
Periovulatory changes in female sexual behavior and patterns of ovarian steroid secretion in...
20
HORMONES AND BEHAVIOR 18, 431-450 (1984) Periovulatory Changes in Female Sexual Behavior and Patterns of Ovarian Steroid Secretion in Group-Living Rhesus Monkeys KIM WALLEN, LINDA A. WINSTON, SUZANNE CAVENTA, MARYANN DAVIS-DASILVA, AND DELWOOD C. COLLINS” Deparment of Psychology, Ye&es Regional Primate Research Cerlter, atrd “Department of Medicine, Ernoty University, and VA Medicai Center, Atlanta. Georgia 3032 The behavior of nine intact group-living adult female rhesus was observed for 30 min daily with each of four adult male rhesus across a verified ovulatory menstrual cycle. Blood samples collected from females daily oe on alternate days were analyzed for estradiol, testosterone, and progesterone. Female patterns of approach, follow, and initiate proximity increased several days prior to the estradiol peak, peaked on the day of the estradiol peak, then declined completely or to very iow frequencies. Mounts, intromissions. and ejaculations increased significantly on the day of the estradiol peak, remained elevated for 2 more days. then declined completely by the fifth day after peak estradiol. Ejaculations never occurred outside of a IO-day period starting 4 days before the estradiol peak and ending 5 days after the estradiol peak. During this period females initiated over 90% of all approaches. Female hand slap, threaten away. and stand up increased significantly on the first day of increased copulation, remained elevated while copulation was significantly elevated, then decreased along with the decline in copulation. Ten of eleven patterns of female behavior correlated signiticantly with estradiol level prior to the estradiol peak. All were significantly inversely correlated with progesterone level after the estradiol peak. No pattern of female behavior correlated significantly with testosterone either before or after the estradiol peak. Similarly, male patterns of behavior correlated with femaie levels of estradiol and progesterone, but not testosterone. These results demonstrate a relationship between increased serum estradiol and increased female initiation of sexual behavior. The finding that some patterns of female behavior increase several days prior to copulation, whereas other behaviors increase coincident with increased copulation suggests that the behavior of group-living rhesus females serves two functions. The first is to communicate sexual interest and the second is to maintain the consort pair and increase the probability that ejaculation will occur. In addition. the strong correlation between preovulatory female behavior and estradiol level suggests that the female’s behavior provides precise information about her re- productive state and could thus coordinate copulation with maximal fertiiity. Q 1984 Academic Press. Inc. Sexual behavior of female rhesus monkeys in semi-free-ranging or captive groups consists of discrete 8- to G-day periods of intense sexnai 431 OOiS-jO6Xi84 51.50 Copyright @J 1984 by Academic Press. Inc. .a1 rights of reproduction in any Eons reserved.
Transcript of Periovulatory changes in female sexual behavior and patterns of ovarian steroid secretion in...

HORMONES AND BEHAVIOR 18, 431-450 (1984)
Periovulatory Changes in Female Sexual Behavior and Patterns of Ovarian Steroid Secretion in Group-Living
Rhesus Monkeys
KIM WALLEN, LINDA A. WINSTON, SUZANNE CAVENTA, MARYANN DAVIS-DASILVA, AND DELWOOD C. COLLINS”
Deparment of Psychology, Ye&es Regional Primate Research Cerlter, atrd “Department of Medicine, Ernoty University, and VA Medicai Center,
Atlanta. Georgia 3032
The behavior of nine intact group-living adult female rhesus was observed for 30 min daily with each of four adult male rhesus across a verified ovulatory menstrual cycle. Blood samples collected from females daily oe on alternate days were analyzed for estradiol, testosterone, and progesterone. Female patterns of approach, follow, and initiate proximity increased several days prior to the estradiol peak, peaked on the day of the estradiol peak, then declined completely or to very iow frequencies. Mounts, intromissions. and ejaculations increased significantly on the day of the estradiol peak, remained elevated for 2 more days. then declined completely by the fifth day after peak estradiol. Ejaculations never occurred outside of a IO-day period starting 4 days before the estradiol peak and ending 5 days after the estradiol peak. During this period females initiated over 90% of all approaches. Female hand slap, threaten away. and stand up increased significantly on the first day of increased copulation, remained elevated while copulation was significantly elevated, then decreased along with the decline in copulation. Ten of eleven patterns of female behavior correlated signiticantly with estradiol level prior to the estradiol peak. All were significantly inversely correlated with progesterone level after the estradiol peak. No pattern of female behavior correlated significantly with testosterone either before or after the estradiol peak. Similarly, male patterns of behavior correlated with femaie levels of estradiol and progesterone, but not testosterone. These results demonstrate a relationship between increased serum estradiol and increased female initiation of sexual behavior. The finding that some patterns of female behavior increase several days prior to copulation, whereas other behaviors increase coincident with increased copulation suggests that the behavior of group-living rhesus females serves two functions. The first is to communicate sexual interest and the second is to maintain the consort pair and increase the probability that ejaculation will occur. In addition. the strong correlation between preovulatory female behavior and estradiol level suggests that the female’s behavior provides precise information about her re- productive state and could thus coordinate copulation with maximal fertiiity. Q 1984 Academic Press. Inc.
Sexual behavior of female rhesus monkeys in semi-free-ranging or captive groups consists of discrete 8- to G-day periods of intense sexnai
431 OOiS-jO6Xi84 51.50
Copyright @J 1984 by Academic Press. Inc. .a1 rights of reproduction in any Eons reserved.

