
Social Conventions in Wild White-faced Capuchin Monkeys
29
241 Current Anthropology Volume 44, Number 2, April 2003 2003 by The Wenner-Gren Foundation for Anthropological Research. All rights reserved 0011-3204/2003/4402-0004$3.00 Social Conventions in Wild White-faced Capuchin Monkeys Evidence for Traditions in a Neotropical Primate 1 by Susan Perry, Mary Baker, Linda Fedigan, Julie Gros-Louis, Katherine Jack, Katherine C. MacKinnon, Joseph H. Manson, Melissa Panger, Kendra Pyle, and Lisa Rose Ten researchers collaborated in a long-term study of social con- ventions in wild white-faced capuchin monkeys, involving exam- ination of a 19,000-hour combined data set collected on 13 social groups at four study sites in Costa Rica over a 13-year period. Five behavior patterns qualified as social traditions, according to the study’s criteria: handsniffing, sucking of body parts, and three types of “games.” Some conventions were independently invented in virtually identical form at multiple sites. The extinc- tion of several conventions was observed during the course of the study; it appears that they rarely last longer than ten years. It is hypothesized that the monkeys are using these group- or clique- specific social conventions to test the quality of their social relationships. susan perry is Director of the Independent Junior Research Group in Cultural Phylogeny at the Max Planck Institute for Evolutionary Anthropology (Inselstrasse 22, D-04103 Leipzig, Germany [[email protected]]) and Assistant Professor of Anthro- pology at the University of California, Los Angeles. mary baker is Visiting Assistant Professor of Anthropology at Whit- tier College. linda fedigan is Professor and Canada Research Chair in the Department of Anthropology, University of Calgary. julie gros-louis is a postdoctoral fellow in psychology at Indi- ana University. katharine jack is Assistant Professor of An- thropology at Appalachian State University. katherine c. mackinnon is Assistant Professor of Anthropology in the De- 1. We are grateful to the Costa Rican National Park Service, the Area de Conservacio ´ n Guanacaste (especially Roger Blanco Segura), the Area de Conservacio ´n Tempisque, the community of San Ra- mo ´ n de Bagaces, Hacienda Pelo ´ n, Rancho Jojoba/Brin D’Amor, and the Schutt family for permission to work in the areas occupied by these monkeys. Assistance in data collection was provided by Laura Sirot, Todd Bishop, Kathryn Atkins, Marvin Cedillos Amaya, Sarah partment of Sociology and Criminal Justice, Saint Louis Univer- sity. joseph h. manson is a scientist in the Cultural Phylogeny Group of the Max Planck Institute for Evolutionary Anthropol- ogy in Leipzig and Associate Professor of Anthropology at the University of California, Los Angeles. melissa panger is a re- searcher in the Department of Anthropology at George Washing- ton University. kendra pyle is a graduate student in biology at the University of Pennsylvania. lisa rose is Assistant Professor in the Department of Anthropology and Sociology, University of British Columbia. The present paper was submitted 10 ix 01 and accepted 30 v 02. Social learning has become a topic of intense research interest for anthropologists, psychologists, and behav- ioral ecologists (McGrew 2001:325). Behavioral ecolo- gists are primarily interested in determining the relative costs and benefits of acquiring behaviors via social learn- ing, asocial learning, and/or genetic transmission under various sets of conditions and in modeling the population dynamics of social transmission (see, e.g., Boyd and Rich- erson 1985; Giraldeau and Caraco 2000; Laland 1996, 1999; Lefebvre and Giraldeau 1996; Dewar n.d.). Psy- chologists generally focus on the proximate mechanisms and developmental processes by which animals engage in socially biased learning and are interested in the cog- nitive aspects of social learning (e.g., Whiten 2000, Whiten and Ham 1992, Custance, Whiten, and Fredman 1999, Byrne 1995; see also chapters in Russon, Bard, and Parker 1996, Heyes and Huber 2000, and Heyes and Galef 1996). Anthropologists are interested in social learning and social traditions (one of the possible outcomes of social learning) because of what they can tell us about the origin of culture in humans (King 1991, 1994; McGrew 1992; Boesch and Tomasello 1998; Boyd and Richerson 1996). When anthropologists turn to the com- parative method for aid in constructing models of human evolution, they generally take one of two approaches: argument by homology or argument by analogy. In an argument by homology, the researchers investigate the qualities of those organisms that are shared by humans and their closest living relatives (i.e., chimpanzees and the other great apes) and infer that these traits were pre- sent in their recent common ancestors. Given the tre- mendous variation in social organization, diet, and cog- nitive abilities among the apes, however, it is difficult to use the behavioral data from the extant apes to re- construct ancestral traits. In an argument by analogy, the researcher searches the zoological record more broadly, seeking instances in which taxa have been confronted by similar adaptive problems and asking whether they solved them in the same way. For example, Harcourt (1995), in seeking to test whether males of species having promiscuous mating systems engage in sperm compe- Carnegie, Alisha Steele, Matthew Duffy, Maura Varley, Ryan Cro- cetto, Hannah Gilkenson, Jill Anderson, Craig Lamarsh, Sasha Gil- more, Dale Morris, Dusty Becker, three expeditions of Earthwatch volunteers, and seven groups of University Research Expedition Program volunteers. Susan Wofsy and Denise Alabart assisted in compiling the data. Barb Smuts, Joan Silk, Simon Reader, Mike Huffman, Dorothy Fragaszy, Bill McGrew, Andy Whiten, and two anonymous reviewers commented on the manuscript, providing many helpful comments. Members of the UCLA Behavior, Evo- lution, and Culture group and the contributors to the “Traditions
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Social Conventions in Wild White-faced Capuchin Monkeys

241
C u r r e n t A n t h ro p o l o g y Volume 44, Number 2, April 2003� 2003 by The Wenner-Gren Foundation for Anthropological Research. All rights reserved 0011-3204/2003/4402-0004$3.00
Social Conventions inWild White-facedCapuchin Monkeys
Evidence for Traditions in aNeotropical Primate1
by Susan Perry, Mary Baker,Linda Fedigan, Julie Gros-Louis,Katherine Jack,Katherine C. MacKinnon,Joseph H. Manson,Melissa Panger, Kendra Pyle, andLisa Rose
Ten researchers collaborated in a long-term study of social con-ventions in wild white-faced capuchin monkeys, involving exam-ination of a 19,000-hour combined data set collected on 13 socialgroups at four study sites in Costa Rica over a 13-year period.Five behavior patterns qualified as social traditions, according tothe study’s criteria: handsniffing, sucking of body parts, andthree types of “games.” Some conventions were independentlyinvented in virtually identical form at multiple sites. The extinc-tion of several conventions was observed during the course of thestudy; it appears that they rarely last longer than ten years. It ishypothesized that the monkeys are using these group- or clique-specific social conventions to test the quality of their socialrelationships.
susan perry is Director of the Independent Junior ResearchGroup in Cultural Phylogeny at the Max Planck Institute forEvolutionary Anthropology (Inselstrasse 22, D-04103 Leipzig,Germany [[email protected]]) and Assistant Professor of Anthro-pology at the University of California, Los Angeles. marybaker is Visiting Assistant Professor of Anthropology at Whit-tier College. linda fedigan is Professor and Canada ResearchChair in the Department of Anthropology, University of Calgary.julie gros-louis is a postdoctoral fellow in psychology at Indi-ana University. katharine jack is Assistant Professor of An-thropology at Appalachian State University. katherine c.mackinnon is Assistant Professor of Anthropology in the De-
1 . We are grateful to the Costa Rican National Park Service, theArea de Conservacion Guanacaste (especially Roger Blanco Segura),the Area de Conservacion Tempisque, the community of San Ra-mon de Bagaces, Hacienda Pelon, Rancho Jojoba/Brin D’Amor, andthe Schutt family for permission to work in the areas occupied bythese monkeys. Assistance in data collection was provided by LauraSirot, Todd Bishop, Kathryn Atkins, Marvin Cedillos Amaya, Sarah
partment of Sociology and Criminal Justice, Saint Louis Univer-sity. joseph h. manson is a scientist in the Cultural PhylogenyGroup of the Max Planck Institute for Evolutionary Anthropol-ogy in Leipzig and Associate Professor of Anthropology at theUniversity of California, Los Angeles. melissa panger is a re-searcher in the Department of Anthropology at George Washing-ton University. kendra pyle is a graduate student in biology atthe University of Pennsylvania. lisa rose is Assistant Professorin the Department of Anthropology and Sociology, University ofBritish Columbia. The present paper was submitted 10 ix 01 andaccepted 30 v 02.
Social learning has become a topic of intense researchinterest for anthropologists, psychologists, and behav-ioral ecologists (McGrew 2001:325). Behavioral ecolo-gists are primarily interested in determining the relativecosts and benefits of acquiring behaviors via social learn-ing, asocial learning, and/or genetic transmission undervarious sets of conditions and in modeling the populationdynamics of social transmission (see, e.g., Boyd and Rich-erson 1985; Giraldeau and Caraco 2000; Laland 1996,1999; Lefebvre and Giraldeau 1996; Dewar n.d.). Psy-chologists generally focus on the proximate mechanismsand developmental processes by which animals engagein socially biased learning and are interested in the cog-nitive aspects of social learning (e.g., Whiten 2000,Whiten and Ham 1992, Custance, Whiten, and Fredman1999, Byrne 1995; see also chapters in Russon, Bard, andParker 1996, Heyes and Huber 2000, and Heyes and Galef1996). Anthropologists are interested in social learningand social traditions (one of the possible outcomes ofsocial learning) because of what they can tell us aboutthe origin of culture in humans (King 1991, 1994;McGrew 1992; Boesch and Tomasello 1998; Boyd andRicherson 1996). When anthropologists turn to the com-parative method for aid in constructing models of humanevolution, they generally take one of two approaches:argument by homology or argument by analogy. In anargument by homology, the researchers investigate thequalities of those organisms that are shared by humansand their closest living relatives (i.e., chimpanzees andthe other great apes) and infer that these traits were pre-sent in their recent common ancestors. Given the tre-mendous variation in social organization, diet, and cog-nitive abilities among the apes, however, it is difficultto use the behavioral data from the extant apes to re-construct ancestral traits. In an argument by analogy, theresearcher searches the zoological record more broadly,seeking instances in which taxa have been confrontedby similar adaptive problems and asking whether theysolved them in the same way. For example, Harcourt(1995), in seeking to test whether males of species havingpromiscuous mating systems engage in sperm compe-
Carnegie, Alisha Steele, Matthew Duffy, Maura Varley, Ryan Cro-cetto, Hannah Gilkenson, Jill Anderson, Craig Lamarsh, Sasha Gil-more, Dale Morris, Dusty Becker, three expeditions of Earthwatchvolunteers, and seven groups of University Research ExpeditionProgram volunteers. Susan Wofsy and Denise Alabart assisted incompiling the data. Barb Smuts, Joan Silk, Simon Reader, MikeHuffman, Dorothy Fragaszy, Bill McGrew, Andy Whiten, and twoanonymous reviewers commented on the manuscript, providingmany helpful comments. Members of the UCLA Behavior, Evo-lution, and Culture group and the contributors to the “Traditions

242 F current anthropology Volume 44, Number 2, April 2003
tition, used type of mating system as the independentvariable (i.e., the adaptive challenge) and testes size asthe dependent variable (i.e., the adaptive outcome), con-trolling for phylogeny to some extent by performing theanalysis at the level of the genus. Although Japaneseresearchers working with Japanese macaques (Macacafuscata) were the first to propose that social traditions(which they termed “pre-culture,” “infrahuman cul-ture,” or “sub-human culture”) were an important partof the behavioral biology of primates (Itani 1958, Itaniand Nishimura 1973, Kawai 1965, Kawamura 1959), themajority of research in the field now known as “culturalprimatology” (McGrew 1998, de Waal 1999) has focusedon the great apes (e.g., McGrew 1992; McGrew and Tutin1978; Whiten et al. 1999; Russon 1996; van Schaik, Fox,and Sitompul 1996; Boesch 1996a, b; Matsuzawa andYamakoshi 1996). Social learning is by no means re-stricted to humans and their closest relatives; in fact,some of the best-documented examples of social learningand traditions are found outside the order Primates (e.g.,feeding traditions in rats [Terkel 1996, Galef n.d.], sociallearning of courtship signals in cowbirds [West and King1996], mate-choice copying in guppies and quail [Du-gatkin 1996, White and Galef 1999], and vocal traditions
in Nonhuman Primates” conference at the University of Geor-gia–Athens provided useful comments as well. Joan Silk wrote thedata-collection program, FOCOBS, used by many of the researchers.Gayle Dower produced the line drawings.
The UCLA Council on Research provided SP with funding forthis project during the data analysis stage, and the Max PlanckInstitute for Evolutionary Anthropology provided funding duringthe write-up stage. Numerous other granting agencies inadvertentlyfunded the fieldwork that gave rise to this project as well: SP thanksthe National Science Foundation (for a graduate fellowship, an NSF-NATO postdoctoral fellowship, and a POWRE grant #SBR-9870429),the L. S. B. Leakey Foundation (for three grants), the National Ge-ographic Society, the University of Alberta (for an I. W. KillamPostdoctoral Fellowship), Sigma Xi, the University of MichiganRackham Graduate School (for four grants), the University of Mich-igan Alumnae Society, and UCLA (for two Faculty Career Devel-opment grants). MB thanks the University of California–RiversideGraduate Division (for three grants), Earthwatch, UREP, and NIH-MIRT (for a grant to E. Rodriguez). LF’s research is funded by anongoing grant (A7723) from the Natural Sciences and EngineeringResearch Council of Canada (NSERCC). JG thanks the Universityof Pennsylvania and NSF for a graduate fellowship and a dissertationimprovement grant. KJ’s research was supported by the NationalGeographic Society, the NSERCC (postgraduate scholarship), theAlberta Heritage Scholarship Fund (Ralph Steinhauer Award), theRuggles-Gates Fund for Biological Anthropology (Royal Anthro-pological Institute), Sigma Xi, the Faculty of Graduate Studies andResearch/Department of Anthropology at the University of Alberta,and the above-mentioned NSERCC grant to LF. KM’s fieldwork wassupported by an NSF dissertation improvement grant (#SBR-9732926) and a University of California Berkeley Social Sciencesand Humanities Research Grant. JM was supported by a Universityof California Los Angeles Faculty Career Development Award. MPthanks the Costa Rican National Park System (park fee exemptiongrant), the Organization for Tropical Studies (especially M. Quesadaand K. Stoner), and the University of California Berkeley’s Anthro-pology Department (three grants) for helping to make her researchpossible. LR was supported by an NSERCC postgraduate scholar-ship, NSF, the National Geographic Society, the L. S. B. LeakeyFoundation, Sigma Xi, and Ammonite Ltd. [Supplementary mate-rials appear on the journal’s web page (http://www.journals.uchicago.edu/CA/home.html).]
in birds and cetaceans [see review by Janik and Slatern.d.]. To understand the socioecological factors that pro-mote social learning in an evolutionary perspective, itis necessary to look beyond the great apes.
The genus Cebus (capuchin monkeys, a type of NewWorld primate) proves to be a particularly interestingsource of data for researchers interested in explaininghuman origins because capuchins have independentlyevolved many traits that are present in humans and/orchimpanzees (Visalberghi and McGrew 1997). Capuchinshave extraordinarily large brain sizes relative to theirbody sizes (Harvey, Martin, and Clutton-Brock 1987, Ste-phan, Baron, and Frahm 1988), they are omnivorous andrely heavily on extractive foraging (Freese 1976, Chap-man and Fedigan 1990, Terborgh 1983), they hunt ver-tebrates (Terborgh 1983, Perry and Rose 1994, Fedigan1990, Rose 1997), they are very tolerant during foragingand sometimes share food (de Waal 1997, 2000; Perryand Rose 1994), they are skilled tool users (Urbani1999,Visalberghi 1990, Ottoni and Mannu 2001, Boinski1988, Westergaard 1994, Westergaard and Suomi 1995),they engage in nonconceptive sex in all age-sex combi-nations (Manson, Perry, and Parish 1997), they exhibitfrequent alloparenting (Manson 1999, O’Brien and Rob-inson 1991), they have high rates of intraspecific lethalaggression (unpublished data), and they have complexsocial relationships characterized by frequent coopera-tion and coalitionary aggression (Perry 1996, 1997,1998a, b, n.d.; Manson et al. 1999; Rose 1992). The cap-uchin’s generalist lifestyle, combined with its large brainsize, tolerance, and intensely social nature, makes thistaxon a likely candidate for prevalent social learning ina variety of behavioral domains (van Schaik, Deaner, andMerrill 1999, Reader n.d., Coussi-Korbel and Fragaszy1995). Previous studies in captivity have demonstratedcapuchins’ abilities to engage in social learning in thebehavioral domains of food choice and food processing(Custance, Whiten, and Fredman 1999, Fragaszy and Vis-alberghi 1996, Visalberghi and Addessi 2000), and thesestudies provide mixed results regarding the complexityof the social learning mechanisms used by capuchins.To date, there is no published literature on social learn-ing or traditions in wild capuchins in any behavioraldomain.
If you ask laypeople or even cultural anthropologiststo describe the cultural differences between two culturesthey have experienced, chances are they will not dwellupon the technological differences between the two pop-ulations but will tell you much about their social cus-toms, perhaps detailing the various faux pas they madewhen attempting to interact with the locals properly.Human cultures are so rich in social conventions (i.e.,dyadic social behaviors or communicative behaviors thatare unique to particular groups or cliques) that it takesimmigrants decades to become enculturated to the pointthat they know the subtle nuances of appropriate inter-action patterns and cease to make social blunders. If so-cial conventions are such an important aspect of humanculture, then it is striking that so little of the literatureto date on culture in nonhumans has dealt with this

