
Role of ethylene and jasmonic acid on rhizome induction and growth in rhubarb ( Rheum rhabarbarum L
11
ORIGINAL PAPER Role of ethylene and jasmonic acid on rhizome induction and growth in rhubarb (Rheum rhabarbarum L.) Usha P. Rayirath • Rajasekaran R. Lada • Claude D. Caldwell • Samuel K. Asiedu • Kevin J. Sibley Received: 31 May 2010 / Accepted: 30 September 2010 / Published online: 20 October 2010 Ó Her Majesty the Queen in Right of Canada 2010 Abstract Experiments were conducted to elucidate the hormonal induction and regulation of rhizome growth in rhubarb (Rheum rhabarbarum L.). It was found that eth- ylene is the key regulator of rhizome induction and development. The role of jasmonic acid (JA) and its interaction with ethylene in rhizome induction and growth were also examined. Both ethylene and JA have a signifi- cant effect on promoting rhizome formation in vitro. Conversely, the ethylene biosynthesis inhibitor aminoeth- oxyvinylglycine (AVG) (1.5 lM) inhibited rhizome induction in multiple-shoot clumps in vitro, and suppressed the stimulatory effects of exogenously applied ethephon (1 mg l -1 ) and JA (10 ng l -1 ) in promoting mini-rhizome formation, further confirming the role of endogenous eth- ylene in the process. In addition, rhizome growth was significantly enhanced in the presence of both ethylene and JA (ethephon 1 mg l -1 and JA 10 ng l -1 ) compared to JA alone. These results suggest that endogenous ethylene is the key regulator of rhizome growth in rhubarb and JA promotes rhizome formation, possibly through inducing endogenous ethylene synthesis. Keywords Ethylene Jasmonic acid Hormonal induction Growth Mini-rhizome Rhubarb Abbreviations JA Jasmonic acid ACC 1-Aminocyclopropane-1-carboxylic acid AVG Aminoethoxyvinylglycine PGRs Plant growth regulators Introduction Rhubarb (Rheum rhabarbarum L.) is a herbaceous cool- season perennial vegetable having high market potential for expansion as a commercial vegetable industry in North America and Europe. The current method of propagation by rhizome division often results in a very low rate of propagule production; thus, enhancing rhizome growth is critical to generate more propagules. The physiological mechanisms and hormonal relationships that determine the growth and enlargement of rhizomes in rhubarb are not known (Rayirath et al. 2009). On the basis of the infor- mation currently available on the role of plant growth regulators (PGRs) in promoting vegetative storage organs, it is expected that ethylene, jasmonic acid (JA), ethylene gibberellic acid (GA 3 ) and abscisic acid (ABA) can all modulate the growth and development of these plant organs (Ja ´sik and de Klerk 2006; Bhatia et al. 1992; Kim et al. 2005; Zaib-Un-Nissa and Rafiq 1980). Determining the role(s) of these PGRs in manipulating rhizome growth in rhubarb will help to identify the key regulators of the rhizome formation process in rhubarb. Plant hormones are the key factors that elicit various physiological changes and modulate plant growth and development under varying environmental conditions (Ecker 1995; Etheridge et al. 2005). In general, exogenous U. P. Rayirath R. R. Lada (&) C. D. Caldwell S. K. Asiedu Department of Plant and Animal Sciences, Nova Scotia Agricultural College, 50 Pictou Road, P.O. Box 550, Truro, NS B2N 5E3, Canada e-mail: [email protected] K. J. Sibley Department of Environmental Sciences, Nova Scotia Agricultural College, Truro, NS B2N 5E3, Canada 123 Plant Cell Tiss Organ Cult (2011) 105:253–263 DOI 10.1007/s11240-010-9861-y
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Role of ethylene and jasmonic acid on rhizome induction and growth in rhubarb ( Rheum rhabarbarum L

ORIGINAL PAPER
Role of ethylene and jasmonic acid on rhizome inductionand growth in rhubarb (Rheum rhabarbarum L.)
Usha P. Rayirath • Rajasekaran R. Lada •
Claude D. Caldwell • Samuel K. Asiedu •
Kevin J. Sibley
Received: 31 May 2010 / Accepted: 30 September 2010 / Published online: 20 October 2010
� Her Majesty the Queen in Right of Canada 2010
Abstract Experiments were conducted to elucidate the
hormonal induction and regulation of rhizome growth in
rhubarb (Rheum rhabarbarum L.). It was found that eth-
ylene is the key regulator of rhizome induction and
development. The role of jasmonic acid (JA) and its
interaction with ethylene in rhizome induction and growth
were also examined. Both ethylene and JA have a signifi-
cant effect on promoting rhizome formation in vitro.
Conversely, the ethylene biosynthesis inhibitor aminoeth-
oxyvinylglycine (AVG) (1.5 lM) inhibited rhizome
induction in multiple-shoot clumps in vitro, and suppressed
the stimulatory effects of exogenously applied ethephon
(1 mg l-1) and JA (10 ng l-1) in promoting mini-rhizome
formation, further confirming the role of endogenous eth-
ylene in the process. In addition, rhizome growth was
significantly enhanced in the presence of both ethylene and
JA (ethephon 1 mg l-1 and JA 10 ng l-1) compared to JA
alone. These results suggest that endogenous ethylene is
the key regulator of rhizome growth in rhubarb and JA
promotes rhizome formation, possibly through inducing
endogenous ethylene synthesis.
Keywords Ethylene � Jasmonic acid � Hormonal
induction � Growth � Mini-rhizome � Rhubarb
Abbreviations
JA Jasmonic acid
ACC 1-Aminocyclopropane-1-carboxylic acid
AVG Aminoethoxyvinylglycine
PGRs Plant growth regulators
Introduction
Rhubarb (Rheum rhabarbarum L.) is a herbaceous cool-
season perennial vegetable having high market potential
for expansion as a commercial vegetable industry in North
America and Europe. The current method of propagation
by rhizome division often results in a very low rate of
propagule production; thus, enhancing rhizome growth is
critical to generate more propagules. The physiological
mechanisms and hormonal relationships that determine the
growth and enlargement of rhizomes in rhubarb are not
known (Rayirath et al. 2009). On the basis of the infor-
mation currently available on the role of plant growth
regulators (PGRs) in promoting vegetative storage organs,
it is expected that ethylene, jasmonic acid (JA), ethylene
gibberellic acid (GA3) and abscisic acid (ABA) can all
modulate the growth and development of these plant
organs (Jasik and de Klerk 2006; Bhatia et al. 1992; Kim
et al. 2005; Zaib-Un-Nissa and Rafiq 1980). Determining
the role(s) of these PGRs in manipulating rhizome growth
in rhubarb will help to identify the key regulators of the
rhizome formation process in rhubarb.
