
A protocol for efficient transformation and regeneration of Carica papaya L
Upload
independentCategory
view
2download
0
ORIGINAL PAPER
Arbuscular mycorrhizal status and root phosphatase activitiesin vegetative Carica papaya L. varieties
Sharda W. Khade • Bernard F. Rodrigues •
Prabhat K. Sharma
Received: 27 July 2009 / Revised: 10 October 2009 / Accepted: 26 November 2009 / Published online: 23 December 2009
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2009
Abstract The arbuscular mycorrhizal (AM) status and
root phosphatase activities were studied in four vegetative
Carica papaya L. varieties viz., CO-1, CO-2, Honey Dew
and Washington. Standard techniques were used to ascertain
information on spore density and species diversity of AM
fungi. Although in case of estimation of root colonization
and root phosphatase activities, the existing methods were
slightly modified. Root colonization and spore density of
AM fungi along with root phosphatase (acid and alkaline)
activities varied significantly in four papaya varieties. The
present study recorded higher acid root phosphatase activity
when compared with alkaline root phosphatase activity
under P-deficient, acidic soil conditions. The present study
revealed that the root colonization of AM fungi influenced
acid root phosphatase activity positively and significantly
under P-deficient, acidic soil conditions. A total of 11 spe-
cies of AM fungi belonging to five genera viz., Acaulospora,
Dentiscutata, Gigaspora, Glomus and Racocetra were
recovered from the rhizosphere of four papaya varieties.
Keywords Arbuscular mycorrhizal fungi �Carica papaya L. � Root colonization �Root phosphatase activity � Spore density
Introduction
Phosphatases represent a broad range of intracellular as well
as soil accumulated activities that catalyze the hydrolysis of
both esters and anhydrides of phosphoric acid (Speir and
Ross 1978). Phosphatase enzymes are also directly involved
in the acquisition of phosphorus by plants. However, their
importance is not always obvious. The proposition that
plants with lower activities of root phosphatases may gain
and use phosphorus more readily than plants with higher
ones has been put forward by McLachlan (1980) who found
that acid phosphatase activity was lower in plants with more
efficient in P-uptake than grown under P-deficient condi-
tions. During the last few years, evidence has accumulated,
which suggests that microbial activity plays a significant
role in soil phosphorus transformations (Chauhan et al.
1981). Similarly, Helal and Sauerbeck (1984, 1987) have
demonstrated that soil organic phosphorus (Po), which is a
product of microbial activity, is transformed rather rapidly
in the plant rhizosphere. In agreement with this, the plant
availability of phosphorus from organic phosphates added
to soil has been frequently demonstrated. However, the
factors controlling the plant availability of soil Po are not
yet clear. Of special interest is the role of root phosphatases
and their genetic dependence (Helal 1990) in this context.
Because action of phosphatases, release inorganic phos-
phorus (Pi) from Po (Joner et al. 2000). Colonization by soil
arbuscular mycorrhizal (AM) fungi have shown to influence
phosphatase activity (Krishna et al. 1983). These AM fungi
are known to be a major microbial component of plant and
Communicated by W. Filek.
S. W. Khade � B. F. Rodrigues � P. K. Sharma
Department of Botany, Goa University,
Taleigao Plateau, Panaji 403206, Goa, India
B. F. Rodrigues
e-mail: [email protected]
P. K. Sharma
e-mail: [email protected]
Present Address:S. W. Khade (&)
IInd Floor, Darshan Apts, Vidhyanagar Colony,
Carenzalem Post, Miramar, Panaji 403002, Goa, India
e-mail: [email protected]
123
Acta Physiol Plant (2010) 32:565–574
DOI 10.1007/s11738-009-0433-x

soil ecology. They form highly evolved, mutualistic asso-
ciations with plant roots. The AM symbiosis, influences
several aspects of plant physiology such as mineral nutri-
tion, plant development and plant protection (Gianinazzi
et al. 1990). The AM fungi have direct influence on plant
growth because of their effect on soil structure stabilization,
i.e. soil aggregate formation and accumulation of humic
substances (Bethlenfalvay and Linderman 1992; Bethlen-
falvay and Schuepp 1994). The primary effect of AM
symbiosis is to increase the supply of mineral nutrients
particularly those whose ionic forms have a poor mobility
rate or those which are present in low concentration in the
soil solution. This mainly concerns phosphate, ammonium,
zinc and copper (Barea 1991). The interconnected network
of external hyphae acts as additional catchments and
absorbing surface in the soil beyond the depletion zone
that would otherwise remain inaccessible (Rhodes and
Gerdemann 1975).
The association between AM fungi and papaya has been
reported (Khade et al. 2002; Khade and Rodrigues 2008a,
b, 2009). Papaya belongs to family Caricaceae. The genus
Carica has about 48 species of which only Carica papaya
L. is grown for its edible fruits. This paper presents results
of preliminary work, where commercially available varie-
ties of papaya were screened for their root phosphatase
activities in relation to their AM status. The activity of the
enzyme phosphatase is a physiological characteristic rela-
ted to plant efficiency in relation to P acquisition and uti-
lization, and is genetically variable (Machado and Furlani
2004). Therefore, as part of a study on papaya genotype
characterization, experiments were set-up to measure
phosphatase activity in roots of four dioecious papaya
varieties and to investigate its AM status.
