
Petiole-Lamina Transition Zone: A Functionally Crucial ... - MDPI

Plastid x3-fatty acid desaturase-dependent accumulation of asystemic acquired resistance inducing activity in petioleexudates of Arabidopsis thaliana is independent ofjasmonic acid
Ratnesh Chaturvedi1,2,‡, Kartikeya Krothapalli2,‡, Ragiba Makandar1,2, Ashis Nandi2,†, Alexis A. Sparks2,
Mary R. Roth2, Ruth Welti2 and Jyoti Shah1,2,*
1Department of Biological Sciences, University of North Texas, Denton, TX 76203, USA, and2Division of Biology and the Molecular Cellular and Developmental Biology Program, Kansas State University,
Manhattan, KS 66506, USA
Received 22 October 2007; accepted 6 December 2007.*For correspondence (fax +1 940 565 4136; e-mail [email protected]).†Present address: School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India.‡These authors contributed equally to this work.
Summary
Systemic acquired resistance (SAR) is an inducible defense mechanism that is activated throughout the plant,
subsequent to localized inoculation with a pathogen. The establishment of SAR requires translocation of an
unknown signal from the pathogen-inoculated leaf to the distal organs, where salicylic acid-dependent
defenses are activated. We demonstrate here that petiole exudates (PeXs) collected from Arabidopsis leaves
inoculated with an avirulent (Avr) Pseudomonas syringae strain promote resistance when applied to
Arabidopsis, tomato (Lycopersicum esculentum) and wheat (Triticum aestivum). Arabidopsis FATTY ACID
DESATURASE7 (FAD7), SUPPRESSOR OF FATTY ACID DESATURASE DEFICIENCY1 (SFD1) and SFD2 genes are
required for accumulation of the SAR-inducing activity. In contrast to Avr PeX from wild-type plants, Avr PeXs
from fad7, sfd1 and sfd2 mutants were unable to activate SAR when applied to wild-type plants. However, the
SAR-inducing activity was reconstituted by mixing Avr PeXs collected from fad7 and sfd1 with Avr PeX from
the SAR-deficient dir1 mutant. Since FAD7, SFD1 and SFD2 are involved in plastid glycerolipid biosynthesis and
SAR is also compromised in the Arabidopsis monogalactosyldiacylglycerol synthase1 mutant we suggest that
a plastid glycerolipid-dependent factor is required in Avr PeX along with the DIR1-encoded lipid transfer
protein for long-distance signaling in SAR. FAD7-synthesized lipids provide fatty acids for synthesis of
jasmonic acid (JA). However, co-infiltration of JA and methylJA with Avr PeX from fad7 and sfd1 did not
reconstitute the SAR-inducing activity. In addition, JA did not co-purify with the SAR-inducing activity
confirming that JA is not the mobile signal in SAR.
Keywords: inducible defense, long-distance signaling, salicylic acid, systemic acquired resistance, jasmonic
acid.
Introduction
Plants have pre-formed and inducible mechanisms that
protect them from pathogens. Systemic acquired resistance
(SAR) is an inducible defense mechanism that confers
enhanced resistance against a broad-spectrum of pathogens
(Chaturvedi and Shah, 2006; Durrant and Dong, 2004;
Metraux et al., 2002). SAR is activated in the healthy leaves
of a plant subsequent to the exposure of another leaf to a
pathogen. The pathogen-inoculated leaf is the source of a
long-distance SAR signal that is translocated to the other
organs (Dean and Kuc, 1986). Allograft experiments between
cucumber (Cucumis sativus), muskmelon (Cucumis melo)
and watermelon (Citrullus lanatus) demonstrated that the
SAR signal can function across Cucurbit species (Kuc, 1982).
Furthermore, girdling experiments in cotton (Gossypium
106 ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd
The Plant Journal (2008) 54, 106–117 doi: 10.1111/j.1365-313X.2007.03400.x

hirsutum) suggested that the SAR-signal is translocated
through the phloem (Guedes et al., 1980). However, experi-
ments with Arabidopsis thaliana have indicated that phloem
is not the exclusive conduit for translocation of the SAR
signal; some signal can be translocated via other routes
(Kiefer and Slusarenko, 2003).
The activation of SAR is accompanied by an increase in
the level of salicylic acid (SA) and elevated expression of the
pathogenesis-related PR1 gene in the pathogen inoculated
and the distal organs in which SAR is manifested (Chaturv-
edi and Shah, 2006; Durrant and Dong, 2004; Metraux et al.,
2002). Studies in tobacco (Nicotiana tabaccum) and Arabid-
opsis have confirmed an important role for SA in the
manifestation of SAR (Chaturvedi and Shah, 2006; Durrant
and Dong, 2004; Metraux et al., 2002). For example, appli-
cation of SA induces a SAR-like mechanism. Furthermore,
preventing accumulation of SA by either blocking its
synthesis, as in the Arabidopsis salicylic acid-induction
deficient2 (sid2) and enhanced disease susceptibility5
(eds5) mutants, or promoting its breakdown by expression
of the bacterial nahG-encoded salicylate hydroxylase in
transgenic plants compromised SAR (Nawrath and Metraux,
1999; Vernooij et al., 1994). SAR was also attenuated in the
Arabidopsis nonexpresser of PR genes1 (npr1) mutant,
which is defective in SA signaling (Maldonado et al., 2002;
Nandi et al., 2004). Although SA moves from the pathogen-
inoculated organ to the distal organs (Molders et al., 1996;
Shulaev et al., 1995), leaf excision experiments in cucumber
and grafting experiments involving tobacco NahG plants
suggested that SA is not the primary mobile signal that is
translocated from the pathogen-inoculated leaf to the distal
organs for establishment of SAR (Rasmussen et al., 1991;
Vernooij et al., 1994). However, Darby et al. (2000) demon-
strated that nahG expression when targeted to the phloem
tissue compromised SAR. Furthermore, a recent study with
SA-binding protein2 (SABP2), which possesses methylSA
(MeSA) esterase activity, indicated that MeSA is required for
long-distance signaling associated with Tobacco mosaic
virus-induced SAR in N. tabaccum (Park et al., 2007).
Jasmonic acid (JA) is another important signal molecule
in plant stress responses (Devoto and Turner, 2003; Howe
and Schilmiller, 2002; Pieterse et al., 2002). In tomato, JA
functions as a graft transmissible signal in the wound
response (Li et al., 2002) and in Arabidopsis, JA signaling
is required for systemic disease resistance induced by root-
colonizing bacteria (Devoto and Turner, 2003; Pieterse et al.,
2002). A recent study showed that JA signaling is also
involved in SAR that was induced by localized inoculation
with an avirulent (Avr) strain of Pseudomonas syringae
(Truman et al., 2007); systemic resistance was attenuated in
the JA-response jasmonate insensitive1 (jin1) mutant and
the oxophytodienoic acid reductase3 (opr3) mutant, which is
JA deficient. However, two other studies showed that SAR
was not compromised in the JA-insensitive Arabidopsis
coronatine-insensitive1 (coi1) and jasmonate resistant1
(jar1) mutants (Cui et al., 2005; Mishina and Zeier, 2007).
Genetic studies in Arabidopsis have suggested the
involvement of lipids in long-distance signaling in the
establishment of SAR (Chaturvedi and Shah, 2006; Durrant
and Dong, 2004; Shah, 2005). The DEFECTIVE IN INDUCED
RESISTANCE1 (DIR1) gene, which encodes a putative apo-
plastic protein with homology to non-specific lipid transfer
proteins (nsLTPs), is required for the activation of SAR
(Maldonado et al., 2002). Unlike petiole exudates (PeXs)
collected from Avr pathogen inoculated wild-type (WT)
leaves, Avr PeX collected from the dir1 mutant were unable
to induce PR1 expression when applied to WT leaves.
However, the dir1 mutant leaves were responsive to the
PR1 expression-inducing activity present in Avr PeXs from
WT leaves. These results suggest that DIR1 is required for
the synthesis and/or translocation of the SAR signal. In
contrast to dir1, SAR was constitutively manifested in the
Arabidopsis suppressor of SA-insensitivity2 (ssi2) mutant
that constitutively expressed PR1 and exhibited heightened
resistance to bacterial, viral and oomycete pathogens
(Kachroo et al., 2003; Nandi et al., 2003; Sekine et al., 2004;
Shah et al., 2001). SSI2 encodes a stearoyl-acyl carrier
protein desaturase involved in lipid metabolism (Kachroo
et al., 2001). ssi2-conditioned constitutive PR1 expression
and resistance were attenuated by the suppressor of fatty
acid desaturase deficiency1 (sfd1) mutant allele in the ssi2
sfd1 double-mutant plants (Nandi et al., 2003). Studies with
the sfd1 single mutant indicated that SFD1 is essential for
SAR, but not basal resistance to pathogen (Nandi et al.,
2004). Although SA accumulation was not adversely affected
in pathogen-infected sfd1 leaves, SAR-associated accumu-
lation of SA in the distal leaves was blocked in sfd1,
suggesting that SFD1 is required prior to accumulation of
SA in the organs exhibiting SAR (Nandi et al., 2004). SFD1
encodes a dihydroxyacetonephosphate (DHAP) reductase
that catalyzes the interconversion between DHAP and glyc-
erol-3-phosphate (G3P). Glycerol-3-phosphate provides car-
bon for a variety of macromolecules including the glycerol
backbone in complex lipids. Leaf galactolipid composition
was altered in the sfd1 mutant. The level of the 34:6 (18:3
plus 16:3 acyl composition)-monogalactosyldiacylglycerol
(MGD) was 45% lower, while the level of 36:6 (18:3 plus 18:3
acyl composition)-MGD was two-fold higher in the leaves of
the sfd1 mutant than in the WT plant (Nandi et al., 2004).