Plant hormones are the key factors that elicit various
physiological changes and modulate plant growth and
development under varying environmental conditions
(Ecker 1995; Etheridge et al. 2005). In general, exogenous
U. P. Rayirath � R. R. Lada (&) � C. D. Caldwell � S. K. Asiedu
Department of Plant and Animal Sciences, Nova Scotia
Agricultural College, 50 Pictou Road, P.O. Box 550,
Truro, NS B2N 5E3, Canada
e-mail: [email protected]
K. J. Sibley
Department of Environmental Sciences, Nova Scotia
Agricultural College, Truro, NS B2N 5E3, Canada
123
Plant Cell Tiss Organ Cult (2011) 105:253–263
DOI 10.1007/s11240-010-9861-y

application of PGRs can positively regulate the formation,
growth and development of various plant organs by acting
as chemical signals that alter source–sink relationships,
long-distance photoassimilate translocation, leaf senes-
cence and other developmental processes (Wardlaw 1990;
Morris 1996). The role of PGRs like JA, GA3, ABA, auxin
and cytokinin in inducing the formation and development
of storage structures like tubers and corms is well dem-
onstrated in several crop species (Bhatia et al. 1992; Jasik
and Mantell 2000; Jasik and de Klerk 2006; Caldiz 1996;
Kim et al. 2005; El-Antably et al. 1967; Zaib-Un-Nissa and
Rafiq 1980; Romanov et al. 2000).
In addition to its role as a key wounding hormonal signal
(O’Donnell et al. 1996; Leon et al. 2001), ethylene also
participates in regulating a variety of developmental pro-
cesses in plants, including seed germination, lateral bud
stimulation, adventitious rooting, overcoming dormancy
and organ senescence and abscission (Abeles et al. 1992).
Exogenous application of ethylene induces tuberisation in
potatoes and enhances root bulking in carrots (Vreugenhil
and van Dijk 1989; Bhatia et al. 1992; Neuteboom et al.
2002). These results suggest that ethylene could be a pos-
sible rhizome-inducing hormonal signal in rhubarb. How-
ever, the specific role of ethylene in plant morphogenetic
processes such as the induction of vegetative storage
structures has not been reported so far and it has been
suggested that its stimulatory effect on shoot and root
growth depends on the plant species (Dimasi-Theriou et al.
1993).
The plant defence-related signals, JA and its methyl
esters, regulate many physiological and developmental
processes, such as root growth, tuberisation, fruit ripening,
senescence and pollen development (Schaller et al. 2005).
During wound stress, jasmonates activate mechanisms
involved in the healing processes through enhanced cell
division and proliferation (Leon et al. 2001). Jasmonates
are also potent inducers of vegetative storage protein (VSP)
gene expression (Lorenzo and Solano 2005) and play a
significant role in the formation of vegetative storage
organs (Jasik and de Klerk 2006). While the exogenous
application of JA has been reported to induce vegetative
structures like tubers (Koda and Kikuta 1991; Debeljak
et al. 2002; Zhang et al. 2006) and bulbs (Ravnikar et al.
1993; Santos and Salema 2000), it is not known whether
jasmonates induce rhizome growth or whether there is any
interaction between ethylene and JA in this process. Based
on all of this information, it was hypothesised that ethylene
and JA induce and regulate rhizome development in
rhubarb.
To explore the hormonal physiology involved in the
induction and growth of rhizomes in rhubarb, we investi-
gated the role of ethylene and JA in inducing miniature
rhizomes in rhubarb shoots in vitro.
Materials and methods
Plant culture and treatment application
Clonally propagated 7-year-old plants of Rheum rhabarba-
rum L. cv., Sutton were used for obtaining explants to pro-
duce multiple shoots in vitro. Multiple-shoot clumps were
produced in vitro from meristem tips from lateral buds borne
on the rhizome. The buds were surface-sterilised by
immersing in 2% (NaOCl) sodium hypochlorite for 10 min,
followed by four rinses in sterile distilled water. Surface-
sterilised lateral buds were dissected under a microscope to
separate the meristem tips from the outer tissues. These
meristem tips were cultured in sterile Murashige and Skoog
(MS) (1962) basal medium supplemented with 1 mg l-1
each of 6-benzylaminopurine (BAP) and indole-3-butyric
acid (IBA), 20 g l-1 of sucrose and solidified with 8 g l-1
agar (pH 5.5) (Roggemans and Claes 1979; Walkey and
Mathews 1979). A shoot clump refers to the bunch of off-
shoots that arise from a single bud on the rhizome. Seven-
week-old in-vitro-cultured shoot clumps, each consisting of
3–4 miniature shoots, were transferred to MS (1962) basal
medium modified with varying concentrations of PGRs and
30 g l-1 sucrose solidified with 7 g l-1 agar after adjusting
the pH to 5.5. The experiment was conducted in three stages.
In the first stage, selected PGRs, GA3 (10, 20, 40 and
80 mg l-1), ABA (0.1, 1, 10 and 100 lg l-1), JA (10 ng l-1,
1 lg l-1, 10 lg l-1 and 1 mg l-1) and the ethylene-releas-
ing compound, ethephon (2-chloroethylphosphonic acid) (1,
10 and 50 mg l-1), were screened in vitro to investigate
their role in inducing mini-rhizomes in multiple shoots. The
term mini-rhizome as used in this paper refers to a miniature
rhizome produced in vitro from cultured shoot explants. In
order to further confirm the results of the first stage, in the
second stage, the ethylene-releasing compound ethephon (2-
chloroethylphosphonic acid) (1, 10 and 50 mg l-1), the
ethylene precursor ACC (1-aminocyclopropane-1-car-
boxylic acid) (0.5, 1.0, 1.5 mM) and the ethylene biosyn-
thesis inhibitor AVG (aminoethoxyvinylglycine) (0.5, 1.0,
1.5 lM) were used to determine their roles as signals in
inducing mini-rhizomes. While JA and ethephon induced
mini-rhizomes (see the results of the first and second stages),
it was not clear whether JA-induced mini-rhizome formation
is through promoting endogenous ethylene. Also, it was
important to confirm if mini-rhizomes can be induced when
endogenous ethylene synthesis is prevented. To investigate
the interaction between JA and ethylene and the role of
ethylene biosynthesis inhibitor in the process, multiple-
shoot clumps were cultured in MS basal medium modified
with the most effective concentrations of JA, ethephon
(Fig. 1a) and their combination in the presence or absence of
AVG (1.5 lM) with 30 g l-1 sucrose solidified with 7 g l-1
agar after adjusting the pH to 5.5.
