
Ethylene involvement in vegetative bud formation in tobacco thin layers
11
Protoplasma (1998) 202:134-144 PROTOPLASMA Springer-Verlag 1998 Printed in Austria Ethylene involvement in vegetative bud formation in tobacco thin layers S. Biondi 1'*, S. ScaramagW, F. Capitani 2, G. Marino 3, M. M. Altamura 2, and P. Torrigiani 1 IDipartimento di Biologia and 3Dipartimento di Colture Arboree, Universit~ di Bologna, Bologna and 2Dipartimento di Biologia Vegetale, Universit~ di Roma "La Sapienza", Roma Received January 20, 1998 Accepted March 5, 1998 Summary. The role of ethylene in vegetative bud regeneration was studied in cultured tobacco (Nicotiana tabacum L. cv Samsun) thin- layer explants. The experimental approach consisted in supplement- ing the bud-inducing medium with an inhibitor of ethylene biosyn- thesis, aminoethoxyvinylglycine (AVG), an ethylene antagonist, sil- ver thiosulphate (STS), or an ethylene-releasing compound, 2- chloroethylphosphonic acid (CEPA), at various concentrations. The organogenic response was assessed both macroscopically (percent- age of bud-forming explants, final number of buds per explant) and cytohistologically (number, characteristics, and localisation of meristemoids and bud primordia). The time course of ethylene pro- duction during culture was also evaluated. At the end of culture (day 27) all the explants treated with these compounds had a lower num- ber of buds compared to controls. STS was detrimental to meriste- moid initiation at all the concentrations tested. In contrast, 0.5 ~tM AVG, which strongly inhibited ethylene production, provoked a large increase in the formation of meristemoids early in culture and the appearance of anomalous ("twin") buds. CEPA reduced meriste- moid formation but, at the lower concentrations (1 and 10 p~M) speeded up bud emergence. On the whole it mainly favoured disor- ganised growth and xylogenesis. The results of this work highlight the contrasting effects of ethylene in relation to the two critical stages of the organogenic process, i.e., meristemoid formation and bud pri- mordium development. Keywords: Ethylene; Ethylene inhibitors; Meristemoids; Nicotiana tabacum; Thin layers; Vegetative buds. Abbreviations: ACC 1-aminocyclopropane-l-carboxylic acid; AVG aminoethoxyvinylglycine; STS silver thiosulphate; CEPA 2- chloroethylphosphonic acid; IAA indole-3-acetic acid; BA benzyl- adenine; HF hormone-free. *Correspondence and reprints: Dipartimento di Biologia e.s., Uni- versit~t di Bologna, via Imerio 42, 1-40126 Bologna, Italy. Introduction Plant tissue culture depends on the external control of morphogenesis by chemical factors, particularly hor- mones such as auxins, cytokinins, and to a lesser extent abscisic acid and gibberellins, which are inctuded in the culture medium. Ethylene is not usu- ally deliberately supplied but necessarily accumu- lates, together with other gases, in the culture vessel. The amount of ethylene that accumulates will depend on the rate of production by the plant tissue but also on the rate at which it escapes from the vessel. Although containers routinely used for tissue culture may lose ethylene quite rapidly (Biddington and Robinson 1991), ethylene production often exceeds leakage and results in considerable accumulation (Jackson et al. 1991). As there are numerous reports on the effects of ethylene on unorganised and organ- ised growth in plant tissue culture, especially shoot regeneration (Huxter et al. 1981, Chi and Pua 1989, Chraibi et al. 1991, Taylor et al. 1994) and somatic embryogenesis (Nissen 1994; Roustan et al. 1989, 1990), the role of this hormone on morphogenesis cannot be ignored. It can be evaluated either by study- ing means of controlling accumulation in the culture vessel, by measuring its rate of biosynthesis, and/or by using inhibitors of its biosynthesis or action. The role of ethylene in in vitro morphogenesis is far from being clear. Several reports indicate that shoot regeneration is stimulated by inhibiting ethylene syn- thesis or action (Chraibi et al. 1991, Purnhauser et al. 1987). In these studies, control explants had a low
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Ethylene involvement in vegetative bud formation in tobacco thin layers

Protoplasma (1998) 202:134-144 PROTOPLASMA �9 Springer-Verlag 1998 Printed in Austria
Ethylene involvement in vegetative bud formation in tobacco thin layers
S. Biondi 1'*, S. ScaramagW, F. Capitani 2, G. Marino 3, M. M. Altamura 2, and P. Torrigiani 1
IDipartimento di Biologia and 3Dipartimento di Colture Arboree, Universit~ di Bologna, Bologna and 2Dipartimento di Biologia Vegetale, Universit~ di Roma "La Sapienza", Roma
Received January 20, 1998 Accepted March 5, 1998
Summary. The role of ethylene in vegetative bud regeneration was studied in cultured tobacco (Nicotiana tabacum L. cv Samsun) thin- layer explants. The experimental approach consisted in supplement- ing the bud-inducing medium with an inhibitor of ethylene biosyn- thesis, aminoethoxyvinylglycine (AVG), an ethylene antagonist, sil- ver thiosulphate (STS), or an ethylene-releasing compound, 2- chloroethylphosphonic acid (CEPA), at various concentrations. The organogenic response was assessed both macroscopically (percent- age of bud-forming explants, final number of buds per explant) and cytohistologically (number, characteristics, and localisation of meristemoids and bud primordia). The time course of ethylene pro- duction during culture was also evaluated. At the end of culture (day 27) all the explants treated with these compounds had a lower num- ber of buds compared to controls. STS was detrimental to meriste- moid initiation at all the concentrations tested. In contrast, 0.5 ~tM AVG, which strongly inhibited ethylene production, provoked a large increase in the formation of meristemoids early in culture and the appearance of anomalous ("twin") buds. CEPA reduced meriste- moid formation but, at the lower concentrations (1 and 10 p~M) speeded up bud emergence. On the whole it mainly favoured disor- ganised growth and xylogenesis. The results of this work highlight the contrasting effects of ethylene in relation to the two critical stages of the organogenic process, i.e., meristemoid formation and bud pri- mordium development.
Keywords: Ethylene; Ethylene inhibitors; Meristemoids; Nicotiana tabacum; Thin layers; Vegetative buds.
Abbreviations: ACC 1-aminocyclopropane-l-carboxylic acid; AVG aminoethoxyvinylglycine; STS silver thiosulphate; CEPA 2- chloroethylphosphonic acid; IAA indole-3-acetic acid; BA benzyl- adenine; HF hormone-free.
*Correspondence and reprints: Dipartimento di Biologia e.s., Uni- versit~t di Bologna, via Imerio 42, 1-40126 Bologna, Italy.
