
Mastitis Vaccine - Pakistan Research Repository
169
i
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Mastitis Vaccine - Pakistan Research Repository

i

ii
Laboratory and Field Evaluation of a Locally Prepared
Montanide® Adjuvanted Combined Hemorrhagic Septicemia-
Mastitis Vaccine
By
QUDRATULLAH
D.V.M., M. Phil. (CMS) (UAF)
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS
FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN
CLINICAL MEDICINE & SURGERY
DEPARTMENT OF CLINICAL MEDICINE & SURGERY
FACULTY OF VETERINARY SCIENCE
UNIVERSITY OF AGRICULTURE,
FAISALABAD,
PAKISTAN.
(2016)

iii
To,
The Controller of Examinations,
University of Agriculture,
Faisalabad, Pakistan.
We, the Supervisory Committee, certify that the contents and form of thesis submitted
by Mr. Qudratullah (Regd. No. 2003-ag-2087) have been found satisfactory and
recommend that it be processed for evaluation by the External Examiner(s) for the award of
the degree.
SUPERVISORY COMMITTEE:
Chairman:
(Prof. (R) Dr. Ghulam Muhammad)
Member:
(Dr. Muhammad Saqib)
Member:
(Dr. Muhammad Qamar Bilal)

iv
DEDICATED
TO
The sweet memories of my late father Hon. Capt. (R) Muhammad
Tahir and Evere-praying affectionate Mother, sweet daughter (Nehal
Khan) and my caring wife (Tanzila)

v
DECLARATION
I hereby declare that the contents of the thesis “Laboratory and Field Evaluation of a
Locally Prepared Montanide® Adjuvanted Combined Hemorrhagic Septicemia-
Mastitis Vaccine” are product of my own research and no part has been copied from any
published source (except the references, standard mathematical or generic models/ equations/
formulae/ protocols etc.). I further declare that this work has not been submitted for award of
any other diploma/ degree. The University may take action if the information provided is
found inaccurate at any stage. (In case of any default, the scholar will be proceeded against as
per HEC plagiarism policy).
--------------------------------------
QUDRATULLAH (2003-ag-2087)

vi
ACKNOWLEDGEMENT
I would like to take this opportunity to sincerely thank all of the people who have helped me
with my Ph.D. work. I am grateful to Prof. (R) Dr. Ghulam Muhammad Department of
Clinical Medicine and Surgery and Chairman Supervisory Committee for generously
providing me a conducive environment that enabled me to concentrate on, and successfully
complete my Ph.D. studies. His supervision and able guidance in planning study program and
timely completion of work are highly appreciated. I also thank Dr. Muhammad Saqib,
Department of Clinical Medicine and Surgery for moral and technical help in research. I
thank sincerely, Dr. Muhammad Qamar Bilal for his suggestions, constructive criticism,
and interest in my academic pursuits. I am thankful to all colleagues, particularly Dr. Immad
Rashid for their help and moral support during the course of my studies. Dr. Muhammad
Latif, Director Farms and the staff of the Livestock Research Station responded cheerfully
whenever called upon to assist in research and data collection. I would like to appreciate the
help extended by staff members of Department of Clinical Medicine and Surgery, Nuclear
Institute of Agriculture and Biology (NIAB) Faisalabad, Ayub Agriculture Research
Institute, Faisalabad, and owners of private dairy farms Raja Dairy Farm, Rajawala,
Faisalabad; Bilal Rashid Dairy Farm, Aminpur Road, Faisalabad, and Mehar Anayat Dairy
Farm, Green View Colony, Faisalabad. Last but not the least deepest appreciation are
extended to my ever praying affectionate Father (Late) and Mother for their financial help,
moral supports, my Brothers, Sisters, Wife, and all the family members for their affection,
cooperation and amicable attitude. I feel this endeavor justifies their faith in me.
I am thankful to Dr. Siraj Ahmad Kakar, Chairman Board of intermediate and Secondary
Eductaion, Quetta and Dr. Habib-ur-Rehman Veterinary Officer, Quetta for their kind
support and encouragement throughout the research work.
I am also thankful to the Higher Education Commission, Islamabad, for financial assistance
provided under HEC indigenous 5000 Ph.D. fellowship Batch VII. Above all, I have enjoyed
tremendous health, motivation, intellect, wisdom, protection, and facilitating circumstances,
which made it all possible, for which I am forever grateful to Almighty Allah who gave me
the courage to complete this work.
Qudratullah
vii
CONTENTS
CHAPTER TITLE PAGE #
1 INTRODUCTION 1-3
2 REVIEW OF LITERATURE 4-27
3 MATERIALS AND METHODS 28-44
4 RESULTS 45-100
5 DISCUSSION 101-114
6 SUMMARY 115-119
LITERATURE CITED 120-139

viii
TABLE OF CONTENTS
CHAPTER NO. TITLE
Page No
1 INTRODUCTION 1
2 REVIEW OF LITERATURE 4
2.1 Economic impact of animal diseases in developed and
developing countries
4
2.2 Mastitis 4
2.3 General aspects of hemorrhagic septicemia ( HS) 5
2.3.1 Carrier state 6
2.3.2 Transmission 6
2.3.3 Association of HS with other diseases 6
2.3.4 Pathogenesis 7
2.3.5 Rainy seasonal occurrence 7
2.3.6 Clinical signs and post mortem findings 8
2.3.7 Treatment. 8
2.3.8 Morbidity, mortality and case fatality rates 10
2.4 Prevalence and economic significance of HS in Pakistan 11
2.5 Hemorrhagic septicemia vaccines 13
2.6 Importance of transferrin binding protein A as an
Immunogen
18
2.7 Montanide® adjuvanted HS vaccines 19
2.8 Prevalence and economic significance of mastitis in Pakistan 20
2.9 Role of Staphylococcus aureus and Streptococcus agalactiae
in mastitis
21
2.10 Mastitis vaccines 22
2.10.1 Montanide® (Seppic, France) adjuvanted mastitis vaccines 23
2.11 Need for a combined HS–Mastitis vaccine (‘combo’ or
‘cocktail’ HS –mastitis vaccine)
24
2.12 Mastitis in heifers and justification of use of combined HS-
mastitis vaccine
26
3 MATERIALS AND METHODS 28
Phase I Preparation and Evaluation of Montanide® Adjuvanted
Combined Hemorrhagic Septicemia -Mastitis Vaccine in
Laboratory Settings
28
3.1 Collection and culture of blood samples for isolation of HS
pathogen
28
3.1.1 Sources and types of samples for isolation of P. multocida: 28
3.1.2 Isolation and biochemical and molecular confirmation of P.
multocida
29
3.1.3 Synthetic oligonucleotide primers 30
3.1.4 PCR Reaction 30

ix
3.1.5 Pathogenicity (virulence) of 3 candidate vaccinal isolates of P.
multocida
30
3.2 Sources of milk samples, isolation of candidate vaccinal S.
aureus and Str. agalactiae isolates and their partial
characterization.
31
3.2.1 Molecular confirmation of S. aureus 32
3.2.1.1 Detection of nuc gene by polymerase chain reaction (PCR) 32
3.2.1.2 Preparation of genomic DNA from S. aureus 33
3.2.1.3 PCR amplification 33
3.3 Detection of biofilm production trait of S. aureus 34
3.3.1 Qualitative determination by tube method (TM) 34
3.3.2 Qualitative determination by Congo red agar method (CRA) 34
3.3.3 Quantitative determination of biofilm production by
spectrophotometric method
35
3.3.4 Pathogenicity of 4 candidate vaccinal isolates of S. aureus 36
3.3.5 Pathogenicity of 2 candidate vaccinal isolates of Str. agalactiae 36
3.4 Immunogenicity testing of vaccinal isolates of P. multocida,
S. aureus and Str. agalactiae
37
3.5 Antimicrobial susceptibility testing of candidate bacterial
isolates (Antibiogram)
37
3.6 Selection of vaccinal P. multocida isolate 37
3.7 Selection of vaccinal S. aureus isolate 38
3.8 Selection of vaccinal Str. agalactiae isolate 38
3.9 Preparation of Montanide® adjuvanted combined HS-
Mastitis vaccine
39
3.9.1 Preparation of antigen of P. multocida 39
3.9.2 Preparation of antigens of S. aureus and Str. agalactiae 39
3.9.3 Preparation of combined HS-Mastitis vaccine 40
3.10 Evaluation of quality of vaccine 40
3.10.1 Sterility 40
3.10.2 Safety and side effects 40
3.10.3 Challenge- protection assay (Vaccination- challenge study) 40
3.10.4 Stability 41
Phase II Field Evaluation of Montanide® Adjuvanted Combined HS-
Mastitis Vaccine in Target Dairy Species
41
3.11 Animals, their management and treatment-based grouping 41
3.12 Evaluation parameters 43
3.12.1 Hemorrhagic septicemia 43
3.12.1.1 ELISA based antibody titers against P. multocida 43
3.12.1.2 Incidence of HS 43
3.12.1.3 Local and systemic reactions 43

x
3.13 Mastitis 43
3.13.1 ELISA based antibody titers against S. aureus and Str.
agalactiae
43
3.13.2 Incidence and prevalence of S. aureus and Str. agalactiae
mastitis
43
3.13.3 Mastitis screening tests based prevalence and incidence of
mastitis
43
3.13.4 Severity of mastitis in vaccinated and control animals 43
3.13.5 Isolation and identification of pathogens from clinically mastitic
animals
44
3.13.6 Milk somatic cell count 44
3.13.7 Effect of vaccination on milk yield 44
3.13.8 Vaccine efficacy 44
3.13 Statistical Analysis 44
4 RESULTS 45
PHASE-I 45
4.1 Pasteurella multocida isolates recovered from peripheral
blood, femur, and heart blood of HS affected animals
45
4.2 Cultural and biochemical characteristics of suspected
isolates of P. multocida
45
4.2.1 Biochemical reactions of P. multocida isolates 46
4.2.2 PCR based confirmation of P. multocida isolates 47
4.2.3 Antimicrobial susceptibility testing of 3 candidate vaccinal P.
multocida isolates 47
4.2.4 Pathogenicity of the 3 candidate vaccinal isolates of P.
multocida in mice
47
4.3 Staphylococcus aureus isolates recovered from peracute/acute
clinically mastitic quarters (n=37) of dairy cattle and buffaloes
48
4.3.1 Confirmation and characterization of 4 candidate vaccinal
S. aureus isolates
48
4.3.2 Antimicrobial susceptibility of candidate vaccinal S. aureus
isolates
50
4.3.3 Pathogenicity of the 4 candidate vaccinal isolates of S. aureus in
rabbits
50
4.3.4 Biofilm production by candidate vaccinal S. aureus isolates
(n=4)
51
4.3.4.1 Tube method 51
4.3.4.2 Spectrophotometric method 51
4.3.4.3 Congo red agar-method 52

xi
4.3.4.4 PCR based confirmation of S. aureus isolates 52
4.4 Streptococcus agalactiae isolates recovered from
peracute/acute clinical mastitis from dairy cattle and
buffaloes.
53
4.4.1 Confirmation and characterization of candidate vaccinal Str.
agalactiae isolate
53
4.4.2 Seven-digit numerical biotype (biochemical) characteristics of
candidate vaccinal isolates
53
4.4.3 Antimicrobial susceptibility of candidate vaccinal Str.
agalactiae isolates
54
4.4.4 Pathogenicity of the 2 candidate vaccinal isolates of Str.
agalactiae in rabbits
55
4.4.5 Immunogenicity of the 3 candidate vaccinal isolates of P. multocida
in rabbits 55
4.4.6 Immunogenicity of the 4 candidate vaccinal isolates of S.
aureus in rabbits
56
4.4.7 Immunogenicity of the 2 candidate vaccinal isolates of Str.
agalactiae in rabbits
56
4.5 Final selection of vaccinal isolates from amongst the
candidate vaccinal isolates of P. multocida, S. aureus and Str.
agalactiae for vaccine production
57
4.6 Evaluation of quality of vaccine 59
4.6.1 Sterility 59
4.6.2. Stability 60
4.6.2.1 Effect of temperature and duration of storage on the physical
stability of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine.
60
4.6.3 Safety and side effects 60
4.6.3.1 Comparative safety of Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine in cattle and buffaloes 60
4.6.4 Challenge- protection assay (Vaccination- challenge study) 62
4.6.4.1 Vaccination challenge protection assay of Montanide®
adjuvanted combined hemorrhagic septicemia-mastitis vaccine
in mice.
62
4.6.5 ELISA- based antibody titers generated by Montanide®
adjuvanted combined HS-mastitis vaccine against P. multocida,
S. aureus and Str. agalactiae in cows and buffaloes
63

xii
4.7 Incidence of HS 65
4.7.1 Number of new HS cases in vaccinated and control cows,
buffaloes and calves during 180 days post- vaccination
65
4.8 Incidence and prevalence of S. aureus and Str.
agalactiae mastitis
68
4.8.1 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter-based incidence of Str. agalactiae and S.
aureus IMIs in cows and buffaloes initially culture negative
and CMT/ SFMT negative
68
4.8.2 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter- based incidence of Str. agalactiae and S.
aureus IMIs in cows and buffaloes initially culture positive and
CMT/ SFMT positive
70
4.8.3 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter- based time points prevalence of Str.
agalactiae and S. aureus IMIs in cows and buffaloes initially
culture positive and CMT/SFMT positive
72
4.9 Mastitis screening tests based prevalence and
incidence of mastitis
75
4.9.1 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT and SFMT based quarter incidence of mastitis
in cows and buffaloes initially culture negative for S. agalactiae
and S. aureus and CMT/ SFMT negative
75
4.9.2 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT/SFMT- based quarter time points prevalence
of mastitis in cows and buffaloes initially negative for Str.
agalactiae and S. aureus plus CMT/SFMT negative
77
4.9.3 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT and SFMT based quarter incidence of mastitis
80

xiii
in cows and buffaloes initially culture positive for S. agalactiae
and S. aureus and CMT/ SFMT positive
4.9.4 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT/SFMT- based time points quarter prevalence
of mastitis in cows and buffaloes initially positive for Str.
agalactiae and S. aureus plus CMT/SFMT positive
82
4.10 Severity of mastitis in vaccinated and control
animals
85
4.10.1 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the severity of mastitis.
85
4.11 Milk somatic cell count 87
4.11.1 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the quarter milk somatic cell count (× 105
ml of
milk) in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased and control-diseased groups of cows.
87
4.12 Effect of vaccination on milk yield 91
4.12.1 Milk yield (Mean ±SE; L/24 hour) in vaccinated and control
cows and buffaloes at different post-vaccination days
91
4.13 Cost- benefit analysis of Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine
95
4.13.1 HS related cost -benefit calculations 95
4.13.2 Mastitis related cost-benefit calculations 97
4.13.2.1
Mastitis control related benefits accruing from use of
Montanide® adjuvanted combined hemorrhagic septicemia-
mastitis vaccine
97
4.14 Calculation of vaccine efficacy 99
4.14.1 Calculation of vaccine efficacy for HS in cows, buffaloes and
calves
99
4.14.2 Calculation of vaccine efficacy for mastitis in cow sand
buffaloes
100
5 DISCUSSION 101
xiv
5.1 Preparation of combined HS-mastitis vaccine and its evaluation
against HS
101
5.2 Evaluation of combined HS-mastitis vaccine against mastitis 105
6 SUMMARY 115
LITERATURE CITED 120
APPENDICES 140

xv
LIST OF TABLES
Table No. Title Page No. 2.1 Cure rate of 9 different treatment protocols used to manage
an outbreak of HS (Pasteurella multocida serotype B2) that
occurred in the wake of SAT2 FMD infection in cow and
buffalo fattening calves in Alexandria Egypt.
9
2.2 Reported prevalence of mastitis in cows and buffaloes in
some selected studies conducted in Pakistan.
20
3.1 The oligonucleotide primer pair used for PCR based
confirmation of Pasteurella multocida
30
3.2 Sequence of nuc gene primers used for confirmation of S.
aureus and PCR product size
33
3.3 Composition of PCR reaction mixture used for amplification
of nuc (S. aureus specific) gene
34
3.4 Composition and preparation of Congo red agar medium
(Mathur et al., 2006). Ingredients per liter (L):
35
3.5 Selection of vaccinal isolate by biofilm detection methods
and other traits
38
3.6 Grouping of animals, their experimental treatments, and
HS/mastitis status at the time of initiation of field trial of
combined HS- mastitis vaccine.
42
4.1 Pasteurella multocida isolates recovered from peripheral
blood, femur, and heart blood of HS affected animals
45
4.2 Biochemical profiles of P. multocida isolates (n=3)recovered
from femur bone marrow samples and heart blood samples
46
4.3 Antibiotic susceptibility profiles of 3 candidate vaccinal isolates
of P. multocida against 11 antibiotics/antimicrobials
47
4.4 Pathogenicity of the 3 candidate vaccinal isolates of P. multocida in mice
48
4.5 Seven-digit numerical biotype (biochemical) profiles of 4
candidate vaccinal S. aureus isolates determined by using
API® STAPH
48
4.6 Antibiotic susceptibility profiles of 4 candidate vaccinal isolates of
S. aureus against 11 antibiotics/antimicrobials 50
4.7 Pathogenicity of the 4 candidate vaccinal isolates of S. aureus
in rabbits
51
4.8 Tube method-based scores of biofilm production by
candidate vaccinal S. aureus isolates (n=4)
51
4.9 Spectrophotometric method based optical densities (OD
values) for biofilm production of 4 candidate vaccinal S.
aureus isolates
52

xvi
4.10 Degrees of biofilm production trait of 4 candidate vaccinal S.
aureus isolates on Congo red agar (CRA)
52
4.11 Seven-digit numerical (biochemical) profiles of 2 candidate
vaccinal Str. agalactiae isolates determined by API® 20
STREP (bioMerieux, France)
54
4.12 Antibiotic susceptibility profiles of 2 candidate vaccinal
isolates of Str. agalactiae against 11
antibiotics/antimicrobials
55
4.13 Pathogenicity of the 2 candidate vaccinal isolates of Str.
agalactiae in rabbits
55
4.14 Indirect ELISA-based serum antibody titers in rabbits to 3
candidate vaccinal isolates of P. multocida
56
4.15 Indirect ELISA-based serum antibody titers in rabbits to 4
candidate vaccinal isolates of S. aureus
56
4.16 Indirect ELISA-based serum antibody titers in rabbits to 2
candidate vaccinal isolates of Str. agalactiae
57
4.17 Composition of Montanide® adjuvanted combine HS-
mastitis vaccine.
57
4.18 Indirect ELISA-based serum antibody titers in rabbits to
vaccinal isolates of P. multocida, S. aureus and
Str.agalactiae
58
4.19 Effect of temperature and duration of storage on the physical
stability of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine
60
4.20 Comparative safety of Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine in cattle and
buffaloes
61
4.21 Comparative safety of Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine in mice
61
4.22 Vaccination challenge protection assay of Montanide®
adjuvanted combined hemorrhagic septicemia-mastitis
vaccine in mice
62
4.23 ELISA-based antibody titers generated by Montanide®
adjuvanted combined HS-mastitis vaccine against P.
multocida, S. aureus and Str. agalactiae in cows and
buffaloes
64
4.24 Number of new HS cases in vaccinated and control cows,
buffaloes and calves during 180 days post- vaccination period
66
4.25 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter-based incidence of Str. agalactiae and S.
aureus IMIs in cows and buffaloes initially culture negative
and CMT/ SFMT negative
69
4.26 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter- based incidence of Str. agalactiae and S.
aureus IMIs in cows and buffaloes initially culture positive
and CMT/ SFMT positive
70

xvii
4.27 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on quarter- based time points prevalence of Str.
agalactiae and S. aureus IMIs in cows and buffaloes initially
culture positive and CMT/ SFMT positive
73
4.28 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT and SFMT-based quarter incidence of
mastitis in cows and buffaloes initially culture negative for S.
agalactiae and S. aureus and CMT/ SFMT negative
75
4.29 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT/SFMT- based quarter time points
prevalence of mastitis in cows and buffaloes initially negative
for Str. agalactiae and S. aureus plus CMT/SFMT negative
78
4.30 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT and SFMT based quarter incidence of
mastitis in cows and buffaloes initially culture positive for S.
agalactiae and S. aureus and CMT/ SFMT positive
80
4.31 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on CMT/SFMT- based time points quarter
prevalence of mastitis in cows and buffaloes initially positive
for Str. agalactiae and S. aureus plus CMT/SFMT positive
83
4.32 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the severity of mastitis in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased,
control-diseased groups of cows and buffaloes
85
4.33 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the quarter milk somatic cell count (× 105
ml of
milk) in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of cows
88
4.34 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the quarter milk somatic cell count (× 105
ml of
milk) in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of cows
88
4.35 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the quarter milk somatic cell count (× 105
ml of
milk) in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of buffaloes
90
4.36 Effect of Montanide® adjuvanted combined HS-mastitis
vaccine on the quarter milk somatic cell count (× 105
ml of
milk) in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of buffaloes
90
4.37 Milk yield (Mean ±SE; L/24 hours) of vaccinated and control
cows and buffaloes at different post-vaccination days
94
4.38 Number of HS cases in vaccinated and control cows,
buffaloes and calves during 180 days post- vaccination period
and pecuniary losses based on market prices of affected
animals
96
xviii
4.39 Mastitis control related benefits accruing from use of
Montanide® adjuvanted combined hemorrhagic septicemia-
mastitis vaccine
98
4.40 Calculation of vaccine efficacy for HS in cows, buffaloes and
calves
99
4.41 Calculation of vaccine efficacy for mastitis in cows and
buffaloes
100

xix
LIST OF FIGURES Fig. No. Title Page No.
4.1 Indirect ELISA-based serum antibody titers in rabbits to vaccinal isolates
59
4.2 ELISA-based antibody titers generated by Montanide® adjuvanted combined HS-mastitis vaccine against P. multocida, S. aureus and Str. agalactiae in cows and buffaloes
65
4.3 Number of new HS cases in vaccinated and control cows, buffaloes and calves during 180 days post- vaccination
67
4.4 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter-based incidence of Str. agalactiae and S. aureus IMIs in cows and buffaloes initially culture negative and CMT/ SFMT negative
69
4.5 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter- based incidence of Str. agalactiae and S. aureus IMIs in cows and buffaloes initially culture positive and CMT/ SFMT positive
71
4.6 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter- based time points prevalence of Str. agalactiae and S. aureus IMIs in cows and buffaloes initially culture positive and CMT/ SFMT positive
74
4.7 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and SFMT-based quarter incidence of mastitis in cows and buffaloes initially culture negative for S. agalactiae and S. aureus and CMT/ SFMT negative
76
4.8 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT/SFMT based quarter time points prevalence of mastitis in cows and buffaloes initially negative for Str. agalactiae and S. aureus plus CMT/SFMT negative
79
4.9 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and SFMT based quarter incidence of mastitis in cows and buffaloes initially culture positive for S. agalactiae and S. aureus and CMT/ SFMT positive
81
4.10 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT/SFMT- based time points quarter prevalence of mastitis in cows and buffaloes initially positive for Str. agalactiae and S. aureus plus CMT/SFMT positive
84
4.11 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the severity of mastitis in vaccinated–non-diseased, control-non-diseased, vaccinated –diseased, control-diseased groups of cows and buffaloes
86
4.12 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the quarter milk somatic cell count (× 10
5 ml of milk) in
vaccinated–non-diseased, control-non-diseased, vaccinated –diseased, control-diseased groups of cows
89
4.13 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the quarter milk somatic cell count (× 10
5 ml of milk) in
vaccinated–non-diseased, control-non-diseased, vaccinated –diseased, control-diseased groups of buffaloes
91
4.14 Milk yield (Mean ±SE; L/24 hours) in vaccinated and control cows and buffaloes at different post-vaccination days
91
xx
LIST OF PLATS
Plate No. Title Page No.
1 Growth of a P. multocida on blood agar plate 46
2 Growth of a P. multocida on Casein-sucrose yeast agar 46
3 Seven-digit numerical biotype profile of a candidate vaccinal
S. aureus isolates determined by API® STAPH.
49
4 Latex agglutination test positive and negative reactions using
Staphytect plus® kit
49
5 Growth of a Str. agalactiae on a blood agar plate 53
6 Seven-digit numerical biotype profile of a candidate vaccinal
Str. agalactiae isolates determined by API® 20 STREP.
54
7 Montanide® adjuvanted combined hemorrhagic septicemia-
mastitis vaccine
58
xxi
LIST OF APPENDICES
App. No. Title Page No.
I Composition of casein-sucrose-yeast (CSY) agar and method
of its preparation
101
II Preparation of blood agar medium and collection of
defibrinated blood
101
III Preparation of culture media and composition 102
A Blood agar plates 102
B Staph-110 agar plates 102
C MacConkey Agar Plates 103
D Nutrient Broth 103
E Brain Heart Infusion Broth (BHI) 104
F Tryptone Soya Broth 104
IV Collection, processing and preparation of rabbit plasma for
tube coagulase test to differentiate coagulase positive and
coagulase negative Staphylococci
105
V Preparation of sterile milk whey 106
VI CAMP (Christie, Atkins and Munch-Peterson)-Esculine test 106
VII Somatic cell count 106

xxii
ABSTRACT The present study represents a maiden attempt to develop and evaluate a combined hemorrhagic
septicemia (HS) and mastitis vaccine in cows and buffaloes. The study was compartmentalized into
two phases. In phase I (laboratory settings), isolates of Pasteurella multocida, Staphylococcus aureus
and Streptococcus agalactiae isolated from field cases of HS and mastitis were scrutinized for
virulence/pathogenicity and immunogenicity in laboratory animals. Bacterin- toxoids of S. aureus and
Str. agalactiae were blended with prepared antigen of P. multocida, Montanide® ISA 201 VG,
thimerosal and sodium azide to prepare combined HS- mastitis vaccine that was evaluated for
sterility, safety and side effects under in vitro conditions and in vivo in cattle, buffaloes and mice. A
challenge-protection assay conducted in immunized mice indicated 100% survival of challenged
mice. The vaccine was physically stable in terms of pH, sedimentation, color, appearance, and
syringibility for 6 months observation period at 37°C. In Phase II (field evaluation), the combined
vaccine was evaluated in adult cattle and buffaloes and calves of cattle and buffaloes. To this end, a
total of 70 S. aureus and Str. agalactiae free lactating buffaloes (n=45) and cows (n=25), 50 lactating
cows (n=25) and buffaloes (n=25) positive for S. aureus/Str. agalactiae and dairy calves (buffalo
calves n=70; cow calves n=50) aged up to 1 year were treated with 2 doses of combined HS-mastitis
vaccine at 21 day interval and evaluated (where relevant) for 6 months in terms of ELISA based
antibody titers against P. multocida, S. aureus and Str. agalactiae, incidence of HS, local and
systemic reactions, incidence and prevalence of S. aureus and Str. agalactiae mastitis, severity of
mastitis, milk somatic cell count, milk yield, cost effectiveness and vaccine efficacy. ELISA based
antibody titers against P. multocida, S. aureus and Str. agalactiae were significantly (P˂0.05) higher
in vaccinated groups than in unvaccinated groups. Two cases of HS were recorded in vaccinated
animals vis-à-vis 7 cases in un-vaccinated animals. Incidence of S. aureus and Str. agalactiae over
180 days in vaccinated and un-vaccinated cows and buffaloes initially cultural –ve for these
pathogens was 3 and 10, respectively; the corresponding figures in groups initially culture +ve for
these pathogens being 2 and 12, respectively. Cumulative mean somatic cell counts in vaccinated
groups were significantly lower (P<0.05) than those in respective unvaccinated controls. Milk yield
was significantly higher (P<0.05) in vaccinated cows and buffaloes than in un-vaccinated controls.
Mastitis severity scores were significantly lower (P<0.05) in vaccinated groups than in unvaccinated
controls. The vaccine tested had a vaccine efficacy 84.78 and 90.25% against HS and mastitis,
respectively with a financial benefit worth Rs 2,060,300. In sum, Montanide® adjuvanted combined
HS-mastitis vaccine had preventative role against HS and both preventative and curative role against
S. aureus and Str. agalactiae associated mastitis. In view of the preliminary nature of the study,
additional work involving much larger number of cows, buffaloes and calves is clearly warranted.

1
CHAPTER 1
INTRODUCTION
Field surveys of major dairy animal diseases in Pakistan have indicated that
hemorrhagic septicemia (colloquially known as "gulghotoo") and mastitis (colloquially
known as "Sauroo") are the two most rife, endemic and economically important diseases of
buffalo and cattle (Khan et al., 1991; Anonymous, 1996; Hussain et al., 2005) Hemorrhagic
septicemia (HS) is an infectious, highly fatal bacterial disease caused by Pasteurella
multocida serotype B: 2 or Robert's type 1 (De Alwis, 1984; OIE, 2012). This organism was
also incriminated as the etiologic agent of a respiratory syndrome in buffaloes in Landhi
Dairy Colony, Karachi (Zahur et al., 2007). Accounting for annual losses of Rs. 2.17 billions
in Punjab only, this disease was ranked number one among the economically important
livestock diseases in Pakistan (Anonymous, 1996) and it was further appraised that even a
50% reduction in HS incidence would be sufficient to bridge the gap between growing milk
demand and supply. Milk demands will double itself by 2030 (Upton, 2001). Control of
infectious diseases of epidemic proportion ought to be the priority number one in the animal
sector in developing countries (Arye-Smith, 1971). Launching a National program for control
of HS has been identified as an animal health imperative in Pakistan (Afzal, 2009).
Vaccination using different types of vaccines (e.g. formalin-killed alum precipitated bacterin,
oil adjuvanted vaccines) is the single most important control measure against HS in Pakistan
and in other Asian , African, South European and Middle East countries where this disease is
endemic (Benkirane and De Alwis, 2002; Sotoodehnia et al., 2005). The vaccines are quite
effective when used as per the recommendations of manufacturers and extension agencies.
Alum precipitated bacterin is still the most commonly used HS vaccine in Pakistan but it
gives immunity for a short duration i.e. 3 months (Muneer et al., 2005) and its protective
efficacy is about 60% (FAO, 1998). Comparative field evaluation of alum precipitated (APV)
and oil adjuvanted HS vaccines has demonstrated a justification for the replacement of the
former by the later (Muneer et al., 2005).
Mastitis (inflammation or swelling of milk producing organ) is another
common dairy disease which although not fatal, causes colossal economic losses to our
resource-poor dairy farmers and milk processing industry (Khan et al., 1991; Blood and

2
Studdert, 1999; Deb et al., 2013). Research conducted over the past four decades has shown
that at the very least 20% of cows and buffaloes are afflicted with this disease. The huge
affected population of buffaloes and cattle not only sustains nearly a 25% reduction in their
milk yield (Arshad et al., 1995; Radostits et al., 2007) but the milk produced is also
unwholesome for human consumption as it contains pathogenic bacteria, toxins and other
harmful substances some of which are not destroyed by UHT treatment (Verdi, 1988;
Barbano, 1989; Heeschen, 2005) utilized by the milk processing industry in Pakistan. Control
of this disease is therefore, imperative for economically viable dairy farming as well as to
produce milk which conforms to the standards of WTO Accord.
Over the past nine years, researchers at the Department of Clinical Medicine and
Surgery, University of Agriculture, Faisalabad have demonstrated that a cost-effective
control of mastitis is an attainable dairy health objective with the use of locally prepared
mastitis vaccines containing two most prevalent mastitis pathogens (Staphylococcus aureus
and Streptococcus agalactiae) associated with mastitis in local dairy cattle and buffaloes
(Shakoor et al., 2006; Athar et al., 2007). Montanides® are ready-to- use mineral oil
adjuvants. These are reported to elicit superior immune response as compared to aluminium
hydroxide in cattle when used as an adjuvant in HS vaccine (Reddy et al., 1995). These
retain potency for longer period than conventional aqueous formulation following storage at
4°C and have no evidence of toxicity in cattle even after booster dose.
At present, a "do nothing" policy is in place virtually everywhere in Pakistan as far
as mastitis control is concerned. As mentioned earlier, the use of HS vaccine is a fairly well
established practice in Pakistan and the mastitis vaccination can conceivably be integrated
into the vaccination program already in vogue by developing a combined HS- mastitis
Montanide®vaccine. The use of a combined HS-mastitis vaccine would theoretically be better
than the use of either vaccine alone because:
1) Control of HS and mastitis in tandem with a single vaccine would be achieved,
2) Less frequent handling of animals required for vaccination against HS and mastitis.
3) Combined HS-mastitis vaccination may potentially be more protective and cost-
effective by virtue of its Montanide® adjuvant.
4) Initiation of control of mastitis against which a "do nothing" policy is in place
virtually throughout the country

3
The present study was therefore planned to prepare and evaluate a Montanide®
adjuvanted combined hemorrhagic septicemia-mastitis vaccine under laboratory settings and
under field conditions
Objectives:
a. To prepare and evaluate a Montanide® adjuvanted combined hemorrhagic septicemia-
mastitis vaccine in laboratory settings and experimental animals
b. To evaluate Montanide®adjuvanted combined hemorrhagic septicemia-mastitis
vaccine under field conditions.
c. To determine the cost-benefit ratio of hemorrhagic septicemia and mastitis control
through the use of a Montanide® adjuvanted combined hemorrhagic septicemia-
mastitis vaccine.

4
CHAPTER 2
REVIEW OF LITERATURE
2.1 Economic impact of animal diseases in developed and developing countries
Animal diseases, in particular infectious diseases, have major adverse effects on
livestock productivity as well as animal welfare worldwide. The costs of animal diseases are
around 17% of turner-over of the livestock sector in the developed countries while it is 35-
50% in the developing countries (Bishop et al., 2010; Holden et al., 1996). Infectious
diseases pose a significant challenge to all animal production systems, which sustain
significant production losses from these diseases. Thus animal diseases are a significant
threat to food security worldwide. In addition, several infectious diseases do not respect the
species barriers and about 70% of all infections are common to man and animals (zoonosis).
Several options available to curtail the colossal economic losses associated with animal
diseases include immunization, chemotherapy, improved management, nutritional and
genetic improvement, diagnosis and removal of infected animals. Implementation of each of
these options can add to solving the problems of productivity losses and zoonosis caused by
these diseases (Bishop et al., 2010). Investment in animal health research has been shown to
pay rich dividend by alleviating poverty in developing countries (Perry et al., 2002).
According to Garcia et al. (2003), a comparison of average milk yield across different
countries would reveal that one New Zealand cow produces as much milk as 3 dairy animals
in Pakistan, whereas one American cow produces milk that is nearly equal to milk produced
by 7 Pakistani cows. This dramatic difference in the milk yield of Pakistani dairy animals vis-
à-vis dairy cows of advanced countries can be attributed to a verity of factors including
genetics, nutrition, management and diseases etc.
2.2 Mastitis
According to Bishop et al (2010), in terms of economic losses, mastitis is the single
most important disease in cows in developed counties and is also of major importance in
developing countries. Avenues of economic losses include milk loss, premature culling, and
cost incurred on replacements, occasional mortality, treatment costs and the cost of discarded
milk. Treatment of clinical cases not only involves the costs of antibiotics and other
therapeutic agents, but may also create opportunities of antibiotic and drug residues in the

5
milk of treated animals. Furthermore, in livestock production systems where farmers are
paid according to milk quality, elevated milk somatic cell count (SCC) as a result of clinical
and sub clinical mastitis may affect the profitability of dairying. Selection against subclinical
mastitis using SCC is now widely practiced in countries where dairying is well developed.
Thus, selection for resistance against clinical mastitis is one of the important components of
dairy cattle breeding programmes in Scandinavian countries.
Khan and Chaudhry (2001) analyzed the data on 2704 lactations of 993 Nili-Ravi
buffaloes to investigate the frequency and behavior of short and complete lactations.
Lactation milk yield up to 44 weeks (308 days) was included and lactations with less than 56
days were excluded from the analysis. Of 2107 lactations of less than 14 weeks durations, the
causes of drying could be determined from the farm record for 534 lactations. Mastitis
accounted for 31% of these lactations for drying during the course of shorter than normal
lactations. A meta-analysis of investigations conducted for one year period (1994-1995) on
economically important diseases of livestock in Punjab, Pakistan ranked mastitis at the 5th
slot. This disease was responsible for 8% of the total pecuniary losses inflicted by all
livestock diseases in this province (Khan et al., 1994-95).
2.3 General aspects of hemorrhagic septicemia (HS)
Hemorrhagic septicemia (HS) is an acute highly fatal, septicemic disease occurring
primarily in water buffaloes (Bubalus bubalis) and cattle but occasionally in other domestic
and wild mammals (De Alwis, 1992; Adlakha and Sharma, 1992). Water buffalo is far more
susceptible to this disease than cattle (De Alwis, 1999; Khan et al., 2006). The etiologic
agent of HS is a Gram negative bacterium, Pasteurella multocida (Tabatabaei et al., 2007;
Carter and De Alwis, 1989). Pasteurella multocida has at least five serotypes designated as
A, B, C, D and E. Serotypes B: 2 and E: 2 are two common serotypes of P. multocida
associated with HS in animals in Asia and Africa, respectively (Benkirane and De Alwis,
2002). In serotype classification, the letter is used to denote the capsular antigen that was
determined originally by Carter (1984) with the use of indirect hemagglutination test. The
numeral 2 denotes the somatic or O antigen that was determined originally by Namioka and
Bruner (1963) by using agar gel diffusion precipitin test developed by Heddelston and
associates.

6
Hemorrhagic septicemia (HS) is one of the most important diseases of bovines in
South Asian and Middle Eastern countries. Epidemiological studies conducted in India over a
period of approximately 13 years (1974-1986) indicated that HS was placed mortality wise
first and morbidity wise second as compared to 4 other epizootic diseases namely, foot and
mouth disease (FMD), rinderpest, anthrax and black quarter (Dutta et al., 1990).
2.3.1 Carrier state
Carriers animals are the source of the organism for other animals. Cattle and buffaloes
are considered to be the principal carriers of P. multocida, but other animals such as pigs,
sheep, goats and horses can also act as carriers. The incidence of HS has been shown to be
directly proportional to the number of carriers in the animal population. Thus in India in low
incidence areas, no carriers were detected, whereas in the moderate and high incidence areas,
the carrier rates were 1.9 and 5-6%, respectively (Adlakha and Sharma, 1992). Regular
vaccination has been shown to eliminate the carrier status. (De Alwis, 1984).
2.3.2 Transmission
Pasteurella multocida is spread by inhalation, ingestion of contaminated feed, water,
direct contact, and fomites etc. The causative agent of HS is mostly shed into the oropharynx,
lymphatic tissue associated with the upper respiratory tract, and periodically in nasal
secretions during stress (poor food supply, close herding, and wet conditions). HS pathogen
does not remain alive for log time, but can survive for a few hours or days in damp soil. The
favorable condition for transmission is rainy season and humid environment (OIE, 2012).
2.3.3 Association of HS with other diseases
Some other infectious diseases may predispose to occurrence of hemorrhagic
septicemia. Foot and mouth disease causes immunosuppression (Golde et al., 2014; Maddur
et al., 2010) and thus can precipitate an attack of HS or some other bacterial infections.
Elshemey and Abd-Elrahman( 2013) reported that 56.32% (n=2600) of 4616 cow and buffalo
fattening calves infected with SAT2 serotype of FMD virus showed signs of hemorrhagic
septicemia (P. multocida B2 serotype) with a case fatality rate of 13.92%. All of 5630 calves
had been vaccinated with an inactivated FMD vaccine containing antigens of serotype A and
O. Kumar et al. (2007) reported outbreaks of concurrent pasteurellosis (P. multocida
serotype B: 2) and classical swine fever that occurred in Indian Punjab. Overall mortality,
morbidity, and case fatality rate were 77.5, 88.2, and 87.8%, respectively, in pigs ≤3 months

7
of age, and 8.2, 20.5, and 40%, respectively in older pigs. More marked pneumonic lesions
were recorded in cases from which P. multocida was isolated.
2.3.4 Pathogenesis
Hemorrhagic septicemia is associated with P. multocida serotype B: 2. It is a Gram
negative bacterium that secretes lipopolysaccharide (LPS), also named as endotoxin, in blood
stream of the host on its lysis. All manifestations of the disease are due to these endotoxins
which lead to endotoxemia associated sepsis (De Alwis, 1992; Horadagoda et al., 2001).
Endotoxins induce many complex deleterious biological reactions in the host. They stimulate
a cascade of endogenous mediators such as the coagulation and arachidonic acid systems.
Endotoxin also activate polymorphonuclear cells, monocytes and macrophages. When
monocytes and macrophages sense the presence of endotoxin, they release special peptides
called cytokines.
Horadagoda et al (2002) studied the clinical changes along with acute phase
responses (including tumour necrosis factor; TNF α) in 6 buffalo calves following iv
administration of P. multocida B:2 endotoxin @ of 1µg /kg in 10 ml of phosphate - buffered
saline. There was a rapid onset of clinical signs (dullness, sternal recumbency, fever,
excessive salivation and respiratory difficulty) following administration of endotoxin. These
clinical signs lasted for about 12 hours. Serum TNF α levels increased rapidly within one
hour post inoculation of endotoxin, reaching their peak levels (8-150 ng/ml) at 1-2 hours post
inoculation and then declined rapidly to baseline values at 3-5 hours post inoculation. Other
acute phase responses included leukopenia, decreased serum levels of iron and zinc. In
addition, there was a delayed but prolonged increase in haptoglobin from 12 hours post
inoculution that reached its peak from 60 hours post inoculation. Control buffalo calves (n=3)
showed neither clinical signs nor the acute phase serum changes mentioned above. The
results of this study confirmed the notion that endotoxin has a cardinal role in the
pathogenesis of P. multocida B:2 associated pasteurellosis of buffalos, commonly knowns as
hemorrhagic septicemia.
2.3.5 Rainy seasonal occurrence
Although, occurrence of HS has been reported throughout the year (Dutta et al.,
1990), its incidence much higher in hot and humid weather (Sivakumar et al., 2012).
Chowdhury et al. (2014) recently reported an outbreak of HS in a buffalo herd in the month

8
of December. High humidity coupled with high environmental temperature and stressful
conditions prompts growth of the P. multocida and shortens the incubation period of the
pathogen by nearly 30 hours, resulting in HS outbreaks with high case fatality rate (De
Alwis, 1995).
2.3.6 Clinical signs and post mortem findings
Hussain et al. (2014) recorded an outbreak of pneumonic pasteurellosis (P.
multocida) in cattle and buffaloes in district Sahiwal of Pakistani Punjab. Clinical signs
observed included fever, anorexia, brisket edema, profuse salivation, mucopurulent nasal
discharge, protruded tongue with open mouth breathing, submandibular edema and dyspnea
with respiratory grunts. Necropsy of the dead animals reveled severe pneumonia with
consolidation of lungs, froth in trachea, clotted blood in heart, intense pleural adhesions,
accumulation of serosanguinous fluid in pericardial and peritoneal cavities. Acute HS with
severe respiratory distress has also been reported in wild ruminents (Dhoot and Upadhye,
2001)
2.3.7 Treatment
Owing to generally a peracute nature of the disease, treatment of HS is not effective,
and there is need to find effective therapeutic agents for the treatment of HS. This disease.
According to Shahzad et al. (2013), treatment was effective only when instituted at the early
stage of the disease. These workers also reported that ciprofloxacin, ceftiofur hydrochloride +
tylosin were the most effective antibiotics for treatment. Nonetheless, the workers did not
mention the cure rates of these antibiotics. Egyptian workers (Elshemey and Abd-Elrahman,
2013), treated an outbreak of HS that occurred in the wake of SAT2 FMD infection in cow
and buffalo fattening calves. Table 2.1 gives the cure rate of 9 different treatment protocols
used to manage this HS outbreak.
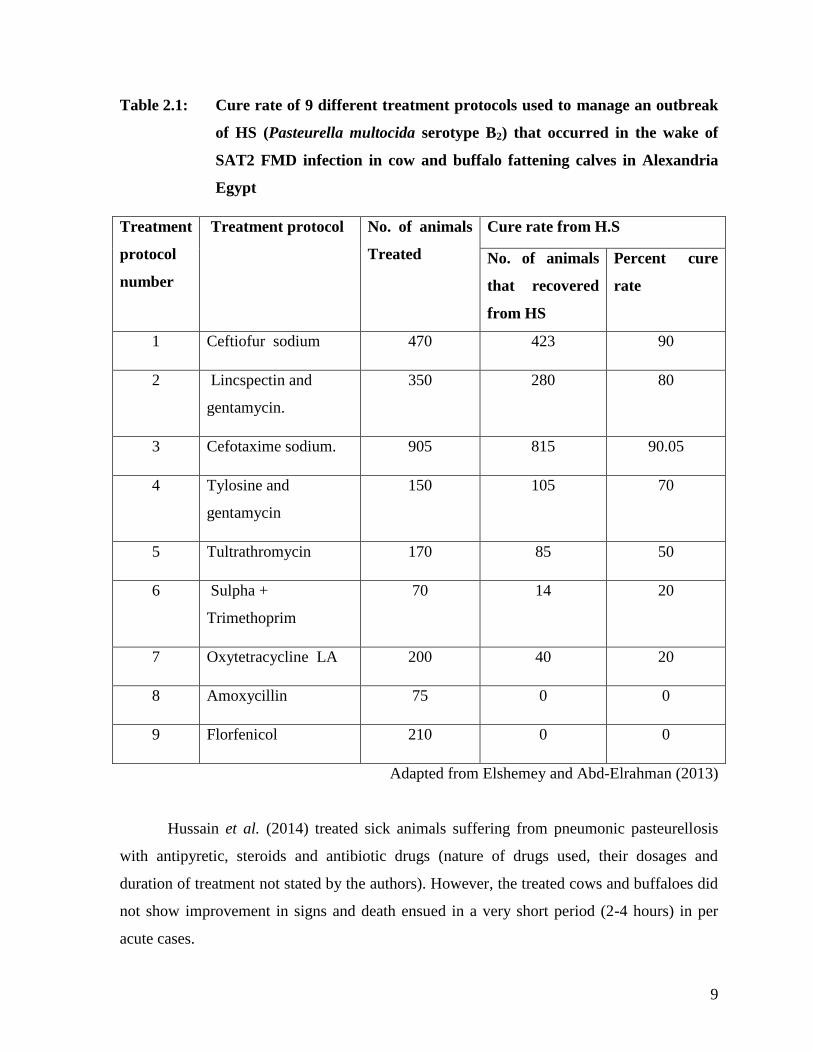
9
Table 2.1: Cure rate of 9 different treatment protocols used to manage an outbreak
of HS (Pasteurella multocida serotype B2) that occurred in the wake of
SAT2 FMD infection in cow and buffalo fattening calves in Alexandria
Egypt
Treatment
protocol
number
Treatment protocol No. of animals
Treated
Cure rate from H.S
No. of animals
that recovered
from HS
Percent cure
rate
1 Ceftiofur sodium 470 423 90
2 Lincspectin and
gentamycin.
350 280 80
3 Cefotaxime sodium. 905 815 90.05
4 Tylosine and
gentamycin
150 105 70
5 Tultrathromycin 170 85 50
6 Sulpha +
Trimethoprim
70 14 20
7 Oxytetracycline LA 200 40 20
8 Amoxycillin 75 0 0
9 Florfenicol 210 0 0
Adapted from Elshemey and Abd-Elrahman (2013)
Hussain et al. (2014) treated sick animals suffering from pneumonic pasteurellosis
with antipyretic, steroids and antibiotic drugs (nature of drugs used, their dosages and
duration of treatment not stated by the authors). However, the treated cows and buffaloes did
not show improvement in signs and death ensued in a very short period (2-4 hours) in per
acute cases.

