
An auxin signaling network translates low-sugar-state input ...
Upload
independentCategory
view
0download
0
JMB—MS 688 Cust. Ref. No. PEW 076/95 [SGML]
J. Mol. Biol. (1995) 251, 533–549
The PS-IAA4/5 -like Family of Early Auxin-induciblemRNAs in Arabidopsis thaliana
Steffen Abel, Minh D. Nguyen and Athanasios Theologis*
Plant Gene Expression The plant hormone auxin transcriptionally activates early genes. We haveisolated a 14-member family of DNA sequences complementary toCenter, 800 Buchanan Streetindoleacetic acid (IAA)-inducible transcripts in Arabidopsis thaliana. TheAlbany, CA 94710, USAcorresponding genes, IAA1 to IAA14, are homologs of PS-IAA4/5 andPS-IAA6 from pea, Aux22 and Aux28 from soybean, ARG3 and ARG4 frommungbean, and AtAux2-11 and AtAux2-27 from Arabidopsis. The membersof the family are differentially expressed in mature Arabidopsis plants.Characterization of IAA gene expression in etiolated seedlings demonstratesspecificity for auxin inducibility. The response of most family members toIAA is rapid (within 4 to 30 minutes) and insensitive to cycloheximide.Cycloheximide alone induces all the early genes. Auxin-induction of two lategenes, IAA7 and IAA8, is inhibited by cycloheximide, indicating requirementof protein synthesis for their activation. All IAA genes display a biphasic doseresponse that is optimal at 10 mM IAA. However, individual genes responddifferentially between 10 nM and 5 mM IAA. Expression of all genes isdefective in the Arabidopsis auxin-resistant mutant lines axr1, axr2 and aux1.
The encoded polypeptides share four conserved domains, and seveninvariant residues in the intervening regions. The spacers vary considerablyin length, rendering the calculated molecular mass of IAA proteins to rangefrom 19 kDa to 36 kDa. Overall sequence identity between members of thefamily is highly variable (36 to 87%). Their most significant structuralfeatures are functional nuclear transport signals, and a putative baa-foldwhose modeled three dimensional structure appears to be compatible withthe prokaryotic b-ribbon DNA recognition motif. The data suggest that auxininduces in a differential and hierarchical fashion a large family of early genesthat encode a structurally diverse class of nuclear proteins. These proteinsare proposed to mediate tissue-specific and cell-type restricted responses tothe hormone during plant growth and development.
7 1995 Academic Press Limited
Keywords: auxin action; early response genes; multigene family; nuclearproteins; Arabidopsis thaliana*Corresponding author
Introduction
The growth hormone auxin typified by IAA,appears to control a plethora of processes during thelife-cycle of a plant such as apical dominance,tropisms, vascular tissue differentiation, or lateral
and adventitious root formation (Went & Thimann,1937). The hormone profoundly effects cell turgor,cell elongation, cell division and cell differentiation,the shaping forces in morphogenesis and oncogene-sis (Went & Thimann, 1937; Estelle, 1992). At themolecular level, auxin exerts its regulatory role mostlikely by modulating gene expression (Guilfoyle,1986; Theologis, 1986) and membrane function(Blatt & Thiel, 1993; Assmann, 1993). Despite itsfundamental role in plant growth and development,the molecular mechanisms of auxin action are notunderstood.
Auxin-mediated cell elongation is one of the fastesthormonal responses known (with a lag period of 15to 25 minutes) and is associated with rapid changes
Abbreviations used: ABA, abscisic acid; ACC,1-aminocyclopropane-1-carboxylate; a-NAA,naphthalene-1-acetic acid; BA, benzyl adenine; CHX,cycloheximide; EST, expressed sequence tag; GA,gibberellic acid, GUS, b-glucuronidase; IAA,indole-3-acetic acid; NLS, nuclear localization signal;ORF, open reading frame; PAA, phenylacetic acid; PCR,polymerase chain reaction; SAUR, small auxin up RNA;2,4-D, 2,4-dichlorophenoxyacetic acid.
0022–2836/95/340533–17 $12.00/0 7 1995 Academic Press Limited

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis534
in the expression of a select set of early genes(Guilfoyle, 1986; Theologis, 1986). We have beenfocusing our efforts to understand how auxinpromotes plant cell growth using two earlyauxin-induced genes, PS-IAA4/5 and PS-IAA6, frompea. Early genes have successfully been used asbifocal molecular probes to elucidate signaltransduction pathways of growth factors in animalcells (Herschman, 1991). Their promoter responseelements provide a platform for exploring theintervening steps between gene activation and signalperception, whereas their encoded products mostlikely play a crucial role in downstream signalingevents (Herschman, 1991).
The PS-IAA4/5 and PS-IAA6 genes qualify asprimary response genes (Theologis et al., 1985). Bothgenes have been structurally characterized (Oelleret al., 1993), and the auxin-responsive promoterelement of PS-IAA4/5 has been functionallyidentified (Ballas et al., 1993, 1995). The encodedproteins, PS-IAA4 and PS-IAA6, share similarphysical properties and extensive amino acidsequence identity in four domains, which areconserved in polypeptides encoded by related earlyauxin-inducible genes, Aux22 and Aux28 fromsoybean (Ainley et al., 1988), ARG3 and ARG4 frommung bean (Yamamoto et al., 1992), and AtAux2-11and AtAux2-27 from Arabidopsis (Conner et al., 1990).PS-IAA4 and PS-IAA6 are short-lived nuclearproteins that contain functional NLS and a putativebaa-fold reminiscent of the b-ribbon DNA recog-nition motif of prokaryotic repressor polypeptides(Abel et al., 1994; Abel & Theologis, 1995; Oeller &Theologis, 1995). These properties suggest aregulatory function of PS-IAA4-like polypeptides inauxin-induced events responsible for plant cellgrowth. However, no direct evidence has beenobtained thus far.
As for any protein, the most appropriate approachto study the biological function of PS-IAA4-likepolypeptides is complete inhibition of their ex-pression by targeted gene replacement (Scherer &Davis, 1979). Since this technique is not applicable yetfor plants, reverse genetic approaches such asconstruction of mutants by overexpressing sense orantisense RNA to manipulate levels of geneexpression (Gray et al., 1992; Napoli et al., 1990) or byinsertional mutagenesis (Osborne & Baker, 1995) arethe current methods used. To study the function ofPS-IAA4/5-like gene products by genetic and reversegenetic means, Arabidopsis is the plant of choice(Meyerowitz, 1989).
As a first step toward this long-term goal, wedescribe the isolation, structure and expressioncharacteristics of a large family of PS-IAA4/5-liketranscripts in Arabidopsis thaliana, designated IAA1 toIAA14 for IAA-inducible. This family provides asource of diverse probes to elucidate the function ofPS-IAA4-like proteins in auxin-regulated growthprocesses by reverse genetic, molecular andbiochemical approaches. The putative function ofthis divergent family of nuclear proteins in auxinsignal transduction is discussed.
Results
Identification of a PS-IAA4/5 -like gene family inArabidopsis
A recent immunochemical study indicates thatPS-IAA4-like proteins are encoded by a smallmultigene family in various plant species (Oeller &Theologis, 1995). To identify additional PS-IAA4/5-like genes in Arabidopsis, we applied a PCR-basedapproach using degenerate oligonucleotides, DC2and DC4, corresponding to discrete and highlyconserved regions in domain II and domain IV ofPS-IAA4-like polypeptides (see Figure 10).
Amplification of PS-IAA4/5-like sequences fromvarious plant species
A complex but similar pattern of multiple DNAfragments is amplified in a PCR with genomic DNAfrom various plant species (Figure 1, panel IA). ThePCR products of Arabidopsis, cauliflower, pea,tomato, maize and rice range in size from 240 bp toabout 1 kb (lanes 2 to 7). The product pattern isnotably similar between Arabidopsis and cauliflower,members of the Cruciferae family (compare lane 2with lane 3), or between maize and rice, members ofthe Graminae family (compare lane 6 with lane 7).Southern blot hybridization was performed with adegenerate oligomer, DC3, which corresponds to themotif KVSM/V/I/DG in domain III of PS-IAA4-likeproteins (see Figure 10). The probe recognizes adistinct set of amplified DNA fragments for eachplant species tested (Figure 1, panel IB, lanes 2 to 7).This indicates that PS-IAA4-like polypeptides areencoded by multigene families in monocotyledonousand dicotyledonous plants.
Classification of the Arabidopsis PCR products
Twelve major DNA fragments ranging in size from250 bp to 550 bp, were reproducibly amplified in aPCR with Arabidopsis genomic DNA (Figure 1, panelIA, lane 2). DNA inserts of 72 transformants werecharacterized by cross-hybridization at high strin-gency, revealing 12 classes of non-cross-hybridizingDNA fragments. DNA sequencing identified 11unique fragments related to PS-IAA4/5-like genes.Using a pool of these 11 genomic fragments as aprobe, substractive screening by colony hybridiz-ation of an additional 280 transformants identifiedanother three unique DNA fragments of thePS-IAA4/5-type. The amino acid sequence deducedfrom each of the 14 genomic DNA products is similarto the region between domain II and domain IV ofPS-IAA4-like proteins (see Figure 10). Five PS-IAA4/5-type genomic fragments contain a single ORF,whereas a putative intron interrupts the codingregion of the other nine gene fragments at aconserved position (Table 1; and see Figure 10).
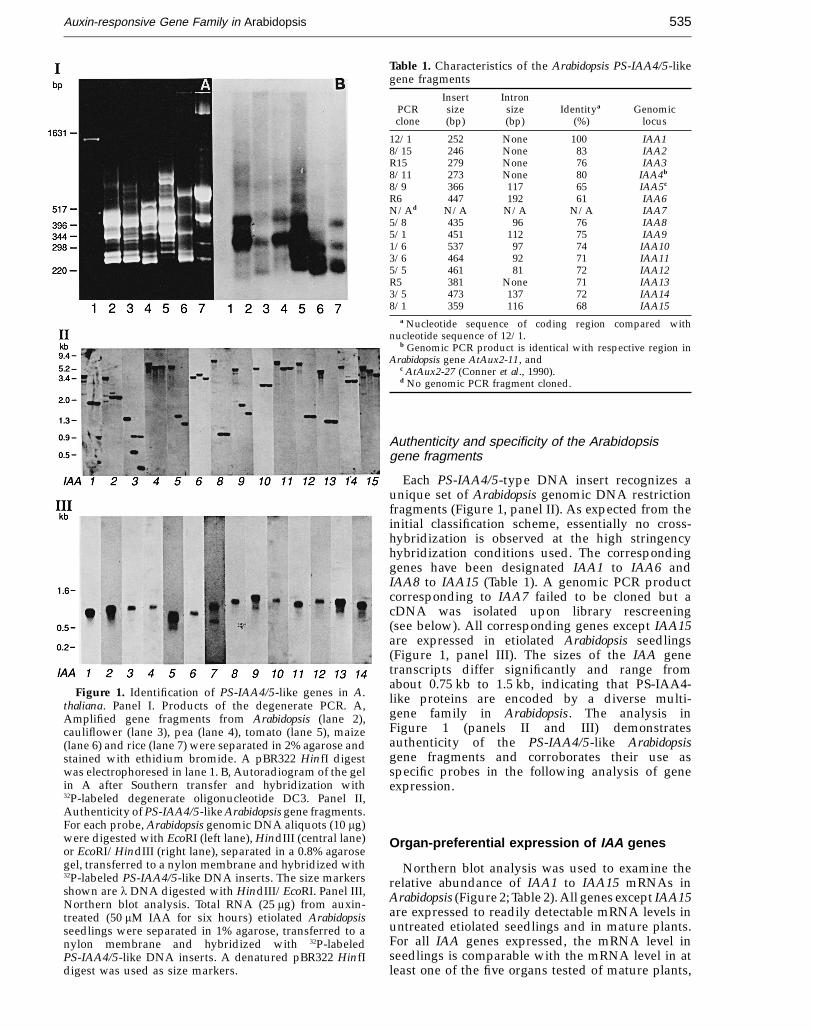
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 535
Figure 1. Identification of PS-IAA4/5-like genes in A.thaliana. Panel I. Products of the degenerate PCR. A,Amplified gene fragments from Arabidopsis (lane 2),cauliflower (lane 3), pea (lane 4), tomato (lane 5), maize(lane 6) and rice (lane 7) were separated in 2% agarose andstained with ethidium bromide. A pBR322 HinfI digestwas electrophoresed in lane 1. B, Autoradiogram of the gelin A after Southern transfer and hybridization with32P-labeled degenerate oligonucleotide DC3. Panel II,Authenticity of PS-IAA4/5-like Arabidopsis gene fragments.For each probe, Arabidopsis genomic DNA aliquots (10 mg)were digested with EcoRI (left lane), HindIII (central lane)or EcoRI/HindIII (right lane), separated in a 0.8% agarosegel, transferred to a nylon membrane and hybridized with32P-labeled PS-IAA4/5-like DNA inserts. The size markersshown are l DNA digested with HindIII/EcoRI. Panel III,Northern blot analysis. Total RNA (25 mg) from auxin-treated (50 mM IAA for six hours) etiolated Arabidopsisseedlings were separated in 1% agarose, transferred to anylon membrane and hybridized with 32P-labeledPS-IAA4/5-like DNA inserts. A denatured pBR322 HinfIdigest was used as size markers.
