
Investigation of UV optical fibers under synchrotron irradiation

ARTICLE IN PRESS
www.elsevier.com/locate/ymvre
YMVRE-02578; No. of pages: 12; 4C:
DTD 5
Microvascular Research
Pericyte adhesion and growth onto polyhydroxymethylsiloxane surfaces
nanostructured by plasma treatment and ion irradiation
Giovanna Asseroa, Cristina Satrianob, Gabriella Lupoa, Carmelina Daniela Anfusoa,
Giovanni Marlettab, Mario Alberghinaa,*
aDepartment of Biochemistry, University of Catania, 95125 Catania, ItalybDepartment of Chemical Sciences, University of Catania, 95125 Catania, Italy
Received 25 May 2004
Abstract
The study deals with the adhesion and proliferation of bovine retina pericytes onto surfaces of poly(hydroxymethylsiloxane) (PHMS)
modified either by cold plasma or by low-energy ion beams. The surface treatment was able to convert the original polymer matrix into SiO2-
like phases for O2-plasma or ion-mixed SiCxOy(Hz) phases for ion irradiation, respectively, with different modification levels of the surface
free energy (SFE) and related surface wettability. Pericytes exhibited a negligible adhesion and proliferation onto untreated PHMS, an
enhanced adhesion but not proliferation on plasma-treated PHMS, and great adhesion and proliferation to full confluence on ion-irradiated
PHMS, as measured by X-ray photoelectron spectroscopy (XPS), atomic force microscopy (AFM), quartz crystal microbalance, and optical
microscopy. On the other hand, the adhesion and proliferation of GP8.39 endothelial cells (EC), which are strongly associated with pericytes
in microvasculature, were very scarce onto both untreated and surface-modified PHMS. The surface-selective pericytal response was related
to changes of physicochemical properties of PHMS film, from hydrophobic/neutral towards hydrophilic/negatively charged polymer layers,
as well as to short- and long-time events of cell–surface interaction. We propose that surface properties can mediate and modulate cell–
polymer matrix adhesion through the establishment of stereospecific chemical interactions and/or electrostatic repulsion, which can also
explain the different behavior of pericytes compared to EC.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Pericytes; Endothelial cells; Poly(hydroxymethylsiloxane); Ion irradiation; Plasma treatment; Surface free energy; X-ray photoelectron
spectroscopy; Atomic force microscopy
Introduction
The cell adhesion and proliferation on synthetic surfaces
are fundamental processes in view of emerging medical
applications of biomaterials, including scaffolds for tissue
engineering, biosensors, medical devices, and bioelectronics
(Dee and Bizios, 1996; Grayson et al., 2004; Willner and
Willner, 2001). Among the various employed materials, an
increasing interest is addressed to the use of engineered
0026-2862/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.mvr.2004.08.003
* Corresponding author. Department of Biochemistry, Faculty of
Medicine, University of Catania, Viale Andrea Doria 6, 95125 Catania,
Italy. Fax: +39 95 7384220.
E-mail address: [email protected] (M. Alberghina).
polymers for controlled cell adhesion due to their inherent
advantages, as for instance the biodegradability, the process-
ability, the low cost, and the versatility in shaping (De Santis
et al., 2003; Hunkeler, 1997). Thus, a very large number of
reports deal with adhesion and proliferation of different cells
such as endothelial cells (EC), aortic smooth muscle cells,
chondrocytes, fibroblasts, astrocytes, glial cells, staminal
cells, osteoblasts, etc., on as different polymers as polylactic
acid (Watanabe et al., 2002), lactide-based polymers (Naj-
man et al., 2004), polyurethanes (Wang et al., 2002),
polyesters (Cenni et al., 1993; McFarland et al., 1999;
Ohsawa et al., 2001; Rizzi et al., 2001; Zreiqat et al., 1999),
polysiloxanes (Satriano et al., 1999, 2002a,b, 2001), and
polystyrene (Teare et al., 2000).
