
Antibiotic Prophylaxis in Gastric, Biliary and Colonic Surgery
Upload
independentCategory
view
1download
0
Journal of Autoimmunity 24 (2005) 209e219
www.elsevier.com/locate/issn/08968411
Phylogenetic and immunological definition of four lipoylatedproteins from Novosphingobium aromaticivorans,
implications for primary biliary cirrhosis
Kerstien A. Padgetta, Carlo Selmia,b, Thomas P. Kennya, Patrick S.C. Leunga,David L. Balkwillc, Aftab A. Ansarid, Ross L. Coppele, M. Eric Gershwina,*
aDivision of Rheumatology, Allergy and Clinical Immunology, University of California School of Medicine,
GBSF, 451 E. Health Sciences Drive, Suite 6510, Davis, California 95616, USAbDivision of Internal Medicine, Department of Medicine, Surgery and Dentistry, University of Milan, Milan, Italy
cDepartment of Biological Sciences, Florida State University, Tallahassee, FL, USAdDepartment of Pathology, Emory University School of Medicine, Atlanta, GA, USA
eDepartment of Microbiology, Monash University, Clayton, Vic., Australia
Received 24 November 2004
Abstract
Novosphingobium aromaticivorans, a unique ubiquitous bacterium that metabolizes xenobiotics and activates environmentalestrogens, has been suggested as a pathogenic factor in the development of primary biliary cirrhosis (PBC). To define the molecularbasis of PBC sera reactivity, we investigated the characteristic of the bacterial antigens involved. We cloned and sequenced four genesfrom N. aromaticivorans coding for immunoreactive proteins, arbitrarily named Novo 1 through Novo 4. We subsequently analyzed
these proteins for their homology to known mitochondrial proteins and defined their reactivity using monoclonal antibodies (mAbs),rabbit anti-lipoic acid antibody, and PBC/control sera. Moreover, we studied their phylogenetic relation with the known PBCautoantigens. Novo proteins have an extraordinary degree of amino acid homology with all of the major human mitochondrial
autoantigens PDC-E2 (Novo 1 and 2), OGDC-E2 (Novo 3), and BCOADC-E2 (Novo 4). Moreover, Novo 1e4 contain a lipoylateddomain, are recognized by AMA-positive sera, and react with specific mAbs to mitochondrial antigens. Interestingly, thephylogenetic relation of the proteins emphasizes the conservation of the lipoylated domain. In conclusion, our data provide a high
degree of confidence that N. aromaticivorans may potentiate the breakdown of self tolerance in genetically susceptible individuals.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Molecular mimicry; Protein homology; Pyruvate dehydrogenase; Lipoic acid
1. Introduction
Primary biliary cirrhosis (PBC) is an autoimmunedisease of unknown etiology leading to progressivedestruction of intrahepatic bile ducts and eventuallyliver cirrhosis [1]. Anti-mitochondrial antibodies (AMA)
* Corresponding author. Tel.: C1 530 752 2884; fax: C1 530 752
4669.
E-mail address: [email protected] (M.E. Gershwin).
0896-8411/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jaut.2005.01.012
are uniquely detected in sera of 90e95% of patients withPBC and have been shown to react with the lipoylateddomains of the E2 components of the 2-oxoaciddehydrogenase complex (2-OADC) family, includingthe E-2 subunits of the pyruvate dehydrogenase complex(PDC-E2), 2-oxogluterate dehydrogenase complex(OGDC-E2), and branched-chain 2-oxoacid dehydroge-nase complex (BCOADC-E2) [2].
Although genetic background appears to be importantin determining susceptibility to PBC [3], environmental

210 K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
factors, including xenobiotics [4] andmicrobial infections[5], have been implicated as potential triggers ofautoimmunity in predisposed individuals. Among thelatter it has been shown that sera from patients with PBCrecognize homologs of human autoantigens from a num-ber of microorganisms [5].
Novosphingobium aromaticivorans is a strictly aerobic,gram-negative alpha proteobacterium that has recentlybeen reclassified from the genus Sphingomonas [6].N. aromaticivorans is ubiquitous in the environmentand has been studied by environmental biologists forapplications in bioremediation because of its capabilityto degrade a broad range of aromatic hydrocarbons [7]as well as the naturally occurring steroid hormone 17b-estradiol [8]. We have recently demonstrated, usingcrude bacterial lysates and Western blot techniques, thatsera from patients with PBC (including a groupof AMA-negative sera) recognized proteins fromN. aromaticivorans with 100- to 1000-fold higher titercompared to similar lipoylated proteins from Escher-ichia coli [9].
Based on the molecular weights of these proteins wereasoned that they could be bacterial homologs of themitochondrial autoantigens previously identified astargets of AMAs in PBC. To better define these bacterialhomologs and to ascertain the phylogenetic relationshipin the evolution of these reactive proteins, the genes en-coding the lipoylated proteins from N. aromaticivoranswere cloned, sequenced and analyzed. Each of theserecombinant proteins, arbitrarily referred to as Novo 1,2, 3 and 4, was examined by sequence comparison andidentified as homologs of human 2-OADC enzymes. Wereport herein the definition of the four recombinantNovo proteins using specific monoclonal antibodies aswell as PBC/control sera, and assess them phylogenet-ically to gain insight into the evolutionary relationshipsof these proteins and their human counterparts.
2. Methods
2.1. Culture of N. aromaticivorans and isolation ofgenomic DNA
A bacterial culture of N. aromaticivorans was used tostreak a trypticase soy agar plate and subsequently
incubated at 37 �C for 24 h. Two single colonies wereused to inoculate a 2!10-ml trypticase soy broth andincubated at 37 �C for 24 h. The cultures were thencentrifuged and the bacterial pellets stored at �20 �C.Genomic DNA was isolated by resuspending each frozenpellet in bacterial lysis solution containing 50 mM TrispH 8, 50 mM EDTA pH 8, 1% SDS and 2 mg/mlProteinase K (New England Biolabs, Beverly, MA). Thesamples were then incubated for 1 h at 37 �C, cooled onice and passed through a 1-ml tuberculin syringe severaltimes to reduce viscosity. The DNA was extracted twicewith an equal volume of phenol/chloroform/isoamylalcohol (ratio 25:24:1) and precipitated with 1/10volume of 3 M sodium acetate (pH 5.5) and 3 volumesof 100% ice-cold ethanol. The DNA was then resus-pended in TE pH 8 (10 mM Tris pH 8, 1 mM EDTA pH8) and digested with 1 mg/ml RNase (Roche MolecularSystems, Alameda, CA) for 30 min at 37 �C to eliminatepotential RNA contamination.
