
Chlorophyll fluorescence as a tool to evaluate the ripening of ‘Golden’ papaya fruit
11
Postharvest Biology and Technology 33 (2004) 163–173 Chlorophyll fluorescence as a tool to evaluate the ripening of ‘Golden’ papaya fruit Ilana Urbano Bron a , Rafael Vasconcelos Ribeiro b,∗ , Marisa Azzolini b , Angelo Pedro Jacomino a , Eduardo Caruso Machado c a Departamento de Produção Vegetal, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418-900 Piracicaba, SP, Brazil b Departamento de Ciˆ encias Biológicas, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418-900 Piracicaba, SP, Brazil c Centro de Pesquisa e Desenvolvimento em Ecofisiologia e Biof´ ısica, Instituto Agronˆ omico de Campinas, 13001-970 Campinas, SP, Brazil Received 25 September 2003; accepted 14 February 2004 Abstract Fruit classification based on one or more physical or physiological attributes is important to improve quality during storage and marketing. Our objectives were to evaluate changes in chlorophyll fluorescence of papaya fruit (Carica papaya L. cv. Golden) at different ripeness stages and during the ripening to determine if this non-destructive technique might assist the evaluation of fruit ripeness. Maximal (F m ), minimal (F o ), variable (F v ), variable:maximal (F v /F m ) and maximal:minimal (F m /F o ) chlorophyll fluorescence, fruit firmness, and skin color (H ◦ ) were evaluated at three ripeness stages and during the ripening of papaya fruit. Simple and multiple empirical regressions assessed the relationships among all measured parameters. The three ripeness stages were differentiated using the chlorophyll fluorescence parameters. All chlorophyll fluorescence parameters declined with the progress of fruit ripening, where F m and F o showed the best correlation with changes in skin color and fruit firmness. This suggests a relationship between the fluorescence and the level of papaya fruit senescence. Chlorophyll fluorescence decline was probably due to chlorophyll degradation and also to the degree of chloroplast membrane injury due to fruit ripeness and senescence. Chlorophyll fluorescence associated with chloroplast function, declined in parallel with the decline of other quality characteristics during papaya fruit ripening. The use of a single parameter to determine the overall quality of papaya fruit may be unreliable, and the chlorophyll fluorescence technique may be a helpful, complementary, non-destructive technology that can be combined with other quality measurement techniques. © 2004 Elsevier B.V. All rights reserved. Keywords: Carica papaya; Firmness; Fruit quality; Non-destructive testing; Ripening ∗ Corresponding author. Tel.: +55-19-3429-4458; fax: +55-19-3434-8295. E-mail address: [email protected] (R. Vasconcelos Ribeiro). 1. Introduction During sorting of papaya at packing-houses, each fruit is generally segregated according to size and vi- sual skin color. However, this classification does not 0925-5214/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.postharvbio.2004.02.004
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Chlorophyll fluorescence as a tool to evaluate the ripening of ‘Golden’ papaya fruit

Postharvest Biology and Technology 33 (2004) 163–173
Chlorophyll fluorescence as a tool to evaluate theripening of ‘Golden’ papaya fruit
Ilana Urbano Brona, Rafael Vasconcelos Ribeirob,∗, Marisa Azzolinib,Angelo Pedro Jacominoa, Eduardo Caruso Machadoc
a Departamento de Produção Vegetal, Escola Superior de Agricultura “Luiz de Queiroz”,Universidade de São Paulo, 13418-900 Piracicaba, SP, Brazil
b Departamento de Ciˆencias Biológicas, Escola Superior de Agricultura “Luiz de Queiroz”,Universidade de São Paulo, 13418-900 Piracicaba, SP, Brazil
c Centro de Pesquisa e Desenvolvimento em Ecofisiologia e Biof´ısica, Instituto Agronˆomico de Campinas,13001-970 Campinas, SP, Brazil
Received 25 September 2003; accepted 14 February 2004
Abstract
Fruit classification based on one or more physical or physiological attributes is important to improve quality during storage andmarketing. Our objectives were to evaluate changes in chlorophyll fluorescence of papaya fruit (Carica papayaL. cv. Golden)at different ripeness stages and during the ripening to determine if this non-destructive technique might assist the evaluation offruit ripeness. Maximal (Fm), minimal (Fo), variable (Fv), variable:maximal (Fv/Fm) and maximal:minimal (Fm/Fo) chlorophyllfluorescence, fruit firmness, and skin color (H◦) were evaluated at three ripeness stages and during the ripening of papaya fruit.Simple and multiple empirical regressions assessed the relationships among all measured parameters. The three ripeness stageswere differentiated using the chlorophyll fluorescence parameters. All chlorophyll fluorescence parameters declined with theprogress of fruit ripening, whereFm andFo showed the best correlation with changes in skin color and fruit firmness. Thissuggests a relationship between the fluorescence and the level of papaya fruit senescence. Chlorophyll fluorescence declinewas probably due to chlorophyll degradation and also to the degree of chloroplast membrane injury due to fruit ripeness andsenescence. Chlorophyll fluorescence associated with chloroplast function, declined in parallel with the decline of other qualitycharacteristics during papaya fruit ripening. The use of a single parameter to determine the overall quality of papaya fruit maybe unreliable, and the chlorophyll fluorescence technique may be a helpful, complementary, non-destructive technology that canbe combined with other quality measurement techniques.© 2004 Elsevier B.V. All rights reserved.
Keywords: Carica papaya; Firmness; Fruit quality; Non-destructive testing; Ripening
∗ Corresponding author. Tel.:+55-19-3429-4458;fax: +55-19-3434-8295.
E-mail address:[email protected] (R. Vasconcelos Ribeiro).
1. Introduction
During sorting of papaya at packing-houses, eachfruit is generally segregated according to size and vi-sual skin color. However, this classification does not
0925-5214/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.postharvbio.2004.02.004

164 I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173
consider intrinsic characteristics such as fruit firmness,sugar content, and flavor (Song et al., 1997). Thus, thecapacity to accurately classify fruit is limited whenonly external attributes are evaluated. On the otherhand, the technique to measure fruit firmness is de-structive and it could lead to inadequate evaluation offruit lots inasmuch as this technique is only suitablefor small samples. Non-destructive technologies to as-sess one or more attributes of quality can be helpfulto sort fruit correctly and thereby improve the qualityof fruit delivered to the consumer (Song et al., 1997).