xx (2004) xxx–xxx

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx2
Among the various cell types, pericytes are very interest-
ing in view of their capability to act as progenitor cells
differentiating into a variety of different types including
osteoblasts (Couch, 1990; Diaz-Flores et al., 1992), macro-
phages and fibroblasts (Balabanov and Dore-Duffy, 1998;
Thomas, 1999), and adipocytes (Richardson et al., 1982). In
particular, it has been demonstrated that the process of bone
formation from osteoblasts cells already present in the
periosteum is enhanced by proliferation and differentiation
of pericytes, which contribute a supplementary population of
osteoprogenitor cells (Brighton et al., 1992). Furthermore,
microvascular pericytes exhibit in vitro phenotypic expres-
sions that are similar to that of in vitro bone cells (Reilly et al.,
1998). In this context, numerous studies have been carried out
to optimize culturing of osteoblast-like cells on various
biomaterials, for the development of the tissue engineering
techniques, and (in a larger perspective) also for reconstruc-
tive surgery. The growth of osteoblast-like cells on surface-
modified titanium (Yang et al., 2002), on titanium and titania
or hydroxyapatite surfaces (Ramires et al., 2002), on a
bioactive glass and a glass ceramic (Ohgushi et al., 1996), and
on hydroxyapatite (Cerroni et al., 2002; Dong et al., 2001;
Kilpadi et al., 2001) has been widely investigated, but to the
best of our knowledge, no studies have been reported on the
growth of pericytes on polymeric surfaces.
To study the cell adhesion on polymeric surfaces two
main strategies are currently pursued: the first one involves
the chemical modification of the surfaces (Craighead et al.,
2001; Zhang et al., 1999), while the second one points to
topographical structuring of the surfaces (Curtis and
Wilkinson, 1997). A relatively unconventional way to
induce controlled chemical modifications on the polymer
surfaces involves the use of high-energy density deposition
methods, like keV-MeV ion beams (Curtis and Wilkinson,
1997; Kusakabe et al., 1995; Marletta and Satriano, 2004;
Satriano et al., 2003a,b,c) and RF or microwave cold
plasmas (Dewez et al., 1999; Pu et al., 2002).
Previous studies have shown that cell adhesion enhance-
ment may be related to as different surface properties as the
increase of polar basic character of the irradiated surfaces
(Satriano et al., 2003b), the formation of very complex
conductive hydrogenated amorphous carbon layers for
carbon-based polymers (Marletta and Satriano, 2004), or
the formation of an insulating silica-like phase for silicon-
based polymers (Satriano et al., 2002a,b). This large spread
of physical properties and chemical surface structures has
not yet been unified in a single exhaustive model, but it
appears that the response of the various cell lines is
substantially affected by the nature and structure of the
adsorbed protein layer, acting as the mediating agent
between surface and cell membrane (Satriano et al.,
2003a). Thus, different cell types could exhibit different
adhesion and spreading behavior on a surface of given
composition and properties.
Accordingly, the present study reports the comparison
between the cell response of pericytes from bovine retina
microcapillaries and immortalized endothelial cells
(GP8.39) from rat brain microcapillaries onto surfaces of a
poly(hydroxymethylsiloxane) (PHMS) modified by O2-
plasma treatments and 50 keV Ar+ ion beams. It appeared
very promising to study the different sensitivity of these two
types of cells with respect to a model activated polymeric
surface due to the fact that microcapillary pericytes are
strongly associated with EC and share a common basement
membrane with them in microvasculature where muscular
cells are absent, playing the key role of maintaining the
vasal tone.
The polysiloxane has been chosen as model polymer to
test the basic process of cell–surface interaction for these
specific cells because previous works in our laboratory
demonstrated that irradiated PHMS exhibit a dramatic
enhancement of adhesion and proliferation of fibroblasts.
Furthermore, the properties of surface free energy and the
related wettability of PHMS may be easily modified in a
controlled and graded way by means of surface irradiation
techniques (Satriano et al., 1999, 2002a,b, 2003a, 2001), at
variance of what it has been found for other polymer surfaces
such as poly(ethyleneterephtalate) and poly(caprolactone),
which do not exhibit such a behavior (Satriano et al., 2003d).
Materials and methods
Polymer film preparation and modification
Poly(hydroxymethylsiloxane) (PHMS, HoneyWell) thin
films were deposited by spin coating (3000 rpm, 60 s, room
temperature) from solutions on either p-doped silicon (100)
wafers, glass or gold-covered quartz crystals. The structure
formula of the polymer is reported below.
