
(PDF) Textile dye wastewater characteristics and constituents ...
Upload
independentCategory
view
0download
0
Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2013 Article ID 371813 15 pageshttpdxdoiorg1011552013371813
Research ArticlePomegranate Bioactive Constituents Suppress CellProliferation and Induce Apoptosis in an Experimental Model ofHepatocellular Carcinoma Role of Wnt120573-CateninSignaling Pathway
Deepak Bhatia1 Roslin J Thoppil1 Animesh Mandal1 Karishma A Samtani1
Altaf S Darvesh1 and Anupam Bishayee2
1 Cancer Therapeutics and Chemoprevention Group Department of Pharmaceutical Sciences College of PharmacyNortheast Ohio Medical University Rootstown OH 44272 USA
2Department of Pharmaceutical Sciences School of Pharmacy American University of Health Sciences Signal Hill CA 90755 USA
Correspondence should be addressed to Anupam Bishayee abishayeeauhsedu
Received 3 January 2013 Accepted 12 February 2013
Academic Editor Ari M Mackler
Copyright copy 2013 Deepak Bhatia et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related death worldwide and chemoprevention representsa viable approach in lowering the mortality of this disease Pomegranate fruit an abundant source of anti-inflammatoryphytochemicals is gaining tremendous attention for its wide-spectrum health benefitsWe previously reported that a characterizedpomegranate emulsion (PE) prevents diethylnitrosamine (DENA)-induced rat hepatocarcinogenesis though inhibition of nuclearfactor-kappaB (NF-120581B) Since NF-120581B concurrently induces Wnt120573-catenin signaling implicated in cell proliferation cell survivaland apoptosis evasion we examined antiproliferative apoptosis-inducing and Wnt120573-catenin signaling-modulatory mechanismsof PE during DENA rat hepatocarcinogenesis PE (1 or 10 gkg) was administered 4 weeks before and 18 weeks following DENAexposure There was a significant increase in hepatic proliferation (proliferating cell nuclear antigen) and alteration in cell cycleprogression (cyclinD1) due toDENA treatment and PE dose dependently reversed these effects PE substantially induced apoptosisby upregulating proapoptotic protein Bax and downregulating antiapoptotic protein Bcl-2 PE dose dependently reduced hepatic120573-catenin and augmented glycogen synthase kinase-3120573 expression Our study provides evidence that pomegranate phytochemicalsexert chemoprevention of hepatic cancer through antiproliferative and proapoptotic mechanisms by modulating Wnt120573-cateninsignaling PE thus targets two interconnected molecular circuits (canonical NF-120581B and Wnt120573-catenin pathways) to exertchemoprevention of HCC
1 Introduction
Hepatocellular carcinoma (HCC) the major primary malig-nant tumor of the liver is one of the most life-threateninghuman cancers in the world resulting in almost one milliondeaths every year [1] There has been a drastic surge (70increase) in the incidence of HCC in the United States duringthe last quarter century with nearly 29000 new cases andmore than 20000 deaths expected to occur in 2012 alone[2] Lack of effective diagnostic tools for early detection andlimited treatment options for patients with advanced HCC
contribute to a dismal prognosis coupled with high mortalityfor this disease A critical need exists for the discoveryand development of novel preventive as well as therapeuticstrategies to combat the current morbidity and mortalityassociated with HCC [3ndash6]
Persistent oxidative stress and unresolved inflammationare two primary driving forces behind the development andexacerbation of HCC [7ndash9] Lingering infection with hepati-tis B virus (HBV) and hepatitis C virus (HCV) represents themajor risk factor forHCC [10] Chronic liver disease has beenproposed to be a predisposing factor for at least 80 of HCC
2 Evidence-Based Complementary and Alternative Medicine
cases [11] As a matter of fact any condition that is linked todevelopment of fibrosis and cirrhosis is strongly associatedwith the occurrence of HCC In response to hepatocyteinjury due to various factors including inflammation andoxidative stress hepatic stellate cells and portal fibroblastsundergo activation and transformation resulting in fibrosisand ultimately cirrhosis [12] Under these circumstances thesurviving hepatocytes proliferate to regenerate the injuredliver This cellular proliferation in the background of sus-tained inflammation and oxidative stress embodies a drivingforce for hepatic tumorigenesis [13] Identification of cellularpathways necessary for the proliferation and survival ofmalignant cells in HCC not only aids in understandingthe pathophysiology diagnosis and progression but alsoprovides a valuable tool in designing effective prevention andintervention of the disease One such relevant pathway isthe Wnt120573-catenin signaling cascade which plays a decisiverole in cell fate favoring cell proliferation over apoptosisAccumulating evidence has shown that the canonicalWnt120573-catenin pathway is frequently activated in HCC and respon-sible for initiation and progression of the disease [13ndash16]and this pathway could be an attractive and viable target forchemoprevention and therapy of HCC [17ndash19]
The pomegranate (Punica granatum Punicaceae) aunique mystical and distinctive fruit is a native of theHimalayas in India and extensively cultivated in India IsraelSpain and United States In addition to its ancient historicaluses pomegranate is used in several systems of medicinefor a wide variety of ailments [20 21] The ldquosuperfruitrdquopomegranate is receiving substantial importance becauseof its powerful antioxidant and anti-inflammatory proper-ties attributed to polyphenolic components (such as antho-cyanins) hydrolysable tannins (eg ellagitannins and gal-lotannins) and condensed tannins (proanthocyanidins) [22ndash26] Pomegranate constituents have been shown to possessexceptional health effects such as protection against andortreatment of cancer neurodegenerative diseases inflam-mation ulcers diabetes dental ailments high cholesterolcardiovascular disease obesity bacterial infections erectiledysfunction and male infertility (reviewed in [27 28])Pomegranate extracts and purified phytochemicals suppressthe proliferation of human breast prostate lung and coloncancer cells in vitro as well as prevent andor treat breast skinlung colon and prostate tumors in preclinical animalmodels(reviewed in [24 29ndash31]) Several phase II clinical trialshave linked oral consumption of pomegranate juice withsignificant prolongation of prostate-specific antigen (PSA)doubling time for men with prostate carcinoma with noaccompanying serious adverse effects [32 33]
Although pomegranate products have shown promisingantitumor activities in various organs of animals the chemo-preventive potential of pomegranate has not been investi-gated against the preclinical model of hepatic tumorigenesisuntil very recently Our laboratory has provided evidence forthe first time that pomegranate-derived phytoconstituentsexert a significant chemopreventive efficacy against dietaryhepatocarcinogen diethylnitrosamine (DENA)-induced livertumorigenesis in rats by potent antioxidant mechanismsmediated by upregulation of hepatic antioxidant and phase
2 genes [34] Very recently we have shown that suppressionof the inflammatory cascade through modulation of nuclearfactor-kappaB (NF-120581B) signaling pathway represents a novelmechanism of liver tumor inhibitory effects of pomegranatephytochemicals against experimental hepatocarcinogenesis[35] It is well known that NF-120581B the cardinal regulatorof inflammation acts as a hub for a number of intercon-nected signaling pathways implicated in malignancy suchas cell proliferation cell survival differentiation apoptosisinvasion angiogenesis and metastasis [36] Emerging evi-dence strongly suggests that NF-120581B may exhibit oncogenicpotential by coordinately activating the canonical Wnt120573-catenin signaling pathway [37ndash39] Functional proteomicsanalysis revealed that enhanced expression of Wnt-1 proteinassociated with NF-120581B might be an important mechanismof hepatitis B- and C-related HCC [40] Based on multipletargets of NF-120581B and the possibility of crosstalk between NF-120581B andWnt120573-catenin signaling pathways as well as previousreports from this laboratory [34 35] we have hypothesizedthat (i) pomegranate-mediated chemoprevention of exper-imental hepatocarcinogenesis could be achieved by inhibi-tion of abnormal hepatocyte proliferation and promotionof apoptosis and (ii) antiproliferative effects pomegranatephytochemicals may be linked to suppression of activatedWnt120573-catenin signaling The current study was thereforeinitiated to investigate the extent of cell proliferation andapoptosis during DENA-induced chemical rat liver carcino-genesis in the presence or absence of pomegranate treatmentThe involvement of Wnt120573-catenin signaling under the sameexperimental condition has also been examined by monitor-ing the expression of several key components of this pathwayas a possiblemechanism of pomegranate chemoprevention ofexperimental hepatic malignancy
2 Materials and Methods
21 Materials Pomegranate emulsion (PE) was purchasedfromRimonest Ltd Haifa IsraelWe previously provided thedetailed description of the preparation of this formulation[34] The chemical analyses of this product showed thepresence of mixed octadecatrienoic acids sterols and steroids(such as 17-120572-estradiol) tocol 120574-tocopherol in the lipid phaseand caffeic acid corilagin ellagic acid ferulic acid gallic acid5-hydroxymethylfurfural protocatechuic acid punicalagins(A and B) and trans-p-coumaric acid in the aqueous phase[34] Paraformaldehyde was procured from Ted Pella IncRedding CA Primary antibodies such asmousemonoclonalproliferating cell nuclear antigen (PCNA sc-56) rabbit poly-clonal cyclin D1 (sc-753) mouse monoclonal Bax (sc-70407)mouse monoclonal Bcl-2 (sc-7382) rabbit polyclonal 120573-catenin (sc-7199) rabbit polyclonal glycogen synthase kinase-3120573 (GSK-3120573 sc-9166) 120573-actin (sc-47778) and mouse andrabbit ABC staining systems were purchased from SantaCruz Biotechnology (Santa Cruz CA) TdT-FragEL DNAfragmentation detection assay kit was procured from EMDBiosciences Inc (San Diego CA) Pierce BCA protein assaykit was obtained from Thermo Scientific (Rockford IL)Quick RNAmini Prep kit and Verso cDNA synthesis kit were
Evidence-Based Complementary and Alternative Medicine 3
purchased from Zymo Research Corporation (Irvine CA)andThermo Scientific (Waltham MA) respectively
22 In Vivo Experimental Protocol and Tissue HarvestingLiver tissue used for all assays in this study was harvestedfrom our previously published chemopreventive study inwhich male Sprague-Dawley rats (Harlan Laboratories Indi-anapolis IN) fed with oral PE at a dose of 1 or 10 gkg bodyweight exhibited 26 or 50 inhibition of hepatic noduleincidence respectively [34] The animal experimentationwas carried out at the Northeast Ohio Medical University(Rootstown OH) following the animal protocol approved bythe Institutional Animal Care and Use Committee In brieffollowing an acclimatization period (7 days) the animalswere randomly divided into five groups While group Aanimals were maintained as untreated normal control groupB animals were administered with a sham emulsion (Rimon-est Ltd Haifa Israel) via oral gavage at a dose of 10 gkgthree timesweek (Monday Wednesday and Friday) Threeother animal groups (groups C D and E) were similarly fedwith PE at 1 gkg (groups C) or 10 gkg (groups D and E)The aforementioned oral feeding was continued for 4 weeksand then hepatocarcinogenesis was initiated in animals fromgroups B C and D by single intraperitoneal (ip) injectionof DENA (200mgkg) Following a recovery period of twoweeks phenobarbital (PB a well-known promoter of ratliver carcinogenesis) was introduced in the drinking water ofDENA-exposed animals at a concentration of 005 (wv)Oral feeding of rats with PE or sham emulsion was continuedtill the end of the animal experiment All animals weresacrificed 18 weeks following the DENA administration thatis 22 weeks after commencement of the study Liver tissuesfrom various rat groups were collected and either preservedin paraformaldehyde for immunohistochemical work orimmediately snap frozen in liquid nitrogen stored at ndash70∘Cand used for molecular studies as described in the following
23 Immunohistochemical Determination Serial sections ofliver tissue were prepared using a cryostat (Leica Microsys-tems Nussloch Germany) and used for immunohistochem-ical analysis of PCNA cyclin D1 Bax Bcl-2 120573-catenin andGSK-3120573 protein expressions following our published meth-ods [41] The presence of apoptotic cells in liver sections wasdetected by TdT-FragEL DNA fragmentation detection assaykit following the manufacturerrsquos protocol as we describedearlier [42]The immunohistochemical slides were visualizedunder a light microscope and 1000 hepatocytesanimalwere analyzed The labeling index (LI) and apoptotic index(AI) were expressed as the number of PCNA-positive andapoptotic cells per 100 hepatocytes respectively All otherimmunohistochemical results were expressed as percentageof immunopositive cells
24 Western Blot Analysis Frozen liver tissue samples werehomogenized in ice-cold RIPA lysis buffer (containing50mM Tris-HCl 1 Nonidet P-40 05 sodium deoxy-cholate 40mM NaF 10mM NaCl 10mM Na
3VO4 1 mM
phenylmethanesulfonyl fluoride and 10mM dithiothreitol)