432 WALLEN ET AL.
activity followed by complete sexual inactivity (Carpenter, 1942; Gordon, 1981; Wilson, Gordon, and Collins, 1982a). Sexual activity is initiated with increasing preovulatory estradiol and terminates when luteal pro- gesterone is above 2-3 rig/ml (Gordon, 1981; Wilson et al., 1982a). In addition to increased mounts, intromissions, and ejaculations by males, females display increased following, and initiation of proximity to males (Carpenter, 1942; Cochran, 1979). Increases in female behavior appear related to periovulatory hormonal changes (Cochran, 1979), but this pos- sibility has not been explicitly investigated.
The pattern of mating behavior seen in pairs of rhesus monkeys differs from that of multifemale groups (Wallen and Winston, 1984). Although a periovulatory peak in copulation is often seen in rhesus monkeys tested in pairs (Goy, 1979; Goy and Resko, 1972; Michael and Bonsall, 1979; Michael and Welegalla, 1968; Michael and Zumpe, 1970;), it sometimes is not (Johnson and Phoenix, 1978), and complete luteal cessation of mating, typical of multianimal groups (Gordon, 1981), has not been reported. Similarly, the relationship between ovarian steroids and female behavior in pair tests is unclear. Female initiation of proximity and solicitations peak at midcycle, in both freely interacting pairs (Czaja and Bielert, 1975; Goy and Resko, 1972) and when female behavioral change is max- imized by restricting male mobility (Czaja and Bielert, 1975; Pomerantz and Goy, 1983). However, in pair tests, female hormonal condition does not reliably influence female presentations (Michael and Welegalla, 1968; Michael, Zumpe, and Bonsall, 1982; Johnson and Phoenix, 1978). In addition, while ovariectomy reduces or completely eliminates female initiation of proximity and solicitations it does not consistently reduce female presentations (Johnson and Phoenix, 1976; Michael and Zumpe, 1970; Wallen and Goy, 1977). Thus pair-test studies suggest an ovarian steroid influence on rhesus sexual behavior. However, the influence appears to be highly variable and the extent to which female sexual behavior varies with hormonal state remains unclear.
The lack of a strong relationship between female behavior and ovarian steroids in pair tests has led to an emphasis upon steroid-induced changes in female attractivity (Herbert and Trimble, 1967; Beach, 1976) as an explanation for the midcycle peak in copulation (Michael and Keverne, 1968; Herbert, 1978; Keverne, 1976, 1982; Michael and Bonsall, 1979). In this view estrogens stimulate the production of vaginal secretions which increase female attractiveness stimulating male sexual initiation. Female behavior is not seen as critical in determining the pattern of rhesus sexual behavior. An alternative view is that the female’s hormonal state influences her initiation of sexual behavior (proceptivity: Beach, 1976) and that female behavioral changes communicate her motivation to copulate (Cochran, 1979; Czaja and Bielert, 1975; Goldfoot, 1981, 1982; Goy and Resko, 1972; Peretz, Goy, and Resko, 1971). Pair tests

OVARIAN STEROIDS IN RHESUS MONKEYS 433
provide supporting evidence for both views, and studies of groups, while demonstrating cyclic changes in female behavior, have not had the hormonal measurements which allow assessment of hormonal influences on female behavior (Carpenter, 1942; Cochran, 1979).
The recent finding that increases in the space and social complexity of the testing environment increase the extent to which a rhesus female’s hormonal state influences the occurrence of copulation (Wallen, 1982; Wallen and Winston, 1984) suggests that reliance upon testing rhesus pairs in small areas has produced information about hormonal influences on rhesus sexual behavior relevant only to that specific testing environment. The present study investigated changes in female behavior and ovarian steroids in a single-malelmultifemale social group. This type of group exhibits a pattern of copulatory behavior comparable to that seen in groups with multiple males (Wallen and Winston, 1984) and allows detailed capture of interactions between the male and all group females. We present evidence that the behavior of female rhesus, under these ob- servational conditions, is highly correlated with her hormonal state and could provide the male with sufficient information about her reproductive condition to coordinate copulation with maximal fertility.
MATERIALS AND METHODS
Subject. Nine adult female rhesus between 5 and 12 years of age and four adult vasectomized rhesus between 7 and 13 years of age served as subjects. Females were reared through adulthood in the same social group at the Yerkes Primate Center Field Station. Six months prior to behavioral observations females were removed from their natal group and housed in a 15.2 x 15.2-m compound with attached 3.1 x 4.9-m heated and ventilated indoor quarters. Three of the four males were reared in social groups at the Yerkes Primate Center Field Station. The fourth male, obtained from the Wisconsin Regional Primate Research Center at 12 years of age, was reared in a semi-free-ranging group at La Parguerra, Puerto Rico until 7 years of age. Females lived continuously in their compound while males were housed individually in a building separate from the females between testing periods. Both males and females had continuous access to monkey chow and water during behaviora. observations and received oranges daily following observations.
Social adaptation. Three months after the females were moved to the smaller compound each of the four males was introduced singly into the females’ home compound once per week for a 24-hr period of accli- matization. This continued until each male had spent 4 days with the females. For the month prior to data collection the introduction procedure for behavioral observations (see below) was followed 3 days per week. Thus prior to data collection males had 2 months of social experience with group females. No fighting was observed between males and females after the first week of introductions.

434 WALLEN ET AL.
Testing procedure. From October 25 until December 20, 1982 each of the four vasectomized males was individually introduced once daily into the females’ compound for a minimum of 30 min. Introduction order was rotated so each male received an equal number of first, second. third, and fourth introductions across a 28-day period, AU tests occurred between 830 and 1630 hr at ambient temperature and lighting. Several times per week males were observed for 30 min with a separate group of females at least 1 hr prior to their introduction to the study females.
Tests started with the release of a male from a visually isolated holding area into the females’ compound and terminated after 30 min unless the male obtained an intromission during the last 5 min of the test. When this occurred the test was continued until either the male ejaculated or a 5-min period passed without an intromission.
Behavioral data were recorded using an actor: behavior: recipient format entered into a Datamyte 900 electronic data recorder. The Datamyte automatically attached to each behavioral entry an elapsed time from the start of the test. Data were collected by two trained observers seated in a 5.5-m tower providing an unobstructed view of all areas of the compound, with one calling the behavior and the other entering the codes into the Datamyte. During an observation period all interactions between the male and any group female were recorded. Interactions scored between females were limited to grooming, agonistic behavior, and the occurrence of sitting within 25 cm of another female.
Behavioral dejnitions. A behavioral lexicon of 42 behavioral patterns was employed. The coding scheme captured both female distance from and orientation to the male within 1 m, and distance at 2 m or beyond. The present analysis is limited to the following behavioral patterns of which approach, hip touch, mount, intromission, ejaculation, noncontact present, and hand slap have been previously defined, or are referenced, in Wallen and Winston (1984). The following additional behaviors were recorded:
Follow. The actor proceeds, within 2 m, on exactly the same route as the animal being followed. Following was also scored when the two animals traveled beside each other if separated by 25 cm or less.
Back prox. Similar to back-to-partner (Wallen and Goy, 1977), this was scored when the female sat within 25 cm of the male oriented with her back toward the male and not more than 45” off an imaginary line perpendicular to his ventrum.
Stand up. Moving from any sitting position to a standing position when 2.5 cm or less separated the two animals.
Proximity. Sitting or standing within 25 cm of another animal. Duration does not include time spent grooming or being groomed.
SnifJ Movement of the male’s muzzle within 10 cm of a female’s perineal area sometimes while holding the female’s tail aside.