Materials and methods
Study site and management regimes
Papaya plantations in agricultural farm located at Old Goa
(North Goa) was selected for the study. Four dioecious
varieties viz., CO-1, CO-2, Honey Dew and Washington
were planted in monoculture. Papaya plantations were
managed conventionally. Generally, papaya seedlings are
raised in the nursery for 2 months and then they are
transplanted in the field. The early vegetative stage mainly
comprises of tap root system with feeble primary branches
making it difficult to collect root (feeder roots) samples for
mycorrhizal studies. Hence, the study was initiated during
late vegetative stage (here after referred to as ‘vegetative
stage’) when the plants were 6-month old and exhibited
extensive root system with ample of feeder roots during the
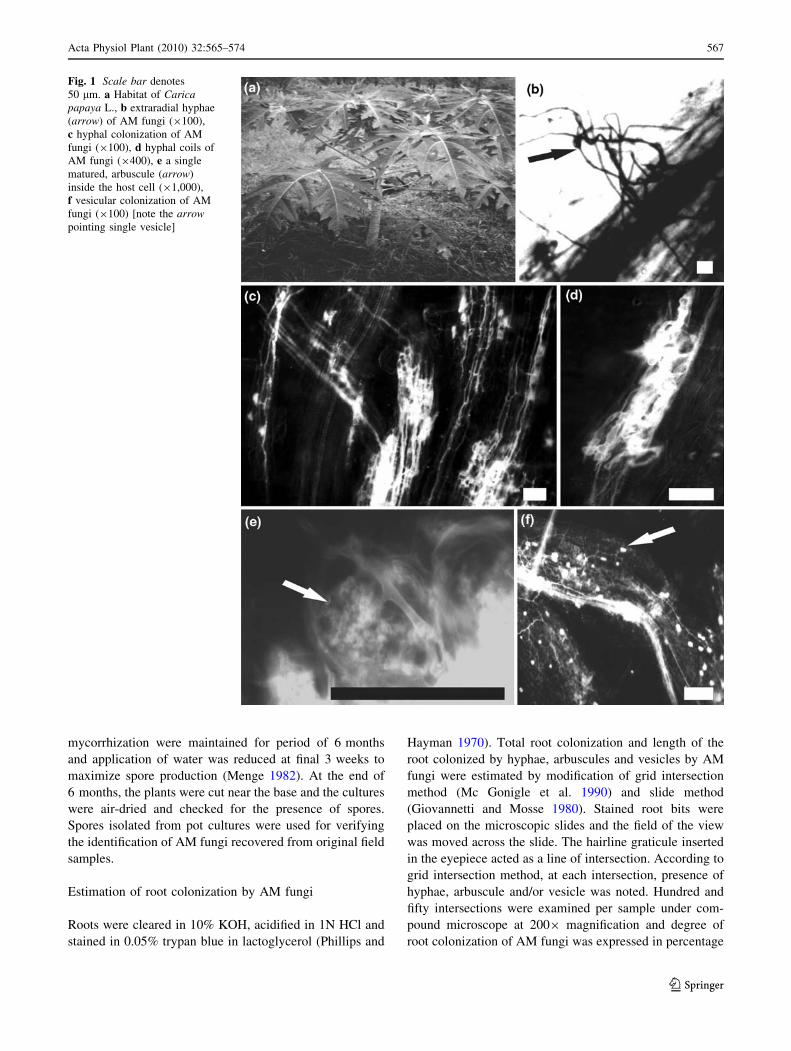
monsoons (Fig. 1a).
Collection of samples
During vegetative stage, two healthy plants per variety were
randomly selected for the collection of rhizosphere soil and
root samples. For each plant, three random soil cores were
collected from within 60 cm of each plant, at the depth of
0–25 cm. Sub-samples collected from both plants were then
combined to make composite sample after thorough mixing.
From this composite sample, five sub-samples were made
for quantification of spore density, two sub-samples for
nutrient analysis and two sub-samples for establishment of
pot cultures. Roots were packed separately for assay of
phosphatases and for estimation of root colonization per
variety in the field. Root samples collected for assay of
phosphatases were transported in ice to the laboratory.
These root samples (feeder roots) were washed in ice-cold
water and rinsed with double distilled water, packed in
polyethylene bags, labeled and stored at -80�C. For esti-
mation of root colonization, the root samples of both the
plants were combined. From this composite sample, five
sub-samples were made and they were utilized for estima-
tion of degree of root colonization and these root samples
were processed freshly. Sampling procedures were carried
out according to Tews and Koske (1986) except that the
core size was bigger (15 cm in diameter). This was carried
out to avoid non-normal distribution of spores recorded in
counts from small core samples (St. John and Koske 1988).
Soil sample analysis
Two rhizosphere soil samples per variety were employed
for analysis of nine edaphic factors during the vegetative
stage. Soil pH was measured in 1:2 soil water suspension
using pH meter (LI-120 Elico, India). Electrical conduc-
tivity was measured at room temperature in 1:5 soil sus-
pension using conductivity meter (CM-180 Elico, India).
Standard soil analysis techniques viz., micro-Kjeldahl
method (Jackson 1971) and Bray and Kurtz (1945) method
were employed for the determination of total nitrogen and
available phosphorus, respectively. Available potassium
was estimated by ammonium acetate method (Hanway and
Heidel 1952) using Flame Photometer (CL-361, Elico,
India). Available zinc, copper, manganese and iron were
quantified by DTPA–CaCl2–TEA method (Lindsay and
Norvell 1978) using Atomic Absorption Spectrophotometer
(AAS 4139, Electronic Corporation of India Ltd, India).
Establishment of pot cultures
Baiting of native AM fungi were carried out using open pot
cultures (Gilmore 1968). Two pot cultures were maintained
per variety. The roots of host species were checked for AM
colonization after 45 days. Pots showing successful
566 Acta Physiol Plant (2010) 32:565–574
123
mycorrhization were maintained for period of 6 months
and application of water was reduced at final 3 weeks to
maximize spore production (Menge 1982). At the end of
6 months, the plants were cut near the base and the cultures
were air-dried and checked for the presence of spores.
Spores isolated from pot cultures were used for verifying
the identification of AM fungi recovered from original field
samples.
Estimation of root colonization by AM fungi
Roots were cleared in 10% KOH, acidified in 1N HCl and
stained in 0.05% trypan blue in lactoglycerol (Phillips and
Hayman 1970). Total root colonization and length of the
root colonized by hyphae, arbuscules and vesicles by AM
fungi were estimated by modification of grid intersection
method (Mc Gonigle et al. 1990) and slide method
(Giovannetti and Mosse 1980). Stained root bits were
placed on the microscopic slides and the field of the view
was moved across the slide. The hairline graticule inserted
in the eyepiece acted as a line of intersection. According to
grid intersection method, at each intersection, presence of
hyphae, arbuscule and/or vesicle was noted. Hundred and
fifty intersections were examined per sample under com-
pound microscope at 2009 magnification and degree of
root colonization of AM fungi was expressed in percentage
Fig. 1 Scale bar denotes
50 lm. a Habitat of Caricapapaya L., b extraradial hyphae
(arrow) of AM fungi (9100),
c hyphal colonization of AM
fungi (9100), d hyphal coils of
AM fungi (9400), e a single
matured, arbuscule (arrow)
inside the host cell (91,000),
f vesicular colonization of AM
fungi (9100) [note the arrowpointing single vesicle]
Acta Physiol Plant (2010) 32:565–574 567
123

according to slide method. Five replicates were considered
per plant per variety
Assay of phosphatase enzyme activities
This experiment comprises two sets of observations with
five replications each per type of assay per variety.