Introduction
Plant tissue culture depends on the external control of morphogenesis by chemical factors, particularly hor- mones such as auxins, cytokinins, and to a lesser extent abscisic acid and gibberellins, which are inctuded in the culture medium. Ethylene is not usu- ally deliberately supplied but necessarily accumu- lates, together with other gases, in the culture vessel. The amount of ethylene that accumulates will depend on the rate of production by the plant tissue but also on the rate at which it escapes from the vessel. Although containers routinely used for tissue culture may lose ethylene quite rapidly (Biddington and Robinson 1991), ethylene production often exceeds leakage and results in considerable accumulation (Jackson et al. 1991). As there are numerous reports on the effects of ethylene on unorganised and organ- ised growth in plant tissue culture, especially shoot regeneration (Huxter et al. 1981, Chi and Pua 1989, Chraibi et al. 1991, Taylor et al. 1994) and somatic embryogenesis (Nissen 1994; Roustan et al. 1989, 1990), the role of this hormone on morphogenesis cannot be ignored. It can be evaluated either by study- ing means of controlling accumulation in the culture vessel, by measuring its rate of biosynthesis, and/or by using inhibitors of its biosynthesis or action. The role of ethylene in in vitro morphogenesis is far from being clear. Several reports indicate that shoot regeneration is stimulated by inhibiting ethylene syn- thesis or action (Chraibi et al. 1991, Purnhauser et al. 1987). In these studies, control explants had a low

S. Biondi et al.: Ethylene involvement in vegetative bud formation 135
regenerative capacity (10% at most). A similar effect has been reported for recalcitrant genotypes of Bras- sica campestris (Chi and Pua 1989, Chi et al. 1991). These authors put forward the hypothesis that ethyl- ene is involved in the regulation of cell recalcitrance with respect to in vitro shoot formation. In other tis- sue culture systems, endogenous levels of ethylene are considered to be suboptimal. In such cases, low concentrations of exogenously supplied ethylene can promote organogenesis. For example, low concen- trations of 1-aminocyclopropane-l-carboxylic acid (ACC), 2-chloroethylphosphonic acid (CEPA) or eth- ylene stimulated somatic embryogenesis in carrot ceil suspensions and ethylene inhibitors retarded it (Nis- sen 1994) but these results are in contrast to those of Roustan et al. (1990) who used another cell line, pos- sibly producing more ethylene. A morphogenic role for exogenously supplied ethylene was also reported in hypocotyl cultures of Digitalis obscura (Perez- Bermudez et al. 1985) as well as in Pinus radiata cotyledons (Kumar et al. 1987). It is evident that the effect of ethylene is variable depending on species, tissue, and presence of other plant growth regulators, all of which probably affect endogenous ethylene pro- duction or cell sensitivity to this hormone (Salisbury et al. 1988, Liu and Reid 1990). Thin-layer explants of tobacco are a versatile in vitro system with which to study the hormonal control of organogenesis inasmuch as they can be easily induced to form either vegetative buds, floral buds or roots essentially by varying the type and the concentration of auxin and cytokinin supplied in the medium (Tran Thanh Van et al. 1974, Smulders et al. 1990, Van der Krieken etal. 1990, Heylen and Vendrig 199i, Peeters et al. 1991). Smulders and coworkers (1990) also examined the role of ethylene with regard to flower bud formation in this system. They stimulated the biosynthesis of this hormone by supplying its immediate precursor, ACC, inhibited it with amino- ethoxyvinylglycine (AVG), and blocked its action with silver nitrate. Based on their interpretation of the results, ethylene seems to reduce tissue sensitivity to auxin as well as the regenerative capacity of the explants with respect to this organogenic process. Several physiological studies, pertaining especially to polyamine accumulation and metabolism, have already been conducted by our group using this tissue culture model system (Torrigiani et al. 1987, 1989, 1993; Scaramagli et al. 1995). Detailed cytohistolog- ical analyses have also been performed in relation to root and shoot neoformation (Altamura et al. 1991,
1995). The objective of the present work was to attempt to answer the following question: to what extent and at which developmental stages is ethylene involved in vegetative bud regeneration in tobacco thin-layer explants. Inhibitors of ethylene biosynthe- sis (AVG) or action (silver thiosutphate, STS) and an ethylene-releasing compound (CEPA) were added to the bud-inducing medium and organogenesis was evaluated, macroscopically and cytohistologically, at various intervals during culture.
Material and methods Plant material and tissue culture
Plants of Nicotiana tabacum L. cv. Samsun, were grown in a growth chamber under a 12h photoperiod (Philips TLD 36W/33, 1.87 W/m 2) and under day/night temperatures of 25/19 ~ respec- tively, up to the vegetative stage of 17 + 1 internodes. Thin-layer explants (2 mm • 10 ram) were excised aseptically from intemodes 6 to 9 counting from the shoot apex, and cultured on Murashige and Skoog (1962) basal medium supplemented with 0.55 mM myo-inos- itol, 1.2 ~tM thiamine, 30 g of glucose per liter and solidified with 0.8% (w/v) Bios agar (Biolife, Milano, Italy). Vegetative bud-induc- ing (control) medium also contained 1 ~tM IAA and 10 ~M BA (Altamura et al. 1995). For comparative purposes, explants were also cultured on nonorganogenic hormone-free (HF) medium. The fol- lowing compounds, filter-sterilised and added to the bud-inducing medium after autoclaving, were tested: 0.1, 0.5, 1, and 10 ~M AVG; 1, 10 and 100 ~tM STS; 1, 10, and 100 ~tM CEPA.
Macroscopic and histological analyses
The time course of organogenesis (percentage bud-forming explants) was evaluated macroscopically at weekly intervals up to day 27 in culture when the number of buds per explant was also counted under a dissecting microscope. Two to five randomly chosen explants were fixed in FAA (70% ethanol : glacial acetic acid : formalin, 18 : 1 : 1, v/v) on days 8 and 13 (controls, CEPA, STS) or on days 5, 9, and 15 (controls, 0.5 [xM AVG), dehydrated, embedded in paraffin (melting point 52-54 ~ Merck, Darmstadt, Federal Republic of Germany) and sectioned at 8 ~tm intervals with a Top-super S-150 microtome (Pabisch, Milano, Italy). After staining with eosin and Carazzi's haemalum (Altamura et al. 1991) using a Top-stainer LX-100 (Pabisch), meristemoids and primordia were counted in the explants and expressed as mean number (with standard error) per explant.
Statistical analysis
Each experiment was repeated at least once with 150-200 explants per treatment every time. The morphogenic response followed the same trend in each experiment and the results of one of these are reported here. Differences between percentages of bud-forming explants were analysed by the X 2 test, and differences between means (number of meristemoids, primordia or buds, explant fresh weight) by Student's t-test (significance at 0.01 or 0.05 as specified).
Ethylene analysis by gas chromatography
At different times in culture (1, 2, 8, and 15 days) approx. 200-300 nag fresh weight of thin-layer explants were transferred to 50 ml flasks containing 2 ml liquid medium having the same compo-
136
sition as the one they were growing on and left standing for half an hour under the laminar flow hood. Flasks were then sealed with air- tight serum caps and placed in a growth cabinet at 23 ~ in the light. 2 ml gas samples were withdrawn from the headspace at 1 h intervals for a total of 3 h and injected into a Dani DS 86.01 (Dani, Milano, Italy) packed-gas chromatograph equipped with a Porapak Q column and a flame ionization detector. The carrier gas was nitrogen at a flow rate of 16 ml/min. The temperature was set at 80 ~ for the col- umn and at 150 ~ for the injector and flame ionization detector. Eth- ylene concentration was determined using the following standard (SIAD, Bologna, Italy): 3.4 ppm C2H4, 300 ppm CO2, and 21.11% 02 in N2. Four replicate flasks per treatment were used and the exper- iment was repeated once.
