
Porteresia coarctata (Roxb.) Tateoka, a wild rice: a potential model for studying salt-stress...
17
Porteresia coarctata (Roxb.) Tateoka, a wild rice: a potential model for studying salt-stress biology in riceSONALI SENGUPTA & ARUN LAHIRI MAJUMDER Plant Molecular and Cellular Genetics, Bose Institute, P-1/12, C I T Scheme VII M, Kolkata: 700 054, India ABSTRACT Porteresia coarctata (Syn = Oryza coarctata) is a tetrap- loid wild rice growing abundantly in the coastal region of India and some other Asian countries. The salt tolerance property of this mangrove associate has been dealt with by a number of workers earlier. The distinct morphology and leaf architecture enabling the plant to exclude salt is a char- acteristic feature of Porteresia in comparison with Oryza sp. A number of genes have been isolated and characterized from Porteresia that are related to the salt-tolerance prop- erty of the plant. Evidence have accumulated that some pathways critical to salt tolerance are in operation in Por- teresia of which the inositol metabolic pathway has been recently elaborated. Some of the enzymes of Porteresia have been shown to function as salt-tolerant under in vitro studies giving a clue that this wild halophytic rice may have evolved genes and proteins capable of functioning under a salt environment. Bioprospecting of such genes and pro- teins coupled with genomic and proteomic approaches remain an exciting area of research in evaluating this plant as a model for salt tolerance for the rice plant. Key-words: salt hairs; salt tolerance; wild rice. INTRODUCTION Soil with an electrical conductivity (ECe) above 4 dS m -1 (ª 40 mm NaCl) is defined as saline soil. Increasing soil salinity is the most critical factor regulating agricultural productivity worldwide. Accumulation of soluble salts of various types, mainly chlorides of sodium, calcium, magne- sium and sulphates or carbonates, makes groundwater con- taminated. The high osmotic potential of contaminated water and accompanying ionic toxicity create imbalance in the supplies of water and nutrient solutes. Such imbalance disturbs normal ecosystem and also generates perturbation in designed agriculture industry. NaCl accumulation is one of the major sources of such perturbation. Sodium salinity represents a considerable threat to food provisions of world population resulting from limited growth and low harvest- able yield of major crop species such as wheat, rice, corn, potato, cotton or sorghum in a global scale. Most of the common grain crops and vegetables are highly susceptible to soil salinity at the soil ECe range 4 dS m -1 . However, a salt-tolerant or salt-loving species (halophyte) can tolerate salinity up to 20–40 dS m -1 or more in sharp contrast to its glycophytic kinfolks. This broad range of tolerance proves the existence of a very high genetic variation among plant species for the salt-tolerance trait. Exploring this genetic variation is necessary for setting up a model halophytic system for crops. Fortunately, the tools of genetic engineering totally supplements sustainable agriculture in the modern era and there are more reason- able approaches towards raising genetically modified (GM) crops without residual harmful effects.Towards the practi- cal end, plant stress biology of the day is under constant demand of entirety in our knowledge of plant response to salinity in a species/variety specific manner, which is necessary to ascertain the exact traits to improve resistance of a crop without altering its adaptability, status in the evolutionary process and the normal genetic complement. Setting up a model system for salinity tolerance improve- ment in a crop is thus an arduous task and claims several layers of consideration at taxonomic, genetic and molecular level. ADVERSE EFFECTS OF SALINITY ON PLANTS Physiologically, sodium chloride-based salinity has several implications on plant growth and development. It imposes an initial water deficit that results from the relatively high solute concentrations in the soil, causes ion-specific stresses resulting from altered K + /Na + ratio and builds up detrimen- tal levels of cellular Na + and Cl - concentrations. Growth and survival of vascular plants at high salinity depends on adap- tation to both low water potentials and high sodium con- centrations, as high salinity in the external solution of plant cells produces a variety of negative consequences. Consid- ering sodium chloride as the primary inflictor of salinity, the penalty of a high extracellular NaCl concentration on plant is obvious. Cellular ionic imbalance generated from excess sodium and chloride ions affect life processes negatively. Na + enters cell efficiently through K + transport channels, effectively replaces monovalent cations like K + as cofactors of several enzymatic processes, and in doing so inhibits such enzyme activities. Therefore, plant survival and growth depends on adaptations to re-establish ionic homeostasis (Hasegawa et al. 2000). Hyperosmotic shock is another result of high salinity. Chemical activity of water is decreased causing a loss of cell turgor. Salt induced reduction in chloroplast stromal Correspondence: A. L. Majumder. Fax: +852-2879-5170; e-mail: [email protected]; [email protected] Plant, Cell and Environment (2010) 33, 526–542 doi: 10.1111/j.1365-3040.2009.02054.x © 2009 Blackwell Publishing Ltd 526
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Porteresia coarctata (Roxb.) Tateoka, a wild rice: a potential model for studying salt-stress...

Porteresia coarctata (Roxb.) Tateoka, a wild rice: apotential model for studying salt-stress biology in ricepce_2054 526..542
SONALI SENGUPTA & ARUN LAHIRI MAJUMDER
Plant Molecular and Cellular Genetics, Bose Institute, P-1/12, C I T Scheme VII M, Kolkata: 700 054, India
ABSTRACT
Porteresia coarctata (Syn = Oryza coarctata) is a tetrap-loid wild rice growing abundantly in the coastal region ofIndia and some other Asian countries. The salt toleranceproperty of this mangrove associate has been dealt with bya number of workers earlier. The distinct morphology andleaf architecture enabling the plant to exclude salt is a char-acteristic feature of Porteresia in comparison with Oryzasp. A number of genes have been isolated and characterizedfrom Porteresia that are related to the salt-tolerance prop-erty of the plant. Evidence have accumulated that somepathways critical to salt tolerance are in operation in Por-teresia of which the inositol metabolic pathway has beenrecently elaborated. Some of the enzymes of Porteresiahave been shown to function as salt-tolerant under in vitrostudies giving a clue that this wild halophytic rice may haveevolved genes and proteins capable of functioning under asalt environment. Bioprospecting of such genes and pro-teins coupled with genomic and proteomic approachesremain an exciting area of research in evaluating this plantas a model for salt tolerance for the rice plant.
Key-words: salt hairs; salt tolerance; wild rice.
INTRODUCTION
Soil with an electrical conductivity (ECe) above 4 dS m-1
(ª 40 mm NaCl) is defined as saline soil. Increasing soilsalinity is the most critical factor regulating agriculturalproductivity worldwide. Accumulation of soluble salts ofvarious types, mainly chlorides of sodium, calcium, magne-sium and sulphates or carbonates, makes groundwater con-taminated. The high osmotic potential of contaminatedwater and accompanying ionic toxicity create imbalance inthe supplies of water and nutrient solutes. Such imbalancedisturbs normal ecosystem and also generates perturbationin designed agriculture industry. NaCl accumulation is oneof the major sources of such perturbation. Sodium salinityrepresents a considerable threat to food provisions of worldpopulation resulting from limited growth and low harvest-able yield of major crop species such as wheat, rice, corn,potato, cotton or sorghum in a global scale.
Most of the common grain crops and vegetables arehighly susceptible to soil salinity at the soil ECe range
4 dS m-1. However, a salt-tolerant or salt-loving species(halophyte) can tolerate salinity up to 20–40 dS m-1 or morein sharp contrast to its glycophytic kinfolks. This broadrange of tolerance proves the existence of a very highgenetic variation among plant species for the salt-tolerancetrait. Exploring this genetic variation is necessary for settingup a model halophytic system for crops. Fortunately, thetools of genetic engineering totally supplements sustainableagriculture in the modern era and there are more reason-able approaches towards raising genetically modified (GM)crops without residual harmful effects. Towards the practi-cal end, plant stress biology of the day is under constantdemand of entirety in our knowledge of plant responseto salinity in a species/variety specific manner, which isnecessary to ascertain the exact traits to improve resistanceof a crop without altering its adaptability, status in theevolutionary process and the normal genetic complement.Setting up a model system for salinity tolerance improve-ment in a crop is thus an arduous task and claims severallayers of consideration at taxonomic, genetic and molecularlevel.
ADVERSE EFFECTS OF SALINITY ON PLANTS
Physiologically, sodium chloride-based salinity has severalimplications on plant growth and development. It imposesan initial water deficit that results from the relatively highsolute concentrations in the soil, causes ion-specific stressesresulting from altered K+/Na+ ratio and builds up detrimen-tal levels of cellular Na+and Cl- concentrations. Growth andsurvival of vascular plants at high salinity depends on adap-tation to both low water potentials and high sodium con-centrations, as high salinity in the external solution of plantcells produces a variety of negative consequences. Consid-ering sodium chloride as the primary inflictor of salinity, thepenalty of a high extracellular NaCl concentration on plantis obvious. Cellular ionic imbalance generated from excesssodium and chloride ions affect life processes negatively.Na+ enters cell efficiently through K+ transport channels,effectively replaces monovalent cations like K+ as cofactorsof several enzymatic processes, and in doing so inhibits suchenzyme activities. Therefore, plant survival and growthdepends on adaptations to re-establish ionic homeostasis(Hasegawa et al. 2000).
Hyperosmotic shock is another result of high salinity.Chemical activity of water is decreased causing a loss ofcell turgor. Salt induced reduction in chloroplast stromal
Correspondence: A. L. Majumder. Fax: +852-2879-5170; e-mail:[email protected]; [email protected]
Plant, Cell and Environment (2010) 33, 526–542 doi: 10.1111/j.1365-3040.2009.02054.x
© 2009 Blackwell Publishing Ltd526

