
Characterization of a New Defensin from Cowpea (Vigna unguiculata (L.) Walp
Upload
independentCategory
view
0download
0
Abstract Plants exude a variety of substances
through their roots, germinating seeds and aerial
parts. Some of these released compounds seem to
have an inhibitory effect against pathogens. The
aim of this work was to investigate and identify
antifungal proteins present in exudates from
imbibed cowpea seeds (Vigna unguiculata (L.)
Walp). The obtained exudation was analyzed in
regard to specific protein activities by enzymatic
or immunological assays for plant defense pro-
teins, from 4 h to 48 h of seed imbibition. Our
results show that cowpea seeds exudates present
several defense related proteins characterized as
b-1,3-glucanases, cystatins, vicilins and lipid
transfer proteins (LTPs), as well as a storage
vacuole membrane a-TIP protein, since the very
first hours of imbibition. These exudates also have
an ‘‘in vitro’’ inhibitory effect on the growth of
the fungus Fusarium oxysporum f. sp. phaseoli.
Our results suggest that seed exudates should
promote seed protection from soil pathogens.
Keywords LTP Æ b-1,3-glucanases Æ Vicilin ÆCystatin Æ Fusarium oxysporum f. sp. phaseoli ÆSeed exudates Æ Soil pathogen Æ Cowpea
Introduction
Cowpea (Vigna unguiculata) is a tropical legume
originated from Africa, which is cultivated in
most tropical regions of the world (Ehlers and
Hall 1997). In Brazil, cowpea is mainly produced
in the Northeastern part of the country and, in
this region, their grains are the main protein
source for the great majority of the population.
Cowpea seeds are severely damaged both in the
field and during storage by the cowpea weevil
(Callosobruchus maculatus), and by a great
number of pathogens such as virus, bacteria and
mainly fungi (Allen 1982; Rios 1988). Among the
fungal pathogens Fusarium oxysporum is an
important pathogen causing Fusarium wilt a
serious disease in Brazil (Ehler and Hall 1997). It
is thought that this high susceptibility of seeds of
T. L. Rose Æ L. A. Okorokov Æ A. O. Carvalho ÆV. M. Gomes (&)Laboratorio de Fisiologia e Bioquımica deMicrorganismos, Centro de Biociencias eBiotecnologia, Universidade Estadual do NorteFluminense, Av. Alberto Lamego 2000, 28013-600Campos dos Goytacazes, RJ, Brazile-mail: [email protected]
A. da S. Conceicao Æ F. Marty Æ D. Marty-MazarsLaboratoire de PhytoBiologie Cellulaire, Universitede Bourgogne, BP 47870, 21078 Dijon Cedex, France
J. Xavier-Filho Æ K. V. S. FernandesLaboratorio de Quımica e Funcao de Proteınas ePeptıdeos, Centro de Biociencias e Biotecnologia,Universidade Estadual do Norte Fluminense, Av.Alberto Lamego 2000, 28013-600 Campos dosGoytacazes, RJ, Brazil
Plant Soil (2006) 286:181–191
DOI 10.1007/s11104-006-9036-0
123
ORIGINAL PAPER
Defense proteins from Vigna unguiculataseed exudates: characterization and inhibitoryactivity against Fusarium oxysporum
Tatiana L. Rose Æ Alexandre da Silva Conceicao Æ Jose Xavier-Filho ÆLev A. Okorokov Æ Katia V. S. Fernandes Æ Francis Marty ÆDaniele Marty-Mazars Æ Andre O. Carvalho Æ Valdirene M. Gomes
Received: 16 November 2005 / Accepted: 12 May 2006 / Published online: 11 August 2006� Springer Science+Business Media B.V. 2006

cowpea to pests is associated with inadequate
defenses, either constitutive or induced (Singh
and Rachie 1985). Nevertheless, several cowpea
cultivars are able to express defense components
against many viruses, bacteria, fungi and insects
(Singh et al. 1985; Gomes and Xavier-Filho 1994)
and many of these defense compounds were
identified as proteins such as vicilins (7S globu-
lins), chitinases, b-1,3-glucanases, and the two
antimicrobial peptides, a defensin and a lipid
transfer protein (LTP) (Macedo et al. 1993;
Gomes et al. 1996; Fernandes and Xavier-Filho
1998; Carvalho et al. 2001). These proteins have
also been found in seeds of different plant species
(Kader 1996; Fernandes and Xavier Filho 1998;
Flores et al. 2001; Thomma et al. 2002).
Germinating seeds and roots release com-
pounds that may interfere with the development
of surrounding microorganisms (Nobrega et al.
2005; Okubara and Paulitz 2005). Several com-
pounds exuded from seeds were identified and
some of them have the ability to inhibit pathogen
growth, thus preventing seed infection, and others
have direct beneficial effects on the germination
itself (Barbour et al. 1991). The best-described
groups of defense compounds released from seeds
are soluble sugars, flavonoids, sterols and proteins
(Casey et al. 1998; Terras et al. 1995). Volatile
compounds are also among the well-known seed
exuded components and are chemically classified
as aldehydes, alcohols, ethylene, CO2 and fatty
acids (Gorecki et al. 1985; Duvick et al. 1992).
Emmert et al. (1998) revealed that the exudation
of the amino acid canavanine from alfalfa seeds
influenced negatively the growth of Bacillus cer-
eus, which is a beneficial microorganism known as
a suppressor of seedling damping off of alfalfa.
Terras et al. (1995) showed that exudates of rad-
ish seed contain as a principal component a pep-
tide identified as a plant defensin. These exuded
defensins are responsible for causing fungal
growth inhibition around the germinating seed,
thus providing a safe environment on seed
periphery during this vulnerable phase of plant
development.
In the last years, several proteins such as
trypsin and papain inhibitors (Xavier-Filho et al.