Results
Time course of organogenesis: macroscopic observations
As shown in Table 1, on day 7 in culture no macro- scopic vegetative buds were observed on the explants in any of the treatments. Most explants were visibly hypertrophic, with the exception of 50% of those growing on hormone-free (HF) medium and in the presence of 10 ~M AVG or 100 gM CEPA which remained in the initial stage (alive but without visible growth). On day 15 explants with buds were observed in all treatments (except HF) but in different percent- ages: 0.1 BM AVG and 1 ~xM CEPA significantly (P < 0.01) enhanced the percentage of bud-forming ex- plants; in the other treatments this value was lower than or, with 10 ~tM STS or 10 ~xM CEPA, similar to
Table 1. Time course of organogenesis in tobacco thin layers cultured at different concentrations.
S. Biondi et al.: Ethylene involvement in vegetative bud formation
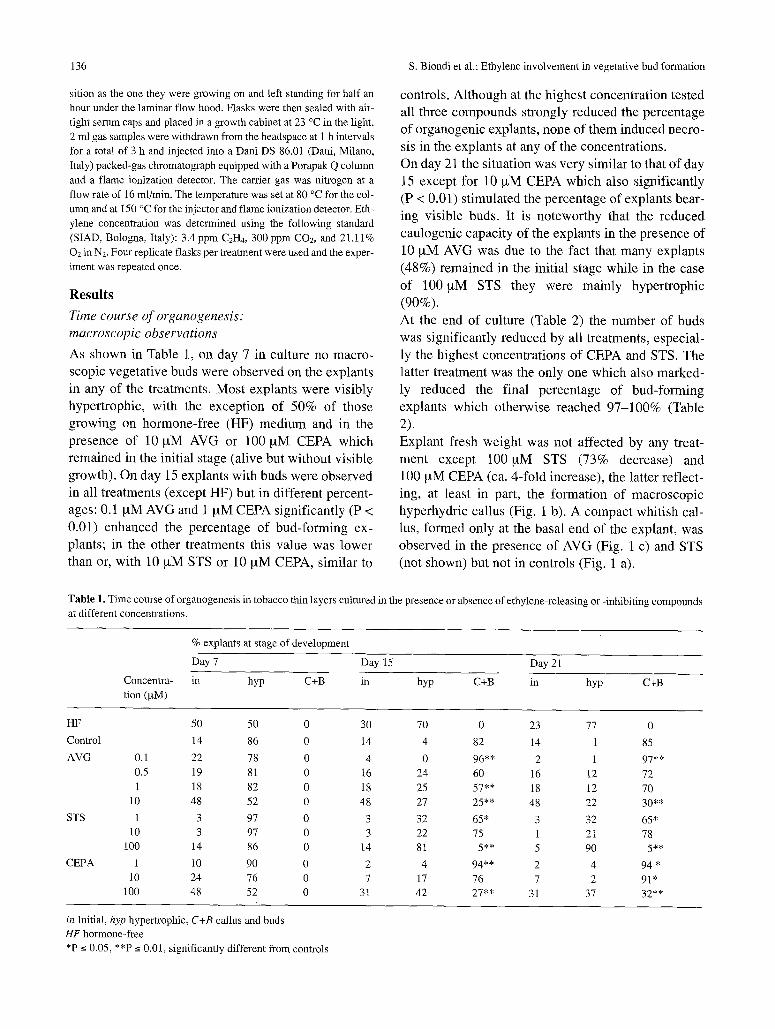
controls. Although at the highest concentration tested all three compounds strongly reduced the percentage of organogenic explants, none of them induced necro- sis in the explants at any of the concentrations. On day 21 the situation was very similar to that of day 15 except for 10 ~xM CEPA which also significantly (P < 0.01) stimulated the percentage of explants bear- ing visible buds. It is noteworthy that the reduced caulogenic capacity of the explants in the presence of 10 ~tM AVG was due to the fact that many explants (48%) remained in the initial stage while in the case of 100 ~tM STS they were mainly hypertrophic (90%). At the end of culture (Table 2) the number of buds was significantly reduced by all treatments, especial- ly the highest concentrations of CEPA and STS. The latter treatment was the only one which also marked- ly reduced the final percentage of bud-forming explants which otherwise reached 97-100% (Table 2). Explant fresh weight was not affected by any treat- ment except 100 ~tM STS (73% decrease) and 100 ~tM CEPA (ca. 4-fold increase), the latter reflect- ing, at least in part, the formation of macroscopic hyperhydric callus (Fig. 1 b). A compact whitish cal- lus, formed only at the basal end of the explant, was observed in the presence of AVG (Fig. 1 c) and STS (not shown) but not in controls (Fig. 1 a).
in the presence or absence of ethylene-releasing or -inhibiting compounds
% explants at stage of development
Day 7 Day 15 Day 21
Concentra- in hyp C+B in hyp C+B in hyp C+B tion (~tM)
HF
Control
AVG
STS
CEPA
50 50 0 30 70 0 23 77 0
14 86 0 14 4 82 14 1 85
0.1 22 78 0 4 0 96** 2 1 97** 0.5 19 81 0 16 24 60 16 12 72 1 18 82 0 18 25 57** 18 12 70
10 48 52 0 48 27 25** 48 22 30**
1 3 97 0 3 32 65* 3 32 65* 10 3 97 0 3 22 75 1 21 78
100 14 86 0 14 81 5** 5 90 5**
1 10 90 0 2 4 94** 2 4 94 * 10 24 76 0 7 17 76 7 2 91"
100 48 52 0 31 42 27** 31 37 32**
in Initial, hyp hypertrophic, C+B callus and buds HF hormone-free *P <- 0.05, **P <_ 0.01, significantly different from controls
S. Biondi et al.: Ethylene involvement in vegetative bud formation
Table 2. Fresh weight and organogenic response at the end of culture (day 27) in tobacco thin layers cultured on bud-inducing medium without or with AVG, or STS, or CEPA at different concentrations
Concentra- FW/explant % organo- number of tion (gM) (mg) genic buds/explant
explants
HF
Control
AVG
STS
CEPA
110+60 0 -
340 + 189 100 26.0 _+ 12.0
0.1 443 _+ 324 100 16.2 + 5.9* 0.5 201 _+ 51" 100 18.9 -+ 10.2" 1 214 + 102' 100 17.6 + 9.5*
10 345 +_ 214 97 17.0 + 7.8*
1 314 + 129 100 14.3 + 6.0*
10 391 + 138 100 17.2+_7.3" 100 91 + 59* 62 4.2 + 4.7*
1 599 + 350* 100 19.3 _+ 7.7* 10 353 + 255 100 19.7 -+ 10.0"
100 1238 + 894* 97 11.7 _+ 7.8*
Explant fresh weight and number of buds are means +- SD *P < 0.01, significantly different from controls HF Hormone-free
137
Time course of ethylene production
Ethy lene p roduc t ion rate by cont ro l explan ts g rowing
on b u d - i n d u c i n g med ium, af ter an ini t ia l increase
be tween day 1 and 2, r ema ined cons tant up to day 15
(Fig. 2). Explan ts g rowing in the p resence o f 0.5 ~ M
AVG exh ib i t ed the same increase up to day 2, when
e thy lene p roduc t ion was a l r eady 40% less than in
controls ; subsequen t ly p roduc t ion d ropped wel l
b e l o w cont ro l levels and was to ta l ly abo l i shed by day
15. S ince this concen t ra t ion of A V G was the lowes t one
which p r o v o k e d a dras t ic r educ t ion in e thy lene for-
ma t ion (data not shown) , it was chosen for the h i s to-
log ica l analysis .