Activities were assayed separately for acid phosphatase
and alkaline phosphatase. In all 40 readings, each was
recorded for acid and alkaline phosphatase, respectively.
One gram of root tissue was weighed per set. Methods for
quantification of activity of root phosphatases were stan-
dardized for papaya based on the procedures provided by
Kapoor et al. (1989) and Sukhada (1992).
Extraction of enzyme
Enzyme was extracted by macerating 1 g of detached root
tissue at 4�C using 20 ml of phosphate (0.1 M KH2PO4, pH
6.6) buffer. The homogenate was filtered through muslin
cloth and the filtrate was centrifuged at 5,000 rpm for 15 min
using a cooling centrifuge (Remy C-24, India). The super-
natant was stored at 4�C and further employed for protein
estimation of enzyme and assay of root phosphatases.
Protein estimation
Protein content of the enzyme extract was estimated by
Lowry’s method (Lowry et al. 1951) using bovine serum
albumin as standard.
Assay of root phosphatases
For alkaline root phosphatase activity, 5 lg protein equiv-
alent enzyme extract was incubated with 2 ml of 15 mM
p-nitrophenyl phosphate (pNPP) and 0.8 ml 0.25 M
Tris–HCl (buffer pH 9.8). For acid root phosphatase activity,
5 lg protein equivalent enzyme extract was incubated with
2 ml of 15 mM p-nitrophenyl phosphate (pNPP) and 0.8 ml
0.25 M sodium acetate (buffer pH 6.0). The reactions in both
the above-mentioned cases were terminated by adding 2 ml
of 1 N NaOH. For each set, T00 (zero time of incubation) and
T030 (30 min after incubation) were taken separately for
alkaline and acid root phosphatase activity. Optical density
was read at 410 nm using uv-1201 Shimadzu spectropho-
tometer (Japan) against a blank solvent (distilled water).
Root phosphatase activity was expressed in terms of n moles
of p-nitrophenol released per min per lg protein.
Quantification of spore density of AM fungi
Spores and sporocarps of AM fungi were isolated by wet
sieving and decanting method (Gerdemann and Nicolson
1963) and quantification of spore density of AM fungi was
carried out by method described by Gaur and Adholeya
(1994). Five replicates were considered per plant per
variety.
Identification of AM fungi
Diagnostic slides containing intact and crushed spores and
sporocarps of AM fungi were prepared in polyvinyl alcohol
lactoglycerol (Koske and Tessier1983). Spore morphology
and wall characteristics were considered for the identifi-
cation of AM fungi and these characteristics were ascer-
tained using compound microscope. The AM fungi were
identified to species level using manual identification of
AM fungi by Schenck and Perez (1990). Taxonomic
identification of spores was also carried out by matching
the descriptions provided by International Collection of
Vesicular-Arbuscular Mycorrhizal fungi (http://invam.
caf.wvu.edu) and Oehl et al. (2008).
Frequency of occurrence
Frequency of occurrence of AM fungi was calculated using
the following formula (Beena et al. 2000).
Relative abundance
Relative abundance of AM fungi was calculated using the
following formula (Beena et al. 2000).
Frequency ð%Þ ¼ Number of soil samples that possess spores of particular species
Total number of soil samples analyzed� 100:
568 Acta Physiol Plant (2010) 32:565–574
123

Statistical analysis
Data on root colonization, spore density and root phos-
phatase activity was subjected to one-way analysis of
variance to investigate the variations in these parameters
with respect to four varieties of papaya. Before analysis of
variance, root colonization values were subjected to arcsine
transformations. In addition, multiple linear regression
analysis was carried out to determine the relationship
between root colonization and root phosphatase activity.
Data were statistically analyzed using Web Agri Stat
Package (WASP) 1.0.
Results
Edaphic factors
Data on edaphic factors are recorded in Table 1. The soil
pH was acidic, while electrical conductivity was ranging
from low to optimum. Available phosphorus recorded low
levels, whereas total nitrogen and potassium levels were
ranging from low to optimum. Micronutrients in general
were present in high concentrations.