Since, SFD1 function is required in the plastids it was
suggested that a plastid-synthesized lipid may be critical for
SAR (Nandi et al., 2004).
In this study, we demonstrate that in addition to SFD1, the
Arabidopsis FATTY ACID DESATURASE7 (FAD7), SFD2
and MONOGALACTOSYLDIACYLGLYCEROL SYNTHASE1
(MGD1) genes, which are also involved in the synthesis of
plastidic glycerolipids, are required for the activation of SAR.
These genes are vital for the accumulation of a factor that,
Plant defense signaling 107
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

along with the DIR1-encoded nsLTP, is required in PeXs for
the activation of SAR. Although FAD7-synthesized lipids
provide fatty acids for JA synthesis, we provide evidence
that JA is not a component of this SAR inducing activity in
Arabidopsis PeXs.
Results
SAR is compromised in the Arabidopsis fad7, sfd2 and
mgd1 mutants
To determine whether a plastid-synthesized lipid is involved
in SAR, we studied the activation of SAR-conferred resis-
tance in the Arabidopsis fad7 and sfd2 mutants in which the
composition of plastid-synthesized phosphatidylglycerol
(PG), MGD and digalactosyldiacylglycerol (DGD) are altered
relative to the WT plant (Tables S1 and S2) (Nandi et al.,
2003; Wallis and Browse, 2002). In addition, the fad7 muta-
tion also affects plastid synthesized sulfoglycerolipid (SL)
composition (Wallis and Browse, 2002). Figure 1 summa-
rizes the involvement of the FAD7 and SFD2 genes in Ara-
bidopsis lipid biosynthesis. To monitor SAR-conferred
resistance, 3 days after the primary inoculation with the Avr
pathogen, P. syringae pv. tomato DC3000 expressing
avrRpt2, four upper leaves of the plant were challenged
(secondary inoculation) with the virulent pathogen P. sy-
ringae pv. maculicola ES4326 (Psm) and bacterial numbers
monitored 3 days post the secondary inoculation. Psm-
challenged plants that received a primary inoculation with
10 mM MgCl2 (Mock) served as the control. As expected of
SAR, Psm numbers were lower in leaves of WT accessions
Nossen (No) and Columbia (Col), and Col containing the
glabra1 allele (gl) that received a primary inoculation of the
Avr pathogen, than in plants that received a Mock primary
inoculation (Figure 2a). In contrast, comparable numbers
of Psm were present in the upper leaves of the Mock and
Avr pathogen-treated fad7 and sfd2 mutants (Figure 2a).
SAR-conferred induction of PR1 expression in the upper
leaves was also attenuated in the fad7 and sfd2 mutants
(Figure 2b). Growth of Psm in fad7 and sfd2 that received a
Mock primary inoculation was comparable to that in the
Col-gl and No, which contain the WT FAD7 and SFD2 alleles,
suggesting that these mutations do not have any effect on
basal resistance against Psm. Furthermore, the onset of
the hypersensitive response (HR) induced in response to
primary inoculation with the Avr pathogen was comparable
between the WT and the mutant plants (data not shown).
The fad7 mutant allele also attenuated the ssi2-conditioned
constitutive PR1 expression and resistance to Psm in two
independent ssi2 fad7 double-mutant lines (Figure 2c),
confirming that FAD7 is required for SAR. Our previous
studies had showed that the ssi2-conferred constitutive
SAR phenotype was similarly attenuated in the ssi2 sfd1
and ssi2 sfd2 double-mutant plants (Nandi et al., 2003).
The level of 34:4 PG, the major PG species in Arabidopsis
leaves is 30% lower in the fad7 mutant than in the WT plant
(Table S1) (Wallis and Browse, 2002). Similarly, in compar-
ison to the WT, levels of PG species containing polyunsat-
urated fatty acids were lower in the sfd2 mutant. To
determine whether PG contributes to SAR, SAR was
evaluated in the fad4 mutant in which 34:4 PG level is
reduced by 99% and total PG content is 40% lower than in
the WT plant; however, the fad4 allele does not affect
galactolipid and SL content (Table S1) (Wallis and Browse,
2002). As shown in Figure 2(a), SAR-conferred resistance to
Psm was comparable between the WT accession Col and
the fad4 mutant, indicating that PG is not important
for SAR.
The involvement of plastid glycerolipids in SAR was
further tested by studying SAR-conferred resistance against
Psm in the Arabidopsis mgd1 mutant (Jarvis et al., 2000).
MGD1 encodes a MGD synthase activity that catalyzes the
transfer of a D-galactose moiety from UDP-galactose to
diacylglycerol to synthesize MGD in the plastids (Figure 1)
(Arai et al., 2001; Jarvis et al., 2000). Levels of MGD in the
leaves of the mgd1 mutant are reduced by 75% compared
with the WT (Jarvis et al., 2000). However, levels of PG and
SL are not reduced in the mgd1 mutant (Jarvis et al., 2000).
Similar to the fad7, sfd1 and sfd2 mutants, SAR, but not
basal resistance, was compromised in the mgd1 mutant
(Figure 2a). The above results establish that a plastid
synthesized glycerolipid, most likely a galactolipid, is
required for the establishment of SAR.
sfd1
DHAP
G3P LPA PA DAG
MGDPG
mgd1
fad4sfd2
fad7
MGDPG
fad7
DGD
DGD
fad7
sfd2
SL
fad7
sfd2
Figure 1. Schematic of steps in phosphatidylglycerol (PG), galactolipid and
sulfoglycerolipid (SL) synthesis in plastids affected by the sfd1, sfd2, fad4,
fad7 and mgd1 mutants.
Steps leading to synthesis of PG, the galactolipids monogalactosyldiacyl-
glycerol (MGD) and digalactosyldiacylglycerol (DGD) and SL in the plastids
are shown. The acyl chains on PG, MGD, DGD and SL are desaturated by
specific fatty acyl desaturases. SFD1 encodes a dihydroxyacetonephosphate
(DHAP) reductase that provides glycerol-3-phosphate (G3P) for glycerolipid
synthesis in the plastids. MGD1 encodes an enzyme that transfers a
D-galactose moiety onto diacylglycerol (DAG) to yield MGD. FAD4-encoded
desaturase primarily acts on 16:0 acyl chains at the sn2 position in PG to yield
16:1. The identity of the SFD2 gene is not known. However, lipid profiling
indicates that the sfd2 allele affects the desaturation of 16:0 acyl chains
present in MGD and DGD to 16:1. The FAD7-encoded desaturase catalyzes the
conversion of 18:2 and 16:2 acyl chains in PG, MGD, DGD and SL to 18:3 and
16:3, respectively.
LPA, lysophosphatidic acid; PA, phosphatidic acid.
108 Ratnesh Chaturvedi et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

Petiole exudates from avirulent pathogen-inoculated
wild- type leaves contain a SAR-inducing activity
Previously, Maldonado et al. (2002) showed that PeXs, which
are enriched in phloem sap, collected from Avr pathogen-
inoculated leaves of WT Arabidopsis contained an activity
that, when infiltrated into Arabidopsis leaves, induced accu-
mulation of PR1 transcript. However, whether these exudates
(Avr PeX) from Avr pathogen-inoculated leaves promote a
SAR-like mechanism was not known. We similarly observed
that in comparison to PeX collected from 10 mM MgCl2-
infiltrated (Mock PeX) WT leaves, Avr PeX contained an
activity that enhanced PR1 transcript accumulation in the
infiltrated leaf (Figure 3a). In addition, PR1 transcripts also
accumulated at elevated levels in the upper leaves of Avr
PeX-treated plants. More significantly, resistance to Psm was
enhanced in the local Avr PeX-infiltrated (Figure 3b) and
upper leaves of WT plants (Figure 4a–c and Figure S3). When
applied to healthy plants, Avr PeX did not induce necrosis.
Furthermore, Avr PeX did not exhibit bacteriostatic activity
against Psm (data not shown), suggesting that Avr PeX-
conferred suppression of P. syringae growth in planta was
due to the activation of plant defense responses. The ability of
Avr PeX to promote disease resistance required SA synthesis
and signaling in the recipient plant; Avr PeX did not promote
resistance in the SA-deficient Arabidopsis sid2 and eds5
mutants, and the SA-insensitive npr1 mutant plants (Fig-
ure 3b). The Avr PeX from WT leaves also restricted Fusarium
head blight disease of wheat caused by the fungal pathogen
Fusarium graminearum (Figure 3c) and Psm growth in
tomato leaves (Figure S1). These results indicate that Avr
PeX from WT Arabidopsis leaves contains a SAR-inducing
activity. No difference in the salicylate content between
the Avr PeX (0.26 � 0.10 ng per leaf) and Mock PeX
(0.27 � 0.10 ng per leaf) were found.
