
Fruit Development, Ripening and Quality Related Genes in the Papaya Genome
32
Fruit Development, Ripening and Quality Related Genes in the Papaya Genome Robert E. Paull & Beth Irikura & Pingfang Wu & Helen Turano & Nancy Jung Chen & Andrea Blas & John K. Fellman & Andrea R. Gschwend & Ching Man Wai & Qingyi Yu & Gernot Presting & Maqsudul Alam & Ray Ming Received: 5 November 2008 / Accepted: 12 November 2008 / Published online: 18 December 2008 # Springer Science + Business Media, LLC 2008 Abstract Papaya (Carica papaya L.) is the first fleshy fruit with a climacteric ripening pattern to be sequenced. As a member of the Rosids superorder in the order Brassicales, papaya apparently lacks the genome duplication that occurred twice in Arabidopsis. The predicted papaya genes that are homologous to those potentially involved in fruit growth, development, and ripening were investigated. Genes homologous to those involved in tomato fruit size and shape were found. Fewer predicted papaya expansin genes were found and no Expansin Like-B genes were predicted. Compared to Arabidopsis and tomato, fewer genes that may impact sugar accumulation in papaya, ethylene synthesis and response, respiration, chlorophyll degradation and carotenoid synthesis were predicted. Similar or fewer genes were found in papaya for the enzymes leading to volatile production than so far determined for tomato. The presence of fewer papaya genes in most fruit development and ripening categories suggests less subfunctionalization of gene action. The lack of whole genome duplication and reductions in most gene families and biosynthetic pathways make papaya a valuable and unique tool to study the evolution of fruit ripening and the complex regulatory networks active in fruit ripening. Keywords Fruit growth . Expansins . Ethylene . Respiration . Carotenoids . Chlorophyll . Volatiles . Sugar accumulation Introduction The cultivated papaya (Carica papaya L.) is the most important economic species in Caricaceae and is the only member of the genus Carica [19, 20]. Papaya and Arabidopsis have both been placed in the Brassicales [8, 9]. This classification is supported by morphological and molecular characteristics [160, 161, 165, 166]. Whereas the core Brassicales (including Arabidopsis and Brassica) and Moringaceae (Moringa oleifera) have dry siliques or capsules, papaya has a fleshy berry fruit. Papaya is Tropical Plant Biol. (2008) 1:246–277 DOI 10.1007/s12042-008-9021-2 Communicated by Dr. Paul Moore Electronic supplementary material The online version of this article (doi:10.1007/s12042-008-9021-2) contains supplementary material, which is available to authorized users. R. E. Paull (*) : B. Irikura : P. Wu : H. Turano : N. J. Chen : C. M. Wai Department of Tropical Plant and Soil Sciences, University of Hawaii at Manoa, Honolulu, HI 96822, USA e-mail: [email protected] A. Blas : G. Presting Department of Molecular Biosciences and Bioengineering, University of Hawaii at Manoa, Honolulu, HI 96822, USA J. K. Fellman Department of Horticulture and Landscape Architecture, Washington State University, Pullman, WA 99164, USA A. R. Gschwend : R. Ming Department of Plant Biology, University of Illinois at Urbana-Champaign, Urbana, IL 61801, USA Q. Yu Hawaii Agriculture Research Center, Aiea, HI 96701, USA M. Alam Advanced Studies in Genomics, Proteomics and Bioinformatics, University of Hawaii, Honolulu, HI 96822, USA
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Fruit Development, Ripening and Quality Related Genes in the Papaya Genome

Fruit Development, Ripening and Quality Related Genesin the Papaya Genome
Robert E. Paull & Beth Irikura & Pingfang Wu &
Helen Turano & Nancy Jung Chen & Andrea Blas &
John K. Fellman & Andrea R. Gschwend &
Ching Man Wai & Qingyi Yu & Gernot Presting &
Maqsudul Alam & Ray Ming
Received: 5 November 2008 /Accepted: 12 November 2008 /Published online: 18 December 2008# Springer Science + Business Media, LLC 2008
Abstract Papaya (Carica papaya L.) is the first fleshy fruitwith a climacteric ripening pattern to be sequenced. As amember of the Rosids superorder in the order Brassicales,papaya apparently lacks the genome duplication thatoccurred twice in Arabidopsis. The predicted papaya genes
that are homologous to those potentially involved in fruitgrowth, development, and ripening were investigated.Genes homologous to those involved in tomato fruit sizeand shape were found. Fewer predicted papaya expansingenes were found and no Expansin Like-B genes werepredicted. Compared to Arabidopsis and tomato, fewergenes that may impact sugar accumulation in papaya,ethylene synthesis and response, respiration, chlorophylldegradation and carotenoid synthesis were predicted. Similaror fewer genes were found in papaya for the enzymes leadingto volatile production than so far determined for tomato. Thepresence of fewer papaya genes in most fruit developmentand ripening categories suggests less subfunctionalization ofgene action. The lack of whole genome duplication andreductions in most gene families and biosynthetic pathwaysmake papaya a valuable and unique tool to study theevolution of fruit ripening and the complex regulatorynetworks active in fruit ripening.
Keywords Fruit growth . Expansins . Ethylene .
Respiration . Carotenoids . Chlorophyll . Volatiles .
Sugar accumulation
Introduction
The cultivated papaya (Carica papaya L.) is the mostimportant economic species in Caricaceae and is the onlymember of the genus Carica [19, 20]. Papaya andArabidopsis have both been placed in the Brassicales [8,9]. This classification is supported by morphological andmolecular characteristics [160, 161, 165, 166]. Whereas thecore Brassicales (including Arabidopsis and Brassica) andMoringaceae (Moringa oleifera) have dry siliques orcapsules, papaya has a fleshy berry fruit. Papaya is
Tropical Plant Biol. (2008) 1:246–277DOI 10.1007/s12042-008-9021-2
Communicated by Dr. Paul Moore
Electronic supplementary material The online version of this article(doi:10.1007/s12042-008-9021-2) contains supplementary material,which is available to authorized users.
R. E. Paull (*) : B. Irikura : P. Wu :H. Turano :N. J. Chen :C. M. WaiDepartment of Tropical Plant and Soil Sciences,University of Hawaii at Manoa,Honolulu, HI 96822, USAe-mail: [email protected]
A. Blas :G. PrestingDepartment of Molecular Biosciences and Bioengineering,University of Hawaii at Manoa,Honolulu, HI 96822, USA
J. K. FellmanDepartment of Horticulture and Landscape Architecture,Washington State University,Pullman, WA 99164, USA
A. R. Gschwend : R. MingDepartment of Plant Biology,University of Illinois at Urbana-Champaign,Urbana, IL 61801, USA
Q. YuHawaii Agriculture Research Center,Aiea, HI 96701, USA
M. AlamAdvanced Studies in Genomics, Proteomics and Bioinformatics,University of Hawaii,Honolulu, HI 96822, USA

polygamous and is a herbaceous, single-stemmed and erectplant. Other than Carica papaya L., the other edible speciesin the family are Vasconcella candamarcensis Hook. f., V.monoica Desf., V. erythrocarpa Heilborn, V. goudotianaSolms-Laubach and V. quercifolia Benth. and Hook [187].These fruit are normally dry and lacking the juicy flesh ofC. papaya are mostly eaten cooked. Another edible species,V. pentagona, is called ‘babaco’. Isozyme and AFLPanalysis indicates that C. papaya is only distantly relatedto the Vasconcella species. The greatest diversity in C.papaya exists in the Yucatan-San Ignacio-Peter-Rio Mota-gua area of Central America. The wild population in thisarea has greater diversity than domesticated populations[134, 205].
The papaya fruit is a fleshy berry and fruit growthfollows a single sigmoid growth curve [224]. Duringdevelopment, all tissues in the gynoecium less than 1 mmare meristematic [170]. Later, the outer layer of theepidermis increases in size while the subepidermal layercontinues to divide both anticlinally and periclinally. Thecentral parenchyma of the pericarp increases in size anddivides with the placenta forming opposite the marginalvascular bundles. This meristematic activity lasts 28–42 days and determines final fruit size. Fruit growth showstwo major phases. The first lasts about 80 days post-anthesis, with a large increase in dry weight occurring justbefore fruit maturity. Fruit development takes 150–164 daysthat is extended by another 14–21 days in Hawaii in thecolder months [145, 155]. Mesocarp growth parallels seedand total fruit growth.
Papaya fruit shape is a sex-linked character and rangesfrom spherical to ovoid in female flowers to long,cylindrical or pyriform (pear-shaped) in hermaphroditeflowers. The fruit is normally composed of five carpelsunited to form a central ovarian cavity that is lined with theplacenta carrying numerous black seeds. Placentation isparietal with the seeds attached by 0.5–1 mm stalks. Theovarian cavity is larger in female fruit than hermaphrodite.The shape of the cavity at the transverse cut ranges fromstar-shape with five to seven furrows to smooth and circular[45].
Papaya ripening is climacteric with the rise in ethyleneproduction occurring at the same time as the respiratory rise[146]. Respiration is a critical factor in fruit growth anddevelopment, especially as it relates to fruit ripening. Fruitsare divided into two broad groups based on the role ofethylene in the ripening process and its relationship to therespiratory pattern. Climacteric fruits (e.g., banana, papaya,peach, tomato) demonstrate a peak in respiration andethylene production during ripening, including autostimu-latory (so-called System 2) ethylene production [27]. Thetiming of the autostimulatory ethylene peak [23, 108] andof other ripening events (skin color changes, carotenoid
synthesis, flavor development, softening) varies widelybetween species and cultivars [27, 35, 38, 86]. Non-climacteric fruit a gradual decline in the respiration rate,no marked peak in ethylene production and gradualchanges in other ripening parameters.
The papaya genome has been recently sequenced [132].The sequences represented 75% (277.4 Mb) of the(372 Mb) genome and 90% of the euchromatic regions.The assembled EST unigene set contained 16,362 unigeneswith 92.1% matching of the assembled whole shotgunsequences (WGS). The transcribed sequences matched only3.6% (13.4 Mb) of the whole genome and 48% of the genicregion in WGS [132]. This resource allows us to determinethe number of genes that are potentially involved in fruitgrowth and development, and in fruit ripening. Anunderstanding of the predicted gene number in each familyis the first step in determining which members of eachfamily are expressed and at what level during fruitdevelopment. In this review, we provide an overview ofthe number of predicted genes potentially involved in thedifferent phases of fruit growth, development and ripening.
Results and Discussion
Fruit Development
The papaya plant is a herb that can grow up to 9–10 m inheight and generally has only a single stem. Lateralbranching does occur if the apex is damaged or when theplant is tall. The lack of lateral shoot growth in plants hasbeen associated with auxin inhibition, abscisic acid (ABA)and strigolactone [82, 115, 221]. The plant growth regulatorABA plays a crucial role in the plant’s adaptation to stressand in seed maturation and dormancy, and fruit ripeningand senescence [221]. ABA and strigolactone are productsfrom the cleavage of carotenoids at one of its double bonds.Carotenoid cleavage is carried out by two groups of relatedenzymes. One group uses multiple carotenoid substratesand is referred to as carotenoid cleavage dioxygenase(CCD). The other group has C40-9-cis-epoxycarotenoid asthe preferred substrate and referred to as 9-cis-epoxycar-otenoid dioxygenase (NCED). The NCEDs are thought tobe involved in ABA synthesis while the role of CCDs isless clear, with some evidence to suggest a role in lateralshoot growth [14, 15], possibly by cleavage of carotenoidsto strigolactone or a related compound [82].
Two Arabidopsis genes CCD7/MAX3 and CCD8/MAX4control lateral shoot growth [14, 31, 82]. The citrushomolog gene (CsNCED1), likely plays a role in ABAsynthesis in leaves and fruits [163]. ABA has also beengiven a role in the regulation of citrus fruit skin coloration[162]. Citrus is a non-climacteric fruit and normally
Tropical Plant Biol. (2008) 1:246–277 247247

responds to ethylene by induced fruit coloration [3]. Papayais climacteric fruit though fruit mesocarp carotenoiddevelopment and fruit degreening are ethylene independent[121]. ABA via CCD and/or NCED may therefore play arole in fruit mesocarp carotenoid development and skindegreening. One CCD (CpCCD1) and two NCEDs(CpNCED1, CpNCED2) are predicted in papaya (Table 1).A partial open reading frame (ORF) was found for anotherpossible CpCCD (ABIM01019146.1) that had 67% identity(E-value 5e-37) to Arabidopsis CCD1 (NP_191911.1). Apapaya EST was not found for CpCCD1 or CpNCED1 anda chloroplast transit peptide was not found for CpNCED1.The Arabidopsis carotenoid cleavage family has fiveputative NCEDs and four putative CCDs [15] and tomatohas two (LeCCD1A and LeCCD1B) [180]. CpCCD1 had69% identity (E-value 0.0) with Arabidopsis AtNCED4,CpNCED1 has 73% identity (E-value 0.0) with ArabidopsisAtNCED5. CpNCED2 had 77% identity (E-value 0.0) withAtNECED3. The CpCCD and the CpNCEDs were allpredicted to have a carotenoid oxygenase motif.
Fruit Growth
The papaya fruit is a fleshy berry and fruit growth follows asingle sigmoid growth curve [224] and can weigh from 0.2 kgto 10 kg [136]. This plasticity in size provides a uniqueopportunity to study fruit development control. Tomato fruitcan similarly vary from small berries (~2 g) to large fruit(1,000 g) [113] with a single gene ORFX (QTL loci fw 2.2)being responsible for 30% of the difference [24, 52, 75].ORFX appears to act at or near the plasma membrane βsubunit of CKII kinase [52] and exerts its control throughearly fruit cell division [113]. One homolog to ORFX waspredicted in papaya (CpORFX; ABIM01013288.1) with 63%identity (E-value 2e-49) with ORFX from Solanum pennellii(AF261775.1) and 62% (E-value 1e-49) with Solanumlycopersicum (AAF74286.1). CpORFX shows homology tothe cysteine-rich PLAC8 domain of unknown function that isfound in animals and plants [123]. No papaya ESTs were
detected for CpORFX. The function of CpORFX in papaya isunknown though the wide range of papaya fruit size presentthe possibility it may have a similar role to that in tomato.
Fruit Shape
The shape of papaya fruit varies with fruit from femaleflowers being spherical to ovoid, while fruit from hermaph-rodite flowers being cylindrical or pear shaped [136]. Amutation in the quantitative trait locus (QTL), OVATE,changes the shape of tomato from round to pear-shaped[114]. This regulatory gene apparently has its impact earlyin flower development [104, 105]. A similar QTL ineggplant has a similar effect on shape [104]. OVATE is anegative regulatory hydrophilic protein with a putativebipartite nuclear localization signal [114]. Three OVATEproteins were predicted in the papaya genome with ho-mology to OVATE sequences in Arabidopsis (AAY78676.1)and Solanum lycopersicum (AAN17752.1). Papaya ESTswere only found for one of the gene (CpOVATE1,ABIM01006651.1) that had about 38% identity (E-value4e-06) to Solanum lycopersicum OVATE (AAN17752.1)and 39% identity (E-value 4e-35) with Arabidopsis(NP_178114.1). All three predicted papaya genes had the~70 aa plant specific motif (DUF623) of unknown func-tion, also reported for the tomato OVATE gene [114].CpOVATE 2 (ABIM01006789.1) has 31% identity (E-value2e-25) with Arabidopsis Ovate family (AtOFP8,NP_197466.1) and 58% identity (E-value 5e-21) with tomato(AAN17752.1). CpOVATE3 (ABIM01013535.1) which wason the same linkage group as the papaya sex related gene has67% identity (E-value 6e-22) with tomato OVATE protein(AAN17752.1) and an OVATE-like protein from tobacco(ABW05088) with 65% identity (E-value 2e-21).
Another major gene controlling the elongated fruit shapeof tomato is SUN that acts in a dosage dependent manner[214]. The gene belongs to the IQD family with AtIQD1being the only member with a known function. A homologto SUN was found in papaya (ABIM01022664.1). This
Table 1 Carotenoid cleavage dioxygenases predicted in the papayagenome, the whole genome shotgun sequence accession number inNCBI (WGS Accession), coding sequence for amino acids (CDS),amino acids (aa), whether a chloroplast transit peptide was predicted,presence of discrete portion of protein possessing its own function and
the superfamily (domain), similarity in peptide sequence with anotherspecies (homology), extent the sequences were invariant (identity), E-value was the expectation value for homology, and the number ofpapaya expressed sequence tags (EST) detected with at least 500 basesand 99% identity
WGS Accession cds aa Transit Peptide Domains Homology Identity, E-value ESTs
CpCCD1 ABIM01016290.1 1275 592 Yes Carotenoid cleavagedioxygenase 4a,
Citrus clementinaABC26011.1
72, 0.0 0
CpNCED1 ABIM01023760.1 1524 507 None 9-cis-epoxycartenoiddioxygenase 5,
Citrus clementinaABC26010.1
79, 0.0 0
CpNCED2 ABIM01015050.1 1857 618 Yes 9-cis-epoxycartenoiddioxygenase 3,
Citrus clementinaABC26013.1
77, 0.0 1
248 Tropical Plant Biol. (2008) 1:246–277

papaya homolog had four introns, the same as SUN andwas 446 aa long versus 405 aa for SUN [214]. Thehomology of the predicted papaya gene was moderate(37%, E-value 4e-34) for SUN and 50% (E-value 5e-103)for Arabidopsis IQD11 (NP_196850.1) and 35% (E-value3e-32) for IQD12 (NP_196016.1). Another predictedpapaya homolog (ABIM01018776.1) was two-thirdsthe length and had only two introns possibly due to agap between the two sequenced contigs. This secondpredicted gene was 45% (E-value 4e-27) homologousto SUN and 53% (E-value 2e-29) to Arabidopsis IQD12(NP_196016.1). The variation in papaya shape from roundto elongated presents a unique model to ascertain the roleof SUN homologs in determining fruit shape.
Expansins
The cell wall proteins termed expansin are involved in cellwall relaxation and growth [129]. A number of expansingenes are recognized [56]. The expansins have been dividedinto two classes based on sequence homology: expansin A(EXPA), expansin B (EXPB) with additional groups that areEXPA-Like and EXPB-Like proteins (EXLA, EXLB). EXPAand EXPB normally have a secretory sequence of 20–30 aa,a EG-45 domain (~120 aa) with distant homology toglycoside hydrolases (GH45) though having no enzymaticactivity, and a potential carbohydrate binding domain (CBD)of 98 aa [49, 173].
Expansins have been shown to be expressed duringtomato fruit growth [168] and different isoforms areexpressed during fruit ripening [34, 167]. Papaya fruit havealso been shown to express an expansin (CpEXPA1). Fourhave been reported in banana fruit [12]. Immunoblots havealso detected expansins in ripening pear, persimmon, kiwifruit, strawberry and pineapple but not detected in pepper[168]. The papaya genome contains at least 15 CpEXPA,three CpEXPB and one CpEXPLA (Supplementary Table 1).Secretory sequences were not predicted on four of theCpEXPAs. All CpEXPs had similar intron positions andlengths to those described for Arabidopsis [49].
The papaya expansins were part of the large superfamilyof at least three subfamilies (Fig. 1) but did not appear tohave members in three of the clades (VI, VII, X) [173] andno EXLB were found. Sampedro et al., [173, 175] proposedthat the number in the last common ancestor for expansinEXLB was four, though none were predicted for papaya(Table 2). The monocot/dicot ancestor had 15 to 17expansin genes [173]. Papaya is classified as a basal cladein the Order Brassicales [165] with at least 19 expansingenes in close to the number predicted for the monocot/dicot ancestor (15) [173].
The expansins are believed to have arisen and diversi-fied early if not before colonization on land [49, 110].
Expansins are found in monocots, pines, ferns and mosses.The closely related expansin-like sequences are similarlywidely found and the absence of any EXLB predicted inpapaya was unexpected. One EXLB is found in Arabidopsisand rice, and four in poplar [173] and one predicted in pine[174] suggesting that EXLBs were present before theangiosperms/gymnosperms separation. The Arabidopsis(At4g17030) and Pinus EXLB (TC89171) are distantlyrelated [174] but no match was found to any predictedpapaya gene sequences or in the papaya EST database.When predicted papaya protein models were queried usingthe Arabidopsis expansin like-B sequence, homology wasfound to EXPLA and EXPB sequences predicted in papaya.A number of other papaya sequences were predicted with aEG45 motif though these peptides had less than 100 aminoacids and had no CBD motif.
Skin Color Changes
The pathway of chlorophyll breakdown during fruitripening is expected to be similar to that occurring duringsenescence and involves the removal of the phytol residueand the central Mg by chlorophyllase and a dechelatase.The product of these two activities is pheophorbide-a whichis the substrate for pheophorbide-a oxygenase (PAO), andred chlorophyll catabolite reductase (RCCR) [92]. Thebreakdown product from PAO and RCCR is then exportedfrom the degenerating chloroplast for further breakdown inthe cytoplasm. The predicted Mg dechelatase gene has notbeen identified [106].
One chlorophyllase (CpCLH) gene (ABIM01011000.1)was predicted in the papaya genome versus two genes inArabidopsis thaliana and three in Brassica oleracea [92].The papaya CpCLH protein had a predicted chloroplasttransit peptide of 21 aa and had 60% homology (E-value9e-18) to B. oleraceae BoCLH3 protein (Fig. 2) and 61%(E-value 4e-17) to AtCLH2. The CpCLH has a 2Fe-2Sferrodoxin, iron sulfur binding site and was expressed inpapaya based on ESTs. A magnesium cheltase subunitgene with three ESTs was predicted in papaya(ABIM01016494.1+ABIM01016495.1). This gene had91% identity (E-value 0.0) with soybean (CAA04526.1).
PAO genes have been identified by functional genomics.A single PAO gene is predicted for papaya (CpPAO,ABIM01011326.1). The predicted gene is expressed basedupon EST data and has a 72-aa chloroplast transit peptide.CpPAO was similar (E-value 2e-131) to the ArabidopsisAtPAO gene (At3g44880), first described as AcceleratedCell Death (ACD1) in Arabidopsis. ACD1 has homology totomato lethal leaf spot 1-like protein (AAL32300) and aputative cell death suppressor protein in Oryza sativa(AAK98735). All three species’s PAO genes contain apredicted iron-sulfur binding domain.
Tropical Plant Biol. (2008) 1:246–277 249249

Fig. 1 Phylogenetic tree ofexpansin super family proteinsequences from Arabidopsis,Popular and papaya aligned withClustalW and neighbor-joiningvalues. The tree was was con-structed with MEGA 4.0. Theexpansin and popular sequenceswere downloaded from http://www.bio.psu.edu/expansins/index.htm. Accessed 2008, Sept04. CpEXPA1 had been previ-ously described (ABD65309)and was used to number thepredicted CpEXPA. EXPAorthologous gene clades are in-dicated by bars and Romannumerals from [174]
250 Tropical Plant Biol. (2008) 1:246–277

