
Ethylene stimulates the accumulation of ripening-related mRNAs in tomatoes
9
Platit, Cell atid Etivironment (1987) 10, 177-184 Ethylene stimulates the accumulation of ripening-related mRNAs in tomatoes M. J. MAUNDERS, M. J. HOLDSWORTH, A. SLATER*, J. E. KNAPP, C. R. BIRDf, W. SCHUCHf & D. GRIERSON Department of Physiology and Environmental Science, University of Nottingham, School of Agriculture, Sutton Bonington, Loughborough, Leicestershire, U.K. Received 17 September 1986; accepted for publication 26 October 1986 Abstract. Seven cDNA clones for ripening-related mRNAs were used as hybridization probes to study the regulation of gene expression during tomato {Lyeopersieon eseulentum Mill) ripening. The mRNAs corresponding to each clone were detected by Northern blotting and the sizes of the transcripts compared with the molecular weights of polypeptides identified previously by hybrid-release translation. Dot hybridization to poIy(A)-t- mRNA from leaves, roots, unripe and ripe tomatoes established that five of the clones encoded mRNAs that were ripening specific and two encoded mRNAs also expressed in other organs. Appearance of these ripening-specific mRNAs was correlated with ethylene production as ripening commenced and occurred befote lycopene synthesis began. Thereafter, the mRNAs increased to maximum levels at the orange stage and then declined slowly as fruit turned red. When tnature green fruit were picked before any increase in ethylene synthesis occurred and placed in air plus 10 cm-* m^-* ethylene, substantial changes in trans- latable tnRNA were noted within 30 h. Five mRNAs that increased in amount directed the synthesis in vitro of poiypeptides similar in size to those encoded by the five ripening-specific clones. Several different patterns of mRNA accumulation were observed, with increases in concentration ofthe order of 300-fold. Key-words: Lycopersicon escutetttutn Mill; Solanaceae; tomato; ripening; ethylene; mRNA; hybridization. Introduction Fruit ripening is a co-ordinated process during which colour, texture, fiavour, aroma and other attributes change in a tegulated manner. These changes are caused by alterations in enzyme activity (Sacher, 1973), which are in turn thought to be detertnined •Present address: School of Life Sciences, Scraptoft Campus. Leieester Polytechnic, Leicester LE9 7SU. U.K. tPre.sent address: Plant Biotechnology Group, ICI pic. PO Box 1 I, The Heath. Runcorn. Cheshire WA7 4QE. U.K. Correspondence: Dr D. Grierson, Department of Physiology and Environmental Seience. University of Nottingham. School of Agriculture, Sutton Bonington, Loughborough. Leicestershire LE12 5RD, U.K. by alterations in specific gene expression. De novo enzytne synthesis has been demonstrated for tomato {Lyeopersieon eseulentutn Mill) invertase (Iki et ai, 1978) and polygalacturonase (Tucker, Robertson & Grierson 1980; Tucker & Grierson, 1982), and avocado {Per.sea atnerieana Mill) cellulase (Christoffersen, Warm & Laties, 1982). During ripening, climacteric fruit produce significant quantities of ethylene frotn methionine, catalysed by I-aminocyclo-propane-1-carboxylic acid (ACC) synthase (Adams & Yang, 1979) and the 'ethylene-forming enzyme'. Administration of exogenous ethylene to mature unripe fruit can initiate or accelerate ripening (Sacher, 1973; McGIasson, Wade & Adato, 1978), and stimulate autocatalytic ethylene synthesis. In tomatoes, this effect is dependent on the fruit being of sufficient maturity to respond to ethylene. At the level of colour, texture and flavour changes, the artificially induced ripening sequence is indistinguishable from the natural process. A similar pattern of changes is also observed at the biochetnical level with respect to tespiration (Theologis & Laties, 1982), mRNA profile, polysome levels and enzyme activities (;ChristoiTersen, Tucker & Laties, 1984; Grierson & Tucker, 1983). Tomato fruit actively synthesize RNA (Rattanapanone, Grierson & Stein, 1977; Rattanapanone, Speirs & Grierson, 1978) and protein (Grierson et cd., 1985) throughout the ripening period, and a nutnber of changes in the pattern of protein synthesis during development and ripening can be observed by two-dimensional gel electrophotesis (Baker, Anderson & Hruschka, 1985). Specific changes occur in the mRNA population (Rattanapanone et ai, 1978; Speirs et ai, 1984; Grierson, 1984; Grierson et ai, 1985). /;; vitro translation studies have shown that some mRNAs disappear while eight or more new species appear after the onset of ripening (Grierson et ai, 1985). Ripening-related clones were identified in a tomato cDNA library by Slater etal. (1985) using a differential hybridization technique. This procedure revealed at least 19 different mRNAs that were present in ripe fruit, but wete undetectable or present at a reduced level in unripe fruit. Five of these 177
-
Upload
nottingham -
Category
Documents
-
view
0 -
download
0
Transcript of Ethylene stimulates the accumulation of ripening-related mRNAs in tomatoes

Platit, Cell atid Etivironment (1987) 10, 177-184
Ethylene stimulates the accumulation of ripening-relatedmRNAs in tomatoes
M. J. MAUNDERS, M. J. HOLDSWORTH, A. SLATER*, J. E. KNAPP, C. R. BIRDf, W. SCHUCHf & D.GRIERSON Department of Physiology and Environmental Science, University of Nottingham, School ofAgriculture, Sutton Bonington, Loughborough, Leicestershire, U.K.