volume and generation of reactive oxygen species (ROS)also plays an important role in decreasing plant photosyn-thetic capacity and therefore growth. There are severalillustrious and knowledgeable reviews on the salt-tolerancemechanisms of plants (Ingram & Bartels 1996; Hasegawaet al. 2000), the elaboration of which is beyond the scopeof current discussion. Considering the diverse deleteriouseffects of salinity on plants, it is easy to understand thatadaptation to salinity in a plant is a summation of severalmetabolic modifications, new mechanisms and alteredexpression levels. It is also conceivable that raising a salt-tolerant crop would involve modification of a large numberof parameters and therefore a holistic approach towardsexploring the salt-tolerance mechanism inherent of ahalophyte is needed.
HALOPHYTES ARE GOOD MODELS FORSTUDYING SALINITY STRESS
Halophytes are generally a small number of plant lineagesthat have evolved structural, phenological, physiologicaland biochemical mechanisms for salt tolerance and the evo-lution is convergent in numerous related families. Desertand coastal halophytes possess similar mechanisms fordealing with salt toxicity, salt stress and water deficit andsometimes share phylogenetic ancestry. There are alsomarine phanerogams (seed-bearing plants) that live com-pletely submerged in seawater.
Among the angiosperms, Chenopodiaceae and Poaceaeshow the largest number of halophytes. The Chenopodi-aceae has over 380 halophytic species and the same forPoaceae is over 140 (Flowers, Hajibagheri & Clipson 1986).Halophytic succulence is the common trait in the halophyticgenera of the family Chenopodiaceae such as Allenrolfea,Arthrocnemum, Halimione, Nitrophil, Salicornia, Suaedaand Artiplex. True succulents such as Aizoon and Mesem-bryanthemum belonging to the ice plant family Aizoaceaeare widely studied (Adams et al. 1998). The large centralvacuole in thin-walled cells of their succulent leaves andstems accumulate substantial quantities of salts. By depos-iting ions of salts in vacuoles, the toxicity is partitioned fromthe cytoplasm and organelles of the cells.
For the Poaceae, most of the halophytic plants are typicalsalt extruder, which includes three tribes of grasses,Sporoboleae, Chlorideae and Aeluroideae. Some membersof these are the well-known tidal saltmarsh plants as thecordgrasses (Spartina alterniflora, S. patens), alkali grass(Puccinellia phryganodes), saltgrass (Distichlis spicata) andshoregrass (Monanthochloe littoralis). Short, two-celled tri-chomes serve as collecting chambers for salts, principallybetween the cuticle and the wall of the cap cell. From thereit is forced through tiny pores in the stretched cuticle.
Among the halophytes, the seagrasses, which are sub-merged plants of shallow marine meadow, have true salttolerance, because the osmolality of cytoplasm is adjustedto match the osmolality of the seawater, so that desiccationof the plant does not occur. This contrasts the strategies ofexcluders and succulents, which are primarily salt-avoiding
mechanisms that attempt to reduce salt concentrationswithin the cytoplasm of photosynthetic cells.
Response of a multicellular halophytic plant towards saltstresses could be dissected in three levels – cellular level,tissue level and organismal level. Again, a halophyte couldbe obligate towards saline environment, that is, it may notbe able to grow in a glycophytic mode. Such halophyticplants exhibit a great range of unique physiological adap-tations, such as viviparous germination, pneumatophores orelaborate salt glands, which are not possible to transfer intocrop plants. These plants are best suited for saline environ-ment, but it is difficult to decipher the code of such adapta-tions. So, one looks for facultative halophytes, which show avariable range of affinity towards salt and may also grow ina glycophytic mode.
For a crop improvement program, it is a common practiceto look among the wild relatives of domesticated crops thatcarry a good gene pool albeit compromising yield and cropquality. The current trend of stress biotechnology researchis to improve the tolerance level of domestic crops bytransferring genes for individual functional enzymes, or theentire metabolic pathways, which might have evolved tofunction in presence of high sodium in halophytes. Thisrequires ample understanding of life processes underlyingsalt stress biology as well as establishment of a comparabledomestic and halophytic model eligible for gene transfer orbreeding.
IN SEARCH OF A MONOCOT MODEL SYSTEMFOR SALT TOLERANCE
The complexity of higher plants causes problems in experi-mentation and interpretation of results, making it difficultto decipher the exact effect that salt exerts on plants. Ourfocus has been the grain crops, which are monocots belong-ing mostly to the grass family. The domesticated grass cropsencompass an extraordinarily diverse set of species distrib-uted around the world, including the most important cropsfor human subsistence, such as wheat (Triticum aestivum),rice (Oryza sativa), sorghum (Sorghum bicolor), maize (Zeamays), barley (Hordeum vulgare), rye (Secale cereale) andoat (Avena sativa). Some of them are temperate grasses, likewheat, maize and oat and some, are tropical like rice. Rice(O. sativa L.) is an important crop in eastern Asia and alsois the staple food of one-third of world population. A vastnumber of rice cultivars as well as wild species of rice arewidely grown, and their genetic and molecular make-up arebeing actively investigated (Sasaki & Sederoff 2003).
Rice is considered a model plant in monocots because ofthe relatively small size of its genome. The rice genomeconsists of about 430 million base pairs, and about 30 000genes can be expressed in rice plant tissue (Komatsu et al.2003).The nuclear genome sequences of rice have now beendeciphered (Yu et al. 2002). These sequences have affordedplant scientists an unprecedented opportunity to investi-gate the genome organization in plants and have also pro-vided a wealth of information that can be used to unravelthe function of many new genes.
Porteresia as model for studying salt-stress biology 527
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

Genome sequence of the dicot model Arabidopsis andthe monocot model rice provide contrasting opportunitiesto study abiotic stress tolerance genes and to identify novelbiotechnological approaches to crop improvement. It hasoften been proved fruitful to exploit genomic discoveriesacross distant plant groups. For example, the sequence of anArabidopsis gene involved in gibberellin responsivenessserved as the starting point to isolate Rht dwarfing genes inwheat that have contributed to major yield gains in this crop(Flintham et al. 1997). Although Arabidopsis is not particu-larly an attractive system for salt tolerance, relatives ofArabidopsis with better tolerance strategies have beenexploited generously. Thellungiella halophila, a close rela-tive of Arabidopsis, has been widely used as a model forsalt tolerance (Bressan et al. 2001; Zhu 2001; Inan et al.2004; Taji et al. 2004; Amtmann, Bohnert & Bressan 2005)although salt tolerance in Thellungiella is not comparablewith that of other dicot halophytes (Vera-Estrella et al.2005). Another species in addition to T. halophila, which iswell-established as a model in halophyte research, is theice-plant Mesembryanthemum crystallinum (Adams et al.1998; Bohnert & Cushman 2000), a species with bladdercells that accumulate ions. This species tolerates high con-centrations of salt, and accumulates inositol and its methy-lated derivatives as an immediate response to salt stress. Itis also able to switch the photosynthetic carbon assimilationfrom a C3 mode to crassulacean acid metabolism pathway(Adams et al. 1998). M. crystallinum belongs to the orderCaryophyllales, which includes the Chenopodiaceae andAizoaceae, families with significant numbers of halophyticspecies. Investigations on the mechanism of salt tolerance inSuaeda also seem worth to pursue. Suaeda is representativeof a group of very tolerant halophytes with highly succulentleaves and able to accommodate ions without the need forsecretion via salt glands.
However, considering the responses to salinity, none ofthese dicot plants is ideal as a halophytic model to monocotplants including the domesticated grass crops. As pointedout earlier, the mechanisms of tolerance to abiotic stressescan exhibit fundamentally different characteristics betweenmonocotyledons and dicotyledons. For example, whengrown in saline soils, many dicotyledonous halophytes accu-mulate much higher concentrations of Na+ in their shootsthan monocotyledonous halophytes, a feature that may berelated to the observation that succulence is observed morecommonly in dicotyledons than monocotyledons, particu-larly the grasses. Transferring knowledge from the dicotyle-donous model plants to the major crops (primarily thegrasses, both cereals and forage species) thus may not turnout to be the best strategy.
Such conclusion is supported by evolutionary distancebetween the two major groups. Modern monocot and dicotplant groups diverged from a common ancestor between120 and 200 Ma (Wolfe et al. 1989). The estimated rate ofstructural chromosomal mutation predicted that within a3 cM long region, 50% of the genes would be retained bychance between monocots and dicots (Paterson et al. 1996).The study of Devos et al. (1999) based on similarities
between Arbidopsis genomic sequences and rice expressedsequence tag (ESTs) suggested that synteny between Ara-bidopsis and rice has been eroded during evolution so thatit is no longer detectable using a comparative mappingstrategy. Microsynteny is observed between rice and Arabi-dopsis, but the ancient degree of speciation between thesetwo species makes cross-comparison difficult.
Thus, for researchers interested in studying rice for its salttolerance characteristics (Gao et al. 2007), a halophyticmonocot model is desirable. Moreover, rice is separated by~50 Ma of evolution from many important cool season grasscrops such as wheat. The estimates suggest the grass familyoriginated roughly ~77 Ma. The divergence between Erhar-toideae (rice) and the Poeoideae (oats, barley and wheat) isestimated at 46 Ma (Gaut 2002). Although the tropicalmonocot group akin to rice could use rice as a model, forcool season temperate monocots rice is not a favourablemodel system. Thus a significant void exists in mergingcereal crop improvement in a single bracket owing to thefact that there is no unified model system akin to Arabidop-sis available for grass crops and even within grass cropsthere is a gap between cool season temperate crops andtropical crops. Thus, the temperate and tropical cropsshould be addressed separately in terms of salt-tolerancephenomenon.
Considering the tolerance against salt, domesticated cul-tivars show little genetic variation. Domestication of cropsis a co-evolutionary process that is highly influenced byenvironmental closure of natural selection and humanselection of desirable phenotypes over low-yield, low-quality phenotypes. Such selection evoke changes in thepopulation’s genotypes rendering them more useful tohumans and better adapted to human intervention(Clement 1999). Varying degrees of genetic change can berecognized among the world’s crops, reflecting the duration,intensity and genetic gain realized from human selection.However such biased selection creates a pressure on thetraits like tolerance to pathogens and environmentalextremities that are often not required in a good and idealfield condition. The extant tolerant genes are thus oftenaccumulated in the wild species, which had not been shapedgenetically by human selection over thousands of years.Such genetic remodelling may include subtle changes inorthologs or gross changes in chromosomal or even ploidylevel, which are prevalent among cereal crops. Identificationof extant tolerance genes within cereal germplasms needs abetter understanding of basic tolerance mechanisms in wildmonocots, which is unfortunately only poorly studied.
Because of the importance of members of the Triticeae ascrops, the halophytic relatives of wheat and barley (speciesof Thinopyrum and Hordeum) could be investigated asmodels. Other potential monocotyledonous models consid-ered are species of Distichlis, Spartina and Puccinellia. Brac-hypodium distachyon is another plant currently proposedas a model system for genomics of monocots because ofits small genome (approximately 350 M bp), diploid eco-type (2n = 10), annual lifecycle, self-fertility and a close rela-tionship with the temperate cereals or cool-season grasses
528 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