As with a single gene in Arabidopsis (At4g37000),described as ACD2 [117, 213], a single RCCR gene waspredicted in papaya (CpRCCR, ABIM01012476.1). Thepapaya CpRCCR protein has a predicted chloroplasttransit peptide (61 aa) and papaya ESTs. RCCR hasslightly greater identity (74%, E-value 4e-92) to thetomato RCCR [154] than to the Arabidopsis ACD2(71%, E-value 1e-80).
Carotenoid Biosynthesis
Carotenoids serve several functions in plants [58, 76].During fruit ripening as chlorophyll is degraded, theunderlying carotenoids are unmasked and de novo biosyn-thesis occurs in the papaya mesocarp chromoplasts.Carotenoids are derived from the 5-carbon compoundisopentenyl pyrophosphate (IPP). The C40 backbone of allcarotenoids is assembled from two C20 geranylgeranylpyrophosphate (GGPP) molecules which in turn are eachderived from four C5 IPP molecules (see volatile productionbelow).
Synthesis of phytoene from two GGPP molecules is thefirst step in the carotenoid-specific biosynthesis pathwayand is catalyzed by phytoene synthase (PSY). PSY is a keyregulator in carotenoid biosynthesis and has been found tobe the rate-limiting enzyme in ripening tomato fruits,canola seeds and marigold flowers [10, 33, 73, 74, 91].PSY is expected to have close membrane-association asphytoene is lipid-soluble and is localized, along with itssubsequent end-products, inside the chloroplasts andchromoplasts [58]. Additionally, two forms of PSY, achromoplast- and chloroplast-specific form, have beenfound in tomato: PSY-1 and PSY-2, respectively [73].Desaturation of phytoene into z-carotene and lycopene ismediated by phytoene desaturase (PDS) and ζ-carotenedesaturase (ZDS), respectively. This desaturation convertsthe colorless phytoene into the pink-hued lycopene [58].Like PSY, the desaturases PDS and ZDS are closelyassociated with the plastid membrane but are not integralmembrane proteins [177]. Plastid transit peptides weredetected for PSY, PDS, ZDS and LCY-b for the papayagenome predicted genes. The published sequences for bothPDS and ZDS appear to be lacking part of the leadersequences, and do not start with a methionine as required bythe transit peptide prediction software. When the predictedpeptide sequences for PSY and ZDS from the papayagenome were used, both of which start with methionine,transit peptides were predicted.
Cyclization of lycopene via lycopene e-cyclase (LCY-e)or lycopene b-cyclase (LCY-b) results in a a- or b-carotene,respectively. Yamamoto [218] showed that 59.3% of thetotal carotenoids in yellow-fleshed papaya are comprised ofb-carotene or its derivatives, i.e., cryptoxanthin, indicatingthe b-ring cyclization pathway for synthesis of the yellow-pigmented carotenoids. The family of lycopene cyclases inplants shows evidence of multiple gene duplication eventsand divergence of catalytic function. Plant lycopene cyclasesshare a common phylogenetic origin with bacterial crtY andcrtL lycopene cyclases. Plant and bacterial lycopene cyclasesare polypeptides of ~400 aa, however, the plant lycopenecyclases have an additional 100 aa N-terminal transitsequence. Five regions of conserved amino acid sequencehave been identified: one putative dinucleotide-bindingregion and four motifs of unknown function [10, 58].
Several papaya genes that encode enzymes in thecarotenoid biosynthesis pathway have been previouslyidentified: phytoene synthase (PSY), phytoene desaturase(PDS), ζ-carotene desaturase (ZDS) and a chloroplast-specific lycopene β-carotene (LCY-b) (Table 3). BLASTnsearch of an EST database generated from a multiple-tissuetype cDNA library supports the activity of these singlegenes in papaya. Putative papaya homologs were identifiedfor lycopene ε-cyclase (LCY-e), β-ring carotene hydroxy-lase (CRTR) and zeaxanthin epoxidase (ZE) based on amino
CpCLH1
BoCLH3
AtCLH2
BoCLH2
SoyCLH1
SoyCLH3
CitrusCLH
SoyCLH2
AtCLH1
BoCLH1
0.000.050.100.150.200.250.300.35
Fig. 2 Phylogenetic relationship between predicted papaya chloro-phyllase and published sequences from NCBI aligned with ClustalWand neighbor-joining values. Tree was constructed with MEGA 4.0
Table 2 Expansin families in different plants, gene numbers and thelast common ancestor from Sampedro and Cosgrove [171]. Theestimates for papaya do not include incomplete sequences
Species EXPA EXPB EXLA EXLB
Last Common Ancestor 12 2 1 2Papaya 15 3 1 0Arabidopsis 26 6 3 1Popular 27 2 2 4Rice 34 19 4 1
Tropical Plant Biol. (2008) 1:246–277 251251
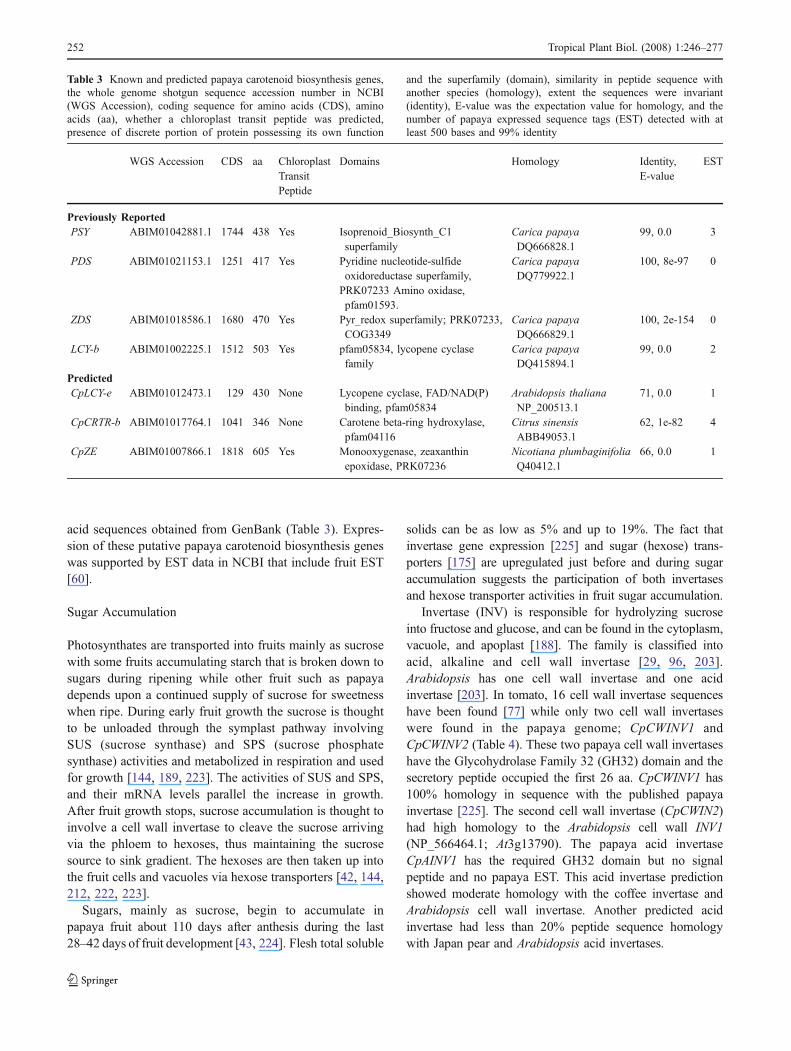
acid sequences obtained from GenBank (Table 3). Expres-sion of these putative papaya carotenoid biosynthesis geneswas supported by EST data in NCBI that include fruit EST[60].
Sugar Accumulation
Photosynthates are transported into fruits mainly as sucrosewith some fruits accumulating starch that is broken down tosugars during ripening while other fruit such as papayadepends upon a continued supply of sucrose for sweetnesswhen ripe. During early fruit growth the sucrose is thoughtto be unloaded through the symplast pathway involvingSUS (sucrose synthase) and SPS (sucrose phosphatesynthase) activities and metabolized in respiration and usedfor growth [144, 189, 223]. The activities of SUS and SPS,and their mRNA levels parallel the increase in growth.After fruit growth stops, sucrose accumulation is thought toinvolve a cell wall invertase to cleave the sucrose arrivingvia the phloem to hexoses, thus maintaining the sucrosesource to sink gradient. The hexoses are then taken up intothe fruit cells and vacuoles via hexose transporters [42, 144,212, 222, 223].
Sugars, mainly as sucrose, begin to accumulate inpapaya fruit about 110 days after anthesis during the last28–42 days of fruit development [43, 224]. Flesh total soluble
solids can be as low as 5% and up to 19%. The fact thatinvertase gene expression [225] and sugar (hexose) trans-porters [175] are upregulated just before and during sugaraccumulation suggests the participation of both invertasesand hexose transporter activities in fruit sugar accumulation.
Invertase (INV) is responsible for hydrolyzing sucroseinto fructose and glucose, and can be found in the cytoplasm,vacuole, and apoplast [188]. The family is classified intoacid, alkaline and cell wall invertase [29, 96, 203].Arabidopsis has one cell wall invertase and one acidinvertase [203]. In tomato, 16 cell wall invertase sequenceshave been found [77] while only two cell wall invertaseswere found in the papaya genome; CpCWINV1 andCpCWINV2 (Table 4). These two papaya cell wall invertaseshave the Glycohydrolase Family 32 (GH32) domain and thesecretory peptide occupied the first 26 aa. CpCWINV1 has100% homology in sequence with the published papayainvertase [225]. The second cell wall invertase (CpCWIN2)had high homology to the Arabidopsis cell wall INV1(NP_566464.1; At3g13790). The papaya acid invertaseCpAINV1 has the required GH32 domain but no signalpeptide and no papaya EST. This acid invertase predictionshowed moderate homology with the coffee invertase andArabidopsis cell wall invertase. Another predicted acidinvertase had less than 20% peptide sequence homologywith Japan pear and Arabidopsis acid invertases.
Table 3 Known and predicted papaya carotenoid biosynthesis genes,the whole genome shotgun sequence accession number in NCBI(WGS Accession), coding sequence for amino acids (CDS), aminoacids (aa), whether a chloroplast transit peptide was predicted,presence of discrete portion of protein possessing its own function
and the superfamily (domain), similarity in peptide sequence withanother species (homology), extent the sequences were invariant(identity), E-value was the expectation value for homology, and thenumber of papaya expressed sequence tags (EST) detected with atleast 500 bases and 99% identity
WGS Accession CDS aa ChloroplastTransitPeptide
Domains Homology Identity,E-value
EST
Previously ReportedPSY ABIM01042881.1 1744 438 Yes Isoprenoid_Biosynth_C1
superfamilyCarica papayaDQ666828.1
99, 0.0 3
PDS ABIM01021153.1 1251 417 Yes Pyridine nucleotide-sulfideoxidoreductase superfamily,PRK07233 Amino oxidase,pfam01593.
Carica papayaDQ779922.1
100, 8e-97 0
ZDS ABIM01018586.1 1680 470 Yes Pyr_redox superfamily; PRK07233,COG3349
Carica papayaDQ666829.1
100, 2e-154 0
LCY-b ABIM01002225.1 1512 503 Yes pfam05834, lycopene cyclasefamily
Carica papayaDQ415894.1
99, 0.0 2
PredictedCpLCY-e ABIM01012473.1 129 430 None Lycopene cyclase, FAD/NAD(P)
binding, pfam05834Arabidopsis thalianaNP_200513.1
71, 0.0 1
CpCRTR-b ABIM01017764.1 1041 346 None Carotene beta-ring hydroxylase,pfam04116
Citrus sinensisABB49053.1
62, 1e-82 4
CpZE ABIM01007866.1 1818 605 Yes Monooxygenase, zeaxanthinepoxidase, PRK07236
Nicotiana plumbaginifoliaQ40412.1
66, 0.0 1
252 Tropical Plant Biol. (2008) 1:246–277
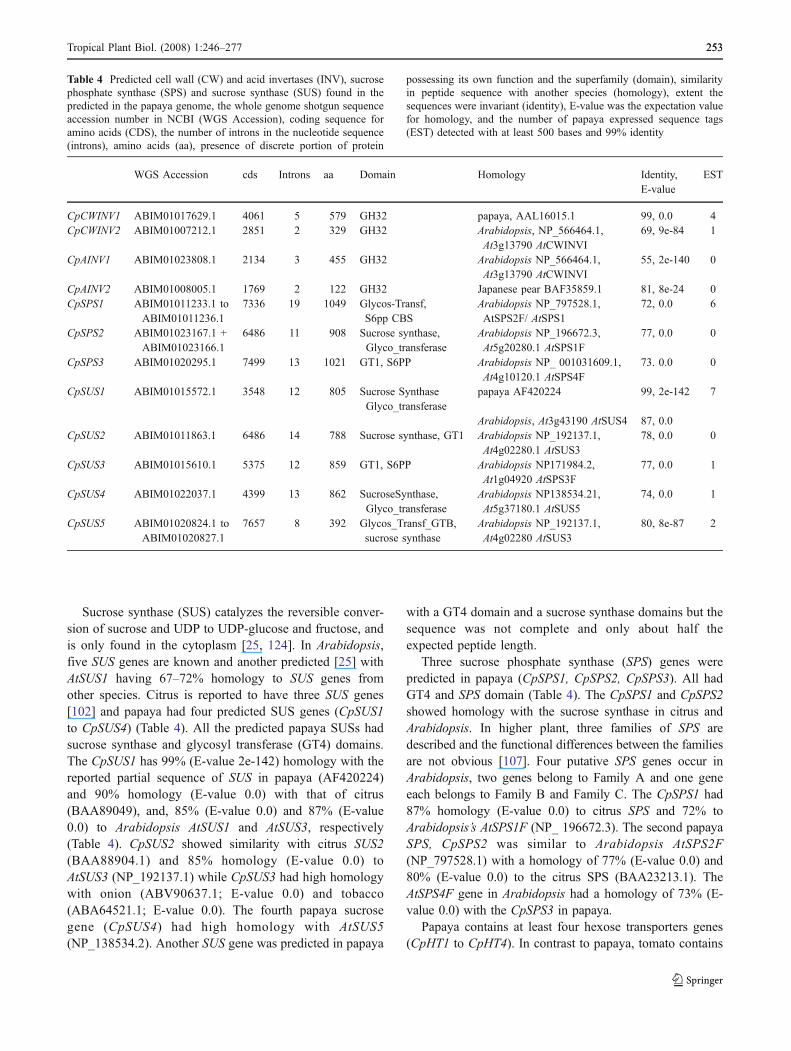
Sucrose synthase (SUS) catalyzes the reversible conver-sion of sucrose and UDP to UDP-glucose and fructose, andis only found in the cytoplasm [25, 124]. In Arabidopsis,five SUS genes are known and another predicted [25] withAtSUS1 having 67–72% homology to SUS genes fromother species. Citrus is reported to have three SUS genes[102] and papaya had four predicted SUS genes (CpSUS1to CpSUS4) (Table 4). All the predicted papaya SUSs hadsucrose synthase and glycosyl transferase (GT4) domains.The CpSUS1 has 99% (E-value 2e-142) homology with thereported partial sequence of SUS in papaya (AF420224)and 90% homology (E-value 0.0) with that of citrus(BAA89049), and, 85% (E-value 0.0) and 87% (E-value0.0) to Arabidopsis AtSUS1 and AtSUS3, respectively(Table 4). CpSUS2 showed similarity with citrus SUS2(BAA88904.1) and 85% homology (E-value 0.0) toAtSUS3 (NP_192137.1) while CpSUS3 had high homologywith onion (ABV90637.1; E-value 0.0) and tobacco(ABA64521.1; E-value 0.0). The fourth papaya sucrosegene (CpSUS4) had high homology with AtSUS5(NP_138534.2). Another SUS gene was predicted in papaya
with a GT4 domain and a sucrose synthase domains but thesequence was not complete and only about half theexpected peptide length.
Three sucrose phosphate synthase (SPS) genes werepredicted in papaya (CpSPS1, CpSPS2, CpSPS3). All hadGT4 and SPS domain (Table 4). The CpSPS1 and CpSPS2showed homology with the sucrose synthase in citrus andArabidopsis. In higher plant, three families of SPS aredescribed and the functional differences between the familiesare not obvious [107]. Four putative SPS genes occur inArabidopsis, two genes belong to Family A and one geneeach belongs to Family B and Family C. The CpSPS1 had87% homology (E-value 0.0) to citrus SPS and 72% toArabidopsis’s AtSPS1F (NP_ 196672.3). The second papayaSPS, CpSPS2 was similar to Arabidopsis AtSPS2F(NP_797528.1) with a homology of 77% (E-value 0.0) and80% (E-value 0.0) to the citrus SPS (BAA23213.1). TheAtSPS4F gene in Arabidopsis had a homology of 73% (E-value 0.0) with the CpSPS3 in papaya.
Papaya contains at least four hexose transporters genes(CpHT1 to CpHT4). In contrast to papaya, tomato contains
Table 4 Predicted cell wall (CW) and acid invertases (INV), sucrosephosphate synthase (SPS) and sucrose synthase (SUS) found in thepredicted in the papaya genome, the whole genome shotgun sequenceaccession number in NCBI (WGS Accession), coding sequence foramino acids (CDS), the number of introns in the nucleotide sequence(introns), amino acids (aa), presence of discrete portion of protein
possessing its own function and the superfamily (domain), similarityin peptide sequence with another species (homology), extent thesequences were invariant (identity), E-value was the expectation valuefor homology, and the number of papaya expressed sequence tags(EST) detected with at least 500 bases and 99% identity
WGS Accession cds Introns aa Domain Homology Identity,E-value
EST
CpCWINV1 ABIM01017629.1 4061 5 579 GH32 papaya, AAL16015.1 99, 0.0 4CpCWINV2 ABIM01007212.1 2851 2 329 GH32 Arabidopsis, NP_566464.1,
At3g13790 AtCWINVI69, 9e-84 1
CpAINV1 ABIM01023808.1 2134 3 455 GH32 Arabidopsis NP_566464.1,At3g13790 AtCWINVI
55, 2e-140 0
CpAINV2 ABIM01008005.1 1769 2 122 GH32 Japanese pear BAF35859.1 81, 8e-24 0CpSPS1 ABIM01011233.1 to
ABIM01011236.17336 19 1049 Glycos-Transf,
S6pp CBSArabidopsis NP_797528.1,AtSPS2F/ AtSPS1
72, 0.0 6
CpSPS2 ABIM01023167.1 +ABIM01023166.1
6486 11 908 Sucrose synthase,Glyco_transferase
Arabidopsis NP_196672.3,At5g20280.1 AtSPS1F
77, 0.0 0
CpSPS3 ABIM01020295.1 7499 13 1021 GT1, S6PP Arabidopsis NP_ 001031609.1,At4g10120.1 AtSPS4F
73. 0.0 0
CpSUS1 ABIM01015572.1 3548 12 805 Sucrose SynthaseGlyco_transferase
papaya AF420224 99, 2e-142 7
Arabidopsis, At3g43190 AtSUS4 87, 0.0CpSUS2 ABIM01011863.1 6486 14 788 Sucrose synthase, GT1 Arabidopsis NP_192137.1,
At4g02280.1 AtSUS378, 0.0 0
CpSUS3 ABIM01015610.1 5375 12 859 GT1, S6PP Arabidopsis NP171984.2,At1g04920 AtSPS3F
77, 0.0 1
CpSUS4 ABIM01022037.1 4399 13 862 SucroseSynthase,Glyco_transferase
Arabidopsis NP138534.21,At5g37180.1 AtSUS5
74, 0.0 1
CpSUS5 ABIM01020824.1 toABIM01020827.1
7657 8 392 Glycos_Transf_GTB,sucrose synthase
Arabidopsis NP_192137.1,At4g02280 AtSUS3
80, 8e-87 2
Tropical Plant Biol. (2008) 1:246–277 253253