1989), variant vicilins (7S globulin storage pro-
teins) (Fernandes and Xavier-Filho 1998),
glucan hydrolases represented by chitinases and
b-1,3-glucanase (Gomes et al. 1996), antimicro-
bial peptides identified as lipid transfer proteins
(LTP) and defensins (Carvalho et al. 2001,
2004) were found in seeds of cowpea. These
proteins are commonly linked to plant defense
mechanisms. They are induced and accumulate
in the infected tissues systemically and also
exhibit antimicrobial activity (Van Loon and
Van Strien 1999). These proteins have different
described mechanisms of action. For example,
vicilins present affinity to chitin, binding to it
and interfering with fungal growth by an un-
known mechanism (Gomes et al. 1997, 1998).
Chitinases and b-1,3-glucanases are hydrolases
that break the b-1,3 linked glucans and b-1,4
linked N-acetyl-D-glucosamine, respectively,
which are the two major structural components
of the fungal cell wall, thus inhibiting their
development (Carlile et al. 2001). Defensins and
LTPs interact with fungal membrane causing its
permeabilization (Thevissen et al. 1999; Regente
et al. 2005).
The aim of this work was to identify defense
proteins in exudates from germinating cowpea
seeds and test the ability of such exudates for
inhibition of the fungal pathogen, Fusarium oxy-
sporum. Herein we report the identification,
partial characterization and antifungal activity of
several defense proteins exuded from imbibed
cowpea seeds. We consider such information as a
prerequisite to further understand the physiolog-
ical significance of their presence in the micro-
environment surrounding germinating seeds and
how microorganisms dwelling in it are affected by
them.
Materials and methods
Plant material
Cowpea (Vigna unguiculata L. Walp.) seeds of
the EPACE-10 cultivar, which are susceptible to
the bruchid insect Callosobruchus maculatus
(Xavier-Filho et al. 1989), were supplied by the
Centro de Ciencias Agrarias, Universidade Fed-
eral do Ceara, Fortaleza, Brazil where they were
developed.
182 Plant Soil (2006) 286:181–191
123

Fungi
The fungal isolate Fusarium oxysporum forma
specialis phaseoli, causing wilting of beans, were
kindly supplied by CNPAF/EMBRAPA, Goia-
nia, Goias, Brazil. The fungi were maintained on
Sabouraud agar (1.0% peptone, 2.0% glucose and
1.7% agar–agar).
Obtaining of seed exuded proteins and protein
determination
Seed exudate was obtained from cowpea using a
method similar to that described by Nelson and
Hsu (1994). To prepare the seed exudates, 50
seeds, with no cracks or other injuries on the pill,
were superficially disinfested for 5 min in 25 ml
0.5% sodium hypochloride containing one drop
of Tween-20 and finally rinsed three times with
sterile water. The disinfested seeds were added to
sterile flasks containing 50 ml of sodium acetate
buffer (50 mM, pH 4.5), water (pH 6.0), or
sodium phosphate buffer (100 mM, pH 7.5), at
27�C on a rotary shaker at 70 rpm for 4, 8, 16, 24,
32 or 48 h. At each time flask contents were fil-
tered, and the exuded proteins were recovered by
precipitation with ammonium sulfate (0–90%)
followed by dialysis against water and freeze-
drying. Protein content was determined as
described by Bradford (1976) using BSA as a
standard protein.
Fungal growth inhibition
Fungal growth inhibition was assayed as described
by Broekaert et al. (1990a) with some modifica-
tions. For the preparation of conidia of F. oxy-
sporum f. sp. phaseoli, fungal culture was
transferred to Petri dishes containing Sabouraud
agar for 12 days; after this period sterile distilled
water (10 ml) was added to the dish, and it was
gently shaken for 1 min for conidia liberation,
with the help of a Drigalski loop. Conidia were
quantified in a Neubauer chamber to ensure
appropriate dilutions in further inhibition assays.
Initially conidia suspension (20,000 in 1 ml of
Sabouraud broth) were incubated at 28�C with the
exudate solutions (200 ll at 100 mg ml–1), for
60 h, in microplates. Optical readings were taken
at 600 nm at each 6 h interval. All the experiments
were run in triplicate and the reading averages,
the standard errors and coefficients of variation
were calculated. A general control without addi-
tion of exudates was also utilized. After 60 h, fungi
cells were separated from the growth medium by
centrifugation, washed in 0.1 M Tris–HCl buffer
(pH 8.0) and plated for observation with a light
microscope (Axioplan Zeiss).
Gel electrophoresis
Sodium dodecyl sulfate electrophoresis (SDS-
PAGE) was carried out according to Laemmli
(1970). SDS-Tricine-gel electrophoresis was per-
formed according to Schagger and Von Jagow
(1987). Samples (1 mg of lyophilized exudate)
were solubilized in 0.1 ml Tris–HCl buffer
(100 mM, pH 8.0) containing 10% SDS, 5%
sucrose and 0.001% bromophenol blue, at 100�C
for 5 min and loaded (20 ll) on the gels.
Western blotting
Polyclonal antibodies specific against vicilins,
cystatins and LTPs from seeds of cowpea were
raised in New Zealand white rabbits according to
Macedo et al. (1993), Flores et al. (2001), and
Carvalho et al. (2001), respectively. Polyclonal
antibodies against a-TIP were prepared as
described by Johnson et al. (1989).
Western blottings were carried out using nitro-
cellulose membranes according to the method
described by Towbin et al. (1979). Blots were
incubated overnight with primary antiserum
against vicilins, LTPs and cystatins (at 1:2000) and
against a-TIP (at 1:3000) diluted in the blocking
buffer. Membranes were then washed (4 times,
during 10 min at each time) with 0.1 M phosphate
buffer containing 0.15 M NaCl, pH 7.3 (PBS). They
were incubated with peroxidase-conjugated goat
anti-rabbit secondary antibody (Sigma Immuno
Chemicals) (at 1:2000) diluted in blocking buffer,
for 2 h, at room temperature and then, washed as
above. Antibody binding was revealed using a
chemiluminescence detection Kit (ECL-Western
blotting detection reagents/RPN 2209) according
to the manufacturer’s instructions. The chemilu-
minescence signal was recorded on an X-ray film.