Chlorophyll fluorescence is a technique character-ized by relatively fast measurement and with greatprecision (DeEll et al., 1999). Since chlorophyll flu-orescence at room temperature is almost exclusivelyoriginated from light-excited chlorophylla moleculesassociated with photosystem II (PSII), it reflectsprimary photosynthetic reactions in thylakoid mem-branes (Papargeorgiou, 1975; Schreiber and Bilger,1987). After light absorption, excited chlorophyllmolecules can return to the ground level (de-excited)by fluorescence emission, excitation transfer to neigh-boring molecules, photochemical reaction, and bythermal dissipation (Krause and Weis, 1991; Schreiberet al., 1994, 1998). The fluorescence is a small portionof absorbed energy re-irradiated and not used in pho-tochemical reactions of photosynthesis (Krause andWeis, 1991; Maxwell and Johnson, 2000). Minimalfluorescence is referred asFo (PSII is able to pass onalmost all of the electrons excited by light). When thefluorescence intensity is at a maximum it is referredasFm (PSII is able to pass on only a fraction of theelectrons excited by light). Thus, the variable fluores-cence (Fv) expresses the difference betweenFm andFo (Fv = Fm − Fo), permitting the calculation of thechlorophyll fluorescence parameterFv/Fm, which isa measure of the efficiency in energy transfer processand chloroplast activity (van Kooten and Snel, 1990).
Chlorophyll fluorescence has been used as anon-intrusive indicator of in vivo photosynthetic re-actions and environmental stresses (Schreiber et al.,1994; Gray et al., 1997) of various crop plants. Sinceprotein–chlorophyll complexes are associated withthe thylakoid membrane, the fluorescence is an intrin-sic indicator of the fluidity, stability, and organizationof membranes (Schreiber and Bilger, 1987). WhenPSII is damaged by some kind of stress, the fluores-cence characteristics are altered. Thus, chlorophyll
fluorescence is an indirect measurement of the phys-iological status of green tissues (Krause and Weis,1984, 1991; Schreiber and Bilger, 1987; Maxwell andJohnson, 2000), being used in both green leaves andseveral chlorophyll-containing fruits (Smillie et al.,1987; Song et al., 1997; DeEll et al., 1998; Mir et al.,1998b; DeEll and Toivonen, 2000). Concerning theevaluation of changes in fruit tissues, chlorophyllfluorescence measurement has the advantage of de-tecting cellular injury due to natural senescence orenvironmental stresses in advance to the developmentof visible symptoms (Smillie et al., 1987; Sanxteret al., 1992; DeEll et al., 1996, 1998).
Fruit ripening and senescence processes may affectthe chlorophyll fluorescence yield by loss of pho-tosynthetic activity and by decrease in chlorophyllcontent due to reduced PSII activity and increasedchlorophyllase activity, respectively (Smillie et al.,1987; Sanxter et al., 1992; Tucker, 1993). Decline inchlorophyll fluorescence during the ripening of ba-nana and mango is related to the loss of chlorophyllcontent and chloroplast competence (Smillie et al.,1987). During apple ripening, fluorescence declinecorrelates well with fruit firmness and hue angle,and appears to reflect some loss of chloroplast func-tion with advancing senescence (Mir et al., 1998b).Chlorophyll fluorescence has been used to evalu-ate injuries caused by low temperatures in mango(Smillie et al., 1987), cucumber (van Kooten et al.,1992) and apples (DeEll et al., 1996; Mir et al.,1998a), physiological disorders related to low oxygenand high carbon dioxide concentrations (DeEll et al.,1998), and low-oxygen tolerance (Prange et al., 2002,2003). In addition,DeEll et al. (1996)suggested thatFv at harvest might be a tool to predict the scalddevelopment in ‘Golden Delicious’ apples.
However, one should be careful in assigningchanges in chlorophyll fluorescence as being dueto ripening, considering the complexity existing be-tween PSII activity and the complete stress response(van Kooten et al., 1992). In fact, some studies haveshown that sample size, cultivars (DeEll et al., 1997)and environmental conditions (Song et al., 1997)during ripening and senescence can affect the rela-tionship between chlorophyll fluorescence emissionand fruit quality characteristics, such as firmness.These findings show that measurements of chloro-phyll fluorescence, if used in conjunction with other

I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173 165
conventional quality evaluations such as skin color,appear to be a promising tool to evaluate the ripeningand senescence of chlorophyll-containing fruit.
One of the major problems of papaya fruit af-ter harvest is the irregular ripening of fruit of thesame lot, as a consequence of inadequate sorting atpacking-houses. Similarly, in apple skin color char-acteristics does not always accurately predict fruitquality (Song et al., 1997). Trying to minimize theseproblems, this study was designed to (i) determinepossible relationships between chlorophyll fluores-cence parameters and fruit quality indices, such asskin color and fruit firmness and (ii) provide a de-scription of chlorophyll fluorescence changes duringripening of papaya fruit.
2. Material and methods
2.1. Plant material
‘Golden’ papaya fruit were harvested from a com-mercial orchard on Linhares County, ES, Brazil, inJanuary 2002. The fruit were transported by refrig-erated truck at 10◦C for 20 h to the Laboratory ofPostharvest Physiology (“Luiz de Queiroz” College ofAgriculture, Piracicaba, SP, Brazil) where each fruitwas grouped according to its size and color.
2.2. Chlorophyll fluorescence measurements
A pulse amplitude-modulated fluorometer (FMS1,Hansatech, King’s Lynn, UK) was used to obtain theminimal and maximal chlorophyll fluorescence (Foand Fm, respectively), variable chlorophyll fluores-cence (Fv = Fm − Fo), and potential quantum yieldof photosystem II (Fv/Fm) (terminology according tovan Kooten and Snel, 1990). Before chlorophyll fluo-rescence measurements, fruits were dark-adapted for30 min using a dark towel. The fiber optic was fit-ted with a fiber-optic adapter to fix the distance be-tween the fiber optic terminus and the fruit exocarp.Fowas measured with a measuring beam at a light in-tensity less than 0.05�mol m−2 s−1. Fm was obtainedby measuring chlorophyll fluorescence during a 2.5-spulse of saturating light (18 000�mol m−2 s−1). Mea-surements were taken in two opposite positions of eachfruit at the same location in the fruit surface and then
averaged. This same procedure was applied to fruitfirmness and skin color measurements.
2.3. Fruit firmness and skin color
The firmness of fruit was measured manually us-ing a digital penetrometer (53200-Samar, Tr-Turoni,Forli, Italy) fitted with an 8-mm diameter probe. Oneskin disc was removed at the previously marked posi-tion (same position where the chlorophyll fluorescenceand skin color measurements were taken). Readingson firmness were recorded as newtons. Skin color of‘Golden’ papaya fruit was measured as hue angle (H◦)with a colorimeter (Minolta Chromameter 300, Mi-nolta Camera Co., Osaka, Japan).