Table 1. Characteristics of the Arabidopsis PS-IAA4/5-likegene fragments
Insert IntronPCR size size Identitya Genomicclone (bp) (bp) (%) locus
12/1 252 None 100 IAA18/15 246 None 83 IAA2R15 279 None 76 IAA38/11 273 None 80 IAA4b
8/9 366 117 65 IAA5c
R6 447 192 61 IAA6N/Ad N/A N/A N/A IAA75/8 435 96 76 IAA85/1 451 112 75 IAA91/6 537 97 74 IAA103/6 464 92 71 IAA115/5 461 81 72 IAA12R5 381 None 71 IAA133/5 473 137 72 IAA148/1 359 116 68 IAA15
a Nucleotide sequence of coding region compared withnucleotide sequence of 12/1.
b Genomic PCR product is identical with respective region inArabidopsis gene AtAux2-11, and
c AtAux2-27 (Conner et al., 1990).d No genomic PCR fragment cloned.
Authenticity and specificity of the Arabidopsisgene fragments
Each PS-IAA4/5-type DNA insert recognizes aunique set of Arabidopsis genomic DNA restrictionfragments (Figure 1, panel II). As expected from theinitial classification scheme, essentially no cross-hybridization is observed at the high stringencyhybridization conditions used. The correspondinggenes have been designated IAA1 to IAA6 andIAA8 to IAA15 (Table 1). A genomic PCR productcorresponding to IAA7 failed to be cloned but acDNA was isolated upon library rescreening(see below). All corresponding genes except IAA15are expressed in etiolated Arabidopsis seedlings(Figure 1, panel III). The sizes of the IAA genetranscripts differ significantly and range fromabout 0.75 kb to 1.5 kb, indicating that PS-IAA4-like proteins are encoded by a diverse multi-gene family in Arabidopsis. The analysis inFigure 1 (panels II and III) demonstratesauthenticity of the PS-IAA4/5-like Arabidopsisgene fragments and corroborates their use asspecific probes in the following analysis of geneexpression.
Organ-preferential expression of IAA genes
Northern blot analysis was used to examine therelative abundance of IAA1 to IAA15 mRNAs inArabidopsis (Figure 2; Table 2). All genes except IAA15are expressed to readily detectable mRNA levels inuntreated etiolated seedlings and in mature plants.For all IAA genes expressed, the mRNA level inseedlings is comparable with the mRNA level in atleast one of the five organs tested of mature plants,

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis536
Figure 2. Expression of IAA genes in Arabidopsis plants.Poly(A)+ RNA (2.5 mg) isolated from untreated etiolatedseedlings (5 days old) and from various organs oflight-grown mature plants (25 days old) were separated in1% agarose, transferred to a nylon membrane andhybridized with 32P-labeled PS-IAA4/5-like DNA inserts orwith a 32P-labeled 17S rDNA probe. The lanes are: 1,etiolated seedlings; 2, roots; 3, stems; 4, leaves; 5, flowers;and 6, siliques. Exposure times: six hours, IAA7, IAA8,IAA9 and IAA13; 24 hours, IAA1, IAA2, IAA4, IAA6,IAA10, IAA11, IAA12 and IAA14; 72 hours, IAA3 andIAA5; 144 hours, IAA15.
abundance in roots. Low levels of mRNA arefound for IAA3 and IAA5 to IAA7 in leaves, andfor IAA14 in stems. Members of the IAA gene familyfall into four groups according to their organ-prefer-ential expression profile: (1) IAA3, IAA5 to IAA8and IAA11 are highly abundant in stems andflowers; (2) IAA1, IAA4, IAA9 and IAA12 arepreferentially expressed in stems, leaves and flowers;(3) IAA2 and IAA10 are more abundant in vegetativethan in reproductive organs; and (4) the expressionpatterns of IAA13 and IAA14 differ from the otherIAA genes.
Response of the gene family to IAA and CHX
The effect of IAA and of the protein synthesisinhibitor CHX was examined on IAA gene expression(Figure 3). Treatment of intact etiolated Arabidopsisseedlings with 20 mM IAA for one hour (Figure 3A,lane 3) increases steady-state mRNA levels for allIAA genes tested, except for IAA7 and IAA8, whencompared with untreated (lane 1) or mock-treated(lane 2) seedlings. Accumulation of IAA mRNAsranges from a two-fold increase for IAA9 to a 25-foldincrease for IAA5. Incubation with 50 mM CHXresults in accumulation of high levels of IAA mRNA,again with the exception of IAA7 and IAA8(compare lane 2 with lane 4). At this concen-tration, CHX effectively prevents protein biosyn-thesis (data not shown). In general, IAA mRNAaccumulation is more profound in response toCHX than to IAA (compare lanes 3 and 4). CHXdoes not interfere with the effect of IAA (comparelane 3 with lane 5). Insensitivity of the auxinresponse to CHX is best revealed by an augmentedresponse to both chemicals for some of the IAAgenes, such as IAA1, IAA2, IAA5 or IAA6 (comparelane 4 with lane 5). These results qualify most of theIAA genes as primary response genes. Theirexpression is independent of de novo proteinsynthesis.
The IAA mRNA accumulation profiles inresponse to IAA and/or CHX treatment for one hour(Figure 3A) are essentially maintained whenseedlings are treated for six hours (panel B).However, two significant variations are noticed.Firstly, mRNA levels of IAA7 and IAA8 increase(about threefold and 1.5-fold, respectively) inresponse to IAA (compare lane 2 with lane 3 in panelB). CHX alone has no effect on IAA7 and IAA8(compare lane 2 with lane 4) but inhibits the responseto auxin when administered together with thehormone (compare lane 3 with lane 5). IAA7 andIAA8 are late genes whose expression appears to bedependent on de novo protein synthesis. Secondly,IAA does not significantly increase IAA3 mRNA orIAA6 mRNA levels after a six hour treatment.Interestingly, the transcript levels are comparablewith the mRNA concentrations of the mock control(compare lane 2 with lane 3 in both panels). Thisindicates transient response of gene expression toauxin.
i.e. in roots, stems, leaves, flowers or siliques (Fig-ure 2, compare lane 1 with lanes 2 to 6). In general,IAA1 to IAA14 mRNAs are highly abundant inflowers but are significantly less abundant indeveloping siliques (compare lanes 5 and 6).Interestingly, a comparison of the expressionpattern between the family members revealsdifferential preferences for the other threeorgans tested. For example, the mRNAs of IAA1,IAA3 to IAA7 and IAA12 are in relatively low
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 537
Table 2. Characteristics of the Arabidopsis IAA cDNAsGenBank Insert RUUUR/ Relative Amino Calculated
cDNA accession size Integrity DST abundance acids molecularclone numbera (bp) of cDNA motifs (mRNA)b (no.) mass (kDa)
IAA1 L15448 504 N/Ac 1/0d + + 168 19.1IAA2 L15449 522 N/Ac 2/0d + + 174 19.9IAA3 U18406 1062 Poly(A) 1/0 + 189 21.5IAA4 L15450 558 N/Ac 0/0 + + 186 21.0IAA5 U18407 578 Truncated 0/0 + 174 19.6IAA6 U18408 879 No poly(A) 2/0 + 189 21.0IAA7 U18409 947 No poly(A) 0/0 + + 243 26.4IAA8 U18410 1342 Poly(A) 2/0 + + + 321 35.1IAA9 U18411 1450 Poly(A) 0/0 + + + 338 36.4IAA10 U18412 1268 Poly(A) 1/0 + + 261 27.9IAA11 U18413 1013 Poly(A) 1/0 + + 246 26.5IAA12 U18414 1174 Poly(A) 3/1 + 239 26.3IAA13 U18415 1341 No poly(A) 1/1 + + 246 26.6IAA14 U18417 754 5'-Truncated/Poly(A) 1/0 + N/A N/A
a Additional Accession Numbers are: U18418 (genomic PCR fragment of IAA15); T04508, T20545, and Z47584 (novel PS-IAA4/5-likeArabidopsis EST’s).
b Based on Figure 2.c Cloning of coding region by PCR.d Based on inverse PCR product.
Specificity of the hormonal response
The specificity of the hormonal response isdocumented for the IAA1 gene in Figure 4.Qualitatively the same pattern of expression isfound for genes IAA2 to IAA14 (data not shown).IAA15 mRNA was not detected under anyconditions. The effect of various IAA analogs(20 mM each) is shown for 2,4-D (lane 6), a-NAA (lane7), PAA (lane 8) and L-tryptophan (lane 9). Ascompared with the response to 20 mM IAA(compare lane 1 or lane 2 with lane 3; or lane 13 withlane 14), 2,4-D and a-NAA are highly effective ininducing accumulation of IAA1 mRNAs. PAA hasonly a slight effect (compare lane 8 with lane 2),whereas the structural analog tryptophan isineffective. Other plant hormones such as 20 mMABA (lane 15), 20 mM GA (lane 16), 20 mM BA(lane 17) or 10 ppm ethylene (compare lane 21 withlane 23) do not significantly increase IAA1 mRNAsabove control levels (lane 13). A combination of20 mM IAA with 20 mM BA neither augments norattenuates the response to auxin (compare lane 13with lane 18).
The effect of several stress conditions on IAAgene expression was also studied. Wounding(lane 10), osmotic shock (lane 11), heat shock(lane 12), anaerobiosis (lane 22), and phosphatestarvation (lane 25) fail to induce IAA1 transcriptaccumulation. LiCl, which is known to inter-fere with phosphatidylinositol metabolism andsignaling (Berridge et al., 1989), has no effect,either when applied alone (lane 20) or incombination with 20 mM IAA and 20 mM benzyladenine (lane 19). The only other inducer found isinhibition of protein synthesis by CHX (lane 3 andlane 4).