(Draper et al. 2001). There is, however, no report of anyabiotic stress-tolerance property from Brachypodium. Thegenome sequence of first C4 crop and second food cropSorghum is currently published (Sasaki & Antonio 2009),which is an important reference for drought tolerance anddry land crops.Various projects are in order to trace the highbiomass and C4 mode of photosynthesis in Sorghum, as wellas comparing its genetic colinearity with rice.
In this communication, we discuss the potential of a halo-phytic model for studying rice salt tolerance. Although riceserves the purpose as a domesticated crop model in theperspective of recent advances in rice genomics, the salinitytolerance limit in most of the cultivars of rice is low. Henceidentification of tolerant landraces closely related to ricewith high homology in genetic content is needed.
PORTERESIA COARCTATA, THE WILDHALOPHYTIC RICE
Wild rice species have proven to be tremendous gene res-ervoirs to increase domesticated rice yield, quality, andresistance to diseases and insects. Wild rice species havefurnished genes for the hybrid rice revolution, exhibit yield-enhancing traits and have shown tolerance to biotic andabiotic stress. Among the wild relatives of rice, P. coarctata,which grows in abundance along the eastern and westerncoasts of India, Pakistan and Bangladesh shows con-siderable adaptation to salinity. Porteresia, a tetraploid(2n = 4x = 48) distant wild rice relative, is a monotypicgenus occurring as an associate of mangroves along thecoastal belts (Fig. 1, Porteresia vegetation). A comparative
(a)
(c) (d) (e)
(b)
(f)
Figure 1. Porteresia coarctata: Habitat, morphology and anatomy. (A) Native Porteresia vegetation: (a) inundated condition; (b) non-inundated condition; (c) close-up view of the same. (B) Porteresia growing in green house alongside rice plants. (C) (a) Transverse sectionof Porteresia leaves (adapted from Flowers et al. 1990); (b) salt hairs; scaning electron micrograph; upper surface; (c) salt hairs; scanningelectron micrograph; lower surface. (D) Na : K ratio in Porteresia leaves after 3 d of salt stress. (E) Whole and dehusked seeds of Oryzasativa: (a) variety IR64 (b); variety Pokkali; (c) P. coarctata, Porteresia seeds are not viable in maturity. (F) O. sativa and P. coarctata afterapplication of 14 d NaCl stress (0.2 mm); (a) growth of the plant (Oryza, IR64 variety (salt-sensitive); early ripening of the panicle isobserved as in (b); drying of the culm portion after stress is magnified in (c); (d–f); the same for Oryza sativa (Pokkali, little tolerant tosalt); (g–i); the same for Porteresia coarctata. The difference in the growth of plant from that of rice is easily observed (g). It does notsuffer from drying after stress [as magnified in (i)]. The panicles, however dehisce even without stress, and stress imposed little effect onthat (h).
Porteresia as model for studying salt-stress biology 529
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

pictorial account of the details of Porteresia and Oryzaunder salinity stress has been provided in Fig. 1. Thoughthe salt-tolerance range of Porteresia is yet to be verifiedexperimentally, the natural vegetations are inundated twicea day with saline river or seawater of 20 to 40 dS m-1 (Bal &Dutt 1986). P. coarctata act as pioneer species in the succes-sion process of mangrove formation along the estuaries ofIndia (Jagtap, Bhosale & Singh 2006). Porteresia forms avast population as mangrove associates throughout the rootmatrices of tree mangroves, binds the soil and prevents thecoastline from erosion. The plant is distributed along theeast and west coastlines of India, the major habitats beingthe Sunderbans, Goa, Chennai coastline and Bhitorkonikareserve forest in Orissa.
GENETIC RELATIONSHIPS OF P. COARCTATAWITH O. SATIVA
The genus Oryza comprising of 24 species with two culti-vated ones, is distributed in Asia, Africa, Australia, Centraland South America. Cytogenetic analyses distinguished 10different genome types based on chromosome pairingbehaviour at meiosis in interspecific hybrids (Ge et al.1999). They are classified as AA, BB, CC, EE, FF and GGfor diploid genomes, and BBCC, CCDD, HHJJ and HHKKfor allotetraploid genomes (Table 1). Although severalspecies of Oryza have not been domesticated, with theexception of O. sativa and O. glaberrima, they containseveral genes for tolerance to both abiotic and bioticstresses (Stich et al. 1990). For the Oryza Map AlignmentProject (OMAP), representatives from each of these 10genome types were selected for bacterial artificial chromo-some (BAC) library construction, BAC end sequencing
and physical map construction (http://www.omap.org).From analysis of the OMAP data, Oryza comparativegenomics has given novel insights into Oryza genome evo-lution and genome size variation (Ammiraju et al. 2008).
P. coarctata Tateoka (= O. coarctata) is a tetraploid(2n = 4x = 48) monotypic genus.Tateoka (1965) classified O.coarctata as P. coarctata based on morphology and embryoanatomy (Flowers et al. 1990; Finch, Mcroberts & Bal-akrishna 1997). Garcia (1992) postulated its relation to O.meyeriana based on isozyme data. Latha et al. (1998) usedAFLP and RAPD molecular markers to assess the geneticrelationship between cultivated and wild rice species and P.coarctata. By generating and comparing two nuclear genetrees (Adh1 and Adh2) and a chloroplast gene tree (matK)of all rice species, Ge et al. (1999) inferred that Porteresiashows genome type HHKK supporting its inclusion inOryza species.According to the placing of P. coarctata in thedendrogram of Ge et al. (1999), a more ancient origin thanthat of O. sativa is suggested. The results are partly in accor-dance with phylogenetic reconstruction of rice genomes asascertained by Ge et al. (1999), which clearly shows that theclosest ancestral species to KK (P. coarctata) genome areDD and HH genomes.
Recent comparison of Monoculm1 (MOC1) genomicregions suggest that O. coarctata (or P. coarctata) has aunique genome type (Lu et al. 2009). Although most Oryzagenome types were determined by traditional genome ormolecular analysis (Li et al. 1964; Aggarwal et al. 1997), O.coarctata was designated as an HHKK genome type basedsolely on its phylogenetic position (Ge et al. 1999). Whenthe HH subgenomes in O. coarctata (HHKK) and O. ridleyi(HHJJ) were compared, no homology was observed inthe intergenic regions. These findings contrast with other
Table 1. Origin and genome types ofOryza genera analysed so far (Ge et al.1999)
Serial Species Genome Accession Origin
1 P. coarctata (Syn. O. coarctata) HHKK (?) MSSR007 India2 O. sativa AA IR64 IRRI3 O. glaberrima AA 100792 Senegal4 O. nivara AA 106185 India5 O. rufipogon AA 105908 Thailand6 O. longistaminata AA 103886 Tanzania7 O. punctata BB 100937 Ghana8 O. officinalis CC 101116 Philippines9 O. rhizomatis CC 105448 Sri Lanka
10 O. minuta BBCC 100880 Philippines11 O. eichingeri CC 105408 Sri Lanka12 O. malampuzhaensis BBCC 105328 India13 O. alta CCDD 100025 Surinam14 O. grandiglumis CCDD 105156 Brazil15 O. latifolia CCDD 105139 Guatemala16 O. australiensis EE 105272 Australia17 O. brachyantha FF 101232 Sierra Leone18 O. longiglumis HHJJ 105146 Indonesia19 O. ridleyi HHJJ 100820 Thailand20 O. granulata GG 101084 Sri Lanka21 O. meyeriana GG 106473 Philippines22 O. indandamanica unknown 105694 India
IRRI, International Rice Research Institute.
530 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

subgenome comparisons that show homologous sequencesand shared transposable elements in intergenic regions,such as the BB and CC genome types (Lu et al. 2009).Moreover, the gene sequence differences between the pre-dicted HH subgenome types in O. coarctata and O. ridleyiwere more different from AA and BB genome types. Bothof these subgenomes were estimated to have diverged fromeach other ~11 Ma. Hence, HH subgenomes in O. coarctataand O. ridleyi are likely to belong to different genome types.To avoid confusion in future research, Lu et al. (2009)suggested O. coarctata should be designated as KKLL.
ECOLOGICAL STATUS OF PORTERESIA
P. coarctata, though of great significance to estuarine anddeltaic environments, is very poorly understood ecologi-cally. Temporal and spatial patterns in growth and biomassproduction of P. coarctata were evaluated at selected locali-ties along the banks of Mandovi estuary, Goa, India (Jagtapet al. 2006). Considering ecological significance, tolerance towide salinity range and adaptability to sandy and muddysubstrate, P. coarctata is of a great value in protection, con-servation and restoration of estuarine and creek systems inIndia. The habitats of this plant in the country have beencategorized under ecologically sensitive zone, and pro-tected vide Coastal Regulation Zone (CRZ) Act of 1990,along with mangroves. However, Porteresia habitats con-tinue to be under constant threat from ever increasinganthropogenic demands, and hence warrants for strictimplementation of CRZ rule for their protection.
Studies into this wild relative of rice are now receivingattention because of its inherent capacity of tolerating highlevels of salinity. Being a wild rice relative it could serve asan intermediate agent for studying the model system inmonocotyledonous plants, expression and behavioralpattern during salinity stress. Specially, being tolerant tosea-water salinity in contrast to rice, it can serve as a ben-eficial ‘gene pool’ for generation of GM salt-tolerant ricethat will be a great help to the agriculture in Southeast Asia.
MORPHOLOGICAL FEATURES OFP. COARCTATA
P. coarctata has a distinctly different morphology from thatof cultivated rice. It has a highly differentiated undergroundrhizome system from the nodes of which a number of leafyshoots arise to make a complex association. Developmentof mechanical tissue is higher in Porteresia than in otherOryza species but the detailed anatomy has not beenstudied. Rhizoid-like thin rootlets arise from the rhizomeserving the purpose of absorption and anchorage. Thoughthe root system of this species does not penetrate deeplybecause of poor aeration of the substratum, the extensiverhizomatous system serves well to cope with absorptionneeds under high salinity. The leaves of Porteresia are waxyunlike those of rice, and the surfaces display alternate ridgesand furrows.The plant growth is more luxurious in its nativehabitat on the coastal mud flooded by saline water twice a
day. Under laboratory conditions, we have been able tosuccessfully propagate the plant on non-saline soil albeit ata lower growth rate. Porteresia grow well submerged inflowing water indicating that it has submergence tolerancetrait similar to that of rice. The seeds produced by the plantdehisce prematurely. Normally the plant propagates invegetative mode.
While propagated in the controlled environment of theplant growth chamber using non-saline soil as growthmedium, Porteresia exhibits growth characteristics compa-rable with rice. However, while grown in different concen-trations of NaCl, Porteresia exhibits a considerably betterperformance in terms of growth, chlorophyll content andphotosynthetic efficiency compared with rice (Sengupta &Majumder 2009).
SALT HAIRS AND SALT EXCLUSION
Like other salt marsh grasses, Porteresia also excludes saltthrough salt hairs (Flowers et al. 1990). These hairs arestructurally simple unicellular trichomes without any dis-tinct basal and cap cells. X-ray analysis by Flowers et al.(1990) showed that the hair contents are actually the sourceof NaCl secretion. Two different types of hair are actuallyfound on upper and lower surface of leaves in Porteresia.The upper surface hairs are finger-shaped whereas thelower surface hairs are peg-like. We observed that the hairson upper surface of the leaf did not rupture at high saltconcentration though they continued to secrete higheramount of salt forming visible crystals on leaf surface. Thehair on the lower surface of leaf swell, rupture and collapsein high salt concentration, and regrow when there is less saltin the medium (Fig. 1). We hypothesized that during thesaline water level fluctuation in the estuarine ecosystem,there is a continuous and often diurnal variation of saltconcentration in the rhizosphere of the plant, which causesthe lower surface hairs fall and regrow. Though the twotypes of hair function differently, they essentially functiontogether in keeping the intercellular sodium ion concentra-tion and sodium: potassium ratio low as reflected in theirion content profile (Sengupta & Majumder 2009). Manymangrove species and halophytes have salt glands (Liphs-chitz et al. 1974) but unlike those, the salt hairs of wild riceare unicellular and bursts on high salinity. According toFlowers et al. (1990), Porteresia plants accumulate Na+ andCl- ions in leaves, but maintain a Na :K ratio as low as 0.7even after 6 weeks of growth in 25% artificial saline water(ASW) where the Na : K ratio was 34. This ability to main-tain a low Na : K ratio in the leaves is in part a consequenceof the secretion of ions from the leaves (Bal & Dutt 1986).The salt secreted by the gland is a significant proportion ofthe ions reaching the leaf.
ATTEMPTS OF IN VITRO REGENERATION OFP. COARCTATA
Various attempts of regeneration of P. coarctata has beenmade as the resources of wild rice germplasm are under
Porteresia as model for studying salt-stress biology 531
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