2.4. Experiment I—assessment of the relationshipsbetween chlorophyll fluorescence, skin color andfruit firmness
This experiment was divided in three parts. Firstlywe used fruit sorted into three different ripenessstages based on the skin color: upon detection of ini-tial yellow color formation (S1), three-quarters yellow(S2), and full yellow (S3). Ten papaya fruits of eachripeness stage were used to evaluate chlorophyll fluo-rescence, fruit firmness, and skin color. In the secondpart of this experiment all characteristics stated abovewere evaluated for fruit of the same ripeness stagefrom two different companies. The third portion ofthe study focused on establishing a relationship be-tween chlorophyll fluorescence, skin color and fruitfirmness on 57 fruits of various ripeness stages usingmultiple regressions. These relationships were testedusing the measurements for fruit from the first part ofthis experiment.
2.5. Experiment II—evaluation of changes inchlorophyll fluorescence during ripening
Ten papaya fruits of the same ripeness stage (S1)were maintained at room conditions to simulatecommercial conditioning where the air temperature(24± 1◦C) and the relative humidity (80± 4%) weremonitored with a psychrometer (Marin et al., 2001).Changes in chlorophyll fluorescence and skin colorwere measured during the ripening at 24-h intervalsfor 7 days. Fruit firmness was measured only at the

166 I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173
end of the period (7th day). Each papaya fruit wasnumbered and all measurements were taken at thesame marked surface point on each half of 10 fruitsto ensure that subsequent readings would be takenfrom the same tissues.
2.6. Data analyses
The experimental design was completely ran-domized, with replications varying according to theexperiment. Data were analyzed using the ANOVAprocedure and the Tukey test (P < 0.05) to comparemeans (ESTAT, version 2.0, UNESP-FCAV, Jabotica-bal, SP, Brazil). Fruit firmness and skin color regressedagainst chlorophyll fluorescence parameters (Fv/Fm,Fo, Fm, andFv) and the relationships between chloro-phyll fluorescence parameters, skin color and fruitfirmness (multiple regressions) were tested for sig-nificance using commercial software (Origin, version6.0, Microcal Software, Northampton, MA, USA).
3. Results and discussion
3.1. Experiment I—assessing the relationshipsamong papaya fruit characteristics
3.1.1. Part I: fruit characteristics and ripeness stagesThe three ripeness stages were differentiated using
both hue angle and chlorophyll fluorescence param-eters, but not fruit firmness, which was more vari-able (Fig. 1A–D). Since the firmness is an importantattribute of fruit quality and its changes during fruitripening are associated with cell wall dissociation un-der the control of many enzymes (Huber, 1983), wecould expect a higher variation. Fruit firmness showedthe highest CV (43.8%) resulting in no statistical dif-ference between S2 and S3, even though S2 firmnesswas approximately twice S3 firmness (Fig. 1A). Giventhe high variability of this assessment, only firmnessresults would be unreliable for larger fruit population.In addition, several factors such as excessive moistureloss (shrivel) and injury influence the correct evalua-tion of fruit firmness.
The values ofFv/Fm, Fm, Fo, Fv, and H◦ de-creased with the ripeness stage increase (P < 0.05)(Fig. 1B–D). Generally, decreases inH◦ result fromgreenness loss due to chlorophyll degradation during
0
20
40
60
80
100
120
140
bb
a(B)(A)
Fru
it f
irm
ness
(N
)
70
80
90
100
110
120
c
b
a
Hue
ang
le (
Hº)
S1 S2 S30.3
0.4
0.5
0.6
0.7
c
b
a(C)
Fv/F
m
Maturity stages
S1 S2 S30
100
200
300
400
500
600
700
c ccb
b
ba
a
a
(D)
Chl
orop
hyll
fluo
resc
ence
(re
l. un
its)
Maturity stages
Minimal (Fo) Variable (Fv) Maximal (Fm)
Fig. 1. Fruit firmness (A), hue angle (B), and chlorophyll fluores-cence parameters (C and D) at three ripeness stages of ‘Golden’papaya fruit: upon detection of initial yellow color formation (S1),three-quarters yellow (S2), and full yellow (S3). Different lettersrepresent statistical differences between ripeness stages (n = 10)by the Tukey test (P < 0.05).
ripening (Holden, 1961; Tucker, 1993). It is reason-able to expect that chlorophyll fluorescence declineswhen chlorophyll content decreases in papaya fruit, asreported bySanxter et al. (1992). However, it wouldbe also due to membrane degradation during ripening(Marangoni et al., 1996). The drop inFv/Fm, Fm, Fo,andFv also reflect a loss in chloroplast function withadvancing ripening and senescence as reported forapple (Mir et al., 1998b).
The decrease inFv/Fm was associated with de-creases in bothFo and Fm. These parameters repre-sent the fluorescence yield when all reaction centers ofPSII are open and closed, respectively (van Kooten andSnel, 1990). Decreases inFm indicate an increase inheat formation in both the reaction center and antennaof PSII (Schreiber et al., 1998) whereas changes inFoare due to structural alterations in the thylakoid mem-branes (Kitao et al., 2000). Therefore, changes inFmandFo parameters reported here suggest losses occurin chlorophyll content and photosynthetic membraneactivity with advancing ripeness/senescence stages ofpapaya fruit.