Received 17 September 1986; accepted for publication 26 October 1986
Abstract. Seven cDNA clones for ripening-relatedmRNAs were used as hybridization probes to studythe regulation of gene expression during tomato{Lyeopersieon eseulentum Mill) ripening. ThemRNAs corresponding to each clone were detectedby Northern blotting and the sizes of the transcriptscompared with the molecular weights of polypeptidesidentified previously by hybrid-release translation.Dot hybridization to poIy(A)-t- mRNA from leaves,roots, unripe and ripe tomatoes established that fiveof the clones encoded mRNAs that were ripeningspecific and two encoded mRNAs also expressed inother organs. Appearance of these ripening-specificmRNAs was correlated with ethylene production asripening commenced and occurred befote lycopenesynthesis began. Thereafter, the mRNAs increased tomaximum levels at the orange stage and thendeclined slowly as fruit turned red. When tnaturegreen fruit were picked before any increase inethylene synthesis occurred and placed in air plus10 cm-* m^-* ethylene, substantial changes in trans-latable tnRNA were noted within 30 h. FivemRNAs that increased in amount directed thesynthesis in vitro of poiypeptides similar in size tothose encoded by the five ripening-specific clones.Several different patterns of mRNA accumulationwere observed, with increases in concentration oftheorder of 300-fold.
Key-words: Lycopersicon escutetttutn Mill; Solanaceae; tomato;ripening; ethylene; mRNA; hybridization.
Introduction
Fruit ripening is a co-ordinated process during whichcolour, texture, fiavour, aroma and other attributeschange in a tegulated manner. These changes arecaused by alterations in enzyme activity (Sacher,1973), which are in turn thought to be detertnined
•Present address: School of Life Sciences, Scraptoft Campus.Leieester Polytechnic, Leicester LE9 7SU. U.K.
tPre.sent address: Plant Biotechnology Group, ICI pic. PO Box1 I, The Heath. Runcorn. Cheshire WA7 4QE. U.K.
Correspondence: Dr D. Grierson, Department of Physiologyand Environmental Seience. University of Nottingham. School ofAgriculture, Sutton Bonington, Loughborough. LeicestershireLE12 5RD, U.K.
by alterations in specific gene expression. De novoenzytne synthesis has been demonstrated for tomato{Lyeopersieon eseulentutn Mill) invertase (Iki et ai,1978) and polygalacturonase (Tucker, Robertson &Grierson 1980; Tucker & Grierson, 1982), andavocado {Per.sea atnerieana Mill) cellulase(Christoffersen, Warm & Laties, 1982).
During ripening, climacteric fruit producesignificant quantities of ethylene frotn methionine,catalysed by I-aminocyclo-propane-1-carboxylic acid(ACC) synthase (Adams & Yang, 1979) and the'ethylene-forming enzyme'. Administration ofexogenous ethylene to mature unripe fruit caninitiate or accelerate ripening (Sacher, 1973;McGIasson, Wade & Adato, 1978), and stimulateautocatalytic ethylene synthesis. In tomatoes, thiseffect is dependent on the fruit being of sufficientmaturity to respond to ethylene. At the level ofcolour, texture and flavour changes, the artificiallyinduced ripening sequence is indistinguishable fromthe natural process. A similar pattern of changes isalso observed at the biochetnical level with respect totespiration (Theologis & Laties, 1982), mRNAprofile, polysome levels and enzyme activities(;ChristoiTersen, Tucker & Laties, 1984; Grierson &Tucker, 1983).
Tomato fruit actively synthesize RNA(Rattanapanone, Grierson & Stein, 1977;Rattanapanone, Speirs & Grierson, 1978) andprotein (Grierson et cd., 1985) throughout theripening period, and a nutnber of changes in thepattern of protein synthesis during development andripening can be observed by two-dimensional gelelectrophotesis (Baker, Anderson & Hruschka,1985). Specific changes occur in the mRNApopulation (Rattanapanone et ai, 1978; Speirs et ai,1984; Grierson, 1984; Grierson et ai, 1985). /;; vitrotranslation studies have shown that some mRNAsdisappear while eight or more new species appearafter the onset of ripening (Grierson et ai, 1985).
Ripening-related clones were identified in a tomatocDNA library by Slater etal. (1985) using adifferential hybridization technique. This procedurerevealed at least 19 different mRNAs that werepresent in ripe fruit, but wete undetectable or presentat a reduced level in unripe fruit. Five of these
177

178 M. J. MAUNDERS ET AL.
mRNAs encoded polypeptides similar to those thathad previously been found to increase duringripening (Grierson et ai, 1985), and which werepresumed to be related to the ripening process.
In this study, gene expression during ripening wasinvestigated using seven cDNA clones, thetranslation products of which have been identified byhybrid select translation (Slater et ai, 1985). Wehave investigated the validity of defining the mRNAsas 'ripening specific' by measuring expression inorgans other than ripe fruit, and have monitored thecourse of mRNA accumulation over the ripeningperiod. Expression has also been measured indetached fruit induced to ripen in an atmospherecontaining ethylene to examine the role of the gas instimulating ripening, and to determine whether ornot it induces a similar pattern of ripening at thelevel of gene expression.
Materials and methods
Plant material
Tomato plants {Lyeopersieon eseulentum Mill. cv.Ailsa Craig) were grown from surface-sterilized seedsunder conditions minimizing the possibifity of viralcontamination (Grierson et ai, 1985). Fruit wereharvested when the appropriate stage of ripeness hadbeen reached. Prior to RNA extraction, the ethyleneeffusion rate was measured by the method ofGrierson & Tucker (1983), and the lycopene contentwas assayed from a small portion of each fruit(Tomes, 1963). Some fruit were harvested at themature green stage (7 weeks after fertilization) andwere placed in an atmosphere containing 10 cm^m^^ ethylene. Samples were taken for RNAextraction at intervals over a period of 6 d. Leaftissue was harvested from partly expanded axillaryshoots whenever required for RNA extraetion. Rootswere induced on excised axillary shoots byimmersion in 10"^ mol m"^ naphthyl aeetic acid for6 d. After transfer to distilled water for 7-10 d theadventitious roots were excised and the RNAextracted.