to produce a 10 wv tissue homogenate The samplewas then centrifuged at 4∘C at 14000timesg for 20min Thesupernatant was collected and protein concentration wasestimated using the Pierce BCA protein assay kit followingthe accompanying instructions Equal amounts of proteinsamples were loaded and run on a 10 Tris-HCl gel (Bio-RadLaboratories Hercules CA) transferred onto a nitrocellulosemembrane and separately probed with anti-cyclin D1 anti-120573-catenin anti-GSK-3120573 or anti-120573-actin (all 1 1000 dilution)antibody The transferred proteins were visualized by anenhanced chemiluminescence detection system (Thermo Sci-entific Rockford IL) and analyzed using a Kodak analyzerLoading of equal amounts of protein was ensured by 120573-actin
25 Gene Expression Studies Total RNA fromsim20mg of liversample was extracted using Quick RNA mini Prep kit as perthe vendorrsquos protocol Complementary DNA (cDNA) wassynthesized from 1 120583g of total RNA using the Verso cDNAsynthesis kit following the manufacturerrsquos instructions Poly-merase chain reaction (PCR) was performed using specificprimers for rat cyclin D1 120573-catenin GSK-3120573 and glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) using followingprimer pairs cyclin D1 sense 51015840-GCGTACCCTGACACC-AATCT-31015840 antisense 51015840-GGCTCCAGAGACAAGAAA-CG 120573-catenin sense 51015840-GCCAGTGGATTCCGTACTGT-31015840 antisense 51015840-GAGCTTGCTTTCCTGATTGC GSK-3120573sense 51015840-CAAGCAGACACTCCCTGTGA-31015840 antisense 51015840-GTGGCTCCAAAGATCAGCTC GAPDH sense 51015840-AGA-CAGCCGCATCTTCTTGT-31015840 and antisense 51015840-TACTCA-GCACCAGCATCACC-31015840 These primer sequences weredesigned utilizing the Primer3 program and synthesized byEurofins MWG Operon (Huntsville AL) The PCR productswere analyzed by agarose gel electrophoresis and visualizedusing ethidium bromide staining GAPDH was used as thehousekeeping gene
26 Statistical Analysis All quantitative data reported hereare presented as mean plusmn standard error of mean (SEM)Significant differences among various animal groups weredetected by one-way ANOVA with post hoc analysis per-formed by the Student-Neuman-Keuls test A 119875 value lessthan 005 was considered statistically significant Statisticalanalysis and graphical representation of data were performedusing SigmaStat 31 software (Systat software Inc San JoseCA)
3 Results
31 Antiproliferative Cellular Mechanism of Pomegranatein DENA-Induced Hepatocarcinogenesis As shown in(Figure 1(A)andashd) the immunohistochemical staining depictsthe differential expression levels of the proliferative markerPCNA in liver sections obtained from the various experi-mental rat groups Near to complete absence of PCNA-positive cells was observed in both normal control(Figure 1(A)a) and only PE-treated (10 gkg) (figure notshown) groups An increase in hepatic PCNA expression wascontrastingly observed in animals treated with DENA alone
4 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(c) (d)(B)
PCN
A la
belin
g in
dex
()
0
10
20
30
40
50a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
Figure 1 Effects of PE on PCNA expression during DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemicallocalization of PCNA (magnification 100x) Arrows indicate immunohistochemical staining of PCNA Several groups are (a) normal control(b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of PCNA-positive cells based on 1000hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with normalgroup b119875 lt 0001 as compared with DENA control
(Figure 1(A)b) while PE treatment (1 gkg) in conjunctionwith DENA exposure resulted in insignificant decreases inPCNA-positive expression (Figure 1(A)c) These elevatedPCNA levels were shown to be subsequently reversedin the sections obtained from rats treated with PE at10 gkg in addition to DENA treatment (Figure 1(A)d)Figure 1(B) represents the quantitative analysis performedon the immunohistochemical data which shows significant(119875 lt 0001) elevation in the frequency of PCNA expressinghepatocytes (PCNA LI) in the DENA-treated animalscomparing to the normal control In comparison to theDENA-control rats PE treatment is shown to decrease thenumber of PCNA-positive cells Nevertheless a significant(119875 lt 0001) reduction was only achieved in rats treated withPE at the highest dose of 10 gkg
32 Differential Expression of Cell Cycle Specific GenemdashCyclinD1mdashin the Presence and Absence of Pomegranate Treatment inHepatocarcinogenesis Represented in Figure 2(A)andashd is theimmunocytochemical data depicting the expression of cellcycle specific gene cyclin D1 in hepatic sections procuredfrom several experimental animals Cyclin D1 was found tobe highly expressed in the DENA-control rats (Figure 2(A)b)whereas this observation was not made in the rat liversections obtained from the normal control (Figure 2(A)a)as well as PE control (figure not shown) groups A drasticreduction in the numbers of cyclin D1 immunoreactive cellswas recorded in rats treatedwith PE at 10 gkg (Figure 2(A)d)The subsequent quantitative analysis performed denotes asignificant (119875 lt 0001) increase in the percentage of cyclin
D1-positive cells in the rats administered with DENA withrespect to the normal control rats (Figure 2(B)) Elevatedimmunohistochemical expression of cyclin D1 was signif-icantly (119875 lt 0001) abrogated only in the rats treatedwith PE at the highest dose of 10 gkg when comparedto the DENA-challenged rats although reduction in thepercentage of immunopositive cells was noted in all PE-treated groups These data are further confirmed utilizingWestern blotting technique and RT-PCR (Figures 2(C) and2(D)) Hepatic cyclin D1mRNA and protein expression levelswere clearly upregulated in the DENA-treated animals whilePE treatment at both doses was found to consistently decreasetranscriptional and translational levels of cyclin D1
33 Proapoptotic Potential of Pomegranate in DENA-EvokedHepatocarcinogenesis The DNA fragmentation assay per-formed on the rat liver sections indicated almost no apop-totic cells in both the normal control group (Figure 3(A)a)and PE-administered group (figure not shown) Animalsexposed to DENA alone (Figure 3(A)b) were similarly foundto have absolutely no immunopositive cells which upontreatment with PE at both doses was found to be increased(Figure 3(A)c-d) Interestingly a substantial increase inapoptotic cells was noticed in liver sections obtained fromanimals treated with PE at 10 gkg (Figure 3(A)d) Quantita-tive analysis of immunohistochemical data revealed that thefrequency of apoptotic cells were significantly (119875 lt 0001)elevated in the rats subjected to PE treatment at 10 gkg incomparison to the animals administered with DENA alone(Figure 3(B))
Evidence-Based Complementary and Alternative Medicine 5
Cycli
n D
1 la
belin
g in
dex
()
0
2
4
6
8
10
12a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b(a)
(A)
(C) (D)
(B)
(b)
(c) (d)
Cyclin D1
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
Cyclin D1
GAPDH
Figure 2 Effects of PE on cyclin D1 expression during DENA-mediated hepatocarcinogenesis in rats (A) Representative immunohisto-chemical localization of cyclin D1 (magnification 100x) Arrows indicate immunohistochemical staining of cyclin D1 Several groups are (a)normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of cyclin D1-positive cellsbased on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared withnormal group b119875 lt 0001 as compared with DENA control (C) RepresentativeWestern blot indicating protein expression levels of cyclin D1in various experimental groups and (D) representative RT-PCR analysis of cyclin D1 expression in various groups of rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCR analysis using specific primer sequence TheGAPDH was used as the housekeeping gene
34 Regulation of Apoptosis-Related Proteins by Pomegranateduring Experimental Rat Liver Carcinogenesis Liver sectionsobtained from the different experimental rat groups wereimmunostained for the proapoptotic protein Bax and theantiapoptotic protein Bcl-2 and the results are shown inFigure 4(A)andashf Sections from rats exposed to DENA alonewere shown to have very limited expression of the proapop-totic protein Bax (Figure 4(A)a) while treatment with PE at1 gkg (Figure 4(A)b) and 10 gkg (Figure 4(A)c) reflected anincrease in the expression of Bax In stark contrast immunos-taining for the antiapoptotic protein Bcl-2 in the DENA-control liver sections yielded very high expression levels ofthe protein (Figure 4(A)d) which was subsequently reducedwith PE exposure at both doses (Figure 4(A)e-f) Figures 4(B)and 4(C) represent the affirmative corresponding quantitativeanalysis of the immunohistochemical data associated withBax and Bcl-2 respectively Although an increase in theproapoptotic protein Bax is seen in the PE-treated (1 gkg)rats significant (119875 lt 0001) increases in the percentageof Bax-positive cells were clearly observed in the animals
exposed to the highest dose of PE (10 gkg) when com-pared to the DENA-exposed rats (Figure 4(B)) Figure 4(C)indicates a very high expression of the antiapoptotic proteinBcl-2 in the hepatocytes of DENA-control rats as well as inthose of PE-treated (1 gkg) animals however treatment withPE at the highest dose (10 gkg) significantly (119875 lt 0001)decreased the increased expression of Bcl-2 A significant(119875 lt 0001) elevation in the BaxBcl-2 ratio was observedin the liver tissue harvested from the PE-treated (10 gkg) ratsin comparison to the DENA-exposed animals (Figure 4(D))
35 Reversal of DENA-Mediated Induction of Hepatic 120573-Catenin Expression by Pomegranate Based on immunohis-tochemical analysisvariations in the nuclear and cytoso-lic expressions of 120573-catenin were clearly observed in theliver sections harvested from several groups of animals(Figure 5(A)andashd) Expression of immunopositive cells wasclearly absent in sections obtained from both the normalcontrol (Figure 5(A)a) as well rats treated with only PE
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Evidence-Based Complementary and Alternative Medicine
cases [11] As a matter of fact any condition that is linked todevelopment of fibrosis and cirrhosis is strongly associatedwith the occurrence of HCC In response to hepatocyteinjury due to various factors including inflammation andoxidative stress hepatic stellate cells and portal fibroblastsundergo activation and transformation resulting in fibrosisand ultimately cirrhosis [12] Under these circumstances thesurviving hepatocytes proliferate to regenerate the injuredliver This cellular proliferation in the background of sus-tained inflammation and oxidative stress embodies a drivingforce for hepatic tumorigenesis [13] Identification of cellularpathways necessary for the proliferation and survival ofmalignant cells in HCC not only aids in understandingthe pathophysiology diagnosis and progression but alsoprovides a valuable tool in designing effective prevention andintervention of the disease One such relevant pathway isthe Wnt120573-catenin signaling cascade which plays a decisiverole in cell fate favoring cell proliferation over apoptosisAccumulating evidence has shown that the canonicalWnt120573-catenin pathway is frequently activated in HCC and respon-sible for initiation and progression of the disease [13ndash16]and this pathway could be an attractive and viable target forchemoprevention and therapy of HCC [17ndash19]
The pomegranate (Punica granatum Punicaceae) aunique mystical and distinctive fruit is a native of theHimalayas in India and extensively cultivated in India IsraelSpain and United States In addition to its ancient historicaluses pomegranate is used in several systems of medicinefor a wide variety of ailments [20 21] The ldquosuperfruitrdquopomegranate is receiving substantial importance becauseof its powerful antioxidant and anti-inflammatory proper-ties attributed to polyphenolic components (such as antho-cyanins) hydrolysable tannins (eg ellagitannins and gal-lotannins) and condensed tannins (proanthocyanidins) [22ndash26] Pomegranate constituents have been shown to possessexceptional health effects such as protection against andortreatment of cancer neurodegenerative diseases inflam-mation ulcers diabetes dental ailments high cholesterolcardiovascular disease obesity bacterial infections erectiledysfunction and male infertility (reviewed in [27 28])Pomegranate extracts and purified phytochemicals suppressthe proliferation of human breast prostate lung and coloncancer cells in vitro as well as prevent andor treat breast skinlung colon and prostate tumors in preclinical animalmodels(reviewed in [24 29ndash31]) Several phase II clinical trialshave linked oral consumption of pomegranate juice withsignificant prolongation of prostate-specific antigen (PSA)doubling time for men with prostate carcinoma with noaccompanying serious adverse effects [32 33]
Although pomegranate products have shown promisingantitumor activities in various organs of animals the chemo-preventive potential of pomegranate has not been investi-gated against the preclinical model of hepatic tumorigenesisuntil very recently Our laboratory has provided evidence forthe first time that pomegranate-derived phytoconstituentsexert a significant chemopreventive efficacy against dietaryhepatocarcinogen diethylnitrosamine (DENA)-induced livertumorigenesis in rats by potent antioxidant mechanismsmediated by upregulation of hepatic antioxidant and phase