I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173 167
80
90
100
110
120
0.3
0.4
0.5
0.6
0.7
0 30 60 90 120 150 180
0
150
300
450
600
750
(A)
Hue
ang
le (
Hº)
(B)
Fv/F
m
(C)Chl
orop
hyll
fluo
resc
ence
(rel
. uni
ts)
Fruit firmness (N)
Minimal (Fo)
Maximal (Fm)
Variable (Fv)
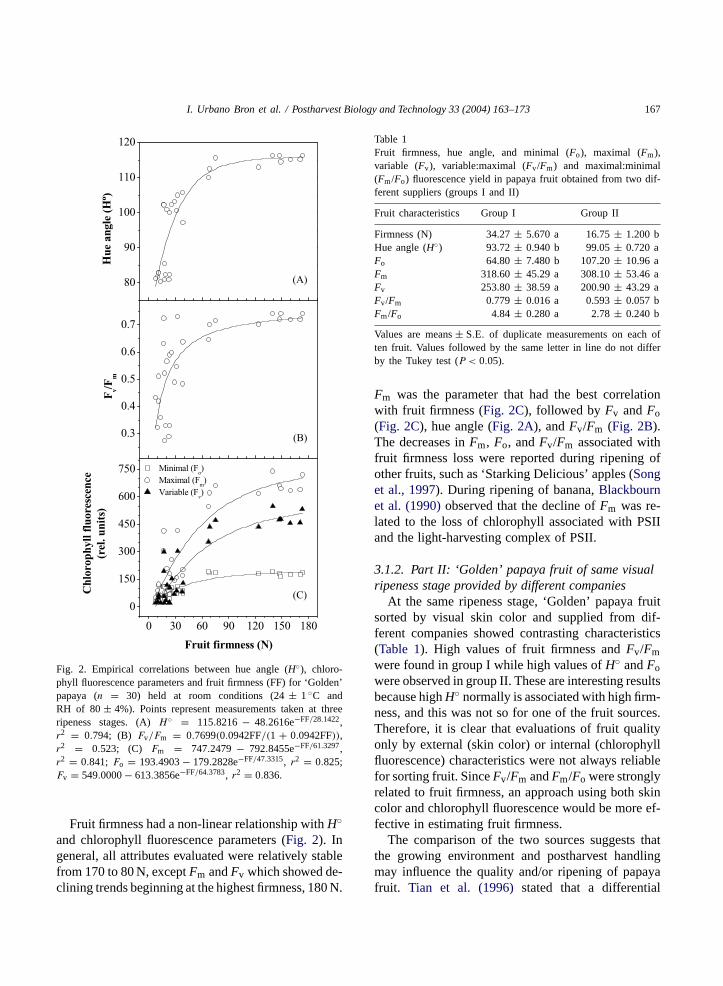
Fig. 2. Empirical correlations between hue angle (H◦), chloro-phyll fluorescence parameters and fruit firmness (FF) for ‘Golden’papaya (n = 30) held at room conditions (24± 1◦C andRH of 80± 4%). Points represent measurements taken at threeripeness stages. (A)H◦ = 115.8216 − 48.2616e−FF/28.1422,r2 = 0.794; (B) Fv/Fm = 0.7699(0.0942FF/(1 + 0.0942FF)),r2 = 0.523; (C) Fm = 747.2479 − 792.8455e−FF/61.3297,r2 = 0.841; Fo = 193.4903− 179.2828e−FF/47.3315, r2 = 0.825;Fv = 549.0000− 613.3856e−FF/64.3783, r2 = 0.836.
Fruit firmness had a non-linear relationship withH◦and chlorophyll fluorescence parameters (Fig. 2). Ingeneral, all attributes evaluated were relatively stablefrom 170 to 80 N, exceptFm andFv which showed de-clining trends beginning at the highest firmness, 180 N.
Table 1Fruit firmness, hue angle, and minimal (Fo), maximal (Fm),variable (Fv), variable:maximal (Fv/Fm) and maximal:minimal(Fm/Fo) fluorescence yield in papaya fruit obtained from two dif-ferent suppliers (groups I and II)
Fruit characteristics Group I Group II
Firmness (N) 34.27± 5.670 a 16.75± 1.200 bHue angle (H◦) 93.72± 0.940 b 99.05± 0.720 aFo 64.80± 7.480 b 107.20± 10.96 aFm 318.60± 45.29 a 308.10± 53.46 aFv 253.80± 38.59 a 200.90± 43.29 aFv/Fm 0.779± 0.016 a 0.593± 0.057 bFm/Fo 4.84 ± 0.280 a 2.78± 0.240 b
Values are means± S.E. of duplicate measurements on each often fruit. Values followed by the same letter in line do not differby the Tukey test (P < 0.05).
Fm was the parameter that had the best correlationwith fruit firmness (Fig. 2C), followed byFv andFo(Fig. 2C), hue angle (Fig. 2A), andFv/Fm (Fig. 2B).The decreases inFm, Fo, andFv/Fm associated withfruit firmness loss were reported during ripening ofother fruits, such as ‘Starking Delicious’ apples (Songet al., 1997). During ripening of banana,Blackbournet al. (1990)observed that the decline ofFm was re-lated to the loss of chlorophyll associated with PSIIand the light-harvesting complex of PSII.
3.1.2. Part II: ‘Golden’ papaya fruit of same visualripeness stage provided by different companies
At the same ripeness stage, ‘Golden’ papaya fruitsorted by visual skin color and supplied from dif-ferent companies showed contrasting characteristics(Table 1). High values of fruit firmness andFv/Fmwere found in group I while high values ofH◦ andFowere observed in group II. These are interesting resultsbecause highH◦ normally is associated with high firm-ness, and this was not so for one of the fruit sources.Therefore, it is clear that evaluations of fruit qualityonly by external (skin color) or internal (chlorophyllfluorescence) characteristics were not always reliablefor sorting fruit. SinceFv/Fm andFm/Fo were stronglyrelated to fruit firmness, an approach using both skincolor and chlorophyll fluorescence would be more ef-fective in estimating fruit firmness.
The comparison of the two sources suggests thatthe growing environment and postharvest handlingmay influence the quality and/or ripening of papayafruit. Tian et al. (1996)stated that a differential
168 I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173

I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173 169
response of chlorophyll fluorescence and skin colorof broccoli florets to a heat treatment may be due togenetic or environmental factors. Among apple culti-vars,DeEll et al. (1997)found significant correlationbetween chlorophyll fluorescence and firmness for‘McIntosh’ but non-significant for ‘Delicious’ and‘Golden Delicious.’