three named loci (HT1, HT2, and HT3) and seven putativeloci [61, 79], Arabidopsis contains four named loci (SGB1,TMT1, TMT2, TMT3) and five putative loci [39]. A fifthpapaya hexose transporter was indicated, but the genomicsequence of the ORF was incomplete. Papaya ESTs werefound for two of the predicted hexose transporters CpHT2and CpHT3 (Table 5). The number of introns varied widely,CpHT1, CpHT2, CpHT3, and CpHT4 had two, 12, 11, andone intron, respectively. The predicted peptide for CpHT3had a 60% chance of being localizing to the plastid. Theremaining papaya hexose transporters did not show specificlocalization or export. Papaya contains at least threeincomplete hexose transporter genes or pseudogenes thathad only seven to 11 of the 12 expected transmembranedomains while having domains for MFS and sugartransporter. One of the pseudogenes (ABIM01025390.1)had four ESTs identified. Two of the three pseudogenes hadsecretory sequences for the endoplasmic reticulum.
Respiration
Plant respiration differs from most animal respiration inpossessing a number of additional components including i)the presence of an alternative oxidase to Cytochrome Coxidase that is cyanide insensitive, ii) an internal rotenone-insensitive NAD(P)H non-proton pumping dehydrogenase,and iii) external NAD(P)H non-proton pumping dehydro-genases [64, 118, 206]. Alternate oxidase (AOX) is aterminal quinol oxidase that is non-proton pumping, andtransfers an electron to oxygen while dissipating the energyas heat [128]. AOX is resistant to cyanide [90] and sensitiveto salicylhydroxamic acid (SHAM) [178]. AOX activity istaxonomically widespread and found in all kingdoms [128].In dicots, two types of AOX, AOX1 and AOX2 are found,while monocots seem to posses only AOX1 [54]. AOXs
have been associated with a number of physiologicalfunctions such as thermogenesis in Sacred Lotus duringflowering [210]. Other roles include balancing carbonmetabolism and electron transport, as may happen duringclimacteric fruit ripening [53, 195], control of reactiveoxygen species generation, O2 scavenging and resistance totoxins and pathogenicity [128, 133]. Though AOX1 andAOX2 are present in multigene families in most plants [128]only two genes for alternative oxidase AOX1 was predictedfor papaya, CpAOX1 (ABIM01002926.1), and CpAOX2(ABIM010 10551.1 + ABIM01010553.1). The predictedCpAOX1 protein had a predicted signal peptide of 22 aminoacids, two transmembrane regions, and was supported byfinding papaya ESTs. The two characteristic AOX conservedcysteine residues occurred at Cys 105 and Cys 155. Twointrons similar to Oryza sativa AOX (AB004813) werepresent in the gene. The predicted CpAOX1 gene has 79%(E-value 1e-138) and 71% (E-value 1e-139) identity to thetwo Arabidopsis genes Q9ZRT8 and Q39219, respectively.CpAOX2 similarly has two transmembrane regions. Nosecretory sequence was predicted though EST data suggestedit is expressed. CpAOX2 has 88% identity (E-value 1e-109)to Vigna unguiculata AOX (Q93X12) and 82% (E-value 1e-102) with Arabidopsis (O22049).
Plant mitochondria can oxidize NADH and NADPHwithout proton pumping [80, 130]. These dehydrogenasesare referred to as Type II NAD(P)H [130, 131]. Type IIdehydrogenases operate in parallel to Type I proton-pumping multi-subunit complex I. Both types are found inthe electron transfer chains of several bacteria, and infungal and plant mitochondria. Type II dehydrogenases arefound on the external (NDB) and internal (NDA) faces ofthe inner mitochondrial membrane and transfer electronsfrom NAD(P)H to quinone. It has been proposed that thereare four distinct types of NAD(P)H dehydrogenases: two on
Table 5 Predicted hexose transporters in the papaya genome, thewhole genome shotgun sequence accession number in NCBI (WGSAccession), coding sequence for amino acids (CDS), the number ofintrons in the nucleotide sequence (introns), amino acids (aa), whethera secretory peptide was predicted (secretory) and predicted location(ER endoplasmic retriculum), presence of discrete portion of protein
possessing its own function and the superfamily (domain), similarityin peptide sequence with another species (homology), extent thesequences were invariant (identity), E-value was the expectation valuefor homology, and the number of papaya expressed sequence tags(EST) detected with at least 500 bases and 99% identity
WGS Accession cds Introns aa Secretory Domains Homology E value EST
CpHT1 ABIM01004384.1 2700 2 503 ER Sugar transporter conserved site_2,Sugar transporter conserved site_1,12TMHMM
Q07423, Ricinuscommunis
0 2
CpHT2 ABIM01021724.1,ABIM01021723.1
6879 12 543 ER Sugar transporter conserved site_2,12TMHMM
AAF74569,Arabidopsis thaliana,
0 3
CpHT3 ABIM01019606.1 10663 11 537 Plastid Sugar transporter conserved site_2,MFS, 12TMHMM
AAF74567, Solanumtuberosum
1e-114 1
CpHT4 ABIM01025307.1 1569 1 522 ER Sugar:hydrogen ion symporter,Sugar transporter, 12TMHMM
NP_172592, Arabidopsisthaliana,
0 0
254 Tropical Plant Biol. (2008) 1:246–277
either side of the inner membrane. One to oxidize NADHand the other for NADPH [130, 159]. However, Rasmussonet al., [158] reported two Type II dehydrogenases for potatolocated on the inner and outer surfaces of the innermitochondrial membrane. Only three ORF with similarityto Type II dehydrogenase genes were found in the papayagenome. The ORF for an internal NAD(P)H dehydroge-nase, CpNDA1 (ABIM01007569.1) had 67% identity (E-value 0.0) to Arabidopsis NDA2 (At2g29990, NP_180560.1). A papaya EST was found for CpNDA1. Onegene was also predicted, CpNDB1 (ABIM0101 0022.1 +ABIM01010023.1) that had 79% identity (E-value 0.0) toArabidopsis NDB4 (At2g20800, NP_179673.1). A mito-chondrial transit peptide was predicted for the CpNDB1protein but not for CpNDA1. All had predicted ORFs motifsfor the pyridine-redox superfamily.
Ethylene Synthesis
Arabidopsis and papaya show evolutionary similarity in thenumber of genes associated with ethylene synthesis, receptorsand response pathways. Papaya had four S-adenosyl-L-
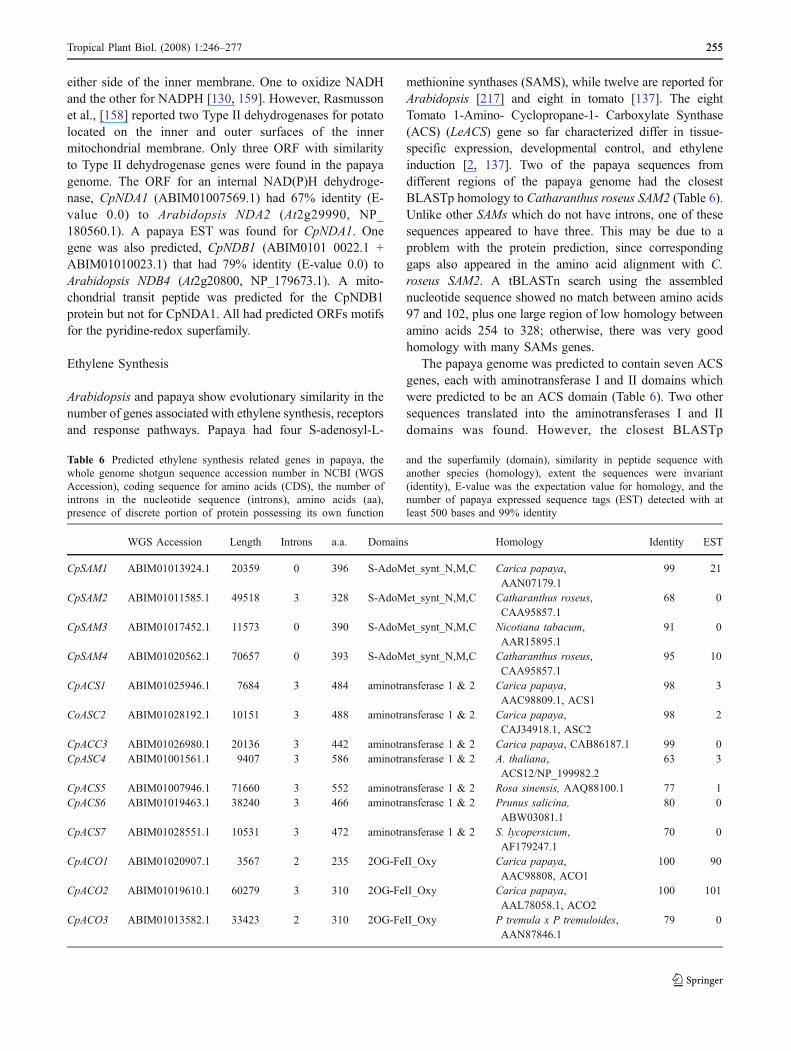
methionine synthases (SAMS), while twelve are reported forArabidopsis [217] and eight in tomato [137]. The eightTomato 1-Amino- Cyclopropane-1- Carboxylate Synthase(ACS) (LeACS) gene so far characterized differ in tissue-specific expression, developmental control, and ethyleneinduction [2, 137]. Two of the papaya sequences fromdifferent regions of the papaya genome had the closestBLASTp homology to Catharanthus roseus SAM2 (Table 6).Unlike other SAMs which do not have introns, one of thesesequences appeared to have three. This may be due to aproblem with the protein prediction, since correspondinggaps also appeared in the amino acid alignment with C.roseus SAM2. A tBLASTn search using the assemblednucleotide sequence showed no match between amino acids97 and 102, plus one large region of low homology betweenamino acids 254 to 328; otherwise, there was very goodhomology with many SAMs genes.
The papaya genome was predicted to contain seven ACSgenes, each with aminotransferase I and II domains whichwere predicted to be an ACS domain (Table 6). Two othersequences translated into the aminotransferases I and IIdomains was found. However, the closest BLASTp
Table 6 Predicted ethylene synthesis related genes in papaya, thewhole genome shotgun sequence accession number in NCBI (WGSAccession), coding sequence for amino acids (CDS), the number ofintrons in the nucleotide sequence (introns), amino acids (aa),presence of discrete portion of protein possessing its own function
and the superfamily (domain), similarity in peptide sequence withanother species (homology), extent the sequences were invariant(identity), E-value was the expectation value for homology, and thenumber of papaya expressed sequence tags (EST) detected with atleast 500 bases and 99% identity
WGS Accession Length Introns a.a. Domains Homology Identity EST
CpSAM1 ABIM01013924.1 20359 0 396 S-AdoMet_synt_N,M,C Carica papaya,AAN07179.1
99 21
CpSAM2 ABIM01011585.1 49518 3 328 S-AdoMet_synt_N,M,C Catharanthus roseus,CAA95857.1
68 0
CpSAM3 ABIM01017452.1 11573 0 390 S-AdoMet_synt_N,M,C Nicotiana tabacum,AAR15895.1
91 0
CpSAM4 ABIM01020562.1 70657 0 393 S-AdoMet_synt_N,M,C Catharanthus roseus,CAA95857.1
95 10
CpACS1 ABIM01025946.1 7684 3 484 aminotransferase 1 & 2 Carica papaya,AAC98809.1, ACS1
98 3
CoASC2 ABIM01028192.1 10151 3 488 aminotransferase 1 & 2 Carica papaya,CAJ34918.1, ASC2
98 2
CpACC3 ABIM01026980.1 20136 3 442 aminotransferase 1 & 2 Carica papaya, CAB86187.1 99 0CpASC4 ABIM01001561.1 9407 3 586 aminotransferase 1 & 2 A. thaliana,
ACS12/NP_199982.263 3
CpACS5 ABIM01007946.1 71660 3 552 aminotransferase 1 & 2 Rosa sinensis, AAQ88100.1 77 1CpACS6 ABIM01019463.1 38240 3 466 aminotransferase 1 & 2 Prunus salicina,
ABW03081.180 0
CpACS7 ABIM01028551.1 10531 3 472 aminotransferase 1 & 2 S. lycopersicum,AF179247.1
70 0
CpACO1 ABIM01020907.1 3567 2 235 2OG-FeII_Oxy Carica papaya,AAC98808, ACO1
100 90
CpACO2 ABIM01019610.1 60279 3 310 2OG-FeII_Oxy Carica papaya,AAL78058.1, ACO2
100 101
CpACO3 ABIM01013582.1 33423 2 310 2OG-FeII_Oxy P tremula x P tremuloides,AAN87846.1
79 0
Tropical Plant Biol. (2008) 1:246–277 255255

matches to these two sequences were not annotated as ACS,and their homology to non-ACS aminotransferases washigh and were not included in the papaya ACS list. Threesequences matched papaya sequences already deposited inGenbank, including CpACS1 and CpACS2 (98% and 98%identity, respectively). All seven putative ACS sequencescontained the conserved dodeca-peptide binding site [219],although, one had an Asn substitution for the key Lys thatbound either PLP or the 2-aminobutyrate of adenosylme-thionine [219]. The possible existence of nonfunctionalsequences with ACS homology in the papaya genomewould not be novel; Arabidopsis has at least one nonfunc-tional gene (AtACS1) shown to encode a protein with noenzyme activity [202] that is nevertheless included in thegene family. In addition, AtACS3 is not even transcribed,and AtACS10 and AtACS12 encode aminotransferasedomains but have no ACS activity; none of these areincluded in the ACS family [217].
A minimum of three ACO genes were predicted inpapaya (Table 6), with six additional sequences that showedpartial homology to ACO-like genes or genes-encoding the2-oxoglutarate FeII oxygenase domain. This domainencompasses more than ACO’s, but it is the motif thatdescribes ACO’s. As with the ACS, papaya has two well-studied CpACO genes, with multiple Genbank representa-tives. The CpACO genes identified (Table 6) reflectBLASTp matches to CpACO1 and CpACO2 from C.papaya, plus a translated supercontig with 79% identity toa poplar protein shown to have ACO activity [7]. The
sequence that matched CpACO1, lacks the first 83 aa fromACO1 (AAC98808), due to sequencing ambiguities, butotherwise matched the remaining 235 aa. All threesequences retained the three conserved subdomains ofACOs and 2-oxoglutarate FeII oxygenases [201], as wellas the HxD site for enzymatic activity. CpACOs share themotif KxxR within the conserved C-terminal site that wasidentified as essential for Petunia hybrida ACO1 enzymeactivity [220]. The protein encoded on (ABIM01013582.1)CpACO3 differs at this site by three amino acids, which ismore than any of the ACOs from 24 different plants,including papaya (LPKEPRFR vs. the consensusQAKEPRFE). Five other predicted models had between42 and 71% homology to Arabidopsis proteins with 2-oxoglutarate FeII oxygenase domains, but these Arabidop-sis proteins are not clearly involved in ACO activity.
Ethylene Response
Papaya ripening has both ethylene insensitive and ethylenesensitive components [47, 122] as has been found fortomato [150]. At least three ethylene receptor genes(CpETR1 to CpETR3) were predicted in the papayagenome. All three encode a protein with the expectedprotein domains, though CpETR1 had no receiver domain(Table 7). CpETR1 was homologous (100%, E-value 0.0)to the papaya receptor cDNA sequence (AF311942.1)deposited by the late Dr. Hamid Lazan and his group(2000) from Malaysia. This sequence has 78% homology
Table 7 Predicted papaya ethylene receptors, structure and phylogenetic relationship with known Arabidopsis ethylene receptors. The whole genomeshotgun sequence accession number in NCBI (WGS Accession), coding sequence for amino acids (CDS), the number of introns in the nucleotidesequence (introns), amino acids (aa), presence of discrete portion of protein possessing its own function and the superfamily (domain), similarity inpeptide sequence with another species (homology), extent the sequences were invariant (identity), E-value was the expectation value for homology,and the number of papaya expressed sequence tags (EST) detected with at least 500 bases and 99% identity
WGSAccession
cds Intron aa Domains Homology Identity,E-value
ESTs
CpETR1 ABIM01020836 25248 4 629 GAF domain, His Kinase A (phosphoacceptor) domain, 3 transmembranedomains
Carica papaya AAG41977 100%, 0.0 9
CpETR2 ABIM01015727 56390 5 738 GAF Domain Clp amino terminal domain, His Kinase A (phosphoacceptor)domain, Response regulator receiver domain, 3 transmembrane domains
Prunus persica ethylenereceptor Q9M7M1 ETR1
88%, 0.0 4
CpETR3 ABIM01030637 5560 9 767 GAF domain, Response regulatory domain, His-kinase domain, 3transmembrane domains
Lycopersicon esculentumethylene receptor homologAF118844_1
67%, 0.0 8
AtERS1
CpETR1
CpETR2
AtETR1
CpETR3
AtEIN4
AtETR2
0.05
Table 7 Predicted papaya ethylene receptors, structure and phyloge-netic relationship with known Arabidopsis ethylene receptors. Thewhole genome shotgun sequence accession number in NCBI (WGSAccession), coding sequence for amino acids (CDS), the number ofintrons in the nucleotide sequence (introns), amino acids (aa), presenceof discrete portion of protein possessing its own function and the
superfamily (domain), similarity in peptide sequence with anotherspecies (homology), extent the sequences were invariant (identity), E-value was the expectation value for homology, and the number ofpapaya expressed sequence tags (EST) detected with at least 500 basesand 99% identity
256 Tropical Plant Biol. (2008) 1:246–277

(E-value 0.0) with the Arabidopsis ERS1 (At2g40940,NP_181626.1) (Table 7). Arabidopsis ETR1 (At1g66340,NP_176808.3) has the greatest homology with CpETR2(86%, E-value 0.0). Based upon gene and protein structure,both CpETR1 and CpETR2 fall into the ethylene receptorsubfamily 1 [94] with CpETR3 being in subfamily 2.Subfamily 1 receptors maybe required for most ethyleneresponses [208]. Originally, we reported four two-compo-nent ethylene receptors [132], however, a more thoroughanalysis indicated that one of the previously reportedCpETRs lacked the expected GAF domain and a fullhistidine domain [88, 208]. This partial sequences(ABIM01010980) was 139 aa long peptide with 75%homology (E-value 3e-75) to apple ethylene receptor. Afifth potential gene (ABIM01025304.1) was short (224 aa).It only had the DUF623 domain of unknown function andlow homology to the nearest ethylene receptor(AAQ56281) of Litchi chinensis. Papaya’s three predictedethylene receptors were fewer than the five ETRs found inArabidopsis [176] and the six in tomato [100]. Sinceethylene receptors seem to be a negative regulator of action[93], degradation plays a significant role in the control ofripening [99]. The relatively small complement of papayaethylene receptors means that fewer interactions may occurbetween receptors and that their role in ripening should bemore easily discerned.
Ethylene receptors are disulfide linked dimers and ethylenebinding involves a copper cofactor [164]. The gene RAN(Response to Antagonist) plays a role in making the ethylenereceptor apoprotein functional by transporting a copper ion tothe ethylene receptor site in the membrane spanning regions.As with Arabidopsis, only one RAN homolog was found inpapaya (Table 8), CpRAN had 76% homology (E-value 0.0)to Arabidopsis RAN1 (NP_ 200892.1).
The ethylene response pathway following ethylenebinding involves a RAF-related kinase CTR1 that has arole in the negative response by forming a complex with thereceptor [93]. Ethylene binding inhibits CTR1 kinaseactivity [141] and thus relieves the repression of theethylene response pathway [5, 6]. A signal is thentransmitted from the positive regulator EIN2 to EIN3/EILsand induces transcription of ethylene response factors(ERF). AtCTR1 is one of six Arabidopsis MAPK kinasesand three LeCTR1-like genes in tomato [1, 78] with noevidence that more than one (AtCTR1, LeCTR1) beinginvolved in the ethylene signal transduction pathway. Atleast one CpCTR1 gene was found in the papaya genome(ABIM01019395 + ABIM01040871) that had 56% homol-ogy with LeCTR1 (E-value 0.0). A second, but incompleteCTR sequence, was also found (ABIM01001640) (Table 8).One possible CpEIN2 gene was found, although thenucleotide and amino acid sequence were short (Table 8),
Table 8 Ethylene response genes predicted in the papaya genomeThe whole genome shotgun sequence accession number in NCBI(WGS Accession), coding sequence for amino acids (CDS), thenumber of introns in the nucleotide sequence (introns), amino acids(aa), presence of discrete portion of protein possessing its own
function and the superfamily (domain), similarity in peptide sequencewith another species (homology), extent the sequences were invariant(identity), E-value was the expectation value for homology, and thenumber of papaya expressed sequence tags (EST) detected with atleast 500 bases and 99% identity
WGS Accession cds Introns aa Domains Homology Identity,E-value
EST
CpRAN1 ABIM01008271 9019 8 ATPase, coupled totransmembrane movementof ions, phosphorylativemechanism Heavy-metal-associateddomain
NP_199292.1 RAN1Arabidopsis thaliana
76%, 0.0 4
CpCTR1 ABIM01019395 17660 13 793 CTR1-like protein tyrosine kinase AAR89821 Lycopersiconesculentum
56%, 0.0 4
ABIM01040871 1813CpCTR2 ABIM01001640 44125 7 1022 Protein tyrosine kinase ABE80154 Medicago
truncatula88%, 2e-114 0
CpEIN1 ABIM01024839 14603 7 1294 EIN2 AAR08678Petunia x hybrida
86%, 1e-22 2
CpEIN2 ABIM01006170 34454 1 601 EIL2 Ethylene insensitive 3 ABK35086 Prunus persica 61%, 1e-87 0CpEIN2 ABIM01011208 9335 1 665 EIN3-binding F-box protein 2 ABC24972 Lycopersicon
esculentum57%, 0.0 7
CpEIL1 ABIM01017539 9817 0 603 EIL4, Ethylene insensitive 3 AAP04000Nicotiana tabacum
70%, 0.0 10
CpEIL2 ABIM01012325 17077 1 601 EIL2, Ethylene insensitive 3 ABK35086 Prunus persica 70%, 0.0 2
Tropical Plant Biol. (2008) 1:246–277 257257

the alignments with CTR1-like kinase and the proteinkinase family had fewer gaps and about one hundred aminoacid matches. Four possible EIN3/EIL1 genes were alsofound, with two (CpEIL1, CpEIN3) having high EIN2/EILhomologies to published sequences and these wereexpressed as papaya ESTs. Arabidopsis has nine EIN3and EIN3-like genes [28] and tomato has at least five [186].
Eight ERF genes were found with high homlogy andanother twelve predicted ERF-like sequences that had lowhomology matches or had no significant protein domains(Supplementary Table 2). One hundred and twenty-twoethylene responsive binding factors in the AP2/ERFsuperfamily have been found in Arabidopsis [135].CpERF1 falls into group IX that has 17 members inArabidopsis with eight in group VIII. Using the conserved60 amino acid sequences for AP2/ERF [135], an additional92 potential genes with this sequence were found in papayathat suggested more ERF maybe present in the papayagenome (data not shown).
Isothiocyanates
The glucosinolates are found almost exclusively in theorder Brassicales [161] and have a sulfonated oxime with aβ-thioglucose residue. Papaya synthesizes high levels ofbenzylglucosinolates (BG) and benzylisothiocyanates(BITC) with BG being the only glucosinolate found inpapaya [26]. BITC is produced by the removal of theglucose moiety by the action of a β-thioglucosidase(myrosinase). Myrosinase is separated from the isothiocya-nate, in specialized myrosin cells where myrosinase isbrought into contact with its substrate BG followingphysical damage. Myrosinase expression and activityincreases are fourfold at harvest, then decline slightlyduring ripening [169]. Both BG and BITC levels arehighest in the seeds, followed by the peel and pulp, anddecrease during fruit development [119, 169, 193].
Genes were predicted for all steps except for the firsthydroxylase in the biosynthesis of BG. In Arabidopsis, theP450 monooxygenase CYP79A2 is involved in the conver-sion of phenylalanine to the aldoximine [83, 101, 211]. Nopapaya gene with homology to CYP79A2 was found. WhenArabidopsis CYP79A2 was used to search the papayadatabase, homology was only found to two closely relatedP450s, CYP79B2 and CYP79B3 (Table 9). CYP79B2 andCYP79B3 both act on tryptophan and may be involved inauxin synthesis [101]. It is possible that the papayaCYP79B2 and CYP79B3 homologs have broader substratespecificity as found for some other P450 monooxygenases[83], and that can convert in papaya phenylalanine to thealdoximine. A single P450 gene (CpCYP83B1) waspredicted in papaya for the reduction and cysteine additionstep with 70% homology (E-value 0.0) to Arabidopsis
CYP83B1 gene. After these two reduction steps, twopredicted aminotransferases C-S lyases (CpCSL1, CpCSL2)were found with high homology to the Arabidopsis homologand Brassica napus SUR1. A single glucosyl transferasegene was found in papaya with high homology (55%, E-value 2e-138) to Brassica napus UGT74B1 [84]. The nextstep, which involves sulfate transfer, had two predictedgenes (CpPAPS1, CpPAPS2), though CpPAPS2 had only apartial sulfotransferase domain characteristic of this genefamily [152].
Two myrosinase genes have been reported in Arabidop-sis [157, 215] though only one (CpMYR) was predicted forpapaya, which is in agreement with a recent report [169].The predicted CpMYR had 98% homology (E-value 3e-146) to the published papaya myrosinase sequence with aglucosyl hydrolase family 1 (GH1) domain. Only onemyrosinase binding protein was found with low homology(37%, E-value 4e-20) to the Arabidopsis MBP2. A singlemyrosinase-associated protein was also predicted in papaya,as for Brassica napus [157].
Laticifers and Protease
The papaya fruit, as do the other aerial parts of this plant, has adense network of laticifers [170]. The articulated laticifersbegin as a column of cells that form vessel-like structuresthat retain the usual organelles. The milky latex is about 85%water with the 15% being composed of 25% insoluble matterof unknown composition. The soluble fraction containscarbohydrates (~10%), salts (~10%), lipids (~5%), andbiomolecules mainly proteins (~40%) [65]. The biomole-cules include cysteine proteinases (papain, chymopapain),cystatin, β-1,3-glucanase, chitinase, lipases and other pro-teins. The latex is thought to function as an induced defensemechanism [172, 182]. The latex is harvested from largefruit green varieties by scratching the skin, allowing theexuded latex to coagulate and dry, then the dried latex iscollected by scrapping. The latex is purified and itsproteolytic activity is used for meat tenderization and chill-proofing of beer [65]. During fruit ripening proteinaseactivity declines [147] and some of that laticifers break andrelease latex under the cuticle disrupting this barrier andincreasing fruit water loss rate [148].
Four cysteine proteinases account for 80% of theenzyme fraction [65]. The proteinases are papain, chymo-papain, caricain (proteinase omega) and glycyl endopepti-dase (proteinase IV). Papaya proteinases are synthesized asproezymes with a signal sequence. The prosequence iscleaved and proteinase activated. All four proteinases aremembers of the peptidase C1A subfamily of cysteineproteinases. Papain has been most intensively studied [65]though a minor component (5–8%) of the endopeptidases inpapaya latex [18, 21]. One gene for a propapain (CpPAPA)
258 Tropical Plant Biol. (2008) 1:246–277