10
Ashraf et al. (2009) compared florfenicol, florfenicol + flunixin meglumine,
amoxicillin and amoxicillin + flunixin meglumine for the treatment of HS in buffalo calves (a
total of 40 buffalo calves aged 6-18 months). The recovery rate (survival) in calves treated
with florfenicol + flunixin meglumine was 90%, whereas in calves treated with amoxicillin +
flunixin meglumine was 60%. Calves treated with florfenicol alone had a recovery rate of
80%, whereas 50% of the calves treated with amoxicillin alone survived. Chowdhury et al.
(2014) treated HS affected buffaloes with (a) Inj. Ceftiofur sodium @ 1gm I/M for 5 days,
(b) Inj. Meloxicam @ 20 ml I/M for 5 days, (c) Inj. Prednisolone @10 ml I/M for 5 days (in
tapering doses). The buffaloes with early stage of disease responded to the treatment.
Raza et al. (2000) investigated a total of 50 animals (39 buffaloes and 11 cattle)
suffering from hemorrhagic septicemia (HS), selected from the field and divided into 3
groups (A, B and C). Group A (20 animals) was treated with protocol A i.e. norfloxacin
(Norfloxillin; Tarobina) + diclofenac sodium +Novacoc forte (Richter Pharma). Group B (20
animals) was treated with protocol B which was essentially the same as protocol A except
that Novacac forte was omitted. Group C (10 animals, control) was treated with one of
conventional therapies of HS, i.e gentamicin + dipyron. The disease severity index of
experimental animals under each treatment protocol was recorded at 0, 12, 24, 48, and 72
hours from the start of treatment. All treatments were repeated at 12, 24, and 48 hours after
start of treatment as the sitaution warranted. The survival percent was 85, 80 and 30 amongst
animals treated respectively with protocol A, B and C. Three animals in group A and 2
animals in group B died after recovering completely. Excluding these animals from the
recovered ones, net survival (percentage) was 70, 70, and 30 in groups A, B and C,
respectively. In terms of reduction of severity of disease, there was significance difference
(P≤ 0.05) between protocol C and A, between protocol C and B at 12 and 48 hours after
treatment but there was non-significant difference at hours 24. Both treatment protocols A
and B exploiting the use of a quinolone (norfloxacine) plus a non–steroidal anti-
inflammatory drug (diclofenac sodium) with or without a toxin neutralizer and circulatory–
stimulant (Novacoc forte) were more effective than the conventional treatment i.e gentamicin
+ dipyron for the treatment of HS.

11
2.3.8 Morbidity, mortality and case fatality rates
Hemorrhagic septicemia or pasteurellosis is probably the most serious disease of
buffaloes and there are outbreaks which cause heavy mortality and morbidity (Dhillion et al.,
1971; Saini et al., 1991; Joshiet et al., 1987; Dhand et al., 2002). The death rate in buffaloes
is three times more than in cattle (Bain et al., 1982). Field observations of 9 districts of
Punjab, Pakistan showed 9% mortality, 11% incidence, and 87% case fatality rates of
hemorrhagic septicemia in buffalo, whereas these values were 2.5, 4, and 62% in cattle
(Sheikh et al., 1996)
Taking cues from the previously reported observation that fever increases the survival
rate in reptiles, Kluger and Vaughn (1978) investigated the relationship between fever and
survival in rabbits infected with Pasteurella multocida. A statistically significant correlation
was recorded between the fever magnitude and survival. As fever increased by 2.25 ºC, the
survival rate increased.
2.4 Prevalence and economic significance of HS in Pakistan
Hemorrhagic septicemia is a common and important infectious disease of buffaloes
and cattle in Pakistan (Khan et al., 1991; Ashraf et al., 2011). With a mortality rate near
70% (May et al., 2001), this disease was responsible for causing pecuniary losses worth Rs
2.17 billion per annum in 1996 in the Punjab province alone (Anonymous, 1996). Owing to
an increase in animal population (cattle n=41.2 millions; buffalo n= 35.6 millions) in 2014-
2015, and escalation of prices of animals, the current monitory losses may be at least 2
magnitude higher than these figures. Using an active surveillance system, Khan et al. (1991)
reported an estimated economic loss of Rs 689900 due to HS in 10 randomly selected
villages of Tehsil Lahore. During 2000-2005, field workers of FAO project entitled “Support
for Emergency Prevention and Control of Main Trans-boundary Diseases in Pakistan”
conducted a Participatory Disease Surveillance. An active surveillance of trans-boundary
animal diseases and other important diseases throughout the country (10% villages were
randomly selected throughout Punjab, Sindh, Khyber Pakhtunkhwa, Azad Jammu and
Kashmir and Northern Areas) was conducted by 17 teams comprising of 5 active field
veterinarians each. The highest prevalence (49%) of HS was recorded in district Khanewal,
Punjab. On the other hand, the lowest prevalence (0.78%) was recorded in Jamshaid Saddar
town of Karachi. The highest prevalence (75.64%) was recorded in Faisalabad district, while

12
lowest (1.53%) in Bajaur agency (Farooq et al., 2007). Based on information gleaned from
veterinary officers in 9 districts of Pakistani Punjab, Sheik et al. (1996) documented 9%
mortality, 11% incidence, and 78% case fatality rates of hemorrhagic septicemia in buffalo,
whereas these values were 2.5, 4, and 62% in cattle. Disease incidence was higher in 0 to 24
month old animals and groups of less than 10 animals. Farooq et al. (2011) determined the
sero-surveillance, morbidity, mortality and case fatality rate of HS in cattle and buffaloes in
Dera Ghazi Khan district, Punjab, Pakistan. The average geometric mean titers (GMT)
recorded against HS in diseased buffaloes and cattle were 5.7 and 6.1, respectively. The
morbidity, mortality and case fatality rates respectively were 57.58, 52.30 and 90.83% in
young buffalo calves. The corresponding values for adult buffaloes were 3.17, 1.92 and
60.65%, respectively. In case of young cattle calves, morbidity, mortality and case fatality
rates were 8.63, 5.27 and 61.11%, respectively; the corresponding values in adult cattle being
4.83, 2.18 and 45.23%, respectively. This study demonstrated that the mortality, morbidity
and case fatality rates due to HS were higher in young calves than in adults both in buffaloes
and cattle. It also reinforced the notion that buffaloes are more susceptible to HS than cattle.
Recently, Khan et al. (2011) reported an outbreak of HS in buffalo calves (aged 6-11 months;
n=54) kept at the Livestock Experimental Station, University of Agriculture, Faisalabad,
Pakistan. This outbreak occurred in August and September, 2009. Morbidity and mortality
rates in this outbreak were 100% and 31.48%, respectively. Mortality peaked on 8th
day
despite institution of a theoretically effective treatment (Amoxicillin long acting, ceftiofur
sodium, prednisolone acetate + dexamethasone sodium phosphate and meloxicame). It is
pertinent to note that this outbreak occurred despite vaccination in June with a reputed oil
based HS vaccine inoculated as per the manufacturer‟s recommendation.
Khan et al. (2006) reported the findings of a comparative sero-surveillance and
clinical epidemiological study conducted on HS in cattle and buffaloes in district Malakand,
NWFP (current name of the province is Khyber Pakhtoon Khwa). The average geometric
mean titer (GMT) recorded against HS in buffaloes ranged from 4.12-46.98, whereas in cattle
the corresponding values ranged from 4.45-46.40. In young buffalo calves (the age not
mentioned by the authors), mortality rate, morbidity rate and incidence rate, were 21.19,
95.25 and 22.25%, respectively. In adult buffaloes, the corresponding values were morbidity,
mortality and case fatality rates were 5.49, 1.65 and 30%, respectively. In case of young

13
cattle calves, mortality, morbidity, and case fatality rates were lower than those in young
buffalo calves, being 1.77, 3.94 and 45%, respectively. The values for morbidity, mortality
and case fatality rates in adult cattle were 2.51, 0.39 and 15.79%, respectively.
Ahmad and Naz (2000) reported HS incidence of 6.8% in buffalo calves maintained
at the Livestock Experiment Station, Bahadurnagar, Okara, Pakistan. Riaz et al. (1992)
conducted an active surveillance from September 1989 to August 1990 in district Gujrat,
Pakistan. A total of 1025 farmers were interviewed. HS incidence of 1.31% in cattle and
7.24% in buffaloes was recorded.
With the establishment of Directorate for Animal Disease Surveillance and Reporting
System in Punjab under the administrative control of Director General (Ext), Livestock and
Dairy Development, Punjab, the disease incidence reports communicated by field
veterinarians through District Livestock Officers/ District Disease Reporting Officers across
Punjab during the month of July to September, 2009 were incorporated to prepare the first
quarter report on the distribution of notifiable animal diseases in the province (Anonymous,
2009). FMD (foot and mouth disease) PPR (Peste des petitis ruminants), CCPP (contagious
caprine pleuropneumonia), ET (enterotoxemia), HS, BQ (black quarter) and ecthyma are the
diseases among all the contagious diseases that were reported during the reporting period. A
total of 208 outbreaks of 23 contagious diseases were reported for different diseases
including HS (82), FMD (38), ET (19), PPR (12), BQ (9), and CCPP (9) as main prevalent
diseases in the reported area. HS, FMD, ET, PPR, BQ and CCPP respectively affected 247,
148, 74, 50, 9 and 159 heads of animals while 95, 22, 16, 13, 5 and 12 died, respectively.
Prevalence and incidence of HS, FMD, ET, PPR, BQ and CCPP were found to be higher than
other contagious diseases. HS and BQ were reported from Faisalabad, Kasur, Rahim Yar
khan and Chakwal, DG Khan, Rahim Yar Khan, respectively. FMD was found to be present
in district Bahawalpur, Dera Ghazi Khan and Faisalabad. FMD is a disease that affected
highest number of animal heads in Punjab with a low mortality rate. PPR and ET were
reported from district Bahawalpur, DG Khan, Faisalabad, Lahore and Bahawalpur, Chakwal,
DG Khan, and Faisalabad, respectively. It was concluded that HS, FMD, ET, PPR, BQ and
CCPP were the diseases of main concern in the reported areas of Punjab during the months of
July to September, 2009.

14
Khan (2013) reported that no case of HS was brought to the notice of District Diagnostic
Laboratory Rahim Yar Khan during the period from May – July, 2012.
2.5 Hemorrhagic septicemia vaccines
Use of vaccines for the control of pasteurellosis in animals is since long in vogue.
Shilston (1923) while reviewing the information on hemorrhagic septicemia (pasteurellosis)
(that was published in the book edited by Wooldridge, 1923) cited several authors and
investigators who used pasteurellosis vaccines for the control of this group of infectious
diseases associated with P. multocida. For want of availability of these archaic reports, the
work of some of these investigators has been reproduced almost verbatim from Shilston
(1923) in the ensuing paragraph Oreste and Armmani (1887) attenuated the Bacillus.
Bubalisepticus (P. multocida) by passage through pigeons and employed the blood of these
birds after they had died from the inoculation of the organism, for the immunization of the
buffaloes. Three injection of 0.1 ml of the blood were given. By growing the organism at
temperature of 300C to 32
0C, reduced the virulence of P. multocida so that the cultures could
be employed to give protection to the buffaloes and sheep. Lignieres (1902) prepared a
polyvalent vaccine with strains of the hemorrhagic septicemia bacillus from cattle, sheep,
horses, pigs, fowls, and dogs. Cultures of the organisms of uniform virulence were kept at a
temperature of 420
to 430 C. for five days to produce the first vaccine and for two days for the
second vaccine. The vaccines were injected subcutaneously in doses of from 0.15 to 1 ml
according to the size of the animals, at interval of 12-15 days and were assessed protective
against the disease in all species of animals for a period of about one year. Mohler and
Eichhorn (1912) employed vaccines prepared by Lignieres method of protecting the buffalo
herd in the Yellowstone Park against hemorrhagic septicemia which was causing a heavy
mortality. The organisms were injected subcutaneously with 1 ml of each vaccine at an
interval of ten days, and no further cases of disease occurred during the succeeding 12
months. These investigation also tested the immunization power of vaccines prepared in the
same way as those of Lignieres but to which 5% carbolic acid had been added, thus making
them dead vaccines. They found that the carbolized vaccine gave protection against an
inoculation of the virulent organism, but, as judged by complement fixation test of serum of
the treated animals, this protection was of shorter duration than after injections of
uncarbolized vaccines. Holmes tested the immunity conferred by dead vaccines prepared in

15
various ways from cultures of P. multocida. Broth cultures of a virulent strain of the
organism heated to 650
C. for half an hour and carbolized was found to protect young cattle
and buffaloes, when injected subcutaneously in dose of 5-10 ml for a period of 6 weeks
ageist an inoculation of the virulent organism. The vaccine was considered harmless, only
causing in some cases slight swelling which disappeared in a few days, protection was not
established until four days after the injection of vaccine. After 150,000 doses of vaccine were
employed annually in India as a prophylactic measure in areas where the disease is enzootic
at periodic, inoculations, being carried out at the beginning of the winter rains and monsoon
season. Muktesar Laboratory in India adopted serum-live culture inoculation method for the
control of HS. This method is briefly as follows: Buffaloes and hill cattle are first injected
subcutaneously with a protective dose of immune serum and 4 hours later receive 0.1 ml of
48 hours old broth culture of virulent hemorrhagic septicemia bacilli; the initial protection
may also be conferred by an injection of dead vaccine followed in ten days by 1ml of virulent
culture. A temperature reaction and local edema usually follows the inoculation of the
culture; when this has subsided, after 8-10 days an injection of 1ml broth culture was given.
Further injections of 5, 25, 100 and 500 ml of culture were given at intervals of ten days and
the animals were given at intervals of 10 days. The animals are then bled for serum; the
injections and bleedings were repeated, the dose of culture being gradually increased up to
100 ml. An injection of the serum confers an immediate immunity lasting for a period of 3-4
weeks. It has been very effective when employed during outbreaks of hemorrhagic
septicemia, the mortality being at once arrested; the serum also possesses some curative
power.
Aggressins were one of the earliest immunizing agents used against HS, blackleg,
anthrax and certain other human and animal diseases. These are the substances (including but
not limited to capsular material, bacterial protein, secretions, excretions, enzymes and toxins)
secreted and excreted by certain organisms under favorable conditions of growth, which have
the property of inhibiting phagocytosis by a specific action on the leucocytes and reticulo-
endothelial system. Owing to their deleterious effect on the tissues of the host and leucocytes
and reticulo-endothelial system, they may facilitate the rapid development of normally
sublethal infections of disease- producing organisms, resulting in death. Injection of
aggressins into the body causes the production of specific anti-aggressins (Scott, 1931).

16
Gochenour, (1924) cited by Scott, (1931) developed an aggressin for the immunization of
animals against hemorrhagic septicemia. The hemorrhagic septicemia aggressin is probably
the least efficient, due to the peracute nature of the disease. Losses from shipping fever were
reportedly nearly twice as great among cattle vaccinated with HS aggressins, or other
vaccines including bacterins at the stock yards or on arrival at the farm as among the cattle
not treated with these immunizing agents (Scott, 1931).
Currently, vaccination against HS is the single most important control intervention in
countries plagued by this disease. A variety of vaccines containing either whole local isolates
of P. multocida or their components have been developed and tested under control and filed
conditions (Verma and Jaiswal, 1998; Shivachandra et al., 2011). A few of these vaccines
have found their wide spread field use. One of the hallmarks of these vaccines is a
considerable variation in duration of immunity conferred (Shivachandra et al., 2011). Plain
broth formalin killed bacterins, alum precipitated and aluminium hydroxide gel adjuvanted
vaccines have been used until recently. Their immunity lasts for 4-6 months and their
protective efficacy is about 60% (FAO, 1998; Verma and Jaiswal, 1998; Tasneem et al.,
2009; Shivachandra et al., 2011). Qureshi and Saxena, (2014) recently reported that cattle
vaccinated with conventional alum precipitated HS bacterin did not develop and sustain
adequate levels of antibody for long duration. Owing to their shorter duration of immunity,
these vaccines are now being supplanted by oil adjuvant vaccines that give both a higher
degree of protection and a longer duration of immunity up to a year (Verma and Jaiswal,
1998; Tasneem et al., 2009). Oil adjuvant HS vaccines have the drawback of high viscosity
and consequently difficulty in injecting (De Alwis, 1984). In order to address these
difficulties, a double emulsion and multiple emulsion vaccines endowed with thin viscosity
and long stability have been developed that gave immunity similar to oil adjuvant vaccines
(Muneer and Afzal, 1989; Muneer et al.,1994; Verma and Jaiswal, 1997; Verma and Jaiswal,
1998). A score of studies have investigated live streptomycin-dependent P. multocida (Wei
and Carter, 1978; De Alwis et al., 1980; Lu and Pakes, 1981; Verma and Jaiswal, 1998;
Dagleish et al., 2007; Ataei et al., 2009) and P. multocida + P. haemolytica vaccine (Catt et
al., 1985). De Alwis et al. (1980) investigated a live mutant strain of P. multocida that was
administered as a single dose containing 1010
to 1011
viable organism. This vaccine
immunized 75 % of cattle and 100% of buffalo. The number of live microorganism used in

17
this vaccine is seemingly so high that it precludes the practical utility of this vaccine in the
immunological control of HS. In a subsequent study, De Alwis, (1981) reported that natural
P. multocida infection of buffalo calves give considerably higher immunity than that
conferred by vaccine which can be interpreted to mean that a live vaccine, using a suitable
mutant, may prove superior to the existing ones(Adlakha and Sharma, 1992).A live HS
vaccine containing P. multocida serotype B: 3, 4 was administered by intranasal aerosol in
Myanmar (Burma) to more than 15 million cattle and buffaloes between 1989 and 1999
(Myint et al., 2005). Field observations of veterinary officers in 9 districts of Pakistan,
Punjab have pointed out the occurrence of field outbreaks of HS despite vaccination with
alum- precipitated P. multocida bacterin prepared in public sector (Sheik et al., 1996). In
order to obtain maximum protection from HS vaccine, it is recommended that the vaccines
be prepared from P. multocida strains circulating in the regions of intended use. FAO
recommends a live avirulent HS vaccine prepared from a P. multocida serotype B: 3 of
fallow deer origin for use in Southeast Asia (Carter, 2005). Despite vaccination and
improved management, occurrence of disease outbreaks is a regular feature each year,
especially in endemic areas of the country (Afzal and Muneer, 1990). Pakistani workers
(Shahzad et al., 2013) reported an outbreak of HS in buffalo calves that had been vaccinated
with alum precipitated HS vaccine two months earlier. Arif et al. (2013) recently
documented that subunit or whole bacterin based P. multocida vaccines were less
immunogenic than an anti-idiotypic vaccine prepared against P. multocida B: 2.
Hemorrhagic septicemia is predominantly a disease of young cattle and buffaloes.
How soon after birth, the calves should be vaccinated against this disease is a very pertinent
question in the immunization program of dairy animals. Japanese workers (Otomaru et al.,
2015) in a bid to answer this question investigated the dynamics and duration of antibody
titers against P. multocida in Japanese black calves. To this end, 20 unvaccinated calves from
two Japanese black breeding farms in Japan were investigated. Serum samples were collected
from these calves at 1, 4, 8, 12, 16 and 20 weeks after birth. Similarly, serum samples were
also obtained from their dams once at 1 week after calving. Serum antibody titers against P.
multocida in calves at 1 week of age correlated very well with those in their dams at 1 week
after calving. Maternally derived antibody titers against P. multocida in experimental calves
fell to their lowest values at 4 weeks of age. One of the findings relevant to immunization

18
against P. multocida was that calves started producing antibodies against this organism by
themselves between 4 and 8 weeks of age. In the light of the findings of this Japanese study,
vaccination against P. multocida can probably be started after the calves have attained an age
of 4 weeks.
Nadeem et al. (2010) conducted a study to determine the effect of prophylactic
application of levamisole in hemorrhagic septicemia (HS) vaccinated buffalo calves. A total
of 60, 5 month old buffalo-calves were randomly selected from the Livestock Production
Research Institute Bhadurnagar (LPRI) Okara, Pakistan and divided into 3 groups (G1, G2
and G3) of 20 calves each. Animals in G1 group were vaccinated with 3ml (I/M) injection of
a commercially available oil based HS bacterin (NIAB-HS™) while animals in group G2
received levamisole @ 0.5mg/kg body weight two days prior to vaccination with this
bacterin. Group G3 served as unvaccinated and non-medicated control. IHA based geometric
mean titers (GMT) against P. multocida and Lymphocytes Proliferation Assay (LPA) were
used as evaluation criteria. Geometric mean titers (GMT) were significantly higher (P>0.05)
in calves of group G2 (21.5) as compared to GMT values (14.79) recorded in calves of group
G1. Similarly, the values (0.317±0.041) of LPA in calves of group G2 were significantly
higher than the corresponding values (0.043±0.002) recorded in calves of group G1. The
values of GMT and LPA in non–vaccinated, non–medicated calves (group G3) were 2.5 and
0.043±0.002, respectively. It was concluded that prophylactic application of levamisole helps
in mounting a better humoral and cellular immune response to P. multocida vaccine.
2.6 Importance of transferrin binding protein A as an immunogen
IROMPs (Iron regulated outer membrane proteins), in particular TbpA (transferrin
binding protein A) has been proposed as a vaccine candidate immunogen for P. haemolytica,
Neisseria meningitides, and Haemophilus influenza. The rationale for targeting TbpA as an
immunogen is three folds (Singh et al., 2011). Firstly, this is one of the important IROMPs
involved in acquisition of iron from the host and is essential for overcoming the iron
restriction imposed by the host iron binding i.e. transferrin. Secondly, it is present at the cell
surface of the bacterial cell and expressed by bacteria when growing inside host body in
response to iron depleted conditions. Its expression is enhanced by incorporation of iron
chelators like 2, 2-dipyridyl in the culture media (Srivastava, 1998). Transferrin binding
protein is one of the important virulence factors of P. multocida. Thus, Veken et al. (1996)

19
reported a positive association of TbpA with HS causing strains of P. multocida in bovine
whereas the strains of non HS serotypes did not express this protein. Singh et al. (2011)
evaluated the potential of TbpA as a DNA vaccine against HS in a mouse model. The TbpA
gene of P. multocida serotype B:2 was cloned in a mammalian expression vector alone and
along with murine IL2 gene as immunological adjuvant to produce monocistronic and
bicistronic DNA vaccine constructs, respectively. The immune response to DNA vaccines
was determined on the basis of serum antibody titers and lymphocyte proliferation assay.
Mice vaccinated with DNA vaccines showed significantly higher antibody titers and cell
mediated immune response than the unvaccinated mice. The bicistronic DNA vaccine
elicited a superior immune response and protection following challenge as compared to
monocistronic construct. These investigators concluded that DNA vaccine offers the promise
of an effective immunological approach for the control of HS.
2.7 Montanide® adjuvanted HS vaccines
In a bid to develop a stable, easily injectable HS vaccine with a long duration of
immunity, Pakistani workers (Muneer and Afzal, 1989) prepared and evaluated two oil-
adjuvant vaccines viz. water-in-oil emulsion (final product incorporating Marcol 52,
Montanide® 103 and antigen in ratios of 6:1:3) and double emulsion (Marcol 52, Arlacel A
and Tween 80 plus antigen) of P. multocida Robert's type 1. These preparations were
evaluated on only 10 buffalo calves aged ≥ 4 months which showed sustained antibody
response throughout the entire duration of experiment (=240 days). The pitfall of small
number of test subjects (buffalo calves) in this study were addressed by Muneer et al. (1994)
by evaluating three oil adjuvant vaccines of P. multocida 6: B. in terms of level and duration
of antibody levels in buffalo calves (n=45). The three vaccines evaluated were (a) water-in-
oil emulsion (= single emulsion; OAV1) containing Marcol 52, Montanide®
888 and antigen
incorporated at a ratio of 6:1:3, (b) double emulsion (OAV2) formulated to contain Marcol
52 (63%), Arlacel A (7%) and Tween 80 (1.5%) + antigen (28.5%), (c) vitamin E adjuvant
vaccine (OAV3) incorporating Vit. E (62%), Montanide® 888 (8%) and antigen (30%).
ELISA-based P. multocida antibody titers elicited by these vaccines remained substantially
higher than those recorded by using traditional alum precipitated bacterin following
sensitizing and booster vaccination spaced 2 months apart. All three vaccines induced
immune response in buffalo calves that persisted beyond 270 days post-vaccination. On
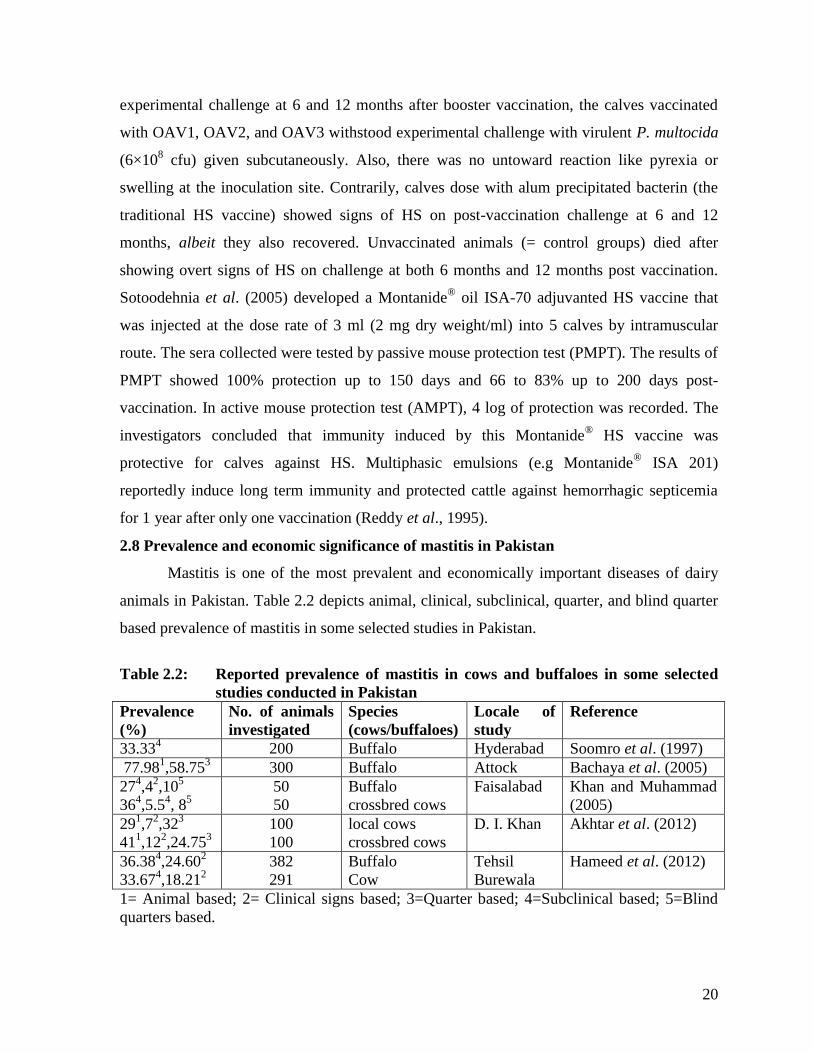
20
experimental challenge at 6 and 12 months after booster vaccination, the calves vaccinated
with OAV1, OAV2, and OAV3 withstood experimental challenge with virulent P. multocida
(6×108 cfu) given subcutaneously. Also, there was no untoward reaction like pyrexia or
swelling at the inoculation site. Contrarily, calves dose with alum precipitated bacterin (the
traditional HS vaccine) showed signs of HS on post-vaccination challenge at 6 and 12
months, albeit they also recovered. Unvaccinated animals (= control groups) died after
showing overt signs of HS on challenge at both 6 months and 12 months post vaccination.
Sotoodehnia et al. (2005) developed a Montanide® oil ISA-70 adjuvanted HS vaccine that
was injected at the dose rate of 3 ml (2 mg dry weight/ml) into 5 calves by intramuscular
route. The sera collected were tested by passive mouse protection test (PMPT). The results of
PMPT showed 100% protection up to 150 days and 66 to 83% up to 200 days post-
vaccination. In active mouse protection test (AMPT), 4 log of protection was recorded. The
investigators concluded that immunity induced by this Montanide®
HS vaccine was
protective for calves against HS. Multiphasic emulsions (e.g Montanide®
ISA 201)
reportedly induce long term immunity and protected cattle against hemorrhagic septicemia
for 1 year after only one vaccination (Reddy et al., 1995).
2.8 Prevalence and economic significance of mastitis in Pakistan
Mastitis is one of the most prevalent and economically important diseases of dairy
animals in Pakistan. Table 2.2 depicts animal, clinical, subclinical, quarter, and blind quarter
based prevalence of mastitis in some selected studies in Pakistan.
Table 2.2: Reported prevalence of mastitis in cows and buffaloes in some selected
studies conducted in Pakistan
Prevalence
(%)
No. of animals
investigated
Species
(cows/buffaloes)
Locale of
study
Reference
33.334
200 Buffalo Hyderabad Soomro et al. (1997)
77.981,58.75
3 300 Buffalo Attock Bachaya et al. (2005)
274,4
2,10
5
364,5.5
4, 8
5 50
50
Buffalo
crossbred cows
Faisalabad Khan and Muhammad
(2005)
291,7
2,32
3
411,12
2,24.75
3 100
100
local cows
crossbred cows
D. I. Khan Akhtar et al. (2012)
36.384,24.60
2
33.674,18.21
2 382
291
Buffalo
Cow
Tehsil
Burewala
Hameed et al. (2012)
1= Animal based; 2= Clinical signs based; 3=Quarter based; 4=Subclinical based; 5=Blind
quarters based.

21
Hussain et al. (2005) reported the findings of a farmers‟ participatory surveillance
survey of livestock diseases in Islamabad capital territory. They used the technique of
proportional piling to estimate the relative prevalence of livestock diseases in the area. In the
execution of the study, 100 beans (or pebbles in some cases) were dispensed to the farmers
and they were asked to make piles according the relative incidence of 5 most prevalent
diseases. The results indicated that HS, FMD and mastitis were the 3 most common disease
conditions observed by the participating farmers. Gender based stratification of the disease
incidence data revealed that 15.8 and 16.3% of the male and female livestock keepers,
respectively in Islamabad capital territory had noticed occurrence of mastitis in their cows
and buffaloes.
A recently reported study conducted under Pakistan Strategy Support Program of
USAID by Ashfaq et al. (2014 ) estimated and evaluated on the basis of prevalence of main
livestock diseases that effect on diary animal productivity and farm incomes in five tehsils of
district Faisalabad. Three villages were selected from each tehsil and ten livestock farmers
from each village picked randomly to collect information. Farmers categories into three on
the basis of adult cows and buffaloes owned were as follows: small (1-3 animals), medium
(4-6 animals), and large farmers (more than 6 animals). The study focused on the negative
impact on milk production and farm incomes due to mastitis, parturient hemoglobinuria,
FMD, and tick infestation. The morbidity or mortality rate, incidence rate, and case fatality
rates of each disease were determined. The economic losses related with these diseases were
appraised. In addition, economic returns associated with controlling these 4 diseases were
calculated in the form of benefit-cost ratios. Overall, 9.01% of total economic losses
associated with these 4 diseases were caused by mastitis. Small, medium and large farmers,
respectively sustained 14.09, 8.34 and 9.59% of the total economic losses caused by 4
diseases selected for survey information. Riaz et al. (1992) conducted an active surveillance
from September 1989 to August 1990 in district Gujrat, Pakistan. A total of 1025 farmers
were interviewed. Mastitis incidence of 4.04% in cattle and 5.48% in buffaloes was recorded.
A meta-analysis of investigations conducted over a one year period (1994-1995) on
economically important diseases of livestock in Punjab (Pakistan) ranked mastitis at the 5th
slot, accounting for 8% of the total pecuniary losses inflicted by all livestock diseases in this
providence (Khan et al., 1994-1995).

22
2.9 Role of Staphylococcus aureus and Streptococcus agalactiae in mastitis
In countries lacking application of mastitis control programs, contagious organisms
(S. aureus and Str. agalactiae) are the two most predominant mastitis pathogens (Pankaj et
al., 2013). Collectively, these two pathogens are responsible for nearly 75% cases of this
disease in cows and buffaloes (Allore, 1993). Pakistani worker (Shakoor, 2006) while
reviewing the reported prevalence of pathogens associated with mastitis in Pakistani studies
(n=16, conducted between 1966-2002) concluded that S. aureus was the most prevalent
mastitis pathogen in buffaloes and cows followed by Str. agalactiae , E. coli, and mixed
infections.
2.10 Mastitis vaccines
High prevalence, poor response to antibiotics, development of antibiotic resistance
and consumers concern about residues of antibiotics/antibacterials in milk and meat of the
treated animals have spurred interest in the control of mastitis through vaccination
(Tollersrud, 2001). The idea of use of mastitis vaccine for control and treatment is not new.
According to Wooldridge, (1923), vaccines of various kinds e.g. stock vaccines containing a
mixture of streptococci, staphylococci, and Bacterium pyogenes, were in wide use in 1920s
to prevent or treat bovine mastitis. This author also stated that in herds in which there has
been a series of clinical cases, favorable results have been reported from the use of so called
stall-specific vaccines (autogenous vaccines).The production of autogenous mastitis vaccines
entails isolation of the organism (s) responsible for the outbreak from several cows, its mass
cultivation followed by treatment of the whole of the milking herd with this vaccine made
therewith. In order to prevent outbreaks, it was recommended that doses of dead vaccine
should be given during the dry period. In the subsequent period, investigations by French
(Richou and Thienlin., 1955), and English workers Derbyshire et al. (1961), demonstrated
that use of staphylococcal mastitis vaccines had a definite increase in immunity against
certain strains of staphylococci as reflected by a decrease in the spread of this infection
coupled by a clear reduction of the number of acute cases of mastitis. An equal effect against
all strains of staphylococci was, however, not noticed. Notwithstanding this limited success,
as emphasized by Derbyshire et al. 1961) further research into mastitis vaccinology is clearly
warranted. (Heidrich and Renk, 1967). During the past 3 decades, a variety of mastitis

23
vaccines were investigated and a few of these are now commercially available (Ahmad,
2009; Pereira et al., 2011; Rashid, 2011). Monovalent and polyvalent mastitis vaccines
developed and evaluated at the Mastitis Research Lab. Department of Clinical Medicine and
Surgery, University of Agriculture, Faisalabad, Pakistan were found effective in mastitis
control and treatment (Shakoor, 2006; Athar, 2007; Yousaf et al., 2009; Ahmad, 2009;
Rashid, 2011). Recently, Leitner et al. (2011) reported the efficacy and safety of a
recombinant Montanide® ISA 206 S. aureus mastitis vaccine in dairy cows. This vaccine
was based on Target of RNAIII Activating Protein (TRAP), which is a membrane associated
167AA protein constitutively expressed in all strains of S. aureus and coagulase negative
staphylococci. This recombinant TRAP (rTRAP) S. aureus mastitis vaccine was safe,
immunogenic and elicited a humoral immune response that remained high for at least 160
days post booster shot.
2.10.1 Montanide® (Seppic, France) adjuvanted mastitis vaccines
Montanide® ISA are fairly a new generation of ready-to-use adjuvants which allow
the manufacture of different type of emulsions: water in oil (W/O), oil in water (O/W), or
water in oil in water (W/O/W). They can be based on mineral oil, non-mineral oil or their
mixture. Adjuvants designed for veterinary and human vaccines were reviewed by
Aucouturier et al. (2001). According to these authors, W/O/W (e.g. Montanide® ISA 201
VG, a multiphasic emulsions) can induce long and short term immune response. Low
viscosity (thus an easy syringibility) is another desirable attribute of this class of Montanide®.
After vaccination, the antigen in the external water phase is immediately available to the
immune system like aqueous formulations while antigen entrapped in the internal aqueous
phase is slowly released like W/O emulsions. Water in oil in water (W/O/W) emulsions elicit
superior immune response as compared with aluminium hydroxide in cattle. They are well
tolerated and induce a strong stimulation of cellular immune response. According to
Aucouturier et al. (2001) screening studies in mice with viral, bacterial or parasitic antigens
indicated that W/O/W emulsions induce higher IgG2a antibody levels than other type of
emulsions. IgG2 is the most desirable antibody for protection against S. aureus mastitis
(Watson, 1975) because neutrophils have receptors for this antibody. Tollersrud et al. (2002)
reported that S. aureus capsular polysaccharide type expressed on the surface of
formaldehyde inactivated whole cells emulsified in the Montanide® gave a strong and long

24
lasting immune response. Tollersrud, (2001) reported that Montanide® ISA-70 enhances a
stronger immune response to α, β-toxins than carbopol vaccine tested in sheep. Yousaf et al.
(2009) evaluated the effect of a Montanide®
ISA 206 adjuvanted S. aureus bacterin-toxoid on
the prevalence and incidence of mastitis in cows. A total of 60 California Mastitis Test and
surf field mastitis test negative cows in their first and second month of lactation were
selected and assigned randomly to 3 groups (C1, C2, and C3). Montanide® ISA-206
adjuvanted S. aureus bacterin –toxoid (in 5 ml doses) was injected I.M twice with 4 weeks
spacing to animals of group C1 and C2. Levamisole (2.5mg/kg body weight) was
administered per os to cows of group C2 after first and second shots of vaccine. Cows in
group C3 served as control (non -vaccinated and non- medicated with levamisole).
Prevalence and incidence rates of mastitis were determined. The results showed that
maximum number of quarters were found positive in group C3 with maximum cumulative
prevalence (27.5%) while minimum cumulative prevalence (11.25%) was noted in group C2
followed by group C1 (13.75 %). There was no significant variation in cumulative incidence
between group C1 and C2. Montanide® ISA-25 and ISA-206 (Seppic, France) as adjuvants in
cattle and pigs are much better than conventional aqueous formulations to retain potency and
to enhance the antibody response (Barnett et al., 1996). No toxicity (even after booster dose)
by the use of these vaccines can be seen (Castrucci et al., 1993). It is reported that
Montanide® adjuvanted vaccine is less viscous, less irritant and easily injected than
conventional Freund‟s adjuvanted vaccine (Cook et al., 1990).The Montanide® adjuvanted
vaccine elicited superior and rapid immune response at any time period which was
maintained for longer time ( Patil et al., 2002). Ahmad and Muhammad, (2008) demonstrated
that a locally prepared Staphylococcus aureus and Streptococcus agalactiae aluminium
hydroxide adjuvanted mastitis vaccine was immunogenic when tested in rabbits.
2.11 Need for a combined HS–Mastitis vaccine (‘combo’ or ‘cocktail’ HS –mastitis
vaccine)
Combined vaccines („Combo vaccines‟) are vaccines containing combination of two
or more vaccinal antigens from different pathogens blended in one vaccine vial/bottle. These
vaccines usually target more than one disease. The use of combination vaccines is a
pragmatic strategy to overcome the constraints and problems associated with multiple

25
injections of individual vaccines. Combined vaccines containing bivalent, trivalent as well as
polyvalent microorganisms (bacterial and /or viral) including P. multocida have been
developed to reduce the number of vaccine doses and reduce the cost of production as well
as to expand the spectrum of protective immunity against multiple infections (Verma and
Jaiswal,1998; De Alwis,1999). Combined vaccines targeting HS, black quarter, FMD,
infectious bovine rhinotracheitis, adeno virus type-3 parainfluenza and some other
respiratory pathogens have been developed and tested.(Srivastava et al., 1975; Osman et al.,
1990).
The use of combined vaccines lessens the cost for extra health care visits, facilitates the
addition of new vaccines and diseases into immunization programs (CDC, 2011). Geering
and Lubroth, (2002) opined that during visits by FMD vaccination teams to livestock farming
communities, it may be desirable to combine FMD vaccination with vaccination against
other diseases (e.g. HS, contagious bovine pleuropneumonia, sheep and goat pox). Combined
vaccination not only conserves resources but is also helpful in achieving a higher level of
farmer cooperation in the FMD control program. According to Hussain et al. (2005), HS and
mastitis are among the 5 most important diseases that affect the local animals in Pakistan. An
immunization program based on the use of combined HS- mastitis vaccine may potentially
be associated with the following advantages:
Less frequent handling of the candidate vaccinate cows and buffalos
Inclusion of mastitis in the vaccination program. By and large a „do nothing‟ policy is in
place in Pakistan as far as mastitis control is concerned.
Less cost of vaccination
Efficacy of a bivalent vaccine viz. FMD and HS was compared to individual FMD
and HS vaccines in rabbits by Altaf et al. (2012). The investigators concluded that this
combined vaccine induced higher antibody titers against both FMD and HS than the
individual FMD (prepared from O serotype) and HS vaccine (alum precipitated). Srinivasan
et al. (2001) compared the serological response of cattle to a combined FMD, rabies, HS,
black quarter vaccine to individual component vaccines containing FMD, rabies, P.
multocida, and Clostridium chauvoei antigens. A statistically non- significant variation in the
serological response was elicited by individual component vaccines and combined vaccine

26
containing antigens of all four disease pathogens. As far as could be ascertained from the
available literature, a combined HS-mastitis vaccine has not been evaluated thus far.
According to Chakrabarti, (2002), a combined H.S-black quarter vaccine is available in
India. It is a combination of inactivated antigen of P. multocida type-I and Clostridium
chauvoei. This is stabilized and adjuvanted with BAIF aluminium hydroxide gel aluminox.
The manufacturer claims that this vaccine protects animals against both hemorrhagic
septicemia and black quarter.
In Pakistan, because of extremely small herd size (more than 80% animals kept in
groups of 3-4 animals/family; Jost, 1980; Tuefel, 1998), widely rampant poverty and
illiteracy, lack of any milk quality premium program adoption of the standard mastitis control
practices i.e. post milking antiseptic teat dipping and dry period antibiotic therapy as
advocated by the National Mastitis Council Inc, USA (Nickerson, 1994) is conceivably
difficult. Even on well organized private dairy farms and those in public sector, mastitis
control is not in place. Against this backdrop, vaccination against the common mastitis
pathogens holds the promise of an adjunct/alternative mastitis control strategy (Koiranen,
1977).
2.12 Mastitis in heifers and justification of use of combined HS-mastitis vaccine
Future of a dairy herd hinges to a considerable extent on the milk producing potential
of replacement animals i.e. heifers. Investment in general management and health of the
heifers is reflected in good herd replacements. Managmental and health deficiencies during
the period intervening between calf birth and its first calving can drastically decrease the
milk yield of heifer as she enters the lactating herd. On many farms, calves suffer from the
vice of suckling each other‟s navel and developing udder .The occurrence of subclinical
intramammary infections (IMI) in non-lactating heifers was once considered rare but in
recent years, studies have indicated that the prevalence of IMI in heifers can be as high as
97% in some herds (Trinidad et al., 1990b). These infections are important because
infections acquired during the prepartum period may damage developing secretary tissue,
increase the SCC (Trinidad et al., 1990a; Trinidad et al., 1990b; Nickerson et al., 1995;
Oliver et al., 2003) and reduce lifetime milk production (Oliver et al., 2003, De Vliegher,
2004). Therefore, the control of IMIs in heifers is receiving increased attention.
Staphylococci (primarily coagulase-negative Staphylococci but also Staphylococcus aureus)

27
are the most frequent isolates recovered from heifers but environmental pathogens such as
Streptococci are also frequently recovered (De Vliegher, 2004). Use of a combined HS–
mastitis vaccine incorporating antigens of P. multocida, S. aureus and Str. agalactiae in
heifers may not only potentially reduce the incidence of HS but also reduce the incidence and
prevalence of S. aureus and Str. agalactiae associated mastitis.
Suboptimal immune response to bovine vaccines is an important problem and several
interventions can be used to overcome this problem. Arthington and Havenga, (2011)
demonstrated that an injectable trace minerals supplement (MultiMi®, Fort Collins,
Colorado, USA) containing 15, 40, 10 and 5 mg/ml of CU, Zn, Mn ( all as disodium EDTA
salts),and Se ( as Na selenite) when administered concurrently with a bovine viral vaccine
(Titanium® 5; AgriLabs, St. Joseph, Missouri, USA) containing bovine herpes virus
(BHV1),bovine viral diarrhea virus (BVDV1), BVDV-2, bovine parainfluenza virus type
3,and bovine respiratory syncytial virus did not impair humoral immune responses in beef
calves. In addition, concurrent administration of injectable trace minerals supplement and
BHV-1 vaccine enhanced the production of neutralizing antibodies to BHV-1 in previously
naïve beef calves. No such investigation seems to have been conducted on hemorrhagic
septicemia vaccine.