3.1.3. Part III: correlations between skin color,chlorophyll fluorescence and fruit firmness
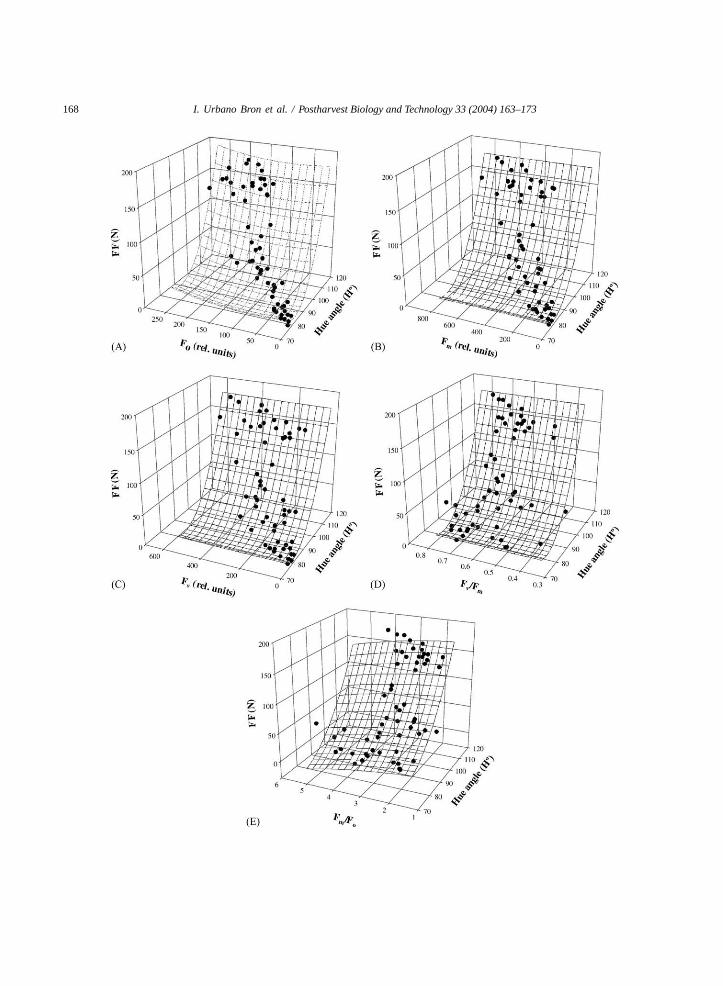
Multiple regressions between evaluated parametersshowed that the use of pairs of parameters to estimatefruit firmness was reliable (Fig. 3). In general, corre-lation coefficients (r2) were between 0.771 (data notshown) and 0.726 (Fig. 3E) when fruit firmness wasestimated using onlyH◦ andH◦ with Fm/Fo, respec-tively. Although the estimation of firmness based onH◦ had highr2 for a single fruit lot, estimating firm-ness usingH◦ to predict firmness is likely not practicalfor fruit from different sources, as shown inTable 1.
After testing these relationships with the measure-ments taken in papaya fruit at three ripeness stages(part I of this experiment), we obtained good relation-ships between measured and estimated fruit firmness(Table 2). In a broad sense, all correlations overesti-mated the fruit firmness, as indicated by the angularcoefficients (b > 1). Fruit firmness was overestimatedthe most using onlyH◦ (b = 1.3987) in spite of thehighestr2 value. On the other hand, the estimation offirmness using bothH◦ andFm/Fo had the lowestr2
value (0.727) (Fig. 3E), but it was more accurate inestimating firmness (b = 1.2425) (Table 2).
The relationships between chlorophyll fluorescenceparameters and fruit firmness have also been found inother fruits, such as kiwifruit (Kempler et al., 1992)and peach (Beaudry et al., 1997). However, using onlyone fruit characteristic to evaluate the fruit firmnessor the overall fruit quality is not always suitable, asdemonstrated inTable 1and suggested byKingston
Fig. 3. Empirical multiple regressions between hue angle (H◦), chlorophyll fluorescence parameters and fruit firmness (FF) for ‘Golden’papaya (n = 57) held at room conditions (24± 1◦C and RH of 80± 4%). (A) relation betweenH◦, minimal fluorescence yield (Fo)and FF as given by FF= (H◦ − 84.122)2/2.2532 + (Fo − 84.122)2/46.5582, r2 = 0.756 (P < 0.0001); (B) relation betweenH◦,maximal fluorescence yield (Fm) and FF as given by FF= (H◦ − 82.625)2/2.3502 − (Fm/1362.561)2, r2 = 0.754 (P < 0.0001); (C)relation betweenH◦, variable fluorescence yield (Fv), and FF as given by FF= (H◦ − 82.574)2/2.3592 + (Fv + 2.359)2/494.8082,r2 = 0.754 (P < 0.0001); (D) relation betweenH◦, variable:maximal fluorescence yield (Fv/Fm) and FF as given byFF = (H◦ − 82.372)2/2.3722 + (Fv/Fm − 82.372)2/1290382.0892, r2 = 0.754 (P < 0.0001); (E) relation betweenH◦, maximal:minimalfluorescence yield (Fm/Fo) and FF as given by FF= (H◦ − 70.597)2/3.3242 − (Fm/Fo)
2, r2 = 0.726 (P < 0.0001).
Table 2Angular (b) and correlation (r2) coefficients of linear regressiona
(n = 30) between measured (MF) and estimated (EF) fruit firmnessusing the relations between firmness, hue angle, and chlorophyllfluorescence parameters
Used relationshipsb b r2 (P value)
Firmness andH◦ 1.3987 0.826 (<0.0001)Firmness,H◦, andFo 1.3917 0.824 (<0.0001)Firmness,H◦, andFm 1.3790 0.808 (<0.0001)Firmness,H◦, andFv 1.3814 0.808 (<0.0001)Firmness,H◦, andFv/Fm 1.3770 0.806 (<0.0001)Firmness,H◦, andFm/Fo 1.2425 0.732 (<0.0001)
a Through the origin (EF= b × MF).b Relationships obtained from part III of Experiment I (Fig. 3)
and tested with the data set shown in part I (Fig. 2).
(1991). A similar finding was made in broccoli whenToivonen and Vidaver (1988)detected a decrease inchloroplast function without yellowing. We suggestthat an erroneous fruit quality evaluation may be min-imized using more than one parameter, as shown inTable 2.