254 Plant Cell Tiss Organ Cult (2011) 105:253–263
123

The concentrations of the PGRs used in the experiments
were determined based on the previous similar in vitro
studies conducted in similar crop species producing rhi-
zomes and mini-tubers and in rhubarb (Bjornseth 1946) and
also based on initial trial experiments conducted in wide
concentration ranges to determine the optimum concentra-
tion range without damaging the explant tissue, depending
on the active ingredient present in the formulations of these
PGRs (data not provided). The details on the PGR prepa-
rations are given in the Appendix. The cultures were grown
in GA-7 Magenta vessels of size H 77 9 77 9 97 mm
(Sigma-Aldrich, Ontario, Canada) with 50 ml of nutrient
media and maintained at 20 ± 1�C day/night temperature
under a 16-h photoperiod. The light intensity was main-
tained between 100 and 120 lmol m-2 s-1 with cool white
fluorescent lights.
The induction of mini-rhizomes at the base of the clumps
and their growth were monitored in the cultures. After
6 weeks of culture initiation, the plantlets were taken out.
Observations were made on the length of the shoots, shoot
growth, root growth, presence of mini-rhizomes and the
width of the micro-rhizomes produced. For measuring the
shoot growth and root growth, a standardised grade scale
was used (Rayirath 2008). Data on shoot growth and root
growth are not shown. One of the ten replicates in those
treatments which induced mini-rhizomes was sacrificed in
order to take microscopic sections of the mini-rhizomes.
Microscopy
In order to confirm that the growth at the base of in-vitro-
cultured shoot clumps is not a callus tissue but a mini-
Fig. 1 a Mini-rhizome width
(cm) in multiple-shoot clumps.
The multiple-shoot clumps were
incubated in Murashige and
Skoog (MS) basal medium with
various concentrations of plant
growth regulators (PGRs) for
6 weeks. The values represent
the means of ten replicates and
the error bars represent the
standard error (SE) at a = 0.05.
b Left to right: rhizome
induction (initiation) in
ethephon 1 mg l-1 after 1 week
in culture (picture taken from
the bottom of the culture jar);
mini-rhizomes in the explants
cultured with ethephon 1 mg l-1
for 6 weeks; absence of mini-
rhizome growth in control (with
distilled water) explants
Plant Cell Tiss Organ Cult (2011) 105:253–263 255
123

rhizome, microscopic sections of the in-vitro-produced
mini-rhizome and the mother rhizome were compared and
studied. Fresh transverse sections of the in-vitro-produced
mini-rhizomes were made using sterile scalpel blades and
viewed using an Olympus BH-2 compound transmitted
light microscope (Carson Medical and Scientific Co. Ltd.,
Markham, Ontario, Canada) under standard bright-field
conditions and photographed using an Olympus OM-2S
35-mm camera. The sections were not stained since
staining could interfere with the starch which might be
present in the tissues and lead to misinterpretation. Sections
of the in vitro mini-rhizomes were compared with the
transverse sections of the original rhizome obtained from a
live plant grown in the greenhouse. The anatomical struc-
tures of the two different sections were characterised and
compared.
Pot culture and mini-rhizome growth measurements
of in-vitro-cultured rhubarb plantlets
After 6 weeks growth in the culture media with various
PGRs, the shoot clumps which produced mini-rhizomes at
their base and the respective controls were planted in 15 cm
pots filled with the commercial peat-based potting mixture
ProMix BX (Premier Horticulture, Quebec, Canada). The
transplants were kept in a mist chamber for a week to
harden off and were maintained for evaluating mini-rhi-
zome growth further under in vivo conditions in a green-
house at the Nova Scotia Agricultural College (NSAC),
Truro, Nova Scotia, Canada (45� 220 N; 63� 160 W) at 20�/
15�C (day/night) under a 16-h photoperiod. The natural
daylight was supplemented with light from high-pressure
sodium bulbs (400 W). Since one of the replicates from the
in vitro plant outs was used for microscopy study, the
remaining nine replicates were transplanted into the pots.
The plants were watered every third day until the soil was
saturated. After 4 months in pot culture, the plantlets were
gently uprooted and observations were made on the signal
induced mini-rhizome growth and enlargement. The mini-
rhizomes were weighed and the rhizome width and length
were measured. The number of buds produced on the mini-
rhizomes and the number of rhizome branches per plantlet
were also counted. Observations on the root growth, shoot
growth, shoot length and number of leaves were also taken.
Data on root and shoot growth are not shown.
Foliar application of ethephon on ex vitro plants
A pot culture experiment was conducted to confirm the
effect of ethephon at 1 mg l-1 [the most effective PGR in
the in vitro studies (Fig. 1)], as a foliar spray on newly
planted rhubarb plant cultures under greenhouse condi-
tions. Uniform size rhizome parts weighing 60 g and
bearing at least two viable buds were planted in large-sized
pots (30 cm diameter). There were ten replicates and a
control without the PGR treatment was also maintained.
The plant cell-tested commercial formulation of ethephon
(Sigma-Aldrich, Ontario, Canada) along with the surfactant
Tween 20 (*0.1%) was applied as a foliar spray using a
hand-pump sprayer, Super Spray GX60 (Green Cross,
Division of CIBA-GEIGY, Canada Ltd.) of capacity
1.25 L. One hundred millilitres of the spray solution was
applied per plant at 8 weeks and 12 weeks after sprout
emergence. The control plants were sprayed with distilled
water containing the same amount of surfactant. The plants
were placed in tall cardboard boxes (cut open at the top) to
prevent drifting of the spray solution to the neighbouring
plants and were sprayed until the leaves were dripping.
After 5 months, the plants were harvested and measure-
ments were made on the rhizome growth.
Experimental design and data analysis
The in vitro and pot culture experiments utilised a com-
pletely randomised design (CRD). All of the experiments
were replicated ten times and were repeated twice. The
general linear models (GLM) procedure (SAS Institute Inc.
1999) was used for the analysis of variance (ANOVA),
with Tukey’s multiple means comparison test being used
for determining differences between the means.
Results
Effect of ethephon and JA on inducing mini-rhizomes
in vitro
Among the PGRs tested, the ethylene-releasing compound,
ethephon, and JA induced mini-rhizomes in in-vitro-cul-
tured rhubarb multiple-shoot clumps. As shown in Fig. 1a,
exogenously applied ethephon and JA induced mini-rhi-
zomes at lower concentrations tested. Within 1 week after
culturing (*5 days), the cultures incubated in the media
with ethephon (ranging from 1 to 50 mg l-1) and JA
(ranging from 10 ng l-1 to 1 mg l-1) exhibited rhizome
induction with a noticeable bulging at the base of the shoot
clumps (Fig. 1b). The width of the induced mini-rhizomes
increased as the length of time in the culture progressed.