DNA-RNA hybridizations
One /ig samples of poly(A) -I- RNA were used for do thybridizations to •'^P-labelled cloned probes asdescribed by Slater et ai (1985). Following au to -radiography on pre-flashed Kodak X-Omat AT filmat — 75°C, the signals were quantified by scanningthe film on a Joyce Loebl Chromoscan 3 densito-meter and comparing to standard radioactive D N Asamples. This method was sensitive to 0 5 nopoly(A)-h RNA. *̂ ^
For Northern hybridizations, 12/ig samples ofglyoxal-denatured poly(A)-|- RNA were electro-phoresed through 1% agarose gels in 12.5 mol m~3sodium phosphate buffer, pH 6.5 at 20 V/cm. Sizestandard markers consisting of 0.5 /(g Hind mdigested lambda DNA were also run. The markertracks were excised and stained with 1 ^g cm~3ethidium bromide to visualize the bands. ThepoIy(A)+ RNA tracks were transferred toGenescreen transfer membrane (New EnglandNuclear) under conditions described by the suppliersAfter baking at 80 °C for 2-5 h, the filters wereprehybridized and hybridized in the same manner asthe dot hybridizations.
Results
Organ specificity of the eDNA eloties
Seven of the 'ripening-related' cDNA clones isolatedpreviously (Slater et ai, 1985) were used in thehybridization experiments: pTOM 5, pTOM 6pTOM 13, pTOM 36, pTOM 75, pTOM 99 andpTOM 137. None of these clones hybridized to eachother (Slater et ai, 1985). Hybridization of each todots of poly(A)-f RNA isolated from ripe fruit,unripe fruit, roots and leaves, showed that themRNAs for five of the clones were only present inripe fruit (Fig. 1). After long exposure of the X-rayfilm, slight hybridization of pTOM 13 to poly(A)-fRNA from unripe fruit was also detected, but thiswas approximately 300-fold less than to poly(A)4-RNA from ripe fruit.
mRNA preparation and in vitro protein synthesis
Poly(A) -I- RNA was prepared from tomato pericarpas described by Slater et al. (1985). Total RNA fromleaves and roots was prepared by the method ofCovey & Grierson (1976) and poly(A)-f RNAprepared from this in the same manner as forpericarp RNA.
A mRNA-dependent rabbit reticulocyte lysatesystem prepared by the method of Pelham & Jackson(1976) was obtained from Dr M. Wilson (John InnesInstitute, Norwich, U.K.). This was used for thetranslation of poly(A)-l- RNA (0.5 ng RNA/10assay) as described by Slater el ai (1985).
Ripe
Unripe
fruit
fruit
Root
Leaf
•
5
...
6 13
•
36 Ta 99
pTOM number
•
0#137
Figure 1. Expression in different organs of mRNAs homologous toripening-related cDNAs. -'^P-labelled nick-translated cDNAprobes were hybridized to I fig lots of poly(A)+ RNA fromvarious tomato plant tissues. The numbers refer to the cDNAclones (see Table I).
ETHYLENE STIMULATES RIPENING-RELATED mRNA ACCUMULATION 179
13 36 75 99
cDNA clone number
137
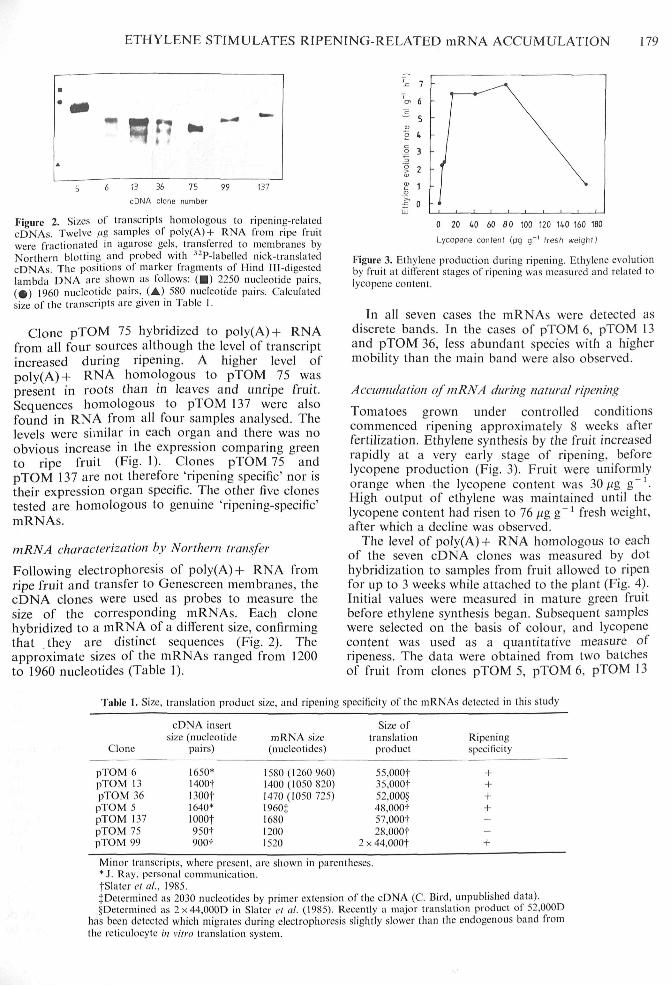
Figure 2. Sizes of transcripts homologous to ripening-relatedcDNAs. Twelve /(g .samples of poly(A)+ RNA from ripe fruitwere fractionated in agarose gels, transferred to membranes byNorthern blotting and probed with -"P-labelled nick-translatedcDNAs. The positions of marker fragments of Hind Ill-digestedlambda DNA arc shown as follows: ( • ) 2250 nucleotide pairs,( 9 ) 1960 nucleotide pairs. (A) 5H0 nucleotide pairs. Calculatedsize of the transcripts are given in Table 1.
