
Methicillin‐resistant Staphylococcus aureus colonization ...
Upload
independentCategory
view
8download
0
RESEARCH ARTICLE
Genomic and proteomic characterization of
Staphylococcus aureus mastitis isolates of bovine origin
Carmen Wolf1, Harald Kusch1�, Stefan Monecke2, Dirk Albrecht1, Silva Holtfreter 3�, Christofvon Eiff 4�, Wolfram Petzl5, Pascal Rainard6, Barbara M. Broker3 and Susanne Engelmann1
1 Institute for Microbiology, University of Greifswald, Greifswald, Germany2 Institute of Medical Microbiology and Hygiene, Technical University of Dresden, Dresden, Germany3 Institute for Immunology and Transfusion Medicine, University of Greifswald, Greifswald, Germany4 Institute for Medical Microbiology, University Hospital M .unster, M .unster, Germany5 Clinic for Ruminants, Ludwig-Maximilians-University, Munich, Germany6 INRA, UR Infectiologie Animale et Sante Publique, Nouzilly, France
Received: November 1, 2010
Revised: March 11, 2011
Accepted: March 14, 2011
Staphylococcus aureus colonizes and infects humans as well as animals. In the present study,
17 S. aureus strains isolated from cows suffering from mastitis were characterized. The well-
established multilocus sequence typing (MLST) technique and a diagnostic microarray
covering 185 S. aureus virulence and resistance genes were used for genetic and epidemio-
logical analyses. Virulence gene expression studies were performed by analyzing the
extracellular protein pattern of each isolate on 2-D gels. By this way, a pronounced hetero-
geneity of the extracellular proteome between the bovine isolates has been observed
which was attributed to genome plasticity and variation of gene expression. Merely 12
proteins were expressed in at least 80% of the isolates, i.e. Atl, Aur, GlpQ, Hla, LtaS, Nuc,
PdhB, SAB0846, SAB2176, SAB0566, SspA, and SspB forming the core exoproteome. Fifteen
extracellular proteins were highly variably expressed and only present in less than 20% of the
isolates. This includes the serine proteases SplB, C, and F, and the superantigens SEC-bov,
SEL and TSST-1. Compared to human isolates we identified at least six proteins with
significantly different expression frequencies. While SAB0846 was expressed more frequently
in bovine isolates, LytM, EbpS, Spa, Geh, and LukL1 were seen less frequently in these
isolates.
Keywords:
Bovine mastitis / Exoproteome / Microbiology / Staphylococcus aureus
1 Introduction
Staphylococcus aureus is an important pathogenic bacterium
with a broad host range. It colonizes and infects humans as
well as farm and wild animals. In farm animals, S. aureus is
among the most common etiologic agents of bovine mastitis
and one of the pathogens most frequently isolated from raw
milk [1]. Mastitis is characterized by an inflammation of the
mammary gland that can be induced in all mammalians in
response to (intramammary) bacterial infections. As an
endemic disease it causes big financial/economic loss to the
dairy industry worldwide, mainly due to reduced milk
production and the need to discard contaminated milk [2, 3].
Abbreviations: CV, core-variable; MLST, multilocus sequence
typing; MSCRAMM, microbial surface components recognizing
adhesive matrix molecule; PBMC, peripheral blood mononuclear
cell; SAg, superantigen; ST, sequence type; TSB, tryptic soy
broth
�Current addresses: Harald Kusch, Institute for Microbiology and,
Genetics, University of Gottingen, Gottingen, Germany,
Silva Holtfreter, Department of Molecular Medicine and Pathology,
University of Auckland, Auckland, New Zealand,
Christof von Eiff, Pfizer Pharma, Berlin, Germany
Correspondence: Dr. Susanne Engelmann, Institut f .ur Mikro-
biologie, Ernst-Moritz-Arndt-Universit .at, F.-L.-Jahn-Str. 15,
D-17487 Greifswald, Germany
E-mail: [email protected]
Fax: 149-3834-864202
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2011, 11, 2491–2502 2491DOI 10.1002/pmic.201000698

In contrast to mastitis caused by environmental patho-
gens such as Escherichia coli, S. aureus mastitis is contagious.
Bacteria are transmitted mainly during the milking process
[4]. Most of these infections take a subclinical course with
mild symptoms or they persist unnoticed, often resulting in
a long-lasting chronic form, which is particularly difficult to
treat [5].
The severity and clinical outcome of S. aureus infections
strongly depends on host factors such as the immune status
of the infected host. In addition, the virulence potential of
S. aureus that is characterized by its virulence gene equip-
ment may contribute to the course of infection [6]. S. aureushas the capacity to produce a large variety of virulence
factors, such as toxins, cell-surface-associated adhesins and
secreted exoproteins. It has been proposed that any S. aureusgenotype possesses the potential to transform into a life-
threatening pathogen, although strains from some clonal
lineages are more virulent than others [7].
Numerous typing methods have been applied to investi-
gate the genetic heterogeneity of S. aureus and revealed the
clonal population structure of the species [8]. They enable
epidemiological investigations to ascertain the source
and chain of infection in disease outbreaks [9, 10].
Molecular epidemiological analysis and comparative geno-
mics of bovine S. aureus demonstrated that a small number
of clonal types with a broad geographical distribution
caused the majority of mastitis cases [11, 12] and that a
limited number of genes or gene combinations confer host
specificity [13].
The genome of S. aureus consists of conserved and variable
regions [14]. The ‘‘core genome’’ is present in more than 95%
of the S. aureus strains and comprises approximately 75% of
all genes that are mainly associated with housekeeping (or
growth) and survival functions. It is subdivided into the so-
called ‘‘core-stable’’ genome and the ‘‘core-variable’’ (CV)
genome. CV genes make up approximately 10% of the
genome and show substantial variation between different
S. aureus strains, but they are typically stable and transferred
vertically [15]. Some of the CV genes encode virulence factors
involved in pathogenesis, e.g. toxins, superantigens (SAgs),
exoenzymes, and regulatory elements.
The remaining 25% of S. aureus genes encode non-
essential functions, ranging from virulence, resistance and
miscellaneous metabolic functions. Most of these ‘‘variable
or accessory’’ genes are carried on mobile (or once mobile)
genetic elements (MGEs). These elements, including
bacteriophages, S. aureus pathogenicity islands (SaPI),
staphylococcal cassette chromosomes (SCC), plasmids, and
transposons, are likely to transfer horizontally among
staphylococci [14] and contribute to the considerable genetic
heterogeneity of S. aureus and consequently to its enhanced
niche adaptation and virulence [16].
Whole genome sequencing of the common bovine
S. aureus strain RF122 identified a set of genetic features
that distinguishes this successful bovine clone from the
numerous sequenced human S. aureus strains. It was
proposed that specialized bovine clones diverged from
common human-associated ancestors through acquisition
and loss of DNA [17]. Kozytska et al. showed some genes are
specific for bovine isolates, but none of them was present in
all bovine isolates [18].
In the present study, 17 S. aureus isolates were subjected
to comprehensive genetic and proteomic analysis to reveal
virulence factors characteristic for bovine mastitis. Genetic
variability was determined by multilocus sequence typing
(MLST) and DNA microarrays. In addition we performed
2-D protein gel analyses, since we have previously shown
that differences in protein expression greatly contribute to
the diversity of the species S. aureus beyond the variability
that is conferred at the gene level. Finally, we analyzed
RNAIII transcription and studied T-cell-stimulating activ-
ities of the strains using both human and bovine peripheral
blood mononuclear cells (PBMC).