Differential kinetics of mRNA accumulation inresponse to IAA
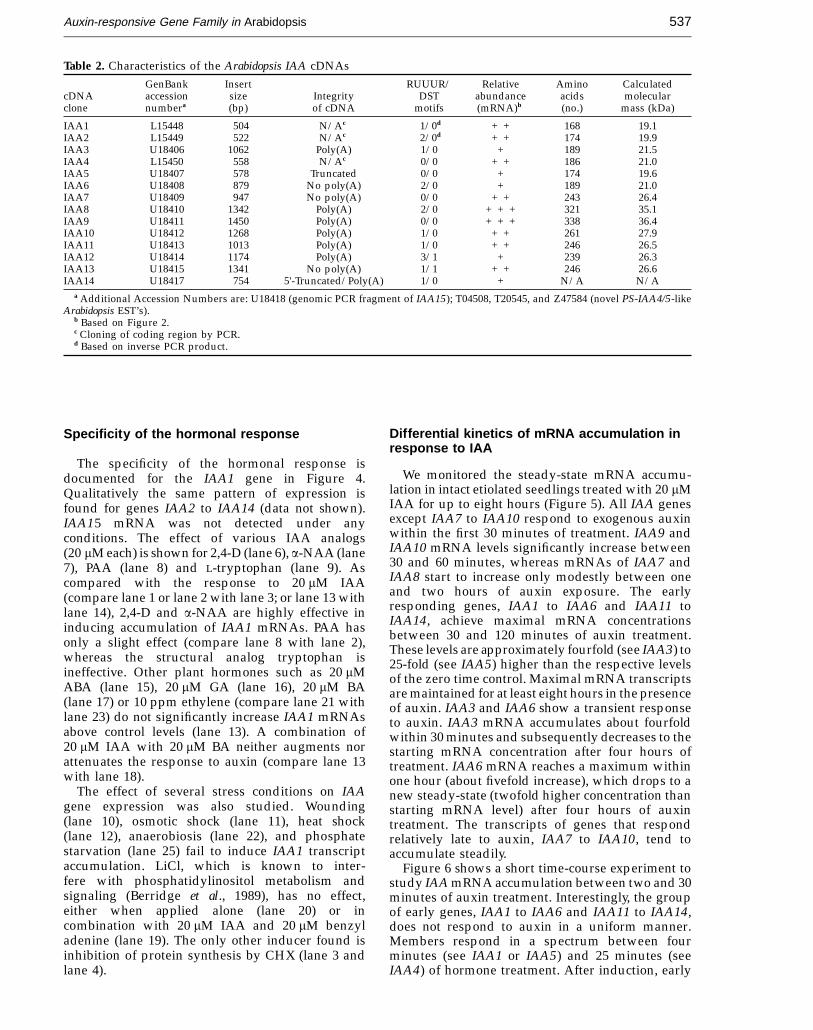
We monitored the steady-state mRNA accumu-lation in intact etiolated seedlings treated with 20 mMIAA for up to eight hours (Figure 5). All IAA genesexcept IAA7 to IAA10 respond to exogenous auxinwithin the first 30 minutes of treatment. IAA9 andIAA10 mRNA levels significantly increase between30 and 60 minutes, whereas mRNAs of IAA7 andIAA8 start to increase only modestly between oneand two hours of auxin exposure. The earlyresponding genes, IAA1 to IAA6 and IAA11 toIAA14, achieve maximal mRNA concentrationsbetween 30 and 120 minutes of auxin treatment.These levels are approximately fourfold (see IAA3) to25-fold (see IAA5) higher than the respective levelsof the zero time control. Maximal mRNA transcriptsare maintained for at least eight hours in the presenceof auxin. IAA3 and IAA6 show a transient responseto auxin. IAA3 mRNA accumulates about fourfoldwithin 30 minutes and subsequently decreases to thestarting mRNA concentration after four hours oftreatment. IAA6 mRNA reaches a maximum withinone hour (about fivefold increase), which drops to anew steady-state (twofold higher concentration thanstarting mRNA level) after four hours of auxintreatment. The transcripts of genes that respondrelatively late to auxin, IAA7 to IAA10, tend toaccumulate steadily.
Figure 6 shows a short time-course experiment tostudy IAA mRNA accumulation between two and 30minutes of auxin treatment. Interestingly, the groupof early genes, IAA1 to IAA6 and IAA11 to IAA14,does not respond to auxin in a uniform manner.Members respond in a spectrum between fourminutes (see IAA1 or IAA5) and 25 minutes (seeIAA4) of hormone treatment. After induction, early

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis538
Figure 3. Response to IAA and CHX. Total RNA (25 mg)isolated from intact etiolated seedlings (5 days old) wasprobed as in Figure 2. The seedlings were treated either forone hour (A) or for six hours (B). The lanes are: 1, untreated;2, control-treated; 3, 20 mM IAA; 4, 50 mM CHX; 5, 20 mMIAA and 50 mM CHX after 30 minutes pretreatment with50 mM CHX only.
Figure 4. Specificity of the hormonal response. TotalRNA (25 mg) from six days old etiolated seedlings treatedfor one hour with various chemicals and conditions, if nototherwise indicated, were hybridized with a 32P-labeledIAA1-specific probe. The lanes are: 1, untreated; 2 and 13,control-treated; 3 and 14, 20 mM IAA; 4, 50 mM CHX; 5,20 mM IAA and 50 mM CHX after 30 minutes pretreatmentwith 50 mM CHX only; 6, 20 mM 2,4-D; 7, 20 mM a-NAA; 8,20 mM PAA; 9, 20 mM L-tryptophan; 10, wounding; 11,0.5 M sorbitol; 12, heat treatment at 42°C for 15 minutesfollowed by 45 minutes recovery at room temperature; 15,20 mM ABA; 16, 20 mM GA; 17, 20 mM BA; 18, 20 mM IAAand 20 mM BA; 19, 20 mM IAA/20 mM BA and 50 mM LiCl;20, 50 mM LiCl; 21, 10 ppm ethylene; 22, N2; 23, air control;24, control (six hours in 10 mM phosphate); 25, phosphatestarvation (six hours, no phosphate present).
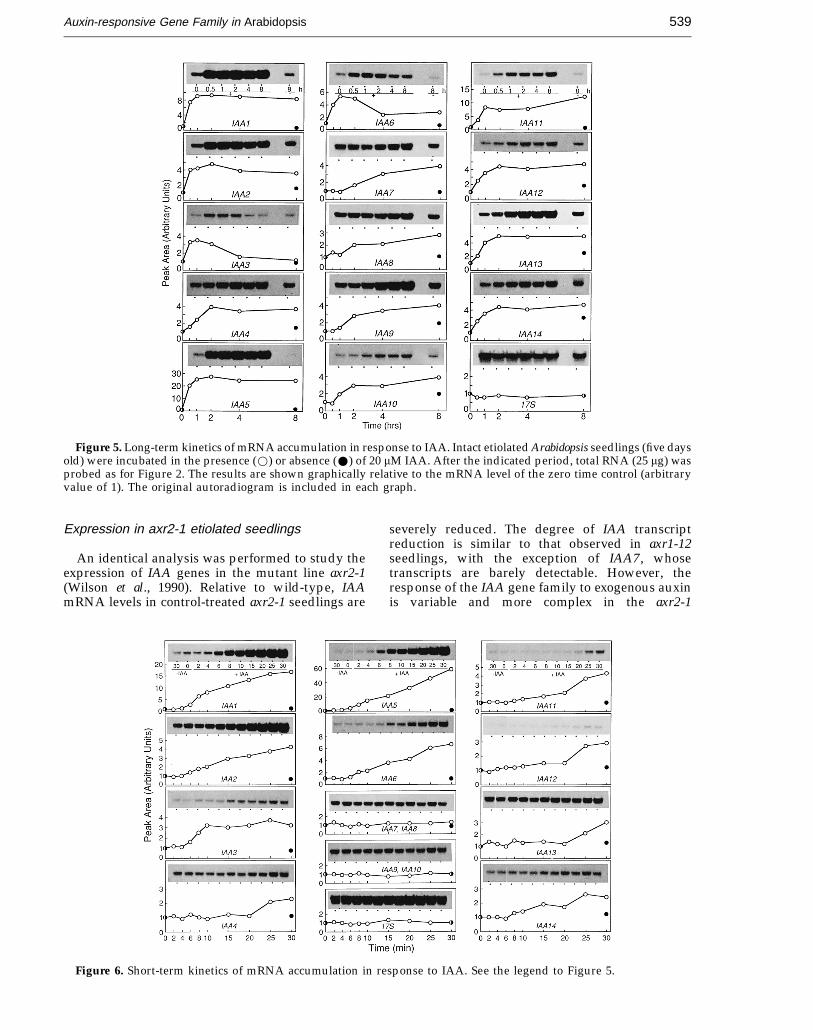
completely inhibits steady-state mRNA accumu-lation. However, at auxin concentrations below1 × 10−5 M IAA, individual members of the IAA genefamily respond in a differential fashion. For example,steady-state mRNA levels of IAA1, IAA5, IAA6 orIAA13 start to increase at an IAA concentration aslow as 1 × 10−8 M and continue to accumulate over aconcentration range of three orders of magnitude.The other IAA genes are gradually less sensitive intheir response, and some of them begin to respondat IAA concentrations only as high as 5 × 10−6 M (seeIAA3, IAA7 and IAA14).
Expression in auxin-resistant mutants ofArabidopsis
Expression in axr1-12 etiolated seedlings
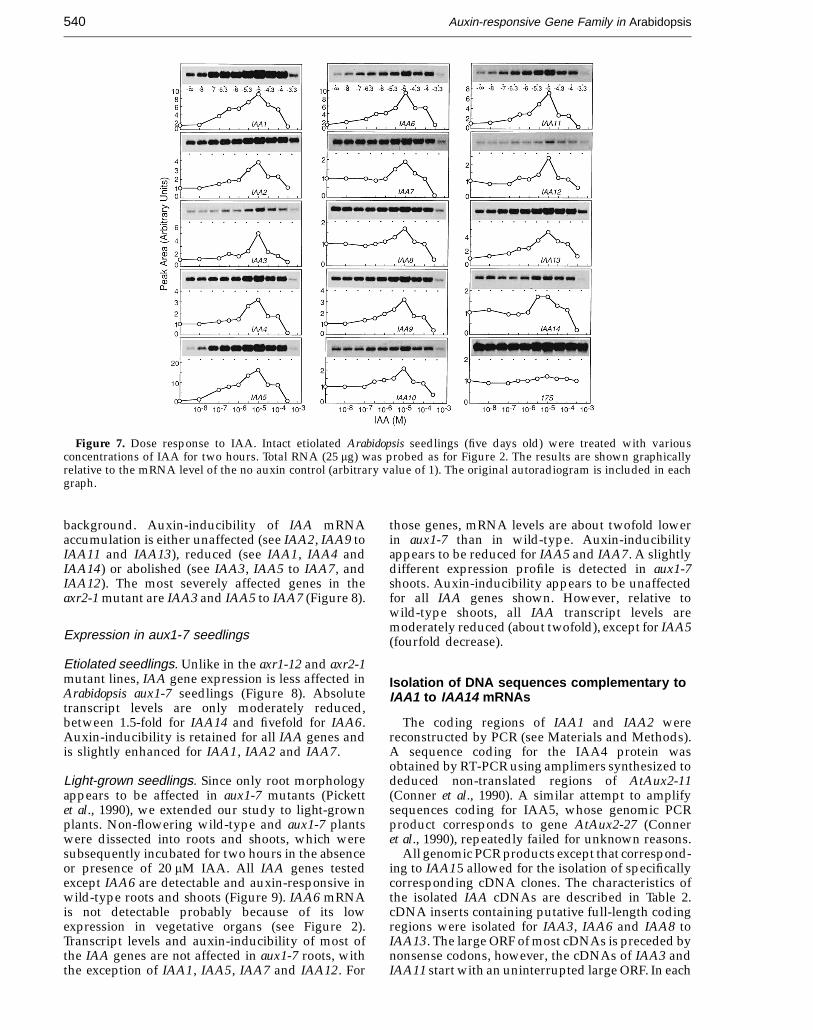
Expression of IAA genes was studied in intactetiolated seedlings of the Arabidopsis mutant linearx1-12 (Lincoln et al., 1990). Wild-type seedlings andmutant seedlings were treated in the presence orabsence of 20 mM IAA for two hours. Relative tomock-treated, wild-type tissue, steady-state IAAmRNA levels are severely reduced in respectivelytreated axr1-12 seedlings, between threefold forIAA12 or IAA14 and 15-fold for IAA7 (Figure 8).Transcripts of IAA5 and IAA6 are not detectable bythis analysis in control-treated mutant seedlings.Interestingly, when compared with the hormonalresponse in wild-type, auxin-inducibility of IAAmRNA accumulation in axr1-12 appears not to beaffected. Defective gene expression in axr1-12 is mostsevere for IAA5 and IAA6 when based on acomparison between auxin-treated seedlings(Figure 8).