Clone pTOM 75 hybridized to poly(A)-|- RNAfrom all four sources although the level of transcriptincreased during ripening. A higher level ofpoly(A)-l- RNA hotnologous to pTOM 75 waspresent in roots than in leaves and unripe fruit.Sequences homologous to pTOM 137 were alsofound in RNA from all four samples analysed. Thelevels were similar in each organ and there was noobvious increase in the expression cotnparing greento ripe fruit (Fig. 1). Clones pTOM 75 andpTOM 137 are not therefore 'ripening specific' nor istheir expression organ specific. The other five clonestested are homologous to genuine 'ripening-specific'mRNAs.
mRNA eharaeterization by Northern tran.sfer
Following electrophoresis of poly(A)-l- RNA fromripe fruit and transfer to Genescreen tnembranes, thecDNA clones were used as probes to measure thesize of the corresponding mRNAs. Each clonehybridized to a mRNA of a different size, confirmingthat they are distinct sequences (Fig. 2). Theapproximate sizes of the niRNAs ranged from 1200to 1960 nueleotides (Table 1).
0 20 40 60 BQ 100 120 UO 160 160
Lycopene confenf (pg g~' fresh weight)
Figure 3. Ethylene production during ripening. Ethylene evolutionby fruit at different stages of ripening was measured and related tolycopene content.
In all seven cases the mRNAs were detected asdiscrete bands. In the cases of pTOM 6, pTOM 13and pTOM 36, less abundant species with a highermobility than the main band were also observed.
Aeeuntulatiott of mRNA during natural ripening
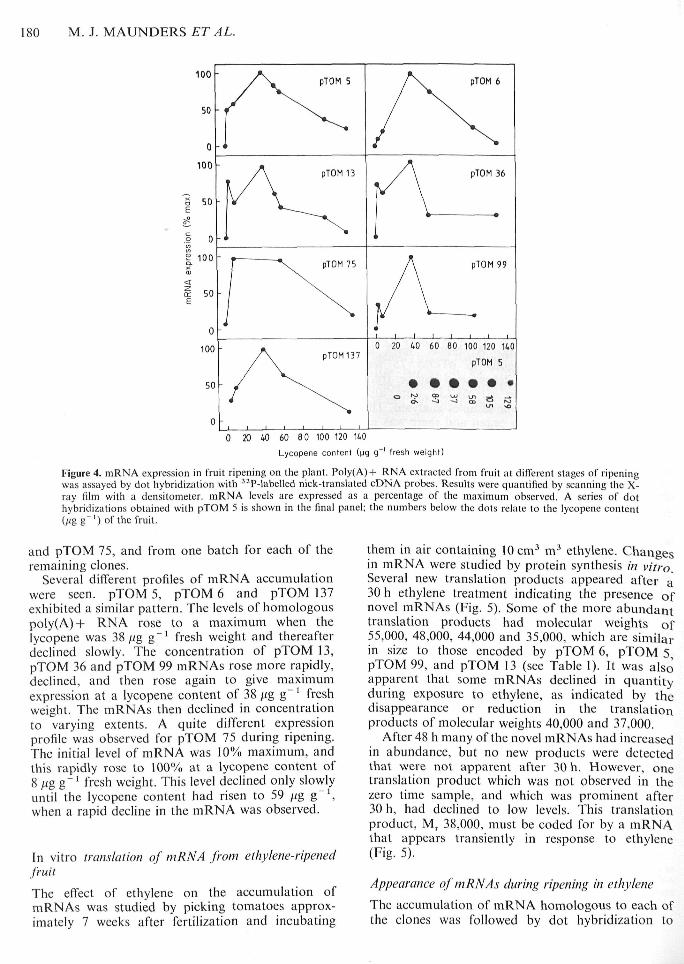
Tomatoes grown under controlled conditionscommenced ripening approxitnately 8 weeks afterfertilization. Ethylene synthesis by the fruit increasedrapidly at a very early stage of ripening, beforelycopene production (Fig. 3). Fruit were uniformlyorange when the lycopene content was 30//g g~'.High output of ethylene was maintained until thelycopene content had risen to 76 |ig g^' fresh weight,after which a decline was observed.
The level of poly(A)-l- RNA homologous to eachof the seven cDNA clones was measured by dothybridization to samples from fruit allowed to ripenfor up to 3 weeks while attached to the plant (Fig. 4).Initial values were measured in mature green fruitbefore ethylene synthesis began. Subsequent sampleswere selected on the basis of colour, and lycopenecontent was used as a quantitative tneasure ofripeness. The data were obtained from two batchesof fruit from clones pTOM 5, pTOM 6, pTOM 13
Table 1, Size, translation product size, and ripening speeilicity ofthe mRNAs detected in this study
Clone
pTOM 6pTOM UpTOM 36
pTOM 5pTOM 137pTOM 75pTOM 99
cDNA insertsize (nucleotide
pairs)
1650*14001l300t1640*lOOOt950t900t
mRNA .size(nueleotides)
1580 (1260 960)1400 (1050 820)1470(1050 725)1960t168012001520
Size oftranslation
product
55.000t35.000t52.000§48,000t57.000t28.000t
2 X 44.000t
Ripeningspeciticity
++++-—+
Minor transcripts, where pre.scnt. are shown in parentheses.*J. Ray. personal communication.tSlater (V «/., 1985.tDelermined as 2030 nueleotides by primer extension ofthe cDNA (C. Bird, unpublished data).§Deteimined as 2 x 44.000D in Slater et al. (1985). Reeently a major translation produet of 52.O0OD
has heen detected which migrates during electrophoresis slightly slower than the endogenous band frotnthe reticulocyte itt vitro translation .system.