28
CHAPTER 3
MATERIALS AND METHODS
The present study is the first-ever attempt towards development and evaluation of a
Montanide® adjuvanted combined hemorrhagic septicemia -mastitis vaccine in dairy
buffaloes and cattle. It was executed keeping in view the animal welfare concepts of 3R‟s.
The objectives of the study were pursued by compartmentalizing the entire work into
following 2 phases
Phase I: Preparation and evaluation of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine in laboratory settings
Phase II: Field evaluation of Montanide® adjuvanted combined HS-Mastitis vaccine in
target dairy species
Phase I: Preparation and Evaluation of Montanide® Adjuvanted
Combined Hemorrhagic Septicemia-Mastitis Vaccine in
Laboratory Settings
3.1 Collection and culture of blood samples for isolation of P. multocida
3.1.1 Sources and types of samples for isolation of P. multocida
Peripheral blood samples of 5 buffalo calves and 2 adult buffaloes suffering from HS
were collected aseptically and processed for the isolation of P. multocida as per OIE, 2012.
Briefly, in each case, 0.2 ml of collected blood sample was eluted into 2 ml sterile normal
saline and an aliquot was cultured on casein-sucrose-yeast (CSY) agar plate containing 5%
calf blood and blood agar plates within 3 hours of collection. The inoculated plates were
incubated at 370C for 48 hours. The composition and method of preparation of casein-
sucrose-yeast (CSY) agar is given in the Appendix- I.
Isolation of P. multocida was also attempted from the femur of 4 buffaloes that died
of HS during the past 48-72 hours. To this end, the femur of the carcass was exposed with the
help of a necropsy knife, followed by swabbing with methyl alcohol, split opening of the
bone and aseptic aspiration of approximately 2 grams of the bone marrow. The bone marrow
sample was admixed in 10 ml of sterile normal saline in a sterile caped test tube. The tube
containing bone marrow–normal saline admixture was vigorously shaken for 2 minutes.

29
Finally, 0.1 ml aliquots of bone marrow-normal saline admixture were cultured on casein-
sucrose-yeast (CSY) agar plates containing 5% calf blood (OIE, 2012; Ali et al., 2000) and
blood agar plates.
In addition to peripheral blood samples and femur marrow samples, isolation of P.
multocida was also attempted from the heart of 4 additional buffalo calves immediately after
their slaughtering prompted by their moribund condition with overt clinical HS (De Alwis,
1992). Following slaughter, the heart was removed, washed with clean water, placed in a
sterile tray, and swabbed thoroughly with 70% alcohol. A sterile 18 gauge needle attached to
a disposable sterile 20 ml syringe was pierced into the left ventricle to withdraw about 5 ml
of blood that was mixed with 5 ml of sterile normal saline. Heart blood- sterile normal saline
mixture was dispensed into 10 ml of enrichment transport medium (BHI broth) for
transportation to the Mastitis Research Lab. Department of Clinical Medicine & Surgery,
University of Agriculture, Faisalabad. Immediately after arrival at this Lab., 0.1 ml aliquots
of heart blood –normal saline –transport medium were cultured on casein-sucrose-yeast
(CSY) agar plates containing 5% calf blood and blood agar plates. In addition to aspirated
heart blood samples, swab samples of heart blood were collected by drenching sterile swab
(IMPROSWAB™, Microbiological Transport Swab, IMPROVE®, Guangzhou Improve
Medical Instruments Co., Ltd.) with heart blood (OIE, 2012). The swabs were cultured on
casein-sucrose-yeast (CSY) agar plates and blood agar plates.
3.1.2 Isolation and biochemical and molecular confirmation of P. multocida
The following features were considered for identification of P. multocida. It forms
smooth grayish glistening translucent colonies of nearly 1 mm diameter on blood agar after
24 hours incubation at 37°C. It did not grow on MacConkey‟s agar. On Gram staining, the
organism appeared as Gram –ve, short, ovoid bipolar staining coccoid with pleomorphism
(OIE, 2012). Further confirmation of the organism and type (i.e., group B) was done by
biochemical reactions (Butt et al., 2003; Ashraf et al., 2011; Ekundayo et al.,2008) and by
using PCR primers (Townsend et al., 1998; OIE, 2012).

30
3.1.3 Synthetic oligonucleotide primers
Table 3.1: The oligonucleotide primer pair used for PCR based confirmation of
Pasteurella multocida (Townsend et al., 1998; OIE, 2012)
PCR amplification using the aforementioned primers was carried out as described by
Townsend et al. (1998). Primers (KTT72 and KTS P61) were got synthesized from Gene
Link, USA. These primers specifically produce products of approximately 620 bp from HS-
causing type B isolates of P. multocida.
3.1.4 PCR Reaction
Suspect colonies were sub-cultured and subjected to biochemical testing (Butt et al.,
2003; Ashraf et al., 2011). After confirmation, the colony was streaked onto sheep blood
agar plate. Following overnight incubation at 37 °C, a single colony was picked to run the
PCR reaction. PCR was carried out in a 50µl reaction volume using Techne Touchgene
Gradient PCR Thermal Cycler (Bibby Scientific Limited, UK). Single bacterial colony was
added to 48µl of PCR reaction mixture consisting of: 35µl DD-water; 5µl, 10XPCR buffer;
5µl MgCl2 (25mM), 0.2µl TaqDNA polymerase (5U/µl), 1µl deoxynucleotide triphosphate
(dNTP) mix (10mM each), 1µl Forward primer (25 p moles/µl), 1 µl Reverse primer (25 p
moles/µl). Cycling profile used for PCR reaction was as under: The program used for
diagnosis of P. multocida with primer pair KTT72/ KTS P61: 94 °C for 5 min (1 cycle); 95
°C for 1 min., 55 °C for 1 min, 72 °C for 1 min (30 cycles); 72 °C for10 min (1 cycle). PCR
reaction product were subjected to gel electrophoresis (1.5 agarose gel, 85 volts for 20
minutes), stained by Ethidium bromide and visualized by gel doc system (Bio-Rad, USA).
3.1.5 Pathogenicity (virulence) of 3 candidate vaccinal isolates of P. multocida
LD50 (virulence) of candidate vaccinal P. multocida isolates (n= 3) was determined in
six weeks old Swiss albino mice following the procedure described by Butt et al. (2003). One
Primer Sequence Product Size Primer Specificity
KTT72 5‟-GGCTCGTTTGGATTAGTAAG-3‟
618 bp
HS-causing type-B-specific
KTS P61 5‟-ATCCGCTAACACACTCTC-3‟

31
tenth of an ml (i.e. 0.1 ml) of each tenfold dilution (10-1
to 10-10
in sterile normal saline) of 18
hours old cultures grown in 1% nutrient broth (Merck™
, Darmstadt, Germany) was
inoculated intraperitoneally in mice using 10 mice for each dilution. A set of another 10
mice, for each dilution was kept as control and injected with sterile normal saline. Mortality
and time of death associated with each dilution was recorded to calculate LD50. Blood smears
prepared from the cardiac blood of dead mice were stained with Giemsa‟s stain. In addition,
the cardiac blood samples of dead mice were cultured on blood agar and CYS agar plates for
reisolation of P. multocida (Naz et al., 2012).
3.2 Sources of milk samples, isolation of candidate vaccinal S. aureus
and Str. agalactiae isolates and their partial characterization
Milk samples from 37 quarters of cows (n=10) and buffaloes (n=9) managed at the
Livestock Experimental Station, University of Agriculture, Faisalabad, Ayub Agriculture
Research Institute, Faisalabad, Raja Asghar dairy farm, Mehar Anayat dairy farm, Raja wala
and Bilal Rashid dairy farm, and suffering from peracute/acute clinical mastitis (Radostitset
al., 2007; Schalm et al., 1971) were sampled aseptically following the procedures described
by Hogan et al. (1999). Immediately after collection, milk samples were placed on crushed
ice in a thermos chest (Coleman®; the Coleman Company Ltd, USA.) and transported to the
Mastitis Research Lab. Department of Clinical Medicine and Surgery, University of
Agriculture, Faisalabad. On arrival at this lab, using a platinum-rhodium inoculation loop,
0.01 ml (i.e. one hundredth of a milliliter) of each milk sample was inoculated onto blood
agar (Oxoid Ltd., Basingstoke, Hampshire, England) plate containing 5% defibrinated ovine
blood and MacConkey‟s agar plate (Hogan et al., 1999; National Mastitis Council, Inc. USA,
1987; Claxton and Ryan, 1993). Four milk samples were cultured on each agar plate of 100
mm diameter. After inoculation of milk samples, blood agar and MacConkey‟s agar plates
were incubated aerobically at 37ºC for 48 hours, followed by examination of the growth (if
present) by Gram staining, catalase reaction and other relevant tests (Appendix II and
Appendix III). β hemolytic, Gram positive, catalase positive coccal isolates that grew on
blood agar plates were presumptively identified as staphylococci and subjected to tube
coagulase test. (Hogan et al., 1999; National Mastitis Council, Inc. USA, 1987). Briefly, a
loop-full of growth of each isolate presumptively identified as a Staphylococcus was

32
inoculated into 0.5 ml of rabbit plasma diluted 1:1 with sterile normal saline (Appendix IV)
and incubated at 37oC for 24 hours. Staphylococcal isolates with positive tube coagulase test
at 4 hours were provisionally identified as S. aureus, whereas staphylococcal isolates
negative in tube coagulase test till 24 hours were categorized as coagulase negative
staphylococci (National Mastitis Council, Inc. USA, 1987; National Mastitis Council, Inc.
USA, 1990). Four selected S. aureus isolates that displayed wide hemolytic zones on blood
agar plates (candidate vaccinal isolates) were biotyped using a commercially available
Staphylococcus kit (API® STAPH; bioMerieux, France). Latex Slide Agglutination test was
performed using Staphytect Plus® kit (Oxoid Ltd, Basingstoke, UK) for the determination of
clumping factor, protein A and certain polysaccharides found only in S. aureus (Rashid,
2011; Raza, 2012).
Gram positive, catalase-negative, esculin-negative, CAMP- positive (Appendix VI),
and β-hemolytic cocci (i.e. clear hemolysis) were considered as isolates of Streptococcus
agalactiae (Altwegg and Bockemühl, 1998). Blood agar with 0.1% esculine and 0.01%
ferric citrate was used for testing the isolates for CAMP reaction (National Mastitis Council,
Inc. USA, 1990). Three isolates of Streptococcus agalactiae were biotyped using a
commercially available kit of genus Streptococcus (API® 20-STREP kit bioMerieux,
France). Isolates other than those of Staphylococcus and Streptococcus that grew on blood
agar plates were not further processed.
3.2.1 Molecular confirmation of S. aureus
3.2.1.1 Detection of nuc gene by polymerase chain reaction (PCR)
The vaccinal isolate of S. aureus was confirmed by PCR by targeting ‘nuc’ a
chromosomal gene encoding extracellular thermo-stable nuclease activity (a specific target
for S. aureus; Brakstad et al., 1992), using a commercially available Multiplex PCR kit
(BactReadyTM
Multiplex PCR system; Genescript, USA). Sequence of forward and reverse
primer used and size of PCR product have been mentioned in Table 3.2. All primers were
purchased from GeneLinkTM
(USA) through a local vendor (The Worldwide Scientific,
Lahore, Pakistan).

33
Table 3.2: Sequence of nuc gene primers used for confirmation of S. aureus and PCR
product size
Targeted
genes
Sequence Product
Size
Reference
nuc forward
nuc reverse
Nuc 5‟- GCG ATT GAT GGT GAT ACG GTT-3‟
Nuc 5‟-AGC CAA GCC TTG ACG AAC TAA AGC-3
279bp Brakstad et al.
(1992)
3.2.1.2 Preparation of genomic DNA from S. aureus
For PCR, genomic DNA was prepared using extraction buffer (BR-A buffer)
available with BactReady Multiplex PCR system (Genescript, USA). Briefly, single, isolated,
18-hour old colony from blood agar plate was picked and suspended in 25 µl of DNase free
water and 1µl of cell suspension was mixed with 20 µl of BR-A/extraction buffer. The lysate
was spun down by short centrifugation (2000 rpm for 1 minute) and the supernatant
containing gDNA was used directly for amplification.
3.2.1.3 PCR amplification
The reaction was performed using BactReadyTM
Multiplex PCR system following the
manufacturer instructions. The reaction mixture (20 μl) was prepared in thin walled with flat
cape, DNase-RNase free 0.2 ml thermo-tubes (Thermo-scientific, UK) with 1 μl of template
DNA. Amplifications were performed using a micro-processed controlled SwiftTM Maxi
Thermal Cycler Block (ESCO Technologies Inc. France) under the following conditions:
activation of ScriptTM DNA polymerase at 94 ºC for 15 minutes, followed by 35 cycles of
denaturation (95ºC for 1 min), annealing (55ºC for 1 min), extension (72ºC for 1 min), and a
final extension step at 72ºC for 3 minutes. The composition of reaction mixture is given in
the Table 3.3. The amplicons were analyzed by agrose gel electrophoresis using a horizontal
mini agarose gel electrophoresis system (ENDUROTM Labnet International Inc.,
Woodbridge, New Jersey, USA). A mixture of undiluted PCR products (5 μl) and 5X loading
dye (1 μl; Fermentas Thermo Fischer Scientific Inc., UK) was loaded to 1.2% agrose gel
(multipurpose agarose, low EEO, multipurpose, Fischer Scientific Ltd, Loughborough, UK)
containing ethidium bromide (0.5 μg./ml; Fischer Scientific Ltd, Loughborough, UK) for
DNA staining. The gels were run in 1X TAE buffer (50X TAE Buffer, Fischer Scientific Ltd,
Loughborough, UK) at 70 volts (80mA) for 1 hour. The amplicons were visualized on a
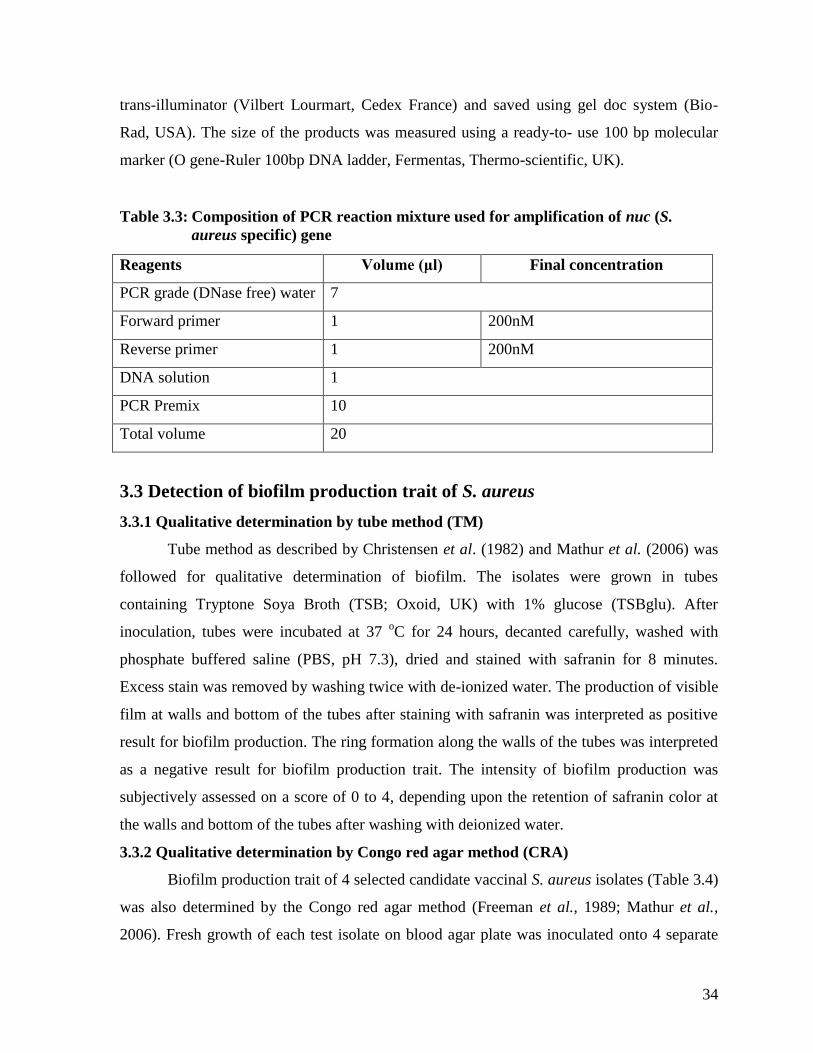
34
trans-illuminator (Vilbert Lourmart, Cedex France) and saved using gel doc system (Bio-
Rad, USA). The size of the products was measured using a ready-to- use 100 bp molecular
marker (O gene-Ruler 100bp DNA ladder, Fermentas, Thermo-scientific, UK).
Table 3.3: Composition of PCR reaction mixture used for amplification of nuc (S.
aureus specific) gene
Reagents Volume (μl) Final concentration
PCR grade (DNase free) water 7
Forward primer 1 200nM
Reverse primer 1 200nM
DNA solution 1
PCR Premix 10
Total volume 20
3.3 Detection of biofilm production trait of S. aureus
3.3.1 Qualitative determination by tube method (TM)
Tube method as described by Christensen et al. (1982) and Mathur et al. (2006) was
followed for qualitative determination of biofilm. The isolates were grown in tubes
containing Tryptone Soya Broth (TSB; Oxoid, UK) with 1% glucose (TSBglu). After
inoculation, tubes were incubated at 37 oC for 24 hours, decanted carefully, washed with
phosphate buffered saline (PBS, pH 7.3), dried and stained with safranin for 8 minutes.
Excess stain was removed by washing twice with de-ionized water. The production of visible
film at walls and bottom of the tubes after staining with safranin was interpreted as positive
result for biofilm production. The ring formation along the walls of the tubes was interpreted
as a negative result for biofilm production trait. The intensity of biofilm production was
subjectively assessed on a score of 0 to 4, depending upon the retention of safranin color at
the walls and bottom of the tubes after washing with deionized water.
3.3.2 Qualitative determination by Congo red agar method (CRA)
Biofilm production trait of 4 selected candidate vaccinal S. aureus isolates (Table 3.4)
was also determined by the Congo red agar method (Freeman et al., 1989; Mathur et al.,
2006). Fresh growth of each test isolate on blood agar plate was inoculated onto 4 separate

35
Congo red agar (CRA) plates and incubated for 24 hours at 37oC. Interpretation of biofilm
production by this method was as follows (Mathur et al., 2006).
Positive result: Appearance of black colored colonies having dry crystalline
consistency.
Weak biofilm production: Pink colonies with no dry crystalline consistency (although
occasionally darkening at the center).
Intermediate biofilm production: Darkening of the colonies with the absence of a dry
crystalline colonial morphology.
Table. 3.4: Composition and preparation of Congo red agar medium (Mathur et al.,
2006)
Ingredients Quantity (g/l)
Brain heart infusion broth (BHI, Oxoid, UK) 37 g
Congo red stain (BDH Chemical Ltd, Poole, England) 0.8 g
Sucrose 50 g
Agar technical (Oxoid, UK) 10 g
Congo red stain was dissolved in distilled water, followed by sterilization at 121oC
for 15 minutes. Mixture of brain heart infusion broth (BHI), sucrose and agar technical was
sterilized in a separate container at 1210C for 15 minutes and cooled to approximately 55
0C
before mixing it with autoclaved Congo red stain. The final mixture was then quickly
dispensed into sterilized Petri plates of 100 mm diameter.
3.3.3 Quantitative determination of biofilm production by spectrophotometric method
Quantitative evaluation of biofilm production trait of 4 candidate vaccinal
staphylococcal isolates was performed spectrophotometerically on flat bottom 96 well
ELISA plate (Greiner® Germany) by the method of Mathur et al. (2006) modified by Rashid
(2011). Each well of the plate was filled with 5 μl culture broth (i.e. Tryptone Soya Broth
with 1% glucose inoculated with the test isolate) and 95 μl of fresh Tryptone Soya Broth
(TSB; Oxoid, UK) with 1% glucose (TSBglu). As negative controls, 3 wells were filled with
100 μl of TSBglu and 2 wells with 100 μl of de-ionized water to check the nonspecific
binding of the culture medium. Plates were incubated for 18 hours at 37oC. Following
incubation, the contents of the plates were discarded by sharply striking the inverted ELISA

36
plate against the top of a disposal tray. Using a multichannel micro-dispenser, each well was
washed 4 times with 0.3 ml of phosphate buffered saline (PBS; pH 7.2). Biofilms formed by
adherent „sessile‟ organisms in plates were fixed with sodium acetate (2%) and stained for 10
minutes with crystal violet (0.1% w/v). Excess stain was removed by thorough washing with
de-ionized water and plates were left for drying. It was noticed that adherent staphylococcal
cells formed biofilm on all sides of wells and were stained uniformly with crystal violet.
Optical density (OD) of wells of ELISA plate was determined by ELISA auto reader (BioTek
Instruments, Highland Park, USA) at 570 nm (OD 570 nm) wavelength. These OD values
were considered as an index of adherence of bacteria to the surface and reflected the intensity
of biofilm production. Experiment was performed in duplicate and repeated two times. The
average was calculated both from cultured wells and negative control wells. In order to
recompense for background absorbance, OD readings from sterile culture medium (Tryptone
Soya Broth), fixative (sodium acetate 2%) and dye (crystal violet 0.1% w/v) were averaged
and subtracted from all individual test values. The mean OD value obtained from media
control well was subtracted from all the test OD values (Mathur et al., 2006; Rashid, 2011).
3.3.4 Pathogenicity of 4 candidate vaccinal isolates of S. aureus
Pathogenicity of candidate vaccinal isolates of S. aureus (n= 4) was determined in
rabbits by subcutaneous inoculation of 0.2 ml of bacterial suspensions (1x1010
cells / ml)
grown in nutrient broth. Suspension of each isolate was inoculated separately in 5 rabbits.
The morbidity/mortality was observed for up to 48 hours post inoculation (Shakoor et al.,
2006; Ahmad, 2009; Hirsch and Strauss, 1964).
3.3.5 Pathogenicity of 2 candidate vaccinal isolates of Str. agalactiae
Pathogenicity of candidate vaccinal isolates of Str. agalactiae (n= 2) was determined in
rabbits by subcutaneous inoculation of 0.2 ml of bacterial suspensions (1x109
cells / ml)
grown in nutrient broth. Suspension of each isolate was inoculated separately in 5 rabbits.
The morbidity/mortality was observed for up to 48 hours post inoculation (Shakoor et al.,
2006; Ahmad, 2009; Hirsch and Strauss, 1964). Dead rabbits were subjected to necropsy
examination.

37
3.4 Immunogenicity testing of vaccinal isolates of P. multocida, S. aureus
and Str. agalactiae
Immunogenic response of vaccinal isolates was determined in rabbits. To this end, 30
adult male rabbits were randomly divided into 3 equal groups (RaA, RaB and RaC) of 10
animals each. Rabbits in each of the 3 groups were respectively inoculated with 0.2 ml
formalin killed suspensions (1×106 per ml) of P. multocida (n=3), S. aureus (n=4) and Str.
agalactiae (n=2) subcutaneously twice at an interval of 7 days (Athar, 2007). Serum
antibody titers were determined by in-house indirect ELISA. Briefly, each of the vaccinal
isolates was grown on nutrient-whey broth, followed by inactivation with formalin (0.1%).
The growths were washed, sonicated separately (Sajjad-ur-Rehman et al., 2004) and the
supernatants were estimated for total protein contents by using Bradford reagent. ELISA
plates were coated with bicarbonate buffer (Sigma, USA) containing 15 µg protein
(antigen)/ml and blocked with bovine serum albumin (0.1%). Antibodies were detected by
protein G-HRP conjugate (LSI Vet, France). TMB (3, 3‟, 5, 5‟-Tetramethylbenzidine) was
finally used as substrate to detect the conjugation and reaction was stopped with sulphuric
acid (1M H2SO4). Plates were read at 450 nm wave length for optical density.
3.5 Antimicrobial susceptibility testing of candidate bacterial isolates
(Antibiogram)
The susceptibility of 9 bacterial isolates (P. multocida, n= 3; S. aureus, n=4 and Str.
agalactiae, n=2) to 11 different antibiotics was tested by disc diffusion method on Mueller –
Hinton medium (Oxoid Ltd., Basingstoke, Hampshire, England), according to the guidelines
of Clinical Laboratory Standards Institute (CLSI, 2012). For Streptococci, Mueller –Hinton
medium was enriched with 5% ovine blood. The plates were incubated at 37°C for 24 hours
and inhibition zones were measured with the antibacterial zone gauge. Staphylococcus
aureus ATCC (American Type Culture Collections- Rockville, Maryland USA) 25923 was
used as a sensitive quality control.
3.6 Selection of vaccinal P. multocida isolate
From a pool of three candidate vaccinal P. multocida isolates, the isolate No. 1 with a
strong pathogenicity (LD50=10-7.75
) and immunogenicity was selected as a vaccinal isolate for
the production of Montanide® adjuvanted combined HS-Mastitis vaccine.

38
3.7 Selection of vaccinal S. aureus isolate
The selection was based primarily on the level of biofilm production, pathogenicity
and immunogenicity of the candidate isolates. A beta hemolytic S. aureus isolate number 1
with the strongest biofilm production ability in tube method and in spectrophotometric
method was selected as a vaccinal isolate for preparation of montanide® adjuvanted
combined hemorrhagic septicemia-mastitis vaccine (Table 3.5). The selected isolate showed
highest values in tube method and spectrophotometric method but intermediate in CRA. The
selection was dependent on tube method and spectrophotometric methods because CRA was
an additional test. This isolate was strongly pathogenic and immunogenic.
Table 3.5: Selection of vaccinal isolate by biofilm detection methods and other traits
Isolate I.D. Spectrophoto
metric OD
Tube
method
score
Biofilm
production
degree on
Congo red
agar (CRA)
Pattern of
hemolysis
on blood
agar
Pathog
enicity
Immuno
genicity
1* 0.31 4 Strong Beta 100 1.23
2 0.19 2 Intermediate Beta 80 1.11
3 0.17 2 Intermediate Beta 80 1.12
4 0.222 2 Intermediate Beta 60 1.14
* Isolate was selected on the basis of tube method score, spectrophotometric method score
and pattern and intensity of hemolysis. The current designation of this isolate is Mastitis
Research Lab is 2 as it originated from an acutely mastitic cross bred cow managed at the
Livestock Experimental Station, University of Agriculture, Faisalabad.
3.8 Selection of vaccinal Str. agalactiae isolate
From a pool of three candidate vaccinal Str. agalactiae isolates, the isolate (SA-100)
with strong pathogenicity and immunogenicity and displaying API® 20 STREP biochemical
profile of 3462414 was selected as a vaccinal isolate for the production of Montanide®
adjuvanted combined HS-Mastitis vaccine. This isolate was recovered from a cow suffering
from periodic flare ups of clinical mastitis. The cow belonged to Livestock Experimental
Station, University of Agriculture, Faisalabad.

39
3.9 Preparation of Montanide® adjuvanted combined HS-Mastitis
vaccine
3.9.1 Preparation of antigen of P. multocida
Procedure adopted by Nuclear Institute of Agriculture and Biology (NIAB)
Faisalabad (OIE, 2012) for production of NIAB HS vaccine was used. A single colony of P.
multocida was picked from nutrient agar plate, inoculated into 50 ml of CSY broth and
incubated for 24 hours at 37 °C. One liter of CSY broth was inoculated with 12.5 ml of P.
multocida pure culture, and incubated at 37°C for 36 hours with constant agitation. The
bacterial sediment (from one liter broth) was harvested by centrifugation at 7000 rpm for 20
minutes. The bacterial sediment was dissolved in 50 ml normal saline and the bacterial
suspension inactivated by addition of 0.4 % of 37 % formaldehyde. The inactivated bacterial
suspension was placed at 37 °C for 24 hours. The sterility of this suspension was checked by
inoculation onto blood agar, nutrient agar, MacConkey‟s agar and citrate agar plates at 37°C.
The inactivated bacterial sediment was harvested by centrifugation and supernatant
discarded. Dilution factor was determined to attain a transmittance of 20% with the help of
spectrophotometer at 540 nm. Normal saline was added to whole bacterial sediment to attain
20% transmittance. The resulting final solution was designated as prepared antigen and was
used for vaccine preparation.
3.9.2 Preparation of antigens of S. aureus and Str. agalactiae
The selected vaccinal isolates of S. aureus and Str. agalactiae were grown
independently in modified nutrient broth (nutrient broth supplemented with 10% sterile
bubaline whey) at 37 ºC for 48 hours on an orbital shaker set at 60 rpm under aerobic
conditions. The suspensions were confirmed for culture purity by Gram‟s staining technique
and inoculation onto blood agar and MacConkey‟s agar plates. Pure cultures of S. aureus and
Str. agalactiae in broth were inactivated by addition of formalin (0.4% V/V) and then
centrifuged at 6000xg for 1 hour at 4°C and resuspended in PBS (pH 7.2). Bacterial
concentrations of each of the vaccinal organisms were adjusted to 1× 109
cells/ml
spectrophotometrically (Hirsch and Strauss, 1964). For incorporation of crude extract of S.
aureus and Str. agalactiae, supernatant fluid collected from 48-hours old broth cultures of
respective isolate were autoclaved at 121 °C for 20 minutes. The autoclaved supernatant was
added to the vaccine preparation at the concentration of 5 mg of dry weight per dose (Athar

40
et al., 2007).
3.9.3 Preparation of combined HS-Mastitis vaccine
Montanide® adjuvanted bacterin-toxoid was prepared by adding Montanide® ISA
201 VG (Seppic, France) in 50/50 (W/W) ratio to a suspension of plain bacterins of S.
aureus, Str. agalactiae, P. multocida and crude extracts of S. aureus and Str. agalactiae. The
beaker containing oil phase was fitted to a homogenizer and stirred at 200 rpm. The antigens
in aqueous phase were then added slowly over a period of 30 seconds. Stirring was increased
to 2000 rpm for 10 minutes. The 5 ml of Montanide® adjuvanted bacterin- toxoid finally
contained 5x 109
cells each of S. aureus, Str. agalactiae and P. multocida, toxin extracts of S.
aureus, Str. agalactiae, formalin, thimerosal, sodium azide and Montanide® ISA 201 VG to
make a total volume of 5 ml (Athar, 2007; Yousaf et al., 2009; Giraudo et al., 1997).
3.10 Evaluation of quality of vaccine
3.10.1 Sterility
The prepared vaccine was cultured on blood agar and MacConkey‟s agar plates for
checking its sterility. The inoculated plates were incubated at 37 °C for 48 hours and
observed for any growth.
3.10.2 Safety and side effects
Four cows and buffaloes were vaccinated with double the recommended dose of
vaccine (10 ml) and monitored for 14 days for local and systemic reactions if any. In
addition, 5 mice were inoculated intramuscularly with 0.2 ml of the vaccine and observed for
5 days for occurrence of any untoward local and systemic reaction (OIE, 2012).
3.10.3 Challenge- protection assay (Vaccination- challenge study)
Fifty mice divided randomly into 5 groups (M-1 thru M-5) were immunized
intraperitoneally with 0.1 ml doses of the vaccine administered at 15 day interval. The mice
were then challenged with inocula 30 days after administration of the second dose of vaccine.
The procedure for preparation and administration of inocula is as under:
One ml inoculum containing 109 CFU ml
-1 of each of the vaccinal isolates (S. aureus, Str.
agalactiae and P. multocida) grown in modified nutrient broth for 24 hours at 37 oC,
centrifuged at 6,000×g for 30 minutes at 4°C, and resuspended in peptone solution (0.5% w/v
of peptone and 0.5% NaCl), was inoculated intraperitoneally in vaccinated and control mice.

41
The numbers of dead mice within 3 days of the challenge were recorded and compared to
calculate the percent survival rate (Giraudo et al., 1997; Athar, 2007)
3.10.4 Stability
The physical stability of vaccine was evaluated by storing vaccine vials at 4 °C and
7° C for 6 months and parameters (pH, sedimentation, color, appearance, syringibility) were
recorded at monthly intervals (OIE, 2012; Athar, 2007)
Phase II: Field Evaluation of Montanide® Adjuvanted Combined HS-
Mastitis Vaccine in Target Dairy Species
3.11 Animals, their management and treatment-based grouping
A total of 70 mastitis free lactating animals (buffaloes n=45; cows n=25), 50 lactating
cows and buffaloes microbiologically positive for S. aureus/Str. agalactiae (buffaloes n=25,
cow n=25) and dairy calves aged up to 1 year (buffalo calves n=70; cow calves n=50) were
utilized for the field evaluation of combined HS-mastitis vaccine. The animals were selected
from those maintained at government institutes (Livestock Experimental Station, University
of Agriculture, Faisalabad, Ayub Agriculture Research Institute, Faisalabad, etc.,) and two
private dairy farms (Raja dairy farm, Samundri road, Faisalabad and Mehar Anayat dairy
farm, Raja wala, Faisalabad). Non vaccination status against HS during the last 6 months and
low or no P. multocida antibody titers (ELISA- based) were another criterion for selecting
animals for field trial. The grouping of these animals, their experimental treatments, and
HS/mastitis status at the time of initiation of trial is depicted in Table 3.6. General principles
for the design of clinical trials with special reference to mastitis and HS described by
Thorburn (1990) and OIE (2012) were followed.

42
Table 3.6: Grouping of animals, their experimental treatments, and HS/mastitis status
at the time of initiation of field trial of combined HS- mastitis vaccine
Groups Experimental
treatment
Dairy species and
number (n) of
animals
HS-mastitis status at the time of
initiation of trial
A Vaccination twice
at 21 day interval
Buffalo (n=30)
Cow (n=10)
Milk cultu re –ve for S. aureus and
Str agalactiae, CMT/ SFMT –ve;
no history of vaccination against
HS during the last six months, no
or low antibody titers against P.
multocida
B (control) No vaccination Buffalo (n=15)
Cow (n=15)
Milk culture –ve for S. aureus and
Str. agalactiae, CMT/ SFMT –ve;
no history of vaccination against
HS during the last six months, no
or low antibody titers against P.
multocida
C (Sub
clinically
mastitic
buffaloes and
cows)
Vaccination twice
at 21 days
interval
Buffalo (n=15)
Cow (n=15)
Milk culture +ve for S. aureus/Str.
agalactiae, CMT/ SFMT +ve in
one or more quarter but no clinical
signs of mastitis
D (Sub
clinically
mastitic
buffaloes and
cows)
Infected
control
No vaccination Buffalo (n=10)
Cow (n=10)
Milk culture +ve for S. aureus/Str.
agalactiae, CMT/ SFMT +ve in
one or more quarter but no clinical
signs of mastitis
E
Vaccination twice
at 21 days
interval
Buffalo calves aged
up to 1 year (n=50)
Cow calves aged up
to 1 year (n=25)
No history of vaccination against
HS during the last six months
together with no or very low
antibody titer against P. multocida
F (Control)
No
vaccination
Buffalo calves aged
up to 1 year (n=20)
Cow calves aged up
to 1 year (n=25)
No history of vaccination against
HS during the last six months
together with no or very low
antibody titer against P. multocida
IMI=Intrammamry infection; SFMT=Surf field mastitis test; CMT=California mastitis test

43
3.12 Evaluation parameters
3.12.1 Hemorrhagic septicemia
3.12.1.1 ELISA- based antibody titers against P. multocida
ELISA-based antibody titers against P. multocida were determined from 20 % of
randomly selected vaccinated and control animals at day 0, 60, 120, and 180 post
vaccination. Serum antibody titers were determined by in-house indirect ELISA, as described
in the preceding section 3.4.
3.12.1.2 Incidence of HS
Incidence of HS in vaccinated and control animals over a period of 180 days if any was
recorded. The diagnosis of HS was based on typical signs (De Alwis, 1992).
3.12.1.3 Local and systemic reactions
Local and systemic reactions in vaccinated animals (OIE, 2012) were monitored.
3.13 Mastitis
3.13.1 ELISA-based antibody titers against S. aureus and Str. agalactiae
ELISA-based antibody titers against S. aureus, and Str. agalactiae were determined
from at least 20 % of randomly selected vaccinated and control animals at day 0, 60, 120, and
180 post vaccination (Qureshi and Saxena, 2014). Serum antibody titers were determined by
in-house indirect ELISA, as described in the preceding section 3.4.
3.13.2 Incidence and prevalence of S. aureus and Str. agalactiae mastitis
Incidence and prevalence of S. aureus and Str. agalactiae mastitis over a period of
180 days were determined by microbiological examination of aseptically collected quarter
foremilk samples as described by Hogan et al. (I999).
3.13.3 Mastitis screening tests based prevalence and incidence of mastitis
Surf Field Mastitis test (Muhammad et al., 2010) and California Mastitis Test
(Schalm et al., 1971) based prevalence and incidence of mastitis was determined at day 60,
120, and 180 post vaccination.
3.13.4 Severity of mastitis in vaccinated and control animals
Severity of clinical mastitis in vaccinated and control groups was determined as per
Athar (2007).

44
3.13.5 Isolation and identification of pathogens from clinically mastitic animals
Clinically mastitic quarters of vaccinated and unvaccinated animals were subjected to
microbiological examination of aseptically collected milk samples to determine the nature of
mastitis causing pathogens (Hogan et al., I999).
3.13.6 Milk somatic cell count
Somatic cell count was monitored in vaccinated and control groups at day 60, 120,
and 180 post vaccination (Singh and Ludri, 2000; Shailja and Singh, 2002; Athar et al., 2007
and Rashid, 2011).
3.13.7 Effect of vaccination on milk yield
Mean milk yields (liters per 24 hours) were recorded on monthly basis for six months post
vaccination. Similar observations were made in unvaccinated groups (Nickerson, 1994)
3.13.8 Vaccine efficacy
Proportionate reduction in S. aureus, Str. agalactiae mastitis and HS attack rates
(AR) between the unvaccinated (ARU) and vaccinated (ARV) study cohorts were calculated
from the relative risk (RR) of these diseases among the vaccinated group using the following
formulas (Weinberg and Szilagyi, 2010).
ARU - ARV
Efficacy = -------------------------- × 100
ARU
Efficacy= (1-RR) × 100
3.14 Statistical Analysis
The data collected were subjected to statistical analysis using ANOVA and other
appropriate design software package (Anonymous, 2004). Mammary quarters were
considered as unit of concern for prevalence, incidence and somatic cell count. For serum
and milk whey antibody titers buffaloes and cows were the unit of concern. Significance was
declared as P≤0.05 (Steel et al., 1997). Cost benefit analysis was performed by partial
budgeting technique/spread sheet cost- benefit analysis (Miller, 1986).
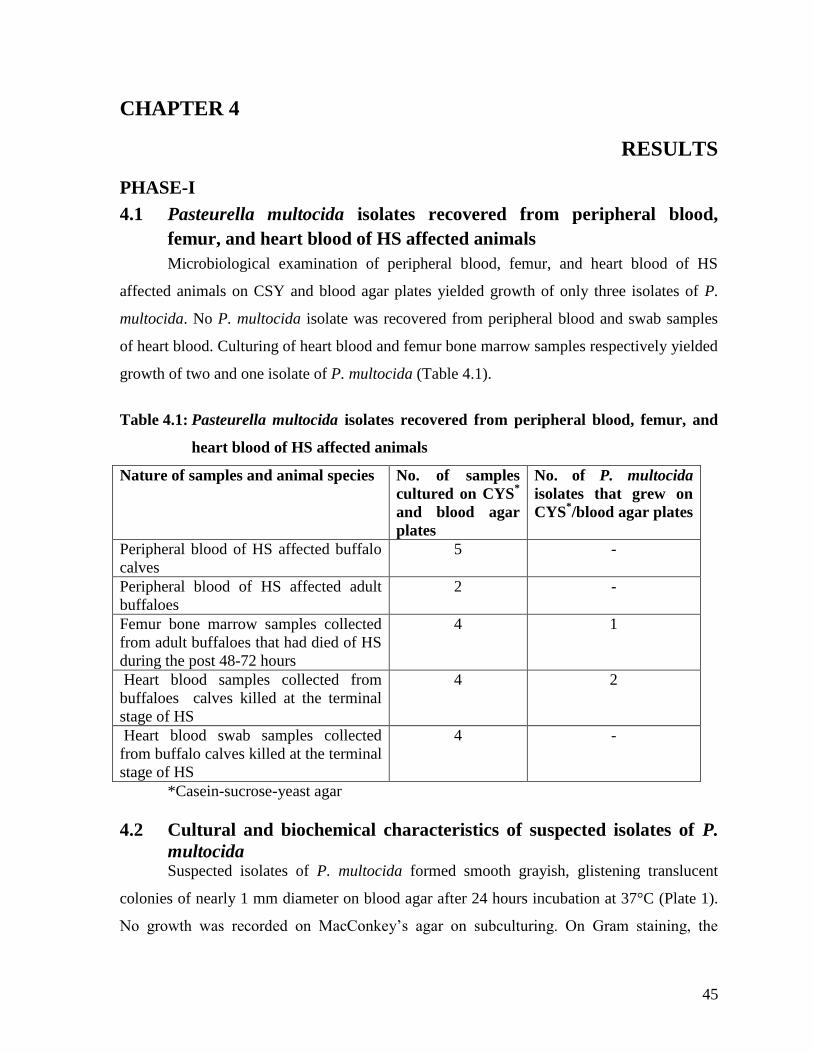
45
CHAPTER 4
RESULTS
PHASE-I
4.1 Pasteurella multocida isolates recovered from peripheral blood,
femur, and heart blood of HS affected animals
Microbiological examination of peripheral blood, femur, and heart blood of HS
affected animals on CSY and blood agar plates yielded growth of only three isolates of P.
multocida. No P. multocida isolate was recovered from peripheral blood and swab samples
of heart blood. Culturing of heart blood and femur bone marrow samples respectively yielded
growth of two and one isolate of P. multocida (Table 4.1).
Table 4.1: Pasteurella multocida isolates recovered from peripheral blood, femur, and
heart blood of HS affected animals
Nature of samples and animal species No. of samples
cultured on CYS*
and blood agar
plates
No. of P. multocida
isolates that grew on
CYS*/blood agar plates
Peripheral blood of HS affected buffalo
calves
5 -
Peripheral blood of HS affected adult
buffaloes
2 -
Femur bone marrow samples collected
from adult buffaloes that had died of HS
during the post 48-72 hours
4 1
Heart blood samples collected from
buffaloes calves killed at the terminal
stage of HS
4 2
Heart blood swab samples collected
from buffalo calves killed at the terminal
stage of HS
4 -
*Casein-sucrose-yeast agar
4.2 Cultural and biochemical characteristics of suspected isolates of P.
multocida Suspected isolates of P. multocida formed smooth grayish, glistening translucent
colonies of nearly 1 mm diameter on blood agar after 24 hours incubation at 37°C (Plate 1).
No growth was recorded on MacConkey‟s agar on subculturing. On Gram staining, the

46
organism appeared as Gram –ve, short, ovoid bipolar coccoid, with some degree of
pleomorphism. Growth on CYS agar was smooth, yellowish, mucoid and larger in size than
that on blood agar (Plate 2).
Plate 1. Growth of a P. multocida Plate 2. Growth of a P.multocida on a
Casein- on a blood agar plate sucrose-yeast agar plate
4.2.1 Biochemical reactions of P. multocida isolates
All three P. multocida isolates were positive for indol, ODC, glucose, fructose, and
catalase, negative for H2S production, lactose, D-dulcitol and maltose. Variable results were
recorded for D-sorbitol and arabinose (Table 4.2).
Table 4.2: Biochemical profiles of P. multocida isolates (n=3) recovered from femur
bone marrow and heart blood samples
Biochemical test No. of P. multocida isolates showing + ve or –ve
reaction
Indol All three isolates showed a +ve reaction
ODC* All three isolates showed a +ve reaction
H2S on TSI All three isolates showed a -ve reaction
D-sorbitol Two isolates showed +ve reaction and one isolate
showed -ve reaction
D-dulcitol All three isolates showed a -ve reaction
Arabinose
One isolate showed +ve reaction and two isolates
showed-ve reaction
Glucose All three isolates showed a +ve reaction
Fructose All three isolates showed a +ve reaction
Lactose All three isolates showed a -ve reaction
Catalase All three isolates showed a +ve reaction
Maltose All three isolates showed a -ve reaction *ODC = Ornithine decarboxylase activity.