The thickness of the deposited films was 500 F 50 nm,
as measured from an alpha-step profilometer. The surfaces
modification was performed with low-energy ion irradiation
or cold plasma treatment. The ion irradiation was done with
a Danfysik ion implanter, with Ar+ ions at an energy of 50
keV. The beam was rastered over the samples to keep the
thermal load as low as possible and the ion dose was
controlled at 1015 ions/cm2. Plasma treatments were carried
out in a March Instrument solid-state PlasmodR unit
(Concord, CA, USA) supplied with an RF generator with
an excitation frequency of 13.56 MHz. The treatment
conditions were as follows: 99.95% minimum purity oxy-
gen; power, 100 W; pressure, 66.6 Pa; treatment time, 1
min. After the plasma exposure the samples were aged in

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx 3
laboratory atmosphere for periods ranging from 24 h up to 1
week, that is, the periods observed for the surfaces to exhibit
a steady behavior. The effective thickness of the modified
layer can be estimated to be higher than roughly 9 nm
because the XPS analysis of the fresh plasma-modified
samples does not show any significant traces of the
characteristic carbon signal coming from the original methyl
groups of the polymer.
Surface characterization
X-ray photoelectron spectroscopy (XPS)
XPS analysis was carried out with a Kratos HX AXIS
spectrometer equipped with a dual Al/Mg anode and a
hemispherical analyzer. The spectra were obtained in fixed
analyzer transmission mode (pass energy 40 eV) by using
the Mg Ka1,2 radiation. The estimated sampling depth is
about 9 nm, according to an attenuation length of 3.0 nm for
Si 2p peak in organic materials (Suzuki et al., 1997). Such
value is actually comparable to the estimated thickness of
the ion- and plasma-modified layers (see above). XPS
spectra were analyzed by using an iterative least squares
fitting routine based on Gaussian peaks and the Shirley
background subtraction (Seah and Brown, 1999). Binding
energies (BEs) of all the spectra were referenced to the
intrinsic (before irradiation treatment) hydrocarbon-like C
1s peak set at 284.6 eV or to the adventitious one set at
285.0 eV (after the plasma treatments) (Hongbing and
Hamers, 1998).
Atomic Force Microscopy (AFM)
The surface microtopography and the morphology of the
surfaces were measured with a Multimode/Nanoscope IIIA
Atomic Force Microscope (Veeco) in tapping mode in air
with a standard silicon tip. The relative room humidity was
30% and the room temperature was 238C. Data were
acquired on square frames having edges of 10 Am, 1 Am,
and 350 nm. Images were recorded using height, phase-
shift, and amplitude channels with 512 � 512 measurement
points (pixels). Measurements were made twice or three
times on different zones of each sample.
Surface free energy (SFE) measurements
Measurements of surface free energy were performed by
evaluating both static and dynamic contact angles of three
different liquids onto the untreated and treated surfaces. Half
automatic video-based measurements of contact angle were
performed at 258C and 65% relative humidity by using an
OCA30 instrument (Dataphysics). By using the sessile drop
method, liquid drops of 2 Al of volume were applied on
different zones of each sample surface; and by digital image
analysis, the static contact angles (hs) were measured on
both sides of the two-dimensional projection of the droplet.
The advancing (hadv) and the receding (hrec) contact angleswere measured by the needle-syringe method (Erbil et al.,
1999). At least five measurements were made for each
sample and then averaged. The surface free energies, in
terms of apolar Lifshitz–van der Waals (cLW) and polar
Lewis acid (c+) and basic (c�) components, were evaluated
by using the Good-van Oss model (van Oss, 2002), with the
three following liquids: ultrapure Millipore water, glycerol,
and tricresyl phosphate (Aldrich).
Pericytes isolation and in vitro culture
Microvessel pericytes were extracted from bovine retinas
as previously described (Lupo et al., 2001). Cells were
characterized by negative staining for factor VIII-related
antigen, positive staining for smooth muscle a-actin
monoclonal antigen, morphological features including
absence of contact inhibition and g-glutamyltranspeptidase
activity (Lupo et al., 2001). The isolated cells were then
cultured in DMEM supplemented with 10% Fetal Calf
Serum (FCS), 100 U/ml penicillin, and 100 Ag/ml strepto-
mycin. Cells were incubated at 378C in a 5% CO2 incubator
and the medium was changed every 2 days. When the cells
reached the plateau phase of growth, they were harvested by
trypsinization, followed by addition of fresh culture medium
to create a new single cell suspension with desired seeding
cell number per cm2. Pericytes were identified by their
characteristic polygonal shape and through smooth muscle
a-actin staining.