Plant Soil (2006) 286:181–191 183
123

Antibodies and ELISA test
Immunodetection ELISA assays were performed
on micro plates (96 wells; Nunc, Denmark), which
were initially coated with 100 ll of seed exudates
for 16 h. Antigen-coated plates were then washed
twice in PBS/0.05% Tween-20 and then blocked
with 1% BSA/PBS/0.05% Tween-20 for 1 h.
After one wash with PBS/0.05% Tween-20, vicilin
and LTP antibodies diluted at 1:5000 in 1% BSA/
PBS/0.05% Tween-20 were added. Plates were
incubated for 2 h at 37�C, and then washed four
times with PBS/0.05% Tween-20. Anti-IgG con-
jugated to alkaline phosphatase (at 1:2000 in 1%
BSA/PBS/0.05% Tween-20) was added and after
1 h of incubation at 37�C plates were washed four
times with PBS/0.05% Tween-20 and one last
time with PBS. The reaction was revealed using
ortho-phenylenediamine (OPD) as substrate.
Reaction intensity was measured using an ELISA
colorimetric plate reader at 410 nm. Negative
controls were performed by employing the IgG–
alkaline phosphatase complex without addition of
the anti-vicilin/anti-LTP IgGs and a blank well
with no added antibodies.
Enzymatic assays
b-1,3-glucanase was assayed by the method of
Fink et al. (1988), which employs laminarin
(Sigma Chemicals Co., St. Louis) as the substrate.
Exudates (50 ll) were incubated at 37�C, for
12 h, with 0.125 ml of a 2 mg ml–1 laminarin
solution in 0.05 M acetate buffer, pH 5.0 in a final
volume of 0.2 ml. Color was developed by the
copper–arsenomolybdate method (Nelson 1944;
Somogy 1952) and the absorbance was monitored
at 500 nm. One unit of activity was defined as the
concentration of enzyme that produces an
absorbance variation of 0.001, in relation to that
from the control (prepared by replacing the sub-
strate with buffer) reaction tube.
Chitinase activity was determined by incubat-
ing 50 ll of exudate with 0.25 M of the fluorogenic
substrate 4-methylumbelliferyl-N,N¢,N¢¢-tria-
cetylchitotrioside (Sigma Chemical Co.) in 2 ml of
25 mM phosphate buffer pH 5.0. The fluorescent
methylumbelliferyl (4-MU-sodium sat) released
during the reaction was measured using a Hitachi
F4500 fluorescence spectrophotometer, equipped
with a 320 nm primary filter and a 460 nm sec-
ondary filter. In order to relate fluorescence out-
put to the concentration of the released product a
calibration curve was plotted using 4-MU-sodium
salt. One unit of chitinase activity will liberate
1 nmol of methylumbeliferone (MU) per min.
Controls were made by replacing the substrate
with buffer. This methodology was described by
O’Brien and Colwell (1987).
Inhibitory activity assay
The inhibitory activity of the seed exuded cystatin,
against papain, was measured essentially as de-
scribed by Abe et al. (1994). The substrate for this
assay was the N-benzoyl-L-arginine-2-naphthyla-
mide (BANA, Sigma). The inhibitory activity was
detected as the reduction of BANA hydrolysis
activity of papain, per ml of inhibitor solution, and
expressed as the OD540 h–1 ml–1. Negative con-
trols were performed with no BANA or no seed
sample added to the reaction media. In a positive
control seed exudate was absent.
Results
Fungal growth inhibition
The incubation of F. oxysporum f. sp. phaseoli,
with the exudates obtained from cowpea seeds
imbibed in phosphate (pH 7.5), water (pH 6.0)
and acetate (pH 4.5) caused inhibition of 45.2,
82.1 and 75.7%, respectively, of fungal growth, in
comparison with the control (Fig. 1).
The micrographs taken from the fungal myc-
elia at the final time of the growth inhibition assay
demonstrated that fungal growth was inhibited by
all exudates in relation to the control and also
that the exudates were interfering with fungal
germination, germ tube elongation and formation
of hyphae (data not shown).
Characterization of proteins present on the
exudates
Analysis by SDS-PAGE electrophoresis showed
significant differences in the pattern of proteins
184 Plant Soil (2006) 286:181–191
123
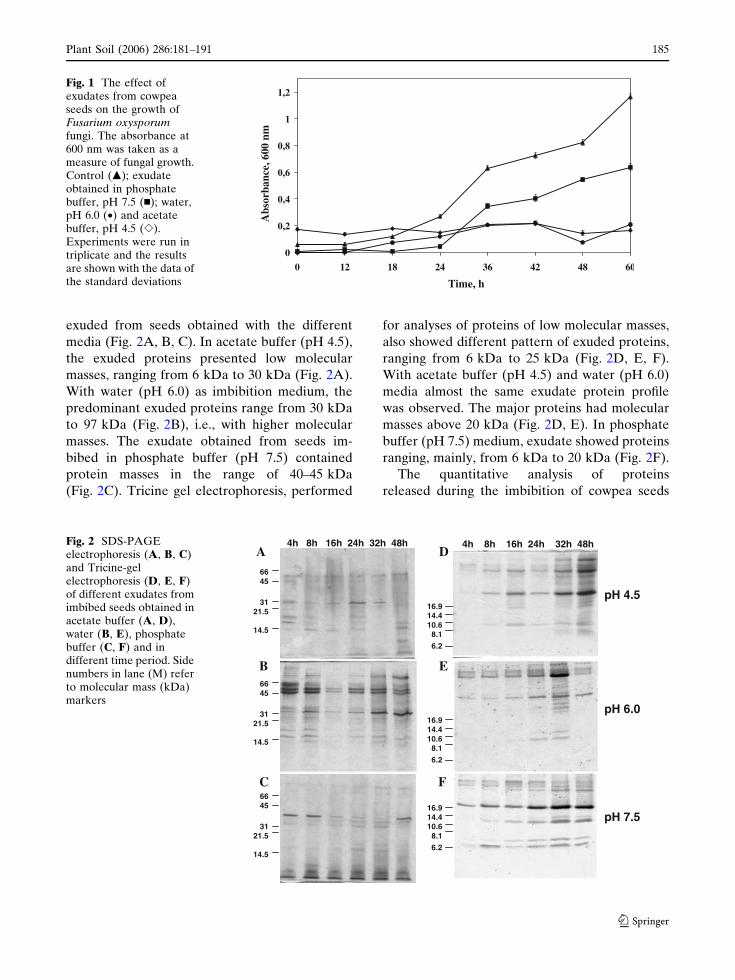
exuded from seeds obtained with the different
media (Fig. 2A, B, C). In acetate buffer (pH 4.5),
the exuded proteins presented low molecular
masses, ranging from 6 kDa to 30 kDa (Fig. 2A).