Time course of organogenesis: histological analysis
Explan ts t rea ted with 1, 10, and 100 ,uM CEPA and
STS were h i s to log ica l ly ana lysed on days 8 and 13,
i.e., in the t ime in terva l in which the first m a c r o s c o p -
ic buds appeared , and c o m p a r e d wi th controls . In the
latter, a mean n u m b e r o f 66 mer i s t emoids and 5.5 bud
p r imord i a pe r explan t were de tec ted on day 8. Pri-
m o r d i a were to ta l ly absent in the o ther t rea tments ,
wi th the excep t ion o f 1 ~tM CEPA (about 1 pe r
explant ) , and mer i s t emoids were s ign i f i can t ly less
numerous (Fig. 3 a). The leve l o f m e r i s t e m o i d organ-
Fig. 1. Tobacco thinqayer explants cultured for 27 days on auxin- and cytokinin-containing bud-inducing medium alone (a) and sup- plemented with 100 ~tM CEPA (b) or 1 ~tM AVG (c). Bars: 1 cm
15
'7 10
5
0
0 2 4 6 8 10 12 14 16
Time (days)
Fig. 2. Time course of ethylene production by tobacco thin layers cultured on bud-inducing medium without (A) or with 0.5 vM AVG (11). Bars indicate standard deviation
isat ion, which was h igher in control than in CEPA-
and STS- t r ea t ed explants , is also e x e m p l i f i e d in Fig. 4
a and b. The mean n u m b e r o f me r i s t emo ids was not
s ign i f ican t ly d i f ferent when compar ing di f ferent
138 S. Biondi et al.: Ethylene involvement in vegetative bud formation
t00
= 80
60
40 E = 20
@ i
A
100
80
"~ 60
40
20 1 Control C1 CIO C 100 S1 S10
B
s lOO
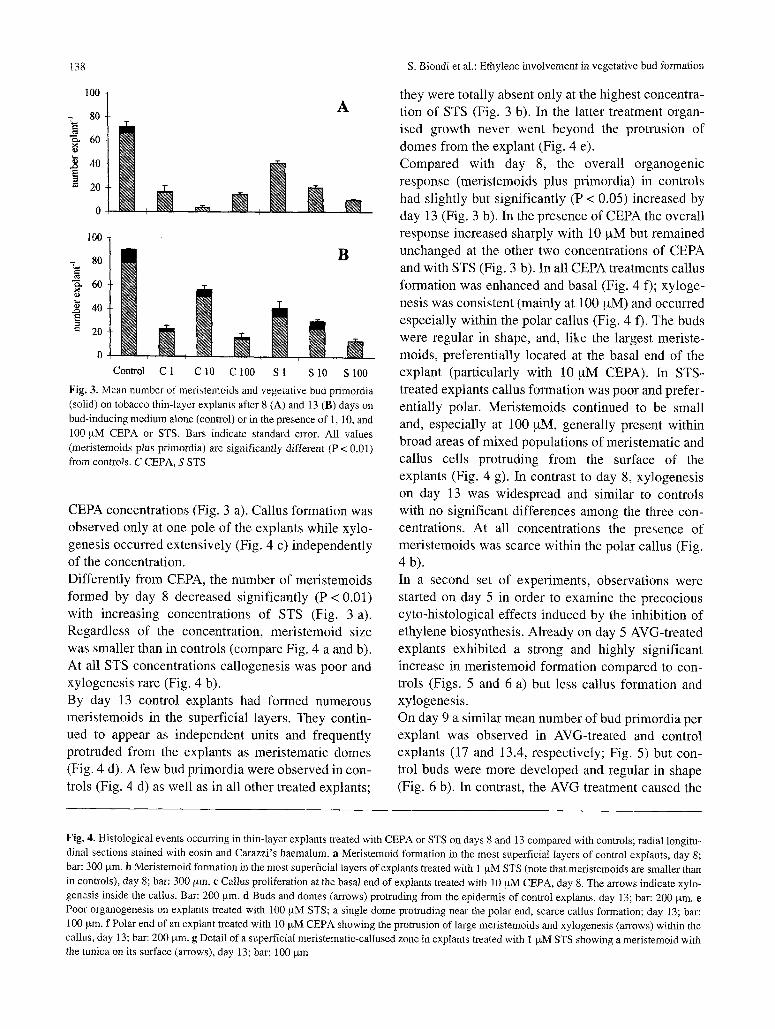
Fig. 3. Mean nmnber of meristemoids and vegetative bud primordia (solid) on tobacco thin-layer explants after 8 (A) and 13 (B) days on bud-inducing medium alone (control) or in the presence of 1, 10, and 100 ~uM CEPA or STS. Bars indicate standard error. All values (meristemoids plus primordia) are significantly different (P < 0.01) from controls. C CEPA, S STS
CEPA concentrations (Fig. 3 a). Callus formation was
observed only at one pole of the explants while xylo- genesis occurred extensively (Fig. 4 c) independently of the concentration.
Differently from CEPA, the number of meristemoids
formed by day 8 decreased significantly (P < 0.01) with increasing concentrations of STS (Fig. 3 a).
Regardless of the concentration, meristemoid size was smaller than in controls (compare Fig. 4 a and b). At all STS concentrations callogenesis was poor and xylogenesis rare (Fig. 4 b).
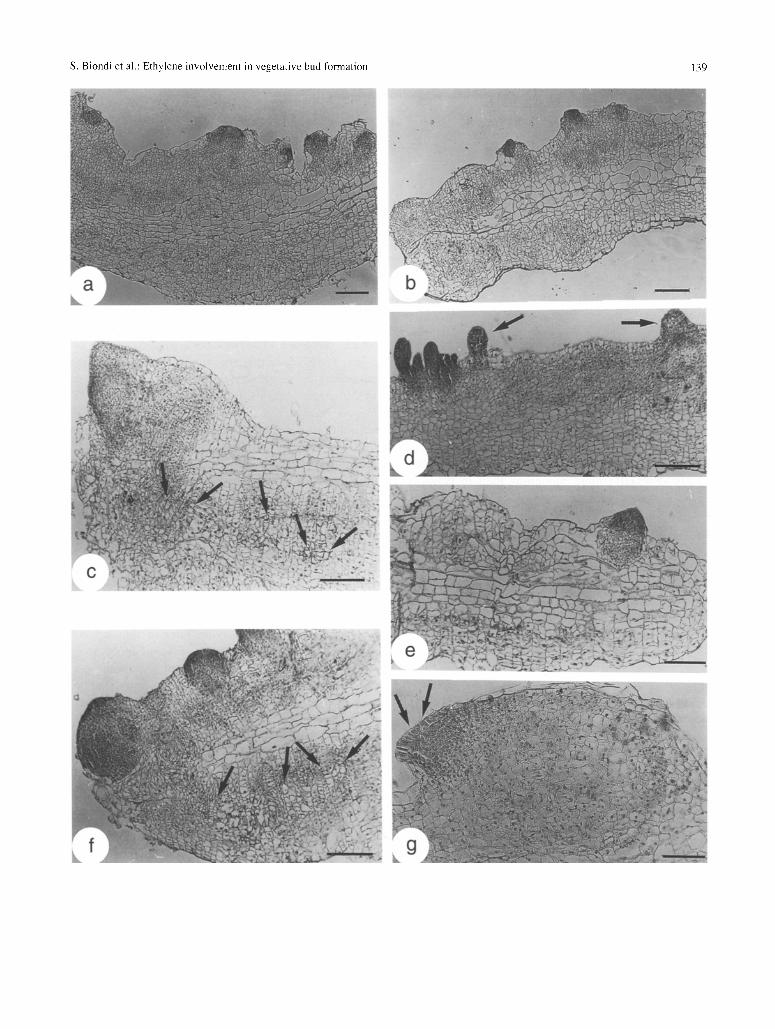
By day 13 control explants had formed numerous meristemoids in the superficial layers. They contin- ued to appear as independent units and frequently protruded from the explants as meristematic domes (Fig. 4 d). A few bud primordia were observed in con-
trols (Fig. 4 d) as well as in all other treated explants;
they were totally absent only at the highest concentra- tion of STS (Fig. 3 b). In the latter treatment organ- ised growth never went beyond the protrusion of
domes from the explant (Fig. 4 e).