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 243
behavioral domain. Is it the case that most nonhumanslack interpopulation variability in social conventions?Or has there merely been a dearth of research effort inthis domain?
Most researchers who have attempted to documentsocial traditions or social learning in mammals have fo-cused on foraging techniques (e.g., Itani 1958; Kawai1965; Scheurer and Thierry 1985; Watanabe 1989, 1994;McGrew 1992; Matsuzawa 1994; Boesch 1996a; Matsu-zawa and Yamakoshi 1996; Fragaszy and Visalberghi1996; Galef 1996; Terkel 1996; Boran and Heimlich 1999;Byrne 1999; Hudson, Schaal, and Bilks 1999; Kitchener1999; Laland 1999; Nel 1999; van Schaik, Deaner, andMerrill 1999; Wilkinson and Boughman 1999; chaptersin Fragaszy and Perry n.d.a; see also, for a review of therole of social learning in foraging in vertebrates generally,Galef and Giraldeau 2001), and therefore most of thetheory that has been developed concerning the emer-gence and maintenance of traditions has focused on thisbehavioral domain. However, there are also some inter-site or intergroup differences reported for some speciesthat can best be described as social conventions. Someexamples are the various styles of grooming in Japanesemacaques (e.g., alternative forms of louse-egg-handlingtechniques) and chimpanzees (e.g., social scratching, leafgrooming, and hand-clasp grooming) that are unique toparticular matrilines or sites (Nakamura et al. 2000;McGrew and Tutin 1978; Tanaka 1995, 1998; Boesch1996a, b; Whiten et al. 1999; McGrew et al. 2001). Somesocial signals (e.g., leaf-clipping) are found at multiplesites in almost identical forms but are used for differentpurposes at different sites (Boesch 1996a, b). Other sig-nals are different in their form but can serve the samefunction at different sites (e.g., knuckle-knocking andleaf-clipping are both used as courtship gestures by chim-panzees at different sites [Boesch 1996b]).
In this paper, we will focus exclusively on traditionsin the behavioral domain of social conventions. Usingdata collected at four study sites over 13 years, we willdescribe several social conventions that seem to qualifyas traditions in wild white-faced capuchin monkeys (Ce-bus capucinus), and we will attempt to determine thefunctions of these behaviors. Three classes of behaviorswill be discussed: handsniffing, sucking on body parts,and “games.” We will go on to consider the putativefunctions of such signals, the design features that areoptimal for such signals, and the transmission dynamicsof social conventions. Most models of social transmis-sion are about behaviors performed by single individualsthat have a clear adaptive function (e.g., food choice, tooluse, food processing). Given that (a) social conventionsare necessarily performed dyadically rather than by sin-gle individuals and (b) the role that social conventionsplay in the behavioral biology of the animals is quitedifferent from the role of foraging-related behaviors, it isto be expected that the transmission dynamics of socialconventions will be different from those of behaviorsthat are more frequently modeled.
Identifying a Tradition
We define a “behavioral tradition” as a practice that isrelatively long-lasting and shared among members of agroup, each new practitioner relying to some extent uponsocial influence to learn to perform it. We imposed thefollowing three criteria on the data to determine whethera behavior pattern qualified as a “tradition”: (a) Inter-group variation: The behavior in question must be pre-sent in at least one social group and absent in at leastone group. To qualify as “present” or common in a par-ticular group, the behavior must be seen at a rate of atleast once per 100 hours of observation and must be per-formed by at least three individuals; to qualify as “ab-sent,” the behavior must never have been seen, and thegroup must have been observed for at least 250 hours.(b) Expansion: The behavior must exhibit an expansionin the number of performers over time (unless all groupmembers perform it). Ideally, we tried to document morethan two links in a social transmission chain, but be-cause methodological difficulties such as gaps in obser-vation time precluded the use of such a stringent crite-rion in many cases we also allowed behaviors to qualifyif there was simply an increase in the number of per-formers over time. (c) Durability: The behavior must bedurable. We arbitrarily coded behaviors as durable if theywere observed spanning at least a six-month period.
These criteria are ways of operationalizing traditions,rules of thumb that are designed to detect traditions witha minimum of error in a messy collection of datasets. Itis possible to think of ways in which they could be vi-olated even while the behavior qualified as a traditionaccording to our definition. For example, animals at allsites could conceivably learn to identify a particular spe-cies as a predator through some combination of trial-and-error learning and social transmission; this would be atrue tradition, though it would violate the intergroup-variation criterion. However, in the behavioral domainof social conventions, use of the variation criterion isless likely to rule out true traditions (see discussion). Itis also possible to think of ways in which some of theabove criteria might be met even if social learning werenot the mechanism for acquiring the trait (i.e., the be-havior was not a true tradition). For example, regardingthe expansion criterion, it is possible to conceive of someecological condition that would cause an expansion innumber of performers regardless of whether social learn-ing was the mechanism for expansion; again, while thisis quite possible for foraging-related behaviors, it is im-probable in the domain of social conventions. The du-rability criterion is entirely arbitrary, as even a veryshort-lived “fad” can be purely the consequence of sociallearning. This criterion was imposed primarily to makethe datasets more comparable across sites and to givesome indication of whether the behavior had become aregular part of the group’s behavioral repertoire.
Previous research programs designed to detect tra-ditions in wild populations (e.g., Whiten et al. 1999) em-phasize the importance of excluding the possibility that

244 F current anthropology Volume 44, Number 2, April 2003
Fig. 1. Costa Rica, showing the capuchin study sites.
interpopulation genetic and/or ecological differences un-derlie behavioral differences between populations. Webelieve the genetic and ecological differences betweenour four study sites to be fairly trivial (see below). Fur-thermore, we are not convinced that this “process ofelimination” approach is a necessary or sufficient meansof detecting traditions in wild populations, although itis quite useful as a first step in isolating phenomena forfuture research. In our view (Fragaszy and Perry n.d.b)the most critical feature of a tradition is a substantialrole of social influence in the acquisition of the trait bynew practitioners. There may well be genetic differencesbetween individuals (e.g., in their perceptual abilities ortemperaments) that make some individuals more likelythan others to exhibit particular behaviors, but as longas social influence is demonstrated to be important inthe acquisition of the trait it may still qualify as atradition.
The Study Species
Capuchin monkeys are long-lived animals, living 40�years in captivity and having a prolonged juvenile period(Harvey, Martin, and Clutton-Brock 1987, Fragaszy, Vis-alberghi, and Robinson 1990, Ross 1988). Among white-faced capuchins (Cebus capucinus), females first givebirth at about six to eight years of age (Fedigan and Perry,unpublished data), whereas males are considered sexu-
ally mature at about eight years (Freese and Oppenhei-mer 1981). These monkeys live in multimale, multifem-ale groups. Females are philopatric, whereas malesmigrate multiple times in their lifetime and often form“migration alliances” of two to four males that haveaffiliative relationships and migrate together, invadingnew groups and evicting some or all of the resident males(Fedigan, Rose, and Avila 1996, Fedigan 1993, Fediganand Jack 2001). Group sizes tend to average around 17–19individuals at Santa Rosa (SR) and Palo Verde (PV),though the study groups at Lomas Barbudal (LB) are larger(ranging from 20–37) (Fedigan, Rose, and Avila 1996, Fe-digan and Jack 2001, Panger 1997, Perry, Manson, andGros-Louis, unpublished data). Sex ratios range from 1male to 1.2 females at PV and SR (Panger 1997, Fediganand Jack 2001) to 1 male to 2 females at LB (Perry, Man-son and Gros-Louis, unpublished data). Female-femaledyads typically have the closest bonds, as measured byproximity, grooming frequencies, and reliable coalition-ary aid (Perry 1996, Manson et al. 1999). The alpha maleplays a central role in group politics as the preferred part-ner of both males and females in coalitionary aggressionand the primary male recipient of females’ grooming(Perry 1997, 1998a, b). However, subordinate adult malesare also active in the social life of the group and areparticularly active in alloparenting and play sessionswith juveniles (Perry and MacKinnon, unpublished data).Coalitionary aggression both within and between social

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 245
Fig. 2. Core areas of study troops. A, Santa Rosa: NA, Nancite; CA, Cafetal; BH, Bosque Humido; CU, Cuanji-niquil; SE, Sendero; LV, Los Valles; B, Palo Verde: LT, Lagoon Troop; ST, Station Troop; WHT, Water Hole Troop;C, Lomas Barbudal: RA, Rambo’s group; AB, Abby’s group. (The latter two groups’ broadly overlapping rangesextend into the northern tip of the reserve.)
groups is an integral aspect of capuchin social life (Perry1996, 1997, 1998a, b, n.d.).
The Study Sites and Databases
Data were contributed from four study sites in north-western Costa Rica: Lomas Barbudal (LB), Santa Rosa(SR), Palo Verde (PV), and Curu (CU) (see figs. 1 and 2).All of these sites consist largely of tropical dry forest.The first three of these sites have been the subjects ofmany ecological studies over the past few decades, andwe can say with some degree of confidence that they areecologically very similar (Panger et al. 2002). Becausedeforestation has only quite recently (i.e., in the pastthree decades) become a problem in Costa Rica, we con-sider it very unlikely that there are any major geneticdifferences between the study populations, because there
were forest corridors connecting these sites in the recentpast. In fact, forest corridors still connect LB to PV. Table1 details the observation periods for each researcher ateach study site. The number of observation hours in thetable represents the number of contact hours (i.e., thenumber of hours that at least one observer was collectingad lib and/or focal-animal samples [Altmann 1974]). Jackand Fedigan did not record ad lib data, so the numberreported for them is the number of hours of focal sam-ples. The total number of contact hours from which thedata in this paper are drawn is 19,145 (or well over 25,000hours of behavioral records, because there were oftenmultiple observers collecting data simultaneously). Inthis paper, the study groups will be referred to by a codeconsisting of the site name followed by the first twoletters of the group name at that site (see table 1 for thefull names of each study group and site, which can be

246 F current anthropology Volume 44, Number 2, April 2003
table 1Periods of Data Collection on Social Behavior at Each Study Site
Site and Study Group Time Period (months/year) Observation Hours Principal Investigators
Lomas BarbudalAbby’s (AB) 5–8/90 337 SP
5–12/91 619 SP1–12/92 1,850 SP1–5/93 1,234 SP
2/94 72 SP, JM7–8/95 282 SP, JM12/96 48 SP, JM
1–8/97 914 SP, JM, JGL2–5/98 381 JGL1–7/99 356 SP, JM, JGL1–6/00 372 JGL1–6/01 784 SP, JM
Rambo’s (RA) 1–8/97 964 SP, JGL1–5/98 315 JGL1–7/99 759 SP, JGL, JM1–5/00 542 JGL1–6/01 655 SP, JM
Santa RosaSendero (SE) 1–6/86 69 LMF
5–9/92 10 KM1–4/93 35 KM
Cerco de Piedra (CP) 1–6/86 123 LMF1–7/91 285 LR5–9/92 150 KM1–4/93 120 KM
1–9,12/95 405 LR1–7/96 327 LR
1–12/98 471 KM, KJ1–4/99 86 KJ
Los Valles (LV) 1–7/91 260 LR5–9/92 170 KM1–4/93 200 KM
1–9,12/95 656 LR1–7/96 341 LR
1–12/98 673 KM, KJ1–4/99 133 KJ
Nancite (NA) 12/95–6/96 408 LRCuajiniquil (CU) 2–12/98 126 KJ
1–2/99 1 KJCafetal (CA) 3–4/99 56 KJBosque Humido (BH) 2–12/98 305 KJ
1–4/99 131 KJPalo Verde
Station Troop (ST) 4–12/95 852 MP1/96 36 MP
Water Hole Troop (WHT) 3/95 84 MPLagoon Troop (LT) 3–7/95 228 MP
Curu, Bette’s (BE) 8–9/91 189 MB1–6/93 692.5 MB7–8/94 147.5 MB
1–4,6–9/95 665 MB7–9/96 226 MB
used to link these results with information on thesegroups in other publications).
The study of social traditions was never the primaryfocus at any site. The methods of data collection variedfrom site to site and from year to year because the re-search topics varied. Panger (PV) studied handedness andobject manipulation and observed members of all age-sex classes, collecting ten-minute focal-animal samples(including proximity scans at the end of each sample)
and ad lib data on rare behaviors. Baker (CU) focused onfur-rubbing in members of all age-sex classes. All of theLB researchers have studied some aspect of social be-havior. Data collection consisted primarily of ten-min-ute focal samples of individuals of all age-sex classesduring 1992–93 (Perry), 1996–2000 (Gros-Louis), and1999 (Perry). Adult and juvenile males were the focus ofstudy in 1996–97 (Perry), and adult females were the fo-cus of study in 1995–97 and 1999 (Manson), whereas

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 247
Fig. 3. Two females handsniffing.
adults of both sexes were followed in 1990–91 (Perry)and 2001 (Perry and Manson). Ad lib data were recordedon all odd social behaviors observed in any age-sex classthroughout the course of the study, particularly whenthe behavior observed was not on existing ethograms.Fedigan (SR) studied social behavior in adults of bothsexes. Rose (SR) collected data on social and foragingbehaviors in adults and subadults. Although she spentmore time in the field than is indicated in table 1, thetable includes only those observation periods for whichshe is certain handsniffing and other odd ad lib socialbehaviors were reliably recorded. Jack (SR) studied socialbehavior in males (both adult and juvenile). MacKinnon(SR) studied social development, and most of her dataconsisted of ten-minute samples of immatures, thoughshe also collected some ten-minute samples and groupscans of adults. Although each researcher had a differentresearch agenda, all of the researchers at LB, SR, and PVwere in regular contact with one another from the be-ginnings of their studies. We made a point of collectingdata on these odd behaviors (handsniffing, games, andsucking) and comparing notes on them throughout ourstudies, even before we had formal research plans foranalyzing the data. Because of our mutual interest inthese striking behaviors, we are confident that we re-corded all instances of these behaviors in focal data, andwe probably failed to record very few observations evenin ad lib data, though our different sampling schedulesmay have biased observations toward some age-sex clas-ses rather than others.
Handsniffing
Handsniffing (fig. 3) was first reported by Fedigan (1993:861) and described as follows: “One monkey places an-other’s hand or foot over its own face and, with eyesclosed, inhales deeply and repeatedly over ≥1 min.” Thisbehavior has been seen at other sites as well, with someminor alterations. At LB-AB, where the behavior hasbeen most common, one monkey takes the hand of an-other monkey and either covers its own nose and mouthwith it, as if putting on a gas mask, or else inserts theother monkey’s fingers up its own nostrils. This is oftena mutual behavior, with each monkey simultaneouslyhaving the other’s hand on or in its own nose. Partici-pants will sit in this pose for up to several minutes withtrancelike expressions on their faces, sometimes sway-ing. Grooming typically precedes and/or follows hand-sniffing. Sometimes the sniffers sneeze, blowing the fin-gers out of their noses, but they always rapidly reinsertthe partner’s fingers when this happens. At LB, it is al-ways the hands, rather than the feet, that are used. Onefemale at LB typically sniffed her own hand while in-serting the fingers of her other hand up her partner’snose. In SR-CU and also in PV-ST, handsniffing was oftencombined with finger-sucking rather than insertingfingers up the nose. Visual images of handsniffing beha-vior, as well as some of the other social conventionsdescribed in this paper, can be viewed at the following
web site: http://www.sscnet.ucla.edu/02S/anthro128a-1/cebustraditions.htm.
The researchers who made the initial reports of hand-sniffing assumed that handsniffing was a species-typicalbehavior and speculated that some sort of olfactory com-munication was occurring, perhaps involving commu-nication about reproductive states via pheromones inurine, bonding via transmission of semiochemicals (seeNicholson 1984), or information exchange about recentforaging or social activities (Fedigan 1993, Perry 1995).However, subsequent research has revealed that hand-sniffing is by no means a universal feature of capuchincommunication. If olfactory communication is occur-ring along with handsniffing, then the chemical mes-sages being conveyed are likely to be different in differentgroups, because the distribution of handsniffing acrossage-sex classes is strikingly different in different groupsand at different times. For example, the pheromonestransmitted by female urine would not necessarily bethe same ones transmitted via male urine; also, the typesof pheromonally transmitted information about conspe-cifics that would be useful to females are not necessarilythe same pieces of information that would be interestingto males.
Table 2 details the distribution of handsniffing obser-vations across study sites and years. Handsniffing wasobserved on a fairly regular basis in 5 out of 14 studygroups and three out of four sites. It appears that it isnot universal across study groups at any site (though dataare scant for some groups at some study sites). Thus,handsniffing appears to have been invented or imported(i.e., transmitted socially by a migrant) multiple times.Once it is established, it does not remain in the repertoirepermanently. The behavior disappeared for a period ofseveral years at SR-CP before being reinvented. It mayhave disappeared at SR-SE as well, though there wereinsufficient hours of observation to confirm this. Afterbeing a frequent behavior at LB-AB for at least sevenyears, handsniffing vanished from the repertoire in 1998,at the time the most avid handsniffer disappeared. Al-though the form of handsniffing was roughly consistent