2.2. PCR amplification of genomic DNA
All PCR reactions were carried out in 50 ml volumewith 500 ng of N. aromaticivorans genomic DNA, 10pmol of each primer, 250 mMof each nucleotide and 1.25to 2 U of TaqGold hotstart enzyme (Applied Biosys-tems, Foster City, CA). Tables 1 and 2 illustrate theconditions used for the amplification of Novo genes andthe sequences of the oligonucleotides used, respectively.The high GC content of Novo 1, 3 and 4 necessitated theaddition of Betaine (Sigma Chemicals, St. Louis, MO) toa final concentration of 0.5 M to the PCR reactions inorder to facilitate the separation of template strands andthe stability of the DNA/primer duplex. The obtainedPCR products were visualized on a 1% TAEeagarosegel and column purified before cloning.
2.3. Construction of Novo clones
We inserted PCR products of Novo 1 and 2 into theTopo pCR2.1 cloning vector (Invitrogen, Carlsbad, CA)and then subcloned the genes directionally into bothpRSET A (Invitrogen) and pCal-n-Flag (Stratagene, LaJolla, CA) expression vectors using BamHI and HindIIIrestriction sites. Novo 3 and 4 PCR fragments were
Table 1
Specific conditions and product size for Novo genes amplification
Hot start 3 cycles 25 cycles Expected product
size (bp)
Novo 1 10 min/95 �C 95 �C/1 min, 64 �C/30 s, 72 �C/2 min 95 �C/20 s, 68 �C/2 min 1389
Novo 2 10 min/95 �C 95 �C/1 min, 58 �C/30 s, 72 �C/2 min 95 �C/15 s,59 �C/20 s
72 �C/1.5 min
1405
Novo 3 10 min/95 �C 95 �C/30 s, 60 �C/1 min, 72 �C/1.5 min 95 �C/15 s, 68 �C/2 min 1239
Novo 4 10 min/95 �C 95 �C/30 s, 54 �C/1 min, 72 �C/2 min 95 �C/15 s, 68 �C/2 min 1361

211K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
Table 2
Primer sequences for Novo genomic DNA amplification
Forward primer Reverse primer
Novo 1 AATAGGATCCATGCGCTGCGACATGGCG AATAAAGCTTCAGGCGACCAGGCCGAGCG
Novo 2 AATAGGATCCATGGCAATCGAACTGAAG AAAAAAGCTTCAGCGATAGCAGACCTTG
Novo 3 AAAGGATCCGTGCTGAACGAACTGCGCATC AAAGAATTCTCACGGCCGTTCCAGCGTTG
Novo 4 AAATGGATCCATGGGAACCTACACATTCC AAATGAATTCTCAGTCCGCCAGCAGCAGCAC
cloned directly into the pCal-n-Flag expression vectorusing BamHI and EcoRI restriction sites. Because ofsolubility issues, Novo1 and Novo3 were furthersubcloned into the pET32a bacterial expression vector,which contains the S-tag and 6!His purification tag.All DNAs were transformed into BL21DE3 Goldcompetent E. coli cells (Stratagene) and grown overnightat 37 �C on solid Luria Bertani (LB) mediumsupplemented with 200 mg/ml ampicillin (Sigma). Single,well-isolated colonies were then grown overnight inliquid LB/amp medium and the plasmid DNA isolatedusing the Qiagen plasmid purification kit (Qiagen,Valencia, CA). The resulting clones were analyzed bydiagnostic restriction digests, nucleotide sequencing andprotein expression studies. The pCal-n-Flag expressionvector was chosen for several reasons. First, the vector isinducible with protein expression being repressed undernormal growth conditions and induced by addingisopropyl-b-D-thiogalactopyranoside (IPTG) to thegrowth medium. Second, the vector contains a small,26-amino-acid calmodulin-binding peptide (CBP) affin-ity tag for protein purification, a FLAG epitope forfusion protein detection, and an enterokinase cleavagesite for the removal of the N-terminal fusion tag indownstream applications, if needed. The pET32aexpression vector (Novagen, Madison, WI) was chosenbecause it contains the thioredoxin (TRX) polypeptidewhich has been reported to enhance the solubility of itsfusion partner. Furthermore, this vector is inducible andcarries the S-tag and 6!His purification tags which arearranged in tandem. These two tags allow fusionproteins to be purified under native and/or denaturingconditions.
2.4. Protein expression studies
Bacterial cells harboring the different Novo con-structs were grown overnight in 2 ml of LB/amp brothsupplemented with 400 mg/ml ampicillin. Cells werediluted 1:20 into fresh LB/ampicillin broth and grownfor 2 h at 37 �C. A 100 ml aliquot of cells was removedand stored for later analysis. The remaining culture wasinduced with 1 mM IPTG grown at 37 �C and also atroom temperature to limit the possibility of forminginclusion bodies. One-milliliter aliquots were thenremoved at 1, 2 and 3 h after induction for proteinanalysis. The aliquots were centrifuged for 5 min at full
speed and the cell pellets were lysed with CelLytic lysissolution (Sigma-Aldrich, St. Louis, MO). Fusion pro-tein expression was detected by Coomassie-stainedprotein gels and confirmed by Western blot analysesusing either anti-FLAG M2 monoclonal antibody(Stratagene) or Tetra-His monoclonal antibody (Qia-gen) depending on the fusion tag, in addition tomonoclonal antibodies directed against mitochondrialantigens as described below.
2.5. Protein purification
Five hundred milliliters of exponentially growingbacterial cells harboring the fusion constructs wasinduced with IPTG and grown for 3e4 h at roomtemperature to reduce formation of inclusion bodies.The induced cells were centrifuged at 5000!g and thecell pellets were stored overnight at �20 �C. Thebacterial cells were lysed with CelLytic lysis buffer(Sigma-Aldrich). The Novo1 and Novo3 fusion proteinswere purified via interaction of their 6!His purificationtag with NieNTA resin (Qiagen). The Novo2 andNovo4 fusion proteins were purified using the Calmod-ulin purification system (Stratagene). All Novo fusionproteins, except Novo1, were purified under nativeconditions.