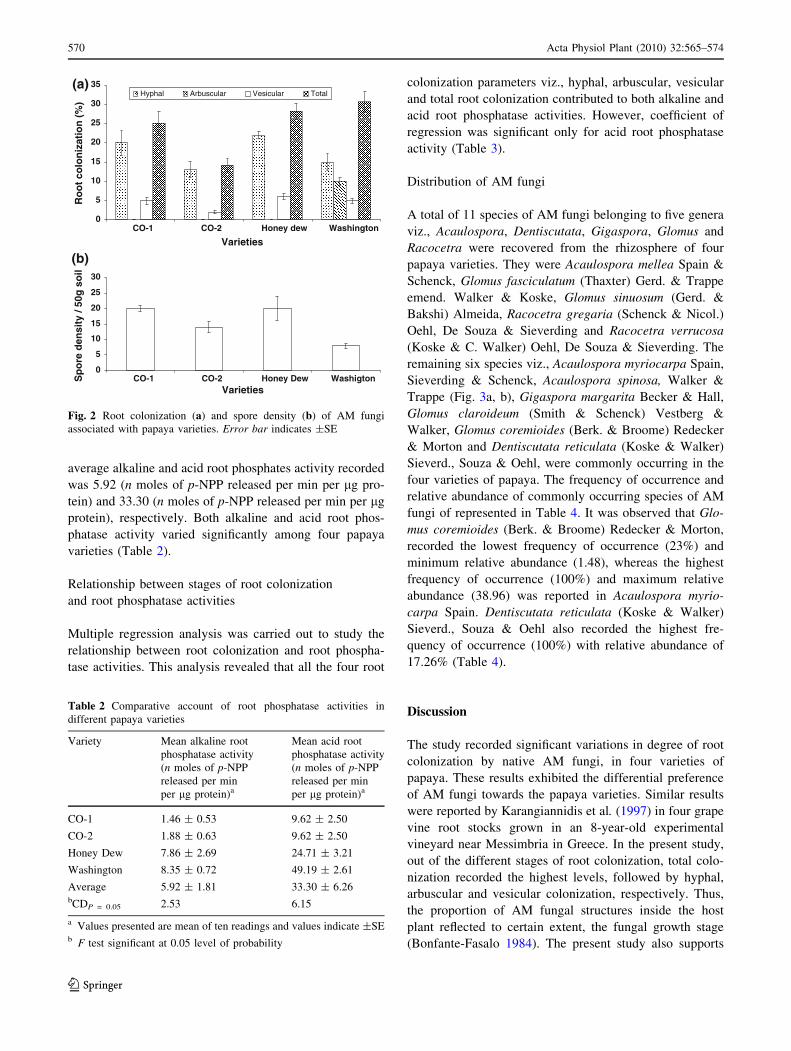
Root colonization and spore density of AM fungi
All the root samples collected during the study period
exhibited the presence of AM colonization that was char-
acterized by the presence of extramatrical hyphae
(Fig. 1b), hyphae (Fig. 1c), hyphal coils (Fig. 1d), arbus-
cules (Fig. 1c, e) and vesicles (Fig. 1f). Data on root col-
onization of AM fungi in four papaya varieties are
presented in Fig. 2a. In the present study, CO-2 variety
recorded the lowest hyphal (13%), vesicular (2%) and total
(14.2%) root colonization of AM fungi. Although the
highest hyphal (22%) and vesicular (6%) colonization was
recorded in Honey Dew variety. The highest total root
colonization (30.6%) was recorded in Washington variety.
In the study, arbuscular colonization was recorded only in
Washington variety (6%) (Fig. 2a). The average hyphal,
arbuscular, vesicular and total root colonization recorded
during the study was 17.5, 2.45, 4.3 and 24.5%, respec-
tively. Further, the hyphal (CD = 3.488; P = 0.05),
arbuscular (CD = 1.202; P = 0.05), vesicular (CD =
2.599; P = 0.05) and total root colonization (CD = 4.067;
P = 0.05) of AM fungi varied significantly in four varieties
of papaya during vegetative stage.
Data on spore density of AM fungi in four papaya
varieties are presented in Fig. 2b. The lowest spore density
was recorded in Washington (8 spores 50 g-1 soil) and the
highest was recorded in CO-1 and Honey Dew (20 spores
50 g-1 soil) variety, respectively. The average spore den-
sity recorded during the study was 15.5 spores 50 g-1 soil.
Spore density of AM fungi also varied significantly within
the four varieties of papaya during vegetative stage
(CD = 25.961; P = 0.05).
Root phosphatase activity
Comparative account of alkaline and acid root phosphatase
activities in four papaya varieties is recorded in Table 2. In
the present study, Washington variety recorded the highest
root phosphatase (alkaline and acid) activity followed by
Honey Dew, CO-2 and CO-1 variety (Table 2). In addition,
in the present study, acid root phosphatase activity was
consistently higher as compared to alkaline root phospha-
tase activity in four papaya varieties (Table 2). The
Table 1 Comparative account of the edaphic factors in papaya varieties
Varieties aEdaphic factors
pH EC
(m mos cm-1)
Total N (%) Available P
(kg Ha-1)
Available K
(kg Ha-1)
Zn
(ppm)
Cu
(ppm)
Fe
(ppm)
Mn
(ppm)
CO-1 4.80 (0.218) 0.08 (0.001) 0.51 (0.002) 6.00 (0.861) 110.00 (5.486) 2.29 (0.000) 5.31 (0.088) 64.28 (0.451) 67.64 (1.337)
CO-2 4.90 (0.165) 0.40 (0.032) 0.35 (0.001) 4.00 (0.014) 129.00 (4.448) 1.27 (0.001) 4.31 (0.841) 22.94 (0.469) 97.41 (1.580)
Honey Dew 5.00 (0.222) 0.29 (0.035) 0.51 (0.000) 4.00 (0.028) 200.00 (3.231) 1.48 (0.002) 5.69 (0.457) 16.88 (0.587) 10.70 (1.452)
Washington 4.40 (0.234) 0.42 (0.033) 0.51 (0.008) 6.00 (0.027) 200.00 (4.520) 2.03 (0.003) 5.11 (0.358) 34.64 (0.521) 78.49 (1.741)
a Values presented are mean of two readings and values in parenthesis indicate ±SE
Relative abundance (% ) ¼ Number of AM fungal spores of particular species
Total number of AM fungal spores of all species� 100:
Acta Physiol Plant (2010) 32:565–574 569
123
average alkaline and acid root phosphates activity recorded
was 5.92 (n moles of p-NPP released per min per lg pro-
tein) and 33.30 (n moles of p-NPP released per min per lg
protein), respectively. Both alkaline and acid root phos-
phatase activity varied significantly among four papaya
varieties (Table 2).
Relationship between stages of root colonization
and root phosphatase activities
Multiple regression analysis was carried out to study the
relationship between root colonization and root phospha-
tase activities. This analysis revealed that all the four root
colonization parameters viz., hyphal, arbuscular, vesicular
and total root colonization contributed to both alkaline and
acid root phosphatase activities. However, coefficient of
regression was significant only for acid root phosphatase
activity (Table 3).
Distribution of AM fungi
A total of 11 species of AM fungi belonging to five genera
viz., Acaulospora, Dentiscutata, Gigaspora, Glomus and
Racocetra were recovered from the rhizosphere of four
papaya varieties. They were Acaulospora mellea Spain &
Schenck, Glomus fasciculatum (Thaxter) Gerd. & Trappe
emend. Walker & Koske, Glomus sinuosum (Gerd. &
Bakshi) Almeida, Racocetra gregaria (Schenck & Nicol.)