IAA mRNAs accumulate steadily during theremaining time of the experiment. However, thetransiently expressed IAA3 mRNAs achieve a nearlymaximal level within the first ten minutes of auxintreatment, which remains essentially unchanged forup to 30 minutes.
Differential sensitivity of gene expressionto IAA
Next, we studied the effect of different IAAconcentrations on the expression of genes IAA1 toIAA14. Since all expressed IAA genes respond toexogenous auxin within two hours of treatment (seeFigure 5), a dose response curve ranging from1 × 10−8 M to 5 × 10−4 M of IAA was obtained after atwo hour auxin treatment of intact seedlings(Figure 7). The dose response of all IAA genesstudied shows a characteristic bimodal shape. Allgenes respond maximally to exogenous IAA at aconcentration of 1 × 10−5 M. Higher concentrations ofIAA are suboptimal, and IAA at 5 × 10−4 M
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 539
Figure 5. Long-term kinetics of mRNA accumulation in response to IAA. Intact etiolated Arabidopsis seedlings (five daysold) were incubated in the presence (w) or absence (W) of 20 mM IAA. After the indicated period, total RNA (25 mg) wasprobed as for Figure 2. The results are shown graphically relative to the mRNA level of the zero time control (arbitraryvalue of 1). The original autoradiogram is included in each graph.
Expression in axr2-1 etiolated seedlings
An identical analysis was performed to study theexpression of IAA genes in the mutant line axr2-1(Wilson et al., 1990). Relative to wild-type, IAAmRNA levels in control-treated axr2-1 seedlings are
severely reduced. The degree of IAA transcriptreduction is similar to that observed in axr1-12seedlings, with the exception of IAA7, whosetranscripts are barely detectable. However, theresponse of the IAA gene family to exogenous auxinis variable and more complex in the axr2-1
Figure 6. Short-term kinetics of mRNA accumulation in response to IAA. See the legend to Figure 5.
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis540
Figure 7. Dose response to IAA. Intact etiolated Arabidopsis seedlings (five days old) were treated with variousconcentrations of IAA for two hours. Total RNA (25 mg) was probed as for Figure 2. The results are shown graphicallyrelative to the mRNA level of the no auxin control (arbitrary value of 1). The original autoradiogram is included in eachgraph.
background. Auxin-inducibility of IAA mRNAaccumulation is either unaffected (see IAA2, IAA9 toIAA11 and IAA13), reduced (see IAA1, IAA4 andIAA14) or abolished (see IAA3, IAA5 to IAA7, andIAA12). The most severely affected genes in theaxr2-1 mutant are IAA3 and IAA5 to IAA7 (Figure 8).
Expression in aux1-7 seedlings
Etiolated seedlings. Unlike in the axr1-12 and axr2-1mutant lines, IAA gene expression is less affected inArabidopsis aux1-7 seedlings (Figure 8). Absolutetranscript levels are only moderately reduced,between 1.5-fold for IAA14 and fivefold for IAA6.Auxin-inducibility is retained for all IAA genes andis slightly enhanced for IAA1, IAA2 and IAA7.
Light-grown seedlings. Since only root morphologyappears to be affected in aux1-7 mutants (Pickettet al., 1990), we extended our study to light-grownplants. Non-flowering wild-type and aux1-7 plantswere dissected into roots and shoots, which weresubsequently incubated for two hours in the absenceor presence of 20 mM IAA. All IAA genes testedexcept IAA6 are detectable and auxin-responsive inwild-type roots and shoots (Figure 9). IAA6 mRNAis not detectable probably because of its lowexpression in vegetative organs (see Figure 2).Transcript levels and auxin-inducibility of most ofthe IAA genes are not affected in aux1-7 roots, withthe exception of IAA1, IAA5, IAA7 and IAA12. For
those genes, mRNA levels are about twofold lowerin aux1-7 than in wild-type. Auxin-inducibilityappears to be reduced for IAA5 and IAA7. A slightlydifferent expression profile is detected in aux1-7shoots. Auxin-inducibility appears to be unaffectedfor all IAA genes shown. However, relative towild-type shoots, all IAA transcript levels aremoderately reduced (about twofold), except for IAA5(fourfold decrease).
Isolation of DNA sequences complementary toIAA1 to IAA14 mRNAs
The coding regions of IAA1 and IAA2 werereconstructed by PCR (see Materials and Methods).A sequence coding for the IAA4 protein wasobtained by RT-PCR using amplimers synthesized todeduced non-translated regions of AtAux2-11(Conner et al., 1990). A similar attempt to amplifysequences coding for IAA5, whose genomic PCRproduct corresponds to gene AtAux2-27 (Conneret al., 1990), repeatedly failed for unknown reasons.
All genomic PCR products except that correspond-ing to IAA15 allowed for the isolation of specificallycorresponding cDNA clones. The characteristics ofthe isolated IAA cDNAs are described in Table 2.cDNA inserts containing putative full-length codingregions were isolated for IAA3, IAA6 and IAA8 toIAA13. The large ORF of most cDNAs is preceded bynonsense codons, however, the cDNAs of IAA3 andIAA11 start with an uninterrupted large ORF. In each
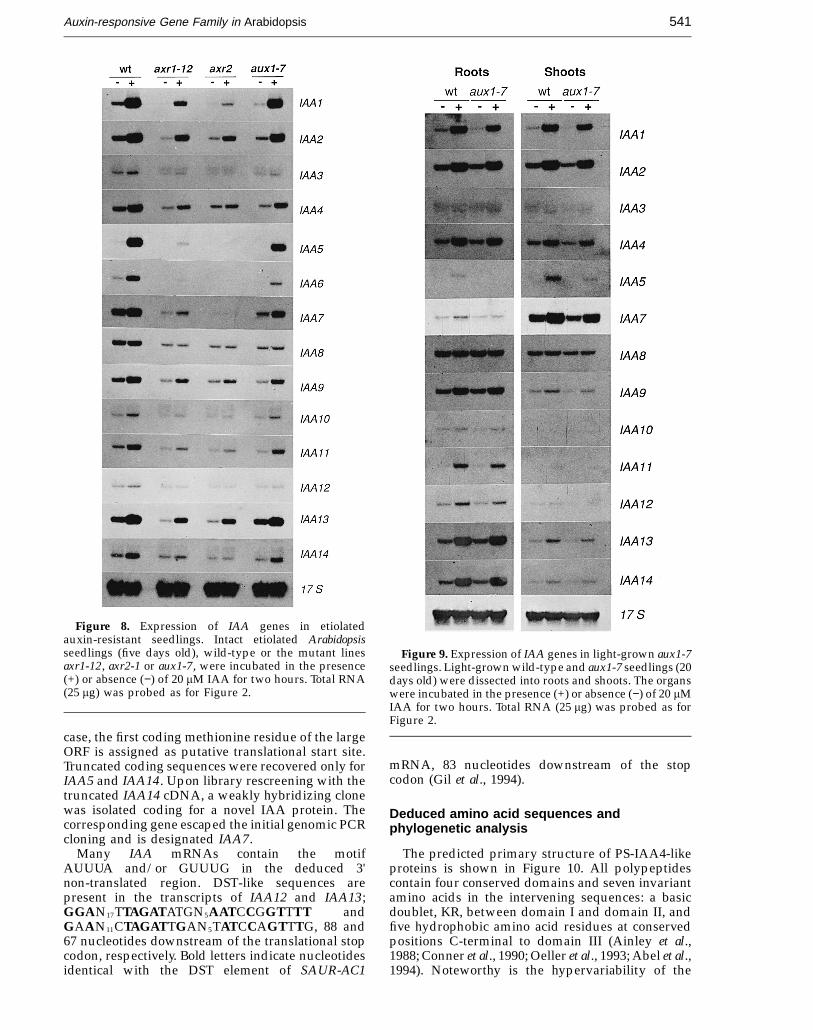
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 541
Figure 8. Expression of IAA genes in etiolatedauxin-resistant seedlings. Intact etiolated Arabidopsisseedlings (five days old), wild-type or the mutant linesaxr1-12, axr2-1 or aux1-7, were incubated in the presence(+) or absence (−) of 20 mM IAA for two hours. Total RNA(25 mg) was probed as for Figure 2.
Figure 9. Expression of IAA genes in light-grown aux1-7seedlings. Light-grown wild-type and aux1-7 seedlings (20days old) were dissected into roots and shoots. The organswere incubated in the presence (+) or absence (−) of 20 mMIAA for two hours. Total RNA (25 mg) was probed as forFigure 2.
case, the first coding methionine residue of the largeORF is assigned as putative translational start site.Truncated coding sequences were recovered only forIAA5 and IAA14. Upon library rescreening with thetruncated IAA14 cDNA, a weakly hybridizing clonewas isolated coding for a novel IAA protein. Thecorresponding gene escaped the initial genomic PCRcloning and is designated IAA7.
Many IAA mRNAs contain the motifAUUUA and/or GUUUG in the deduced 3'non-translated region. DST-like sequences arepresent in the transcripts of IAA12 and IAA13;GGAN17TTAGATATGN5AATCCGGTTTT andGAAN11CTAGATTGAN5TATCCAGTTTG, 88 and67 nucleotides downstream of the translational stopcodon, respectively. Bold letters indicate nucleotidesidentical with the DST element of SAUR-AC1
mRNA, 83 nucleotides downstream of the stopcodon (Gil et al., 1994).
Deduced amino acid sequences andphylogenetic analysis
The predicted primary structure of PS-IAA4-likeproteins is shown in Figure 10. All polypeptidescontain four conserved domains and seven invariantamino acids in the intervening sequences: a basicdoublet, KR, between domain I and domain II, andfive hydrophobic amino acid residues at conservedpositions C-terminal to domain III (Ainley et al.,1988; Conner et al., 1990; Oeller et al., 1993; Abel et al.,1994). Noteworthy is the hypervariability of the

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis542
Figu
re10
.Seq
uenc
eal
ignm
enta
nddo
mai
nst
ruct
ure
ofpr
imar
yau
xin-
resp
onsi
vege
nepr
oduc
ts.T
hem
ulti
ple
sequ
ence
alig
nmen
tobt
aine
dby
DA
RW
IN(G
onne
teta
l.,19
92)
was
man
ually
impr
oved
.Ide
ntic
alan
dco
nser
ved
amin
oac
idre
sidu
es(a
tle
ast
11of
21m
atch
es)
are
shad
ed(o
rang
e)an
dap
pea
rin
the
cons
ensu
s(c
apit
alan
dsm
all
lett
ers,
resp
ecti
vely
).C
onse
rved
dom
ains
are
unde
rlin
edan
din
dic
ated
byR
oman
num
eral
s.Si
mila
ram
ino
acid
resi
dues
cons
erve
dw
ithi
nd
isti
nctl
inea
ges
are
colo
ured
(sm
all,
G,A
;ac
idic
,D,E
;bas
ic,H
,R,K
;gen
eral
lyhy
dro
phob
ic,A
,C,V
,I,L
,M,F
,Y,W
;pol
ar,S
,T;a
ndN
,Q).