With water (pH 6.0) as imbibition medium, the
predominant exuded proteins range from 30 kDa
to 97 kDa (Fig. 2B), i.e., with higher molecular
masses. The exudate obtained from seeds im-
bibed in phosphate buffer (pH 7.5) contained
protein masses in the range of 40–45 kDa
(Fig. 2C). Tricine gel electrophoresis, performed
for analyses of proteins of low molecular masses,
also showed different pattern of exuded proteins,
ranging from 6 kDa to 25 kDa (Fig. 2D, E, F).
With acetate buffer (pH 4.5) and water (pH 6.0)
media almost the same exudate protein profile
was observed. The major proteins had molecular
masses above 20 kDa (Fig. 2D, E). In phosphate
buffer (pH 7.5) medium, exudate showed proteins
ranging, mainly, from 6 kDa to 20 kDa (Fig. 2F).
The quantitative analysis of proteins
released during the imbibition of cowpea seeds
0
0,2
0,4
0,6
0,8
1
1,2
0 12 18 24 36 42 48 60
Time, h
Abs
orba
nce,
600
nm
Fig. 1 The effect ofexudates from cowpeaseeds on the growth ofFusarium oxysporumfungi. The absorbance at600 nm was taken as ameasure of fungal growth.Control (m); exudateobtained in phosphatebuffer, pH 7.5 (n); water,pH 6.0 (•) and acetatebuffer, pH 4.5 (e).Experiments were run intriplicate and the resultsare shown with the data ofthe standard deviations
4h 8h 16h 24h 32h 48h
16.914.410.6
8.1
6.2
4h 8h 16h 24h 32h 48h
6645
3121.5
14.5
pH 4.5
16.914.410.6
8.1
6.2
6645
3121.5
14.5
pH 6.0
16.914.410.6
8.1
6.2
6645
3121.5
14.5
pH 7.5
A D
E
F
B
C
Fig. 2 SDS-PAGEelectrophoresis (A, B, C)and Tricine-gelelectrophoresis (D, E, F)of different exudates fromimbibed seeds obtained inacetate buffer (A, D),water (B, E), phosphatebuffer (C, F) and indifferent time period. Sidenumbers in lane (M) referto molecular mass (kDa)markers
Plant Soil (2006) 286:181–191 185
123

demonstrated that their content increased with
time (from 4 h to 48 h). Results also demonstrate
that the highest levels of exuded proteins oc-
curred from 24 h to 48 h for all conditions studied
(Fig. 3).
Specific assay to determine the b-1,3-glucanase
activity revealed its presence at all exudates since
the very beginning of imbibition, at 4 h. The level
of b-1,3-glucanase activity increased significantly
and was highest at 48 h of imbibition. At this
imbibition stage it reached up to six times of the
initial value found for all three media (Fig. 4).
Chitinase activity was analyzed by a fluorogenic
technique but no chitinase activity was detected
in the different analyzed exudates (data not
shown).
The qualitative analysis of the exuded proteins
revealed the presence of proteins immunorelated
to vicilins (66 kDa) (Fig. 5A), cystatins (96 kDa)
(Fig. 5C) and the antimicrobial LTP peptide
(10 kDa) (Fig. 5B) by Western blotting technique
using specific antisera against these proteins.
In order to quantify the specific amount of
vicilins and LTPs in the exudates, ELISA tests
were performed. The results showed that vicilins
represented ca. 3–10% of the total exuded seed
proteins and quantification of LTPs revealed that
they correspond to ca. 0.7–5.6% of total exuded
seed proteins. Vicilins and LTPs concentrations
reached a maximum after 4–8 h of imbibition and
remained almost constant until 48 h (Fig. 5). The
quantity of cystatins was determined by papain
inhibition assay, which showed that there were no
significant differences in cystatin activities among
all studied exudates. Maximum levels of exuda-
tion were already present at the early beginning
of imbibition for all times (data not shown).
The presence of the a-TIP, an aquaporin mem-
ber protein, was investigated also by Western
blotting technique by the use of a specific antibody
against it. Results revealed a cross-reactive band
of 31 kDa in exudates obtained with the three
different media with different kinetics (Fig. 6).
In the acetate buffer and water exudates the
presence of a-TIP was detected after 8 h and 16 h
of imbibition, respectively (Figs. 6A, B) with the
strongest response obtained in water exudates
derived from water imbibition (Fig. 6B). In phos-
phate buffer, the immunodetection was observed
earlier, at 4 h after imbibition (Fig. 6C).