OVARIAN STEROIDS IN RHESUS MONKEYS 435
Tlzrear anq. Scored similarly to Altmann (1962), and Zumpe and Michael (1970). This behavior is similar to a threat except that it involves rapid alteration of gaze between the animals being threatened and the male, who is usually behind the female showing the behavior.
Gr-uom. Scored according to Altmann (1962). Blood coZL~fion. Females were checked daily for menstruation using
a moistened cotton swab inserted into the vagina, with the first day of bleeding being cycle Day 1. A 5-ml blood sample was cotiected after behavioral testing (between 1500 and 1700 hr) as previously described (Wallen and Winston, 1984), on cycle Days 3, 5, 7,9-20, and then every 2 days until menstruation. Blood was collected in evacuated serum sep- aration tubes (Vacutainer 651 I. B&D), clotted at ambient temperature. centrifuged, and serum stored at -20°C until assayed.
Radioimmunoassay, Serum samples were assayed. using previously validated procedures, for estradiol (E2; Wright. Collins, and Preedy , 1978), progesterone (P; Wilson, Gordon and Collins, 1982b), an tosterone (T; Perachio, Alexander, Marr, and Collins, 1977). Estradiol determinations used duplicate 200~~1 serum samples extracted in 2 ml of benzene. The E2 standard curve ranged from 2 to 80 pg with a sens~~~~~ty of 2.0 pg. The inter- and intraassay coefficients of variation (CV) were 10.2 and 6.3%. Radioimmunoassay of progesterone used duplicate 50-,tiLLi serum samples extracted in 2 ml of anesthesia grade ether. The P standard curve ranged from 10 to 400 pg with a sensitivity of LO.0 pg. The inter- and intraassay CV were 12.1 and 7.2%. Duplicate 50+1 serum samples were extracted with 2 ml of a 4 : 1 hexane : ether solvent for determ~~at~o~ of T values. The T standard curve ranged from 2 to 75 pg with a sensitivity of 2.0 pg. The intra- and interassay CV were 8.2 and 5.6%.
Menstrz4al cycle selection and alignment. Females started menstrual cycling at different times during the fall breeding season. This resuhe in three females displaying two complete menstrual cycles during the behavioral observation period and the remaining six displaying only one complete cycle. For those females displaying two verified ovulatory cycles. one was randomly selected for behavioral analysis. This resulted in two cases where the female’s second cycle was used for behavioral analysis.
ecause female cycles varied in length from 24 to 31 days (mean 26.7 + 2. A days), only days common to all cycles after alignment on the day of the E2 peak were used for behavioral comparisons. This alignment produced a 24day period starting 8 days prior to the E2 peak and co~tin~~~g through 15 days after the E2 peak.
Data arzalysis. Since test lengths varied all frequencies were converted to hourly rates. Duration measures were calculated for the first 30 mln of each test. Because the emphasis in this study was on changes in female behavior a mean score for each femak across all four males was calculated for each behavioral pattern.

436 WALLEN ET AL.
Changes in behavioral measures across the 24 common test days were analyzed using a one-way analysis of variance for repeated measures with total df equal to (24 - 1, days) x (9 - 1, subjects) = 184 (Bruning and Kintz, 1977). Comparisons between mean values on different cycle days used Newman-Keuls procedure (Bruning and Kintz, 1977). Pearson product-moment correlations were calculated between daily values for each of the three steroids and daily frequencies or durations of selected behavioral measures (Bruning and Kintz, 1977). Individual correlations were converted to Z scores and an overall correlation between daily steroid level and behavior was derived by retransforming the mean Z score to a correlation coefficient (Bruning and Kintz, 1977). The degrees of freedom for this overall correlation were estimated as the number of individual correlations minus two. A probability less than or equal to 0.05 was considered significant.
RESULTS
Hormonal Patterns
All females exhibited patterns of estradiol and progesterone secretion consistent with ovulation (Fig. 1). Progesterone showed a detectable increase on the day of the estradiol peak, rising to peak luteal levels 9 days post-E2 peak and then declining to less than 1 nglml by Day + 1.5. Testosterone varied less than either estradiol or progesterone, showing a moderate increase coincident with the estradiol peak then falling to slightly lower levels, where it remained through the rest of the cycle (Fig. 1).
N= PI 131 I61 141 161 ill 161 171 191 191 IYI 131 191 I91 IYI 191 I81 IPI 161 151 131 1’1 131 161 --8 -8 -4 -2 0 +2 +4 +6 +8 +,o +12 +14
Days From Estradiol Peak
FIG. 1. Mean serum levels of estmdiol (O-O), progesterone (O---O), and testosterone (O-O) during the menstrual cycles of nine rhesus monkeys. N is the number of females sampled on each cycle day.

OVARIAN STEROIDS IN RHESUS MONKEYS 43-T
Behavioral Patterns
Copulations. The percentage of tests in which ejaculation occurred was related to the stage of the female’s cycle as illustrated in Fig, 2. Females never received ejaculations earlier than 4 days prior to (Day - 4)) nor later than 5 days after (Day + 5) the estradiol peak. Eight of nine females received ejaculations on the day of the estradiol peak with a smaller proportion receiving ejaculations on other cycle days. Tests with ejaculation peaked coincident with highest estradiol and testosterone levels and remained elevated for the 2 following days (Days 0, + I, and +2).
Ejaculation frequency per test (IL SEM) varied significantly with female cycle stage (F(23, 184) = 9.2, P < O.OOl), increased significantly to 1.1 r 0.3 on Day 0, peaked at 1.5 2 0.4 on Day + 1, and declined to 0.2 + 0.2, a level not above baseline, on Day + 4. Ejaculations ceased completely 6 days after the estradiol peak. No ejaculations were observed during the 10 days prior to menstruation in unaligned cycles. Thus, copulation to ejaculation was limited to a lo-day midcycle period for all females.
Female behavioral changes. The frequency with which females ap- proached, followed, presented, and groomed males; threatened away toward other females and hand slapped varied with their hormonal COR- dition. Female approaches to within at least 2 m of the male varied significantly with cycle stage (F(23, 184) = 17.1, P < O.OOl), increased signrficantly on Day - 3, peaked on Day 0 at a rate of 63/hr, and declined to baseline by Day +4 (Fig. 3). The female approach frequency for Days -2 through +2 differed significantly from all other cycle days, but not from each other, except for Day 0 which was significantly higher than every other cycle day..
J&-r k -6 -4 -2 ; i ; i i ? t ,l 0 +2 +4 +o +* +10 +12 +,a
Days From Estradiol Peak
FIG. 2. Mean percentage of tests with ejaculation in relation to the day of the estradiol peak.