248 F current anthropology Volume 44, Number 2, April 2003
table 2Temporal Distribution of Handsniffing across Social Groups
1986 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999
SR-SE CD3 – – xxx xxx – – – – – –SR-CP CD3 – 0 xxx xxx – 0 RN1 – CD3 xxxSR-LV – – 0 xxx xxx – 0 RN1 – RD3 0SR-NA – – – – – – – 0 – – –SR-CA – – – – – – – – – – xxxSR-CU – – – – – – – – – CD Cx1
2,4SR-BH – – – – – – – – – 0 0LB-AB — 0 CN2 CD4 CD4 Cx1 Cx3 Cx1 CD3 0 0LB-RAa – – – – – – – xxx 0 0 0PV-LT – – – – – – xxx – – – –PV-WHT – – – – – – xxx – – – –PV-ST – – – – – – CD4 xxx – – –CU-BE – – xxx – 0 xxx 0 xxx – – –
note: First letter indicates frequency, second durability, number spread across social network; –, no data; xxx, some observationtime but insufficient data to confirm absence (i.e., ! 250 contact hours or ! 100 focal hours: ! 6 months); C, common ( 1 1 observa-tion per 100 hours); R, rare ( ! 1 observation/100 hours); 0, absent (no observations); D, durable (behavior observed over a 6�-monthperiod); N, not durable; 1, only one dyad performs the behavior; 2, multiple dyads perform the behavior but all of them have oneindividual in common; 3, multiple dyads perform the behavior; 4, all members of at least one age-sex class perform the behavior.aThere were anecdotal observations of handsniffing in LB-RA in 2000.
table 3Distribution of Handsniffing (percentage) across Sex Classes
Group Male Sniffs Male Male Sniffs Female Female Sniffs Male Female Sniffs Female Na
LB-AB ! 1 ! 1 2 96 168SR-SE 71 14 14 0 7SR-CP 1986 86 10 0 5 21SR-CP 1996–99 0 0 75 25 16SR-CU 64 27 9 0 10PV-STb 33 50 33 0 12
aTotal number of handsniffing observations in the sample.bTotal exceeds 100% because some of these male-female bouts are mutual.
across sites, the distribution of handsniffing across age-sex classes is strikingly different across study groups.Thus, it is worthwhile to describe the direction and dis-tribution of handsniffing among males and females foreach site (table 3), particularly in light of previously pro-posed hypotheses regarding the possible olfactory func-tions of handsniffing. Handsniffing was exhibited almostexclusively by female-female dyads at LB-AB, the sitewhere it was most common, whereas it was seen exclu-sively in male-male and male-female dyads at SR-SE, SR-CU, and PV-ST.
Further information on the form, rate, and distributionof handsniffing across social networks can be summa-rized as follows: (a) the precise form of handsniffing var-ied somewhat from site to site; (b) the successful prop-agation of handsniffing seemed to depend very much onthe social characteristics of its practitioners (e.g., age,sex, rank, position in a social network), and handsniffingvanished from the repertoire when key handsniffers van-ished; and (c) in those groups for which datasets per-mitted analysis, handsniffing was associated with fre-
quent proximity and grooming (for details, see theelectronic edition of this issue on the journal’s web page).
Handsniffing met all of the criteria for a tradition. Itwas common in five groups, rare in two, and completelyabsent in six groups (three of which were observed for asufficient amount of time to be confident that the be-havioral repertoire was adequately sampled). For two ofthese groups, it was possible to document an expansionof the number of performers over time. The behavior wasdurable in six groups.
Sucking of Body Parts
In some groups, pairs of monkeys were observed to spendlong periods of time sucking on one another’s fingers,toes, ears, or tails. This behavior was similar to hand-sniffing with regard to its social context in that it oc-curred in relaxed social contexts, such as resting in con-tact or grooming, while pairs of monkeys were fairlyisolated from the rest of the group. Sucking was partic-

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 249
Fig. 4. Guapo and a juvenile playing the finger-in-mouth game.
ularly prevalent at LB-RA, where it was often a mutualactivity that could last for over an hour at a time. Of 34observations of this behavior spanning a six-month fieldseason, over half involved mutual sucking. This activitywas equally common in male-male and male-female dy-ads but was not seen in female-female dyads. The vastmajority of the observations involved a particular youngadult male. Sucking virtually vanished from the group’sbehavioral repertoire when this male disappeared in July1997.
On rare occasions, tail-sucking was observed at LB-ABas well. Virtually all of it involved a single adult male,Guapo. In 1990–96, Guapo’s tail-sucking behavior wasunreciprocated and seemed to function as a way for himto invite grooming. In 1997 he began to engage in mutualtail-sucking with a new immigrant male, Hongo. As faras we know, mutual sucking of body parts has not beenobserved at sites other than LB.
Finger-sucking has been observed in some of the SRgroups and generally conforms to the same patterns seenfor handsniffing in these groups. In SR-CU, a pair ofmales that migrated together between SR-LV, SR-CU,and SR-CA often engaged in both finger-sucking andhandsniffing, sometimes combining them in the sameinteraction. Finger-sucking was observed once at SR-CP.Occasionally it was observed at CU-BE in the context ofplaying the “toy” game (see below).
Sucking, therefore, met only two of the criteria for atradition. It was common only in two groups and seenoccasionally in three more in slightly different contexts.It was durable at all sites where it was observed. How-ever, documentation of the expansion in the number ofperformers for sucking was difficult, because we are notsure exactly when the behavior entered the repertoireand how individuals’ acquisitions of the behavior coin-cided with the timing of field seasons. There was nomarked expansion of number of performers during ob-servation periods.
Games
What we will refer to as “games,” because they are ofteninitiated in a play context, differ from rough-and-tumbleplay in being more relaxed and quiet. They are dyadic,and the pair performing the behavior tends to be on theperiphery of the group, often during a group siesta.Games are typically preceded by either slow-motionwrestling or grooming, particularly of the mouth andface. With one exception (the “toy” game, which isplayed at CU-BE as well), games are found exclusivelyat LB-AB. Circumstantial evidence suggests that all ofthe games at LB-AB may have been transmitted initiallyby a single individual, Guapo. When the study began in1990, Guapo was a young adult male, probably about8–10 years of age judging from his size. He was a sub-ordinate male for most of 1990–99; he was alpha malefor a few days in 1998 and became alpha male on a morepermanent basis in 1999, until he disappeared from thegroup toward the end of the year 2000. Guapo was the
first individual observed to play each of these games andalso the most frequent game player, especially in theearly years of observation. Aside from Guapo, the mostfrequent participants in games were juvenile males, butsome games were also played by adult females. The groupsize for most of the study of LB-AB was approximately21 individuals (ranging in size from 17 to 31), and eachgame was played by 10–13 individuals.
The most commonly observed game was the finger-in-mouth game (fig. 4), which was first observed in No-vember 1991. In this game, monkey A puts its finger inmonkey B’s mouth—this can start in the context ofgrooming the inside of the mouth. Monkey B clampsdown on A’s fingers, apparently not enough to hurt buthard enough that A cannot readily remove its fingers.Monkey A goes through various contortions to extractits finger—this may involve using both hands and feetto pry open B’s mouth or putting the feet on B’s face forleverage while pulling hard on the captured hand. OnceA’s finger has been successfully extracted, either A rein-serts its hand in B’s mouth for another bout of the gameor B inserts its fingers in A’s mouth so that the gamecontinues with the roles reversed.
Most dyads observed to play this game includedGuapo. Prior to 1993 all 30 finger-in-mouth games ob-served involved Guapo. In 1993 others began to playwithout Guapo, and in the last three field seasons ap-proximately half of the dyads observed to play this gamedid not involve him. Figure 5 shows the social trans-mission pathways (i.e., social transmission chains) forthe finger-in-mouth game. In this figure, arrows desig-nate the probable transfer of the convention from oneindividual to another. Most of the females acquired thegame as adults, whereas the males were all juvenileswhen they acquired it from Guapo. Aside from one pos-sible instance involving two juvenile males, we neverobserved this game in neighboring LB-RA. This gamewas still being played in 1999, and a less elaborate formof the game was seen in two dyads in 2000. The dyadsthat played together after Guapo’s withdrawal from game
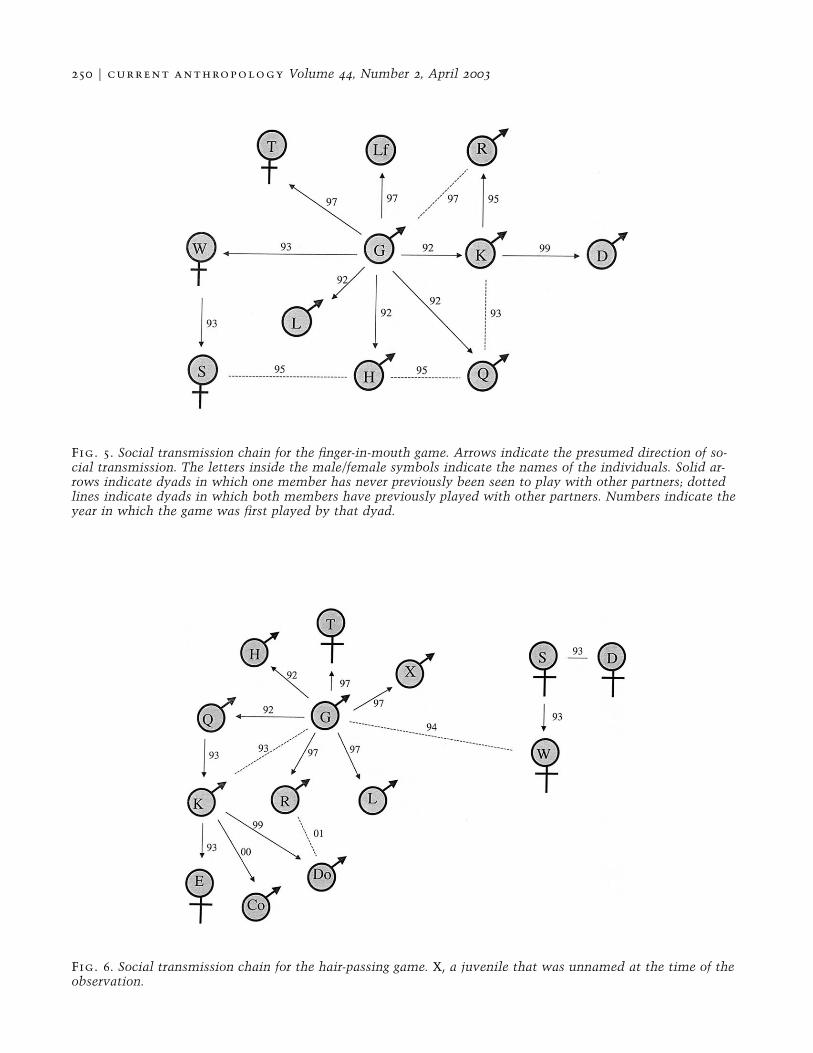
250 F current anthropology Volume 44, Number 2, April 2003
Fig. 5. Social transmission chain for the finger-in-mouth game. Arrows indicate the presumed direction of so-cial transmission. The letters inside the male/female symbols indicate the names of the individuals. Solid ar-rows indicate dyads in which one member has never previously been seen to play with other partners; dottedlines indicate dyads in which both members have previously played with other partners. Numbers indicate theyear in which the game was first played by that dyad.
Fig. 6. Social transmission chain for the hair-passing game. X, a juvenile that was unnamed at the time of theobservation.

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 251
Fig. 7. Social transmission chain for the toy game. Xj, a juvenile that was unnamed at the time of theobservation.
playing subsequently emigrated together, forming an all-male group.
In January 1992, we started seeing a variant of thefinger-in-mouth game that we dubbed the hair-passinggame (fig. 6). In this game, monkey A bites a tuft of hairout of the face or shoulder of monkey B. Then monkeyB attempts to extract the hair from A’s mouth, using thesame techniques described for the finger-in-mouth game.Once monkey B succeeds in recovering the hair, A triesto get the hair back from B’s mouth. The game continues,with A and B reluctantly passing the hair from mouthto mouth, until all the hair has been accidentally swal-lowed or dropped. Then A bites another tuft of hair outof B to start the game anew. Guapo was a member of allhair-passing dyads for the first 13 months in which itwas observed. Then a female-female dyad was observedto play the game without either female’s having playedit with Guapo (to our knowledge)—though one of thesefemales had played similar games with Guapo in thepast. From early 1993 on the hair-passing game wasplayed by other dyads not involving Guapo and was ap-parently transmitted by Guapo’s previous partners tonew monkeys. This game was still being played in1999–2001 by the four males who subsequently emi-grated from the group to form an all-male group together.This game has been observed at no other site. The alphamale of neighboring LB-RA was observed to bite hair outof a juvenile’s face on one occasion, but because the ju-venile did not try to retrieve the hair it was not a full-fledged hair-passing game.
Another variant of the finger-in-mouth game is the toygame. This was the first game to be observed, and it wasfirst seen in June 1991. In this game, monkey A has anonfood object (i.e., a “toy”)—a stick, a leaf, a piece ofbark, or a green or otherwise inedible fruit—in its mouth.Monkey B attempts to pry open the mouth of monkeyA to obtain the toy. Once B successfully removes the toyand puts it in its own mouth, A tries to pry it out of B’smouth. The toy is passed back and forth until it is toomangled to be usable. Neither monkey attempts to eatthe toy. Like the finger-in-mouth and hair-passing games,this game is a quiet, slow-motion play event performedby a dyad, usually on the periphery of the group.
Figure 7 shows the pattern of social transmission forthe toy game. It was less frequently seen than the finger-in-mouth and hair-passing games, but it was spread morewidely across the group. Although Guapo was a partic-ipant in 6 of the initial 7 toy games, 10 of the 22 observedtoy games did not involve him. Like the other games, itseems to have originated with Guapo but was rapidlytransmitted to other monkeys—probably initially to ju-venile males and also to a low-ranking female, who thenplayed the game on three occasions with a high-rankingadult female. A juvenile male (K) who played this gamein 1991 with Guapo was subsequently observed to playit with four other individuals of varying age-sex classes,including the juvenile male R, who was observed in 1999playing it with a very young juvenile. Guapo was notobserved to play this or any other game after he became

252 F current anthropology Volume 44, Number 2, April 2003
table 4Number of Individuals Observed to Play Each Role in the Games
Seen PlayingMouth Role
Seen PlayingRetriever Role
Overlap betweenMouth and
Retriever Role
Mouth RoleRemaining WhenGame Last Played
Retriever RoleRemaining WhenGame Last Played
Overlap betweenMouth and Retriever
Categories in1999–2000
Finger-in-mouth game 8 8 7 4 4 4Hair-passing game 7 9 5 3 5 2Toy game 8 13 8 3 3 3
alpha male in 1999; nor were any previous alpha malesof LB-AB observed playing games.
Baker has observed a slightly modified version of thetoy game at CU-BE. She describes the game as follows:Monkey A sits by monkey B, picks up some object (apiece of bark, leaf, or fruit), and places it in its ownmouth, leaving the lips slightly parted. Monkey B thenuses its fingers and/or mouth to attempt to remove theobject from A’s mouth. They may hold hands, or B mayleave its fingers in A’s mouth. Monkey A sometimesholds B’s hand to prevent transfer of the object. If B suc-cessfully removes the object, A may try to regain it andreinsert it in its mouth. During this game, the monkeysdo not make eye contact but stare off into the distanceas if in a daze. Following this game, the monkeys oftengroom or remain in contact for several minutes with B’sfingers in A’s mouth. This behavior was first observedin 1993, and it was still part of the group’s behavioralrepertoire in 1996. Thirty-eight observations of the be-havior were made over a period of four years (1,920 con-tact hours). Seventeen of these involved juveniles of un-known sexes, but of the remaining 21 observations 3occurred in female-female dyads and 18 involved male-male dyads; all of these observations involved at leastone adult. Thus, the game was never observed to beplayed in opposite-sexed dyads. At least 16 different dy-ads and 14 identifiable individuals (plus some uniden-tified juveniles) were observed to play the game. Thegroup included 20 adults and 14 juveniles, so approxi-mately half of all group members exhibited the behavior.The temporal patterning of the behavior across dyadssuggested that the behavior had been a part of the rep-ertoire well before the 1993 field season began; diversedyads exhibited the behavior in 1993, and the numberof dyads did not increase dramatically over the course ofthe study.
One of the most intriguing aspects of the games wasthe fact that there were two distinct roles in these con-ventions. Because the fact that there are two roles couldbe relevant to the transmission dynamics of the behavior,we decided to investigate the connection between roleplaying and group demography at the time the gamesvanished from the repertoire. Guapo was the most fre-quent player of all three games, and the rate at whichthey were played dropped precipitously when he becamealpha male and stopped playing. Perhaps the failure ofgames to thrive after Guapo’s withdrawal was due to thefact that the remaining players did not know how to
perform both roles. Povinelli, Parks, and Novak (1992)have demonstrated experimentally that role reversal isa challenging cognitive feat for rhesus monkeys, so itseems reasonable to think that capuchins might alsohave difficulty learning both roles. Role reversals werecommon within bouts for some games (27% in the fin-ger-in-mouth game, 48% in the toy game) but relativelyuncommon in the hair-passing game (3%). We will referto the “mouth” role as the role (common to all games)in which the monkey holds the object (finger, hair, ortoy) in its mouth and the “retriever” role as the role inwhich the monkey is trying to remove the object fromthe mouth. The finger-in-mouth game can be initiatedby a monkey playing either the mouth role (by bitingthe partner’s fingers) or the retriever role (by insertingthe fingers in the partner’s mouth), whereas for the othergames the monkey playing the mouth role is the initi-ator. Table 4 shows the number of monkeys observed toplay each role in the three games. The lefthand columnsshow the number of monkeys observed in each role overthe entire study period; the righthand columns show thenumber of game-playing monkeys who were still resid-ing in the group in 1999, when games were last observed(excluding Guapo, who ceased to play games when heassumed alpha rank), or in 2000 in the case of the hair-passing game. Therefore, although only a few individualsdefinitely knew both roles by the time the game-playingtraditions were extinguished, there were at least two perrole for each game, and therefore the disappearance ofthe games from the repertoire cannot be attributed en-tirely to a lack of practitioners of a particular role.
At LB-AB, the most frequent game-playing pairs wereadult male–juvenile male dyads. Dyads that played onegame tended to play other games as well (for details seethe electronic edition of this issue on the journal’s webpage). During the 1991–93 field seasons (for which therichest behavioral data are available), there was a slighttendency for game-playing individuals to spend moretime in close proximity to their game-playing partnersthan to individuals who did not play games with them(Wilcoxon: n p 12, 1.3% time with partners vs. 0.8%time with nonpartners, T p 18, p ! 0.10).
The games meet all of the criteria for a tradition. Twoof the games were observed only in a single group, andone was found in two groups at different sites. All gameswere common (at least during some field seasons) anddurable, remaining in the repertoire for several years, andfor LB-AB it was possible to document a clear expansion