AvrPeX from fad7, sfd1 and sfd2 lack the
SAR-inducing activity
The FAD7, SFD1 and SFD2 genes may be involved in either
the accumulation of a SAR-inducing activity in the Avr
pathogen-inoculated leaf or perception of this activity in the
upper leaves which exhibit SAR. To distinguish between the
above possibilities we collected Avr PeX and Mock PeX from
the fad7, sfd1 and sfd2 mutants and the corresponding WT
plants. Petiole exudates from the WT plant were infiltrated
into leaves of the fad7, sfd1 and sfd2 plants, and as a control
into the WT plant, to determine whether they could promote
fad7WT(Col-gl)
WT(Nö) sfd2C
FU
leaf
dis
c–1WT(Col) fad4
105
106
107
105
106
107
105
104
106
107mgd1WT(Col)
106
107
108(a)
(b) (c)
106
107
108
CF
U le
af d
isc–1
CF
U le
af d
isc–1
WT(Nö)
fad7 ssi2 fad7ssi2
PR1
ACT8
Mock Avr Mock Avr
WT(Col-gl) fad7
PR1
ACT8
Mock Avr Mock Avr
WT(Nö) sfd2
PR1
ACT8
Figure 2. Systemic acquired resistance (SAR) is
compromised in the fad7, mgd1 and sfd2
mutants.
(a) Growth of the virulent pathogen P. syringae
pv. maculicola (Psm) in wild type (WT), fad7,
mgd1, sfd2 and fad4 mutant plants. Pseudomo-
nas syringae pv. tomato DC3000 carrying the
avrRpt2 avirulence gene was infiltrated into three
lower leaves of fad7, sfd2, mgd1 and fad4 mutant
plants and their corresponding WT plants. Plants
similarly treated with 10 mM MgCl2 provided
controls. Three days later, four upper leaves in
each plant were infiltrated with Psm and bacterial
numbers monitored 3 days post-inoculation
(dpi). Each bar represents the average Psm count
in 15 leaf discs � SD. White bars, primary inoc-
ulation with 10 mM MgCl2; black bars, primary
inoculation with Avr pathogen. The mgd1 and
fad4 mutants are in the accession Columbia
(Col), the fad7 mutant is in accession Col con-
taining the glabra allele (gl), and the sfd2 mutant
is in the accession Nossen (No) background.
(b) Reverse transcription-PCR analysis for PR1
expression in the upper leaves of WT, fad7 and
sfd2 mutants, 2 days post-infiltration of three
lower leaves with 10 mM MgCl2 (Mock) or the
avirulent pathogen (Avr).
(c) Upper panel: bacterial counts in Psm-inocu-
lated leaves of WT accession No, the fad7 and
ssi2 single mutant and two ssi2 fad7 double-
mutant lines, 3 dpi. Each bar represents the
average Psm count in 15 leaf discs � SD. Lower
panel: RT-PCR analysis for PR1 expression in
leaves of the WT, fad7, ssi2 and ssi2 fad7 mutant
plants.
In (b) and (c) ACT8 provided a control for RT-PCR.
Plant defense signaling 109
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

resistance against Psm in the upper leaves. In parallel, PeXs
from the mutant leaves were infiltrated into WT leaves, and
as a control into the corresponding mutant plant, to deter-
mine whether they contained a factor that could activate
SAR-associated resistance in WT leaves. Resistance against
Psm was enhanced in the fad7, sfd1 and sfd2 mutant plants
that were treated with Avr PeX collected from the corre-
sponding WT plant (Figure 4a–c), demonstrating that these
mutants are capable of responding to the SAR signal. In
contrast, Avr PeX collected from the fad7, sfd1 and sfd2
CF
U le
af d
isc–1
104
105
106
107
WT(Col)
sid2 eds5 npr1
PR1ACT8
Inf Up
Mock Pe(a)
(b)
(c)
X Avr PeX
Inf UpIn
fect
ed s
pike
lets
(%
)
WT PeX sfd1 PeX
Mock Avr 0
20
40
60
80
3 DPI9 DPI
15 DPI21 DPI
Mock Avr
Mock PeX Avr PeX
Figure 3. Systemic resistance conferred by petiole exudates (PeXs) from
avirulent (Avr) pathogen-inoculated Arabidopsis leaves requires salicylic acid
(SA) signaling.
(a) Petiole exudates (Mock PeX and Avr PeX) were collected from MgCl2- and
Avr pathogen-inoculated leaves of WT accession No plants. The PeXs were
infiltrated into three leaves of each WT plant of accession No. Twenty-four
hours later RNA was extracted from the infiltrated (Inf) and upper (Up) leaves
and analyzed by RT-PCR for PR1 transcript accumulation. Reverse transcrip-
tase-PCR for ACT8 provided control for the quality of RNA.
(b) Petiole exudates from WT accession No leaves were infiltrated into leaves
of the WT accession Col and sid2, eds5 and npr1 mutant plants, which are in
the accession Col background. Three days later, these leaves were challenged
with Pseudomonas syringae pv. maculicola (Psm) and bacterial numbers
determined 3 days post-inoculation (dpi). Each bar represents the average
Psm count in 15 leaf discs � SD. White bars, fractions from Mock PeX; black
bars, fractions from Avr PeX.
(c) The SAR-competent Avr PeXs from Arabidopsis promote Fusarium head
blight resistance in wheat. Top panel: Fusarium graminearum growth on
spikes that were pre-treated with either 10 ll of Mock PeX or Avr PeX from WT
Arabidopsis plants of accession No. A single spikelet on each spike was
treated with PeX and 3 days later inoculated with 300 spores of Fusarium
graminearum. The inoculated spikes were photographed 7 dpi. Bottom panel:
quantification of severity of Fusarium head blight disease in F. graminearum-
inoculated spikes that were pre-treated with Mock PeX and Avr PeX from WT
plants of accession No and sfd1 mutant. Head blight severity was measured
as the percentage of spikelets on each spike showing blight symptoms at 3, 9,
15 and 21 dpi with fungal spores. All values are mean of seven spikes per
treatment.
WT(Col-gl) fad7
WT(Col-gl)
WT(Col-gl)
fad7
fad7104
105
10
(a)
(b)
(c)
6
CF
U le
af d
isc–1
CF
U le
af d
isc–1
CF
U le
af d
isc–1
104
WT(Nö) sfd1
WT(Nö) sfd1 sfd1WT(Nö)
105
106
WT(Nö) sfd2
WT(Nö) sfd2 sfd2WT(Nö)104
105
106
Source of PeX
Source of PeX
Source of PeX
Figure 4. Avirulent (Avr) petiole exudates (PeXs) from fad7, sfd1 and sfd2 lack
a systemic acquired resistance-inducing activity.
Mock PeX and Avr PeX were collected from the fad7, sfd1 and sfd2 mutants
and the corresponding wild-type (WT) plants. These PeXs were infiltrated into
three lower leaves of WT (left panel) and the mutant (right panel) plants. Three
days later, four upper leaves in each plant were challenged with Pseudomo-
nas syringae pv. maculicola (Psm) and bacterial counts determined 3 days
post-inoculation (dpi). Each bar represents the average Psm count in 15 leaf
discs � SD. White bars, fractions from Mock PeX; black bars, fractions from
Avr PeX.
(a) PeXs collected from Col-gl, which contains the WT FAD7 allele, and the
fad7 mutant.
(b) PeXs collected from WT plants of accession No and the sfd1 mutant.
(c) PeXs collected from WT plants of accession No and the sfd2 mutant.
110 Ratnesh Chaturvedi et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

mutants, when applied to the corresponding WT plant did
not promote resistance against Psm (Figure 4a–c). The Avr
PeX from sfd1 did not promote resistance in wheat and
tomato either (Figure 3c and Figure S1). Furthermore,
co-infiltration of fad7 plus sfd1 Avr PeXs and fad7 plus sfd2
Avr PeXs also did not activate SAR in Arabidopsis (Figure 5a
and Figure S2). We conclude that the SAR deficiency of the
fad7, sfd1 and sfd2 mutants is due to their inability to
accumulate a SAR-inducing activity in Avr PeX.
The SAR-activating activity is reconstituted by mixing
Avr petiole exudates from dir1 with Avr petiole
exudates from fad7 and sfd1
Like the fad7, sfd1 and sfd2 mutant alleles, the SAR-incom-
petent dir1 is also epistatic to ssi2. The ssi2-conditioned
constitutive PR1 expression and heightened resistance to
Psm were suppressed in the ssi2 dir1 double mutant
(Figure 6a,b), suggesting that DIR1 and the plastid glycer-
olipid-dependent factor are both required for a common
defense mechanism. Like the sfd1, sfd2 and fad7 mutants,
the dir1 mutant is unable to accumulate a SAR-promoting
activity in Avr PeX (Figure S3) (Maldonado et al., 2002).
However, the dir1 mutant was responsive to the SAR-pro-
moting activity present in Avr PeX collected from WT leaves.