Am
ino
acid
resi
dues
corr
esp
ond
ing
tode
gene
rate
ampl
imer
sD
C2
and
DC
4ar
ein
dic
ated
inth
eIA
A15
sequ
ence
(>>
>an
d<
<<
,res
pec
tive
ly).
An
arro
wde
sign
ates
the
cons
erve
dp
osit
ion
ofan
intr
onin
the
PCR
-gen
erat
edge
nefr
agm
ents
whe
npr
esen
t.C
onse
rved
basi
cre
sidu
esth
atfu
ncti
onas
anN
LS
inPS
-IA
A4
and
PS-I
AA
6(A
bel&
The
olog
is,1
995)
are
ind
icat
edon
top
ofth
eal
ignm
ent.
Star
sin
dic
ate
apa
tter
nof
cons
erve
dle
ucin
ere
sidu
esin
dom
ain
Ian
dof
cons
erve
dpr
olin
ere
sidu
esin
dom
ain
II.S
econ
dar
yst
ruct
ure
pred
icti
ons
(PH
D)
for
IAA
9w
ere
calc
ulat
edac
cord
ing
toR
ost
&Sa
nder
(199
3)an
dar
ein
dic
ated
byh
(hel
ix),
e(e
xten
ded
shee
t),l
(loo
p);a
per
iod
ind
icat
esno
pred
icti
on.A
min
oac
idre
sidu
esth
atm
ayfo
rmhy
dro
phob
icsu
rfac
esin
the
pred
icte
dco
nser
ved
amph
ipat
hic
baa
mot
ifar
ein
dic
ated
by(W
).So
urce
sof
the
sequ
ence
sar
eas
follo
ws:
IAA
1-IA
A15
(Ara
bido
psis
thal
iana
,thi
sst
udy)
;PS-
IAA
4,PS
-IA
A6
(pea
(Pis
umsa
tivu
m),
Oel
ler
etal
.,19
93);
AR
G3
and
AR
G4
(mun
gbe
an(V
igni
ara
diat
a),Y
amam
oto
etal
.,19
92);
Gm
Aux
22an
dG
mA
ux28
(soy
bean
(Gly
cine
max
),A
inle
yet
al.,
1988
).
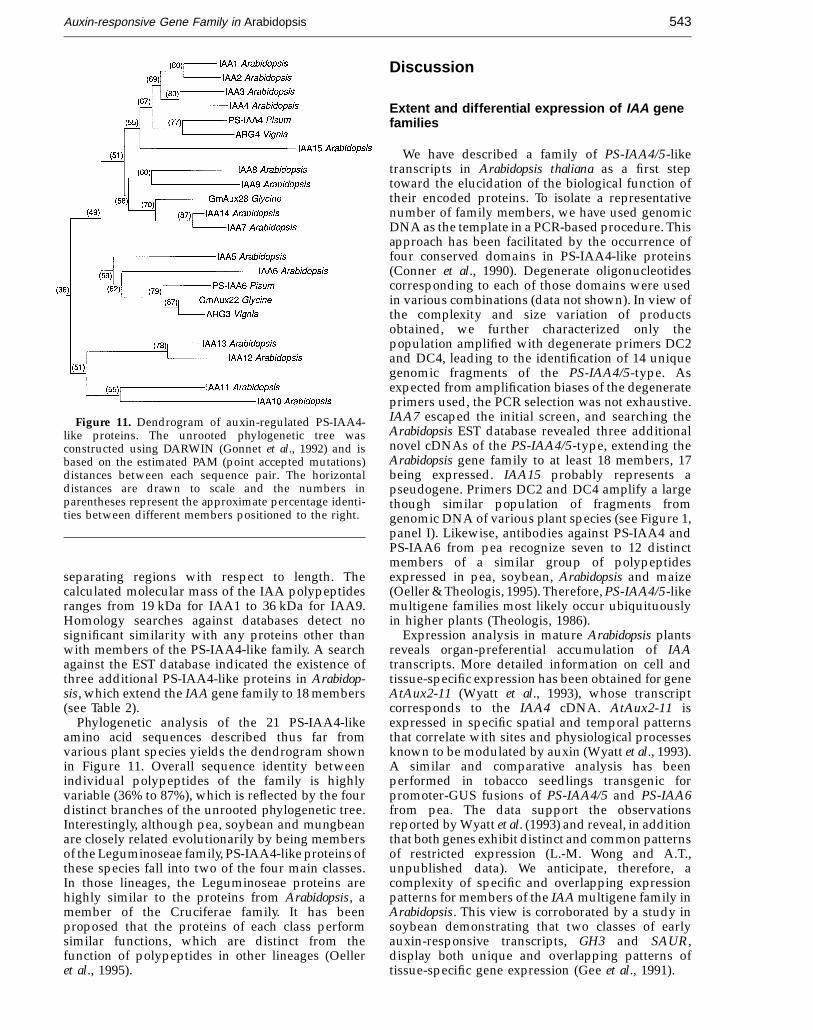
JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 543
Figure 11. Dendrogram of auxin-regulated PS-IAA4-like proteins. The unrooted phylogenetic tree wasconstructed using DARWIN (Gonnet et al., 1992) and isbased on the estimated PAM (point accepted mutations)distances between each sequence pair. The horizontaldistances are drawn to scale and the numbers inparentheses represent the approximate percentage identi-ties between different members positioned to the right.
Discussion
Extent and differential expression of IAA genefamilies
We have described a family of PS-IAA4/5-liketranscripts in Arabidopsis thaliana as a first steptoward the elucidation of the biological function oftheir encoded proteins. To isolate a representativenumber of family members, we have used genomicDNA as the template in a PCR-based procedure. Thisapproach has been facilitated by the occurrence offour conserved domains in PS-IAA4-like proteins(Conner et al., 1990). Degenerate oligonucleotidescorresponding to each of those domains were usedin various combinations (data not shown). In view ofthe complexity and size variation of productsobtained, we further characterized only thepopulation amplified with degenerate primers DC2and DC4, leading to the identification of 14 uniquegenomic fragments of the PS-IAA4/5-type. Asexpected from amplification biases of the degenerateprimers used, the PCR selection was not exhaustive.IAA7 escaped the initial screen, and searching theArabidopsis EST database revealed three additionalnovel cDNAs of the PS-IAA4/5-type, extending theArabidopsis gene family to at least 18 members, 17being expressed. IAA15 probably represents apseudogene. Primers DC2 and DC4 amplify a largethough similar population of fragments fromgenomic DNA of various plant species (see Figure 1,panel I). Likewise, antibodies against PS-IAA4 andPS-IAA6 from pea recognize seven to 12 distinctmembers of a similar group of polypeptidesexpressed in pea, soybean, Arabidopsis and maize(Oeller & Theologis, 1995). Therefore, PS-IAA4/5-likemultigene families most likely occur ubiquituouslyin higher plants (Theologis, 1986).
Expression analysis in mature Arabidopsis plantsreveals organ-preferential accumulation of IAAtranscripts. More detailed information on cell andtissue-specific expression has been obtained for geneAtAux2-11 (Wyatt et al., 1993), whose transcriptcorresponds to the IAA4 cDNA. AtAux2-11 isexpressed in specific spatial and temporal patternsthat correlate with sites and physiological processesknown to be modulated by auxin (Wyatt et al., 1993).A similar and comparative analysis has beenperformed in tobacco seedlings transgenic forpromoter-GUS fusions of PS-IAA4/5 and PS-IAA6from pea. The data support the observationsreported by Wyatt et al. (1993) and reveal, in additionthat both genes exhibit distinct and common patternsof restricted expression (L.-M. Wong and A.T.,unpublished data). We anticipate, therefore, acomplexity of specific and overlapping expressionpatterns for members of the IAA multigene family inArabidopsis. This view is corroborated by a study insoybean demonstrating that two classes of earlyauxin-responsive transcripts, GH3 and SAUR,display both unique and overlapping patterns oftissue-specific gene expression (Gee et al., 1991).
separating regions with respect to length. Thecalculated molecular mass of the IAA polypeptidesranges from 19 kDa for IAA1 to 36 kDa for IAA9.Homology searches against databases detect nosignificant similarity with any proteins other thanwith members of the PS-IAA4-like family. A searchagainst the EST database indicated the existence ofthree additional PS-IAA4-like proteins in Arabidop-sis, which extend the IAA gene family to 18 members(see Table 2).
Phylogenetic analysis of the 21 PS-IAA4-likeamino acid sequences described thus far fromvarious plant species yields the dendrogram shownin Figure 11. Overall sequence identity betweenindividual polypeptides of the family is highlyvariable (36% to 87%), which is reflected by the fourdistinct branches of the unrooted phylogenetic tree.Interestingly, although pea, soybean and mungbeanare closely related evolutionarily by being membersof the Leguminoseae family, PS-IAA4-like proteins ofthese species fall into two of the four main classes.In those lineages, the Leguminoseae proteins arehighly similar to the proteins from Arabidopsis, amember of the Cruciferae family. It has beenproposed that the proteins of each class performsimilar functions, which are distinct from thefunction of polypeptides in other lineages (Oelleret al., 1995).

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis544
Characteristics of the hormonal response
Basal mRNA levels of most IAA genes are readilydetectable in untreated Arabidopsis seedlings, indicat-ing that endogenous auxin concentrations aresufficient in at least some tissues to induce andmaintain their expression. All genes expressed, IAA1to IAA14, respond specifically to natural andsynthetic auxins as the only external inducersidentified, besides CHX. This observation is inagreement with previous results for related genes(Theologis et al., 1985; Hagen & Guilfoyle, 1985;Yamamoto et al., 1992). The SAUR genes and GH3 arethe only other classes of early genes described to bespecifically induced by auxin (Guilfoyle et al., 1993).
Members of the Arabidopsis gene family react toexogenous IAA in a highly differential fashion. Firstdetectable increases in steady-state IAA mRNAsoccur between four and 120 minutes after auxinapplication. Some of the early genes, such as IAA1 toIAA3, have mRNA accumulation kinetics verysimilar to the SAUR genes, the fastest respondinggenes known to be induced by auxin. The SAURtranscripts start to accumulate within two to fiveminutes and achieve maximal levels by 60 minutesof 2,4-D treatment (McClure & Guilfoyle, 1987). Thedose response curves of IAA genes reveal differentialsensitivities over a broad range of IAA concen-trations, covering about three orders of magnitudefrom 10 nM to 5 mM IAA. The differences in kineticsand sensitivity of transcript accumulation betweenindividual IAA genes is likely due to a variety offactors. These may include unknown parameterssuch as tissue-specific auxin permeability andreception, cell-type dependent and differentialregulation of free auxin concentrations, or differentmodes of auxin-dependent transcriptional activationand post-transcriptional regulation. However, allIAA genes respond maximally at about 10 mM IAA.A similar saturable response has been described forPS-IAA4/5 gene expression in pea epicotyl proto-plasts (Ballas et al., 1993), for the PS-IAA4/5-typegene GH1 in soybean seedlings (Hagen & Guilfoyle,1985), or for SAUR promoter-driven GUS expressionin transgenic tobacco (Li et al., 1991). Interestingly,the biphasic dose response to IAA of PS-IAA4/5-likemRNA accumulation is paralleled by similarbimodal characteristics of other auxin-elicitedresponses in plants, such as stem elongation,stomatal opening, or auxin-modulation of theinward-rectifying K+ channel, Ik,in, in stomatal guardcells (Blatt & Thiel, 1994). Intracellular pH andcalcium concentrations have been proposed asopposing second messengers in auxin signaling,which reciprocal interaction evokes the bimodalcontrol of the potassium transporter (Blatt & Thiel,1994). It is tempting to extend this hypothesis toauxin-regulated gene expression.