2 genes [34] Very recently we have shown that suppressionof the inflammatory cascade through modulation of nuclearfactor-kappaB (NF-120581B) signaling pathway represents a novelmechanism of liver tumor inhibitory effects of pomegranatephytochemicals against experimental hepatocarcinogenesis[35] It is well known that NF-120581B the cardinal regulatorof inflammation acts as a hub for a number of intercon-nected signaling pathways implicated in malignancy suchas cell proliferation cell survival differentiation apoptosisinvasion angiogenesis and metastasis [36] Emerging evi-dence strongly suggests that NF-120581B may exhibit oncogenicpotential by coordinately activating the canonical Wnt120573-catenin signaling pathway [37ndash39] Functional proteomicsanalysis revealed that enhanced expression of Wnt-1 proteinassociated with NF-120581B might be an important mechanismof hepatitis B- and C-related HCC [40] Based on multipletargets of NF-120581B and the possibility of crosstalk between NF-120581B andWnt120573-catenin signaling pathways as well as previousreports from this laboratory [34 35] we have hypothesizedthat (i) pomegranate-mediated chemoprevention of exper-imental hepatocarcinogenesis could be achieved by inhibi-tion of abnormal hepatocyte proliferation and promotionof apoptosis and (ii) antiproliferative effects pomegranatephytochemicals may be linked to suppression of activatedWnt120573-catenin signaling The current study was thereforeinitiated to investigate the extent of cell proliferation andapoptosis during DENA-induced chemical rat liver carcino-genesis in the presence or absence of pomegranate treatmentThe involvement of Wnt120573-catenin signaling under the sameexperimental condition has also been examined by monitor-ing the expression of several key components of this pathwayas a possiblemechanism of pomegranate chemoprevention ofexperimental hepatic malignancy
2 Materials and Methods
21 Materials Pomegranate emulsion (PE) was purchasedfromRimonest Ltd Haifa IsraelWe previously provided thedetailed description of the preparation of this formulation[34] The chemical analyses of this product showed thepresence of mixed octadecatrienoic acids sterols and steroids(such as 17-120572-estradiol) tocol 120574-tocopherol in the lipid phaseand caffeic acid corilagin ellagic acid ferulic acid gallic acid5-hydroxymethylfurfural protocatechuic acid punicalagins(A and B) and trans-p-coumaric acid in the aqueous phase[34] Paraformaldehyde was procured from Ted Pella IncRedding CA Primary antibodies such asmousemonoclonalproliferating cell nuclear antigen (PCNA sc-56) rabbit poly-clonal cyclin D1 (sc-753) mouse monoclonal Bax (sc-70407)mouse monoclonal Bcl-2 (sc-7382) rabbit polyclonal 120573-catenin (sc-7199) rabbit polyclonal glycogen synthase kinase-3120573 (GSK-3120573 sc-9166) 120573-actin (sc-47778) and mouse andrabbit ABC staining systems were purchased from SantaCruz Biotechnology (Santa Cruz CA) TdT-FragEL DNAfragmentation detection assay kit was procured from EMDBiosciences Inc (San Diego CA) Pierce BCA protein assaykit was obtained from Thermo Scientific (Rockford IL)Quick RNAmini Prep kit and Verso cDNA synthesis kit were
Evidence-Based Complementary and Alternative Medicine 3
purchased from Zymo Research Corporation (Irvine CA)andThermo Scientific (Waltham MA) respectively
22 In Vivo Experimental Protocol and Tissue HarvestingLiver tissue used for all assays in this study was harvestedfrom our previously published chemopreventive study inwhich male Sprague-Dawley rats (Harlan Laboratories Indi-anapolis IN) fed with oral PE at a dose of 1 or 10 gkg bodyweight exhibited 26 or 50 inhibition of hepatic noduleincidence respectively [34] The animal experimentationwas carried out at the Northeast Ohio Medical University(Rootstown OH) following the animal protocol approved bythe Institutional Animal Care and Use Committee In brieffollowing an acclimatization period (7 days) the animalswere randomly divided into five groups While group Aanimals were maintained as untreated normal control groupB animals were administered with a sham emulsion (Rimon-est Ltd Haifa Israel) via oral gavage at a dose of 10 gkgthree timesweek (Monday Wednesday and Friday) Threeother animal groups (groups C D and E) were similarly fedwith PE at 1 gkg (groups C) or 10 gkg (groups D and E)The aforementioned oral feeding was continued for 4 weeksand then hepatocarcinogenesis was initiated in animals fromgroups B C and D by single intraperitoneal (ip) injectionof DENA (200mgkg) Following a recovery period of twoweeks phenobarbital (PB a well-known promoter of ratliver carcinogenesis) was introduced in the drinking water ofDENA-exposed animals at a concentration of 005 (wv)Oral feeding of rats with PE or sham emulsion was continuedtill the end of the animal experiment All animals weresacrificed 18 weeks following the DENA administration thatis 22 weeks after commencement of the study Liver tissuesfrom various rat groups were collected and either preservedin paraformaldehyde for immunohistochemical work orimmediately snap frozen in liquid nitrogen stored at ndash70∘Cand used for molecular studies as described in the following
23 Immunohistochemical Determination Serial sections ofliver tissue were prepared using a cryostat (Leica Microsys-tems Nussloch Germany) and used for immunohistochem-ical analysis of PCNA cyclin D1 Bax Bcl-2 120573-catenin andGSK-3120573 protein expressions following our published meth-ods [41] The presence of apoptotic cells in liver sections wasdetected by TdT-FragEL DNA fragmentation detection assaykit following the manufacturerrsquos protocol as we describedearlier [42]The immunohistochemical slides were visualizedunder a light microscope and 1000 hepatocytesanimalwere analyzed The labeling index (LI) and apoptotic index(AI) were expressed as the number of PCNA-positive andapoptotic cells per 100 hepatocytes respectively All otherimmunohistochemical results were expressed as percentageof immunopositive cells
24 Western Blot Analysis Frozen liver tissue samples werehomogenized in ice-cold RIPA lysis buffer (containing50mM Tris-HCl 1 Nonidet P-40 05 sodium deoxy-cholate 40mM NaF 10mM NaCl 10mM Na
3VO4 1 mM
phenylmethanesulfonyl fluoride and 10mM dithiothreitol)
to produce a 10 wv tissue homogenate The samplewas then centrifuged at 4∘C at 14000timesg for 20min Thesupernatant was collected and protein concentration wasestimated using the Pierce BCA protein assay kit followingthe accompanying instructions Equal amounts of proteinsamples were loaded and run on a 10 Tris-HCl gel (Bio-RadLaboratories Hercules CA) transferred onto a nitrocellulosemembrane and separately probed with anti-cyclin D1 anti-120573-catenin anti-GSK-3120573 or anti-120573-actin (all 1 1000 dilution)antibody The transferred proteins were visualized by anenhanced chemiluminescence detection system (Thermo Sci-entific Rockford IL) and analyzed using a Kodak analyzerLoading of equal amounts of protein was ensured by 120573-actin
25 Gene Expression Studies Total RNA fromsim20mg of liversample was extracted using Quick RNA mini Prep kit as perthe vendorrsquos protocol Complementary DNA (cDNA) wassynthesized from 1 120583g of total RNA using the Verso cDNAsynthesis kit following the manufacturerrsquos instructions Poly-merase chain reaction (PCR) was performed using specificprimers for rat cyclin D1 120573-catenin GSK-3120573 and glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) using followingprimer pairs cyclin D1 sense 51015840-GCGTACCCTGACACC-AATCT-31015840 antisense 51015840-GGCTCCAGAGACAAGAAA-CG 120573-catenin sense 51015840-GCCAGTGGATTCCGTACTGT-31015840 antisense 51015840-GAGCTTGCTTTCCTGATTGC GSK-3120573sense 51015840-CAAGCAGACACTCCCTGTGA-31015840 antisense 51015840-GTGGCTCCAAAGATCAGCTC GAPDH sense 51015840-AGA-CAGCCGCATCTTCTTGT-31015840 and antisense 51015840-TACTCA-GCACCAGCATCACC-31015840 These primer sequences weredesigned utilizing the Primer3 program and synthesized byEurofins MWG Operon (Huntsville AL) The PCR productswere analyzed by agarose gel electrophoresis and visualizedusing ethidium bromide staining GAPDH was used as thehousekeeping gene
26 Statistical Analysis All quantitative data reported hereare presented as mean plusmn standard error of mean (SEM)Significant differences among various animal groups weredetected by one-way ANOVA with post hoc analysis per-formed by the Student-Neuman-Keuls test A 119875 value lessthan 005 was considered statistically significant Statisticalanalysis and graphical representation of data were performedusing SigmaStat 31 software (Systat software Inc San JoseCA)
3 Results
31 Antiproliferative Cellular Mechanism of Pomegranatein DENA-Induced Hepatocarcinogenesis As shown in(Figure 1(A)andashd) the immunohistochemical staining depictsthe differential expression levels of the proliferative markerPCNA in liver sections obtained from the various experi-mental rat groups Near to complete absence of PCNA-positive cells was observed in both normal control(Figure 1(A)a) and only PE-treated (10 gkg) (figure notshown) groups An increase in hepatic PCNA expression wascontrastingly observed in animals treated with DENA alone
4 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(c) (d)(B)
PCN
A la
belin
g in
dex
()
0
10
20
30
40
50a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
Figure 1 Effects of PE on PCNA expression during DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemicallocalization of PCNA (magnification 100x) Arrows indicate immunohistochemical staining of PCNA Several groups are (a) normal control(b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of PCNA-positive cells based on 1000hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with normalgroup b119875 lt 0001 as compared with DENA control
(Figure 1(A)b) while PE treatment (1 gkg) in conjunctionwith DENA exposure resulted in insignificant decreases inPCNA-positive expression (Figure 1(A)c) These elevatedPCNA levels were shown to be subsequently reversedin the sections obtained from rats treated with PE at10 gkg in addition to DENA treatment (Figure 1(A)d)Figure 1(B) represents the quantitative analysis performedon the immunohistochemical data which shows significant(119875 lt 0001) elevation in the frequency of PCNA expressinghepatocytes (PCNA LI) in the DENA-treated animalscomparing to the normal control In comparison to theDENA-control rats PE treatment is shown to decrease thenumber of PCNA-positive cells Nevertheless a significant(119875 lt 0001) reduction was only achieved in rats treated withPE at the highest dose of 10 gkg
32 Differential Expression of Cell Cycle Specific GenemdashCyclinD1mdashin the Presence and Absence of Pomegranate Treatment inHepatocarcinogenesis Represented in Figure 2(A)andashd is theimmunocytochemical data depicting the expression of cellcycle specific gene cyclin D1 in hepatic sections procuredfrom several experimental animals Cyclin D1 was found tobe highly expressed in the DENA-control rats (Figure 2(A)b)whereas this observation was not made in the rat liversections obtained from the normal control (Figure 2(A)a)as well as PE control (figure not shown) groups A drasticreduction in the numbers of cyclin D1 immunoreactive cellswas recorded in rats treatedwith PE at 10 gkg (Figure 2(A)d)The subsequent quantitative analysis performed denotes asignificant (119875 lt 0001) increase in the percentage of cyclin
D1-positive cells in the rats administered with DENA withrespect to the normal control rats (Figure 2(B)) Elevatedimmunohistochemical expression of cyclin D1 was signif-icantly (119875 lt 0001) abrogated only in the rats treatedwith PE at the highest dose of 10 gkg when comparedto the DENA-challenged rats although reduction in thepercentage of immunopositive cells was noted in all PE-treated groups These data are further confirmed utilizingWestern blotting technique and RT-PCR (Figures 2(C) and2(D)) Hepatic cyclin D1mRNA and protein expression levelswere clearly upregulated in the DENA-treated animals whilePE treatment at both doses was found to consistently decreasetranscriptional and translational levels of cyclin D1
33 Proapoptotic Potential of Pomegranate in DENA-EvokedHepatocarcinogenesis The DNA fragmentation assay per-formed on the rat liver sections indicated almost no apop-totic cells in both the normal control group (Figure 3(A)a)and PE-administered group (figure not shown) Animalsexposed to DENA alone (Figure 3(A)b) were similarly foundto have absolutely no immunopositive cells which upontreatment with PE at both doses was found to be increased(Figure 3(A)c-d) Interestingly a substantial increase inapoptotic cells was noticed in liver sections obtained fromanimals treated with PE at 10 gkg (Figure 3(A)d) Quantita-tive analysis of immunohistochemical data revealed that thefrequency of apoptotic cells were significantly (119875 lt 0001)elevated in the rats subjected to PE treatment at 10 gkg incomparison to the animals administered with DENA alone(Figure 3(B))
Evidence-Based Complementary and Alternative Medicine 5
Cycli
n D
1 la
belin
g in
dex
()
0
2
4
6
8
10
12a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b(a)
(A)
(C) (D)
(B)
(b)
(c) (d)
Cyclin D1
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
Cyclin D1
GAPDH
Figure 2 Effects of PE on cyclin D1 expression during DENA-mediated hepatocarcinogenesis in rats (A) Representative immunohisto-chemical localization of cyclin D1 (magnification 100x) Arrows indicate immunohistochemical staining of cyclin D1 Several groups are (a)normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of cyclin D1-positive cellsbased on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared withnormal group b119875 lt 0001 as compared with DENA control (C) RepresentativeWestern blot indicating protein expression levels of cyclin D1in various experimental groups and (D) representative RT-PCR analysis of cyclin D1 expression in various groups of rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCR analysis using specific primer sequence TheGAPDH was used as the housekeeping gene