438 WALLEN ET AL.
Days From Estradiol Peak
FIG. 3. Mean hourly frequency per male of female initiated approach to within at least 2 m of the male in relation to the day of the estradiol peak.
The frequency of female initiation of proximity @-ox) varied significantly with female cycle stage (F(23, 184) = 11.5, P < O.OOl), increased sig- nificantly to 15.0 + 6.0 initiationsihr on Day -2, peaked at 36.0 + 6.9/ hr on Day 0, remained significantly elevated on Days + 1 through +3, and declined to 5.6 + 2.6/hr by Day +4, which was not significantly different from zero. The mean time females spent in proximity to the male varied with cycle day (F(23, 184) = 11.3, P < O.OOl), but exhibited a different pattern of change in comparison to proximity frequency. The first significant increase, and the peak, in proximity duration occurred on Day 0 (8.1 + 1.5 min), 2 days after the first significant increase in proximity frequency. Like proximity frequency, proximity duration re- mained significantly elevated through Day +3 and then declined to a level not significantly different from zero on Day +4 (1.50 f 0.7 min).
The frequency and time females spent in back proximity varied with cycle stage (frequency: F(23, 184) = 5.2, P < 0.001; duration: F(23, 184) = 9.7, P < 0.001). Back proximity increased significantly and peaked on Day 0 (5.0 +- 1.7/hr), remained significantly elevated through Day +2, and declined to a frequency not significantly different from zero (1.3 f 0.4/hr) by Day +3. Back proximity duration significantly increased and peaked on Day 0 (2.0 & 1.4 min), remained significantly elevated through Day + 3? and declined to a baseline level of 0.2 t 0.1 min by Day t-4.
The tendency of the female to follow the male varied significantly with her cycle stage (F(23, 184) = 5.0, P < O.OOl), increased significantly on Day - 1 to 15.0 t 5.8/hr, peaked on Day 0 at 22.0 + 6.8/hr, and declined to a frequency not different from 0 (2.2 r 1.1) by Day +4. Females followed the males persistently on Days 0, + 1, and +2, with the males’
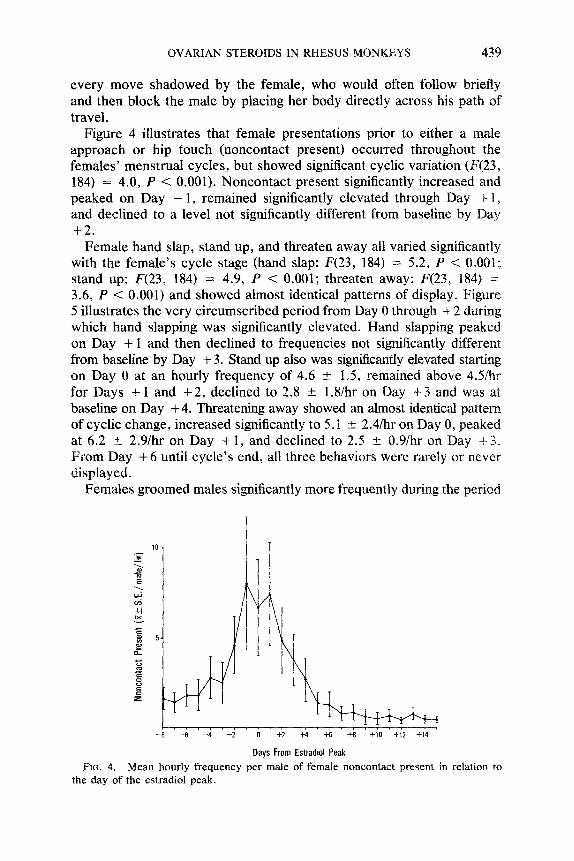
OVARIAN STEROIDS IN RHESUS MONKEYS 439
every move shadowed by the female, who would often follow briefly and then block the male by placing her body directly across his path of travel.
Figure 4 illustrates that female presentations prior to either a male approach or hip touch (noncontact present) occurred throughout the females’ menstrual cycles, but showed significant cyclic variation (F(23, 184) = 4.0, P < 0.001). Noncontact present significantly increased and peaked on Day - 1, remained significantly elevated through Day + 1 7 and declined to a level not significantly different from baseline by Day +2.
Female hand slap, stand up, and threaten away all varied significantly with the female’s cycle stage (hand slap: F(23, 184) = 5.2, P < 0.001: stand up: F(23, 184) = 4.9, P < 0.001; threaten away: F(23, 184) = 3.6, P < 0.001) and showed almost identical patterns of display. Figure 5 illustrates the very circumscribed period from Day 0 through + 2 during which hand slapping was significantly elevated. Hand slapping peaked on Day + 1 and then declined to frequencies not significantly different from baseline by Day +3. Stand up also was significantly elevated starting on Day 0 at an hourly frequency of 4.6 + 1.5, remained above 4.Ybr for Days + 1 and +2, declined to 2.8 t 1.8/hr on Day + 3 and was at baseline on Day +4. Threatening away showed an almost identical pattern of cyclic change, increased significantly to 5.1 + 2.4ihr on Day 0, peaked at 6.2 t 2.9/hr on Day + 1, and declined to 2.5 2 0.9ihr on Day +3. From Day +6 until cycle’s end, all three behaviors were rarely or never displayed.
Females groomed males significantly more frequently during the period
Days From Estradiol Peak
FIG. 4. Mean hourly frequency per male of female noncontact present in relation TV the day of the estradiol peak.