3.2. Experiment II—changes of chlorophyllfluorescence and skin color during ripening
Chlorophyll fluorescence parameters and skin colordeclined during fruit ripening (Fig. 4A–C). Similar re-sponses were also detected during the ripening of ap-ple, mango, and banana (Smillie et al., 1987; Songet al., 1997). BecauseFv/Fm remained relatively con-stant until the 4th day, it is suggested that theFv/Fm pa-rameter was insensible to early changes in the ripenessprocess (Fig. 4C). Sanxter et al. (1992)also observedrelatively stable values ofFv/Fm when fruit ripenessstages were from dead-green to three-quarter ripe. Infact, it was due to an early similar reduction (until 4thday) in bothFm and Fo parameters (Fig. 4B). Songet al. (1997)reported a sharp decline in theFv/Fm
170 I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173
0
150
300
450
600
750
0 1 2 3 4 5 6 7
0.52
0.56
0.60
0.64
0.68
0.72
80
90
100
110
120(A)
Hue
ang
le (
ºh)
(B)
Chl
orop
hyll
fluo
resc
ence
(rel
. uni
ts)
Minimal (Fo)
Maximal (Fm)
Variable (Fv)
(C)
Fv/F
m
Days of ripening
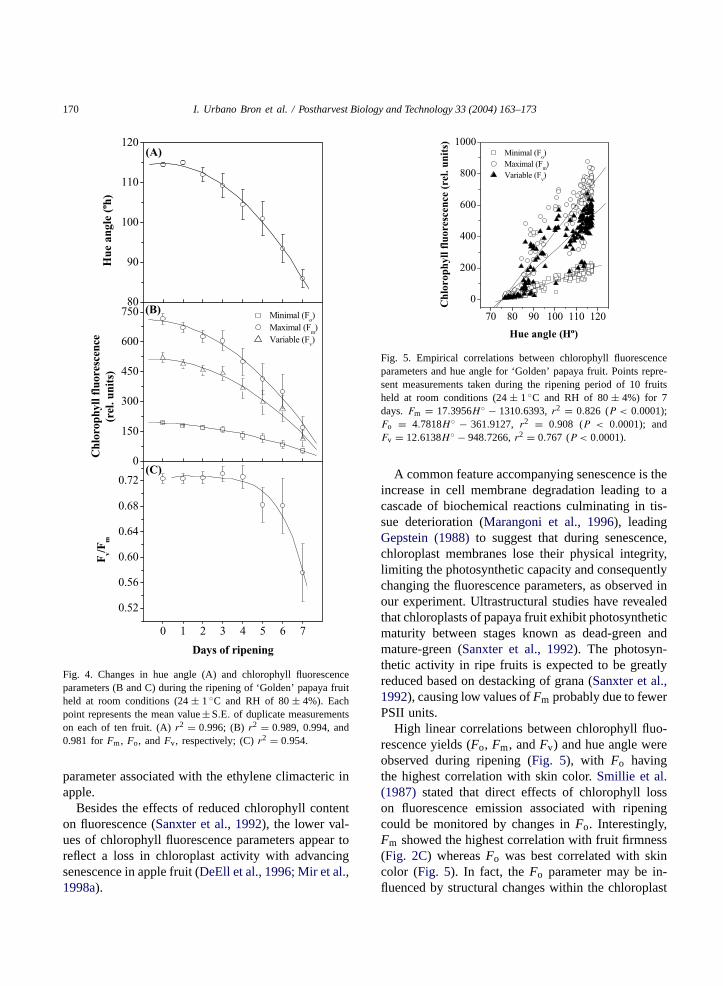
Fig. 4. Changes in hue angle (A) and chlorophyll fluorescenceparameters (B and C) during the ripening of ‘Golden’ papaya fruitheld at room conditions (24± 1◦C and RH of 80± 4%). Eachpoint represents the mean value± S.E. of duplicate measurementson each of ten fruit. (A)r2 = 0.996; (B) r2 = 0.989, 0.994, and0.981 forFm, Fo, andFv, respectively; (C)r2 = 0.954.
parameter associated with the ethylene climacteric inapple.
Besides the effects of reduced chlorophyll contenton fluorescence (Sanxter et al., 1992), the lower val-ues of chlorophyll fluorescence parameters appear toreflect a loss in chloroplast activity with advancingsenescence in apple fruit (DeEll et al., 1996; Mir et al.,1998a).
70 80 90 100 110 120
0
200
400
600
800
1000
Chl
orop
hyll
fluo
resc
ence
(re
l. un
its)
Hue angle (Hº)
Minimal (Fo)
Maximal (Fm)
Variable (Fv)
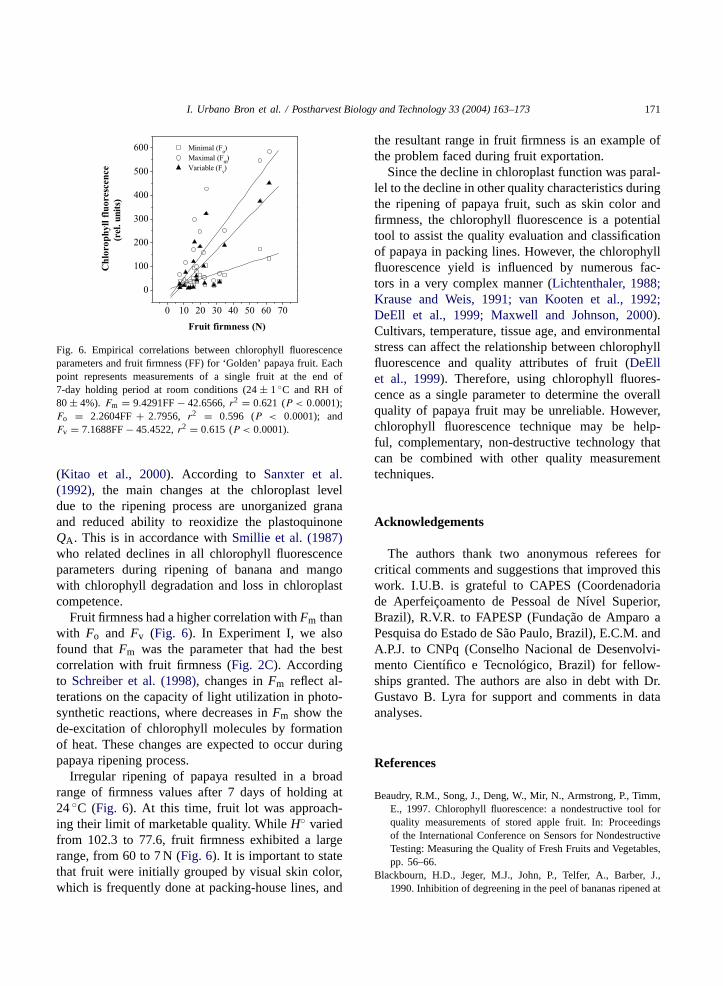
Fig. 5. Empirical correlations between chlorophyll fluorescenceparameters and hue angle for ‘Golden’ papaya fruit. Points repre-sent measurements taken during the ripening period of 10 fruitsheld at room conditions (24± 1◦C and RH of 80± 4%) for 7days.Fm = 17.3956H◦ − 1310.6393, r2 = 0.826 (P < 0.0001);Fo = 4.7818H◦ − 361.9127, r2 = 0.908 (P < 0.0001); andFv = 12.6138H◦ − 948.7266, r2 = 0.767 (P < 0.0001).