2.6. Sequence analysis
Column-purified Miniprep DNA was sequenced fromboth directions by dye-terminator reactions on an ABI3730 capillary sequencer (Applied Biosystems Inc.,Foster City, CA). Sequences that yielded ambiguousresults near the 3# terminus were resequenced withinternal primers. The raw sequence data were convertedto Macintosh format and initially analyzed with theEditView software (Applied Biosystems). Sequenceswere then assembled into contigs using Vector NTIsoftware (InforMax Inc., Frederick, MD) and translatedinto their corresponding amino acid sequence.
2.7. BLAST searches and phylogenetic study
BLAST searches were performed on the nucleotideand deduced amino acid sequences of the constructsusing the non-redundant blast engines and databasesincluding all reported sequences from any organism at

212 K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
NCBI. We investigated the phylogenetic relationship ofthe 4 Novo proteins with known sequences from otherorganisms. The deduced amino acid sequence from eachclone was used in a BLAST search against all organismsthat are present in the NCBI database and the resultinghits were ordered according to taxonomy. The mostrelevant amino acid sequences from various organismswere then aligned as FASTA files in ClustalW anda phylogenetic tree was constructed using Phylodendronphylogenetic tree printer version 0.8d. Both softwareversions are available on the EMBnet website. To obtaina representative dendrogram, we included speciesbelonging to prokaryotes and eukaryotes, includingthe human homologs. PDC-E2 and E1beta of represen-tative species from three subgroups of proteobacteria(i.e. alpha, beta and gamma) were also included. Thesedata also included organisms previously investigated inthe pathogenesis of PBC, such as E. coli[10], Chlamydiapneumoniae[11] and Saccharomyces cerevisiae[12].
2.8. Immunological studies
Western blot analysis was used to further define theimmunological characteristics of the four recombinantNovo proteins. Twenty micrograms of each purifiedNOVO protein was fractionated on a 10% polyacryl-amide gel and then transferred onto nitrocellulose paper[9]. Mouse-derived IgG monoclonal antibodies withspecificity for (a) human PDC-E2 (clone 4C8, 1:100dilution) and (b) against all three previously identifiedmitochondrial autoantigens (clone 2H4, 1:1000 dilution)[17,18] were used as the primary antibody probes. Wealso used rabbit anti-lipoic acid antiserum (1:20,000dilution) [19], and serum from patients with PBC (1:1000dilution) to further characterize the four recombinantNovo proteins. Negative control sera were used through-out the study. Additional controls included 1:10,000diluted mouse monoclonal antibodies detecting Tetra-His residues (for Novo1, 2 and 3 6!His fusion proteins)(Qiagen) and anti-Flag tag (for Novo4 Flag fusionprotein) (Stratagene). HRP-conjugated goat anti-mouse,anti-rabbit or anti-human IgG (Zymed LaboratoriesInc.) at a 10�4 dilution were used as secondary anti-bodies. The PBC serum had previously been character-ized in our laboratory and shown to have high titerreactivity against recombinant PDC-E2, OGDC-E2, andBCOADC-E2 by ELISA and Western blot assays [13].
3. Results
3.1. Nucleotide sequence
Using the PCR conditions described earlier, we clonedfour genes from N. aromaticivorans genomic DNA.Multiple clones of each construct were sequenced in both
directions to assure nucleotide accuracy. The resultingcontigs were assembled into a reference sequence usingVector NTI software and translated into the correspond-ing primary amino acid sequence. BLAST searches of thenucleotide sequences indicated that all four cloned Novogenes belong to the family of 2-OADC. Nucleotidesequence analysis of Novo1 and Novo2 contigs showedseveral base changes from the published sequences (datanot shown). However, their deduced primary amino acidsequences differed only in a single amino acid sub-stitution within the coding sequence (Fig. 1), whichresulted in O99% homology to N. aromaticivoranspublished sequences (GenBank accession numbersZP_00095480 (Novo1), ZP_00094526 (Novo2)). BothNovo3 and Novo4 cloned genes were identical to thepublished bacterial sequence (GenBank accession num-bers ZP_00095006 and ZP_00095507, respectively).
3.2. Protein sequence and lipoylated domains
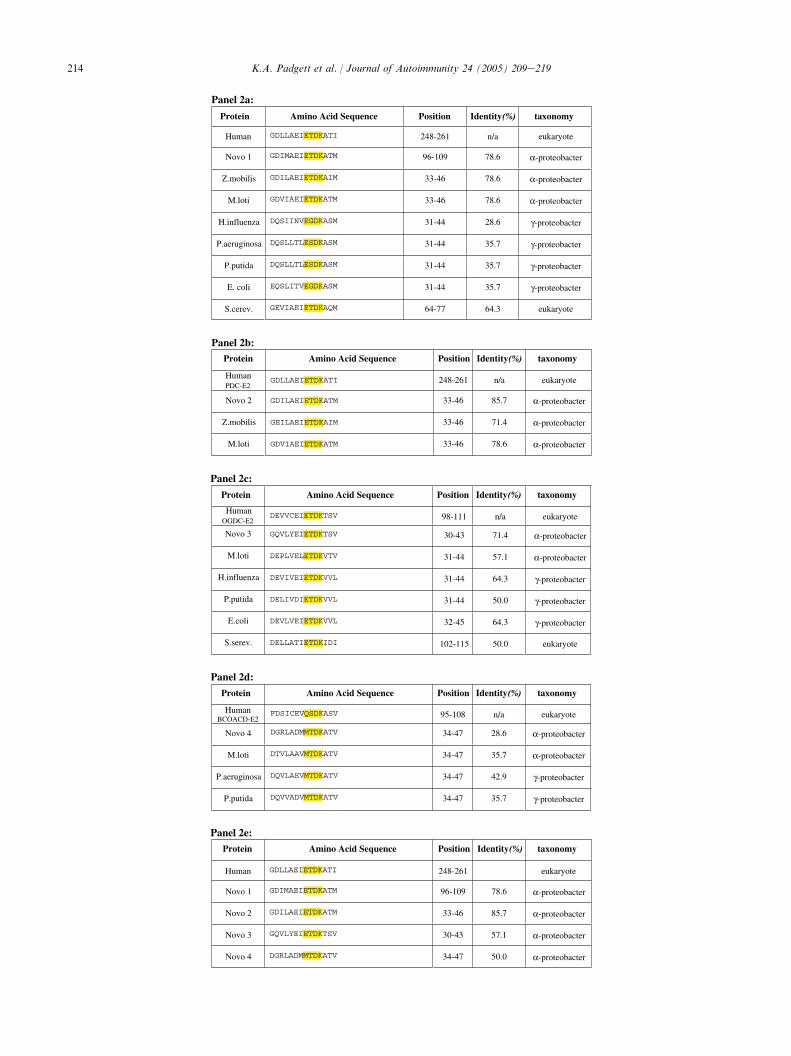