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 253
in the number of performers and to infer that there weretwo or three links in the transmission chain over theyears.
Discussion
As McGrew (2001:235) notes, most interdisciplinary dis-cussions of the evolution of culture center around threeapproaches: (1) Anthropologists most often ask “What isculture?” (which I interpret as meaning “Can we dem-onstrate the presence of social traditions in a given pop-ulation?”) (2) Evolutionary biologists ask “Why is cul-ture?”—in other words, What is the biological signifi-cance of culture? What are the fitness consequences toindividuals exhibiting cultural traits? And why is it moreeffective to have such practices transmitted sociallyrather than genetically? And (3) Psychologists tend toask “How are cultural elements acquired and transmit-ted?” This latter question is too difficult to address withany great sophistication using the methods available infield studies of primates, but we will address the othertwo.
Several behaviors met our criteria for a tradition incapuchins. Whether these collections of traditions qual-ify as “culture” depends on one’s definition of culture.The most commonly used definition of culture in cul-tural primatology is McGrew’s (1992:304) adaptation ofKroeber’s (1928:331) definition. The capuchin data qual-ify according to the first three criteria of this definition(i.e., innovation, transmission, and consistency of form),but the latter three criteria (durability, vertical trans-mission, and spread across social units) are somewhatmore questionable. Thus, the temporal and geographicdistribution of social conventions merits further discus-sion here.
How durable are capuchin social conventions? Someof these traditions (the games) have lasted for as long asten years and have spanned at least one genetic gener-ation and three links in a social transmission chain.Nonetheless, one of the most remarkable things aboutthis dataset is the number of apparent extinctions oftraditions that have been observed in a relatively shorttime. Handsniffing died out after seven years at LB-AB.It died out at SR-CP (reappearing several years later) andalso appears to have vanished from SR-SE. Sucking van-ished from both LB-RA and LB-AB. It appears likely thatthe three games have basically vanished from LB-AB af-ter nine or ten years, since they were observed very rarelyin 1999–2001.
Why do such traditions become extinct so quickly incapuchins? The LB data suggest two possibilities: dem-ographic instability in small groups and, to a far lesserextent, the difficulty of preserving traditions that requirean understanding of two roles. The extinction of hand-sniffing at LB-AB is particularly mysterious because itwas a high-frequency behavior, practiced by all adult fe-males in the group. The decline and disappearance ofhandsniffing may be in part due to the presumed deathsof two avid handsniffers. Sucking was most common at
LB-RA, but a single individual was a participant in 88%of all sucking bouts, despite the fact that 12 other mon-keys participated in sucking and 10 of those took activeroles. Sucking has been seen only three times in the threeyears since the primary sucker’s disappearance. Gameplaying declined precipitously when the presumed in-ventor became alpha male and stopped playing. Addi-tionally, there was a drop in the number of players whoknew both roles (though there were still two to five in-dividuals remaining in the group who definitely kneweach role for each game at the time the games becameextinct). Thus, for all of the social conventions, it appearsthat the disappearance of particularly enthusiastic prac-titioners of a behavior can cause it to vanish from agroup’s repertoire even when several other members ofthe group know how to perform it. Traditions such asthese are particularly fragile in small groups. Demo-graphic changes such as migrations, deaths, and rankreversals pose a major challenge to the stability oftraditions in small groups. If traditions are so readilydisrupted by demographic changes even in capuchins,which have quite low mortality rates (Fedigan, Rose, andAvila 1996) and fairly stable group compositions anddominance hierarchies, it seems likely that they wouldbe even less durable in more demographically fragilespecies.
Why is it that some of these behaviors have appearedat multiple sites when there appears to be no migrationbetween sites today? The geographic patterning ofhandsniffing is difficult to account for. Handsniffing maybe common in one group and completely absent in an-other group with a broadly overlapping home range. Thisfact makes social transmission seem fairly likely andallows ecological differences to be readily ruled out. Yethandsniffing is observed in some groups at three differentsites for which social transmission via migration is im-plausible, which suggests the possibility of independentinvention at each site.
Comparison of the social contexts (i.e., distributionacross age-sex classes) in which handsniffing occurs re-veals extreme differences between groups and also dif-ferences over time (at SR-CP, for example, handsniffingwas observed primarily in male-male dyads in 1986 butprimarily in male-female and female-female dyads in1998). It is possible that the suites of behaviors we arecalling handsniffing and sucking conveyed different so-cial messages in different groups and at different periodsof each group’s history. It seems likely that handsniffingwas independently invented at a number of sites. Thebehavioral elements that make up handsniffing (e.g., put-ting a hand over the nose and mouth, inhaling, stickinga finger up the nose or in the mouth) are already in thebehavioral repertoire in slightly different forms andtherefore readily occur to monkeys (see Huffman andHirata n.d. for similar reasoning regarding the appearanceof similar traditions at multiple Japanese macaque sites).For example, capuchins routinely stick their fingers intosmall holes in the context of extractive foraging for in-sects, and so sticking a finger into another monkey’snostril or mouth probably does not involve a big mental

254 F current anthropology Volume 44, Number 2, April 2003
leap. Likewise, the task of removing a finger, toy, or hairfrom someone else’s mouth involves basically the samemotor skills (prying, digging, pulling, probing) that areroutinely performed in the course of extractive foraging.The restrained biting seen in the games is similar to thatseen in play bouts except that it is of much longer du-ration. Thus, any particular element of these odd behav-ioral patterns is something that would readily occur toa capuchin; all that the monkeys need to do is to assem-ble these elements into chains and induce other indi-viduals to cooperate in the details of performance (e.g.,agree on which parts of the bodies should be involved,decide which roles to play when). With our current data-set, we cannot rigorously address the question of whichsocial learning processes lead to the construction of thesesocial conventions, but the most likely scenario is thatthey come about via a process akin to “ontogenetic rit-ualization” (sensu Tomasello and Call 1997:301; To-masello et al. 1997; Boesch and Tomasello 1998), inwhich the two participants shape one another’s behaviorduring the course of repeated interaction rather than oneanimal’s being the model and the other the learner. Be-cause there are rarely if ever third parties in the im-mediate vicinity when these social conventions are per-formed, it seems unlikely that social learning mechan-isms requiring imitation after observation (as opposed todirect interaction) would be very effective. In chimpan-zees gestural signals have been found to be not readilylearned via imitation; ontogenetic ritualization is farmore effective (Tomasello et al. 1997).
What is the function of these behaviors? To an evo-lutionary biologist attempting to assess the biologicalsignificance of social traditions, the first order of busi-ness is to determine the function of the traits. What aretheir probable fitness consequences, and why is it moreeffective for the trait to be transmitted socially ratherthan genetically? As noted above, it is possible that eachtradition described has a different function—that the be-haviors involved convey different meanings to the mon-keys performing and witnessing them. However, all ofthe behaviors described here have certain features incommon, which may suggest that they are “designed”to communicate similar messages. All of these behaviorsoccur in relaxed social contexts, in which the partici-pants are typically somewhat isolated from the rest ofthe group (perhaps not visually isolated but at least a fewmeters from neighboring monkeys). The participantsmove slowly (which is highly unusual for a capuchin)and have trancelike expressions on their faces, staringout into space or else looking at their fingers (as in theretriever role of the games). Perhaps the most noteworthyfeature that they have in common is that all of theminvolve a certain amount of risk or discomfort to one orboth participants. Handsniffers have one another’s fin-gernails delicately lodged in their nostrils, which re-stricts their movement; tail-sucking and the finger-in-mouth game involve placing a body part between thesharp teeth of the partner (since many capuchins aremissing digits and tail tips, it is reasonable to assumethat this is risky); the hair-passing game involves yank-
ing significant amounts of hair out of the face and shoul-ders, which cannot be very comfortable. Perhaps theseconventions are ways of testing the bonds between in-dividuals, in the same sense in which male savanna ba-boon “diddling” has been proposed to test bonds betweencoalition partners (Smuts and Watanabe 1990).
Zahavi (1977) proposed that stressful stimuli are oftenimposed by one individual (A) on another (B) in order totest the strength of A and B’s social bond. The stressfulsignal is typically something that has a strong sensualcomponent that is perceived by B to be pleasant if thebond is strong or unpleasant if the relationship is notgood. Examples of such a signal would be a French kiss,a child’s using an adult as a climbing toy, or a capuchin’ssticking its fingers up a conspecific’s nose. B’s responseto A’s signal conveys useful information to A about thequality of the relationship: If B tolerates the stressor orresponds positively to it, then this indicates to A that Bis positively disposed to A and presumably likely to besupportive (e.g., when asked for aid in the near future,though this is only one type of possible benefit). If Bresponds negatively, for example, by ignoring A, termi-nating the interaction, or responding aggressively, thenthis indicates to A that B is not currently positively dis-posed toward A and is unlikely to be a reliable source ofbenefits in the near future. The forms of the handsniff-ing, sucking, and game-playing behaviors make themideally suited to be Zahavian bond-testing signals. In or-der to test whether this model is applicable to the be-havioral phenomena we are trying to explain, it wouldbe necessary to have more precise data on the initiationsand terminations of each interaction, baseline data onthe quality of each dyad’s relationship, and data on thedyad’s interactions for a few hours following the inter-action. Regrettably, our datasets do not allow us to testthis hypothesis, because they were designed to answerquite different questions.
Do the culturally variable social signals described forchimpanzees and Japanese macaques exhibit any paral-lels to capuchin social conventions that conform to thisZahavian bond-testing hypothesis? One possibilitycomes to mind: hand-clasp grooming in chimpanzees(McGrew et al. 2001, de Waal and Seres 1997). This is adyadic behavior in which both participants are focusedon grooming one another with one hand while their otherhands are clasped or propping one another up with theirarms extended above their heads. One of the most in-triguing aspects of the patterning of hand-clasp groomingat Mahale is that it is always the subordinate individualwho is supporting the weight of the dominant’s arm(McGrew et al. 2001). It is not clear from the publishedliterature who initiates these interactions, but it is pos-sible that the dominant is using hand-clasps as a test ofthe bond—by imposing the weight of his arm upon thesubordinate he is testing the subordinate’s acknowledg-ment of his higher rank and his willingness to providecontinued political support. Such rank asymmetries arenot apparent in capuchin social conventions, perhaps be-cause capuchins are less obsessed with rank than chim-panzees (and particularly male chimpanzees) are.

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 255
Whereas chimpanzees regularly remind one another oftheir ranks via formalized dominance signals (e.g., thepant grunt, which acknowledges higher rank in the re-ceiver of the signal [de Waal 1982:87]), capuchins do notso clearly advertise rank.
Another body of theory that has potential relevanceto explaining social conventions comes from microso-ciology. Collins (1981, 1993) has proposed that “inter-action rituals” (e.g., human conversations, though vir-tually any type of social interaction would qualify)inform the participants about the status of their rela-tionship with other participants in the interaction. Heassumes that because group-living individuals lack thecomputational capacity to assess their relative compet-itive ability or their place in the constantly fluctuatingpower structure of the group directly, they rely on theemotions generated in interaction rituals in order to es-timate these variables. The form or content of the in-teraction ritual is generally not as important as the emo-tions and corresponding affective displays that aregenerated from them. For example, in terms of the in-teraction’s effect on the quality of that dyad’s future in-teractions, it does not matter whether the content of aconversation was the relative merits of particular foot-ball teams or the weather; nor does it matter whetherthe information conveyed is true. What matters is theamount of enthusiasm, engagement, and coordination/agreement between the human conversational partners;this is what informs them about their willingness tocooperate reliably with one another in the near future.Likewise, it may not be terribly important whether acapuchin sticks its fingers up its partner’s nose or in itsmouth as long as they agree that this is the right thingto do. The real information in the signal comes from theaffective responses of the two partners. On the basis ofCollins’s theory, we would predict that the followingvariables will be positively associated with a dyad’s will-ingness to cooperate in future interactions: duration ofthe current signaling interaction, degree of focus or con-centration on the interaction, and extent of coordinationbetween the participants (i.e., agreement about what el-ements should be included in the task and who shouldplay which role at any given time). Again, our data col-lection protocols did not allow such tests of this model,but future research can perhaps be designed with thesequestions in mind. Collins’s and Zahavi’s models makesome similar predictions about the relevant qualities ofbond-testing interactions. In both types of signals, theaffect of the signal recipient, which tells the signalerabout the current state of the social relationship, is themost critical information. In Zahavi’s model, the cost-liness of the signal (i.e., degree of risk or discomfort tothe recipient) is its most salient feature, whereas in Col-lins’s model the degree of coordination and focus in theinteraction is the most critical feature.
What is the phylogenetic distribution of bond-testingsocial conventions, and why? Another angle to considerwhen discussing the function of the social conventionsthat are the subject of this paper is why similar behaviorshave not often been observed in other primates. What is
remarkable about the invention of these behaviors is notthe motor patterns they exhibit (which, as we note above,are present in other domains of the capuchin behavioralrepertoire) but the extent to which they are coordinatedbetween individuals. Turn taking is quite rare in non-human primates (aside from the contexts of groomingand rough-and-tumble play in some species), and the roleswitching, particularly in the games (which involve theassumption of some fairly novel roles, unlike the morestereotyped behaviors exhibited in grooming and mostrough-and-tumble play), may require cognitive skills thatare lacking or poorly developed in many other primates(e.g., Povinelli, Parks, and Novak 1992). To our knowl-edge, interactions involving role switching are rare inprimates, and thus it is worthwhile to ask why suchinteractions should appear in capuchins but not others.One possibility is that such coordinated interactions pro-vide practice for dyads that need to engage in cooperativeactivities involving complex motor coordination. Onesuch domain of cooperation might be aggressive coali-tions, which are extremely common in capuchins rela-tive to most other primate species. For example, 23% ofconflicts were polyadic during the LB 1997 field season,though at some points in a particular group’s history therate of polyadic conflicts has been as high as 65% (Perry,unpublished data). To put this in comparative perspec-tive, 24% of macaque conflicts are polyadic, 40% ofchimpanzee conflicts are polyadic (de Waal and Harcourt1992), and intragroup coalitionary conflict is essentiallyabsent in the published literature for New World pri-mates outside the genus Cebus. Intragroup coalitionaryconflicts are common both within and between groups,and capuchins also cooperate in defense against predatorsand feeding competitors (Perry n.d.). Cooperation hasalso been documented in tasks requiring motor coordi-nation in captive C. apella (de Waal and Berger 2000). Ifit is the case that turn-taking games in wild capuchinsprovide practice for future cooperative behaviors, thenwe might expect to find social interactions incorporatingrole switching in other species that routinely engage incoalitionary aggression, such as baboons, chimpanzees,macaques, and bottle-nosed dolphins (Harcourt and deWaal 1992). Another domain in which coordination androle reversal might be important is cooperative hunting,as seen in chimpanzees (Boesch 1994a, b; cf. Stanford1995, 1998; Stanford et al. 1994), social carnivores (e.g.,Packer and Ruttan 1988, Scheel and Packer 1991, Stander1992, Kruuk and Turner 1967, Nel 1999), cetaceans(Boran and Heimlich 1999), and perhaps capuchins undervery limited circumstances (Rose 1997). This sort of in-formation is difficult to pull out of the published liter-ature, but some papers suggest that there is an elementof turn taking and role reversal in baboon greetings,which have been suggested to function as communica-tion about future cooperation in coalitionary aggressionfor the purpose of consort takeovers (Smuts and Wata-nabe 1990).
Yet another domain in which cooperation and coor-dination of actions is important is parental care, for mo-nogamous species. Many monogamous species, includ-

256 F current anthropology Volume 44, Number 2, April 2003
ing many birds and also monogamous primates, engagein some form of duetting or joint ritualized display,which occurs not just during courtship but also at manyother times once the pair bond has been formed (seereview by Wachtmeister 2001). At least in some of thesespecies there is considerable complexity, as well as in-terpair variation, in the details of the duetting behavior(Wickler 1980). It has often been proposed (see Wacht-meister 2001 for a history of the debate) that duets andother forms of display in monogamous pairs aid in pairbonding, though the evolutionary logic of why the be-havior should strengthen bonds is typically unclear inthese models. Perhaps such displays, rather thanstrengthening the bond, merely provide current infor-mation about commitment within the pair, which maythen aid the individuals in deciding whether to investmore or less in the relationship. If so, then the degree ofsynchrony and intensity of display should predict theloyalty of the pair when subsequently provided with op-portunities for one partner to desert a mate in favor ofanother. It might also be expected that the frequenciesof such displays would be higher when the pair is in needof information, for example, after separations when onepair member has had opportunities to interact with otherindividuals out of view of the other.
What design features would be optimal for signalsused to test social bonds and practice coordination? Ifthere are indeed signals that function to provide socialpartners with (a) practice in coordinating their actionsor (b) information about emotional engagement/com-mitment as indicated by the attention they pay to co-ordination in a conventional social interaction, the fol-lowing design features may be optimal for such signals:(1) More complex behavioral sequences will be preferableto simple ones because they will require more concen-tration and coordination between partners, thus provid-ing them with more practice and more information aboutemotional engagement. (2) Interactions that involve turntaking and/or role reversal will require more coordina-tion than interactions in which both members have thesame role and perform it simultaneously. (3) Signals thatare flexible and individually idiosyncratic in their form,rather than rigidly stereotyped, will require more con-centration and coordination; partners will have to fa-miliarize themselves with one another’s quirks and ad-just their own behavior slightly to produce an interactionthat is mutually satisfying. Taking the time to do thiswill provide the partners with coordination practice andsimultaneously provide them with the information thatthere is mutual interest in interacting. It is easy to seehow mutual adjustments of idiosyncratic behaviorsmight eventually produce social traditions in social con-ventions among networks of individuals that regularlyengage in cooperative activities.
It is important to emphasize that flexible, idiosyn-cratic signals are expected to emerge only in dyads thatare already somewhat familiar with one another andhave some degree of trust and predictability. Such flex-ible signals will not be optimal for negotiation of rela-tionships in dyads that are in the initial stages of rela-
tionship formation, when the participants are uncertainof one another’s fighting ability, tendency toward ag-gression, etc. Most species that have cooperative long-term relationships already have some more stereotypedsignals (e.g., grooming, various vocal signals, noncon-ceptive sex) for communicating about the status of theirsocial relationships and about their intentions in the nearfuture. Stereotyped signals are easier to interpret thanmore flexible ones and therefore more valuable/reliablefor interpreting the intentions of social partners amongdyads that are just becoming acquainted. Flexible signalslike the ones described in this paper may provide richerinformation about the cooperative potential of a dyad,but they will be too costly to use in the early stages ofrelationship formation because of the potential formisinterpretation.
How might functional hypotheses for capuchin socialconventions be tested? Current data-sets are inadequateto test these hypotheses about the functions of tradi-tional social conventions in capuchins in any sophisti-cated way. However, there is some weak evidence thatsuch behaviors are performed among dyads that havefairly solid social bonds: game players spent slightlymore time with their game-playing partners than withother animals, and handsniffing rates were correlatedwith grooming rates at LB (Perry 1996). If game playingis associated with the development or testing of coop-erative relationships, we might expect an increase ingame playing during periods of social instability or justprior to rank reversals, when monkeys need allies themost. The rates of game playing at LB-AB (as crudelymeasured by number of ad libitum observations per hourof contact with the group) varied from year to year. Inthe first phase of the project (1991–92), group member-ship was completely stable, and there were no rank re-versals. During this phase, games were played at a rateof 1.8 bouts per 100 hours. During the six months fol-lowing a rank reversal and male immigration, there wastremendous social upheaval accompanied by heightenedrates of both aggression and affiliation (Perry 1998b); dur-ing this phase, the game-playing rate was 2.7 bouts/100hours. In the 1997 field season, there was no social up-heaval (though the alpha male was unpopular and waskilled by his groupmates—including some of Guapo’sgame-playing partners—the following year [Gros-Louis,unpublished data]); during this season, games wereplayed at a rate of 3.3 bouts/100 hours. Group member-ship and ranks were stable in the 1999 field season, whenGuapo was alpha male, and then game playing occurredat a rate of 1.0 bout/100 hours. These data are quitedifficult to interpret because of the challenges involvedin operationalizing social instability and accurately es-timating rates of rarely performed behaviors and becauseof the numerous confounding factors, but it is possiblethat games are played more frequently in periods of socialupheaval (or just preceding major social upheaval). It isalso worth noting that the only four game players tocontinue game playing after Guapo’s withdrawal sub-sequently emigrated together and continued to playgames in an all-male group. Also, finger-sucking (another