Deficient expression in auxin-resistant mutants
Expression of IAA genes is severely inhibited inetiolated seedlings of the Arabidopsis auxin-resistant
mutants axr1-12 and axr2-1. These mutants have apleiotropic though auxin-related phenotype (Hobbie& Estelle, 1994). Most aspects of the axr2-1phenotype are due to a dramatic decrease in cellelongation rather than a reduction in cell number,whereas the opposite effect has been described foraxr1-12 mutant plants (Lincoln et al., 1990; Timpteet al., 1992). The alterations of axr1-12 plants havebeen explained by a reduced sensitivity to auxin(Hobbie & Estelle, 1994). This interpretion issupported by dramatically reduced IAA mRNAlevels in etiolated axr1-12 seedlings, thoughauxin-inducibility appears to be maintained(Figure 8). The AXR1 gene reveals significantsimilarity to the ubiquitin-activating enzyme E1,although AXR1 is probably not a functional E1homolog (Leyser et al., 1993). Nonetheless, thepossibility exists that ubiquitin-dependent proteol-ysis may play an important role in auxin responses,for instance, by controlling the level but not auxinsensibility of the short-lived repressor postulated toregulate PS-IAA4/5-like gene expression (Ballas et al.,1993). Expression of IAA genes in axr2-1 seedlings isas severely inhibited as in axr1-12 plants; however,auxin-inducibility is also impaired for most of thegenes. A similar reduced expression in axr2-1seedlings has been described for the SAUR-AC1gene (Gil et al., 1994; Timpte et al., 1994). The axr2-1mutation has recently been characterized as again-of-function mutation and proposed to preventauxin-mediated inactivation of a transcriptionalrepressor (Timpte et al., 1994). Mutant aux1-7 plantsdisplay a severe defect in root gravitropism but areotherwise wild-type in their aerial structures (Pickettet al., 1990). Only a modest reduction of IAA geneexpression is noticed, however, both in roots and inshoots of light-grown seedlings (Figure 9). This is inagreement with the suggested function of AUX1also in the aerial portions of the plant (Hobbie& Estelle, 1994). We have also studied the expressionof an auxin-inducible ACC synthase gene and ofthe SAUR-AC1 gene. Interestingly, in each ofthe three mutant lines tested both genes are inhibitedto a similar extent as the IAA genes (unpublishedresults). Inhibition of three classes of genes withdifferent auxin-responsive elements supportsthe previous notion that either mutation acts earlyin an auxin response pathway (Hobbie & Estelle,1994).
Regulation of IAA gene expression
The expression of PS-IAA4-like proteins appearsto be stringently controlled at the transcriptional andpost-transcriptional level, which is a hallmark ofregulatory proteins (Gottesman & Maurizi, 1992).PS-IAA4/5-like genes are rapidly transcriptionallyactivated by auxin (Theologis et al., 1985; Hagen &Guilfoyle. 1985; Ainley et al., 1988; T. Koshiba andA.T., unpublished results). The auxin-responsivecis-element of the PS-IAA4/5 promoter has beenfunctionally defined, and in vivo titration exper-

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 545
iments indicate binding to positively acting factors(Ballas et al., 1993, 1995).
The transcripts of PS-IAA4/5 and PS-IAA6 arelabile. Their mRNA half-lives, 60 minutes and 75minutes, respectively (T. Koshiba and A.T., unpub-lished results), are comparable with those of manyunstable mRNAs of mammalian early-responsegenes (Atwater et al., 1990), or with the soybeanSAUR transcripts, 40 to 50 minutes, (Franco et al.,1990). The DST sequence element in the 3'-untrans-lated region of the SAUR genes has been shown tosignal rapid mRNA decay (Newman et al., 1993).Likewise, as in mammalian cells (Chen & Shyu,1994), AU-rich elements can confer mRNA instabil-ity in plants (Ohme-Takagi et al., 1993). Consistentwith their expected instability, most of the IAAtranscripts contain AU-rich elements and, IAA12 andIAA13, a DST-like sequence element in the 3'untranslated region (see Table 2). A short half-life ofIAA mRNAs is indicated by the transient accumu-lation of IAA3 and IAA6 transcripts in response toauxin (Figure 5).
The PS-IAA4 and PS-IAA6 proteins are ofextremely low abundance in vivo (Oeller & Theologis,1995). The metabolic half-lifes of both proteins,between six and eight minutes (Abel et al., 1994),are among the shortest known for eukaryoticproteins (Gottesman & Maurizi, 1992). Althoughthe mechanism of PS-IAA4 degradation is as yetunknown, we note that determinants for rapidprotein turnover such as PEST sequences or otherpotential destruction signals (Gottesman & Maurizi,1992) are not discernible in IAA polypeptides.
Effect of CHX
Protein synthesis inhibitors have been used as atool to unmask regulatory mechanisms of early geneactivation. Induction of early genes is oftenindependent of de novo synthesized proteins,indicating a primary response to the stimulusvia modification of pre-existing components (Her-schman, 1991). In addition, inhibition of proteinsynthesis potentiates the strength and duration ofmany signal-induced transcriptional responses byelimination of attenuating factors that are oftensynthesized as part of the response (Hill & Treisman,1995). Induction by auxin of most IAA genes isinsensitive to CHX, qualifying them as primaryauxin-responsive genes (Figure 3). CHX alone or incombination with auxin, superinduces PS-IAA4/5-like genes (Figure 3; Theologis et al., 1985). Thisphenomenon is well described for many early genesand oncogenes in mammalia (Herschman, 1991). Theeffect of CHX on the expression of PS-IAA4/5 hasbeen investigated, implicating two targets ofinhibitor action: mRNA stabilization and transcrip-tional activation presumably by preventing synthesisor activation of a labile repressor polypeptide(T. Koshiba and A.T., unpublished results). Theinduction of negatively acting factors as part of theauxin response is best revealed by the accumulation
kinetics of IAA3 or IAA6 mRNAs. Both genes aretransiently stimulated by IAA, and transcript levelsare adjusted to low basal levels after four hours in thepresence of the hormone (Figure 5). However, in thepresence of IAA and CHX, induced mRNA levels aremaintained for at least six hours (Figure 3). Thisindicates de novo synthesis of a protein that directlyor indirectly attenuates the transcriptional response.Such directly acting factors have been described andinclude the fos protein that trans-represses its owngene (Sassone-Corsi et al., 1988). The effect of CHXon expression of the relatively late auxin genes IAA7and IAA8 suggests dependence on de novo proteinsynthesis of auxin-induction. Auxin-inducibility ofboth genes is inhibited in the presence of CHX, andCHX alone has no effect, indicating that IAA7 andIAA8 are secondary response genes (Figure 3). Thesedata suggest a hierarchical response to auxin of IAAgenes.
Structural motifs of IAA proteins
All deduced PS-IAA4-like protein sequencesdescribed thus far contain the four conserveddomains originally noticed by Ainley et al. (1988).More importantly, seven amino acids are absolutelyconserved in hypervariable regions. These regionsare highly polymorphic and responsible for the widerange of molecular mass calculated, 19 to 36 kDa. Wehave previously shown that PS-IAA4-like polypep-tides are able to direct the GUS reporter protein intothe cell nucleus (Abel et al., 1994; Abel & Theologis,1994). Functional NLSs have been identified inPS-IAA4 and PS-IAA6 (Abel & Theologis, 1995). Theessential NLS is of bipartite structure, comprising theinvariant basic doublet KR and basic amino acids indomain II. A second, SV40-type NLS is located indomain IV (Figure 10). Domain I is the core of aleucine-rich region. Most IAA proteins share theconsensus motif LxLxxxxLxLxL or in IAA10 toIAA13, LxLxLxLxL (overlapping residues are inbold-face type). This motif is reminiscent of theleucine-rich repeat unit that is proposed to mediateprotein-protein interactions (Kobe & Deisenhofer,1994). Domain II is relatively rich in proline residues,and the spacer of IAA8 and IAA9 contains shortsequences, (PXX)n , indicative of a polyproline IIconformation. Proline-rich motifs have been impli-cated in protein-protein interactions (Williamson,1994).
The calculated secondary structures are similar forall members of the IAA family, suggesting that theseproteins have related tertiary folds. The predictedsecondary structure for IAA9 is shown in Figure 10.Most of the spacer sequences are likely to adopt arandom conformation in all IAA proteins analyzed.Domain III and the adjacent five invarianthydrophobic amino acid residues at conservedpositions are predicted to form an amphipathicbaa-fold similar to the b-ribbon DNA recognitiondomain of prokaryotic repressor polypeptides, suchas Arc and MetJ (Phillips, 1994). If experimentally

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis546
proven, IAA polypeptides may represent the firsteukaryotic members of this class of DNA-bindingproteins (Raumann et al., 1994). Arc and MetJ areproteins with similar functions and tertiary struc-tures but with no apparent sequence identities,which is probably a reason why additional membersof this family have not been identified yet.Interestingly, a phylogenetic analysis shows that thebaa-motif of PS-IAA4-like proteins is as distantlyrelated to the prokaryotic motifs (Arc, Mnt, TraY andMetJ) as those are to each other (data not shown). Thetertiary structures of Arc and MetJ have beenresolved (Phillips, 1994). A three-dimensional (3D)compatibility analysis of the IAA baa-fold with theb-ribbon DNA recognition motif of Arc or MetJ as thetertiary template was performed using the 3D profilemethod of Bowie et al. (1991). In both profiles, the3D-1D scores for Arc versus MetJ differ, since theirbaa-domains vary in length. However, the score forthe IAA baa-motif is comparable with the scoresobtained for the other prokaryotic baa-folds in eachprofile (data not shown). This indicates that theputative eukaryotic baa-motif is compatible with theprokaryotic b-ribbon DNA recognition motifs. Wehave proposed an analogous function of the putativebaa-motif in PS-IAA4-like proteins (Abel et al., 1994).
Functional implications
This study demonstrates that the plant growthhormone auxin induces in a highly differential andhierarchical fashion a large family of early genes thatencode a structurally diverse class of short-livednuclear proteins. These polypetides contain aconserved putative DNA-binding motif, and theirexpression is stringently regulated at multiple levels.Therefore, it has been proposed that PS-IAA4-likeproteins play a pivotal role in the regulation ofsubsequent auxin-induced events responsible forplant cell growth (Abel et al., 1994). Thus, the modeof auxin action in plants appears to follow theparadigm of how, for instance, ecdysone (Andres &Thummel, 1992) or other growth factors andextracellular signals (Herschman, 1991; Hill &Treisman, 1995) regulate behavior and developmentof animal cells. Factors that promote long-termcellular responses such as proliferation and differen-tiation coordinate hierarchical changes in the patternof gene expression. Initially, a set of various earlyresponse genes is rapidly and transiently expressed.Many of those early genes encode transcriptionfactors that activate secondary genes in a tissue-specific manner to shape the morphological andfunctional properties of each target tissue (Her-schman, 1991). For instance, members of a large anddiverse family of POU-domain regulatory genes areexpressed in a distinct temporal and spatial patternduring neuronal development (He et al., 1989).However, co-expression of different POU-domainproteins in certain brain regions is evident and thecombinatorial code is suggested to be critical forspecifying mature cellular phenotypes in the
mammalian nervous system (He et al., 1989).Likewise, the tissue coordination model predicts thatecdysone activates tissue-specific sets of early genesencoding ecdysone receptors and regulatory pro-teins. The combination of those leads to activation ofdistinct sets of late genes that define the uniqueproperties of each ecdysone target tissue at eachstage in its development (Karim & Thummel, 1991).The studies done by Gee et al. (1991), Wyatt et al.(1993) and our laboratory (unpublished results)demonstrate that expression of early auxin-respon-sive genes is restricted to a few tissue types at specificdevelopmental stages. Furthermore, the existence ofmultiple auxin receptors has been inferred (Gee et al.,1991). In view of the highly differential mode of IAAgene expression in Arabidopsis, with respect totiming, hormone concentration and spatially re-stricted expression patterns, the prospect arises thatcombinatorial interactions among co-expressed IAAproteins define tissue-specific responses to auxinduring plant growth and development. Defining theexpression profiles of the different IAA transcriptswill provide clues regarding those potentialcombinatorial interactions. These studies should alsoprovide insight into the molecular mechanisms thatcontrol the timing of sequential gene expression inresponse to auxin.