Compared with day 8, the overall organogenic response (meristemoids plus primordia) in controls
had slightly but significantly (P < 0.05) increased by day 13 (Fig. 3 b). In the presence of CEPA the overall response increased sharply with 10 ~xM but remained
unchanged at the other two concentrations of CEPA and with STS (Fig. 3 b). In all CEPA treatments callus
formation was enhanced and basal (Fig. 4 f); xyloge-
nesis was consistent (mainly at 100 gM) and occurred especially within the polar callus (Fig. 4 f). The buds
were regular in shape, and, like the largest meriste- moids, preferentially located at the basal end of the explant (particularly with 10 gM CEPA). In STS-
treated explants callus formation was poor and prefer- entially polar. Meristemoids continued to be small
and, especially at 100 ~tM, generally present within broad areas of mixed populations of meristematic and
callus cells protruding from the surface of the explants (Fig. 4 g). In contrast to day 8, xylogenesis on day 13 was widespread and similar to controls with no significant differences among the three con- centrations. At all concentrations the presence of meristemoids was scarce within the polar callus (Fig. 4 b).
In a second set of experiments, observations were started on day 5 in order to examine the precocious cyto-histological effects induced by the inhibition of ethylene biosynthesis. Already on day 5 AVG-treated explants exhibited a strong and highly significant
increase in meristemoid formation compared to con-
trols (Figs. 5 and 6 a) but less callus formation and xylogenesis.
On day 9 a similar mean number of bud primordia per explant was observed in AVG-treated and control explants (17 and 13.4, respectively; Fig. 5) but con- trol buds were more developed and regular in shape
(Fig. 6 b). In contrast, the AVG treatment caused the
Fig. 4. Histological events occurring in thin-layer explants treated with CEPA or STS on days 8 and 13 compared with controls; radial longitu- dinal sections stained with eosin and Carazzi's haemalum, a Meristemoid formation in the most superficial layers of control explants, day 8; bar: 300 ~tm. b Meristemoid formation in the most superficial layers of explants treated with 1 ptM STS (note that meristemoids are smaller than in controls), day 8; bar: 300/~m. e Callus proliferation at the basal end of explants treated with 10 ~tM CEPA, day 8. The arrows indicate xylo- genesis inside the callus. Bar: 200 gm. d Buds and domes (arrows) protruding from the epidermis of control explants, day 13; bar: 200 ~tm. e Poor organogenesis on explants treated with 100 p.M STS; a single dome protruding near the polar end, scarce callus formation; day 13; bar: 100 gm. f Polar end of an explant treated with 10 p~M CEPA showing the protrusion of large meristemoids and xylogenesis (an'ows) within the callus, day 13; bar: 200 ~tm. g Detail of a superficial meristematic-callused zone in explants treated with 1 ~tM STS showing a meristemoid with the tunica on its surface (arrows), day 13; bar: 100 ~tm
S. Biondi et al.: Ethylene involvement in vegetative bud formation 139
140 S. Biondi et al.: Ethylene involvement in vegetative bud formation
180
150
"7 120
90
30
0
5 9 15 Time (days)
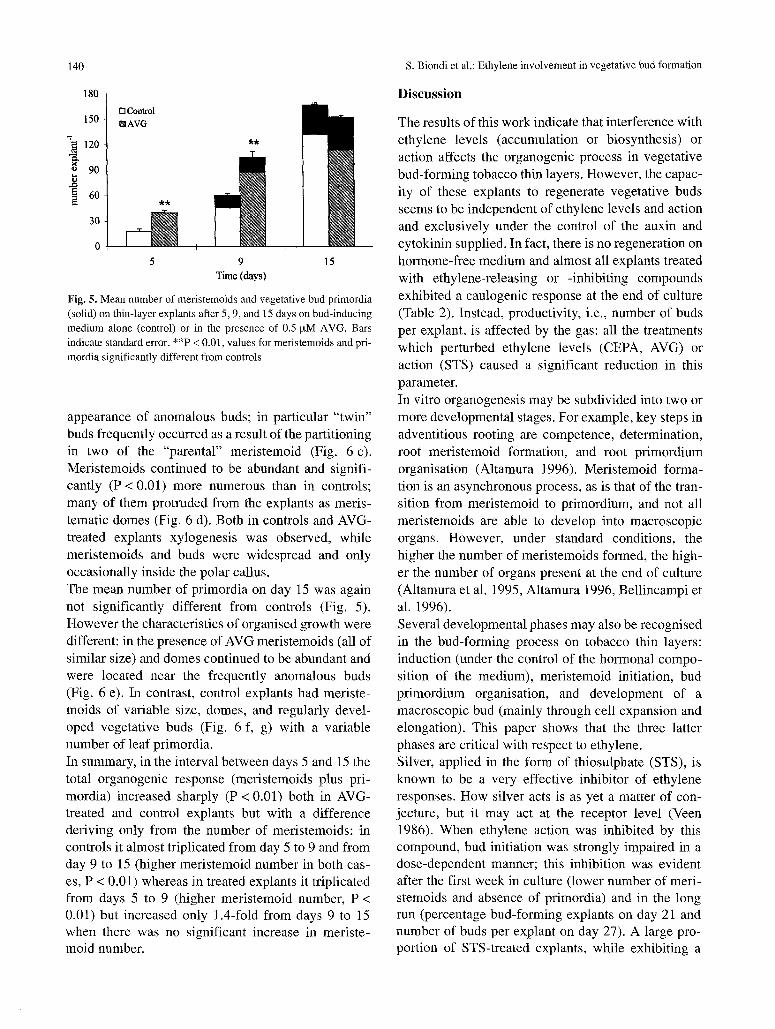
Fig. 5. Mean number of meristemoids and vegetative bud primordia (solid) on thin-layer explants after 5, 9, and 15 days on bud-inducing medium alone (control) or in the presence of 0.5 lxM AVG. Bars indicate standard error. **P < 0.01, values for meristemoids and pri- mordia significantly different from controls
appearance of anomalous buds; in particular "twin" buds frequently occurred as a result of the partitioning in two of the "parental" meristemoid (Fig. 6 c). Meristemoids continued to be abundant and signifi- cantly (P < 0.01) more numerous than in controls; many of them protruded from the explants as meris- tematic domes (Fig. 6 d). Both in controls and AVG- treated explants xylogenesis was observed, while meristemoids and buds were widespread and only occasionally inside the polar callus. The mean number of primordia on day 15 was again not significantly different from controls (Fig. 5). However the characteristics of organised growth were different: in the presence of AVG meristemoids (all of similar size) and domes continued to be abundant and were located near the frequently anomalous buds (Fig. 6 e). In contrast, control explants had meriste- moids of variable size, domes, and regularly devel- oped vegetative buds (Fig. 6 f, g) with a variable number of leaf primordia. In summary, in the interval between days 5 and 15 the total organogenic response (meristemoids plus pri- mordia) increased sharply (P < 0.01) both in AVG- treated and control explants but with a difference deriving only from the number of meristemoids: in controls it almost triplicated from day 5 to 9 and from day 9 to 15 (higher meristemoid number in both cas- es, P < 0.01) whereas in treated explants it triplicated from days 5 to 9 (higher meristemoid number, P < 0.01) but increased only 1.4-fold from days 9 to 15 when there was no significant increase in meriste- mold number.