To characterize the relationship between DIR1 and the plastid
glycerolipid-dependent factor in SAR long-distance signal-
ing we mixed Avr PeX from dir1 with Avr PeX collected from
the sfd1 mutant. These mixed PeXs were then infiltrated into
leaves of WT No plants and after 3 days the upper leaves
were challenged with Psm. As negative controls, Mock PeX
from dir1 was mixed with Mock PeX from sfd1 and similarly
infiltrated into WT No plants. The Mock and Avr PeXs from
the sfd1 and dir1 mutants when injected individually, and
Mock PeXs from the mutants when mixed together, did
not enhance resistance to Psm (Figure 5b). In contrast,
co-application of Avr PeXs from sfd1 plus dir1 to WT No
plants resulted in heightened resistance to Psm (Figure 5b)
and also activated PR1 expression (Figure S4). Similarly,
co-application of Avr PeXs from fad7 plus dir1 also enhanced
resistance to Psm (Figure 5c). These results suggest that the
accumulation of the DIR1-derived factor and the plastid
glycerolipid-dependent factor in Avr PeX is independent of
the other. Furthermore, both factors are required together in
the Avr PeX for long-distance signaling in the establishment
of SAR.
JA is not a component of the SAR-inducing activity
present in Arabidopsis petiole exudates
Recently, it was shown that JA signaling is required for the
SAR-conferred restriction of pathogen growth in Arabidop-
sis (Truman et al., 2007). Moreover, as shown in Figure 7(a),
the JA level was 15-fold higher in Avr PeX than in Mock PeX
of WT leaves, raising the possibility that JA is a component
of the SAR-inducing activity present in Avr PeX. Since FAD7-
synthesized glycerolipids provide fatty acids for the
10 4
10 5
10 6
10 7
WT (Col-gl)
fad7 dir1 fad7 +
dir 1 Source of PeX
Source of PeX
104
10 5
10 6
WT (Nö)
sfd1 dir1 sf d1 +
dir1
WT (Nö)
sf d1 fad7 10 5
10 6
10
(a)
(b)
(c)
7
sf d1 +
fad7 Source of PeX
CF
U le
af d
isc–1
C
FU
leaf
dis
c–1
CF
U le
af d
isc–1
Figure 5. Co-application of avirulent (Avr) petiole exudate (PeX) from sfd1
and fad7 with Avr PeX from dir1 activates systemic acquired resistance in
wild-type (WT) plants.
Mock PeX and Avr PeX were collected from WT and mutant plants. These
PeXs were applied to three lower leaves of WT accession No plant. In addition,
equal volumes of PeXs from the mutant plants were mixed and infiltrated into
leaves of the WT accession No plant. Three days later four upper leaves of
each plant were challenged with Pseudomonas syringae pv. maculicola (Psm)
and bacterial numbers were determined 3 days post-inoculation (dpi). Each
bar represents the average Psm count in 15 leaf discs � SD. White bars,
fractions from Mock PeX; black bars, fractions from Avr PeX.
(a) The PeXs were collected from WT plants of accession No and the sfd1 and
fad7 mutant plants.
(b) The PeXs were collected from No and the sfd1 and dir1 mutant plants.
(c) The PeXs were collected from WT plants of accession Col-gl and the fad7
and dir1 mutant plants.
Plant defense signaling 111
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117
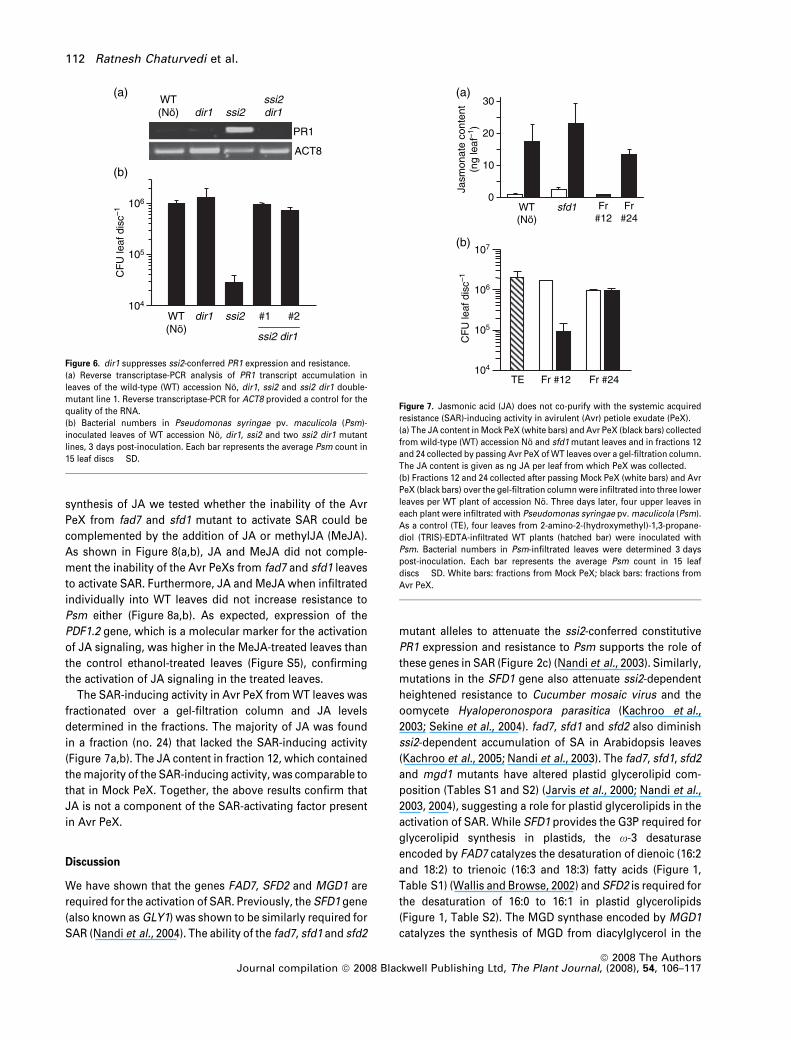
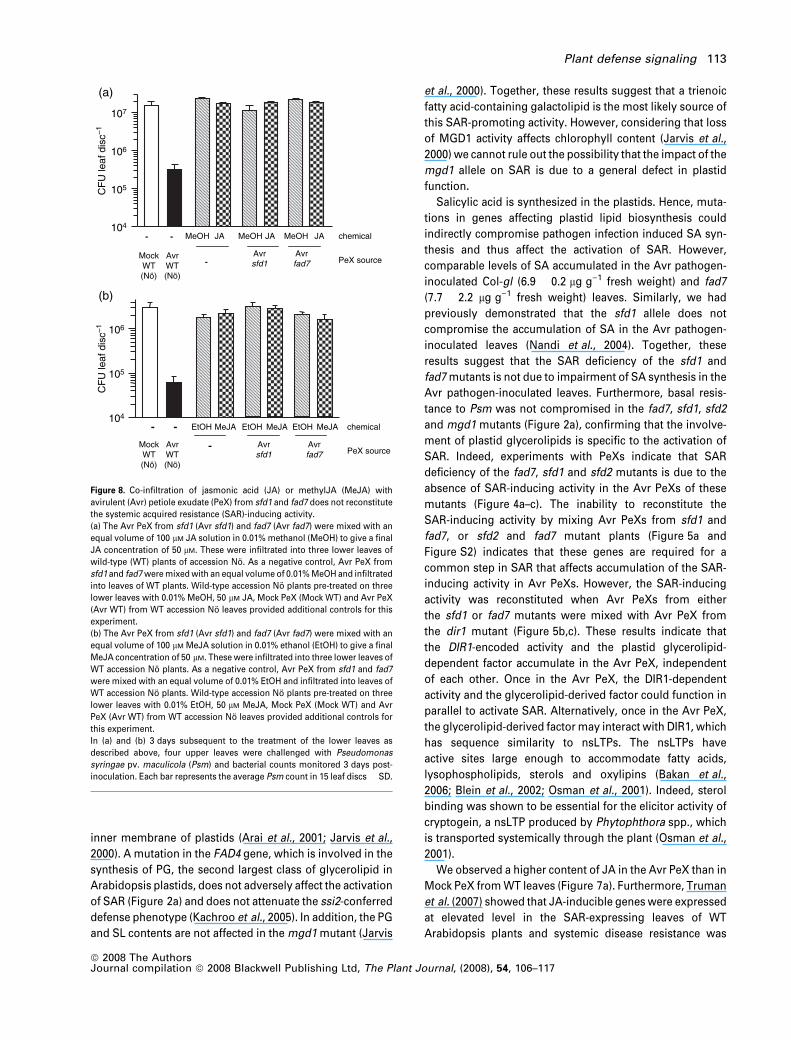
synthesis of JA we tested whether the inability of the Avr
PeX from fad7 and sfd1 mutant to activate SAR could be
complemented by the addition of JA or methylJA (MeJA).
As shown in Figure 8(a,b), JA and MeJA did not comple-
ment the inability of the Avr PeXs from fad7 and sfd1 leaves
to activate SAR. Furthermore, JA and MeJA when infiltrated
individually into WT leaves did not increase resistance to
Psm either (Figure 8a,b). As expected, expression of the
PDF1.2 gene, which is a molecular marker for the activation
of JA signaling, was higher in the MeJA-treated leaves than
the control ethanol-treated leaves (Figure S5), confirming
the activation of JA signaling in the treated leaves.