When the complete deduced amino acid sequence ofthe cloned genes was used in BLAST queries, Novo1and Novo2 were shown to be homologs of PDC-E2 andE1beta, respectively, from prokaryotic and eukaryoticorganisms. Novo3 and Novo4 on the other hand wereshown to be homologs of OGDC-E2 and BCOADC-E2,respectively. A further search within the NCBI con-served domain database revealed that all four clonedNovo proteins contained a single lipoylated domainlocated at the amino terminal portion of the sequence.These motifs (highlighted in Fig. 1) are similar to thoseof the 2-OADC complexes found in other prokaryoticand eukaryotic organisms. Comparison of the aminoacid residues within the lipoylated domains of the clonedN. aromaticivorans proteins with their prokaryotic andeukaryotic counterparts showed variable sequencehomology (Fig. 2). Maximum sequence identities werefound to exist between the lipoylated domains of Novo1and Novo2 (79% and 86%, respectively) and humanPDC-E2 (Fig. 2a,b). Although BLAST searches usingthe complete amino acid sequence of Novo2 showedsignificant homology with the E1beta subunit of PDC,such homology was limited when only the lipoylateddomain was considered. This can be attributed mostlikely to the fact that the human PDC-E1beta does notcontain a canonical lipoylated motif. With 71% aminoacid identity between the domain of Novo3 and humanOGDC-E2, Novo3 was closer in homology to thehuman peptide than its prokaryotic counterparts(Fig. 2c). Interestingly, the lipoylated domain of Novo4,with 50% of the residues identical to human PDC-E2,was found to be more homologous to the inner domainof human PDC-E2 (Fig. 2e) than to the humanBCOADC-E2 with which it had only 29% sequenceidentity (Fig. 2d). However, when we compared Novo4to the BCOADC-E2 lipoylated domain of other

213K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
prokaryotes, we found it to be very similar to both alphaand gamma proteobacteria (Fig. 2d).
3.3. Phylogenetic trees
Fig. 3 (panels 1e4) illustrates the phylogenetic treesobtained for the four Novo proteins. The dendrogramsare shown and the branching pattern represents theevolutionary path of the homologous proteins fromdifferent species. The length of the branches connectingterminal nodes indicates the estimated phylogeneticdistance between homologs from two organisms.
Novo1
MRCDMAICPSRSTGIGKFVCGASGNPLDPTRHAGEMAPRKDAGEVSATAPSSLRYQDTEN
TPMPIAIKMPALSPTMEEGTLAKWLVKVGDKVSSGDIMAEIETDKATMEFEAVDEGTIVS
IDVAEGSEGVKVGTVIATLAGEDEDASAPAPKAVAPAAAPVPVPAPKAEPAPAAVSTPAP
AAASASKGDRVIATPLAKRIAADKGIDLKGVAGSGPNGRIIRADVEGAKPAAAAPVSTVA
PAVASAAAPARAPAAVPDFGIPYEAQKLNNVRKTIARRLTEAKQTIPHIYLTVDIRLDAL
LKLRGDLNKALEAQGVKLSVNDLIIKALAKALMQVPKCNVSFAGDELRSFKRADISVAVA
APSGLITPIIVDAGSKSVSAIATEMKALANKAREGKLQPHEYQGGTASLSNLGMFSIKNF
DAVINPPQAMIMAVGAGEQRPYVIDGALGIATVMSATGSFDHRAIDGADGAELMQAFKNL
IENPLGLVA
Novo2
MAIELKMPALSPTMEEGTLAKWLVKAGDEVRSGDILAEIETDKATMEFEAVDEGVIAEIL
VAEGTEGVKVGTVIATIQGEGEDAAPAAATPAVEQKVEMSEAAPSVEARAAPAVAIAPKV
DAKPAVDPEIPAGTAMVPTTVREALRDAMAEEMRADDRVFVMGEEVAEYQGAYKVTQGLLDEFGPR
RVIDTPITEYGFVGIGAGAAMGGLRPIIEFMTFNFAMQAIDHIINSAAKTNYMS
GGQMRCPIVFRGPNGAASRVGAQHSQNYGPWYANVPGLVVIAPYDSADAKGLMKAAIRSEDPVVFL
ENELVYGRTFDVPQMDDFVLPIGKARIVRQGKDVTIVSYSIGVGLALEAAETLA
AEGIDAEVIDLRTLRPLDKDTVLASLAKTNRLVVAEEGFPVCSIASEIMAICMEDGFDHL
DAPVLRVCDEDVPLPYAANLEKAALIDAGKIAAAVRKVCYR
Novo3
VLNELRIPRMGSVENARLLNWRVQEGEAYEPGQVLYEIETDKTSVEVEAEGPGVLARHLA
AVGDEFKVGDRIGLWALPGTAPATLRAALSPQPMPASEPAPSPSSTLPAAVSAPGLHALR
PVSRDAAGGRRVSPLARRLAAQNGVDLATVTGTGMGGKISGKDVLAASAKPRPAPVPVSP
PRPGSDGEIVPHSLRRRTIAQRMVEAAAIPTLTADMEVDLTALFARRRSVEGNGASVLGM
IAEAAIAALLQHRRLNAHWREDAMVQFGAVHLGIAVDTPEGLVVPVVRNAESLNARGLTDAIAALA
DKARAGTLRPQDMEGGTFTISNPGSMGPVVRAEALLNPPQVALLGLPGIVRAPV
AIKDGDAWAMAVRPLLRLSLSFDHRALDGGPVIAFLNTLKATLERP
Novo4
MGTYTFRLPDIGEGIAEAEIVAWHVKVGDTVEEDGRLADMMTDKATVEMESPVAGKVVSVAGEVGD
VVAIGSALVVIETEGEDEAPAPAAAPAPKAAIVEERIEVETPEPPQPPSPPQPL
FVSREVEAPPAVPATGSGVAPGPRASTAPDTIGGAGAKVLASPAVRQRARDLGIDLSEVR
PSEEGRIRHADLDQFLSYNASGGYRAAGAERGDEVIRVIGMRRRIAENMAASKRHIPHFS
YVEECDVTALEIMREQLNAGRGDKPKLTMLPLLITAICRALPQYPMINARYDDEAGVVTR
YGAVHLGMAAQTPAGLMVPVIRNAQTLNLWQLAREIVRLAEAARSGSAKSDELSGSTLTV
TSLGPLGGVATTPVINRPEVAIIGPNRIVERPMFVSDGMGGERIEKRKLMNISISCDHRV
VDGHDAASFIQAVKKLIETPVLLLAD
Fig. 1. Deduced amino acid sequence of cloned Novo 1 through 4
proteins. Putative lipoylated domains are highlighted. Changes in the
deduced amino sequence compared to the published sequence are
underlined.