Discussion
The results of this work indicate that interference with ethylene levels (accumulation or biosynthesis) or action affects the organogenic process in vegetative bud-forming tobacco thin layers. However, the capac- ity of these explants to regenerate vegetative buds seems to be independent of ethylene levels and action and exclusively under the control of the auxin and cytokinin supplied. In fact, there is no regeneration on hormone-free medium and almost all explants treated with ethylene-releasing or -inhibiting compounds exhibited a caulogenic response at the end of culture (Table 2). Instead, productivity, i.e., number of buds per explant, is affected by the gas: all the treatments which perturbed ethylene levels (CEPA, AVG) or action (STS) caused a significant reduction in this parameter. In vitro organogenesis may be subdivided into two or more developmental stages. For example, key steps in adventitious rooting are competence, determination, root meristemoid formation, and root primordium organisation (Altamura 1996). Meristemoid forma- tion is an asynchronous process, as is that of the tran- sition from meristemoid to primordium, and not all meristemoids are able to develop into macroscopic organs. However, under standard conditions, the higher the number of meristemoids formed, the high- er the number of organs present at the end of culture (Altamura et al. 1995, Altamura 1996, Bellincampi et al. 1996). Several developmental phases may also be recognised in the bud-forming process on tobacco thin layers: induction (under the control of the hormonal compo- sition of the medium), meristemoid initiation, bud primordium organisation, and development of a macroscopic bud (mainly through cell expansion and elongation). This paper shows that the three latter phases are critical with respect to ethylene. Silver, applied in the form of thiosulphate (STS), is known to be a very effective inhibitor of ethylene responses. How silver acts is as yet a matter of con- jecture, but it may act at the receptor level (Veen 1986). When ethylene action was inhibited by this compound, bud initiation was strongly impaired in a dose-dependent manner; this inhibition was evident after the first week in culture (lower number of meri- stemoids and absence of primordia) and in the long run (percentage bud-forming explants on day 21 and number of buds per explant on day 27). A large pro- portion of STS-treated explants, while exhibiting a
S. Biondi et al.: Ethylene involvement in vegetative bud formation 141
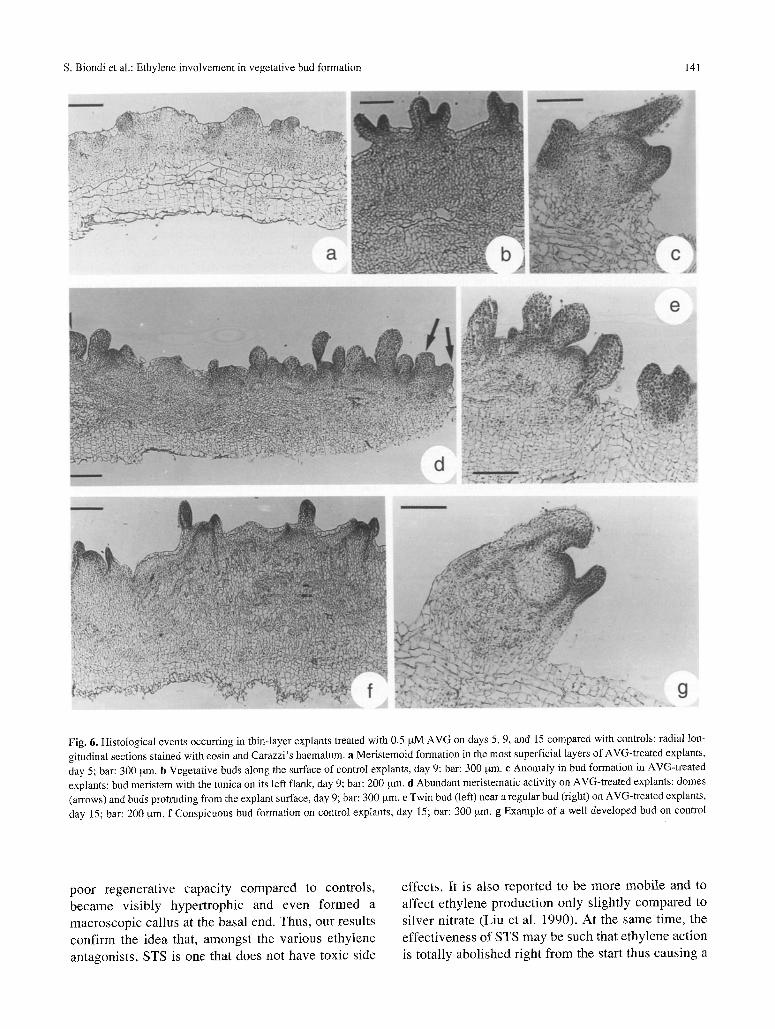
Fig. 6. Histological events occurring in thin-layer explants treated with 0.5 p.M AVG on days 5, 9, and 15 compared with controls; radial lon- gitudinal sections stained with eosin and Carazzi's haemalum, a Meristemoid formation in the most superficial layers of AVG-treated explants, day 5; bar: 300 ~tm. b Vegetative buds along the surface of control explants, day 9; bar: 300 btm. c Anomaly in bud formation in AVG-treated explants: bud meristem with the tunica on its left flank, day 9; bar: 200 [.tm. d Abundant meristematic activity on AVG-treated explants: domes (arrows) and buds protruding from the explant surface, day 9; bar: 300 [*m. e Twin bud (left) near a regular bud (right) on AVG-treated explants, day 15; bar: 200 ~xm. f Conspicuous bud formation on control expIants, day 15; bar: 300 ,u.m. g Example of a well developed bud on control
p o o r regenera t ive capac i ty c o m p a r e d to controls ,
b e c a m e v i s ib ly hype r t roph ic and even fo rmed a
m a c r o s c o p i c cal lus at the basa l end. Thus, our resul ts
conf i rm the idea that, amongs t the va r ious e thy lene
antagonis ts , STS is one that does not have toxic s ide
effects . It is a lso repor ted to be more mob i l e and to
affect e thy lene p roduc t ion only s l ight ly c o m p a r e d to
s i lver ni t rate (Liu et al. 1990). At the same t ime, the
e f fec t iveness o f STS m a y be such that e thy lene ac t ion
is to ta l ly abo l i shed r ight f rom the start thus caus ing a

142
marked inhibition already in the early stages of organogenesis (number and size of meristemoids). Our results also suggest that by blocking ethylene action organised growth is inhibited but not undiffer- entiated proliferative growth. In bud-forming tobacco thin layers AVG provoked very different effects compared to STS. A block of ethylene action was deleterious to organogenesis at all stages whereas a reduction of ethylene synthesis, while resulting in a lower number of buds per explant at the end of culture, accelerated meristemoid forma- tion. This could perhaps be explained by the fact that the small amount of ethylene produced in AVG-treat- ed explants early in culture (60% of controls on day 2) was sufficient to sustain, and even enhance meriste- moid formation up to day 8. Later, as ethylene pro- duction fell to near-zero values, meristemoid forma- tion in AVG-treated explants slowed down and fell below control values. The number of meristemoids that become bud pri- mordia was not significantly affected either by AVG (Fig. 5) or STS (Fig. 3) compared to controls, except at the highest concentration of the latter compound where it was practically null. The last stage, i.e., the transition from primordium to macroscopic, fully developed bud, was, as reflected in the final number of buds per explant (Table 2), inhibited by both com- pounds, suggesting a positive role of ethylene in this phase. Another important difference between the two inhibitors lies in the fact that buds produced in the presence of AVG were frequently anomalous (twin buds); this points to an AVG-induced perturbation at the level of the meristemoid which negatively affects normal bud ontogenesis. The formation of a large macroscopic callus at the basal end of STS- and AVG-treated explants, while only a microscopic callus is formed in controls, may support the contention that ethylene inhibits basipetal auxin transport (Burg and Burg 1967) or that it enhances auxin catabolism (Sagee etal. 1990). Inhibitors of ethylene synthesis or action may there- fore promote polar transport of auxin and/or reduce its breakdown, thus leading to an accumulation of auxin at the basal end. Meristemoids and buds were scarce within the polar callus and uniformly distrib- uted on the rest of the explant. This suggests that aux- in accumulated at the basal end may have reached supraoptimal levels for bud formation and shifted cells towards callus proliferation. The promotive action of auxin on callus formation is well known,