Oehl, De Souza & Sieverding and Racocetra verrucosa
(Koske & C. Walker) Oehl, De Souza & Sieverding. The
remaining six species viz., Acaulospora myriocarpa Spain,
Sieverding & Schenck, Acaulospora spinosa, Walker &
Trappe (Fig. 3a, b), Gigaspora margarita Becker & Hall,
Glomus claroideum (Smith & Schenck) Vestberg &
Walker, Glomus coremioides (Berk. & Broome) Redecker
& Morton and Dentiscutata reticulata (Koske & Walker)
Sieverd., Souza & Oehl, were commonly occurring in the
four varieties of papaya. The frequency of occurrence and
relative abundance of commonly occurring species of AM
fungi of represented in Table 4. It was observed that Glo-
mus coremioides (Berk. & Broome) Redecker & Morton,
recorded the lowest frequency of occurrence (23%) and
minimum relative abundance (1.48), whereas the highest
frequency of occurrence (100%) and maximum relative
abundance (38.96) was reported in Acaulospora myrio-
carpa Spain. Dentiscutata reticulata (Koske & Walker)
Sieverd., Souza & Oehl also recorded the highest fre-
quency of occurrence (100%) with relative abundance of
17.26% (Table 4).
Discussion
The study recorded significant variations in degree of root
colonization by native AM fungi, in four varieties of
papaya. These results exhibited the differential preference
of AM fungi towards the papaya varieties. Similar results
were reported by Karangiannidis et al. (1997) in four grape
vine root stocks grown in an 8-year-old experimental
vineyard near Messimbria in Greece. In the present study,
out of the different stages of root colonization, total colo-
nization recorded the highest levels, followed by hyphal,
arbuscular and vesicular colonization, respectively. Thus,
the proportion of AM fungal structures inside the host
plant reflected to certain extent, the fungal growth stage
(Bonfante-Fasalo 1984). The present study also supports
Table 2 Comparative account of root phosphatase activities in
different papaya varieties
Variety Mean alkaline root
phosphatase activity
(n moles of p-NPP
released per min
per lg protein)a
Mean acid root
phosphatase activity
(n moles of p-NPP
released per min
per lg protein)a
CO-1 1.46 ± 0.53 9.62 ± 2.50
CO-2 1.88 ± 0.63 9.62 ± 2.50
Honey Dew 7.86 ± 2.69 24.71 ± 3.21
Washington 8.35 ± 0.72 49.19 ± 2.61
Average 5.92 ± 1.81 33.30 ± 6.26bCDP = 0.05 2.53 6.15
a Values presented are mean of ten readings and values indicate ±SEb F test significant at 0.05 level of probability
(a)
0
5
10
15
20
25
30
35
CO-1 CO-2 Honey dew Washington
Varieties
Ro
ot
colo
niz
atio
n (
%)
Hyphal Arbuscular Vesicular Total
(b)
0
5
10
15
20
25
30
CO-1 CO-2 Honey Dew WashigtonVarieties
Sp
ore
den
sity
/ 50
g s
oil
Fig. 2 Root colonization (a) and spore density (b) of AM fungi
associated with papaya varieties. Error bar indicates ±SE
570 Acta Physiol Plant (2010) 32:565–574
123

the contention that root colonization is genetically con-
trolled (Kesava Rao et al. 1990; Mercy et al. 1990; Raju
et al. 1990). Throughout the study, feeder roots were
sampled because they generally reported greater incidence
and intensity of colonization than the main root axis
(Mosse 1975; Smith and Walker 1981). In present study,
although the spore density of AM fungi exhibited narrow
fluctuation, it varied significantly in four papaya varieties.
Similar results were reported by Karangiannidis et al.
(1997) in four grape vine root stocks grown in an 8-year-
old experimental vineyard near Messimbria in Greece.
The present study was carried out during the monsoons
that support the findings of Clarholm and Rosengren-
Brinck (1995) who reported that estimations of acid
phosphatase made during moist soil conditions were much
more informative than those made during dry conditions.
Further, in present the study, acid root phosphatase activity
and alkaline root phosphatase activity varied significantly
within different varieties of papaya under P-deficient,
acidic (pH 4.4–5.0) soil conditions. Similarly, Helal (1990)
reported that root phosphatase activity is pH-dependent and
varietal differences in root phosphatase activities are
especially evident in the more acid range (pH 4–5) and
diminish at higher pH (6). Furthermore, Helal (1990)
reported that the pronounced pH dependency of root
phosphatase activity is indicative of the significance of the
rhizosphere pH not only for the availability of inorganic
phosphorus, but also for the activity of root enzymes and
the related turnover of organically bound plant nutrients.
The practical significance of root phosphatase for the
nutritional efficiency of plants under field conditions is not
yet clear. However, the results are indicative that plant
roots with high phosphatase activity obviously have the
potential to utilize soil organic phosphorus. Contradictory
to the findings of present study, Rubio et al. (1990) in a
greenhouse pot experiments conducted at the Universidad
de la Frontera, Tamuco, Chile, reported that root surface
acid phosphatase activity did not vary significantly in
wheat cultivars grown in the typical low phosphorus vol-
canic soils.
The present study reported consistently higher acid root
phosphatase activity when compared with alkaline root
phosphatase activity in four papaya varieties under
P-deficient, acidic soil conditions. Therefore, our study
Fig. 3 Scale bar denotes 50 lm. a Spore of Acaulospora spinosaWalker & Trappe laterally attached to hyaline sporiferous saccule.
b A portion of spore wall of Acaulospora spinosa Walker & Trappe
with spines
Table 4 Frequency of occurrence and relative abundance of AM
fungi common to four papaya varieties
AM fungal species Frequency of
occurrence (%)
Relative
abundance (%)
Acaulospora spinosa 50 2.96
Acaulospora myriocarpa 100 38.96
Gigaspora margarita 50 11.85
Glomus claroideum 62.50 2.90
Glomus coremioides 25 1.48
Dentiscutata reticulata 100 17.26
Table 3 Relationship between root phosphatase activity (Y) and root colonization of AM fungi (X) in papaya varieties
Root phosphatase activity (n moles of p-NPP
released per min per lg protein)
Equations Regression
coefficient (R2)a
Alkaline Y = -5.12 - 0.33H - 0.53 A ? 0.47V ? 1.09T 0.373
Acid Y = 3.75 ? 0.10H ? 1.84A ? 0.83V - 1.73T 0.747a
T total root colonization and root length colonized by H hyphae, A arbuscules, and V vesiclesa Regression significant at 0.05 level of probability
Acta Physiol Plant (2010) 32:565–574 571
123

upholds the view that one of the initial responses of plants
to P deficiency stress is an increase in root acid phospha-
tase activity (Goldstein et al. 1988, Duff et al. 1994)
because its function is the hydrolysis of inorganic phos-
phorous (Pi) from orthophosphate monoesters used for
plant nutrition in the soil (Huttova et al. 2002). Further, the
findings of the present study is in accordance with Sukhada
(1992) who reported higher acid phosphatase activity in
comparison to alkaline phosphatase activity in roots of
4-month-old papayas inoculated with Glomus mosseae
(Nicol. & Gerd.) Gerd. & Trappe and Glomus fasciculatum
(Thaxt.) Gerd. & Trappe in acidic soil conditions. Similarly
Krishna et al. (1983) reported higher acid root phosphatase
activity when compared with alkaline root phosphatase
activity in different growth stages of Arachis hypogea L.