2 Materials and methods
2.1 Bacterial strains
Sixteen S. aureus isolates were derived from cows suffering
from clinical or subclinical mastitis in different geographical
regions (Table 1). All strains were identified as S. aureus by
plating and detection of genes encoding coagulase (coa) and
thermonuclease (nuc). Antibiotic sensitivity was determined
by the automated Vitek 2 system (bioMerieux, N .urtingen,
Germany). The sequenced S. aureus mastitis strain RF122
was used as a reference for genomic and proteomic studies
(Table 1). We used S. aureus COL and RN6390 as reference
strains for RNAIII transcription analyses [19, 20]. Control
strains for detection of the agr-type included COL (agr-1),
N315 (agr-2), TY114 8 (agr-3), and A920210 (agr-4)
[19, 21–23].
2.2 MLST
Total DNA of S. aureus was isolated with the WizardTM
Genomic DNA purification kit (Promega, Madison, WI,
USA) according to the manufacturer’s instructions with
some modifications. Most importantly, the addition of
0.6 mg lysozyme and 0.1 mg lysostaphin per sample resulted
in more efficient cell wall degradation. Isolated chromoso-
mal DNA was resolved in distilled water and stored at
�201C.
The protocol published by Enright et al. [24] was used for
MLST analyses. The resulting DNA sequences were
submitted to the Internet database (www.mlst.net). For each
isolate an allelic profile was generated which results in a
particular sequence type (ST). To classify different STs into
clusters of clonal complexes, the eBURST (based on related
sequences) algorithm software was applied (http://saureus.
mlst.net/eburst/) [25].
2492 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

2.3 Microarray hybridization
The diagnostic DNA microarray (StaphyType; Alere Tech-
nologies, Jena, Germany) used for this study covers about
300 target sequences corresponding to 185 distinct genes
and their allelic variants. A complete list of targets, as well as
the principle of the assay and related procedures, have
previously been described in detail [10, 26].
Additionally, to distinguish between agr types I and IV in
some ambiguous cases we used a multiplex PCR approach
for agr-1 to -4 as described previously [22].
2.4 Split network tree construction
Adapted from Monecke et al. [26], a split network tree was used
to visualize similarities between hybridization patterns. The
results of all array hybridization experiments were arranged in
a matrix where the columns represent the target genes and the
rows represent the experiments. The hybridization results
were converted into ‘‘sequences’’ using ‘‘A’’ for positive, ‘‘T’’
for negative results and ‘‘C’’ for ambiguous. Afterwards, the
matrix was converted into a series of ‘‘sequences.’’ These were
used for tree construction using SPLITSTREE 4.1
(www.splitstree.org) software (parameters: NJ, convex hull).
2.5 Preparation of extracellular protein extracts
Bacteria were cultivated in tryptic soy broth (TSB) under
vigorous agitation at 371C to an optical density (OD540) of 10.
One hundred millilitres of culture were harvested by centri-
fugation (10 min, 41C, 10 000� g). Extracellular proteins were
precipitated from the supernatant with 10% w/v trichloro-
acetic acid (TCA) at 41C overnight. After centrifugation
(45 min, 41C, 10 000� g), the precipitates were washed
several times with 96% v/v ethanol and dried. Protein extracts
were resuspended in an appropriate volume of 8 M urea and
2 M thiourea. The protein concentration was determined by
using Roti-Nanoquant according to the manufacturer’s
instructions (Carl Roth, Karlsruhe, Germany).
2.6 Analytic and preparative 2-D-PAGE
Protein extracts (80 mg) were separated according to their pIon commercially available linear immobilized pH gradient
(IPG) strips in a pI range of 3–10 (GE-Healthcare, Uppsala,
Sweden). 2-D PAGE using the IPG technique was
performed as described previously [27–29]. Protein gels were
stained with KryptonTM Protein Stain (Thermo Scientific,
USA) according to the manufacturer’s instructions (http://
www.piercenet.com/files/1829kc9.pdf), scanned with the
Typhoon 9400 Scanner with a 532 nm laser light source.
Additionally, protein extracts (350 mg) were separated by
preparative 2-D gels as described above and the proteins
were stained with colloidal Coomassie brilliant blue (CBB)
G-250 [30] and scanned with the light scanner. For protein
identification by matrix-assisted laser desorption/ionization-
time of flight mass spectrometry (MALDI-TOF-MS),
proteins were cut from CBB-stained gels by using the spot
cutter Proteome Work (GE-Healthcare), digested and
analyzed as described previously [31, 32].
2.7 Transcriptional analyses
S. aureus cells were cultivated in TSB medium to an OD540
of 10 and total RNA was isolated using the acid–phenol
method with modifications as described previously [33, 34].
Table 1. Characteristics of the S. aureus mastitis isolates used in this study
S. aureus – strain Year of isolation Origin; reference Course of infection agr-type MLST CC
RF122 1993 Ireland [11]; GenBank: AJ938182) Clinical 2 151 705377 n/a W .urzburg, Germany n/a 1 115 97A 2006 Upper Bavaria, Germany Subclinical 2 1274 705B 2006 Upper Bavaria, Germany Subclinical 2 504 705C 2006 Upper Bavaria, Germany Subclinical 2 504 705D 2006 Upper Bavaria, Germany Subclinical 2 1275E 2006 Upper Bavaria, Germany Subclinical 2 479 479F 2006 Upper Bavaria, Germany Subclinical 2 504 7051027 2003 Hannover, Germany [64] Clinical 1 133 133D4-106.06 1961 Yonne, Burgundy, France [65] Subclinical 1 97 97D4-126.29 1977 Aveyron, southern France Probably subclinical 1 71 97D8-628.24 1992 Pays de Loire, western France Probably subclinical 1 389 20D8-644.15 1992 Savoie, Alps, France Probably subclinical 1 389 20D8-684.18 1993 Indre et Loire, Touraine, France Probably subclinical 2 1276D9-780.07 1997 Savoie, eastern France Probably subclinical 1 97 97D9-798.19 2004 Mayenne, Pays de Loir, France Clinical 1 352 97N305 1958 Ontario, Canada [66] Clinical 1 115 97
n/a, not available.
Proteomics 2011, 11, 2491–2502 2493
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Digoxygenin-labeled RNA probes for RNA III were prepared
by in vitro transcription with T7 RNA polymerase by using
PCR-generated fragments as templates. The PCR fragments
were generated by using chromosomal DNA of S. aureusCOL isolated with the chromosomal DNA isolation kit
(Promega) according to the manufacturer’s instructions and
the respective oligonucleotides as described by Ziebandt
et al. [32].
Northern blot analyses were carried out as previously
described [35]. The digoxygenin-labeled RNA marker I
(Roche) was used as a standard to calculate the sizes of the
transcripts.
2.8 T-cell proliferation assay
For preparation of S. aureus supernatants, S. aureus cultures
at OD540 of 10 were centrifuged (21 000� g, 5 min, 41C). Cell
debris was removed from the culture supernatants using
filters with a pore size of 2mm. The cell-free supernatants
were stored at �201C.
Human and bovine PBMC were isolated by density
centrifugation over Ficoll (Biocoll Separating Solution,
Biochrom, Berlin). Human blood was obtained from healthy
blood donors from the Department of Transfusion Medicine
in Greifswald. Bovine blood samples were taken from a
healthy cow. Human and bovine cells were cultured in
96-well flat-bottomed plates as described previously [36]. For
bovine cells Iscove’s medium instead of RPMI was used. The
cells were stimulated for proliferation and harvested accord-
ing to the protocol described by Holtfreter et al. [36]. Incor-
porated [3H]-thymidine was quantified. Two biological
replicates each in technical triplicates and with two different
blood donors were quantified. Standard errors of the means
(SEM) of technical triplicates and SEM of biological replicates
were below 20% except for values of below 1000 cpm.