47
4.2.2 PCR based confirmation of P. multocida isolates
These isolates of P. multocida were confirmed by amplification of P. multocida
specific gene using KTSP61/KTT7 set of primers. These primers yielded a product of 618
bp on electrophoresis which confirmed these isolates as P. multocida type B.
4.2.3 Antimicrobial susceptibility testing of 3 candidate vaccinal P. multocida isolates
All 3 isolates of P. multocida were sensitive to enrofloxacin, amoxicillin, ceftiofur
sodium and oxacillin and resistant to tetracycline, fusidic acid, and penicillin. All three
isolates displayed an intermediate sensitivity to vancomycine. A variable degree of
susceptibility was displayed to lincomycine, septran and gentamicin (Table 4.3).
Table 4.3: Antibiotic susceptibility profiles of 3 candidate vaccinal isolates of P.
multocida against 11 antibiotics/antimicrobials
Designation of P.
multocida isolate
Antibiotics /antimicrobials and their susceptibility profiles
Linco
Enro
Amox
Tetra
Vanco
Sept
Gen
Fus.acid
Pen
Ceft
Ox
P. multocida
isolate No.1
R S S R I R S R R S S
P. multocida
isolate No. 2
S S S R I I I R R S S
P. multocida
isolate No. 3
S S S R I I S R R S S
Linco= Lincomycin; Enro= Enrofloxacin; Amox= Amoxicillin; Tetra= Tetracycline; Van=
Vancomycine; Sept= Septran (Sulfamethoxazole+Trimethoprim; Gen=Gentamicin;
Fus.acid= Fusidic acid; Pen=Penicillin G; Ceft= Ceftiofur sodium; Ox=Oxacillin; S=
Sensitive; I= Intermediate; R= Resistant
4.2.4 Pathogenicity of the 3 candidate vaccinal isolates of P. multocida in mice
Table 4.4 depicts the mouse LD50 of 3 candidate vaccinal isolates of P. multocida. As
can be seen in this table, isolate No.1 (vaccinal isolate) was more virulent than isolates no.2
and 3, the latter two isolates were equal in terms of virulence in mouse. Necropsy
examination of all mouse dying in 24 hours revealed clear septicemic lesions consisting of
necrotic foci on the dorsal surface of liver and hemorrhages in lungs, trachea and spleen.
Giemsa‟s stained smears prepared from the cardiac blood of dead mice revealed the presence
of cocco bacilli. Culturing of heart blood samples on blood agar and CYS agar revealed the
presence of colonies of P. multocida with characteristics as described in section 4.2 above.
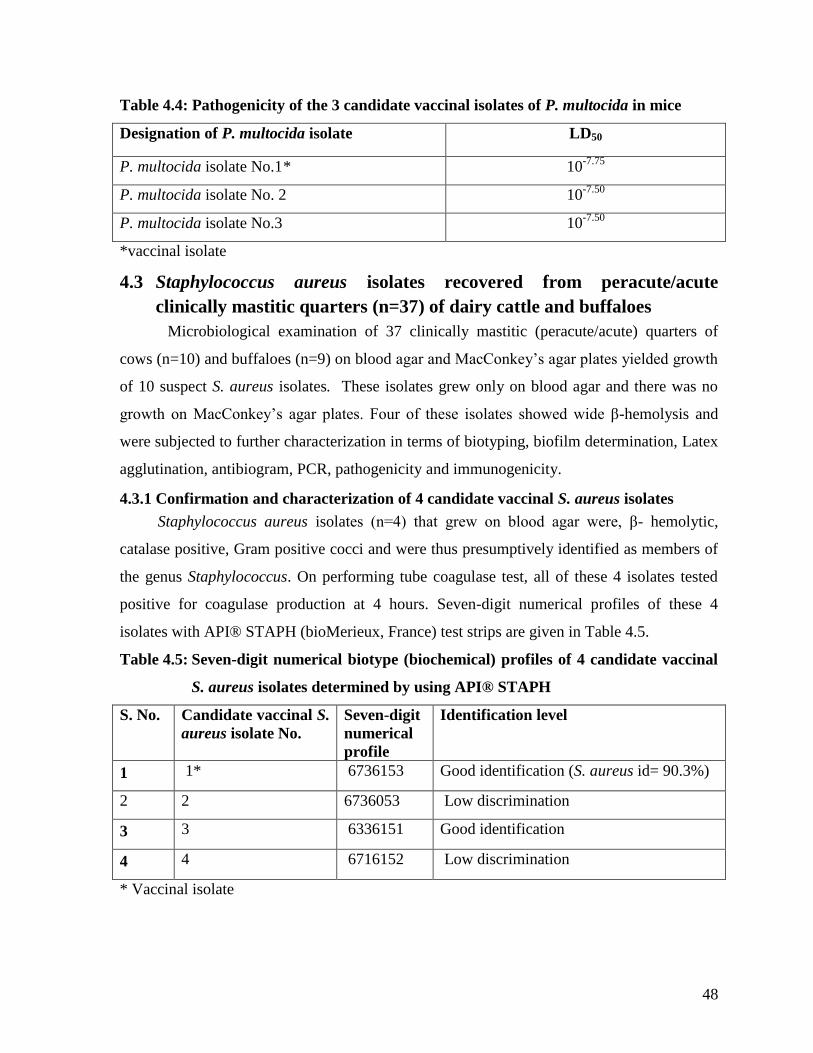
48
Table 4.4: Pathogenicity of the 3 candidate vaccinal isolates of P. multocida in mice
Designation of P. multocida isolate LD50
P. multocida isolate No.1* 10-7.75
P. multocida isolate No. 2 10-7.50
P. multocida isolate No.3 10-7.50
*vaccinal isolate
4.3 Staphylococcus aureus isolates recovered from peracute/acute
clinically mastitic quarters (n=37) of dairy cattle and buffaloes
Microbiological examination of 37 clinically mastitic (peracute/acute) quarters of
cows (n=10) and buffaloes (n=9) on blood agar and MacConkey‟s agar plates yielded growth
of 10 suspect S. aureus isolates. These isolates grew only on blood agar and there was no
growth on MacConkey‟s agar plates. Four of these isolates showed wide β-hemolysis and
were subjected to further characterization in terms of biotyping, biofilm determination, Latex
agglutination, antibiogram, PCR, pathogenicity and immunogenicity.
4.3.1 Confirmation and characterization of 4 candidate vaccinal S. aureus isolates
Staphylococcus aureus isolates (n=4) that grew on blood agar were, β- hemolytic,
catalase positive, Gram positive cocci and were thus presumptively identified as members of
the genus Staphylococcus. On performing tube coagulase test, all of these 4 isolates tested
positive for coagulase production at 4 hours. Seven-digit numerical profiles of these 4
isolates with API® STAPH (bioMerieux, France) test strips are given in Table 4.5.
Table 4.5: Seven-digit numerical biotype (biochemical) profiles of 4 candidate vaccinal
S. aureus isolates determined by using API® STAPH
S. No. Candidate vaccinal S.
aureus isolate No.
Seven-digit
numerical
profile
Identification level
1 1* 6736153 Good identification (S. aureus id= 90.3%)
2 2 6736053 Low discrimination
3 3 6336151 Good identification
4 4 6716152 Low discrimination
* Vaccinal isolate

49
Plate 3 depicts the biochemical reactions of one of the 4 candidate vaccinal S. aureus isolates
Plate 3. Seven-digit numerical biotype profile of a candidate vaccinal S. aureus isolates
determined by API® STAPH.
Latex agglutination test using Staphytect plus® kit was positive for all of the four candidate
vaccinal S. aureus isolates (Plate 4)
Plate 4. Latex agglutination test positive and negative reactions using Staphytect plus® kit

50
4.3.2 Antimicrobial susceptibility of candidate vaccinal S. aureus isolates
All 4 isolates of S .aureus were sensitive to lincomycin, enrofloxacin, amoxicillin,
ceftiofur sodium and oxacillin and resistant to tetracycline. A variable degree of
susceptibility was displayed to vancomycine, septran, gentamicin, fusidic acid and penicillin,
(Table 4.6).
Table 4.6: Antibiotic susceptibility profiles of 4 candidate vaccinal isolates of S. aureus
against 11 antibiotics/antimicrobials
Designation of
S. aureus isolate
Antibiotics /antimicrobials and their susceptibility profiles
Linco
Enro
Amox
Tetra
Vanco
Sept
Gen
Fus.acid
Pen
Ceft
Ox
S. aureus isolate
No.1*
S S S R I I I R R S S
S. aureus isolate
No.2
S S S R S S S S S S S
S. aureus isolate
No.3
S S S R I I S R R S S
S. aureus
isolate No.4 S S S R S S S S S S S
Linco= Lincomycin; Enro= Enrofloxacin; Amox= Amoxicillin; Tetra= Tetracycline; Van=
Vancomycine; Sept= Septran (Sulfamethoxazole+Trimethoprim; Gen=Gentamicin;
Fus.acid= Fusidic acid; Pen=Penicillin G; Ceft= Ceftiofur sodium; Ox=Oxacillin; S=
Sensitive; I= Intermediate; R= Resistant, *vaccinal isolate
4.3.3 Pathogenicity of the 4 candidate vaccinal isolates of S. aureus in rabbits
Table 4.7 depicts the results of pathogenicity testing of 4 candidate vaccinal isolates
of S. aureus in rabbits. As can be seen in this table, 100, 80, 80 and 60% rabbits died within
18 hours of inoculation of isolates No. 1, 2, 3 and 4, respectively. Necropsy examination of
dead rabbits revealed septicemia along with congested lungs and heart and petechiation on
serosal surface of intestine, with straw colored fluid in abdominal cavity. Stained smears of
blood and straw colored fluid revealed S. aureus under microscope. Each isolate was also
isolated from straw colored fluid in the abdominal cavity.

51
Table 4.7: Pathogenicity of the 4 candidate vaccinal isolates of S. aureus in rabbits
Designation of S.
aureus isolate
Percent mortality of rabbits within 18 hours of inoculation of 0.2 ml
of bacterial suspension containing 1x1010
cells / ml of respective
isolate.
S. aureus isolate No.1* 100
S. aureus isolate No.2 80
S. aureus isolate No.3 80
S. aureus isolate No.4 60
*vaccinal isolate
4.3.4 Biofilm production by candidate vaccinal S. aureus isolates (n=4)
4.3.4.1 Tube method
Table 4.8 depicts tube method-based scores of biofilm production by 4 candidate
vaccinal S. aureus isolates. As can be seen in this table, S. aureus isolate number 2, 3 and, 4
showed a moderate level of biofilm production whereas isolate number 1 showed strong
biofilm production.
Table 4.8: Tube method-based scores of biofilm production by candidate vaccinal S.
aureus isolates (n=4)
S. No. Candidate vaccinal S. aureus isolate No. Biofilm production score*
1 1** 4 (strong )
2 2 2 (moderate)
3 3 2 (moderate)
4 4 2 (moderate)
**Vaccinal isolate *As per Christensen et al. (1982).
4.3.4.2 Spectrophotometric method
Table 4.9 depicts the result of spectrophotometric based estimates of biofilm
production traits of 4 candidate vaccinal isolates of S. aureus. All 4 isolates produced
biofilms, however highest OD (0.31nm) was noted in isolate # 1, whilst OD values ranged
between 0.17- 0.22. Thus the isolate #1 was considered as highest producer of biofilm in
spectrophotometric method.
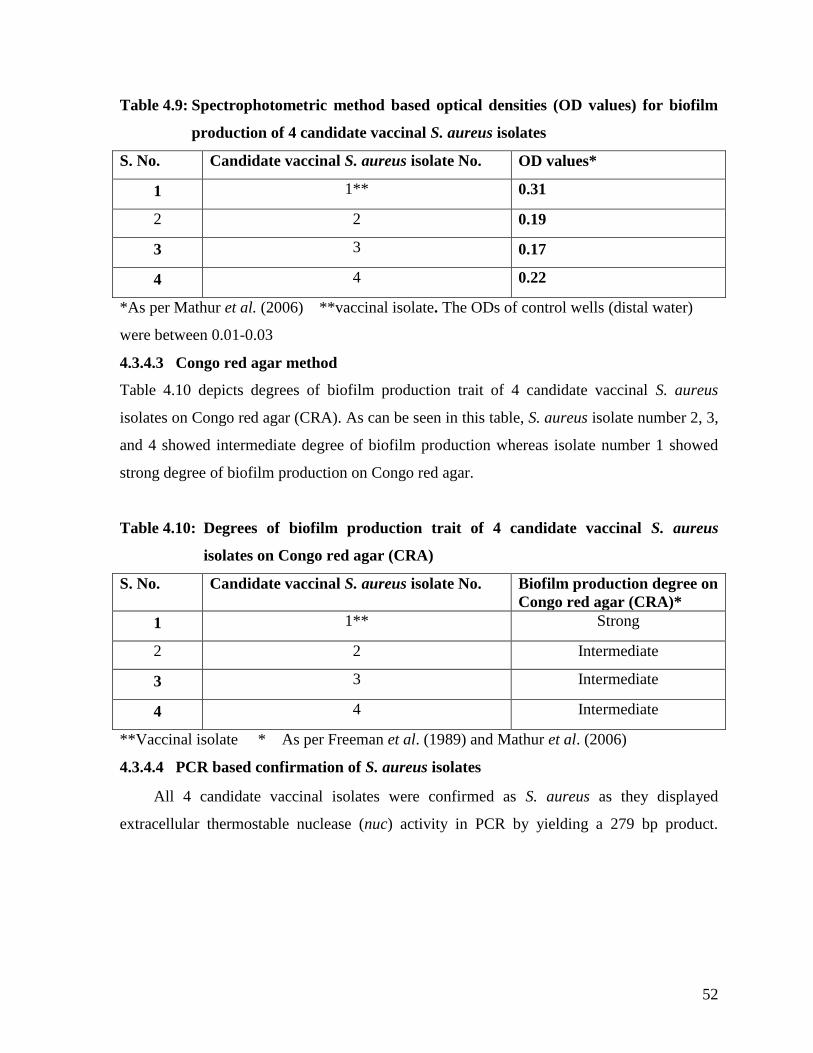
52
Table 4.9: Spectrophotometric method based optical densities (OD values) for biofilm
production of 4 candidate vaccinal S. aureus isolates
S. No. Candidate vaccinal S. aureus isolate No. OD values*
1 1** 0.31
2 2 0.19
3 3 0.17
4 4 0.22
*As per Mathur et al. (2006) **vaccinal isolate. The ODs of control wells (distal water)
were between 0.01-0.03
4.3.4.3 Congo red agar method
Table 4.10 depicts degrees of biofilm production trait of 4 candidate vaccinal S. aureus
isolates on Congo red agar (CRA). As can be seen in this table, S. aureus isolate number 2, 3,
and 4 showed intermediate degree of biofilm production whereas isolate number 1 showed
strong degree of biofilm production on Congo red agar.
Table 4.10: Degrees of biofilm production trait of 4 candidate vaccinal S. aureus
isolates on Congo red agar (CRA)
S. No. Candidate vaccinal S. aureus isolate No. Biofilm production degree on
Congo red agar (CRA)*
1 1** Strong
2 2 Intermediate
3 3 Intermediate
4 4 Intermediate
**Vaccinal isolate * As per Freeman et al. (1989) and Mathur et al. (2006)
4.3.4.4 PCR based confirmation of S. aureus isolates
All 4 candidate vaccinal isolates were confirmed as S. aureus as they displayed
extracellular thermostable nuclease (nuc) activity in PCR by yielding a 279 bp product.

53
4.4 Streptococcus agalactiae isolates recovered from peracute/acute
clinical mastitis from dairy cattle and buffaloes.
Microbiological examination of 37 clinically mastitic (peracute/acute) quarters of cows
(n=10) and buffaloes (n=9) on blood agar yielded growth of 2 suspect Str. agalactiae
isolates.
4.4.1 Confirmation and characterization of candidate vaccinal Str. agalactiae isolate
On blood agar, suspect Str. agalactiae isolates (n=2) displayed β-hemolysis. Microscopic
examination of Gram stained smears of these isolates revealed Gram positive cocci. On
catalase test, these isolates were negative. CAMP test reaction on blood agar with 0.1%
esculine and 0.01% ferric citrate was positive for both the isolates.
Plate 5: Growth of a Str. agalactiae on a blood agar plate
4.4.2 Seven-digit numerical biotype (biochemical) characteristics of candidate vaccinal
isolates
Biotypes 2 candidate vaccinal Str. agalactiae isolates in terms of seven-digit
numerical profiles determined by using a commercially available kit API® 20 STREP,
bioMérieux, France) have been depicted in Table 4.11.
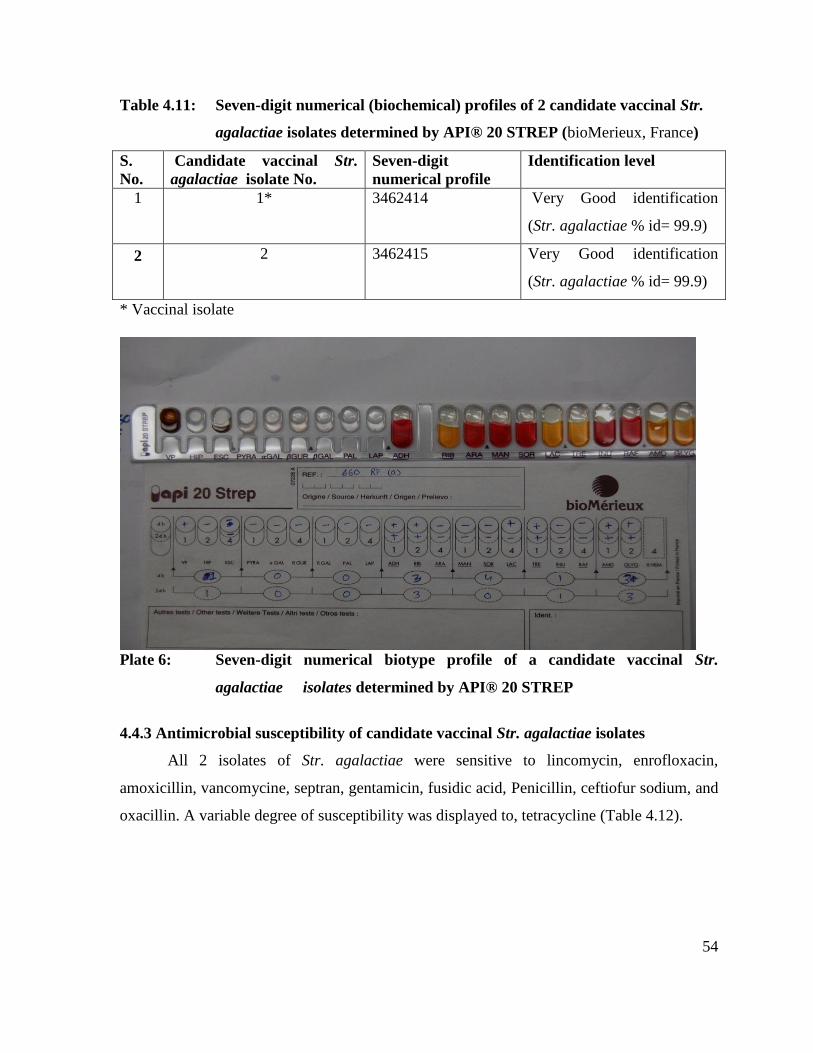
54
Table 4.11: Seven-digit numerical (biochemical) profiles of 2 candidate vaccinal Str.
agalactiae isolates determined by API® 20 STREP (bioMerieux, France)
S.
No.
Candidate vaccinal Str.
agalactiae isolate No.
Seven-digit
numerical profile
Identification level
1 1* 3462414 Very Good identification
(Str. agalactiae % id= 99.9)
2 2 3462415 Very Good identification
(Str. agalactiae % id= 99.9)
* Vaccinal isolate
Plate 6: Seven-digit numerical biotype profile of a candidate vaccinal Str.
agalactiae isolates determined by API® 20 STREP
4.4.3 Antimicrobial susceptibility of candidate vaccinal Str. agalactiae isolates
All 2 isolates of Str. agalactiae were sensitive to lincomycin, enrofloxacin,
amoxicillin, vancomycine, septran, gentamicin, fusidic acid, Penicillin, ceftiofur sodium, and
oxacillin. A variable degree of susceptibility was displayed to, tetracycline (Table 4.12).
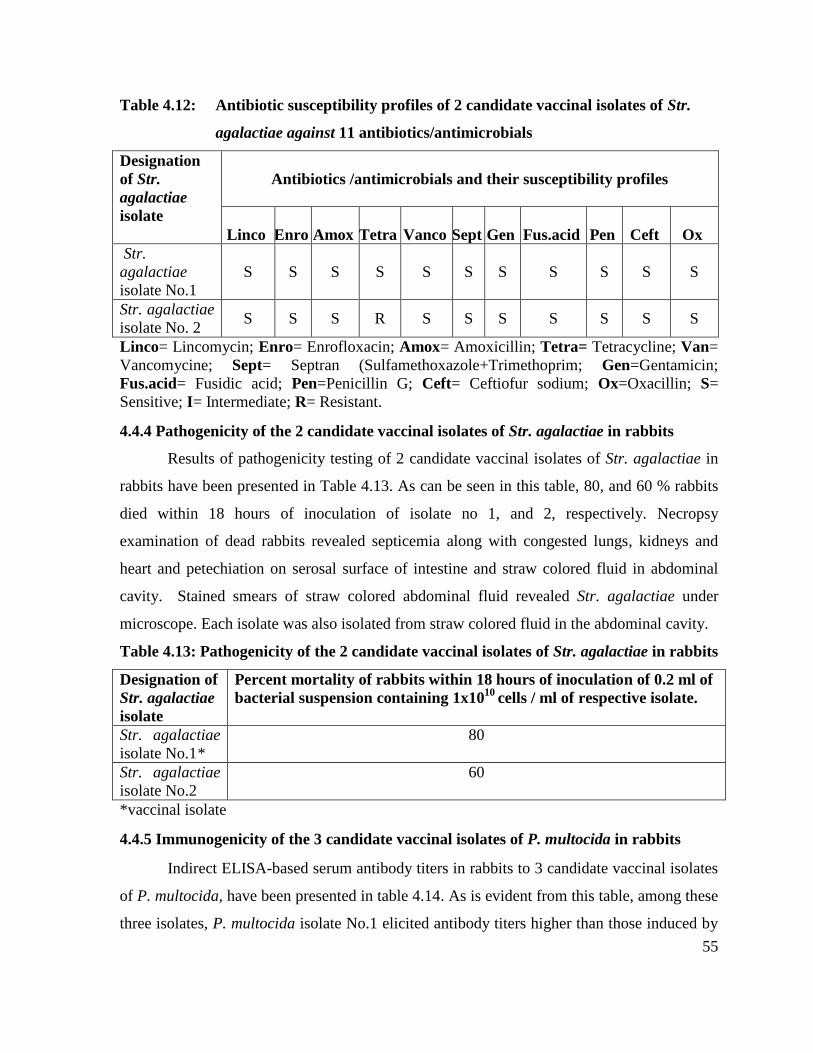
55
Table 4.12: Antibiotic susceptibility profiles of 2 candidate vaccinal isolates of Str.
agalactiae against 11 antibiotics/antimicrobials
Designation
of Str.
agalactiae
isolate
Antibiotics /antimicrobials and their susceptibility profiles
Linco
Enro
Amox
Tetra
Vanco
Sept
Gen
Fus.acid
Pen
Ceft
Ox
Str.
agalactiae
isolate No.1
S S S S S S S S S S S
Str. agalactiae
isolate No. 2 S S S R S S S S S S S
Linco= Lincomycin; Enro= Enrofloxacin; Amox= Amoxicillin; Tetra= Tetracycline; Van=
Vancomycine; Sept= Septran (Sulfamethoxazole+Trimethoprim; Gen=Gentamicin;
Fus.acid= Fusidic acid; Pen=Penicillin G; Ceft= Ceftiofur sodium; Ox=Oxacillin; S=
Sensitive; I= Intermediate; R= Resistant.
4.4.4 Pathogenicity of the 2 candidate vaccinal isolates of Str. agalactiae in rabbits
Results of pathogenicity testing of 2 candidate vaccinal isolates of Str. agalactiae in
rabbits have been presented in Table 4.13. As can be seen in this table, 80, and 60 % rabbits
died within 18 hours of inoculation of isolate no 1, and 2, respectively. Necropsy
examination of dead rabbits revealed septicemia along with congested lungs, kidneys and
heart and petechiation on serosal surface of intestine and straw colored fluid in abdominal
cavity. Stained smears of straw colored abdominal fluid revealed Str. agalactiae under
microscope. Each isolate was also isolated from straw colored fluid in the abdominal cavity.
Table 4.13: Pathogenicity of the 2 candidate vaccinal isolates of Str. agalactiae in rabbits
Designation of
Str. agalactiae
isolate
Percent mortality of rabbits within 18 hours of inoculation of 0.2 ml of
bacterial suspension containing 1x1010
cells / ml of respective isolate.
Str. agalactiae
isolate No.1*
80
Str. agalactiae
isolate No.2
60
*vaccinal isolate
4.4.5 Immunogenicity of the 3 candidate vaccinal isolates of P. multocida in rabbits
Indirect ELISA-based serum antibody titers in rabbits to 3 candidate vaccinal isolates
of P. multocida, have been presented in table 4.14. As is evident from this table, among these
three isolates, P. multocida isolate No.1 elicited antibody titers higher than those induced by

56
isolate No.2 and 3 on all the three post inoculation sampling days (i.e.
day 7, 14, and 21).
Table 4.14: Indirect ELISA-based serum antibody titers in rabbits to 3 candidate
vaccinal isolates of P. multocida
Designation of P. multocida isolate D-0 D-7 D-14 D-21
P. multocida isolate No.1* 0.26 0.96 1.56 1.9
P. multocida isolate No.2 0.20 0.8 1.2 1.4
P. multocida isolate No. 3 0.1 0.86 1.3 1.8
*vaccinal isolate
4.4.6 Immunogenicity of the 4 candidate vaccinal isolates of S. aureus in rabbits
Table 4.15 depicts indirect ELISA-based serum antibody titers in rabbits to 4
candidate vaccinal isolates of S. aureus. As is evident from this table, amonge these three
isolates, S. aureus No.1 elicited antibody titers higher than those induced by isolate No.2, 3
and 4 on all the three post inoculation sampling days (i.e. day 7, 14 and 21).
Table 4.15: Indirect ELISA-based serum antibody titers in rabbits to 4 candidate
vaccinal isolates of S. aureus
Designation of S. aureus isolate D-0 D-7 D-14
D-21
S. aureus isolate No.1* 0.19 0.6 1.3 1.23
S. aureus isolate No.2 0.18 0.5 1.2 1.11
S. aureus isolate No.3 0.17 0.51 1.22 1.12
S. aureus isolate No.4 0.165 0.52 1.24 1.14
*vaccinal isolate
4.4.7 Immunogenicity of the 2 candidate vaccinal isolates of Str. agalactiae in rabbits
Indirect ELISA-based serum antibody titers in rabbits to 2 candidate vaccinal isolates
of Str. agalactiae, have been presented in table 4.16. As is evident from this table, among
these three isolates, Str. agalactiae isolate No.1 elicited antibody titers higher than those
induced by isolate No.2 on all the three post inoculation sampling days (i.e.
day 7, 14, and 21).
57
Table 4.16: Indirect ELISA-based serum antibody titers in rabbits to 2 candidate
vaccinal isolates of Str. agalactiae
Designation of Str. agalactiae isolate D-0 D-7 D-14 D-21
Str. agalactiae isolate No.1* 0.113 0.878 1.13 1.12
Str. agalactiae isolate No.2 0.19 0.6 1.3 1.1
*vaccinal isolate
4.5 Final selection of vaccinal isolates from amongst the candidate
vaccinal isolates of P. multocida, S. aureus and Str. agalactiae for
vaccine production
The most pathogenic (table 4.4) and most immunogenic (table 4.14) P. multocida was
selected for vaccine production. From amongst the 4 candidate S. aureus isolates, the
strongest biofilm producing (table 4.8) the most pathogenic (table 4.7) and most
immunogenic (table 4.15) S. aureus isolate was selected for vaccine production. Similarly,
the more pathogenic (table 4.13) and more immunogenic (table 4.16) of the 2 candidate Str.
agalactiae isolates was selected for vaccine production.
Table 4.17: Composition of Montanide® adjuvanted combine HS-mastitis vaccine.
Ingredients Each 5m ml dose contained
P. multocida 5 × 109 cells
Staphylococcus aureus ( 6736153) 5 × 109 cells
Streptococcus agalactiae (3462414) 5 × 109
cells
Crude toxin extract 1.5 ml
Montanide® ISA 201 VG 2.5 ml
Formalin <0.02 ml
Thimeosal 50 µg=0.00005 g
Sodium azide 50 µg=0.00005 g
PBS Q. S.

58
Plate 7: Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccine
Table 4.18: Indirect ELISA-based serum antibody titers in rabbits to vaccinal isolates of P.
multocida, S. aureus and Str.agalactiae
Vaccinal Isolates D-0 D-7 D-14 D-21
P. multocida isolate No.1* 0.26 0.96 1.56 1.9
S. aureus isolate No.1* 0.19 0.6 1.3 1.23
Str. agalactiae isolate No.1* 0.113 0.878 1.13 1.12
* Vaccinal isolates
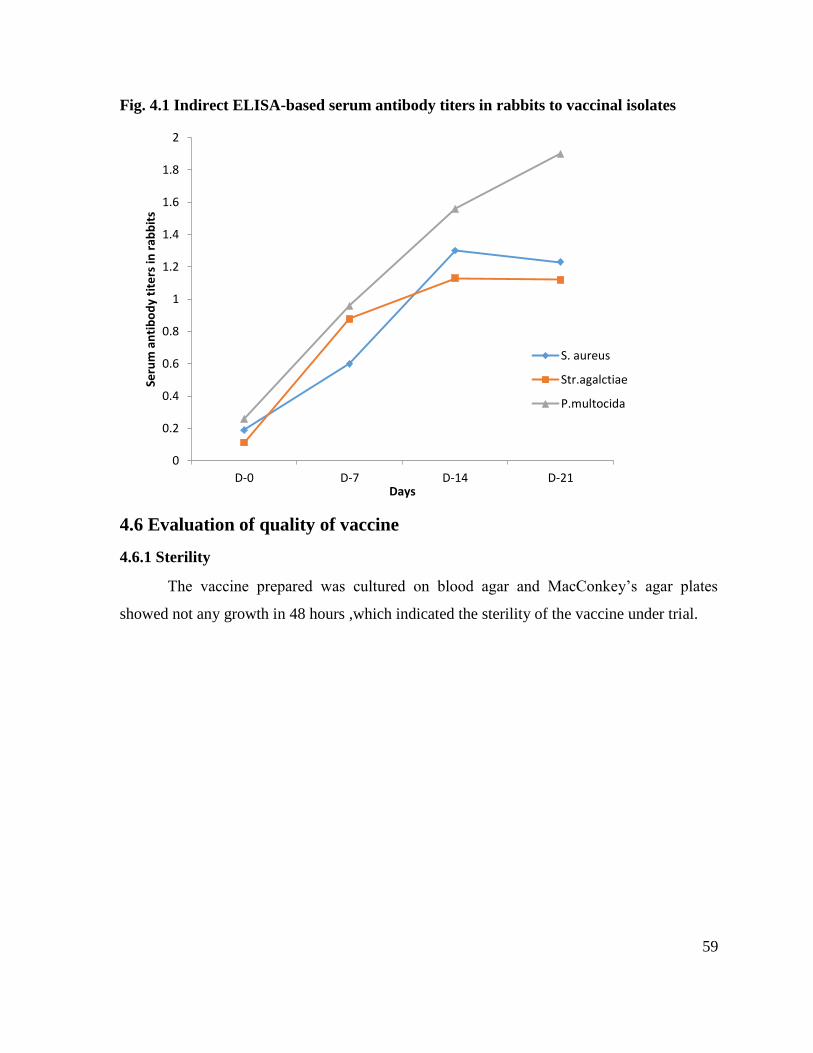
59
Fig. 4.1 Indirect ELISA-based serum antibody titers in rabbits to vaccinal isolates
4.6 Evaluation of quality of vaccine
4.6.1 Sterility
The vaccine prepared was cultured on blood agar and MacConkey‟s agar plates
showed not any growth in 48 hours ,which indicated the sterility of the vaccine under trial.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
D-0 D-7 D-14 D-21
Seru
m a
nti
bo
dy
tite
rs in
rab
bit
s
Days
S. aureus
Str.agalctiae
P.multocida

60
4.6.2 Stability
4.6.2.1 Effect of temperature and duration of storage on the physical stability of
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccine.
The physical stability of vaccine were evaluated by storing vaccine vials at 4°C and
37°C for 6 months and parameters (pH, sedimentation, color, appearance, syringibility) were
recorded at monthly intervals.
Table 4.19: Effect of temperature and duration of storage on the physical stability of
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccine
Temperature Days in Storage
7 90 180
4°C Good Good Satisfactory
25°C Good Good Satisfactory
37°C Good Good Satisfactory
Good = Milky white suspension without any sedimentation; Satisfactory = Milky with a
small portion of water phase temporarily separated which upon shaking readily becomes
suspension
4.6.3 Safety and side effects
4.6.3.1 Comparative safety of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine in cattle and buffaloes.
Subcutaneously administration of vaccine was associated with a varying degree of
mild to moderate swelling at injection site in cows and buffaloes that subsided within 3-7
days while intramuscular administration caused either no or a very mild swelling at the
injection site that subsided within 48 hours. No systemic effects were noted.

61
Table 4.20: Comparative safety of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine in cattle and buffaloes
Groups
of cows
and
buffaloes
Vaccine Number
of
animals
Dose and
Route
Mortality Severity of
swelling at
injection
site
Cows
Montanide®
Adjuvanted Combined
Hemorrhagic
Septicemia-Mastitis
Vaccine
4
10 ml SC Nil Moderate
10 ml IM Nil Mild
Buffaloes
Montanide®
Adjuvanted Combined
Hemorrhagic
Septicemia-Mastitis
Vaccine
4 10 ml SC Nil Mild
10 ml IM Nil Nil
Control Placebo 4 No
treatment
Nil Nil
Nil Nil
SC=Subcutaneously; IM=intramuscularly; Mild: swelling at the injection site, which subside
within 48 hours; Moderate: swelling at the injection site, which subside within 3-7 days
Table 4.21: Comparative safety of Montanide® adjuvanted combined hemorrhagic
septicemia-mastitis vaccine in mice
Groups
of mice
Vaccine dose and route of
inoculation
Mice no. Mortality Morbidity
/patterns
M1 Montanide® Adjuvanted Combined
Hemorrhagic Septicemia-Mastitis
Vaccine
1 Nil Nil
2 Lethargy
3 Nil
4 Nil
5 Nil
M2 Placebo(control) 1 Nil Nil
2 Nil
3 Nil
4 Nil
5 Nil
None of the mice died following intramuscularly administration of 0.2 ml of the vaccine.
Only one mice had shown lethargy during first hours of after injection.
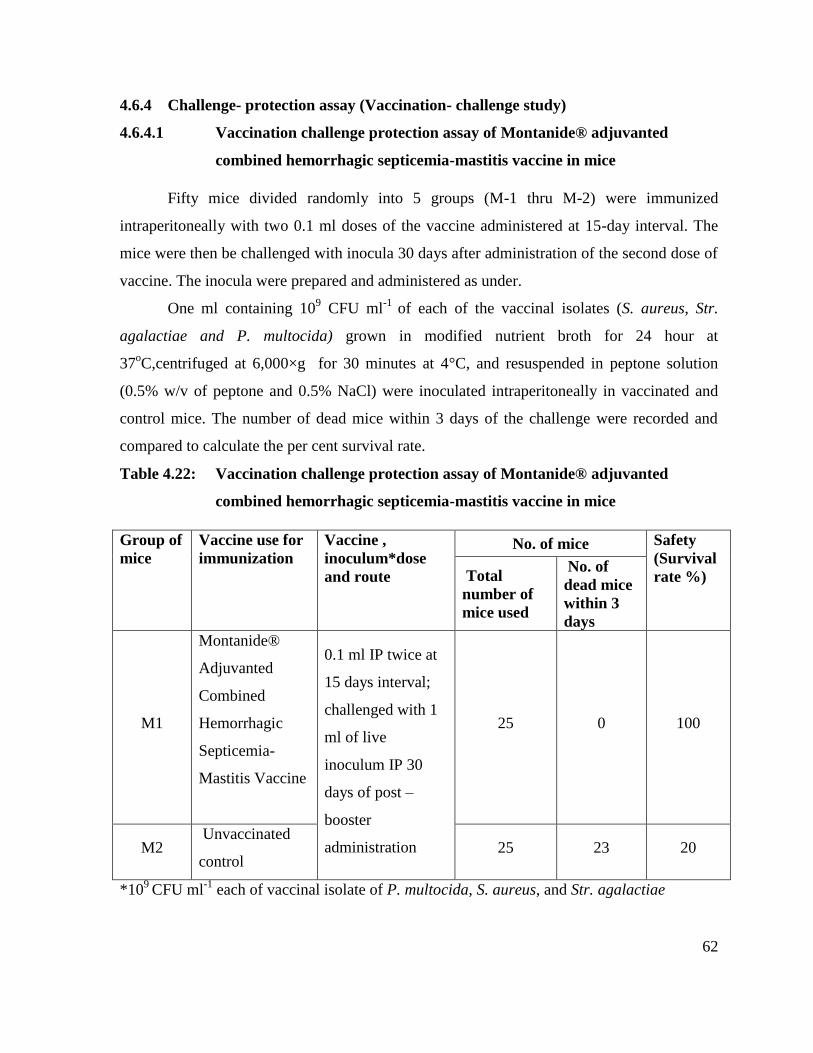
62
4.6.4 Challenge- protection assay (Vaccination- challenge study)
4.6.4.1 Vaccination challenge protection assay of Montanide® adjuvanted
combined hemorrhagic septicemia-mastitis vaccine in mice
Fifty mice divided randomly into 5 groups (M-1 thru M-2) were immunized
intraperitoneally with two 0.1 ml doses of the vaccine administered at 15-day interval. The
mice were then be challenged with inocula 30 days after administration of the second dose of
vaccine. The inocula were prepared and administered as under.
One ml containing 109 CFU ml
-1 of each of the vaccinal isolates (S. aureus, Str.
agalactiae and P. multocida) grown in modified nutrient broth for 24 hour at
37oC,centrifuged at 6,000×g for 30 minutes at 4°C, and resuspended in peptone solution
(0.5% w/v of peptone and 0.5% NaCl) were inoculated intraperitoneally in vaccinated and
control mice. The number of dead mice within 3 days of the challenge were recorded and
compared to calculate the per cent survival rate.
Table 4.22: Vaccination challenge protection assay of Montanide® adjuvanted
combined hemorrhagic septicemia-mastitis vaccine in mice
Group of
mice
Vaccine use for
immunization
Vaccine ,
inoculum*dose
and route
No. of mice Safety
(Survival
rate %) Total
number of
mice used
No. of
dead mice
within 3
days
M1
Montanide®
Adjuvanted
Combined
Hemorrhagic
Septicemia-
Mastitis Vaccine
0.1 ml IP twice at
15 days interval;
challenged with 1
ml of live
inoculum IP 30
days of post –
booster
administration
25 0 100
M2 Unvaccinated
control 25 23 20
*109
CFU ml-1
each of vaccinal isolate of P. multocida, S. aureus, and Str. agalactiae