Exogenous application of ethephon had a stronger effect in
inducing rhizomes and promoting their growth; rhizome
growth was observed at all concentrations of ethephon
tested. After 6 weeks in culture, the highest rhizome width
of 1.56 cm (Fig. 1b) was achieved in cultures with
1 mg l-1 ethephon. The cultures with 10 ng l-1 and
1 lg l-1 JA were the second best after ethephon 1 mg l-1
in mini-rhizome induction. However, higher concentrations
256 Plant Cell Tiss Organ Cult (2011) 105:253–263
123
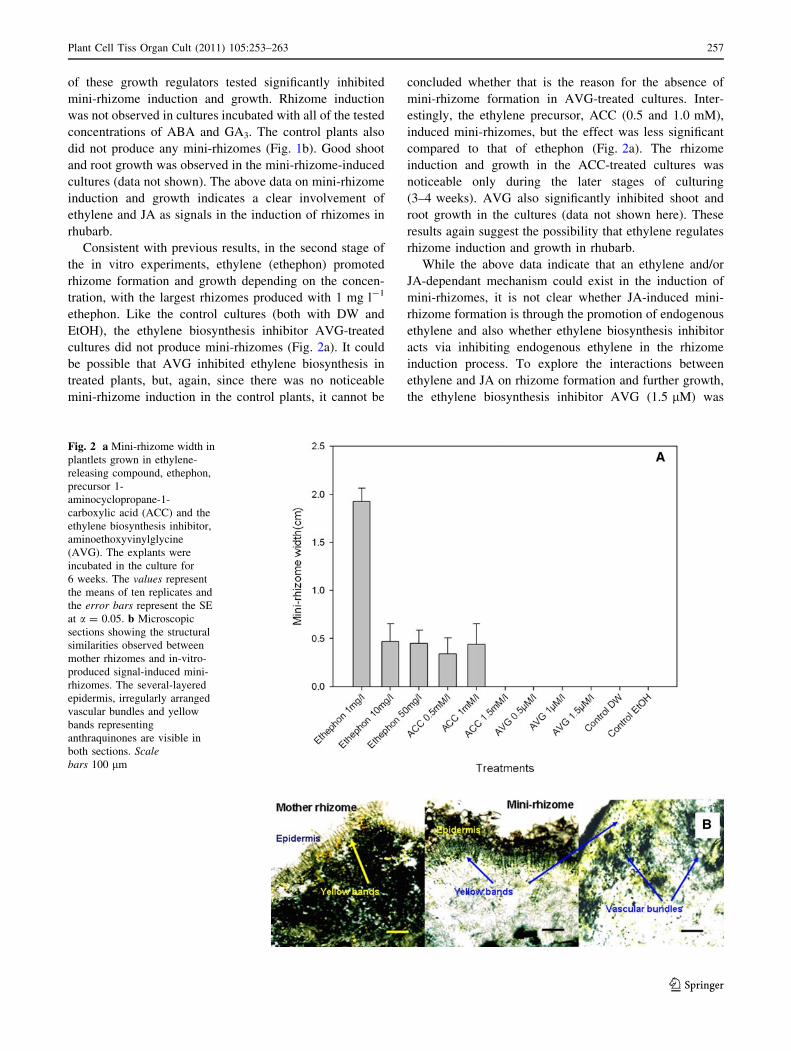
of these growth regulators tested significantly inhibited
mini-rhizome induction and growth. Rhizome induction
was not observed in cultures incubated with all of the tested
concentrations of ABA and GA3. The control plants also
did not produce any mini-rhizomes (Fig. 1b). Good shoot
and root growth was observed in the mini-rhizome-induced
cultures (data not shown). The above data on mini-rhizome
induction and growth indicates a clear involvement of
ethylene and JA as signals in the induction of rhizomes in
rhubarb.
Consistent with previous results, in the second stage of
the in vitro experiments, ethylene (ethephon) promoted
rhizome formation and growth depending on the concen-
tration, with the largest rhizomes produced with 1 mg l-1
ethephon. Like the control cultures (both with DW and
EtOH), the ethylene biosynthesis inhibitor AVG-treated
cultures did not produce mini-rhizomes (Fig. 2a). It could
be possible that AVG inhibited ethylene biosynthesis in
treated plants, but, again, since there was no noticeable
mini-rhizome induction in the control plants, it cannot be
concluded whether that is the reason for the absence of
mini-rhizome formation in AVG-treated cultures. Inter-
estingly, the ethylene precursor, ACC (0.5 and 1.0 mM),
induced mini-rhizomes, but the effect was less significant
compared to that of ethephon (Fig. 2a). The rhizome
induction and growth in the ACC-treated cultures was
noticeable only during the later stages of culturing
(3–4 weeks). AVG also significantly inhibited shoot and
root growth in the cultures (data not shown here). These
results again suggest the possibility that ethylene regulates
rhizome induction and growth in rhubarb.
While the above data indicate that an ethylene and/or
JA-dependant mechanism could exist in the induction of
mini-rhizomes, it is not clear whether JA-induced mini-
rhizome formation is through the promotion of endogenous
ethylene and also whether ethylene biosynthesis inhibitor
acts via inhibiting endogenous ethylene in the rhizome
induction process. To explore the interactions between
ethylene and JA on rhizome formation and further growth,
the ethylene biosynthesis inhibitor AVG (1.5 lM) was
Fig. 2 a Mini-rhizome width in
plantlets grown in ethylene-
releasing compound, ethephon,
precursor 1-
aminocyclopropane-1-
carboxylic acid (ACC) and the
ethylene biosynthesis inhibitor,
aminoethoxyvinylglycine
(AVG). The explants were
incubated in the culture for
6 weeks. The values represent
the means of ten replicates and
the error bars represent the SE
at a = 0.05. b Microscopic
sections showing the structural
similarities observed between
mother rhizomes and in-vitro-
produced signal-induced mini-
rhizomes. The several-layered
epidermis, irregularly arranged
vascular bundles and yellow
bands representing
anthraquinones are visible in
both sections. Scalebars 100 lm
Plant Cell Tiss Organ Cult (2011) 105:253–263 257
123

used in the culture media, along with the most effective
concentrations of ethephon (1 mg l-1) and JA (10 ng l-1)
and their combinations. After 6 weeks of culturing
explants, consistent with previous results, ethephon
(1 mg l-1) and JA (10 ng l-1) induced mini-rhizomes and
significantly promoted their growth (Fig. 3a). The combi-
nation treatment (JA 10 ng l-1 and ethephon 1 mg l-1)
exhibited better mini-rhizome growth than that of JA alone,
but was significantly less than that of ethephon alone
(Fig. 3a, b). This demonstrated that, in combination with
ethephon, the rhizome growth triggering effect of JA was
further enhanced compared to when applied alone.
As shown in Fig. 3a, c, AVG produced an 88% reduc-
tion in mini-rhizome width in cultures with JA (JA and
AVG), an 90% reduction in cultures with ethylene (ethe-
phon and AVG) and an 89% reduction in cultures with both
JA and ethylene (JA and ethephon and AVG), respectively,
when compared with those without AVG. Considerable
reduction in the mini-rhizome width observed in cultures
treated with AVG 1.5 lM despite the presence of exoge-
nous JA and ethylene further suggests that it is an ethylene-
dependent process. Consistent with the previous results, the
control cultures did not produce mini-rhizomes.