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 257
putative bond-testing signal) was exceedingly commonin the pair of males at SR that migrated from group togroup together. Given the high risks associated with mi-gration and takeovers of new groups, it is likely thatmales who choose to migrate together have fairly solidbonds. No doubt even those members of male-male dy-ads that feel comfortable with one another will need totest their bonds frequently as the males stage take-overs of new groups because of the potential for shiftingalliances in the new social context.
To summarize our working hypothesis about the prob-able function of the interactions described in this paper:We suspect that such interactions share the function ofnegotiating the status of social relationships. This maytake the form of practicing coordination and/or using theinteractions to obtain information about partners’ cur-rent disposition toward one another. The precise motordetails of the interactions are expected to be arbitraryand variable between groups or social networks; the util-ity of these interactions is in the coordination of behav-ioral elements between partners. Because such interac-tions have their origins in individually idiosyncraticbehaviors, which are presumably modified over thecourse of time by coordination with other individualswho are members of a social network, traditions are ex-pected to emerge. Because the motor details are fairlyarbitrary, the precise form of these traditions is not ex-pected to be particularly stable, and traditions of this sortare expected to dissolve (or be replaced by similartraditions) when key individuals vanish, causing disrup-tion of social networks. If our hypothesis is correct, thenwe might expect to find similar traditions in other spe-cies that have well-developed cooperative relationshipsand signals that are apparently designed to communicateabout these relationships.
Is the durability of a tradition necessarily a good in-dicator of its biological significance? When thinkingabout the biological importance of traditions, it is im-portant to maintain a conceptual distinction between thebehavior itself and the psychological machinery that isresponsible for producing it. Not all behaviors that areculturally transmitted are necessarily fitness-enhancing(Richerson and Boyd 1989, Boyd and Richerson 1992);nonetheless, the psychological machinery enabling so-cial learning (possibly including particular perceptualskills, emotional proclivities facilitating close associa-tions with others and attentiveness to conspecifics’ ac-tions, and perhaps imitative abilities) will be adaptiveeven if it sometimes causes animals to acquire mal-adaptive behaviors as long as its net effect is to enableacquisition of fitness-enhancing behaviors. One issuethat has long persisted in the study of culture in non-humans (see, e.g., McGrew 1992) is whether the longev-ity of a social tradition is correlated with its biologicalimportance. Some scholars would insist that verticaltransmission (i.e., transmission across generations) is anessential feature of culture and that behaviors remainingin a population’s behavioral repertoire for shorter timeperiods are of lesser biological significance and shouldbe denied the lable of “culture” or “tradition” and re-
labeled “fad.” I will leave aside the question of labels forthe moment and address the question of biological im-portance. For most evolutionary biologists, the biologicalimportance of a trait is measured in terms of fitness con-sequences for the individual bearing the trait. Accordingto this framework, would capuchin social conventionsbe biologically important? Regrettably, we lack data onthe fitness consequences of handsniffing, game playing,etc., and it will be difficult to obtain accurate estimatesof the fitness consequences of practicing any particularsocial convention for any species. Therefore, for the timebeing we will resort to the far less satisfying option ofdoing thought experiments such as the following:
A human couple may have many “couples rituals”that are signals to them of their mutual devotion (e.g.,being ready with a towel when the other is ready to getout of the shower or reciting lines from their mutualfavorite movie on every possible socially appropriate oc-casion). Such behaviors would not seem to have any greatfitness-enhancing value on the surface, but the socialcontext and ontogenetic ritualization process in whichthey were created are what gives them their importance(i.e., they are used to signal commitment to a particularsocial relationship). Consequently, failure to perform oneof these rituals when it is expected can initiate the break-down of a valuable social relationship by signaling in-attentiveness to the partner. The particular behaviors areimportant only in the context of this particular socialrelationship, and the particular behaviors used to signalcommitment may be fairly arbitrary—the members ofthis particular couple may not use these same signals intheir subsequent romantic relationships, though theywill produce other “couples rituals.” Thus, the precisedetails of the particular behaviors involved are fairly ar-bitrary, and the behaviors may be short-lived, but thepropensity to invent and adopt these interaction ritualsmay have great adaptive value and should be the targetof natural selection.
This example deals with two-person “cultures,” andsome anthopologists will object that the term “culture”is more often applied at the population level or at thevery least to social networks larger than two individuals.However, the logic above applies to larger groups as wellas to couples. Whenever there is a situation in whichthere is a need for large coalitions, there should be se-lection also for interaction rituals that help define groupmembership and locate individuals in the coalitionarystructure. The more complex and multilayered the coal-itional structure in the society, the more need thereshould be for such signaling and hence the greater se-lective pressure to invent and acquire from others thesignals used in these interaction rituals. We would pre-dict that dolphins and hamadryas baboons, with theirsupercoalitions and multitiered societies (Connor,Smolker, and Richards 1992, Kummer 1995), might ex-hibit such behaviors. And humans, of course, have themost complex layering of coalitionary networks knownin the animal kingdom, even participating in coalitionswith people they do not know personally.
Conclusion. Despite the fact that so many traits

258 F current anthropology Volume 44, Number 2, April 2003
deemed to be “cultural” in humans fall in the domainof social conventions rather than subsistence technology,most theorizing about the evolution of culture has fo-cused on the technological domain. This is perhaps inpart because technological culture leaves material re-mains that can be traced over long periods of time. Itmay also be because it is far easier to compare the effi-ciencies of alternative technologies than it is to estimatefitness costs and benefits of particular social conven-tions. Nonetheless, the evolution of social conventionsis a fascinating topic that is crying out for a better com-parative database. It is our hope that researchers of othersocial, cooperative animals will begin to collect data onthis issue so that we can better assess the phylogeneticdistribution of such behaviors and the circumstances un-der which they arise. We also hope to stimulate research-ers of human populations to attend to the form (i.e., “sig-nal design”), transmission dynamics, social contexts, andontogeny of social conventions so that there can be morecross-disciplinary exchange on this topic.
Comments
r . l . day, i . coolen, y. van bergen,and k. n . lalandSub-Department of Animal Behaviour, University ofCambridge, Cambridge CB3 8AA, U.K. (Day, Coolen,van Bergen)/ Centre for Social Learning and CognitiveEvolution, School of Biology, St. Andrews University,St. Andrews, Fife KY16 9JU, U.K. (Laland) ([email protected]). 18 x 02
Perry and colleagues are to be congratulated for this val-uable collaborative exercise, which we regard as not onlya milestone in the study of animal traditions but also acontribution to a deeper understanding of the roots ofhuman culture. As they suggest, the study of animalculture has mostly focused on the technological domain,yet much human culture is based on social conventions.Animal conventions are important not just because theyhave been neglected but because, unlike behaviour inthe technological domain, they are not easily explainedaway by differences in ecology (e.g., Tomasello 1994). Weshare the authors’ hope that others will be encouragedto design studies specifically with social conventions inmind in order to create a comparative dataset that goesbeyond apes. It is important that anthropologists beaware of the extent to which the behaviour of a multi-tude of animals, including fish, birds, and mammals, isshaped by socially transmitted traditions. “Culture” canbe regarded as the exclusive domain of humans only ifit is defined according to arbitrary and anthropocentriccriteria.
Yet readers of CA will be well aware of the problemsassociated with defining culture, and to date the animalliterature has been no more successful than its humancounterpart in establishing consensual criteria by which
culture can be classified and analysed. Purported definingcriteria (or necessary conditions) for animal culture in-clude interpopulation variation in behaviour, differencesbetween groups that cannot be attributed to genetic orecological variation, socially transmitted information,transgenerational traditions, universality, uniformity,behavioural innovation, cumulative traditions, teaching,imitation, and others (Tomasello 1994, McGrew 1998,Whiten et al. 1999). Because this is not the place to enterinto a detailed discussion of the merits of such criteria,we will concentrate on two points on which we feel wecan make constructive suggestions.
The first concerns the “durability” criterion empha-sized by Perry et al., which implies that investigationsof animal culture require long-term studies. While weappreciate the value of long-term field observation, weare concerned that this emphasis on durability may becounterproductive. A vast amount of human culture istransient, fickle, and spreads horizontally, but it is noless cultural for that. The same is likely to be true foranimals (Laland, Richerson, and Boyd 1993), and inter-esting cultural phenomena in capuchins and other spe-cies might be neglected if the durability criterion werestrictly applied. Indeed, it is debatable to what extent thecapuchin data genuinely meet the arbitrary six-monthcriterion. Furthermore, short-term studies can contrib-ute to the study of culture too, particularly through con-trolled experimentation. For instance, features of socialtransmission can be investigated using transmission-chain procedures in which the passage of learned behav-iour is tracked as the composition of experimental groupschanges. This design has been successfully applied to fish(Laland and Williams 1997), birds (Curio, Ernst, andVieth 1995, Zeltner, Klein, and Huber-Eicher 2000), andrats (Galef and Allen 1995). While such procedures maybe more difficult with primates, they are surely not im-possible. Ultimately, experimentation provides the onlytruly adequate means of satisfying critics that a partic-ular animal “has culture.”
Second, the data presented by Perry et al. resonate withcurrent thinking on animal innovation. Reader and La-land (2001) invoke the adage “Necessity is the motherof invention” to highlight the fact that poor competitors,such as subordinates, frequently appear to be driven toinnovate while good competitors can satisfice usingtried-and-tested strategies. We note that the innovatorsamongst the capuchins were typically subordinates andthat the innovator, Guapo, ceased performing the novelbehaviour once he achieved alpha status. In species suchas capuchins the alpha male may dominate reproductiveand foraging success, forcing subordinate males to in-novate in order to gain necessary resources. The capu-chin conventions tend to occur on the periphery of thegroup, where presumably low-status individuals will befound. Moreover, it appears that all of the capuchin socialconventions were initiated by males, an observation thatalso corresponds to the findings of a recent survey of theprimate literature (Reader and Laland 2001). Given thegreater variability in male than in female mating success,it may pay low-status males to innovate to gain re-

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 259
sources. Because capuchin females are philopatric, mi-grant males may be expected to be innovative in thesocial domain, as they must establish their social posi-tion in order to achieve alpha status and form coalitionsto avoid fatal conflicts.
We also note that Guapo was the inventor of severalsocial conventions, which brings to mind the multipleinnovations of Imo, the celebrated Japanese macaque. Nodoubt similar plaudits will be landed on Guapo, but wepropose a note of caution. It has never been establishedthat Imo’s achievements are a result of her “innovativepersonality” (as opposed to more mundane motivationalfactors), and even if it were, it is not clear that the rel-evant personality trait is “intelligence” or “creativity”as opposed to, say, “boldness,” “perseverance,” or “non-conformity.” Imo’s fame has led to the widespread butprobably false impression that primate innovators arelikely to be juvenile and female (Reader and Laland 2001).The strength of the kind of comparative analysis carriedout by Perry et al. is that researchers can go beyond themisleading impressions that can emerge from a singlepopulation.
william c. mc grewDepartments of Anthropology and Zoology, MiamiUniversity, Oxford, Ohio 45056, U.S.A. ([email protected]). 24 ix 02
Researchers on white-faced capuchin monkeys in CostaRica have produced an impressive comparative analysisof a set of behavioral patterns that seem to qualify associal conventions. They have done so rigorously andsystematically, by collaboration, collating data-sets fromfour sites. In doing so, they have advanced the emergingfield of cultural primatology beyond any previous cross-populational study, including those on great apes(Whiten et al. 1999). Their results are all the more no-table because these New World primates are phyloge-netically distant from Homo sapiens, showing that ar-gument by analogy can be as fruitful as argument byhomology. This is not always appreciated by anthropol-ogists, who when being comparative tend to focus ongreat apes.
Perry et al. stress that they deal only with social pat-terns, not with subsistence ones such as the elementarytechnology of foraging. Although the former have beenless commonly analyzed, they were the subject of thevery first cross-populational study in primates, that ofthe chimpanzees’ social custom of the grooming hand-clasp (McGrew and Tutin 1978). Obviously, there ismuch more to be done in the social domain (e.g., oncourtship, display, etc.).
The authors rightly emphasize explicit operational cri-teria for what qualifies in their analysis, but all three aredebatable. Variation across groups is likely a sufficientcondition but not a necessary one. One can imagine astudy of a single population’s revealing fascinating cus-toms. Expansion is more problematic, as this is surely amatter of timing, that is, when the observations begin
vis-a-vis the origins of the behavioral pattern. Early ob-servations may yield rapid expansion in the number ofperformers, while later ones may show no expansion ifthe pattern is established. Durability seems to be a mat-ter of “horses for courses.” Students of fads in popularculture would not like to be constrained chronologically,any more than students of traditional culture would behappy with a time-scale that is less than transgener-ational.
The ethnography is fascinating in its richness, but onelooks for common elements underlying the patterns. Inaddition to those proposed by the authors, a commonstrand seems to be that of one individual’s constrainingthe activities of another. This suggests dominance-sub-ordination relations and the possibility that the conven-tional performances not only test or reinforce relation-ships but also signal them to others. (The same mayapply to the grooming hand-clasp of chimpanzees. Whenthe subordinate supports the weight of the joint effort,this is obvious for all to see [McGrew et al. 2001].) Thisidea can be tested by looking more closely at who is oris not the “audience” for behavioral performance.
The distinction between complementarity, turn tak-ing, and role reversal in such conventional interactionsis heuristically useful, especially in going beyond dyadicto polyadic exchanges. This is the stuff of social com-plexity. Perry et al.’s study sets new standards for futureanalyses in cultural primatology.
el isabetta visalberghi and elsa addess iIstituto di Psicologia, Consiglio Nazionale delleRicerche, via U. Aldrovandi 16 b, 00197 Rome, Italy([email protected]). 3 x 02
Do capuchins’ “social conventions” function to test thequality of the relationship between group members, assuggested by Perry and coworkers? Do social conven-tions provide useful information for deciding how to be-have towards group members? We argue that if this kindof information were necessary, then social conventionswould be present most of the time and performed/re-ceived by most individuals, but this is not the case forcapuchins. Social conventions are rarely observed andconfined to a few individuals even though there is noreason to think that the pairs in which they occur arethe only ones in need of testing their bonds, and theirhigh rate of extinction makes them short-lived. Fur-thermore, why do Perry et al. consider grooming andnonconceptive sex to be “stereotyped signals” not flex-ible enough to perform the function of testing bonds?This position is puzzling to those of us who observecapuchins’ behavior in the laboratory, where we havefound a high rate of innovation and marked interindi-vidual behavioral differences in a closely related species(Cebus apella) (e.g., sexual behavior [Carosi and Visal-berghi 2002] and tool use [Visalberghi and Trinca 1989]).Other less demanding explanations may account for theoccurrence of social conventions. For example, it is pos-sible that the smell of someone else’s hands, being in