Results showed that, despite the highly conserved natureof mitochondrial proteins, sequences from Homo sapiensand N. aromaticivorans presented an early phylogeneticseparation during development. However, Novo2 wasshown to be phylogenetically closer to human PDC-E1beta than to beta or gamma proteobacteria, or thebacterium C. pneumoniae.
3.4. Reactivity with specific antibodies and sera
Fig. 4 shows the reactivity patterns observed with thedifferent antibodies using the four recombinant Novoproteins. All recombinant proteins were recognized bythe anti-lipoic acid polyclonal antisera (Fig. 4, lane 1)and by the 2H4 monoclonal antibody (Fig. 4, lane 2).Only Novo1 and Novo2 reacted with the PDC-E2-specific monoclonal antibody 4C8 (Fig. 4, lane 3). PBCserum recognized all four proteins (Fig. 4, lane 4).
4. Discussion
Molecular mimicry by infectious agents has beenproposed as the mechanism responsible for breakingtolerance in a number of autoimmune diseases, in-cluding PBC [5]. We have previously suggested thata recently defined Gram-negative bacterium, N. aroma-ticivorans, is a better candidate for triggering sucha putative mechanism in PBC than other microbialstrains previously proposed [14]. We based this hypoth-esis on the fact that 100% of AMA-positive and a smallnumber of AMA-negative PBC sera showed a signifi-cantly higher reactivity to proteins from N. aromatici-vorans compared to E. coli. This thesis was supported bythe finding that N. aromaticivorans is naturally found inhuman fecal samples [9] and that similar observationswere reported in patients from other geographical areas[15]. We have now cloned and expressed four lipoylatedproteins from N. aromaticivorans and arbitrarily namedthem Novo 1 through 4. We compared the nucleotideand deduced amino acid sequences of such proteins withthose of other species to better define the degree ofhomology between them and 2-OADC proteins fromother organisms. In addition, we investigated thephylogenetic relationship between homologs from hu-man, N. aromaticivorans, and other bacterial species,including those previously suggested in the etiopatho-genesis of PBC.
The 2-OADC family is composed of evolutionarilyconserved multienzyme complexes, including PDC,OGDC, and BCOADC, that are located on the innermembrane of mitochondria. The three multienzymecomplexes share a common architecture being com-posed of multiple copies of 2-oxo acid dehydrogenase(E1 subunit), dihydrolipoamide acyltransferase (E2subunit), and dihydrolipoamide dehydrogenase (E3
214 K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
Panel 2a:
Protein Amino Acid Sequence Position taxonomy
Protein Amino Acid Sequence Position taxonomy
Protein Amino Acid Sequence Position taxonomy
Protein Amino Acid Sequence Position taxonomy
Protein Amino Acid Sequence Position taxonomy
Human GDLLAEIETDKATI 248-261 n/a eukaryote
Novo 1 GDIMAEIETDKATM 96-109 78.6 α-proteobacter
Z.mobilis GDILAEIETDKAIM 33-46 78.6 α-proteobacter
M.loti GDVIAEIETDKATM 33-46 78.6 α-proteobacter
H.influenza DQSIINVEGDKASM 31-44 28.6 γ-proteobacter
P.aeruginosa DQSLLTLESDKASM 31-44 35.7 γ-proteobacter
P.putida DQSLLTLESDKASM 31-44 35.7 γ-proteobacter
E. coli EQSLITVEGDKASM 31-44 35.7 γ-proteobacter
S.cerev. GEVIAEIETDKAQM 64-77 64.3 eukaryote
Panel 2b:
Panel 2c:
Panel 2d:
Panel 2e:
HumanPDC-E2
GDLLAEIETDKATI 248-261 n/a eukaryote
Novo 2 GDILAEIETDKATM 33-46 85.7 α-proteobacter
Z.mobilis GEILAEIETDKAIM 33-46 71.4 α-proteobacter
M.loti GDVIAEIETDKATM 33-46 78.6 α-proteobacter
HumanOGDC-E2
DEVVCEIETDKTSV 98-111 n/a eukaryote
Novo 3 GQVLYEIETDKTSV 30-43 71.4 α-proteobacter
M.loti DEPLVELETDKVTV 31-44 57.1 α-proteobacter
H.influenza DEVIVEIETDKVVL 31-44 64.3 γ-proteobacter
P.putida DELIVDIETDKVVL 31-44 50.0 γ-proteobacter
E.coli DEVLVEIETDKVVL 32-45 64.3 γ-proteobacter
S.serev. DELLATIETDKIDI 102-115 50.0 eukaryote
Human BCOACD-E2
FDSICEVQSDKASV 95-108 n/a eukaryote
Novo 4 DGRLADMMTDKATV 34-47 28.6 α-proteobacter
M.loti DTVLAAVMTDKATV 34-47 35.7 α-proteobacter
P.aeruginosa DQVLAEVMTDKATV 34-47 42.9 γ-proteobacter
P.putida DQVVADVMTDKATV 34-47 35.7 γ-proteobacter
Human GDLLAEIETDKATI 248-261 eukaryote
Novo 1 GDIMAEIETDKATM 96-109 78.6 α-proteobacter
Novo 2 GDILAEIETDKATM 33-46 85.7 α-proteobacter
Novo 3 GQVLYEIETDKTSV 30-43 57.1 α-proteobacter
Novo 4 DGRLADMMTDKATV 34-47 50.0 α-proteobacter

215K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
subunit). The E2 subunits carry lipoic acid, an essentialcofactor, which is covalently bound to the lysine residuewithin the lipoylated domain of the protein. While PDC-E2 presents either one, two or three lipoylated domains(inner and outer) in different species, OGDC-E2 andBCOACD-E2 feature only a single domain. Our grouphas recently reported the important role of lipoic acid indetermining the immunogenicity of mitochondrial anti-gens in PBC [16], and we have also hypothesized thatmodifications of this cofactor are involved in thebreakdown of tolerance in experimental animals [17].