QCM-D experiments and early events of cell–surface
interaction studies
In situ and real time studies of the early events, that is, till
2 h of incubation, of cell–surface interaction in PBS solution
were undertaken by using a Quartz Crystal Microbalance
with Dissipation Monitoring (QCM-D) instrument (Q-Sense
AB, Gothenburg, Sweden), which allowed the simultaneous
measurements of both frequency ( f) and energy dissipation
(D) of the sensor consisting of 5 MHz-crystals (Q-Sense),
spin-coated with PHMS thin films. Baseline curves were
measured with sensors oscillating in phosphate buffer saline
(PBS) solution; then the changes in D and f due to the
addition of a PBS solution containing 104 cells/ml were
monitored for both the fundamental frequency (n = 1, i.e., f
approximately 5 MHz), and the first three overtones (n = 3,
5, and 7, corresponding to f approximately 15, 25, and 35
MHz, respectively). The experiment was performed at 378C.
Pericytes growth studies on various PHMS substrates
Third passage pericytes were used to seed various PHMS
substrates at an initially cell density of 2 � 104 cells/cm2.
Incubation was performed in polystyrene culture dishes. The
PHMS samples were placed in the center of the dishes and
added with 1 ml of cell suspension to allow full attachment
of cells to polymers. Cultivation was conducted for 2, 24, 48
h up to 6 days. Culture media were changed every 2 days.
The cultures were carried out up to 5 days. All experiments
ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx4
were performed in triplicate. After each incubation, cells
were washed with Dulbecco Phosphate Buffer Saline
(DPBS) to remove nonadherent cells, and samples were
observed using an inverse phase light microscope (Zeiss
Axiovert 100) for analysis of cell morphology, spreading
and distribution on the investigated substrates, and collec-
tion of photomicrographs. In order to determine the number
of cells grown on various substrates, at least five random
visual fields per sample were photographed, and cells were
counted on five areas of each sample at 100� magnification.
Quantification of cells was obtained using an image analysis
software (Scion Image) after taking pictures of spreading
cells, evaluating the cell coverage in terms of integrated
density (ID). Cell viability was assessed using a trypan blue
assay. In another set of experiments, various PHMS
substrates were seeded with pericytes and cells were at the
end of the same incubation period washed three times with
DPBS and then treated with trypan blue for 15 min at 378C.Trypan blue assay was employed in this study to quantita-
tively assess the number of viable pericytes attached and
grown on tested polymers.