180 M. J. MAUNDERS ET AL.
100
50
20 40 50 8 0 100 120 140
Lycopene content {\iq g"' fresh weight)
Figure 4. mRNA expression in fruit ripening on the pfant. Pofy(A)-l- RNA extraeled from fruit at difTerent stages of ripeningwas assayed by dot hybridization with ^^P-labelled niek-translated eDNA probes. Results were quantified by seanning the X-ray film with a densitometer. mRNA levels are expressed as a percentage of the maximum observed. A series of dothybridizations obtained with pTOM 5 is shown in the fnial panel; the numbers below the dots relate to the lyeopene eontent
; g" ' ) of the fruit.
and pTOM 75, and from one batch for each of theremaining clones.
Several different profiles of mRNA accumulationwere seen. pTOM 5, pTOM 6 and pTOM 137exhibited a similar pattern. The levels of homologouspoly(A)-l- RNA rose to a maximum when thelycopene was 38/ig g" ' fresh weight and thereafterdeclined slowly. The concentration of pTOM 13,pTOM 36 and pTOM 99 mRNAs rose more rapidly,declined, and then rose again to give maximumexpression at a lycopene content of 38 ;<g g~' freshweight. The mRNAs then declined in concentrationto varying extents. A quite different expressionprofile was observed for pTOM 75 during ripening.The initial level of mRNA was 10% maximum, andthis rapidly rose to 100% at a lycopene content of8 /ig g" ' fresh weight. This level declined only slowlyuntil the lycopene content had risen to 59 pg g" ' ,when a rapid decline in the mRNA was observed.
In vitro translation of mRNA from ethylene-ripenedfruit
The effect of ethylene on the accumulation ofmRNAs was studied by picking tomatoes approx-imately 7 weeks after fertilization and incubating
them in air containing 10 cm^ nv' ethylene. Changesin mRNA were studied by protein synthesis //; vitro.Several new translation products appeared after a30 h ethylene treatment indicating the presence ofnovel mRNAs (Fig. 5). Some of the more abundanttranslation products had molecular weights of55,000, 48,000, 44,000 and 35,000, which are similarin size to those encoded by pTOM 6, pTOM 5pTOM 99, and pTOM 13 (see Table 1). It was alsoapparent that some mRNAs declined in quantityduring exposure to ethylene, as indicated by thedisappearance or reduction in the translationproducts of molecular weights 40,000 and 37,000.
After 48 h many of the novel mRNAs had increasedin abundance, but no new products were detectedthat were not apparent after 30 h. However, onetranslation product which was not observed in thezero time sample, and which was prominent after30 h, had declined to low levels. This translationproduct, Mr 38,000, must be coded for by a mRNAthat appears transiently in response to ethylene(Fig. 5).
Appearance of mRNAs during ripening in ethylene
The accumulation of mRNA homologous to each ofthe clones was followed by dot hybridization to

ETHYLENE STIMULATES RIPENING-RELATED mRNA ACCUMULATION 181
Figure 5. /" vitro translation of mRNAs produeed during ripeningin elhylene. Poly(A)-l- RNA extracted from mature green fruitbefore and after ineubation in air containing 10 em-' m" ' ethylenewas translated in Ihe presenee of -"S-melhionine. The productswere separated on a 10-15% gradient polyacrylamide-SDS gel anddetected by Ifuorography. The positions and sizes (lVl,x 10"-') of('''C)-labelled marfcer proleins are indicated on the left. ( • ) Noveltranslation products appearing after 30 h; (O) translationproducts declining after elhylene Ireatmenl; (A) a noveltranslation produel Ihat deelines within 48 h.
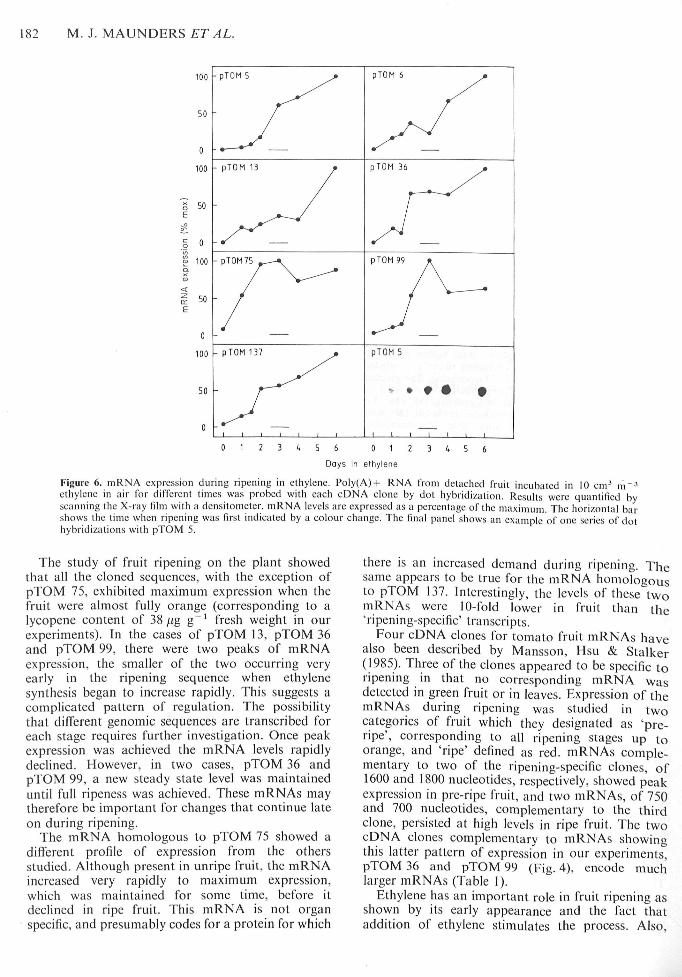
poly(A)-l- RNA extracted from mature green fruitincubated in ethylene over a period of 6 d (Fig. 6).These data are the average of three to sixexperiments for each clone. A number of differentpatterns of gene expression were observed. ThemRNA corresponding to pTOM 5 exhibited a lag of24-36 h before it began to accumulate, followed by arapid and sustained increase. mRNAs homologousto clones pTOM 6 and pTOM 137 showed no lag inexpression, with significant amounts present after24 h. mRNA continued to accumulate to Day 6.pTOM 13 mRNA showed an increase over the first24 h, followed by a plateau in the level of mRNA forthe next 36 h, after which a second, larger increasewas observed. Clone pTOM 36 also showed abiphasic pattern of expression: in this case, the initialrise in mRNA amount continued for 48 h andreached a plateau at a much higher level ofexpression than pTOM 13. Transcripts homologousto clone pTOM 75 exhibited an immediate and rapidrise to a maximum level after 2-3 d; this wasfollowed by a slow decline. In contrast, the initialaccumulation of pTOM 99 mRNA was slow; it thenreached maximum expression wifhin 36 h andsubsequently declined slowly.