Microscopic studies and structural comparison of in-
vitro-produced mini-rhizomes with the mother rhizome
Structural similarities were observed between the rhizomes
(Fig. 2b). The presence of large amounts of starch made it
difficult to obtain good sections of the mother rhizome. As
shown in Fig. 2b, the epidermal layer was thick, with
several layers of densely packed cells. The vascular bun-
dles in both cases (mini-rhizome and mother rhizome) were
irregularly arranged. They lay scattered and, in between,
there were golden yellow bands/rays, which could possibly
be anthraquinones (Fig. 2b) that are abundant in the rhu-
barb rhizome tissues (Gao et al. 2009). Interestingly, cer-
tain green-coloured structures, which are more likely
chloroplasts, were also observed in some sections. These
structural similarities indicate that the growth at the shoot
bases induced by ethylene and JA are miniature rhizomes.
Growth of induced mini-rhizomes in in vitro plant outs
After 5 months of growth of the plant outs in the pot cul-
ture, they were harvested and measurements were made on
the mini-rhizome growth in vivo. Consistent with the in
vitro results, the mini-rhizomes from the cultures with
ethylene, JA and their combination exhibited the same pace
of growth when compared to the controls (DW and EtOH)
(Fig. 4). As per Table 1, exogenous ethephon (1 mg l-1)
application strongly promoted mini-rhizome growth and
the cultures with the highest width, length, branches and
lateral buds. Rhizome growth was significantly higher in
the cultures with JA (10 ng l-1) compared to the control.
Even though JA–ethephon combination treatment exhibited
considerably higher rhizome growth than JA alone, it was
not superior to those with ethephon alone. Lateral buds
were also significantly promoted by exogenous ethylene
(ethephon) and JA. The PGR-treated cultures exhibited
good shoot and root growth compared to the control in vivo
(data not shown).
Effect of ethephon spray on rhizome growth and lateral
bud production in rhubarb in a pot culture
The foliar application of ethephon (1 mg l-1, the most
effective concentration in vitro) on rhubarb plants at 8 and
12 weeks after shoot emergence significantly enhanced the
number of lateral buds on the rhizome after 4 months in pot
culture. As shown in Table 2, the number of lateral buds
increased two-fold in the treated plants compared with the
control, and there was a significant increase in the rhizome
width, even though the fresh weight did not differ signifi-
cantly between the treated and untreated plants.
Discussion
The absence of mini-rhizomes (Figs. 2a and 3c) in AVG-
treated plant cultures and AVG’s antagonistic effect on
mini-rhizome induction and growth in cultures treated with
exogenous ethephon and JA strongly indicate that ethylene
is a key regulator and/or inducer and growth enhancer of
rhizomes in Rheum rhabarbarum plantlets cultured in
vitro. Interestingly, the ethylene precursor, ACC, was also
effective in inducing mini-rhizomes but to a lesser extent
than ethephon. It appears as if, in the treated cultures, AVG
inhibited the formation of ethylene precursor ACC) from
S-adenosylmethionine (SAM) by inhibiting the rate-limit-
ing enzyme ACC synthase (Yang and Hoffman 1984),
limiting endogenous ethylene and inhibiting the rhizome
induction process. Even though JA triggers promoter
activities of ACC synthase genes (Tang et al. 2008),
enhancing ethylene biosynthesis, here, in the cultures with
JA and AVG, AVG might have counteracted JA’s effect by
antagonistically inhibiting endogenous ethylene.
Several previous studies have demonstrated that
exogenous ethylene could induce endogenous ethylene
through the upregulation of ethylene biosynthesis genes
(Alonso et al. 2003; Van Zhong and Burns 2003).
Investigating the role of endogenous ethylene in cotton
fibre cell elongation, Shi et al. (2006) have shown that
exogenously applied ethylene induce the expression of
ACO genes (1-Aminocyclopropane-1-Carboxylic Acid
Oxidase) involved in the oxidation of ACC to ethylene
258 Plant Cell Tiss Organ Cult (2011) 105:253–263
123

(Yang and Hoffman 1984), thereby, inducing endogenous
ethylene synthesis, while the cotton ovules treated with
ethylene biosynthesis inhibitor AVG suppressed fibre cell
elongation through inhibiting endogenous ethylene. The
regulation of the expression of ACO genes, thus, has
functional significance (Prescott and John 1996) and it
Fig. 3 a Mini-rhizome width in
plantlets grown in JA, ethephon
and their most effective
combinations in the presence
and absence of AVG. The
explants were incubated in the
culture for 6 weeks. The valuesrepresent the means of ten
replicates and the error barsrepresent the SE at a = 0.05.
b Plants after 6 weeks growth in
1 mg l-1 ethephon and
10 ng l-1 JA combination and
1 mg l-1 ethephon (top), and
the mini-rhizome growth in
1 mg l-1 ethephon and 10 ng
l-1 JA combination and 1 mg
l-1 ethephon (bottom). The
explants were cultured in MS
basal medium with PGRs for
6 weeks (pictures taken from
the bottom of the culture jars).
c Left to right: mini-rhizome
growth inhibited in explants
incubated with 10 ng l-1 JA and
1.5 lM AVG combination;
1 mg l-1 ethephon and 1.5 lM
AVG combination; 1 mg l-1
ethephon, 10 ng l-1 JA and
1.5 lM AVG combination
treatment
Plant Cell Tiss Organ Cult (2011) 105:253–263 259
123

has also been proved that at least two ACO genes in
Arabidopsis thaliana are ethylene-inducible (Alonso et al.
2003; Van Zhong and Burns 2003). Hence, in this study,
exogenous ethylene (ethephon) appears to induce mini-
rhizomes in treated cultures through enhanced ethylene
biosynthesis, which is inhibited by AVG, resulting in the
suppression of mini-rhizome growth in the combination
cultures. However, the present investigation, being purely
physiological, cannot address the molecular mechanisms
which result in these differential responses to the biosyn-
thetic inhibitors. The results presented in Fig. 1a suggest
that GA3 and ABA did not have any significant role in the
process as supported by previous studies (Obasi and Atanu
2004; Zheng et al. 2005). Even though ABA induces tuber
formation and growth in potato by counteracting the effect
of GA3 (Xu et al. 1998), scientific evidence on its role in
inducing rhizome growth in vegetatively propagated
perennial crops is not available thus far.
The comparison of microscopic sections of in-vitro-
generated mini-rhizomes and mother rhizomes of a normal
plant grown in the greenhouse clearly revealed some
structural similarities (Fig. 2b). These structural similari-
ties strongly support that the growth obtained at the base of
the explants treated with effective concentrations of ethe-
phon and JA is a miniature rhizome. A rhizome is a stem
modification that is a common storage organ in vegeta-
tively propagated crops (Hartmann et al. 2002). In some
herbaceous perennials which over-winter (like rhubarb),
rhizomes serve as carbon reserves for survival (Moore et al.