3 Results
3.1 Clonal relationship of the bovine S. aureus
isolates
First, the 17 bovine S. aureus isolates were assessed by MLST
to ascertain their clonal relationship. They belonged to 12
different STs (Fig. 1). Three of them were new (ST1274,
ST1275, and ST1276) and were submitted to the MLST
database (http://www.mlst.net). The new STs resulted from
new combinations of known alleles (ST1275 and ST1276) or
from the presence of a new allele (tpi-157 in ST1274). The
isolates clustered into five clonal complexes with six isolates
assigned to CC97, five to CC705, two to CC20, and one
strain to CC479 and CC133, respectively. The remaining two
isolates are singletons.
Of the 16 isolates three belong to lineages of known
prevalence in cattle: ST133 (n 5 1), and ST97 (n 5 2)
[13, 37–41]. ST133 represents one of the most common
animal-associated MLST types, and it has been rarely found
in humans so far [41]. A further seven isolates were assigned
to STs closely related to common intercontinental bovine
mastitis lineages: ST115 (CC97), ST71 (CC97), ST352
(CC97), and ST504 (CC705 (formerly CC151)). The two
ST389 isolates (CC20) have so far only been described in
humans albeit at low frequency [42]. Nine isolates were
classified as belonging to agr-type I (CC97, CC133, and
CC20) and six isolates to agr-type II (CC 151, CC479, and the
three singletons) (Table 1).
3.2 Detection of virulence-associated genes using
a diagnostic DNA microarray
The prevalence of 185 genes was determined using a diag-
nostic DNA microarray [26], which covered regulatory genes,
genes encoding virulence factors and microbial surface
components recognizing adhesive matrix molecules
(MSCRAMMs), capsule type-specific genes, as well as
resistance determinants in the different staphylococcal
strains including RF122.
The results are presented in the Supporting Information
Table S1. Altogether, 43 of the virulence genes were
conserved among the bovine S. aureus isolates. These are the
hemolysin-encoding genes hla, hlb, and hld, the two genes hl(SACOL0921) and hlIII (SACOL2160) with homologies to
hemolysin-encoding genes, the protease-encoding genes
aur, sspA, sspB, sspP, splA, and splB, the ssl genes setB, setB2,
50.00
26.79
42.86
24.29
35.71
0.71
10.71
11.43
3.57
7.14
7.14
N305
377
D4-106.06
D9-780.07
D4-126.29
D9-798.19
D9-628.24
D8-644.15
E
D
1027
F
C
B
A
D8-684.18
7.14
7.14
14.29
14.29
14.29
35.71
7.14
1276
705
97
115
115
1274
504
504
504
133 133
1275
479
97
97
97
97
705
705
71
352
97
389
389 20
20
strain ST CC
97
97
RF12216.07
8.93
151 705
479
705
Figure 1. Dendogram of multilocus STs detected among
the bovine S. aureus isolates of this study. The tree was
computed by using concatenated nucleotide sequences of seven
housekeeping genes and was constructed using the tool
provided by the MLST database (http://saureus.mlst.net/sql/
uniquetree.asp?]). Bootstrap values are indicated above the
branches. Clonal complexes were calculated by using the
eBURST algorithm.
2494 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

setB3, setC, ssl1, ssl5, and ssl3, the leukocidin genes hlgA and
hlgB ( 5 lukF) and hlgC ( 5 lukS) and lukX/Y and other
virulence genes as coa, sbi, spa, nuc, hysA1, hysA2, isaB, and
isdA. Moreover, we found that all isolates harbored the
MSCRAMM genes clfA, clfB, ebh, ebpS, eno, fib, fnbA, map,
sdrC, and vwb. Genes encoding capsular polysaccharide
biosynthesis proteins of serotype 5 (capH5/J5/K5) and
serotype 8 (capH8/I8/J8/K8) were similarly distributed
among the strains (eight and seven strains, respectively).
Other virulence genes were more variable. In nine
isolates we detected SAg-encoding genes, the enterotoxin
gene cluster (egc containing seg, sei, sem, sen, seo, and seu)
being most frequent (eight isolates). Two strains addition-
ally contained the bovine variants of tst1 and sec, and sel,indicating that they harbor the pathogenicity island SaPIbov
which is specific for bovine isolates [43]. Moreover, an
enterotoxin-like gene ORF CM14 was detected in five
strains.
Genes encoding the bovine specific virulence genes lukMand lukF’-PV were detected in 50% of the strains.
As expected, specific human S. aureus virulence genes
such as those encoding the Pantone Valentine Leukocidin
(PVL) (lukS-PV1lukF-PV), the exfoliative toxins (eta, etb, and
etd) or the epidermal cell differentiation inhibitors (edinA-C)
were absent from the bovine S. aureus isolates (Supporting
Information Table S1). Also, genes located on hlb-integrat-
ing phages, sak, sea, chp, and scn were not detected [44].
Accordingly, hlb was not disrupted in all isolates.
Genes associated with antibiotic resistance in human
isolates were rarely detected in the bovine isolates
(Supporting Information Table S1). For example, genes
conferring resistance to b-lactam antibiotics (blaZ, blaI, and
blaR) were only found in strain D4-126.29, the vanB gene in
strain D9-780.07 and the fosB gene in strains D8-628.24 and
D8-644.15. Other genes involved in antibiotic resistance
present on the DNA microarray have not been detected in
any of the isolates.
By split network tree construction [26] we compared the
overall hybridization patterns of the DNA-arrays of the
different isolates (Fig. 2). Some isolates cluster together
indicating a very similar virulence and resistance gene
pattern (Fig. 2). In general, these isolates belong to the same
STs (D8-628.24 and D8-644.15) or to closely related STs
(N305, 377, and D4-106.06; D9-780.07 and D9-798.19; A, B,
C, and F, E and D) (Fig. 1).
3.3 The exoproteome of the different bovine
isolates
From previous studies we knew that many virulence factors
in S. aureus are regulated by the quorum sensing system agrand are induced under high ODs [32, 45]. An agr mutant was
impaired in virulence in different mouse models indicating
that the expression of at least some of these proteins was
relevant for infection [46–49]. To get a first impression of
virulence gene expression of different clinical S. aureusisolates, a detailed comparison of the virulence factor patterns
under highly standardized in vitro growth conditions was
required, preferably at high ODs. Since many virulence
factors are known to be secreted, we focused on the exopro-
teome of these isolates. Extracellular proteins of the 17 bovine
S. aureus isolates (including RF122) were prepared from the
supernatants of bacterial cell cultures grown in TSB medium
to OD540 of 10 and separated on 2-D gels.
Altogether 232 different proteins were identified by
MALDI-TOF MS/MS (Supporting Information Table S2).
Seventy-four of them possessed signal sequences specific for
the Sec-dependent translocation machinery. Using PSORTb
(http://www.psort.org/psortb), 44 of these proteins were
predicted to be extracellular and eight cell wall-associated.
The localization of a further 37 proteins has not been exactly
predicted: 18 contained a signal peptide and 19 did not.
Similar to other studies [32, 50], we identified possible
membrane-associated (six proteins) and a considerable
number of cytoplasmic proteins (139 proteins) in the
exoproteomes of the bovine mastitis isolates.