conservation. Attempts have been made to transfer thesalt-tolerant character to cultivated rice through widehybridization procedures (Jena 1994). Establishment ofthe regenerating material in a suitable medium, however,was difficult. Protoplast fusion was attempted in a study ofsomatic hybridization between the two species (Finchet al. 1990; Jelodar et al. 1999). However, the production ofsomatic hybrids of rice and related wild species has beenreported only where suspension cultures are available forboth parents (Hayashi, Kyozuka & Shimamoto 1988), whichwas difficult to establish in Porteresia.The inherent problemwith Porteresia, besides being recalcitrant, is its associationwith a fungal endophyte Acremonium sp. that caused con-siderable problems in establishing cultures (Ramanan, Bal-akrishna & Suryanarayanan 1996). The clonal propagationmethod showed some multiplication in WP-based mediumand has turned out to be ecotype specific (Latha et al. 1998).There are no reports on chromosome complementationstudies in P. coarctata except for the determination ofchromosome number (2n = 48). Somatic hybrid plantswere obtained following the electrofusion of rice (O. sativacv. Taipei) cell suspension-derived protoplast with non-dividing leaf protoplasts of Porteresia (Jelodar et al. 1999).One of the somatic hybrid lines was an amphiploid somatichybrid with 72 chromosomes containing full chromosomecomplement of both O. sativa and P. coarctata althoughadvancement of the line was not reported by the authors.The most fruitful method yet seems to be establishment ofa clonal propagation system from axillary buds as followedby Sengupta et al. (2008). A proper genotype independentregeneration system is a prerequisite in considering Porter-esia as a model system.
METABOLIC PATHWAYS ANDGENES/PROTEINS IN RELATION TO SALTTOLERANCE IN PORTERESIA
Inositol metabolic pathway in relation tostress tolerance
Synthesis and metabolism of inositol is by far the mostwidely studied aspect of Porteresia salt-tolerance biology.Metabolic fates of inositol have been shown to be closelylinked to salt-stress tolerance in this plant. Inositol, a ubiq-uitous six-carbon cyclohexane hexitol, and its methylatedderivative pinitol are implicated in stress responses having arole as osmolytes in several biological systems (Shen et al.1999). In addition to such functions, inositol and its deriva-tives are also implicated in a variety of other cellular pro-cesses including growth regulation, membrane biogenesis,signal transduction, ion channel physiology and membranedynamics (Loewus & Murthy 2000).
In all inositol producing organisms studied to date thisimportant cyclitol is produced via conversion of glucose-6phosphate to inositol-1 phosphate by the L-myo-inositol1-phosphate synthase (MIPS; Enzyme Commission EC5.5.1.4) involving NAD+ followed by dephosphorylationcatalysed by inositol monophosphatase (IMPase; EC
3.1.3.25) (Majumder et al. 2003). The MIPS enzyme or itsstructural gene INO1 has been reported from evolutionarydiverse sources, including higher plants and animals,parasites, fungi, green algae, bacteria and archaea, and thesame has been considered to be an ancient protein/genehaving an evolutionary conserved ‘core catalytic structure’(Majumder et al. 2003; Ghosh Dastidar et al. 2006a).
Several inositol-metabolizing pathways, leading to stressregulatory secondary metabolites play important role instress amelioration, notably in the tolerant genotypes. Forexample, inositol is methylated to pinitol by the inositolmethyl transferase (EC 2.1.1.4) coded by an IMT1 genein an S-adenosyl methionine (SAM)-dependent reaction.Several harsh-environment inhabitated plants are known toaccumulate pinitol in response to salt or drought stress(Sengupta et al. 2008). Another important metabolic fate ofinositol is production of raffinose family oligosaccharides(RFOs), namely the trisaccharide, raffinose and the tet-rasaccharide, stachyose. RFOs have diverse roles in plants,being used for the transport and storage of carbon and ascompatible solutes for protection against abiotic stress(Bachmann, Matile & Keller 1994; Haritatos, Keller &Turgeon 1996; Taji et al. 2002). They are synthesized fromsucrose by the sequential addition of galactose moietiesdonated by galactinol. Galactinol is formed from uridinediphosphate (UDP)-galactose and myo-inositol via theactivity of galactinol synthase (GS), which is considered akey regulator in the pathway (Keller & Pharr 1996). Expres-sion of GS is increased during drought and cold exposure inArabidopsis and GS overexpression leads to increaseddrought tolerance (Liu et al. 1998; Taji et al. 2002; Cunning-ham et al. 2003). In many plant species, raffinose is alsostored in high amounts in the seeds where it is thought tohave an additional role in desiccation tolerance (Obendorf1997). Rather than GS activity alone, the concentrationof the initial substrates like myo-inositol and sucrose areconsidered to control RFO accumulation in seeds (Karneret al. 2004).
L-ascorbic acid (AsA) is the major antioxidant in plantcells as it controls redox potential during environment-induced oxidative stress. In both plants and animals, AsA isimportant as a cofactor for numerous key enzymes involvedin photoprotection, metal and xenobiotic detoxification, cellcycle, cell wall growth and cell expansion (Smirnoff 2000;Smirnoff & Wheeler 2000; Franceschi & Tarlyn 2002). LeafAsA content can also modulate the expression of genesinvolved in plant defense and regulate genes that controldevelopment through hormone signalling (Pastori et al.2003). Production of plant AsA depends on cellular inositolcontent, which through the activity of myo-inositol mono-oxygenase is converted to D-glucuronate, precursor ofAsA. Thus, inositol monooxygenase is another metabolicfate of inositol important in stress amelioration in plant life.Details of the various pathways have been schematicallyrepresented in Fig. 2.
In Porteresia, the inositol metabolic pathways are mostwell-studied and explored through a number of possibleapproaches. Both MIPS coding INO1 and IMT1 genes have
532 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

been cloned from Porteresia and completely characterized.Their expressions under abiotic stresses were determined,and inositol/pinitol are established as the osmopro-tectant(s) of choice in Porteresia. Both the genes explicitlycontribute towards improved salt tolerance in transgenicsystems.
PcINO1, an unique adaptation tosaline environment
The MIPS coding gene from P. coarctata (PcINO1) standsunique among all the INO1 homologs known to date(Majee et al. 2004), being the only salt-tolerant MIPScoding gene known from any eukaryotic source. A37-amino acid stretch outside the catalytic domain hasbeen confirmed as the salt-tolerance determinant domainin PcINO1. A similar situation has been reported for theallene oxide cyclase homolog (‘mangrin’) from the man-grove, Bruguiera sexangula (Yamada et al. 2002), wherein astretch of 70 amino acid residues, different in its organiza-tion from the allene oxide cyclase from Arabidopsis orLycopersicum, conferred salt tolerance to Escherichia coli,yeast and tobacco cells.
PcINO1 is special in its amino acid sequence for a stretchof about 147 amino acids in the mid-portion (betweenamino acids 173 and 320) the other parts being completelyidentical with its O. sativa homolog OsINO1. Additionsof other amino acids in PcINO1 at positions 174–180(WCLSLAS), 185/186 (SS), 239 (Cys), 273 (Ser) and 304(Pro), and deletions of OsINO1 amino acids in PcINO1at positions 184–186 (DVI), 195–200 (NNVIKG) and 245(Asp) make the PcINO1 gene product as composed of atotal of 512 amino acids, longer than that of OsINO1 by 2amino acids.
Recombinant PcINO1 protein has a higher substrateaffinity compared with the recombinant OsINO1 proteins.The expressed PcINO1 protein or the corresponding puri-fied native enzyme exhibits uninhibited enzyme activity invitro up to a concentration of 500 mm NaCl in contrast toOsINO1, which was inhibited in increasing NaCl. The bac-terially expressed mutated gene product with the 37 aminoacid stretch (the so-called ‘salt-tolerance domain’) deletedwas catalytically active as the PcINO1 gene product, butshows inhibition of the enzyme activity in the presence ofincreasing NaCl concentrations similar to the OsINO1protein. Although the OsINO1 protein aggregates with
Figure 2. The different pathways for utilization of inositol in plants that are important in stress. The key genes under active studyin present laboratory involve MIPS itself, genes leading towards RFO family sugar synthesis, pinitol synthesis and L-ascorbic acidsynthesis. IMT, inositol methyl transferase; PHI, phosphohexoisomerase; UDP-glucose/galactose, uridine diphosphate glucose/galactose;P, phosphate; RFO, raffinose family oligosaccharide; MIOX, myo-inositol oxygenase; SuSy, sucrose synthase.
Porteresia as model for studying salt-stress biology 533
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