1998). Anatomically, rhizomes mostly lack a proper stele
pattern and form shoot and root buds from the nodes
(Raven et al. 2005; Hartmann et al. 2002). The presence of
structures, which are more likely chloroplasts (seen as
green-coloured structures), further confirms that it is not a
Fig. 4 Mini-rhizome weight (g) of in vitro plantlets treated with
1 mg l-1 ethephon, 10 ng l-1 JA and their combination (1 mg l-1
ethephon, 10 ng l-1 JA) after 4 months in pot culture. The valuesrepresent the means of nine replicates and the error bars represent the
SE at a = 0.05
Table 1 Growth of in-vitro-induced mini-rhizomes after transplantation to pots
Treatments Rhizome
width (cm)
Rhizome
length (cm)
Rhizome
weight (g)
No. of
rhizome branches
No. of buds
Ethephon 1 mg l-1 6.6a 6.9a 53.5a 5.1a 19.1a
JA 10 ng l-1 4.8b 5.0b 38.2b 3.4b 10.2c
JA 1 lg l-1 5.5b 4.9b 36.2b 3.8b 7.4d
Ethephon 1 mg l-1 and JA 10 ng l-1 5.4b 5.4b 49.4a 3.1b 11.7b
Control DW 2.7c 4.8bc 11.1c 1.4c 3.2e
Control EtOH 1.6c 2.8c 6.9c 0.8c 2.4e
The in-vitro-produced rhubarb plantlets with the induced mini-rhizomes were grown under controlled conditions in the greenhouse for 4 months
and then uprooted and observations were made
EtOH ethanol, DW distilled watera–e Means with the same letters indicate no significant difference based on Tukey’s test at a = 0.05; n = 9
Table 2 Foliar treatment with ethephon on plants in pot culture
Treatments Rhizome width (cm) Rhizome weight (g) No. of buds
Ethephon 1 mg l-1 8.6a 117.6a 22.2a
Control 7.5b 110.9a 11.6b
Measurements were made on the rhizome growth of pot-grown plants 5 months after foliar treatment with ethephona, b Means with the same letters indicate no significant difference based on Tukey’s test at a = 0.05; n = 10
260 Plant Cell Tiss Organ Cult (2011) 105:253–263
123

root structure and the presence of vascular structures also
supports that it is not a mere callus growth but a well-
differentiated plant organ.
To further support the results of the in vitro studies,
exogenously applied ethephon in the form of foliar spray
on in situ rhubarb plants during active vegetative growth (8
and 12 weeks after shoot emergence) significantly
enhanced the number of potential buds on the rhizome.
Being a gaseous hormone, ethylene is not translocated
through the vascular system. Ethylene is neither actively
transported nor degraded (Bleecker and Kende 2000). The
induction of ethylene synthesis by signals such as auxin or
wounding usually occurs through the activation of ACC
synthase through increased gene expression (Bleecker and
Kende 2000). However, the exact molecular mechanism of
ethephon- and JA-induced rhizome growth is yet to be
uncovered. Here, exogenous ethylene appeared to trigger
adventitious bud development in the sprayed plants, pos-
sibly through breaking lateral bud dormancy. Previous
reports indicate that ethylene (and ethylene-releasing
compounds) could positively regulate developmental pro-
cesses like adventitious root formation (Liu et al. 1990),
trigger adventitious bud development releasing lateral bud
dormancy (Reid 1987), enhance cambial activity and
induce radial swelling of storage organs (Reid 1987; Ne-
uteboom et al. 2002).
Our study clearly indicates the specific role of ethylene
in the induction, growth and development of rhizomes in
rhubarb. Although ethylene has a pleiotropic role in various
processes of plant growth and development, specific roles
of this hormonal signal in plant morphogenesis, such as the
formation and development of vegetative storage organs or
vegetative reproductive structures, has not been reported so
far.
In the present study, JA at lower concentrations
(10 ng l-1 and 1 lg l-1) significantly induced mini-rhi-
zomes in rhubarb plantlets (Fig. 1a). However, the rhizome-
inducing effect of JA was suppressed in the presence of
ethylene synthesis inhibitor, AVG, and, at the same time,
further enhanced by ethylene (ethephon) when they were
combined and applied exogenously in the growing media.
This implies that, while JA could promote rhizome induc-
tion, it requires the presence of endogenous ethylene as
AVG inhibited JA-induced rhizome growth. Also, it is
possible that, since JA induces ethylene synthesis (Tang
et al. 2008), the JA-induced mini-rhizome synthesis may be,
perhaps, due to JA-induced ethylene. Though synergistic
interactions between ethylene and JA have been proposed to
contribute to diverse plant responses to abiotic and biotic
stresses (Zhu et al. 2006; Feys and Parker 2000; Glazebrook
2001; Lorenzo et al. 2003), the relationships between JA and
ethylene in regulating developmental processes, other than
plant defence responses, has not been studied intensively.
The exogenous application of JA rapidly enhances ethylene
emission in tomato and supports the indication that they act
synergistically in the expression of proteinase inhibitors
(PIN) genes in response to wound stress (O’Donnell et al.
1996). Ethylene is found to act downstream of JA in the
wound signal transduction pathway (O’Donnell et al. 1996).
Similarly, methyl jasmonate acts synergistically with ACC
in enhancing ethylene production and promoting gum for-
mation in tulips through stimulated ACC oxidase activity
(Saniewski et al. 2003). The interactive role of these hor-
monal signals in developmental processes such as root hair
formation has been clearly demonstrated in recent studies
(Zhu et al. 2006). Studies on the promoter activities of
Arabidopsis ACC synthase genes have recently shown that
exogenous JA could enhance the activity of Arabidopsis
ACC synthase (the rate-limiting step in ethylene biosyn-
thesis) promoter AtACS4 in the wild-type genotype (Tang
et al. 2008). The above explanations indicated that ethylene
is the key player in the process of rhizome induction and
development and JA might act through ethylene induction in
the process. All of these results support a significant role of
the PGRs ethylene and JA in the induction and growth of
rhizomes in Rheum rhabarbarum.
Taken together, the findings of this study strongly suggest
that ethylene is the rhizome induction signal in rhubarb. The
significant antagonistic effect of AVG on rhizome growth
even in the presence of exogenous ethylene and JA further
confirms this and suggests that JA’s role in the process is
possibly through stimulating endogenous ethylene levels
(Zhu et al. 2006; Tang et al. 2008). Further experiments
focusing on the interaction of ethylene and JA utilising
respective biosynthesis and action inhibitors would certainly
disclose the cross-talk between the two hormones in the
regulation of rhizome induction and growth in rhubarb. The
findings of such investigations would improve our under-
standing of how these two hormones regulate growth and
development processes in different crop species. However,
unlike in model plant species like Arabidopsis, in the
absence of hormonal signalling mutants of this plant, it is
very challenging to unveil the hormonal signalling pathway
involved in this developmental process.