Tab
le9
Predicted
papaya
glycosinalates
synthesisgenesandamyrosinasegene.N
oho
molog
toCYP79
A2,
thefirststepin
biosyn
thesis,w
asfoun
din
papaya.T
hree
closelyrelatedCYP79
genes
werepredicted.
The
who
legeno
meshotgu
nsequ
ence
accessionnu
mberin
NCBI(W
GSAccession
),coding
sequ
ence
foram
inoacids(CDS),thenu
mberof
intron
sin
thenu
cleotid
esequ
ence
(introns),am
inoacids(aa),p
resenceof
discretepo
rtionof
proteinpo
ssessing
itsow
nfunctio
nandthesuperfam
ily(dom
ain),sim
ilarity
inpeptidesequ
ence
with
anotherspecies(hom
olog
y),extent
thesequ
enceswereinvariant(identity
),E-value
was
theexpectationvalueforho
molog
y,andthenu
mberof
papaya
expressedsequ
ence
tags
(EST)detected
with
atleast50
0basesand99
%identity
WGSAccession
cds
Intron
saa
Dom
ains
Hom
olog
yIdentity,
E-value
EST
CYP79
ABIM
0102
3062
.117
811
558
cytochromeP45
0,family
79,subfam
ilyB,
polypeptide2;
oxyg
enbind
ing
Arabido
psisthaliana
CYP79
B2,
NP_1
9570
5.1
60,0.0
0
ABIM
0101
9933
.120
501
534
cytochromeP45
0,family
79,subfam
ilyB,
polypeptide3;
oxyg
enbind
ing
Arabido
psisthaliana
CYP79
B3,
NP_1
7982
0.2
58,.0.0
0
ABIM
0102
3066
.117
721
559
cytochromeP45
0,family
79,subfam
ilyB,
polypeptide3,
oxyg
enbind
ing
Arabido
psisthaliana
CYP79
B3,
NP_1
7982
0.2
0
CpC
YP83
B1
ABIM
0100
7450
.116
452
477
P45
0,83
B1
Arabido
psisthaliana
CYP83
B1,
NP_1
9487
8.1
70,0.0
3
C-S
Lyase
ABIM
0101
9595
.142
166
421
Aspartate/ty
rosine/aromatic
aminotransferase
Arabido
psisthaliana
,BAB10
727.1
80,0.0
2CpC
SL1
ABIM
0101
9594
.1CpC
SL2
ABIM
0101
5665
.118
866
420
Aspartate/ty
rosine/aromatic
aminotransferase
Brassicarapa
subsp.
pekinensis
SUR1,
ACH41
754.1
61,4e-144
2
Thioh
ydroximate
S-glycosyltransferaseUGT74
B1
ABIM
0100
2346
.114
751
453
thiohy
drox
imateS-glucosyltransferase
Glycosyltransferasefamily
28N-terminal
domain.
Brassicana
pus,AAL09
350.1
55,2e-138
0
CpPAPS1
ABIM
0102
3999
.110
761
344
sulfotransferase_1
superfam
ilyArabido
psisthaliana
ST2a,
NP_5
6817
7.1
60,6e-126
1
CpPAPS2
ABIM
0100
1554
.120
004
126
Partialsulfotransferase_1
superfam
ily.
Brassicarapa
ST5a,ACH41
751.1
781e-39
0MyrosinaseCpM
YR
ABIM
0100
5474
.136
6313
492
Glucohy
drolaseGH1
Caricapa
paya
thioglucoside
glucoh
ydrolase,ACC95
418.1
98,3e-146
1
MYRBinding
Protein
ABIM
0101
9550
.110
861
191
Jacalin
-likelectin
domain.
Proteins
containing
thisdo
mainarelectins.
Arabido
psisthaliana
MBP2
(Myrosinasebind
ingprotein2),
NP_1
7561
5.1
37,4e-20
MYRAssociatedProtein
ABIM
0100
7310
.113
884
343
SGNH_p
lant_lipase_lik
e,,aplantspecific
subfam
ilyof
theSGNH-fam
ilyof
hydrolases
Arabido
psisthaliana
epith
iospecifier
mod
ifier,ABB90
255.1
52,3e-100
12
Tropical Plant Biol. (2008) 1:246–277 259259

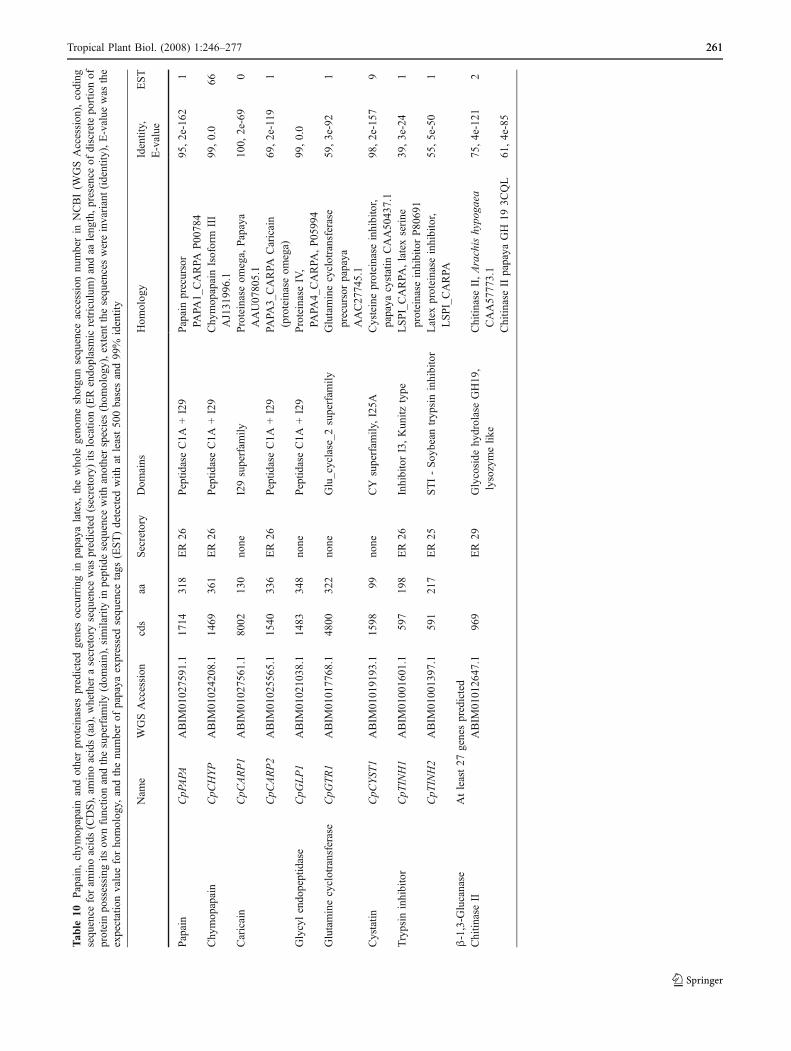
precursor was predicted (ABIM02027591.1). The encodedprotein has a 26 aa ER secretory sequence and 95% identity(E-Value 2e-162) to the published papaya cDNA sequence(P00784) (Table 10). A papaya ESTwas found for CpPAPA.
A single papaya chymopapain gene (CpCHYP) waspredicted that had 99% identity (E-value 0.0) to chymopa-pain isoforms I, III, and V (X97789, AJ131996, andAJ131998 respectively), and 100% to chymopapain iso-form II and IV (AJ0131995 and AJ1319970) (Table 10).Chymopapain is distinguished from papain by the proteo-lytic activity remaining after papain is removed [95]. Fiveprochymopapain isoforms (I to V) were identified fromsequenced leaf cDNAs [194], all the isoforms have a freecysteine at position 251. The translated isoform cDNAsdiffer in one or two amino acid substitutions. At position222, a cysteine is replaced by a tyrosine in isoforms III andV, while at position 266, a valine is replaced by aphenylalanine in isoforms II, III, and V. These single aminoacid substitutions require only a single base change. Thepredicted CpCHYP protein had the nine amino acidsreported for isoforms III, IV and V. It is possible that thechymopapain isoforms differ because of errors in PCR,sequencing or translation. The CpCHY protein, possiblyisoform III, had 66 ESTs. A chymopapain isoform III ESTwas also reported by Devitt et al. [60].
A number of other proteinases have been reported inpapaya latex [16, 65, 182]. Two caricain genes werepredicted (proteinase omega) and one glycyl endopeptidasegene (Table 10). The predicted gene numbers are in agree-ment with those reported [65], though one of the caricaingene cDNA has not been deposited in Genbank. TheCpCARP1 was 100% identical (E-value 2e-69) to the de-scribed sequence, although it is only 130 aa long due toincomplete sequencing of the WGS. A glutamine cyclo-transferase (glutamine cyclase) gene was predicted in papaya(CpGTR) as previously reported from papaya latex [65, 140].The predicted papaya gene (Table 10) was longer than thepredicted 288 aa peptide from the cDNA (AF061240) inGenBank though analysis suggested intron boundaries mayhave not been accurately predicted.
One cystatin (CpCYST) and two kunitz-type trypsininhibitors (CpTINH1, CpTINH2) were predicted in thepapaya genome in agreement with the published data [17].The cystatin encoded the 125A domain and was classifiedas a member of the CY superfamily. Papaya ESTs werefound for both the cystatin and trypsin inhibitors (Table 10).
At least 27 β-1,3-glucanase proteins (GH17) werepredicted in papaya and activity has been detected inpapaya latex [65]. The papaya carbohydrate active enzymeswill be the focus of a paper in preparation. In addition to β-1,3-glucanase, chitinase II (GH19) has been reported forpapaya latex. A gene with 61% identity to the partial cDNA(P81241) was found in the papaya genome (Table 10).
Other than the genes encoding proteinases and thoselaticifer proteins described above, the genes encoding otherproteins potentially found in papaya latex have not beenwell described. A search of the papaya genome for latexassociated protein genes found six genes including one forcaspase reported as a rubber latex abundant protein andlysophospholipase (Supplementary Table 3). In addition, anumber of potential latex genes were found whosepredicted protein had allergen domains.
Lignin Synthesis
Lignins are amorphous heteropolymers resulting from theoxidative coupling of p-hydroxycinnamyl alcohols, p-coumaryl, coniferyl, and sinapyl alcohols by both laccasesand plant peroxidases in the cell wall. This lignificationprocess is crucial to xylem formation as it confers resistanceagainst the tensile forces in the xylem, imparts waterimpermeability, and protection against chemical and bio-logical degradation [51, 63, 126, 143].
The papaya genome had three predicted phenylalanineammonia lyase (PAL) lignin biosynthesis genes, comparedto the four Arabidopsis genes and two in Populus [89, 156].CpPAL1 and CpPAL2 genomic sequences had two predictedexons, whereas CpPAL3 had only one exon. The PAL genesin Arabidopsis have from one to three exons. Papaya PALgene amino acid sequences were ~85% homologous to theArabidopsis sequences and about 88% homologous to thePopulus PAL sequences (Supplementary Table 4). CpPAL1and CpPAL2 both contained the PAL and PAL-HALconserved domains found in Arabidopsis and PopulusPAL genes, but CpPAL3 had only a partial sequence ofthe conserved domain. It was possible that the second exonof CpPAL3, actually has the remainder of the conserveddomain, but it was located on a separate scaffold.
Two candidate trans-cinnamate 4-hydroxylase (C4H)genes were identified in papaya (Supplementary Table 4)while two genes are identified in Populus and one inArabidopsis [89, 156]. As in the Arabidopsis C4H gene,CpC4H1 consists of three predicted exons, while CpC4H2had two. Papaya’s class I C4H gene, CpC4H1, was verysimilar to Arabidopsis and Populus class I C4H genes, withan amino acid sequence homology of 86% and 93%,respectively. CpC4H2, a class II C4H gene, shared signifi-cant homology (83%) to the Populus class II C4H gene.Both papaya C4H genes contained the cytochrome p450conserved domain.
Similar to Arabidopsis, papaya had four candidate 4-coumarate: CoA ligase (4CL) genes, whereas Populus hasone [63, 89, 156]. Cp4CL1, Cp4CL2, and Cp4CL3, allbelong to the class I 4CL genes. The class I papaya 4CLgenes had five predicted exons, similar to the four and fiveexons in the class I 4CL genes in Arabidopsis. The class II
260 Tropical Plant Biol. (2008) 1:246–277
Tab
le10
Papain,
chym
opapainandotherproteinasespredictedgenesoccurringin
papaya
latex,
thewho
legeno
meshotgu
nsequ
ence
accessionnu
mberin
NCBI(W
GSAccession
),coding
sequ
ence
foram
inoacids(CDS),am
inoacids(aa),whether
asecretorysequ
ence
was
predicted(secretory)its
locatio
n(ERendo
plasmic
retriculum
)andaa
leng
th,presence
ofdiscrete
portionof
proteinpo
ssessing
itsow
nfunctio
nandthesuperfam
ily(dom
ain),similarity
inpeptidesequ
ence
with
anotherspecies(hom
olog
y),extent
thesequ
enceswereinvariant(identity
),E-value
was
the
expectationvalueforho
molog
y,andthenu
mberof
papaya
expressedsequ
ence
tags
(EST)detected
with
atleast50
0basesand99
%identity
Nam
eWGSAccession
cds
aaSecretory
Dom
ains
Hom
olog
yIdentity,
E-value
EST
Papain
CpPAPA
ABIM
0102
7591
.117
1431
8ER26
Peptid
aseC1A
+I29
Papainprecursor
PAPA
1_CARPA
P00
784
95,2e-162
1
Chy
mop
apain
CpC
HYP
ABIM
0102
4208
.114
6936
1ER26
Peptid
aseC1A
+I29
Chy
mop
apainIsoform
III
AJ131
996.1
99,0.0
66
Caricain
CpC
ARP1
ABIM
0102
7561
.180
0213
0no
neI29superfam
ilyProteinaseom
ega,
Papaya
AAU07
805.1
100,
2e-69
0
CpC
ARP2
ABIM
0102
5565
.115
4033
6ER26
Peptid
aseC1A
+I29
PAPA
3_CARPA
Caricain
(proteinaseom
ega)
69,2e-119
1
Glycylendo
peptidase
CpG
LP1
ABIM
0102
1038
.114
8334
8no
nePeptid
aseC1A
+I29
ProteinaseIV,
PAPA
4_CARPA
,P05
994
99,0.0
Glutaminecyclotransferase
CpG
TR1
ABIM
0101
7768
.148
0032
2no
neGlu_cyclase_2
superfam
ilyGlutaminecyclotransferase
precursorpapaya
AAC27
745.1
59,3e-92
1
Cystatin
CpC
YST
1ABIM
0101
9193
.115
9899
none
CY
superfam
ily,I25A
Cysteineproteinase
inhibitor,
papaya
cystatin
CAA50
437.1
98,2e-157
9
Trypsin
inhibitor
CpT
INH1
ABIM
0100
1601
.159
719
8ER26
InhibitorI3,Kun
itztype
LSPI_CARPA
,latexserine
proteinase
inhibitorP80
691
39,3e-24
1
CpT
INH2
ABIM
0100
1397
.159
121
7ER25
STI-Soy
bean
tryp
sininhibitor
Latex
proteinase
inhibitor,
LSPI_CARPA
55,5e-50
1
β-1,3-G
lucanase
Atleast27
genespredicted
Chitin
aseII
ABIM
0101
2647
.196
9ER29
Glycoside
hydrolaseGH19
,lysozymelik
eChitin
aseII,Arachishypo
gaea
CAA57
773.1
75,4e-121
2
Chitin
aseIIpapaya
GH
193C
QL
61,4e-85
Tropical Plant Biol. (2008) 1:246–277 261261

papaya 4CL gene had six predicted exons, while theArabidopsis class II 4CL gene have seven exons. The fourpredicted papaya Cp4CL genes encoded proteins with 65–75% amino acid homology to the class I 4CL genes inArabidopsis and 68–78% in Populus. Cp4CL4 belongs tothe class II 4CL genes although it shared less than 50%homology with the class II genes found in Arabidopsis. Allpapaya 4CL genes contained the acetyl-CoA synthaseconserved domain as found in the Arabidopsis and Populus4CL genes. Though Cp4CL4 had low homology withArabidopsis and Populus 4CL genes, it still contained theexpected conserved domain. Further studies are needed toascertain the role of this gene.
Three hydroxycinnamoyl-CoA shikimate/quinatehydroxycinnamoyltransferase (HCT) candidate genes werefound in papaya (Supplementary Table 4), whereas onlyone HCT gene is present in Arabidopsis and Populus [89,156]. CpHCT1 contained two exons, as does the Arabi-dopsis HCT gene, but CpHCT2 has ten predicted exons andCpHCT3 had five. The CpHCT1 predicted protein had 83%homology to both Arabidopsis and Populus HCT aminoacid sequences. CpHCT2 and CpHCT3 had considerablyless homology, below 50%. CpHCT1 and CpHCT2contained a complete transferase conserved domain, butCpHCT3 only has a partial conserved domain. The lowsequence similarity and broken conserved domain sug-gested CpHCT2 and CpHCT3 may have functions otherthan those of bonafide HCT genes.
Expression data showed only one P-coumaroyl shiki-mate 3′-hydroxylase/coumaroyl 3-hydroxylase (C3H) genepresent in papaya, which is fewer than the two C3H genesreported by Ming et al. [132]. Arabidopsis and Populus alsoonly have one C3H gene [63, 89, 156]. CpC3H1 encoded aprotein with 83% homology to the amino acid sequence ofthe Arabidopsis C3H gene and an 89% homology to thePopulus C3H gene. CpC3H1, as with the Arabidopsis C3Hgene, had three predicted exons and also contained thecomplete cytochrome p450 conserved domain.
The papaya genome was predicted to have two trans-caffeoyl-CoA 3-O-methyltransferase (CCOMT) genes, asdoes Populus, but Arabidopsis only contains one [63, 89,132, 156]. CpCCOMT1 was predicted to have five exonsand CpCCOMT2 had one; Arabidopsis CCOMT genecontains four exons. The CpCCOMT1 predicted amino acidsequence only matched about 60% to the amino acid se-quences of CCOMT genes in Arabidopsis and Populus.CpCCOMT2 amino acid sequence had a much higher ho-mology, greater than 90% to both Arabidopsis and Populusamino acid sequences. Both papaya CCOMT genes had theS-adenosylmethionine-dependent-methyltransferase con-served domain, though CpCCOMT2 was a partial domain.
Whereas Arabidopsis has two cinnamoyl-CoA reductase(CCR) genes, only one CCR gene was present in papaya
and Populus [63, 89, 132, 156]. Three exons were predictedfor CpCCR1, but the two Arabidopsis CCR genes have fourand five exons. The CpCCR1 predicted protein had about a75% to 80% homology to the Arabidopsis CCR amino acidsequences and 87% homology to the Populus CCR aminoacid sequence. CpCCR1 contained a partial AdoHcyase, S-adenosyl-L-homocysteine hydrolase conserved domain asalso found in the Arabidopsis and Populus CCR genes.
Four candidate coniferylaldehyde 5-hydroxylase(CA5H), also called ferulate 5-hydroxylase (F5H), geneswere identified in papaya [132]. Populus has two CA5Hgenes and Arabidopsis has one [63, 89, 156]. CpCA5H1and CpCA5H2 both had two exons, CpCA5H3 had three,and CpCA5H4 had seven predicted exons. CpCA5H1 andCpCA5H2 had the closest homology to Arabidopsis andPopulus CA5H amino acid sequences, with 79% and 78%homology, respectively. CpCA5H3 and CpCA5H4 pre-dicted protein amino acid sequences were not as similar,with about 70% similarity to Arabidopsis and PopulusCA5H amino acid sequences. All of the papaya CA5Hgenes contained a complete cytochrome p450 conserveddomain (Supplementary Table 4).
Papaya and Arabidopsis genomes both contained onecaffeic acid 3-O-methyltransferase (COMT) gene while thePopulus genome contains two [89, 132, 156]. CpCOMT1contained five exons, compared to four exons in theArabidopsis COMT gene. The CpCOMT1 predicted aminoacid sequence was about 76% homologous to that of bothArabidopsis and Populus. The BLASTp results showed agap of 16 amino acids in the papaya sequence after aminoacid 264, compared to both the Arabidopsis and Populussequences. CpCOMT1 contained a complete S-adenosyl-methionine-dependent methyltransferase conserved do-main, as well as a dimerisation domain, as do theArabidopsis and Populus COMT genes.
Two candidate cinnamyl alcohol dehydrogenase (CAD)genes were identified in papaya, the same number as foundin Arabidopsis, with Populus having one CAD [63, 89,156]. CpCAD1 had six and CpCAD2 had seven predictedexons. CpCAD1 predicted protein has the highest aminoacid sequence homology to Arabidopsis and Populus CADproteins, with 78% and 81% similarity, respectively.CpCAD2 homology to Arabidopsis and Populus CADamino acid sequences was lower. The Arabidopsis CADgenes have five exons. Both CpCAD1 and CpCAD2 containthe alcohol dehydrogenase GroES-like conserved domainand the zinc-binding dehydrogenase conserved domain, asseen in Arabidopsis and Populus CAD genes.
Polyphenoloxidases and Peroxidases
Flavonoid oxidation occurs widely in plants and linked toprotection against UV damage, a deterrent for herbivores,
262 Tropical Plant Biol. (2008) 1:246–277