Materials and Methods
Arabidopsis strains, growth conditions and otherplant material
Wild-type Arabidopsis thaliana (L.) Heynh. ecotypeColumbia and auxin-resistant mutant lines axr1-3, axr1-12(Lincoln et al. 1990), aux1-7 (Pickett et al. 1990) and axr2(Wilson et al. 1990) were obtained from the BiologicalResource Center at Ohio State University. Etiolatedseedlings were grown as described (Liang et al., 1992).Wild-type and aux1-7 seedlings were grown in Petri dishesat 22°C for three weeks under illumination withfluorescent and incandescent light (16 hour photoperiod).Mature plants were grown in the greenhouse at 22°C to25°C under continuous illumination. Etiolated seedlings ofpea (Pisum sativum cv. Alaska), corn (Zea maize cv. Funk),rice (Oryza sativa L. cv. IR36) and tomato (Lycopersiconesculentum cv. Rutgers) were grown as described (Oeller &Theologis, 1995). Cauliflower was purchased from a localmarket.
Tissue treatment
Intact etiolated seedlings (five to six days old) wereplaced in 0.5 × MS salt solution (pH 5.6) and supplementedwith the appropriate chemicals. The seedlings wereincubated in the dark at room temperature with moderateshaking (50 to 100 rpm). Mock control incubations weresupplemented with an equal amount of the solvent used toprepare the stock solution of the respective chemical. Afterthe indicated time, aliquots (3 to 5 g fresh weight) ofseedlings were removed, immediately frozen with liquidnitrogen and stored at − 80°C. Light-grown wild-type andaux1-7 seedlings were dissected into roots and shoots, andtreated as described above. Different organs from25-day-old light-grown plants were harvested and kept at− 80°C before RNA extraction.

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 547
Isolation of nucleic acids
Genomic DNA of Arabidopsis, cauliflower, pea, tomato,rice, and corn was isolated from etiolated seedlings(Dellaporta et al., 1983). The DNA preparations werefurther purified by equilibrium sedimentation in cesiumchloride gradients (Davis et al., 1980). Total nucleic acidswere prepared from frozen Arabidopsis tissues (Liang et al.,1992). Poly(A)+ RNAs were isolated by affinity chromatog-raphy using oligo(dT)-cellulose (Theologis et al., 1985).
PCR
Two degenerate oligonucleotide primers were syn-thesized, DC2 (512-fold degeneracy) and DC4 (64-folddegenaracy), which correspond to motifs I/VVGWPP indomain II and DKDGDWM in domain IV of PS-IAA4-likeproteins, respectively (Figure 10). The sequence andorientation of both primers is as follows: DC2 5'-GCC GAATTC RTN GTN GGN TGG CCN CC-3'; DC4 5'-GCC AGGCTT CAT CCA RTA NCC RTC YTT RTC-3'; N is A, C, Gor T; R is A or G; Y is C or T. The EcoRI site in DC2 andthe HindIII site in DC4 are underlined. The degeneratePCR (Compton, 1990) reaction mixture contained 100 ng ofArabidopsis genomic DNA, 1 mM of each primer, 0.5 unit ofTaq polymerase (Perkin-Elmer-Cetus) and was incubatedin a thermocycler (Perkin-Elmer-Cetus) for 30 cycles, eachcycle consisting of one minute at 94°C, one minute at 43°Cand one minute at 72°C. The PCR products werefractionated in 3% NuSieve (FMC BioProducts)/1%agarose and recovered by adsorption to Schleicher &Schuell NA-45 paper. Amplified genomic DNA fragmentswere directionally subcloned into pUC19(EcoRI/HindIII)and subsequently characterized by cross-hybridizationand DNA sequencing of both strands.
Poly(A)+ RNA from IAA/CHX-treated Arabidopsisseedlings was used to synthesize circularized cDNA(Huang et al., 1990). To reconstruct the coding region ofIAA1 and IAA2, inverse PCR amplimers were designedusing the sequence information of the DC2-DC4 PCRproducts that correspond to IAA1 and IAA2, respectively.Sequence and orientation of these primers is asfollows: IAA1, 12/1A 5'-TTCACAGTAGGTGAATATTCC-3' (residues 111 to 117) and 12/1B 5'-ACATAACTCAC-GTTTTTGTTG-3' (residues 77 to 70); IAA2, 8/15A5'-AATATGTTCAAAGTCATGATT-3' (residues 109 to 116)and 8/15B 5'-TTTCACGTAGCTCACACTGTT-3' (residues81 to 75). Using these primer sets and circular cDNA,inverse PCR products corresponding to the coding regionof IAA1 and IAA2 were generated. The respective DNAfragments were purified, subcloned into the EcoRV site ofpIC20R and sequenced.
The complete deduced coding region of IAA1 and IAA2was generated by reverse transcription of poly(A)+ RNAsfollowed by PCR (Jones, 1993). Based on sequences of therespective inverse PCR products, the following amplimerscomplementary to non-translated regions of IAA1 cDNAand IAA2 cDNA were synthesized (the EcoRI and HindIIIsites are underlined): IAA1, CD1-1 5'-GCCGAATTCGA-GAGAATATGGAAGTCACC-3' and CD1-2 5'-GCCAAG-CTTCAAAAGGTGTTTTGAGACAAT-3'; IAA2, CD2-15' - GCCGAATTCACAAAGATTGATATTGATCC - 3' andCD2-2 5'-GCCAAGCTTCAAAAGATTCTCAGCTTCTC-3'. The coding region of IAA4 cDNA was obtained by thesame method using amplimers complementary to deducednon-translated sequences of the corresponding geneAtAux2-11 (Conner et al., 1990): CD4-1 5'-GCCGAATTCA-CAAAGAGGTTCTTTCTTTG-3' and CD4-2 5'-GCCAA-GCTTTTATCTCTCAATTAGGTTTG-3'. The amplified cD-
NAs were directionally subcloned into pUC19(EcoRI/HindIII) and both strands were sequenced.
Hybridization analysis and cDNA library screening
PCR products obtained with the degenerate amplimersDC2 and DC4 were electrophoresed on 2% (w/v) agarosegels and transferred to Nytran nylon membrane (Scleicher& Schuell). The degenerate oligonucleotide DC3 (5'-AARGTN AGY RTN GAY GG-3'; N is A, C, G or T; R is A or G;Y is C or T), corresponding to the motif KVSM/V/I/DGin domain III of PS-IAA4-like auxin-regulated polypep-tides (see Figure 10), was used as hyridization probeaccording to Sambrook et al. (1989).
Appropiate enzyme digests of plasmid DNAs or totalArabidopsis genomic DNA were electrophoresed on 2% or0.8% agarose gels, respectively. Southern transfer andhybridization was performed according to Oeller et al.(1993).
Northern analysis was performed according to Ecker &Davis (1987). The autoradiograms were quantified usingan LKB ultrascan laser densitometer (Bromma, Sweden).After exposure, the probe was removed by rinsing thefilters in 0.1% (w/v) SDS at 95 to 100°C for 15 to30 minutes.
An Arabidopsis cDNA library (Elledge et al., 1991) wasscreened with individual PS-IAA4/5-like DNA fragments.IAA cDNA inserts were recovered as described (Elledgeet al., 1991). Both strands of the cDNAs were sequenced.
Other molecular techniques
Standard molecular techniques and colony hybridiz-ations were performed according to Sambrook et al. (1989).Plasmid DNA sequencing was carried out with the chaintermination method (Sanger et al., 1977) using [a-35S]dATPand the modified phage T7 DNA polymerase Sequenase(U.S. Biochemicals).
Protein structure computer analysis
Homology searches were performed in January 1995against protein data banks (SwissProt, GenPept, PIR,PROSITE) and nucleotide data banks (EMBL, GenBank,dBEST) using BLAST programmes (Altschul et al., 1990)through NCBI (The National Center for BiotechnologyInformation; [email protected]). Secondary struc-ture predictions were obtained using a neural networkalgorithm (Rost & Sander, 1993) at the EMBL Heidelberg([email protected]). The phylogenetictree of IAA protein sequences was constructed usingDARWIN (Gonnet et al., 1992) at the public [email protected].
AcknowledgementsWe thank Drs Mark Estelle for seeds of auxin-resistant
mutant lines, Tao Zhang for 3D modeling, David Dilley fornoticing the polyproline motif, Lu-Min Wong, Nurit Ballas,Paul Oeller, Xiawou Liang and Tom Zarembinski for theircontinuous support and critical discussions, and Ron Wellsfor editing the manuscript. This work was supported by agrant from the National Institute of Health (GM-35447) toA.T., and by postdoctoral fellowships from BoehringerIngelheim Fonds, Germany, and the U.S. Department ofAgriculture (5335-21430-002-00D) to S.A.

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis548
References
Abel, S. & Theologis, A. (1994). Transient transformationin Arabidopsis leaf protoplasts: a versatile experi-mental system to study gene expression. Plant J. 5,421–427.
Abel, S. & Theologis, A. (1995). A polymorphic bipartitemotif signals nuclear targeting of early auxin-inducible proteins related to PS-IAA4 from pea (Pisumsativum) Plant J. 8, 101–110.
Abel, S., Oeller P. W. & Theologis, A. (1994). Earlyauxin-induced genes encode short-lived nuclearproteins. Proc. Natl Acad. Sci. USA, 91, 326–330.
Ainley, W. M., Walker, J. C., Nagao, R. & Key, J. L. (1988).Sequence and characterization of two auxin-regulatedgenes from soybean. J. Biol. Chem. 263, 10658–10666.
Altschul, S. F., Gish, W., Miller, W. Myers, E. & Lipman, D. J.(1990). Basic local alignment search tool. J. Mol. Biol.215, 403–410.
Andres, A. J. & Thummel, C. S. (1992). Hormones, puffsand flies: the molecular control of metamorphosis byecdysone. Trends Genet. 8, 132–138.
Assmann, S. M. (1993). Signal transduction in guard cells.Annu. Rev. Cell Biol. 9, 345-375.
Atwater, J. A., Wisdom, R. & Verma, I. M. (1990). RegulatedmRNA stability. Annu. Rev. Genet. 24, 519–541.
Ballas, N., Wong, L.-M. & Theologis, A. (1993). Identifi-cation of the auxin-responsive element, AuxRE, in theprimary indoleacetic acid-inducible gene, PS-IAA4/5,of pea (Pisum sativum). J. Mol. Biol. 233, 580–596.