63
None of the mice in group M-1 (vaccination twice before challenging) died within 3
days of the challenging IP(Intra peritoneal) with I ml of live inoculum containing 109
CFU
ml-1
of each of the vaccinal isolates whereas only two mouse survived from the control
group (which also died later on day 4). This translated into green signal for all the
experimental Montanide® Adjuvanted Combined Hemorrhagic Septicemia-Mastitis Vaccine
to be used safely in the future trials whereas death of the 23 mice in the unvaccinated control
group (within three days) proved its lethality.
4.6.5 ELISA- based antibody titers generated by Montanide® adjuvanted combined
HS-mastitis vaccine against P. multocida, S. aureus and Str. agalactiae in cows
and buffaloes
Prevaccination serum IgG antibody titers (optical density values; OD values) against
P. multocida, S. aureus and Strep agalactiae were not significantly different in vaccinated
and unvaccinated groups (Table 4.23). The highest anti-staphylococcal IgG values (OD
2.56) were recorded in non-mastitic group at day 60 that remained significantly higher
throughout the observation period (180 days) than prevaccination titer (day zero).
The result of present study revealed that the administration of Montanide®
adjuvanted combined HS-mastitis vaccine produced higher antibody titers against P.
multocida, S. aureus and Str. agalactiae in vaccinated group (A and C) compared to un-
vaccinated groups( B and D) as shown in table 4.23 and Fig. 4.2. The antibody titers were
higher in non-diseased vaccinated (group A) animals compared to diseased vaccinated
animals ( group C) at 0, 60, 120 and 180 days of post -vaccination as evident in Table 4.23.
It was observed that the antibody titers against P. multocida, S. aureus and Str.
agalactiae augmented at 60 and 120 days of vaccine administration in non-diseased
vaccinated animals (Table 4.23) and decreased at 180 days of post-vaccination (Table 4.23).
Data was statistically designed by 2 factor CRD and comparison of means was done by using
Tukey‟s test at 5% level of significance.
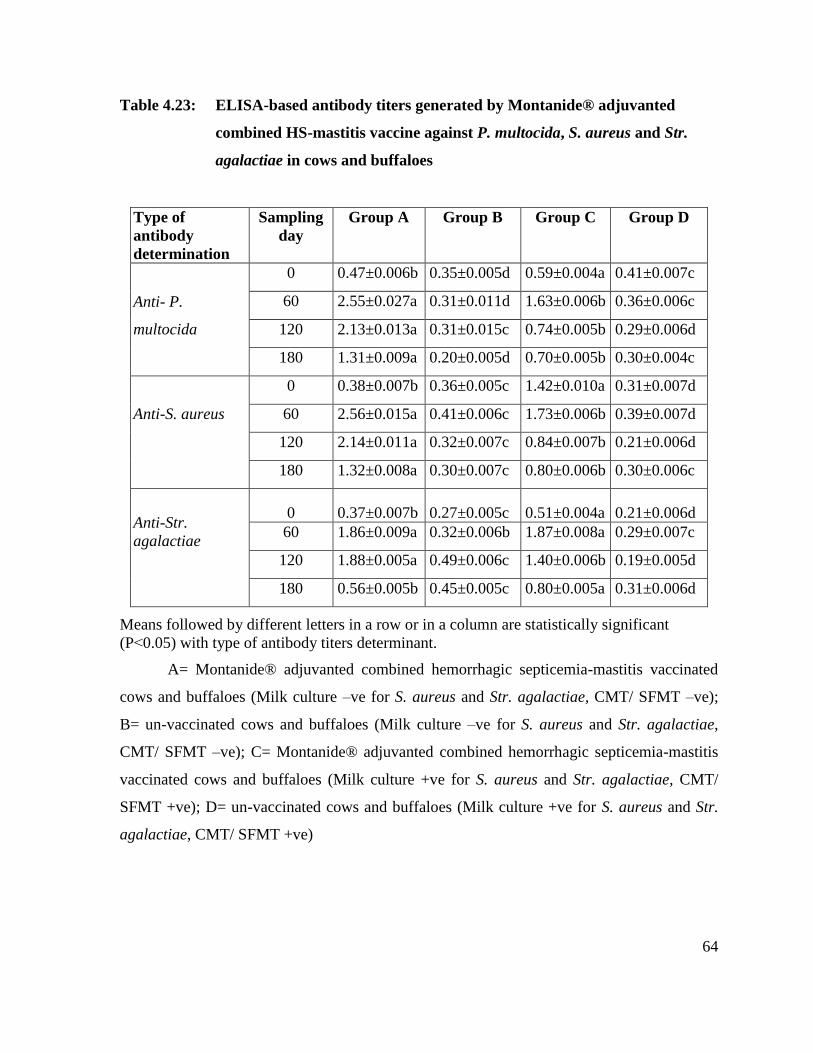
64
Table 4.23: ELISA-based antibody titers generated by Montanide® adjuvanted
combined HS-mastitis vaccine against P. multocida, S. aureus and Str.
agalactiae in cows and buffaloes
Type of
antibody
determination
Sampling
day
Group A Group B Group C Group D
Anti- P.
multocida
0 0.47±0.006b 0.35±0.005d 0.59±0.004a 0.41±0.007c
60 2.55±0.027a 0.31±0.011d 1.63±0.006b 0.36±0.006c
120 2.13±0.013a 0.31±0.015c 0.74±0.005b 0.29±0.006d
180 1.31±0.009a 0.20±0.005d 0.70±0.005b 0.30±0.004c
Anti-S. aureus
0 0.38±0.007b 0.36±0.005c 1.42±0.010a 0.31±0.007d
60 2.56±0.015a 0.41±0.006c 1.73±0.006b 0.39±0.007d
120 2.14±0.011a 0.32±0.007c 0.84±0.007b 0.21±0.006d
180 1.32±0.008a 0.30±0.007c 0.80±0.006b 0.30±0.006c
Anti-Str.
agalactiae
0 0.37±0.007b 0.27±0.005c 0.51±0.004a 0.21±0.006d
60 1.86±0.009a 0.32±0.006b 1.87±0.008a 0.29±0.007c
120 1.88±0.005a 0.49±0.006c 1.40±0.006b 0.19±0.005d
180 0.56±0.005b 0.45±0.005c 0.80±0.005a 0.31±0.006d
Means followed by different letters in a row or in a column are statistically significant
(P˂0.05) with type of antibody titers determinant.
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated
cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve);
B= un-vaccinated cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae,
CMT/ SFMT –ve); C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve); D= un-vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str.
agalactiae, CMT/ SFMT +ve)
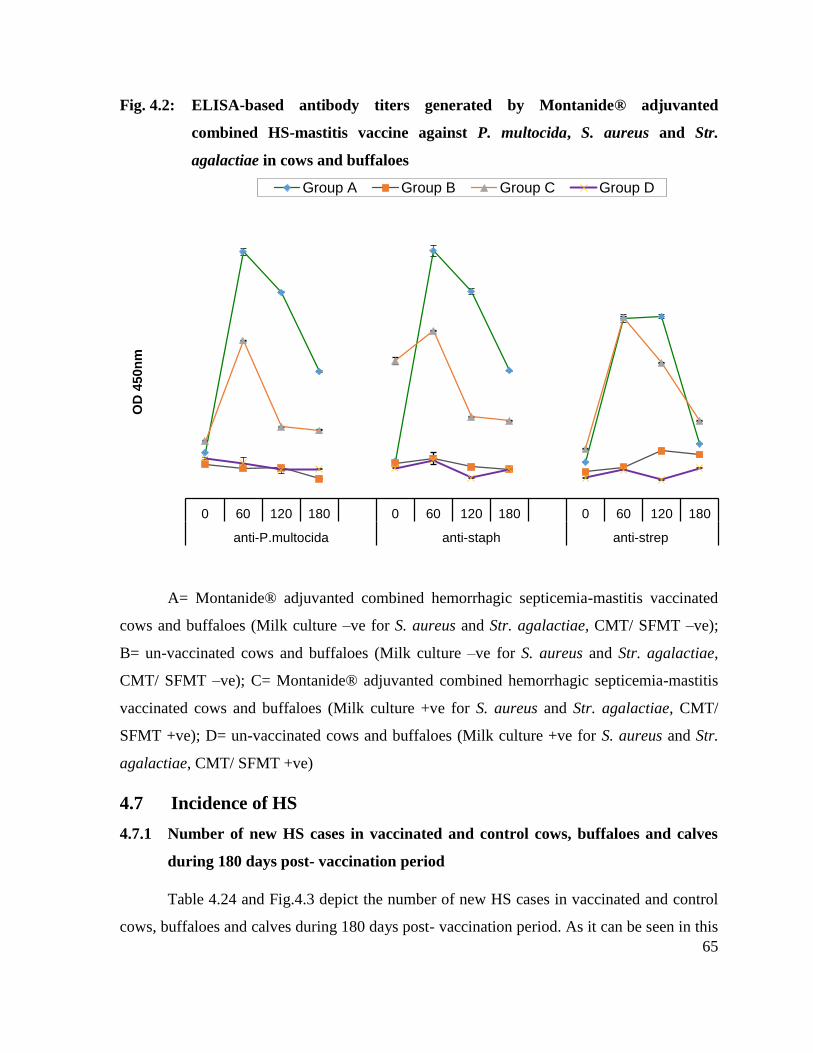
65
Fig. 4.2: ELISA-based antibody titers generated by Montanide® adjuvanted
combined HS-mastitis vaccine against P. multocida, S. aureus and Str.
agalactiae in cows and buffaloes
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated
cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve);
B= un-vaccinated cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae,
CMT/ SFMT –ve); C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve); D= un-vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str.
agalactiae, CMT/ SFMT +ve)
4.7 Incidence of HS
4.7.1 Number of new HS cases in vaccinated and control cows, buffaloes and calves
during 180 days post- vaccination period
Table 4.24 and Fig.4.3 depict the number of new HS cases in vaccinated and control
cows, buffaloes and calves during 180 days post- vaccination period. As it can be seen in this
0 60 120 180 0 60 120 180 0 60 120 180
anti-P.multocida anti-staph anti-strep
OD
450n
m
Group A Group B Group C Group D
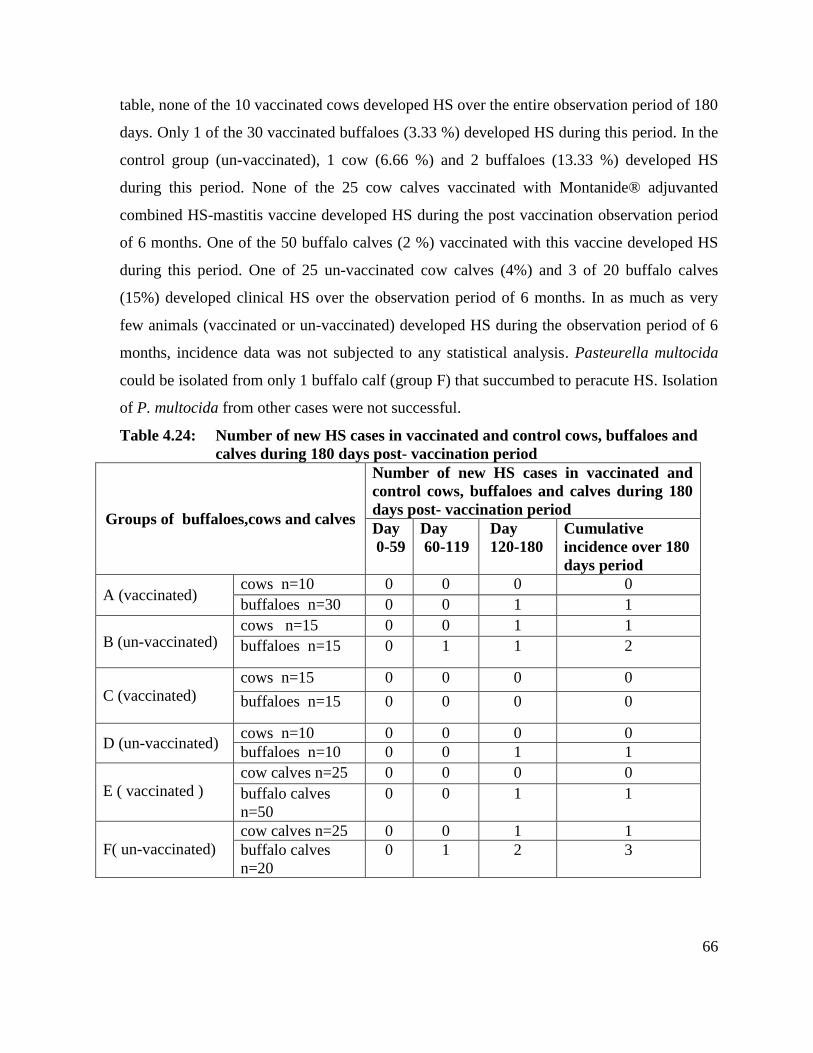
66
table, none of the 10 vaccinated cows developed HS over the entire observation period of 180
days. Only 1 of the 30 vaccinated buffaloes (3.33 %) developed HS during this period. In the
control group (un-vaccinated), 1 cow (6.66 %) and 2 buffaloes (13.33 %) developed HS
during this period. None of the 25 cow calves vaccinated with Montanide® adjuvanted
combined HS-mastitis vaccine developed HS during the post vaccination observation period
of 6 months. One of the 50 buffalo calves (2 %) vaccinated with this vaccine developed HS
during this period. One of 25 un-vaccinated cow calves (4%) and 3 of 20 buffalo calves
(15%) developed clinical HS over the observation period of 6 months. In as much as very
few animals (vaccinated or un-vaccinated) developed HS during the observation period of 6
months, incidence data was not subjected to any statistical analysis. Pasteurella multocida
could be isolated from only 1 buffalo calf (group F) that succumbed to peracute HS. Isolation
of P. multocida from other cases were not successful.
Table 4.24: Number of new HS cases in vaccinated and control cows, buffaloes and
calves during 180 days post- vaccination period
Groups of buffaloes,cows and calves
Number of new HS cases in vaccinated and
control cows, buffaloes and calves during 180
days post- vaccination period
Day
0-59
Day
60-119
Day
120-180
Cumulative
incidence over 180
days period
A (vaccinated) cows n=10 0 0 0 0
buffaloes n=30 0 0 1 1
B (un-vaccinated) cows n=15 0 0 1 1
buffaloes n=15 0 1 1 2
C (vaccinated)
cows n=15 0 0 0 0
buffaloes n=15 0 0 0 0
D (un-vaccinated) cows n=10 0 0 0 0
buffaloes n=10 0 0 1 1
E ( vaccinated )
cow calves n=25 0 0 0 0
buffalo calves
n=50
0 0 1 1
F( un-vaccinated)
cow calves n=25 0 0 1 1
buffalo calves
n=20
0 1 2 3

67
A= Montanide® adjuvanted combined HS-mastitis vaccinated cows and buffaloes
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS
vaccination during the last 180 days); B= un-vaccinated cows and buffaloes (Milk culture –
ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS vaccination during
the last 180 days); C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve); D= un-vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str.
agalactiae, CMT/ SFMT +ve); E= vaccinated cow and buffalo calves; no history of HS
vaccination during the last 180 days F= un-vaccinated cow and buffalo calves; no history of
HS vaccination during the last 180 days.
Fig. 4.3: Number of new HS cases in vaccinated and control cows, buffaloes and
calves during 180 days post- vaccination period
A= Montanide® adjuvanted combined HS-mastitis vaccinated cows and buffaloes
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS
vaccination during the last 180 days); B= un-vaccinated cows and buffaloes (Milk culture –
ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS vaccination during
the last 180 days); E= vaccinated cow and buffalo calves; no history of HS vaccination
0
0.5
1
1.5
2
2.5
3
3.5
A B C D E F
Nu
mb
er o
f n
ew H
S
Day Day Day Cumulative incidence over 180 days period

68
during the last 180 days F= un-vaccinated cow and buffalo calves; no history of HS
vaccination during the last 180 days.
4.8 Incidence and prevalence of S. aureus and Str. agalactiae mastitis
4.8.1 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter-
based incidence of Str. agalactiae and S. aureus IMIs in cows and buffaloes
initially culture negative and CMT/ SFMT negative
Tabel 4.25 and Fig. 4.4 depict the effect of Montanide® adjuvanted combined HS-
mastitis vaccine on quarter-based incidence of Str. agalactiae and S. aureus IMIs in cows
and buffaloes initially culture negative and CMT/ SFMT negative. As can be seen in these
depictions, of the 10 vaccinated cows, only 1 quarter each of two cows developed new Str.
agalactiae and S. aureus intramammary infection (IMI) over the 6 months period. None of
the 30 vaccinated buffaloes developed new IMI with Str. agalactiae whereas 1 quarter
developed new S. aureus IMI. Of the 15 un-vaccinated cows, new IMI with Str. agalactiae
was detected in one quarter whereas 5 quarters of 4 un-vaccinated cows developed new IMI
with S. aureus over a period of 180 days. New IMIs with Str. agalactiae and S. aureus were
detected in 1 and 3 quarters respectively of 15 un-vaccinated buffaloes. Amalgamating the
new Str. agalactiae and S. aureus IMIs in vaccinated cows and buffaloes and comparing the
corresponding figure with that of un-vaccinated cows and buffaloes would indicate that the
quarter–based incidence of these mastitis 2 pathogens over the 6 months observation period
was 3.3 folds higher in un-vaccinated cows and buffaloes.

69
Table 4.25: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter-based incidence of Str. agalactiae and S. aureus IMIs in cows and
buffaloes initially culture negative and CMT/ SFMT negative
Groups of cows and buffaloes Quarter based cumulative incidence over 180
days post vaccination period
A (vaccinated)
Cows n=10 Str. agalactiae 1
2 S. aureus 1
Buffaloes n=30 Str. agalactiae 0
1 S. aureus 1
B (un-
vaccinated)
Cows n=15 Str. agalactiae 1
6 S. aureus 5
Buffaloes n=15 Str. agalactiae 1
4 S. aureus 3
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve) B= un-
vaccinated cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/
SFMT –ve)
Fig 4.4: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter-based incidence of Str. agalactiae and S. aureus IMIs in cows and
buffaloes initially culture negative and CMT/ SFMT negative
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve) B= un-
vaccinated cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/
SFMT –ve)
0
1
2
3
4
5
6
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Cows n=10 Buffaloes n=30 Cows n=15 Buffaloes n=15
A (vaccinated) B (un-vaccinated)
Ov
era
ll n
ew
In
fecti
on
s (
incid
en
ce)
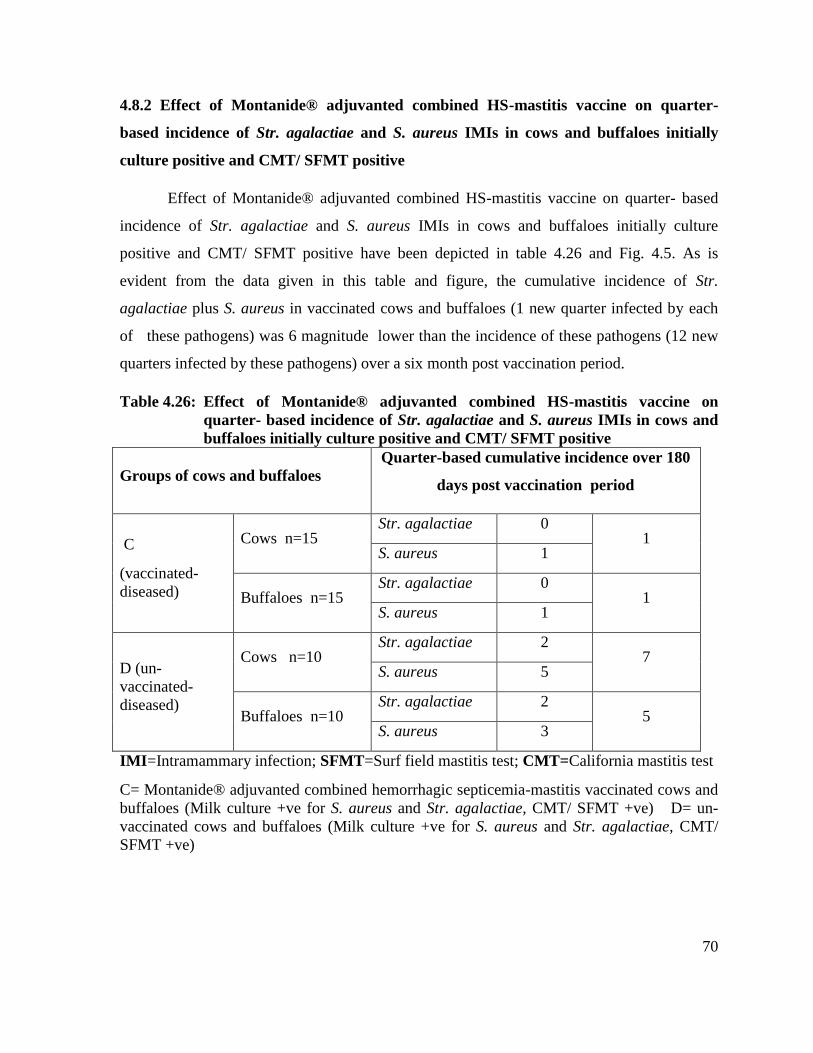
70
4.8.2 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter-
based incidence of Str. agalactiae and S. aureus IMIs in cows and buffaloes initially
culture positive and CMT/ SFMT positive
Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter- based
incidence of Str. agalactiae and S. aureus IMIs in cows and buffaloes initially culture
positive and CMT/ SFMT positive have been depicted in table 4.26 and Fig. 4.5. As is
evident from the data given in this table and figure, the cumulative incidence of Str.
agalactiae plus S. aureus in vaccinated cows and buffaloes (1 new quarter infected by each
of these pathogens) was 6 magnitude lower than the incidence of these pathogens (12 new
quarters infected by these pathogens) over a six month post vaccination period.
Table 4.26: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter- based incidence of Str. agalactiae and S. aureus IMIs in cows and
buffaloes initially culture positive and CMT/ SFMT positive
Groups of cows and buffaloes
Quarter-based cumulative incidence over 180
days post vaccination period
C
(vaccinated-
diseased)
Cows n=15 Str. agalactiae 0
1 S. aureus 1
Buffaloes n=15 Str. agalactiae 0
1 S. aureus 1
D (un-
vaccinated-
diseased)
Cows n=10 Str. agalactiae 2
7 S. aureus 5
Buffaloes n=10 Str. agalactiae 2
5 S. aureus 3
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve) D= un-
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve)

71
Fig: 4.5: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter- based incidence of Str. agalactiae and S. aureus IMIs in cows and
buffaloes initially culture positive and CMT/ SFMT positive
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve) D= un-
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve)
0
1
2
3
4
5
6S
tr. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Cows n=15 Buffaloes n=15 Cows n=10 Buffaloes n=10
C (vaccinated- diseased) D (un-vaccinated-diseased)
Ov
era
ll n
ew
In
fecti
on
s (
incid
en
ce)

72
4.8.3 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on quarter-
based time points prevalence of Str. agalactiae and S. aureus IMIs in cows and
buffaloes initially culture positive and CMT/SFMT positive
Table 4.27 and figure 4.6 show the effect of Montanide® adjuvanted combined HS-
mastitis vaccine on quarter- based time points prevalence of Str. agalactiae and S. aureus
IMIs in cows and buffaloes initially culture positive and CMT/SFMT positive. As can be
seen in these depictions, at day 0, the number of Str. agalactiae and S. aureus infected
quarters of vaccinated –diseased group (group C) was numerically comparable to Str.
agalactiae and S. aureus infected quarters of un-vaccinated –diseased group (i.e. group D).
At day 60, day 120 and day 180 post vaccination, the number of Str. agalactiae and S.
aureus infected quarters in vaccinated –diseased group (group C) was lower than the number
of quarters infected by these two pathogens at day 0. In other words, vaccination reduced the
prevalence of Str. agalactiae and S. aureus IMIs over the entire observation period of 180
days. Towards the end of study period (i.e. day 180), the prevalence of Str. agalactiae and S.
aureus IMIs was slightly higher than the prevalence of these pathogens at day 60 and day
120. In the un-vaccinated- diseased group (group D), the number of Str. agalactiae and S.
aureus infected quarters increased over the observation period of 180 days. Thus the
prevalence of these pathogens at day 60, day 120, and day 180 was higher than their
prevalence at day 0.

73
Table 4.27: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter- based time points prevalence of Str. agalactiae and S. aureus
IMIs in cows and buffaloes initially culture positive and CMT/ SFMT
positive
Groups of cows and
buffaloes
Number of quarters detected
culturally positive for Str.
agalactiae and S. aureus at
different days post vaccination
Day 0 Day 60 Day 120 Day 180
C
(Vaccinated)
Cows
n=15
Str. agalactiae 9 (15%) 4(6.7%) 3(5%) 4(6.7%)
S. aureus 17(28.3%) 10(16.7%) 10(16.7%) 11(18.3%)
Buffaloes
n=15
Str. agalactiae 6(10%) 2(3.3%) 2(3.3%) 3(5%)
S. aureus 12(20%) 6(10%) 6(10%) 7(11.7%)
D (un-
vaccinated)
Cows
n=10
Str. agalactiae 5(12.5%) 7(17.5%) 7(17.5%) 8(20%)
S. aureus 12(30%) 17(42.5%) 16(40%) 17(42.5%)
Buffaloes
n=10
Str. agalactiae 3(7.5%) 5(12.5%) 5(12.5%) 4(10%)
S. aureus 8(20%) 11(27.5%) 11(27.5%) 10(25%)
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvantead combined hemorrhagic septicemia-mastitis vaccinated cows
and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/ SFMT +ve); D= un-
vaccinated cows and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/
SFMT +ve)
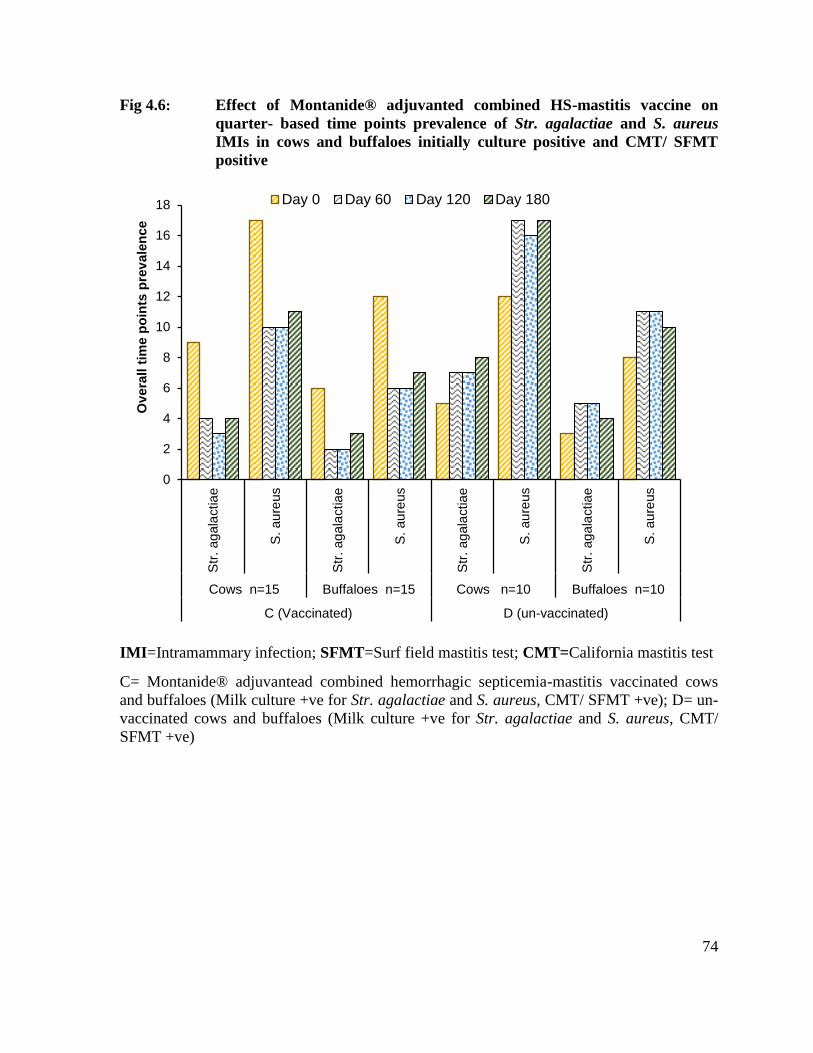
74
Fig 4.6: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter- based time points prevalence of Str. agalactiae and S. aureus
IMIs in cows and buffaloes initially culture positive and CMT/ SFMT
positive
IMI=Intramammary infection; SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvantead combined hemorrhagic septicemia-mastitis vaccinated cows
and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/ SFMT +ve); D= un-
vaccinated cows and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/
SFMT +ve)
0
2
4
6
8
10
12
14
16
18S
tr. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Str
. ag
ala
ctiae
S. au
reus
Cows n=15 Buffaloes n=15 Cows n=10 Buffaloes n=10
C (Vaccinated) D (un-vaccinated)
Ov
era
ll t
ime p
oin
ts p
rev
ale
nce
Day 0 Day 60 Day 120 Day 180

75
4.9 Mastitis screening tests based prevalence and incidence of mastitis
4.9.1 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and
SFMT based quarter incidence of mastitis in cows and buffaloes initially culture
negative for S. agalactiae and S. aureus and CMT/ SFMT negative
Table 4.28 and Figure 4.7 depict the quarter-based incidence rates of new subclinical
and clinical mastitis cases in vaccinated and control cows, and buffaloes (initially culture
negative for S. agalactiae and S. aureus and CMT/SFMT negative) during 180 days post-
vaccination period. As it can be seen in these depictions, 4 of 40 quarters (10%) of
vaccinated cows developed mastitis over the entire observation period of 180 days. Only 2 of
the 120 quarters (1.6 %) of vaccinated buffaloes developed mastitis during this period. In the
control group (un-vaccinated), 8 (13.33%) quarters of 15 cow and 6 (10%) quarters of 15
buffaloes developed mastitis during this period. In as much as a fairly few quarters of
vaccinated or unvaccinated developed subclinical or clinical mastitis during the observation
period of 6 months, incidence data were not subjected to any statistical analysis.
Table 4.28: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and
SFMT-based quarter incidence of mastitis in cows and buffaloes initially culture
negative for S. agalactiae and S. aureus and CMT/ SFMT negative
Groups of cows and buffaloes
Number of new quarters reacting
positive in CMT and SFMT over
180 days post –vaccination period
A ( vaccinated –culture –ve for Str.
agalactiae and S. aureus and
CMT/ SFMT –ve )
Cows n=10 4 (10 % )
Buffaloes
n=30 2 (1.6 % )
B (un- vaccinated –culture –ve for
Str. agalactiae and S. aureus and
CMT/ SFMT –ve)
Cows n=15 8 (13.33 % )
Buffaloes
n=15 6 (10 % )
SFMT=Surf field mastitis test; CMT= California mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture –ve for Str. agalactiae and S. aureus, CMT/SFMT –ve) B= un-
vaccinated cows and buffaloes (Milk culture –ve for Str. agalactiae and S. aureus, CMT
/SFMT –ve)
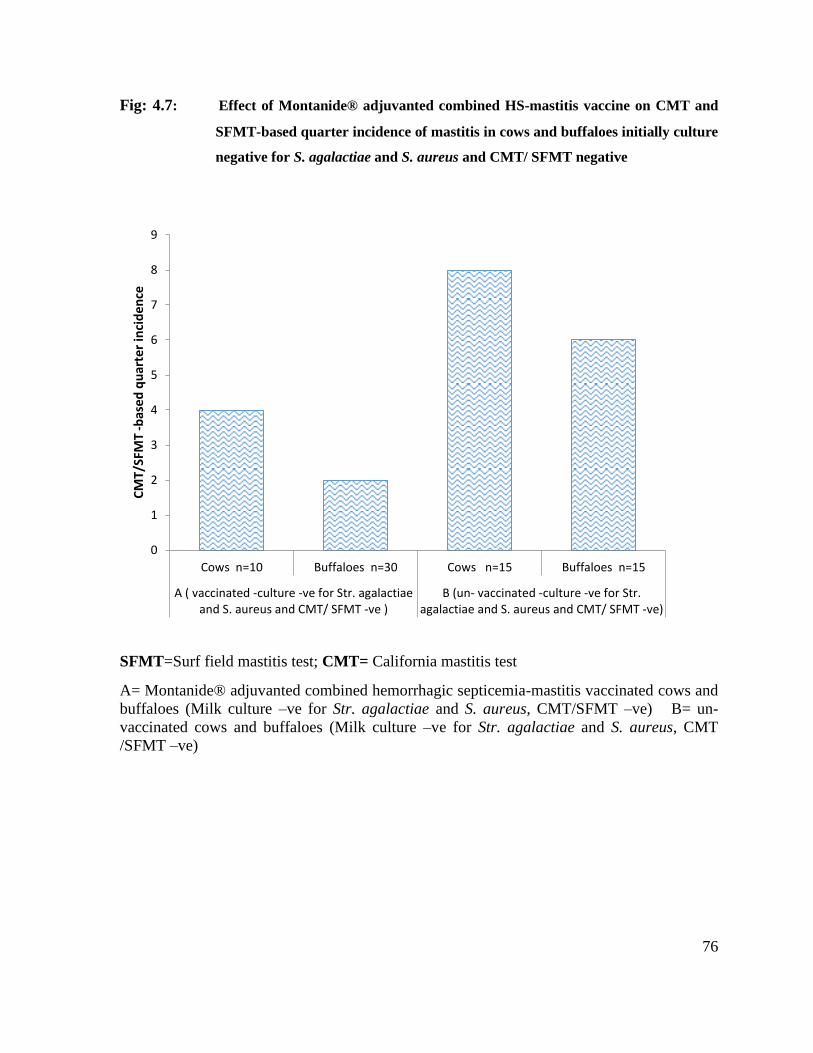
76
Fig: 4.7: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and
SFMT-based quarter incidence of mastitis in cows and buffaloes initially culture
negative for S. agalactiae and S. aureus and CMT/ SFMT negative
SFMT=Surf field mastitis test; CMT= California mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture –ve for Str. agalactiae and S. aureus, CMT/SFMT –ve) B= un-
vaccinated cows and buffaloes (Milk culture –ve for Str. agalactiae and S. aureus, CMT
/SFMT –ve)
0
1
2
3
4
5
6
7
8
9
Cows n=10 Buffaloes n=30 Cows n=15 Buffaloes n=15
A ( vaccinated -culture -ve for Str. agalactiaeand S. aureus and CMT/ SFMT -ve )
B (un- vaccinated -culture -ve for Str.agalactiae and S. aureus and CMT/ SFMT -ve)
CM
T/S
FMT
-bas
ed
qu
arte
r in
cid
en
ce

77
4.9.2 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT- based quarter time points prevalence of mastitis in cows and
buffaloes initially negative for Str. agalactiae and S. aureus plus CMT/SFMT
negative
Table 4.29 and Figure 4.8 show the effect of Montanide® adjuvanted combined HS-
mastitis vaccine on CMT/SFMT- based quarter time points prevalence of mastitis in cows
and buffaloes initially negative for Str. agalactiae and S. aureus plus CMT/SFMT negative.
As can be seen in these depictions, at day 0, the numbers of CMT/SFMT positive quarters of
vaccinated cows and buffaloes (group A) were broadly comparable to the corresponding
numbers in control (un-vaccinated; group B) cows and buffaloes. In the vaccinated cows, an
increase of 2.5 % was noted at day 60 and day 120. At day 180 a further increase of 7.5 % in
the CMT/SFMT quarter prevalence of mastitis was recorded. In vaccinated buffaloes,
CMT/SFMT quarter prevalence of mastitis did not change much; quarter prevalence at day 0,
day 160, day 120, and day 180 being 4.16, 3.33, 4.16, and 5%, respectively. In the case of
un-vaccinated cows (control), increases of 11.67, 13.34 and 13.34 % were registered at day
60, day 120 and day 180, respectively. Un-vaccinated buffaloes witnessed a sharp increase of
8.33% in CMT/SFMT- based quarter prevalence of mastitis at day 60 whereas at day 120 and
180 a steady small further increases of CMT/SFMT-based quarter prevalence of mastitis was
observed.

78
Table 4.29: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT- based quarter time points prevalence of mastitis in cows
and buffaloes initially negative for Str. agalactiae and S. aureus plus
CMT/SFMT negative
Groups of cows and buffaloes
Number of quarters reacting positive in
CMT/SFMT at different days post
vaccination
Day 0 Day 60 Day 120 Day 180
A(vaccinated –
culture –ve for Str.
agalactiae and S.
aureus and CMT/
SFMT –ve )
Cows n=10 3 (7.5%) 4 (10%) 4 (10 %) 6 (15%)
Buffaloes
n=30 5 (4.16% ) 4 (3.33 % ) 5 (4.16 %) 6 (5 %)
B (un- vaccinated –
culture –ve for Str.
agalactiae and S.
aureus and CMT/
SFMT –ve)
Cows
n=15 5 (8.33% ) 12 (20 % ) 13 (21.67%) 13 (21.67 %)
Buffaloes
n=15 3 (5% ) 8 (13.33% ) 9 (15 %) 10 (16.67%)
CMT= California mastitis test; SFMT=Surf field mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture -ve for Str. agalactiae and S. aureus CMT /SFMT -ve); B= un-
vaccinated cows and buffaloes (Milk culture -ve for Str. agalactiae and S. aureus, CMT
/SFMT -ve)
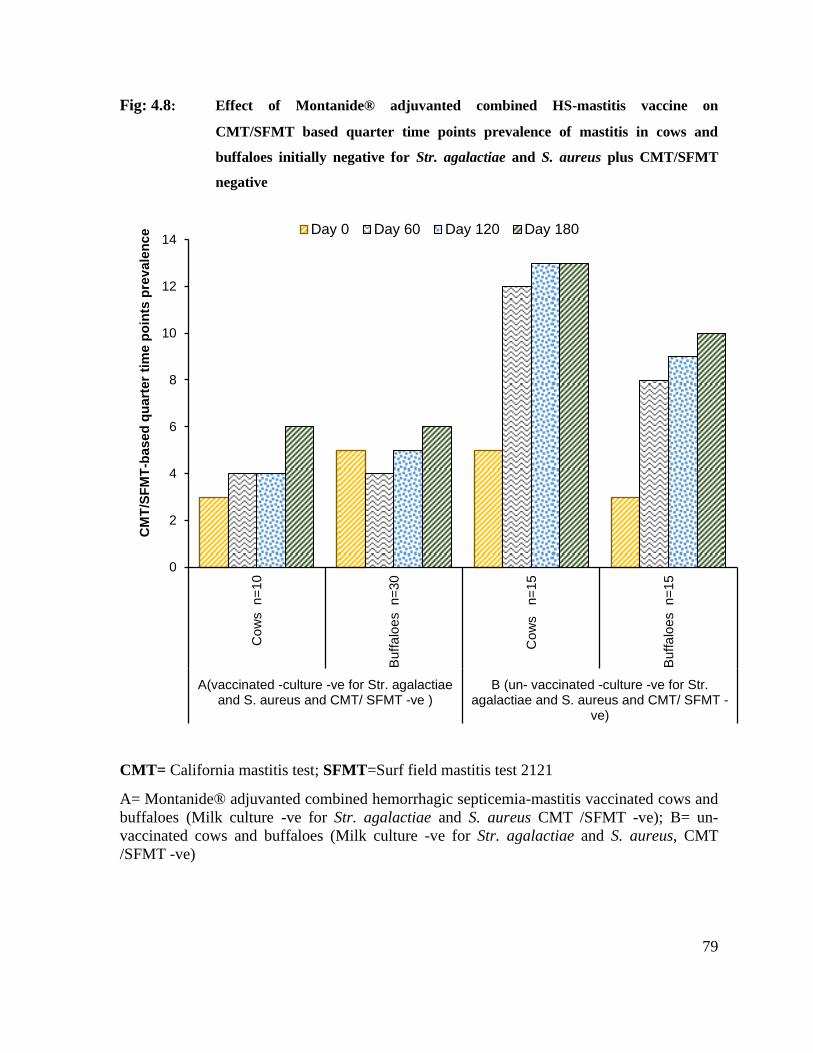
79
Fig: 4.8: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT based quarter time points prevalence of mastitis in cows and
buffaloes initially negative for Str. agalactiae and S. aureus plus CMT/SFMT
negative
CMT= California mastitis test; SFMT=Surf field mastitis test 2121
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture -ve for Str. agalactiae and S. aureus CMT /SFMT -ve); B= un-
vaccinated cows and buffaloes (Milk culture -ve for Str. agalactiae and S. aureus, CMT
/SFMT -ve)
0
2
4
6
8
10
12
14
Co
ws
n=
10
Buffa
loe
s
n=
30
Co
ws
n=
15
Buffa
loe
s
n=
15
A(vaccinated -culture -ve for Str. agalactiaeand S. aureus and CMT/ SFMT -ve )
B (un- vaccinated -culture -ve for Str.agalactiae and S. aureus and CMT/ SFMT -
ve)
CM
T/S
FM
T-b
ased
qu
art
er
tim
e p
oin
ts p
rev
ale
nce
Day 0 Day 60 Day 120 Day 180

80
4.9.3 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT and
SFMT based quarter incidence of mastitis in cows and buffaloes initially culture
positive for S. agalactiae and S. aureus and CMT/ SFMT positive
Three additional quarters of 15 vaccinated- disease cows (group C) reacted positive in
CMT/ SFMT over 180 days post vaccination period. The corresponding figure in 15
vaccinated-diseased buffaloes (group C) was 2. In the un-vaccinated-diseased cows (group
D), 8 new quarters developed CMT/ SFMT positivity over 180 days post vaccination period
whereas the corresponding figure for the un-vaccinated –diseased buffaloes (group D) was 6
( table 4.30 , Figure 4.9).
Table 4.30: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT
and SFMT based quarter incidence of mastitis in cows and buffaloes
initially culture positive for S. agalactiae and S. aureus and CMT/ SFMT
positive
SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve) D= un-
vaccinated- diseased cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae,
CMT/ SFMT +ve)
Groups of cows and buffaloes Quarter-based cumulative incidence (new
cases) over 180 days post vaccination period
C (vaccinated- diseased)
Cows
n=15 3
Buffaloes
n=15 2
D (un-vaccinated-diseased)
Cows
n=10 8
Buffaloes
n=10 6

81
Fig: 4.9: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on CMT
and SFMT based quarter incidence of mastitis in cows and buffaloes
initially culture positive for S. agalactiae and S. aureus and CMT/ SFMT
positive
SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve) D= un-
vaccinated- diseased cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae,
CMT/ SFMT +ve)
0
1
2
3
4
5
6
7
8
9
Cows n=15 Buffaloes n=15 Cows n=10 Buffaloes n=10
C (vaccinated- diseased) D (un-vaccinated-diseased)
CM
T/SF
MT-
bas
ed q
uar
ter
inci
den
ce

82
4.9.4 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT- based time points quarter prevalence of mastitis in cows and
buffaloes initially positive for Str. agalactiae and S. aureus plus CMT/SFMT
positive
Table 4.31 and Figure 4.10 show the effect of Montanide® adjuvanted combined HS-
mastitis vaccine on CMT/SFMT- based quarter time points prevalence of mastitis in cows
and buffaloes initially positive for Str. agalactiae and S. aureus plus CMT/SFMT positive.
As can be seen in these depictions ,at day 0, the number positive quarters of vaccinated –
diseased group (group C) was numerically comparable to positive quarters of un-vaccinated
–diseased group (i.e. group D). At day 60, day 120 and day 180 post vaccination, the number
of positive quarters in vaccinated –diseased group (group C) was lower than the number of
quarters positive at day 0. In other words, vaccination reduced the prevalence over the entire
observation period of 180 days. Towards the end of study period (i.e day 180), the prevalence
was slightly higher than at day 60 and day 120.
In the un-vaccinated- diseased group (group D), the number of positive quarters increased
over the observation period of 180 days. Thus the prevalence of these pathogens at day 60,
day 120, and day 180 was higher than their prevalence at day 0.

83
Table 4.31: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT- based time points quarter prevalence of mastitis in cows
and buffaloes initially positive for Str. agalactiae and S. aureus plus
CMT/SFMT positive
Groups of cows and buffaloes
Number of quarters detected positive by
CMT/SFMT at different post vaccination
days
Day 0 Day 60 Day 120 Day 180
C
(Vaccinated)
Cows n=15 24(40%) 17 (28.33%) 16 (26.67 %) 18 (30%)
Buffaloes n=15 16 (26.67%) 10 (16.67%) 10 (1 6.67%) 12 (20 %)
D (un-
vaccinated)
Cows n=10 17 (42.50 %) 32 (80 %) 31 (77.5 %) 33 (82.5 %)
Buffaloes n=10 11 (27.5 %) 22 (55%) 22 (55%) 20 (50 %)
SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/ SFMT +ve); D= un-
vaccinated cows and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/
SFMT +ve)
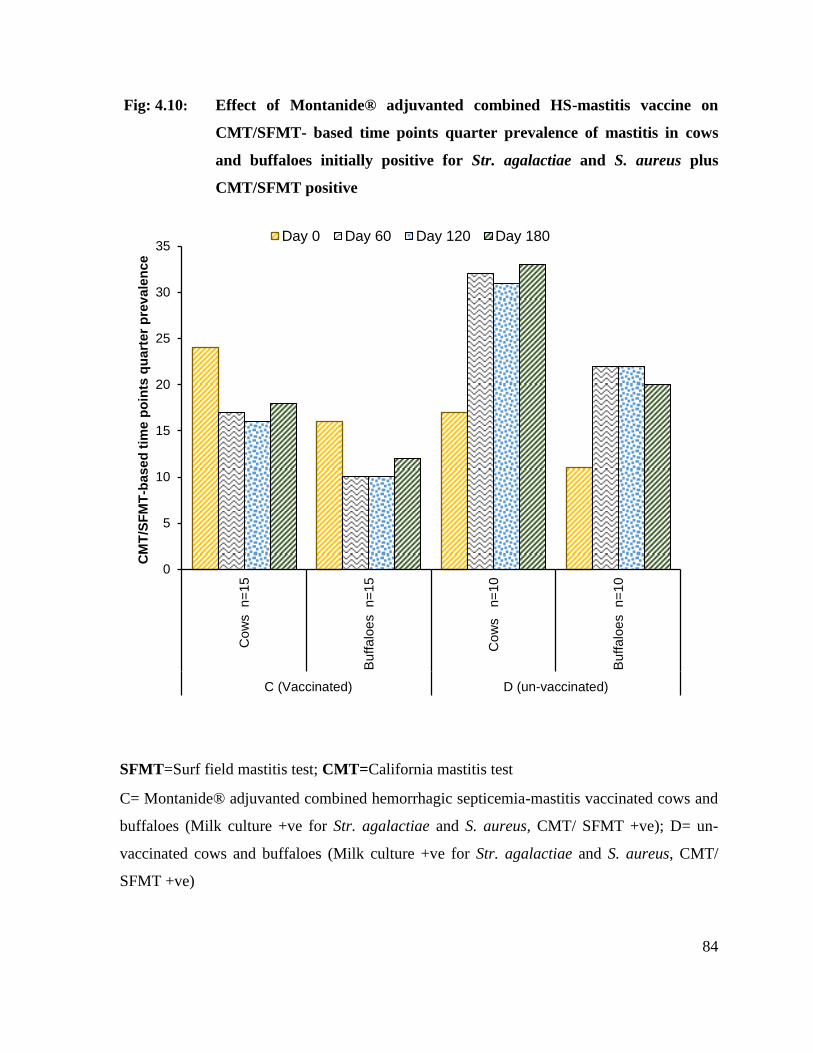
84
Fig: 4.10: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on
CMT/SFMT- based time points quarter prevalence of mastitis in cows
and buffaloes initially positive for Str. agalactiae and S. aureus plus
CMT/SFMT positive
SFMT=Surf field mastitis test; CMT=California mastitis test
C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/ SFMT +ve); D= un-
vaccinated cows and buffaloes (Milk culture +ve for Str. agalactiae and S. aureus, CMT/
SFMT +ve)
0
5
10
15
20
25
30
35C
ow
s
n=
15
Buffa
loe
s
n=
15
Co
ws
n=
10
Buffa
loe
s
n=
10
C (Vaccinated) D (un-vaccinated)
CM
T/S
FM
T-b
ased
tim
e p
oin
ts q
uart
er
pre
vale
nce
Day 0 Day 60 Day 120 Day 180
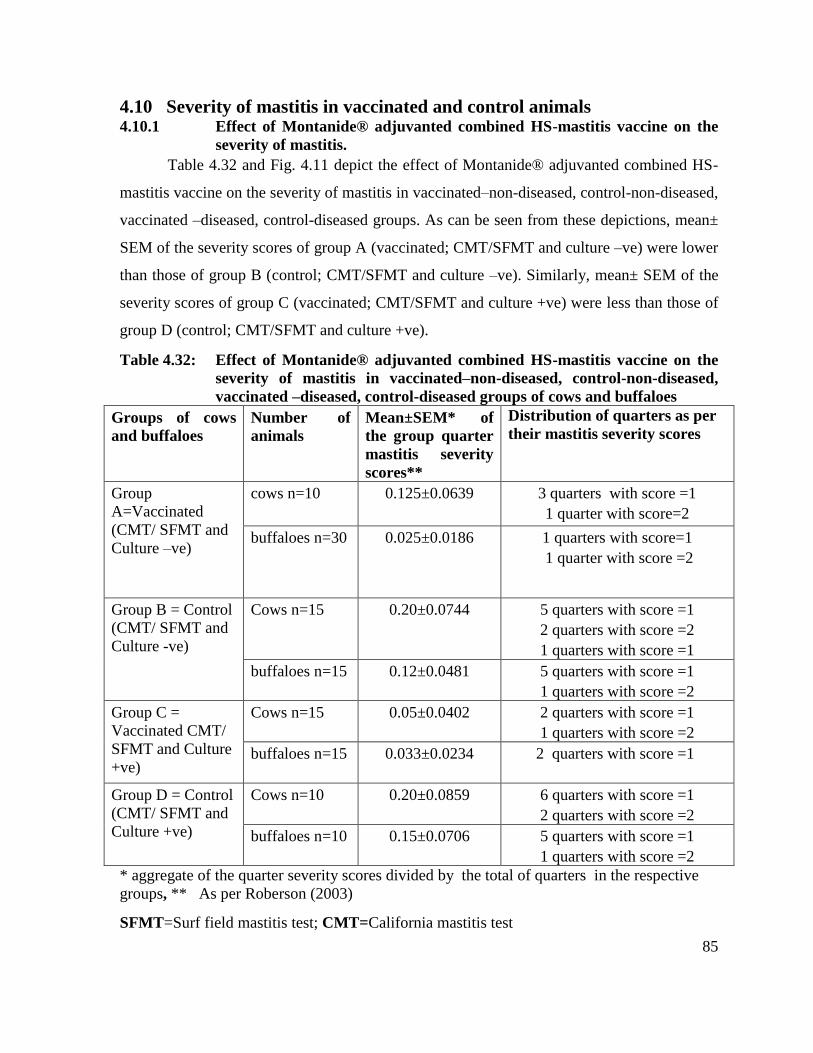
85
4.10 Severity of mastitis in vaccinated and control animals 4.10.1 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
severity of mastitis.
Table 4.32 and Fig. 4.11 depict the effect of Montanide® adjuvanted combined HS-
mastitis vaccine on the severity of mastitis in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups. As can be seen from these depictions, mean±
SEM of the severity scores of group A (vaccinated; CMT/SFMT and culture –ve) were lower
than those of group B (control; CMT/SFMT and culture –ve). Similarly, mean± SEM of the
severity scores of group C (vaccinated; CMT/SFMT and culture +ve) were less than those of
group D (control; CMT/SFMT and culture +ve).
Table 4.32: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
severity of mastitis in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of cows and buffaloes
Groups of cows
and buffaloes
Number of
animals
Mean±SEM* of
the group quarter
mastitis severity
scores**
Distribution of quarters as per
their mastitis severity scores
Group
A=Vaccinated
(CMT/ SFMT and
Culture –ve)
cows n=10 0.125±0.0639
3 quarters with score =1
1 quarter with score=2
buffaloes n=30 0.025±0.0186
1 quarters with score=1
1 quarter with score =2
Group B = Control
(CMT/ SFMT and
Culture -ve)
Cows n=15 0.20±0.0744
5 quarters with score =1
2 quarters with score =2
1 quarters with score =1
buffaloes n=15 0.12±0.0481
5 quarters with score =1
1 quarters with score =2
Group C =
Vaccinated CMT/
SFMT and Culture
+ve)
Cows n=15 0.05±0.0402
2 quarters with score =1
1 quarters with score =2
buffaloes n=15 0.033±0.0234
2 quarters with score =1
Group D = Control
(CMT/ SFMT and
Culture +ve)
Cows n=10 0.20±0.0859
6 quarters with score =1
2 quarters with score =2
buffaloes n=10 0.15±0.0706
5 quarters with score =1
1 quarters with score =2
* aggregate of the quarter severity scores divided by the total of quarters in the respective
groups, ** As per Roberson (2003)
SFMT=Surf field mastitis test; CMT=California mastitis test

86
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture -ve for Str. agalactiae and S. aureus CMT /SFMT -ve); B= un-
vaccinated cows and buffaloes (Milk culture -ve for Str. agalactiae and S. aureus, CMT
/SFMT -ve) C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve) D= un-vaccinated- diseased cows and buffaloes (Milk culture +ve for S. aureus
and Str. agalactiae, CMT/ SFMT +ve)
Fig. 4.11: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
severity of mastitis in vaccinated–non-diseased, control-non-diseased,
vaccinated –diseased, control-diseased groups of cows and buffaloes
SFMT=Surf field mastitis test; CMT=California mastitis test
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture -ve for Str. agalactiae and S. aureus CMT /SFMT -ve); B= un-
vaccinated cows and buffaloes (Milk culture -ve for Str. agalactiae and S. aureus, CMT
/SFMT -ve) C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve) D= un-vaccinated- diseased cows and buffaloes (Milk culture +ve for S. aureus
and Str. agalactiae, CMT/ SFMT +ve)
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
cow
s n
=10
bu
ffalo
es n
=3
0
Co
ws n
=15
bu
ffalo
es n
=1
5
Co
ws n
=15
bu
ffalo
es n
=1
5
Co
ws n
=10
bu
ffalo
es n
=1
0
Group A=Vaccinated(CMT/ SFMT and
Culture -ve)
Group B = Control(CMT/ SFMT and
Culture -ve)
Group C = VaccinatedCMT/ SFMT and
Culture +ve)
Group D = Control(CMT/ SFMT and
Culture +ve) c
Mean
±S
EM

87
4.11 Milk somatic cell count
4.11.1 Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased and control-diseased
groups of cows
A non-significant decrease in somatic cell count was noted at day 60, and day 120 in
vaccinated-non-diseased cows (group A). At day 180 post vaccination, somatic cell counts in
this group was non-significantly higher than at day 0, day 60 and day 120. In unvaccinated–
non-diseased cows (group B), a non-significant increase in SCC was recorded at day 60,120
and 180. Vaccination of diseased cows (group C) affected a significant decrease in SCC on
all post vaccination observation time points (day 60, 120 and 180) albeit at day 180, a slight
increase in SCC was noticed. Somatic cell counts in unvaccinated –diseased cows (group D)
differed non-significantly at day 0, 60, 120, and 180 (Table 4.34; Fig.4.12).
A non-significant decrease in somatic cell count was noted at day 60, and day 120 in
vaccinated-non-diseased buffaloes (group A). At day 180 post vaccination, somatic cell
counts in this group was non-significantly higher than at day 0, day 60 and day 120. In
unvaccinated–non-diseased buffaloes (group B), a non-significant increase in SCC was
recorded at day 60, 120 and 180. Vaccination of diseased buffaloes (group C) affected a
highly significant decrease in SCC on all post vaccination observation time points (day 60,
120 and 180) albeit at day 180, a slight increase in SCC was noticed. Somatic cell counts in
unvaccinated –diseased buffaloes (group D) differed non-significantly at day 0, 60, 120, and
180 (Table 4.36; Fig.4.13).