inoculated with Glomus fasciculatum (Thaxt.) Gerd. &
Trappe under P-deficient, acidic soil conditions. Garcıa-
Gomez et al. (2002) also reported increased acid root
phosphatase activity in papaya plants inoculated with
Glomus claroideum as compared to non-mycorrhizal
plants. Contradictory to this, Allen et al. (2006) reported
that Bouteloua gracilis colonized with Glomus fascicula-
tum and grown in the phytate medium had substantially
higher alkaline phosphatase activity than non-mycorrhizal
plants and that acid phosphatase activity was not affected
by mycorrhizal condition.
Further, it is evident from multiple regression analysis
that the root colonization of AM fungi influenced acid and
alkaline root phosphatase activity positively. However,
acid root phosphatase activity was significantly influenced
by root colonization by AM fungi, while alkaline root
phosphatase activity was marginally influenced by root
colonization of AM fungi. Similarly, McArthur and
Knowles (1993) reported that the specific activities of root
acid phosphatases were enhanced in P-deficient potato
(Solanum tuberosum L.) plants inoculated by Glomus fas-
ciculatum [Thaxt. sensu Gerdemann] Gerdemann and
Trappe). Here, the establishment of AM symbiosis by low-
P plants was essential for efficient P acquisition and a
greater root colonization levels for P-stressed plants indi-
cated increased compatibility to the AM fungus. However,
the AM fungus only, partially alleviated P deficiency stress
and did not completely compensate for inadequate abiotic
P supply. In another studies conducted at the Department of
Botany and Plant Pathology, Michigan State University, E
Lansing, USA, maize (Zea mays cv. Great Lakes 586)
plants were grown under five different levels of soil
phosphorus, either in the presence or absence of formo-
nonetin or the VAM fungus, Glomus intraradices. Here,
the ACP (acid phosphatase) and ALP (alkaline phospha-
tase) activities were closely related to the level of fungal
colonization in maize roots. Acid phosphatase activity in
maize roots responded more to soil phosphorus availability
than ALP activity (38% more). These results suggest that
ACP was involved in the increased uptake of phosphorus
from the soil, while ALP may be linked to active phosphate
assimilation or transport in mycorrhizal roots. Thus, soil
phosphorus directly affected a number of enzymes essen-
tial in host–endophyte interplay (Fries et al. 1998). These
findings of Fries et al. (1998) explains the results of the
present study that recorded higher acid phosphatase activ-
ity in papaya roots when compared with alkaline phos-
phatase activity under P-deficient, acidic soil conditions.
The study recorded eleven species of AM fungi in
association with four papaya varieties. The AM fungal
species belonging to genus Glomus were the most repre-
sentative type. Similarly, maximum numbers of Glomus
species were found to be associated with papayas (Khade
and Rodrigues 2008a, b, 2009), medicinal plants (Khade
et al. 2002), pteridophytes (Khade and Rodrigues 2002) and
forest tree species (Khade and Rodrigues 2003) of Goa,
India. In the present study, few species were frequently
occurring and abundant when compared with other species
of AM fungi. Thus, the occurrence of one species sporu-
lating at the expense of others may be regulated by factors,
such as interspecific competition, spatial restriction and/or
edaphic factors (Gemma et al. 1989). Allen et al. (2003)
have pointed out that AM fungal root colonization is
mediated by interspecific fungal interactions, such as
competition, antagonism and dominance. The results from
other studies (Koske 1987; Sylvia 1986) suggest that indi-
vidual AM fungal species compete for resources through a
combination of strategies resulting in the maintenance of a
diverse AM fungal community. Therefore, a possible
explanation for the patterns found in our experiment is the
competitive interaction among AM fungal species (Schal-
amuk et al. 2006). Open pot culture technique in present
study was not successful for sporocarpic species of Glomus
that were placed earlier under genus Sclerocystis as they
failed to produce new sporocarps. This is accordance with
the findings of Muthukumar and Udaiyan (2002).
In conclusion, production of root phosphatase enzymes
specifically acid root phosphatase are inducible under
P-deficient, acidic soil conditions and this activity consid-
erably varies among four papaya varieties. The study
highlighted considerable varietal differences in root colo-
nization and spore density of AM fungi associated with
papayas. The higher acid and alkaline root phosphatase
activity along with higher total root colonization values in
Honey and Washington variety proves high P demand of
these varieties when compared to CO varieties. The present
study reported consistent higher acid root phosphatase
activity as compared to alkaline root phosphatase activity
in four papaya varieties under P-deficient, acidic soil
conditions. Therefore, the study emphasizes the fact that
tropical soils are acidic and, therefore, P-deficient that
572 Acta Physiol Plant (2010) 32:565–574
123

induces the secretion of root phosphatase activity, espe-
cially high amount of acid phosphatase that results in the
availability of phosphorus and is, therefore, taken up by
extraradical fungal hyphae. This P is translocated passively
by intra-radical fungal hypha or actively transferred to the
plant through arbuscules; the key feature of AM symbiosis.
The study, therefore, shows the relationship between the
root length colonized by hyphae, arbuscules, vesicles and
total root colonization of symbiotic AM fungi with root
phosphatase activity and highlights the fact that under
acidic soil conditions, acidic root phosphatase activity
increases with the root colonization of AM fungi. The
present study has new approach since it was carried out
under field conditions and under the influence of all envi-
ronmental pressures that to in monsoons and still exhibited
a consistency in the findings of data pooled from four
varieties, which were generalized to logical conclusions.