increasing salt concentration, the PcINO1 protein doesnot even at 500 mm NaCl. The effect of the 37-amino acidresidue stretch on the salt-induced structural plasticity orrigidity of PcINO1 was further explained. The presence of21 hydrophobic residues in clusters in OsINO1 comparedwith 15 such scattered residues in PcINO1 generallyexplains the higher surface hydrophobicity of OsINO1over PcINO1 in absence of salt. The excess exposure ofthe hydrophobic clefts in presence of salt in the cases ofOsINO1 makes it susceptible to aggregation as opposed toPcINO1, resistant to such salt-induced aggregation (GhoshDastidar et al. 2006b).As salts screen the electrostatic inter-actions on the protein surface, the hydrophobic interactionsare screened through protein oligomerization interaction.Hence, loss of MIPS activity in OsINO1 in presence ofsalt may be directly related to such oligomerization. Salt-induced surface hydrophobic exposure is accompanied byrearrangements of internally buried groups, the loss ofenzymatic function may also be caused by both restructur-ing of the catalytic site and physical blocking of the accesspath of the substrate by oligomerization (Ghosh Dastidaret al. 2006b). In addition to the identification of the37-amino acid residue stretch as the salt-tolerance determi-nant in PcINO1, it has also been demonstrated that whiledeletion of this stretch can render the PcINO1 protein salt-sensitive (Majee et al. 2004), the same can attribute salt-tolerance property to an otherwise salt-sensitive MIPS byreplacement of the corresponding amino acid stretch. Suchrecombination has resulted in salt tolerance of Os:PcINO1or Bj:PcINO1 hybrid gene products from O. sativa orBrassica juncea INO1 genes (Majee et al. 2004; GhoshDastidar et al. 2006b). In addition, MIPS gene organizationhas been studied in Porteresia in comparison with that ofOryza and unique features were revealed.Several stress-specific cis-elements are also recognized upstream of otherrecently isolated PcINO1 genes in contrast to OsINO1 tosignify the regulatory mechanism of inositol productionunder abiotic stress in Porteresia (unpublished results, thislaboratory).
Further, functional introgression of the PcINO1 geneconfers salt tolerance to evolutionary diverse organismsfrom prokaryotes to eukaryotes including crop plants albeitto a variable extent (Das-Chatterjee et al. 2006). A directcorrelation between unabated increased synthesis of inosi-tol under salinity stress by the PcINO1 gene product andsalt tolerance has been demonstrated for all the systemspointing towards the universality of the application acrossevolutionary divergent taxa. The gene is known to confersalt tolerance in transgenic yeast, tobacco, mustard and rice(Das-Chatterjee et al. 2006).
Sequence comparison between O. sativa and P. coarctatawith respect to INO1 gene sequence to determine the rela-tive divergence of the species, as well as sequence compari-sons between the different genes of O. sativa were made todetermine the relative divergence of the genes. Divergenceat the MIPS coding region between O. sativa and P. coarc-tata was analysed separately at synonymous (KS) and non-synonymous (KA) sites and was compared with that found
in other genes coding for enzymatic proteins sequenced inO. sativa and P. coarctata. The KA estimate for INO1 ascompared with other genes shows that rate of evolution israther high. The high KA values of the INO1 gene providefurther evidence that MIPS is a rapidly evolving enzyme(Ray et al., personal communication). Despite the highevolutionary rate of MIPS protein, active sites, signaturesequences and NAD+ binding domains are conservedamong MIPS from different species or among distantlyrelated lineages (Majumder et al. 2003).
Pinitol production in Porteresia: First IMT1reported from monocots
D-pinitol production in the halophyte Mesembryantheu-mum crystallinum (ice plant) requires increased flux ofcarbon from glucose 6-phosphate to myo-inositol via themyo-inositol 1-phosphate reaction (Ishitani et al. 1996).The first gene of the pathway, inositol-1-phosphatesynthase (MIPS), is transcriptionally upregulated (Ishitaniet al. 1996; Nelson et al. 1998) followed by an inositolmonophosphatase function. Utilizing increased amountsof inositol following stress, the enzyme myo-inositol O-methyltransferase (IMT1), generates D-ononitol (Vernon& Bohnert 1992). Finally, D-ononitol is converted intoD-pinitol by an epimerization reaction termed OEP, whichis yet to be characterized biochemically or genetically. Inthe ice plant, IMT1 is expressed only following salt stress(Vernon & Bohnert 1992; Nelson et al. 1998).
Figure 2 depicts the pathway for synthesis of inositol andits methylation to pinitol with the probable regulation sitesof the pathway. This pathway is innately connected to inosi-tol synthesis and phospholipid biosynthesis (Nikoloff &Henry 1991) and beyond that the synthesis of the methy-lated inositols is connected to the major flux of carbon inphotosynthetic cells (Sengupta et al. 2008).The methylationpathway also removes the product from general metabo-lism to form D-pinitol, an extremely stable end-product.The activities of IMT1 and OEP are not found in tobaccoand Arabidopsis (Vernon, Ostrem & Bohnert 1993; Ishitaniet al. 1996; Sheveleva et al. 1997); in fact, genes for theseenzymes seem to be missing in these species.
Methylation of inositol to pinitol by the IMT1 genecoded inositol methyl transferase (EC 2.1.1.40) is a SAM-dependent reaction and is closely linked to the methyl cycleactivated as a result of elevated photorespiration rate. TheIMT1 gene has so far been isolated only from the halo-phytic ice plant, M. crystallinum and P. coarctata. In both,salt-induced transcriptional induction of the IMT1 is coor-dinated with the INO1 gene for pinitol biosynthesis undersalt stress (Ishitani et al. 1996; Sengupta et al. 2008). In Por-teresia, the action of IMT1 is by relative availability of inosi-tol for the IMT1 reaction over other competing pathways ofutilization and the availability of SAM indirectly regulatedby the methionine production through the homocysteinemethyl transferase (HMT; EC 2.1.1.14). Incidentally HMTis also upregulated in salt-treated Porteresia. Several of
534 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

these steps are thought to play important role in stressperception and amelioration in Porteresia (Sengupta et al.2008).
In P. coarctata leaves, pinitol accumulates under salinitystress to a level ~10-fold of the control condition thatis equivalent to ~30 mmol g-1 fresh weight comparablewith the Nicotiana tabaccum transgenics overexpressingMcIMT1 (Sheveleva et al. 1997). A concomitant increase intranscript and protein levels of both PcINO1 and PcIMT1 isevidenced on stress application. Coordinated expression ofPcINO1 along with PcIMT1 indicates that in P. coarctata,accumulation of pinitol via inositol is a stress-regulatedpathway. In silico analysis confirms the absence of an IMT1homolog in Oryza genome, and PcIMT1 is identified asremotely related phylogenetically to the methyl transferasegene family in rice (Sengupta et al. 2008). The presenceof the ancient pinitol synthesizing protein/gene in a wildhalophytic rice is remarkable indicating that there mightbe other beneficial genes in this species that are lost inrice during the long domestication process.
Chloroplastic fructose-1,6-bisphosphatase(PcCFR) from Porteresia
In higher plants during photosynthetic CO2 fixation, thesynthesized carbohydrate gets balanced in one form orthe other depending on the need of the plant and theenvironmental conditions. Chloroplastic fructose-1,6-bisphosphatase (FruP2ase; EC 3.1.3.11), present in thestroma of the chloroplast, catalyses an irreversible reactionof hydrolysis of fructose-1,6-bisphosphate to fructose-6-phosphate and Pi, to regenerate the primary CO2 acceptormolecule, ribulose-1,5-bisphosphate. Interestingly, alongwith ribulose 1·5-bisphosphate carboxylase/oxygenase(Rubisco), FruP2ase has also been suggested to be one ofthe likely sites of inhibition of photosynthetic activity understress. Reduced activity of the chloroplast FruP2ase lowersthe photosynthetic activity and impairs plant growth.Trans-genic tobacco plants expressing less than 15% of the chlo-roplast FruP2ase compared with the wild-type plantsshowed reduced photosynthetic activity and growth sug-gesting that the enzyme is truly a limiting one in the Calvincycle (Kossmann et al. 1992). Miyagawa, Tamoi & Shigeoka(2001) reported an overexpression of a cyanobacterialFruP2ase protein in tobacco that could enhance photosyn-thesis and growth. Earlier studies in our laboratory showedthat the purified chloroplastic FruP2ase from Porteresiarecords in vitro salt tolerance as opposed to the Oryzaenzyme, which is protected from salt sensitivity by a numberof osmolytes (Ghosh et al. 2001). Further studies in ourlaboratory were carried out for cloning and bacterialexpression of the FruP2ase gene(s) from P. coarctata(Roxb.) Tateoka (PcCFR) and O. sativa (OsCFR), alongwith the biochemical characterization of the gene prod-uct(s) with special reference to their salt-tolerance prop-erty. Comparison between the nucleotide and deducedamino acid sequences of the two proteins revealed a differ-ence of only five amino acid residues, of which three serine
residues in PcCFR are replaced by Glu, Thr, Arg andalanine in OsCFR. The purified recombinant PcCFR isfound to retain its enzymatic activity even in presence of500 mm NaCl in vitro as opposed to OsCFR, which is inac-tivated at such salt concentration. The in vitro generatedpoint mutant proteins of PcCFR involving replacement ofindividual serine residues showed varied degrees of sensi-tivity towards high salt, with the maximum OsCFR likesalt-sensitivity effect in the triple mutant involving all threeserine residues suggesting a possible concerted role of theseserine residues in salt tolerance property of PcCFR. Subse-quently, transgenic tobacco plants with chloroplast targetedPcCFR and OsCFR were raised to evaluate the effect ofintrogression of the genes on growth and photosynthesis ofthese plants during salt stress (Chatterjee et al. unpublishedresults, this laboratory).
Serine-rich protein (PcSrp)
A cDNA clone encoding a serine-rich protein has beenisolated from the cDNA library of salt stressed roots of P.coarctata, and designated as P. coarctata serine-rich-protein(PcSrp) encoding gene. Northern analysis and in situmRNA hybridization revealed expression of PcSrp in thesalt-stressed roots and rhizomes of P. coarctata. However,no such expression was seen in the salt stressed leaves andin the unstressed tissues of root, rhizome and leaf, indicatingthat PcSrp is under the control of a salt-inducible, tissue-specific promoter. The transgenic finger millet plants(Eleusine coracana) and yeast (Saccharomyces cerevisiae),expressing PcSrp, exhibited increased tolerance to salinitystress (Mahalakshmi et al. 2006).
Vacuolar H+-ATPase (V-ATPase)
The ion pump, vacuolar H+-ATPase (V-ATPase) plays amajor role in the maintenance of cellular pH, and influencesthe transport of cations into the vacuoles of plant cells. AcDNA clone (PcVHA-cl) encoding the c subunit ofV-ATPase was isolated from P. coarctata. The DNAsequence of PcVHA-cl showed significant homology withV-ATPase subunit c of rice. The deduced amino acidsequence of PcVHA-cl and other reported c subunits werecompared, and sequence relationships have been drawn toascertain their genetic relatedness. Southern analysis sug-gested presence of multiple coding regions for subunit c inP. coarctata. Northern and western analyses of salt-treatedP. coarctata plants revealed that subunit c of V-ATPase isupregulated by NaCl treatment at both transcriptional andtranslational level (Senthilkumar et al. 2005).
Translation initiation factor 1 (PceIF1)
Accumulation of increased sodium ions are known to affectthe protein synthesis (Wyn Jones & Pollard 1983). Transla-tion initiation factor 1 (PceIF1) cDNA was isolated fromP. coarctata leaves subjected to 150 mm NaCl. Expressionof PceIF1 is upregulated by application of exogenous ABA
Porteresia as model for studying salt-stress biology 535
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