88
Table 4.33: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-diseased, control-
non-diseased, vaccinated –diseased, control-diseased groups of cows
Source of variation Degrees of
freedom
Sum of squares Mean squares F-value
Group
Day
Group x Day
Error
Total
3
3
9
80
95
201.257
7.036
23.537
2.287
234.118
67.0858
2.3454
2.6152
0.0286
2346.31***
82.03***
91.47***
*** = Highly significant (P<0.001)
Table: 4.34: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased, control-diseased
groups of cows
Group of cows Days with reference to first vaccination
Mean 0 60 120 180
A 2.14 ± 0.02c 2.09 ± 0.07
c 2.08 ± 0.03
c 2.31 ± 0.04
c 2.15 ± 0.04
C
B 2.16 ± 0.08c 2.23 ± 0.07
c 2.25 ± 0.08
c 2.38 ± 0.09
c 2.25 ± 0.03
C
C 5.77 ± 0.09a 3.13 ± 0.11
b 3.16 ± 0.09
b 3.26 ± 0.07
b 3.83 ± 0.56
B
D 5.73 ± 0.11a 5.78 ± 0.07
a 5.66 ± 0.07
a 5.71 ± 0.11
a 5.72 ± 0.02
A
Mean 3.95 ± 1.03A 3.31 ± 0.85
B 3.29 ± 0.82
B 3.41 ± 0.79
B
Means sharing similar letter in a row or in a column are statistically non-significant (P>0.05).
Small letters represent comparison among interaction means and capital letters are used for
overall mean.
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= Un vaccinated
cows (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve ); C=
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows (Milk
culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve); D= Un vaccinated cows
(Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve)
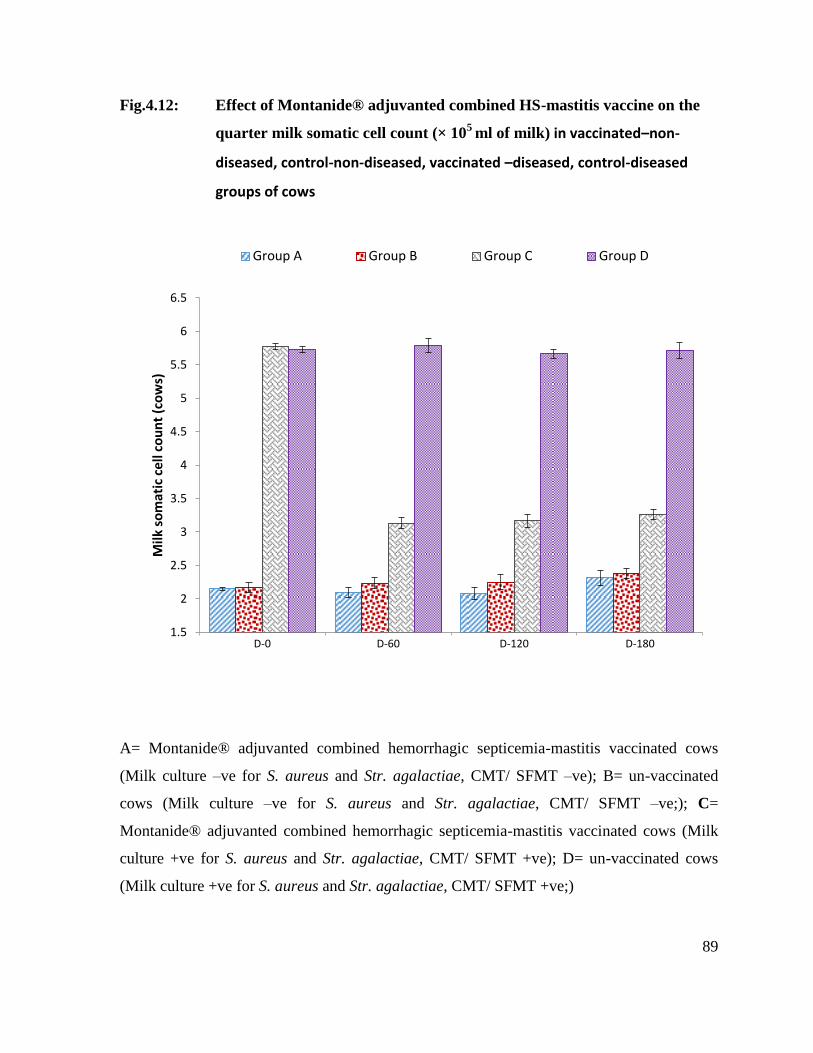
89
Fig.4.12: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased, control-diseased
groups of cows
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= un-vaccinated
cows (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve;); C=
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows (Milk
culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve); D= un-vaccinated cows
(Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve;)
1.5
2
2.5
3
3.5
4
4.5
5
5.5
6
6.5
D-0 D-60 D-120 D-180
Milk
so
mat
ic c
ell
cou
nt
(co
ws)
Group A Group B Group C Group D
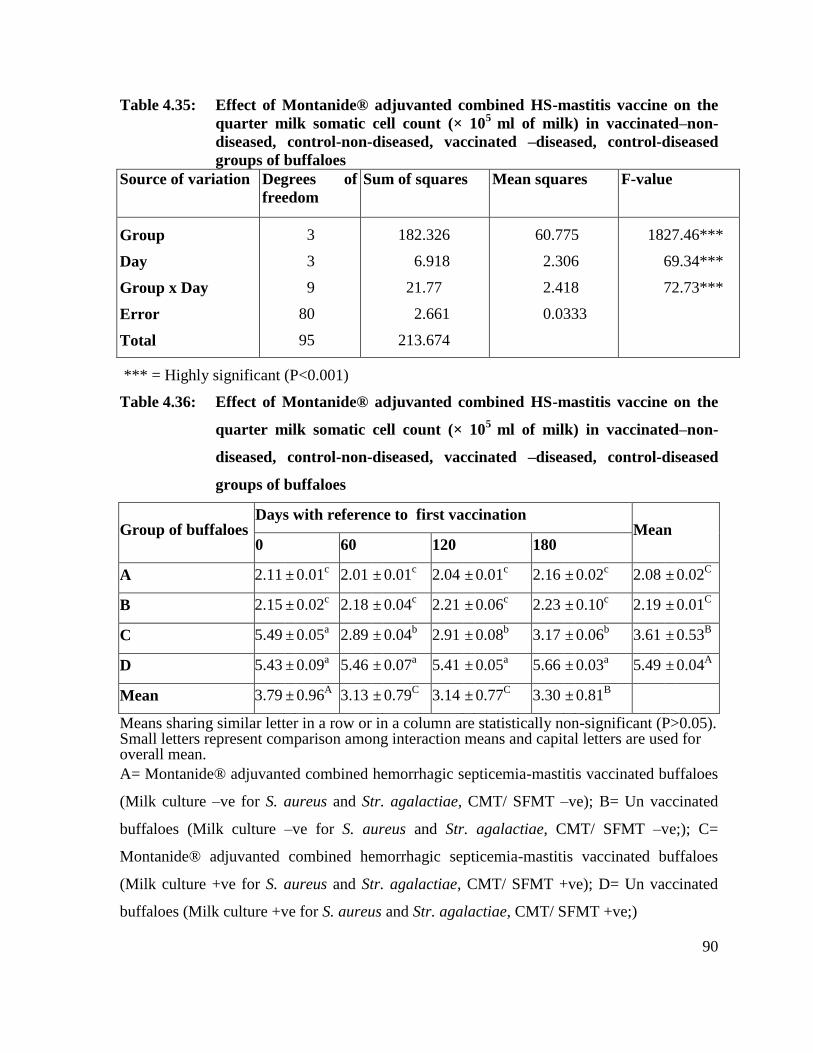
90
Table 4.35: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased, control-diseased
groups of buffaloes
Source of variation Degrees of
freedom
Sum of squares Mean squares F-value
Group
Day
Group x Day
Error
Total
3
3
9
80
95
182.326
6.918
21.77
2.661
213.674
60.775
2.306
2.418
0.0333
1827.46***
69.34***
72.73***
*** = Highly significant (P<0.001)
Table 4.36: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the
quarter milk somatic cell count (× 105
ml of milk) in vaccinated–non-
diseased, control-non-diseased, vaccinated –diseased, control-diseased
groups of buffaloes
Group of buffaloes Days with reference to first vaccination
Mean 0 60 120 180
A 2.11 ± 0.01c 2.01 ± 0.01
c 2.04 ± 0.01
c 2.16 ± 0.02
c 2.08 ± 0.02
C
B 2.15 ± 0.02c 2.18 ± 0.04
c 2.21 ± 0.06
c 2.23 ± 0.10
c 2.19 ± 0.01
C
C 5.49 ± 0.05a 2.89 ± 0.04
b 2.91 ± 0.08
b 3.17 ± 0.06
b 3.61 ± 0.53
B
D 5.43 ± 0.09a 5.46 ± 0.07
a 5.41 ± 0.05
a 5.66 ± 0.03
a 5.49 ± 0.04
A
Mean 3.79 ± 0.96A 3.13 ± 0.79
C 3.14 ± 0.77
C 3.30 ± 0.81
B
Means sharing similar letter in a row or in a column are statistically non-significant (P>0.05). Small letters represent comparison among interaction means and capital letters are used for overall mean.
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= Un vaccinated
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve;); C=
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve); D= Un vaccinated
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve;)
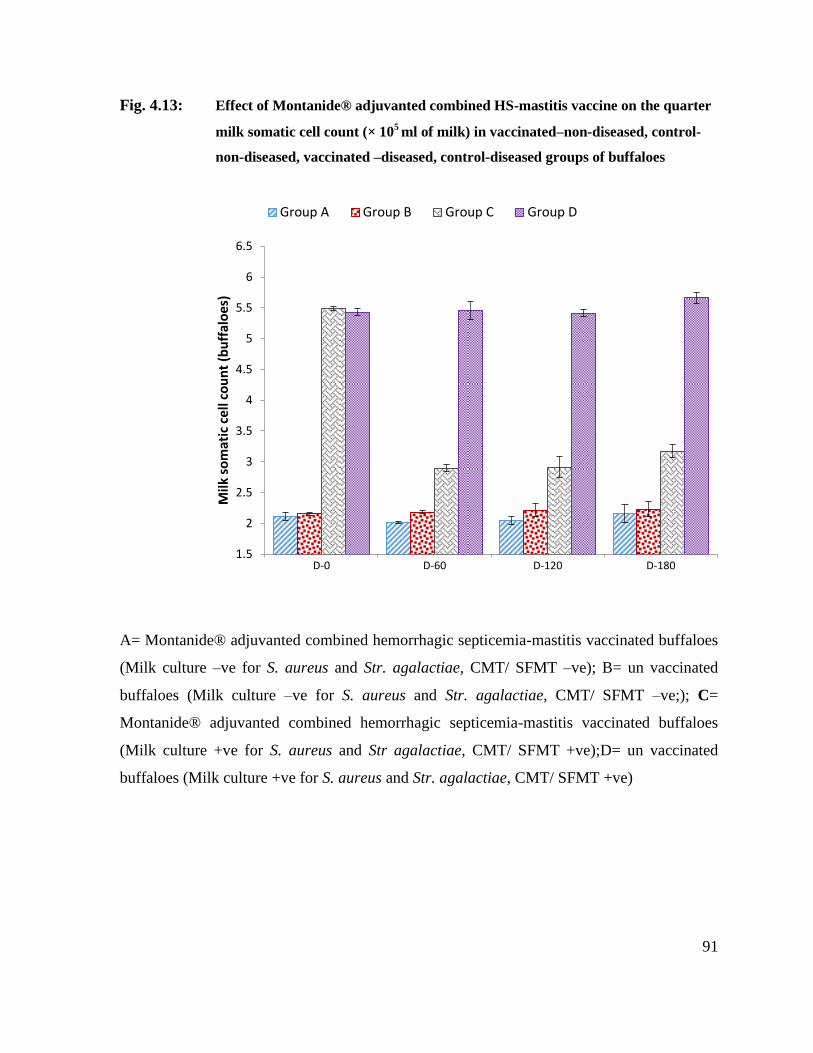
91
Fig. 4.13: Effect of Montanide® adjuvanted combined HS-mastitis vaccine on the quarter
milk somatic cell count (× 105 ml of milk) in vaccinated–non-diseased, control-
non-diseased, vaccinated –diseased, control-diseased groups of buffaloes
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= un vaccinated
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve;); C=
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture +ve for S. aureus and Str agalactiae, CMT/ SFMT +ve);D= un vaccinated
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve)
1.5
2
2.5
3
3.5
4
4.5
5
5.5
6
6.5
D-0 D-60 D-120 D-180
Milk
so
mat
ic c
ell
cou
nt
(bu
ffal
oes
) Group A Group B Group C Group D

92
4.12 Effect of vaccination on milk yield
4.12.1 Milk yield (Mean ±SE; L/24 hours) in vaccinated and control cows and buffaloes
at different post-vaccination days
At Day 0, cows of group A (vaccinated-non-diseased cows) and B (unvaccinated-
non-diseased cows) differed non-significantly in terms of milk yield per day (Table 4.37). On
Day 0, cows of group C (vaccinated-diseased) and group D (unvaccinated diseased) yielded
significantly lesser milk per day than those of group A (vaccinated-non-diseased) and B
(unvaccinated-non-diseased). At Day 60 and Day 120, per day milk yield of cows of group A
(vaccinated-non-diseased) was non-significantly (P ˃ 0.05) higher than their milk yileds at
Day 0. At Day, 60 milk yiled of cows of group A (vaccinated-non-diseased) and B
(unvaccinated-non-diseased) differed non-significantly from each other whereas group C
cows (vaccinated diseased) and group D cows (unvaccinated-diseased) produced
significantly lower milk than those of group A (vaccinated-non-diseased). At this day, group
D cows (unvaccinated diseased cows) yielded significantly lower milk than those of group C
cows (vaccinated-diseased). At Day 120, cows of group A (vaccinated-non-diseased) and B
(unvaccinated-non-diseased) differed non-significantly from each other in their yielded.
Whereas group C cows (vaccinated diseased) and group D cows yielded significantly lower
milk quantity than group A cows (vaccinated-non-diseased) and group B cows
(unvaccinated-non-diseased). Group D cows also yielded significantly lower quantity of milk
than group C cows (vaccinated-diseased). At Day 180, cows of group A had significantly
higher yield than those of group B cows (unvaccinated-non-diseased). However, group C
(vaccinated diseased) was significantly different both from group A (vaccinated-non-
diseased) and B (unvaccinated-non-diseased cows). Group D cows (unvaccinated diseased)
had the lowest yield was significantly lower than those of all of the 3 groups. Over all the
highest yielded was recorded in group A cows (vaccinated-non-diseased) followed by group
B cows (unvaccinated-non-diseased), group C cows (vaccinated-diseased). The lowest
overall yielded was recorded in cows of group D (unvaccinated diseased) that was
significantly lower than those of group A, B and C cows. Entire study period of all groups
differed significantly different from one another in their yieled.

93
Buffaloes of Group A (vaccinated-non-diseased) and group B (unvaccinated-non-
diseased) differed non-significantly from each other in terms of milk yield per day at Day 0.
Similarly, buffaloes of group C (vaccinated diseased) and those of group D (unvaccinated
diseased) yielded non-significantly different quantities of milk per day at day 0 (Table 4.37).
However, group C buffaloes (vaccinated-diseased) and group D buffaloes (unvaccinated-
diseased) had significantly lower yield than those of group A (vaccinated-non-diseased) and
group B (unvaccinated-non-diseased). At Day 60, buffaloes of group A (vaccinated-non-
diseased) and B (unvaccinated-non-diseased) had a non-significant difference among
themselves, group C (vaccinated-diseased-buffaloes) and D (unvaccinated-diseased
buffaloes) was significantly differed from group A and B (unvaccinated-non-diseased
buffaloes). Group D (unvaccinated-diseased buffaloes) also had significantly lower values
than group C (vaccinated-diseased buffaloes). At day 120 buffaloes of group A (vaccinated-
non-diseased buffaloes) had highest milk yield values followed by group B (unvaccinated-
non-diseased buffaloes), C and lowest values observed in group D (unvaccinated diseased
buffaloes). All the groups were significantly different from one another. At day 180 buffaloes
group A (vaccinated-non-diseased buffaloes) had a highest values and was significantly
different from all other group. Group B (unvaccinated-non-diseased buffaloes), C
(vaccinated-diseased buffaloes), and D (unvaccinated-diseased buffaloes) had non-
significantly different yield values from each other but significantly lower from group A
(vaccinated-non-diseased). Over all highest values present in group A (vaccinated-non-
diseased) followed by group B (unvaccinated-non-diseased), C (vaccinated diseased
buffaloes) and lowest values was observed in group D (unvaccinated-diseased buffaloes). All
the groups were significantly different from one another.
Overall milk yield of vaccinated cows and buffaloes (non-diseased or diseased) was
significantly higher than those of unvaccinated cows and buffaloes (non-diseased or
diseased). In other words, vaccination had a positive effect on the milk yield of both healthy
(free from S. aureus and Str. agalactiae) and diseased (infected with S. aureus and Str.
agalactiae) cows and buffaloes.

94
Table 4.37: Milk yield (Mean ±SE; L/24 hours) of vaccinated and control cows and
buffaloes at different post-vaccination days
Animal
groups
Animals and
their numbers
Milk yield (Mean ±SE; L/24 hours) at
different post-vaccination days
Overall
milk yield
(Mean±SE
L/24 hours) Day-0 Day-60 Day-120 Day-180
A
Cows (n= 10) 7.89± 0.15a 8.17± 0.12
a 7.95±0.17
a 5.45±0.16
bc 7.36±0.55
A
Buffaloes (n=
30) 9.13±0.05
ab 9.71±0.06
a 9.52±0.03
a 6.22±0.10
d 8.64± 0.71
A
B
Cows (n= 15) 7.87± 0.19a 7.72± 0.11
a 7.67±0.20
a 4.49±0.22
d 6.93±0.70
B
Buffaloes (n=
15) 9.10±0.18
ab 9.13±0.09
ab 8.57±0.31
b 5.15±0.36
ef 7.99± 0.82
B
C
Cows (n= 15) 5.21± 0.13c 6.13±0.19
b 6.01± 0.14
b 4.76±0.25
cd 5.53±0.28
C
Buffaloes
(n=15) 6.11± 0.25
d 7.10±0.18
c 6.43±0.28
cd 4.72±0.23
f 6.09±0.43
C
D
Cows (n= 10) 5.18± 0.12c 5.10±0.08
cd 4.95± 0.17cd 3.32± 0.11
e 4.64±0.38
D
Buffaloes
(n=10) 6.17± 0.14
d 5.86± 0.13
de 5.27± 0.04
ef 3.79±0.12
f 5.27±0.45
D
Means sharing similar letter in a row or in a column are statistically non-significant (P>0.05).
Small letters represent comparison among interaction means and capital letters are used for
overall mean.
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated cows and
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= un-
vaccinated cows and buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/
SFMT –ve); C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/
SFMT +ve);D= un vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str.
agalactiae, CMT/ SFMT +ve)

95
Fig.4.14: Milk yield (Mean ±SE; L/24 hours) in vaccinated and control cows and
buffaloes at different post-vaccination days
A= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve); B= un vaccinated
buffaloes (Milk culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve;); C=
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccinated buffaloes
(Milk culture +ve for S. aureus and Stragalactiae, CMT/ SFMT +ve);D= un-vaccinated
buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve)
4.13 Cost- benefit analysis of Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine
4.13.1 HS related cost -benefit calculations
Table 4.38 depicts the number of HS cases in vaccinated and control cows, buffaloes
and calves during 180 days post- vaccination period and pecuniary losses based on market
prices of affected animals.
0
2
4
6
8
10
12
cows (10) buffaloes(30)
cows (15) buffaloes(15)
cow (25) buffalo (50) cow (20) buffalo (25)
A B C D
Animal groups
Milk
yie
ld (
per
lact
atio
n)
Day Day Day Day

96
Table 4.38: Number of HS cases in vaccinated and control cows, buffaloes and calves
during 180 days post- vaccination period and pecuniary losses based on
market prices of affected animals
Groups of buffaloes, cows and calves
Cumulative
HS incidence
over 180 days
period
Monitory losses (Rs) based on
the prevailing market prices
(buffalo Rs=125000; cow
Rs=100000; buffalo and cow
calf Rs =25000) of HS affected
animals
A (vaccinated) cows n=10 0 0
buffaloes n=30 1 125,000
B (un-vaccinated)
cows n=15 1 100,000
buffaloes n=15 2 250,000
C (vaccinated)
cows n=15 0 0
buffaloes n=15 0 0
D (un-vaccinated) cows n=10 0 0
buffaloes n=10 1 125,000
E ( vaccinated )
cow calves n=25 0 0
buffalo calves n=50 1 250,00
F( un-vaccinated)
cow calves n=25 1 25,000
buffalo calves n=20 3 75,000
A= Montanide® adjuvanted combined HS-mastitis vaccinated cows and buffaloes (Milk
culture –ve for S. aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS vaccination
during the last 180 days); B= un-vaccinated cows and buffaloes (Milk culture –ve for S.
aureus and Str. agalactiae, CMT/ SFMT –ve; no history of HS vaccination during the last
180 days); C= Montanide® adjuvanted combined hemorrhagic septicemia-mastitis
vaccinated buffaloes (Milk culture +ve for S. aureus and Str. agalactiae, CMT/ SFMT +ve);
D= un vaccinated cows and buffaloes (Milk culture +ve for S. aureus and Str. agalactiae,
CMT/ SFMT +ve); E= vaccinated cow and buffalo calves; no history of HS vaccination
during the last 180 days F= un-vaccinated cow and buffalo calves; no history of HS
vaccination during the last 180 days.

97
Summary
Vaccinated groups
a) Financial losses due to HS associated mortality= Rs. 150,000
b) Cost of two shots of the Montanide® adjuvanted combined HS-mastitis vaccine
( Rs. 60/dose)= Rs. 17,400
Total of a and b = Rs. 167,400
Un-vaccinated groups
a) Financial losses due to HS associated
mortality= Rs. 575,000
Benefits of vaccination (credits) i.e. differential of the debits in un-vaccinated
groups minus debits in vaccinated groups =Rs. 407,600
4.13.2 Mastitis related cost -benefit calculations
4.13.2.1 Mastitis control realted benefits accruing from use of Montanide®
adjuvanted combined hemorrhagic septicemia-mastitis vaccine
Table 4.39 depicts mastitis control realted benefits accruing from the use of
Montanide® adjuvanted combined hemorrhagic septicemia-mastitis vaccine.
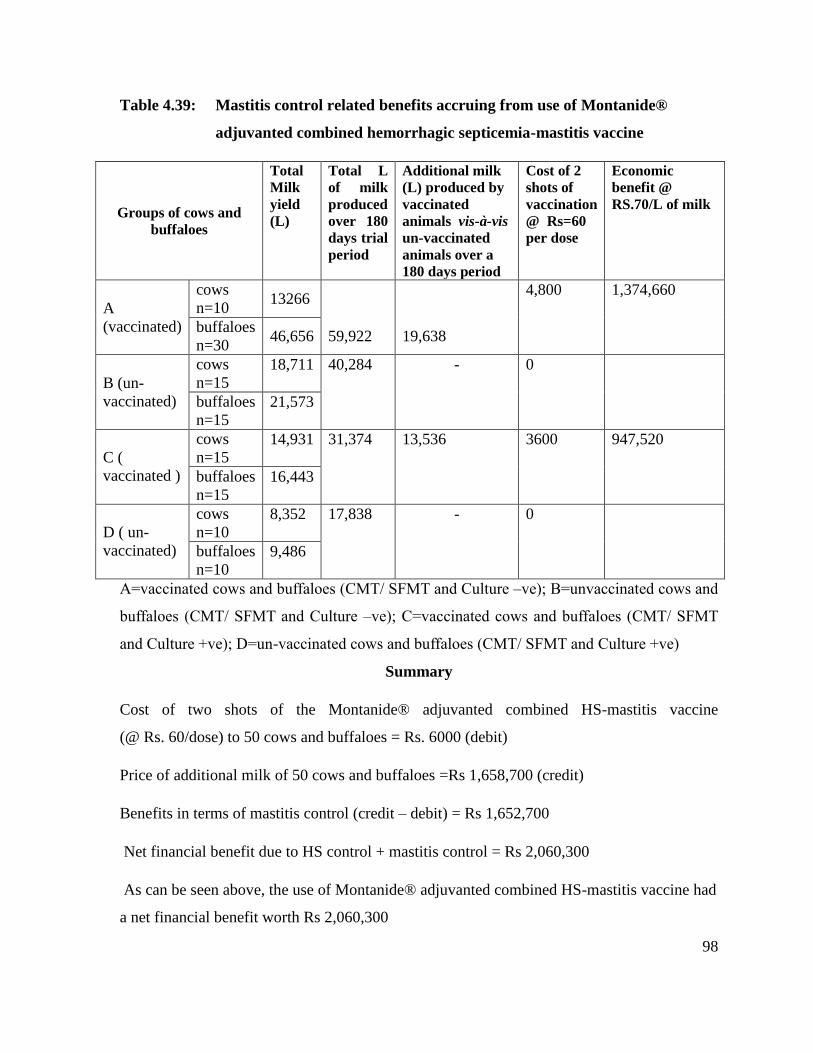
98
Table 4.39: Mastitis control related benefits accruing from use of Montanide®
adjuvanted combined hemorrhagic septicemia-mastitis vaccine
Groups of cows and
buffaloes
Total
Milk
yield
(L)
Total L
of milk
produced
over 180
days trial
period
Additional milk
(L) produced by
vaccinated
animals vis-à-vis
un-vaccinated
animals over a
180 days period
Cost of 2
shots of
vaccination
@ Rs=60
per dose
Economic
benefit @
RS.70/L of milk
A
(vaccinated)
cows
n=10 13266
59,922 19,638
4,800 1,374,660
buffaloes
n=30 46,656
B (un-
vaccinated)
cows
n=15
18,711 40,284 - 0
buffaloes
n=15
21,573
C (
vaccinated )
cows
n=15
14,931 31,374 13,536 3600 947,520
buffaloes
n=15
16,443
D ( un-
vaccinated)
cows
n=10
8,352 17,838 - 0
buffaloes
n=10
9,486
A=vaccinated cows and buffaloes (CMT/ SFMT and Culture –ve); B=unvaccinated cows and
buffaloes (CMT/ SFMT and Culture –ve); C=vaccinated cows and buffaloes (CMT/ SFMT
and Culture +ve); D=un-vaccinated cows and buffaloes (CMT/ SFMT and Culture +ve)
Summary
Cost of two shots of the Montanide® adjuvanted combined HS-mastitis vaccine
(@ Rs. 60/dose) to 50 cows and buffaloes = Rs. 6000 (debit)
Price of additional milk of 50 cows and buffaloes =Rs 1,658,700 (credit)
Benefits in terms of mastitis control (credit – debit) = Rs 1,652,700
Net financial benefit due to HS control + mastitis control = Rs 2,060,300
As can be seen above, the use of Montanide® adjuvanted combined HS-mastitis vaccine had
a net financial benefit worth Rs 2,060,300
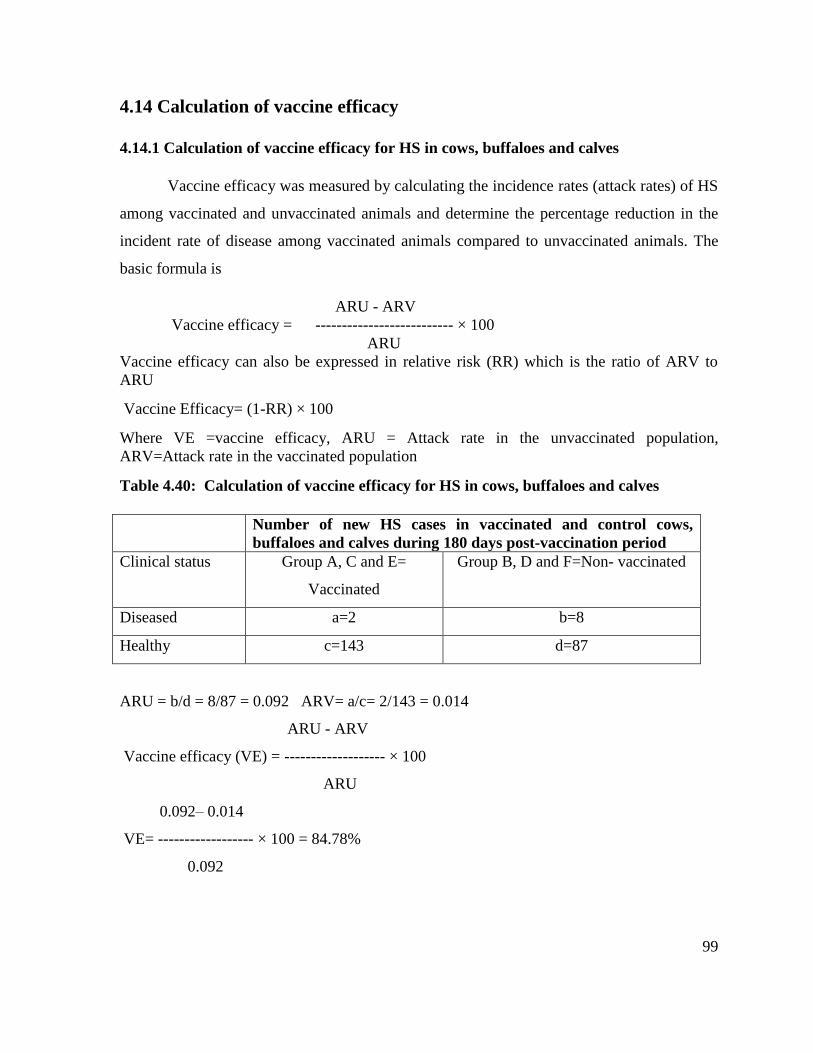
99
4.14 Calculation of vaccine efficacy
4.14.1 Calculation of vaccine efficacy for HS in cows, buffaloes and calves
Vaccine efficacy was measured by calculating the incidence rates (attack rates) of HS
among vaccinated and unvaccinated animals and determine the percentage reduction in the
incident rate of disease among vaccinated animals compared to unvaccinated animals. The
basic formula is
ARU - ARV
Vaccine efficacy = -------------------------- × 100
ARU
Vaccine efficacy can also be expressed in relative risk (RR) which is the ratio of ARV to
ARU
Vaccine Efficacy= (1-RR) × 100
Where VE =vaccine efficacy, ARU = Attack rate in the unvaccinated population,
ARV=Attack rate in the vaccinated population
Table 4.40: Calculation of vaccine efficacy for HS in cows, buffaloes and calves
Number of new HS cases in vaccinated and control cows,
buffaloes and calves during 180 days post-vaccination period
Clinical status Group A, C and E=
Vaccinated
Group B, D and F=Non- vaccinated
Diseased a=2 b=8
Healthy c=143 d=87
ARU = b/d = 8/87 = 0.092 ARV= a/c= 2/143 = 0.014
ARU - ARV
Vaccine efficacy (VE) = ------------------- × 100
ARU
0.092– 0.014
VE= ------------------ × 100 = 84.78%
0.092

100
4.14.2 Calculation of vaccine efficacy for mastitis in cows and buffaloes
Vaccine efficacy was measured by calculating the incidence rates (attack rates) of
mastitis among vaccinated and unvaccinated animals and by determining the percent
reduction in the incidence rate of disease among vaccinated animals compared to
unvaccinated animals
Table 4.41: Calculation of vaccine efficacy for mastitis in cows and buffaloes
Quarter based cumulative incidence over 180 days post
vaccination period
Clinical status Group A and C= Vaccinated Group B and D=Non- vaccinated
Diseased a=5 b=22
Healthy c=65 d=28
ARU = b/d = 22/28 = 0.79 ARV= a/c= 5/65 = 0.077
ARU – ARV
Vaccine efficacy (VE) = ------------------- × 100
ARU
0.79 – 0.077
VE= ------------------ × 100 = 90.25%
0.79

101
CHAPTER 5
DISCUSSION
Hemorrhagic septicemia (HS) and mastitis are two of the 10 most prevalent and
important diseases of dairy animals in Pakistan (Khan et al., 1991; Khan et al., 1994-1995).
Although, vaccination against HS is practiced in routine, the application of mastitis control
measures is almost totally nonexistent, save a small number of large commercial dairy herds.
A combined HS-mastitis vaccine, therefore holds the promise of simultaneous control of
these two rife dairy animal diseases in Pakistan. The present investigations were carried out
in pursuit of this objective by following the internationally acceptable guidelines/
requirements of a vaccine preparation and testing (EMEA, 2000).
5.1 Preparation of combined HS-mastitis vaccine and its evaluation against HS
An attempt was made to use P. multocida isolates, isolated from the field cases of
clinical HS. In the context of the present study, a calf was defined as any bovine or bubaline
animal aged up to one year (http://en.wikipedia.org/wiki/calf). This is the age group of cattle
and buffaloes most susceptible to HS (Farooq et al., 2011; Khan et al., 2011). The septicemia
in HS occurs at the terminal stage of the disease. Therefore, blood samples taken from sick
animals before death may not always contain P. multocida organisms. This microorganism is
also not consistently present in the nasal secretions of sick animals. A blood sample or swab
collected from the heart is satisfactory if it is taken within a few hours of death. If the animal
has been dead for a long time, a long bone, free of tissue, can be taken. If there is no facility
for postmortem examination, blood can be collected from the jugular vein by incision or
aspiration. Blood samples in any standard transport medium should be dispatched on ice and
well packed to avoid any leakage. Blood smears from affected animals are stained with
Gram, Leishman‟s or methylene blue stains. The organisms appear as Gram-negative,
bipolar-staining short cocco-bacilli. No conclusive diagnosis can be made on the basis of
direct microscopic examinations alone. Blood samples, or swabs eluted into 2–3 ml sterile
physiological saline are cultured. Alternatively, the surface of a long bone is swabbed with
alcohol and split open. The marrow is extracted aseptically and cultured. Direct culture is

102
usually satisfactory only if the material is fresh and free from contaminants or post-mortem
invaders that would otherwise overgrow any Pasteurella present.
The failure to recover P. multocida in overwhelming majority of clinical cases in the
present study is broadly congruent with the findings of previous studies (Butt et al., 2003;
Ashraf et al., 2011; Ekundayo et al., 2008). Many blood samples collected from HS
affected animals failed to yield the etiologic agent on cultural examination because
septicemia develops only in the terminal stages of this disease (Center for Food Security and
Public Health and Institute for International Cooperation in Animal Biologics an OIE
Collaborating Center, 2009). This notion was borne out by the findings of the present study
wherein blood samples of all 5 buffalo calves and 2 adult buffaloes tested negative on
cultural examination. Long bones from the suspected carcasses of HS were found to be
sample of choice for isolation of P. multocida. Isolation of P. multocida has been attempted
from a wide range of organs such as kidney, heart, liver, brain, tonsils, and lymph nodes from
experimentally infected calves (Chowdhury et al., 2014; OIE, 2012; Singh et al., 2007).
From the liver and blood samples, recovery of the causative organisms is poor. Gross
pathological changes might have inactivated the causative agents in these samples (Naz et
al., 2012). In the present study, microbiological examination of 7 blood samples (collected
from five HS affected calves and two HS affected adult buffaloes) on CYS agar did not yield
growth of P. multocida. The negative blood culture results in these cases may be attributed to
the notion that P. multocida appears in blood only at the terminal septicemic stages of the
disease (Carter and De Alwis, 1989). A total of 3 isolates of P. multocida could be recovered
on microbiological examination of peripheral blood, femur, and heart blood of HS affected
animals on CSY and blood agar plates. No P. multocida isolate was recovered from
peripheral blood and swab samples of heart blood. Culturing of heart blood and femur bone
marrow samples respectively yielded growth of two and one isolate of P. multocida.
Suspected isolates of P. multocida formed smooth grayish, glistening translucent colonies of
nearly 1 mm diameter on blood agar after 24 hours incubation at 37°C. No growth was
recorded on MacConkey‟s agar on subculturing. On Gram staining, the organism appeared as
Gram –ve, short, ovoid bipolar staining coccobacilli with some degree of pleomorphism.
Growth on CYS agar was smooth, yellowish, mucoid and larger in size than that on blood

103
agar. All three P. multocida isolates were positive for indol, ODC, glucose, fructose, and
catalase, while negative for H2S production, lactose, D-dulcitol and maltose. Variable results
were recorded for D-sorbitol and arabinose. These isolates of P. multocida were confirmed
by amplification of P. multocida specific gene using KTSP61/KTT7 set of primers, which
yielded a product of 618 bp on electrophoresis that confirmed these isolates as P. multocida
type B. Conditions for HS-causing type-B-specific PCR were similar to those described for
P. multocida-specific PCR (Townsend et al., 1998). The usefulness of these primers has been
reported for identification of serogroup B strains. HS-causing type-B-specific PCR primers
can also be used in a Multiplex PCR with the P. multocida-specific primers, dramatically
decreasing the time required for P. multocida detection and presumptive identification of the
HS-serotype. To date, the HS-causing type-B-specific PCR remains 100% specific for HS-
causing type B serotypes of isolated P. multocida (OIE, 2012). Type B cultures with the
predominant somatic antigen being either type 2 or 5 are identified by the amplification of a
~620 bp fragment with the KTSP61/KTT7 set of primers.
All 3 isolates of P. multocida were sensitive to enrofloxacin, amoxicillin, ceftiofur
sodium and oxacillin and resistant to tetracycline, fusidic acid, and penicillin. All three
isolates displayed an intermediate sensitivity to vancomycine. A variable degree of
susceptibility was displayed to lincomycine, Septran, and gentamicin. Kehrenberg et al.
(2001) has reviewed antibiotic susceptibility of P. multocida that causes HS.
LD 50 of the three candidate vaccinal isolates of P. multocida in the present study are
broadly in agreement with those reported by Butt et al. (2003). These workers reported the
mouse LD50 of 19 field isolates of P.multocida. The LD50 of these isolates ranged from 10-7.75
to 10-7.50
. According to OIE (2012), the type B: 2 strains associated with HS are highly
virulent for mice and rabbits with LD50 ranging from 1and10 viable microrganisms.
A high degree of immunogenicity is one of the cardinal considerations in selecting an
isolate for vaccine production. As per Muneer et al (2005) comparativetly high antibody
titers were observed in animals of all age groups vaccinated with oil adjuvant vaccine
throughout the trial period while the titers in animals vaccinated with alum precipited
vacccine declined after 3 months following vaccination and reached at minimal level 180

104
days post vaccination. According to the workers, results justified the replacement of alum
precipited vaccine with the newly developed oil adjuvant vaccine for the control of HS in
animals. In the present study, Montanide® ISA 201 VG was used as an adjuvant. Vaccines
formulated with Montanides® (Seppic, France) are cheaper and these adjuvants are ready-to-
use mineral oil adjuvants. The use of Montanides® in veterinary vaccines as adjuvants has
shown very promising results. The use of a Montanide® adjuvanted mastitis vaccine
reportedly induced significantly high immune response against α and β toxins of S. aureus
and pseudocapsule in sheep and cattle (Nordhaug et al., 1994a; Tollersurd et al., 2001).
Montanide® adjuvanted vaccine is reported to be less viscous and less irritant than traditional
Freund‟s adjuvanted vaccine. Montanide® ISA 201 VG adjuvanted quadrivalent foot-and-
mouth vaccine has been reported to elicit superior immune response compared to that elicited
by aluminium hydroxide gel. The vaccine resulted in rapid immune response that was
maintained for longer duration (Patil et al., 2002). Owing to these desirable attributes, the
combined HS-mastitis vaccine tested in the present study utilized Montanide as an adjuvant.
The number of new HS cases in vaccinated and control cows, buffaloes and calves
during 180 days post-vaccination period was investigated. None of the 10 vaccinated cows
and 25 cow calves developed HS over the entire observation period of 180 days. Only 1 of
the 30 vaccinated buffaloes (3.33%) developed HS during this period. In the control group
(un-vaccinated), 1 cow (6.66 %) and 2 buffaloes (13.33 %) developed HS during this period.
One of the 50 buffalo calves (2%) vaccinated with this vaccine developed HS during this
period. One of 25 un-vaccinated cow calves (4%) and 3 of 20 buffalo calves (15%)
developed clinical HS over the observation period of 6 months. Pasteurella multocida could
be isolated from only 1 buffalo calf (group F) that succumbed to peracute HS. Isolation of P.
multocida from other cases were not successful. The Montanide® adjuvanted combined
hemorrhagic septicemia-mastitis vaccine thus seems protective for HS over 6 months trial
period. This vaccine had a vaccine efficacy of 84.78% against HS. Nevertheless, in view of a
limited number of test animals utilized in the present study, a much larger clinical trial
involving a much larger number of bovines and bubaline subjects is clearly warranted. In as
much as the use of a combined HS-mastitis vaccine has not been reported heretofore, it is not
possible to compare the results of the present study with those of any previous study.