Endothelial cell culture
The immortalized rat brain endothelial cells (GP8.39)
used were generously provided by Dr. J. Greenwood
(Department of Clinical Ophthalmology, University Col-
lege, London). The cell line was already characterized and
our cell cultures were prepared following the procedures
previously described (Anfuso et al., 1999; Greenwood et al.,
1996). EC were grown at confluency in F-10 Ham medium
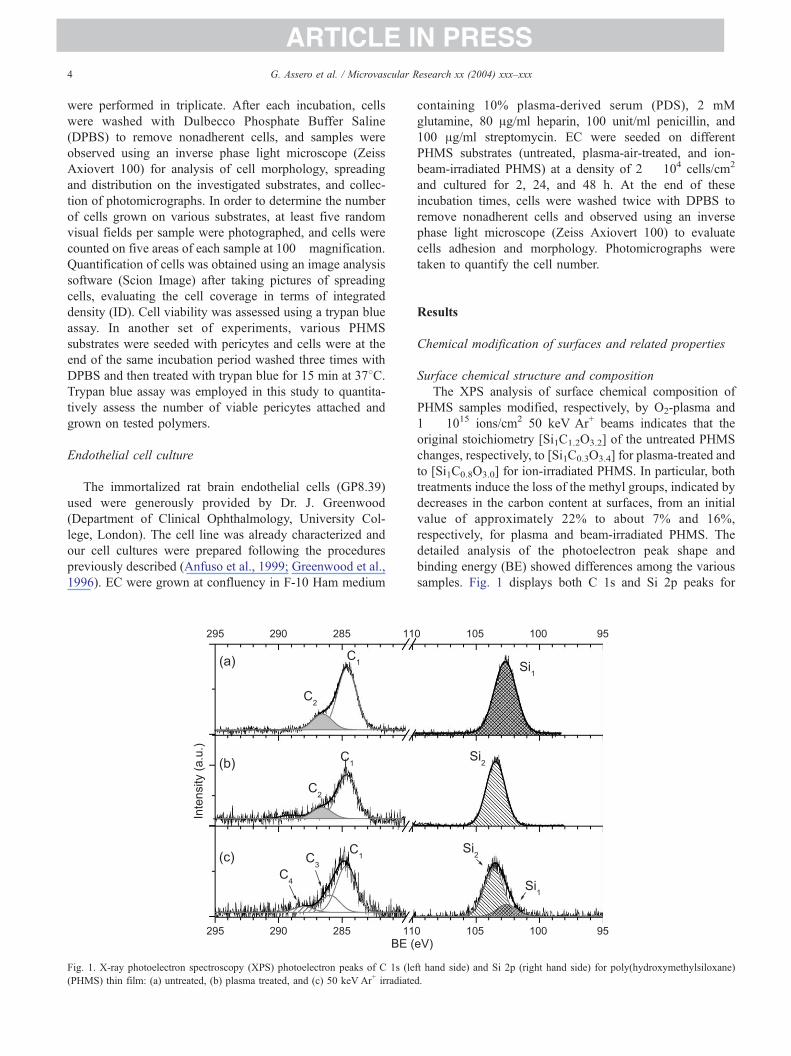
Fig. 1. X-ray photoelectron spectroscopy (XPS) photoelectron peaks of C 1s (le
(PHMS) thin film: (a) untreated, (b) plasma treated, and (c) 50 keV Ar+ irradiate
containing 10% plasma-derived serum (PDS), 2 mM
glutamine, 80 Ag/ml heparin, 100 unit/ml penicillin, and
100 Ag/ml streptomycin. EC were seeded on different
PHMS substrates (untreated, plasma-air-treated, and ion-
beam-irradiated PHMS) at a density of 2 � 104 cells/cm2
and cultured for 2, 24, and 48 h. At the end of these
incubation times, cells were washed twice with DPBS to
remove nonadherent cells and observed using an inverse
phase light microscope (Zeiss Axiovert 100) to evaluate
cells adhesion and morphology. Photomicrographs were
taken to quantify the cell number.
Results
Chemical modification of surfaces and related properties
Surface chemical structure and composition
The XPS analysis of surface chemical composition of
PHMS samples modified, respectively, by O2-plasma and
1 � 1015 ions/cm2 50 keV Ar+ beams indicates that the
original stoichiometry [Si1C1.2O3.2] of the untreated PHMS
changes, respectively, to [Si1C0.3O3.4] for plasma-treated and
to [Si1C0.8O3.0] for ion-irradiated PHMS. In particular, both
treatments induce the loss of the methyl groups, indicated by
decreases in the carbon content at surfaces, from an initial
value of approximately 22% to about 7% and 16%,
respectively, for plasma and beam-irradiated PHMS. The
detailed analysis of the photoelectron peak shape and
binding energy (BE) showed differences among the various
samples. Fig. 1 displays both C 1s and Si 2p peaks for
ft hand side) and Si 2p (right hand side) for poly(hydroxymethylsiloxane)
d.

ARTICLE IN PRESS
Table 1
Root mean square (RMS) and average roughness (Ra) for the various
PHMS surfaces measured in air by AFM in tapping mode on (1 � 1) Am2
scan regions
PHMS RMS (nm) Ra (nm)
Untreated 0.546 F 0.007 0.440 F 0.009
Plasma treated 0.520 F 0.06 0.428 F 0.04
Ion irradiated 0.544 F 0.06 0.434 F 0.04
G. Assero et al. / Microvascular Research xx (2004) xxx–xxx 5
untreated, plasma-treated, and ion-irradiated PHMS. For the
untreated PHMS surfaces (Fig. 1a), the C 1s peak can be
fitted by using only two basic components, both having full
width at half maximum (FWHM) of approximately 1.5 eV.
