
Pre-B-Cell Colony-Enhancing Factor Regulates NAD+Dependent Protein Deacetylase Activity and Promotes...
19
Pre–B-Cell Colony–Enhancing Factor Regulates NAD -Dependent Protein Deacetylase Activity and Promotes Vascular Smooth Muscle Cell Maturation Eric van der Veer, Zengxuan Nong, Caroline O’Neil, Brad Urquhart, David Freeman, J. Geoffrey Pickering Abstract—Conversion of vascular smooth muscle cells (SMCs) from a proliferative state to a nonproliferative, contractile state confers vasomotor function to developing and remodeling blood vessels. Using a maturation-competent human SMC line, we determined that this shift in phenotype was accompanied by upregulation of pre–B-cell colony– enhancing factor (PBEF), a protein proposed to be a cytokine. Knockdown of endogenous PBEF increased SMC apoptosis and reduced the capacity of synthetic SMCs to mature to a contractile state. In keeping with these findings, human SMCs transduced with the PBEF gene had enhanced survival, an elongated bipolar morphology, and increased levels of h-caldesmon, smoothelin-A, smoothelin-B, and metavinculin. Notwithstanding some prior reports, PBEF did not have attributes of a cytokine but instead imparted the cell with increased nicotinamide phosphoribosyltransferase activity. Intracellular nicotinamide adenine dinucleotide (NAD ) content was increased in PBEF-overexpressing SMCs and decreased in PBEF-knockdown SMCs. Furthermore, NAD -dependent protein deacetylase activity was found to be essential for SMC maturation and was increased by PBEF. Xenotransplantation of human SMCs into immunodeficient mice revealed an increased capacity for PBEF-overexpressing SMCs to mature and intimately invest nascent endothelial channels. This microvessel chimerism and maturation process was perturbed when SMC PBEF expression was lowered. These findings identify PBEF as a regulator of NAD -dependent reactions in SMCs, reactions that promote, among other potential processes, the acquisition of a mature SMC phenotype. (Circ Res. 2005;97:25-34.) Key Words: vascular smooth muscle pre–B-cell colony– enhancing factor maturation nicotinamide phosphoribosyltransferase deacetylation C onversion of smooth muscle cells (SMCs) from a pro- liferative, noncontractile state to a nonproliferative, con- tractile state is essential for conferring vasomotor function to developing arteries. 1,2 This shift toward a more mature SMC phenotype is also important for terminating SMC-mediated remodeling of diseased arteries. 2 Recently, we cloned adult vascular SMC lines that, in contrast to other human SMC preparations, could reversibly convert between a spread, proliferative, and synthetic state when cultured in the presence of serum to a highly elongated, nonproliferative state when serum was withdrawn. 3,4 In the latter state, the cells displayed decreased apoptosis, increased contractile protein expression, and the ability to contract in response to vasoactive agonists. This system, therefore, provided us with an opportunity to seek out factors that enabled a proliferative adult SMC to efficiently shift to a more quiescent state specialized to contract. Accordingly, we undertook differential display polymerase chain reaction (PCR) and high-density microarray analyses to identify genes that were differentially expressed as these human SMCs executed this key shift in phenotype. These surveys consistently identified pre–B cell colony– enhancing factor (PBEF) as being upregulated as SMCs shifted toward maturity. PBEF is a 52- to 55-kDa protein that has been proposed to be a cytokine. 5 Reported actions in this regard include synergizing with other cytokines to stimulate the maturation of pre–B cells, 5 stimulating the expression of inflammatory cytokines in epithelial cells, 6 prolonging neu- trophil survival, 7 and acting as an adipokine that lowers plasma glucose levels. 8 However, the contention that PBEF is a secreted cytokine is controversial. PBEF does not have a signal sequence for secretion, and the presence of PBEF in culture media has been suggested to be a consequence of activation-induced cell death, rather than secretion by either a classical or alternative pathway. 9,10 Moreover, PBEF has sequence similarity with bacterial NadV, a protein that Original received April 21, 2005; revision received May 31, 2005; accepted May 31, 2005. From the Robarts Research Institute (Vascular Biology Group) (E.v.d.V., Z.N., C.O., J.G.P.) and London Health Sciences Centre (B.U., D.F., J.G.P.), Departments of Medicine (Cardiology), Biochemistry, Medical Biophysics, Physiology, and Pharmacology, University of Western Ontario, London, Canada. Correspondence to Dr J. Geoffrey Pickering, London Health Sciences Centre, 339 Windermere Rd, London, Ontario N6A 5A5, Canada. E-mail [email protected] © 2005 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000173298.38808.27 25 by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from by guest on July 23, 2016 http://circres.ahajournals.org/ Downloaded from
Transcript of Pre-B-Cell Colony-Enhancing Factor Regulates NAD+Dependent Protein Deacetylase Activity and Promotes...

Pre–B-Cell Colony–Enhancing Factor RegulatesNAD�-Dependent Protein Deacetylase Activity and Promotes
Vascular Smooth Muscle Cell MaturationEric van der Veer, Zengxuan Nong, Caroline O’Neil, Brad Urquhart,
David Freeman, J. Geoffrey Pickering
Abstract—Conversion of vascular smooth muscle cells (SMCs) from a proliferative state to a nonproliferative, contractilestate confers vasomotor function to developing and remodeling blood vessels. Using a maturation-competent humanSMC line, we determined that this shift in phenotype was accompanied by upregulation of pre–B-cell colony–enhancingfactor (PBEF), a protein proposed to be a cytokine. Knockdown of endogenous PBEF increased SMC apoptosis andreduced the capacity of synthetic SMCs to mature to a contractile state. In keeping with these findings, human SMCstransduced with the PBEF gene had enhanced survival, an elongated bipolar morphology, and increased levels ofh-caldesmon, smoothelin-A, smoothelin-B, and metavinculin. Notwithstanding some prior reports, PBEF did not haveattributes of a cytokine but instead imparted the cell with increased nicotinamide phosphoribosyltransferase activity.Intracellular nicotinamide adenine dinucleotide (NAD�) content was increased in PBEF-overexpressing SMCs anddecreased in PBEF-knockdown SMCs. Furthermore, NAD�-dependent protein deacetylase activity was found to beessential for SMC maturation and was increased by PBEF. Xenotransplantation of human SMCs into immunodeficientmice revealed an increased capacity for PBEF-overexpressing SMCs to mature and intimately invest nascent endothelialchannels. This microvessel chimerism and maturation process was perturbed when SMC PBEF expression was lowered.These findings identify PBEF as a regulator of NAD�-dependent reactions in SMCs, reactions that promote, amongother potential processes, the acquisition of a mature SMC phenotype. (Circ Res. 2005;97:25-34.)
Key Words: vascular smooth muscle � pre–B-cell colony–enhancing factor � maturation� nicotinamide phosphoribosyltransferase � deacetylation
Conversion of smooth muscle cells (SMCs) from a pro-liferative, noncontractile state to a nonproliferative, con-
tractile state is essential for conferring vasomotor function todeveloping arteries.1,2 This shift toward a more mature SMCphenotype is also important for terminating SMC-mediatedremodeling of diseased arteries.2
Recently, we cloned adult vascular SMC lines that, incontrast to other human SMC preparations, could reversiblyconvert between a spread, proliferative, and synthetic statewhen cultured in the presence of serum to a highly elongated,nonproliferative state when serum was withdrawn.3,4 In thelatter state, the cells displayed decreased apoptosis, increasedcontractile protein expression, and the ability to contract inresponse to vasoactive agonists. This system, therefore,provided us with an opportunity to seek out factors thatenabled a proliferative adult SMC to efficiently shift to amore quiescent state specialized to contract. Accordingly, weundertook differential display polymerase chain reaction
(PCR) and high-density microarray analyses to identify genesthat were differentially expressed as these human SMCsexecuted this key shift in phenotype.
These surveys consistently identified pre–B cell colony–enhancing factor (PBEF) as being upregulated as SMCsshifted toward maturity. PBEF is a 52- to 55-kDa protein thathas been proposed to be a cytokine.5 Reported actions in thisregard include synergizing with other cytokines to stimulatethe maturation of pre–B cells,5 stimulating the expression ofinflammatory cytokines in epithelial cells,6 prolonging neu-trophil survival,7 and acting as an adipokine that lowersplasma glucose levels.8 However, the contention that PBEF isa secreted cytokine is controversial. PBEF does not have asignal sequence for secretion, and the presence of PBEF inculture media has been suggested to be a consequence ofactivation-induced cell death, rather than secretion by either aclassical or alternative pathway.9,10 Moreover, PBEF hassequence similarity with bacterial NadV, a protein that
Original received April 21, 2005; revision received May 31, 2005; accepted May 31, 2005.From the Robarts Research Institute (Vascular Biology Group) (E.v.d.V., Z.N., C.O., J.G.P.) and London Health Sciences Centre (B.U., D.F., J.G.P.),
Departments of Medicine (Cardiology), Biochemistry, Medical Biophysics, Physiology, and Pharmacology, University of Western Ontario, London,Canada.
Correspondence to Dr J. Geoffrey Pickering, London Health Sciences Centre, 339 Windermere Rd, London, Ontario N6A 5A5, Canada. [email protected]
© 2005 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000173298.38808.27
25 by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