The lipoylated domains from Novo1 (Fig. 2a) andNovo2 (Fig. 2b) present the highest homology withhuman PDC-E2 (in the case of Novo1 the same degreeof homology is also shared by Zymomonas mobilis andMesorhizobium loti). Similarly, the lipoylated domainfrom Novo3 (Fig. 2c) has the highest homology with thehuman OGDC-E2 lipoylated domain. We note thatNovo4, on the other hand, did not show the same degreeof similarity with the human BCOADC-E2 domain(Fig. 2d). Based on these observations, we performed animmunological study using well-established and highlyspecific monoclonal and polyclonal antibodies. Ourfindings demonstrate that all four Novo proteins arelipoylated, as indicated by the strong reactivity with therabbit anti-lipoic acid polyclonal antisera (Fig. 4).Moreover, all four recombinant Novo proteins wererecognized by our 2H4 monoclonal antibody, which hadpreviously been shown to react with all three majormitochondrial antigens (PDC-E2, OGDC-E2, andBCOADC-E2) (Fig. 4). Both Novo1 and Novo2 werein turn recognized by our 4C8 clone, which is specific tohuman PDC-E2.
The homology of three of the four bacterial proteinsis the highest ever reported for the immunodominantlipoylated epitopes in PBC. It could be argued that ourdata are explained by the highly conserved nature ofmitochondrial proteins across species. However, ourphylogenetic study indicates that the bacterial proteinsdescribed herein present a significant phylogeneticdistance from their human homologs, and are in somecases even more divergent than those from otherbacterial species previously studied in PBC (i.e. E. coli),which also showed significantly lower cross-reactivity.
There has been significant work showing a lack ofcorrelation between PBC disease activity and the titer ofAMA [18,19]. These observations have led some tosuggest that the AMA response may be an epiphenom-enon. However, we are reminded that, with the
exception of high-titer antibodies to dsDNA and theirassociation with lupus nephritis [20], there is a lack ofcorrelation between other antibody-mediated diseasesand their respective autoantibody titers; reflected instudies of myasthenia gravis [21], type 1 diabetes [22],and autoimmune thyroiditis [23]. On the other hand, weare more persuaded by the correlation of the autoim-mune response to mitochondrial antigens in PBC basedon the dramatic increase in the frequency of autoantigenspecific CD4C and CD8C cells localized within the liverof patients compared to blood [24]. Hence, we believethat the identification of the original immunogenicmolecule that breaks tolerance to mitochondrial anti-gens is crucial to dissecting the pathogenesis of thedisease.
This discussion on the role of N. aromaticivorans inPBC takes on added significance when the uniquebiological features of this organism are considered. Infact, we submit that the development of PBC not onlyrequires a genetically susceptible host, but also at leasttwo and/or multiple and perhaps simultaneous immu-nological hits that lead to the induction of AMA. Wefurther submit that the induction of AMA is anextraordinarily reliable indicator of the presence orfuture development of overt disease. We base thisassertion on the fact that AMA are absent, by diagnostictests using defined recombinant antigens, in all peopleexcept patients with PBC or those who will develop PBCin the future [25,26]. This degree of specificity impliesthat following the break of tolerance there must be a bileduct-specific injury that promotes the onset of disease.It also suggests that the development of PBC, thevariability in disease severity between patients, andthe AMAePBC latency time will therefore be based onthe break of tolerance and hence a liver specific event.We should also note for completeness that it has beensuggested that PBC is caused by a retrovirus [27] whileour laboratory, using extensive panels of sera and liversamples, have failed to recapitulate the hypothesis [28]while the blast search performed for this study alsoincluded known viral sequences.
The fact that N. aromaticivorans is a normal com-mensal and found in a significant number of otherwisehealthy people [9] implies that the bacterium alone isinsufficient in causing the disease. The ability of thisorganism to ingest and metabolize chemicals [29] couldtherefore facilitate the modification of lipoic acid, whichin turn could lead to the development of AMA. In fact,the presence of N. aromaticivorans in mucosal secretions
Fig. 2. Sequence comparison of lipoylated domains of the human 2-OADC enzymes with those from other organisms. The canonical lipoyl motif
EXDK is highlighted. (a) Amino acid sequence comparison of the human PDC-E2 lipoyl domain with other microorganisms. (b) Amino acid
sequence comparison of the human PDC-E2 lipoyl domain with the PDC-E1b lipoyl domain of alpha proteobacteria. (c) Amino acid sequence
comparison of the human OGDC-E2 lipoyl domain with other microorganisms. (d) Amino acid sequence comparison of the human BCOACD-E2
lipoyl domain with other microorganisms. (e) Amino acid sequence comparison of the human PDC-E2 inner lipoyl domain with the single lipoyl
domain of Novo clones 1 through 4.

216 K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
Fig. 3. Phylogenetic trees comparing Novo proteins with human and other organisms. The 2-OADC protein homologs were identified using BLAST.
Corresponding protein sequences from bacteria and eukaryotes were obtained from GenBank and aligned using the default parameters in the
ClustalW alignment software. The trees were constructed from the aligned sequences using the Phylodendron software, version 0.8d. The scale bar
represents 0.1 substitutions per site.
might even suggest that the breakdown of tolerance doesnot have to originate in the liver. Once tolerance isbroken, the specific characteristics of bile duct cellswould then become the target in the biliary epithelium-specific pathology. However, the biliary cell may bemore than an innocent victim and may even contributeto its own demise via the poly-immunoglobulin receptor,the survival of the inner lipoylated domain of themitochondrial autoantigens in apoptotic cells, and thecross-priming of mitochondrial immune complexes andCD4C and CD8C cells [30]. Further, we note thatno evidence is currently available on the tropism ofN. aromaticivorans toward human cell types, includingbiliary epithelia.