The main one (C1) is centered at 284.6 eV of BE and
assigned to NC–Si bonds, in agreement with literature data
(Satriano et al., 2001); the second peak component (C2) is
found at approximately 286.6 eV of BE, being assigned to
NC–OH and NC–O–C groups belonging either to the
terminal polymer groups or to the solvent residues (Satriano
et al., 2001). The Si 2p peak analysis evidences a symmetric
band, which is well fitted by using a single gaussian
component (Si1) of FWHM approximately 1.7 eV, centered
at about 102.2 eV of BE, assigned to SiO3C clusters, in
agreement with literature (Satriano et al., 2001).
For the plasma-treated and aged PHMS surfaces (Fig.
1b), the C1 component is dramatically reduced with respect
the C2 one; these components basically remain at the same
BE as in unirradiated PHMS, whereas in the freshly treated
samples (not shown) the carbon peak is completely
eliminated. This fact suggests that a partial recovery process
occurs with exposure to atmosphere, basically involving a
small but significant surface segregation of bulk chains
(Satriano et al., 2002a,b). As to Si 2p peak, a symmetric and
narrow band (FWHM approximately 1.5 eV) has been
found at approximately 103.6 eV (Si2 component), charac-
teristic of the formation of an amorphous SiO2-like phase,
predominantly formed by randomly interlinked [SiO4]
clusters.
At variance of the plasma case, for the 50-keV Ar+-
irradiated PHMS (Fig. 1c), two new components, centered at
286.1 eV (C3) and 288.0 eV (C4), were found for the C 1s
peak and respectively assigned to the formation of C–O–Si
moieties (C4 component) and NC = O groups (C5 compo-
nent) (Satriano et al., 2001). Furthermore, traces of newly
formed NCOO� groups could be identified in the high BE
tail of the peak, confirmed by TOF-SIMS measurements
(data not shown). These new components replaced the
pristine C2 component. It is to stress that in this case, the
freshly irradiated samples have similar composition of the
aged ones, showing no evidence of any carbon recovery at
surfaces. Furthermore, Si 2p peak fitting evidences the
coexistence of two components, Si1 and Si2, with a relative
ratio of 4:1 between [SiO3C] and [SiO4] clusters.
Surface free energy
After irradiation, the water contact angle changes from
the original value of 93.08 F 1.78 of the hydrophobic
untreated PHMS to 12.88 F 0.58 (i.e., very hydrophilic) for
plasma-treated and 51.78 F 0.98 (i.e., mildly hydrophilic)
ion-irradiated PHMS surfaces, respectively. It is worthy to
stress that the contact angle modification for the various
irradiated surfaces could in principle be due to radiation-
induced morphology changes. Accordingly, the average
roughness of all the investigated surfaces has been measured
by AFM. Table 1 shows the results in terms of root mean
square (Rrms) and mean (Ra) roughness. It appears that the
various treatments do not induce any significant change in
roughness for both O2-plasma-treated and 50 keV Ar+-
irradiated samples, respectively.
The surface free energy of each sample was evaluated in
terms of the apolar or Lifshitz–van der Waals component,
including the dispersive, inductive, and orientational con-
tributions to the van der Waals interactions and acid and
basic Lewis polar components. Fig. 2 shows the total SFE,
as well as the abovementioned dispersive and polar
components for untreated, plasma-, and ion-irradiated
PHMS surfaces. Both irradiation treatments generally
increase the polar components much more than the
corresponding apolar cLW term.