Discussion
Biological effects of exuded proteins on fungi
growth
Initial infections of seeds and roots by pathogens
may, at one time, be driven by attractive nutritive
exudate molecules, and at the same time, be
restrained by some other defense related exuded
molecules (Nelson 1990). In order to investigate
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
pH 4.5 pH 6.0 pH 7.5
To
tal p
rote
ins
(mg
/ml)
A AA
BC
D
A
BB
C
D
E
AB B
C
DE
4h 8h 16h 24h 32h 48h 4h 8h 16h 24h 32h 48h 4h 8h 16h 24h 32h 48h
pH 4.5 pH 6.0 pH 7.5
Fig. 3 Proteinconcentration of thecowpea seed exudatesfrom different media.Protein content wasdetermined as describedby Bradford, using BSAas protein standard. Thesignificance of the resultswas analyzed by theStudent’s t-test(P < 0.05). Similar lettersindicate non-significantlydifferent values
186 Plant Soil (2006) 286:181–191
123

the cowpea seeds exudates in respect to their
antifungal inhibitory function, growth curves of
the plant pathogen F. oxysporum f. sp. phaseoli in
the presence of the three exudates were done.
The strongest inhibitory effects, obtained at pH
6.0 and 4.5, were noticed as compared to the
control medium (Fig. 1). The inhibitory activity
of fungal growth by seed exudates was shown to
be influenced by pH and the buffer used, which
mimics the natural environmental conditions
found in soils of Brazil. This differential inhibition
might be due to these treatments, which influence
the exudation of different proteins, qualitatively
and quantitatively, as can be observed on the
Figs. 2, 3 and 5. Such exuded substances, as
demonstrated by us and Terras et al. (1995),
interfere with microorganisms and may do so in a
specific way, which could be triggered by specific
stimuli and environmental conditions.
Terras et al. (1995) have also shown that
defensins, a group of antimicrobial peptides, are
also exuded by radish seeds. The exudation of
defensins by radish seeds occurs when their teg-
uments are disrupted either by natural radicles
emergence or by mechanical incisions. Further-
more, radish seeds when germinating in the
presence of defensins (Rs-AFP1 or Rs-AFP2)
suppressed the growth of the fungus Pyrenophora
tritici-repentis. We have previously shown by
protein purification that cowpea seeds also accu-
mulate several types of antimicrobial proteins
(Gomes et al. 1996, 1997; Carvalho et al. 2001).
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
pH 4.5 pH 6.0 pH 7.5
β-1,
3 g
luca
nas
e ac
tivi
ty
(un
its.
mg
-1 p
rote
in)
4h 8h 16h 24h 32h 48h 4h 8h 16h 24h 32h 48h 4h 8h 16h 24h 32h 48h
AA
A A
B
C
A AA
A
B
C
A A
AB
C
D
pH 4.5 pH 6.0 pH 7.5
Fig. 4 b-1,3-glucanaseactivity of cowpea seedexudates of differentimbibition media. Thesignificance of the resultswas analyzed by theStudent’s t-test(P < 0.05). Similar lettersindicate non-significantlydifferent values
4h 8h 16h 24h 32h 48h 4h 8h 16h 24h 32h 48h4h 8h 16h 24h 32h 48h
pH 4.5 pH 7.5pH 6.0
10
96
66
kDa(A)
(B)
(C)
Fig. 5 Western blotting of different exudates fromimbibed seeds using antisera against vicilin (A), LTP (B)and cystatin (C) obtained in acetate buffer (pH 4.5—left),water (pH 6.0—middle), and phosphate buffer (pH7.5—right) at different time
kDa
31
4h 8h 16h 24h 32h 48h
pH 4.5
pH 6.0
pH 7.5
31
31
(A)
(B)
(C)
Fig. 6 Western blotting using antiserum against a-TIP forexudates obtained in acetate buffer (A), water (B), andphosphate buffer (C) after different time of seed imbibi-tion
Plant Soil (2006) 286:181–191 187
123

Identification of the exuded proteins by
electrophoresis
The quality and quantity of materials released
from germinating seeds and developing plant
roots interfere with plant physiology and may be
affected by a number of factors. Some of those,
such as the stage of plant development, the soil
pH, or temperature, have been shown to influence
the release of certain exudate components into
the rhizosphere (Nelson 1990). In order to iden-
tify proteins in exudates obtained from cowpea
seeds, which are differentially induced by specific
environmental conditions, different imbibition
medium were used. Three media were selected
for their ion composition and principally for their
pH values of 4.5, 6.0 and 7.5, which bear a
resemblance to some soil characteristics of the
Atlantic forest and the Northeastern parts of
Brazil where cowpea is produced (Abreu et al.
2003). The analyses showed that many proteins
were exuded from imbibed cowpea seeds (Fig. 2).
These results suggest the state of an intense out-
flow of proteins from the seeds during the very
beginning of germination. The release of antimi-
crobial proteins from seeds during germination
has already been described on the literature
(Terras et al. 1995; Diz et al. 2003). Furthermore
it has been described that some antimicrobial
peptides, such as LTP, are released by dipping the
plant tissues in extraction buffer (Molina and
Garcia-Olmedo 1993) and, in cell cultures are
also secreted to the growth medium (Sterk et al.
1991). We have previously characterized a LTP
exuded from imbibed cowpea seeds (Diz et al.
2003).
The electrophoretic results also suggest that
exuded proteins from cowpea seeds are distinc-
tively expressed and/or extractable according to
the environmental conditions of seed imbibition
and seem to be severely influenced by the pH and
ionic strength of the surrounding environment.
The results also showed significant quantitative
expression differences among exuded proteins
from 4 h to 48 h. These data were confirmed
when total exuded proteins were estimated by the
Bradford (1976) method. In all tested medium,
concentrations of exuded proteins increased with
time (Fig. 3).
Partial characterization of the exuded proteins
Seeds of many legumes contain abundant supplies
of reserve proteins stored in the parenchyma cells
of the cotyledons. These cells are characterized by
the presence of many large protein bodies con-
sisting of an amorphous protein matrix sur-
rounded by a limiting membrane. In Phaseolus
vulgaris the major reserve proteins are vicilins
and phytohemagglutinins, 50% and 10% of the
total protein content of the cotyledons at seed
maturity, respectively (Johnson et al. 1989).