As expected from our genomic studies, the extracellular
protein patterns varied considerably between the different
isolates (Fig. 3). Interestingly, only one protein (LtaS) was
found in the exoproteome of all isolates (Supporting Infor-
mation Table S2). LtaS (formerly YfnI) is a polytopic
membrane protein, which is processed by signal peptidase I
(SspI) as was previously shown for the homologue of
N305, 377D4-106.06
D9-780.07D9-798.19
D4-126.29
D8-644.15D8-628.24
D8-684.18
1027
ED
FC B
ARF122
0.01
Figure 2. Split network tree based on hybridization results for
every single gene on the diagnostic DNA microarray. The tree
was generated by Splitstree version 4.1 (http://www.splitstree.
org/) and shows genetic relationships between different bovine
S. aureus isolates.
Proteomics 2011, 11, 2491–2502 2495
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

BA DC
FE N305 377
1027 D4-106.06 D4-126.29 D8-628.24
D8-644.15 D8-684.18 D9-780.07 D9-789.19
10
pI[pH]
10 3
MW[kDa]
150
B
ApH 10 pH 3
Figure 3. Extracellular proteomes
of different S. aureus isolates
from bovine mastitis infections.
Cells were grown in TSB medium
to the stationary phase. Proteins
of the culture supernatants were
collected by trichloroacetic acid
precipitation and separated by 2-
D protein gel analysis. Protein
spots were detected by staining
with KryptonTM Protein Stain
(Thermo Scientific). Labeled
proteins were identified by MS.
(A) 2-D protein map of secreted
proteins of S. aureus RF122.
Protein spots are labeled with
protein names as listed in the
Supporting Information Table S2
and colored according to their
localization as predicted by
PSortb: extracellular (red),
unknown with signal peptide
(black), cell wall associated
(green), membrane bound
(violet), cytoplasmatic (blue),
unknown (gray). Superscript ‘‘F’’
indicates protein fragments. (B)
2-D protein gels of extracellular
proteins of different mastitis
isolates. The respective isolate is
indicated in the upper left corner
of each gel.
2496 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

B. subtilis [51]. Depletion of ltaS impaired LTA synthesis
and thus caused defects in cell division in S. aureus [52].
Eleven proteins (i.e. Atl, Nuc, SAB0566, GlpQ, Hla, SAB2176,
SspA, SspB, Aur, SAB0846, and PdhB) were expressed
in at least 80% of the isolates, with eight of them being
predicted as secreted proteins (Supporting Information
Table S2). Together with LtaS they represent the core
exoproteome of the bovine S. aureus mastitis isolates.
According to PSORTb prediction, two proteins (SAB0566 and
SAB0846) are of unknown subcellular localization but
possess signal sequences, and only one cytoplasmic protein
(PdhB) belongs to the core exoproteome. Strikingly, the
protein amounts of the eleven members of the core exopro-
teome varied significantly between isolates (Supporting
Information Fig. S1).
Fifteen extracellular proteins were almost unique, i.e.
they were found secreted by at most three of the 17 isolates.
Among them were well-characterized virulence factors such
as SAgs toxic shock syndrome toxin-1, SEC-bov and SEL,
serine proteases SplB, C, and F, FPRL1 inhibitory protein
FLIPr, HysA1and HysA2, Pbp2, Ssl11, and Ssl9 but also
proteins with unknown function (SAB0739, SAB1980c, and
SAB2421c) (Supporting Information Table S2).
As already mentioned, we found four groups of isolates
whose genetic composition was very similar: (i) D8-628.24
and D8-644.15, (ii) N305, 377, and D4-106.06, (iii) D9-780.07
and D9-798.19, and (iv) A, B, C, F, and RF122. Sometimes,
this genetic similarity was reflected by similar protein
expression patterns as exemplified by N305 and 377 of
group II. More frequently, however, even strains with very
similar virulence gene equipment, differed strongly in their
exoproteome patterns. This became impressively clear
when we compared expression of those proteins whose
genes were conserved among the isolates belonging
to the same group as exemplified for isolates A, B, C, and F.
Of the 82 genes that have been shown to be conserved in
these isolates, the gene products of 20 genes were identified
on the extracellular proteome of at least one of the four
isolates. Strikingly, only six proteins (Aur, Hlb, LukD,
LukF’-PV, LukM, and SspA) were secreted by all four
isolates while 14 proteins were found to be expressed
in two or less isolates. Isolates B and F shared the highest
number of proteins (16 proteins) followed by isolate A and C
(13 proteins).
3.4 Mitogenic activity of bovine S. aureus culture
supernatants
To analyze the mitogenic activity of the S. aureusisolates, supernatants from bacterial cultures at an OD at
540 nm (OD540) of 10 were incubated with human and
bovine PBMCs, respectively. T-cell activation was deter-
mined by [3H]-thymidine incorporation. Despite donor-
dependent variations, the titers of the supernatants with
maximal proliferation as well as the (relative) degree
of T-cell stimulation were reproducible between the experi-
ments. Data from one representative experiment are shown
in Fig. 4.
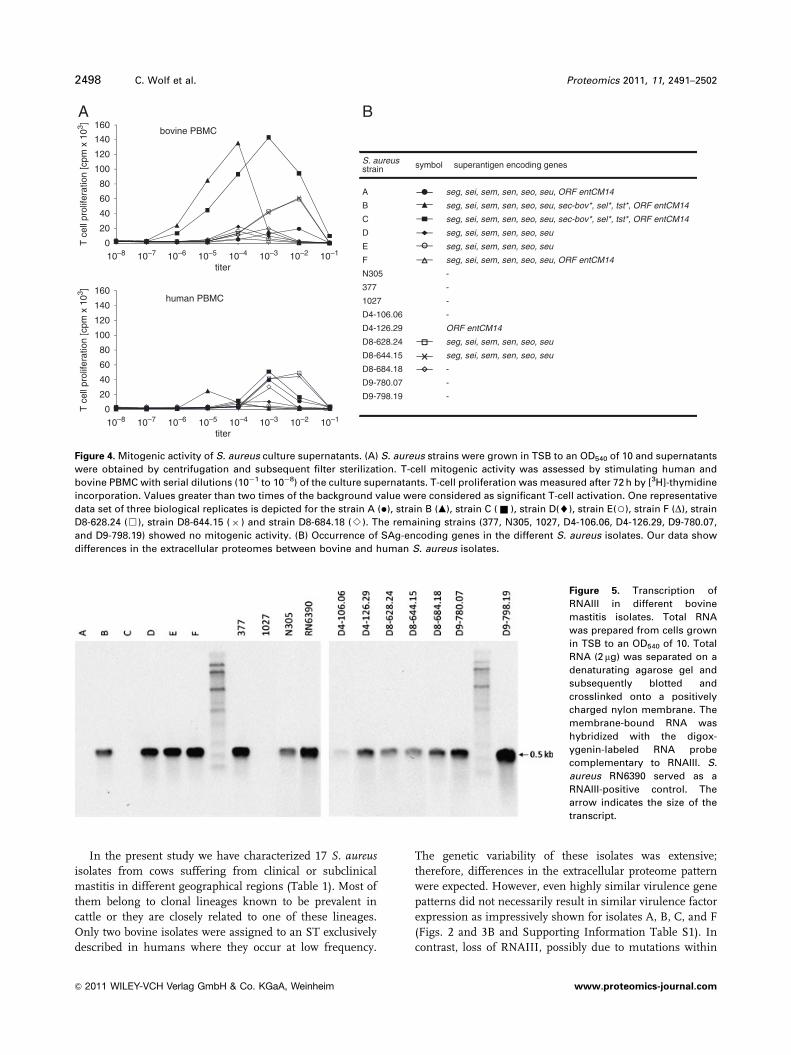
Accordingly, nine isolates (A, B, C, D, E, F, D8-628.24,
D8-644.15, and D8-684.18) showed mitogenic activity, which
correlated well with the presence of SAg genes (Fig. 4).