34 Regulation of Apoptosis-Related Proteins by Pomegranateduring Experimental Rat Liver Carcinogenesis Liver sectionsobtained from the different experimental rat groups wereimmunostained for the proapoptotic protein Bax and theantiapoptotic protein Bcl-2 and the results are shown inFigure 4(A)andashf Sections from rats exposed to DENA alonewere shown to have very limited expression of the proapop-totic protein Bax (Figure 4(A)a) while treatment with PE at1 gkg (Figure 4(A)b) and 10 gkg (Figure 4(A)c) reflected anincrease in the expression of Bax In stark contrast immunos-taining for the antiapoptotic protein Bcl-2 in the DENA-control liver sections yielded very high expression levels ofthe protein (Figure 4(A)d) which was subsequently reducedwith PE exposure at both doses (Figure 4(A)e-f) Figures 4(B)and 4(C) represent the affirmative corresponding quantitativeanalysis of the immunohistochemical data associated withBax and Bcl-2 respectively Although an increase in theproapoptotic protein Bax is seen in the PE-treated (1 gkg)rats significant (119875 lt 0001) increases in the percentageof Bax-positive cells were clearly observed in the animals
exposed to the highest dose of PE (10 gkg) when com-pared to the DENA-exposed rats (Figure 4(B)) Figure 4(C)indicates a very high expression of the antiapoptotic proteinBcl-2 in the hepatocytes of DENA-control rats as well as inthose of PE-treated (1 gkg) animals however treatment withPE at the highest dose (10 gkg) significantly (119875 lt 0001)decreased the increased expression of Bcl-2 A significant(119875 lt 0001) elevation in the BaxBcl-2 ratio was observedin the liver tissue harvested from the PE-treated (10 gkg) ratsin comparison to the DENA-exposed animals (Figure 4(D))
35 Reversal of DENA-Mediated Induction of Hepatic 120573-Catenin Expression by Pomegranate Based on immunohis-tochemical analysisvariations in the nuclear and cytoso-lic expressions of 120573-catenin were clearly observed in theliver sections harvested from several groups of animals(Figure 5(A)andashd) Expression of immunopositive cells wasclearly absent in sections obtained from both the normalcontrol (Figure 5(A)a) as well rats treated with only PE
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 3
purchased from Zymo Research Corporation (Irvine CA)andThermo Scientific (Waltham MA) respectively
22 In Vivo Experimental Protocol and Tissue HarvestingLiver tissue used for all assays in this study was harvestedfrom our previously published chemopreventive study inwhich male Sprague-Dawley rats (Harlan Laboratories Indi-anapolis IN) fed with oral PE at a dose of 1 or 10 gkg bodyweight exhibited 26 or 50 inhibition of hepatic noduleincidence respectively [34] The animal experimentationwas carried out at the Northeast Ohio Medical University(Rootstown OH) following the animal protocol approved bythe Institutional Animal Care and Use Committee In brieffollowing an acclimatization period (7 days) the animalswere randomly divided into five groups While group Aanimals were maintained as untreated normal control groupB animals were administered with a sham emulsion (Rimon-est Ltd Haifa Israel) via oral gavage at a dose of 10 gkgthree timesweek (Monday Wednesday and Friday) Threeother animal groups (groups C D and E) were similarly fedwith PE at 1 gkg (groups C) or 10 gkg (groups D and E)The aforementioned oral feeding was continued for 4 weeksand then hepatocarcinogenesis was initiated in animals fromgroups B C and D by single intraperitoneal (ip) injectionof DENA (200mgkg) Following a recovery period of twoweeks phenobarbital (PB a well-known promoter of ratliver carcinogenesis) was introduced in the drinking water ofDENA-exposed animals at a concentration of 005 (wv)Oral feeding of rats with PE or sham emulsion was continuedtill the end of the animal experiment All animals weresacrificed 18 weeks following the DENA administration thatis 22 weeks after commencement of the study Liver tissuesfrom various rat groups were collected and either preservedin paraformaldehyde for immunohistochemical work orimmediately snap frozen in liquid nitrogen stored at ndash70∘Cand used for molecular studies as described in the following
23 Immunohistochemical Determination Serial sections ofliver tissue were prepared using a cryostat (Leica Microsys-tems Nussloch Germany) and used for immunohistochem-ical analysis of PCNA cyclin D1 Bax Bcl-2 120573-catenin andGSK-3120573 protein expressions following our published meth-ods [41] The presence of apoptotic cells in liver sections wasdetected by TdT-FragEL DNA fragmentation detection assaykit following the manufacturerrsquos protocol as we describedearlier [42]The immunohistochemical slides were visualizedunder a light microscope and 1000 hepatocytesanimalwere analyzed The labeling index (LI) and apoptotic index(AI) were expressed as the number of PCNA-positive andapoptotic cells per 100 hepatocytes respectively All otherimmunohistochemical results were expressed as percentageof immunopositive cells
24 Western Blot Analysis Frozen liver tissue samples werehomogenized in ice-cold RIPA lysis buffer (containing50mM Tris-HCl 1 Nonidet P-40 05 sodium deoxy-cholate 40mM NaF 10mM NaCl 10mM Na
3VO4 1 mM
phenylmethanesulfonyl fluoride and 10mM dithiothreitol)
to produce a 10 wv tissue homogenate The samplewas then centrifuged at 4∘C at 14000timesg for 20min Thesupernatant was collected and protein concentration wasestimated using the Pierce BCA protein assay kit followingthe accompanying instructions Equal amounts of proteinsamples were loaded and run on a 10 Tris-HCl gel (Bio-RadLaboratories Hercules CA) transferred onto a nitrocellulosemembrane and separately probed with anti-cyclin D1 anti-120573-catenin anti-GSK-3120573 or anti-120573-actin (all 1 1000 dilution)antibody The transferred proteins were visualized by anenhanced chemiluminescence detection system (Thermo Sci-entific Rockford IL) and analyzed using a Kodak analyzerLoading of equal amounts of protein was ensured by 120573-actin
25 Gene Expression Studies Total RNA fromsim20mg of liversample was extracted using Quick RNA mini Prep kit as perthe vendorrsquos protocol Complementary DNA (cDNA) wassynthesized from 1 120583g of total RNA using the Verso cDNAsynthesis kit following the manufacturerrsquos instructions Poly-merase chain reaction (PCR) was performed using specificprimers for rat cyclin D1 120573-catenin GSK-3120573 and glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) using followingprimer pairs cyclin D1 sense 51015840-GCGTACCCTGACACC-AATCT-31015840 antisense 51015840-GGCTCCAGAGACAAGAAA-CG 120573-catenin sense 51015840-GCCAGTGGATTCCGTACTGT-31015840 antisense 51015840-GAGCTTGCTTTCCTGATTGC GSK-3120573sense 51015840-CAAGCAGACACTCCCTGTGA-31015840 antisense 51015840-GTGGCTCCAAAGATCAGCTC GAPDH sense 51015840-AGA-CAGCCGCATCTTCTTGT-31015840 and antisense 51015840-TACTCA-GCACCAGCATCACC-31015840 These primer sequences weredesigned utilizing the Primer3 program and synthesized byEurofins MWG Operon (Huntsville AL) The PCR productswere analyzed by agarose gel electrophoresis and visualizedusing ethidium bromide staining GAPDH was used as thehousekeeping gene
26 Statistical Analysis All quantitative data reported hereare presented as mean plusmn standard error of mean (SEM)Significant differences among various animal groups weredetected by one-way ANOVA with post hoc analysis per-formed by the Student-Neuman-Keuls test A 119875 value lessthan 005 was considered statistically significant Statisticalanalysis and graphical representation of data were performedusing SigmaStat 31 software (Systat software Inc San JoseCA)
3 Results
31 Antiproliferative Cellular Mechanism of Pomegranatein DENA-Induced Hepatocarcinogenesis As shown in(Figure 1(A)andashd) the immunohistochemical staining depictsthe differential expression levels of the proliferative markerPCNA in liver sections obtained from the various experi-mental rat groups Near to complete absence of PCNA-positive cells was observed in both normal control(Figure 1(A)a) and only PE-treated (10 gkg) (figure notshown) groups An increase in hepatic PCNA expression wascontrastingly observed in animals treated with DENA alone
4 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(c) (d)(B)
PCN
A la
belin
g in
dex
()
0
10
20
30
40
50a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
Figure 1 Effects of PE on PCNA expression during DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemicallocalization of PCNA (magnification 100x) Arrows indicate immunohistochemical staining of PCNA Several groups are (a) normal control(b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of PCNA-positive cells based on 1000hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with normalgroup b119875 lt 0001 as compared with DENA control
(Figure 1(A)b) while PE treatment (1 gkg) in conjunctionwith DENA exposure resulted in insignificant decreases inPCNA-positive expression (Figure 1(A)c) These elevatedPCNA levels were shown to be subsequently reversedin the sections obtained from rats treated with PE at10 gkg in addition to DENA treatment (Figure 1(A)d)Figure 1(B) represents the quantitative analysis performedon the immunohistochemical data which shows significant(119875 lt 0001) elevation in the frequency of PCNA expressinghepatocytes (PCNA LI) in the DENA-treated animalscomparing to the normal control In comparison to theDENA-control rats PE treatment is shown to decrease thenumber of PCNA-positive cells Nevertheless a significant(119875 lt 0001) reduction was only achieved in rats treated withPE at the highest dose of 10 gkg
32 Differential Expression of Cell Cycle Specific GenemdashCyclinD1mdashin the Presence and Absence of Pomegranate Treatment inHepatocarcinogenesis Represented in Figure 2(A)andashd is theimmunocytochemical data depicting the expression of cellcycle specific gene cyclin D1 in hepatic sections procuredfrom several experimental animals Cyclin D1 was found tobe highly expressed in the DENA-control rats (Figure 2(A)b)whereas this observation was not made in the rat liversections obtained from the normal control (Figure 2(A)a)as well as PE control (figure not shown) groups A drasticreduction in the numbers of cyclin D1 immunoreactive cellswas recorded in rats treatedwith PE at 10 gkg (Figure 2(A)d)The subsequent quantitative analysis performed denotes asignificant (119875 lt 0001) increase in the percentage of cyclin
D1-positive cells in the rats administered with DENA withrespect to the normal control rats (Figure 2(B)) Elevatedimmunohistochemical expression of cyclin D1 was signif-icantly (119875 lt 0001) abrogated only in the rats treatedwith PE at the highest dose of 10 gkg when comparedto the DENA-challenged rats although reduction in thepercentage of immunopositive cells was noted in all PE-treated groups These data are further confirmed utilizingWestern blotting technique and RT-PCR (Figures 2(C) and2(D)) Hepatic cyclin D1mRNA and protein expression levelswere clearly upregulated in the DENA-treated animals whilePE treatment at both doses was found to consistently decreasetranscriptional and translational levels of cyclin D1
33 Proapoptotic Potential of Pomegranate in DENA-EvokedHepatocarcinogenesis The DNA fragmentation assay per-formed on the rat liver sections indicated almost no apop-totic cells in both the normal control group (Figure 3(A)a)and PE-administered group (figure not shown) Animalsexposed to DENA alone (Figure 3(A)b) were similarly foundto have absolutely no immunopositive cells which upontreatment with PE at both doses was found to be increased(Figure 3(A)c-d) Interestingly a substantial increase inapoptotic cells was noticed in liver sections obtained fromanimals treated with PE at 10 gkg (Figure 3(A)d) Quantita-tive analysis of immunohistochemical data revealed that thefrequency of apoptotic cells were significantly (119875 lt 0001)elevated in the rats subjected to PE treatment at 10 gkg incomparison to the animals administered with DENA alone(Figure 3(B))
Evidence-Based Complementary and Alternative Medicine 5
Cycli
n D
1 la
belin
g in
dex
()
0
2
4
6
8
10
12a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b(a)
(A)
(C) (D)
(B)
(b)
(c) (d)
Cyclin D1
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
Cyclin D1
GAPDH
Figure 2 Effects of PE on cyclin D1 expression during DENA-mediated hepatocarcinogenesis in rats (A) Representative immunohisto-chemical localization of cyclin D1 (magnification 100x) Arrows indicate immunohistochemical staining of cyclin D1 Several groups are (a)normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of cyclin D1-positive cellsbased on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared withnormal group b119875 lt 0001 as compared with DENA control (C) RepresentativeWestern blot indicating protein expression levels of cyclin D1in various experimental groups and (D) representative RT-PCR analysis of cyclin D1 expression in various groups of rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCR analysis using specific primer sequence TheGAPDH was used as the housekeeping gene
34 Regulation of Apoptosis-Related Proteins by Pomegranateduring Experimental Rat Liver Carcinogenesis Liver sectionsobtained from the different experimental rat groups wereimmunostained for the proapoptotic protein Bax and theantiapoptotic protein Bcl-2 and the results are shown inFigure 4(A)andashf Sections from rats exposed to DENA alonewere shown to have very limited expression of the proapop-totic protein Bax (Figure 4(A)a) while treatment with PE at1 gkg (Figure 4(A)b) and 10 gkg (Figure 4(A)c) reflected anincrease in the expression of Bax In stark contrast immunos-taining for the antiapoptotic protein Bcl-2 in the DENA-control liver sections yielded very high expression levels ofthe protein (Figure 4(A)d) which was subsequently reducedwith PE exposure at both doses (Figure 4(A)e-f) Figures 4(B)and 4(C) represent the affirmative corresponding quantitativeanalysis of the immunohistochemical data associated withBax and Bcl-2 respectively Although an increase in theproapoptotic protein Bax is seen in the PE-treated (1 gkg)rats significant (119875 lt 0001) increases in the percentageof Bax-positive cells were clearly observed in the animals