In conclusion, our theory on the etiopathogenesisof PBC (Fig. 5) indicates that the ubiquitous bacterium
N. aromaticivorans enters the human system, most likelythrough the digestive mucosa, which is supported by ourprevious findings [9]. The lipoylated bacterial proteinsare then modified by the unique xenobiotic metabolizingcapability of the bacterium to form immunoreactiveprotein adducts, similar to those previously suggested[31]. This modification is sufficient to trigger the innateimmune system to initiate a cascade of local inflamma-tory events, via toll-like receptors for example, whichresults in local dendritic cell activation and processing.The modified Novo proteins are taken up and processedby mucosal antigen-presenting cells (APC), which inturn activate autoreactive T and B cells that can reachthe liver through the portal system. T cells participatedirectly in the autoimmune injury and/or further recruitautoreactive lymphocytes. B cells, on the other hand,

217K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
Fig. 4. Western blot analysis of recombinant Novo proteins. Novo 1 through 4 were fractionated by PAGE, transferred to nitrocellulose, and probed
with anti-lipoic acid rabbit polyclonal antibody (lane 1), mouse monoclonal antibodies reacting with human PDC-E2, BCOADC-E2 and OGDC-E2
(clone 2H4, lane 2), and human PDC-E2 only (clone 4C8, lane 3). The AMA-positive serum from a patient with PBC is shown in lane 4.
secrete AMA, particularly IgA which is transported tothe vascular side and then into the lumen of the biliaryepithelial cells (BECs) by a process known as trans-cytosis. IgA then reacts with the PDC-E2-like molecules
located on the luminal surface of BECs [32] and caninitiate the apoptotic signaling cascade. Based on itsunique features in PBC [33] and the intact PDC-E2resulting from caspase digestion [34], apoptosis of BECs
Fig. 5. Representation of the proposed theory on the etiopathogenesis of PBC based on the suggested role of N. aromaticivorans in combination with
individual susceptibility and environmental factors. The bacterium enters the mucosal system and its lipoylated proteins (Novo) are modified by
xenobiotics thus creating immunoreactive molecules. Modified proteins are then processed by antigen-presenting cells (APC) in the mucosa, thus
activating autoreactive T and B cells. While T cells exert a cytopathic effect directly and/or recruit additional lymphocytes, B cells secrete AMA of the
IgG and IgA isotype. IgA enter the lumen of biliary epithelial cells (BECs) by transcytosis and then react with the PDC-E2-like molecules thus
initiating apoptosis. The intact PDC-E2 liberated from apoptotic cells forms immune complexes with circulating AMA-IgG. Such immune
complexes, the enhanced apoptosis, and the direct cytopathic effect of T cells and possibly AMA eventually lead to the appearance of the histological
signs of PBC, i.e. chronic non-suppurative cholangitis.

218 K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
ultimately contributes to the liver injury. In the samescenario, the immune complexes of intact post-apoptoticPDC-E2 and IgG-AMA as well as the direct cytopathiceffects of autoreactive T cells and possibly AMA alsocontribute to the appearance of the histological lesionstypical of PBC, i.e. chronic non-suppurative cholangitis.Understanding both the inductive and destructivemechanisms leading to the onset and perpetuation ofPBC will be the only way to identify future targets forthe development of new treatments.
References
[1] Talwalkar JA, Lindor KD. Primary biliary cirrhosis. Lancet
2003;362:53e61.
[2] Gershwin ME, Ansari AA, Mackay IR, Nakanuma Y, Nishio A,
Rowley MJ et al. Primary biliary cirrhosis: an orchestrated
immune response against epithelial cells. Immunol Rev
2000;174:210e25.
[3] Selmi C, Mayo MJ, Bach N, Ishibashi H, Invernizzi P, Gish RG
et al. Primary biliary cirrhosis in monozygotic and dizygotic
twins: genetics, epigenetics, and environment. Gastroenterology
2004;127:485e92.
[4] Leung PS, Quan C, Park O, Van de Water J, Kurth MJ,
Nantz MH et al. Immunization with a xenobiotic 6-bromohex-
anoate bovine serum albumin conjugate induces antimitochon-
drial antibodies. J Immunol 2003;170:5326e32.
[5] Van de Water J, Ishibashi H, Coppel RL, Gershwin ME.
Molecular mimicry and primary biliary cirrhosis: premises not
promises. Hepatology 2001;33:771e5.
[6] Takeuchi M, Hamana K, Hiraishi A. Proposal of the genus
Sphingomonas sensu stricto and three new genera. Sphingobium,
Novosphingobium and Sphingopyxis, on the basis of phyloge-
netic and chemotaxonomic analyses. Int J Syst Evol Microbiol
2001;51:1405e17.
[7] Balkwill DL, Drake GR, Reeves RH, Fredrickson JK, White DC,
Ringelberg DB et al. Taxonomic study of aromatic-degrading
bacteria from deep-terrestrial-subsurface sediments and descrip-
tion of Sphingomonas aromaticivorans sp. nov., Sphingomonas
subterranea sp. nov., Sphingomonas stygia sp. nov. Int J Syst
Bacteriol 1997;47:191e201.
[8] Fujii K, Kikuchi S, Satomi M, Ushio-Sata N, Morita N.
Degradation of 17beta-estradiol by a gram-negative bacterium
isolated from activated sludge in a sewage treatment plant in
Tokyo. Japan. Appl Environ Microbiol 2002;68:2057e60.
[9] Selmi C, Balkwill DL, Invernizzi P, Ansari AA, Coppel RL,
Podda M et al. Patients with primary biliary cirrhosis react
against a ubiquitous xenobiotic-metabolizing bacterium. Hepa-
tology 2003;38:1250e7.
[10] Fussey SP, Ali ST, Guest JR, James OF, Bassendine MF,
Yeaman SJ. Reactivity of primary biliary cirrhosis sera with
Escherichia coli dihydrolipoamide acetyltransferase (E2p): char-
acterization of the main immunogenic region. Proc Natl Acad Sci
USA 1990;87:3987e91.