Acknowledgements Financial support from AgriFocus 2000—
Technology Development Program (Nova Scotia Department of
Agriculture and Fisheries) and Knol Farms Ltd., Nova Scotia, Can-
ada, to Dr. Lada is gratefully acknowledged.
Appendix: details of the plant growth regulator
application
Plant-cell-tested commercial formulations of growth regu-
lators from Sigma-Aldrich, Canada, were used. Stock
solutions of GA3 (1,000 mg l-1), ABA (100 mg l-1) and
Plant Cell Tiss Organ Cult (2011) 105:253–263 261
123

JA (1,000 mg l-1) were prepared by dissolving them first
in four drops of 95% ethanol and diluted with distilled
water to the final stock concentrations. ACC, AVG and
ethephon (Sigma-Aldrich, Canada) were dissolved in dis-
tilled water to make the stock solutions of concentrations
1,000 mg l-1, 50 mg l-1 and 1,000 mg l-1, respectively.
ABA and GA3 were autoclaved with other media compo-
nents, whereas all other growth regulators were filter-ster-
ilised and added to the autoclaved culture media after
cooling to 45–50�C. An ethanol control (with four drops of
95% ethanol) and a distilled water control were maintained
in all three sets of experiments, since most of the plant
growth regulators (PGRs) were dissolved in ethanol before
making up the stock solution.
The concentrations of each PGR used in each stage of
the experiments are given below.
Treatments in stage I
10 ng l-1, 1 lg l-1, 10 lg l-1, 1 mg l-1 JA
0.1 lg l-1, 1 lg l-1, 10 lg l-1, 100 lg l-1 ABA
10 mg l-1, 20 mg l-1, 40 mg l-1, 80 mg l-1 GA3
1 mg l-1, 10 mg l-1, 50 mg l-1 ethephon
Control DW—control with distilled water (DW)
Control EtOH—control with ethanol (EtOH)
Treatments in stage II
1 mg l-1, 10 mg l-1, 50 mg l-1 ethephon
0.5 mM, 1.0 mM, 1.5 mM ACC
0.5 lM, 1.0 lM, 1.5 lM AVG
Control DW—control with distilled water (DW)
Control EtOH—control with ethanol (EtOH)
Treatments in stage III
1 mg l-1, 10 mg l-1 ethephon
10 ng l-1, 1 lg l-1 JA
1 mg l-1 ethephon and 1.5 lM AVG
10 ng l-1 JA and 1.5 lM AVG
10 ng l-1 JA and 1 mg l-1 ethephon
10 ng l-1 JA, 1 mg l-1 ethephon and 1.5 lM AVG
Control DW—control with distilled water (DW)
Control EtOH—control with ethanol (EtOH)
References
Abeles FB, Morgan PW, Saltveit ME Jr (1992) Ethylene in plant
biology, 2nd edn. Academic Press, New York, USA, 414 pp
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P,
Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C,
Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L,
Ansari Y, Choy N, Deen H, Geralt M, Hazari N, Hom E, Karnes
M, Mulholland C, Ndubaku R, Schmidt I, Guzman P, Aguilar-
Henonin L, Schmid M, Weigel D, Carter DE, Marchand T,
Risseeuw E, Brogden D, Zeko A, Crosby WL, Berry CC, Ecker
JR (2003) Genome-wide insertional mutagenesis of Arabidopsisthaliana. Science 301:653–657
Bhatia AK, Pandita ML, Khurana SC (1992) Plant growth substances
and sprouting conditions—II. Effect on tuber yield and multi-
plication rate in seed potato production. J Indian Potato Assoc
19:154–156
Bjornseth EH (1946) The effect on yield of freezing and various
ethylene treatments in breaking the dormancy of rhubarb. Proc
Am Soc Hort Sci 48:369-372
Bleecker AB, Kende H (2000) Ethylene: a gaseous signal molecule in
plants. Annu Rev Cell Dev Biol 16:1–18
Caldiz DO (1996) Seed potato (Solanum tuberosum L.) yield and
tuber number increase after foliar applications of cytokinins and
gibberellic acid under field and glasshouse conditions. Plant
Growth Regul 20:185–188
Debeljak N, Regvar M, Dixon KW, Sivasithamparam K (2002)
Induction of tuberisation in vitro with jasmonic acid and sucrose
in an Australian terrestrial orchid, Pterostylis sanguinea. J Plant
Growth Regul 36:253–260
Dimasi-Theriou K, Economou AS, Sfakiotakis EM (1993) Promotion
of petunia (Petunia hybrida L.) regeneration in vitro by ethylene.
Plant Cell Tissue Organ Cult 32:219–225
Ecker JR (1995) The ethylene signal transduction pathway in plants.
Science 268:667–675
El-Antably HMM, Wareing PF, Hillman J (1967) Some physiological
responses to d,l abscisin (dormin). Planta 73:74–90
Etheridge N, Chen Y, Schaller GE (2005) Dissecting the ethylene
pathway of Arabidopsis. Brief Funct Genomic Proteomic 3(4):
372–381
Feys BJ, Parker JE (2000) Interplay of signaling pathways in plant
disease resistance. Trends Genet 16:449–455
Gao XY, Jiang Y, Lu JQ, Tu PF (2009) One single standard substance
for the determination of multiple anthraquinone derivatives in
rhubarb using high-performance liquid chromatography-diode
array detection. J Chromatogr A 1216(11):2118–2123
Glazebrook J (2001) Genes controlling expression of defense
responses in Arabidopsis—2001 status. Curr Opin Plant Biol
4:301–308
Hartmann HT, Kester DE, Davies FT Jr, Geneve RL (2002) Plant
propagation: principles and practices, 7th edn. Prentice Hall,
New Jersey, USA, 880 pp
Jasik J, de Klerk GJ (2006) Effect of methyl jasmonate on
morphology and dormancy development in lily bulblets regen-
erated in vitro. J Plant Growth Regul 25:45–51
Jasik J, Mantell SH (2000) Effects of jasmonic acid and its
methylester on in vitro microtuberisation of three food yam
(Dioscorea) species. Plant Cell Rep 19:863–867
Kim SK, Kim JT, Jang SW, Lee SC, Lee BH, Lee IJ (2005)
Exogenous effect of gibberellins and jasmonate on tuber
enlargement of Dioscorea opposita. Agron Res 3:39–44
Koda Y, Kikuta Y (1991) Possible involvement of jasmonic acid in
tuberization of yam plants. Plant Cell Physiol 32:629–633
Leon J, Rojo E, Sanchez-Serrano JJ (2001) Wound signalling in
plants. J Exp Bot 52:1–9
Liu J, Mukherjee I, Reid DM (1990) Adventitious rooting in
hypocotyls of sunflower (Helianthus annus L.) seedlings. III.