and by mannitol treatment. It has been suggested thatexpression levels of eIF1 transcripts might form a conve-nient indicator for monitoring a stress responsive mecha-nism operative in P. coarctata leaves (Latha et al. 2004).
Generation of ESTs- possible pathwaysoperative during salinity stress
Our interest in Porteresia is primarily supported by twofacts, the first being its salt-tolerance trait and the secondbeing its phylogenetic closeness to cultivated rice. Consid-eration of these two facts makes Porteresia immenselyvaluable to plant stress biologists for search for salt-tolerance genes. However, being genetically a relativelyunexplored species, most of the genes in P. coarctata arenot characterized. In the absence of a whole genomesequencing data, which by itself is a difficult task for thetetraploid species, snapshots of transcripts at a givenstressed condition serve important roles. In our experi-ence, majority of the Porteresia cDNAs exhibit a highsequence homology to Oryza homologs and henceprimers based on rice gene sequence often work well.However, the cDNAs show important sequence variationthat could be subtle (as in PcCFR) or gross (as inPcINO1) and the protein products may show relative tol-erance to salt in vitro in comparison with the ricehomolog. While analysing the upstream sequences ofknown genes like PcINO1, we detected a number ofstress-responsive regulatory elements, which may beinvolved in the regulation of gene expression for survivalunder stress (unpublished results, this laboratory). Suchobservations necessitate having either a cDNA or ESTlibrary at hand that might reflect a general picture ofupregulated transcripts under stress and thus a databaseof important genes playing decisive role in salt tolerancemay be generated. A large number of condition-specificpartial cDNAs could provide a picture of many pathwaysaltered under salt stress and such pathways might turn outto be immensely important in understanding the salt-tolerance mechanism. Many of the genes such as PceIF,V-ATPase and PcSrp have been reportedly isolated fromcDNA libraries of Porteresia and proved to have somerole in its salt tolerance. A salt-subtracted cDNA libraryobtained by the present authors’ group from Porteresia(GenBank Accession Nos. CO805164 to CO805662) is cur-rently being studied in order to identify the functionalgene network operative during salt tolerance.
Proteomics – an approach towardsbioprospecting possiblestress-regulatory genes
Wild rice germplasm is a valuable bioresource for identi-fication of candidate genes that serve an important role forimprovement of salt-tolerance properties of the cultivatedrice through biotechnology. Proteomic approach is anothertool to evaluate its potential as a source of salt-tolerant orupregulated proteins that may serve the plant under saltstress. The first approach of leaf proteomic analysis of P.coarctata as well as any wild rice under salt stress has beenattempted by Sengupta & Majumder (2009). P. coarctata isa facultative halophyte and in experimental conditionsits unabated survival was found to be associated withincreased synthesis of several groups of proteins. The func-tion of the identified proteins are of diverse kinds such as:protection of the photosystems from oxidative/hyperionicdamage and maintaining the ETS function; enhancingavailable catalytic sites of the main carbon-assimilatingenzyme Rubisco under low stromal CO2 concentration;shunting some active oxygen species to photorespiratorycarbon oxidation (PCO) cycle; savings of overall energycosts by favouring low-energy pathways; synthesizingosmotically active compounds; detoxifyng the system byremoving stress-generated alcohols; controlling the tran-scriptional regulatory network through stress-inducedtranscription factors; enhanced synthesis of chaperones touphold normal protein structure that can be altered duringhigh-stress regime; maintaining cellular integrity throughsupplying high amount of cell wall components and thusretaining normal to robust growth under stress (Sengupta& Majumder 2009). Some of the identified proteins canwell be functionally related to the physiology of Porteresiaunder stress (Fig. 3). Since the complete proteomic andgenomic information of rice is now available (Komatsuet al. 2003, Yu et al. 2002) proteomic study of Porteresia inmore detail would serve the purpose of identification ofnewer genes and unique salt tolerance mechanisms in theplant.
CONCLUDING REMARKS
In the immediate future, biotechnological improvement ofrice would require varied sources of new biotic and abioticstress tolerance genes and pathways. Such genes areexpected to come from the stress-tolerant landraces that
Figure 3. Porteresia coarctata is a significant model halophyte important for specially improving tolerance status of rice. (A) Porteresiagrowing in greenhouse along with rice plants. (B) Summary of some of the physiological mechanisms Porteresia adopt under salt stress;(a–c) exclusion of salt using salt hairs on upper (a) and lower (b,c) surface of leaf under high NaCl stress; (d) leaf biomass increases inPorteresia with increasing salt concentration and shows no sign of wilting upto 0.4 M NaCl for 27 d under controlled growth conditions.(e) Photosynthesis in Porteresia remains unabated under high salt concentration as indicated by chlorophyll fluorescence data.(f) Relative water content of Porteresia does not fall under 85% even at 0.4 M NaCl for 27 d. (C) Summary of the importantmolecular mechanisms and effectors for tolerance in Porteresia. Most of the mechanisms and effectors are under elaborate study andmay serve as potential tolerance improvement schemes for rice once fully characterized. EST, expressed sequence tag; PSII, photosystemII; RFO, raffinose family oligosaccharide; Rubisco, ribulose 1·5-bisphosphate carboxylase/oxygenase.
536 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

Porteresia as model for studying salt-stress biology 537
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

needs concerted efforts on selected species. P. coarctata hasthe potential to serve as an important source of salt-tolerance factors for rice, as indicated from the currenttrend of research. However, high ploidy level of Porteresiawith its consequent large genome size put major hindrancetowards accepting this plant as a good system for geneticstudies. Moreover, lack of a very dependable, genotypeindependent and reproducible regeneration system, recal-citrant seeds and vegetative mode of propagation limitthe use of Porteresia for routine laboratory use. However,despite the lack of annual habit to enable ease of experi-mentation, amenability to genetic transformation and plantregeneration, being the only facultative halophyte amongthe rice genera, Porteresia deserves attention as an impor-tant halophyte model akin to rice for future research. Astechnological developments come up, it would be necessaryto ascertain a correct phylogenic and evolutionary status forPorteresia in Poaceae as the existing taxonomic position isnot reliable and is yet to be supported by adequate molecu-lar evidence. Most of the genes isolated from Porteresiashow high sequence homology to the rice genes (Tables 2 &3), which should make the sequencing and annotation ofthis species comparatively easy in near future. From the
limited proteomic and other studies made so far in thislaboratory, we found several unidentified proteins andgenes from Porteresia that turn out to be stress-regulated.This plant could harbor a good number of new genes, as alsoindicated by recent comparison of Monoculm1 (MOC1)genomic regions by Lu et al. (2008) who considered P.coarctata having an unique genome type among the differ-ent Oryza species.
Keeping such unorthodox nature of this plant in mind,ecological study of this species also seems to be necessary,as there are indications of at least two ecotypes existing inIndia itself, that is, around the Sunderbans, Bay of Bengaland the Chennai coastal area in the southern part of thecountry. An efficient embryo rescue process is also neededto conserve the germplasm. Porteresia, as indicated earlier,is an endangered species in India, with constant diminish-ment of normal estuarine ecotone. The vegetation is enor-mously important to keep the mangrove rhizosphere intact.Thus, sustainable conservation of the natural habitat of P.coarctata, accompanied by appropriate steps for bio-prospecting can offer rice stress biology a considerableamount of new information and valuable genes beyondcurrent trend of research with Porteresia.
Table 2. The reported nucleotide sequences from Porteresia (Oryza) coarctata
Gene nomenclature NCBI accession
Oryza coarctata clone a0295K14 Monoculm1, Mlo family protein, aspartic proteinase nepenthesin-1precursor, microtubule-associated protein MAP65-1a, IQ calmodulin-binding motif family protein,EMB2261 putative, polygalacturonase precursor, exopolygalacturonase precursor, and putative RNApolymerase A(I) large subunit genes, complete cds; and unknown gene.
FJ032636
Oryza coarctata clone a0226N01 Monoculm1 gene, complete cds; and unknown genes FJ032635Oryza coarctata chloroplast fructose-1,6-bisphosphatase mRNA, complete cds; nuclear gene for
chloroplast productEU371109
Oryza coarctata inositol-1-phosphate synthase (PcINO1-2) gene, complete cds FJ237300Oryza coarctata inositol-1-phosphate synthase (PcINO1-1) gene, complete cds FJ237299Oryza coarctata inositol methyl transferase mRNA, partial cds EU240449Oryza coarctata chromosome 1 anonymous locus NP2 genomic sequence EF577768Oryza coarctata polA1 gene, intron 19, complete sequence, strain: W0551 AB366710Oryza coarctata alcohol dehydrogenase 2 mRNA, complete cds EU371995Oryza coarctata triosephosphate isomerase mRNA, complete cds EU371994Oryza coarctata phosphoenolpyruvate carboxylase mRNA, partial cds EU371116Oryza coarctata metallothionein type 3 (MT3) gene, complete cds EU121847Porteresia coarctata serine-rich protein mRNA, complete cds AF110148Oryza coarctata tRNA-Leu (trnL) gene, partial sequence; chloroplast AY792522Oryza coarctata promoter region, genomic sequence DQ278911Oryza coarctata voucher 104502 NADH dehydrogease subunit 1 (nad1) gene, intron 2 and partial cds;
mitochondrialAY507935
Porteresia coarctata inositol 1-phosphate synthase (PINO1) gene, complete cds AF412340Porteresia coarctata maturase (matK) gene, partial cds; chloroplast gene for chloroplast product AF148669Porteresia coarctata V-ATPase subunit c (PVA1) mRNA, complete cds AF286464Porteresia coarctata homeobox protein mRNA, partial sequence AF384375Porteresia coarctata translational initiation factor eIF1 mRNA, complete cds AF380357Porteresia coarctata metallothionein mRNA, complete cds AF257465Porteresia coarctata catA gene for catalase, partial cds AB014455Porteresia coarctata cytosolic fructose-1,6-bisphosphatase mRNA, complete cds AF218845Porteresia coarctata alcohol dehydrogenase II (Adh2) gene, exons 3 through 7, partial cds AF148628
AF148627AF148593
Porteresia coarctata histone H3 mRNA, complete cds AF109910Subtracted cDNA library from salt stressed leaf Oryza coarctata cDNA sequence CO805164 to CO805662
538 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542
During the last several years our understanding on plantresponses to a stressful environment mostly centeredaround studies on abiotic or biotic stress signalling and itsperception. Mechanistic insight towards other aspects ofplant stress biology desperately needs to be addressed. Onesuch aspect would be the structural studies on proteinsinvolved in stress adaptation and amelioration in naturallystress-tolerant genotypes in comparison with those from thestress-sensitive ones.The question as to whether all proteinsor at least some of them, playing critical functional roleunder a stress regime, are specifically designed to functionin a cellular environment characterized by tolerance tovarious abiotic stress factors, remains elusive as of now. Aninkling towards the importance of such investigations camefrom the studies of some of the proteins, for example,PcINO1 or PcCFR from Porteresia, which differ signifi-cantly from their Oryza homologs in terms of both thestructure and function. One approach towards this wouldbe a genomic and proteomic comparison of the glycophyteswith their halophytic models as has been attemptedbetween Arabidopsis and its halophytic relative, Thellung-iella (Bressan et al. 2001). A similar comparison betweenOryza and Porteresia is expected to lead to the discovery ofstructural homologs of Oryza genes/proteins that areadapted to functioning under stressed environment. Thishas been documented by the unprecedented reports ofseveral important genes and proteins from Porteresia aselaborated in this discussion.
Studies so far made in our laboratory indicate the patternof physiological and metabolic adaptation of Porteresiaduring salinity stress along with the possible effectorsinvolved in such adaptation (Fig. 3). It must be bornein mind that such studies are only the beginning in thisdirection and to be able to use Porteresia as a halophyticmodel for salt tolerance for rice, it is imperative to considerseveral other important factors. As indicated,some of theseare, analysis of complete sequence of the tetraploid genomeof P. coarctata and its comparison with its completelysequenced diploid counterpart, O. sativa; determination ofthe genetic origin and evolution of Porteresia along withidentification of its diploid parents; a detailed account of thetrancriptomic data under salt stress for Porteresia; identifi-cation of the stress-responsive metabolic pathways anddetailed account of the metabolome and finally, structuraland functional comparison of salt-stress associated proteinsbetween Oryza and Porteresia. All such studies requirea comparative genomic, proteomic and metabolomicapproach for identification of the regulatory circuitryinvolved in salt-stress adaptation of Porteresia, which willserve as a guideline for rice stress biology.
ACKNOWLEDGEMENTS
The work in the authors’ laboratory has been supported byresearch grants to A. L. Majumder from the Departmentof Biotechnology, Government of India. SSG has been
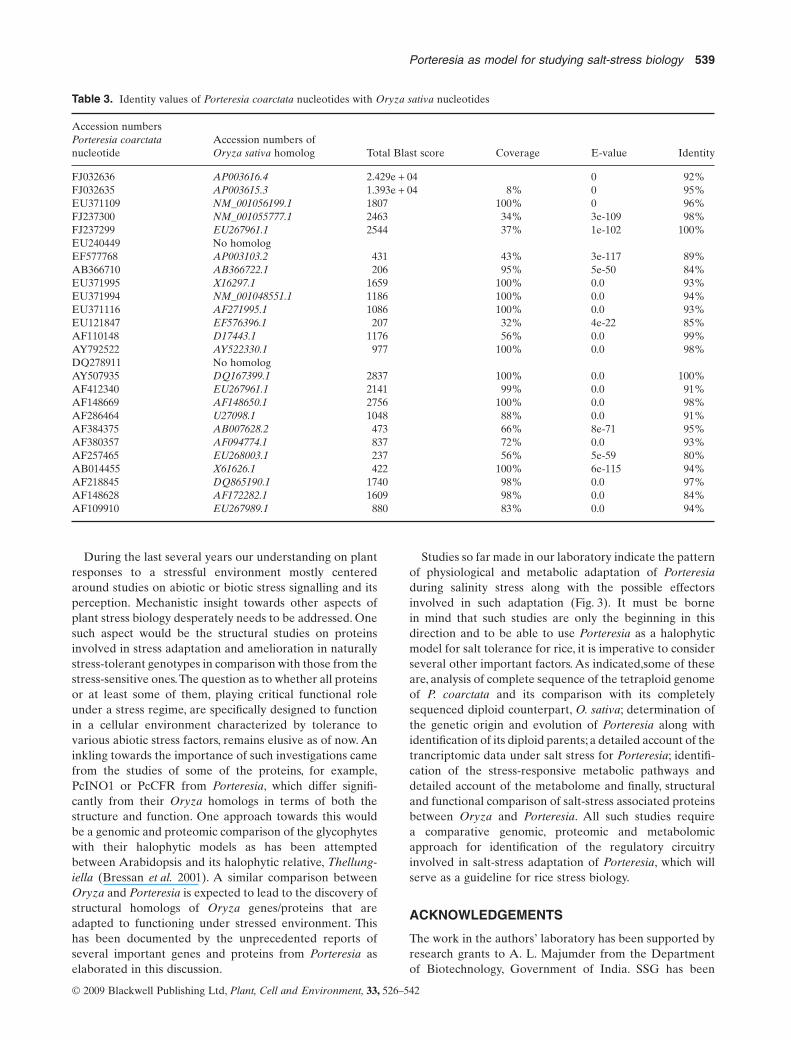
Table 3. Identity values of Porteresia coarctata nucleotides with Oryza sativa nucleotides
Accession numbersPorteresia coarctatanucleotide
Accession numbers ofOryza sativa homolog Total Blast score Coverage E-value Identity
FJ032636 AP003616.4 2.429e + 04 0 92%FJ032635 AP003615.3 1.393e + 04 8% 0 95%EU371109 NM_001056199.1 1807 100% 0 96%FJ237300 NM_001055777.1 2463 34% 3e-109 98%FJ237299 EU267961.1 2544 37% 1e-102 100%EU240449 No homologEF577768 AP003103.2 431 43% 3e-117 89%AB366710 AB366722.1 206 95% 5e-50 84%EU371995 X16297.1 1659 100% 0.0 93%EU371994 NM_001048551.1 1186 100% 0.0 94%EU371116 AF271995.1 1086 100% 0.0 93%EU121847 EF576396.1 207 32% 4e-22 85%AF110148 D17443.1 1176 56% 0.0 99%AY792522 AY522330.1 977 100% 0.0 98%DQ278911 No homologAY507935 DQ167399.1 2837 100% 0.0 100%AF412340 EU267961.1 2141 99% 0.0 91%AF148669 AF148650.1 2756 100% 0.0 98%AF286464 U27098.1 1048 88% 0.0 91%AF384375 AB007628.2 473 66% 8e-71 95%AF380357 AF094774.1 837 72% 0.0 93%AF257465 EU268003.1 237 56% 5e-59 80%AB014455 X61626.1 422 100% 6e-115 94%AF218845 DQ865190.1 1740 98% 0.0 97%AF148628 AF172282.1 1609 98% 0.0 84%AF109910 EU267989.1 880 83% 0.0 94%
Porteresia as model for studying salt-stress biology 539
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