During seedling growth, the reserve proteins are
catabolized within the protein bodies, which are
known as hydrolytic compartments, while their
surrounding membranes remain intact. Legume
seed exudates contain compounds that inhibit the
growth of rhizobia and other bacteria (Materson
and Weaver 1984), and antimicrobial peptides are
also found in those exudates from non-legumi-
nous seeds of Zea mays, Mirabilis jalapa and
Amaranthus caudatus (Broekaert et al. 1990b;
Cammue et al. 1992; Duvick et al. 1992).
To investigate the presence of defense proteins
such as vicilins, LTPs and cystatins in exudates
obtained from cowpea imbibed seeds, we used
Western blotting technique using specific anti-
sera. These proteins were present in all samples
since the first hours of imbibition. Characteriza-
tion and antifungal activities of vicilins (Gomes
et al. 1997), LTPs (Molina et al. 1993; Kader
1996; Diz et al. 2003) and cystatins (Pernas et al.
1999; Siqueira-Junior et al. 2002) have been pre-
viously reported. Molecular masses for the three
protein categories determined in the present work
are in accordance to the literature, stressing that,
in the case of cystatins, the high molecular mass
observed, despite not being compatible with the
most usual sizes for 1 and 2 cystatin families, is
well-matched to those of the multidomain kinin-
ogen type of plant cystatins. Among these are the
potato multicystatin (Walsh and Strickland 1993;
Waldron et al. 1993) and tomato multicystatin
(Siqueira-Junior et al. 2002). Our findings thus
indicate that antimicrobial proteins are released
from seeds during germination. Western blottings
for vicilins, LTPs and cystatins showed no signif-
icant quantitative differences between the three
analyzed exudates. However, pH was seen to
188 Plant Soil (2006) 286:181–191
123

affect the exudation kinetics of these proteins
(Fig. 5).
Plants also produce enzymes such as b-1,
3-glucanases and chitinases (Brunner et al. 1998;
Santos et al. 2004) that can break down pathogen
cell wall components. Given the importance of
these enzymes as potential determinants in the
resistance of plants to fungal diseases (Ji and Kuc
1996; Lawrence et al. 1996), the presence of
b-1,3-glucanases and chitinases in the three exu-
dates from cowpea seeds were also analyzed. The
b-1,3-glucanase activity was detected since the
beginning of the imbibition, at 4 h and increased
significantly and was highest after 48 h of imbi-
bition (Fig. 4). This enzyme is involved in defense
and also participates in physiological and devel-
opmental processes during seed germination
(Sela-Buurlage et al. 1993). Chitinase activity,
tested by using the fluorogenic substrate
4-methylumbelliferyl-N,N¢,N¢¢-triacetylchitotrio-
side, was not detected in any of the different
analyzed exudates.
a-TIP (28–31 kDa) is a protein specific of the
membrane limited storage vacuoles, involved in
the transport of water (Marty 1997). In our
results specific anti-a-TIP antibodies were
employed and revealed a cross-reactive band of
31 kDa in exudates obtained with the three
different media with different kinetic (Fig. 6).
However, further studies are necessary to
understand the origin and the function of this
protein, immunologically related to a-TIP, in
exudates of cowpea seeds.
The work presented here shows that cowpea
seeds exude, during germination, several proteins,
among those some defense related proteins, from
the very first hours of imbibition. These proteins
have an ‘‘in vitro’’ inhibitory effect on the growth
of the F. oxysporum f. sp. phaseoli fungus. The
findings reported in this paper on the presence of
defense proteins in exudates from cowpea seeds
suggest an important role for these proteins in
constitutive host defense mechanisms against
microbial pathogens during seed germination.
Microorganisms, such as soil-borne bacteria and
fungi, dwelling on rhizosphere, and the regulating
balance of such microbial populations is crucial to
plant survival because some microorganisms are
pathogens that can attack and destroy plant
tissues that come in contact with soil, like seeds and
roots, while others are beneficial organisms that
have an imperative role on normal plant develop-
ment such as mycorrhiza. Thus, understanding
what kind of substances are released by seeds and
also roots, in what developmental stage and also in
which environmental conditions these events take
place are the first steps in understanding how ex-
udates might affect the spermosphere and their
population microorganisms (Zhu et al. 1997;
Emmert et al. 1998; Nobrega et al. 2005).
This knowledge is also essential for developing
technologies on how to manipulate the spermo-
sphere and rhizosphere environment in order to
reduce or eliminate seed or root infection by
pathogenic fungi. From these analyses new in-
sights on how to manipulate the spermosphere
may be drawn. Further work will include the
usage of these mechanisms for the development
of biological control approaches of fungal patho-
gens typical for this crop.
Acknowledgements This study is a part of the MSc De-gree Thesis of TLR, carried out in the Universidade Es-tadual do Norte Fluminense through an internationalcollaboration between CAPES and COFECUB. Weacknowledge the financial support of the Brazilian agen-cies CAPES, CNPq and FAPERJ and the InternationalFoundation for Science (IFS), Stockholm, Sweden,through a Grant to C/2806-3F. We are grateful to M.T.Gobo for technical assistance.