Isolate D8-684.18 was exceptional, since no SAg gene
was found but nevertheless the bacterial supernatant
was mitogenic. Point mutations interfering with oligonu-
cleotide binding on the DNA-array or an unknown
SAg could be the reason. In contrast, ORF entCM14 does
not appear to encode a functional SAg, since the super-
natant of strain D4-126.29 was devoid of mitogenic
activity. The isolates B and C, which contained the bovine
pathogenicity island, stimulated bovine PBMCs much
more strongly than human cells. The others, which
only encoded egc SAgs (sometimes together with ORF
entCM14) showed no significant differences in mitogenic
activity when incubated with blood cells from humans or
cattle (Fig. 4).
3.6 Transcription of RNAIII in bovine isolates
Variations in virulence gene expression may be caused
by differential activities of S. aureus regulators such
as RNAIII [32]. As shown previously, a deletion of
RNAIII affects the virulence factor pattern of S. aureusdrastically [32, 45]. RNAIII transcription was analyzed
by Northern blot experiments with RNA prepared from
the different isolates grown to an OD540 of 10 in TSB
medium. Accordingly, RNAIII was not detectable in four
isolates (A, C, 1027, and D4-106.06) (Fig. 5), which is
reflected by a low-level virulence factor expression in these
isolates.
4 Discussion
In the post-genomic era, the availability of the genome
sequences of several S. aureus strains including one bovine
isolate provides the basis for a better understanding of
staphylococcal infection biology. However, despite extensive
comparative genomic studies of bovine and human S. aureusisolates, the molecular basis of host specificity and virulence
of bovine S. aureus mastitis clones is still a matter of debate
[13, 17, 18, 38, 53, 54]. By dissecting the exoproteomes of 25
clinical human S. aureus isolates, we have recently demon-
strated that, within a single bacterial species, the exopro-
teome composition can vary to the extreme. The diversity
was induced not only by genome variations, but also by an
exceptionally high variability in gene expression [32]. These
findings suggest that the combination of genomics and
proteomics will result in a more comprehensive picture of
the virulence potential of a given bacterial strain. The same
may be true for host specialization [32, 45, 55–57].
Proteomics 2011, 11, 2491–2502 2497
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
In the present study we have characterized 17 S. aureusisolates from cows suffering from clinical or subclinical
mastitis in different geographical regions (Table 1). Most of
them belong to clonal lineages known to be prevalent in
cattle or they are closely related to one of these lineages.
Only two bovine isolates were assigned to an ST exclusively
described in humans where they occur at low frequency.
The genetic variability of these isolates was extensive;
therefore, differences in the extracellular proteome pattern
were expected. However, even highly similar virulence gene
patterns did not necessarily result in similar virulence factor
expression as impressively shown for isolates A, B, C, and F
(Figs. 2 and 3B and Supporting Information Table S1). In
contrast, loss of RNAIII, possibly due to mutations within
Figure 5. Transcription of
RNAIII in different bovine
mastitis isolates. Total RNA
was prepared from cells grown
in TSB to an OD540 of 10. Total
RNA (2 mg) was separated on a
denaturating agarose gel and
subsequently blotted and
crosslinked onto a positively
charged nylon membrane. The
membrane-bound RNA was
hybridized with the digox-
ygenin-labeled RNA probe
complementary to RNAIII. S.
aureus RN6390 served as a
RNAIII-positive control. The
arrow indicates the size of the
transcript.
titer
0
20
40
60
80
100
120
140
160A B
titer
10–7 10–6 10–5 10–4 10–3 10–2 10–110–8
10–7 10–6 10–5 10–4 10–3 10–2 10–110–80
20
40
60
80
100
120
140
160
T c
ell p
rolif
erat
ion
[cpm
x 1
03 ]
seg, sei, sem, sen, seo, seu, ORF entCM14
seg, sei, sem, sen, seo, seu, sec-bov*, sel*, tst*, ORF entCM14
seg, sei, sem, sen, seo, seu, sec-bov*, sel*, tst*, ORF entCM14
seg, sei, sem, sen, seo, seu
seg, sei, sem, sen, seo, seu
seg, sei, sem, sen, seo, seu, ORF entCM14
-
-
-
-
ORF entCM14
seg, sei, sem, sen, seo, seu
seg, sei, sem, sen, seo, seu
-
-
-
A
B
C
D
E
F
N305
377
1027
D4-106.06
D4-126.29
D8-628.24
D8-644.15
D8-684.18
D9-780.07
D9-798.19
bovine PBMC
human PBMC
symbol
T c
ell p
rolif
erat
ion
[cpm
x 1
03 ]
superantigen encoding genesS. aureusstrain
Figure 4. Mitogenic activity of S. aureus culture supernatants. (A) S. aureus strains were grown in TSB to an OD540 of 10 and supernatants
were obtained by centrifugation and subsequent filter sterilization. T-cell mitogenic activity was assessed by stimulating human and
bovine PBMC with serial dilutions (10�1 to 10�8) of the culture supernatants. T-cell proliferation was measured after 72 h by [3H]-thymidine
incorporation. Values greater than two times of the background value were considered as significant T-cell activation. One representative
data set of three biological replicates is depicted for the strain A (�), strain B (m), strain C (& ), strain D(~), strain E(3), strain F (D), strain
D8-628.24 (&), strain D8-644.15 (� ) and strain D8-684.18 (B). The remaining strains (377, N305, 1027, D4-106.06, D4-126.29, D9-780.07,
and D9-798.19) showed no mitogenic activity. (B) Occurrence of SAg-encoding genes in the different S. aureus isolates. Our data show
differences in the extracellular proteomes between bovine and human S. aureus isolates.
2498 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

the agr locus, predicted low expression levels of late viru-
lence factors (A and C) (Figs. 3B and 5) that might be
associated with a specific virulence behavior. Consequently,
to understand virulence and host specialization of a given
S. aureus isolate, virulence gene expression has to be
considered besides genomic information.
With regard to specific differences in virulence gene
expression between human and bovine strains, our data
show a core exoproteome of the bovine isolates consisting of
12 proteins. These were found in the supernatant of at least
80% of the bovine isolates (Supporting Information Table
S2). The human S. aureus core exoproteome of 15 clonally
different isolates comprises 26 proteins and is therefore
significantly larger [32]. Since the number of proteins in the
supernatants of the bovine isolates was not different from
that of human isolates, the smaller core exoproteome indi-
cates a higher complexity and diversity of the total exopro-
teome of bovine isolates. Eight proteins were conserved
between bovine and human S. aureus isolates. These are
LtaS (YfnI), Nuc, GlpQ, SspA, Hla, Aur, SAB0566 (SA0570),
and SAB2176. The four proteins SspB, Atl, PdhB, and
SAB0846 (SA0841) were more frequent in bovine isolates.
While SspB, Atl, and PdhB were found in 67–74% of the
human isolates, SAB0846 was expressed by human isolates
with strikingly lower frequency (20%). SAB0846 codes for a
protein highly similar to Map proteins. Typical for the core
exoproteome of the human isolates were LytM, Geh, Spa,
and LukL1, which were found in less than 50% of the bovine
isolates, although the genes encoding LytM, Geh, and Spa
are conserved among bovine isolates.