105
5.2 Evaluation of combined HS-mastitis vaccine against mastitis
With milk production of around 52.6 million tons annually, Pakistan is the fourth largest
milk producing country in the world (Pakistan Economic Survey, 2014-15). Mastitis is a
major production limiting disease of dairy animals all over the world. It not only reduces
milk quantity but also depreciates the milk quality, thus adversely affecting milk production
economics (DeGraves and Fetrow, 1993; Sordillo et al., 1997). On a pattern equivalent to
most other countries, mastitis is the most important health problem of dairy animals in
Pakistan (Cady et al, 1983; Ajmal, 1990; Egenolf, 1990). In Pakistan, about 37 years ago,
losses due to clinical mastitis were estimated at Rs.240 million annually in Punjab province
alone (Chaudhry and Khan, 1978). Staggering economic losses associated with mastitis
warrant a well-planned mastitis control program to curtail losses for profitable dairying.
Dairy farmers in developed countries like USA, Canada and Australia are practicing mastitis
control measures, viz. dry cow therapy, post-milking teat dipping, and culling chronically
mastitic cows since long. Due to small herd size, non-progressive attitude of dairy farmers
and lack of any system of milk quality premium, standard mastitis control measures are
difficult to be adopted in Pakistan. Antibiotic treatment has its own demerits including
emergence of antibiotic resistance and drug residues in milk, thus rendering mastitis control
through antibiotics as an unacceptable approach due to public health hazards. In the absence
of any mastitis control program, high antibiotic resistance and poor therapeutic response,
vaccination against some of the most prevalent mastitis pathogens seems a logical choice to
curtail losses associated with mastitis. In recent studies (Athar et al., 2007; Rashid, 2011),
efforts were made to investigate the role of vaccination as an alternative mastitis control
strategy for farms lacking standard mastitis control program. Although, both live attenuated
and dextran sulphate adjuvanted vaccines evaluated in these studies were found valuable in
the control of mastitis in buffaloes, the limitation of these vaccines including need of a cold
chain for live attenuated vaccine, high production cost of dextran sulphate vaccine (Rs. 500
for 2 shots of dextran sulphate adjuvanted vaccine) and short duration immunological
protection were obvious. There is thus a need not only to curtail the cost of mastitis vaccine
but also to prolong its duration of effectiveness. The principal objective of present study
included the evaluation of Montanide® adjuvanted S. aureus plus Str. agalactiae bacterin-

106
toxoid in an effort to obtain desirable and durable immune response at an economical cost.
From the data generated in the present study, the Montanide® adjuvanted combined HS-
mastitis incorporating S. aureus and Str. agalactiae (the two most prevalent mastitis
pathogens in Pakistan as reviewed by Shakoor, 2006) displayed both preventive and curative
qualities. The preventive and curative attributes of this vaccine (Tables 4.25 to 4.31) are in
agreement with the findings of previous studies (Chishti et al., 1992; Nickerson et al., 1993;
Nordhaug et al., 1994a; Calzolari et al., 1997; Giraudo et al., 1997; Qureshi et al., 2000;
Shakoor, 2006; Shakoor et al., 2006; Athar, 2007; Ashfaq et al., 2014). In addition,
mean±SEM of the group quarter mastitis severity scores (Table 3.32) were lower in
vaccinated groups (group A cows =0.125±0.0639, group A buffaloes =0.025±0.0186; group
C cows =0.05±0.0402, group C buffaloes =0.033±0.0234) than in un-vaccinated groups
(group B cows=0.20±0.0186, group B buffaloes 0.12±0.0481; group D cows=0.20±0.0859,
group D buffaloes= 0.15±0.0706). In this respect, the results of the present study are in
conformity with the results reported by Qureshi et al. (2000), Shakoor (2006), Shakoor et al.
(2006), and Athar (2007). Contrarily, however the results of the present study are discordant
with the results reported by Hoedemaker et al. (2001). These workers determined the effect
of an aluminium hydroxide adjuvanted autogenous S. aureus bacterin on the prevalence of S.
aureus, SCC, and clinical mastitis in a dairy herd beset with infections with this organism.
Animal and quarter based prevalence of S. aureus IMIs did not differ between cows treated
with autogenous vaccine or those treated with a placebo. Although, the combined HS-
mastitis vaccine tested in the present study displayed both preventive and curative effect on
mastitis, its use can not be recommended as a substitute for standard mastitis control
measures such as antiseptic teat dipping, dry cow therapy and culling of chronically infected
animals (Bozhkova et al., 1985). Nonetheless, it can be recommended where a „do nothing‟
policy of mastitis control is in vogue. The increase in milk yield of animals treated with
combined HS-mastitis vaccine in the present study is consistent with the findings of Watson
(1987), Leitner et al. (2003) and Athar (2007). In the present study, concentrations of P.
multocida, S. aureus and Str. agalactiae were adjusted to 1×1010
cells/ml in the montanide
adjuvanted HS-mastitis vaccine. Athar (2007) while evaluating the inactivated polyvalent
vaccines for the control of mastitis in dairy buffaloes reported that a concentration of 1010

107
cells/ml of each of three vaccinal isolates (S. aureus, Str. agalactiae, and E.coli) was more
antigenic as compared to 108
and 1012
cells/ml.
The vaccine developed and tested in the present study harbored antigens not only of P.
multocida (the etiologic agent of a very prevalent disease of calves i.e HS) but also those of
S. aureus and Str. agalactiae (the common etiologic agents of heifer mastitis). Its use in
female calves might have exerted a beneficial role in the control of heifer mastitis. The need
to control mastitis in heifers has since long been recognized (Trinidad et al., 1990a; Trinidad
et al., 1990b; Nickerson et al., 1995; Oliver et al., 2003; Oliver et al., 2003, De Vliegher,
2004) and is increasingly coming into focus in dairy farming.
Mondanide® adjuvanted combined HS-mastitis vaccine evaluated in the present
study was not 100% protective for hemorrhagic septicemia (Table 4.24). This vaccination
failure is consistent with findings of some previous studies. Field observations of veterinary
officers in 9 districts of Pakistani Punjab have indicated the occurrence of field outbreaks of
HS despite vaccination with alum- precipitated P. multocida bacterin prepared in public
sector (Sheik et al., 1996). In order to obtain maximum protection from HS vaccine, it is
recommended that the vaccines be prepared from P. multocida strains circulating in the
regions of intended use. FAO recommends a live avirulent HS vaccine prepared from a P.
multocida serotype B: 3 of fallow deer origin for use in Southeast Asia (Carter, 2005).
Despite vaccination and improved management, occurrence of HS outbreaks is a regular
feature each year, especially in endemic areas of the country (Afzal and Muneer, 1990).
Pakistani workers (Shahzad et al., 2013) reported an outbreak of HS in buffalo calves that
had been vaccinated with alum precipitated HS vaccine two months earlier. Arif et al. (2013)
recently documented that subunit or whole bacterin based P. multocida vaccines were less
immunogenic than an anti-idiotypic vaccine prepared against P. multocida B: 2. Implication
of P. multocida as a mastitis pathogen has also been reported (OIE ,2012)
Isolation, cultural, biochemical characteristics of vaccinal isolates of S. aureus and Str.
agalactiae
Isolation, cultural and biochemical characterization was conducted following the
procedures as describe by brown et al. (1981), National Mastitis Council USA (1990), Athar

108
(2007) and Cengiz et al (2015). The procedures for isolation of S. aureus and Str. agalactiae
are inline with those of Gonzalez et al. (1989). Nikerson (1992), Chaudhry and Azam (1995)
and Fazal-ur-Rehman (1995). Staphylococcus aureus isolates (n=4) that grew on blood agar
were β- hemolytic, catalase positive, Gram positive cocci, 7-digit numerical profile with
api® Staph (bioMerieux, France) test strip = 6736153 and, Latex agglutination test
(Staphytect plus® kit) = positive and were thus presumptively identified as members of the
genus Staphylococcus. On performing tube coagulase test, all of these 4 isolates tested
positive for coagulase production at 4 hours. Staphytect plus® kit (Oxoid, UK) test was used
in the present study for additional confirmation of S. aureus based on the biochemical
differences between S. aureus and other coagulase positive staphylococci (Cohen, 1987).
Several components of S. aureus possess not only antigenic properties (ensuing antibodies
production) but also act as mitogens (i.e. they stimulate cell division) towards B-cells and T-
cells. Protein A is a good B-cell mitogen and techoic acid (a cell wall component) is both a
B-cell and T-cell mitogen. Clumping factor C is one of the surface proteins (adhesions)
which endows adherence properties to S. aureus thus enabling this organism to adhere to
mammary gland tissue (Sears, 2002). In the present study, presence of these cardinal
virulence factors of S. aureus was detected by the Staphytect plus® kit (Oxoid, UK).
Streptococcus agalactiae isolates (n=2) displayed β-hemolysis. Microscopic
examination of Gram stained smears of these isolates revealed Gram positive cocci and
showed api® 20 Strep (3462414). On catalase test, these isolates were negative. CAMP test
reaction on blood agar with 0.1% esculine and 0.01% ferric citrate was positive for both the
isolates.
All 4 isolates of S. aureus and 2 isolates of Str. agalactiae were sensitive to
lincomycin, enrofloxacin, amoxicillin, ceftiofur sodium and oxacillin, while resistant to
tetracycline. Turkish workers (Cengiz et al., 2015) determined the antibiotic resistance of 61
Staphylococcal isolates recovered from subclinically mastitic cows to oxacillin,
erythromycin, tetracycline, nitrocefin, and cefoxitin. Of these isolates (S. aureus, n=34 and
coagulase negative staphylococci, n=27), 26, 29, and 8 displayed resistance to erythromycin,
tetracycline, and oxacillin, respectively.This indicates indiscriminate use of certain

109
antibiotics would have impired resistance in S. ureus and Str. agalactiae and would easily
lead to therapeutic failure therefore antibiotic susceptibility profile is highly warranted.
Biofilm production by candidate vaccinal S. aureus isolates (n=4)
The term biofilm refers to a structured community of bacterial cells enclosed in a self-
produced polymeric matrix and adherent to an inert or living surface (Costerton et al., 1999).
The ability of an organism to produce biofilm (also known as slime production) is
significantly associated with its capability to produce diverse illnesses (Tenover et al., 1998).
Biofilm producing bacteria are known to cause numerous human diseases like cystic fibrosis,
native valve encarditis, otitis media, periodontitis and chronic prostatitis (Rodney, 2002).
Several studies have provided the evidence that S. aureus can form biofilms in the
udder of cows suffering from mastitis. Biofilm production endows upon the bacteria an
ability to impair the action of both the host immune system and antimicrobial agents and
allow the persistence of the infection (Melchior et al., 2006a; Melchior et al., 2009).
Recurrent infections are often attributable to biofilm production by bacteria. Biofilm
formation is accompanied by significant genetic and sub-sequent physiological changes in
the microorganisms resulting, inter alia, in a loss of sensitivity to virtually all classes of
antibiotics (Melchior et al., 2006a). Hensen (2000) reported that in bovine primary mammary
epithelial cell assay, sensitivity for neomycin, neomycin and penicillin, neomycin and
tetracycline, neomycin and bacitracin and tetracycline with bacitracin was higher for non-
adherent S. aureus than for adherent staphylococci. An exopolysaccharide matrix is
constituent of biofilm. Owing to this trait, biofilm resist the access of antibiotics to bacteria
(Stewart, 1996). The exopopolymers forming the biofilm shield the bacteria against
components of the host immune system. Extra-cellular slime (slime is the old term for
biofilm) produced by S. epidermidis protected cells from the phagocytic activity of
macrophages and against opsonins as well as reactive oxygen species in polymorphonuclear
leukocytes. Slime production by S. aureus strains was negatively associated with bactericidal
activity of neutrophils (Melchior et al., 2006a).
In the present study, three methods (tube method, spectrophotometric method and
Congo red agar method) were used for determination of biofilm production by 4

110
staphylococcal isolates. The tube method-based scores of biofilm production by 4 candidate
vaccinal S. aureus isolates. S. aureus isolate number 2, 3 and, 4 showed a moderate level of
biofilm production, whereas isolate number 1 showed strong biofilm production. All 4
isolates produced biofilms, however highest OD (0.31nm) was noted in isolate No. 1, whilst
OD values ranged between 0.17- 0.22. Thus the isolate No.1 was considered as highest
producer of biofilm in spectrophotometric method. On Congo red agar (CRA) S. aureus
isolate number 2, 3 and 4 showed intermediate degree of biofilm production whereas isolate
number 1 showed strong degree of biofilm production.
Turkish workers (Turkyilmaz and Eskiizmirliler, 2006) investigated the production of
slime factor in 180 staphylococcal isolates (90 coagulase positive staphylococcus; CoPS and
90 coagulase negative staphylococcus; CoNS) isolated from various animal specimens
(bovine mastitis, dogs with otitis externa and chickens with various infections) with Congo
red agar (CRA), microplate (MP) and standard tube (ST) tests and the results were compared
to each other. The rate of production of slime factor in all of the staphylococcal species
investigated with CRA, MP and ST tests was 61.1, 55.5 and 50.5%, respectively. Of a total
of 60 CoPS and CoNS isolates associated with mastitis, 40 (66.66%) were positive for slime
factor in CRA method. Frequencies of slime factor positive and slime factor negative isolates
of intramammary origin detected by two other methods (MP and ST) were not stated by these
investigators. According to Keskin et al. (2003), CoNS strains isolated from bovine mastitis
milk samples and chicken lesions were more slime productive and adherent as compared to
the strains isolated from healthy animals. Different methods were used for determination of
slime production. Citak et al. (2003) used CRA method to investigate slime production by
CoNS (32.7%) isolated from raw milk samples. They reported that slime production was an
important virulence factor for identifying pathogenic staphylococci. Mathur et al. (2006)
determined the frequencies and scores of biofilm production by 152 non-repetitive, human
clinical isolates of staphylococcus species isolated from blood, infected devices and skin
surface samples received from different tertiary care centers in India. Of the 152
staphylococcus species, 88 (57.8%) displayed a biofilm positive phenotype under the
optimized conditions in the tissue culture plate method and strains were further classified as
high 22 (14.47%) and moderate 60 (39.4%), while in 70 (46.0%) isolates weak or no biofilm

111
was detected. Though the tube method correlated well with the tissue culture plate test for 18
(11.8%) strongly biofilm producing strains, weak producers were difficult to discriminate
from biofilm negative isolates. Screening on Congo red agar did not correlate well with
either of the two methods for detecting biofilm formation in staphylococci. The presence of
the ICA (inter cellular adhesion, one of the components of the genetic make-up of a
particular strain related to biofilm formation in vitro) locus among S. aureus mastitis isolates
was recently investigated by Vasudevan et al. (2003). All 35 isolates tested possessed the
ICA locus, but not all isolates were positive with Congo red agar or the biofilm assay test.
This indicates that there is a high prevalence of the ICA genes among S. aureus mastitis
isolates. These investigators raised the question whether or not the observation that not all
strains produce biofilms ex vivo is related to specific culture conditions (biofilm growth is
stimulated by hypoxic conditions, but culturing in most cases occurs under oxic conditions).
Additional work in this area is clearly warranted. As indicated by the findings of present
study as well as previous studies (Turkyilmaz and Eskiizmirliler, 2006; Mathur et al., 2006;
Keskin et al., 2003, Melchior et al., 2009), the frequencies of biofilm production by different
isolates in a set of isolates is often a function of the methods used to determine biofilm
production trait.
Evaluation of sterility and safety of the Montanide® adjuvanted combined
HS-mastitis Vaccine
Evaluation of sterility and safety along with efficacy, simplicity of use, compatibility
with immune prophylaxis and affordability are the main components of quality of assurance
of the test vaccine (Soulebot et al., 1997). An attempts was made to follow all the standard
procedures as recommended by ISO (1994), FAO (1998) and OIE (2012), for confirming
sterility and safety of the experimental vaccines in laboratory animals. Albino mice, rabbits,
cattle and buffaloes were used for these preliminary studies and it was found that this vaccine
was safe to inject and free of demonstrable side-effects when given at the standard dose rates
(0.2ml IM for mice and 5 ml for cattle and buffaloes). However, this vaccine produced
sluggish movement in one mice, whereas rabbit showed mild local swelling. Similarly, cattle
and buffaloes exhibited moderate swelling when 5 ml of the vaccine was injected SC. As for
as vaccination–challenge studies were conducted in mice, vaccinated mice showed

112
significantly higher survival percentage as compared to those of control groups. The findings
of the present study are in agreement with those of Giraudo et al. (1997) and Athar (2007).
Effect of HS-mastitis combined vaccine on somatic cell count
The somatic cell count is an indicator of inflammation and milk quality (Sharif et al.,
2007). These cells are secreted during the normal course of lactation and are influenced by a
variety of factors like season, breed, environment and infection. The somatic cell count in the
normal buffalo milk varies from 0.5 × 105 to 3.75 × 10
5 with a mean of 1.4 × 10
5 cells per ml
(Silva and Silva, 1994). In cows, SCC in normal milk is always less than 1 × 105 cells per ml.
The possible reason may be compositional changes of the milk, environmental stress and
quarter udder capacity of the buffalo. In the present study values described by Silva and Silva
(1994) of somatic cell count in normal buffalo milk were used to differentiate normal and
mastitic milk. A non-significant decrease in somatic cell count was noted at days 60, day 120
in vaccinated-non-diseased cows and buffaloes. At day 180 post vaccination, somatic cell
count in this group was non-significantly higher than at days 0, 60 and 120. In unvaccinated–
non-diseased cows and buffaloes, a non-significant increase in SCC was recorded at days 60,
120 and 180. Vaccination of diseased cows and buffaloes caused a significant decrease in
SCC on all post vaccination observation time points (days 60, 120 and 180) albeit at day 180,
a slight increase in SCC was noticed. Somatic cell counts in unvaccinated –diseased cows
and buffaloes differed non-significantly at days 0, 60, 120, and 180. These somatic cell count
at 28 days post-treatment are consistent with that of Owens et al. (1988) and Owens et al.
(1995).
Cost-benefit analysis of Montanide® adjuvanted combined HS-mastitis
vaccine
The use of Montanide® adjuvanted combined HS-mastitis vaccine had a net financial benefit
worth Rs 2,060,300. Profitability can be more visible if more lactation length of the animal is
taken into account. Fabre et al. (2004) proved in a study that treatment could be profitable
during the first 6 months of lactation with expected cure rate of 70%. Its value would more
limited with a lower cure rate. In the calculation of cost-benefit ratio only the monitory value
of additional milk produced is consider. These calculations in this regard overlook the benefit

113
of vaccination in terms of milk quality (e.g. decrease somatic cell count). The results of
the present study are in line with the findings of Amorena et al. (1994) and
Shakoor (2006) that the vaccinated animals had a lower somatic cell count and
higher milk production in relation to the control group, suggesting that the
economic benefit of the vaccine may be 13 times higher than the cost.
Calculation of vaccine efficacy
Vaccine efficacy was measured by calculating the incidence rates (attack rates) of
disease among vaccinated and unvaccinated animals and determine the percentage reduction
in the incident rate of disease among vaccinated animals compared to un- vaccinated animals
(Weinberg and Szilagyi, 2010). Vaccine efficacy by using of Montanide® adjuvanted
combined HS-mastitis vaccine for HS and mastitis is 84.78%.and 90.25% respectively.
Similar result were in a separate study conducted in 3 commercial dairy herds in New
Zealand (Pankey et al., 1983)
In conclusion, the combined HS-mastitis vaccine developed and tested in the present study
indicated a preventative efficacy against HS and preventative as well as a curative efficacy
against mastitis. However, in view of preliminary nature of the present study as well as in
view of a fairly small number of animals utilized, a much broader study involving a much
larger number of bovine and bubaline animals is clearly warranted before a definitive
recommendation can be made for its field use.
Future outlook and recommendations
1. In view of a preliminary nature of the study, external validity (trials to be conducted
at other institutions as well as in the field) of the present study needs to be
established.
2. The role of combined HS-mastitis vaccine in conjunction with other mastitis control
measures (e.g. teat dipping, dry cow therapy, segregation etc.) needs to be
determined.
3. The role of combined HS-mastitis vaccine in the control of HS and heifer mastitis
need to be determined.
4. Larger field trials are needed in particular under field conditions.

114
5. Addition of other bacterial or viral pathogens (e.g Corynebacterium species, E.coli,
leptospira, FMD virus) in the „combo‟ vaccine tested in the present study may be
contemplated.

115
CHAPTER 6
SUMMARY
As per field surveys of major dairy animal diseases in Pakistan, hemorrhagic septicemia (HS)
and mastitis are the two most rife, endemic and economically important diseases of buffalo
and cattle. Hemorrhagic septicemia (HS) is an infectious, highly fatal bacterial disease
caused by Pasteurella multocida serotype B: 2 or Robert's type 1. Vaccination using different
types of vaccines (e.g. formalin-killed alum precipitated bacterin, oil adjuvanted vaccines) is
the single most important control measure against HS in Pakistan and in other Asian,
African, South European and Middle East countries where this disease is endemic. The
vaccines are quite effective when used as per the recommendations of manufacturers and
extension agencies. Alum precipitated bacterin is still the most commonly used HS vaccine
in Pakistan but it gives immunity for a short duration i.e. 3 months and its protective efficacy
is about 60% . Montanide® adjuvanted vaccines are considered to be superior to vaccines
containg other adjuvants. Mastitis is another common dairy disease which although not fatal,
causes colossal economic losses to our resource-poor dairy farmers and milk processing
industry. Research conducted over the past four decades has shown that S. aureus and Str.
agalactiae are the most prevalent mastitis pathogens in dairy animals in Pakistan. Whereas,
vaccination against HS is a fairly well established practice among Pakistan dairy farmers,
adoption of a mastitis control program is virtually nonexistent. Control of this mastitis is
imperative for economically viable dairy farming as well as to produce milk which conforms
to the standards of WTO Accord.
A combined HS-mastitis vaccine may potentially curtail the staggering economic
losses sustained by farmers due to these two diseases. Against this backdrop, the present
study was designed to develop and evaluated a Montanide® adjuvanted combined HS-
mastitis vaccine in laboratory settings and under field conditions. In phase 1 (preparation and
evaluation in laboratory settings), field isolates of P. multocida (n=3), S. aureus (n=4) and
Str. agalactiae (n=2) were scrutinized for their cultural and biochemical charateristics,
antibiograms, pathogenicity/virulence, and immunogenicity. The identity of P .multocida

116
serotype B and S. aureus was confirmed through PCR. The most pathogenic (LD50=10-7.75
; in
mice) and most immunogenic (in rabbits) P. multocida was selected for vaccine production.
From amongst the 4 candidate S. aureus isolates, the strongest biofilm producing (OD value
in spectrophotometric method =0.31) isolate with the highest pathogenicity (percent
mortality in rabbits=100) and immunogenicity (in rabbits) was selected for vaccine
production. Similarly, the more pathogenic (percent mortality in rabbits= 80) and more
immunogenic (in rabbits) of the 2 candidate Str. agalactiae isolates was selected for vaccine
production. Sterile bubaline whey was added @ 10% to nutrient broth for culturing the three
vaccinal organisms with a view to enhance expression of pseudocapsule of S. aureus and to
provide growth factors to Str. agalactiae for vaccine production. A 5ml dose of Montanide®
adjuvanted combine HS-mastitis vaccine contained 5×109 cells each of P. multocida, S.
aureus and Str. agalactiae along with their toxoids, 50μg thimerosal, 50μg sodium azide,
0.02 ml formalin and 2.5 ml Montanide® ISA 201. Evaluation of safety and side effects of
the test vaccine in cows (n=4) and buffaloes (n=4) with its double doses (10ml,
subcutaneously/ intramuscularly) indicated that the vaccine was safe and free of any
clinically significant side effect. The vaccine was found sterile and safe in rabbits. A
challenge-protection assay conducted in mice indicated 100% survival of challenged mice
that had been vaccinated twice 30 days prior to challenge. The vaccine was physically stable
in terms of pH, sedimentation, color, appearance, and syringibility for 6 months observation
period at 37°C.
In Phase II (field evaluation), the combined vaccine was evaluated in cows, buffaloes
and calves. To this end, a total of 70 S. aureus and Str. agalactiae free lactating animals
(buffaloes n=45; cows n=25 divided into vaccinal group A and control group B), 50
lactating cows and buffaloes microbiologically positive for S. aureus/Str. agalactiae
(buffaloes n=25, cow n=25 divided into vaccinal group C and control group D) and dairy
calves (buffalo calves n=70; cow calves n=50 divided into vaccinal group E and control
group F) aged up to 1 year were treated with 2 doses of combined HS-mastitis vaccine at 21
day interval and evaluated for 6 months in terms of ELISA based antibody titers against P.
multocida, S. aureus and Str. agalactiae, incidence of HS, local and systemic reactions,
incidence and prevalence of S. aureus and Str. agalactiae mastitis, mastitis screening tests

117
(Surf field mastitis test and CMT) reaction, severity of mastitis, milk somatic cell count,
effect of vaccination on milk yield, cost benefit analysis and vaccine efficacy.
Administration of Montanide® adjuvanted combined HS-mastitis vaccine produced
significantly (P˂0.05) higher ELISA-based antibody titers against P. multocida, S. aureus
and Str. agalactiae in vaccinated groups (A and C) than in un-vaccinated groups (B and D).
In general, the antibody titers were higher in non-diseased vaccinated (group A) animals
compared to diseased-vaccinated animals (group C) at 0, 60, 120 and 180 days post–
vaccination. It was observed that the antibody titers against P. multocida, S. aureus and Str.
agalactiae augmented at day 60 and 120 of vaccine administration in non-diseased
vaccinated animals and decreased at day 180 of post-vaccination. None of the 10 vaccinated
cows developed HS over the entire observation period of 180 days. Only 1 of the 30
vaccinated buffaloes (3.33%; group A) developed HS during this period. In the control group
(un-vaccinated-non diseased i.e group B), 1 cow (6.66%) and 2 buffaloes (13.33%)
developed HS during this period. None of the 25 cow calves vaccinated with Montanide®
adjuvanted combined HS-mastitis vaccine (group E) developed HS during the post
vaccination observation period of 6 months. One of the 50 buffalo calves (2%) vaccinated
with this vaccine (group E) developed HS during this period. One of 25 un-vaccinated cow
calves (4%; group F) and 3 of 20 buffalo calves (15%; group F) developed clinical HS over
the observation period of 6 months. Pasteurella multocida could be isolated from only 1
buffalo calf (group F) that succumbed to peracute HS. Isolation of P. multocida from other
cases was not successful.
Subcutaneously administration of vaccine was associated with a varying degree of
mild to moderate swelling at injection site in cows and buffaloes that subsided within 3-7
days while intramuscular administration caused either no or a very mild swelling at the
injection site that subsided within 48 hours. No systemic effects were noted (Table 4.20 and
4.21).
Determination of effect of Montanide® adjuvanted combined HS-mastitis vaccine on
quarter-based incidence of S. aureus and Str. agalactiae IMIs in cows and buffaloes
indicated that the quarter–based incidence of mastitis due to these 2 pathogens over the 6
months observation period was 3.3 folds higher in un-vaccinated cows and buffaloes.

118
Mastitis screening tests (Surf field mastitis test and CMT) based incidence of mastitis
in cows and buffaloes initially culture negative for S. aureus and Str. agalactiae indicated
that 4 of 40 quarters (10%) of vaccinated cows developed mastitis over the entire observation
period of 180 days. Only 2 of the 120 quarters (1.6 %) of vaccinated buffaloes developed
mastitis during this period. In the control group (un-vaccinated), 8 (13.33%) quarters of 15
cow and 6 (10%) quarters of 15 buffaloes developed mastitis during this period (Table 4.28).
Analysis of this parameter in cows and buffaloes initially positive for S. aureus and
Str. agalactiae indicated that 3 additional quarters of 15 vaccinated-disease cows (group C)
reacted positive in CMT/ SFMT over 180 days post vaccination period. The corresponding
figure in 15 vaccinated-diseased buffaloes (group C) was 2. In the un-vaccinated-diseased
cows (group D), 8 new quarters developed CMT/ SFMT positivity over 180 days post
vaccination period whereas the corresponding figure for the un-vaccinated –diseased
buffaloes (group D) was 6 ( table 4.30 , Figure 4.9).
The vaccine reduced the prevalence of S. aureus and Str. agalactiae IMIs over the
entire observation period of 180 days. Towards the end of study period (i.e. day 180), the
prevalence of Str. agalactiae and S. aureus IMIs was slightly higher than the prevalence of
these pathogens on day 60 and day 120. In the un-vaccinated- diseased group (group D), the
prevalence of these pathogens on days 60, 120 and 180 was higher than that on at day 0.
Mean± SEM of the mastitis severity score of group A (vaccinated; CMT/SFMT and
culture –ve for S. aureus/Str.agalactiae) was lower than those of group B (control;
CMT/SFMT and culture –ve for S. aureus/Str. agalactiae). Similarly, mean severity scores of
group C (vaccinated; CMT/SFMT and culture +ve for S. aureus/Str. agalactiae) were lower
(Table 4.32) than those of group D (control; CMT/SFMT and culture +ve for S. aureus/Str.
agalactiae).
Cumulative mean somatic cell counts in vaccinated groups (group A and C) were
significantly lower (P˂0.05) than those recorded in respective unvaccinated control groups
(group B and D). The vaccinated groups of both the species differed non-significantly
(P≥0.05) from each other.

119
When compared on the basis of overall milk production for a 6 months study period,
significantly higher (P<0.05) milk yield was recorded in vaccinated cows and buffaloes
(group A and C) as compared to respective unvaccinated control groups ( group B and D).
The vaccinated groups of both species revealed a non-significant difference from each other
with respect to milk yield. Cost-benefit analysis of vaccine indicated a net financial benefit
worth Rs 2,060,300.
The vaccine tested had efficacies of 84.78%.and 90.25% against HS and S.
aureus/Str. agalactiae associated mastitis.
In sum, Montanide® adjuvanted combined HS-mastitis vaccine tested in the present
study demonstrated a preventative role against HS and both preventative and curative role
against S. aureus and Str. agalactiae associated mastitis. In view of the preliminary nature of
the study, establishment of external validity as well as additional work involving much larger
number of cows, buffaloes and calves is clearly warranted.

120
LITERATURE CITED
Adlakha SC and SN Sharma, 1992. Infectious diseases. In: Tulloh NM and JHG. Holmes
(editors) Buffalo Production. Elsevier, Amsterdam, Netherland.
Afzal M and R Muneer, 1990. Development of a combined vaccine for haemorrhagic
septicemia and foot and mouth disease. Pak Vet J, 10: 67-69.
Afzal M, 2009. Improving veterinary service in Pakistan. Pak Vet J, 29: 206-210.
Ahmad R and NA Naz, 2000. Incidence and therapy of pasteurellosis in buffalo calves. Pak
Vet J, 20: 101-102.
Ahmad T and G Muhmmad, 2008. Evaluation of Staphylococcus aureus and Streptococcus
agalactiae aluminium hydroxide adjuvanted mastitis vaccine in rabbits. Pak J Agri
Sci, 45: 353-361.
Ahmad T, 2009. Therapeutic efficacy of an aluminium hydroxide adjuvanted Staphylococcus
aureus and Streptococcus agalactiae bacterin-toxoid alone and in combination with
antibiotics for the treatment of clinical and sub- clinical bubaline mastitis. Ph.D
Thesis, Deptt Clinical Medicine and Surgery, Univ Agri Faisalabad, Pakistan.
Ajmal M, 1990. Livestock wealth of Pakistan. Proc. 3rd International Congress. Pak Vet Med
Assoc and University Grants Commission, Islamabad. Nov 29; pp: 379-392.
Akhtar A, Khairani-Bejo, A Umer, J Tanweer and Habibullah, 2012. Prevalence of mastitis
and identification of causative pathogens in local and crossbred cows in Dera Ismail
Khan. Pak J Sci, 64: 265-268.
Ali L, 2008. Epidemiology of mastitis in dairy buffaloes and cows in Tehsil Samundri of
district Faisalabad. Ph.D. Thesis, Department of Clinical Medicine and Surgery;
University of Agriculture, Faisalabad, Pakistan.
Ali Z, K Muhammad, I Hussain and E Hameed, 2000. Antibody response of buffaloes to
haemorrhagic septicaemia vaccine.International J Agri Biol, 3:183–186.
Allore HG, 1993. A review of the incidence of mastitis in buffaloes and cattle. Pak Vet J, 13:
1–7.

121
Altaf, M Siddique, K Muhammad, M Irshad, MZ Khan, AA Anjum and M Kamran, 2012.
Antibody response of rabbits to combined hemorrhagic septicemia and foot & mouth
disease virus vaccine. J Anim Plant Sci, 22: 501-504.
Altwegg M and J Bockemühl, 1998. Escherichia and Shigella. In: Topley and Wilson‟s
Microbiology and Microbial Infections. 9th Ed Vol II Arnold, London pp, 946–947.
Amorena B, R Baselga and H Albizu, 1994. Use of liposome-immuopotentiated
exopolysaccharide as a component of an ovine mastitis staphylococcal
vaccine.Vaccine, 81: 243-249.
Anonymous, 1996. Economic analysis and survey planning. In: Epidemiology survey,
Punjab, 1994. Pak-German Bilateral Technical Cooperation. Directorate of Planning
and Evaluation. Department of Livestock and Dairy Development, Government of
Punjab-Pakistan, pp: 4-5.
Anonymous, 2004. SAS, Statistical Software Version 9.1. SAS Institute Inc., Cary, NC,
USA.
Anonymous, 2009. Quarterly report on status of notifiable animal diseases in Punjab.
Directorate of animal disease surveillance and reporting system, livestock and dairy
development, Punjab, Pakistan. Diradrns@ punjab.gov.pk.
Arif J, S Rehman, M Arshad, and P Akhter, 2013. Immunopotentiation of outer membrane
protein through anti-idiotype Pasteurella multocida vaccine in rabbits. Biol, 41: 339-
334.
Arshad M, FK Qamar, M Siddique and STAK Sindhu, 1995. Studies on some
epidemiological aspects of bovine mastitis. In: Proc. National Seminar on
Epidemiology of Livestock and Poultry Diseases, Jan. 19-20. College of Vet Sci,
Lahore, Pakistan.
Arthington JD and LJ Havenga, 2011. Effect of injectable trace minerals on the humoral
immune response to multivalent vaccine administration in beef calves. J Anim Sci
90: 1966-1971.
Arye- Smith R, 1971. Principles of increasing livestock production in developing countries.
Trop Anim Hlth Prod, 3: 43-49.

122
Ashfaq M, G Muhammad, Shamsheer-ul-Haq, A Razzaq, 2014. Effects of livestock diseases
on dairy production and In-comes in district Faisalabad, Punjab, Pakistan. Pakistan
Strategy Support Program-International Food Policy Research Institute Working
Paper No.023.
Ashraf A, H Tariq, S Shah, Nadeem, I Manzoor, S Ali, A Ijaz, S Gailani and S Mehboob,
2011. Characterization of Pasteurella multocida strains isolated from cattle and
buffaloes in Karachi, Pakistan. African J Microbiol Res, 5: 4673-4677.
Ashraf S, A Omer, M Ijaz, UN Chaudry and MM Ali, 2009. Efficacy of florfenicol against
hemorrhagic septicemia in buffalo calves. Pak J Zool Supp Ser, 9: 119-122.
Ataei S, R Burchmore, HJ Christopher, A Finucane, R Parton and JG Coote, 2009.
Identification of immunogenic proteins associated with protection against
haemorrhagic septicaemia after vaccination of calves with a live-attenuated aroA
derivative of Pasteurella multocida B: 2. Res Vet Sci, 87: 207-210.
Athar M, 2007. Preparation and evaluation of inactivated polyvalent vaccines for the control
of mastitis in dairy buffaloes. PhD Thesis, Deptt. Clinical Medicine and Surgery,
Univ Agri, Faisalabad, Pakistan.
Athar M, G Muhammad, A shakoor, M Saqib, R Ahmad and A Naureen, 2007. A
preliminary study on the effect of inactivated polyvalent mastitis vaccines on milk
quantity and quality in dairy buffaloes. Pak Vet J, 27: 85-91.
Aucouturier J, L Dupuis, and V Ganne, 2001. Adjuvants designed for veterinary and human
vaccines. Vaccine, 19: 2666–2672.
Bachaya HA, Z Iqbal, G Muhammad, A Yousaf and HM Ali, 2005. Subclinical mastitis in
buffaloes in Attock district of Punjab (Pakistan). Pak Vet J, 25: 134-136.
Bain RVS, MCL De Alwis, GR Carter and BK Gupta, 1982. Haemorrhagic septicemia. FAO
Animal and health paper No.33. FA0, Rome, Italy.
Baker JC, ST Rust, DK Ciszewski, and PH Coc, 1986. A safety trial of a bovine respiratory
syncytial virus vaccine in feedlot calves. In: Proc. 19th Annual Conference American
Association of Bovine Practitioners Nov 18-21, 1986. Iouisville, Kentucky, USA,
pp: 70-72.

123
Barbano, DM, 1989. Impact of mastitis on dairy product quality and yield: Research update.
28th
Annual Meeting, National Mastitis Council, Inc. Tampa, Florida, USA.
Barnett PV, L Pullen, L Williams and TR Doel, 1996. International bank for foot- and-
mouth disease vaccine: Assessment of montanide ISA 25 and ISA 206, two
commercially available oil adjuvants. Vaccine, 14: 1187-1198.
Benkirane A and MCL De Alwis, 2002. Hemorrhagic septicaemia, its significance,
prevention and control in Asia. Vet Med Czech, 47: 234-240.
Bishop S, M de Jong, D Gray, 2010. Opportunities for incorporating genetic elements into
the management of farm animal disease: policy issues. Commission on Genetic
Resources for Food and Agriculture. Background study paper No.18. Accessed from
www.fao.org/ag/magazine/bsp 18-e.pdf.
Blood DC and VP Studdert, 1999. Page 1380 in saunders comprehensive veterinary
dictionary, WB Saunders, Philadelphia, PA, USA.
Bozhkova G, B Bozhilov, Ya Bachvarova, D Stoychev, P Gancheva, Y Gogov, P Petkov, D
Stoichev, I Gogov and J Wisniowshi, 1985. Clinical, cytological and microbiological
investigations on the milk gland at cow mastitis immunity-prophylaxis measures.In:
proV Int Symp. Mastitis Control, pp: 12:1-26.
Brakstad OG, K Aasbakk and JA Maeland, 1992. Detection of Staphylococcus aureus by
polymerase chain reaction amplification of „nuc‟ gene. J Clin Microbiol, 30: 1654-
1660.
Butt AA, 2006. Evaluation of four adjuvanted trivalent vaccines for the control of mastitis in
dairy buffaloes and cows. Ph.D. Thesis, Department of Clinical Medicine and
Surgery, University of Agriculture, Faisalabad, Pakistan.
Butt IA, K Tasneem, A Raza and ZJ Gill, 2003. Biochemical, serological and immunological
properties of Pasteurella multocida strains isolated from natural outbreaks of
haemorrhagic septicaemia. Pak J Vet Res, 1:1-4.
Cady RA, SK Shah, EC Schermerhorn and RE McDowell, 1983. Factors affecting
performance of Nili-Ravi buffaloes in Pakistan. J Dairy Sci, 66: 578-586.
Calzolari A, JA Giraudo, H Rampone, L Odierno, AT Giraudo, C Frigerio, S Bettera, C
Raspanti, J Hernandez, M Wehbe, M Mattea, M Ferrari, Al Larriestra and R Nagel.

124
1997. Field trials of a vaccine against bovine mastitis. 2. Evaluation in two
commercial dairy herds. J Dairy Sci, 80: 854–858.
Carter GR, 1984. Serotyping Pasteurelia multocida. Methods in Microbiol, 16: 247-258.
Carter GR, 2005. Hemorrhagic septicemia. In: Kahn, C. M (ed.). The Merck Veterinary
Manual. 9th
Ed. Merck & Co., Inc. Whitehouse Station, New Jersey, USA. pp: 607-
609.
Carter GR, and MCL De Alwis, 1989. Haemorrhagic septicaemia. In: Pasteurella and
Pasteurellosis, Adlam C and JM Rutter (eds.) Academic Press, London. pp: 131-160.
Castrucci G, M Ferrari, V Angelillo, F Rigonat and L Capodicasa, 1993. Field evaluation of
the efficacy of romovac 50, a new inactivated, adjuvanted bovine rotavirus vaccine.
Comp Immunol Microbiol Infect Dis, 16: 235-239.
Catt DM, MM Chengappa, WL Kadel, and CE Herren, 1985. Preliminary studies with a live
streptomycin-dependent Pasteurella multocida and Pasteurella haemolytica vaccine
for the prevention of bovine pneumonic pasteurellosis. Can J Comp Med, 49: 366-
371.
CDC, 2011. Morbidity and mortality weekly report (MMWR), general recommendations on
immunization, and recommendations of the advisory committee on immunization
practices (ACIP), 60: 1-64.
Cengiz S, G Dinc and M Cengiz, 2015. Evaluation of antimicrobial resistance in
Staphylococcus spp. isolated from subclinical mastitis in cows. Pak Vet J, 35: 334-
338.
Chakrabarti A, 2002. Hemorrhagic septicemia.A text book of preventive veterinary medicine.
pp: 255-262.
Chaudhry NA and BB Khan, 1978. Estimation of economic losses due to animal diseases in
Punjab (Final report of research project). Univ Agri, Faisalabad, Pakistan.
Chhabra R, R Sharma and NK Kakker, 2004. Comparative immunogenicity of foot and
mouth disease virus antigens in FMD–haemorrhagic septicemia combined vaccine
and FMD vaccine alone in buffalo calves. Indian J Exp Biol, 42:259-264.
Chisti, MA, M Afzal and R Muneer, 1992. Effect of immunopotentiating on subclinical
mastitis in cattle and buffaloes. Asian Aust J Anim Sci, 5: 733-736.

125
Chowdhury M, J Mitra, S Sarkar, T Samanta and BB Roy, 2014. PCR and electron
microscopy based diagnosis of an outbreak of haemorrhagic septicemia in buffalo
and its control in a farm of West Bengal, India. Explor Anim Med Res, 4: 86-94.
Christensen GD, WA Simpson, AL Bisno and EH Beachey, 1982. Adherence of slime
producing strains of Staphylococcus epidermidis to smooth surfaces. J Infect Immun,
37: 318-326.
Citak S, O Varlik and N Gundogan, 2003. Slime production and DNase activity of
staphylococci isolated from milk. J. Food Safety, 23: 219-292.
Claxton PD and D Ryan, 1993. Bovine mastitis bacteriology. In: Jeans K. (Managing ed.)
Australian Standard Diagnostic Techniques for Animal Diseases. Commonwealth of
Australia and each of its States and Territories. CSIRO Information Services,
Victoria, Australia, pp: 3-8.
Clinical Laboratory Standards Institute, 2012. Performance Standards for Antimicrobial
Susceptibility Testing; Twenty-Fifth Informational Supplement (M100-S25)
Cohen JO, 1987. Staphylococcosis. J Am Vet Med Assoc, 190: 150-152.
Cook DR, HT Hill and DR Kinker, 1990. Efficacy of a killed gpX deleted pseudorabies virus
vaccine. Can J Vet Res, 54: 438-445.
Costerton JW, PS Stewart, and EP Greenberg, 1999. Bacterial biofilm: a common cause of
persistent infectious. Science, 284: 1318-1322.
Dagleish MP, JC Hodgson, S Ataei, A Finucane, J Finlayson, J Sales, R Parton and JG
Coote, 2007. Safety and protective efficacy of intramuscular vaccination with a live
aroA derivative of Pasteurella multocida B: 2 against experimental hemorrhagic
septicemia in calves. Infect Immun, 75: 5837-5844.
De Alwis MCI, 1981. Mortality among cattle and buffaloes in Sir Lanka due to haemorrhagic
septicaemia. Trop Anim Hlth Prod, 13:195-202.
De Alwis MCL, 1984. Haemorrhagic septicaemia in cattle and buffaloes. Rev Sci Tech Off
Int Epiz, 3: 707-730.
De Alwis MCL, 1992. Haemorrhagic septicaemia. A general review. Br Vet J, 148:99-112.

