). However, to our knowledge, this is the first
report, which describes bacteriocin production in
these four latter staphylococcal species.
The resistance profile of the 12 CNS strains to 14
different antibiotics was also evaluated. In general, the
strains did not present a great number of resistance
phenotypes. These results contrast with those
described for clinical isolates of CNS which generally
carry multiple drug-resistance determinants (Aires de
Sousa et al., 1998; Kohner et al., 1999; Santos et al.,
1999). Only strain 3024 was shown to be resistant to
six of the antibiotics tested, mainly b-lactamic drugs.
The mecA gene, detected in this strain by PCR, is
probably involved in resistance to the latter drugs.
In Gram-positive bacteria, the genetic determinants
involved in the Bac production have been found either
on plasmids or on the bacterial chromosome (Jack
et al., 1995). Amongst the 12 Bac+ CNS strains
associated with bovine mastitis studied in this work,
all strains presented at least one plasmid form. Except
for S. hominis 3419, the genetic determinants
encoding the bacteriocins could be either on the
chromosome or on plasmids carried by them. In strain
3419, the bacteriocin genetic determinant is probably
chromosomally encoded since the only plasmid found
in this strain is too small to carry all genes generally
required for bacteriocin expression (Ennahar et al.,
2000; McAuliffe et al., 2001). Three strains (3154,
3528, 3576) were shown to possess a plasmid with a
size similar to that of pRJ6 (8.0 kb), an S. aureus
plasmid which encodes aureocin A70. PCR experi-
ments were then carried out to test if the CNS strains
presented the structural genes involved in aureocin
A70 production. The amplification of the expected
fragment of 525 bp was observed only in strains 3154,
3528 and 3576. These results indicate that the DNA of
these strains possesses similar sequences to that found
in the operon aurABCD. However, despite the
amplification of the 525 bp fragment, it is not possible
to assure that the amplified sequence is identical to
that of pRJ6. This could be confirmed by DGGE
(denaturing gradient gel electrophoresis) experiments,
whose technique allows the detection of differences of
even a single base-pair in DNA fragments with the
same size (Muyzer et al., 1998). These experiments
are currently in progress.
To confirm that the 8.0 kb plasmids found in strains
3154, 3528 and 3576 are involved in bacteriocin
production, experiments of plasmid cure or transfer
are required. However, hybridisation experiments with
the HindIII-A fragment of pRJ6, the amplification of
the bacteriocin operon as well as the amplification of
the genes orfA and orfB, probably involved in the
immunity to aureocin A70, are strong evidences that
these plasmids are responsible for bacteriocin produc-
tion, being either related or identical to pRJ6. For this
reason, a more detailed characterisation of these
plasmids was not performed. Previous studies
(Giambiagi-deMarval et al., 1990; Oliveira et al.,
1998a; Gamon et al., 1999; Nascimento et al., 2002)
have shown that bacteriocinogenic plasmids of 8.0 kb
are spread among the S. aureus population. However,
this is the first report on the presence of this plasmid
also among coagulase-negative staphylococci. Such
results suggest that there is a certain level of transfer of
plasmids related to pRJ6 among different species of
this genus. Taking together these results support the
conclusion that the bacteriocins produced by the CNS
strains 3154, 3528 and 3576 are either identical or
similar to aureocin A70.
In relation to the remaining strains, the immunity/
resistance and PCR experiments showed that they
produce bacteriocins which seem to be different from
the most studied staphylococcins. However, experi-
ments aiming the characterisation of these bacter-
iocins were not performed yet.
The bacteriocins produced by the CNS strains
presented a narrow spectrum of activity against Gram-
positive bacteria when compared to the bacteriocins
produced by the S. aureus strains previously studied
by our group (Giambiagi-deMarval et al., 1990;
Oliveira et al., 1998a; Gamon et al., 1999). Only four
strains (3154, 3528, 3576 and 3577) were able to
inhibit L. monocytogenes, an important food-borne
pathogen. However, when the ability of these strains to
inhibit S. agalactiae strains involved in bovine
mastitis was investigated, five CNS strains inhibited
at least 50% of the indicator strains. These results
suggest that the bacteriocins produced by some of
these CNS strains, especially by S. simulans 3299,
may be developed into useful antimicrobial drug for
either treatment or prevention of bovine mastitis
caused by Streptococcus, the second more important
pathogen associated to this disease. However, applied
studies must be done to confirm their effectiveness ‘‘in
vivo’’. These studies must be preceded by bacteriocin

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–7170
purification and analysis of its biochemical structure.
Since the bacteriocin produced by S. simulans 3299
seems to be the best candidate for prevention of
streptococcal mastitis, the experiments aiming to its
purification and characterisation are currently in
progress.
Acknowledgements
We thank Dr. Hans-Georg Sahl for sending us the S.
epidermidis strains, which produce the bacteriocins
Pep5, epidermin, epilancin K7 and epicidin 280. This
work was supported by grants from CNPq, FAPERJ
and PRONEX to M.C.F.B.