A common feature accompanying senescence is theincrease in cell membrane degradation leading to acascade of biochemical reactions culminating in tis-sue deterioration (Marangoni et al., 1996), leadingGepstein (1988)to suggest that during senescence,chloroplast membranes lose their physical integrity,limiting the photosynthetic capacity and consequentlychanging the fluorescence parameters, as observed inour experiment. Ultrastructural studies have revealedthat chloroplasts of papaya fruit exhibit photosyntheticmaturity between stages known as dead-green andmature-green (Sanxter et al., 1992). The photosyn-thetic activity in ripe fruits is expected to be greatlyreduced based on destacking of grana (Sanxter et al.,1992), causing low values ofFm probably due to fewerPSII units.
High linear correlations between chlorophyll fluo-rescence yields (Fo, Fm, andFv) and hue angle wereobserved during ripening (Fig. 5), with Fo havingthe highest correlation with skin color.Smillie et al.(1987) stated that direct effects of chlorophyll losson fluorescence emission associated with ripeningcould be monitored by changes inFo. Interestingly,Fm showed the highest correlation with fruit firmness(Fig. 2C) whereasFo was best correlated with skincolor (Fig. 5). In fact, theFo parameter may be in-fluenced by structural changes within the chloroplast
I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173 171
0 10 20 30 40 50 60 70
0
100
200
300
400
500
600
Fruit firmness (N)
Chl
orop
hyll
fluo
resc
ence
(re
l. un
its)
Minimal (Fo)Maximal (Fm)Variable (Fv)
Fig. 6. Empirical correlations between chlorophyll fluorescenceparameters and fruit firmness (FF) for ‘Golden’ papaya fruit. Eachpoint represents measurements of a single fruit at the end of7-day holding period at room conditions (24± 1◦C and RH of80± 4%). Fm = 9.4291FF− 42.6566, r2 = 0.621 (P < 0.0001);Fo = 2.2604FF+ 2.7956, r2 = 0.596 (P < 0.0001); andFv = 7.1688FF− 45.4522, r2 = 0.615 (P < 0.0001).
(Kitao et al., 2000). According to Sanxter et al.(1992), the main changes at the chloroplast leveldue to the ripening process are unorganized granaand reduced ability to reoxidize the plastoquinoneQA. This is in accordance withSmillie et al. (1987)who related declines in all chlorophyll fluorescenceparameters during ripening of banana and mangowith chlorophyll degradation and loss in chloroplastcompetence.
Fruit firmness had a higher correlation withFm thanwith Fo and Fv (Fig. 6). In Experiment I, we alsofound thatFm was the parameter that had the bestcorrelation with fruit firmness (Fig. 2C). Accordingto Schreiber et al. (1998), changes inFm reflect al-terations on the capacity of light utilization in photo-synthetic reactions, where decreases inFm show thede-excitation of chlorophyll molecules by formationof heat. These changes are expected to occur duringpapaya ripening process.
Irregular ripening of papaya resulted in a broadrange of firmness values after 7 days of holding at24◦C (Fig. 6). At this time, fruit lot was approach-ing their limit of marketable quality. WhileH◦ variedfrom 102.3 to 77.6, fruit firmness exhibited a largerange, from 60 to 7 N (Fig. 6). It is important to statethat fruit were initially grouped by visual skin color,which is frequently done at packing-house lines, and
the resultant range in fruit firmness is an example ofthe problem faced during fruit exportation.
Since the decline in chloroplast function was paral-lel to the decline in other quality characteristics duringthe ripening of papaya fruit, such as skin color andfirmness, the chlorophyll fluorescence is a potentialtool to assist the quality evaluation and classificationof papaya in packing lines. However, the chlorophyllfluorescence yield is influenced by numerous fac-tors in a very complex manner (Lichtenthaler, 1988;Krause and Weis, 1991; van Kooten et al., 1992;DeEll et al., 1999; Maxwell and Johnson, 2000).Cultivars, temperature, tissue age, and environmentalstress can affect the relationship between chlorophyllfluorescence and quality attributes of fruit (DeEllet al., 1999). Therefore, using chlorophyll fluores-cence as a single parameter to determine the overallquality of papaya fruit may be unreliable. However,chlorophyll fluorescence technique may be help-ful, complementary, non-destructive technology thatcan be combined with other quality measurementtechniques.
Acknowledgements
The authors thank two anonymous referees forcritical comments and suggestions that improved thiswork. I.U.B. is grateful to CAPES (Coordenadoriade Aperfeiçoamento de Pessoal de Nıvel Superior,Brazil), R.V.R. to FAPESP (Fundação de Amparo aPesquisa do Estado de São Paulo, Brazil), E.C.M. andA.P.J. to CNPq (Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnológico, Brazil) for fellow-ships granted. The authors are also in debt with Dr.Gustavo B. Lyra for support and comments in dataanalyses.
References
Beaudry, R.M., Song, J., Deng, W., Mir, N., Armstrong, P., Timm,E., 1997. Chlorophyll fluorescence: a nondestructive tool forquality measurements of stored apple fruit. In: Proceedingsof the International Conference on Sensors for NondestructiveTesting: Measuring the Quality of Fresh Fruits and Vegetables,pp. 56–66.
Blackbourn, H.D., Jeger, M.J., John, P., Telfer, A., Barber, J.,1990. Inhibition of degreening in the peel of bananas ripened at

172 I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173
tropical temperatures. IV. Photosynthetic capacity of ripeningbananas and plantains in relation to changes in the lipidcomposition of ripening banana peel. Ann. Appl. Biol. 117,163–174.
DeEll, J.R., Toivonen, P.M.A., 2000. Chlorophyll fluorescence asa nondestructive indicator of broccoli quality during storage inmodified-atmosphere packaging. HortScience 35, 256–259.
DeEll, J.R., Prange, R.K., Murr, D.P., 1996. Chlorophyllfluorescence of Delicious apples at harvest as a potentialpredictor of superficial scald development during storage.Postharvest Biol. Technol. 9, 1–6.
DeEll, J.R., Prange, R.K., Murr, D.P., 1997. Chlorophyllfluorescence as an indicator of apple fruit firmness. In:Proceedings of the International Conference on Sensors forNondestructive Testing: Measuring the Quality of Fresh Fruitsand Vegetables, pp. 67–73.
DeEll, J.R., Prange, R.K., Murr, D.P., 1998. Chlorophyllfluorescence techniques to detect atmospheric stress in storedapples. Acta Hortic. 464, 127–131.
DeEll, J.R., van Kooten, O., Prange, R.K., Murr, D.P.,1999. Applications of chlorophyll fluorescence techniques inpostharvest physiology. Hortic. Rev. 23, 69–107.