exposed to the highest dose of PE (10 gkg) when com-pared to the DENA-exposed rats (Figure 4(B)) Figure 4(C)indicates a very high expression of the antiapoptotic proteinBcl-2 in the hepatocytes of DENA-control rats as well as inthose of PE-treated (1 gkg) animals however treatment withPE at the highest dose (10 gkg) significantly (119875 lt 0001)decreased the increased expression of Bcl-2 A significant(119875 lt 0001) elevation in the BaxBcl-2 ratio was observedin the liver tissue harvested from the PE-treated (10 gkg) ratsin comparison to the DENA-exposed animals (Figure 4(D))
35 Reversal of DENA-Mediated Induction of Hepatic 120573-Catenin Expression by Pomegranate Based on immunohis-tochemical analysisvariations in the nuclear and cytoso-lic expressions of 120573-catenin were clearly observed in theliver sections harvested from several groups of animals(Figure 5(A)andashd) Expression of immunopositive cells wasclearly absent in sections obtained from both the normalcontrol (Figure 5(A)a) as well rats treated with only PE
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
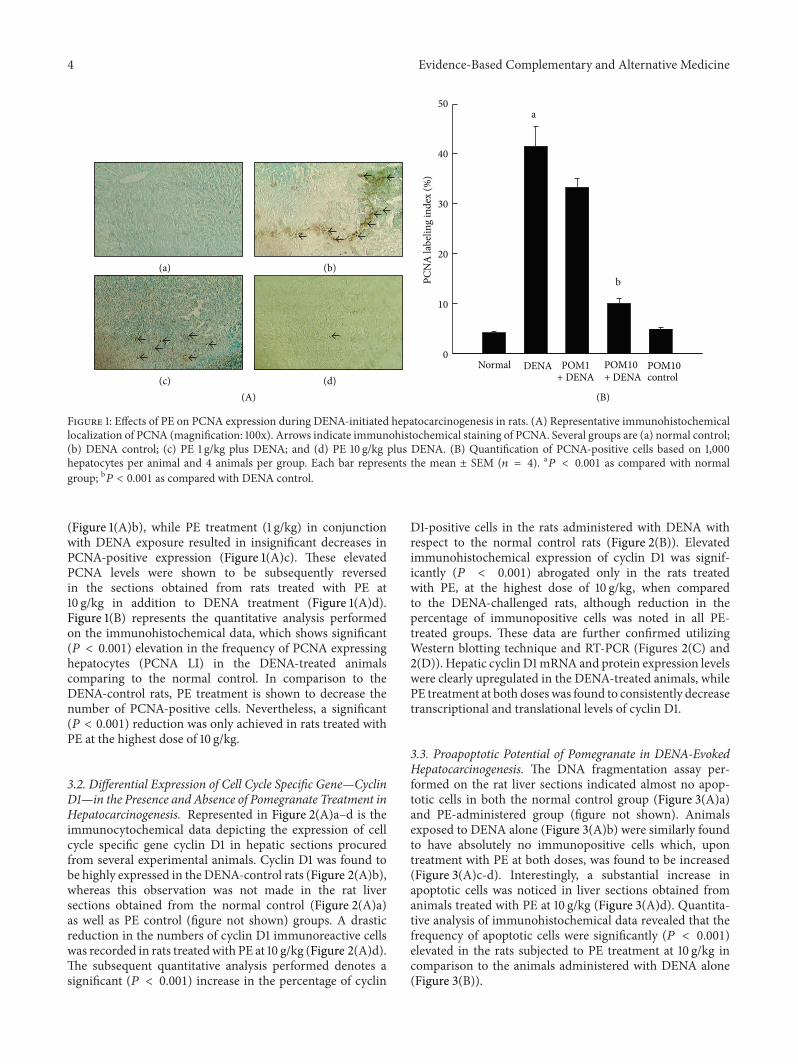
4 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(c) (d)(B)
PCN
A la
belin
g in
dex
()
0
10
20
30
40
50a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
Figure 1 Effects of PE on PCNA expression during DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemicallocalization of PCNA (magnification 100x) Arrows indicate immunohistochemical staining of PCNA Several groups are (a) normal control(b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of PCNA-positive cells based on 1000hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with normalgroup b119875 lt 0001 as compared with DENA control
(Figure 1(A)b) while PE treatment (1 gkg) in conjunctionwith DENA exposure resulted in insignificant decreases inPCNA-positive expression (Figure 1(A)c) These elevatedPCNA levels were shown to be subsequently reversedin the sections obtained from rats treated with PE at10 gkg in addition to DENA treatment (Figure 1(A)d)Figure 1(B) represents the quantitative analysis performedon the immunohistochemical data which shows significant(119875 lt 0001) elevation in the frequency of PCNA expressinghepatocytes (PCNA LI) in the DENA-treated animalscomparing to the normal control In comparison to theDENA-control rats PE treatment is shown to decrease thenumber of PCNA-positive cells Nevertheless a significant(119875 lt 0001) reduction was only achieved in rats treated withPE at the highest dose of 10 gkg
32 Differential Expression of Cell Cycle Specific GenemdashCyclinD1mdashin the Presence and Absence of Pomegranate Treatment inHepatocarcinogenesis Represented in Figure 2(A)andashd is theimmunocytochemical data depicting the expression of cellcycle specific gene cyclin D1 in hepatic sections procuredfrom several experimental animals Cyclin D1 was found tobe highly expressed in the DENA-control rats (Figure 2(A)b)whereas this observation was not made in the rat liversections obtained from the normal control (Figure 2(A)a)as well as PE control (figure not shown) groups A drasticreduction in the numbers of cyclin D1 immunoreactive cellswas recorded in rats treatedwith PE at 10 gkg (Figure 2(A)d)The subsequent quantitative analysis performed denotes asignificant (119875 lt 0001) increase in the percentage of cyclin
D1-positive cells in the rats administered with DENA withrespect to the normal control rats (Figure 2(B)) Elevatedimmunohistochemical expression of cyclin D1 was signif-icantly (119875 lt 0001) abrogated only in the rats treatedwith PE at the highest dose of 10 gkg when comparedto the DENA-challenged rats although reduction in thepercentage of immunopositive cells was noted in all PE-treated groups These data are further confirmed utilizingWestern blotting technique and RT-PCR (Figures 2(C) and2(D)) Hepatic cyclin D1mRNA and protein expression levelswere clearly upregulated in the DENA-treated animals whilePE treatment at both doses was found to consistently decreasetranscriptional and translational levels of cyclin D1
33 Proapoptotic Potential of Pomegranate in DENA-EvokedHepatocarcinogenesis The DNA fragmentation assay per-formed on the rat liver sections indicated almost no apop-totic cells in both the normal control group (Figure 3(A)a)and PE-administered group (figure not shown) Animalsexposed to DENA alone (Figure 3(A)b) were similarly foundto have absolutely no immunopositive cells which upontreatment with PE at both doses was found to be increased(Figure 3(A)c-d) Interestingly a substantial increase inapoptotic cells was noticed in liver sections obtained fromanimals treated with PE at 10 gkg (Figure 3(A)d) Quantita-tive analysis of immunohistochemical data revealed that thefrequency of apoptotic cells were significantly (119875 lt 0001)elevated in the rats subjected to PE treatment at 10 gkg incomparison to the animals administered with DENA alone(Figure 3(B))
Evidence-Based Complementary and Alternative Medicine 5
Cycli
n D
1 la
belin
g in
dex
()
0
2
4
6
8
10
12a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b(a)
(A)
(C) (D)
(B)
(b)
(c) (d)
Cyclin D1
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
Cyclin D1
GAPDH
Figure 2 Effects of PE on cyclin D1 expression during DENA-mediated hepatocarcinogenesis in rats (A) Representative immunohisto-chemical localization of cyclin D1 (magnification 100x) Arrows indicate immunohistochemical staining of cyclin D1 Several groups are (a)normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of cyclin D1-positive cellsbased on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared withnormal group b119875 lt 0001 as compared with DENA control (C) RepresentativeWestern blot indicating protein expression levels of cyclin D1in various experimental groups and (D) representative RT-PCR analysis of cyclin D1 expression in various groups of rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCR analysis using specific primer sequence TheGAPDH was used as the housekeeping gene
34 Regulation of Apoptosis-Related Proteins by Pomegranateduring Experimental Rat Liver Carcinogenesis Liver sectionsobtained from the different experimental rat groups wereimmunostained for the proapoptotic protein Bax and theantiapoptotic protein Bcl-2 and the results are shown inFigure 4(A)andashf Sections from rats exposed to DENA alonewere shown to have very limited expression of the proapop-totic protein Bax (Figure 4(A)a) while treatment with PE at1 gkg (Figure 4(A)b) and 10 gkg (Figure 4(A)c) reflected anincrease in the expression of Bax In stark contrast immunos-taining for the antiapoptotic protein Bcl-2 in the DENA-control liver sections yielded very high expression levels ofthe protein (Figure 4(A)d) which was subsequently reducedwith PE exposure at both doses (Figure 4(A)e-f) Figures 4(B)and 4(C) represent the affirmative corresponding quantitativeanalysis of the immunohistochemical data associated withBax and Bcl-2 respectively Although an increase in theproapoptotic protein Bax is seen in the PE-treated (1 gkg)rats significant (119875 lt 0001) increases in the percentageof Bax-positive cells were clearly observed in the animals
exposed to the highest dose of PE (10 gkg) when com-pared to the DENA-exposed rats (Figure 4(B)) Figure 4(C)indicates a very high expression of the antiapoptotic proteinBcl-2 in the hepatocytes of DENA-control rats as well as inthose of PE-treated (1 gkg) animals however treatment withPE at the highest dose (10 gkg) significantly (119875 lt 0001)decreased the increased expression of Bcl-2 A significant(119875 lt 0001) elevation in the BaxBcl-2 ratio was observedin the liver tissue harvested from the PE-treated (10 gkg) ratsin comparison to the DENA-exposed animals (Figure 4(D))
35 Reversal of DENA-Mediated Induction of Hepatic 120573-Catenin Expression by Pomegranate Based on immunohis-tochemical analysisvariations in the nuclear and cytoso-lic expressions of 120573-catenin were clearly observed in theliver sections harvested from several groups of animals(Figure 5(A)andashd) Expression of immunopositive cells wasclearly absent in sections obtained from both the normalcontrol (Figure 5(A)a) as well rats treated with only PE
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
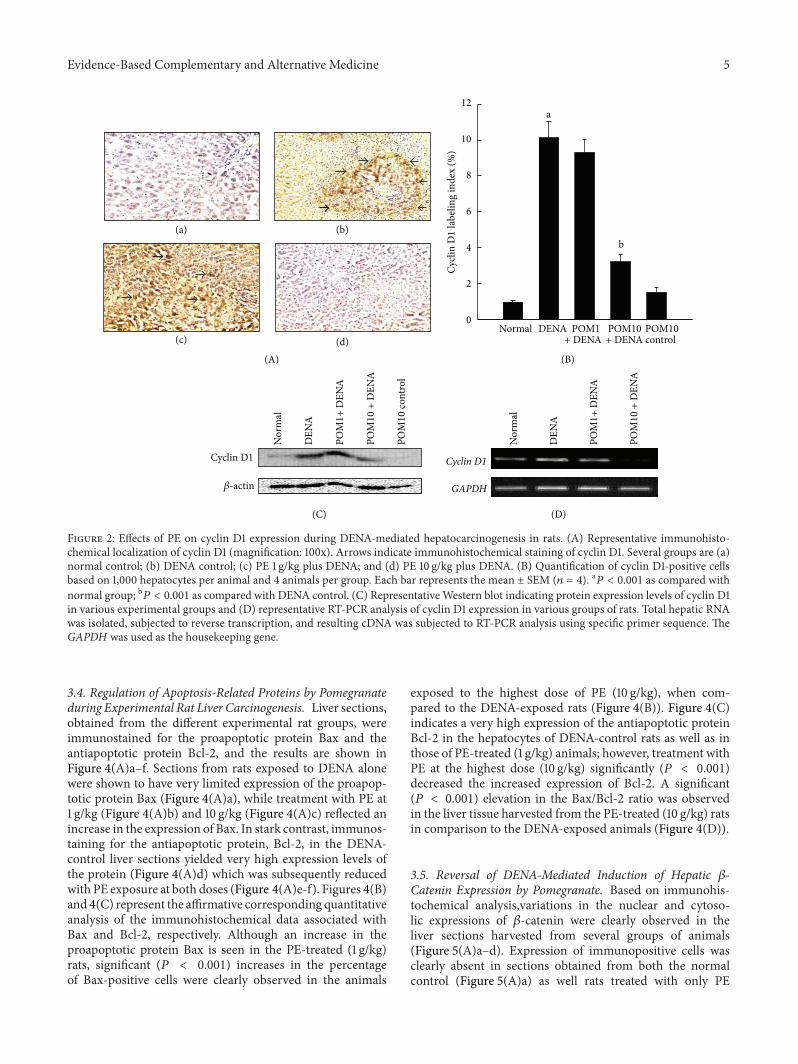
Evidence-Based Complementary and Alternative Medicine 5
Cycli
n D
1 la
belin
g in
dex
()
0
2
4
6
8
10
12a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b(a)
(A)
(C) (D)
(B)
(b)
(c) (d)
Cyclin D1
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
Cyclin D1
GAPDH
Figure 2 Effects of PE on cyclin D1 expression during DENA-mediated hepatocarcinogenesis in rats (A) Representative immunohisto-chemical localization of cyclin D1 (magnification 100x) Arrows indicate immunohistochemical staining of cyclin D1 Several groups are (a)normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of cyclin D1-positive cellsbased on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared withnormal group b119875 lt 0001 as compared with DENA control (C) RepresentativeWestern blot indicating protein expression levels of cyclin D1in various experimental groups and (D) representative RT-PCR analysis of cyclin D1 expression in various groups of rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCR analysis using specific primer sequence TheGAPDH was used as the housekeeping gene
34 Regulation of Apoptosis-Related Proteins by Pomegranateduring Experimental Rat Liver Carcinogenesis Liver sectionsobtained from the different experimental rat groups wereimmunostained for the proapoptotic protein Bax and theantiapoptotic protein Bcl-2 and the results are shown inFigure 4(A)andashf Sections from rats exposed to DENA alonewere shown to have very limited expression of the proapop-totic protein Bax (Figure 4(A)a) while treatment with PE at1 gkg (Figure 4(A)b) and 10 gkg (Figure 4(A)c) reflected anincrease in the expression of Bax In stark contrast immunos-taining for the antiapoptotic protein Bcl-2 in the DENA-control liver sections yielded very high expression levels ofthe protein (Figure 4(A)d) which was subsequently reducedwith PE exposure at both doses (Figure 4(A)e-f) Figures 4(B)and 4(C) represent the affirmative corresponding quantitativeanalysis of the immunohistochemical data associated withBax and Bcl-2 respectively Although an increase in theproapoptotic protein Bax is seen in the PE-treated (1 gkg)rats significant (119875 lt 0001) increases in the percentageof Bax-positive cells were clearly observed in the animals
exposed to the highest dose of PE (10 gkg) when com-pared to the DENA-exposed rats (Figure 4(B)) Figure 4(C)indicates a very high expression of the antiapoptotic proteinBcl-2 in the hepatocytes of DENA-control rats as well as inthose of PE-treated (1 gkg) animals however treatment withPE at the highest dose (10 gkg) significantly (119875 lt 0001)decreased the increased expression of Bcl-2 A significant(119875 lt 0001) elevation in the BaxBcl-2 ratio was observedin the liver tissue harvested from the PE-treated (10 gkg) ratsin comparison to the DENA-exposed animals (Figure 4(D))
35 Reversal of DENA-Mediated Induction of Hepatic 120573-Catenin Expression by Pomegranate Based on immunohis-tochemical analysisvariations in the nuclear and cytoso-lic expressions of 120573-catenin were clearly observed in theliver sections harvested from several groups of animals(Figure 5(A)andashd) Expression of immunopositive cells wasclearly absent in sections obtained from both the normalcontrol (Figure 5(A)a) as well rats treated with only PE
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
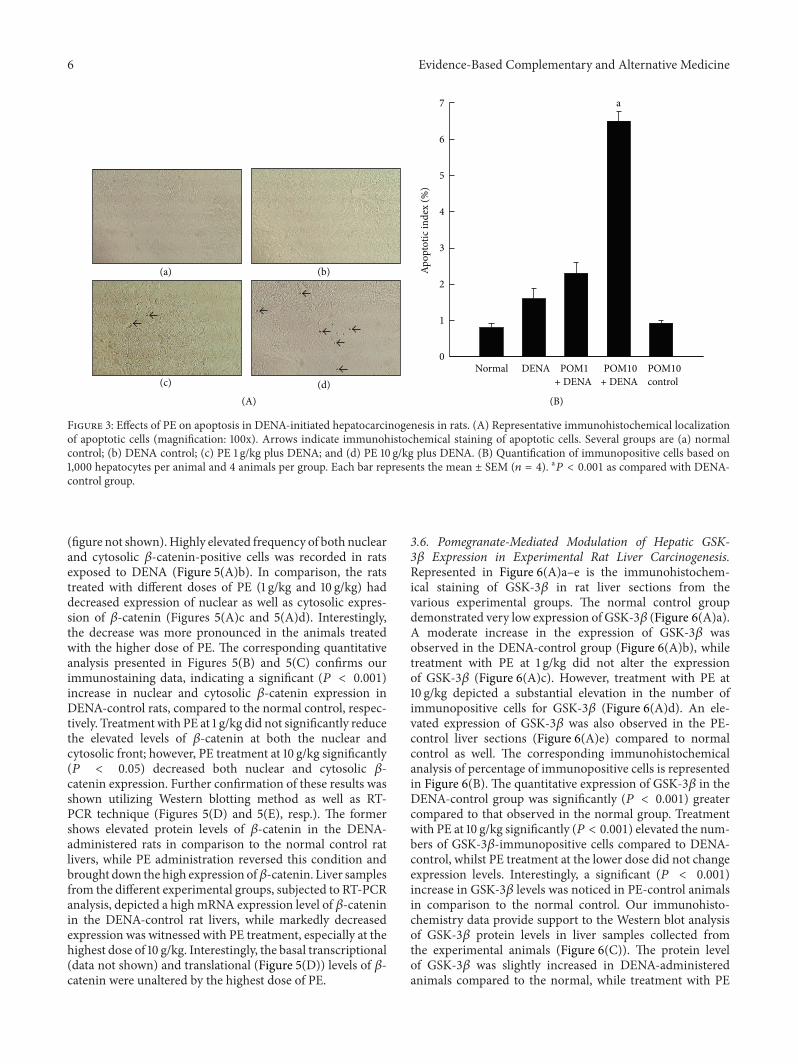
6 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c) (d)
Apop
totic
inde
x (
)
0
1
2
3
4
5
6
7 a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
Figure 3 Effects of PE on apoptosis in DENA-initiated hepatocarcinogenesis in rats (A) Representative immunohistochemical localizationof apoptotic cells (magnification 100x) Arrows indicate immunohistochemical staining of apoptotic cells Several groups are (a) normalcontrol (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of immunopositive cells based on1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA-control group
(figure not shown)Highly elevated frequency of both nuclearand cytosolic 120573-catenin-positive cells was recorded in ratsexposed to DENA (Figure 5(A)b) In comparison the ratstreated with different doses of PE (1 gkg and 10 gkg) haddecreased expression of nuclear as well as cytosolic expres-sion of 120573-catenin (Figures 5(A)c and 5(A)d) Interestinglythe decrease was more pronounced in the animals treatedwith the higher dose of PE The corresponding quantitativeanalysis presented in Figures 5(B) and 5(C) confirms ourimmunostaining data indicating a significant (119875 lt 0001)increase in nuclear and cytosolic 120573-catenin expression inDENA-control rats compared to the normal control respec-tively Treatment with PE at 1 gkg did not significantly reducethe elevated levels of 120573-catenin at both the nuclear andcytosolic front however PE treatment at 10 gkg significantly(119875 lt 005) decreased both nuclear and cytosolic 120573-catenin expression Further confirmation of these results wasshown utilizing Western blotting method as well as RT-PCR technique (Figures 5(D) and 5(E) resp) The formershows elevated protein levels of 120573-catenin in the DENA-administered rats in comparison to the normal control ratlivers while PE administration reversed this condition andbrought down the high expression of120573-catenin Liver samplesfrom the different experimental groups subjected to RT-PCRanalysis depicted a highmRNA expression level of 120573-cateninin the DENA-control rat livers while markedly decreasedexpression was witnessed with PE treatment especially at thehighest dose of 10 gkg Interestingly the basal transcriptional(data not shown) and translational (Figure 5(D)) levels of 120573-catenin were unaltered by the highest dose of PE