Discussion
The ripening of tomato fruit is accompained bycolour changes. These are due in part to thedegradation of chlorophyll, and to an increase inlycopene content, beginning 7-8 weeks after fertiliza-
tion, and continuing for 7-10 d when the fruit arefully ripe. At the same time a large increase insynthesis and evolution of ethylene by the fruit isobserved. This increase is very rapid, reaching highlevels early in ripening, but synthesis declines beforemaximum lycopene content is achieved (Fig. 3). Therise in ethylene synthesis and other ripeningphenomena such as softening can be stimulated, andto some extent synchronized in different fruit, by theapplication of exogenous ethylene. Ethylene has beenshown to stimulate gene expression in ripeningavocado fruit (Tucker & Laties, 1984) and in otherplant tissues (Nichols & Laties, 1984). In these cases,a general increase in expression of 'constitutive'genes is observed, and some specific responses suchas the appearance of avocado cellulase mRNA havealso been detected (Tucker & Laties, 1984).
The genes studied in this work have all beenshown to be ripening-related in that their expressionchanges throughout the ripening period. ThemRNAs corresponding to clones pTOM 5, pTOM 6,pTOM 36 and pTOM 99 were undetectablc in unripefruit, roots and leaves. Transcripts related topTOM 13 could be detected in unripe fruit at a levelof 0.3% of that in ripe fruit, but the mRNAshomologous to the other four clones were present atlevels less than this. Thus, the expression of mRNAshomologous to these cDNA clones appears to bespecific to ripening. It would be interesting to extendthe range of cell types investigated to determinewhether any of them are expressed in cellsundergoing senescence processes such as those inabscission zones and senescing leaves. pTOM 75and pTOM 137 both hybridized strongly topoly(A)-f mRNA from all four sources examined,but expression in green fruit was 5-10% themaximum observed during ripening. Our resultsindicate the importance of investigating severaldifferent stages of development and ripening. Thus,although mRNA homologous to pTOM 137 doeschange substantially in concentration during ripening(Fig. 4), this is not apparent if samples are onlytaken from unripe and fully ripe fruit (Fig. 1).
Analysis of the mRNAs by Northern transfer andhybridization showed that distinct mRNAshybridized to each cDNA clone. This is consistentwith earlier results that none of the clones cross-hybridize with each other (Slater et al, 1985). Inmost cases a single band of mRNA was detected,but pTOM 6, pTOM 13 and pTOM 36 eachhybridized to several RNA bands (Fig. 2, Table I). Itis perhaps significant that //) vitro translation of themRNA hybrid selected by pTOM 6, pTOM 13, andpTOM 36 yields some minor polypeptides of smallermolecular weights than the major band (Slater et al,1985; A. Slater, unpublished data). The multipleRNA species are presumably due either to discretedegradation of the major mRNAs, or totranscription of other partially-homologous ripening-related genes.
182 M. J. MAUNDERS ET AE.
100
50
0
100
S 50
9 0
S 100Q.
i 50E
0
100
50
0
- pTOM 13 •
- pTOM75 ^ ^ - \
^ pTOM 137 .
1 1 1 1 I 1 1
pTOM 6 .
pTOM 36 .
pTOM99 n
pTOM 5
V . • • •
! 1 1 ~ 1 1 1
2 3 5 6 0 1
Days in ethyiene
2 3
Figure 6. mRNA expression during ripening in ethylene. Poly(A)-(- RNA from detaehed fruit ineubated in 10 em' m~^ethylene in air for different limes was probed with each cDNA clone by dot hybridization. Results were quanlilied byscanning the X-ray film with a densitometer. mRNA levels are expressed as a percentage of the maximum The horizontal barshows the time when ripening was first indieated by a colour change. The fmal panel shows an example of one series of dothybridizations wilh pTOM 5.
The study of fruit ripening on the plant showedthat all the cloned sequences, with the exception ofpTOM 75, exhibited maximum expression when thefruit were almost fully orange (corresponding to alycopene content of 38,ug g~' fresh weight in ourexperiments). In the cases of pTOM 13, pTOM 36and pTOM 99, there were two peaks of mRNAexpression, the smaller of the two occurring veryearly in the ripening sequence when ethylenesynthesis began to increase rapidly. This suggests acomplicated pattern of regulation. The possibilitythat different genomic sequences are transcribed foreach stage requires further investigation. Once peakexpression was achieved the mRNA levels rapidlydeclined. However, in two cases, pTOM 36 andpTOM 99, a new steady state level was maintaineduntil full ripeness was achieved. These mRNAs maytherefore be important for changes that continue lateon during ripening.
The mRNA homologous to pTOM 75 showed adifferent profile of expression from the othersstudied. Although present in unripe fruit, the mRNAincreased very rapidly to maximum expression,which was maintained for some time, before itdeclined in ripe fruit. This mRNA is not organspecific, and presumably codes for a protein for which
there is an increased demand during ripening. Thesame appears to be true for the inRNA homologousto pTOM 137. Interestingly, the levels of these twomRNAs were 10-fold lower in fruit than the'ripening-specific' transcripts.