confers bacteria with the ability to grow in nicotinamideadenine dinucleotide (NAD�)-deficient conditions.11 In keep-ing with this, Rongvaux et al have reported that mouse PBEFfunctions within the cell as a nicotinamide phosphoribosyl-transferase.9 This enzyme catalyzes the rate-limiting step inthe salvage pathway for NAD� biosynthesis, whereby nico-tinamide that is generated during NAD�-consuming reactionsis used to regenerate NAD�.12,13
We report here that human PBEF is not a cytokine but anintracellular regulator of NAD�-consuming reactions, includ-ing NAD�-dependent protein deacetylation. We further reportthat PBEF can promote the efficient acquisition of a stableand mature SMC phenotype and can increase the capacity ofSMCs to invest developing blood vessels.
Materials and MethodsAn expanded Materials and Methods can be found in the online datasupplement available at http://circres.ahajournals.org
Cell CultureExperiments were performed using the maturation-competent humanvascular SMC lines HITB5 and HITC6 generated from the humaninternal thoracic artery, as described previously.3,4
Overexpression of PBEF in Human SMCsA retroviral gene–delivery system was used to generate humanSMCs stably overexpressing PBEF, using methods described previ-ously.14 A PBEF-IRES-EGFP bicistronic fragment was cloned intothe retroviral expression vector pLNCX2 (Clontech). A secondretroviral expression construct was generated by inserting PBEFcDNA into pQCXIP-IRES-PURO (Clontech). Stable transductantswere selected and overexpression of PBEF was confirmed beforeeach experiment by Western blot analysis.
Western Blot AnalysisExpression of PBEF and SMC differentiation markers was assessedby Western blot analysis with chemiluminescence detection, asdescribed.14
ApoptosisSMC apoptosis was assessed by fluorescence in situ end-labeling ofDNA fragments, as described.3
Knockdown of PBEF by RNA InterferencePBEF knockdown was accomplished by infecting human SMCs withretrovirus-containing sequences encoding hairpin small interfering (siRNA)fragments. Three different targeting sequences were used, each consisting of19 nucleotides starting at nucleotides 147, 384, and 1278 of the PBEFcoding sequence (siRNA147, 5�-GGAAGGTGAAATATGAGGA-3�;siRNA384, 5�-ATGTTCTCTTCACGGTGGA-3�; siRNA1278, 5�-AGG-G CCGATTATCTTTACA-3�). Each sequence was separated by a 9-nu-cleotide noncomplementary spacer from the reverse complement of thesame 19-nucleotide sequence. Blast search confirmed that only the PBEFgene was targeted. Control inserts contained the gene-specific 19-nucleotidesequence and hairpin loop sequence but not the antisense component.
Real-Time Reverse Transcription–PCRProbe (5�-CAGTTGCTGATCCCA- 3�) and flanking primers (5�-primer, 5�-TGCAGCTATGTTGTAACCAATGG-3�; 3�-primer, 5�-ACAAAAGGTCGAAAAAGGGCC-3�) were used for TaqManreal-time reverse transcription (RT)-PCR for PBEF. GAPDH tran-script abundance was used as an endogenous RNA control to whichPBEF transcript abundance was normalized.
Nicotinamide Phosphoribosyltransferase ActivityCell lysates were reacted with 5 �mol/L [carbonyl-14C]nicotinamidein the presence of 0.5 mmol/L phosphoribosylpyrophosphate andlabeled, acetone-precipitable nicotinamide mononucleotide (NMN)quantified by scintillation counting.9
NAD� Analysis by High-PerformanceLiquid ChromatographyCellular nucleotides were extracted using perchloric acid, neutralizedwith KOH, and deproteinized cell lysate residues were analyzed byhigh-performance liquid chromatography (HPLC). NAD� retentiontime, determined from a NAD� standard, was 10 minutes.
Histone Deacetylase AssayHistone H4 peptide (Upstate) was labeled with [3H]acetyl coenzymeA (ICN) using PCAF histone acetylase. Cell lysates were incubatedwith [3H]acetyl histone and released [3H]acetate assessed by scintil-lation counting (SCID, Charles River Laboratories, Wilmington,Mass).
Human-Mouse Chimeric Angiogenesis In VivoHITC6 SMCs in M199 with 10% FBS and fibroblast growth factor(FGF)-2 were mixed with an equal volume of growth factor-reducedmatrigel (BD Discovery Labware) yielding final concentrations of5�105 cells/mL and 250 ng/mL FGF-2. The cell-matrigel suspension(500 �L) was subcutaneously injected into the abdomen of mice withsevere combined immunodeficiency syndrome (SCID). After 8 days,implants were harvested, fixed for 8 hour in Tris-buffered zinc,15 andparaffin-embedded tissues were sectioned at 5 �m. Sections weredouble-immunostained for CD31 (BD Biosciences) and enhancedgreen fluorescent protein (EGFP) (BD Clontech) using diaminoben-zidine (Vector Laboratories Ltd) and red alkaline phosphatasesubstrates, respectively, and counterstained with Harris hematoxylin.Apoptosis of human SMCs in matrigel was assessed by terminaldeoxynucleotidyltransferase-mediated dUTP nick end labeling(TUNEL) assay, after immunostaining for EGFP and incubatingsections with proteinase K (100 �g/mL).
Algorithms for quantifying EGFP-positive human SMCs,TUNEL-positive human SMCs, microvessels, and SMC investmentof microvessels are described in the online data supplement. HumanSMCs were deemed to be investing a microvessel if they assumedcontiguous and close apposition with an endothelial cell(s) andoriented their long axis along a contour of the vessel wall.
StatisticsValues are expressed as mean�SEM. Comparisons were made by ttest or analysis of variance with Bonferroni post hoc test. Signifi-cance of fold-changes was assessed by the Wilcoxon signed-ranktest. Statistical significance was set at P�0.05.
ResultsPBEF Is Upregulated During SMC MaturationThe generation of clonal populations of SMCs from thehuman internal thoracic artery, designated HITB5 andHITC6, that can convert from a proliferative to a contractileSMC3,4 enabled us to screen for endogenous factors involvedin this phenotype conversion. These screens suggested thatPBEF was upregulated as SMCs shifted to a mature, contrac-tile state. To verify this finding, HITB5 SMCs were analyzedfor PBEF mRNA and protein expression by Northern andWestern blot analysis, respectively. As shown in Figure 1, 6days after serum withdrawal, HITB5 SMCs converted fromspread cells variably oriented on the dish to highly elongatedcells that had crawled in a directed fashion into multilayeredcell aggregates. Concurrently, the 3 major transcripts ofPBEF (4.8, 2.9, and 2.2 kb) were markedly upregulated
26 Circulation Research July 8, 2005
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