The SAR-inducing activity in Avr PeX from WT leaves was
fractionated over a gel-filtration column and JA levels
determined in the fractions. The majority of JA was found
in a fraction (no. 24) that lacked the SAR-inducing activity
(Figure 7a,b). The JA content in fraction 12, which contained
the majority of the SAR-inducing activity, was comparable to
that in Mock PeX. Together, the above results confirm that
JA is not a component of the SAR-activating factor present
in Avr PeX.
Discussion
We have shown that the genes FAD7, SFD2 and MGD1 are
required for the activation of SAR. Previously, the SFD1 gene
(also known as GLY1) was shown to be similarly required for
SAR (Nandi et al., 2004). The ability of the fad7, sfd1 and sfd2
mutant alleles to attenuate the ssi2-conferred constitutive
PR1 expression and resistance to Psm supports the role of
these genes in SAR (Figure 2c) (Nandi et al., 2003). Similarly,
mutations in the SFD1 gene also attenuate ssi2-dependent
heightened resistance to Cucumber mosaic virus and the
oomycete Hyaloperonospora parasitica (Kachroo et al.,
2003; Sekine et al., 2004). fad7, sfd1 and sfd2 also diminish
ssi2-dependent accumulation of SA in Arabidopsis leaves
(Kachroo et al., 2005; Nandi et al., 2003). The fad7, sfd1, sfd2
and mgd1 mutants have altered plastid glycerolipid com-
position (Tables S1 and S2) (Jarvis et al., 2000; Nandi et al.,
2003, 2004), suggesting a role for plastid glycerolipids in the
activation of SAR. While SFD1 provides the G3P required for
glycerolipid synthesis in plastids, the x-3 desaturase
encoded by FAD7 catalyzes the desaturation of dienoic (16:2
and 18:2) to trienoic (16:3 and 18:3) fatty acids (Figure 1,
Table S1) (Wallis and Browse, 2002) and SFD2 is required for
the desaturation of 16:0 to 16:1 in plastid glycerolipids
(Figure 1, Table S2). The MGD synthase encoded by MGD1
catalyzes the synthesis of MGD from diacylglycerol in the
TE104
105
106
107
Fr #12 Fr #24
Jasm
onat
e co
nten
t(n
g le
af–1
)
0
10
20
30(a)
(b)
WT(Nö)
sfd1 Fr#12
Fr#24
CF
U le
af d
isc–1
Figure 7. Jasmonic acid (JA) does not co-purify with the systemic acquired
resistance (SAR)-inducing activity in avirulent (Avr) petiole exudate (PeX).
(a) The JA content in Mock PeX (white bars) and Avr PeX (black bars) collected
from wild-type (WT) accession No and sfd1 mutant leaves and in fractions 12
and 24 collected by passing Avr PeX of WT leaves over a gel-filtration column.
The JA content is given as ng JA per leaf from which PeX was collected.
(b) Fractions 12 and 24 collected after passing Mock PeX (white bars) and Avr
PeX (black bars) over the gel-filtration column were infiltrated into three lower
leaves per WT plant of accession No. Three days later, four upper leaves in
each plant were infiltrated with Pseudomonas syringae pv. maculicola (Psm).
As a control (TE), four leaves from 2-amino-2-(hydroxymethyl)-1,3-propane-
diol (TRIS)-EDTA-infiltrated WT plants (hatched bar) were inoculated with
Psm. Bacterial numbers in Psm-infiltrated leaves were determined 3 days
post-inoculation. Each bar represents the average Psm count in 15 leaf
discs � SD. White bars: fractions from Mock PeX; black bars: fractions from
Avr PeX.
104
105
106
ssi2 dir1
WT(Nö)
dir1 ssi2 #1 #2
WT(a)
(b)
(Nö) dir1 ssi2ssi2dir1
PR1
ACT8
CF
U le
af d
isc–1
Figure 6. dir1 suppresses ssi2-conferred PR1 expression and resistance.
(a) Reverse transcriptase-PCR analysis of PR1 transcript accumulation in
leaves of the wild-type (WT) accession No, dir1, ssi2 and ssi2 dir1 double-
mutant line 1. Reverse transcriptase-PCR for ACT8 provided a control for the
quality of the RNA.
(b) Bacterial numbers in Pseudomonas syringae pv. maculicola (Psm)-
inoculated leaves of WT accession No, dir1, ssi2 and two ssi2 dir1 mutant
lines, 3 days post-inoculation. Each bar represents the average Psm count in
15 leaf discs � SD.
112 Ratnesh Chaturvedi et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117
inner membrane of plastids (Arai et al., 2001; Jarvis et al.,
2000). A mutation in the FAD4 gene, which is involved in the
synthesis of PG, the second largest class of glycerolipid in
Arabidopsis plastids, does not adversely affect the activation
of SAR (Figure 2a) and does not attenuate the ssi2-conferred
defense phenotype (Kachroo et al., 2005). In addition, the PG
and SL contents are not affected in the mgd1 mutant (Jarvis
et al., 2000). Together, these results suggest that a trienoic
fatty acid-containing galactolipid is the most likely source of
this SAR-promoting activity. However, considering that loss
of MGD1 activity affects chlorophyll content (Jarvis et al.,
2000) we cannot rule out the possibility that the impact of the
mgd1 allele on SAR is due to a general defect in plastid
function.
Salicylic acid is synthesized in the plastids. Hence, muta-
tions in genes affecting plastid lipid biosynthesis could
indirectly compromise pathogen infection induced SA syn-
thesis and thus affect the activation of SAR. However,
comparable levels of SA accumulated in the Avr pathogen-
inoculated Col-gl (6.9 � 0.2 lg g)1 fresh weight) and fad7
(7.7 � 2.2 lg g)1 fresh weight) leaves. Similarly, we had
previously demonstrated that the sfd1 allele does not
compromise the accumulation of SA in the Avr pathogen-
inoculated leaves (Nandi et al., 2004). Together, these
results suggest that the SAR deficiency of the sfd1 and
fad7 mutants is not due to impairment of SA synthesis in the
Avr pathogen-inoculated leaves. Furthermore, basal resis-
tance to Psm was not compromised in the fad7, sfd1, sfd2
and mgd1 mutants (Figure 2a), confirming that the involve-
ment of plastid glycerolipids is specific to the activation of
SAR. Indeed, experiments with PeXs indicate that SAR
deficiency of the fad7, sfd1 and sfd2 mutants is due to the
absence of SAR-inducing activity in the Avr PeXs of these
mutants (Figure 4a–c). The inability to reconstitute the
SAR-inducing activity by mixing Avr PeXs from sfd1 and
fad7, or sfd2 and fad7 mutant plants (Figure 5a and
Figure S2) indicates that these genes are required for a
common step in SAR that affects accumulation of the SAR-
inducing activity in Avr PeXs. However, the SAR-inducing
activity was reconstituted when Avr PeXs from either
the sfd1 or fad7 mutants were mixed with Avr PeX from
the dir1 mutant (Figure 5b,c). These results indicate that
the DIR1-encoded activity and the plastid glycerolipid-
dependent factor accumulate in the Avr PeX, independent
of each other. Once in the Avr PeX, the DIR1-dependent
activity and the glycerolipid-derived factor could function in
parallel to activate SAR. Alternatively, once in the Avr PeX,
the glycerolipid-derived factor may interact with DIR1, which
has sequence similarity to nsLTPs. The nsLTPs have
active sites large enough to accommodate fatty acids,
lysophospholipids, sterols and oxylipins (Bakan et al.,
2006; Blein et al., 2002; Osman et al., 2001). Indeed, sterol
binding was shown to be essential for the elicitor activity of
cryptogein, a nsLTP produced by Phytophthora spp., which
is transported systemically through the plant (Osman et al.,
2001).
We observed a higher content of JA in the Avr PeX than in
Mock PeX from WT leaves (Figure 7a). Furthermore, Truman
et al. (2007) showed that JA-inducible genes were expressed
at elevated level in the SAR-expressing leaves of WT
Arabidopsis plants and systemic disease resistance was
Avrfad7
Avrsfd1
-MockWT(Nö)
AvrWT(Nö)
- - MeJAEtOH MeJAEtOH MeJAEtOH chemical104
105
106
104
105
106
10
(a)
(b)
7
- -
AvrWT(Nö)
MockWT(Nö)
Avrsfd1
Avrfad7-
JAMeOH chemicalJAMeOH JAMeOH
PeX source
PeX source
CF
U le
af d
isc–1
CF
U le
af d
isc–1
Figure 8. Co-infiltration of jasmonic acid (JA) or methylJA (MeJA) with
avirulent (Avr) petiole exudate (PeX) from sfd1 and fad7 does not reconstitute
the systemic acquired resistance (SAR)-inducing activity.