126
De Alwis MCL, 1995. Haemorrhagic septicemia in cattle and buffaloes.In:W.Donachie et al.
(ed.), Haemophilus, Actinobacillus and Pasteurella. Plenum Press, New York, pp.9-
24.
De Alwis MCL, 1999. Haemorrhagic septicaemia. Australian Center for international
Agriculture Research (ACIAR) monograph no.57, Canberra, Austalia, pp: 1-34.
De Alwis MCL, GR Carter, and MM Chengappa, 1980. Production and characterization of
streptomycin dependent mutants of Pasteurella multocida from bovine haemorrhagic
septicaemia. Can J Comp Med, 44: 418-422.
De Belder AN, 1985. Dextran .Pharmacia AB, Uppsala, Sweden pp: 71.
De Vliegher S, 2004. Udder Health in Dairy Heifers - some epidemiological and
microbiological aspects. Ph.D. Thesis, Dept. of Repro, Obstetrics and Herd Health,
Fac Vet Med Ghent, University, Belgium.
Deb R, A Kumar, S Chakraborty, AK Verma, R Tiwari, K Dhama, U Singh and S Kumar
2013. Trends in diagnosis and control of bovine mastitis: a review. Pak J Biol sci, 23:
1653-1661.
DeGraves FJ and J Fetrow, 1993. Economics of mastitis and mastitis control. Vet Clin North
Am, Food Anim Pract, 9: 421-434.
Derbyshire JB, I Davidson and CD Wilson, 1961. Symposium on staphylococcal mastitis.
Vet Rec, 73: 1011-1033.
Dhand NK, RM Gopal, J Singh, DR Sharma, KS Sandhu and PK Sindhu, 2002. Disease
outbreaks in the field and emerging disease an overview. Proceedings of the animal
husbandry officers workshop department of veterinary and animal husbandry
extension, agricultural university (ed KB Sing, H.K Verma) pp: 1-8.
Dhillion SS, MS Kwatra and PN Dhingra, 1971. In intestinal form of pasteurellosis following
rinderpest vaccination in Punjab. Bulletin of international epizootics 75:335-341.
Dhoot VM and SV Upadhye, 2001. Pasteurellosis in a chital deer (Axis Axis) in captivity.
Zoos Print J, 16: 428-429.
Dutta J, BS Rathore, SG Mulick, R Singh and GC Sharma, 1990. Epidemiological studies on
occurrence of hemorrhagic septicemia in Inida. Ind Vet J, 67: 893-899.

127
Egenolf P, 1990. Health and production problems of smallholder ruminants in the Punjab,
Pakistan. Animal Res. Development, Instt for Scientific Cooperation, Tubingen,
Germany, 32: 104-111.
Ekundayo SO, MO Odugbo, AO Olabode and PA Okewole, 2008. Phenotypic variability
among strains of Pasteurella multocida isolated from avian, bovine, caprine,
leporineand ovine origin.African J. Biotechnol. 7: 1347-1350.
Elshemey, TM and AH Abd-Elrahman, 2013. Hemorrrhagic septicemia outbreak as a
consequence of SAT2 FMD infection in buffalo and cattle in Alexandria province,
Egypt. Life Sci J, 10: 816-822.
EMEA, 2000. The European Agency for the Evaluation of Medical Products Evaluation of
Medicines for Veterinary Use .CVMP/IWP/52/97-Final, page: 1-6.
FAO, 1998. Production Yearbook. Food and Agri. Organization, Rome, Italy.
Farooq U, M Hussain, H Irshad, N Badar, R Munir and Q Ali, 2007. Status of haemorrhagic
septicaemia based on epidemiology in Pakistan. Pak Vet J, 27: 67-72.
Farooq U, Z Saeed, MA Khan, Ali and MF Qamar, 2011. Sero-surveillance of hemorrhagic
septicemia in buffaloes and cattle in Southern Punjab, Pakistan. Pak Vet J, 31: 254-
256.
Freeman DJ, FR Falkiner and CT Keane, 1989. New method for detecting slime production
by coagulase negativttfxse staphylococci. J Clini Pathol, 42: 872-874.
Garcia O, K Mahmood and T Hemme, 2003. A review of milk production in Pakistan with
particular emphasis on small-scale producers. PPLPI working paper no.3.
International Farm Comparison Network (IFCN) of FAO. P: 5.
Geering WA, and J Lubroth. 2002. Preparation of Foot-and- Mouth Disease Contingency
Plans .FAO Animal Health Manual. No.16. FAO, Rome Italy P: 51.
Giraudo JA, A Calzolari, H Rampone, AT Giraudo, C Bogni, A Larriestra and R Nagel,
1997. Field trials of a vaccine against bovine mastitis. I. Evaluation in heifers. J
Dairy Sci, 80:845-853.
Golde WT, T deLosSantos, L Robinson, MJ Grubman, N Sevilla, A Summerfield and B
Charleston. 2011. Evidence of activation and suppression during the early immune
response to foot-and-mouth disease virus. Transbound Emerg Dis, 4:283-290.

128
Hameed S, M Arshad, M Ashraf, M Avais and MA Shahid, 2012. Cross-sectional
epidemiological studies on mastitis in cattle and buffaloes of tehsil Burewala,
Pakistan. J Anim Plant Sci, 22: 371-37.
Heeschen, WH, 2005. Somatic cells as indicator of milk hygiene, scientific basis and the EU
approach. Proc. 44th
National Mastitis Council Annual Meeting.Orlando, FL, USA,
pp.52-72.
Heidrich HJ and W Renk, 1967. Diseases of the Mammary Glands of Domestic Animals.
Translated from German by LW Van, Den Heever, WB Saunders Co. Philadelphia
USA.P. 205.
Hensen SM, 2000. In bovine Staphylococcus aureus mastitis bacterial adhesion and invasion
in relation to pathogenesis and antimicrobial sensitivity.University of Utrecht,
Utrecht, pp. 113-129.
Hirsch JG and B Strauss, 1964. Studies on heat-labile opsonin in rabbit serum. J Immunol,
92: 145-154.
Hoedemaker M, B Koff, B Edler, M Emmert and E Bleckman, 2001. Dynamics of
Staphylococcus aureus infections during vaccination with an autogenous bacterin in
dairy cattle. J Vet Med, 48:373-383.
Hogan JS, RN Gonzalez, RJ Harmon, SC Nickerson, SP Oliver, JW Pankey, and KL Smith
1999. Laboratory Handbook on Bovine Mastitis Revised ed. National Mastitis
Council, Wisconsin.
Holden S, S Ashley, and P Bazeley, 1996. Improving the delivery of animal health services
in developing countries-A literature review. Accessed from:
http://www.theidlgroup.com/documents/improving the delivery of Animal Health
Services in Developing Countries. Pdf.
Horadagoda NU, JC Hodgson, GM Moon, TG Wijewardana and PD Eckersall, 2001. Role of
endotoxin in the pathogenesis of haemorrhagic septicemia in the buffalo, Microbial
Pathogenesis, 30: 171-178.
Horadagoda NU, JC Hodgson, GM Moon, TG Wijewardana and PD Eckersall, 2002.
Development of clinical syndrome resembling haemorrhagic septicemia in the
buffalo following intravenous inoculation of Pasteurella multocida B: 2 endotoxin

129
and the role of tumour necrosis factor-alpha. Res. Vet Sci, 72: 194-200.
Hussain M, MA Malik, Z Fatima, and MR Yousaf, 2005. Participatory surveillance of
livestock diseases in Islamabad capital territory. Intl J Agric Biol, 7: 567-570.
Hussain R, F Mahmood, A Khan, M Z Khan, and A B Siddique, 2014. Pathological and
molecular based study of pneumonic pasteurellosis in cattle and buffalo (Bubalus
bubalis). Pak J Agri Sci, 51: 235-240.
ISO, 1994. Quality management and quality assurance. Int Org for Standardization
Vocabulary, ISO8402; Geneva, Switzerland.
Joshi VB, KK Baxi and DS Sambyal, 1987. Prevalence of pasteurella multocida in domestic
animals and birds of Punjab, Journal of research, Punjab agricultural university 24:
484-486.
Jost NP, 1980. Livestock Production and Marketing Survey Punjab. Report # 5. Production
and Sales of Milk and Ghee. LDC, LEP/AI and GTZ PN 74: 2041-2047.
Kehrenberg C, G Schulze-Tanzil, JL Martel, EC Dancla and S Schwarz, 2001. Antimicrobial
resistance in pasteurella and mannheimia: epidemiology and genetic basis. Vet Res,
32: 323–339.
Keskin O, G Altay and M Akan, 2003. Farkli hayvansal kaynaklardan izole edilen koagulaz
negatif stafilokoklarda slime uretimi ve adherans. Turk. J Vet Anim Sci, 27: 253-
257.
Khan A, KS Saleemi, MZ Khan, ST Gul, M Irfan, S Qamar, 2011. Hemorrhagic septicemia
in buffalo (Bubalus bubalis) calves under sub-tropical conditions in Pakistan. Pak J
Zool, 43: 295-302.
Khan A, U Saddique, R Ahmad, H Khan, Y Mohammad and M Zubair, 2006. Sero-
serveillance of hemorrhagic septicemia in cattle and buffaloes in district Malakand,
NWFP. J Agri Biol Sci, 1:11-14.
Khan AZ, and G Muhammad, 2005. Quarter-wise comparative prevalence of mastitis in
buffaloes and crossbred cows. Pak Vet J, 25: 9-12.
Khan FM. 2013. Second Quarterly Notifiable Disease Forecast Report of District Rahim Yar
Khan. 58-60/ADIO/RYK/02.05.21.1-8.

130
Khan MA, M Ajmal, M Yamin, MS Khan and MA Athar, 1991. Epidemiological and
economical based ranking order of buffalo and cattle diseases through active
surveillance system. Pak J Livestock Res, 1: 38-43.
Khan MA, MS Khan, HR Chaudhry, J Khan and S Saleem, 1994-1995. Meta analysis of
epidemiological and economical losses and ranking order of buffalo and cattle
disease in Punjab. Project report on Epidemiological of Major Livestock Diseases in
Pakistan. pp: 73-90.
Khan MS and HZ Chaudhry, 2001. Short and atypical lactations in Nili-Ravi buffaloes.
Pakistan Vet J, 2: 124-127.
Kluger MJ and LK Vaughn, 1978. Fever and survival in rabbits infected with Pasteurella
multocida. J Physiol, 282: 243-251.
Koiranen L, 1977. Significance of vaccination for prevention of mastitis. Suomen
Elainlaakarilehti. 83: 77-78.
Kumar H, V Mahajan and S Sharma, 2007. Concurrent pasteurellosis and clinical swine fever
in Indian pigs. J Swine Health Prod, 15: 279-283.
Leitner G, E Lubashevsky, A Glickman, M Winker, A Saran and Z Trainin, 2003.
Development of a Staphylococcus aureus vaccine against mastitis in dairy cows. I
Challenge trials. Vet Immunol Immunopathol, 93:31-38.
Leitner G, O Krifucks, MD Kiran and N Balaban, 2011. Vaccine development for the
prevention of staphylococcal mastitis in dairy cows. Vet Immunol Immunopathol,
142: 25-35.
Lignieres and spitz, 1902. Bul Soc Cent Med Vet, 20: 487.
Lu YS, and SP Pakes, 1981. Protection of rabbits against experimental pasteurellosis by a
streptomycin-dependent Pasteurella multocida serotype 3: A live mutant vaccine.
Infec and Immun, 34: 1018-1024.
Maddur MS, S Kishore, AK Chockalingam, S Gopalakrishna, N Singh, VVS Suryanarayana,
and MR Gajendragad, 2010. The relationship between cellular immune response to
foot-and-mouth disease virus Asia 1 and viral persistence in Indian cattle (Bos
indicus). Res in Vet, 89: 36-40.

131
Mathur T, S Singhal, S Khan, DJ Upadhyay, T Fatma and A Rattan, 2006. Detection of
biofilm formation among the clinical isolates of staphylococci: an evaluation of three
different screening methods. Indian J Med Microbiol, 24: 25-29.
May BJ, QT, Zhang and V Kapur, 2001. Complete genomic sequence of Pasteurella
multocida, Pm70. Proceedings of the National Academy of Sciences of the United
States of America, 98: 3460-3465.
Melchior MB, H Vaarkamp and JF Gremmels, 2006a. Biofilms: a role in recurrent mastitis
infections. Vet J, 171: 398-407.
Melchior MB, MHJV Ocsh, RM Graat, EV Duijkeren, DJ Mevius, M Nielen, W Gaastra and
JF Gremmels, 2009. Biofilm formation and genotyping of Staphylococcus aureus
bovine mastitis isolate: Evidence for lack of penicillinresistance in Agr-type II
strains. Vet Microbiol, 137: 83-89.
Miller GY, 1986. The economic of mastitis control. Bovine Pract, 21: 79-81.
Muhammad G, A Naureen, MN Asi, M Saqib and Fazal-ur-Rehman, 2010. Evaluation of a
3% surf solution (surf field mastitis test) for the diagnosis of subclinical bovine and
bubaline mastitis. Trop Anim Hlth Prod, 42: 457-464.
Muneer R and M Afzal, 1989. Preliminary studies on improved oil-adjuvant vaccine for
haemorrhagic septicaemia in buffalo calves. Rev Sci Tech Off Int Epiz, 8: 999-1004.
Muneer R, M Hussain and AB Zahur, 2005. Efficacy of oil based haemorrhagic septicaemia
vaccine: A field trial. lntl. J Agri Biol, 7: 571-573.
Muneer R, S Akhtar and M Afzal, 1994. Evaluation of three oil-adjuvant vaccines against
Pasteurella multocida in buffalo calves. Rev Sci Tech Off Int Epiz, 13: 837-843.
Myint A, TO Jones, and HH Nyunt, 2005. Safety, efficacy and cross-protectivity of a live
intranasal aerosol haemorrhagic septicaemia vaccine. Vet Rec, 256: 41 –45.
Nadeem M, I Hussain, T Ahmad, AS Qureshi and A Yousaf, 2010. Effect of prophylactic
application of levamisole in buffalo-calves immunized with haemorragic septicemia
vaccine. In: Crudeli GA, EM Patino and JL Konard ( Guest Editors).Proc. 9th
World
Buffaloes Congress , Buenos Aires, Argentina, pp: 451-454.
Namioka S, and DW Bruner. 1963. Serological studies on Pasteurella multocida, IV. Type
distribution of the organisms on the basis of their capsule and O groups. Cornell Vet,

132
53:41-53.
National Mastitis Council Inc. 1987. Laboratory and Field Handbook on Bovine Mastitis.
National Mastitis Council Inc., Arlington, Virginia, USA.
National Mastitis Council, Inc., 1990. Microbiological Procedures for the Diagnosis of
Bovine Udder Infection. National Mastitis Council, Inc., Arlington, Virginia, USA.
Naz S, A Hanif, A Maqbool, S Ahmed and K Muhammand, 2012. Isolation, characterization
and monitoring of antibiotic Resistance in Pasteurella multocida isolates from
buffalo (Bubalus Bubalis) herds around Lahore. J Anim Plant Sci, 22(3 Suppl.): 242-
245.
Nickerson SC, 1994. Progress in the development of mastitis vaccine. Proc. National Matitis
Council, Inc, Arlington, USA, PP: 133-134.
Nickerson SC, WE Owens and RL Boddi, 1993. Effect of Staphyloccocus aureus bacterin on
serum antibody, new infections and mammary histology in non-lactating cows. J
Dairy Sci, 76:1290-1297.
Nickerson SC, WE Owens and RL Boddie, 1995. Mastitis in dairy heifers: initial studies on
prevalence and control. J Dairy Sci, 78: 1607-1618.
Norcross NL and K Kenny, 1986. Evaluation of a vaccine for mastitis caused by
Staphylococcus aureus. Proc. 14th World Congress on Diseases of Cattle. Dublin.
Pp: 1094-1099.
Nordhaug ML, LL Nesse, NL Norcross, and R Gudding, 1994. A field trial with an
experimental vaccine against Staphylococcus aureus mastitis in cattle. 1. Clinical
parameters. J Dairy Sci, 77: 1267–1275.
OIE, 2012. Haemorrhagic Septicaemia. In: OIE Manual of Standards for Diagnostic Tests
and Vaccines, 3rd
Ed. Office International De Epizootics, Paris, pp: 739-751.
Oliver SP, MJ Lewis, BE Gillespie, H H Dowlen, EC Janicke and RK Robers. 2003. Milk
production, milk quality and economics benefit associated with prepartum antibiotic
treatment of heifers. J Diary Sci, 86:1187-1193.
Oreste and Armani, 1887. Cited by Bain et al. 1982.
Osman OA, RS Abdul, S Makeen, RA Dinvitiri, MA Monaz and RR Youswf, 1990. The
simultaneous inoculation of cattle with rinderpest and haemorrhagic septicaemia

133
vaccines. Australian Vet Med J, 23: 57–63.
Otomaru KS, Kubota and M Tokimori 2015. Maternally and naturally acquired antibodies to
Pasteurella multocida in Japanase black calves. Pak Vet J, 35: 108-110.
Owens WE, CH Ray, RL Boddie, NT Boddie, CA Pratt, SC Nickerson, JW Hallberg and AP
Belchner, 1995. Efficacy of sequential pirlimycin intramammary treatment against
chronic Staphylococcus aureus intramammary infection. In Proc Natl Mastitis
Counc, Inc Arlington, VA, pp: 144-145.
Owens WEJL, RL Watts, SC Boddie and S.C. Nikerson, 1988. Treatment of mastitis:
Comparison of intramammary and intramammary plus intramuscular therapies. J
Dairy Sci, 7: 3143-3147.
Pankaj A Sharma, R Chhabra and N Sindhu, 2013. Sub-clinical mastitis in Murrah buffaloes
with special reference to prevalence, etiology and antibiogram. Buffalo Bulletin. 32:
107- 115.
Pankey JW, G Murray and A Twomey, 1983. Evaluation of a commercial bacterin against S.
aureus mastitis in New Zealand. Dairy Res Report, 1983. Hill Far res Stn, Homer,
LA, pp:157-161.
Patil PK, J Bayry, C Ramakrishna, B Hugar, LD Misra and C Natarajan, 2002. Immune
responses of goats against foot and mouth disease quadrivalent vaccine: Comparison
of double oil emulsion and aluminum hydroxide vaccines in eliciting immunity.
Vaccine, 20: 2781-2789.
Pereira UP, DG Oliveira, LR Mesquita, GM Costa, and LJ Pereira, 2011. Efficacy of
Staphylococcus aureus vaccines for bovine mastitis: a systematic review. Vet
Microbiol, 148:117-124.
Perry BD, McDermott, JJ, Randolp TF, KR Sones, and Thornton, PK. 2002.Investing in
animal health research to alleviate poverty. International Livestock Research Institute
(ILRI), Nairobi, Kenya.
Qureshi S, and HM Saxena, 2014. Estimation of titers of antibody against Pasteurella
multocida in cattle vaccinated with haemorrhagic septicemia alum precipitated
vaccine. Vet World, 7: 224-228.

134
Qureshi, ZI, LA Lodhi, H Jamil and M Nawaz, 2000. Effect of levamisole hydrochloride on
serum and colostral antibody titres against foot and mouth disease virus in vaccinated
buffaloes (Bubalus bubalis). Vet Arch, 70: 59-66.
Radostits OM, CC Gay, KW Hinchcliff, and PD Constable, 2007. Veterinary Medicine - A
textbook of diseases of cattle, sheep, pigs, goats and horses. 10th
Ed., WB Saunders
Co, Philadelphia, USA.
Rashid I, 2011. A short-term field evaluation of a mastitis vaccine prepared from a biofilm
producing local isolate of Staphylococcus aureus, M Phil Thesis, Deptt Clinical
Medicine and Surgery Univ Agri Faisalabad, Pakistan.
Raza A, 2011.Evaluation of a bacterin-toxoid mastitis vaccine prepared from a biofilm
producing isolate of Staphylococcus aureus in rabbits. M Phil Thesis Deptt of
Clinical Medicine and Surgery; University of Agriculture, Faisalabad.
Raza MA, G Muhammad, M Athar and M. Zargham Khan, 2000. Comparative efficacy of
three protocols for the treatment of haemorrhagic septicemia in buffaloes and cattle.
Pak Vet J, 20: 35-39.
Reddy GS, KA Rao and VA Srinivasin, 1995. Immunity conferred by oil-adjuvant
haemorrhagic septicaemia vaccine. Indian J Anim Sci, 66: 703-704.
Refai M, SA Youssef, NA Saleh, SA Attia, HM Zaki, W Tawakkol and BS Moshref, 2005.
Studies of virulence factors of staphylococcus aureus isolatesd from mastitic
buffaloes.Vet Med J Giza. 2: 253-265.
Riaz M, MS Khan, MA Khan, AA Rabbani and M Arshad. 1992. Investigation on
epidemiology and economic losses of major livestock diseases in district Gujrat. Pak
Vet J 12: 86-88.
Richou R and G Thienlin, 1955. Sur la prevention et le traitment des mammities. Recueil
Med Vet, 131: 73-85.
Roberson JR 2003. Establishing treatment protocols for clinical mastitis.Vet Clin Food Anim
19,:223-234.
Rodney MD, 2002. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis., 8: 881- 890.
Saini SS, DR Sharma, BS Gill, MS Kwatra, J Singh, JK Sharma, SS Dhillion and Ramneek
(1991). Reemergence of hemorrhagic septicemia in Punjab. Ind J ani sci 61: 1178-

135
1180.
Sajjad-ur-Rahman, M Athar, A Shakoor, G Muhammad and AA Butt, 2004. Standardization
of indirect haemagglutination test for titration of antibodies against Staphylococcus
aureus, Streptococcus agalactiae and Escherichia colie isolated from bubaline
mastitis. Intl J Agri Biol, 7: 35-40.
Schalm OW, EJ Carrol and NC Jain, 1971. Bovine Mastitis. Lea and Febiger, P hiladelphia,
USA.
Scott JP. 1931. Aggressins: An outline of the development of the theory and notes on the use
of these products. J Bacteriol, 22: 323-337.
Sears PM, and KK McCarthy, 2003. Management and treatment of staphylococcal mastitis.
Vet Clin of North America-Food Anim Pract, 19: 171-185.
Shahzad W, R Munir, M Asif, MS Sagar, M Altaf, W Aslam, G Akbar and F Mehmood,
2013. Prevalence, molecular diagnosis treatment of field isolates of toxogenic
Pasteuralla multocida in a hemorrhagic septicemia outbreak in Nili-Ravi Buffalo
calves at Livestock Experiment Station, Bahadurnugar, Okara, Pakistan.
Shailja and M Singh, 2002. Effect of post milking teat dip on somatic cell count, milk
production and composition in cows and buffaloes. Asian-Aust J Anim Sci, 15:1517-
1522.
Shakoor A, 2006. Preparation and evaluation of Staphylococcus aureus vaccines for the
control of mastitis in dairy buffaloes (Bubalus bubalis), Ph.D Thesis, Deptt Clinical
Medicine and Surgery, Univ Agri Faisalabad, Pakistan.
Shakoor A, M Athar, G Muhammad, SU Rahman, AA Butt, I Hussain and R Ahmad, 2006.
Effect of different Staphylococcus aureus mastitis vaccines on the milk yield, fat,
protein and somatic cell count in buffaloes. Pakistan Vet J, 26: 67-72.
Sharif A, T Ahmad, MQ Bilal, A Yousaf, G Muhmmad, SU Rehman and FM Pansota, 2007.
Estimation of milk lactose and somatic cells for the diagnosis of subclinical mastitis
in dairy buffaloes. Int J Agri Biol, 9: 267-270.
Sheikh MA, M Anzam and AR Shakoori, 1996. Observations on haemorrhagic septicemia in
Pakistan Livestock. J of Vete Med, Series B.43: 293-304.

136
Shilston AW, 1923. Haemorrhagic septicemia (pasteurellosis). In: Wooldridge, GH. (editor)
Encyclopedia of Veterinary Medicine, Surgery and Obstetrics. Vol 1 (Veterinary
Medicine). Henry Frowde and Hodder and Stoughton, Lodon. PP: 103-112.
Shivachandra SB, KN Viswas and AA Kumar, 2011. A review of hemorrhagic septicemia in
cattle and buffalo. Anim Hlth Res Rev, 12: 67-82.
Silva ID, and KFST Silva, 1994. Total and differential cell counts in buffalo milk. Buffalo
Journal 2: 133-137.
Singh KP, D Chandra, R Rathore, AB Pandey, AG Telang, 2007. Investigation of outbraks of
Pasteurellosis in cattle and buffaloes. Indian J Vet Pathol, 31:40-44.
Singh M, and RS Ludri, 2000. Somatic cell count in Murrah buffaloes during different stages
of lactation, parity and season. Asian-Aust. J Anim Sci, 14:189-192.
Singh S, VP Singh, PS Cheema M Sandey, R Ranjan, SK Gupta, and B Sharma,2011.
Immune response to dna vaccine expressing transferrin binding protein a gene of
Pasteurella multocida.Braz J Miceobiol.42 (2): 750-760.
Sivakumar T, A Thennarasu, and JSI Rajkumar.2012. Effect of season on the incidence of
infectious diseases of bovine in Tamilnadu. Elixir Meteorol 47: 8874-8875.
Soomro SA, KB Mirbahar, MA Memon, and MI Memon, 1997. Prevalence of clinical and
sub-clinical mastitis in buffalo at Hyderabad, Sindh. Pak J Agri Agril Engg Vet Sci,
13: 28-30.
Sordillo LM, K Shafer-Weaver and D DeRosa, 1997. Immunobiology of the mammary
gland. J Dairy Sci, 80: 1851-1865.
Sotoodehnia A, G Moazeni, S Ataei and B Omidi, 2005.Study on immunity of an
experimental oil adjuvant haemorrhagic septicaemia vaccine in cattle Arch Razi Inst,
59: 95-101.
Soulebot JP, VJ Polyya, M Rweyemomu and D Sylla, 1997. Quality Assurance and Good
Manufacturing Practice. FAO of UN, Rome, pp: 297-15.
Srinivasan VA, GS Reddy, KA Rao and U Kihm , 2001. Serological response of bovines to
combined vaccine containing foot and mouth disease virus, rabies virus, Pasteurella
multocida and Clostridium chauvoei antigens. Vet Arhiv, 71: 37-45.

137
Srivastava NC, PC Harbola and SS Khera, 1975. Preliminary observations on combined
vaccination against haemorrhagic septicaemia and black quarter. Indian Vete J, 53:
168–172.
Srivastava SK. 1998.Immunogenicity of Pasteurella multocida grown in iron restricted
medium. J Appl Anim Res, 13: 137-144.
Steel RGD, JH Torrie and D Dickey, 1997. Principles and Procedures of Statistics: A
Biometrical Approach; 3rd Ed. McGraw Hill Book Co Inc, New York.
Stewart PS, 1996. Theoretical aspects of antibiotic diffusion into microbial biofilms.
Antimicrob Agents Chemother, 40: 2517-2522.
Sultana S, 2012. Studies on the comparative immunogenicity of duck cholera vaccine
produced by bau and lri in jinding ducks. Master of Science (M. S.) Thesis,
Department of Microbiology and Hygiene Bangladesh Agricultural University
Mymensingh, Bangladesh Agricultural University, Bangladesh.
Tabatabaei M, GRM Jula, AR Jabbari and M Esmailzadeh. 2007. Vaccine efficacy in cattle
against hemorrhagic septicemia with live attenuated aroA mutant of Pasteurella
multocida B: 2 strain. J Cell Anim Biol, 1: 62-65.
Tasneem k, B Zamir, S Ali, ZJ Gill and A Raza, 2009. Haemorrhagic septicaemia: A review.
Pak J Sci, 61: 10-13.
Tenover FC, MV Lancaster, and BC Hill, 1998. Characterization of staphylococci with
reduced susceptibilities to vancomycin and other glycopeptides. J Clin Microbiol, 36:
1020-1027.
The Center for Food Security and Public Health and Institute for International Cooperation in
Animal Biologics- an OIE Collaborating Center.2009. Hemorrhagic septicemia.
Accessed from web pp: 1-5.
Thorburn MA, 1990. General principles for the design of clinical trials with special reference
to mastitis therapy. Bull Intern Dairy Feder, 247: 40-51.
Tollersrud T, 2001. Staphylococcus aureus mastitis: Bacterial characteristics and host
immune responses. Ph.D Thesis, Natl Vet Instt Oslo, Norway. 2010 Mar doi:
10.1007/s11250-009-9443-3. Epub 2009 Sep 3.

138
Tollersrud T, L Zernichow, SR. Andersen, K Kenny and A Lund, 2001. Staphylococcus
aureus capsular polysaccharide type 5 conjugate and whole cell vaccines stimulate
antibody responses in cattle. Vaccine, 19: 3896-3903.
Tollersrud T, PE Norstebo, JP Engvik, SR Anderson LJ Reitan and A Lund, 2002. Antibody
responses in sheep vaccinated against Staphylococcus aureus mastitis: a comparison
of two experimental vaccines containing different adjuvants. Vet Res Commun,
26:587-600.
Townsend KM, AJ Frost, CW Lee, JM Papadimitriou and HJS Dawkins, 1998. Development
of PCR assays for species- and type-specific identification of Pasteurella multocida
isolates. J Clin. Microbiol, 36:1096–1100.
Trinidad P, SC Nickerson and TK Alley, 1990a. Prevalence of intramammary infection and
teat canal colonization in unbred and primigravid dairy heifers. J Dairy Sci.:73:107-
14.
Trinidad PSC. Nickerson and TK Alley, 1990b. Efficacy of intramammary treatment in
unbred and primigravid dairy heifers. J Am Vet Med Assoc 197:107-114.
Tuefel N, 1998. Village milk production in central Punjab (Pakistan).Buffalo Newsletter 10:
4-9.
Turkyilmaz S and S Eskiizmirliler, 2006. Detection of slime factor production and antibiotic
resistance in staphylococcus strains isolated from various animal clinical samples.
Turk J Vet Anim Sci, 30: 201-206.
Upton M, 2001. Trade in livestock and livestock products: International regulation and role
for economic development. Livestock information and policy branch AGAL, Food
and Agriculture Organization.
Van der Meer C, FMA Hofhuis and JMN Willers, 1997. Killed Listeria monocytogenes
vaccines becomes protective on addition of polyanions. Nature, 269: 594-595.
Vasudevan P, MK Nair, T Annamalai and KS Venkitanarayanan, 2003. Phenotypic and
genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for
biofilm formation. Vet Microbial, 92: 179-185.
Veken JW, NH, Shah, P Klaasen, B Oudega and FK DeGraaf. 1996. Binding of host iron-
binding proteins and expression of iron –regulated membrane proteins by different

139
serotypes of Pasteurella multocida causing haemorrhagic septicemia, Microb.
Pathog. 21: 59-64.
Verdi, RJ, 1988. Endogenous proteases bovine milk and the effect of thermal processing on
their activity. Ph.D.Thesis, Cornell University, Ithaca., New York, USA.
Verma R and TN, Jaiswal, 1997. Protection, humoral and cell- mediated immune responses
in calves immunized with multiple emulsion haemorrhagic septicaemia vaccine.
Vaccine, 15: 1254-1260.
Verma R and TN, Jaiswal, 1998. Haemorrhagic septicaemia vaccines. Vaccine. 16: 1184-
1192.
Watson DL and HI Davies, 1993. Influence of adjuvants on the immune response of sheep to
a novel Staphylococcus aureus vaccine. Vet Microbiol, 34: 139-153.
Watson DL, 1975. Cytophylic attachment of ovine IgG2 to autologous polymorphonuclear
leucocytes. Aust J Exp Biol Med Sci, 53:527-529.
Watson DL, 1987. Serolgical response of sheep to live and killed Staphylococcus aureus
vaccine .Vaccine, 5: 275-288.
Watson, DL, 1988. Vaccination against experimental staphylococcal mastitis in ewes.Res.
Vet Sci, 45: 16.
Wei BD, GR Carter,1978. Live streptomycin-dependent Pasteurella multocida vaccine for
the prevention of hemorrhagic septicemia. Am J Vet Res, 39: 1534-7.
Weinberg GA and PG Szilagyi, 2010. Vaccine epidemiology: Efficacy, effectiveness, and the
translational research roadmap. J Infect Dis, 201: 1607-1610.
Wooldridge GH, 1923. Encyclopedia of Veterinary Medicine, Surgery and Obstetrics Vol.II,
Veterinary Surgery and Obstetrics, 2nd
Ed. Henry Frowde & Hoddrer and Stoughton,
London.P:1559.
Yousaf A, G Muhammad, Sajjad ur Rahman, M Siddique and MZ Masood, 2009. Effect of
montanide adjuvanted Staphylococcus aureus bacterin-toxoid on prevalence and
incidence of mastitis in cows. Pak J Agri Sci, 4: 119-123
Zahur AB, U Farooq, M Hussain, SH Hashmi, and R Muneer, 2007. Respiratory syndrome: a
major threat to the livestock farmers and its economic impact. Pak Vet J, 27: 189-
193.

140
APPENDICES
Appendix 1: Composition of casein-sucrose-yeast (CSY) agar and
method of its preparation
Composition of Casein-Sucrose-Yeast (CSY) agar
Sr# Ingredients Ingredients (g/l)
1 Casein hydrolysate 3g
2 Sucrose 3g
3 Yeast extract 5g
4 Sodium chloride 5g
5 Anhydrous dipotassium hydrogen orthophosphate 3g
7 Distilled water 1 liter
8 Agar 15g
Method of preparation
The above given ingredients were dissolved in water. The pH was adjusted to 7.3–7.4, after
which 1.5% agar was added. The medium was autoclaved at 1 bar for 15 minutes. After
cooling to 45–50°C, 5% calf blood (antibody-free P. multocida) was added. Finally the
medium was dispensed into sterile petri plate. (0IE, 2012)
Appendix II: Preparation of blood agar medium and collection of
defibrinated blood
Blood from sheep was collected by adopting the procedure described by National Mastitis
Council Inc. (1990). That was
i. Juglar vein was cleared by clipping the hairs over the skin, swabbed with alcohol
followed by tincture iodine.
ii. Blood from the vein was collected in sterile disposable 50 ml syringe fitted with 18
gauge needle by adopting all aseptic precautions.

141
iii. Needle from syringe was removed after blood collection and poured gently into
sterilized flask of 250ml with glass beads previously sterilized by dry heat at 27oC for
40 minutes in hot air oven.
iv. As the blood pour into the flask, it was gently rotated.
v. To defibrinate the blood, flask was vigorously shaken for 10 minute after recapping.
Appendix III: Preparation of culture media and composition
A. Blood agar plates
Blood agar base 40g
Distilled water 1000 ml
Blood agar base manufactured by Difco Laboratories, Detroit, Michigan, USA was used in
this study. In order to tropicalize the medium 4 additional gram of agar technical was added.
Ingredient per litter
Ingredients Ingredients (g/l)
Beef heart infusion from 500.00 g
Bacto tryptose 10.00 g
Sodium chloride 5.00 g
Bacto agar 15.00 g
Additional agar 4.00 g
i. Following the instructions described by National Mastitis Council Inc. (1987)
medium was dissolved, sterilized by autoclaving and cooled to about 500C before
adding 5 percent defibrinated blood
ii. Medium poured into the plates, cooled and incubated at 370C overnight to check the
sterility. In order to preserve the moisture in the plates they stored in the refrigerator
in polythene bags. All plates were used within the period of two weeks.
B. Staph-110 agar plates
Staph-110 agar powder 40g
Distilled water 1000ml

142
i. Staph-110 agar was manufactured by Oxoid Ltd, Basingstoke; Hampshire England
was used in this study. In order to tropicalize the medium 4 additional gram of agar
technical was added.
Ingredients (g/ liter)
Ingredients g/ liter
Yeast extract 2.5
Tryptone 10.0
Lactose 2.0
Mannitol 10.0
Sodium chloride 75.0
Di-potassium hydrogen phosphate 5.0
Gelatin 30.0
Agar 15.0
Additional agar 4.0
ii. Following the instructions described by National Mastitis Council Inc. (1987)
medium was dissolved, sterilized by autoclaving and cooled to about 50oC.
iii. Medium poured into the plates, cooled and incubated at 37oC overnight to check the
sterility. In order to preserve the moisture in the plates they stored in the refrigerator
in polythene bags. All plates were used within the period of two weeks.
C.MacConkey Agar Plates
MacConkey Agar Powder 52g
Distilled water 1000ml
i. MacConkey agar was manufactured by Oxoid Ltd, Basingstoke, Hampshire England
was used in this study. In order to tropicalize the medium 4 additional gram of agar
technical was added.

143
Ingredients (g/ liter)
Peptone 20.0
Lactose 10.0
Bile salts 5.0
Sodium chloride 5.0
Neutral red 0.075
Agar 12.0
Additional agar 4.0
ii. Following the instructions described by National Mastitis Council Inc. (1987)
medium was dissolved, sterilized by autoclaving and cooled to about 50oC.
iii. Medium poured into the plates, cooled and incubated at 37oC overnight to check the
sterility. In order to preserve the moisture in the plates they stored in the refrigerator
in polythene bags. All plates were used within the period of two weeks.
D. Nutrient Broth
i) Nutrient broth powder was manufactured by Merck, Germany.
Nutrient broth powder 8 grams
Distilled water 1000 ml
Ingredients (g/ liter)
Peptone from meat 5
Meat extracts 3
ii) Following the instructions described by National Mastitis Council Inc., (1987) nutrient
broth was dissolved in distilled water, then sterilized by autoclave and cooled to 500C.
E. Brain Heart Infusion Broth (BHI)
Brain heart infusion broth powder 37g
Distilled water 1000 ml
i) Brain heart infusion broth powder was manufactured by Oxoid Ltd, Basingstoke,
Hampshire, England.
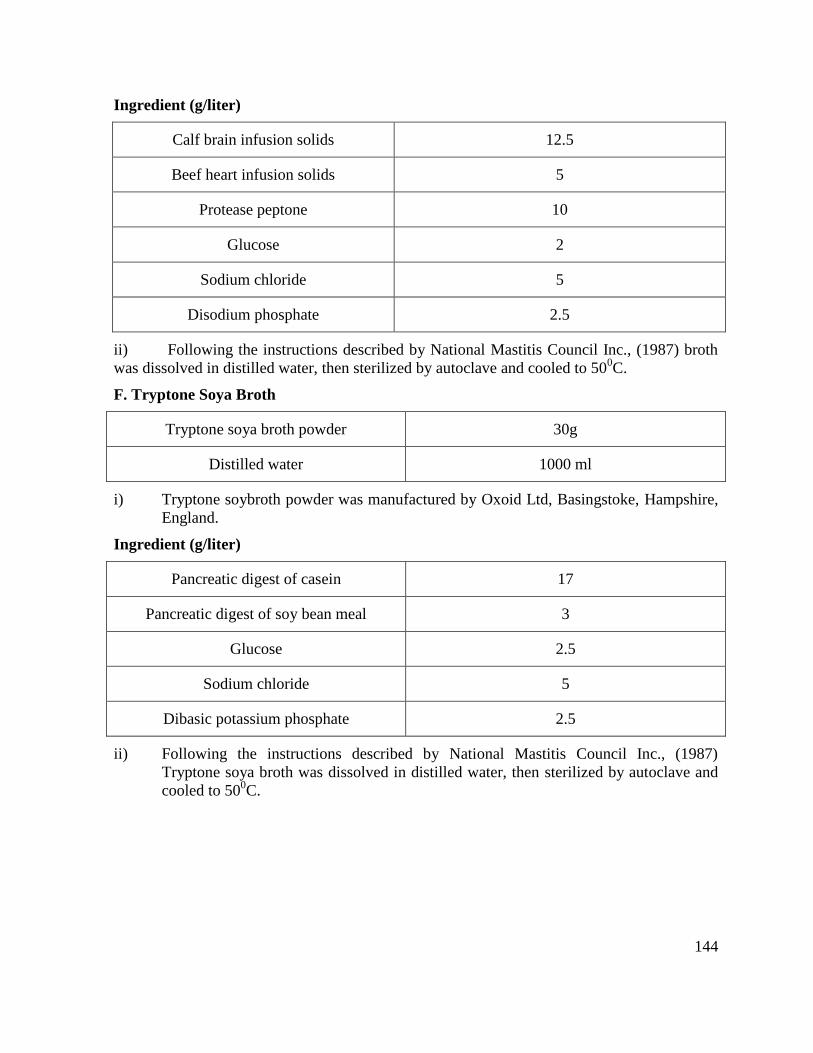
144
Ingredient (g/liter)
Calf brain infusion solids 12.5
Beef heart infusion solids 5
Protease peptone 10
Glucose 2
Sodium chloride 5
Disodium phosphate 2.5
ii) Following the instructions described by National Mastitis Council Inc., (1987) broth
was dissolved in distilled water, then sterilized by autoclave and cooled to 500C.
F. Tryptone Soya Broth
Tryptone soya broth powder 30g
Distilled water 1000 ml
i) Tryptone soybroth powder was manufactured by Oxoid Ltd, Basingstoke, Hampshire,
England.
Ingredient (g/liter)
Pancreatic digest of casein 17
Pancreatic digest of soy bean meal 3
Glucose 2.5
Sodium chloride 5
Dibasic potassium phosphate 2.5
ii) Following the instructions described by National Mastitis Council Inc., (1987)
Tryptone soya broth was dissolved in distilled water, then sterilized by autoclave and
cooled to 500C.

145
Appendix IV: Collection, processing and preparation of rabbit plasma
for tube coagulase test to differentiate coagulase positive
and coagulase negative Staphylococci
A tube coagulase test was used to differentiate coagulase positive and coagulase
negative Staphylococci. Tube coagulase positive test at 4 hours confirmed the isolates as
those of S. aureus.
Material required
Rabbit plasma (diluted 1:1 with normal saline). Fresh overnight culture of Gram
positive, catalase positive coccus. Glass tubes
Collection of rabbit plasma
A rabbit of approximately 1.1 kg body weight yielded about 30-38ml of blood. Five percent
solution of sodium EDTA was used as an anticoagulant for the collection of plasma from
rabbit blood.
Preparation of rabbit before slaughtering
The rabbit was restrained in lateral recumbency. The neck of the rabbit was swabbed
with a cotton swab moistened with normal saline to remove the dirt and broken hair attaching
to the body of the animal.
Preparation of EDTA solution
Five percent EDTA solution was prepared at a concentration of 1 g in 20 ml distilled
water. One and a half ml of this solution was gradually added to the blood that gushed from
the neck of the rabbit on slaughtering.
Collection of blood & preparation of plasma
Rabbit was slaughtered with a sharp knife and blood was collected in a beaker placed
under the neck of rabbit. EDTA solution was poured along the blade of the knife for proper
mixing of it with the blood while shaking the underlying beaker gently. The blood from the
beaker was poured into centrifugation tubes which were than centrifuge at 2000-2500rpm for
25 minutes. The plasma was then aspirated with the help of sterilized glass pipette. The
plasma thus separated was diluted 1:1 with sterilized normal saline. Aliquots of plasma were
dispensed into sterile eppendorf tubes which were then stored at -20oC.

146
Test Procedure
1. Take rabbit plasma
2. Make 1:1 dilution of the plasma
3. Then 24 hours fresh growth of the isolate was inoculated
4. Tube was placed in incubator at 37oC for 4 hours
5. The plasma was changed into a coagulum after 4 hours in case of positive test and
there was no change in case of negative result.
Appendix V: Preparation of sterile milk whey (Watson and Watson, 1989)
To prepare one liter whey by rennin precipitation.
1) Rennin suspension was prepared by dissolving 0.5mg of rennin in 27ml of saline.
2) The suspension was then added to 1.5 liter defatted bovine milk and kept at 37ºC for
30 minutes
3) The samples were then centrifuged at 10,000Xg for 20 minutes.
4) Whey thus obtained was sterilized by passing first through 0.45 μm membrane filters
and then through 0.22 μm filter.
Appendix VI: CAMP (Christie, Atkins and Munch-Peterson)-Esculine test
(National Mastitis Council, Inc. USA, 1987)
Inoculate a CAMP-esculine plate by streaking a culture of beta-hemolysis producing
S. aureus in a straight line across the center. Picked suspect Streptococcal colonies and
streaked perpendicular to and within 2 to 3 mm of the S. aureus streak. Ten isolates may be
tested per plates, with five on a side incubate the plate inverted for 18-24 hours at 37ºC.
Interpretation: CAMP reaction –complete clearing in a half –circle fashion (arrow –head) in
the partial lysis zone of the S. aureus. Esculine hydrolysis –browning around organism
growth
Appendix VII: Somatic cell count
(Singh and Ludri, 2002; Shailja and Sing, 2002; Athar, 2007; Tanveer, 2009).
Somatic cell count was performed using the technique as modified by Athar (2007) from
Schalm et al. (1971)
1) 10 μl of milk samples were spread on slides having a marked area of 10 mm x 10 mm
using a micropipette, dried at temperature 30-40oC in an oven.

147
2) The slides were dipped in xylene for 1-2 minutes to remove fat globules and dried
subsequently
3) The slides were then stained with Newman Lamperts stain for 15 minutes, took out
and dried at room temperature.
4) The dried slides were rinsed with tap water to remove excess of stain from the smear
and dried again at room temperature
5) The poorly stained smears were further stained with blue aliquot of Diff Quick Stain
(Difco Labs., Michigan, USA) for 10-15 seconds rinsed with tap water and dried.
6) Counting of cells at 100X in 50 fields which were multiplied by microscopic factor
(MF) to get the number of cells per ml of milk
MF =d x b
20 x 100
d = diameter of the microscopic field
b = number of microscopic fields
The number was then multiplied by 1000 to calculate number of somatic cells/ml of milk.




