(Figure 1B). Intracellular PBEF protein abundance alsoincreased in maturing HITB5 and HITC6 SMCs (3.9�0.5-fold after 6 days), as did the expression of the SMC contractileapparatus proteins h-caldesmon and smoothelin A (Figure 1C).PBEF was not detected in concentrated culture media at anystage of the maturation program (data not shown).
Overexpression of PBEF PromotesSMC MaturationTo determine whether PBEF was capable of promoting amore mature SMC phenotype, HITB5 SMCs were infectedwith retrovirus containing cDNA encoding PBEF and EGFP,or cDNA encoding EGFP alone, and stable transductantswere selected. As shown in Figure 2, SMCs overexpressingPBEF developed a bipolar morphology and were longer andthinner than vector-infected SMCs. This relative elongationpersisted as the SMCs further elongated in response to serumwithdrawal (Figure 2B) and was also observed with HITC6SMCs and primary SMCs expressing PBEF (data not shown).To determine whether the capacity for spatial organization ofSMCs was impacted by PBEF, cells were plated at higherdensities (12 000 cells/cm2) and subjected to serum with-drawal. As shown in Figure 2, by 3 days PBEF-overexpressing SMCs had aggregated and organized intodiscrete, multilayered ridges and nodules. In contrast, theextent of patterning in control SMCs was modest.
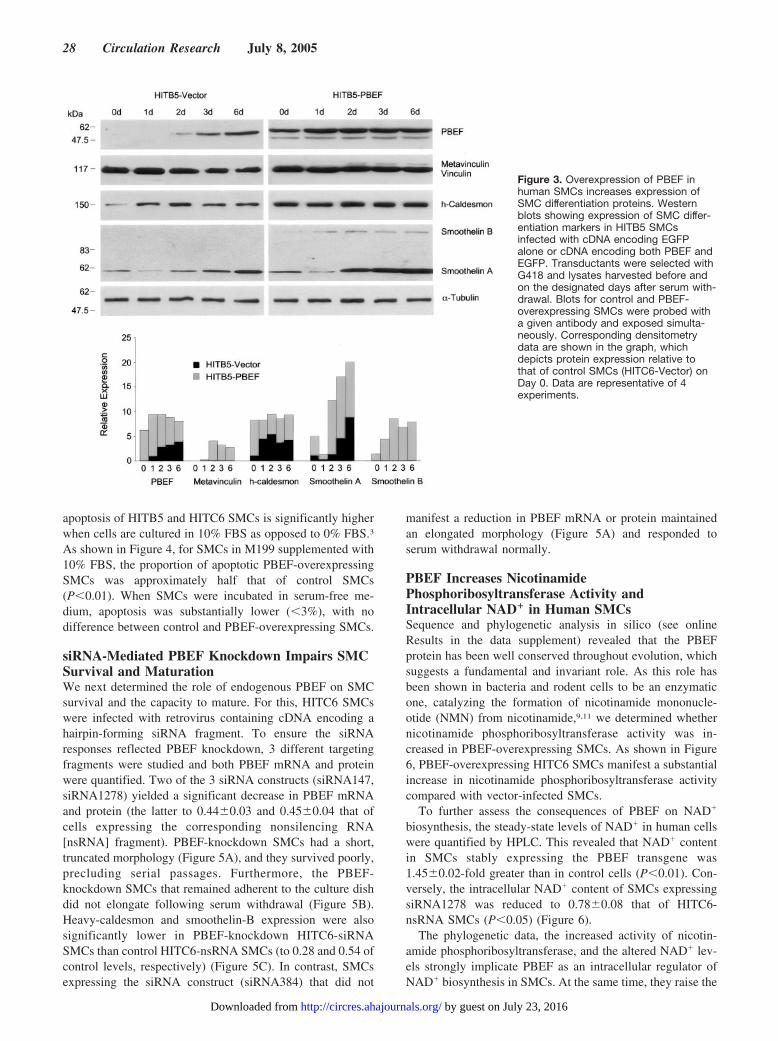
PBEF-overexpressing SMCs also displayed increased lev-els of h-caldesmon, smoothelin-A, and smoothelin-B com-pared with control SMCs assessed simultaneously (Figure 3).Withdrawal of serum from cultures of HITB5-PBEF SMCsled to further upregulation of smoothelin-A, and metavinculinexpression was induced, whereas the latter remained unde-tected in control SMCs. Thus, augmented expression ofPBEF shifted both the morphological and biochemical phe-notype of SMCs closer to that of mature, contractile SMCs.
Overexpression of PBEF Reduces SMC ApoptosisDifferentiation of SMCs has been associated with a decline inSMC apoptosis.3,16 To determine whether SMC survival wasaffected by PBEF, apoptosis was assessed using TUNEL. Wepreviously established that, in contrast to primary cells,
Figure 1. PBEF is upregulated during maturation of HITB5 andHITC6 human vascular SMCs. A, Hoffman-modulated contrastimages of HITB5 SMCs in M199 supplemented with 10% FBS(left) and 6 days after culturing SMCs in serum-free M199 (right).Bar, 50 �m. B, Northern blot showing upregulation of the 3major transcripts of PBEF in HITB5 SMCs following withdrawalof serum from cultures. C, Western blot of cell lysates harvestedfrom HITC6 SMCs before and after withdrawal of serum fromcultures.
Figure 2. Overexpression of PBEF in human SMCs induces cel-lular elongation and aggregation of cells into multilayered ridges.A, Hoffman-modulated contrast images of subconfluent (top) orpostconfluent (middle) HITB5 SMCs transduced with retroviruscontaining cDNA encoding EGFP alone (left) or PBEF and EGFP(right). Transgene expression is indicated by EGFP fluorescence(bottom). Bar, 50 �m. B, Quantification of length-width ratios ofcontrol and PBEF-overexpressing SMCs cultured in the pres-ence of serum and 3 days after serum withdrawal. The dimen-sions of 100 randomly selected SMCs from cultures at 40% to50% confluence were determined using Northern Eclipse soft-ware. *P�0.01 vs HITB5-EGFP SMCs.
van der Veer et al PBEF and Vascular Smooth Muscle 27
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from
apoptosis of HITB5 and HITC6 SMCs is significantly higherwhen cells are cultured in 10% FBS as opposed to 0% FBS.3
As shown in Figure 4, for SMCs in M199 supplemented with10% FBS, the proportion of apoptotic PBEF-overexpressingSMCs was approximately half that of control SMCs(P�0.01). When SMCs were incubated in serum-free me-dium, apoptosis was substantially lower (�3%), with nodifference between control and PBEF-overexpressing SMCs.
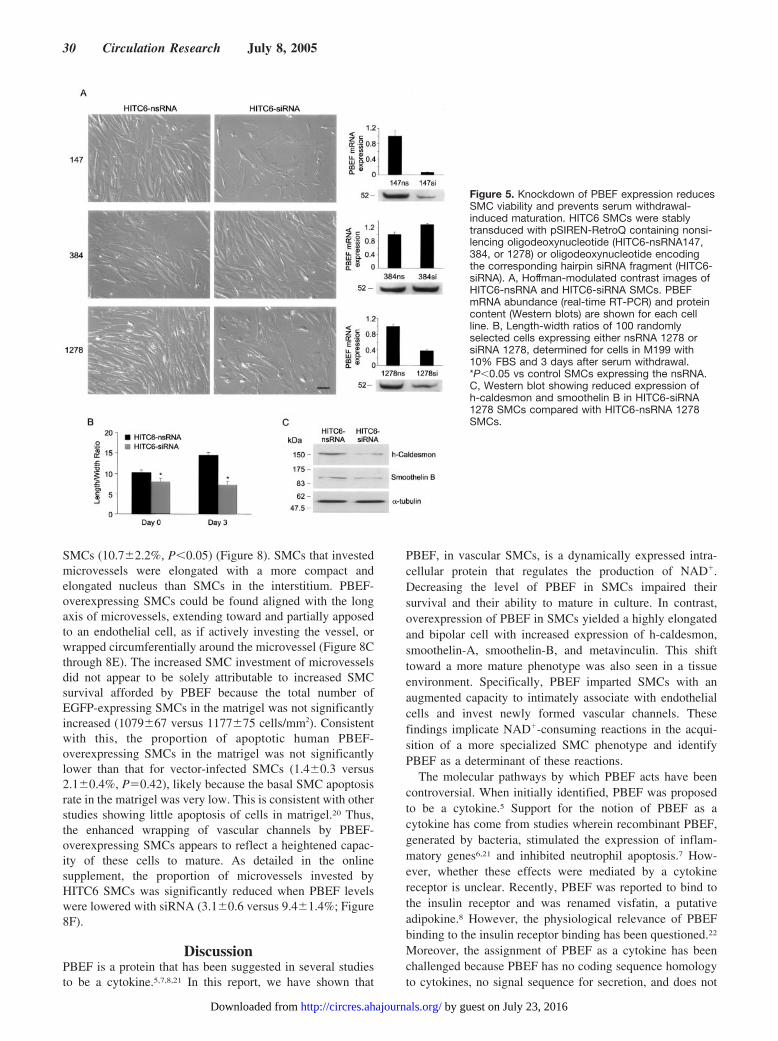
siRNA-Mediated PBEF Knockdown Impairs SMCSurvival and MaturationWe next determined the role of endogenous PBEF on SMCsurvival and the capacity to mature. For this, HITC6 SMCswere infected with retrovirus containing cDNA encoding ahairpin-forming siRNA fragment. To ensure the siRNAresponses reflected PBEF knockdown, 3 different targetingfragments were studied and both PBEF mRNA and proteinwere quantified. Two of the 3 siRNA constructs (siRNA147,siRNA1278) yielded a significant decrease in PBEF mRNAand protein (the latter to 0.44�0.03 and 0.45�0.04 that ofcells expressing the corresponding nonsilencing RNA[nsRNA] fragment). PBEF-knockdown SMCs had a short,truncated morphology (Figure 5A), and they survived poorly,precluding serial passages. Furthermore, the PBEF-knockdown SMCs that remained adherent to the culture dishdid not elongate following serum withdrawal (Figure 5B).Heavy-caldesmon and smoothelin-B expression were alsosignificantly lower in PBEF-knockdown HITC6-siRNASMCs than control HITC6-nsRNA SMCs (to 0.28 and 0.54 ofcontrol levels, respectively) (Figure 5C). In contrast, SMCsexpressing the siRNA construct (siRNA384) that did not
manifest a reduction in PBEF mRNA or protein maintainedan elongated morphology (Figure 5A) and responded toserum withdrawal normally.
PBEF Increases NicotinamidePhosphoribosyltransferase Activity andIntracellular NAD� in Human SMCsSequence and phylogenetic analysis in silico (see onlineResults in the data supplement) revealed that the PBEFprotein has been well conserved throughout evolution, whichsuggests a fundamental and invariant role. As this role hasbeen shown in bacteria and rodent cells to be an enzymaticone, catalyzing the formation of nicotinamide mononucle-otide (NMN) from nicotinamide,9,11 we determined whethernicotinamide phosphoribosyltransferase activity was in-creased in PBEF-overexpressing SMCs. As shown in Figure6, PBEF-overexpressing HITC6 SMCs manifest a substantialincrease in nicotinamide phosphoribosyltransferase activitycompared with vector-infected SMCs.
To further assess the consequences of PBEF on NAD�
biosynthesis, the steady-state levels of NAD� in human cellswere quantified by HPLC. This revealed that NAD� contentin SMCs stably expressing the PBEF transgene was1.45�0.02-fold greater than in control cells (P�0.01). Con-versely, the intracellular NAD� content of SMCs expressingsiRNA1278 was reduced to 0.78�0.08 that of HITC6-nsRNA SMCs (P�0.05) (Figure 6).
The phylogenetic data, the increased activity of nicotin-amide phosphoribosyltransferase, and the altered NAD� lev-els strongly implicate PBEF as an intracellular regulator ofNAD� biosynthesis in SMCs. At the same time, they raise the
Figure 3. Overexpression of PBEF inhuman SMCs increases expression ofSMC differentiation proteins. Westernblots showing expression of SMC differ-entiation markers in HITB5 SMCsinfected with cDNA encoding EGFPalone or cDNA encoding both PBEF andEGFP. Transductants were selected withG418 and lysates harvested before andon the designated days after serum with-drawal. Blots for control and PBEF-overexpressing SMCs were probed witha given antibody and exposed simulta-neously. Corresponding densitometrydata are shown in the graph, whichdepicts protein expression relative tothat of control SMCs (HITC6-Vector) onDay 0. Data are representative of 4experiments.
28 Circulation Research July 8, 2005
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