supported by Research Fellowships from the Council ofIndustrial Research and the Department of Biotechnology,Government of India.
REFERENCES
Adams P., Nelson D.E., Yamada S., Chmara W., Jensen R.G.,Bohnert H.J. & Griffiths H. (1998) Growth and development ofMesembryanthemum crystallinum (Aizoaceae). New Phytologist138, 171–190.
Aggarwal R.K., Brar D.S. & Khush G.S. (1997) Two new genomesin the Oryza complex identified on the basis of molecular diver-gence analysis using total genomic DNA hybridization. Molecu-lar and General Genetics 254, 1–12.
Ammiraju J.S.S., Lu F., Sanyal A., et al. (2008) Dynamic evolutionof Oryza genomes is revealed by comparative genomic analysisof a genus-wide vertical data set. The Plant Cell 20, 3191–3209.
Amtmann A., Bohnert H.J. & Bressan R.A. (2005) Abiotic stressand plant genome evolution. Search for new models. PlantPhysiology 138, 127–130.
Bachmann M., Matile P. & Keller F. (1994) Metabolism of theraffinose family oligosaccharides in leaves of Ajuga reptans L.Cold acclimation, translocation, and sink to source transition:discovery of a chain elongation enzyme. Plant Physiology 105,1335–1345.
Bal A.R. & Dutt S.K. (1986) Mechanisms of salt tolerance in wildrice (Oryza coarctata Roxb. Plant and Soil 92, 399–404.
Bressan R.A., Zhang C., Zhang H., Hasegawa P.M., Bohnert H.J. &Zhu J-K. (2001) Learning from the Arabidopsis experience. Thenext gene search paradigm. Plant Physiolology 127, 1354–1360.
Clement C.R. (1999) 1492 and the loss of Amazonian crop geneticresources. Part 1. The relation between domestication andhuman population decline. Economic Botany 53, 188–202.
Cunningham S.M., Nadeau P., Castonguay Y., Laberge S. &Volenec J.J. (2003) Raffinose and stachyose accumulation, galac-tinol synthase expression, and winter injury of contrasting alfalfagermplasms. Crop Science 43, 562–570.
Cushman J.C. & Bohnert H.J. (2000) Genomic approaches to plantstress tolerance. Current Opinion in Plant Biology 3, 117–124.
Das-Chatterjee A., Goswami L., Maitra S., Ghosh Dastidar K., RayS. & Majumder A.L. (2006) Introgression of a novel salt-tolerantL-myo-inositol 1-phosphate synthase from Porteresia coarctata(Roxb.) Tateoka (PcINO1) confers salt tolerance to evolutionarydiverse organisms. FEBS Letters 580, 3980–3988.
Devos K.M., Beales J., Nagamura Y. & Sasaki T. (1999)Arabidopsis-rice: will colinearity allow gene prediction acrossthe Eudicot-Moncot Divide? Genome Research 9, 825–829.
Draper J., Mur L.A., Jenkins G., Ghosh-Biswas G.C., Bablak P.,Hasterok R. & Routledge A.P. (2001) Brachypodium dis-tachyon.A new model system for functional genomics in grasses.Plant Physiology 127, 1539–1555.
Finch R.P., Slamet I.H. & Cocking E.C. (1990) Production ofheterokaryons by the fusion of mesophyll protoplasts of P. coarc-tata and cell suspension-derived protoplasts of O. sativa: A newapproach to somatic hybridization in rice. Journal of PlantPhysiology 136, 592–598.
Finch R.P., Mcroberts N. & Balakrishna P. (1997) Molecular analy-sis of diversity in Porteresia coarctata. Rice BiotechnologyQuarterly 29, 10–15.
Flintham J.E., Börner A., Worland A.J. & Gale M.D. (1997) Opti-mizing wheat grain yield: effects of Rht (gibberellin-insensitive)dwarfing genes. The Journal of Agricultural Science 128, 11–25.
Flowers T.J., Hajibagheri M.A. & Clipson N.J.W. (1986) Halo-phytes. The Quarterly Review of Biology 61, 313–337.
Flowers T.J., Flowers S.A., Hajibagheri M.A. & Yeo A.R. (1990)Salt tolerance in the halophytic wild rice, Porteresia coarctata. T.New Phytologist 114, 675–684.
Franceschi V.R. & Tarlyn N.M. (2002) l-Ascorbic acid is accumu-lated in source leaf phloem and transported to sink tissues inplants. Plant Physiology 130, 649–656.
Gao J.-P., Chao D.-Y. & Lin H.-X. (2007) Understanding abioticstress tolerance mechanisms: recent studies on stress response inrice. Journal of Integrative Plant Biology 49, 742–750.
Garcia A. (1992) Salt tolerance in the halophytic wild rice, Porter-esia coarctata Tateoka. PhD Thesis, University of Sussex.
Gaut B.S. (2002) Evolutionary dynamics of grass genomes. NewPhytologist 154, 15–28.
Ge S., Sang T., Lu B.R. & Hong D.Y. (1999) Phylogeny of ricegenomes with emphasis on origins of allotetraploid species.Proceedings of the National Academy of Sciences of the UnitedStates of America 96, 14400–14405.
Ghosh S., Bagchi S. & Majumder A.L. (2001) Chloroplast fructose-1,6-bisphosphatase from Oryza differs in salt tolerance propertyfrom the Porteresia enzyme and is protected by osmolytes. PlantScience 160, 1171–1181.
Ghosh Dastidar K., Chatterjee A., Chatterjee A. & Majumder A.L.(2006a) Evolutionary divergence of L-myo-inositol 1-phosphatesynthase: significance of a ‘Core catalytic structure’. In Biology ofInositols and Phosphoinositides (eds A.L. Majumder & B.B.Biswas), Subcellular Biochemistry, Vol. 39, Chapter 13, pp. 313–340. Springer: The Netherlands.
Ghosh Dastidar K., Maitra S., Goswami L., Roy D., Das K.P. &Majumder A.L. (2006b) An insight into the molecular basis ofsalt-tolerance of L-myo inositol 1-phosphate synthase (PcINO1)from Porteresia coarctata (Roxb.) Tateoka, a halophytic wild rice.Plant Physiology 140, 1279–1296.
Haritatos E., Keller F. & Turgeon R. (1996) Raffinose oligosaccha-ride concentrations measured in individual cell and tissue typesin Cucumis melo L. leaves: implications for phloem loading.Planta 198, 614–622.
Hasegawa P.M., Bressan R.A., Zhu J.-K. & Bohnert H.J. (2000)Plant cellular and molecular responses to high salinity. AnnualReview of Plant Physiology and Plant Molecular Biology 51,463–499.
Hayashi Y., Kyozuka J. & Shimamoto K. (1988) Hybrids of rice(Oryza sativa L.) and wild Oryza species obtained by cell fusion.Molecular and General Genetics 214, 6–10.
Inan G., Zhang Q., Li P., et al. (2004) Salt cress. A halophyte andcryophyte arabidopsis relative model system and its applicabilityto molecular genetic analyses of growth and development ofextremophiles. Plant Physiology 135, 1–20.
Ingram J. & Bartels D. (1996) The molecular basis of dehydrationtolerance in plants. Annual Review of Plant Physiology and PlantMolecular Biology 47, 377–403.
Ishitani M., Majumder A.L., Bornhouser A., Michalowski C.B.,Jensen R.G. & Bohnert H.J. (1996) Coordinate transcriptionalinduction of myo-inositol metabolism during environmentalstress. Plant Journal 9, 537–548.
Jagtap T.G., Bhosale S. & Singh C. (2006) Characterization of Por-teresia coarctata beds along the Goa coast, India. Aquatic Botany84, 37–44.
Jelodar N.B., Blackhall N.W., Hartman T.P.V., Brar D.S., KhushG.S., Davey M.R., Cocking E.C. & Power J.B. (1999) Intergenericsomatic hybrids of rice (Oryza sativa L. + Porteresia coarctata(Roxb.) Tateoka). Theoretical and Applied Genetics 99, 570–577.
Jena K.K. (1994) Production of intergeneric hybrid between Orzyasativa L. and Porteresia coarctata T. Current Science 67, 744–746.
Karner U., Peterbauer T., Raboy V., Jones D.A., Hedley C.L. &Richter A. (2004) Myo-inositol and sucrose concentrations
540 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