Ballas, N., Wong, L. M., Ke, M. & Theologis, A. (1995). Twoauxin responsive domains interact positively toinduce expression of the early indoleacetic acid-inducible gene, PS-IAA4/5. Proc. Natl Acad. Sci. USA,92, 3483–3487.
Berridge, M. J., Downes, C. P. & Hanley, M. R. (1989).Neural and developmental actions of lithium: aunifying hypothesis. Cell, 59, 411–419.
Blatt, M. R. & Thiel, G (1993). Hormonal control of ionchannel gating. Annu. Rev. Plant Physiol. Plant Mol.Biol. 44, 543–567.
Blatt, M. R. & Thiel, G (1994). K+ channels of stomatal guardcells: bimodal control of the K+ inward-rectifierevoked by auxin. Plant J. 5, 55–68.
Bowie, J. U., Luthy, R. & Eisenberg, D. (1991). A method toidentify protein sequences that fold into a knownthree-dimensional structure. Nature, 253, 164–170.
Chen, C.-Y. A. & Shyu, A.-B. (1994). Selective degradationof early-response gene mRNAs: functional analyses ofsequence features of the AU-rich element. Mol. Cell.Biol. 14, 8471–8482.
Compton, T. (1990). Degenerate primers for DNAamplification. In: PCR Protocols: A Guide to Methods andApplications (Innis, M. A., Gelfand, D. H., Sninsky, J. J.& White, T. J., eds.), pp. 39–45, Academic Press,New York.
Conner, T. W., Goekjian, V. H., LaFayette, P. R. & Key, J. L.(1990). Structure and expression of two auxin-inducible genes from Arabidopsis. Plant Mol. Biol. 9,2487–2492.
Davis, R. W., Botstein, D. & Roth, J. R. (1980). AdvancedBacterial Genetics, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
Dellaporta, S. L., Wood, J. & Hicks, J. B. (1983). A plantDNA minipreparation: version II. Plant Mol. Biol. Rep.1, 19–21.
Ecker, J. R. & Davis, R. W. (1987). Plant defense genes areregulated by ethylene. Proc. Natl Acad. Sci. USA, 84,5202–5206.
Elledge, S. J., Mulligan, J. T., Ramer, S. W., Spottswood, M.& Davis R. W. (1991). lYES: a multifunctional cDNAexpression vector for the isolation of genes bycomplementation of yeast and Escherichia colimutations. Proc. Natl Acad. Sci. USA, 88, 1731–1735.
Estelle, M. (1992). The plant hormone auxin: insight insight. Bioessays, 14, 439–443.
Franco, A. R., Gee, M. A. & Guilfoyle, T. J. (1990). Inductionand superinduction of auxin-responsive mRNAs withauxin and protein synthesis inhibitors. J. Biol. Chem.265, 15845–15849.
Gee, M., Hagen, G. & Guilfoyle, T. J. (1991). Tissue-specificand organ-specific expression of soybean auxin-responsive transcripts GH3 and SAURs. Plant Cell, 3,419–430.
Gil, P., Liu, Y., Orbovic, V., Verkamp, E., Poff, K. L. & Green,P. (1994). Characterization of the auxin-inducibleSAUR-AC1 gene for use as a genetic tool in Arabidopsis.Plant Physiol. 104, 777–784.
Gonnet, G. M., Cohen, M. A. & Benner, S. A. (1992).Exhaustive matching of the entire protein sequencedatabase. Science, 256, 1443–1445.
Gottesman, S. & Maurizi M. R. (1992). Regulation ofproteolysis: Energy-dependent proteases and theirtargets. Microbiol. Rev. 56, 592–621.
Gray, J., Pictor, S., Shabbeer, J., Schuch, W. & Grierson, D.(1992). Molecular biology of fruit ripening and itsmanipulation with antisense genes. Plant Mol. Biol. 19,69–87.
Guilfoyle, T. J. (1986). Auxin-regulated gene expression inhigher plants. CRC Crit. Rev. Plant Sci. 4, 247–276.
Guilfoyle, T. J., Hagen, G., Li, Y., Ulmasov, T., Lui, Z.,Strabala, T. & Gee, M. (1993). Auxin-regulatedtranscription. Aust. J. Plant Physiol. 20, 489–502.
Hagen, G. & Guilfoyle, T. J. (1985). Rapid induction ofselective transcription by auxins. Mol. Cell. Biol. 5,1197–1203.
He, X., Treacy, M. N., Simmons, D. M., Ingraham,H. A., Swanson, L. W. & Rosenfeld, M. G. (1989).Expression of a large family of POU-domainregulatory genes in mammalian brain development.Nature, 340, 35–42.
Herschman, H. R. (1991). Primary response genes inducedby growth factors and tumor promoters. Annu. Rev.Biochem. 60, 281–319.
Hill, C. S. & Treisman, R. (1995). Transcriptional regulationby extracellular signals: mechanisms and specificity.Cell, 80, 199–211.
Hobbie, L. & Estelle, M. (1994). Genetic approaches toauxin action. Plant Cell Environ. 17, 525–540.
Huang, S. H., Hu, Y. Y., Wu, C. H. & Holcenberg, J. (1990).A simple method for direct cloning cDNA sequencethat flanks a region of known sequence from totalRNA by applying the inverse polymerase chainreaction. Nucl. Acids Res. 18, 1992.
Jones, M. D. (1993). Reverse transcription of mRNA byThermus aquaticus DNA polymerase followed bypolymerase chain reaction amplification. MethodsEnzymol. 218, 413–419.
Karim, F. D. & Thummel, C. S. (1991). Ecdysonecoordinates the timing and amounts of E74A and E74Btranscription in Drosophila. Genes Dev. 5, 1067–1079.
Kobe, B. & Deisenhofer, J. (1994). The leucine-rich repeat:a versatile binding motif. Trends Biochem. Sci. 19,415–421.
Leyser, H. M. O., Lincoln, C., Timpte, C., Lammer, D.,Turner, J. & Estelle, M. (1993). The auxin-resistancegene AXR1 of Arabidopsis encodes a protein related toubiquitin-activating enzyme E1. Nature, 364, 161–164.

JMB—MS 688
Auxin-responsive Gene Family in Arabidopsis 549
Li, Y., Hagen, G. & Guilfoyle, T. J. (1991). Anauxin-responsive promoter is differentially inducedby auxin gradients during tropisms. Plant Cell, 3,1167–1175.
Liang, X., Abel, S., Keller, J. A., Shen, N. F. & Theologis, A.(1992). The 1-aminocyclopropane-1-carboxylate syn-thase gene family of Arabidopsis thaliana. Proc. NatlAcad. Sci. USA, 89, 11046–11050.
Lincoln, C., Britton, J. & Estelle, M. (1990). Growth anddevelopment of the axr1 mutants of Arabidopsis. PlantCell, 2, 1071–1080.
McClure, B. A. & Guilfoyle, T. J. (1987). Characterizationof a class of small auxin-inducible soybeanpolyadenylated RNAs. Plant Mol. Biol. 9, 611–623.
Meyerowitz, E. M. (1989). Arabidopsis, a useful weed. Cell,56, 263–269.
Napoli, C., Lemieux, C. & Jorgensen, R. (1990).Introduction of a chimeric chalcone synthase gene intopetunia results in reversible co-suppression ofhomologous genes in trans. Plant Cell, 2, 279–289.
Newman, T. C., Ohme-Takagi, M., Taylor, C. B. & Green P. J.(1993). DST sequences, highly conserved among plantSAUR genes, target reporter transcripts for rapiddecay in tobacco. Plant Cell, 5, 701–714.
Oeller, P. W. & Theologis, A. (1995). Induction kinetics ofthe nuclear proteins encoded by the early indoleaceticacid-inducible genes, PS-IAA4/5 and PS-IAA6, in pea(Pisum sativum L.). Plant J. 7, 37–48.
Oeller, P. W., Keller, J. A., Parks J. E., Silbert, J. E. &Theologis, A. (1993). Structural characterization of theearly indoleacetic acid-inducible genes, PS-IAA4/5and PS-IAA6, of pea (Pisum sativum L.) J. Mol. Biol. 233,789–798.
Ohme-Takagi, M., Taylor, C. B., Newman, T. C. & Green P. J.(1993). The effect of sequences with high AU contenton mRNA stability in tobacco. Proc. Natl Acad. Sci.USA, 90, 11811–11815.
Osborne, B. I. & Baker, B. (1995). The maize transposonsare moving and mutagenizing in other genomes. Curr.Opin. Cell Biol. In the press.
Phillips, S. E. V. (1994). The b-ribbon DNA re-cognition motif. Annu. Rev. Biophys. Biomol. Struct. 23,671–701.
Pickett, F. B., Wilson, A. K. & Estelle, M. (1990). The aux1mutation of Arabidopsis confers both auxin andethylene resistance. Plant Physiol. 94, 1462–1466.
Raumann, B. E., Brown, B. M. & Sauer, R. T. (1994). Majorgroove DNA recognition by b-sheets: the ribbon-
helix-helix family of gene regulatory proteins. Curr.Op. Struct. Biol. 4, 36–43.
Rost, B. & Sander, C. (1993). Improved prediction ofprotein secondary structure by use of sequenceprofiles and neural networks. Proc. Natl Acad. Sci.USA, 90, 7558–7562.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). MolecularCloning: A Laboratory Manual. 2nd edit., Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Sanger, F., Nicken, S. & Coulson, A. R. (1977). DNAsequencing with chain terminating inhibitors. Proc.Natl Acad. Sci. USA, 74, 5463–5467.
Sassone-Corsi, P., Sisson, J. C. & Verma, I. M. (1988).Transcriptional autoregulation of the proto-oncogenefos. Nature, 334, 314–319.
Scherer, S. & Davis, R. W. (1979). Replacement ofchromosome segments with altered DNA sequencesconstructed in vitro. Proc. Natl Acad. Sci. USA, 76,4951–4955.
Theologis, A. (1986). Rapid gene regulation by auxin.Annu. Rev. Plant Physiol. 37, 407–438.
Theologis, A., Huynh, T. V. & Davis R. W. (1985). Rapidinduction of specific mRNAs by auxin in pea epicotyltissue. J. Mol. Biol. 183, 53–68.
Timpte, C., Wilson, A. K. & Estelle, M. (1992). Effects of theaxr2 mutation of Arabidopsis on cell shape in hypocotyland inflorescence. Planta, 188, 271–278.
Timpte, C., Wilson, A. K. & Estelle, M. (1994). The axr2-1mutation of Arabidopsis thaliana is a gain-of functionmutation that disrupts an early step in auxin response.Genetics, 138, 1239–1249.
Went, F. W. & Thimann, K. V. (1937). Phytohormones,Macmillan, New York.
Williamson, M. P. (1994). The structure and function ofproline-rich regions in proteins. Biochem. J. 297,249–260.
Wilson, A. K., Pickett, F. B., Turner, J. & Estelle, M. (1990).A dominant mutation in Arabidopsis confers resistanceto auxin, ethylene and abscisic acid. Mol. Gen. Genet.222, 377–383.
Wyatt, R. E., Ainley, W. M. Ainley, Nagao, R. T., Conner,T. W. & Key J. L. (1993). Expression of the ArabidopsisAtAux2-11 auxin-responsive gene in transgenicplants. Plant Mol. Biol. 22, 731–749.
Yamamoto, K. T., Mori, H. & Imaseki, H. (1992). cDNAcloning of indole-3acetic acid-regulated genes: Aux22and SAUR from mung bean (Vignia radiata) hypocotyltissue. Plant Cell Physiol. 33, 93–97.
Edited by N. H. Chua
(Received 28 March 1995; accepted 30 May 1995)
Copyright © 2022 FDOKUMEN




