question as to whether the upregulation of PBEF in maturingSMCs might, in part, be a response to suboptimal in vitroconcentrations of NAD� precursors, potentially exacerbatedby removal of FBS. That this was not the case was demon-strated by strong upregulation of PBEF following serumwithdrawal even when the medium was supplemented withnicotinic acid and nicotinamide, at levels up to 5000-foldgreater than those found in M199 (Figure 6C). Thus, PBEFupregulation in maturing SMCs appears to be an inherentfeature of the HIT SMC maturation program, rather than aresponse to substrate limitation, and suggests that NAD� isconsumed during SMC maturation.
NAD�-Dependent Deacetylase Activity Is Requiredfor SMC Maturation and Is Increased inPBEF-Overexpressing SMCsRecently, a number of vital NAD�-dependent reactions havebeen identified that depend on the ability of the cell toregenerate NAD� from nicotinamide. These reactions includedeacetylation of certain histones and other proteins, reactionscritical to gene silencing and cell survival.17,18 To determinewhether SMC maturation was dependent on NAD�-dependent deacetylase activity, we studied the maturation of
HITC6 SMCs in the presence or absence of sirtinol, anoncompetitive inhibitor of NAD�-dependent proteindeacetylases that has no known effect on NAD�-independenthistone deacetylases (HDACs).19 As shown in Figure 7,10 �mol/L sirtinol had no discernable effect on the morphol-ogy or growth of HITC6 SMCs in the presence of 10% FBS.However, following serum withdrawal, SMCs incubated withsirtinol failed to elongate. This effect was reversible, andelongation was restored following sirtinol washout. Westernblot analysis revealed that PBEF expression increased 1.9-fold, 4 days after serum withdrawal, associated with upregu-lation of h-caldesmon and smoothelin-B expression. How-ever, in the presence of sirtinol, h-caldesmon, andsmoothelin-B levels were substantially lower than in di-methyl sulfoxide–treated control SMCs (Figure 7C).
To determine whether PBEF influenced HDAC activity inSMCs, lysates from control and PBEF-overexpressing SMCswere incubated with [3H]-acetylated histone H4 peptide andHDAC activity quantified. As shown in Figure 7D, totalHDAC activity was significantly greater in HITC6-PBEFSMCs than HITC6-Vector SMCs. Sirtinol significantly inhib-ited HDAC activity in lysates of PBEF-overexpressingSMCs, with a more modest inhibition of HDAC activity incontrol SMCs such that there was no longer a difference inHDAC activity between control and PBEF-overexpressingcells. We also examined the effect of 40 nmol/L trichostatinA, an inhibitor of class I and II HDACs, but not theNAD�-dependent (class III) HDACs. This substantially in-hibited HDAC activity in both control and PBEF-overexpressing SMCs. However, the residual trichostatinA–independent HDAC activity remained significantly greaterin PBEF-overexpressing SMCs than in control SMCs. Takentogether, the findings establish the following: (1) that vascu-lar SMC maturation requires NAD�-dependent proteindeacetylase activity; (2) that PBEF increases HDAC activityin SMCs; and (3) that most, if not all, of the PBEF-inducedHDAC activity in SMCs can be attributed to NAD�-dependent HDAC reactions.
Investment of Newly Formed Blood Vessels WithSMCs Is Dependent on PBEFTo assess the effect of PBEF on SMC phenotype in vivo, wedeveloped a human-mouse chimeric model of angiogenesis,whereby PBEF expression was manipulated exclusively inSMCs. Growth factor–reduced matrigel, mixed with FGF-2and either PBEF-overexpressing SMCs (HITC6-PBEF-EGFP) or vector-transduced SMCs (HITC6-EGFP), wasinjected subcutaneously into the abdominal regions of SCIDmice. After 8 days, mice were euthanized and zinc-fixedsections were studied histologically. Microvessel density,assessed by quantifying CD31-positive channels, was notsignificantly different in implants containing PBEF-overexpressing SMCs (287�18 versus 268�24 vessels/mm2). However, the proportion of microvessels that wereinvested by human SMCs, determined by double-immunolabeling for mouse endothelial cells and humanSMCs (anti-EGFP), was significantly higher in matrigelimplants containing PBEF-overexpressing SMCs(17.8�2.5%) than in implants loaded with vector-infected
Figure 4. PBEF reduces SMC apoptosis. Fluorescence imagesof control and PBEF-overexpressing SMCs stained withHoechst 33258 to identify nuclei (top) and for apoptotic nucleiby incubating with dUTP fluorescein (middle). Bottom, Graphthat depicts the averaged results from 3 separate experiments,with 900 to 1050 cells counted per experiment. *P�0.01 vs con-trol HITB5 SMCs.
van der Veer et al PBEF and Vascular Smooth Muscle 29
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from
SMCs (10.7�2.2%, P�0.05) (Figure 8). SMCs that investedmicrovessels were elongated with a more compact andelongated nucleus than SMCs in the interstitium. PBEF-overexpressing SMCs could be found aligned with the longaxis of microvessels, extending toward and partially apposedto an endothelial cell, as if actively investing the vessel, orwrapped circumferentially around the microvessel (Figure 8Cthrough 8E). The increased SMC investment of microvesselsdid not appear to be solely attributable to increased SMCsurvival afforded by PBEF because the total number ofEGFP-expressing SMCs in the matrigel was not significantlyincreased (1079�67 versus 1177�75 cells/mm2). Consistentwith this, the proportion of apoptotic human PBEF-overexpressing SMCs in the matrigel was not significantlylower than that for vector-infected SMCs (1.4�0.3 versus2.1�0.4%, P�0.42), likely because the basal SMC apoptosisrate in the matrigel was very low. This is consistent with otherstudies showing little apoptosis of cells in matrigel.20 Thus,the enhanced wrapping of vascular channels by PBEF-overexpressing SMCs appears to reflect a heightened capac-ity of these cells to mature. As detailed in the onlinesupplement, the proportion of microvessels invested byHITC6 SMCs was significantly reduced when PBEF levelswere lowered with siRNA (3.1�0.6 versus 9.4�1.4%; Figure8F).
DiscussionPBEF is a protein that has been suggested in several studiesto be a cytokine.5,7,8,21 In this report, we have shown that
PBEF, in vascular SMCs, is a dynamically expressed intra-cellular protein that regulates the production of NAD�.Decreasing the level of PBEF in SMCs impaired theirsurvival and their ability to mature in culture. In contrast,overexpression of PBEF in SMCs yielded a highly elongatedand bipolar cell with increased expression of h-caldesmon,smoothelin-A, smoothelin-B, and metavinculin. This shifttoward a more mature phenotype was also seen in a tissueenvironment. Specifically, PBEF imparted SMCs with anaugmented capacity to intimately associate with endothelialcells and invest newly formed vascular channels. Thesefindings implicate NAD�-consuming reactions in the acqui-sition of a more specialized SMC phenotype and identifyPBEF as a determinant of these reactions.
The molecular pathways by which PBEF acts have beencontroversial. When initially identified, PBEF was proposedto be a cytokine.5 Support for the notion of PBEF as acytokine has come from studies wherein recombinant PBEF,generated by bacteria, stimulated the expression of inflam-matory genes6,21 and inhibited neutrophil apoptosis.7 How-ever, whether these effects were mediated by a cytokinereceptor is unclear. Recently, PBEF was reported to bind tothe insulin receptor and was renamed visfatin, a putativeadipokine.8 However, the physiological relevance of PBEFbinding to the insulin receptor binding has been questioned.22
Moreover, the assignment of PBEF as a cytokine has beenchallenged because PBEF has no coding sequence homologyto cytokines, no signal sequence for secretion, and does not
Figure 5. Knockdown of PBEF expression reducesSMC viability and prevents serum withdrawal-induced maturation. HITC6 SMCs were stablytransduced with pSIREN-RetroQ containing nonsi-lencing oligodeoxynucleotide (HITC6-nsRNA147,384, or 1278) or oligodeoxynucleotide encodingthe corresponding hairpin siRNA fragment (HITC6-siRNA). A, Hoffman-modulated contrast images ofHITC6-nsRNA and HITC6-siRNA SMCs. PBEFmRNA abundance (real-time RT-PCR) and proteincontent (Western blots) are shown for each cellline. B, Length-width ratios of 100 randomlyselected cells expressing either nsRNA 1278 orsiRNA 1278, determined for cells in M199 with10% FBS and 3 days after serum withdrawal.*P�0.05 vs control SMCs expressing the nsRNA.C, Western blot showing reduced expression ofh-caldesmon and smoothelin B in HITC6-siRNA1278 SMCs compared with HITC6-nsRNA 1278SMCs.
30 Circulation Research July 8, 2005
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