pathogenicity, scavengers of free radicals, and lignindegradation [51, 55, 109, 125, 126, 153, 196, 197]. Theoxidation, especially of o-diphenols in the presence ofmolecular oxygen, leads to the highly reactive semiqui-nones and quinones that interact with phenols, amino acidand proteins yielding brown products. The brown productsoccur in seed coats, and in other tissues followingenvironmental stress and physical injury, and are a majorproblem during food processing. Polyphenol oxidases(PPO) and peroxidases (POD) are the two major groupsof enzymes involved in oxidation. PPOs are divided intotyrosinase and laccase, with the tyrosinases includingmonophenolase and cathechol oxidase that oxidizes o-diphenols. Laccase oxidizes o-diphenol and p-diphenols.Cathechol oxidase is localized in the chloroplast whilelaccases and peroxidases are secreted [51, 126, 153].
In papaya, browning reactions occur during seed coatmaturation, in the skin following chilling injury andfreezing, and occasionally on the cut mesocarp surfacefollowing minimal processing. Phenolic compounds inpapaya skin decline during ripening to one quarter of thatat the mature green stage [192]. Chilling injury of colorbreak fruit is associated with the development of a darkolive skin scald while in riper fruit a lighter browncondition occurs [46] possibly due to lower amount ofphenol substrate in riper fruit. During papaya processinginto juice [44] and with frozen chunks [40, 41], colorchanges have been reported with increases in PPO andPOD activities.
Nine laccase genes were predicted in the papaya genome(Table 11). The papaya laccases (CpLAC1 to CpLAC9) allhave the three laccase Cu-oxidase domains and werepredicted to be secreted. All but four of the predictedlaccase genes had papaya ESTs and greater than 66% to79% identity with known laccase sequences. CpLAC2 andCpLAC3 were found on the same WGS sequence and were91% identical, and may be either a recent duplication or asequencing error. CpLAC9 was predicted to encode aprotein of 431 aa versus 545–593 aa for CpLAC1 toCpLAC8. CpLAC9 maybe only a partial sequence, since ithad only two and not three Cu-oxidase domains normallyfound in laccase genes [51, 126, 127].
The brown reactions reported during chilling injury andprocessing is probably due to PPO activity [41] and at leastfive PPO genes (CpPPO1 to CpPPO5) were predicted(Table 11). The activity of PPO declines slightly duringripening [41] and six isoforms were reported for hermaph-rodite fruit with only four isoforms found for female papayafruit. The difference between the number of isoforms [40]and the predicted gene number may reflect alternativesplicing, post-translation processing or one of the partialPPO-like sequences may be a real gene (data notpresented). All CpPPOs predicted proteins had predictedor potential chloroplast transit peptides of 43–46 aa. Intomato, PPOs are encoded by a seven-member PPOmultigene family on chromosome 8 [196]. The seventomato PPOs show differential expression during develop-ment and following stress [197]. Papaya PPOs were also
Table 11 Polyphenol oxidase (tyrosinase, dicopper containing) and laccase (multi copper oxidase) genes predicted in the papaya genome, thewhole genome shotgun sequence accession number in NCBI (WGS Accession), coding sequence for amino acids (CDS), the number of introns inthe nucleotide sequence (introns), amino acids (aa), whether a secretory sequence was predicted (secretory) its location (ER endoplasmicretriculum) and aa length, similarity in peptide sequence with another species (homology), extent the sequences were invariant (identity), E-valuewas the expectation value for homology, and the number of papaya expressed sequence tags (EST) detected of at least 500 bases and 99% identity
WGS Accession CDS aa Secretory Homology Identity,E-value EST
Laccase Cu-oxidase SuperfamilyCpLAC1 ABIM01016852.1 2124 545 None LAC6 Arabidopsis NP182180.1 70, 0.0 0CpLAC2 ABIM01015850.1 2224 547 ER, 23 Diphenol oxidase Tobacco AAC49536.1 77, 0.0 2CpLAC3 ABIM010158201 2473 558 ER, 23 Diphenol oxidase Tobacco AAC49536.1 78, 0.0 1CpLAC4 ABIM01015697.1 2741 557 ER, 22 Diphenol oxidase Tobacco AAC49536.1 78, 0.0 0CpLAC5 ABIM01007467.1 2162 562 ER LAC11 Arabidopsis NP_195946.2 78, 0.0 1CpLAC6 ABIM01005823.1 2483 579 Poss ER, 31 LAC5 Arabidopsis NP_181568.1 79, 0.0 1CpLAC7 ABIM01012580.1 2829 566 ER, 23 Tomato laccase ACB10229.1 69, 0.0 0CpLAC8 ABIM01015449.1 2376 593 ER, 22 Liriodendron tulipifera laccase AAB17194.1 66, 0.0 0CpLAC9 ABIM01001394.1 2361 431 ER, 32 LAC13 Arabidopsis NP_196330.3, Only 2 Cu oxidase motifs 75, 2e-150 1Tyrosinase SuperfamilyCpPPO1 ABIM01007553.1 1809 602 chloro, 43 Apple polyphenol oxidase BAA21676.1 57, 2e-172 0CpPPO2 ABIM01002511.1 1848 615 chloro, 46 Poplar polyphenol oxidase AAU12257.1 61, 0.0 0CpPPO3 ABIM01025488.1 1794 597 chloro, 43 Poplar polyphenol oxidase AAK53414.1 56, 0.0 1CpPPO4 ABIM01013357.1 1459 365 Poss Plastid Cherimoya polyphenol oxidase ABJ96144.1 56, 7e-74 1CpPPO5 ABIM01013359.1 1305 365 Poss Plastid Cherimoya polyphenol oxidase ABJ96144.1 40, 5e-101 1
Table 11 Polyphenol oxidase (tyrosinase, dicopper containing) andlaccase (multi copper oxidase) genes predicted in the papaya genome,the whole genome shotgun sequence accession number in NCBI(WGS Accession), coding sequence for amino acids (CDS), thenumber of introns in the nucleotide sequence (introns), amino acids(aa), whether a secretory sequence was predicted (secretory) its
location (ER endoplasmic retriculum) and aa length, similarity inpeptide sequence with another species (homology), extent thesequences were invariant (identity), E-value was the expectation valuefor homology, and the number of papaya expressed sequence tags(EST) detected with at least 500 bases and 99% identity
Tropical Plant Biol. (2008) 1:246–277 263263

clustered with two on each supercontigs 27 and 62.Supercontig 27 and 62 are not on the same linkage groupsuggesting that papaya PPO’s may not be located in a smalllocus on the same chromosome. Papaya PPOs did showsignificant homology with published PPO sequences and fitreadily within the superfamily (Fig. 3).
Peroxidases (POD) are secreted glycoproteins that areinvolved in cell elongation, cell wall modification andpathogen defenses [143]. The Arabidopsis genome has 73genes for POD [204] and rice has 138 [143] while only 33were predicted for papaya (Supplementary Table 5). Allpapaya PODs, CpPOD1 to CpPOD33 had a 22 to 26 aminoacid secretory sequences and molecular weights from 279to 437 amino acids with 47 to 83% identity to knownPODs. The presence of the secretory sequence to theapoplast suggested they would be classified as Class IIIPODs. Most of the predicted papaya PODs fell into therange 320–360 aa, similar to that reported for other plantPODs. No papaya ESTs were found for 18 of the predictedpapaya genes. The absence of ESTs did not mean that thegenes were not expressed, many Arabidopsis PODs areonly expressed at low levels and often in all organs [204].RT-PCR does detect the Arabidopsis PODs that have noESTs reported in the databases. Five papaya POD isoformshave been reported to increase during ripening [181] with aMW range similar to that of the predicted genes.
Volatile Production
All plant parts emit volatiles [57] with multiple functions.Of the many roles in plants, ecological interactions such asdefense against herbivores, pathogens, and as attractants foranimals to disperse pollen and seeds are presumed to beprincipal functions. [148, 149, 199]. Additionally, volatilesas components of flavor and aroma perception substantially
contribute to the utility of plant parts as human foodstuffs.Odorants are volatile and interact with human olfactoryreception [36] but they must be somewhat lipohilic as wellas water soluble, possesses adequate vapor pressure, andoccur in sufficient concentration for olfactory receptorinteraction. At least one hundred and sixty-six compoundshave been identified in papaya fruit volatiles [70, 71, 119,151, 179]. The most commonly identified papaya volatileshave been methyl butanoate, ethyl butanoate, 3-methyl-1-butanol and 1-butanol. The esters of lower fatty acids areconsidered to contribute much to the typical papaya flavor[151]. The amounts and relative content of volatiles hasbeen shown to vary with the stage of ripeness [71, 98], forexample, linalool production increases nearly 400 fold withonly a sevenfold increase in benzyl isothiocyanate duringripening [71]. When the fruit has reached the edible ripestage butanol, 3-methylbutanol, benzyl alcohol and α-terpineol are at their maximum concentrations [4]. One ofthe two volatiles that most closely resembled those ofpapayas is linalool, a product of the plastid synthesispathway. The other papaya volatile is methyl benzoate,described as having papaya qualities on odor assessment[119]. The sweaty odor quality of some papaya cultivarsis due probably to the production of methyl butanoate[119]. The Hawaii variety had very little methyl butanoate(0.06%) [70] while the Sri Lankan variety had 48.3%[119]. In addition, benzyl isothiocyanate contributes apungent off-odor. The amount of each volatile componentvaries both with cultivar and locality [72, 119].
Most volatile esters are described as fruity [37] and themost likely precursors are amino acids and lipids. Theprecursors of volatile esters, with branched alkyl chain, arevaline, isoleucine and other amino acids from threonine[138, 216]. Aliphatic esters and alcohols are producedfrom free fatty acids such as linoleic and linolenic [22].
Fig. 3 Polyphenoloxidase rela-tionship with other describedpolyphenol oxidases (PPO) fromNCBI, aligned with ClustalWand neighbor-joining values.The tree was constructed withMEGA 4.0
264 Tropical Plant Biol. (2008) 1:246–277

The pathways come together in the formation of aldehydesthat are reduced to alcohols by alcohol dehydrogenase[185] and ester formation by alcohol acyltransferase [59,69].
Single genes were found in the papaya genome for allthe enzymes in the biosynthesis from threonine to 3-keto-3-methylvalerate (Table 12). At least three branched chainamino transferase genes were found that could convert 3-keto-3-methylvalerate via isoleucine to 3-oxo-methylpenta-noic acid. The next step to 2-methyl butanal involvespyruvate decarboxylase and three genes encoding thisenzyme were predicted from the genome data; all werepredicted to have the characteristic domains. Four branchedchain alpha keto acid dehydrogenases (alcohol dehydro-genases) were predicted with three that had papaya EST.The interconversion between 3-methyl butanol and 2-methylbutyl acetate possibly involved a single predictedalcohol acyl transferase gene and four predicted carbox-ylesterases genes (Table 12).
Straight chain ester biosynthesis commonly occurs fromfatty acid with linoleic acid being a common precursor. Thefirst enzyme is lipoxygenase of which nine genes werepredicted in papaya with another possible partial sequencefound (Table 13). Except for two of the predicted lip-oxygenase, all had papaya ESTs. The predicted gene for thepreviously described papaya lipoxygenase (CpLOX1) hadeight ESTs and a chloroplast transit peptide. The otherCpLOXs did not have predicted chloroplast transit peptides,suggesting they may be located in the cytoplasm (Table 13).
One papaya hydroperoxide lyase gene (P450 superfamilymember) was found and no isomerasewas predicted (Table 13).Hex-3-enal to hex-2-enal isomerase has been reported to beunstable and no sequences are in the gene databases. Theisomerase possibly occurs in the cytoplasm [140, 226]. Thedehydrogenation of hex-3-enal to hex-3-enoic acid by alde-hyde dehydrogenase could involve upwards of five genes. Twoof the aldehyde dehydrogenase had predicted mitochondrialtransit peptides and may be involved in volatile production. Anumber of alcohol dehydrogenase genes were predicted thatcould convert hex-3-enal to hex-3-enol (Table 13).
Two volatiles with fruity/floral properties are 2-phenyl-acetaldehyde and 2-phenylethanol derived from phenylala-nine. The first enzyme in the pathway is a bifunctionalphenylacetaldehyde synthase possibly combining decarbox-ylation-amine oxidation leading to phenylacetaldehyde[97]. The last step is the reduction of two phenylacetalde-hyde to 2-phenylethanol [199]. The reduction involvesNADPH and 2-phenylacetaldehyde reductase. Genes werepredicted in papaya for both steps in biosynthesis(Table 14). Two predicted phenylacetaldehyde synthase(CpPALDS1, CpPALDS2) genes were found having highhomology to the deposited gene sequences for Rose andPetunia (Table 14). A single gene with moderate homology
(48%, E-value 3e-68) to the tomato gene was predicted forthe reduction to 2-phenylethanol [199].
Terpenoids, widely distributed in lichens, algae, andhigher plants [13, 30, 62, 178, 190, 99], represent the mostdiverse family of natural products, as there are over 40,000different structures identified thus far. Many terpenoids arenon-volatile and constitute important elements of plantfunctional processes, i.e. plant growth regulation, redoxreactions, membrane structure, and photosynthesis [56].Volatile and flavor compounds from essential oils ofvarious terpene classes are produced in leaves and fruitwhile carotenoids give the fruit skin and flesh its uniquecolor. Biosynthesis occurs through two different pathways:the acetate-mevalonate pathway in the cytoplasm [30] andthe non-mevalonate pathway in the plastids [111, 112]. Themevalonate pathway leads to sesquiterpenes and triterpenes(sterols) while monoterpenes, diterpenes, tetraterpenes(carotenoids) and polyterpenes are formed in the plastids.
Genes were predicted for most of the steps in terpenebiosynthesis occurring both in the plastids (Table 15) and inthe cytoplasm (Table 16). Two DXP synthases werepredicted with the shorter predicted peptide lacking achloroplast transit peptide (Table 15). Another partial DXPsynthase, one-third the length of the expected peptide wasfound with 78% homology (E-value 3e-77) to the rubberTPP enzyme domain (BAF98289.1). A full length linaloolsynthase gene (ABIM01025787.1) was predicted with twopartial sequences (ABIM0102980.1, ABIM01019472.1). Nogeranyl-geranyl diphosphate (GGPP) synthase was foundthat had a chloroplast transit peptide. A short sequence(ABIM01003036.1) had low homology (61%, E-value 1e-26, 105 aa) to AAQ84170.1 GGPP synthase that catalyzesthe farnesyl diphosphate to GGPP conversion was found.
Cytoplasmic terpene synthesis from mevalonate todimethylally diphosphate has a single gene for each step(Table 16). Two geranyl diphosphate genes were predictedwith significant homology to published sequences. Two fulllength squalene synthases were predicted along with twopartial sequences (ABIM01018750.1, 191 aa; ABIM010207788.1, 261 aa) that had high homology to a tomatohomologue (ABU40771.1) and Panax quinquefolius se-quence (CAJ58418.1).
The cleavage of carotenoids derived from the terpenesynthesis pathway can lead to the production of volatiles.Carotenoid cleavage can occur at any conjugated doublebonds by various enzymes such as CCD and NCEDs(discussed above in the Fruit Growth section) to form analdehyde or ketone in each product. The products includeabscisic acid, strigolactone and various aroma compounds[207]. While Arabidopsis contains nine CCD, five of whichare directly involved in abscisic acid synthesis, the otherCCDs lead to volatiles and non-volatile apocarotenoids [183]with one CCD able to cleave multiple substrates at different
Tropical Plant Biol. (2008) 1:246–277 265265

Tab
le12
Predicted
papaya
genesforthebranched
chainestersbiosynthesispathway.T
thewholegenomeshotgunsequence
accessionnumberin
NCBI(W
GSAccession),coding
sequence
foram
ino
acids(CDS),am
inoacids(aa),presenceof
discreteportionof
proteinpossessing
itsow
nfunctio
nandthesuperfam
ily(dom
ain),sim
ilarityinpeptidesequence
with
anotherspecies
(hom
ology),extentthe
sequenceswereinvariant(identity),E
-value
was
theexpectationvalueforhomology,andthenumberof
papaya
expressedsequence
tags
(EST
)detected
with
atleast5
00basesand99%
identity
Sub
strate
Enzym
eWGSAccession
cds
aaDom
ain
Hom
olog
yIdentity,
E-value
EST
Threonine
Threon
inedeaminase
ABIM
0100
2164
.147
3959
8PA
LP,
PRK09
224,
Arabido
psisthaliana
NP_1
8761
6.1
76,0.0
12-ketobu
tyrate Acetolactatesynthase
ABIM
0123
825.1
6524
257
ACT,
iLvH
,Arabido
psisthaliana
NP_8
5017
2.1
smallsubu
nit
82,8e-124
0
2-acetoh
ydroxy
butyrate
Acetohyd
roxy
acid
reductoisomerase
ABIM
0102
2972
.131
3845
9Ado
Hcyase,
IlvC
Sup
erfamily,
Arabido
psisthaliana
NP_1
9142
0.1
85,0.0
0
2,3-dihy
drox
y-3-methy
lvalerate
ABIM
0101
2960
.189
5261
7PRK00
911,
IlvD
-EDD
Arabido
psisthaliana
NP_1
8903
6.1
83,0.0
02-keto-3-m
ethy
lvalerate
Aminotransferase
isoleucine
Aminotransferase
ABIM
0101
9595
.119
5133
7BranchedchainAA
transferase,,
PLPDE-IV
Sup
erfamily
RiceNP_0
0104
2537
.165
,1e-126
7
ABIM
010115
18.1
1472
315
PLPDE-IV,Aminotransferase,
Arabido
psisthaliana,NP_2
0059
3.2
69,8e-126
1ABIM
0101
9463
.159
3041
7BCAT_b
eta_family,PLPDE_IV
Branched
Chain
Aminotransferase
Arabido
psisthaliana
NP_5
6692
3.1
76,0.0
1
2-ox
o-3-methy
lpentano
icacid
Pyruva
tedecarbox
ylase
ABIM
0101
8822
.124
9164
0COG39
61,TPP_enzym
e_N
Sup
erfamily,
TPP_P
DC_IPDC,.1
Arabido
psisthaliana
NP_1
9503
366
,0.0
2
ABIM
0100
2684
.133
2658
9TPP_enzym
e_N
Sup
erfamily,
TPP_P
DC_IPDC,
Citrus
sinensisAAZ05
069.1
87,0.0
1
ABIM
0101
8795
.124
9160
5TPP_enzym
e_N,TPP_P
DC_IPDC,
Arabido
psisthaliana
NP_2
0030
7.1
85,0.0
72-methy
lbutanal
Alcoh
oldehyd
rogenase
ABIM
0102
3823
.123
9935
4Alkenal
redu
ctase,
Arabido
psisthaliana
NP_1
9719
9.1
73,4e-152
1branched
chainalpha
ketoacid
dehyd
rogenase
ABIM
0100
2736
.128
5146
5TPP_enzym
esSup
erfamily,
TPP_E
1_PDC_A
DC_B
CADC
RiceNP_0
0106
6324
.172
,5e-180
1
ABIM
0101
0666
.141
3347
7AcoA,TPP_enzym
esSup
erfamily,
TPP_E
1_PDC_A
DC_B
CADC,
RiceABA95
968.2-alph
asubu
nit
76,0.0
0
ABIM
010118
66.1
+ABIM
010118
65.1
5264
501
2-ox
oacid_
dhsuperfam
ily,PRK1185
6Arabido
psisthaliana
NP_8
5052
7.1
62,7e-171
3
3-methy
lbutanol
⇓Alcoh
olacyl
tran
sferase
ABIM
0102
1279
.119
7635
5BCAT_b
eta_family,PLPDE_IV
Sup
erfamily.
Arabido
psisthaliana
NP_1
7247
8.1
6.75
0e-139
1
ABIM
0100
7428
.113
7445
7Transferase
Sup
erfamily,AtAcetyl
transferaselik
eArabido
psisthaliana
NP_1
8923
3.1
36,1e-79
0
⇑Carbox
ylesterase
ABIM
0102
0297
.110
0233
3Esterase_lip
aseSup
erfamily,CXE
Actinidia
deliciosa
ABB89
020.1
57,4e-110
0ABIM
0102
2048
.110
4134
6Esterase_lip
aseSup
erfamily,CXE
Actinidia
deliciosa
ABB89
015.1
62,3e-117
3ABIM
0102
3942
.194
231
3Esterase_lip
aseSup
erfamily,CXE
Malus
pumila
ABB89
002.1
67,4e-125
1ABIM
0101
3294
.197
532
4Esterase_lip
aseSup
erfamily,CXE
Malus
pumila
,ABB89
010.1
58,2e-101
32-methy
lbutylacetate
266 Tropical Plant Biol. (2008) 1:246–277
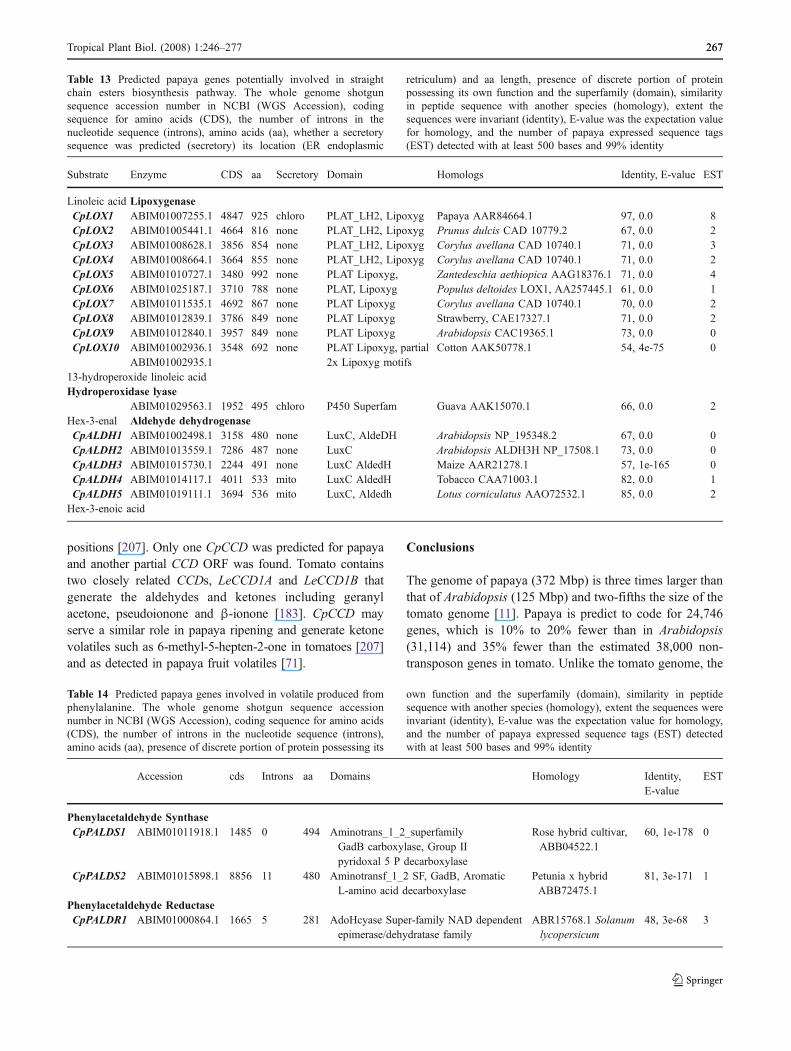
positions [207]. Only one CpCCD was predicted for papayaand another partial CCD ORF was found. Tomato containstwo closely related CCDs, LeCCD1A and LeCCD1B thatgenerate the aldehydes and ketones including geranylacetone, pseudoionone and β-ionone [183]. CpCCD mayserve a similar role in papaya ripening and generate ketonevolatiles such as 6-methyl-5-hepten-2-one in tomatoes [207]and as detected in papaya fruit volatiles [71].
Conclusions
The genome of papaya (372 Mbp) is three times larger thanthat of Arabidopsis (125 Mbp) and two-fifths the size of thetomato genome [11]. Papaya is predict to code for 24,746genes, which is 10% to 20% fewer than in Arabidopsis(31,114) and 35% fewer than the estimated 38,000 non-transposon genes in tomato. Unlike the tomato genome, the
Table 14 Predicted papaya genes involved in volatile produced fromphenylalanine. The whole genome shotgun sequence accessionnumber in NCBI (WGS Accession), coding sequence for amino acids(CDS), the number of introns in the nucleotide sequence (introns),amino acids (aa), presence of discrete portion of protein possessing its
own function and the superfamily (domain), similarity in peptidesequence with another species (homology), extent the sequences wereinvariant (identity), E-value was the expectation value for homology,and the number of papaya expressed sequence tags (EST) detectedwith at least 500 bases and 99% identity
Accession cds Introns aa Domains Homology Identity,E-value
EST
Phenylacetaldehyde SynthaseCpPALDS1 ABIM01011918.1 1485 0 494 Aminotrans_1_2_superfamily
GadB carboxylase, Group IIpyridoxal 5 P decarboxylase
Rose hybrid cultivar,ABB04522.1
60, 1e-178 0
CpPALDS2 ABIM01015898.1 8856 11 480 Aminotransf_1_2 SF, GadB, AromaticL-amino acid decarboxylase
Petunia x hybridABB72475.1
81, 3e-171 1
Phenylacetaldehyde ReductaseCpPALDR1 ABIM01000864.1 1665 5 281 AdoHcyase Super-family NAD dependent
epimerase/dehydratase familyABR15768.1 Solanumlycopersicum
48, 3e-68 3
Table 13 Predicted papaya genes potentially involved in straightchain esters biosynthesis pathway. The whole genome shotgunsequence accession number in NCBI (WGS Accession), codingsequence for amino acids (CDS), the number of introns in thenucleotide sequence (introns), amino acids (aa), whether a secretorysequence was predicted (secretory) its location (ER endoplasmic
retriculum) and aa length, presence of discrete portion of proteinpossessing its own function and the superfamily (domain), similarityin peptide sequence with another species (homology), extent thesequences were invariant (identity), E-value was the expectation valuefor homology, and the number of papaya expressed sequence tags(EST) detected with at least 500 bases and 99% identity
Substrate Enzyme CDS aa Secretory Domain Homologs Identity, E-value EST
Linoleic acid LipoxygenaseCpLOX1 ABIM01007255.1 4847 925 chloro PLAT_LH2, Lipoxyg Papaya AAR84664.1 97, 0.0 8CpLOX2 ABIM01005441.1 4664 816 none PLAT_LH2, Lipoxyg Prunus dulcis CAD 10779.2 67, 0.0 2CpLOX3 ABIM01008628.1 3856 854 none PLAT_LH2, Lipoxyg Corylus avellana CAD 10740.1 71, 0.0 3CpLOX4 ABIM01008664.1 3664 855 none PLAT_LH2, Lipoxyg Corylus avellana CAD 10740.1 71, 0.0 2CpLOX5 ABIM01010727.1 3480 992 none PLAT Lipoxyg, Zantedeschia aethiopica AAG18376.1 71, 0.0 4CpLOX6 ABIM01025187.1 3710 788 none PLAT, Lipoxyg Populus deltoides LOX1, AA257445.1 61, 0.0 1CpLOX7 ABIM01011535.1 4692 867 none PLAT Lipoxyg Corylus avellana CAD 10740.1 70, 0.0 2CpLOX8 ABIM01012839.1 3786 849 none PLAT Lipoxyg Strawberry, CAE17327.1 71, 0.0 2CpLOX9 ABIM01012840.1 3957 849 none PLAT Lipoxyg Arabidopsis CAC19365.1 73, 0.0 0CpLOX10 ABIM01002936.1 3548 692 none PLAT Lipoxyg, partial Cotton AAK50778.1 54, 4e-75 0
ABIM01002935.1 2x Lipoxyg motifs13-hydroperoxide linoleic acidHydroperoxidase lyase
ABIM01029563.1 1952 495 chloro P450 Superfam Guava AAK15070.1 66, 0.0 2Hex-3-enal Aldehyde dehydrogenaseCpALDH1 ABIM01002498.1 3158 480 none LuxC, AldeDH Arabidopsis NP_195348.2 67, 0.0 0CpALDH2 ABIM01013559.1 7286 487 none LuxC Arabidopsis ALDH3H NP_17508.1 73, 0.0 0CpALDH3 ABIM01015730.1 2244 491 none LuxC AldedH Maize AAR21278.1 57, 1e-165 0CpALDH4 ABIM01014117.1 4011 533 mito LuxC AldedH Tobacco CAA71003.1 82, 0.0 1CpALDH5 ABIM01019111.1 3694 536 mito LuxC, Aldedh Lotus corniculatus AAO72532.1 85, 0.0 2Hex-3-enoic acid
Tropical Plant Biol. (2008) 1:246–277 267267