References
Abe M, Abe K, Iwabuchi K, Domoto C, Arai S (1994)Corn cystastin I expressed in Escherichia coli: inves-tigation of its inhibitory profile and occurrence in cornkernels. J Biochem 116:488–492
Abreu Jr CH, Muraoka T, Lavorante AF (2003) Rela-tionship between acidity and chemical properties ofBrazilian soils. Sci Agric 60:337–343
Allen DJ (1982) The pathology of tropical food legumes;disease resistance in crop improvement. John Wiley,Chinchester, pp 431–445
Barbour WM, Hatterman DR, Stacey G (1991) Chemo-taxis of Bradyrhizobium japonicum to soybean exu-dates. Appl Environ Microbiol 57:2625–2639
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of proteinutilizing the principle of protein-dye binding. AnalBiolchem 72:248–254
Broekaert WF, Terras FRG, Cammue BPA, Vanderley-den J (1990a) An automated quantitative for fungalgrowth inhibition. FEMS Microbiol Lett 69:55–60
Plant Soil (2006) 286:181–191 189
123

Broekaert WF, Lee H-I, Kush A, Chua N-H, RaikhelN (1990b) Wound-induced accumulation of mRNAcontaining a hevein sequence in laticifers of rub-ber tree (Hevea brasiliensis). Proc Natl Acad Sci87:7633–7637
Brunner F, Stintzi A, Fritig B, Legrand M (1998) Substrateof tobacco chitinases. Plant J 14:225–234
Cammue BPA, Deboll MFC, Terras FRG, Proost P,Vandamme J, Rees SB, Vanderlevden J, BroekaertWF (1992) Isolation and characterization of a novelclass of plant antimicrobial peptides from Mirabilisjalapa L. seeds. J Biol Chem 267:2228–2233
Carlile MJ, Watkinson SC, Gooday GW (2001) The fungi.Academic Press, USA
Carvalho AO, Tavares OLM, Santos IS, Da Cunha M,Gomes VM (2001) Antimicrobial peptide and immu-nolocalization of a LTP in Vigna unguiculata seeds.Plant Physiol Biochem 39:137–146
Carvalho AO, Teodoro CES, Da Cunha M, Okorokova-Facanha AL, Okorokov LA, Fernandes KVS, GomesVM (2004) Localization of a lipid transfer protein inVigna unguiculata seeds. Physiol Plant 122:328–336
Casey CE, O’Sullivan OB, O’Gara F, Glennon JD (1998)Ion chromatographic analysis of nutrients in seedexudate for microbial colonization. J Chromatogr804:311–318
Diz MSS, Carvalho AO, Gomes VM (2003) Purificationand molecular mass determination of a lipid transferprotein exuded from Vigna unguiculata seeds. Braz JPlant Physiol 15:171–175
Duvick JP, Rood T, Rao AG, Marshak DR (1992) Isola-tion and characterization of a novel class of plantantimicrobial peptides from maize (Zea mays) ker-nels. J Biol Chem 267:18814–18820
Ehlers JD, Hall AE (1997) Cowpea (Vigna unguiculata L.Walp.). Field Crops Res 53:187–204
Emmert EAB, Milner JL, Lee JC, Pulvermacher KL,Olivares HA, Clardy J, Handelsman J (1998) Effect ofcanavanine from alfafa seeds on the population biol-ogy of Bacillus cereus. Appl Environ Microbiol64:4683–4688
Fernandes KVS, Xavier-Filho J (1998) The biological rolesof legume seed vicilins (7s storage proteins). TrendsComp Biochem Physiol 2:241–245
Fink W, Liefland M, Mendgen K (1988) Chitinases andb-1,3-glucanases in the apoplastic compartment of oatleaves (Avena sativa L.). Plant Physiol 88:270–275
Flores VMQ, Louro RP, Xavier Filho J, Barratt DHP,Shewry PR, Fernandes KVS (2001) Temporal andtissue localization of a cowpea (Vigna unguiculataWalpers cv. pituba) cystatin. Physiol Plant111:195–199
Gomes VM, Xavier-Filho J (1994) Biochemical defencesof plants. Braz Arq Biol Tecnol 37:371–383
Gomes VM, Oliveira AEA, Xavier-Filho J (1996) Achitinase and a b-1,3-glucanase isolated from theseeds of cowpea (Vigna unguiculata L. Walp) inhibitthe growth of fungi and insect pests of the seeds. J SciFood Agric 72:86–90
Gomes VM, Mosqueda M-I, Blanco-Labra A, Sales MP,Fernandes KVS, Cordeiro RA, Xavier-Filho J (1997)Vicilin storage proteins from Vigna ungiculata(Legume) seeds inhibit fungal growth. J Agric FoodChem 45:4110–4115
Gomes VM, Okorokov LA, Rose TL, Fernandes KVS,Xavier-Filho J (1998) Legume vicilins 7S storageglobulins inhibit yeast growth and glucose stimulatedacidification of the medium by yeast cells. BiochimBiophys Acta 1379:207–216
Gorecki RJ, Harman GE, Mattick LR (1985) The volatileexudates from germinating pea seeds of differentviability and vigor. Can J Bot 63:1035–1039
Ji C, Kuc J (1996) Antifungal activity of cucumber b-1,3-glucanase and chitinase. Physiol Mol Plant Pathol.49:257–265
Johnson KD, Herman EM, Chrispeels MJ (1989) Anabundant, highly conserved tonoplast protein in seeds.Plant Physiol 91:1006–1013
Kader J-C (1996) Lipid-transfer protein in plants. AnnuRev Plant Physiol Plant Mol Biol 47: 627–654
Laemmli UK (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature227:680–685
Lawrence CB, Joosten MHA, Tuzun S (1996) Differentialinduction of plant pathogenesis-related protein intomato by Alternaria solani and the association ofbasic chitinase isozyme with resistence. Physiol MolPlant Pathol 48:361–371
Macedo MLR, Andrade LBS, Moraes RA, Xavier-Filho J(1993) Vicilin variants and the resistance of cowpea(Vigna unguiculata) seeds to the cowpea weevil(Callosobruchus maculatus). Comp Biochem Physiol105C:89–94