appear to be secreted from cells by either a classical oralternative pathway.9,10 We were unable to detect PBEFprotein in the concentrated conditioned media from PBEF-overexpressing cell lines despite substantial upregulation ofintracellular protein.
The alternative proposal is that PBEF is a nicotinamidephosphoribosyltransferase, the rate-limiting, intracellular en-zyme for generating NAD� from nicotinamide.9 Sequenceand phylogenetic data are entirely consistent with the identityof human PBEF as a nicotinamide phosphoribosyltransferase.This assignment of PBEF is further strengthened by ourfinding that overexpression of PBEF in human SMCs sub-stantially increased nicotinamide phosphoribosyltransferaseactivity. Moreover, overexpression of PBEF increased thelevel of NAD� in SMCs, whereas a decrease in PBEFexpression was associated with reduced NAD� levels. Wepropose that the sequence data, the close evolutionary rela-tionship with prokaryotic orthologs, the fact that PBEFstimulates the conversion of nicotinamide to NMN andupregulates NAD�, and previous complementation and en-zyme analyses of mouse PBEF9 bring the weight of evidence
to the conclusion that human PBEF is a nicotinamide phos-phoribosyltransferase and not a cytokine.
The current study also reveals that the mechanism bywhich PBEF impacts SMC behavior likely relates to intra-cellular reactions that consume NAD�. In contrast to redoxreactions, which use, but do not consume NAD�, a number ofenzymatic reactions within the nucleus and cytoplasm con-sume NAD� as a cosubstrate. Degradation of NAD� in thesereactions liberates nicotinamide, from which NAD� can beregenerated via a 2-step salvage pathway. Nicotinamidephosphoribosyltransferase/PBEF catalyzes the conversion ofnicotinamide to nicotinamide mononucleotide. NMN is thenconverted to NAD� by NMN adenylyltransferase-1(Nmnat1).12,13 Both steps are necessary for sustaining NAD�-consuming reactions, including the deacetylation of histonesand nonhistone proteins.23,24 Interestingly, Nmnat1 has re-cently been found to prevent axonal degeneration in ex-planted mouse neurons,24 a finding concordant with theobserved anti-apoptotic actions of PBEF.
We found that SMC maturation was dependent on NAD�-dependent protein deacetylase activity. HITC6 SMCs failed
Figure 6. PBEF increases nicotinamidephosphoribosyltransferase activity andthe level of intracellular NAD�. A, Nicotin-amide phosphoribosyltransferase activityin control (no lysate) and in 100-ng ho-mogenates of rat liver or HITC6 SMCsstably transduced with either pQCXIP(HITC6-Vector) or pQCXIP-PBEF (HITC6-PBEF). Values depict production of 14C-NMN. *P�0.01 vs HITC6-Vector SMCs.B, Nucleotide extracts of HITC6-VectorSMCs or HITC6-PBEF SMCs (left) orHITC6-nsRNA1278 SMCs or HITC6-siRNA1278 SMCs (right) analyzed byHPLC. NAD� eluted from the column at�10 minutes (arrows). The smaller peakbefore the NAD� peak seen on thePBEF-overexpressing tracing was notconsistently observed in repeated experi-ments. Representative PBEF expressionfor control and PBEF-overexpressing orPBEF-knockdown cells is shown in theWestern blot insets. Quantitative data forNAD� levels for PBEF overexpression (5experiments) and PBEF knockdown (7experiments) are shown below therespective chromatograms. *P�0.01 vscontrol cells. C, Western blot showingupregulation of PBEF in HITC6 SMCs 6days following serum withdrawal, despitethe addition of nicotinic acid (NA) andnicotinamide (NAm).
van der Veer et al PBEF and Vascular Smooth Muscle 31
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

to elongate and failed to upregulate SMC contractile proteinsin the presence of the NAD�-dependent deacetylase inhibitorsirtinol. Conversely, overexpression of PBEF not only pro-moted SMC survival and the capacity to mature but stimu-lated NAD�-dependent HDAC activity. Interestingly, NAD�
salvage pathway genes in yeast and mammals have been
found to activate Sir2 (or the mammalian ortholog SIRT1), anNAD�-dependent deacetylase involved in transcriptional si-lencing and lifespan extension in yeast23,25,26 and survival ofmammalian cells.27 Further studies will be required to deter-mine the extent to which specific sirtuins, or other NAD�-dependent enzymes, regulate SMC function. However, the
Figure 7. NAD�-dependent proteindeacetylase activity is required for SMCmaturation and NAD�-dependent HDACactivity is augmented by PBEF. A,Hoffman-modulated contrast images ofHITC6 SMCs before and 4 days afterserum withdrawal in the presence orabsence of 10 �mol/L sirtinol. Sirtinolprevented serum withdrawal-inducedSMC elongation and alignment (FBS�
DMSO-Day 4 vs FBS� Sirtinol-Day 4) buthad no discernable effect on SMCs pro-liferating in medium with 10% FBS(FBS� Sirtinol-Day 4). B, Graph depictingthe effect of sirtinol on SMC elongation 4days following serum withdrawal.Sirtinol-treated SMCs failed to elongate,but 6 days after sirtinol-washout theelongation response was restored.*P�0.01 vs cells incubated with 10%FBS. C, Western blots showing reducedexpression of h-caldesmon and smooth-elin B in sirtinol-treated HITC6 SMCssubjected to 4 days of serum with-drawal. D, HDAC activity in humanSMCs. HITC6 SMCs were stably trans-duced with pQXCIP-PURO (HITC6-Vector) or pQXCIP-PBEF-PURO (HITC6-
PBEF) and HDAC activity determined using [3H]histone H4 peptide. Deacetylation reactions were performed in the presence of vehicle,50 �mol/L sirtinol, or 40 nmol/L trichostatin A. *P�0.01 vs vector-infected SMCs for the same assay condition. DMSO indicatesdimethyl sulfoxide.
Figure 8. Effect of PBEF expression onmicrovessel chimerism and SMC invest-ment in vivo. Control and PBEF-modifiedHITC6 SMCs expressing EGFP weremixed with matrigel and 250 ng/mLFGF-2, transplanted beneath the skin ofSCID mice, and harvested 8 days later.A through E, Sections of matrigelimplants double-immunolabeled for en-dothelial cells (anti-mouse CD31) andhuman SMCs (anti-GFP). Bound anti-CD31 antibody was identified using dia-minobenzidine chromogen (brown) andbound anti-EGFP antibody was visual-ized using red alkaline phosphatase sub-strate (red). Some newly formed bloodvessels are invested by human SMCsand this is especially prominent forimplants containing PBEF-overexpressing SMCs (arrows). Cthrough E, High-magnification imagesshowing EGFP-positive, PBEF-overexpressing human SMCs investingmouse microvessels. C, Xenotrans-planted SMC aligned parallel to an en-dothelial cell-lined vessel containing redblood cells and leukocytes. D, Apparentleading edge of an elongated humanSMC apposed to an endothelial cell,suggesting active vessel investment. E,
Corresponds to box in B, a human SMC circumferentially wrapped around a mouse microvessel. Bar, 50 �m. F, Graph showing theproportion of microvessels invested by EGFP-positive SMCs in gels containing PBEF-overexpressing or PBEF knockdown SMCs.*P�0.05 vs HITC6-nsRNA SMC-loaded matrigel, †P�0.05 vs HITC6-Vector SMC-loaded matrigel.
32 Circulation Research July 8, 2005
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