S. Biondi et al.: Ethylene involvement in vegetative bud formation
also for tobacco thin layers (Altamura et al. 1995). In flower-forming tobacco thin layers, organ distribution also depends on the concentration of applied auxin: with increasing concentration the number of flowers at the basal end progressively declines and callus is formed instead (Smulders et al. 1988). On the con- trary, in bulb-scale explants of Lilium speciosum AVG increases the percentage of basal buds formed while TIBA, an inhibitor of polar auxin translocation, coun- teracts this effect as does ethylene (van Aartrijk et al. 1985). Based on the above data, the authors conclud- ed that auxin and auxin transport are the key factors regulating bud induction in these explants. Our results show that in vegetative bud-forming tobacco thin lay- ers organogenesis is mainly cytokinin dependent. In fact, when cultured in the presence of auxin alone or cytokinin alone, only the latter treatment leads to meristemoid initiation (Altamura et al. 1995). CEPA also caused a reduction in meristemoid forma- tion (Fig. 3), and this reduction was reflected in the final number of buds produced (Table 2). The transi- tion from meristemoid to primordium seemed to be only slightly affected by enhanced ethylene levels. In fact, with 1 ~tM CEPA primordia were histologically observed, like in controls, on day 8; with 10 and 100 txM primordia only appeared on day 13, but by then the primordium-to-meristemoid ratio was similar to that of controls. Finally, compared to controls, bud development was accelerated in the presence of the two lower concentrations of CEPA (Table 1). Thus, cells that must undergo dedifferentiation in order to produce meristemoids are more sensitive to ethylene levels than meristematic cells that must develop into a bud. A low level of ethylene might promote a meri- stematic condition, whereas higher levels might be important for normal bud ontogenesis. In fact, differ- ent from AVG, at all concentrations of CEPA buds were always regular in shape. Increasing ethylene levels in the culture vessel by including 100 ~tM CEPA in the medium promoted a dramatic increase in fresh weight associated with abundant callus formation and xylogenesis. Suppres- sion of organised growth by CEPA in carrot suspen- sion cultures was reported by Wochok and Wetherell (1971) while Huxter et al. (1981) showed that fast- growing non-shoot-forming tobacco callus produced far more ethylene than the shoot-forming one. They concluded that low ethylene levels were associated with organ formation and that ethylene seems to play a role in maintaining tobacco tissues in the undiffer- entiated state.

S. Biondi et al.: Ethylene involvement in vegetative bud folanation 143
Ethylene is also known to play a positive role on xylem differentiation. In Lactuca sativa pith explants the addition of ethylene precursors (L-methionine and S-adenosylmethionine) or CEPA to an auxin- and cytokinin-containing medium enhanced xylogenesis, whereas ethylene inhibitors retarded it (Miller and Roberts 1984). Our present results with CEPA and STS (at least up to day 8) confirm these findings. The present study confirms the notion that the role of ethylene can be quite different depending on the developmental stage. AVG accelerated meristemoid formation early in culture but this process later slowed down resulting in a reduced organogenic response. Instead, blocking ethylene action was dele- terious at all stages. During flower bud formation on tobacco thin layers, Smulders et al. (1990) found that AVG and silver nitrate reduced the number of buds per explant on day 7, but that after 14 days there were no longer any differences. The immediate precursor of ethylene, ACC, on the other hand, increased the number of buds detectable on day 7 but decreased their final number, suggesting that it made the buds emerge earlier. Similar results by Huxter et al. (1981) indicated that in tobacco callus a low concentration of ethylene speeded up primordia emergence but did not affect the final number of shoots formed. Such find- ings on ethylene promotion of early bud outgrowth are in agreement with our results. In summary, under our culture conditions, tobacco thin layers have an excellent regenerative capacity (as indicated by the percentage of organogenic explants and by the high number of buds per explant). A fur- ther improvement may be difficult to obtain since even the faster rate of meristemoid formation induced by AVG is uninfluential on the final number of buds. Thus modification of ethylene synthesis/action is probably necessary in tissue culture systems where the regenerative capacity is suboptimal. From a prac- tical point of view, a two-phase manipulation of eth- ylene levels may be useful to improve shoot regener- ation, especially in recalcitrant species: an early application of AVG in order to promote meristemoid formation followed by an enhancement of ethylene levels (with CEPA, ACC, or other) in order to opti- raise bud development. Generally speaking the present study reveals that eth- ylene plays a role in several cytohistological events, such as meristemoid formation and bud ontogenesis, and confirms its effects on xylogenesis and callus proliferation.
Acknowledgements The authors wish to thank Prof. Nello Bagni for critical revision of the manuscript and Mr. Giovanni Bugamelli for technical assistance. This research was supported by funds from M.U,R.S.T. (60%), Italy.