The use of a diagnostic DNA microarray [26] facilitated a
comprehensive genomic profiling (Supporting Information
Table S1). In total, we identified 27 virulence genes that
belong to the core genome of the 17 bovine as well as the 15
human isolates (Supporting Information Table S1) [32]. In
the secretomes of these isolates, the gene products of 15 of
these genes were observed, most of them with almost the
same frequency in human and bovine isolates (Supporting
Information Table S2). The sole exceptions were EbpS and
Spa. Both proteins EbpS and Spa were frequent in the
secretomes of human isolates (72 and 88%) but rare in the
secretomes of bovine isolates (0 and 11.7%). There are at
least two explanations for this phenomenon: either the
respective genes represent pseudogenes in bovine isolates (i)
or they are repressed (ii).
RF122 is the only sequenced bovine isolate thus far. This
strain also lacks the ability to express protein A, EbpS, Geh,
and LytM. Notably, the spa gene of RF122 contains premature
stop codons and thus is a pseudogene. The same is true for
spa in other successful bovine isolates such as ET3 isolates
different from RF122 and non-ET3 isolates [17]. The lack of
protein A in the supernatant of bovine isolates observed in
the present study supports the idea that premature trunca-
tions of the spa gene might be very common in origin S.aureus strains isolated from cattle. In contrast the ebpS, geh,
and lytM genes are intact in RF122 but obviously not
expressed. This might be induced by mutations in the regu-
latory regions of these genes and/or varying activities of
regulatory molecules. A very detailed comparison of the
upstream regions of lytM and geh, however, revealed no
significant sequence differences between strain RF122 and
strain COL which that abundantly express both genes [55].
The present data clearly show that virulence gene expres-
sion studies add an important dimension to our under-
standing of host specificity of S. aureus isolates which is not
covered by genomic studies. This has now been shown for the
extracellular as well as surface-associated proteins. In view of
the pronounced specific differences in the expression of
surface-associated proteins such as Spa, LytM, and EbpS
between bovine and human isolates, new techniques are now
required to analyze surface-associated virulence factors in
more detail. The method applied in this work to investigate
the extracellular proteome does not efficiently cover surface-
associated proteins such as MSCRAMMs, SERAMs and
lipoproteins. MS-based approaches, which rely on the
separation of complex protein or peptide mixtures by liquid
chromatography or 1-D SDS gel electrophoresis, allow the
identification of proteins in complex protein mixtures and
circumvent the obstacle of separation of proteins by 2-D gels
[58]. With specific targeting of the surface-associated
proteome, an even more comprehensive picture of the
features determining host specificity can be expected.
Despite extensive efforts in functional characterization of
virulence factors in S. aureus, the overall understanding of
the pathogenesis of S. aureus infection is limited. The
functions of at least 60% of the putative secreted proteins, as
derived from the genome sequence, are completely
unknown. In the secretome of the 17 bovine S. aureusisolates, at least 25 of these proteins have been identified
indicating that they are expressed. Functional characteriza-
tion of these particular proteins will be a challenging task for
future studies.
Previous studies suggest that SAgs are important during
mastitis pathogenesis due to their immunmodulatory effects
[59, 60]. SAg-encoding genes were identified in ten of the
isolates with three of them encoding the bovine variants of
SEC and TSST. Only three strains (RF122, B, and C) produced
the SAgs SEC and TSST in amounts detectable on 2-D gels.
However, T cells proved to be a more sensitive read out since
all ten SAg-positive strains induced T-cell proliferation. To test
whether these SAgs induce T-cell proliferation in humans and
cows with comparable intensities, human and bovine PBMCs
were compared. Two strains (B and C) stimulated bovine cells
much more efficiently than human cells suggesting some
degree of species specificity in SAg action. This is in line with
the previous observation that three amino acid differences
between SEC variants from bovine and ovine origin result in a
host-dependent superantigenic activity [61]. To ascertain
whether the SEC and TSST variants of bovine origin are
indeed more active in bovine than in human T cells, detailed
analyses have to be performed with recombinant SAg variants.
The egc SAgs appeared to be comparably active in bovine or
Proteomics 2011, 11, 2491–2502 2499
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

human PBMCs. ORF entCM14 probably does not encode a
functional SAg.
In summary, our data show differences in the extracellular
proteomes between bovine and human S. aureus isolates.
While EbpS, Geh, LytM, Spa, and LukL1 were more
frequently expressed in human isolates, for SAB0846 the
opposite was true. Most probably, the varying extracellular
protein patterns observed under highly standardized in vitro
conditions reflect a similar degree of variability in vivo.
Comparative genome sequencing of ruminant S. aureusisolates belonging to CC133 provided first evidence that
mechanisms of host adaptation involve gene decay and
diversification of proteins important in host pathogen inter-
action [17, 62, 63]. We propose that variability in the regula-
tion of virulence gene expression (in particular the loss of
expression of defined virulence genes) is also linked to the
host specificity of S. aureus.
We are indebted to Anita Harang and Thomas Meier forexcellent technical assistance. We thank Elke Lange (Friedrich-Loffler Institute, Riems, Germany) for providing bovine bloodsamples. The study was supported by research grants from theDFG (FOR 585, GRK 840, and SFB-TRR34), BMBF (ERA-NET Pathogenomics Network: sncRNAomics project) and EU(StaphDynamics: LSHM-CT-2006-019064).
The authors have declared no conflict of interest.
5 References
[1] Barkema, H. W., Green, M. J., Bradley, A. J., Zadoks, R. N.,
The role of contagious disease in udder health. J. Dairy Sci.
2009, 92, 4717–4729.
[2] Miller, G. Y., Bartlett, P. C., Lance, S. E., Anderson, J., Heider,
L. E., Costs of clinical mastitis and mastitis prevention in
dairy herds. J. Am. Vet. Med. Assoc. 1993, 202, 1230–1236.
[3] Seegers, H., Fourichon, C., Beaudeau, F., Production effects
related to mastitis and mastitis economics in dairy cattle
herds. Vet. Res. 2003, 34, 475–491.
[4] Bradley, A., Bovine mastitis: an evolving disease. Vet.
J. 2002, 164, 116–128.
[5] Sutra, L., Poutrel, B., Virulence factors involved in the
pathogenesis of bovine intramammary infections due to
Staphylococcus aureus. J. Med. Microbiol. 1994, 40, 79–89.
[6] Peacock, S. J., Moore, C. E., Justice, A., Kantzanou, M. et al.,
Virulent combinations of adhesin and toxin genes in natural
populations of Staphylococcus aureus. Infect. Immun. 2002,
70, 4987–4996.
[7] Melles, D. C., Gorkink, R. F., Boelens, H. A., Snijders, S. V.
et al., Natural population dynamics and expansion of
pathogenic clones of Staphylococcus aureus. J. Clin. Invest.
2004, 114, 1732–1740.
[8] Feil, E. J., Cooper, J. E., Grundmann, H., Robinson, D. A.
et al., How clonal is Staphylococcus aureus? J. Bacteriol.
2003, 185, 3307–3316.
[9] Maiden, M. C., Bygraves, J. A., Feil, E., Morelli, G. et al.,
Multilocus sequence typing: a portable approach to the iden-
tification of clones within populations of pathogenic micro-
organisms. Proc. Natl. Acad. Sci. USA 1998, 95, 3140–3145.