(a) The Avr PeX from sfd1 (Avr sfd1) and fad7 (Avr fad7) were mixed with an
equal volume of 100 lM JA solution in 0.01% methanol (MeOH) to give a final
JA concentration of 50 lM. These were infiltrated into three lower leaves of
wild-type (WT) plants of accession No. As a negative control, Avr PeX from
sfd1 and fad7 were mixed with an equal volume of 0.01% MeOH and infiltrated
into leaves of WT plants. Wild-type accession No plants pre-treated on three
lower leaves with 0.01% MeOH, 50 lM JA, Mock PeX (Mock WT) and Avr PeX
(Avr WT) from WT accession No leaves provided additional controls for this
experiment.
(b) The Avr PeX from sfd1 (Avr sfd1) and fad7 (Avr fad7) were mixed with an
equal volume of 100 lM MeJA solution in 0.01% ethanol (EtOH) to give a final
MeJA concentration of 50 lM. These were infiltrated into three lower leaves of
WT accession No plants. As a negative control, Avr PeX from sfd1 and fad7
were mixed with an equal volume of 0.01% EtOH and infiltrated into leaves of
WT accession No plants. Wild-type accession No plants pre-treated on three
lower leaves with 0.01% EtOH, 50 lM MeJA, Mock PeX (Mock WT) and Avr
PeX (Avr WT) from WT accession No leaves provided additional controls for
this experiment.
In (a) and (b) 3 days subsequent to the treatment of the lower leaves as
described above, four upper leaves were challenged with Pseudomonas
syringae pv. maculicola (Psm) and bacterial counts monitored 3 days post-
inoculation. Each bar represents the average Psm count in 15 leaf discs � SD.
Plant defense signaling 113
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

attenuated in the JA-insensitive jin1 and JA-biosynthesis
opr3 mutants. These observations raise the hypothesis that
JA may be the plastid glycerolipid-derived factor that is
required in Avr PeX for the establishment of SAR. However,
several lines of evidence counter this possibility. First, the JA
content of Avr PeX from sfd1 leaves was comparable to that
in Avr PeX from WT leaves (Figure 7a). Second, co-infiltra-
tion of JA or MeJA with Avr PeXs collected from the fad7 and
sfd1 mutant leaves did not alter the inability of these PeXs to
activate SAR (Figure 8a,b), indicating that the SAR defi-
ciency of the fad7 and sfd1 mutants is not a result of JA
deficiency. Furthermore, JA and MeJA when individually
applied to WT plants did not activate PR1 expression and the
SAR-associated systemic resistance to Psm (Figure 8a and
Figure S5). This is in striking contrast to the ability of Avr
PeX from WT leaves to increase PR1 transcript accumula-
tion (Figure 3a) and enhance systemic resistance to Psm
(Figures 3b, 4a–c and 5a–c). Finally, JA did not co-purify
with the SAR-inducing activity that was obtained after
fractionation of Avr PeX over a gel-filtration column
(Figure 7a,b). Together, these results argue against a role
for JA as the primary mobile signal in SAR. However, they
are not in disagreement with the involvement of JA-depen-
dent defenses in restricting pathogen growth in the distal
leaves subsequent to the perception of the SAR signal by
these organs. But, since the coi1 and jar1 mutants are SAR-
competent (Cui et al., 2005; Mishina and Zeier, 2007), the
requirement for JA in SAR-conferred resistance seems to be
independent of COI1 and JAR1 but dependent on JIN1.
When Avr PeX collected from WT leaves was infiltrated
into leaves of the sid2, eds5 and npr1 mutant plants, which
are defective in SA synthesis/signaling, SAR was not
observed (Figure 3b). Rather, on the contrary, bacterial
growth was slightly higher in these Avr Pex-infiltrated sid2,
eds5 and npr1 plants than in the corresponding Mock PeX-
infiltrated plants. The increased growth of Psm in the Avr
Pex-infiltrated sid2, eds5 and npr1 mutants as compared to
the corresponding Mock PeX-infiltrated plants could be a
result of systemic induced susceptibility (SIS), a phenome-
non that previously has been reported in Arabidopsis–
P. syringae interaction (Cui et al., 2005). The prior inocula-
tion of Arabidopsis with coronatine-producing P. syringae
results in increased susceptibility of the distal organs to
subsequent challenge by pathogen. Infiltration of purified
coronatine into the apoplast was sufficient to induce SIS in
the untreated leaves of the same plant (Cui et al., 2005).
Systemic induced susceptibility and SAR are counteracting
processes. The Avr PeX used in our experiments may
contain a SIS-inducing factor, presumably coronatine, in
addition to the SAR-inducing factor. The effects of SIS may
become more prominent when the plant is unable to activate
SAR, as is the case with the sid2, eds5 and npr1 mutants.
In conclusion, we have shown that plastid glycerolipid
metabolism is required for the accumulation of a
SAR-inducing activity in the Avr PeX of Arabidopsis leaves.
This glycerolipid-dependent factor is required along with the
DIR1-encoded nsLTP in Avr PeX for the establishment of
SAR in Arabidopsis. The identification of this plastid glyc-
erolipid-dependent factor will aid in understanding systemic
signaling in plants and in promoting resistance to a broad
spectrum of pathogens.
Experimental procedures
Plant and pathogen cultivation
Arabidopsis was grown at 22�C in growth chambers programmedfor 14 h light (100 lE m)2 sec)1) and 10 h dark cycle. Approximately4-week-old plants were used for all experiments. Pseudomonassyringae pv. maculicola ES4326 and P. syringae pv. tomato DC3000containing the avrRpt2 avirulence gene were propagated at 28�C onKing’s B medium containing appropriate antibiotics (Nandi et al.,2003, 2004). Cultivation of tomato, wheat and F. graminearumwere as previously described (Makandar et al., 2006; Tang et al.,1999).
Bacterial and fungal inoculations
Systemic acquired resistance was activated by infiltrating threelower fully expanded leaves with a suspension (107 colony-forming units (CFU) ml)1) in 10 mM MgCl2 of the avirulentpathogen P. syringae pv. tomato DC3000 avrRpt2. Plants infil-trated with 10 mM MgCl2 provided the mock controls. Three dayslater, four upper leaves were challenged with a suspension(105 CFU ml)1) of the virulent pathogen P. syringae pv. maculi-cola ES4326 (Psm). The Psm-challenged leaves were harvested3 days post inoculation (dpi) and rinsed in 10 mM MgCl2. A corkborer was used to punch out leaf discs (area = 0.28 cm2), whichwere ground in 10 mM MgCl2, and serial 10-fold dilutions platedon King’s medium containing streptomycin (100 lg ml)1). Plateswith bacteria were incubated at 28�C for 2 days before countingthe bacterial colonies. A total of 15 leaf disks (three replicationsof five leaf disks in each sample) were analyzed per treatment.Fusarium graminearum inoculation of wheat and Fusarium headblight disease evaluation were done as previously described(Makandar et al., 2006).
Arabidopsis mutants
The sfd1-1, sfd2-2 and ssi2 mutants are in the accession Nossen(No) (Nandi et al., 2003, 2004; Shah et al., 2001), the dir1 mutant(contains a T-DNA insertion) is in the accession Wassilewskija(Ws) background (Maldonado et al., 2002), the fad7-1 mutant is inthe accession Columbia (Col) glabra1, and the mgd1 mutant is inthe accession Col background (Jarvis et al., 2000). To generatethe ssi2 fad7-1 double mutant, the ssi2 mutant was crossed withthe fad7-1 mutant plant and the F2 progeny were screened fordouble-mutant plants. The ssi2 mutant allele was followed byPCR (see below) and plants homozygous for fad7-1 allelewere identified based on their lipid profile (reduction in 34:6- and36:6-MGD levels). The ssi2 dir1 plants were identified from a F2
population derived from a cross between the ssi2 and dir1mutant plants. Segregants homozygous for ssi2 and dir1 wereidentified by PCR (see below).
114 Ratnesh Chaturvedi et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

Lipid and phytohormone analysis
Lipid extraction and electrospray ionization tandem mass spec-trometry analysis of leaf lipids were performed as previouslydescribed (Devaiah et al., 2006; Welti et al., 2002). The JA and SAlevels were determined by gas chromatography-mass spectrometry(Schmelz et al., 2003).
RNA extraction and RT-PCR analysis
Leaf tissue was ground under liquid nitrogen, and RNA wasextracted using acid guanidinium thiocyanate-phenol-chloroformas previously described (Chomczynski and Sacchi, 1987). The iso-lated RNA was purified and used in the reverse-transcription poly-merase chain reactions (RT-PCR) (Pegadaraju et al., 2007). The PCRprimers for the ACT8 gene (At1g49240) are as previously described(Pegadaraju et al., 2005). The At-PR1-F 5¢-GCTCTTGTAGGTGCTC-TTGTTC-3¢ and At-PR1-R 5¢-CAGCTCTTATTTGTATTATTTG-3¢primers were used for PCR amplification of PR1 (At2g14610). ThePCR was performed with the following conditions: 95�C for 5 minfollowed by 25 cycles of 95�C for 30 sec, 60�C for 45 sec, 72�C for1 min, with final extension at 72�C for 10 min.