References Altamura MM (1996) Root histogenesis in herbaceous and woody
explants cultured in vitro: a critical review. Agronomie 16: 589-602
- Capitani F, Serafini-Fracassini D, Torrigiani P, Falasca G (1991) Root histogenesis from tobacco thin cell layers. Protoplasma 161:31-42
- Capitani F, Falasca G, Galleli R, Scaramagli S, Bueno M, Torri- giani P, Bagni N (1995) Morphogenesis in cultured thin layers and pith explants of tobacco I: effect of putrescine on cell size, xylogenesis and meristemoid organization. J Plant Physiol 147: 101-106
Bellincampi D, Cardarelli M, Zaghi D, Serino G, Salvi G, Gatz C, Cervone F, Altamura MM, Costantino P, De Lorenzo G (1996) Oligogalacturonides prevent rhizogenesis in rolB-transformed tobacco explants by inhibiting auxin-induced expression of the rolB gene. Plant Cell 8:477-487
Biddington NL, Robinson HT (1991) Ethylene production during anther culture of Brussels sprouts (Brassica oleracea var. gem- mifera) and its relationship with factors that affect embryo pro- duction. Plant Cell Tissue Organ Cult 25:169-177
Burg SP, Burg EA (1967) Inhibition of polar auxin transport by eth- ylene. Plant Physiol 42:1224-1228
Chi GL, Pua EC (1989) Ethylene inhibitors enhanced de novo shoot regeneration from cotyledons of Brassica campestris ssp. chi- nensis (Chinese cabbage) in vitro. Plant Sci 64:243-250
- - Goh CJ (1991) Role of ethylene on de novo shoot regeneration from cotyledonary explants of Brassica campestris ssp. pekinen-
sis (Lour) Olsson in vitro. Plant Physiol 96:178-183 Chraibi KMB, Latche A, Roustan JP, Fallot J (1991) Stimulation of
shoot regeneration from cotyledons of Helianthus annuus by the ethylene inhibitors, silver and cobalt. Plant Cell Rep 10: 204-207
Heylen C, Vendrig JC (1991) Further studies on the effects of differ- ent auxins and cytokinins on flower formation in thin cell layers of Nicotiana tabacum: the effects of zeatin riboside, dihy- drozeatin and dihydrozeatin riboside. Physiol Plant 83:574-577
Huxter TJ, Thorpe TA, Reid DM (1981 ) Shoot initiation in light- and dark-grown tobacco callus: the role of ethylene. Physiol Plant 53:319-326
Jackson MB, Abbot AJ, Belcher AR, Hall KC, Butler R, Cameron J (1991) Ventilation in plant tissue cultures and effects of poor aeration on ethylene and carbon dioxide accumulation, oxygen depletion and explant development. Ann Bot 67:229-237
Kumar PP, Reid DM, Thorpe TA (1987) The role of ethylene and carbon dioxide in differentiation of shoot buds in excised cotyle- dons of Pinus radiata in vitro. Physiol Plant 69:244-252
Liu J-H, Reid DM (1990) Auxin and ethylene-stimulated adventi- tious rooting in relation to tissue sensitivity to auxin and ethyl- ene production in sunflower hypocotyls. J Exp Bot 43: 1191-1198
- Mukherjee I, Reid DM (1990) Adventitious rooting in hypo- cotyls of sunflower (Helianthus annuus) seedlings III: the role of ethylene. Physiol Plant 78:268-276

144
Miller RA, Roberts LW (1984) Ethylene biosynthesis and xylogene- sis in Lactuca pith explants cultured in vitro in the presence of auxin and cytokinin: the effect of ethylene precursors and inhibitors. J Exp Bot 35:691-698
Murashige T, Skoog F (1962) Revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant 15:473-497
Nissen P (1994) Stimulation of somatic embryogenesis in carrot by ethylene: effects of modulators of ethylene biosynthesis and action. Physiol Plant 92:397.403
Peeters AJM, Gerards W, Barendse GWM, Wullems GJ (1991) In vitro flower bud formation in tobacco: interaction of hormones. Plant Phys 97:402.408
Perez-Bermudez P, Comejo MJ, Segura J (1985) A morphogenic role for ethylene in hypocotyl cultures of Digitalis obscura L.
Plant Cell Rep 4:188-190 Pumhauser L, Medgyesy P, Czako M, Dix PJ, Morton L (1987)
Stimulation of shoot regeneration in Triticum aestivum and Nicotiana ptumbaginifolia Viv.: tissue cultures using the ethyl- ene inhibitor AgNO3. Plant Cell Rep 6:1 .4
Roustan JP, Latche A, Fallot J (1989) Stimulation of Daucus carota
somatic embryogenesis by inhibitors of ethylene synthesis: cobalt and nickel. Plant Cell Rep 8:182-185
- - - (1990) Control of carrot somatic embryogenesis by AgNO3, an inhibitor of ethylene action: effect on arginine decarboxylase activity. Plant Sci 67:89-95
Sagee O, Riov J, Goren R (1990) Ethylene-enhanced catabolism of [14C]indole-3-acetic acid to indole-3-carboxylic acid in citrus leaf tissues. Plant Physiol 91: 5 4 6 0
Salisbury FB, Gillespie L, Rorabaugh PA (1988) Gravitropism in higher plant shoots V: changing sensitivity to auxin. Plant Phys- iol 88:1186-1194
Scaramagli S, Bueno M, Torrigiani P, Altamura MM, Capitani F, Bagni N (1995) Morphogenesis in cultured thin layers and pith explants of tobacco II: emly hormone-modulated polyamine biosynthesis. J Plant Physiol 147:113-117
Smulders MJM, Croes AF, Wullems GJ (1988) Polar transport of 1-
S. Biondi et al.: Ethylene involvement in vegetative bud formation
naphthaleneacetic acid determines the distribution of flower buds on explants of tobacco. Plant Physiol 88:752-756
- Kemp A, Barendse GWM, Croes AF, Wullems GJ (1990) Role of ethylene in auxin-induced flower bud formation in tobacco explants. Physiol Plant 78:167-172
Taylor PWJ, Hian-Lien K, Fraser TA, Masel N, Adkins SW (1994) Effect of silver nitrate on sugarcane cell suspension growth, pro- toplast isolation, ethylene production and shoot regeneration from cell suspension cultures. J Exp Bot 45:1163-1168
Torrigiani P, Altamura MM, Pasqua G, Monacelli B, Seraflni-Fra- cassini D, Bagni N (I987) Free and conjugated polyamines dur- ing de novo floral and vegetative bud formation in thin cell lay- ers of tobacco. Physiol Plant 70:453.460
- - Capitani F, Serafini-Fracassini D, Bagni N (1989) De novo root formation in thin cell layers of tobacco: changes in free and bound polyamines. Physiol Plant 77:294-301
- - Scaramagli S, Capitani F, Falasca G, Bagni N (1993) Regula- tion of rhizogenesis by polyamines in tobacco thin layers. J Plant Physiol 142:81-87
Tran Thanh Van K, Dien NT, Chlyah A (1974) Regulation of organogenesis in small explants of superficial tissue of Nico- tiana tabacum L. Planta 119:149-159
Van Aartrijk J, Blom-Barnhoorn GJ, Bruinsma J (1985) Adventi- tious bud formation from bulb-scale explants of Lilium specio- sum Thunb. in vitro: effects of aminoethoxyvinylglycine, 1- aminocyclopropane-l-carboxylic acid, and ethylene. J Plant Physiol 117:401-410
Van der Krieken WM, Croes AF, Smulders M/M, Wullems GJ (1990) Cytokinins and flower bud formation in vitro in tobacco: role of the metabolites. Plant Physiol 92:565-569
Veen H (1986) A theoretical model for anti-ethylene effects of silver thiosulphate and 2,5-norbornadiene. Acta Hortic 181:129-134
Wochok ZS, Wetherell DF (1971) Suppression of organized growth in cultured wild carrot tissue by 2-chloroethylphosphonic acid. Plant Cell Physiol 12:771-774




