current data suggest that the capacity with which NAD� canbe regenerated (or nicotinamide cleared) is a key determinantof SMC behavior, including the acquisition of a quiescentphenotype.
The more mature attributes of PBEF-overexpressing SMCscould be functionally linked to their increased survival. Thatis, the apoptosis-resistant state imparted by PBEF could allowcultured SMCs to reassume more “physiological” character-istics, including a slender morphology and increased expres-sion of SMC-associated proteins. Although such a relation-ship is likely, the more mature characteristics of PBEF-overexpressing SMCs were also observed undercircumstances where there was no difference in apoptosisbetween control and PBEF-overexpressing cells. In serum-free media, both control and PBEF-overexpressing SMCclones displayed equally low apoptosis prevalence, yet therewere substantially higher levels of smoothelin-A,smoothelin-B, caldesmon, and metavinculin in the PBEF-overexpressing SMCs. Furthermore, the increased investmentof microvessels in vivo by PBEF-overexpressing SMCscompared with control SMCs occurred under conditionswhere SMC apoptosis was low and not different betweencontrol and PBEF-overexpressing SMCs. Interestingly, PBEFexpression has been observed to be upregulated duringdifferentiation/maturation of other cell types, including den-dritic cells and B-lymphocytes.5,28
The relationship between PBEF and specialized SMCperformance was supported in an in vivo context. In order fornewly formed vascular networks to survive, they must beensheathed by SMCs for support and stabilization.1,29 Invest-ment by SMCs also enables the vessels to respond tovasoactive stimuli and thereby appropriately distribute blood.We capitalized on this in vivo, integrative response as afunctional readout for SMC maturation. Human SMCs withreduced PBEF expression displayed a reduced ability toinvest nascent microvessels. In contrast, SMCs overexpress-ing PBEF responded to the angiogenic environment byintimately wrapping around new microvessels and assumingan elongated morphology with a compact nucleus. It remainspossible that the enhanced microvessel investment was attrib-utable, in part, to increased directed migration of PBEF-overexpressing SMCs. Indeed, maturation of HITB5/C6SMCs in culture was closely linked with directed SMCmigration and, in vivo, SMC migration to endothelial chan-nels may similarly be integrated with SMC maturation. Theinvestment of microvessels with maturing SMCs is importantin ischemic tissues, where neovessels are prone to regression.Furthermore, tumor vasculature inadequately invested bySMCs can result in leaky vessels that are predisposed totumor shedding.30 The current findings thus suggest thatmanipulating NAD� biosynthesis pathways may be relevantto these problems.
In summary, PBEF is not a cytokine but an intracellularprotein that regulates NAD� biosynthesis, NAD�-dependentprotein deacetylation, vascular SMC survival, and the capac-ity of SMCs to mature. Augmentation of NAD� regeneration,via PBEF, could have therapeutic potential for vascularconditions that depend on stress-resistant SMCs that canefficiently differentiate.
AcknowledgmentsThis work was supported by grants from the Heart and StrokeFoundation of Canada (T4458), the Heart and Stroke Foundation ofOntario (HSFO) (PRG 4854), the Canadian Institutes of HealthResearch (MT11715), and the Heart and Stroke Foundation ofOntario Career Investigator award (to J.G.P.).
References1. Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat
Med. 2000;6:389–395.2. Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular
smooth muscle cell differentiation in development and disease. PhysiolRev. 2004;84:767–801.
3. Li S, Sims S, Jiao Y, Chow LH, Pickering JG. Evidence from a novelhuman cell clone that adult vascular smooth muscle cells can convertreversibly between noncontractile and contractile phenotypes. Circ Res.1999;85:338–348.
4. Li S, Fan YS, Chow LH, Van Den Diepstraten C, van der Veer E, SimsSM, Pickering JG. Innate diversity of adult human arterial smooth musclecells: cloning of distinct subtypes from the internal thoracic artery. CircRes. 2001;89:517–525.
5. Samal B, Sun Y, Stearns G, Xie C, Suggs S, McNiece I. Cloning andcharacterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol Cell Biol. 1994;14:1431–1437.
6. Ognjanovic S, Bryant-Greenwood GD. Pre-B-cell colony-enhancingfactor, a novel cytokine of human fetal membranes. Am J Obstet Gynecol.2002;187:1051–1058.
7. Jia SH, Li Y, Parodo J, Kapus A, Fan L, Rotstein OD, Marshall JC. Pre-Bcell colony-enhancing factor inhibits neutrophil apoptosis in experimentalinflammation and clinical sepsis. J Clin Invest. 2004;113:1318–1327.
8. Fukuhara A, Matsuda M, Nishizawa M, Segawa K, Tanaka M, KishimotoK, Matsuki Y, Murakami M, Ichisaka T, Murakami H, Watanabe E,Takagi T, Akiyoshi M, Ohtsubo T, Kihara S, Yamashita S, Makishima M,Funahashi T, Yamanaka S, Hiramatsu R, Matsuzawa Y, Shimomura I.Visfatin: a protein secreted by visceral fat that mimics the effects ofinsulin. Science. 2005;307:426–430.
9. Rongvaux A, Shea RJ, Mulks MH, Gigot D, Urbain J, Leo O, Andris F.Pre-B-cell colony-enhancing factor, whose expression is up-regulated inactivated lymphocytes, is a nicotinamide phosphoribosyltransferase, acytosolic enzyme involved in NAD biosynthesis. Eur J Immunol. 2002;32:3225–3234.
10. Kitani T, Okuno S, Fujisawa H. Growth phase-dependent changes in thesubcellular localization of pre-B-cell colony-enhancing factor. FEBS Lett.2003;544:74–78.
11. Martin PR, Shea RJ, Mulks MH. Identification of a plasmid-encoded genefrom Haemophilus ducreyi which confers NAD independence. J Bac-teriol. 2001;183:1168–1174.
12. Rongvaux A, Andris F, Van Gool F, Leo O. Reconstructing eukaryoticNAD metabolism. Bioessays. 2003;25:683–690.
13. Magni G, Amici A, Emanuelli M, Orsomando G, Raffaelli N, Ruggieri S.Enzymology of NAD� homeostasis in man. Cell Mol Life Sci. 2004;61:19–34.
14. Rocnik EF, van der Veer E, Cao H, Hegele RA, Pickering JG. Functionallinkage between the endoplasmic reticulum protein Hsp47 and pro-collagen expression in human vascular smooth muscle cells. J Biol Chem.2002;277:38571–38578.
15. Ismail JA, Poppa V, Kemper LE, Scatena M, Giachelli CM, Coffin JD,Murry CE. Immunohistologic labeling of murine endothelium. Car-diovasc Pathol. 2003;12:82–90.
16. Slomp J, Gittenger-de Groot AC, Gluknova MA, van Munsteren JC,Kockx MM, Schwartz SM, Koteliansky VE. Differentiation, dedifferen-tiation, and apoptosis of smooth muscle cells during the development ofthe human ductus arteriosus. Arterioscler Thromb Vasc Biol. 1997;17:1003–1009.
17. Imai S, Johnson FB, Marciniak RA, McVey M, Park PU, Guarente L.Sir2: an NAD-dependent histone deacetylase that connects chromatinsilencing, metabolism, and aging. Cold Spring Harb Symp Quant Biol.2000;65:297–302.
18. Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA.Nicotinamide and PNC1 govern lifespan extension by calorie restrictionin Saccharomyces cerevisiae. Nature. 2003;423:181–185.
19. Grozinger CM, Chao ED, Blackwell HE, Moazed D, Schreiber SL.Identification of a class of small molecule inhibitors of the sirtuin family
van der Veer et al PBEF and Vascular Smooth Muscle 33
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