36 Pomegranate-Mediated Modulation of Hepatic GSK-3120573 Expression in Experimental Rat Liver CarcinogenesisRepresented in Figure 6(A)andashe is the immunohistochem-ical staining of GSK-3120573 in rat liver sections from thevarious experimental groups The normal control groupdemonstrated very low expression of GSK-3120573 (Figure 6(A)a)A moderate increase in the expression of GSK-3120573 wasobserved in the DENA-control group (Figure 6(A)b) whiletreatment with PE at 1 gkg did not alter the expressionof GSK-3120573 (Figure 6(A)c) However treatment with PE at10 gkg depicted a substantial elevation in the number ofimmunopositive cells for GSK-3120573 (Figure 6(A)d) An ele-vated expression of GSK-3120573 was also observed in the PE-control liver sections (Figure 6(A)e) compared to normalcontrol as well The corresponding immunohistochemicalanalysis of percentage of immunopositive cells is representedin Figure 6(B) The quantitative expression of GSK-3120573 in theDENA-control group was significantly (119875 lt 0001) greatercompared to that observed in the normal group Treatmentwith PE at 10 gkg significantly (119875 lt 0001) elevated the num-bers of GSK-3120573-immunopositive cells compared to DENA-control whilst PE treatment at the lower dose did not changeexpression levels Interestingly a significant (119875 lt 0001)increase in GSK-3120573 levels was noticed in PE-control animalsin comparison to the normal control Our immunohisto-chemistry data provide support to the Western blot analysisof GSK-3120573 protein levels in liver samples collected fromthe experimental animals (Figure 6(C)) The protein levelof GSK-3120573 was slightly increased in DENA-administeredanimals compared to the normal while treatment with PE
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
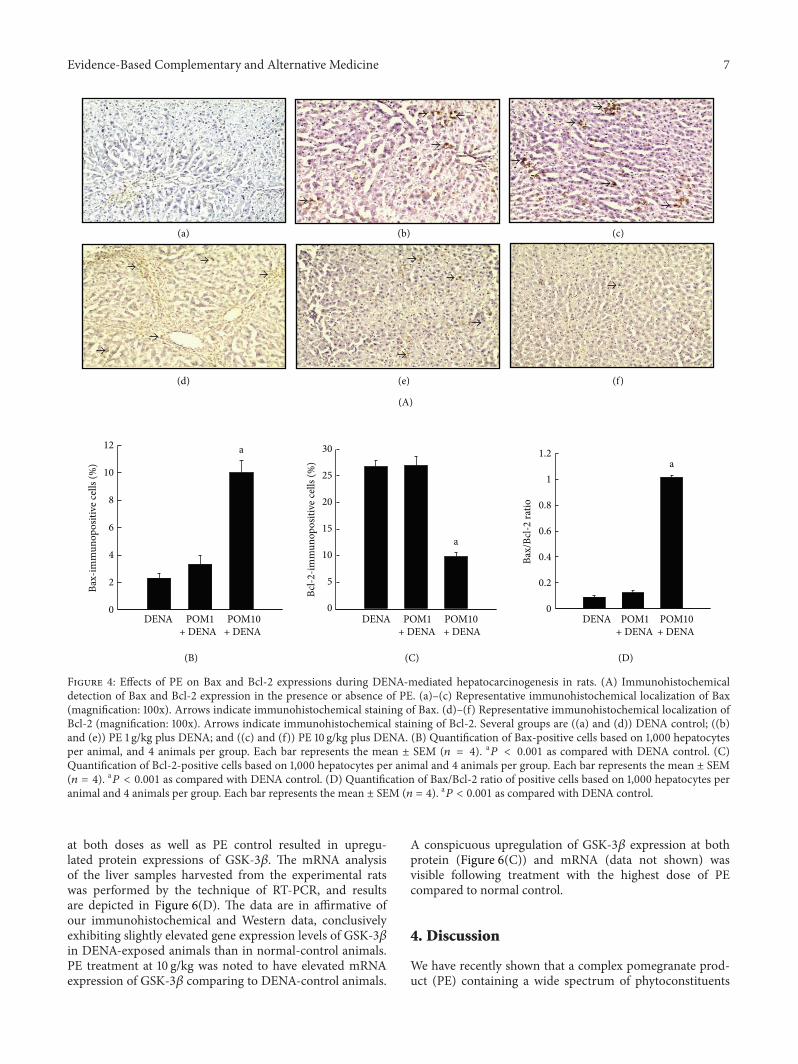
Evidence-Based Complementary and Alternative Medicine 7
(a)
(A)
(b) (c)
(d) (e) (f)
(B)
Bax-
imm
unop
ositi
ve ce
lls (
)
0
2
4
6
8
10
12
DENA
a
POM1 + DENA
POM10 + DENA
(C)
Bcl-2
-imm
unop
ositi
ve ce
lls (
)
0
5
10
15
20
25
30
DENA
a
POM1 + DENA
POM10+ DENA
(D)
Bax
Bcl-2
ratio
0
02
04
06
08
1
12
DENA
a
POM1+ DENA
POM10 + DENA
Figure 4 Effects of PE on Bax and Bcl-2 expressions during DENA-mediated hepatocarcinogenesis in rats (A) Immunohistochemicaldetection of Bax and Bcl-2 expression in the presence or absence of PE (a)ndash(c) Representative immunohistochemical localization of Bax(magnification 100x) Arrows indicate immunohistochemical staining of Bax (d)ndash(f) Representative immunohistochemical localization ofBcl-2 (magnification 100x) Arrows indicate immunohistochemical staining of Bcl-2 Several groups are ((a) and (d)) DENA control ((b)and (e)) PE 1 gkg plus DENA and ((c) and (f)) PE 10 gkg plus DENA (B) Quantification of Bax-positive cells based on 1000 hepatocytesper animal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a
119875 lt 0001 as compared with DENA control (C)Quantification of Bcl-2-positive cells based on 1000 hepatocytes per animal and 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared with DENA control (D) Quantification of BaxBcl-2 ratio of positive cells based on 1000 hepatocytes peranimal and 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared with DENA control
at both doses as well as PE control resulted in upregu-lated protein expressions of GSK-3120573 The mRNA analysisof the liver samples harvested from the experimental ratswas performed by the technique of RT-PCR and resultsare depicted in Figure 6(D) The data are in affirmative ofour immunohistochemical and Western data conclusivelyexhibiting slightly elevated gene expression levels of GSK-3120573in DENA-exposed animals than in normal-control animalsPE treatment at 10 gkg was noted to have elevated mRNAexpression of GSK-3120573 comparing to DENA-control animals
A conspicuous upregulation of GSK-3120573 expression at bothprotein (Figure 6(C)) and mRNA (data not shown) wasvisible following treatment with the highest dose of PEcompared to normal control
4 Discussion
We have recently shown that a complex pomegranate prod-uct (PE) containing a wide spectrum of phytoconstituents
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
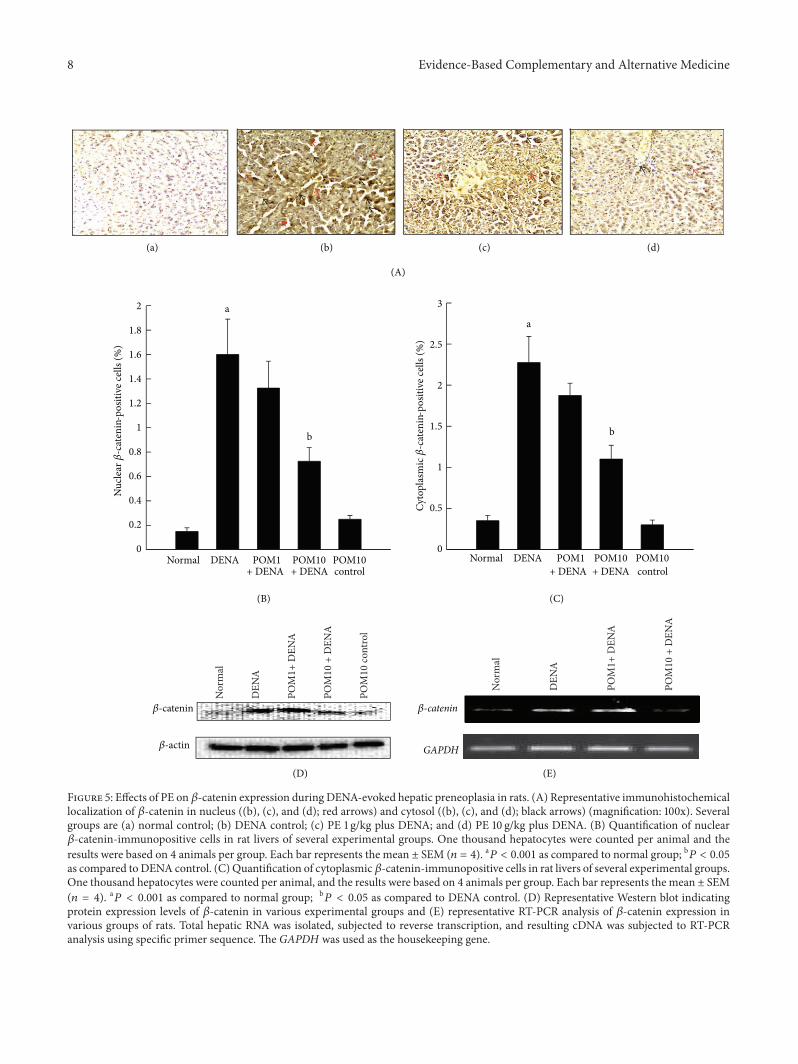
8 Evidence-Based Complementary and Alternative Medicine
(a)
(A)
(b)
(B)
(c)
(C)
(E)
(d)
(D)
0
02
04
06
08
1
12
14
16
18
2
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
0
05
1
15
2
25
3
b
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
120573-actin
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GAPDH
120573-catenin120573-catenin
Cyto
plas
mic120573
-cat
enin
-pos
itive
cells
()
Nuc
lear120573
-cat
enin
-pos
itive
cells
()
Figure 5 Effects of PE on 120573-catenin expression during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization of 120573-catenin in nucleus ((b) (c) and (d) red arrows) and cytosol ((b) (c) and (d) black arrows) (magnification 100x) Severalgroups are (a) normal control (b) DENA control (c) PE 1 gkg plus DENA and (d) PE 10 gkg plus DENA (B) Quantification of nuclear120573-catenin-immunopositive cells in rat livers of several experimental groups One thousand hepatocytes were counted per animal and theresults were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4) a119875 lt 0001 as compared to normal group b119875 lt 005as compared to DENA control (C) Quantification of cytoplasmic 120573-catenin-immunopositive cells in rat livers of several experimental groupsOne thousand hepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM(119899 = 4) a119875 lt 0001 as compared to normal group b
119875 lt 005 as compared to DENA control (D) Representative Western blot indicatingprotein expression levels of 120573-catenin in various experimental groups and (E) representative RT-PCR analysis of 120573-catenin expression invarious groups of rats Total hepatic RNA was isolated subjected to reverse transcription and resulting cDNA was subjected to RT-PCRanalysis using specific primer sequence The GAPDH was used as the housekeeping gene
Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 9
(a)
(A)
(b)
(B)
(e)
(c)
(C)
(d)
(D)
0
1
2
3
4
5
6
7
a
DENA POM1+ DENA
POM10+ DENA
POM10control
Normal
b
a
DEN
A
POM
1+ D
ENA
POM
10 +
DEN
A
POM
10 co
ntro
l
Nor
mal
120573-actinD
ENA
POM
1+ D
ENA
POM
10 +
DEN
A
Nor
mal
GSK-3120573GSK-3120573
GSK
-3120573
-pos
itive
cells
()
GAPDH
Figure 6 Effects of PE on GSK-3120573 expressions during DENA-evoked hepatic preneoplasia in rats (A) Representative immunohistochemicallocalization ofGSK-3120573 (magnification 100x) in different rat groups Several groups are (a) normal control (b)DENAcontrol (c) PE 1 gkg plusDENA (d) PE 10 gkg plus DENA and (e) PE 10 gkg control (B) Quantification of hepatic GSK-3120573-immunopositive cells One thousandhepatocytes were counted per animal and the results were based on 4 animals per group Each bar represents the mean plusmn SEM (119899 = 4)a119875 lt 0001 as compared to normal group b119875 lt 0001 as compared to DENA control (C) Representative Western blot indicating proteinexpression levels of GSK-3120573 in various experimental groups and (D) representative RT-PCR analysis of GSK-3120573 expression in various groupsof rats Total hepatic RNAwas isolated subjected to reverse transcription and resulting cDNAwas subjected to RT-PCR analysis using specificprimer sequence The GAPDH was used as the housekeeping gene
afforded a striking chemoprevention against chemicallyinduced hepatocarcinogenesis in rats [34] It is plausible thatvarious phytochemicals present in PE may target severalinterconnectedmolecular pathways to inhibit liver tumorige-nesis Supporting this possibility we have provided evidencethat PE simultaneously target Nrf2-regulated redox signal-ing and NF-120581B-mediated inflammatory pathway to achievechemoprevention of experimental hepatocarcinogenesis [3435] In the present study we set ourselves at the aim ofinvestigating additional and related mechanisms as regardsto our previous findings to provide a comprehensive under-standing of pleiotropicmechanisms of action of pomegranatephytochemicals in preventing liver cancer
Cell proliferation plays a fundamental role in the multi-step carcinogenesis process including initiation promotionand progression and is considered to be an importantmechanism for the pathogenesis of HCC [43] Hence explo-ration of agents that can affect abnormal proliferation of
hepatocytes may be of immense value in the prevention ofHCC PCNA functions as a cofactor of DNA polymeraseand thus is directly involved in DNA replication The pos-itive expression of PCNA is considered a common indexfor hepatocyte proliferation at late G1- and early S-phasePCNA represents an important cellular marker for assessingthe proliferation during hepatocarcinogenesis [44] In ourstudy the expression of PCNA was examined immuno-histochemically in livers from several animal groups Theelevated expression of PCNA resulting in a drastic increasein PCNA LI in the liver sections of DENA-treated animalsindicates accelerated cell proliferation during an early phaseof rat liver tumorigenesis which supports our previousobservations [41 42] Significant decreases in the numberof PCNA-positive hepatocytes and consequently PCNA LIdue to PE treatment clearly indicate a decrease in hepatocyteproliferation possibly through inhibition of DNA synthesisduring experimental hepatocarcinogenesis It is mostly likely
10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Evidence-Based Complementary and Alternative Medicine
that the antiproliferative mechanisms of pomegranate phy-tochemicals eventually contributed to inhibition of DENA-induced tumorigenesis as we previously observed [34] Ourdata are in agreement of several studies reported from otherlaboratories showing in vitro and in vivo antiproliferativeeffects of various pomegranate phytochemicals in severalcancer models [24 45]
CyclinD1 a cell cycle regulatory protein is responsible forthe transition fromG1- to S-phase [46] As cyclinD1 possessesa critical role for cell cycle progression dysregulated expres-sion of cyclin D1 gene may contribute to cellular genomicinstability and malignancy [47] Elevated expressions ofcyclin D1 gene and protein disrupting G1S regulatory pointof the cell cycle have been found to lead abnormal cell pro-liferation during DENA-induced sequential hepatocarcino-genesis in rats [48ndash50] As increased expression of cyclin D1protein has been found in HCC patients [51] and correlatedwith poor overall survival inHCCpatients [52] it can be usedas a prognostic marker and therapeutic target The results ofour immunohistochemical as well as Western blot analysisclearly demonstrate an upregulation of cyclin D1 proteinexpression in the livers isolated from DENA control animalsThese findings are supported by the increased expression ofcyclin D1 gene following DENA treatment Collectively ourdata underscore the critical role of cell cycle regulatory pro-tein in DENA hepatocellular carcinogenesis in rats under ourexperimental conditions Interestingly a similar transcrip-tional or translational upregulation of cyclin D1 in responseto DENA exposure in rodents has been reported by variousother laboratories [53ndash55] A near normal level of cyclin D1expression was observed due to PE treatment Reversal ofdysregulation of a critical checkpoint of the cell cycle couldbe one of the possible mechanisms of pomegranate-mediatedinhibition of heptotumorigenesis and it underscores thevalue of targeting a cell cycle progression protein to achievechemoprevention of hepatocellular cancer
Apoptosis or programmed cell death a highly pre-served cellular mechanism is involved in tissue homeostasisthrough targeted elimination of singe cells without disruptingphysiological function of a tissue Apoptosis is marked bydistinct morphological alterations characterized by cytoplas-mic as well as nuclear condensation DNA fragmentationphosphatidylserine externalization and plasma membraneblebbing [56] Dysregulation of apoptosis disturbing thebalance between cell proliferation and cell death contributesto development and progression of hepatic malignancy [5758] Apoptosis detection in tumor mass has emerged as animportant diagnostic tool and induction of tumor cell deathby apoptosis has been accepted as one of the fundamentalobjectives of liver cancer therapy [59] Promotion of apop-tosis in tumor-target tissues has been identified as a novelmechanismof potential chemopreventive drug In the presentstudy DNA fragmentation was detected to identify cellsundergoing apoptosis Our results show an extremely loworder of DNA fragmentation in DENA-exposed rat liversindicating apoptosis evasion PE treatment showed a drasticincrease in the formation of DNA fragments which providessubstantial evidence of induction of cell death by apoptosisand consequently reduction of proportion of transformed
hepatocytes in DENA-treated animals Our data showingapoptosis induction during experimental hepatocarcinogen-esis in rats by pomegranate phytoconstituents are concordantwith a large number of studies showing proapoptotic effectsof pomegranate-derived agents in various in vitro and in vivopreclinical cancer models [reviewed in ref [31]]