440 WALLEN ET AL.
Days From Estradiol Peak
FIG. 5. Mean hourly frequency per male of female hand slap in relation to the day of the estradiol peak.
of increased estradiol secretion than at other parts of the cycle (F(23, 184) = 4.9, P < 0.001). This was not the result of a general increase in grooming at midcycle as female grooming of other females showed no significant cyclic variation (F(23, 184) = 0.8, P > 0.10). The first significant increase in grooming of males occurred on Day 0 (1.8 ? 0.7/hr), peaked on Day +2 (6.7 -+ 2.O/hr), and then fell to baseline on Day +3 (2.9 -t 0.9/hr). Females were never observed to groom males from Day + 7 until the end of the cycle.
Male behavioral changes. Male approach, sniff, hip touch, mount, and intromission all varied significantly with the female’s cycle. Male ap- proaches resulting in the male standing within 25 cm of the female were the only type which varied significantly with the female’s cycle (F(23, 184) = 8.9, P < 0.001). These approaches increased significantly on Day - 1 (2.0 + 0.6/hr), peaked on Day 0 (3.7 _t 0.6/hr), remained significantly elevated through Day +5 (1.3 & 0.4/hr), and fell to baseline (0.3 + O.l/ hr) on Day +6. Males had a markedly lower frequency of approaches of all types than females (male peak: 5.3 + 0.9/hr; female peak: 63.0 +- 10.2/hr) with females initiating over 90% of all approaches during the period from Day - 4 to Day + 4. Approaches were initiated about equally by males and females at other times, except during the luteal phase, when males initiated more than 70% of approaches, although approach frequencies were very low at this time.
Male sniffing of the females’ genitals occurred relatively infrequently and with great variability between males, but did vary significantly with the females’ cycle (F(23, 184) = 1.7, P < 0.05). The peak frequency

OVARIAN STEROIDS IN RHESUS MONKEYS 442
(0.8 + 0.3/hr) occurred on the day of the estradiol peak, but this frequency was not statistically different from any other cycle day.
Hip touch, mount, and intromission all varied with the female’s cycle (hip touch: F(23, 184) = 9.9, P < 0.001; mount: F(23, 184) = 10.5, P K 0.001; intromission: F(23, 184) = 10.3, P < 0.001). Mount frequency (Fig. 6) typifies the pattern found for all three of these behavioral patterns, which increased significantly on Day 0, peaked on Day + I, and declined to baseline by Day +4. These behaviors differed only in the magnitude of the frequency displayed on each cycle day, with hip touch occurring more frequently than mount, which occurred more frequently than intromission.
Temporal pattern of behavioral change. Table 1 presents the timing of the onset of significant increases and decreases in male and female initiated patterns of behavior. The data suggest an orderly progression of behavioral change, with changes in female behavior preceding changes in male behavior. Thus female approaches increased 3 days prior to the estradiol peak and 2 days before male approaches increased. Female approach to proximity increased a day later (Day - 2) and female follow and noncontact present increased significantly the next day (Day - 1). On Day - 1 male sexual invitations and approaches to proximity were significantly elevated. Several measures of female approach and proximity peaked on the day of the estradiol peak, coincident with the first significant increases in male hip touch, mount, intromission, and ejaculation? and 1 day prior to the peaks in these behaviors. Peak frequencies of female hand slap, stand up, and threaten away coincided with peak frequencies of male copulatory behavior. Threaten away, hand slap, groom. and back
Days From Estradiol Peak FIG. 6. Mean hourly frequency per female of male mount in relation to the day of the
estradiol peak.

P R
TABLE 1 Timing of Significant Increases in Male and Female Iniliated Behavioral Patterns in Relation to the Females’ Estradiol Peak
Days from estradiol peak
Behavior
Female initiated Female approach Approach to proximity Follow male Noncontact present Prox duration Back prox Back prox duration Threaten away Hand slap Groom male Stand up
Male initiated Sniff Approach to proximity Hip touch Mount Intromission Ejaculation
-4 -3 -2 -1 0 +I +2 +3 +4
0 0 0 0 0 0 0 0 0 0 0
+ 0 0 0 0 0 0 0 0 0 0
+ + 0 0 0 0 0 0 0 0 0
+ + t
+++ 0 0 0 0 0 0 0
+++
+t+
+++ +
-c-Ii- +++
i-t+
+
+ +
+
+ + + + + + +
+++ f++
+ +
+ + + 0 + + + + +
+++ +++
+ 0 + 0 + 0 0 0 + 0 5
0 0 F: +
O E i- 0 0 0 rl
0 0 $ + 0 .
0 0 0 0 0 0 0 0 0 0 0 0 + +++ + + + + 0 0 0 0 + +++ + + 0 0 0 0 0 + +++ + + 0 0 0 0 0 + +++ f + 0 0 0 0 0 + t++ + + 0
Note. 0 = No behavior or not significantly different from baseline. + = Significantly above baseline. + + + = Day of peak frequency 01 duration.

OVARIAN STEROIDS IN RHESUS MONKEYS 443
proximity all declined significantly the day before male copulatory behavior returned to baseline levels. All male and female behavioral patterns, except male approach to proximity were not significantly different from zero 4 days after the estradiol peak. Male approaches declined 2 days later.
Correlation between behavioral changes and steroid levels. Table 2 illustrates the overall correlations between daily levels of estradiol, tes- tosterone, and progesterone in relation to daily behavioral frequencies. For this analysis the menstrual cycle was divided into two segments, one from Day -8 through the estradiol peak, and the other from the first day after the estradiol peak (Day + 1) through 8 days after (Day +8). Ten of 11 female initiated behaviors and all six male initiated behaviors were significantly correlated with the daily estradiol level between Day - 8 and the estradiol peak. In contrast, during the same period none of the female or male initiated behavioral patterns were significantly correlated with daily testosterone levels. Correlations were not done with progesterone because it remained below the limits of the radioim- munoassay during this time. All 11 female initiated and five of six male initiated behavioral patterns were significantly inversely correlated with rising progesterone levels after the estradiol peak, but none of the behavioral patterns were significantly correlated with the level of either estradiol or testosterone. Thus the behavioral patterns presented here are positively correlated with increasing estradiol, but not testosterone levels in the first half of the cycle, and inversely correlated with increasing progesterone levels after the midcycle estradiol peak.
DISCUSSION
This study demonstrates that rhesus female proceptive behavior varies predictably with changes in estradiol and progesterone in multifemale groups. Our finding that increased female proceptivity preceded increased copulation by 2 days, and was strongly correlated with estradiol levels, supports the idea that changes in female behavior are involved in the coordination of copulation with maximal fertility. The correlation between the preovulatory behavior of a female and her estradiol level is strong enough so that a male mating on the day of a female’s peak behavior would almost always be mating within a day or two of ovulation.
Most previous investigations of rhesus sexual behavior concentrated upon either copuIation or male patterns of behavior, which often vary more reliably with the female’s hormona! state in pair tests than do female behavioral patterns (Michael et al., 1982). Results of pair-test studies support the view that rhesus males are always ready to copulate, even with nonfertile females (Michael and Zumpe, 1970; Herbert, 1978). In our multifemale group males were highly selective about the reproductive state of their sexual partners and never copulated with luteal or early