affect the accumulation of raffinose family oligosaccharides inseeds. Journal of Experimental Botany 55, 1981–1987.
Keller F. & Pharr D.M. (1996) Metabolism of carbohydrates insinks and sources: galactosyl-sucrose oligosaccharides. In Photo-assimilate Distribution in Plants and Crops (eds E. Zamski &A.A. Schaffer), pp. 115–184. Marcel Dekker, New York, NY,USA.
Komatsu S., Konishi H., Shen S. & Yang G. (2003) Rice proteomics:a step toward functional analysis of the rice genome. Molecular& Cellular Proteomics 2, 2–10.
Kossmann J., Muller-Rober B., Dyer T.A., Raines C.A., SonnewaldU. & Willmitzer L. (1992) Cloning and expression analysis of theplastidic fructose-1,6-bisphosphatase coding sequence frompotato: circumstantial evidence for the import of hexoses intochloroplasts. Planta 188, 7–12.
Latha R., Anand A.K., Rao C.S., Eganathan P. & Balakrishna P.(1998) In vitro propagation of salt tolerant wild rice relative,Porteresia coarctata Tateoka. Plant Growth Regulation 17, 231–235.
Latha R., Hosseini Salekdeh G., Bennett J. & Swaminathan M.S.(2004) Molecular analysis of a stress-induced cDNA encodingthe translation initiation factor, eIF1, from the salt-tolerant wildrelative of rice, Porteresia coarctata. Functional Plant Biology 31,1035–1042.
Li H.W., Chen C.C., Wu H.K. & Lu K.C.L. (1964) Rice Genetics andCytogenetics (eds S. Tsunoda & N. Takahashi), pp. 118–131.Elsevier, Amsterdam, The Netherlands.
Liphschitz N., Shomer-Ilan A., Eshel A. & Waisel Y. (1974) Saltglands on leaves of Rhodes grass (Chloris gayana Kth.). Annalsof Botany 38, 459–462.
Liu J.J.J., Krenz D.C., Galvez A.F. & de Lumen B.O. (1998) Galac-tinol synthase (GS): increased enzyme activity and levels ofmRNA due to cold and desiccation. Plant Science 134, 11–20.
Loewus F.A. & Murthy P.P.N. (2000) myo-inositol metabolism inplants. Plant Science 150, 1–19.
Lu F., Ammiraju J.S.S., Sanyal A., et al. (2009) Comparativesequence analysis of MONOCULM1-orthologous regions in14 Oryza genomes. Proceedings of the National Academy ofSciences of the United States of America 106, 2071–2076.
Mahalakshmi S., Christopher G.S., Reddy T.P., Rao K.V. & ReddyV.D. (2006) Isolation of a cDNA clone (PcSrp) encoding serine-rich-protein from Porteresia coarctata T. and its expression inyeast and finger millet (Eleusine coracana L.) affording salttolerance. Planta 224, 347–359.
Majee M., Maitra S., Ghose Dastidar K., Pattanaik S., ChatterjeeA., Hait N., Das K.P. & Majumder A.L. (2004) A novel salt-tolerant L-myo-Inositol-1-phosphate synthase from Porteresiacoarctata (Roxb.) Tateoka, a halophytic wild rice. The Journal ofBiological Chemistry 279, 28539–28552.
Majumder A.L., Chatterjee A., Ghosh Dastidar K. & MajeeM. (2003) Diversification and evolution of L-myo-inositol 1-phosphate synthase. FEBS Letters 53, 3–10.
Miyagawa Y., Tamoi M. & Shigeoka S. (2001) Overexpression of acyanobacterial fructose-1,6-/sedoheptulose-1,7-bisphosphatasein tobacco enhances photosynthesis and growth. Nature Biotech-nology 19, 965–969.
Nelson D.E., Rammesmayer G. & Bohnert H.J. (1998) Regulationof cell-specific inositol metabolism and transport in plant salinitytolerance. The Plant Cell 10, 753–764.
Nikoloff D.M. & Henry S.A. (1991) Genetic analysis of yeast phos-pholipid biosynthesis. Annual Review of Genetics 25, 559–583.
Obendorf R.L. (1997) Oligosaccharides and galactosyl cyclitols inseed desiccation tolerance. Seed Science Research 7, 63–74.
Pastori G.M., Kiddle G., Antoniw J., Bernard S., Veljovic-JovanovicS., Verrier P.J., Noctor G. & Foyer C.H. (2003) Leaf vitamin Ccontents modulate plant defense transcripts and regulate genes
that control development through hormone signaling. The PlantCell 15, 939–951.
Paterson A.H., Lan T.H., Reischman K.P., et al. (1996) Towards aunified genetic map of higher plants, transcending the monocot-dicot divergence. Nature Genetics 14, 380–382.
Ramanan B.V., Balakrishna P. & Suryanarayanan T.S. (1996)Search for seed borne endophytes in rice (Oryza sativa) and wildrice (Porteresia coarctata). Rice Biotechnology Quarterly 27, 7–8.
Sasaki T. & Antonio B.A. (2009) Plant genomics: sorghum insequence. Nature 457, 547–548.
Sasaki T. & Sederoff R.R. (2003) The rice genome and comparativegenomics of higher plants. Current Opinion in Plant Biology 6,97–100.
Sengupta S. & Majumder A.L. (2009) Insight into the salt tolerancefactors of a wild halophytic rice, Porteresia coarctata: a physi-ological and proteomic approach. Planta 229, 911–929.
Sengupta S., Patra B., Ray S. & Majumder A.L. (2008) Inositolmethyl transferase from a halophytic wild rice, Porteresia coarc-tata Roxb.(Tateoka): regulation of pinitol synthesis under abioticstress. Plant, Cell & Environment 31, 1442–1459.
Senthilkumar P., Jithesh M.N., Parani M., Rajalakshmi S., PraseethaK. & Parida A. (2005) Salt stress effects on the accumulation ofvacuolar H+-ATPase subunit c transcripts in wild rice, Porteresiacoarctata (Roxb.) Tateoka. Current Science 89, 1386–1394.
Shen B., Hohmann S., Jensen R.G. & Bohnert H.J. (1999) Roles ofsugar alcohols in osmotic stress adaptation. Replacement of glyc-erol by mannitol and sorbitol in yeast. Plant Physiology 121,45–52.
Sheveleva E., Chmara W., Bohnert H.J. & Jensen R.G. (1997)lncreased salt and drought tolerance by D-ononitol productionin transgenic Nicotiana tabacum. Plant Physiology 115, 1211–1219.
Smirnoff N. (2000) Ascorbic acid: metabolism and functions of amultifaceted molecule. Current Opinion in Plant Biology 3, 229–235.
Smirnoff N. & Wheeler G.L. (2000) Ascorbic acid in plants: bio-synthesis and function. Critical Reviews in Biochemistry andMolecular Biology 351, 291–314.
Stich L.A., Dalmacio R.D., Elloran E., Romero G.O., AmanteA.D., Leung H., Nelson R. & Khush G.S. (1990) Wide hybrid-ization for rice improvement (abstract). Proceedings of the 4thannual meeting of the Rockefeller Foundation InternationalProgram on Rice Biotechnology. IRRI, Philippines, May 9–12.
Taji T., Seki M., Satou M., Sakurai T., Kobayashi M., Ishiyama K.,Narusaka Y., Narusaka M., Zhu J.K. & Shinozaki K. (2004)Comparative genomics in salt tolerance between Arabidopsisand Arabidopsis-related halophyte salt cress using Arabidopsismicroarray. Plant Physiology 135, 1697–1709.
Taji T., Chieko O., Satoshi I., Motoaki S., Mie K., Masatomo K.,Yamaguchi-Shinozaki K. & Shinozaki K. (2002) Important rolesof drought- and cold-inducible genes for galactinol synthase instress tolerance in Arabidopsis thaliana. The Plant Journal 29,417–426.
Tateoka T. (1965) Porteresia, a new genus of Graminae. Bulletin ofNational Science Museum, Tokyo 8, 405–406.
Vera-Estrella R., Barkla B.J., Garcia-Ramirez L. & Pantoja O.(2005) Salt stress in Thellungiella halophila activates Na+ trans-port mechanisms required for salinity tolerance. Plant Physiol-ogy 139, 1507–1517.
Vernon D.M. & Bohnert H.J. (1992) A novel methyl transferaseinduced by osmotic stress in the facultative halophyte M. crys-tallinum. EMBO Journal 11, 2077–2085.
Vernon D.M., Ostrem J.A. & Bohnert H.J. (1993) Stress perceptionand responses in a facultative halophyte: the regulation ofsalinity-induced genes in Mesembryanthemum crystallinum.Plant, Cell & Environment 16, 437–444.
Porteresia as model for studying salt-stress biology 541
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542

Wolfe K.H.,Gouy M.,Yang Y.W.,Sharp P.M. & Li W.H. (1989) Dateof the monocot-dicot divergence estimated from chloroplastDNA sequence data. Proceedings of the National Academy ofSciences of the United States of America 86, 6201–6205.
Wyn Jones R.G. & Pollard A. (1983) Proteins enzymes andPseudomonas syringae. In Encyclopedia of Plant Physiology(eds A. Lauchi & R.L. Bieleski), Springer- Gillaspie, eds, pp.528–562. Martinus Nijhoff Publishers Inc., College Park, MD,USA; Verlag, Berlin, Germany.
Yamada A., Saitoh T., Mimura T. & Ozeki Y. (2002) Expression
of mangrove allene oxide cyclase enhances salt tolerance inEscherichia coli, yeast, and tobacco cells. Plant Cell Physiology43, 903–910.
Yu J., Hu S., Wang J., et al. (2002) A draft sequence of the ricegenome (Oryza sativa L. ssp. indica). Science 296, 79–92.
Zhu J.-K. (2001) Plant salt tolerance. Trends in Plant Science 6,66–71.
Received 30 June 2009; received in revised form 20 September 2009;accepted for publication 22 September 2009
542 S. Sengupta & A. L. Majumder
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 33, 526–542




