Marty F (1997) The biogenesis of vacuoles: insights frommicroscopy. Adv Plant Res 25:1–42
Materson LA, Weaver RW (1984) Survival of Rhizobiumtrifolii on toxic and non-toxic arrowleaf clover seeds.Soil Biol Biochem 16:533–535
Molina A, Garcıa-Olmedo F (1993) Developmental andpathogen-induced expression of three barley genesencoding lipid transfer proteins. Plant J 4:983–991
Molina A, Segura A, Garcia-Olmedo F (1993) Lipidtransfer proteins (nsLTP) from barley and maizeleaves are potent inhibitors of bacterial and fungalplant pathogens. FEBS Lett 2: 119–122
Nelson NA (1944) Photometric adaptation of the Somogymethod for the determination of glucose. J Biol Chem153:375–380
Nelson EB (1990) Exudate molecules initiating fungalresponses to seeds and roots. Plant Soil 129:61–73
Nelson EB, Hsu JST (1994) Nutritional factors affectingresponses of sporangia of Pythium ultimum to ger-mination stimulants. Phytopathology 84:677–683
Nobrega FM, Santos IS, Da Cunha M, Carvalho AO,Gomes VM (2005) Antimicrobial proteins from cow-pea root exudates: inhibitory activity against Fusari-um oxysporum and purification of a chitinase-likeprotein. Plant Soil 272:223–232
190 Plant Soil (2006) 286:181–191
123

O’Brien M, Colwell RR (1987) A rapid test for chitinaseactivity that uses 4-methylumberiferyl-N-acetyl-D-glucosaminide. Appl Environ Microbiol 7:1718–1720
Okubara PA, Paulitz TC (2005) Root defense responses tofungal pathogens: a molecular perspective. Plant Soil274:215–226
Pernas M, Lopez-Solanilla E, Sanchez-Monge R, SalcedoG, Rodrıguez-Palenzuela P (1999) Antifungal activityof a plant cystatin. Mol Plant Microbe Interac12:624–627
Regente MC, Giudici AM, Villalaın J, de la Canal L(2005) The cytotoxic properties of a plant lipidtransfer protein involve membrane permeabilizationof target cells. Lett Appl Microbiol 40:183–189
Rios GP (1988) Doencas fungicas e bacterianas do caupı.Em: O caupı no Brasil. In: Watt EE, Araujo JPP (eds)EMBRAPA-IITA, Idaban, Nigeria, pp 551–589
Santos IS, Machado OLT, Da Cunha M, Gomes VM(2004) A chitinase from Adenanthera pavonina L.seeds: purification, characterization and immunolo-calization Plant Sci 167:1203–1210
Schagger H, Von Jagow G (1987) Tricine-sodium dode-cylsulfate polyacrylamide gel eletrophoresis for sepa-ration of proteins in the range from 1 to 100 kDa.Anal Biochem 166:368–379
Sela-Buurlage MB, Ponstein AS, Bres-Vloemans SA,Melchers LS, van den Elzen PJM, Cornelissen BJC(1993) Only specific tobacco (Nicotiana tabacum)chitinases and b-1,3-glucanases exhibit antifungalactivity. Plant Physiol 101:857–863
Singh SR, Rachie KO (1985) In: Cowpea, research, pro-duction and utilization. John Wiley & Sons, Chiche-ster
Singh BB, Singh SR, Adjadi O (1985) Bruchid resistancein cowpea. Crop Sci 25:736–739
Siqueira-Junior CL, Fernandes KVS, Machado OLT, DaCunha M, Gomes VM, Moura D, Jacinto T (2002)87 kDa tomato cystatin exhibits properties of adefense protein and forms protein crystals in prosy-stemin overexpressing transgenic plants. Plant PhysiolBiochem 40:247–254
Somogy M (1952) Notes on sugar determination. J BiolChem 19:19–23
Sterk P, Booij H, Schellekens GA, Van Kammen A, DeVries SC (1991) Cell-specific expression of the carrotEP2 lipid transfer protein gene. Plant Cell 3:907–921
Terras FRG, Eggermont K, Kovaleva V, Raikhel NV,Osborn RW, Kester A, Rees SB, Torrekens S, VanLeuven F, Vanderleyden J, Cammue BPA, BroekaertWF (1995) Small cysteine-rich antifungal proteinsfrom radish: their role in host defense. Plant Cell7:573–588
Thevissen K, Terras FRG, Broekaert WF (1999) Per-meabilization of fungal membranes by plant defensinsinhibits fungal growth. Appl Environ Microbiol65(12):5451–5458
Thomma BPHJ, Cammue BPA, Thevissen K (2002) Plantdefensins. Planta 216:193–202
Towbin H, Stachelin NT, Gordon J (1979) Eletrophoretictransfer of proteins from polyacrylamide gels tonitrocellulose sheets: procedures and some applica-tions. Proc Natl Acad Sci 176:4350–4354
Van Loon LC, Van Serien EA (1999) The families ofpathogenesis related proteins, theirs activities, andcomparative analysis of PR-1 type proteins. PhysiolMol Plant Pathol 55:85–97
Waldron C, Wegrich LM, Merlo PAO, Walsh TA (1993)Characterization of a genomic sequence coding forpotato multicystatin, an eight-domain cysteine pro-teinase inhibitor. Plant Mol Biol 23:801–812
Walsh TA, Strickland JA (1993) Proteolysis of the85-kilodalton crystalline cysteine proteinase inhibitorfrom potato releases functional cystatin domain. PlantPhysiol 103:1227–1234
Xavier-Filho J, Campos FAP, Ary MB, Silva CP, CarvalhoMMM, Macedo MLR, Lemos FJA, Grant G (1989)Poor correlation between the levels of proteinaseinhibitors found in seeds of differente cultivars ofcowpea (Vigna unguiculata) and the resistance/sus-ceptibility to predation by Callosobruchus maculates.J Agric Food Chem 37:1139–1143
Zhu Y, Pierson III LS, Hawes MC (1997) lnduction ofmicrobial genes for pathogenesis and symbiosis bychemicals from root border cells. Plant Physiol115:1691–1698
Plant Soil (2006) 286:181–191 191
123
Copyright © 2022 FDOKUMEN




