Acknowledgments Shri Waman M. Khade Ex-director of Agri-
culture Department and Directorate of Agriculture, State Government
of Goa are thanked for their assistance to carry out research work.
References
Allen EB, Swenson W, Querejeta JI, Egerton-Waburton LM, Treseder
KK et al (2003) Ecology of mycorrhizae: a conceptual frame-
work for complex interactions among plants and fungi. Ann Rev
Phytopathol 41:271–303
Allen MF, Sexton JC, Moore TS Jr, Christensen M et al (2006)
Influence of phosphate source on vesicular-arbuscular mycor-
rhizae of Bouteloua gracilis. New Phytol 87(4):687–694
Barea JM (1991) Vesicular-arbuscular mycorrhizae as modifiers of
soil fertility. In: Stewart BA (ed) Advances in soil science.
Springer, New York, pp 1–40
Beena KR, Raviraja NS, Arun AD, Sridhar KR et al (2000) Diversity
of arbuscular mycorrhizal fungi on coastal sand dunes of the
West Coast of India. Curr Sci 79(10):1459–1465
Bethlenfalvay GJ, Linderman RG (1992) Mycorrhizae in sustainable
agriculture. ASA Special Publication, Madison, p 124
Bethlenfalvay GJ, Schuepp H (1994) Arbuscular mycorrhizas and
agrosystem stability. In: Gianinazzi S, Scheupp H (eds) Impact
of arbuscular mycorrhiza on sustainable agriculture and natural
ecosystems. ALS, Birkhauser, Basel, pp 117–131
Bonfante-Fasalo P (1984) Anatomy and morphology. In: Powell CL,
Bagyaraj DJ (eds) VA mycorrhiza. CRC Press Inc, Boca Raton,
pp 5–33
Bray RH, Kurtz LT (1945) Determination of total organic carbon and
available forms of phosphorus in soils. Soil Sci 59:39–45
Chauhan BS, Stewart JWB, Paul EA (1981) Effect of labile inorganic
phosphate status and organic carbon additions on the microbial
uptake of phosphorus in soils. Can J Soil Sci 61:373–385
Clarholm M, Rosengren-Brinck U (1995) Phosphorous and nitrogen
fertilization of a Norway spruce forest-effect on needle concen-
trations and acid phosphatase activity in the humus layer. Plant
Soil 175:239–249
Duff SMG, Sarath G, Plaxton WC (1994) The role of acid
phosphatase in plant phosphorus metabolism. Physiol Plant
90:791–800
Fries LLM, Pacovsky RS, Safir GR, Kaminski J et al (1998)
Phosphorus effect on phosphatase activity in endomycorrhizal
maize. Physiol Plant 103(2):162–171
Garcıa-Gomez R, Chavez-Espinosa J, Mejıa-Chavez A, Duran BC
et al (2002) Short term effects of Glomus claroideum and
Azospirillum brasilense on growth and root acid phosphatase
activity of Carica papaya L. under phosphorus stress. Rev
Latinoam Microbiol 44(1):31–37
Gaur A, Adholeya A (1994) Estimation of VAM spores in the soil—a
modified method. Mycorrhiza News 6(1):10–11
Gemma JN, Koske RE, Carreiro M et al (1989) Seasonal dynamics of
selected species of VA mycorrhizal fungi in a sand dune. Mycol
Res 92:317–321
Gerdemann JW, Nicolson TH (1963) Spore density of Endogonespecies extracted from soil wet sieving and decanting. Trans Bri
Mycol Soc 46:235–244
Gianinazzi S, Trouvelot A, Gianiazzi-Pearson V et al (1990) Role and
use of mycorrhizas in horticulture crop production. XXII.
International Horticulture Congress Florence, pp 25–30
Gilmore AE (1968) Phycomycetous mycorrhizal organisms collected
by open pot cultures. Hilgardia 39:87–105
Giovannetti M, Mosse B (1980) An evaluation of techniques for
measuring vesicular arbuscular mycorrhizal infection in roots.
New Phytol 84:489–500
Goldstein AH, Baertlein DA, Mcdaniel RG (1988) Phosphate
starvation inducible metabolism in Lycopersicon esculentum. I.
Excretion of acid phosphatase by tomato plants and suspension-
cultured cells. Plant Physiol 87:711–715
Hanway JJ, Heidel H (1952) Soil analysis method as used in
Iowa State College Soil Testing Laboratory. Iowa Agric
57:1–31
Helal HM (1990) Varietal differences in root phosphatase activity as
related to the utilization of organic phosphates. Plant Soil
123:161–163
Helal HM, Sauerbeck DR (1984) Influence of plant roots on carbon
and phosphorus metabolism in soil. Plant Soil 76:175–182
Helal HM, Sauerbeck DR (1987) Direct and indirect influences of
plant roots on organic matter and phosphorus turnover in soil.
INTECOL Bull 15:49–58
Huttova J, Tamas L, Mistrık I (2002) Aluminum induced acid
phosphatase activity in roots of Al-sensitive and Al-tolerant
barley varieties. Rostlinna Vyroba 48(12):556–559
Jackson ML (1971) Soil chemical analysis. Prentice Hall, New Delhi
Joner EJ, Van Aarle IM, Vosatka M et al (2000) Phosphatase activity
of extra- radical arbuscular mycorrhizal hyphae: a review. Plant
Soil 226:199–210
Kapoor A, Singh VP, Mukerji KG (1989) Studies on the phosphatase
of mycorrhizal and non mycorrhizal Trigonella roots. In:
Mahadeva A, Raman A, Natarajan K et al (eds) Mycorrhizae
for Green Asia. CAS, Madras, pp 125–127
Karangiannidis N, Velmis D, Stravropoulos N et al (1997) Root
colonization and spore population by VA-mycorrhizal fungi in
four grapevine rootstock. Vitis 36(2):57–60
Kesava Rao PS, Tilak KVBR, Arunachalam V et al (1990) Genetic
variation of mycorrhiza-dependent phosphate mobilization in
ground nut (Arachis hypogea L.). Plant Soil 121:291–294
Khade SW, Rodrigues BF (2002) Arbuscular mycorrhizal fungi
associated with some pteridophytes from Western Ghat region of
Goa. Trop Eco 43(2):251–256
Khade SW, Rodrigues BF (2003) Occurrence of arbuscular mycor-
rhizal fungi in tree species from Western Ghats of Goa India.