DNA extraction and PCR analysis
Arabidopsis genomic DNA from leaf tissue was isolated as previ-ously described (Konieczny and Ausubel, 1993). ssi2 was distin-guished from SSI2 using a derived-cleaved amplified polymorphicsequence (dCAPS). The primers, ssi2dCAPS-F 5¢-TTGTTTTGGTGGGGGACATGATCACAGAAGGTGCA-3¢ and ssi2dCAPS-R 5¢-TCGATCTGCCTCATGTCAACACG-3¢ were used in the PCR reaction. ThePCR was performed with the following conditions: 95�C for 5 minfollowed by 35 cycles of 95�C for 45 sec, 65�C for 45 sec, 72�C for45 sec, with final extension at 72�C for 5 min. The 200-bp PCRproduct derived from WT DNA contains one ApaL1 (New EnglandBiolabs, http://www.neb.com/) site, which on restriction with Apal1yields two products of 175 and 25 bp. In contrast, the PCR productderived from the ssi2 allele lacks the ApaL1 site. To identify dir1homozygous plants the T-DNA left border primer T-DNA-LBb15¢-GCGTGGACCGCTTGCTGCAACT-3¢ was used along with aprimer designed to the 3¢ untranslated region (UTR) of DIR1(5¢-GGGGTTAACCCATAAACGCCATTTGGCAG-3¢).
Collection of petiole exudates
The PeXs were collected essentially as previously described(Maldonado et al., 2002). A suspension (107 CFU ml)1)ofP. syringae pv. tomato containing the avrRpt2 gene wasinfiltrated into the abaxial surface of leaves. For the controls(Mock), 10 mM MgCl2 was similarly infiltrated into leaves. Plantswere placed in the growth chamber, covered with a plastic dome.Eight hours after infiltration, leaves were harvested for collectionof PeX. Leaves were cut at the base of their petiole using sharpscissors and the petiole immediately dipped in 50% ethanol fol-lowed by 0.0005% bleach and finally in 1 mM EDTA (pH 8.0)solution. The base of each petiole was immersed in 2.5 mlcollection solution (1 mM EDTA, pH 8.0) contained in a well of a24-well tissue culture plate (ICN Biomedical Inc.; http://www.mpbio.com). Five harvested leaves were placed in eachwell. Wet paper towels were placed under the tissue culture traysand the entire set-up was covered with a transparent plasticdome and incubated in a growth chamber exposed to a 14/10 h
light/dark period. For each treatment 50–100 leaves were used. Atthe end of the exudation period, PeXs from multiple wells werepooled. The absence of any contaminating P. syringae pv.tomato DC3000 avrRpt2 in the PeXs was tested by plating analiquot of the exudates on King’s medium (King et al., 1954). Thebiological activity of the PeX was determined by infiltratingexudates diluted two-fold in sterile water into leaves of healthyplants. The treated plants were placed in the growth chamber for3 days to allow for SAR to be activated. At the end of theincubation, the upper uninfiltrated leaves of the plant werechallenged with Psm and bacterial counts determined 3 dayslater. For experiments that involved mixing PeXs from twodifferent genotypes, equal volumes of each PeX were mixedand diluted two-fold in sterile water before infiltration intoArabidopsis leaves.
Gel-filtration fractionation of SAR-inducing activity
The PeX collected from 10 mM MgCl2- and Avr pathogen-inocu-lated plants were lyophilized at )70�C. The lyophilized powderwas reconstituted in 1 ml 2-amino-2-(hydroxymethyl)-1,3-propanediol (TRIS)-EDTA buffer (10 mM TRIS-HCl, 1 mM EDTA,pH 7.5) and centrifuged at 12 000g for 5 min to sediment par-ticulate matter. The supernatant was loaded on a 2 · 25 cm col-umn packed with Bio-Gel A-5m matrix (Bio-Rad, http://www.bio-rad.com/) that was pre-equilibrated with TRIS-EDTA buffer.The column was eluted with TRIS-EDTA buffer. One-milliliterfractions were collected at a flow rate of 1 ml min)1. Biologicalactivity of individual fractions was tested by infiltrating two-folddiluted fractions into leaves of 4-week-old WT plants. Threedays after infiltration, upper leaves of the infiltrated plantswere challenged with Psm and bacterial growth monitored asdescribed above.
Acknowledgements
We thank Fred Ausubel, Robin Cameron and Henrik Aronsson forproviding the sid2, dir1 and mgd1 mutant, respectively, and theABRC stock center at Ohio State University for providing the fad7-1mutants. We thank Eric Schmelz for training AAS in the measure-ment of phytohormones. This work was supported at varied timesby grants from the National Science Foundation (IOB0543862 andMCB0455318) and awards from the NIH Grant Number P20RR016475 from the INBRE Program of the National Center forResearch Resources. Additional support of the Kansas LipidomicsResearch Center was from NSF grants (EPS-0236913 andDBI0521587), Kansas Technology Enterprise Corporation andKansas State University. This is Kansas Agricultural ExperimentalStation contribution 07-151J.
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. Systemic acquired resistance competent petioleexudates from Arabidopsis promote resistance to Pseudomonassyringae in tomato.Figure S2. Co-application of avirulent petiole exudates collectedfrom the Arabidopsis sfd2 and fad7 mutants does not activate SARin wild-type plants.Figure S3. Avirulent petiole exudates from dir1 lack a SAR inducingactivity.
Plant defense signaling 115
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

Figure S4. PR1 expression in wild-type leaves infiltrated with petioleexudates from sfd1 and dir1 mutants.Figure S5. PR1 and PDF1.2 transcript accumulation in methyljasmonic acid-infiltrated leaves.Table S1. Comparison of digalactosyldiacylglycerol (DGD), monog-alactosyldiacylglycerol (MGD) and phosphatidylglycerol (PG) com-position (nmol mg)1 dry weight) in leaves of wild-type Arabidopsisaccession Columbia and the fad4 and fad7 mutants.Table S2. Comparison of digalactosyldiacylglycerol (DGD),monogalactosyldiacylglycerol (MGD) and phosphatidylglycerol(PG) composition (nmol mg)1 dry weight) in leaves of wild-typeArabidopsis accession Nossen and the sfd2-2 mutant.This material is available as part of the online article from http://www.blackwell-synergy.comPlease note: Blackwell Publishing are not responsible for the contentor functionality of any supplementary materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
References
Arai, K., Marechal, E., Block, M.A., Brun, D., Masuda, T., Shimada,
H., Takamiya, K.-I., Ohta, H. and Joyard, J. (2001) Two types ofMGDG synthase genes, found widely in both 16:3 and 18:3 plants,differentially mediated galactolipid syntheses in photosyntheticand nonphotosynthetic tissues in Arabidopsis thaliana. Proc. NatlAcad. Sci. USA, 98, 10960–10965.
Bakan, B., Hamberg, M., Perrocheau, L., Maume, D., Rogniaux, H.,
Tranquet, O., Rondeau, C., Blein, J.-P., Ponchet, M. and Marion,
D. (2006) Specific adduction of plant lipid transfer protein by anallene oxide generated by 9-lipoxygenase and allene oxidesynthase. J. Biol. Chem. 281, 38981–38988.
Blein, J.P., Coutos-Thevenot, P., Marion, D. and Ponchet, M. (2002)From elicitins to lipid transfer proteins: a new insight in cellsignalling involved in plant defence mechanisms. Trends PlantSci. 7, 293–296.
Chaturvedi, R. and Shah, J. (2006) Salicylic acid in plant diseaseresistance. In Salicylic Acid-A Plant Hormone (Hayat, S. andAhmad, A., eds). Dordrecht, The Netherlands: Springer, pp.335–370.
Chomczynski, P. and Sacchi, N. (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Anal. Biochem. 162, 156–159.
Cui, J., Bahrami, A.K., Pringle, E.G., Hernandez-Guzman, G.,
Bender, C.L., Pierce, N.E. and Ausubel, F.M. (2005) Pseudomonassyringae manipulates systemic plant defenses against pathogensand herbivores. Proc. Natl Acad. Sci. USA, 102, 1791–1796.
Darby, R.M., Maddison, A.L., Mur, L.A., Bi, Y.-M. and Draper, J.
(2000) Cell-specific expression of salicylate hydroxylase in anattempt to separate localized HR and systemic signallingestablishing SAR in tobacco. Mol. Plant Pathol. 1, 115–123.
Dean, R.A. and Kuc, J. (1986) Induced systemic protection incucumbers: the source of the ‘signal’. Physiol. Plant. Pathol. 28,227–233.
Devaiah, S.P., Roth, M.R., Baughman, E., Li, M., Tamura, P.,
Jeannotte, R., Welti, R. and Wang, X. (2006) Quantitative profilingof polar glycerolipid species from organs of wild-type Arabidop-sis and a PHOSPHOLIPASE Da1 knockout mutant. Phytochemis-try, 67, 1907–1924.
Devoto, A. and Turner, J.G. (2003) Regulation of jasmonate-medi-ated plant response in Arabidopsis. Ann. Bot. 92, 329–337.
Durrant, W.E. and Dong, X. (2004) Systemic acquired resistance.Annu. Rev. Phytopathol. 42, 185–209.