Tab
le15
Predicted
papaya
genesfortheterpeneplastid
biosyn
thesispathway.The
who
legeno
meshotgu
nsequ
ence
accessionnu
mberin
NCBI(W
GSAccession
),coding
sequ
ence
foram
ino
acids(CDS),am
inoacids(aa),whether
atransitsequ
ence
was
predicted(Transit)
itslocatio
n,presence
ofdiscrete
portionof
proteinpo
ssessing
itsow
nfunctio
nandthesuperfam
ily(dom
ain),
similarity
inpeptidesequ
ence
with
anotherspecies(hom
olog
y),extent
thesequ
enceswereinvariant(identity
),E-value
was
theexpectationvalueforho
molog
y,andthenu
mberof
papaya
expressedsequ
ence
tags
(EST)detected
with
atleast50
0basesand99
%identity
Sub
strate
Enzym
eWGSAccession
cds
aaTransit
Dom
ain
Hom
olog
yIdentity,
E-value
EST
Pyruv
ate+Glyceraldehyd
e-3-ph
osph
ate
↓DXPsynthase
ABIM
0100
5151
.11161
718
chloro
TPP-D
XS
Tom
atoAAD38
941.1
85,0.0
0ABIM
0101
6399
.159
8854
9no
neTPP-D
XS.TPP_enzym
eAtDXPS3
68,9e-163
10ABIM
0101
6400
.1NP_0
0107
8570
.11-deox
y-D-xylulose-5-ph
osph
ate
↓DXPreductoisomerase
ABIM
0101
4958
.141
3138
4no
neDXP_reductoisom
_sup
erfamily,
PRK05
447
Rub
berBAF98
290.1
92,0.0
0
2-C-m
ethy
l-d-erythrito
l-4-ph
osph
ate
↓2C
-methylerythritol,
2,4-cyclod
iphosphatesnthase
ABIM
0102
5949
.174
7523
6chloro
MeC
DP_syn
thase,
Rub
berBAF98
295.1
74,1e-95
9
ABIM
0102
5947
.1ABIM
0102
5948
.1Isop
entyldipho
sphate
(IPP),dimethy
lally
diph
osph
ate
↓Isom
erase
Geranyl
diph
osph
ate
↓Linaloo
lsynthase
ABIM
0102
5787
.139
3085
2Isop
reno
id_B
iosyn_
C1
Arabido
psisthaliana
NP56
4772
.146
,0.0
0Linaloo
lGeranyl
diph
osph
ate
↓farnesyl
diphosphatesynthase
ABIM
0100
7640
.138
0934
2Trans_IPPS_H
T,Isop
reno
id_b
iosyn_
C1
Rub
berAAB07
264.1
85,4e-178
0Farnesyldiph
osph
ate
↓GGPPsynthase
Geranylgerany
ldiph
osph
ate
↓Terpenesynthase
ABIM
0100
7557
.140
5chloro
Isop
reno
id_b
iosyn_
C1
Arabido
psisthaliana
NP_5
6477
2.1
51,3e-81
1Diterpenes
-Phy
toene-Chlorop
hyll&
Carotenoids
268 Tropical Plant Biol. (2008) 1:246–277
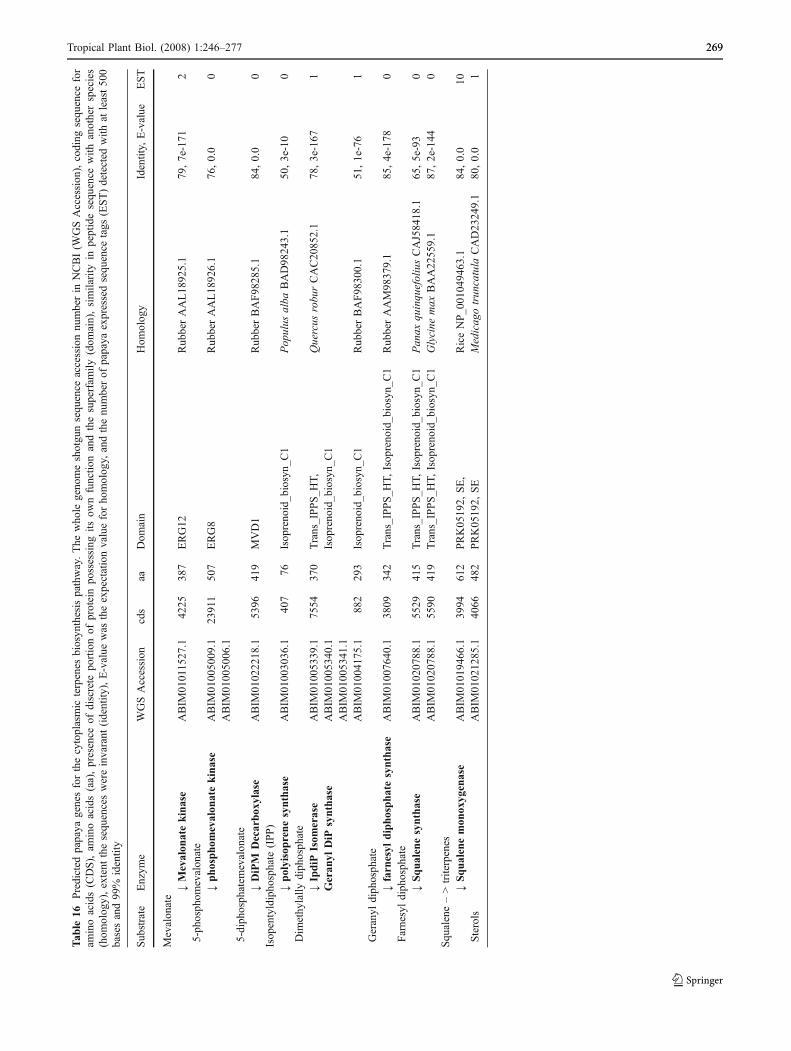
Tab
le16
Predicted
papaya
genesforthecytoplasmic
terpenes
biosyn
thesispathway.The
who
legeno
meshotgu
nsequ
ence
accessionnu
mberin
NCBI(W
GSAccession),coding
sequ
ence
for
aminoacids(CDS),am
inoacids(aa),presence
ofdiscrete
portionof
proteinpo
ssessing
itsow
nfunctio
nandthesuperfam
ily(dom
ain),similarity
inpeptidesequ
ence
with
anotherspecies
(hom
olog
y),extent
thesequ
enceswereinvarant
(identity
),E-value
was
theexpectationvalueforho
molog
y,andthenu
mberof
papaya
expressedsequ
ence
tags
(EST)detected
with
atleast50
0basesand99
%identity
Sub
strate
Enzym
eWGSAccession
cds
aaDom
ain
Hom
olog
yIdentity,
E-value
EST
Mevalon
ate↓Mevalon
atekinase
ABIM
010115
27.1
4225
387
ERG12
Rub
berAAL18
925.1
79,7e-171
25-ph
osph
omevalon
ate
↓phosphom
evalon
atekinase
ABIM
0100
5009
.123
911
507
ERG8
Rub
berAAL18
926.1
76,0.0
0ABIM
0100
5006
.15-diph
osph
atem
evalon
ate
↓DiPM
Decarbox
ylase
ABIM
0102
2218
.153
9641
9MVD1
Rub
berBAF98
285.1
84,0.0
0Isop
entyldipho
sphate
(IPP)
↓polyisoprenesynthase
ABIM
0100
3036
.140
776
Isop
reno
id_b
iosyn_
C1
Pop
ulus
alba
BAD98
243.1
50,3e-10
0Dim
ethy
lally
diph
osph
ate
↓IpdiP
Isom
erase
ABIM
0100
5339
.175
5437
0Trans_IPPS_H
T,Quercus
robu
rCAC20
852.1
78,3e-167
1Geran
ylDiP
synthase
ABIM
0100
5340
.1Isop
reno
id_b
iosyn_
C1
ABIM
0100
5341
.1ABIM
0100
4175
.188
229
3Isop
reno
id_b
iosyn_
C1
Rub
berBAF98
300.1
51,1e-76
1Geranyl
diph
osph
ate
↓farnesyl
diphosphatesynthase
ABIM
0100
7640
.138
0934
2Trans_IPPS_H
T,Isop
reno
id_b
iosyn_
C1
Rub
berAAM98
379.1
85,4e-178
0Farnesyldiph
osph
ate
↓Squalenesynthase
ABIM
0102
0788
.155
2941
5Trans_IPPS_H
T,Isop
reno
id_b
iosyn_
C1
Pan
axqu
inqu
efoliusCAJ584
18.1
65,5e-93
0ABIM
0102
0788
.155
9041
9Trans_IPPS_H
T,Isop
reno
id_b
iosyn_
C1
Glycine
max
BAA22
559.1
87,2e-144
0Squ
alene–>triterpenes
↓Squalenemon
oxyg
enase
ABIM
0101
9466
.139
9461
2PRK05
192,
SE,
RiceNP_0
0104
9463
.184
,0.0
10Sterols
ABIM
0102
1285
.140
6648
2PRK05
192,
SE
Medicag
otrun
catula
CAD23
249.1
80,0.0
1
Tropical Plant Biol. (2008) 1:246–277 269269

papaya genome is largely euchromatic [132, 209]. Thelower gene number in papaya may reflect that, unlikeArabidopsis [32], the papaya genome has not undergone arecent whole-genome duplication and has had feweropportunities for fractionation of linkage arrangements bypost-duplication gene loss [132, 142]. This suggests thatgene structure, function, and arrangement in papaya mayresemble the ancestral angiosperm more closely than dopreviously sequenced angiosperms [116]. Comparison ofthe five sequenced clade members suggests a minimalangiosperm gene set of 13,311. Fewer introns have beenfound in papaya genes than in their orthologs in Arabi-dopsis (8,319 versus 11,718) and rice (6,874 versus 7,011).Approximately 2,000 transcription factors in over 60families have been identified in Arabidopsis [85], withpapaya having fewer members in the majority of families,but a significantly higher number of predicted MADS-boxproteins (171 versus 141). The comparison of geneexpression and activity during fruit ripening suggests thatthe same regulatory pathways and similar controls occur[81]. The lack of whole genome duplication and reductionsin most papaya gene families and biosynthetic pathwaysmakes papaya a valuable tool for the study of the complexregulatory networks active in fruit ripening. The example,fewer gene numbers for ethylene synthesis, receptors and inthe ethylene response pathways may provide insights as tohow ripening is controlled.
Materials and Methods
Gene Prediction
Papaya unigenes from complementary DNA were alignedand used in gene prediction software [133]. Spliced align-ments of proteins from the plant division of GenBank, andtranscripts from related angiosperms, were generated. Genepredictions were combined with spliced alignments ofproteins and transcripts to produce a reference gene set.Gene prediction software Augustus, GlimmerHMM andSNAP were used. Ab initio gene prediction softwareFgenesh, Genscan and TWINSCAN were trained onArabidopsis. Predicted proteins were aligned to the plantdivision of GenBank and transcripts from related angio-sperms (Arabidopsis thaliana, Glycine max, Gossypiumhirsutum, Medicago truncatula, Nicotiana tabacum, Oryzasativa, Zea mays). Gene predictions were combined withspliced alignments of proteins and transcripts to produce areference gene set using the evidence-based combinerEVidenceModeler (EVM). Where predicted genes werenot indicated and to confirm a prediction, known cDNAand protein sequences were downloaded for Arabidopsis,poplar, tomato and other species as required. Local searches
were made using the papaya WGS, EST and predicted genedatabases using the BLAST search application in BioEdit[87]. The potential papaya candidate genes indicated in theBioEdit search were then cross-checked against NCBIBLAST and homology confirmed and conserved domainsdetermined.
Similarity Searches and Alignment
Similarity searches were carried out by online BLASTsearch [120]. Best ORF predicted were BLASTp againstnon-redundant database to determine similarity with otherproteins. Proteins with low E-score of ≥50 were acceptedfor comparison, with emphasis on proteins with E-scores≤100 favored. Similar proteins that had published annota-tion that was not predicted were identified.
Sequences were aligned using the multiple sequencealignment program, CLUSTAL W version 1.6 [200] usingdefault parameters (gap penalty of 15, gap length penalty of6.66). Gaps and positions with ambiguities were excludedfrom the phylogenetic analysis. Phylogenetic analysis wasperformed using MEGA 4.0 [191] using neighbor-joiningmethods [171]. The corresponding whole genome shotgunreads (WGS) sequence was identified by using the BLASTroutine on the genomic sequence identifying the ORF withBLASTn.
Predicted Peptide Domains
Protein domains were predicted using InterProScan againstprotein databases (PRINTS, Pfam, ProDom, PROSITE,SMART).
Papaya ESTs
TheGenBank accession numbers of the papaya ESTs from thegenome project are EX227656 to EX303501 [132]. TheNCBI GenBank contains 77,128 papaya EST entries (2008September 08). The genes reported herein, were doublechecked against this database. EST sequences were identifiedby taking the genomic nucleotide sequence of the regionidentified for the ORF and using BLASTn against the ESTdatabase for Caricia papaya. Matches were screened byaligning the EST sequence with the genomic nucleotidesequence using the ALIGN program. Matches of greater than99% for more than 500 sequential bases were generally used.EST matches were further screened by Prosite and Inter-proscan to insure that they encoded the expected gene.
Targeting Sequences
Predotar was used as the primary screen for N-terminalputative targeting sequences [184] for mitochondrial,
270 Tropical Plant Biol. (2008) 1:246–277

plastid and ER targeting. MitoProt [50], ChloroP [66, 67]and TargetP [67] were also used to verify predictions withall programs having a low false positive rate. The absenceof a predicted transit or secretion sequence did not meanthat it was not present, as the programs only predicts about85% of the sequences [68].
References
1. Adams-Phillips L, Barry C, Kannan P, Lecercq J, Bouzayen M,Giovannoni JJ (2004) Ctr1-mediated ethylene signal transductionin tomato is encoded by a multigene family whose membersdisplay distinct regulatory features. Plant Mol Biol 54:387–404,doi:10.1023/B:PLAN.0000036371.30528.26
2. Alba R, Payton P, Fei Z, McQuinn R, Debbie P, Martin GB,Tanksley SD, Giovannoni JJ (2005) Transcriptome and selectedmetabolite analysis reveal multiple points of ethylene controlduring tomato fruit development. Plant Cell 17:2954–2965,doi:10.1105/tpc.105.036053
3. Alferez F, Zacarias L (1999) Interaction between ethylene andabscisic acid in the regulation of citrus fruit maturation. In:Kanellis AK, Chang C, Klee H, Blecker AB, Pech JC, Grierson D(eds) Biology and biotechnology of the plant hormone ethylene.II. Kluwer Academic Publishers, Amsterdam, pp 183–184
4. Almora K, Pino JA, Hernandez M, Duarte C, Gonzalez J, RoncalE (2004) Evaluation of volatiles from ripening papaya (Caricapapaya L., var. Maradol roja). Food Chem 86:127–130,doi:10.1016/j.foodchem.2003.09.039
5. Alonso JM, Stepanova AN (2004) The ethylene signalingpathway. Science 306:1513–1515, doi:10.1126/science.1104812
6. Alonso JM, Stepanova AN, Solano R, Wisman E, Ferrari S,Ausubel FM, Ecker JR (2003) Five components of the ethylene-response pathway identified in a screen for weak ethylene-insensitive mutants in Arabidopsis. Proc Natl Acad Sci USA100:2992–2997, doi:10.1073/pnas.0438070100
7. Andersson-Gunneras S, Hellgren JM, Bjorklund S, Regan S,Moritz T, Sundberg B (2003) Asymmetric expression of a poplarACC oxidase controls ethylene production during gravitationalinduction of tension wood. Plant J 34:339–349, doi:10.1046/j.1365-313X.2003.01727.x
8. APG (Angiosperm Phylogeny Group) (1998) An ordinalclassification for the families of flowering plants. Ann MissBot Gard 85:531–553, doi:10.2307/2992015
9. APG (Angiosperm Phylogeny Group) II (2003) An update of theAngiosperm Phylogeny Group classification for the orders andfamilies of flowering plants: APG II. Bot J Linnean Soc141:399–436. Updated Stevens PF (2001 onwards). AngiospermPhylogeny Website. Version 9, June 2008 http://www.mobot.org/MOBOT/research/APweb/
10. Armstrong GA, Hearst JE (1996) Carotenoids 2: genetics andmolecular biology of carotenoid pigment biosynthesis. FASEB J10:228–237
11. Arumuganathan K, Earle ED (1991) Nuclear DNA content ofsome important plant species. Plant Mol Biol Rep 9:208–218,doi:10.1007/BF02672069
12. Asha VA, Sane AP, Sane PN (2007) Multiple forms of α-expansin genes are expressed during banana fruit ripening anddevelopment. Postharvest Biol Technol 45:184–192,doi:10.1016/j.postharvbio.2007.03.003
13. Aubourg S, Lecharny A, Bohlmann J (2002) Genomic analysisof the terpenoid synthase (AtTPS) gene family of Arabidopsis
thaliana. Mol Genet Genomics 267:730–745, doi:10.1007/s00438-002-0709-y
14. Auldridge ME, Block A, Vogel JT, Dabney-Smith C, Mila I,Bouzayen M, Magallanes-Lundback M, DellaPenna D, McCartyDR, Klee HJ (2006a) Characterization of three members of theArabidopsis carotenoid cleavage dioxygenase family demon-strates the divergent roles of this multifunctional enzyme family.Plant J 45:982–993, doi:10.1111/j.1365-313X.2006.02666.x
15. Auldridge M, McCarty DR, Klee HJ (2006b) Plant carotenoidcleavage oxygenases and their apocarotenoid products. CurrOpin Plant Biol 9:315–321, doi:10.1016/j.pbi.2006.03.005
16. Azarkan M, Dibiani R, Baulard C, Baeyens-Volant D (2006)Effects of mechanical wounding on Carica papaya cysteineendopeptidases accumulation and activity. Int J Biol Macromol38:216–224, doi:10.1016/j.ijbiomac.2006.02.021
17. Azarkan M, Dibiani R, Goormaghtigh E, Raussens V, Baeyens-Volant D (2006) The papaya Kunitz-type trypsin inhibitor is ahighly stable β-sheet glycoprotein. Biochim Biophy Acta -Proteins & Proteomics 1764:1063–1072
18. Azarkan M, El Moussaoui A, van Wuytswinkel D, Dehon G,Looze Y (2003) Fractionation and purification of the enzymesstored in the latex of Carica papaya. J Chromat B 790:229–238,doi:10.1016/S1570-0232(03)00084-9
19. Badillo VM (1993) Caricaceae segundo esquema. Alcance 43.Universidad Central de Venezuela, Facultad de Agronoma.p111
20. Badillo VM (2000) Carica L. vs. Vasconcella St. Hil. (Carica-ceae): con la rehabilitación de este último. Ernstia 10:74–79
21. Baines BS, Brocklehurst K (1982) Characterization of papayapeptidase A as a cysteine proteinase of Carica papaya L. withactive center properties that differ from those of papain by using2,2′-dipyridyl disulfide and 4-chloro-7-nitrobenzofurazan asreactivity probes. Biochem J 205:205–211
22. Baldwin EA, Scott JW, Shewmaker CK, Schuch W (2000)Flavor trivia and tomato aroma: biochemistry and possiblemechanisms for control of important aroma components.HortScience 35:1013–1022
23. Barry CS, Llop-Tous MI, Grierson D (2000) The regulation of1-aminocyclopropane-1-carboxylic acid synthase gene expres-sion during the transition from system-1 to system-2 ethylenesynthesis in tomato. Plant Physiol 123:979–986, doi:10.1104/pp.123.3.979
24. Bartley GE, Ishida BK (2003) Developmental gene regulationduring tomato fruit ripening and in-vitro sepal morphogenesis.BMC Plant Biology 3, 4 (http://www.biomedcentral.com/ 1471-2229/3/4)
25. Baud S, Vaultier MN, Rochat C (2004) Structure and expressionprofile of the sucrose synthase multigene family in Arabidopsis.J Exp Bot 55:397–409, doi:10.1093/jxb/erh047
26. Bennett RN, Kiddle G, Wallsgrove RM (1997) Biosynthesis ofbenzylglucosinolate, cyanogenic glucosides and phenylpropa-noids in Carica papaya. Phytochemistry 45:59–66, doi:10.1016/S0031-9422(96)00787-X
27. Biale JB (1964) Growth, maturation, and senescence in fruits.Science 146:880–888, doi:10.1126/science.146.3646.880
28. Binder BM, Walker JM, Gagne JM, Emborg TJ, Hemmann G,Bleecker AB, Vierstra RD (2007) The Arabidopsis EIN3 bindingF-Box proteins EBF1 and EBF2 have distinct but overlappingroles in ethylene signaling. Plant Cell 19:509–523, doi:10.1105/tpc.106.048140
29. Bocock PN, Morse AM, Dervinis C, Davis JM (2008) Evolutionand diversity of invertase genes in Populus trichocarpa. Planta227:565–576, doi:10.1007/s00425-007-0639-3
30. Bohlmann J, Meyer-Gauen G, Croteau R (1998) Plant terpenoidsynthases: molecular biology and phylogenetic analysis. ProcNatl Acad Sci USA 95:4126–4133, doi:10.1073/pnas.95.8.4126
Tropical Plant Biol. (2008) 1:246–277 271271