The apoptotic signal in hepatocytes is believed to betransmitted through the complex interaction of intercellularproteinsThe transcription factor NF-120581B andmembers of theBcl-2 family consisting of both proapoptotic and antiapop-totic proteins contribute to the regulation of apoptosis in hep-atocytes [58] Many of the genetic alterations of HCC lead toan imbalance in the proapoptotic and antiapoptotic proteinsof the Bcl-2 family [60]The antiapoptotic proteins includingBcl-2 and Bcl-xL are known to inhibit mitochondrial apop-totic pathway by blocking the release and oligomerization ofproapoptotic proteins and are overexpressed inHCC [61] Onthe other hand proapoptotic members of the Bcl-2 familysuch as Bax Bad and Bid initiate mitochondrial apoptosisby facilitating pore formation and cytochrome 119888 release fromthe inner mitochondrial membrane with subsequent activa-tion of caspases resulting in cell death These proapoptoticmembers are downregulated in HCC [62] An alteration inBaxBcl-2 ratio plays a crucial role in deciding whether a cellshould switch towards proliferation or apoptosis [63] Ourpresent study revealed that PE registered an increase in theexpression of proapoptotic gene Bax and decrease in the levelof antiapoptotic gene Bcl-2 resulting in an extraordinarysurge in BaxBcl-2 ratio Based upon these results it is clearthat pomegranate bioactive phytochemicals induce intrinsicapoptosis and facilitate elimination of transformed cellsduring hepatocellular carcinogenesis by targeting the Bcl-2family members
The Wnt120573-catenin signaling pathway is implicated inliver physiology and pathology through regulation of variousfundamental cellular events including proliferation differ-entiation survival inflammation oxidative stress morpho-genesis and regeneration [15 64 65] In canonical Wntpathway themultifunctional protein120573-catenin represents thekey signaling intermediate Under normal physiological con-ditions free cytoplasmic 120573-catenin undergoes degradationthrough a coordinated action of a multiprotein destructioncomplex consists of GSK-3120573 adenomatous polyposis colicasein kinase 1120572 (CK1120572) and Axin [66] It is known that 120573-catenin is first phosphorylated at serine-45 by CK1120572 facil-itating its subsequent phosphorylation at serine-33 serine-37 and threonine-41 by GSK-3120573 [67 68] The phosphory-lated 120573-catenin first undergoes ubiquitination by the cellularE3 ubiquitin ligase complex and then degradation by 26Sproteasome [69] In the event of Wnt ligands binding withthe transmembrane Frizzled (FZD) receptor and coreceptorlow-density lipoprotein receptor-related protein 5 or 6 (LRP56) activation of the canonical Wnt pathway is initiatedIn this process FZD recruits cytoplasmic scaffolding proteinDishevelled (Dvl) which subsequently recruits Axin andGSK-3120573 to LRP 56 [70] Consequently LRP 56 is phospho-rylated by CK1120572 and GSK-3120573 resulting in disruption of theformation of the destruction complex and inhibition of GSK-3120573-mediated phosphorylation of 120573-catenin The net result is
Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 11
the stabilization and accumulation of 120573-catenin in the cytosolfollowed by its translocation to the nucleus Once insidethe nucleus 120573-catenin interacts with transcription factor T-cell factorlymphoid enhancer factor and eventually activatesthe transcription of various Wnt target genes implicatedin a number of important biological functions includingproliferation (eg c-myc) cell cycle control (eg cyclinD1) apoptosis (eg survivin) and inflammation (eg COX-2) [71ndash74] Aberrant constitutive activation of the Wnt120573-catenin pathway leads to uncontrolled cell proliferationgrowth and survival promoting various human malignan-cies including HCC [13 16] Emerging evidence based onexperimental as well as clinical observations indicate theinvolvement of activated Wnt120573-catenin signaling at var-ious stages of hepatic neoplasia making it an attractiveand viable target for prevention and treatment of HCC[reviewed in refs [17ndash19]] Accumulation of 120573-catenin inthe cytoplasm and nucleus has previously been reportedduring stepwise progression of DENA-initiated hepatocar-cinogenesis in rats [75] and DENAPB-mediated HCC inmice [76] Our immunohistochemical study also reveals thathepatocytes from DENA-control animals exhibit a markedoverexpression of 120573-catenin in the cytoplasm andor nucleusconfirming activation of Wnt120573-catenin signaling pathwayat an early stage of DENA hepatocarcinogenesis in ratsThe accompanying Western blot and RT-PCR data providefurther support to our observation regarding induction of thecanonical Wnt120573-catenin pathway A dose-dependent inhibi-tion of 120573-catenin expression in transcriptional and transla-tional levels by pomegranate constituents clearly underscoresthe impairment of the canonical Wnt120573-catenin oncogenicsignaling in rat liver carcinogenesis
Reduced expression ofGSK-3120573mRNAhas been observedin human hepatic cancer linked to HCV infection [77]HBV-X protein upregulated 120573-catenin with an inactivationof GSK-3120573 in liver cancer [78] Targeted downregulation of120573-catenin protein has been associated with increased GSK-3120573 protein expression in HCC [79] A parallel induction ofhepatic GSK-3120573 protein following pomegranate treatmenthas been an interesting finding of our study which indicatespossible degradation of 120573-catenin protein via upregulationof GSK-3120573 The activation of GSK-3120573 may be responsibleat least in part for the attenuation of oncogenic Wnt120573-catenin signaling and its effect on downstream gene expres-sion While the involvement of GSK-3120573 in downregulationof 120573-catenin protein following pomegranate treatment isapparent from this study it is not clear how pomegranatephytochemicals affect the expression of 120573-catenin at thetranscriptional level and may suggest other mechanismsPomegranate bioactive compounds may have direct effectson transcriptional regulation of120573-catenin andGSK-3120573 genesThis is in line with a recent study that shows reductionin mRNA expressions of 120573-catenin and GSK-3120573 in humanhepatocarcinoma cells following treatment with nimbolide anatural compoundderived fromneem tree [80] Additionallymethylated chrysin a constituent of pepper tree leaves hasbeen found to abrogate the elevated expression of 120573-cateninmRNA in preneoplastic lesions induced byDENA in rats [81]Although emerging studies have provided credible evidence
on the potential of a gamut of bioactive food components inthe prevention and treatment of various neoplastic diseasesby targeting the wingless signaling [reviewed in [82ndash84]]very limited information is available on chemoprevention ofHCC by inhibiting activated Wnt120573-catenin signaling path-way using nutraceuticals and dietary components [reviewedin [19]]
5 Conclusion
The results of our present study undoubtedly establish thatpomegranate bioactive constituents suppress cell prolifera-tion (PCNA) regulate cell cycle progression (cyclin D1) andinduce programmed cell death (apoptosis) during DENA-initiated andPB-promoted hepatic tumorigenesis in Sprague-Dawley rats Pomegranate-mediated apoptogenic signal dur-ing experimental hepatocarcinogenesis may be transmit-ted through up-relegation of proapoptotic protein Bax anddownmodulation of antiapoptotic protein Bcl-2 We alsopresent data that show for the first time that pomegranatephytochemicals diminish cell proliferation and survival pos-sibly through blockade of activated canonical Wnt120573-cateninsignaling in hepatocarcinogen-exposed animalsThe interfer-ence of Wnt120573-catenin signaling may be achieved at least inpart through degradation of 120573-catenin protein the cardinaleffector of Wnt pathway by up-regulation of GSK-3120573 Thisis further supported by the diminished hepatic expressionof cyclin D1 a 120573-catenin-dependent target gene and anindicator of activation of Wnt120573-catenin pathway Thus ourstudy provides encouraging preclinical evidence and novelmechanism of targeting activated Wnt120573-catenin signalingby pomegranate products to achieve chemoprevention ofhepatocellular cancer Previously we have shown modula-tion of the NF-120581B pathway by pomegranate constituents inDENA hepatocarcinogenesis in rats [35] As functional crossregulation between the NF-120581B and Wnt120573-catenin signalingpathways has emerged as a pivotal mechanism for the regu-lation of a diverse array of genes involved in tumorigenesisincluding HCC [37ndash40] results reported here in conjunctionwith our earlier communication [35] indisputably estab-lish the ldquoproof-of-principlerdquo of simultaneously targeting twointerconnected molecular circuits namely canonical NF-120581B and Wnt120573-catenin pathways to achieve prevention ofhepatocellular cancer (summarized in Figure 7) Obviouslythese do not exclude the possibility of other mechanismsOverall our present results based on a clinically relevantmodel of liver cancer coupled with an excellent safetyprofile of pomegranate bioactive substances may facilitatethe development of pomegranate-derived agents as complexsynergistic drug for the prevention and therapy of livercancer which represents a complex disease
Conflict of Interests
The authors declare that no conflict of interests exists
Acknowledgments
A part of this work was carried out at the Northeast OhioMedical University (Rootstown OH) supported by a new
12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Evidence-Based Complementary and Alternative Medicine
Preneoplastic lesions
HCC
1 2
Bax
COX-2
DENA
Wnt
Pomegranate Pomegranate
Cyclin D1 Bcl-2
Cell proliferation
Apoptosisevasion
120573-cateninNF-120581B
Figure 7 Schematic representation of the possible molecular mechanisms of pomegranate-mediated chemoprevention of experimentalhepatocarcinogenesis in rats COX-2 cyclooxygenase-2 DENA diethylnitrosamine HCC hepatocellular carcinoma NF-120581B nuclear factor-120581B DENA simultaneously activates Wnt120573-catenin and NF-120581B signaling pathways which in turn result in an elevated expression of targetgenes including cyclin D1 COX-2 and Bcl-2 and reduced expression of Bax All these features contribute to accelerated hepatocyteproliferation and evasion of apoptosis which results in development of preneoplastic lesions and subsequently HCC Pomegranatephytochemicals block interconnected Wnt120573-catenin and NF-120581B signaling pathways which lead to antiproliferative and proapoptotic effectscontributing to chemoprevention of HCC darr Activation and perp downregulation 1 and 2 indicate results of this study and that from [35]respectively
faculty start-up research Grant to A Bishayee The authorssincerely thank Danielle M Petit and Thomas Mbimba forassistance with the immunohistochemical studies
References
[1] M M Center and A Jemal ldquoInternational trends in livercancer incidence ratesrdquo Cancer Epidemiology Biomarkers andPrevention vol 20 pp 2362ndash2368 2011
[2] R Siegel D Naishadham and A Jemal ldquoCancer statisticsrdquo CACancer Journal For Clinicians vol 62 pp 10ndash29 2012
[3] M B Thomas D Jaffe M M Choti et al ldquoHepatocellular car-cinoma consensus recommendations of the National CancerInstitute Clinical Trials Planning Meetingrdquo Journal of ClinicalOncology vol 28 pp 3994ndash4005 2010
[4] T R Morgan ldquoChemoprevention of hepatocellular carcinomain chronic hepatitis Crdquo Recent Results in Cancer Research vol188 pp 85ndash99 2011
[5] A Bishayee T Politis and A S Darvesh ldquoResveratrol in thechemoprevention and treatment of hepatocellular carcinomardquoCancer Treatment Reviews vol 36 no 1 pp 43ndash53 2010
[6] A Bishayee ldquoEditorial recent advances in the prevention andtherapy of hepatocellular carcinomardquo Current Cancer DrugTargets vol 12 no 9 pp 1043ndash1044 2012
[7] S Kawanishi Y Hiraku S Pinlaor and N Ma ldquoOxidativeand nitrative DNA damage in animals and patients withinflammatory diseases in relation to inflammation-related car-cinogenesisrdquo Biological Chemistry vol 387 no 4 pp 365ndash3722006
[8] H L Ha H J Shin M A Feitelson and D Y Yu ldquoOxidativestress and antioxidants in hepatic pathogenesisrdquoWorld Journalof Gastroenterology vol 16 pp 6035ndash6043 2010
[9] M Marra I M Sordelli A Lombardi et al ldquoMolecular targetsand oxidative stress biomarkers in hepatocellular carcinoma anoverviewrdquo Journal of Translational Medicine vol 9 p 171 2011
[10] J Gao L Xie W S Yang et al ldquoRisk factors of hepatocellularcarcinomamdashcurrent status and perspectivesrdquoAsian Pacific Jour-nal of Cancer Prevention vol 13 pp 743ndash752 2012
[11] P A Farazi and R A DePinho ldquoHepatocellular carcinomapathogenesis from genes to environmentrdquo Nature ReviewsCancer vol 6 no 9 pp 674ndash687 2006
[12] C M Wong and O L Ng ldquoMolecular pathogenesis of hepa-tocellular carcinomardquo Liver International vol 28 pp 160ndash1742008
Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 13
[13] K N Nejak-Bowen and S P Monga ldquoBeta-catenin signalingliver regeneration and hepatocellular cancer sorting the goodfrom the badrdquo Seminars in Cancer Biology vol 21 pp 44ndash582011
[14] H C Lee M Kim and J R Wands ldquoWntFrizzled signalingin hepatocellular carcinomardquo Frontiers in Bioscience vol 11 pp1901ndash1915 2006
[15] Y Takigawa andAMC Brown ldquoWnt signaling in liver cancerrdquoCurrent Drug Targets vol 9 pp 1013ndash1024 2008
[16] S Fatima N P Lee and J M Luk ldquoDickkopfs and Wnt120573-catenin signaling in liver cancerrdquo World Journal of ClinicalOncology vol 2 pp 311ndash325 2011