Guedes, M.E.M., Richmond, S. and Kuc, J. (1980) Induced systemicresistance to anthracnose in cucumber as influenced by thelocation of the inducer inoculation with Colletotrichum lagenari-um and the onset of flowering and fruiting. Physiol. Plant Pathol.17, 229–233.
Howe, G. and Schilmiller, A.L. (2002) Oxylipin metabolism inresponse to stress. Curr. Opin. Plant Biol. 5, 230–236.
Jarvis, P., Dormann, P., Peto, C.A., Lutes, J., Benning, C. and Chory,
J. (2000) Galactolipid deficiency and abnormal chloroplastdevelopment in the Arabidopsis MGD synthase 1 mutant. Proc.Natl Acad. Sci. USA, 97, 8175–8179.
Kachroo, P., Shanklin, J., Shah, J., Whittle, E.J. and Klessig, D.F.
(2001) A fatty acid desaturase modulates the activation of defensesignaling pathways in plants. Proc. Natl Acad. Sci. USA, 98, 9448–9453.
Kachroo, P., Kachroo, A., Lapchyk, L., Hildebrand, D. and Klessig,
D.F. (2003) Restoration of defective cross talk in ssi2 mutants: roleof salicylic acid, jasmonic acid, and fatty acids in SSI2-mediatedsignaling. Mol. Plant Microbe Interact. 16, 1022–1029.
Kachroo, P., Venugopla, S.C., Navarre, D.A., Lapchyk, L. and
Kachroo, A. (2005) Role of salicylic acid and fatty acid desatura-tion pathways in ssi2-mediated signaling. Plant Physiol. 139,1717–1735.
Kiefer, I.W. and Slusarenko, A.J. (2003) The pattern of systemicacquired resistance induction within the Arabidopsis rosette inrelation to the pattern of translocation. Plant Physiol. 132, 840–847.
King, E.O., Ward, M.K. and Raney, D.E. (1954) Two simple media forthe demonstration of phycocyanin and fluorescein. J. Lab. Clin.Med. 44, 301–307.
Konieczny, A. and Ausubel, F.M. (1993) A procedure for mappingArabidopsis mutations using co-dominant ecotype-specific PCRbased markers. Plant J. 4, 403–410.
Kuc, J. (1982) Induced immunity to plant disease. Bioscience, 32,854–860.
Li, L., Li, C., Lee, G.I. and Howe, G.A. (2002) Distinct roles for jas-monic acid synthesis and action in the systemic wound responseof tomato. Proc. Natl Acad. Sci. USA, 99, 6416–6421.
Makandar, R., Essig, J.S., Schapaugh, M.A., Trick, H.N. and Shah, J.
(2006) Genetically engineered resistance to Fusarium head blightin wheat by expression of Arabidopsis NPR1. Mol. Plant MicrobeInteract. 19, 123–129.
Maldonado, A.M., Doerner, P., Dixon, R.A., Lamb, C.J. and Camer-
on, R.K. (2002) A putative lipid transfer protein involved in sys-temic resistance signaling in Arabidopsis. Nature, 419, 399–403.
Metraux, J.-P., Nawrath, C. and Genoud, T. (2002) Systemicacquired resistance. Euphytica, 124, 237–243.
Mishina, T.E. and Zeier, J. (2007) Pathogen-associated molecularpattern recognition rather than development of tissue necrosiscontributes to bacterial induction of systemic acquired resistancein Arabidopsis. Plant J. 50, 500–513.
Molders, W.A., Buchala, A. and Metraux, J.-P. (1996) Transport ofsalicylic acid in tobacco necrosis virus-infected cucumber plants.Plant Physiol. 112, 787–792.
Nandi, A., Krothapalli, K., Buseman, C.M., Li, M., Welti, R.,
Enyedi, A. and Shah, J. (2003) Arabidopsis sfd mutantsaffect plastidic lipid composition and suppress dwarfing, celldeath, and the enhanced disease resistance phenotypes resultingfrom the deficiency of a fatty acid desaturase. Plant Cell, 15, 2383–2398.
Nandi, A., Welti, R. and Shah, J. (2004) The Arabidopsis thalianadihydroxyacetone phosphate reductase gene SUPPRESSOR OFFATTY ACID DESATURASE DEFICIENCY1 is required for glycer-
116 Ratnesh Chaturvedi et al.
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117

olipid metabolism and for the activation of systemic acquiredresistance. Plant Cell, 16, 465–477.
Nawrath, C. and Metraux, J.P. (1999) Salicylic acid induction-deficient mutants of Arabidopsis express PR-2 and PR-5 andaccumulate high levels of camalexin after pathogen inoculation.Plant Cell, 11, 1393–1404.
Osman, H., Vauthrin, S., Mikes, V., Milat, M.L., Panabieres, F.,
Marais, A., Brunie, S., Maume, B., Ponchet, M. and Blein, J.-P.
(2001) Mediation of elicitin activity on tobacco is assumed byelicitin-sterol complexes. Mol. Biol. Cell, 12, 2825–2834.
Park, S.-W., Kaimoyo, E., Kumar, D., Mosher, S. and Klessig, D.F.
(2007) Methyl salicylate is a critical mobile signal for plantsystemic acquired resistance. Science, 318, 113–116.
Pegadaraju, V., Knepper, C., Reese, J. and Shah, J. (2005) Prematureleaf senescence modulated by the Arabidopsis thaliana PAD4gene is associated with defense against the phloem-feedinggreen peach aphid. Plant Physiol. 139, 1927–1934.
Pegadaraju, V., Louis, J., Singh, V., Reese, J.C., Bautor, J., Feys,
B.J., Cook, G., Parker, J.E. and Shah, J. (2007) Phloem-basedresistance to green peach aphid is controlled by ArabidopsisPHYTOALEXIN DEFICIENT4 without its signaling partnerENHANCED DISEASE SUSCEPTIBILITY1. Plant J. 52, 332–341.
Pieterse, C.M.J., Van Wees, S.C.M., Ton, J., Van Pelt, J.A. and Van
Loon, L.C. (2002) Signalling in rhizobacteria-induced systemicresistance in Arabidopsis thaliana. Plant Biol. 4, 535–544.
Rasmussen, J.B., Hammerschmidt, R. and Zook, K.N. (1991) Sys-temic induction of salicylic acid accumulation in cucumber afterinoculation with Pseudomonas syringae pv syringae. PlantPhysiol. 97, 1342–1347.
Schmelz, E.A., Engelberth, J., Alborn, H.T., O’Donnell, P.,
Sammons, M., Toshima, H. and Tumlinson, J.H., III (2003)Simultaneous analysis of phytohormones, phytotoxins, andvolatile organic compounds in plants. Proc. Natl Acad. Sci. USA,100, 10552–10557.
Sekine, K.T., Nandi, A., Ishihara, T., Hase, S., Ikegami, M., Shah, J.
and Takahashi, H. (2004) Enhanced resistance to Cucumbermosaic virus in the Arabidopsis thaliana ssi2 mutant is mediatedvia an SA-independent mechanism. Mol. Plant Microbe Interact.17, 623–632.
Shah, J. (2005) Lipids, lipases and lipid modifying enzymes in plantdisease resistance. Annu Rev. Phytopathol. 43, 229–260.
Shah, J., Kachroo, P.K., Nandi, A. and Klessig, D.F. (2001) A reces-sive mutation in the Arabidopsis SSI2 gene confers SA- andNPR1-independent expression of PR genes and resistance againstbacterial and oomycete pathogens. Plant J. 25, 563–574.
Shulaev, V., Leon, J. and Raskin, I. (1995) Is salicylic acid a trans-located signal of systemic acquired resistance in tobacco? PlantPhysiol. 116, 387–392.
Tang, X., Xie, M., Kim, Y.J., Zhou, J., Klessig, D.F. and Martin, G.B.
(1999) Overexpression of P to activates defense responses andconfers broad resistance. Plant Cell, 11, 15–30.
Truman, W., Bennett, M.H., Kubigsteltig, I., Turnbull, C. and Grant,
M. (2007) Arabidopsis systemic immunity uses conserveddefense signaling pathways and is mediated by jasmonates. Proc.Natl Acad. Sci. USA, 104, 1075–1080.
Vernooij, B., Friedrich, L., Morse, A., Reist, R., Kolditz-Jawhar, R.,
Ward, E., Uknes, S., Kessmann, H. and Ryals, J. (1994) Salicylicacid is not the translocated signal responsible for inducing sys-temic acquired resistance but is required in signal transduction.Plant Cell, 6, 959–965.
Wallis, J.G. and Browse, J. (2002) Mutants of Arabidopsisreveal many roles for membrane lipids. Prog. Lipid Res. 41,254–278.
Welti, R., Li, W., Li, M., Sang, Y., Biesiada, H., Zhou, H.-E.,
Rajashekar, C.B., Williams, T.D. and Wang, X. (2002) Profilingmembrane lipids in plant stress response. Role of phospholipaseDa in freezing-induced lipid changes in Arabidopsis. J. Biol.Chem. 277, 31994–32002.
Plant defense signaling 117
ª 2008 The AuthorsJournal compilation ª 2008 Blackwell Publishing Ltd, The Plant Journal, (2008), 54, 106–117
Copyright © 2022 FDOKUMEN




