of NAD-dependent deacetylases by phenotypic screening. J Biol Chem.2001;276:38837–38843.
20. Buckley S, Barsky L, Driscoll B, Weinberg K, Anderson KD, WarburtonD. Apoptosis and DNA damage in type 2 alveolar epithelial cells culturedfrom hyperoxic rats. Am J Physiol. 1998;274:L714–L720.
21. Ognjanovic S, Tashima LS, Bryant-Greenwood GD. The effects of pre-B-cell colony-enhancing factor on the human fetal membranes bymicroarray analysis. Am J Obstet Gynecol. 2003;189:1187–1195.
22. Hug C, Lodish HF. Medicine. Visfatin: a new adipokine. Science. 2005;307:366–367.
23. Blander G, Guarente L. The Sir2 family of protein deacetylases. Annu RevBiochem. 2004;73:417–435.
24. Araki T, Sasaki Y, Milbrandt J. Increased nuclear NAD biosynthesis andSIRT1 activation prevent axonal degeneration. Science. 2004;305:1010–1013.
25. Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Cohen H, Lin SS,Manchester JK, Gordon JI, Sinclair DA. Manipulation of a nuclearNAD� salvage pathway delays aging without altering steady-stateNAD� levels. J Biol Chem. 2002;277:18881–18890.
26. Revollo JR, Grimm AA, Imai S. The NAD biosynthesis pathwaymediated by nicotinamide phosphoribosyltransferase regulates Sir2activity in mammalian cells. J Biol Chem. 2004;279:50754–50763.
27. Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B,Howitz KT, Gorospe M, De Cabo R, Sinclair DA. Calorie restrictionpromotes mammalian cell survival by inducing the SIRT1 deacetylase.Science. 2004;305:390–392.
28. Tureci O, Bian H, Nestle FO, Raddrizzani L, Rosinski JA, Tassis A,Hilton H, Walstead M, Sahin U, Hammer J. Cascades of transcriptionalinduction during dendritic cell maturation revealed by genome-wideexpression analysis. FASEB J. 2003;17:836–847.
29. Hirschi KK, Rohovsky SA, D’Amore PA. PDGF, TGF-beta, and het-erotypic cell-cell interactions mediate endothelial cell-inducedrecruitment of 10T1/2 cells and their differentiation to a smooth musclefate. J Cell Biol. 1998;141:805–814.
30. Eberhard A, Kahlert S, Goede V, Hemmerlein B, Plate KH, Augustin HG.Heterogeneity of angiogenesis and blood vessel maturation in humantumors: implications for antiangiogenic tumor therapies. Cancer Res.2000;60:1388–1393.
34 Circulation Research July 8, 2005
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

Geoffrey PickeringEric van der Veer, Zengxuan Nong, Caroline O'Neil, Brad Urquhart, David Freeman and J.
Activity and Promotes Vascular Smooth Muscle Cell Maturation-Dependent Protein Deacetylase+Enhancing Factor Regulates NAD−B-Cell Colony−Pre
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2005 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000173298.38808.272005;97:25-34; originally published online June 9, 2005;Circ Res.
http://circres.ahajournals.org/content/97/1/25World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2005/06/09/01.RES.0000173298.38808.27.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 23, 2016http://circres.ahajournals.org/Downloaded from

1
PRE-B-CELL COLONY-ENHANCING FACTOR REGULATES NAD+-
DEPENDENT PROTEIN DEACETYLASE ACTIVITY AND PROMOTES
VASCULAR SMOOTH MUSCLE CELL MATURATION
ONLINE DATA SUPPLEMENT
Eric van der Veer, Zengxuan Nong, Caroline O’Neil, Brad Urquhart, David Freeman, and
J. Geoffrey Pickering
EXPANDED MATERIALS AND METHODS
Cell Culture
Experiments were performed using the maturation-competent human vascular
SMC lines, HITB5 and HITC6, generated from the human internal thoracic artery, as
described previously.1,2 SMCs were maintained in M199 (GibcoBRL) supplemented with
the designated concentration of FBS (Hyclone).
Overexpression of PBEF in Human SMCs
A retroviral gene delivery system was used to generate human SMCs stably
overexpressing PBEF. Full-length cDNA encoding PBEF was amplified from HITB5
SMC mRNA by RT-PCR and subcloned into the pIRES-EGFP vector (Clontech). The
PBEF-IRES-EGFP bicistronic fragment was excised (XhoI, NotI) and inserted into the
retroviral expression vector pLNCX2 (Clontech). A second retroviral expression
construct was generated by inserting PBEF cDNA into pQCXIP-IRES-PURO (Clontech).
Retrovirus containing the cDNA of interest was obtained by calcium phosphate-mediated

2
transfection of the Phoenix-amphotropic retrovirus packaging cell line (kindly provided
by Dr. G Nolan, Stanford University Medical School, CA, distributed by ATCC,
Manassas, VA) as described previously.3 Virus-containing supernatant was added to
proliferating SMCs and stable transductants were selected with 500 µg/ml G418 for 14
days, for pLNCX2-based constructs, and with 3 µg/ml puromycin for 48 hours, for
pQCXIP-based constructs. Overexpression of PBEF was confirmed before each
experiment by Western blot analysis.
Western Blot Analysis
Expression of PBEF and SMC differentiation markers was assessed by Western
blot analysis with chemiluminescence detection, as described.3 PBEF was detected using
a polyclonal rabbit antibody against human PBEF (1857, 1:5000, kindly provided by
Amgen). Monoclonal antibodies were used to detect heavy (h)-caldesmon (clone hHCD,
1:1000, Sigma), smoothelin isoforms A and B (clone MAB3242, 1:500, Chemicon),
vinculin-metavinculin (clone VIN-11-5, 1:2000, Sigma), and α-tubulin (clone B-5-1-2;
1:16000, Sigma). Bands were quantified by laser scanning densitometry (GS-700
Imaging Densitometer, BioRad).
Apoptosis
SMC apoptosis was assessed by fluorescence in situ end-labeling of DNA
fragments, as described.1
Knockdown of PBEF by RNA Interference
PBEF knockdown was accomplished by infecting human SMCs with retrovirus
containing sequences encoding hairpin siRNA fragments. Complementary
oligodeoxynucleotides were synthesized, annealed, and inserted between the BamH1 and
EcoR1 sites of the retroviral expression vector pSIREN-RetroQ (in vitro experiments) or

3
the BglII and HindIII sites of the retroviral expression vector pSUPER-EGFP/Neo (in
vivo experiments). Three different targeting sequences were used, each consisting of 19
nucleotides starting at nucleotides 147, 384, and 1278 of the PBEF coding sequence
(siRNA147 5’-GGAAGGTGAAATATGAGGA-3’; siRNA384 5’-ATGTTCT-
CTTCACGGTGGA-3’; siRNA1278 5’-AGGGCCGATTATCTTTACA-3’). Each
sequence was separated by a 9-nucleotide noncomplementary spacer from the reverse
complement of the same 19-nucleotide sequence. Blast search confirmed that only the
PBEF gene was targeted. Control inserts contained the gene-specific 19-nucleotide
sequence and hairpin loop sequence but not the antisense component. Infection of SMCs
was performed as described above and cells were selected using 3 µg/ml puromycin for
48 hours or 500 µg/ml G418 for 2 days and 250 µg/ml for 2 days. The latter selection
protocol was deliberately gentle, as it was observed that PBEF knockdown SMCs were
susceptible to nonspecific culture stresses and EGFP-positive transductants survived
poorly with more aggressive selection protocols.
Real-time RT-PCR
RNA was harvested using RNeasy mini-column and reagents (Qiagen) and
subjected to DNaseI treatment. Probe (5’-CAGTTGCTGATCCCA-3’) and flanking
primers (5’-primer 5’-TGCAGCTATGTTGTAACCAATGG-3’; 3’-primer 5’-
ACAAAAGGTCGAAAAAGGGCC-3’) for Taqman real-time RT-PCR for PBEF were
designed using Primer Express software (Applied Biosystems). Reactions were
performed using an ABI-Prism 7900 Sequence Detection System (Applied Biosystems).
Optimum signal was obtained with concentrations of 200nM, 300nM and 50nM for the
probe, 5’ primer, and 3’ primer, respectively. Standard curves were generated using
RNA derived from human aortic SMCs, enabling correlation of the determined threshold
cycle to transcript abundance. GAPDH transcript abundance was used as an endogenous