TABLE 2 Correlations between Daily Occurrence of Female and Male Initiated Behaviors and Daily Steroid Levels during Two Portions
of the Menstrual Cycle
f: P
Cycle day
-8 to 0 +I to +8
Hormone E2 P T
Female initiated behavior Approach Approach to proximity Follow Noncontact present Proximity duration Back proximity Back proximity duration Threaten away Hand slap Groom male Stand up
Male initiated behavior Sniff Approach to proximity Hip touch Mount Intromission Ejaculation
0.87* 0.86* 0.85* 0.46 0.89* 0.80* 0.87* 0.89* 0.84* 0.89* 0.83*
- 0.16 - 0.28 - 0.31 - -0.25 - 0.44 - 0.16 - 0.39 - 0.44 - 0.26 - 0.40 - 0.26
0.67* - 0.05 0.89* - 0.16 0.93* - 0.39 0.91* - 0.44 0.92* - 0.51 0.93* - 0.45
E2 P T
0.21 - 0.89” 0.35 -0.87* 0.40 -0.83* 0.19 -0.68* 0.46 -0.76* 0.22 -0.82* 0.40 -0.79* 0.10 -0.78* 0.43 -0.80* 0.04 -0.79*
-0.02 -0.71*
0.42 0.42 0.43 q
0.07 l? 0.24 0.28
K z
0.31 0.36 2
0.39 $ 0.30 0.27
0.31 -0.28 0.08 0.24 -0.72* 0.05 0.36 -0.79* 0.34 0.48 -0.76* 0.33 0.46 -0.77* 0.33 0.19 -0.90* 0.22
Note. Abbwvintions: E2 = estradiol, P = progesterone, T = testosterone. * P < 0.05, df = 7.

OVARIAN STEROIDS IN RHESUS MONKEYS 445
follicular females. In addition, over 65% of all copulations occurred on Day 0, + 1, or +2 of the female’s cycle, even though females were attracted to and actively pursued males during a broader period; initiating more than 90% of all approaches from 5 days before to 3 days after presumed ovulation. However, in the present sample at least one female ‘was always between Days -5 and +4, suggesting that male selectivity might be limited to choosing the female closest to ovulation. Recently we have had several instances in this group where no female was near midcycle and in all cases copulation did not occur (unpublished observations).
The strong correlations between most measures of female initiated behavior and estradiol levels suggest that males could obtain substantial information about a female’s hormonal state from the daily pattern of behavioral change. This is supported by the finding that male copulatory patterns are also highly correlated with preovulatory estradiol level, sug- gesting that the male may be using the female’s behavior to track her endocrine cycle. However, our finding that increased male behavior lags 1 or 2 days behind increased female behavior and that copulation continues to increase after female approaching behavior starts to decline suggests that female behavior may be serving more than one function. Starting several days prior to ovulation, female behavior facilitates the formation of a consort pair. During this period females actively signal their attraction to the male by repeatedly approaching and persistently pursuing the male. often blocking his path if he moves. Once the consort pair ‘has been formed and copulation starts the female’s behavior promotes the con- tinuation of the copulatory series and influences the timing of intermount intervals (Wallen and Davis-DaSilva, in prep.). Thus female approach, establishment of proximity, and follows increase until the first day of frequent copulation (Day 0) and then start to decline. In contrast, hand slap, stand up, and threaten away increase significantly for the first time on Day 0 and then completely mirror the pattern of hip toucl~, momt, intromission, and ejaculation. Thus, the decline in female approaches and following during the period of peak copulation probably does not indicate decreased sexual interest, but reflects the fact that the female is now actively copulating with rather than pursuing the male.
An alternative view is that males do not use female behavior to’ determine when to mate, but instead respond to changes in female attractiveness resulting from increasing amounts of vaginally produced aliphatic acids (Herbert, 1978; Keverne, 1982; Michael and Bonsall, 1979). This specific possibility seems unlikely since the best biochemical evidence suggests that these compounds peak midluteally preventing them from’serving as an ovulatory cue (Goldfoot, Kravetz, Goy, and Freeman, 1976: Michael and Bonsall, 1977a). Although the involvement of some other olfactory cue cannot be ruled out, it seems unlikely that rhesus males would depend solely upon an olfactory cue when females display unambiguous behavioral

446 WALLEN ET AL.
evidence of their reproductive condition. A multiple-channel model of sexual communication (Goldfoot, 1982) would suggest that males would use all available information in determining appropriate times for mating. In social groups of rhesus the behavior of the female would be a principle source of information.
Our finding that complete luteal cessation of copulation occurs in a single-male multifemale group suggests that this social condition is sufficient to produce the reproductive coordination characteristic of age-graded multimale/multifemale groups (Gordon, 1981; Wilson et al., 1982a). This supports our previous finding of no luteal copulation in single-male mul- tifemale groups, based upon less frequent testing (Wallen and Winston, 1984). The relationship seen in this study between female sexual behavior changes and ovarian steroid changes is more pronounced than previously reported and may result in part from the single-male testing environment. Wilson and co-workers (1982a) reported that estradiol and progesterone levels were highly predictive of whether or not a female would mate, but did not strongly predict her copulatory rate on a given day. However in this study, copulatory rate was highly correlated with estradiol level prior to the estradiol peak and inversely with progesterone after the estradiol peak. The presence of a single male probably enhanced female competition so that mating only occurred when female motivation to mate was highest. In a group with multiple males daily fluctuations in copulatory rate may be influenced by nonhormonal events, such as com- petition between males for sexually active females and a reduction in female competition by providing more potential mates. The view that a single male produces sexual behavior limited to a smaller portion of the female’s cycle is supported by the fact that females in the present study mated an average of 4.2 days, as compared to 11 .O days in a multimale group (Wilson et al., 1982a). Thus limiting the female’s sexual opportunities may have enhanced the coupling of hormonal status to rhesus female sexual behavior.
The correlational nature of the present study prevents determination of which hormone is actually responsible for the pattern of female sexual behavior found in this study. However, it is striking that none of the patterns of female behavior correlated significantly with testosterone level either before or after the estradiol peak. However, all patterns of female sexual behavior, except one, were highly correlated with estradiol prior to the estradiol peak, and all were inversely correlated with pro- gesterone after the estradiol peak. The lack of a correlation between testosterone and female behavior agrees with the previous finding that serum testosterone levels of group-living females do not predict the onset and offset of female mating (Wilson et al., 1982a). In addition, testosterone is not required for reinstatement of sexual behavior in pair tests with ovariectomized females (Michael et al., 1978; Zumpe, Bonsall, and Michael, 1983). These findings suggest that cyclic fluctuations in female behavior