[17] R Dahmani P A Just and C Perret ldquoTheWnt120573-catenin path-way as a therapeutic target inhuman hepatocellular carcinomardquoClinics and Research on Hepatology and Gastroenterology vol35 pp 709ndash713 2011
[18] S B Nambotin J RWands andM Kim ldquoPoints of therapeuticintervention along the wnt signaling pathway in hepatocellularcarcinomardquo Anti-Cancer Agents in Medicinal Chemistry vol 11no 6 pp 549ndash559 2011
[19] A Bishayee ldquo120573-Catenin a novel biomarker and therapeutictarget in liver cancerrdquo in Cancer Biomarkers A GeorgakilasEd CRC Press Boca Raton Fla USA 2012
[20] V V Sivarajan and I Balachandran Ayurvedic Drugs and TheirPlant Sources IBH Publishing Oxford UK 1994
[21] A Saxena and N K Vikram ldquoRole of selected Indian plants inmanagement of type 2 diabetes a reviewrdquo Journal of Alternativeand Complementary Medicine vol 10 no 2 pp 369ndash378 2004
[22] M I Gil F A Tomas-Barberan B Hess-Pierce D M Holcroftand A A Kader ldquoAntioxidant activity of pomegranate juiceand its relationship with phenolic composition and processingrdquoJournal of Agricultural and Food Chemistry vol 48 no 10 pp4581ndash4589 2000
[23] A Faria R Monteiro N Mateus I Azevedo and C CalhauldquoEffect of pomegranate (Punica granatum) juice intake onhepatic oxidative stressrdquo European Journal of Nutrition vol 46no 5 pp 271ndash278 2007
[24] E P Lansky and R A Newman ldquoPunica granatum(pomegranate) and its potential for prevention and treatmentof inflammation and cancerrdquo Journal of Ethnopharmacologyvol 109 no 2 pp 177ndash206 2007
[25] A Faria and C Conceicao ldquoThe bioactivity of pomegranateimpact on health and diseaserdquo Critical Reviews in Food Scienceand Nutrition vol 51 no 7 pp 626ndash634 2011
[26] D Heber ldquoChapter 10 pomegranate ellagitanninsrdquo in HerbalMedicine Biomolecular and Clinical Aspects I F F Benzie andS Wachtel-Galor Eds CRC Press Boca Raton Fla USA 2ndedition 2011
[27] J Jurenka ldquoTherapeutic applications of pomegranate (Punicagranatum L) a reviewrdquoAlternativeMedicine Review vol 13 no2 pp 128ndash144 2008
[28] A Basu and K Penugonda ldquoPomegranate juice a heart healthyfruit juicerdquo Nutrition Reviews vol 67 pp 49ndash56 2009
[29] N Khan F Afaq and H Mukhtar ldquoCancer chemopreventionthrough dietary antioxidants progress and promiserdquo Antioxi-dants and Redox Signaling vol 10 no 3 pp 475ndash510 2008
[30] V M Adhami N Khan and H Mukhtar ldquoCancer chemo-prevention by pomegranate laboratory and clinical evidencerdquoNutrition and Cancer vol 61 no 6 pp 811ndash815 2009
[31] A Bishayee and A S Darvesh ldquoPomegranate-derived con-stituents as inducers of cell death implications in cancer
prevention and therapyrdquo in Natural Compounds As Inducers ofCell Death M Diederich Ed Springer New York NY USA2012
[32] A J Pantuck J T Leppert N Zomorodian et al ldquoPhase IIstudy of pomegranate juice formenwith rising prostate-specificantigen following surgery or radiation for prostate cancerrdquoClinical Cancer Research vol 12 no 13 pp 4018ndash4026 2006
[33] C J Paller X Ye P J Wozniak et al ldquoA randomized phase IIstudy of pomegranate extract formenwith rising PSA followinginitial therapy for localized prostate cancerrdquoProstate Cancer andProstatic Diseases vol 16 no 1 pp 50ndash55 2013
[34] A Bishayee D Bhatia R J Thoppil A S Darvesh E Nevoand E P Lansky ldquoPomegranate-mediated chemopreventionof experimental hepatocarcinogenesis involves Nrf2-regulatedantioxidantmechanismsrdquoCarcinogenesis vol 32 no 6 pp 888ndash896 2011
[35] A Bishayee R J Thoppil A S Darvesh V Ohanyan JG Meszaros and D Bhatia ldquoPomegranate phytoconstituentsblunt the inflammatory cascade in a chemically-induced rodentmodel of hepatocellular carcinogenesisrdquo Journal of NutritionalBiochemistry vol 24 no 1 pp 178ndash187 2013
[36] B B Aggarwal and B Sung ldquoNF-120581B in cancer a matter of lifeand deathrdquo Cancer Discovery vol 1 pp 469ndash471 2011
[37] QDu andDAGeller ldquoCross-regulation betweenWnt andNF-120581B signaling pathwaysrdquo Forum on Immunopathological Diseasesand Therapeutics vol 1 pp 155ndash181 2010
[38] M Moreau S Mourah and C Dosquet ldquo120573-Catenin and NF-120581B cooperate to regulate the uPAuPAR system in cancer cellsrdquoInternational Journal of Cancer vol 128 no 6 pp 1280ndash12922011
[39] M M Chaturvedi B Sung V R Yadav R Kannappan and BB Aggarwal ldquoNF-120581B addiction and its role in cancer one sizedoes not fit allrdquo Oncogene vol 30 no 14 pp 1615ndash1630 2011
[40] T H Lee D I Tai C J Cheng et al ldquoEnhanced nuclearfactor-kappa B-associated Wnt-1 expression in hepatitis B- andC-related hepatocarcinogenesis identification by functionalproteomicsrdquo Journal of Biomedical Science vol 13 no 1 pp 27ndash39 2006
[41] A Bishayee and N Dhir ldquoResveratrol-mediated chemopre-vention of diethylnitrosamine-initiated hepatocarcinogenesisinhibition of cell proliferation and induction of apoptosisrdquoChemico-Biological Interactions vol 179 no 2-3 pp 131ndash1442009
[42] A Bishayee T Mbimba R J Thoppil et al ldquoAnthocyanin-rich black currant (Ribes nigrum L) extract affords chemo-prevention against diethylnitrosamine-induced hepatocellularcarcinogenesis in ratsrdquo Journal of Nutritional Biochemistry vol22 pp 1035ndash1046 2011
[43] N C Teoh ldquoProliferative drive and liver carcinogenesis toomuch of a good thingrdquo Journal of Gastroenterology and Hep-atology vol 24 no 12 pp 1817ndash1825 2009
[44] L X Qin and Z Y Tang ldquoThe prognostic molecular markersin hepatocellular carcinomardquoWorld Journal of Gastroenterologyvol 8 pp 385ndash392 2002
[45] D Heber ldquoMultitargeted therapy of cancer by ellagitanninsrdquoCancer Letters vol 269 pp 262ndash268 2008
[46] R Donnellan and R Chetty ldquoCyclin D1 and human neoplasiardquoMolecular Pathology vol 51 pp 1ndash7 1998
[47] K Sugimachi S I Aishima K I Taguchi et al ldquoThe roleof overexpression and gene amplification of cyclin D1 inintrahepatic cholangiocarcinomardquo Journal of Hepatology vol35 no 1 pp 74ndash79 2001
14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

14 Evidence-Based Complementary and Alternative Medicine
[48] M Sundarrajan S Gupta and K V K Rao ldquoOverexpressionof cyclin D1 is associated with the decondensation of chro-matin during den-induced sequential hepatocarcinogenesisrdquoCell Biology International vol 26 no 8 pp 699ndash706 2002
[49] P Parekh and K V K Rao ldquoOverexpression of cyclin D1 isassociated with elevated levels of MAP kinases Akt and Pak1during diethylnitrosamine-induced progressive liver carcino-genesisrdquoCell Biology International vol 31 no 1 pp 35ndash43 2007
[50] D H Park J W Shin S K Park et al ldquoDiethylni-trosamine (DEN) induces irreversible hepatocellular carcino-genesis through overexpression of G1S-phase regulatory pro-teins in ratrdquo Toxicology Letters vol 191 no 2-3 pp 321ndash3262009
[51] A A Bahnassy A R Zekri S A Loutfy et al ldquoThe roleof cyclins and cyclin dependent kinases in development andprogression of hepatitis C virus-genotype 4-associated hepatitisand hepatocellular carcinomardquo Experimental and MolecularPathology vol 91 pp 643ndash652 2011
[52] Y Che F Ye R Xu et al ldquoCo-expression of XIAP and cyclinD1 complex correlates with a poor prognosis in patients withhepatocellular carcinomardquo American Journal of Pathology vol180 pp 1798ndash1807 2012
[53] Y Qian and C Q Lin ldquoPreventive effect of Ganfujian granuleon experimental hepatocarcinoma in ratsrdquo World Journal ofGastroenterology vol 10 no 5 pp 755ndash757 2004
[54] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[55] B Cui Y Chen S Liu et al ldquoAntitumour activity of Lyciumchinensis polysaccharides in liver cancer ratsrdquo InternationalJournal of Biology and Macromolecules vol 5 pp 314ndash318 2012
[56] C Burz I Berindan-Neagoe O Balacescu and A IrimieldquoApoptosis in cancer key molecular signaling pathways andtherapy targetsrdquo Acta Oncologica vol 48 no 6 pp 811ndash8212009
[57] I Fabregat ldquoDysregulation of apoptosis in hepatocellular carci-noma cellsrdquoWorld Journal of Gastroenterology vol 15 pp 513ndash520 2009
[58] J M SchattenbergM Schuchmann and P R Galle ldquoCell deathand hepatocarcinogenesis dysregulation of apoptosis signalingpathwaysrdquo Journal of Gastroenterology and Hepatology vol 26no 1 pp 213ndash219 2011
[59] E Karamitopoulou L Cioccari S Jakob et al ldquoActive caspase 3and DNA fragmentation as markers for apoptotic cell death inprimary and metastatic liver tumoursrdquo Pathology vol 39 no 6pp 558ndash564 2007
[60] J L Mott and G J Gores ldquoPiercing the armor of hepatobiliarycancer Bcl-2 homology domain 3 (BH3) mimetics and celldeathrdquo Hepatology vol 46 no 3 pp 906ndash911 2007
[61] T Takehara X Liu J Fujimoto S L Friedman and H Taka-hashi ldquoExpression and role of Bcl-xL in human hepatocellularcarcinomasrdquo Hepatology vol 34 no 1 pp 55ndash61 2001
[62] W Beerheide Y J Tan E Teng A E Ting A Jedpiyawongseand P Srivatanakul ldquoDownregulation of proapoptotic proteinsBax and Bcl-X(s) in p53 overexpressing hepatocellular carci-nomasrdquo Biochemical and Biophysical Research Communicationsvol 273 no 1 pp 54ndash61 2000
[63] M DrsquoAmelio E Tino and F Cecconi ldquoThe apoptosomeemerging insights and new potential targets for drug designrdquoPharmaceutical Research vol 25 no 4 pp 740ndash751 2008
[64] M D Thompson and S P Monga ldquoWNTbeta-catenin signal-ing in liver health and diseaserdquo Hepatology vol 45 pp 1298ndash1305 2007
[65] AG Lade and S P SMonga ldquoBeta-catenin signaling in hepaticdevelopment and progenitors whichway does theWNTblowrdquoDevelopmental Dynamics vol 240 no 3 pp 486ndash500 2011
[66] C Sakanaka ldquoPhosphorylation and regulation of beta-cateninby casein kinase I epsilonrdquo Journal of Biochemistry vol 132 pp697ndash703 2002
[67] S Amit A Hatzubai Y Birman et al ldquoAxin-mediated CKIphosphorylation of 120573-catenin at Ser 45 a molecular switch forthe Wnt pathwayrdquo Genes and Development vol 16 no 9 pp1066ndash1076 2002
[68] C Liu Y Li M Semenov et al ldquoControl of beta-cateninphosphorylationdegradation by a dual-kinase mechanismrdquoCell vol 108 pp 837ndash847 2002
[69] H Aberle A Bauer J Stappert A Kispert and R Kemler ldquo120573-catenin is a target for the ubiquitin-proteasome pathwayrdquo TheEMBO Journal vol 16 no 13 pp 3797ndash3804 1997
[70] X Zeng H Huang K Tamai et al ldquoInitiation ofWnt signalingcontrol of Wnt coreceptor Lrp6 phosphorylationactivation viafrizzled dishevelled and axin functionsrdquo Development vol 135no 2 pp 367ndash375 2008
[71] T C He A B Sparks C Rago et al ldquoIdentification of c-MYCas a target of the APC pathwayrdquo Science vol 28 pp 1509ndash15121998
[72] M Shtutman J Zhurinsky I Simcha et al ldquoThe cyclin D1 geneis a target of the 120573-cateninLEF-1 pathwayrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 96 no 10 pp 5522ndash5527 1999
[73] T Zhang T Otevrel Z Gao et al ldquoEvidence that APC regulatessurvivin expression a possible mechanism contributing to thestem cell origin of colon cancerrdquo Cancer Research vol 61 no24 pp 8664ndash8667 2001
[74] Y Araki S Okamura S P Hussain et al ldquoRegulation ofcyclooxygenase-2 expression by the Wnt and ras pathwaysrdquoCancer Research vol 63 pp 728ndash734 2003
[75] Q M Wang K M Yang H Y Zhou Z H Yu X Li andH J Yang ldquoRole of 120573-catenin in hepatocarcinogenesis of ratsrdquoHepatobiliary and Pancreatic Diseases International vol 5 no1 pp 85ndash89 2006
[76] D F Calvisi S Ladu V M Factor and S S ThorgeirssonldquoActivation of 120573-catenin provides proliferative and invasiveadvantages in c-mycTGF-120572 hepatocarcinogenesis promoted byphenobarbitalrdquoCarcinogenesis vol 25 no 6 pp 901ndash908 2004
[77] A R N Zekri A A Bahnassy S A Abdel-Wahab et alldquoExpression of pro- and anti-inflammatory cytokines in relationto apoptotic genes in Egyptian liver disease patients associatedwith HCV-genotype-4rdquo Journal of Gastroenterology and Hepa-tology vol 24 no 3 pp 416ndash428 2009
[78] Q Ding W Xia J C Liu et al ldquoErk associates with and primesGSK-3b for its inactivation resulting in upregulation of beta-cateninrdquoMolecular Cell vol 19 pp 159ndash170 2005
[79] X H Wang X W Meng H Xing et al ldquoSTAT3 and beta-catenin signaling pathway may affect GSK-3beta expression inhepatocellular carcinomardquo Hepatogastroenterology vol 58 pp487ndash491 2011
[80] K Kavitha R Vidya Priyadarsini P Anitha et al ldquoNimbolidea neem limonoid abrogates canonical NF-120581B andWnt signalingto induce caspase-dependent apoptosis in human hepatocarci-noma (HepG2) cellsrdquo European Journal of Pharmacology vol681 pp 6ndash14 2012
Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 15
[81] M S Khan D Halagowder and S N Devaraj ldquoMethylatedchrysin induces co-ordinated attenuation of the canonicalWnt and NF-kB signaling pathway and upregulates apoptoticgene expression in the early hepatocarcinogenesis rat modelrdquoChemico-Biological Interactions vol 193 no 1 pp 12ndash21 2011
[82] F H Sarkar Y Li Z Wang and D Kong ldquoThe role ofnutraceuticals in the regulation ofWnt andHedgehog signalingin cancerrdquoCancer andMetastasis Reviews vol 29 no 3 pp 383ndash394 2010
[83] M H Teiten F Gaascht M Dicato and M Diederich ldquoTar-geting the wingless signaling pathway with natural compoundsas chemopreventive or chemotherapeutic agentsrdquoCurrent Phar-maceutical Biotechnology vol 13 pp 245ndash254 2012
[84] R S Tarapore I A Siddiqui and H Mukhtar ldquoModulationof Wnt120573-catenin signaling pathway by bioactive food compo-nentsrdquo Carcinogenesis vol 33 pp 483ndash491 2012
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Copyright © 2022 FDOKUMEN




