4
RNA control (Assays on Demand Hs9999905, Applied Biosystems) to which PBEF
transcript abundance was normalized.
Nicotinamide Phosphoribosyltransferase Activity
Cultured SMC or rat liver lysates were reacted with 5 µM [carbonyl-14C]
nicotinamide for 2 h at 37oC in the presence of 0.5 mM phosphoribosylpyrophosphate, 5
mM MgCl2, 2 mM ATP, and 50 mM Tris, pH8.8, as described.4,5 Nicotinamide
mononucleotide was retrieved by acetone precipitation and filtration through an acetone-
soaked Whatman GF/A filter and labeled product was quantified by scintillation
counting.
NAD+ analysis by HPLC
Cellular nucleotides were extracted using perchloric acid, neutralized with KOH,
and stored at -80°C. The deproteinized cell lysate residues were then analyzed by HPLC
using a mobile phase of 10mM KH2PO4, 0.12% di-n-butylamine, pH, 3.0. Sample was
injected onto a Prodigy C8 column (150 X 3.2 mm, 5µm) (Phenomenex) by a Hewlett
Packard 1090 chromatograph. The column temperature and flow rate were maintained at
40°C and 0.5 ml/min, respectively. The effluent was monitored at 260 nm by a Hewlett
Packard 1050 UV-VIS detector and NAD+ retention time, determined from an NAD+
standard, was 10 minutes.
Histone Deacetylase (HDAC) Assay
Histone H4 peptide (Upstate) was labeled with [3H]-acetyl Coenzyme A (ICN)
using PCAF histone acetylase. Cell lysates were harvested using Passive Lysis Buffer
(Promega) and 25 µg of protein was incubated at 37°C for 6 hours with 50,000 cpm of
[3H]-acetyl histone and 1mM PMSF in HDAC Assay Buffer (Upstate). Released [3H]-

5
acetate was extracted with ethyl acetate6 and counted on a Beckman LS3801 scintillation
counter.
Human-Mouse Chimeric Angiogenesis in vivo
HITC6 SMCs in M199 with 10% FBS and fibroblast growth factor-2 (FGF-2)
were mixed with an equal volume of growth factor-reduced matrigel (BD Discovery
Labware) yielding final concentrations of 5 x 105 cells/ml and 250 ng/ml FGF-2. The
cell-matrigel suspension (500µl) was subcutaneously injected into the abdomen of mice
with severe combined immunodeficiency syndrome (SCID). After 8 days, implants were
harvested, fixed for 8h in Tris-buffered zinc,7 and paraffin-embedded tissues were
sectioned at 5µm. After deparaffinization, endogenous peroxidases were quenched with
0.3% H2O2, nonspecific binding was blocked with 5% goat serum and sections were
immunostained for CD31 (BD Biosciences), and GFP (BD Clontech). To simultaneously
visualize mouse endothelial cells and human SMCs, sections were incubated overnight at
4oC with biotinylated rat anti-mouse CD31 and bound antibody reacted with ABC
reagent (Vector Laboratories Ltd.) and diaminobenzidine (DAB, Vector Laboratories
Ltd.). Tissue was then incubated with rabbit anti-GFP (1:200, BD Clontech) and bound
primary antibody detected with alkaline phosphatase-conjugated goat anti-rabbit
secondary antibody and visualized with red alkaline phosphatase substrate (Vector
Laboratories Ltd.). Single and double immunolabeled sections were counterstained with
Harris’ hematoxylin. Apoptosis of human SMCs in matrigel was assessed by TUNEL
assay, after immunostaining for EGFP and incubating sections with proteinase K (100
µg/ml) for 10 min at 37oC.
The density of all EGFP-positive human SMCs and of TUNEL-positive human
SMCs was determined from 12-15 randomly selected high-powered (x40 objective)
fields. For each implant, 3 sections were quantified corresponding to 25% sectioning
depth, mid-plane, and 75% depth and the values were averaged. Quantification of

6
microvessel density and SMC wrapping were assessed from 10-12 contiguous high-
powered (x40 objective) fields that constituted the zone of matrigel closest to the adjacent
skeletal muscle. This zone, which accounted for about 30% of the matrigel area, was the
region of matrigel that contained the most microvessels and displayed the most consistent
angiogenic response. It was thus predefined as the analysis region for microvessel
quantification. For each implant, 3 sections were quantified as described above. Human
SMCs were deemed to be investing a microvessel if they assumed contiguous and close
apposition with an endothelial cell(s) and oriented their long axis along a contour of the
vessel wall itself. Animals and slides were coded and read independently by 2
investigators and the values were averaged. Animal experiments were performed under a
protocol approved by the Animal Care and Use Committee, University of Western
Ontario.
Statistics
Values are expressed as mean ± standard error of the mean. Comparisons were
made by t-test or analysis of variance with Bonferroni’s post-hoc test. Significance of
fold-changes was assessed by the Wilcoxon signed-rank test. Statistical significance was
set at p<0.05.
SUPPLEMENTAL RESULTS
Sequence and phylogenetic analysis of PBEF in silico
In view of the controversy over PBEF function and whether it acts as a secreted
cytokine or an intracellular enzyme, we generated a phylogenetic tree using protein
sequences similar to human PBEF, derived from a Blast search of the NCBI sequence
database. Multiple sequence alignment revealed that sequences from diverse species,

7
bacterial and eukaryotic, were similar in length, contained strongly conserved regions
with respect to helix propensity and hydrophobicity, and a conserved
phosphoribosyltransferase domain. The phylogenetic tree revealed low point-assisted
mutation scores (below 0.35), reflecting the short distances amongst all nodes and leaves
of the tree. Thus, revealed the PBEF protein has been well conserved throughout
evolution, suggesting a fundamental and invariant role.
Investment of Newly Formed Blood Vessels With SMCs Requires PBEF
The in vivo behavior of SMCs with reduced levels of PBEF was assessed using
SMCs infected with retrovirus containing control (pSUPER-nsRNA1278-EGFP) or
PBEF knockdown (pSUPER-siRNA1278-EGFP) constructs. Addition of SMCs
expressing siRNA1278 to FGF2-matrigel mixtures did not affect the density of
microvessels within the tissue construct, 8 days after implantation (283±26 vs 266±25
vessels/mm2). Apoptosis of SMCs expressing PBEF siRNA was also not significantly
different from that of control SMCs (1.9±0.3 vs 1.6±0.3%), which may reflect the
survival stimuli arising from the matrigel-FGF-2 environment. However, the proportion
of microvessels invested by HITC6-siRNA SMCs was significantly reduced (3.1±0.6 vs
9.4±1.4%. p<0.05).
REFERENCES
1. Li S, Sims S, Jiao Y, Chow LH, Pickering JG. Evidence from a novel human cell
clone that adult vascular smooth muscle cells can convert reversibly between
noncontractile and contractile phenotypes. Circ Res. 1999;85:338-48.
2. Li S, Fan YS, Chow LH, Van Den Diepstraten C, van Der Veer E, Sims SM,
Pickering JG. Innate diversity of adult human arterial smooth muscle cells:

8
cloning of distinct subtypes from the internal thoracic artery. Circ Res.
2001;89:517-25.
3. Rocnik EF, van der Veer E, Cao H, Hegele RA, Pickering JG. Functional linkage
between the endoplasmic reticulum protein Hsp47 and procollagen expression in
human vascular smooth muscle cells. J Biol Chem. 2002;277:38571-8.
4. Elliott GC, Ajioka J, Okada CY. A rapid procedure for assaying nicotinamide
phosphoribosyltransferase. Anal Biochem. 1980;107:199-205.
5. Rongvaux A, Shea RJ, Mulks MH, Gigot D, Urbain J, Leo O, Andris F. Pre-B-
cell colony-enhancing factor, whose expression is up-regulated in activated
lymphocytes, is a nicotinamide phosphoribosyltransferase, a cytosolic enzyme
involved in NAD biosynthesis. Eur J Immunol. 2002;32:3225-34.
6. Landry J, Sternglanz R. Enzymatic assays for NAD-dependent deacetylase
activities. Methods. 2003;31:33-9.
7. Ismail JA, Poppa V, Kemper LE, Scatena M, Giachelli CM, Coffin JD, Murry
CE. Immunohistologic labeling of murine endothelium. Cardiovasc Pathol.
2003;12:82-90.




