References
Aires de Sousa, M., Sanches, I.S., Ferro, M.L., Vaz, M.J., Saraiva, Z.,
Tendeiro, T., Serra, J., Lencastre, H., 1998. Intercontinental spread
of a multidrug-resistant methicillin-resistant Staphylococcus aur-
eus (MRSA) clone. J. Clin. Microbiol. 36, 2590–2596.
Augustin, J., Rosenstein, R., Wieland, B., Schneider, U., Schnell, N.,
Engelke, G., Entian, K.-D., Gotz, F., 1992. Genetic analysis of
epidermin biosynthesis genes and epidermin-negative mutants of
Staphylococccus epidermidis. Eur. J. Biochem. 204, 1149–1154.
Bastos, M.C.F., Murphy, E., 1988. Transposon Tn554 encodes three
products required for transposition. EMBO J. 7, 2935–2941.
Bierbaum, G., Reis, M., Szekat, C., Sahl, H.-G., 1994. Construction
of an expression system for mutagenesis of the lantibiotic Pep5.
Appl. Environ. Microbiol. 60, 2876–2883.
Bradley, A.J., 2002. Bovine mastitis: an evolving disease. Vet. J.
164, 116–128.
Ennahar, S., Sashihara, T., Sonomoto, K., Ishizaki, A., 2000. Class
IIa bacteriocins: biosynthesis, structure, and activity. FEMS
Microbiol. Rev. 24, 85–106.
Gamon, M.R., Moreira, E.C., Oliveira, S.S., Teixeira, L.M., Bastos,
M.C.F., 1999. Characterization of a novel bacteriocin-encoding
plasmid found in clinical isolates of Staphylococcus aureus.
Antonie van Leeuwenhoek 75, 233–243.
Giambiagi-deMarval, M., Mafra, M.A., Penido, E.G.C., Bastos,
M.C.F., 1990. Distinct groups of plasmids correlated with
bacteriocin production in Staphylococcus aureus. J. Gen. Micro-
biol. 136, 1591–1599.
Gruet, P., Maincent, P., Berthelot, X., Kaltsatos, V., 2001. Bovine
mastitis and intramammary drug delivery: review and perspec-
tives. Adv. Drug Deliv. Rev. 50, 245–259.
Harmon, R.J., Eberhart, R.J., Jasper, D.E., et al., 1990. Micro-
biological Procedures for the Diagnosis of Bovine Udder Infec-
tion. National Mastitis Council, Arlington, 34 pp.
Heidrich, C., Pag, U., Josten, M., Metzger, J., Jack, R.W., Bierbaum,
G., Jung, G., Sahl, H.-G., 1998. Isolation, characterization, and
heterologous expression of the novel lantibiotic epicidin 280 and
analysis of its biosynthetic gene cluster. Appl. Environ. Micro-
biol. 64, 3140–3146.
Jack, R.W., Tagg, J.R., Ray, B., 1995. Bacteriocins of Gram-positive
bacteria. Microbiol. Rev. 59, 171–200.
van de Kamp, M., van den Hooven, H.W., Konings, R.N., Bierbaum,
G.A., Sahl, H.-G., Kuipers, O.P., Siezen, R.J., de Vos, W.M.,
Hilbers, C.W., van de Ven, F.J., 1995. Elucidation of the primary
structure of the lantibiotic epilancin K7 from Staphylococccus
epidermidis K7 Cloning and characterisation of the epilancin-
K7-encoding gene and NMR analysis of mature epilancin K7.
Eur. J. Biochem. 230, 587–600.
Kaletta, C., Entian, K.-D., Kellner, R., Jung, G., Reis, M., Sahl, H.-
G., 1989. Pep5, a new lantibiotic: structural gene isolation and
prepeptide sequence. Arch. Microbiol. 152, 16–19.
Kloos, W.E., Schleifer, K.H., 1986. Genus Staphylococcus. In:
Sneath, P.H.A., Mair, N.S., Sharpe, M.E., Holt, J.G. (Eds.),
Bergey’s Manual of Systematic Bacteriology, vol. 2. Williams
& Wilkins, Baltimore, MD, pp. 1013–1035.
Kohner, P., Uhl, J., Kolbert, C., Persing, D., Cockerill, F., 1999.
Comparison of susceptibility testing methods with mecA gene
analysis for determining oxacillin (methicillin) resistance in
clinical isolates of Staphylococcus aureus and coagulase-nega-
tive Staphylococcus spp. J. Clin. Microbiol. 37, 2952–2961.
Lammers, A., van Vorstenbosch, C.J., Erkens, H.F., Smith, H.E.,
2001. The major bovine mastitis pathogens have different cell
tropisms in cultures of bovine mammary gland cells. Vet.
Microbiol. 80, 255–265.
Leitner, G., Lubashevsky, E., Trainin, Z., 2003. Staphylococcus
aureus vaccine against mastitis in dairy cows, composition
and evaluation of its immunogenicity in a mouse model. Vet.