260 F current anthropology Volume 44, Number 2, April 2003
very intimate contact, or being involved in a social ob-ject-play situation provides pleasurable feedback that re-wards both partners. In short, to attribute a bond-testingfunction to social conventions seems unwarranted to us,and to persevere in this direction might turn out to bea Sisyphean effort.
Field (and laboratory [see below]) experiments provideinsights for interpreting culture and social traditions. Onthe one hand, ecological differences between field sitesshould be carefully scrutinized when examining a can-didate case of cultural behavior; on the other hand, it isalmost impossible not to underestimate the ecologicaldifferences among sites. An enlightening example of howecological differences may affect cultural behaviorcomes from recent field experiments on chimpanzees’ant-dipping at Bossou. Humle and Matsuzawa (2002)found that prey behavior (i.e., gregariousness and/or ag-gressiveness) and the ant-dipping context (e.g., whetherthe ants are at the nest or migrating) influence the lengthof the tool and the technique used to dip. Therefore,differences between sites in prey-species behavior andthe context in which dipping takes place may result indifferent tool lengths and ant-dipping techniques.
Perry et al. limit their analyses to social conventions,disregarding the food-related behaviors that are also fun-damental to human cultures (Rozin 1996). Laboratoryresearch may provide insights in this domain. We ex-amined the widespread assumption that observationallearning from experienced group members is quicker andsafer than individual learning as a way of overcomingneophobia (i.e., caution about eating something nevertasted before) (see Visalberghi and Addessi n.d.). If thiswere the case, influences provided by the social contextcould lead to safe feeding only if the target food en-countered by naıve individuals and the food eaten byknowledgeable group members were the same. Tuftedcapuchin monkeys (Cebus apella), a species closely re-lated to C. capucinus, were tested to assess the extentto which social influences on feeding behavior foster asafe diet. Using an observer-demonstrator(s) paradigm inwhich a naıve observer and experienced group member(s)could see each other through a Plexiglas window, wefound that the observers ate significantly more of a novelfood when demonstrators were eating by the windowthan when alone. Surprisingly, however, this occurredregardless of what food the demonstrators were eating(Visalberghi and Addessi 2000, 2001). Moreover, whenthe observer could choose between two colored novelfoods only one of which matched the food being eatenby the demonstrators, it did not eat the matching foodmore than the other (Visalberghi and Addessi 2001). Inshort, in tufted capuchins social influences were not di-rected to a specific food target, and the behavior of groupmembers was not used to assess, without risk, whethera novel food was safe. In another experiment we assessedthe effect of social (encountering novel foods with groupmembers) or non-social (encountering the same novelfoods when alone) treatment on individuals’ preferencesfor the novel foods presented in a binary-choice test andfound that whereas feedback from nutrients and expo-
sure affected food preferences, the treatment did not (Sab-batini, Stammati, and Visalberghi n.d.). These findingsshould caution field observers against assuming that so-phisticated social learning processes underlie feedinghabits. Instead, we should consider how very simplemechanisms bias individual learning (Fragaszy and Vis-alberghi 2001).
andrew whitenCentre for Social Learning and Cognitive Evolutionand Scottish Primate Research Group, School ofPsychology, University of St. Andrews, St. AndrewsKY16 9JU, Scotland ([email protected]). 7 x 02
Perry et al. have here assembled a very special and im-portant contribution to our knowledge of traditionswithin the animal kingdom. In particular, their impres-sively detailed documentation supports their main hy-pothesis that the behaviour patterns they describe areindeed traditions defined by their social content and alsotheir supplementary hypothesis that the function ofthese particular behaviours lies in social bond-testing.Neither phenomenon has been been so well supportedbefore for non-human species, and accordingly this newdatabase should have a significant impact on our con-ceptual map of the subject. The successful collaborativeeffort of these researchers provides a model of what canbe achieved by this route as decades of research offerincreasing opportunities to construct the large-scale per-spective required for a “cultural primatology” (de Waal1999, Whiten 2003). It is also encouraging to see suchan excellent example of true discovery—data that, what-ever the issues of interpretation that remain to be de-bated, can fairly straightforwardly be described as a clus-ter of exciting new facts about behaviour, contrastingwith the disappointingly sterile theoretical and meth-odological hand-wringing of so much contemporary ef-fort in mainstream “cultural anthropology.”
In the context of these celebratory comments, I havejust two main critical comments to make. The first con-cerns inconsistencies in the analysis of criteria for iden-tifying traditions under natural conditions. Perry et al.say that the exclusion of genetic and ecological differ-ences is not “necessary or sufficient,” but this is alsotrue of their own three operational criteria—intergroupvariation, expansion, and durability. A stable traditioncertainly need not meet the first two requirements;equally, none of the three is sufficient to guarantee thata tradition exists. Isn’t it rather the case that the threecriteria of Perry et al. plus the other two provide a setof criteria that offer stronger circumstantial evidence fortraditions the more of them are met? For the five be-haviour patterns they highlight, is there in fact evidencethat appeals to all five criteria?
Second, can “ontogenetic ritualization” really explainthe social transmission implied? Ontogenetic ritualiza-tion, as described by Tomasello and Call (1997), is meantto explain how an action of animal A towards animal B(e.g., arm raise) that initially has no communicative in-

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 261
tent comes to have such intent through anticipatorylearning (e.g., animal A eventually raises its arm inten-tionally to elicit play after both B and A have come toanticipate that this often leads to play). To my knowl-edge, evidence that could prove that such a process op-erated in a particular instance of behavioural develop-ment has not yet been forthcoming. But in any case,ontogenetic ritualization cannot explain how the criticalaspect of reciprocation is acquired. For example, wheremonkey A begins the finger-in-mouth game by clampingits teeth onto B’s fingers, how does B acquire the recip-rocal “idea” of clamping its own teeth onto A’s fingers?Or how does A get to the reciprocal idea of getting B’sfingers in its nose, given that A has its fingers in B’snose? Using the principles of ontogenetic ritualizationalone, it is not clear that this leap can be made. To thecontrary, the reciprocation leap appears an inherentlyimitative one, sophisticated insofar as, in the case ofteeth-clamping, monkey B has to translate from havingits fingers clamped in A’s mouth, which it can feel butperhaps not see (?), to clamping A’s fingers in its ownmouth, which it certainly cannot see.
Two sets of recent findings may be worth bearing inmind in tackling the mystery that I am arguing still ex-ists here. First, there is now evidence that Neotropicalprimates, including capuchins, are capable of more im-itative social learning than the reviews of Visalberghiand Fragaszy (1990, 2002) would suggest (Voelkl andHuber 1999, Fredman and Whiten 2002). Second, thecharacteristics of “mirror neurons” in monkeys suggestthat perceptions of others’ actions are coded motoricallyor at some supramodal level (e.g., Rizzolatti et al. 2002),a phenomenon that might help to explain how the kindof “reciprocation leap” discussed above could beachieved by a monkey. To be sure, the mirror neuronsdescribed to date are all concerned with linkage of ac-tions with matching visual input (e.g., the neuron fireswhen one performs a precision grip or when one seesanother monkey doing this). Is it possible that othersexist for touch, such that the brain of a monkey thatfeels its fingers clamped in another’s mouth registers thisin a neural representation of clamping the other’s fingersin its own mouth? Whatever the answer to such spec-ulations, we would be wise not to underestimate thedifficulty of explaining the social learning and diffusionof reciprocal conventions.
amotz zahaviInstitute of Nature Conservation Research, Tel-AvivUniversity, 69978 Tel-Aviv, Israel ([email protected]). 14 xi 02
Perry et al. present Collins’s theory and mine (Zahavi1977) as different, but they are not. My theory suggestedthat the strength of the bond may be assessed by im-posing a test on a social partner. The information ac-quired by the test reflects the strength of the bond andhelps one to decide whether to maintain the bond orbreak it. Collins’s suggestion that interaction rituals
function to inform participants about the social rela-tionship between them is not essentially different. Weagree that what strengthens the bond is not the socialinteraction itself but the information acquired throughthat interaction—that is, that although many bonds arestrengthened as a consequence of the interaction, someare disrupted. Collins further suggested that it is changein emotions that affects the bond, while I only discussedthe logic of the patterns of the interaction by which theinformation is acquired. The decision to break a bond orimprove it may be a consequence of one’s emotionalstate, a cognitive decision, or some other mechanism.
I do not think that there is a functional difference be-tween a genetically acquired ritual and one which is de-veloped as a culture. Many birds, fishes, and reptileslearn to assess the danger presented by predators or con-specific rivals in a particular place and vary in their re-action to them by fright or mobbing according to thecircumstances. Learning to understand the readiness ofothers to bond, which is learning to assess a positiveresponse, need not be different from learning to avoid aparticular species or an individual predator. I thereforeexpect that detailed studies of social interactions of otheranimals besides primates may reveal learned and pos-sibly cultural differences in the patterning of their socialrituals.
The cooperative birds that I am studying, Arabian bab-blers (Turdoides squamiceps), are faced with social prob-lems similar to those of cebus monkeys. They have de-veloped analogous social interactions; they allopreen,they play games (Posis 1984), they clump, and they havesocial communal dancing (Ostreiher 1995, Zahavi andZahavi 1997). There are quantitative and sometimesqualitative differences in the way in which they interactsocially. Some groups mob certain predators while othersdo not, and some groups allopreen or dance more thanothers (Zahavi, unpublished observations). Posis ob-served social plays that were invented in a particulargroup and were absent in others. I would not be surprisedif part of that variation were found to be cultural ratherthan innate.
Reply
susan perryLeipzig, Germany. 25 xi 02
Because my many collaborators are currently scatteredaround the world, busy with fieldwork or end-of-termteaching commitments, I am unable to coordinate a jointreply in time to meet the deadline. I will therefore takethe liberty of responding myself.1 I thank Zahavi for ex-plaining his perspective on the ways in which his and
1. I thank Joe Manson and Clark Barrett for comments on thismanuscript and the Max Planck Institute for Evolutionary Anthro-pology for support of my research.

262 F current anthropology Volume 44, Number 2, April 2003
Collins’s theories articulate and for confirming the valueof using this theoretical approach for cultural as well as“genetically acquired” rituals. His comment that suchsocial conventions are likely to be present also in thecooperative Arabian babblers he studies is encouraging,and we hope that researchers will investigate this pos-sibility further. I also thank McGrew, Whiten, and Dayet al. for their cogent critiques of the various methodo-logical issues involved in identifying traditions and inpinpointing the mechanisms by which social transmis-sion can occur. The commentators’ criticisms fall intofour categories.
Operational criteria for identifying traditions. Day etal., McGrew, and Whiten comment on our operationalcriteria for identifying traditions. As we state in the ar-ticle, these were intended primarily as “rules of thumbthat are designed to detect traditions with a minimumof error in a messy collection of datasets,” and this setof criteria is neither necessary nor sufficient to pinpointsocial learning as the mechanism for the spread of thetrait in question. To the extent that this set of criteriaproduces errors in the identification of traditions, it islikely to be overly conservative (i.e., to overlook truetraditions). As Whiten points out, despite the fact thatneither my three criteria nor his “non-genetic” and“non-ecological” criteria are necessary or sufficient con-ditions for attribution of social learning, the more ofthese criteria that a behavior meets, the greater the like-lihood that it is a true tradition.
McGrew objects that intergroup variation is not nec-essary to conclude that a behavior is “traditional.” In asense, I agree: for example, if a behavior is highly adap-tive, multiple groups are likely to converge on the samebehavior even if social learning is pivotal in its acqui-sition (as in the case of response to particular predators).But for the class of behaviors which we term social con-ventions (as defined in this article), I see no reason toexpect such a precise convergence of form across groups;in fact, a certain degree of arbitrariness of form is to beexpected. Hence, if all groups exhibit the same behavior,then there are serious grounds for suspicion that the be-havior is part of the species-typical repertoire and sociallearning is unlikely to be required for its acquisition. Butperhaps what McGrew is envisioning is a long-termstudy of a single population in which, after many yearsof study, a new social convention appears, is propagated,and perhaps eventually vanishes. Under such circum-stances, I would agree with him that there may be suf-ficient grounds to term this a tradition in the absence ofdata on interpopulation variation.
I agree with McGrew that the “expansion of perform-ers” criterion is a difficult one, since the detection of itdepends very much on the timing of the study relativeto the timing of the introduction of the trait. Certainlyinsistence on this criterion will produce some “false neg-atives” in the identification of traditions (as was almostcertainly the case for sucking of body parts). However, Ido think that attention to the process of social trans-mission is extremely important and is something that is
sorely lacking in much of the published literature onanimal traditions to date.
Durability is, as I said in the article, the most arbitraryof our criteria. I agree with Day et al. that even a brieffad can be termed “culture” if it is spread via social learn-ing. My main reason for including durability, in com-bination with frequency and intragroup distribution ofthe trait, was to prevent the false attribution of a tra-dition to a group in which a particular behavior was aone-time experiment by a single individual or dyad. Asit happens, there were no candidates for traditions thatwere both frequent across multiple individuals and veryshort-term, so I do not think that we mislabeled any truetraditions as nontraditions by employing the six-monthcriterion.
I sympathize also with Day et al.’s concern that I amoveremphasizing the necessity of long-term field obser-vations. Certainly I agree that field experiments (whichcan be accomplished in relatively short time frames)have much to contribute to the study of social learning.Field experiments are, of course, more tractable in somespecies than in others (e.g., they work extremely well infish and rodents and less well in species such as primatesthat have highly differentiated social relationships,strong social bonds, and sometimes extreme xenophobiathat makes translocation experiments ethically ques-tionable). For some behavioral domains, such as foodchoice, reactions to predators, or possibly even food pro-cessing, it should be possible to introduce an experi-mental apparatus (e.g., a feeding platform with novelfoods, a model predator, or a novel food-processing chal-lenge such as Whiten’s artificial fruits) and then docu-ment the transmission pattern of the relevant behaviorsfrom the beginning. It is difficult to imagine, however,how such an experiment could be implemented for socialconventions. How could wild animals be prompted byhuman experimenters to produce a novel social signal?Time-consuming as it is, the best way to see whethercapuchins transmit their favorite social conventions toother groups is to wait until males migrate, follow themto their new groups, and pray that the migrants do notget killed in the process of migrating as has happened totwo practitioners of the games described in this paper.The two remaining practitioners of the games have fi-nally succeeded in migrating, and we are now waitingto see whether they will transmit the games to their newgroupmates once their relationships with them havestabilized.
Are they testing bonds? Visalberghi and Addessi dis-courage the attribution of any function to social con-ventions because they suspect that the reason capuchinsare performing the behavior is to get pleasurable feed-back. Here they are confusing proximate and ultimateexplanations for behavior, making the all too commonmistake of claiming that if a proximate explanation islikely to be true, then ultimate explanations have beenfalsified. We do not dispute that the monkeys might getpleasurable feedback from the performance of these be-haviors (at least, when the quality of their relationshipsis good), but acknowledging that the experience is pleas-

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 263
urable begs the question why it is pleasurable for somebut not all capuchins, bringing us back to the necessityof asking about the behavior’s evolved function. NikoTinbergen (1963), one of the founders of ethology, co-gently described the four complementary levels at whichbehavior can be explained: function, cause, development,and evolution (or, to use the terms favored by more re-cent writers, ultimate, proximate, developmental, andphylogenetic). Explaining a behavior at one level doesnot eliminate the need to explain it at other levels. Afull understanding of the evolutionary biology of a be-havior must take all four levels of explanation into ac-count, and this is true for behaviors deemed to be tra-ditional as well as for others (Fragaszy and Perry n.d.).Of course, some traits have no function and are merelyby-products of other traits, and traits may acquire sec-ondary functions, but the refusal even to test hypothesesabout adaptive functions will set the field back ratherthan advancing understanding (Alcock 2001).
I am just as puzzled by Visalberghi and Addessi’s claimthat a functional explanation is consistent only with be-havioral uniformity as I am by their claim that a prox-imate explanation such as “It feels good” would ade-quately explain a pattern that exhibits considerableinterindividual variation. First let us consider their gen-eral claim that if social conventions were useful theywould be present in most or all individuals most of thetime. Surely they would not dispute the fact that thereare numerous human conventions (e.g., marriage cere-monies, swearing-in ceremonies of parliamentarians,and authority-related rituals) that, although they clearlyprovide information about how to behave toward groupmembers, are performed as infrequently as once in a life-time and perhaps only by a small subset of the popula-tion. Regarding capuchin social conventions specifically,their proposed proximate explanation gives us no way ofgenerating hypotheses about the observed variation. Whyare these behaviors seen in only some primate species,in only some dyads, and only at some times? Why aren’tthey equally pleasurable to all dyads of all species at alltimes? To explain a behavior by saying that it is pleas-urable is tautological: “They do it because it’s pleasur-able, and we know it’s pleasurable because we see themdo it.” Operationalizing an independent measure ofpleasure sounds even more Sisyphean than testing thefunctional explanation that we have proposed.
Integration of multiple levels of analysis is clearly theroute to a productive research program for the study ofsignaling about social relationships, and Joseph Mansonand I have already launched a research program designedto test many of the theoretical issues raised in this paper.Our more recent datasets involve all-day follows of in-dividual monkeys in which we meticulously record in-itiations and terminations of interactions as well as sub-tleties of posture and arousal levels. Such data, incombination with our baseline data on the quality ofeach dyad’s relationship as relationship quality shiftsover time, allow us to examine each dyad’s interactionsfor several hours preceding and following putative bond-testing signals so that we can more accurately test hy-
potheses generated by Zahavi’s and Collins’s theoriesand hypotheses about the assessment of competitiveability (e.g., Silk 1994) and low-cost signals of benignintent (Silk, Kaldor, and Boyd 2000). The capuchin vocaland gestural repertoire is rich in signals that appear tofunction to negotiate aspects of social relationships.These range from signals such as grooming, which havea long phylogenetic history, are highly stereotyped, andthus appear to be readily interpreted by the recipients ofthe action, to the social conventions described in thispaper, which are idiosyncratic, more complex, and lessreadily interpreted because they are not part of the spe-cies-typical repertoire and most likely involve sensationsof both pleasure and pain. As we have said, even if bothgrooming and these more novel and flexible social con-ventions serve approximately the same function—providing information about the quality of a bond—they still differ in important ways such as interpretabil-ity and signal reliability and therefore are not inter-changeable signals. Easily misinterpreted signals such ashandsniffing or hair-biting games should not be used inearly stages of relationship formation, because they maytrigger severe aggression if the recipient does not under-stand the signaler’s intent. However, these idiosyncraticsocial conventions may provide more reliable (i.e., moredifficult to fake) signals of commitment to a relationshipthan grooming does because they require more trust,more focus in each interaction, and willingness to spenda lot of time working out the details of who does whatto whom such that the interaction is mutually satisfying.In the past few months we have been documenting theinitial stages of some new social conventions in LB-AB,and it is apparent that much time and effort go intoconstructing a convention for which both members ofthe dyad are content to perform their roles.
As is the case with C. apella, the C. capucinus sexualrepertoire is rich, involving numerous vocalizations andgestures which can be combined in various ways, al-though each behavioral element is stereotyped and partof the species-typical repertoire. Visalberghi and Addessimake the claim that there is a high rate of innovationin female sexual behavior in C. apella, but I do not seethis variation (which consists essentially of interindi-vidual differences in the frequencies of performing thesame stereotyped species-typical behavioral elements[Carosi and Visalberghi 2002]) as being highly innovative.At any rate, if we are to understand the role that thesebehaviors play in negotiating relationships (either in co-ordinating a mating or in bond testing, as in nonconcep-tive sexual interactions), we must understand the role ofeach element separately. At Lomas Barbudal, our prelim-inary analyses of the frequencies of signal types acrossdyads with different demographic characteristics haverevealed consistent patterns across dyads as to whichbehaviors emerge in which social contexts. The dance(pacing, spinning, piloerection, and direct gaze) is com-mon to courtship and relationship negotiation in dyadsof all stages of relationship formation. However, the vo-calizations used differ according to the stage of relation-ship formation, reproductive state, and the current