OVARIAN STEROIDS IN RHESUS MONKEYS 447
probably do not result from fluctuations in testosterone level. However, normal levels of testosterone may provide a basal level of female sexual interest which is increased by estradiol stimulation.
The pattern of female behavioral variation found in this study suggests that increased behavior resulted from rising estradiol and decreased be- havior resulted from rising progesterone. An alternative view, that both increases and decreases result from changes in estradiol (Zumpe et al., 1983), is consistent with the data presented here. However, the striking inverse correlation between declines in female behavior and increases in progesterone suggests that the decrease in behavior seen in the intact female may result from both a decrease in estrogen stimulation and an increased progesterone inhibition. Resolution of this question requires experimental manipulation of these hormones in females living in social groups.
Evidence from the present study and others supports the generalization that the tighest coupling between female hormonal changes and rhesus sexual behavior occurs in multifemale social groups in generous spatial conditions (Gordon, 1981; Wilson et al., 1982a; Wallen and Winston, 1984). A similar conclusion has been reached from studies of rats under a variety of social and environmental conditions (McClintock, 1981). However, duplication of all aspects of the rhesus social environment is not necessary to demonstrate hormonal influences as shown by the similar results obtained in this study and in those using age-graded multimale groups. Similarly, evidence of hormonal influences on female sexual motivation can be obtained in laboratory pair tests when the environment is modified to increase female control over the sexual interaction either by restraining the male (Czaja and Bielert, 197.5; Pomerantz and Goy, 1983) or by requiring the female to perform an operant to gain access to the male (Keveme, 1976; Michael and Bonsall, 1977b).
Controlled investigations of rhesus sexual behavior have been done under environmental conditions ranging from pair tests in small areas to outdoor-housed multimale/multifemale groups. In the first condition hor- monal influences are difficult to demonstrate and most easily detected in the male’s behavior. The latter condition is characterized by clearly delimited periods of mating and rapid changes in male and female behavior which closely follow the female’s ovarian cycle. The availability of testing environments where hormones influence rhesus behavior to differing degrees, provides the experimenter with the opportunity to select the testing environment appropriate to the question being investigated. For example some problems, such as the effect of novel odors, on rhesus sexual behavior (Goldfoot, 1981, 1982), can probably only be demonstrated in a testing environment where hormonal influences on behavior are muted. Similarly, analysis of the responses of untreated ovariectomized females to male approaches (Goldfoot, 1982) must use results from pair tests because males rarely approach ovariectomized females in social

448 WALLEN ET AL.
group tests (Wallen and Gaventa, unpublished). On the other hand socially complex environments provide advantages for investigating the relationship between endogenous hormone changes and reproductive behavior.
Models of rhesus sexual behavior based upon pair tests have emphasized the role of the male in determining the pattern of sexual behavior (Herbert, 1978; Keveme, 1982). This study demonstrates that female rhesus actively communicate their sexual interest in males through behavioral patterns whose expression is tightly coupled to fluctuations in estradiol and pro- gesterone. Integration of the data from both pair tests and socially complex groups will be necessary to provide a complete understanding of the hormonal control of rhesus sexual behavior.
ACKNOWLEDGMENTS This work was supported by NSF Grant BNS 81-1627 to K. W. and Grant RR-00165
awarded by the Animal Resources Program of the NationaI Institutes of Health to the Yerkes Regional Primate Research Center. The authors thank Tom Gordon, Margaret Walker, and Mark Wilson for commenting on a previous draft of this manuscript. The expert technical assistance of Mary Alice Smith in performing the steroid assays is gratefully acknowledged. In addition, Peggy Wentworth and Katherine Rogers are thanked for their assistance in animal handling and behavioral observation.
REFERENCES Altmann, S. A. (1962). A field study of the sociobiology of rhesus monkeys, Macaca
mulatta. Ann. N. Y. Acad. Sci. 102, 338-345. Beach, F. A. (1976). Sexual attractivity, proceptivity, and receptivity in female mammals.
Harm. Behav. 7, 105-138. Bruning, J. L., and Kintz, B. L. (1977). Computational Handbook of Statistics, Scott
Foresman, and Co., Glenview. Carpenter, C. R. (1942). Sexual behavior of free ranging rhesus monkeys (Macaca mulatta).
II. Periodicity of estrus, homosexual, autoeroticism and noncomformist behavior. J. Comp. Psychol. 33, 143-162.
Cochran, C. G. (1979). Proceptive patterns of behavior throughout the menstrual cycle in female rhesus monkeys. Behav. Neurol. Biol. 27, 342-353.
Czaja, J. A., and Bielert, C. F. (1975). Female rhesus sexual behavior and distance to a male partner: Relation to stage of the menstrual cycle. Arch. Sex. Behav. 4, 583-597.
Goldfoot, D. A. (1981). Olfaction, sexual behavior and the pheromone hypothesis in rhesus monkeys: A critique. Amer. 2001. 21, 153-164.
Goldfoot, D. A. (1982). Multiple channels of sexual communication in rhesus monkeys: Role of olfactory cues. In C. T. Snowdon, C. H. Brown, and M. R. Petersen (Eds.), Primate Communication, pp. 413-428. Cambridge Univ. Press, Cambridge, Mass.
Goldfoot, D. A., Kravetz, M. A., Goy, R. W., and Freeman, S. K. (1976). Lack of effect of vaginal Iavages and aliphatic acids on ejaculatory responses in rhesus monkeys: Behavioral and chemical analyses. Horm. Behav. 7, 1-27.
Gordon, T. P. (1981). Reproductive behavior in the rhesus monkey: Social and endocrine variables. Amer. Zool. 21, 185-195.
Goy, R. W. (1979). Sexual compatibility in rhesus monkeys: Predicting sexual performance of oppositely sexed pairs of adults. Sex, Hormones and Behaviour, Ciba Foundation Symposium 62 (new series), pp. 227-255. Elsevier/North-Holland, Amsterdam.
Goy, R. W., and Resko, J. A. (1972). Gonadal hormones and behavior of normal and pseudohermaphroditic nonhuman female primates. Rec. Prog. Horm. Res. 28, 707- 733.