The role of ethylene. Physiol Plant 78:268–276
Lorenzo O, Solano R (2005) Molecular players regulating the
jasmonate signalling network. Curr Opin Plant Biol 8:532–540
Lorenzo O, Piqueras R, Sanchez-Serrano JJ, Solano R (2003)
ETHYLENE RESPONSE FACTOR1 integrates signals from
262 Plant Cell Tiss Organ Cult (2011) 105:253–263
123

ethylene and jasmonate pathways in plant defense. Plant Cell
15:165–178
Moore R, Clark WD, Vodopich DS (1998) Botany, 2nd edn.
McGraw-Hill, New York, USA, 512 pp
Morris D (1996) Hormonal regulation of source–sink relationship: an
overview of potential control mechanisms. In: Zamski E,
Schaffer AA (eds) Photoassimilate distribution in plants and
crops: source–sink relationship. M. Dekker, New York,
pp 441–465
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Neuteboom CE, Lada RL, Caldwell CD, Eaton L, Havard P (2002)
Ethephon and Spermidine enhance bulking in dicer carrots
(Daucus carota var. sativus). PGRSA Q 30:77
O’Donnell PJ, Calvert C, Atzorn R, Wasternack C, Leyser HMO,
Bowles DJ (1996) Ethylene as a signal mediating the wound
response of tomato plants. Science 274:1914–1917
Obasi MO, Atanu SO (2004) Effect of growth regulators on growth,
flowering and rhizome yield of ginger (Zingiber officinate Rosc).
Niger J Hort Sci 9:69–73
Prescott AG, John P (1996) DIOXYGENASES: molecular structure
and role in plant metabolism. Annu Rev Plant Physiol Plant Mol
Biol 47:245–271
Raven PH, Evert RF, Eichhorn SE (2005) Biology of plants, 7th edn.
W.H. Freeman and Company, New York, 944 pp
Ravnikar M, Zel J, Plaper I, Spacapan A (1993) Jasmonic acid
stimulates shoot and bulb formation of garlic in vitro. J Plant
Growth Regul 12:73–77
Rayirath UP (2008) Physiology of rhizome growth and development
and propagule production technologies in rhubarb (Rheumrhabarbarum L.). Dissertation, NSAC, Dalhousie University,
Canada
Rayirath UP, Lada RR, Caldwell CD, Asiedu SK, Sibley KJ, Adams
AD (2009) CCC and Prohexadione-Ca enhance rhizome growth
and lateral bud production in rhubarb (Rheum rhabarbarum L.).
J Plant Growth Regul 28:137–146
Reid MS (1987) Ethylene in plant growth, development, and
senescence. In: Davies PJ (ed) Plant hormones and their role
in plant growth and development, 3rd edn. Kluwer Academic
Publishers, Boston, USA, pp 257–279
Roggemans J, Claes M-C (1979) Rapid clonal propagation of rhubarb
by in vitro culture of shoot-tips. Sci Hortic 11:241–246
Romanov GA, Aksenova NP, Konstantinova TN, Golyanovskaya SA,
Kossmann J, Willmitzer L (2000) Effect of indole-3-acetic acid
and kinetin on tuberisation parameters of different cultivars and
transgenic lines of potato in vitro. Plant Growth Regul
32:245–251
Saniewski M, Kawa-Miszczak L, Wegrzynowicz-Lesiak E, Misczcak
A, Ueda J, Miyamoto K (2003) Interaction of methyl jasmonate
and 1-aminocyclopropane-1-carboxylic acid (ACC) in regulation
of some physiological processes in uncooled and cooled tulip
bulbs. In: Proceedings of the XXVI International Horticultural
Congress: Elegant Science in Floriculture, Toronto, Canada,
August 2002, ISHS Acta Horticulturae, vol 622, pp 233–241
Santos I, Salema R (2000) Promotion by jasmonic acid of bulb
formation in shoot cultures of Narcissus triandrus L. Plant
Growth Regul 30:133–138
SAS Institute Inc. (1999) Proprietary Software Version 8. Cary, NC,
USA
Schaller F, Schaller A, Stintzi A (2005) Biosynthesis and metabolism
of jasmonates. J Plant Growth Regul 23:179–199
Shi YH, Zhu SW, Mao XZ, Feng JX, Qin YM, Zhang L, Cheng J,
Wei LP, Wang ZY, Zhu YX (2006) Transcriptome profiling,
molecular biological, and physiological studies reveal a major
role for ethylene in cotton fiber cell elongation. Plant Cell
18:651–664
Tang X, Chang L, Wu S, Li P, Liu G, Wang NN (2008) Auto-
regulation of the promoter activities of Arabidopsis 1-aminocy-
clopropane-1-carboxylate synthase genes AtACS4, AtACS5, and
AtACS7 in response to different plant hormones. Plant Sci
175:161–167
Van Zhong G, Burns JK (2003) Profiling ethylene-regulated gene
expression in Arabidopsis thaliana by microarray analysis. Plant
Mol Biol 53:117–131
Vreugenhil D, van Dijk W (1989) Effects of ethylene on the
tuberization of potato (Solanum tuberosum) cuttings. Plant
Growth Regul 8:31–39
Walkey DGA, Mathews KA (1979) Rapid clonal propagation of
rhubarb (Rheum rhaponticum L.) from meristem-tips in tissue
culture. Plant Sci Lett 14:287–290
Wardlaw IF (1990) The control of carbon partitioning in plants. New
Phytol 116:341–381
Xu X, van Lammeren AAM, Vermeer E, Vreugdenhil D (1998) The
role of gibberellin, abscisic acid, and sucrose in the regulation of
potato tuber formation in vitro. Plant Physiol 117:575–584
Yang SF, Hoffman NE (1984) Ethylene biosynthesis and its
regulation in higher plants. Annu Rev Plant Physiol Plant Mol
Biol 35:155–189
Zaib-Un-Nissa A, Rafiq A (1980) Effect of ABA and GA3 on
tuberization and some chemical constituents of potato. Plant Cell
Physiol 21:1343–1346
Zhang ZJ, Zhou WJ, Li HZ, Zhang GQ, Subrahmaniyan K, Yu JQ
(2006) Effect of jasmonic acid on in vitro explant growth and
microtuberization in potato. Biol Plant 50:453–456
Zheng CS, Zheng XS, Ohno H, Hara T, Matsui S (2005) Involvement
of ethylene and gibberellin in the development of rhizomes and
rhizome-like shoots in oriental cymbidium hybrids. J Jpn Soc
Hortic Sci 74:306–310
Zhu C, Gan L, Shen Z, Xia K (2006) Interactions between jasmonates
and ethylene in the regulation of root hair development in
Arabidopsis. J Exp Bot 57:1299–1308
Plant Cell Tiss Organ Cult (2011) 105:253–263 263
123




