264 F current anthropology Volume 44, Number 2, April 2003
amount of stress on the social relationship. Likewise,the outcome of the dance (i.e., the likelihood that it willend in mounting and thrusting and other physical-con-tact sexual gestures) depends on the stage of relationshipformation, recent events in the dyad’s interaction his-tory, and the amount of pressure the dyad is receivingfrom audience members trying to break up the dance.Analysis of the new datasets (the all-day follows, whichprovide more microcontextual information) is currentlyunder way to pinpoint the types of information that eachsignal is likely to provide the animals and the conse-quences for the dyad of performing these behaviors. Atany rate, it is clear that different types of social signals,by virtue of their differing design features and phyloge-netic histories, serve different purposes. No single rela-tionship negotiation signal, whether it is stereotyped ortraditional, will be appropriate for all dyads in all con-texts, and hence it is not reasonable to assume that socialconventions do not provide useful information for guid-ing future interactions simply because of a lack of uni-formity in the expression of the behavior across dyadsand across time, as Visalberghi and Addessi suggest. So-cial relationships are dynamic. This is why they need tobe tested.
McGrew suggests that, rather than (or in addition to)signaling to each other about their relationship quality,the members of the dyad are using the convention as adisplay by which they signal their relationship quality(not only rank relations but also perhaps commitmentto an alliance) to other members of the group. I had con-sidered this line of reasoning myself in trying to makesense of the patterns I was seeing but ended up rejectingit. If the purpose of the signal is to advertise somethingto other group members, it should be conspicuous andperformed in front of an audience. Neither of these con-ditions is true: the behaviors are performed on the pe-riphery of the group and are very quiet (in contrast withthe more stereotyped wheeze dance, which may verywell serve to announce relationship status and inviteattempts to break up alliances [Perry 1995]). Even whenother monkeys are near the performers of the social con-vention, they show no interest at all in the behavior,which suggests that it is not providing them with usefulinformation.
Are capuchin conventions transmitted via imitation?Although I initially doubted that imitation was involvedin the acquisition of capuchin conventions because ofthe difficulty Tomasello et al. (1997) had in getting chim-panzees to imitate gestures, I find Whiten’s argumentsquite persuasive. It really is difficult to see how the re-ciprocal element of these gestures could arise via onto-genetic ritualization alone, and the recent data on theimitative capacities of brown capuchins and callitrichidsmake Whiten’s imitation interpretation all the moreplausible. Unfortunately, I cannot conceive of a way toaddress this issue rigorously using observational fielddata.
Who innovates? I agree with Day et al., as well asReader and Laland (2001), that “necessity is the motherof invention.” Nonetheless, this tendency is better sup-
ported in some behavioral domains (e.g., food choice andfood-processing techniques) than in others (e.g., socialconventions). First of all, it is somewhat less clear thatsubordinates need allies more than dominants do. Sub-ordinates need allies to increase their rank, but domi-nants need them to maintain their rank; it is hard to saywhich is more important. Also, I would like to correctthe impression that it is just low-rankers and males whoengaged in social conventions in the capuchin dataset.Guapo, though he was not the alpha male at the time ofhis innovations, was nonetheless tied for the second rankin the group. Also, handsniffing was invented and prac-ticed by females, many of them high-ranking, in LB-ABgroup and in SR-CP group. I am intrigued by the hy-pothesis that capuchin males will be more innovative inthe social domain than females because of their greaterneed to form coalitions. Certainly males’ social rela-tionships are more dynamic than those of females, suchthat frequent testing of the bonds would be desirable,and the consequences of having no allies are more dire(i.e., eviction from groups, severe wounding and killingby male conspecifics). At the same time, males, relativeto females, more rarely reach a stage of relationship for-mation that is secure enough to risk employing thesemore flexible and intimate bond-testing signals.
References Citeda l c o c k , j o h n . 2001. The triumph of sociobiology. Oxford:
Oxford University Press.a l t m a n n , j . 1974. Observational study of behavior: Sampling
methods. Behaviour 49:227–67.b o e s c h , c h r i s t o p h e . 1994a. Chimpanzees–red colobus
monkeys: A predator-prey system. Animal Behaviour 47:1135–48.
———. 1994b. Cooperative hunting in wild chimpanzees. Ani-mal Behaviour 48:653–67.
———. 1996a. “The emergence of cultures among wild chimpan-zees,” in Evolution of social behaviour patterns in primatesand man. Edited by W. G. Runciman, John Maynard Smith,and R. I. M. Dunbar, pp. 251–68. Proceedings of the BritishAcademy 88.
———. 1996b. “Three approaches for assessing chimpanzee cul-ture,” in Reaching into thought: The minds of the great apes.Edited by A. E. Russon, K. Bard, and S. T. Parker, pp. 404–29.Cambridge: Cambridge University Press.
b o e s c h , c h r i s t o p h e , a n d m i c h a e l t o m a s e l l o .1998. Chimpanzee and human cultures. current anthropol-ogy 39:591–614.
b o i n s k i , s . 1988. Use of a club by a wild white-faced capu-chin (Cebus capucinus) to attack a venomous snake (Bothropsasper). American Journal of Primatology 14:177–79.
b o r a n , j . r . , a n d s . l . h e i m l i c h . 1999. “Social learn-ing in cetaceans: Hunting, hearing, and hierarchies,” in Mam-malian social learning: Comparative and ecological perspec-tives. Edited by H. O. Box and K. R. Gibson, pp. 282–307.Cambridge: Cambridge University Press.
b o y d , ro b e r t , a n d p e t e r j . r i c h e r s o n . 1985. Cul-ture and the evolutionary process. Chicago: University of Chi-cago Press.
———. 1992. Punishment allows the evolution of cooperation (oranything else) in sizeable groups. Ethology and Sociobiology13:171–95.

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 265
———. 1996. “Why culture is common, but cultural evolution israre,” in Evolution of social behaviour patterns in primatesand man. Edited by W. G. Runciman, John Maynard Smith,and R. I. M. Dunbar, pp. 77–93. Proceedings of the BritishAcademy 88.
b y r n e , r i c h a r d . 1995. The thinking ape: Evolutionary ori-gins of intelligence. Oxford: Oxford University Press.
———. 1999. “Cognition in great ape ecology: Skill-learning abil-ity opens up foraging opportunities,” in Mammalian sociallearning: Comparative and ecological perspectives. Edited byH. O. Box and K. R. Gibson, pp. 333–50. Cambridge: Cam-bridge University Press.
c a ro s i , m o n i c a , a n d e l i s a b e t t a v i s a l b e r g h i .2002. Analysis of tufted capuchin (Cebus apella) courtship andsexual behavior repertoire: Changes throughout the female cy-cle and female inter-individual differences. American Journalof Physical Anthropology 118:11–24. [ev, ea]
c h a p m a n , c . , a n d l . f e d i g a n . 1990. Dietary differencesbetween neighboring Cebus capucinus groups: Local traditions,food availability, or responses to food profitability? Folia Pri-matologica 54:177–86.
c o l l i n s , r . 1981. On the microfoundations of macrosociol-ogy. American Journal of Sociology 86:984–1014.
———. 1993. Emotional energy as the common denominator ofrational action. Rationality and Society 5:203–30.
c o n n o r , r i c h a r d c . , r a c h e l a . s m o l k e r , a n d a n -d r e w f . r i c h a r d s . 1992. “Dolphin alliances and coali-tions,” in Coalitions and alliances in humans and other ani-mals. Edited by Alexander H. Harcourt and Frans B. M. deWaal, pp. 415–43. Oxford: Oxford Science Publications.
c o u s s i - k o r b e l , s . , a n d d . f r a g a s z y. 1995. On the re-lation between social dynamics and social learning. AnimalBehaviour 50:1441–53.
c u r i o , e . , u . e r n s t , a n d w. v i e t h . 1995. The adaptivesignificance of avian mobbing. 2. Cultural transmission of en-emy recognition in blackbirds: Effectiveness and some con-straints. Zeitschrift fur Tierpsychologie 48:184–202. [rld, ic,yv, knl]
c u s t a n c e , d e b o r a h , a n d r e w w h i t e n , a n d t a m a rf r e d m a n . 1999. Social learning of an artificial fruit task incapuchin monkeys (Cebus apella). Journal of Comparative Psy-chology 113:13–23.
d e w a a l , f . b . m . 1982. Chimpanzee politics. New York:Harper and Row.
———. 1997. Food transfers through mesh in brown capuchins.Journal of Comparative Psychology 11:370–78.
———. 1999. Cultural primatology comes of age. Nature 399:635–36.
———. 2000. Attitudinal reciprocity in food sharing amongbrown capuchin monkeys. Animal Behaviour 60:253–61.
d e w a a l , f r a n s b . m . , a n d m i c h e l l e l . b e r g e r .2000. Payment for labour in monkeys. Nature 404:563.
d e w a a l , f r a n s b . m . , a n d a l e x a n d e r h . h a r -c o u r t . 1992. “Coalitions and alliances: A history of etholog-ical research,” in Coalitions and alliances in humans andother animals. Edited by Alexander H. Harcourt and Frans B.M. de Waal, pp. 1–19. Oxford: Oxford Science Publications.
d e w a a l , f r a n s b . m . , a n d m i c h a e l s e r e s . 1997.Propagation of handclasp grooming among captive chimpan-zees. American Journal of Primatology 43:339–46.
d e w a r , g w e n . n.d. “The cost-benefit approach to socialtransmission: Designing tests for adaptive traditions,” in Thebiology of traditions: Models and evidence. Edited by DorothyFragaszy and Susan Perry. Cambridge: Cambridge UniversityPress. In press.
d u g a t k i n , l . 1996. “Copying and mate choice,” in Sociallearning in animals: The roots of culture. Edited by Cecilia M.Heyes and Bennett G. Galef, pp. 85–106. San Diego: AcademicPress.
f e d i g a n , l . m . 1990. Vertebrate predation in Cebus capuci-nus: Meat eating in a neotropical monkey. Folia Primatologica54:196–205.
———. 1993. Sex differences and intersexual relations in adult
white-faced capuchins (Cebus capucinus). International Jour-nal of Primatology 14:853–77.
f e d i g a n , l . m . , a n d k . m . j a c k . 2001. Neotropical pri-mates in a regenerating Costa Rican dry forest: A comparisonof howler and capuchin populations. International Journal ofPrimatology 22:689–713.
f e d i g a n , l i n d a m . , l . m . ro s e , a n d r . m . a v i l a .1996. “See how they grow: Tracking capuchin monkey popula-tions in a regenerating Costa Rican dry forest,” in Adaptive ra-diations of Neotropical primates. Edited by M. Norconck, A.Rosenberger, and P. Garber, pp. 289–307. New York: PlenumPress.
f r a g a s z y, d o ro t h y, a n d s u s a n p e r ry. Editors n.d.a.The biology of traditions: Models and evidence. Cambridge:Cambridge University Press. In Press.
———. n.d.b. “Towards a biology of traditions,” in The biologyof traditions: Models and evidence. Edited by Dorothy Fra-gaszy and Susan Perry. Cambridge: Cambridge UniversityPress. In press.
f r a g a s z y, d . , a n d e . v i s a l b e r g h i . 1996. “Social learn-ing in monkeys: Primate ‘primacy’ reconsidered.” in Sociallearning in animals: The roots of culture. Edited by Cecilia M.Heyes and Bennett G. Galef Jr., pp. 65–84. San Diego: Aca-demic Press.
———. 2001. Recognizing a swan: Socially-biased learning. Psy-chologia 44:82–98. [ev, ea]
f r a g a s z y, d o ro t h y m . , e . v i s a l b e r g h i , a n d j . g .ro b i n s o n . 1990. Variability and adaptability in the genusCebus. Folia Primatologica 54:114–18.
f r e d m a n , t a m a r , a n d a n d r e w w h i t e n . 2002. Thenature and function of observational learning of tool use bycapuchin monkeys (Cebus apella) in relation to human andconspecific models. Poster prepared for the conference “Per-spectives on Imitation,” Royaumont Abbey, France, April. [aw]
f r e e s e , c . h . 1976. Food habits of white-faced capuchins Ce-bus capucinus L. (Primates: Cebidae) in Santa Rosa NationalPark, Costa Rica. Brenesia 10/11:43–56.
f r e e s e , c . h . , a n d j . r . o p p e n h e i m e r . 1981. “The cap-uchin monkey, genus Cebus,” in Ecology and behaviour ofNeotropical primates, vol. 1. Edited by A. F. Coimbra-Filho andR. H. Mittermeier, pp. 331–90. Rio de Janeiro: AcademiaBrasilia.
g a l e f , b e n n e t t g . , j r . 1996. “Social enhancement of foodpreferences in Norway rats: A brief review,” in Social learningin animals: The roots of culture. Edited by Cecilia M. Heyesand Bennett G. Galef Jr., pp. 49–64. San Diego: AcademicPress.
———. n.d. “ ‘Traditional’ foraging behaviors of brown and blackrats (Rattus rattus and Rattus norvegicus),” in The biology oftraditions: Models and evidence. Edited by Dorothy Fragaszyand Susan Perry. Cambridge: Cambridge University Press. Inpress.
g a l e f , b . g . , j r . , a n d c . a l l e n . 1995. A model systemfor studying animal traditions. Animal Behaviour 50:705–17.[rld, ic, yv, knl]
g a l e f , b e n n e t t g . , j r . , a n d l u c - a l a i n g i r a l -d e a u . 2001. Social influences on foraging in vertebrates:Causal mechanisms and adaptive functions. Animal Behaviour61:3–15.
g i r a l d e a u , l u c - a l a i n , a n d t h o m a s c a r a c o . 2000.Social foraging theory. Princeton: Princeton University Press.
h a r c o u r t , a . h . 1995. Sexual selection and sperm competi-tion in primates: What are male genitalia good for? Evolution-ary Anthropology 4:121–29.
h a r c o u r t , a . h . , a n d f . b . m . d e w a a l . Editors.1992. Coalitions and alliances in humans and other animals.Oxford: Oxford University Press.
h a rv e y, p . , r . d . m a r t i n , a n d t . h . c l u t t o n -b ro c k . 1987. “Life histories in comparative perspective,” inPrimate societies. Edited by B. B. Smuts, D. L. Cheney, R. M.Seyfarth, R. W. Wrangham, and T. T. Struhsaker, pp. 181–96.Chicago: University of Chicago Press.
h e y e s , c e c i l i a m . , a n d b e n n e t t g . g a l e f j r . Edi-