Four cDNA clones for tomato fruit mRNAs havealso been described by Mansson, Hsu & Stalker(1985). Three of the clones appeared to be specific toripening in that no corresponding mRNA wasdetected in green fruit or in leaves. Expression of themRNAs during ripening was studied in twocategories of fruit which they designated as 'pre-ripe', corresponding to all ripening stages up toorange, and 'ripe' defined as red. mRNAs comple-mentary to two of the ripening-specific clones, of1600 and 1800 nucleotides, respectively, showed peakexpression in pre-ripe fruit, and two mRNAs, of 750and 700 nucleotides, complementary to the thirdclone, persisted at high levels in ripe fruit. The twocDNA clones complementary to mRNAs showingthis latter pattern of expression in our experiments,pTOM 36 and pTOM 99 (Fig. 4), encode muchlarger mRNAs (Table 1).
Ethylene has an important role in fruit ripening asshown by its early appearance and the fact thataddition of ethylene stimulates the process. Also,

ETHYLENE STIMULATES RIPENING-RELATED mRNA ACCUMULATION 183
ripening is inhibited by aminoethoxy vinyl glycinewhich prevents ethylene synthesis (Yang & Hoffman,1984), and silver ions which interfere with ethyleneaction (Hobson et al., 1984). It has been shownpreviously that ethylene applied to mature green fruitstimulates the synthesis of lycopene and poly-galacturonasc (Gricrson & Tucker, 1983).Accordingly, we examined the effects of exogenousethylene on the appearance of ripening-specificmRNAs in mature green fruit. Translation in vitroshowed that ethylene stimulated the accumulation ofat least eight mRNAs within 30 h (Fig. 5). Thetranslation products of most of these mRNAscorrespond in size to those previously shown toappear during natural ripening (Grierson et al,1985). Five arc similar to translation products ofmRNAs hybrid-selected by tipening-specific clones(Table 1). The levels of these mRNAs were measuredby hybridization during the first 6 d of ethylene-induced ripening. In most cases significant expressionwas observed within 1 d, although visible signs ofripening were not observed until 60-80 h after thestart of the experiment. In most cases, mRNA levelscontinued to increase throughout the 6 d reachinglevels similar to those found during natural ripening.Part of the explanation for the slow rise may berelated to the sequential pattern of ripening, noted inall our experiments, that occurs in different parts ofthe fruit, with more and more cells being recruited toripening with time. This is similar to the situationthat occurs if fruit are left to ripen naturally. Incontrol experiments, mature green fruit incubatedwithout added ethylene began to show an increase inendogenous ethylene synthesis after 4-7 d. Once thisoccured the accumulation of ripening-relatedmRNAs could also be detected. The time scale forthe appearance of new mRNAs in response toethylene was similar to that observed for avocado byTucker & Laties (1984). However, the fact thatmRNAs homologous to most of the ripening-specificclones were still increasing in concentration after 6 dsuggests that, although important for the progress ofripening, these transcripts are not likely to beinvolved in the initial event triggering ripening.
The hybridization experiments did not detect anymRNAs that reached maximum expression rapidlyin response to ethylene. This may be explained by thefact that the cDNA clones used as probes in thisstudy were prepared from mRNA from fully ripenedfruit. Early transcripts produced within the first fewhours of exposure to ethylene may well have declinedas ripening got underway and would not berepresented in the 'ripening-related' cDNA library.The existence of at least one such transient mRNAwas shown by the //; vitro translation experiments(Fig. 5). Thus, Ihe preparation of cDNA clones fromtomato fruit at this early stage might assist inidentifying 'early genes' implicated in ripeninginitiation.
The identities of some mRNAs that respond to
ethylene treatment in other plant cells have beendetermined. Cellulase mRNA increases 50-fold inripening avocado (Christoffersen et al, 1984; Tucker& Laties, 1984) and phenylalanine ammonia lyasemRNA increases in carrot (Daucus carota L.) roots(Christoffersen & Laties, 1982). During tomatoripening, the cell wall degrading enzyme poly-galacturonase is synthesized de novo in response toethylene (Grierson & Tucker, 1983). Immuno-precipitation of translation products hybrid-selectedby tomato ripening cDNAs indicated that pTOM 5encodes tomato polygalacturonase (Slater et al,1985). However, there is disagreement about the sizeof the polygalacturonase mRNA translation productin vitro (Sato et al, 1984; Grierson et al, 1985). Inorder to identify unequivocally the clone forpolygalacturonase, we have sequenced the clonedinserts in pTOM 5, pTOM 6 and pTOM 13. Thisallows us to predict the amino acid sequence of theproteins encoded by the corresponding mRNAs.Comparison of these sequences with the amino acidsequence determined for the N-terminus of tomatopolygalacturonase indicates that pTOM 6 encodestomato polygalacturonase. For a full account ofthese results, see Grierson et al, 1986. The otherripening-specific mRNAs may encode enzymesinvolved in pigment synthesis, starch degradationand ethylene synthesis itself.
Acknowledgments
This work was supported by grants to D. Griersonfrom the Agriculture and Food Research Counciland a co-operative research grant with ICI fromthe Science and Engineering Research Council.M. .1. Holdsworth is the recipient of a CASE awardf>om the Science and Engineering Research Council.
References
Adams. D.O. & Yang. S.F. (1979) Ethylene biosynthesis:idenlilieation of 1-aminoeyclopropane-l-carboxylie acid as anintermediate in the eonversion of niethionine lo ethylene.Proeeedings of the National Aeadeniy of Seiences, U.S.A., 76,170-174.'
Baker. .I.E.. Anderson. J.D. & Hrnsehka. W.R. (1985) Proteinsynthesis in tomato fruil periearp tissue during npennig.Charaeteristies of amino acid incorporation. Jiwrnal i>J PlantPhy.siology. 120, 167-179.