OVARIAN STEROIDS IN RHESUS MONKEYS 449
Herbert, J. (1978). Nemo-hormonal integration of sexual behavior in female primates. In J. B. Hutchison (Ed.), Biological Determinants of Sexual Behavior, pp, 467-491. Wiley, New York.
Herbert, J., and Trimble, M. R. (1967). Effect of oestradiol and testosterone on the sexual receptivity and attractiveness of the female rhesus monkey. Nafure (London) 216, 165-166.
Johnson, D. F., and Phoenix, C. H. (1976). Hormonal control of female sexual attractiveness, proceptivity, and receptivity in rhesus monkeys. J. Comp. Phpsiol. Psychol. 90, 473- 483.
Johnson, D. F., and Phoenix, C. H. (1978). Sexual behavior and hormone levels during the menstrual cycles of rhesus monkeys. Horm. Behav. 11, 160-174.
Keveme, E. B. (1976). Sexual receptivity and attractiveness in the female rhesus monkey. In D. S. Lehrman, R. A. Hinde, and E. Shaw (Eds.), Advances in rhe Study of Behavior, Vol. 7, pp. 155-200, Academic Press, New- York.
Keverne, E. B. (1982). Olfaction and the reproductive behavior of nonhuman primates. In C. T. Snowdon, C. H. Brown, and M. R. Petersen (Eds.), Primate Communication, pp. 396-412, Cambridge Univ. Press, Cambridge, Mass.
McClintock, M. K. (1981). Simplicity from complexity: A naturalistic approach to behavior and neuroendocrine function. In I. Silverman (Ed.), Laboratory and Life: New Directions of Methodology in Social and Behavioral Research, pp. I-19, Jossey-Bass, San Francisco.
Michael, R. P., and Bonsall, R. W. (1977a). Chemical signals and primate behavior. In M. M. Mozell (Ed.), Chemical Signals in Vertebrares, pp. 251-271, Plenum, New York.
Michael. R. P., and Bonsall, R. W. (1977b). Periovulatory synchronization of behaviour in male and female rhesus monkeys. Nature (London) 265, 463-465.
Michael, R. P., and Bonsall, R. W. (1979). Hormones and the sexual behavior of rhesus monkeys. In C. Beyer (Ed.), Endocrine Control of Sexual Behavior, pp. 279-302, Raven Press, New York.
Michael, R. P., and Keverne, E. B. (1968). Pheromones and the cormnunication of sexual status in primates. Nature (London) 218, 746-749.
Michael, R. P., Richter, M. C., Cain, J. A., Zumpe, D., and Bonsall, R. W. (1978). -4rtificial menstrual cycles and behavior: The role of androgens in female rhesus monkeys. Nature (London) 275, 439-440.
Michael, R. P., and Welegalla, J. (1968). Ovarian hormones and the sexual behavior of the female rhesus monkey (Macaca mulatta) under laboratory conditions. J. Endocrinol. 41, 407-420.
Michael, R. P., and Zumpe, D. (1970). Sexual initiating behavior by female rhesus monkeys (Mucaca mulatta) under laboratory conditions. Behaviow 36, 168-186.
Michael, R. P., Zumpe, D., and Bonsall, R. W. (1982). Behavior of rhesus during artificial menstrual cycles. .I. Comp. Physiol. Psycho/. 96(6), 875-885.
Perachio, A. A., Alexander, M., Marr, L. D., and Collins, D. C. (1977). Diurnal variations of serum testosterone levels in intact and gonadectomized male and female rhesus monkeys. Steroids 29, 21-32.
Peretz, E., Coy, R. W., Phoenix, C. H., and Resko, J. A. (1971). Influence of gonadal hormones on the development and activation of the nervous system of the rhesus monkey. Proc. Int. Sot. Psychoneuroendocrinology, 401-411.
Pomerantz, S. M., and Goy, R. W. (1983). Proceptive behavior of female rhesus monkeys during tests with tethered males. Horm. Behav. 17, 237-248.
Wallen, K. (1982). Influence of female hormonal state on rhesus sexual behavior varies with space for social interaction. Science (Washington, D.C.) 217, 375-377.
Wallen, K., and Goy, R. W. (1977). Effects of estradiol benzoate, estrone, and propionates of testosterone or dihydrotestosterone on sexual and related behaviors of rhesus monkeys. Horm. Beizav. 9. 228-248.

450 WALLEN ET AL.
Wallen, K., and Winston, L. A. (1984). Social complexity and hormonal influences on sexual behavior in rhesus monkeys (Macaca mulutra). Physiol. Behav. 32(4), 629- 637.
Wilson, M. E., Gordon, T. P., and Collins, D. C. (1982a). Variation in ovarian steroids associated with the annual mating period in female rhesus (Mucaca mulurra). Biol. Reprod. 27(3), 530-539.
Wilson, M. E., Gordon, T. P., and Collins, D. C. (1982b). Serum 17@estradiol and pro- gesterone associated with mating behavior during early pregnancy in female rhesus monkeys. Horm. Behav. 16, 94-106.
Wright, K., Collins, D. C., and Preedy, J. R. K. (1978). The use of specific radioimmunoassays to determine the renal clearance rates of estrone and 17@estradiol during the menstrual cycle. J. Clin. Endocrinol. Metab. 47, 1084-1091.
Zumpe, D., Bonsall, R. W., and Michael, R. P. (1983). Behavior of rhesus monkeys during 28-day cycles of estrogen treatment. Behav. Nenrosci. 97(4), 615-623.




