J Trop For Sci 15(2):320–331
Khade SW, Rodrigues BF (2008a) Ecology of arbuscular mycorrhizal
fungi associated with Carica papaya L. in agro-based ecosystem
of Goa, India. Trop Subtrop Agroecosyst 8:265–278
Acta Physiol Plant (2010) 32:565–574 573
123

Khade SW, Rodrigues BF (2008b) Spatial variations in arbuscular
mycorrhizal (AM) fungi associated with Carica papaya L. in a
tropical agro-based ecosystem. Bio Agric Hort 26:149–174
Khade SW, Rodrigues BF (2009) Arbuscular mycorrhizal fungi
associated with varieties of Carica papaya L. in tropical agro-
based ecosystem of Goa, India. Trop Subtrop Agroecosyst
10(3):369–381
Khade SW, Bukhari MJ, Jaiswal V, Gaonkar UC, Rodrigues BF et al
(2002) Arbuscular mycorrhizal status of medicinal plants: a field
survey of AM fungal association in shrubs and trees. J Eco Tax
Bot 26(3):571–578
Koske RE (1987) Distribution of VA mycorrhizal fungi along a
latitudinal temperature gradient. Mycologia 79:55–68
Koske RE, Tessier B (1983) A convenient permanent slide mounting
medium. Mycol Soc Am Newslett 34:59
Krishna KR, Bagyaraj DJ, Papavinashasundaram KG et al (1983)
Acid and alkaline phosphatase activities in mycorrhizal and
uninfected roots of Arachis hypogaea L. Ann Bot 51:551–553
Lindsay WL, Norvell WA (1978) Development of DPTA soil test for
zinc, iron, manganese and copper. Am Soil Sci Soc J 42:421–488
Lowry OH, Rosebrough NJ, Fan AL, Randall RJ et al (1951) Protein
measurement with the Folin-phenol reagent. J Biol Chem
193:265–275
Machado CTT, Furlani AMC (2004) Root phosphatase activity, plant
growth and phosphorus accumulation of maize genotypes. Sci
Agric 61(2):216–223
Mc Gonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA et al
(1990) A new method which gives an objective measure of
colonization of roots by vesicular-arbuscular mycorrhizal fungi.
New Phytol 115:495–501
McArthur DAJ, Knowles NR (1993) Influence of vesicular-arbuscular
mycorrhizal fungi on the response of potato to phosphorus
deficiency. Plant Physiol 101(1):147–160
McLachlan KE (1980) Acid phosphate activity of intact roots and
phosphorous nutrition in plants II: variation among wheat roots.
Aust J Agric Res 31:441–448
Menge JA (1982) Utilization of vesicular arbuscular mycorrhizal
fungi in agriculture. Can J Bot 61:1015–1024
Mercy MA, Shivashanker G, Bagyaraj DJ et al (1990) Mycorrhizal
colonization in cowpea is dependent and heritable. Plant Soil
121:291–294
Mosse B (1975) A microbiologist’s view of root anatomy. In: Walker
N (ed) Soil microbiology: a critical review. Butterworths,
London, pp 39–66
Muthukumar T, Udaiyan K (2002) Arbuscular mycorrhizal fungal
composition in semi-arid soils of Western Ghats, southern India.
Curr Sci 82(6):625–628
Oehl F, de Souza FA, Sieverding E (2008) Revision of Scutellosporaand description of five new genera and three new families in the
arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon
106:311–360
Phillips JM, Hayman DS (1970) Improved procedure for clearing
roots and staining of mycorrhizal fungi for rapid assessment of
infection. Trans Bri Mycol Soc 55:158–161
Raju PS, Clark RB, Duncan JR, Maranville JW et al (1990) Benefit
and cost analysis and phosphorus efficiency of VA-mycorrhizal
fungi colonization with Sorghum (Sorghum bicolor) genotypes
grown at varied phosphorus levels. Plant Soil 124:199–204
Rhodes LH, Gerdemann JW (1975) Phosphate uptake zones of
mycorrhizal and non-mycorrhizal onions. New Phytol 75:555–
561
Rubio R, Moraga E, Borie F et al (1990) Acid phosphatase activity
and vesicular-arbuscular mycorrhizal infection associated with
roots of 4 wheat cultivars. J Plant Nutr 13(5):585–598
Schalamuk S, Velazquez H, Cabello CM et al (2006) Fungal spore
diversity of arbuscular mycorrhizal fungi associated with spring
wheat: effects of tillage. Mycologia 98(1):16–22
Schenck NC, Perez Y (1990) Manual for identification of VA
mycorrhizal fungi. INVAM, University of Florida, Gainesville,
USA, pp 1–283
Smith SE, Walker NA (1981) A qualitative study of mycorrhizal
plants in Trifolium: separate determination of the rates of
infection and of mycelial growth. New Phytol 89:225–240
Speir TW, Ross DJ (1978) Soil phosphatase and sulphatase. In: Burns
RG (ed) Soil enzymes. Academic Press, New York, USA,
pp 197–250
St. John TV, Koske RE (1988) Statistical treatment of endogonaceous
spore counts. Trans Bri Mycol Soc 91:117–121
Sukhada M (1992) Effect of VAM inoculation on plant growth,
nutrient level and root phosphatase activity in papaya (Caricapapaya cv. Coorg Honey Dew). Fert Res 31:263–267
Sylvia DM (1986) Spatial and temporal distribution of vesicular-
arbuscular mycorrhizal fungi associated with Uniola paniculatain Florida foredunes. Mycologia 78:728–734
Tews LL, Koske RE (1986) Towards a sampling strategy for vesicular
arbuscular mycorrhizas. Trans Bri Mycol Soc 87(8):353–358
574 Acta Physiol Plant (2010) 32:565–574
123
Copyright © 2022 FDOKUMEN




