266 F current anthropology Volume 44, Number 2, April 2003
tors. 1996. Social learning in animals: The roots of culture.San Diego: Academic Press.
h e y e s , c e c i l i a , a n d l u d w i g h u b e r . Editors. 2000. Theevolution of cognition. Cambridge: MIT Press.
h u d s o n , r . , b . s c h a a l , a n d a . b i l k o . 1999. “Trans-mission of olfactory information from mother to young in theEuropean rabbit,” in Mammalian social learning: Comparativeand ecological perspectives. Edited by H. O. Box and K. R.Gibson, pp. 1441–57. Cambridge: Cambridge University Press.
h u f f m a n , m i c h a e l , a n d y. h i r a t a . n.d. “Biologicaland ecological foundations of primate behavioral traditions,” inThe biology of traditions: Models and evidence. Edited by Do-rothy Fragaszy and Susan Perry. Cambridge: Cambridge Uni-versity Press. In press.
h u m l e , t a t y a n a , a n d m a t s u z a w a t e t s u ro . 2002.Chimpanzees, safari ants, tools, and technique: A new perspec-tive. Paper presented at the 19th Congress of the InternationalPrimatological Society, Beijing, China, August 4–9. [ev, ea]
i t a n i , j . 1958. “On the acquisition and propagation of a newfood habit in the troop of Japanese monkeys at Takasakiyama,”in Japanese monkeys: A collection of translations. Edited by K.Imanishi and S. Altmann, pp. 52–65. Edmonton: University ofAlberta Press.
i t a n i , j . , a n d a . n i s h i m u r a . 1973. “The study of infra-human culture in Japan,” in Symposia of the Fourth Interna-tional Congress of Primatology, vol. 1. Edited by E. W. MenzelJr., pp. 26–60. Basel: Karger.
j a n i k , v i n c e n t m . , a n d p e t e r j . s l a t e r . n.d.“Traditions in mammalian and avian vocal communication,”in The biology of traditions: Models and evidence. Edited byDorothy Fragaszy and Susan Perry. Cambridge: Cambridge Uni-versity Press. In press.
k a w a i , m . 1965. Newly acquired pre-cultural behavior of anatural troop of Japanese monkeys on Koshima Island. Pri-mates 6:1–30.
k a w a m u r a , s . 1959. The process of sub-human culture prop-agation among Japanese macaques. Primates 2:43–60.
k i n g , b a r b a r a j . 1991. Social information transfer in mon-keys, apes, and hominids. Yearbook of Physical Anthropology34:97–115.
———. 1994. The information continuum: Evolution of social in-formation transfer in monkeys, apes, and hominids. Santa Fe:SAR Press.
k i t c h e n e r , a . c . 1999. “Watch with mother: A review of so-cial learning in the Felidae,” in Mammalian social learning:Comparative and ecological perspectives. Edited by H. O. Boxand K. R. Gibson, pp. 236–58. Cambridge: Cambridge Univer-sity Press.
k ro e b e r , a . l . 1928. Sub-human culture beginnings. Quar-terly Review of Biology 3:325–42.
k ru u k , h . , a n d m . t u r n e r . 1967. Comparative notes onpredation by lion, leopard, cheetah, and wild dog in the Seren-geti area, East Africa. Mammalia 31:1–27.
k u m m e r , h a n s . 1995. In quest of the sacred baboon: A sci-entist’s journey. Princeton: Princeton University Press.
l a l a n d , k . 1996. “Developing a theory of animal social learn-ing,” in Social learning in animals: The roots of culture. Ed-ited by Cecilia M. Heyes and Bennett G. Galef Jr., pp. 129–54.New York: Academic Press.
———. 1999. “Exploring the dynamics of social transmissionwith rats,” in Mammalian social learning: Comparative andecological perspectives. Edited by in H. O. Box and K. R. Gib-son, pp. 174–87. Cambridge: Cambridge University Press.
l a l a n d , k . n . , p . j . r i c h e r s o n , a n d r . b o y d . 1993.Animal social learning: Towards a new theoretical approach.Perspectives in Ethology 10:249–77. [rld, ic, yv, knl]
l a l a n d , k . n . , a n d k . w i l l i a m s . 1997. Shoaling gener-ates social learning of foraging information in guppies. AnimalBehaviour 53:1161–69. [rld, ic, yv, knl]
l e f e b v r e , l o u i s , a n d l u c - a l a i n g i r a l d e a u . 1996.“Is social learning an adaptive specialization?” in Social learn-ing in animals: The roots of culture. Edited by Cecilia M.
Heyes and Bennett G. Galef Jr., pp. 107–28. New York: Aca-demic Press.
m c g r e w, w. c . 1992. Chimpanzee material culture: Impli-cations for human evolution. Cambridge: Cambridge Univer-sity Press.
———. 1998. Culture in non-human primates? Annual Review ofAnthropology 27:301–28.
———. 2001. “The nature of culture: Prospects and pitfalls ofcultural primatology,” in Tree of origin: What primate behav-ior can tell us about human social evolution. Edited by Fransde Waal, pp. 231–54. Cambridge: Harvard University Press.
m c g r e w, w. c . , l . f . m a r c h a n t , s . e . s c o t t , a n dc . e . g . t u t i n . 2001. Intergroup differences in a social cus-tom of wild chimpanzees: The grooming hand-clasp of the Ma-hale Mountains. current anthropology 42:148–53.
m c g r e w, w. c . , a n d c . t u t i n . 1978. Evidence for a so-cial custom in wild chimpanzees? Man 13:234–51.
m a n s o n , j . h . 1999. Infant handling in wild Cebus capuci-nus: Testing bonds between females? Animal Behaviour 57:911–21.
m a n s o n , j . h . , s . e . p e r ry, a n d a . r . p a r i s h . 1997.Nonconceptive sexual behavior in bonobos and capuchins. In-ternational Journal of Primatology 18:767–86.
m a n s o n , j . h . , l . ro s e , s . p e r ry, a n d j . g ro s -l o u i s . 1999. Dynamics of female-female social relationshipsin wild Cebus capucinus: Data from two Costa Rican sites. In-ternational Journal of Primatology 20:679–706.
m a t s u z a w a , t . 1994. “Field experiments on use of stonetools by chimpanzees in the wild,” in Chimpanzee cultures.Edited by R. W. Wrangham, W. C. McGrew, F. B. M. de Waal,and P. G. Heltne, pp. 351–70. Cambridge: Harvard UniversityPress.
m a t s u z a w a , t . , a n d g . y a m a k o s h i . 1996. “Compari-son of chimpanzee material culture between Bossou andNimba, West Africa,” in Reaching into thought: The minds ofthe great apes. Edited by A. E. Russon, K. Bard, and S. T. Par-ker, pp. 211–34. Cambridge: Cambridge University Press.
n a k a m u r a , m i c h i o , w i l l i a m c . m c g r e w, l i n d af . m a r c h a n t , a n d t o s h i s a d a n i s h i d a . 2000. Socialscratch: Another custom in wild chimpanzees? Primates 41:237–48.
n e l , j a n a . j . 1999. “Social learning in canids: An ecologicalapproach,” in Mammalian social learning: Comparative andecological perspectives. Edited by H. O. Box and K. R. Gibson,pp. 259–78. Cambridge: Cambridge University Press.
n i c h o l s o n , b . 1984. Does kissing aid human bonding by se-miochemical addiction? British Journal of Dermatology 111:623–27.
o ’ b r i e n , t . g . , a n d j . g . ro b i n s o n . 1991. Allomater-nal care by female wedge-capped capuchin monkeys: Effects ofage, rank, and relatedness. Behaviour 119:30–50.
o s t r e i h e r , r . 1995. Influence of the observer on the fre-quency of the “morning dance” in the Arabian babbler. Ethol-ogy 100:320–30. [az]
o t t o n i , e d u a r d o b . , a n d m a s s i m o m a n n u . 2001.Semi-free-ranging tufted capuchin monkeys (Cebus apella)spontaneously use tools to crack open nuts. International Jour-nal of Primatology 22:347–58.
p a c k e r , c . , a n d l . ru t t a n . 1988. The evolution of coop-erative hunting. American Naturalist 132:159–98.
p a n g e r , m e l i s s a . 1997. Hand preference and object-use infree-ranging white-faced capuchins (Cebus capucinus) in CostaRica. Ann Arbor: University Microfilms International.
p a n g e r , m e l i s s a a . , s u s a n p e r ry, l i s a ro s e , j u l i eg ro s - l o u i s , e r i n v o g e l , k a t h e r i n e c . m a c k i n -n o n , a n d m a ry b a k e r . 2002. Cross-site differences inthe foraging behavior of white-faced capuchins (Cebus capuci-nus). American Journal of Physical Anthropology. 119:52–66
p e r ry, s u s a n . 1995. Social relationships in wild white-facedcapuchin monkeys, Cebus capucinus. Ann Arbor: UniversityMicrofilms International.
———. 1996. Female-female social relationships in wild white-

perry et al . Social Conventions in Wild White-faced Capuchin Monkeys F 267
faced capuchin monkeys, Cebus capucinus. American Journalof Primatology 40:167–82.
———. 1997. Male-female social relationships in wild white-faced capuchin monkeys, Cebus capucinus. Behaviour 134:477–510.
———. 1998a. Male-male social relationships in wild white-facedcapuchins, Cebus capucinus. Behaviour 135:1–34.
———. 1998b. A case report of a male rank reversal in a group ofwild white-faced capuchins (Cebus capucinus). Primates 39:51–69.
———. n.d. “Coalitionary aggression in white-faced capuchins,”in Animal social complexity: Intelligence, culture, and indi-vidualized societies. Edited by F. B. M. de Waal and P. Tyack.Cambridge: Harvard University Press. In press.
p e r ry, s . , a n d l . ro s e . 1994. Begging and transfer of coatimeat by white-faced capuchin monkeys, Cebus capucinus. Pri-mates 35:409–15.
p o s i s , o . 1984. Social play in babblers (in Hebrew with Eng-lish summary). M.Sc. thesis, Tel-Aviv University, Tel-Aviv,Israel.
p o v i n e l l i , d a n i e l j . , k a t h l e e n a . p a r k s , a n dm e l i n d a a . n o v a k . 1992. Role reversal by rhesus mon-keys, but no evidence of empathy. Animal Behaviour 44:269–81.
r e a d e r , s i m o n . n.d. “Relative brain size and the distributionof innovation and social learning across the nonhuman pri-mates,” in The biology of traditions: Models and evidence. Ed-ited by Dorothy Fragaszy and Susan Perry. Cambridge: Cam-bridge University Press. In press.
r e a d e r , s . m . , a n d k . n . l a l a n d . 2001. Primate inno-vation: Sex, age and social rank differences. International Jour-nal of Primatology 22:787–806. [rld, ic, yv, knl]
r i c h e r s o n , p e t e r j . , a n d ro b e r t b o y d . 1989. Therole of evolved predispositions in cultural evolution, or, Hu-man sociobiology meets Pascal’s Wager. Ethology and Socio-biology 10:195–219.
r i z z o l a t t i , g i a c o m o , l u c i a n o f a d i g a , l e o n a r d of o g a s s i , a n d v i t t o r i o g a l l e s e . 2002. “From mirrorneurons to imitation: Facts and speculations,” in The imitativemind. Edited by Andrew Meltzoff and Wolfgang Prinz, pp.247–66. Cambridge: Cambridge University Press. [aw]
ro s e , l . 1992. Benefits and costs of resident males to femalesin white-faced capuchins, Cebus capucinus. American Journalof Primatology 32:235–48.
———. 1997. Vertebrate predation and food sharing in Cebus andPan. International Journal of Primatology 18:727–65.
ro s s , c . 1988. The intrinsic rate of natural increase and repro-ductive effort in primates. Journal of Zoology 241:199–219.
ro z i n , p . 1996. “Sociocultural influences on human food se-lection,” in Why we eat what we eat: The psychology of eat-ing. Edited by E. D. Capaldi, pp. 233–63. Washington, D.C.:American Psychological Association. [ev, ea]
ru s s o n , a . 1996. “Imitation in everyday use: Matching andrehearsal in the spontaneous imitation of rehabilitant orangu-tans (Pongo pygmaeus),” in Reaching into thought: The mindsof the great apes. Edited by A. E. Russon, K. Bard, and S. T.Parker, pp. 152–76. Cambridge: Cambridge University Press.
ru s s o n , a n n e e . , k i m a . b a r d , a n d s u e t a y l o rp a r k e r . 1996. Reaching into thought: The minds of the greatapes. Cambridge: Cambridge University Press.
s a b b a t i n i , g l o r i a , m a r g h e r i t a s t a m m a t i , a n de l i s a b e t t a v i s a l b e r g h i . n.d. Social influences on pref-erences towards novel foods in tufted capuchin monkeys. FoliaPrimatologica. In press. [ev, ea]
s c h e e l , d . , a n d c . p a c k e r . 1991. Group hunting behav-ior of lions: A search for cooperation. Animal Behaviour 41:697–709.
s c h e u r e r , j . , a n d b . t h i e r ry. 1985. A further food-washing tradition in Japanese macaques (Macaca fuscata). Pri-mates 26:491–94.
s i l k , j o a n b . 1994. Social relationships of male bonnet ma-caques: Male bonding in a matrilineal society. Behaviour 130:
271–91.s i l k , j o a n b . , e . k a l d o r , a n d r . b o y d . 2000. Cheap
talk when interests conflict. Animal Behaviour 59:423–32.s m u t s , b a r b a r a b . , a n d j o h n m . w a t a n a b e . 1990.
Social relationships and ritualized greetings in adult male ba-boons (Papio cynocephalus anubis). International Journal ofPrimatology 11:147–72.
s t a n d e r , p . e . 1992. Co-operative hunting in lions: The roleof the individual. Behavioral Ecology and Sociobiology 29:445–54.
s t a n f o r d , c r a i g b . 1995. The influence of chimpanzee pre-dation on group size and anti-predator behaviour in red colobusmonkeys. Animal Behaviour 49:577–87.
———. 1998. Chimpanzee and red colobus: The ecology of pred-ator and prey. Cambridge: Harvard University Press.
s t a n f o r d , c r a i g b . , j a n e t t e w a l l i s , e s l o mm p o n g o , a n d j a n e g o o d a l l . 1994. Hunting decisionsin wild chimpanzees. Behaviour 131:1–18.
s t e p h a n , h . , g . b a ro n , a n d h . d . f r a h m . 1988.“Comparative size of brains and brain components,” in Com-parative primate biology, vol. 4. Edited by H. D. Steklis and J.Erwin, pp. 1–39. New York: Liss.
t a n a k a , i . 1995. Matrilineal distribution of louse egg-handlingtechniques during grooming in free-ranging Japanese macaques.American Journal of Physical Anthropology 98:197–201.
———. 1998. Social diffusion of modified louse egg-handlingtechniques during grooming in free-ranging Japanese macaques.Animal Behaviour 56:1229–1236.
t e r b o r g h , j . 1983. Five New World primates: A study incomparative ecology. Princeton: Princeton University Press.
t e r k e l , j . 1996. “Cultural transmission of feeding behaviourin the black rat (Rattus rattus),” in Social learning in animals:The roots of culture. Edited by Cecilia M. Heyes and BennettG. Galef Jr., pp. 17–48. San Diego: Academic Press.
t i n b e r g e n , n . 1963. On aims and methods of ethology. Zeit-schrift fur Tierpsychologie 20:410–33.
t o m a s e l l o , m . 1994. “The question of chimpanzee culture,”in Chimpanzee cultures. Edited by R. W. Wrangham, W. C.McGrew, F. B. M. de Waal, and P. G. Heltne, pp. 301–17. Cam-bridge: Harvard University Press. [rld, ic, yv, knl]
t o m a s e l l o , m i c h a e l , a n d j o s e p c a l l . 1997. Primatecognition. Oxford: Oxford University Press.
t o m a s e l l o , m i c h a e l , j o s e p c a l l , j e n n i f e r w a r -r e n , g . t h o m a s f ro s t , m a l i n d a c a r p e n t e r , a n dk a t h e r i n e n a g e l l . 1997. The ontogeny of chimpanzeegestural signals: A comparison across groups and generations.Evolution of Communication 1:223–59.
u r b a n i , b e r n a r d o . 1999. Spontaneous use of tools bywedge-capped capuchin monkeys (Cebus olivaceus). Folia Pri-matologica 70:172–74.
v a n s c h a i k , c . p . , r . o . d e a n e r , a n d m . y. m e r -r i l l . 1999. The conditions for tool use in primates: Implica-tions for the evolution of material culture. Journal of HumanEvolution 36:719–41.
v a n s c h a i k , c . p . , e . a . f o x , a n d a . f . s i t o m p u l .1996. Manufacture and use of tools in wild Sumatran orang-utans. Naturwissenschaften 83:186–88.
v i s a l b e r g h i , e . 1990. Tool use in Cebus. Folia Primatolo-gica 54:146–54.
v i s a l b e r g h i , e l i s a b e t t a , a n d e l s a a d d e s s i . 2000.Seeing group members eating a familiar food enhances the ac-ceptance of novel foods in capuchin monkeys. Animal Behav-iour 60:69–76.
———. 2001. Acceptance of novel foods in Cebus apella: Do spe-cific social facilitation and visual stimulus enhancement play arole? Animal Behaviour 62:567–76.
———. n.d. “Food for thoughts: Social learning and the feedingbehavior in capuchin monkeys, insights from the laboratory,”in Traditions in non-human animals: Models and evidence.Edited by D. Fragaszy and S. Perry. Cambridge: CambridgeUniversity Press. In press.
v i s a l b e r g h i , e l i s a b e t t a , a n d d o ro t h y f r a g a s z y.1990. “Do monkeys ape?” in “Language” and intelligence inmonkeys and apes. Edited by Sue Taylor Parker and Kathleen

268 F current anthropology Volume 44, Number 2, April 2003
R. Gibson, pp. 247–73. Cambridge: Cambridge UniversityPress. [aw]
———. 2002. “Do monkeys ape? Ten years after,” in Imitationin animals and artifacts. Edited by Kerstin Dautenhahn andChrystopher L. Nehaniv, pp. 471–99. Cambridge: MIT Press.[aw]
v i s a l b e r g h i , e l i s a b e t t a , a n d w i l l i a m c . m cg r e w. Editors. 1997. Cebus meets Pan. International Journalof Primatology 18.
v i s a l b e r g h i , e l i s a b e t t a , a n d l o r e d a n a t r i n c a .1989. Tool use in capuchin monkeys: Distinguishing betweenperforming and understanding. Primates 30:511–21. [ev, ea]
v o e l k l , b e r n h a r d , a n d l u d w i g h u b e r . 2000. Trueimitation in marmosets. Animal Behaviour 60:195–202. [aw]
w a c h t m e i s t e r , c . - a . 2001. Display in monogamous pairs:A review of empirical data and evolutionary explanations. Ani-mal Behaviour 61:861–68.
w a t a n a b e , k . 1989. Fish: A new addition to the diet of Japa-nese macaques on Koshima Island. Folia Primatologica 52:124–31.
———. 1994. “Precultural behavior of Japanese macaques: Longi-tudinal studies of the Koshima troops,” in The ethologicalroots of culture. Edited by R. A. Gardner, A. B. Chiarelli, B. T.Gardner, and F. X. Plooij, pp. 182–92. Dordrecht: Kluwer Aca-demic Publishers.
w e s t , m . , a n d a . k i n g . 1996. “Social learning: Synergyand songbirds,” in Social learning in animals: The roots of cul-ture. Edited by Cecilia M. Heyes and Bennett G. Galef Jr., pp.155–78. San Diego: Academic Press.
w e s t e r g a a r d , g r e g o ry c . 1994. The subsistence technol-ogy of capuchins. International Journal of Primatology 15:899–906.
w e s t e r g a a r d , g r e g o ry c . , a n d s t e p h e n j . s u o m i .
1995. The stone tools of capuchins (Cebus apella). Interna-tional Journal of Primatology 16:1017–24.
w h i t e , d . j . , a n d b . g . g a l e f . 1999. Mate choice copy-ing and conspecific cueing in Japanese quail, Coturnix coturnixjaponica. Animal Behaviour 57:465–73.
w h i t e n , a n d r e w. 2000. Primate culture and social learning.Cognitive Science 24:477–508.
———. 2003. Cultural panthropology. Evolutionary Anthropol-ogy. In press. [aw]
w h i t e n , a . , j . g o o d a l l , w. c . m c g r e w, t . n i s h -i d a , v. r e y n o l d s , t . s u g i y a m a , c . e . g . t u t i n ,r . w. w. w r a n g h a m , a n d c . b o e s c h . 1999. Culturesin chimpanzees. Nature 399:682–85.
w h i t e n , a . , a n d r . h a m . 1992. “On the nature and evolu-tion of imitation in the animal kingdom: Reappraisal of a cen-tury of research,” in Advances in the study of behaviour, vol.21. Edited by P. J. B. Slater, J. S. Rosenblatt, C. Beer, and M.Milinski, pp. 239–83. San Diego: Academic Press.
w i c k l e r , w o l f g a n g . 1980. Vocal dueting and the pairbond. Zeitschrift fur Tierpsychologie 52:201–9.
w i l k i n s o n , g . , a n d j . b o u g h m a n . 1999. “Social influ-ences on foraging in bats,” in Mammalian social learning:Comparative and ecological perspectives. Edited by H. O. Boxand K. R. Gibson, pp. 188–204. Cambridge: Cambridge Univer-sity Press.
z a h a v i , a . 1977. The testing of a bond. Animal Behaviour 25:246–47.
z a h a v i , a . , a n d a . z a h a v i . 1997. The handicap principle.New York: Oxford University Press. [az]
z e l t n e r , e . , t . k l e i n , a n d b . h u b e r - e i c h e r . 2000. Isthere social transmission of feather pecking in groups of layinghen chicks? Animal Behaviour 60:211–16. [rld, ic, yv, knl]





