[10] Monecke, S., Jatzwauk, L., Weber, S., Slickers, P., Ehricht,
R., DNA microarray-based genotyping of methicillin-resis-
tant Staphylococcus aureus strains from Eastern Saxony.
Clin. Microbiol. Infect. 2008, 14, 534–545.
[11] Fitzgerald, J. R., Meaney, W. J., Hartigan, P. J., Smyth, C. J.,
Kapur, V., Fine-structure molecular epidemiological analy-
sis of Staphylococcus aureus recovered from cows. Epide-
miol. Infect. 1997, 119, 261–269.
[12] Kapur, V., Sischo, W. M., Greer, R. S., Whittam, T. S.,
Musser, J. M., Molecular population genetic analysis of
Staphylococcus aureus recovered from cows. J. Clin.
Microbiol. 1995, 33, 376–380.
[13] Sung, J. M., Lloyd, D. H., Lindsay, J. A., Staphylococcus
aureus host specificity: comparative genomics of human
versus animal isolates by multi-strain microarray. Micro-
biology 2008, 154, 1949–1959.
[14] Lindsay, J. A., Holden, M. T., Staphylococcus aureus:
superbug, super genome? Trends Microbiol. 2004, 12, 378–385.
[15] Lindsay, J. A., Moore, C. E., Day, N. P., Peacock, S. J. et al.,
Microarrays reveal that each of the ten dominant lineages of
Staphylococcus aureus has a unique combination of
surface-associated and regulatory genes. J. Bacteriol. 2006,
188, 669–676.
[16] Hacker, J., Carniel, E., Ecological fitness, genomic islands
and bacterial pathogenicity. A Darwinian view of the
evolution of microbes. EMBO Rep. 2001, 2, 376–381.
[17] Herron-Olson, L., Fitzgerald, J. R., Musser, J. M., Kapur, V.,
Molecular correlates of host specialization in Staphylo-
coccus aureus. PLoS One 2007, 2, e1120.
[18] Kozytska, S., StauX, D., Pawlik, M. C., Hensen, S. et al.,
Identification of specific genes in Staphylococcus aureus
strains associated with bovine mastitis. Vet. Microbiol.
2010, 145, 360–365.
[19] Shafer, W. M., Iandolo, J. J., Genetics of staphylococcal
enterotoxin B in methicillin-resistant isolates of Staphylo-
coccus aureus. Infect. Immun. 1979, 25, 902–911.
[20] Novick, R., Properties of a cryptic high-frequency transdu-
cing phage in Staphylococcus aureus. Virology 1967, 33,
155–166.
[21] Kuroda, M., Ohta, T., Uchiyama, I., Baba, T. et al., Whole
genome sequencing of meticillin-resistant Staphylococcus
aureus. Lancet 2001, 357, 1225–1240.
[22] Holtfreter, S., Grumann, D., Schmudde, M., Nguyen, H. T.
et al., Clonal distribution of superantigen genes in clinical
Staphylococcus aureus isolates. J. Clin. Microbiol. 2007, 45,
2669–2680.
[23] Jarraud, S., Peyrat, M. A., Lim, A., Tristan, A. et al., egc, a
highly prevalent operon of enterotoxin gene, forms a
putative nursery of superantigens in Staphylococcus
aureus. J. Immunol. 2001, 166, 669–677.
[24] Enright, M. C., Day, N. P., Davies, C. E., Peacock, S. J., Spratt, B.
G., Multilocus sequence typing for characterization of methi-
2500 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

cillin-resistant and methicillin-susceptible clones of Staphylo-
coccus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015.
[25] Feil, E. J., Li, B. C., Aanensen, D. M., Hanage, W. P., Spratt, B.
G., eBURST: inferring patterns of evolutionary descent among
clusters of related bacterial genotypes from multilocus
sequence typing data. J. Bacteriol. 2004, 186, 1518–1530.
[26] Monecke, S., Slickers, P., Ehricht, R., Assignment of
Staphylococcus aureus isolates to clonal complexes based
on microarray analysis and pattern recognition. FEMS
Immunol. Med. Microbiol. 2008, 53, 237–251.
[27] B .uttner, K., Bernhardt, J., Scharf, C., Schmid, R. et al.,
A comprehensive two-dimensional map of cytosolic proteins
of Bacillus subtilis. Electrophoresis 2001, 22, 2908–2935.
[28] Bernhardt, J., B .uttner, K., Scharf, C., Hecker, M., Dual
channel imaging of two-dimensional electropherograms in
Bacillus subtilis. Electrophoresis 1999, 20, 2225–2240.
[29] Eymann, C., Dreisbach, A., Albrecht, D., Bernhardt, J. et al.,
A comprehensive proteome map of growing Bacillus
subtilis cells. Proteomics 2004, 4, 2849–2876.
[30] Candiano, G., Bruschi, M., Musante, L., Santucci, L. et al.,
Blue silver: a very sensitive colloidal Coomassie G-250
staining for proteome analysis. Electrophoresis 2004, 25,
1327–1333.
[31] Wolf, C., Hochgr .afe, F., Kusch,H., Albrecht, D. et al.,
Proteomic analysis of antioxidant strategies of Staphylo-
coccus aureus: diverse responses to different oxidants.
Proteomics 2008, 8, 3139–3153.
[32] Ziebandt, A. K., Kusch, H., Degner, M., Jaglitz, S. et al.,
Proteomics uncovers extreme heterogeneity in the Staphylo-
coccus aureus exoproteome due to genomic plasticity and
variant gene regulation. Proteomics 2010, 10, 1634–1644.
[33] Gertz, S., Engelmann, S., Schmid, R., Ohlsen, K. et al.,
Regulation of sigmaB-dependent transcription of sigB and
asp23 in two different Staphylococcus aureus strains. Mol.
Gen. Genet. 1999, 261, 558–566.
[34] Fuchs, S., Pane-Farre, J., Kohler, C., Hecker, M., Engelmann,
S., Anaerobic gene expression in Staphylococcus aureus. J.
Bacteriol. 2007, 189, 4275–4289.
[35] Wetzstein, M., Volker, U., Dedio, J., Lobau,S. et al., Cloning,
sequencing, and molecular analysis of the dnaK locus from
Bacillus subtilis. J. Bacteriol. 1992, 174, 3300–3310.
[36] Holtfreter, S., Bauer, K., Thomas, D., Feig, C. et al., egc-
encoded superantigens from Staphylococcus aureus are
neutralized by human sera much less efficiently than are
classical staphylococcal enterotoxins or toxic shock
syndrome toxin. Infect. Immun. 2004, 72, 4061–4071.
[37] Smith, E. M., Green, L. E., Medley, G. F., Bird, H. E. et al.,
Multilocus sequence typing of intercontinental bovine
Staphylococcus aureus isolates. J. Clin. Microbiol. 2005, 43,
4737–4743.
[38] Monecke, S., Kuhnert, P., Hotzel, H., Slickers, P., Ehricht, R.,
Microarray based study on virulence-associated genes and
resistance determinants of Staphylococcus aureus isolates
from cattle. Vet. Microbiol. 2007, 125, 128–140.
[39] Rabello, R. F., Moreira, B. M., Lopes, R. M., Teixeira, L. M.
et al., Multilocus sequence typing of Staphylococcus aureus
isolates recovered from cows with mastitis in Brazilian dairy
herds. J. Med. Microbiol. 2007, 56, 1505–1511.