31. Booker J, Auldridge ME, Wills S, McCarty DR, Klee HJ, LeyserO (2004) MAX3/CCD7 is a carotenoid cleavage dioxygenaserequired for the synthesis of a novel plant signaling molecule.Curr Biol 14:1–20, doi:10.1016/j.cub.2004.06.061
32. Bowers JE, Chapman AB, Rong J, Paterson AH (2003)Unraveling angiosperm genome evolution by phylogeneticanalysis of chromosomal duplication events. Nature 422:433–438, doi:10.1038/nature01521
33. Bramley P, Teulieres C, Blain I, Bird C, Schuch W (1992)Biochemical characterization of transgenic tomato plants inwhich carotenoid synthesis has been inhibited through theexpression of antisense RNA to pTOM5. Plant J 2:343–349,doi:10.1111/j.1365-313X.1992.00343.x
34. Brummell DA, Harpster MH, Civello PM, Palys JM, BennettAB, Dunsmuir P (1999) Modification of expansin proteinabundance in tomato fruit alters softening and cell wall polymermetabolism during ripening. Plant Cell 11:2203–2216
35. Bruinsma J, Paull RE (1984) Respiration during postharvestdevelopment of soursop fruit. Annona muricata L. Plant Physiol76:131–138
36. Buck L, Axel R (1991) A novel multigene family may encodeodorant receptors: a molecular basis for odor recognition. Cell65:175–187, doi:10.1016/0092-8674(91)90418-X
37. Burdock GA (2002) Fenaroli’s handbook of flavor ingredients,4th edn. CRC Press, Boca Raton
38. Burg SP, Burg EA (1965) Relationship between ethyleneproduction and ripening in bananas. Bot Gaz 126:200–204,doi:10.1086/336320
39. Büttner M (2007) The monosaccharide transporter(-like) genefamily in Arabidopsis. FEBS Lett 581:2318–2324, doi:10.1016/j.febslet.2007.03.016
40. Cano MP, De Ancos B, Lobo G (1995) Peroxidase andpolyphenoloxidase activities in papaya during postharvestripening and after freezing-thawing. J Fd Sci 60:815–820,doi:10.1111/j.1365-2621.1995.tb06236.x
41. Cano MP, Lobo MG, De Ancos B, Galeazzi MAM (1996)Polyphenol oxidase from Spanish hermaphrodite and femalepapaya fruits (Carica papaya Cv. Sunrise, Solo Group). J AgricFood Chem 44:3075–3079, doi:10.1021/jf960119k
42. Caspari T,Will A, OpekarovaM, Sauer N, TannerW (1994) Hexose/H + symporters in lower and higher plants. J Exp Biol 196:483–491
43. Chan HT (1979) Sugar composition of papaya during develop-ment. HortScience 14:140–141
44. Chan HT Jr, Flath RA, Forrey RR, Cavaletto CG, NakayamaTOM, Brekke JE (1973) Development of off-odors and off-flavors in papaya puree. J Agric Food Chem 21:566–570,doi:10.1021/jf60188a018
45. Chan YK, Paull RE (2007) Papaya Carica papaya L., Caricaceae.In: Janick J, Paull RE (eds) Encyclopedia of Fruit and Nuts.CABI, Wallingford, United Kingdom, pp 237–247
46. Chen NM, Paull RE (1985) Development and prevention ofchilling injury in papaya fruit. J Am Soc Hortic Sci 111:639–643
47. Chen NJ, Paull RE (2003) Endoxylanase expressed duringpapaya fruit ripening: purification, cloning and characterization.Funct Plant Biol 30:433–441, doi:10.1071/FP02208
48. Chevalier T, De Rigal D, Mbeguie-A-Mbeguie D, Gauillard F,Richard-Forget F, Fils-Lycaon BR (1999) Molecular cloning andcharacterization of apricot fruit polyphenol oxidase. PlantPhysiol 119:1261–1269, doi:10.1104/pp.119.4.1261
49. Choi D, Cho DT, Lee Y (2006) Expansins: expandingimportance in plant growth and development. Physiol Plant126:511–518
50. Claros MG, Vincens P (1996) Computational method to predictmitochondrially imported proteins and their targeting sequences.Eur J Biochem 241:779–786, doi :10 .1111/ j .1432-1033.1996.00779.x
51. Claus H (2004) Laccases: structure, reactions, distribution.Micron 35:93–96, doi:10.1016/j.micron.2003.10.029
52. Cong B, Tanksley SD (2006) FW2.2 and cell cycle control indeveloping tomato fruit: a possible example of gene co-option inthe evolution of a novel organ. Plant Mol Biol 62:867–880,doi:10.1007/s11103-006-9062-6
53. Considine MJ, Daley DO, Whelan J (2001) The expression ofalternative oxidase and uncoupling protein during fruit ripeningin Mango. Plant Physiol 126:1619–1629, doi:10.1104/pp.126.4.1619
54. Considine MJ, Holtzapffel RC, Day DA, Whelan J, Millar AH(2002) Molecular distinction between alternative oxidase frommonocots and dicots. Plant Physiol 129:949–953, doi:10.1104/pp.004150
55. Constabel CP, Yip L, Patton JJ, Christopher ME (2000)Polyphenol oxidase from hybrid poplar. cloning and expressionin response to wounding and herbivory. Plant Physiol 124:285–296, doi:10.1104/pp.124.1.285
56. Cosgrove DJ (2007) Expansin Home Page. http://www.bio.psu.edu/expansins/index.htm. Cited 22 March 2008
57. Croteau R, Lutchan TM, Lewis NG (2000) Natural Products(secondary metabolites). In: Biochemistry and Molecular Biolo-gy of Plants, Buchanan B, Gruissem W, Jones R (eds.) RockvilleMD, Amer. Soc. Plant Physiol. pp1250–1318
58. Cunningham FX Jr, Gantt E (1998) Genes and enzymes ofcarotenoid biosynthesis in plants. Annu Rev Plant Physiol PlantMol Biol 49:557–583, doi:10.1146/annurev.arplant.49.1.557
59. D’Auria JC (2006) Acyltransferases in plants: a good time to beBAHD. Curr Opin Plant Biol 9:331–340, doi:10.1016/j.pbi.2006.03.016
60. Devitt LC, Sawbridge T, Holton TA, Mitchelson K, Dietzgen RG(2006) Discovery of genes associated with fruit ripening inCarica papaya using expressed sequence tags. Plant Sci170:356–363, doi:10.1016/j.plantsci.2005.09.003
61. Dibley SJ, Gear ML, Xiao Y, Rosche EG, Offler CE, McCurdyDW, Patrick JW (2005) Temporal and spatial expression ofhexose transporters in developing tomato (Lycopersicon escu-lentum) fruit. Funct Plant Biol 32:777–785, doi:10.1071/FP04224
62. Dudareva N, Pichersky E, Gershenzon J (2004) Biochemistry ofplant volatiles. Plant Physiol 135:1893–1902, doi:10.1104/pp.104.049981
63. Ehlting J, Mattheus N, Aeschliman DS, Li E, Hamberger B,Cullis IF, Zhuang J, Kaneda M, Mansfield SD, Samuels L,Ritland K, Ellis B, Bohlmann J, Douglas CJ (2005) Globaltranscript profiling of primary stems from Arabidopsis thalianaidentifies candidate genes for missing links in lignin biosynthesisand transcriptional regulators of fiber differentiation. Plant J42:618–640, doi:10.1111/j.1365-313X.2005.02403.x
64. Elhafez D, Murcha MW, Clifton R, Soole KL, Day DA,Whelan J (2006) Characterization of mitochondrial alternativeNAD(P)H dehydrogenases in Arabidopsis: intraorganelle loca-tion and expression. Plant Cell Physiol 47:43–54, doi:10.1093/pcp/pci221
65. El Moussaoui A, Nijs M, Paul C, Wintjens R, Vincentelli J,Azarkan M, Looze Y (2001) Revisiting the enzymes stored inthe laticifers of Carica papaya in the context of their possibleparticipation in the plant defence mechanism. Cell Mol Life Sci58:556–570, doi:10.1007/PL00000881
66. Emanuelsson O, Nielsen H, von Heijne G (1999) ChloroP, aneural network-based method for predicting chloroplast transitpeptides and their cleavage sites. Protein Sci 8:978–984
67. Emanuelsson O, Nielsen H, Brunak S, von Heijne G (2000)Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J Mol Biol 300:1005–1016,doi:10.1006/jmbi.2000.3903
272 Tropical Plant Biol. (2008) 1:246–277

68. Emanuelsson O, Brunak S, von Heijne G, Nielsen H (2007)Locating proteins in the cell using TargetP, SignalP, and relatedtools. Nat Protocols 2:953–971, doi:10.1038/nprot.2007.131
69. Fellman JK, Miller TW, Mattison DS, Mattheis JP (2000)Factors that influence biosynthesis of volatile flavor compoundsin apple fruits. HortScience 35:1026–1033
70. Flath RA, Forrey RR (1977) Volatile components of papaya(Carica papaya L., Solo variety). J Agric Food Chem 25:103–109, doi:10.1021/jf60209a051
71. Flath RA, Light DM, Jang EB, Mon TR, John JO (1990)Headspace examination of volatile emissions form ripeningpapaya (Carica papaya L., Solo variety). J Agric Food Chem38:1060–1063, doi:10.1021/jf00094a032
72. Franco MRB, Rodriguez ADB (1993) Volatile components oftwo pawpaw cultivars. Arq Biol Tecnol 36:613–632
73. Fraser PD, Truesdale MR, Bird CR, Schuch W, Bramley PM(1994) Carotenoid biosynthesis during tomato fruit development.Plant Physiol 105:405–413
74. Fraser PD, Kiano JW, Truesdale MR, Schuch W, Bramley PM(1999) Phytoene synthase-2 enzyme activity in tomato does notcontribute to carotenoid synthesis in ripening fruit. Plant MolBiol 40:687–698, doi:10.1023/A:1006256302570
75. Frary A, Nesbitt C, Frary A, Grandillo S, van der Knaap E, CongB, Liu J, Meller J, Elber R, Alpert KB, Tanksley SD (2000)fw2.2: a quantitative trait locus key to the evolution of tomatofruit size. Science 289:85–88, doi:10.1126/science.289.5476.85
76. Frank HA, Brudvig GW (2004) Redox functions of carotenoidsin photosynthesis. Biochem 43:8607–8615, doi:10.1021/bi0492096
77. Fridman M, Zamir D (2003) Functional divergence of a syntenicinvertase gene family in tomato, potato, and Arabidopsis. PlantPhysiol 131:603–609, doi:10.1104/pp.014431
78. Frye CA, Tang D, Innes RW (2001) Negative regulation ofdefense responses in plant by conserved MAPKK kinase. ProcNatl Acad Sci USA 98:373–378, doi:10.1073/pnas.011405198
79. Gear ML, McPhillips ML, Patrick JW, McCurdy DW (2000)Hexose transporters of tomato: molecular cloning, expressionanalysis and functional characterization. Plant Mol Biol 44:687–697, doi:10.1023/A:1026578506625
80. Geisler DA, Broselid C, Hederstedt L, Rasmusson AG (2007)Ca2 + -binding and Ca2 + -independent respiratory NADH andNADPH dehydrogenases of Arabidopsis thaliana. J Biol Chem282:28455–28464, doi:10.1074/jbc.M704674200
81. Giovannoni JJ (2004) Genetic regulation of fruit developmentand ripening. Plant Cell 16:S170–S180, doi:10.1105/tpc.019158
82. Gomez-Roldan V, Fermas S, Brewer PB, Puech-Pages V, DunEA, Pillot JP, Letisse F, Matusova R, Danoun S, Portais JC,Bouwmeester H, Becard G, Beveridge CA, Rameau C,Rochange SF (2008) Strigolactone inhibition of shoot branching.Nature 455:189–194, doi:10.1038/nature07271
83. Grubb CD, Abel S (2006) Glucosinolate metabolism and itscontrol. Trends Plant Sci 11:89–100, doi:10.1016/j.tplants.2005.12.006
84. Grubb CD, Zipp BJ, Ludwig-Müller J, Masuno MN, MolinskiTF, Abel S (2004) Arabidopsis glucosyltransferase UGT74B1functions in glucosinolate biosynthesis and auxin homeostasis.Plant J 40:893–908, doi:10.1111/j.1365-313X.2004.02261.x
85. Guo AY, He K, Liu D, Bai SN, Gu XC, Wei LP, Luo JC (2005)DATF: a database of Arabidopsis transcription factors. Bioinfor-matics 21:2568–2569, doi:10.1093/bioinformatics/bti334
86. Gussman CD, Goffredz JC, Gianfagna TJ (1993) Ethyleneproduction and fruit-softening rates in several apple fruitripening variants. HortScience 28:135–137
87. Hall TA (1999) BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symp Ser 41:95–98
88. Hall BP, Shakeel SN, Schaller GE (2007) Ethylene receptors:Ethylene perception and signal transduction. J Plant GrowthRegul 26:118–130, doi:10.1007/s00344-007-9000-0
89. Hamberger B, Ellis M, Friedmann M, de Azevedo Souza C,Barbazuk B, Douglas CJ (2007) Genome-wide analysis ofphenylpropanoid-related genes in Populus trichocarpa, Arabi-dopsis thaliana, and Ozyza sativa: the Populus lignin toolbox andconservation and diversification of angiosperm gene families.Can J Bot 85:1182–1201, doi:10.1139/B07–098
90. Henry MF, Nyns EJ (1975) Cyanide-insensitive respiration. Analternative mitochondrial pathway. Subcell Biochem 4:1–65
91. Hirschberg J (2001) Carotenoid biosynthesis in flowering plants. CurrOpin Plant Biol 4:210–218, doi:10.1016/S1369-5266(00)00163-1
92. Hortensteiner S (2006) Chlorophyll degradation during senes-cence. Annu Rev Plant Biol 57:55–77, doi:10.1146/annurev.arplant.57.032905.105212
93. Hua J, Meyerowitz EM (1998) Ethylene responses are negativelyregulated by a receptor gene family in Arabidopsis thaliana. Cell94:261–271, doi:10.1016/S0092-8674(00)81425-7
94. Hua J, Sakai H, Nourizadeh S, Chen QG, Bleecker AB, EckerJR, Meyerowitz EM (1998) EIN4 and ERS2 are members of theputative ethylene receptor gene family in Arabidopsis. Plant Cell10:1321–1332
95. Jansen EF, Balls AK (1941) Chymopapain: a new crystallineproteinase from papaya latex. J Biol Chem 137:459–460
96. Ji X, Van den Ende W, Van Laere A, Cheng S, Bennett J (2005)Structure, evolution, and expression of the two invertase genefamilies of rice. J Mol Evol 60:615–634, doi:10.1007/s00239-004-0242-1
97. Kaminaga Y, Schnepp J, Peel G, Kish CM, Ben-Nissan G, WeissD, Orlova I, Lavie O, Rhodes D, Wood K, Porterfield DM,Cooper AJL, Schloss JV, Pichersky E, Vainstein A, Dudareva N(2007) Plant phenylacetaldehyde synthase is a bifunctionalhomotetrameric enzyme that catalyzes phenylalanine decarbox-ylation and oxidation. J Biol Chem 281:23357–23366,doi:10.1074/jbc.M602708200
98. Katague DB, Kirch ER (1965) Chromatographic analysis ofvolatile components of papaya fruits. J Pharm Sci 54:891–894,doi:10.1002/jps.2600540616
99. Kevany B, Taylor M, Dal Cin V, Klee HJ (2007) Ethylene receptordegradation controls the timing of ripening in tomato fruit. Plant J51:458–467, doi:10.1111/j.1365-313X.2007.03170.x
100. Klee H, Tieman D (2002) The tomato ethylene receptor genefamily: form and function. Physiol Plant 115:336–341,doi:10.1034/j.1399-3054.2002.1150302.x
101. Kliebenstein DJ, Kroymann J, Mitchell-Olds T (2005) Theglucosinolate-myrosinase system in an ecological and evolution-ary context. Curr Opin Plant Biol 8:264–271, doi:10.1016/j.pbi.2005.03.002
102. Komatsu A, Moriguchi T, Koyama K, Omura M, Akihama T(2002) Analysis of sucrose synthase genes in citrus suggestsdifferent roles and phylogenetic relationships. J Exp Bot 53:61–71, doi:10.1093/jexbot/53.366.61
103. Ku HM, Doganlar S, Chen KY, Tanksley S (1999) The geneticbasis of pear-shaped tomato fruit. TAG 9:844–850, doi:10.1007/s001220051304
104. Ku HM, Grandillo G, Tanksley SD (2000a) fs8.1, a major QTL,sets the pattern of tomato carpel shape well before anthesis. TAG101:873–878, doi:10.1007/s001220051555
105. Ku HM, Vision T, Liu JP, Tanksley SD (2000b) Comparingsequenced segments of the tomato and Arabidopsis genomes:large-scale duplication followed by selective gene loss creates anetwork of synteny. Proc Natl Acad Sci USA 97:9121–9126,doi:10.1073/pnas.160271297
106. Kunieda T, Amano T, Shioi Y (2005) Search for chlorophylldegradation enzyme, Mg-dechelatase, from extracts of Chenopo-
Tropical Plant Biol. (2008) 1:246–277 273273

dium album with native and artificial substrates. Plant Sci169:177–183, doi:10.1016/j.plantsci.2005.03.010
107. Langenkamper G, Fung RWM, Newcomb RD, Atkinson RG,Gardner RC, MacRae EA (2002) Sucrose phosphate synthasegenes in plants belong to three different families. J Mol Evol54:322–332
108. Lelievre JM, Latche A, Jones B, Bouzayen M, Pech JC (1997)Ethylene and fruit ripening. Physiol Plant 101:727–739,doi:10.1111/j.1399-3054.1997.tb01057.x
109. Li L, Steffens JC (2002) Overexpression of polyphenol oxidasein transgenic tomato plants results in enhanced bacterial diseaseresistance. Planta 215:239–247, doi:10.1007/s00425-002-0750-4
110. Li Y, Darley CP, Ongaro V, Fleming A, Schipper O, Baldauf SL,McQueen-Mason SJ (2002) Plant expansins are a complexmultigene family with an ancient evolutionary origin. PlantPhysiol 128:854–864, doi:10.1104/pp.010658
111. Lichtenthaler HK (1999) The 1-deoxy-D-xylulose-5-phosphatepathway of isoprenoid biosynthesis in plants. Annu Rev PlantPhysiol Plant Mol Biol 50:47–65, doi:10.1146/annurev.arplant.50.1.47
112. Lichtenthaler HK (2000) Non-mevalonate isoprenoid biosynthe-sis: Enzymes, genes and inhibitors. Biochem Soc Trans 28:785–789, doi:10.1042/BST0280785
113. Lippman Z, Tanksley SD (2001) Dissecting the genetic pathwayto extreme fruit size in tomato using a cross between the small-fruited wild species Lycopersicon pimpinellifolium and L.esculentum var. Giant Heirloom. Genetics 158:413–422
114. Liu J, Eck JV, Cong B, Tanksley SD (2002) A new class ofregulatory genes underlying the cause of pear-shaped tomatofruit. Proc Natl Acad Sci USA 99:13302–13306, doi:10.1073/pnas.162485999
115. López-Ráez JA, Charnikhova T, Gómez-Roldán V, Matusova R,Kohlen W, De Vos R, Verstappen F, Puech-Pages V, Bécard G,Mulder P, Bouwmeester H (2008) Tomato strigolactones arederived from carotenoids and their biosynthesis is promoted byphosphate starvation. New Phytol 178:863–874, doi:10.1111/j.1469-8137.2008.02406.x
116. Lyons E, Pedersen B, Kane J, Freeling M (2008) The value ofnonmodel genomes and an example using SynMap within CoGeto dissect the hexaploidy that predates the Rosids. Trop PlantBiol 1: (this issue)
117. Mach JM, Castillo AR, Hoogstraten R, Greenberg JT (2001) TheArabidopsis accelerated cell death gene ACD2 encodes redchlorophyll catabolite reductase and suppresses the spread ofdisease symptoms. Proc Natl Acad Sci USA 98:771–776,doi:10.1073/pnas.021465298
118. Mackenzie S, McIntosh L (1999) Higher plant mitochondria.Plant Cell 11:571–586
119. MacLeod AJ, Pieris NM (1983) Volatile components of papaya(Carica papaya L.) with particular reference to glucosinolateproducts. J Agric Food Chem 31:1005–1008, doi:10.1021/jf00119a021
120. Madden TL, Tatusov RL, Zhang J (1996) Application of networkBLAST server. Methods Enzymol 266:131–141, doi:10.1016/S0076-6879(96)66011-X
121. Manenoi A, Bayogan ERV, Thumdee S, Paull RE (2007) Utilityof 1-methylcyclopropene as a papaya postharvest treatment.Postharvest Biol Technol 44:55–62, doi:10.1016/j.postharv-bio.2006.11.005
122. Manenoi A, Paull RE (2007) Papaya fruit softening, endoxyla-nase gene expression, protein and activity. Physiol Plant131:470–480, doi:10.1111/j.1399-3054.2007.00967.x
123. Marchler-Bauer A et al (2005) CDD: a Conserved DomainDatabase for protein classification. Nucleic Acids Res 33:192–196, doi:10.1093/nar/gki069
124. Martin T, Frommer WB, Salanoubat M, Willmitzer L (1993)Expression of an Arabidopsis sucrose synthase gene indicates arole in metabolization of sucrose both during phloem loadingand in sink organs. Plant J 4:367–377, doi:10.1046/j.1365-313X.1993.04020367.x
125. Marusek CM, Trobaugh NM, Flurkey WH, Inlow JK (2006)Comparative analysis of polyphenol oxidase from plant andfungal species. J Inorg Biochem 100:108–123, doi:10.1016/j.jinorgbio.2005.10.008
126. Mayer AM, Staples RC (2002) Laccase: new functions for an oldenzyme. Phytochem 60:551–565, doi:10.1016/S0031-9422(02)00171-1
127. McCaig BC, Meagher RB, Dean JFD (2005) Gene structure andmolecular analysis of the laccase-like multicopper oxidase(LMCO) gene family in Arabidopsis thaliana. Planta 221:619–636, doi:10.1007/s00425-004-1472-6
128. McDonald AE (2008) Alternative oxidase: an inter-kingdomperspective on the function and regulation of this broadlydistributed ‘cyanide-resistant’ terminal oxidase. Funct Plant Biol35:535–552, doi:10.1071/FP08025
129. McQueen-Mason S, Durachko DM, Cosgrove DJ (1992) Twoendogenous proteins that induce cell wall expansion in plants.Plant Cell 4:1425–1433
130. Melo AMP, Bandeiras TM, Teixeira M (2004) New Insights intoType II NAD(P)H:quinone oxidoreductases. Microbiol Mol BiolRev 68:603–616, doi:10.1128/MMBR.68.4.603-616.2004
131. Michalecka AM, Svensson AS, Johansson FI, Agius SC,Johanson U, Brennicke A, Binder S, Rasmusson AG (2003)Arabidopsis genes encoding mitochondrial type II NAD(P)Hdehydrogenases have different evolutionary origin and showdistinct responses to light. Plant Physiol 133:642–652,doi:10.1104/pp.103.024208
132. Ming R et al (2008) Genome of the transgenic tropical fruit treepapaya (Carica papaya Linnaeus). Nature 452:991–996,doi:10.1038/nature06856
133. Moore AL, Albury MS, Crichton PG, Affourtit C (2002) Functionof the alternative oxidase: is it still a scavenger? Trends Plant Sci7:478–481, doi:10.1016/S1360-1385(02)02366-X
134. Morshidi M (1996) Genetic variability in Carica papaya andrelated species. Dissertation, University of Hawaii at Manoa
135. Nakano T, Suzuki K, Fujimura T, Shinshi H (2006) Genome-wide analysis of the ERF gene family in Arabidopsis and Rice.Plant Physiol 140:411–432, doi:10.1104/pp.105.073783
136. Nakasone HY, Paull RE (1998) Tropical Fruits. CAB Interna-tional, Wallingford, England
137. Nakatsuka A, Murachi S, Okunishi H, Shiomi S, Nakano R, KuboY, Inaba A (1998) Differential expression and internal feedbackregulation of 1-aminocyclopropane-1-carboxylate synthase, of 1-aminocyclopropane-1-carboxylase oxidase and ethylene receptorgenes in tomato fruit during development and ripening. PlantPhysiol 118:1295–1305, doi:10.1104/pp.118.4.1295
138. Newcomb RD, Crowhurst RN, Gleave AP, Rikkerink EHA,Allan AC, Beuning LL, Bowen JH, Kim E, Jamieson R, JanssenBJ, Laing WA, McArtney S, Nain B, Ross GS, Snowden KC,Souleyre EJF, Walton EF, Yauk YK (2006) Analyses ofexpressed sequence tags from Apple. Plant Physiol 141:147–166, doi:10.1104/pp.105.076208
139. Noordermeer MA, Veldink GA, Vliegenthart JFG (1999) Alfalfacontains substantial hydroperoxide lyase activity and a 3Z:2E-enal isomerase. FEBS Lett 443:201–204, doi:10.1016/S0014-5793(98)01706-2
140. Oberg KA, Ruysschaert JM, Azarkan M, Smolders N, ZerhouniS, Wintjens R, Amrani A, Looze Y (1998) Papaya glutaminecyclase, a plant enzyme highly resistant to proteolysis, adopts anall-b conformation. Eur J Biochem 258:214–222, doi:10.1046/j.1432-1327.1998.2580214.x
274 Tropical Plant Biol. (2008) 1:246–277