Gepstein, S., 1988. Photosynthesis. In: Nooden, L.D., Leopold,A.C. (Eds.), Senescence and Aging in Plants. Academic Press,San Diego, pp. 85–109.
Gray, G.R., Chauvin, F.S., Huner, N.P.A., 1997. Cold acclimationand freezing tolerance. A complex interaction of light andtemperature. Plant Physiol. 114, 467–474.
Holden, M., 1961. The breakdown of chlorophyll bychlorophyllase. Biochem. J. 78, 359–364.
Huber, J., 1983. The role of cell wall hydrolases in fruit softening.Hortic. Rev. 5, 169–219.
Kempler, C., Kabaluk, J.T., Toivonen, P.M.A., 1992. Effect ofenvironment and harvest date on maturation and ripening ofkiwifruit in British Columbia. Can. J. Plant Sci. 72, 863–869.
Kingston, C.M., 1991. Maturity indices for apple and pear. Hortic.Rev. 13, 407–432.
Kitao, M., Lei, T.T., Koike, T., Tobita, H., Maruyama,Y., Matsumoto, Y., Ang, L., 2000. Temperature responseand photoinhibition investigated by chlorophyll fluorescencemeasurements for four distinct species of dipterocarp trees.Physiol. Plant. 109, 284–290.
Krause, G.H., Weis, E., 1984. Chlorophyll fluorescence as a toolin plant physiology. II. Interpretation of fluorescence signals.Photosynth. Res. 5, 139–157.
Krause, G.H., Weis, E., 1991. Chlorophyll fluorescence andphotosynthesis: the basics. Annu. Rev. Plant. Physiol. PlantMol. Biol. 42, 313–349.
Lichtenthaler, H.K. (Ed.), 1988. Application of ChlorophyllFluorescence. Kluwer, Dordrecht.
Marangoni, A.G., Palma, T., Stanley, D.W., 1996. Membraneeffects in postharvest physiology. Postharvest Biol. Technol. 7,193–217.
Marin, F.R., Angelocci, L.R., Coelho Filho, M.A., Villa Nova,N.A., 2001. Construção e avaliação de psicrometro aspiradode termopar. Sci. Agric. 58, 839–844.
Maxwell, K., Johnson, G.N., 2000. Chlorophyll fluorescence—apractical guide. J. Exp. Bot. 51, 659–668.
Mir, N., Wendorf, M., Perez, R., Beaudry, R.M., 1998a.Chlorophyll fluorescence in relation to superficial scalddevelopment in apple. J. Am. Soc. Hortic. Sci. 123, 887–892.
Mir, N.A., Perez, R., Beaudry, R.M., 1998b. Chlorophyllfluorescence and whole fruit senescence in ‘Golden Delicious’apple. Acta Hortic. 464, 121–126.
Papargeorgiou, G., 1975. Chlorophyll fluorescence: an intrinsicprobe of photosynthesis. In: Govindjee (Ed.), Bioenergetics ofPhotosynthesis. Academic Press, New York, pp. 319–371.
Prange, R.K., De Long, J.M., Leyte, J.C., Harrison, P.A., 2002.Oxygen concentration affects chlorophyll fluorescence inchlorophyll-containing fruit. Postharvest Biol. Technol. 24,201–205.
Prange, R.K., De Long, J.M., Harrison, P.A., Leyte, J.C.,McLean, S.D., 2003. Oxygen concentration affects chlorophyllfluorescence in chlorophyll-containing fruit and vegetables. J.Am. Soc. Hortic. Sci. 128, 603–607.
Sanxter, S.S., Yamamoto, H.Y., Fisher, D.G., Chan, H.T., 1992.Development and decline of chloroplasts in exocarp ofCaricapapaya. Can. J. Bot. 70, 364–373.
Schreiber, U., Bilger, W., 1987. Rapid assessment of stress effectson plant leaves by chlorophyll fluorescence measurements.In: Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel,W.C. (Eds.), Plant Response to Stress-Functional Analysis inMediterranean Ecosystems. NATO Advanced Science InstituteSeries. Springer-Verlag, Berlin, Heidelberg, New York, pp.27–53.
Schreiber, U., Bilger, W., Neubauer, C., 1994. Chlorophyllfluorescence as an non-intrusive indicator for rapid assessmentof in vivo photosynthesis. In: Schulze, E.D., Caldwell, M.M.(Eds.), Ecophysiology of Photosynthesis. Springer-Verlag,Berlin, pp. 49–70.
Schreiber, U., Bilger, W., Hormann, H., Neubauer, C., 1998.Chlorophyll fluorescence as a diagnostic tool: basics andsome aspects of practical relevance. In: Ragavendra, A.S.(Ed.), Photosynthesis: A Comprehensive Treatise. CambridgeUniversity Press, Cambridge, pp. 320–336.
Smillie, R.M., Hetherington, S.E., Nott, R., Chaplin, G.R., Wade,N.L., 1987. Application of chlorophyll fluorescence to thepostharvest physiology and storage of mango and banana fruitand the chilling tolerance of mango cultivars. Asean Food J.39, 55–59.
Song, F., Deng, W., Beaudry, R.M., 1997. Changes in chlorophyllfluorescence of apple fruit during maturation, ripening, andsenescence. HortScience 32, 891–896.
Tian, M.S., Woolf, A.B., Bowen, J.H., Ferguson, I.B., 1996.Changes in color and chlorophyll fluorescence of broccoliflorets following hot water treatment. J. Am. Soc. Hortic. Sci.121, 310–313.
Toivonen, P., Vidaver, W., 1988. Variable chlorophyllafluorescence and CO2 uptake in water-stressed white spruceseedlings. Plant Physiol. 86, 744–748.
Tucker, G.A., 1993. Introduction. In: Seymour, G.B., Taylor, J.E.,Tucker, G.A. (Eds.), Biochemistry of Fruit Ripening. Chapman& Hall, London, pp. 1–51.

I. Urbano Bron et al. / Postharvest Biology and Technology 33 (2004) 163–173 173
van Kooten, O., Snel, J.F.H., 1990. The use of chlorophyllfluorescence nomenclature in plant stress physiology.Photosynth. Res. 25, 147–150.
van Kooten, O., Mensink, M.G.J., Otma, E.C., van Schaik,A.C.R., Schouten, S.P., 1992. Chilling damage of dark
stored cucumbers (Cucumis sativusL.) affects the maximumquantum yield of photosystem 2. In: Murata, N. (Ed.),Progress in Photosynthesis Research, vol. 4. Kluwer, Dordrecht,pp. 161–164.




