[11] Abdulkarim AS, Petrovic LM, Kim WR, Angulo P, Lloyd RV,
Lindor KD. Primary biliary cirrhosis: an infectious disease caused
by Chlamydia pneumoniae? J Hepatol 2004;40:380e4.
[12] Muratori P, Muratori L, Guidi M, Maccariello S, Pappas G,
Ferrari R et al. Anti-Saccharomyces cerevisiae antibodies (ASCA)
and autoimmune liver diseases. Clin Exp Immunol 2003;132:
473e476.
[13] Miyakawa H, Tanaka A, Kikuchi K, Matsushita M, Kitazawa E,
Kawaguchi N et al. Detection of antimitochondrial autoantibodies
in immunofluorescent AMA-negative patients with primary
biliary cirrhosis using recombinant autoantigens. Hepatology
2001;34:243e8.
[14] Selmi C, Gershwin ME. Bacteria and human autoimmunity: the
case of primary biliary cirrhosis. Curr Opin Rheumatol
2004;16:406e10.
[15] Olafsson S, Gudjonsson H, Selmi C, Amano K, Invernizzi P,
Podda M et al. Antimitochondrial antibodies and reactivity to N.
aromaticivorans proteins in Icelandic patients with primary
biliary cirrhosis and their relatives. Am J Gastroenterol 2004;99:
2143e6.
[16] Bruggraber SF, Leung PS, Amano K, Quan C, Kurth MJ,
Nantz MH et al. Autoreactivity to lipoate and a conjugated form
of lipoate in primary biliary cirrhosis. Gastroenterology
2003;125:1705e13.[17] Palmer JM, Robe AJ, Burt AD, Kirby JA, Jones DE. Covalent
modification as a mechanism for the breakdown of immune
tolerance to pyruvate dehydrogenase complex in the mouse.
Hepatology 2004;39:1583e92.
[18] Van Norstrand MD, Malinchoc M, Lindor KD, Therneau TM,
Gershwin ME, Leung PS et al. Quantitative measurement of
autoantibodies to recombinant mitochondrial antigens in patients
with primary biliary cirrhosis: relationship of levels of autoanti-
bodies to disease progression. Hepatology 1997;25:6e11.
[19] Vleggaar FP, van Buuren HR. No prognostic significance of
antimitochondrial antibody profile testing in primary biliary
cirrhosis. Hepatogastroenterology 2004;51:937e40.[20] Vitali C, Bencivelli W, Isenberg DA, Smolen JS, Snaith ML,
Sciuto M et al. Disease activity in systemic lupus erythematosus:
report of the Consensus Study Group of the European Workshop
for Rheumatology Research. II. Identification of the variables
indicative of disease activity and their use in the development of
an activity score. The European Consensus Study Group for
Disease Activity in SLE. Clin Exp Rheumatol 1992;10:541e7.[21] Souan ML, Geffard M, Vieillemaringe J, Lebrun-Grandie P,
Orgogozo JM. Anti-acetylcholine antibodies and the pathogenesis
of myasthenia gravis. Ann N Y Acad Sci 1987;505:423e38.
[22] Falorni A, Calcinaro F. Humoral responses in type 1 diabetes
mellitus. Rev Endocr Metab Disord 2003;4:281e90.
[23] Orgiazzi J. Anti-TSH receptor antibodies in clinical practice.
Endocrinol Metab Clin North Am 2000;29:339e355, vii.[24] Kita H, Imawari M, Gershwin ME. Cellular immune response in
primary biliary cirrhosis. Hepatol Res 2004;28:12e7.
[25] Metcalf JV, Mitchison HC, Palmer JM, Jones DE,
Bassendine MF, James OF. Natural history of early primary
biliary cirrhosis. Lancet 1996;348:1399e402.
[26] Mattalia A, Quaranta S, Leung PS, Bauducci M, Van de Water J,
Calvo PL et al. Characterization of antimitochondrial antibodies
in health adults. Hepatology 1998;27:656e61.[27] Xu L, Shen Z, Guo L, Fodera B, Keogh A, Joplin R et al. Does
a betaretrovirus infection trigger primary biliary cirrhosis? Proc
Natl Acad Sci USA 2003;100:8454e9.
[28] Selmi C, Ross SR, Ansari AA, Invernizzi P, Podda M, Coppel RL
et al. Lack of immunological or molecular evidence for a role of
mouse mammary tumor retrovirus in primary biliary cirrhosis.
Gastroenterology 2004;127:493e501.[29] Shi T, Fredrickson JK, Balkwill DL. Biodegradation of polycyclic
aromatic hydrocarbons by Sphingomonas strains isolated from
the terrestrial subsurface. J Ind Microbiol Biotechnol
2001;26:283e9.[30] Nishio A, Keeffe EB, Gershwin ME. Immunopathogenesis of
primary biliary cirrhosis. Semin Liver Dis 2002;22:291e302.
[31] Long SA, Quan C, Van de Water J, Nantz MH, Kurth MJ,
Barsky D et al. Immunoreactivity of organic mimeotopes of
the E2 component of pyruvate dehydrogenase: connecting
xenobiotics with primary biliary cirrhosis. J Immunol 2001;167:
2956e63.

219K.A. Padgett et al. / Journal of Autoimmunity 24 (2005) 209e219
[32] Fukushima N, Nalbandian G, Van de Water J, White K,
Ansari AA, Leung P et al. Characterization of recombinant
monoclonal IgA anti-PDC-E2 autoantibodies derived from
patients with PBC. Hepatology 2002;36:1383e92.
[33] Odin JA, Huebert RC, Casciola-Rosen L, LaRusso NF, Rosen A.
Bcl-2-dependent oxidation of pyruvate dehydrogenase-E2, a
primary biliary cirrhosis autoantigen, during apoptosis. J Clin
Invest 2001;108:223e32.
[34] Matsumura S, Van de Water J, Kita H, Coppel RL, Tsuji T,
Yamamoto K et al. Contribution to antimitochondrial antibody
production: cleavage of pyruvate dehydrogenase complex-E2 by
apoptosis-related proteases. Hepatology 2002;35:14e22.
Copyright © 2022 FDOKUMEN




