141. Ouaked F, Rozhon W, Lecourieux D, Hirt H (2003) A MAPKpathway mediates ethylene signaling in plants. EMBO J22:1282–1288, doi:10.1093/emboj/cdg131
142. Paquette SM, Bak S, Feyereisen R (2000) Intron-exon organi-zation and phylogeny in a large superfamily, the paralogouscytochrome P450 genes of Arabidopsis thaliana. DNA Cell Biol19:307–317, doi:10.1089/10445490050021221
143. Passardi F, Longet D, Penel C, Dunand C (2004) The class IIIperoxidase multigenic family in rice and its evolution in landplants. Phytochem 65:1879–1893, doi:10.1016/j.phytochem.2004.06.023
144. Patrick JW (1997) Phloem unloading: sieve element unloadingand post-sieve element transport. Annu Rev Plant Physiol PlantMol Biol 48:191–222, doi:10.1146/annurev.arplant.48.1.191
145. Paull RE (1993) Pineapple and Papaya. In: Seymour G, Taylor J,Tucker G (eds) Biochemistry of Fruit Ripening. Chapman &Hall, London, United Kingdom, pp 291–323
146. Paull RE, Chen NJ (1983) Postharvest variation in cell wall-degrading enzymes of papaya (Carica papaya L.) during fruitripening. Plant Physiol 72:382–385
147. Paull RE, Chen NJ (1989) Waxing and plastic wraps influencewater loss from papaya fruit during storage and ripening. J AmSoc Hortic Sci 114:937–942
148. Phillips DR, Rasbery JM, Bartel B, Matsuda SPT (2006)Biosynthetic diversity in plant triterpene cyclization. Curr OpinPlant Biol 9:305–314, doi:10.1016/j.pbi.2006.03.004
149. Pichersky E, Gershenzon J (2002) The formation and function ofplant volatiles: perfumes for pollinator attraction and defense.Curr Opin Plant Biol 5:237–243, doi:10.1016/S1369-5266(02)00251-0
150. Picton S, Gray J, Barton S, AbuBakar U, Lowe A, Grierson D(1993) cDNA cloning and characterisation of nove1 ripening-related mRNAs with altered patterns of accumulation in theripening inhibitor (rin) tomato ripening mutant. Plant Mol Biol23:193–207, doi:10.1007/BF00021431
151. Pino JA, Almora K, Marbot R (2003) Volatile components ofpapaya (Carica papaya L., Maradol variety) fruit. FlavourFragrance J 18:492–496, doi:10.1002/ffj.1248
152. Piotrowski M, Schemenewitz A, Lopukhina A, Müller A,Janowitz T, Weiler EW, Oecking C (2004) Desulfoglucosinolatesulfotransferases from Arabidopsis thaliana catalyze the finalstep in the biosynthesis of the glucosinolate core structure. J BiolChem 279:50717–50725, doi:10.1074/jbc.M407681200
153. Pourcel L, Routaboul JM, Cheynier V, Lepiniec L, Debeaujon I(2006) Flavonoid oxidation in plants: from biochemical proper-ties to physiological functions. Trends Plant Sci 12:29–36,doi:10.1016/j.tplants.2006.11.006
154. Pruzinska A, Anders I, Aubry S, Schenk A, Tapernoux-Luthe E,Muller T, Krautler B, Hortensteiner S (2007) In vivo participa-tion of red chlorophyll catabolite reductase in chlorophyllbreakdown. Plant Cell 19:369–387, doi:10.1105/tpc.106.044404
155. Qiu YX, NishinaMS, Paull RE (1995) Papaya fruit growth, calciumuptake and fruit ripening. J Am Soc Hortic Sci 120:246–253
156. Raes J, Rohde A, Chrisrensen JH, Van der Peer Y, Boerjan W(2003) Genome-wide characterization of the lignification toolboxin Arabidopsis. Plant Physiol 133:1051–1071, doi:10.1104/pp.103.026484
157. Rask L, Andréasson E, Ekbom B, Eriksson S, Pontoppidan B,Meijer J (2000) Myrosinase: gene family evolution andherbivore defense in Brassicaceae. Plant Mol Biol 42:93–113,doi:10.1023/A:1006380021658
158. Rasmusson AG, Svensson AS, Knoop V, Grohmann L, Bren-nicke A (1999) Homologues of yeast and bacterial rotenone-insensitive NADH dehydrogenases in higher eukaryotes: twoenzymes are present in potato mitochondria. Plant J 20:79–87,doi:10.1046/j.1365-313X.1999.00576.x
159. Roberts TH, Fredlund KM, Moller IM (1995) Directevidence for the presence of two external NAD(P)Hdehydrogenases coupled to the electron transport chain inplant mitochondria. FEBS Lett 373:307–309, doi:10.1016/0014-5793(95)01059-N
160. Rodman JE, Karol KG, Price RA, Sytsma KJ (1996) Molecules,morphology, and Dahlgren’s expanded order Capparales. SystBot 21:289–307, doi:10.2307/2419660
161. Rodman JE, Soltis PS, Soltis DE, Sytsma KJ, Karol KG (1998)Parallel evolution of glucosinolate biosynthesis inferred fromcongruent nuclear and plastid gene phylogenies. Am J Bot85:997–1006, doi:10.2307/2446366
162. Rodrigo MJ, Marcos JF, Alférez F, Mallent MD, Zacarías L(2003) Characterization of Pinalate, a novel Citrus sinensismutant with a fruit-specific alteration that results in yellowpigmentation and decreased ABA content. J Exp Bot 54:727–738, doi:10.1093/jxb/erg083
163. Rodrigo MJ, Alquezar B, Zacarias L (2006) Cloning andcharacterization of two 9-cis-epoxycarotenoid dioxygenasegenes, differentially regulated during fruit maturation and understress conditions, from orange (Citrus sinensis L. Osbeck). J ExpBot 57:633–643, doi:10.1093/jxb/erj048
164. Rodríguez FI, Esch JJ, Hall AE, Binder BM, Schaller GE,Bleecker AB (1999) A copper cofactor for the ethylene receptorETR1 from Arabidopsis. Science 283:996–998, doi:10.1126/science.283.5404.996
165. Ronse de Craene LP, Haston E (2006) The systematic relation-ships of glucosinolate-producing plants and related families: acladistic investigation based on morphological and molecularcharacters. Bot J Linn Soc 151:453–494, doi:10.1111/j.1095-8339.2006.00580.x
166. Ronse de Craene LP, Smets EF (1999) The floral developmentand anatomy of Carica papaya (Caricaceae). Can J Bot 77:582–298, doi:10.1139/cjb-77-4-582
167. Rose JKC, Lee HH, Bennett AB (1997) Expression of adivergent expansin gene is fruit-specific and ripening regulated.Proc Natl Acad Sci USA 94:5955–5960, doi:10.1073/pnas.94.11.5955
168. Rose JKC, Cosgrove DJ, Albersheim P, Darvill AG, Bennett AB(2000) Detection of expansin proteins and activity during tomatofruit ontogeny. Plant Physiol 123:1583–1592, doi:10.1104/pp.123.4.1583
169. Rossetto MRM, Oliveira do Nascimento JR, Purgatto E, Fabi JP,Lajolo FM, Cordenunsi BR (2008) Benzylglucosinolate, benzy-lisothiocyanate, and myrosinase activity in papaya fruit duringdevelopment and ripening. J Agric Food Chem 56:9592–9599,doi:10.1021/jf801934x
170. Roth I, Clausnitzer I (1972) Desarrollo y anatomía del fruto y lasemilla de Carica papaya L. (Lechosa). Acta Bot Venez 7:187–206
171. Saito N, Nei M (1987) The neighbor-joiningmethod: a newmethodfor constructing phylogenetic trees. Mol Biol Evol 4:406–425
172. Salas CE, Gomes MTR, Hernandez M, Lopes MTP (2008) Plantcysteine proteinases: Evaluation of the pharmacological activity.Phytochem 69:2263–2269, doi:10.1016/j.phytochem.2008.05.016
173. Sampedro J, Lee Y, Carey RE, de Pamphilis C, Cosgrove DJ(2005) Use of genomic history to improve phylogeny andunderstanding of births and deaths in a gene family. Plant J44:409–419, doi:10.1111/j.1365-313X.2005.02540.x
174. Sampedro J, Carey RE, Cosgrove DJ (2006) Genome historiesclarify evolution of the expansin superfamily: new insights fromthe poplar genome and pine ESTs. J Plant Res 119:11–21,doi:10.1007/s10265-005-0253-z
175. Sangwanangkul P, Paull RE (2005) The role of hexosetransporter in sugar accumulation of papaya fruit duringmaturation and ripening. Acta Hortic 740:313–316
Tropical Plant Biol. (2008) 1:246–277 275275

176. Schaller GE, Kieber JJ (2002) Ethylene. In: Somerville C,Meyerowitz E (eds) The Arabidopsis Book. Rockville, MD:American Society of Plant Biologists
177. Schledz M, al-Babili S, von Lintig J, Haubruck H, Rabbani S,Kleinig H, Beyer P (1996) Phytoene synthase from Narcissuspseudonarcissus: functional expression, galactolipid requirement,topological distribution in chromoplasts and induction duringflowering. Plant J 10:781–792, doi:10.1046/j.1365-313X.1996.10050781.x
178. Schonbaum GR, Bonner WD Jr, Storey BT, Bahr JT (1971)Specific inhibition of the cyanide-insensitive respiratory pathwayin plant mitochondria by hydroxamic acids. Plant Physiol47:124–128
179. Schreier P, Lehr M, Heidlas J, Idstein H (1985) Volatiles frompapaya (Carica papaya L.) fruit: indication of precursors ofterpene compounds. A Lebensm - Unters Forsch 180:297–302
180. Schwab W, Davidovich-Rikanati R, Lewinsohn E (2008)Biosynthesis of plant-derived flavor compounds. Plant J54:712–732, doi:10.1111/j.1365-313X.2008.03446.x
181. Silva E, Lourenço EJ, Neves VA (1990) Soluble and boundperoxidase from papaya fruit. Phytochem 29:1051–1056,doi:10.1016/0031-9422(90)85401-Z
182. Shindo T, Van Der Hoorn RAL (2008) Papain-like cysteineproteases: key players at molecular battlefields employed by bothplants and their invaders. Mol Plant Pathol 9:119–125
183. Simkin A, Schwartz S, Auldridge M, Taylor M, Klee HJ (2004)The tomato carotenoid cleavage dioxygenase 1 genes contribute tothe formation of the flavor volatiles B-ionone, pseudoionone andgeranylacetone. Plant J 40:882–892, doi:10.1111/j.1365-313X.2004.02263.x
184. Small I, Peeters N, Legeai F, Lurin C (2004) Predotar: a tool forrapidly screening proteomes for N-terminal targeting sequences.Proteomics 4:1581–1590, doi:10.1002/pmic.200300776
185. Speirs J, Lee E, Holt K, Yong-Duk K, Scott NS, Loveys B,Schuch W (1998) Genetic manipulation of alcohol dehydroge-nase levels in ripening tomato fruit affects the balance of someflavor aldehydes and alcohols. Plant Physiol 117:1047–1058,doi:10.1104/pp.117.3.1047
186. Stepanova AN, Alonso JM (2005) Ethylene Signaling Pathway.Sci STKE 276:cm3, doi:10.1126/stke.2762005cm3
187. Storey WB (1969) Papaya (Carica papaya L.). In: Ferwerda P,Wit F (eds) Outline of perennial crop breeding in the tropics. H.Veenman & Zonen, Wageningen, pp 389–407
188. Sturm A (1999) Invertases. Primary structures, functions, androles in plant development and sucrose partitioning. PlantPhysiol 121:1–8, doi:10.1104/pp.121.1.1
189. Sturm A, Tang G-Q (1999) The sucrose-cleaving enzymes ofplants are crucial for development, growth and carbon partition-ing. Trends Plant Sci 4:401–407, doi:10.1016/S1360-1385(99)01470-3
190. Takita MA, Berger IJ, Basílio-Palmieri AC, Borges KM, DeSouza JM, Targon MLNP (2007) Terpene production in the peelof sweet orange fruits. Genet Mol Biol 30:841–847, doi:10.1590/S1415-47572007000500012
191. Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4:molecular evolutionary genetics analysis (MEGA) softwareversion 4.0. Mol Biol Evol 24:1596–1599, doi:10.1093/molbev/msm092
192. Tan SC, Lam PF (1985) Effect of gamma irradiation on PALactivity and phenolic compounds in papaya (Carica papaya L.)and mango (Mangifera inidca L.) fruits. ASEAN Food J. 1:134–136
193. Tang CS (1970) Benzyl isothiocyanate of papaya fruit. Phyto-chem 10:117–121, doi:10.1016/S0031-9422(00)90258-9
194. Taylor MAJ, Al-sheikh M, Revell DF, Sumner IG, Connerton IF(1999) cDNA cloning and expression of Carica papaya prochy-
mopapain isoforms in Escherichia coli. Plant Sci 145:41–47,doi:10.1016/S0168-9452(99)00068-0
195. Theologis A, Laties GG (1978) Respiratory contribution of thealternate path during various stages of ripening in avocado andbanana fruits. Plant Physiol 62:249–255
196. Thipyapong P, Joel DM, Steffens JC (1997) Differentialexpression and turnover of the tomato polyphenol oxidase genefamily during vegetative and reproductive development. PlantPhysiol 113:707–718
197. Thipyapong P, Hunt MD, Steffens JC (2004) Antisense down-regulation of polyphenol oxidase results in enhanced diseasesusceptibility. Planta 220:105–117, doi:10.1007/s00425-004-1330-6
198. Tholl D (2006) Terpene synthases and the regulation, diversityand biological roles of terpene metabolism. Curr Opin Plant Biol9:297–304, doi:10.1016/j.pbi.2006.03.014
199. Tieman DM, Loucas HM, Kim JY, Clark DG, Klee HJ (2007)Tomato phenylacetaldehyde reductases catalyze the last step inthe synthesis of the aroma volatile 2-phenylethanol. Phytochem68:2660–2669, doi:10.1016/j.phytochem.2007.06.005
200. Tompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res 22:4673–4680, doi:10.1093/nar/22.22.4673
201. Trentmann SM, Kende H (1995) Analysis of Arabidopsis cDNAthat shows homology to the tomato E8 cDNA. Plant Mol Biol29:161–166, doi:10.1007/BF00019127
202. Tsuchisaka A, Yamagami T, Alonso JM, Ecker JR, Theologis A(2007) The Arabidopsis 1-Aminocyclopropane-1-CarboxylicAcid Synthase (ACS) Gene Family. Conference Proc, 18thInternatl Conference on Arabidopsis Research, 2007. TAIRPublication:501722104
203. Tymowska-Lalanne Z, Kreis M (1998) Expression of theArabidopsis thaliana invertase gene family. Planta 207:259–265, doi:10.1007/s004250050481
204. Valerio L, De Meyer M, Penel C, Dunand C (2004) Expressionanalysis of the Arabidopsis peroxidase multigenic family.Phytochem 65:1331–1342, doi:10.1016/j.phytochem.2004.04.017
205. Van Droogenbroeck B, Breyne P, Goetghebeur P, Romeijn-Peeters E, Kyndt T, Gheysen G (2002) AFLP analysis of geneticrelationships among papaya and its wild relatives (Caricaceae)from Ecuador. TAG 105:289–297, doi:10.1007/s00122-002-0983-4
206. Vanlerberghe GC, McIntosh L (1997) Alternative oxidase: Fromgene to function. Annu Rev Plant Physiol Plant Mol Biol48:703–734, doi:10.1146/annurev.arplant.48.1.703
207. Vogel JT, Tan B-C, McCarty DR, Klee HJ (2008) Thecarotenoid cleavage dioxygenase 1 enzyme has broad substratespecificity, cleaving multiple carotenoids at two different bondpositions. J Biol Chem 283:11364–11373, doi:10.1074/jbc.M710106200
208. Wang W, Hall AE, O’Malley R, Bleecker AB (2003) Canonicalhistidine kinase activity of the transmitter domain of the ETR1ethylene receptor from Arabidopsis is not required for signaltransmission. Proc Natl Acad Sci USA 100:352–357, doi:10.1073/pnas.0237085100
209. Wang Y, Tang XM, Cheng ZK, Mueller L, Giovannoni J,Tanksley SD (2006) Euchromatin and pericentromeric hetero-chromatin: Comparative composition in the tomato genome.Genetics 172:2529–2540, doi:10.1534/genetics.106.055772
210. Watling JR, Robinson SA, Seymour RS (2006) Contribution ofthe alternative pathway to respiration during thermogenesis inflowers of the sacred lotus. Plant Physiol 140:1367–1373,doi:10.1104/pp.105.075523
276 Tropical Plant Biol. (2008) 1:246–277

211. Wittstock U, Halkier BA (2002) Glucosinolate research in theArabidopsis era. Trends Plant Sci 7:263–270, doi:10.1016/S1360-1385(02)02273-2
212. Wu G-L, Zhang X-Y, Zhang L-Y, Pan Q-H, Shen Y-Y, Zhang D-P(2004) Phloem unloading in developing walnut fruit is symplasmicin the seed pericarp and apoplasmic in the fleshy pericarp. PlantCell Physiol 45:1461–1470, doi:10.1093/pcp/pch169
213. Wüthrich KL, Bovet L, Hunziker PE, Donnison IS, HörtensteinerS (2000) Molecular cloning, functional expression and character-isation of RCC reductase involved in chlorophyll catabolism.Plant J 21:189–198, doi:10.1046/j.1365-313x.2000.00667.x
214. Xiao H, Jiang N, Schaffner E, Stockinger EJ, van der Knaap E(2008) A retrotransposon-mediated gene duplication underliesmorphological variation of tomato fruit. Science 319:1527–1530,doi:10.1126/science.1153040
215. Xue J, Jørgensen M, Pihlgren U, Rask L (1995) The myrosinasegene family in Arabidopsis thaliana: gene organization, expres-sion and evolution. Plant Mol Biol 27:911–922, doi:10.1007/BF00037019
216. Yabumoto K, Yamaguchi M, Jennings WG (1977) Biosynthesis ofsome volatiles constituents of `muskmelon Cucumis melo. Chemi-sche Mikrobiologische Technologische Lebensraum 5:53–56
217. Yamagami T, Tsuchisaka A, Yamada K, Haddon WF, HardenLA, Theologis A (2003) Biochemical diversity among the 1-amino-cyclopropane-1-carboxylate synthase isozymes encodedby the Arabidopsis gene family. J Biol Chem 278:49102–49112,doi:10.1074/jbc.M308297200
218. Yamamoto HY (1964) Comparison of the carotenoids in yellow-and red-fleshed Carica papaya. Nature 201:1049–1050, doi:10.1038/2011049a0
219. Yip WK, Dong JG, Kenny JW, Thompson GA, Yang SF (1990)Characterization and sequencing of the active site of 1-amino-cyclopropane-1-carboxylate synthase. Proc Natl Acad Sci USA87:7930–7934, doi:10.1073/pnas.87.20.7930
220. Yoo A, Seo YS, Jung JW, Sung SK, Kim WT, Lee W, Yang DR(2006) Lys296 and Arg299 residues in the C-terminus of MD-ACO1 are essential for a 1-aminocyclopropane-1-carboxylateoxidase enzyme activity. J Struct Biol 156:407–420, doi:10.1016/j.jsb.2006.08.012
221. Zeevart JAD, Creelman RA (1988) Metabolism and physiologyof abscisic acid. Ann Rev Plant Physiol Plant Mo1 Biol 39:439–473
222. Zhang L-Y, Peng Y-B, Pelleschi-Travier S, Fan Y, Lu Y-F, Lu Y-M, Gao X-P, Shen Y-Y, Delrot S, Zhang D-P (2004) Evidencefor apoplasmic phloem unloading in developing apple fruit. PlantPhysiol 135:574–586, doi:10.1104/pp.103.036632
223. Zhang X-Y, Wang X-L, Wang X-F, Xia G-H, Pan Q-H, Fan R-C,Wu F-Q, Yu X-C, Zhang D-P (2006) A shift of phloemunloading from symplasmic to apoplasmic pathway is involvedin developmental onset of ripening in grape berry. Plant Physiol142:220–232, doi:10.1104/pp.106.081430
224. Zhou LL, Paull RE (2001) Sucrose metabolism during papaya(Carica papaya) fruit growth and ripening. J Am Soc Hortic Sci126:351–357
225. Zhou L, Chen CC, Ming R, Christopher DA, Paull RE (2003)Apoplastic invertase and its enhanced expression and post-translational control during papaya fruit maturation and ripening.J Am Soc Hortic Sci 128:628–635
226. Zimmerman DC, Vick BA, Borg TK (1974) Intracellulardistribution of hydroperoxide isomerase. Plant Physiol 53:1–4
Tropical Plant Biol. (2008) 1:246–277 277277




