[40] Jorgensen, H. J., Mork, T., Caugant, D. A., Kearns, A.,
Rorvik, L. M., Genetic variation among Staphylococcus
aureus strains from Norwegian bulk milk. Appl. Environ.
Microbiol. 2005, 71, 8352–8361.
[41] Smyth, D. S., Feil, E. J., Meaney, W. J., Hartigan, P. J. et al.,
Molecular genetic typing reveals further insights into the
diversity of animal-associated Staphylococcus aureus.
J. Med. Microbiol. 2009, 58, 1343–1353.
[42] Enright, M. C., Robinson, D. A., Randle, G., Feil, E. J. et al.,
The evolutionary history of methicillin-resistant Staphylo-
coccus aureus (MRSA). Proc. Natl. Acad. Sci. USA 2002, 99,
7687–7692.
[43] Fitzgerald, J. R., Monday, S. R., Foster, T. J., Bohach, G. A.
et al., Characterization of a putative pathogenicity island
from bovine Staphylococcus aureus encoding multiple
superantigens. J. Bacteriol. 2001, 183, 63–70.
[44] Coleman, D. C., Sullivan, D. J., Russell, R. J., Arbuthnott,
J. P. et al., Staphylococcus aureus bacteriophages
mediating the simultaneous lysogenic conversion of beta-
lysin, staphylokinase and enterotoxin A: molecular
mechanism of triple conversion. J. Gen. Microbiol. 1989,
135, 1679–1697.
[45] Ziebandt, A. K., Becher, D., Ohlsen, K., Hacker, J. et al., The
influence of agr and sigmaB in growth phase dependent
regulation of virulence factors in Staphylococcus aureus.
Proteomics 2004, 4, 3034–3047.
[46] Abdelnour, A., Arvidson, S., Bremell, T., Ryden, C.,
Tarkowski, A., The accessory gene regulator (agr) controls
Staphylococcus aureus virulence in a murine arthritis
model. Infect. Immun. 1993, 61, 3879–3885.
[47] Cheung, A. L., Eberhardt, K. J., Chung, E., Yeaman, M. R.
et al., Diminished virulence of a sar-/agr- mutant of
Staphylococcus aureus in the rabbit model of endocarditis.
J. Clin. Invest. 1994, 94, 1815–1822.
[48] Gillaspy, A. F., Hickmon, S. G., Skinner, R. A., Thomas, J. R.
et al., Role of the accessory gene regulator (agr) in patho-
genesis of staphylococcal osteomyelitis. Infect. Immun.
1995, 63, 3373–3380.
[49] Wright, J. S., 3rd, Jin, R., Novick, R. P., Transient interference
with staphylococcal quorum sensing blocks abscess forma-
tion. Proc. Natl. Acad. Sci. USA 2005, 102, 1691–1696.
[50] Tjalsma, H., Bolhuis, A., Jongbloed, J. D., Bron, S., van Dijl,
J. M., Signal peptide-dependent protein transport in Bacil-
lus subtilis: a genome-based survey of the secretome.
Microbiol. Mol. Biol. Rev. 2000, 64, 515–547.
[51] Antelmann, H., Tjalsma, H., Voigt, B., Ohlmeier, S. et al., A
proteomic view on genome-based signal peptide predic-
tions. Genome Res. 2001, 11, 1484–1502.
[52] Gr .undling, A., Schneewind, O., Synthesis of glycerol
phosphate lipoteichoic acid in Staphylococcus aureus. Proc.
Natl. Acad. Sci. USA 2007, 104, 8478–8483.
[53] Ben Zakour, N. L., Sturdevant, D. E., Even, S., Guinane,
C. M. et al., Genome-wide analysis of ruminant Staphylo-
coccus aureus reveals diversification of the core genome.
J. Bacteriol. 2008, 190, 6302–6317.
Proteomics 2011, 11, 2491–2502 2501
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

[54] Herron, L. L., Chakravarty, R., Dwan, C., Fitzgerald, J. R.
et al., Genome sequence survey identifies unique sequen-
ces and key virulence genes with unusual rates of amino
acid substitution in bovine Staphylococcus aureus. Infect.
Immun. 2002, 70, 3978–3981.
[55] Rogasch, K., R .uhmling, V., Pane-Farre, J., Hoper, D. et al.,
Influence of the two-component system SaeRS on global
gene expression in two different Staphylococcus aureus
strains. J. Bacteriol. 2006, 188, 7742–7758.
[56] Burlak, C., Hammer, C. H., Robinson, M. A., Whitney, A. R.
et al., Global analysis of community-associated methicillin-
resistant Staphylococcus aureus exoproteins reveals
molecules produced in vitro and during infection. Cell
Microbiol. 2007, 9, 1172–1190.
[57] Pocsfalvi, G., Cacace, G., Cuccurullo, M., Serluca, G. et al.,
Proteomic analysis of exoproteins expressed by enter-
otoxigenic Staphylococcus aureus strains. Proteomics 2008,
8, 2462–2476.
[58] Hempel, K., Pane-Farre, J., Otto, A., Sievers, S. et al.,
Quantitative cell surface proteome profiling for SigB-
dependent protein expression in the human pathogen
Staphylococcus aureus via biotinylation approach. J.
Proteome Res. 2010, 9, 1579–1590.
[59] Schuberth, H. J., Krueger, C., Zerbe, H., Bleckmann, E.,
Leibold, W., Characterization of leukocytotoxic and super-
antigen-like factors produced by Staphylococcus aureus
isolates from milk of cows with mastitis. Vet. Microbiol.
2001, 82, 187–199.
[60] Fueyo, J. M., Mendoza, M. C., Rodicio, M. R., Muniz, J. et al.,
Cytotoxin and pyrogenic toxin superantigen gene profiles
of Staphylococcus aureus associated with subclinical
mastitis in dairy cows and relationships with macrorestric-
tion genomic profiles. J. Clin. Microbiol. 2005, 43,
1278–1284.
[61] Dinges, M. M., Orwin, P. M., Schlievert, P. M., Exotoxins of
Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13,
16–34.
[62] Guinane, C. M., Ben Zakour, N. L., Tormo-Mas, M. A.,
Weinert, L. A. et al., Evolutionary genomics of Staphylo-
coccus aureus reveals insights into the origin and molecular
basis of ruminant host adaptation. Genome Biol. Evol. 2010,
2, 454–466.
[63] Viana, D., Blanco, J., Tormo-Mas, M. A., Selva, L. et al.,
Adaptation of Staphylococcus aureus to ruminant and
equine hosts involves SaPI-carried variants of von Will-
ebrand factor-binding protein. Mol. Microbiol. 2010, 77,
1583–1594.
[64] Petzl, W., Zerbe, H., G .unther, J., Yang, W. et al., Escherichia
coli, but not Staphylococcus aureus triggers an early
increased expression of factors contributing to the innate
immune defense in the udder of the cow. Vet. Res. 2008,
39, 18.
[65] Poutrel, B., Lerondelle, C., Induced staphylococcal infec-
tions in the bovine mammary gland. Influence of the month
of lactation and other factors related to the cow. Ann. Rech.
Vet. 1978, 9, 119–128.
[66] Prasad, L. B., Newbould, F. H., Inoculation of the bovine teat
duct with Staphylococcus aureus: the relationship of teat
duct length, milk yield and milking rate to development of
intramammary infection. Can. Vet. J. 1968, 9, 107–115.
2502 C. Wolf et al. Proteomics 2011, 11, 2491–2502
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Copyright © 2022 FDOKUMEN




