Immunol. Immunopathol. 93, 159–167.
McAuliffe, O., Ross, R.P., Hill, C., 2001. Lantibiotics: structure,
biosynthesis and mode of action. FEMS Microbiol. Rev. 25,
285–308.
Muyzer, G., Brinkhoff, T., Nubel, U., Santegoeds, C., Schafer, H.,
Wawer, C., 1998. Denaturing gradient gel electrophoresis
(DGGE) in microbial ecology. Mol. Microbiol. Ecol. 3, 1–27.
Nascimento, J.S., Santos, K.R.N., Gentilini, E., Sordelli, D., Bastos,
M.C.F., 2002. Phenotypic and genetic characterisation of bac-
teriocin-producing strains of Staphylococcus aureus involved in
bovine mastitis. Vet. Microbiol. 95, 133–144.
National Committee for Clinical Laboratory Standards (NCCLS),
2002. Performance Standards for Antimicrobial Sensitivity
Testing. NCCLS, Wayne, PA.
Netz, D.J.A., Sahl, H.-G., Marcolino, R., Nascimento, J.S., Oliveira,
S.S., Soares, M.B., Bastos, M.C.F., 2001. Molecular character-
isation of aureocin A70, a multi-peptide bacteriocin from Sta-
phylococcus aureus. J. Mol. Biol. 311, 939–949.
Netz, D.J.A., Pohl, R., Beck-Sickinger, A.G., Selmer, T., Pierik,
A.J., Bastos, M.C.F., Sahl, H.-G., 2002. Biochemical character-
isation and genetic analysis of aureocin A53, a new, atypical
bacteriocin from Staphylococcus aureus. J. Mol. Biol. 319, 745–
756.
Nunes, E.L.C., Santos, K.R.N., Mondino, P.J.J., Bastos, M.C.F.,
Giambiagi-deMarval, M., 1999. Detection of ileS-2 gene encod-
ing mupirocin resistance in methicillin-resistant Staphylococcus

J.S. Nascimento et al. / Veterinary Microbiology 106 (2005) 61–71 71
aureus by multiplex PCR. Diagn. Microbiol. Infect. Dis. 34, 77–
81.
Oliveira, S.S., Povoa, D.C., Nascimento, J.S., Pereira, M.S.V.,
Siqueira Jr., J.P., Bastos, M.C.F., 1998a. Antimicrobial sub-
stances produced by Staphylococcus aureus strains isolated from
cattle in Brazil. Lett. Appl. Microbiol. 27, 229–234.
Oliveira, S.S., Abrantes, J., Cardoso, M., Sordelli, D., Bastos,
M.C.F., 1998b. Staphylococcal strains involved in bovine mas-
titis are inhibited by Staphylococcus aureus antimicrobial pep-
tides. Lett. Appl. Microbiol. 27, 287–291.
Riffon, R., Sayasith, K., Khalil, H., Dubreuil, P., Drolet, M., Lagace,
J., 2001. Development of a rapid and sensitive test for identifica-
tion of major pathogens in bovine mastitis by PCR. J. Clin.
Microbiol. 39, 2584–2589.
Sahl, H.-G., Bierbaum, G., 1998. Lantibiotics: biosynthesis and
biological activities of uniquely modified peptides from Gram-
positive bacteria. Annu. Rev. Microbiol. 52, 41–79.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning:
A Laboratory Manual. Cold Spring Harbor Laboratory Press,
New York.
Santos, K.R.N., Teixeira, L.M., Leal, G.S., Fonseca, L.S., Gontijo
Filho, P.P., 1999. DNA typing of methicillin-resistant Staphy-
lococcus aureus: isolates and factors associated with nosocomial
acquisition in two Brazilian university hospitals. J. Med. Micro-
biol. 48, 17–23.
Soltys, J., Quinn, M.T., 1999. Selective recruitment of T-cell subsets
to the udder during staphylococcal and streptococcal mastitis:
analysis of lymphocyte subsets and adhesion molecule expres-
sion. Infect. Immun. 67, 6293–6302.
Sordelli, D.O., Buzzola, F.R., Gomez, M.I., Steele-Moore, L., Berg,
D., Gentilini, E., Catalano, M., Reitz, A.J., Tollersrud, T., Dena-
miel, G., Jeric, P., Lee, J.C., 2000. Capsule expression by bovine
isolates of Staphylococcus aureus from Argentina: genetic and
epidemiologic analyses. J. Clin. Microbiol. 38, 846–850.
Younis, A., Krifucks, O., Heller, E.D., Samra, Z., Glickman, A.,
Saran, A., Leitner, G., 2003. Staphylococcus aureus exosecre-
tions and bovine mastitis. J. Vet. Med. 50, 1–7.
Zhang, S., Maddox, C.W., 2000. Cytotoxic activity of coagulase-
negative staphylococci in bovine mastitis. Infect. Immun. 68,
1102–1108.




