Covey. S.N. & Grierson. D. (1976) The measurement of plantpolyadenylie aeid by hybridisation with radioaetive polyuridyhcacid. Pliwta, 131, 75-79. .
Chtistodcrsen. R.E. & Lalies. G.G. (1982) Elhylene regulation olgene expression in carrots. Proeeedings of the Nattonal Acailetnyof .Scietiees, U.S.A., 79, 4060-4063.
Christoffersen. R.E.. Tucker. M.L. & Laties. G.G. (1984)Cellulase gene expression in ripening avoeado Iruit: tlieaeeumiilation of celhila.se mRNA and protein as demonstratedby eDNA hybridisation and immunodeleelion. Plant MolecttlarBiologv. 3, .i;85-391.
Chrislollbtsen. R.E.. Warm. E. & Lalies. G.G. (1982) Geneexpression during fruit ripening in avoeado. Planta, 155, 5.-57.
Grierson. D. (1984) Nueleie aeid and protein synthesis during fruitripening and seneseenee. In Cell Ageittg and Cell Death (eds

184 M. J. MAUNDERS ET AE,
I. Davies and D.C. Sigee), pp. 189-202. Cambridge UniversityPress, Cambridge.
Grierson. D., Slater. A.. Speirs. .1. & Tucker. G.A. (1985) Theappearance of polygalacturonase mRNA in tomatoes: one of aseries of ehanges in gene expression during development andripening. Planta, 163, 263-271.
Grierson, D. & Tueker, G.A. (1983) Timing of ethylene andpolygalacluronase .synthesis in relation to the eontrol of tomatofruit ripening. Planta, 157, 174-179.
Grierson, D.. Tucker. G.A.. Keen. J., Ray, J., Bird. C.R. &Sehuch. W. (1986) Sequencing and identification of a eDNAclone lor tomato polygalaeturonase. Nueleie Aeids Researeh, 14,8595-8603.
Hobson, G.E., Niehols, R., Davies. J.N. & Atkey, P. (1984) Theinhibition of tomato fruil ripening by silver. Journal of PlantPhy.siology, \\(>,2\-29.
Iki. K.. Sekiguehi, K., Kurata, K., Tada, T., Nakagawa, M.,Ogura, N. & Takehana, H. (1978) Immunologieal propertiesof /i-fruetoturanosidase from ripening tomato fruit. Phyto-ehemistry, 17, 311-312.
McGfasson. W.B., Wade. N.L. & Adato, I. (1978) Phytohormonesand fruit ripening. In Phytohormones and Related Compounds—a Comprehensive Treatise (eds D.S. Lethani, P.B. Goodwin andT.J.V. Higgins). Vol. 2, pp. 447^93. Elsevier/Norlh-Holland,Amsterdam.
Mansson, P.E.. Hsu. D. & Stalker, D. (1985) Charaeterisation offruit specific cDNAs from tomato. Molecular and GeneralGeneties, 200, 356-361.
Nichols, S.E. & Laties, G.D. (1984) Ethylene-regulated genetranseription in carrot roots. Plant Molecular Biology, 3,393^01.
Pelham, H.R.B. & Jaekson. R.J. (1976) An effieient mRNA-dependent translation system from reticulocyte lysates.European Journat of Biochemistry, 67, 347-256.
Rattanapanone, N.. Grierson, D. & Stein, M. (1977) Ribonueleicacid metabolism during the development and ripening oftomato fruits. Phytochemistry, 16, 629-633.
Raltanapanone. N.. Speirs. J. & Grierson. D. (1978) Evidence forchanges in messenger RNA eontent related lo lonialo fruitripening. Phytoehemistry, 17, 1485-1486.
Saeher. J.A. (1973) Seneseenee and postharvest physiology. AnnualReview of Plant Physiohgy, 24, 197-224.
Sato, T., kusaba. S., Nakagawa. H. & Ogura. N. (1984) Cell-freesynthesis of a putative preeursor ofpolygalacturona.se in tomatofruits. Platit atid Cell Physiology, 25, 1069-1071.
Slater, A.. Maunders. M.J.. Edwards. K.. Sehueh. W. & GriersonD. (1985) Isolation and characterisation of cDNA clones fortomato polygalaeturonase and other ripening-related proteinsPlant Moleettlar Biology, 5, 137-147.
Speirs, J., Brady, C.J.. Grierson, D. & Lee. E. (1984) Changes inribosome organisation and messenger RNA abundance inripening tomato fruits. Australian Jiwrtial of Plant Physiology,11,225-233. • 'Ry.
Theologis. A. & Laties, G.G. (1982) Potentiating efleet of pureoxygen on the enhancement of respiration f:)y ethylene in plantstorage organs: a comparative study. Plant Physiology fio1031-1035. ^.>. oy,
Tomes, M.L. (1963) Temperalure inhibition of carotene syntfiesisin tomato. Botanieal Gazette, 124, 180-185.
Tucker. G.A. & Grierson, D. (1982) Synthesis ofpolygalacluronase during tomato fruil ripening. Planta 1«;<;64-67. ' '
Tucker. M. & Laties, G.G. (1984) Interrelationship of geneexpression, polysome prevalence, and respiration duringripening of elhylene and/or eyanide-treated avoeado fruit PlatiiPhysiology, 74, 307-315.
Tueker. G.A.. Robertson, N.G. & Grierson. D. (1980) Changes inpolygalacturonase isoenzymes during the 'ripening' of normaland mutant tomato fruit. European Journal of Bii>ehetnistry,112,119-124. • •*•
Yang. S.E. & Hoffman. N.E. (1984) Ethylene biosynthesis and itsregulation in higher planls. Atttiual Revie\y of Plant Phy.sioloBv35, 155-189. • *"-̂ '





