Short-term events in cell–surface interaction
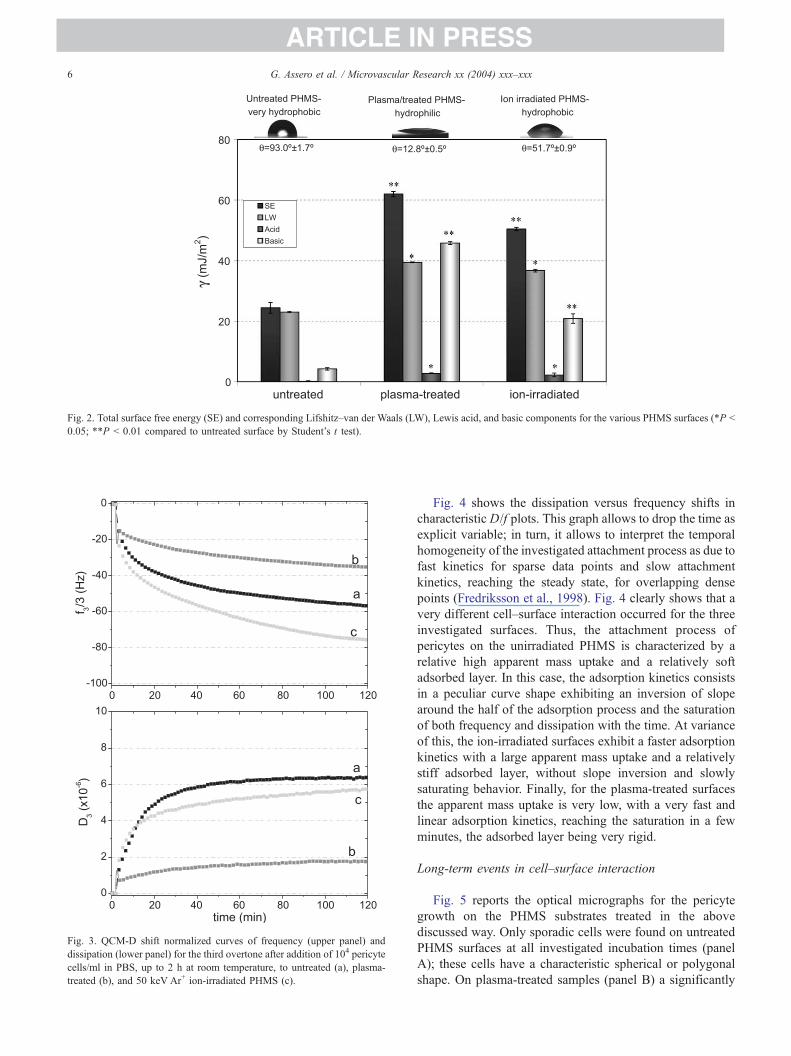
Fig. 3 shows the frequency and dissipation plots for cell
attachment events onto untreated (a), O2-plasma-treated (b),
and 50 keV Ar+ ion-irradiated PHMS (c), respectively. The
experiments have been ran for 2 h in PBS solution, allowing
just an early interaction of the cells on the various polymer
surfaces. The frequency ( f) and dissipation (D) plots show
that PHMS surfaces trigger different pericyte responses
according to various treatments. In fact, both untreated and
ion-irradiated PHMS surfaces exhibited a huge frequency
shift of approximately �70 H �80 Hz due to a quite high
beffectiveQ cellular mass attachment on the electrode surface
and a corresponding dissipation shift of approximately 6 �10�6, that is, a large dissipative response of the attached
cellular matter, corresponding to a very pronounced cell
viscoelastic character. A different cell behavior was found in
the plasma-treated PHMS surfaces where the observed shift
in frequency was about�25 Hz, and the dissipation of about
approximately 1 � 10�6, that is, a lower effective cell mass
on the surface and a lower viscoelasticity of the adhered
matter. It is to point out that the frequency shift observed in
the case of cell–substrate interaction in the QCM-D experi-
ments is not directly proportional to real mass uptake
because the shear stress wave measuring the adsorbed mass
penetrates no more than 0.25 Am in the overlaying matter.
Therefore, the technique does not bsenseQ the whole cellularbody, but only the interaction of cellular membrane region
with the substrate (Fredriksson et al., 1998), and provides
just an estimation of beffectiveQ mass, which is not directly
proportional to the number of adhered cells, but to the
relative substrate coverage.
ARTICLE IN PRESS
Fig. 3. QCM-D shift normalized curves of frequency (upper panel) and
dissipation (lower panel) for the third overtone after addition of 104 pericyte
cells/ml in PBS, up to 2 h at room temperature, to untreated (a), plasma-
treated (b), and 50 keV Ar+ ion-irradiated PHMS (c).
Fig. 2. Total surface free energy (SE) and corresponding Lifshitz–van der Waals (LW), Lewis acid, and basic components for the various PHMS surfaces (*P b
0.05; **P b 0.01 compared to untreated surface by Student’s t test).
G. Assero et al. / Microvascular Research xx (2004) xxx–xxx6
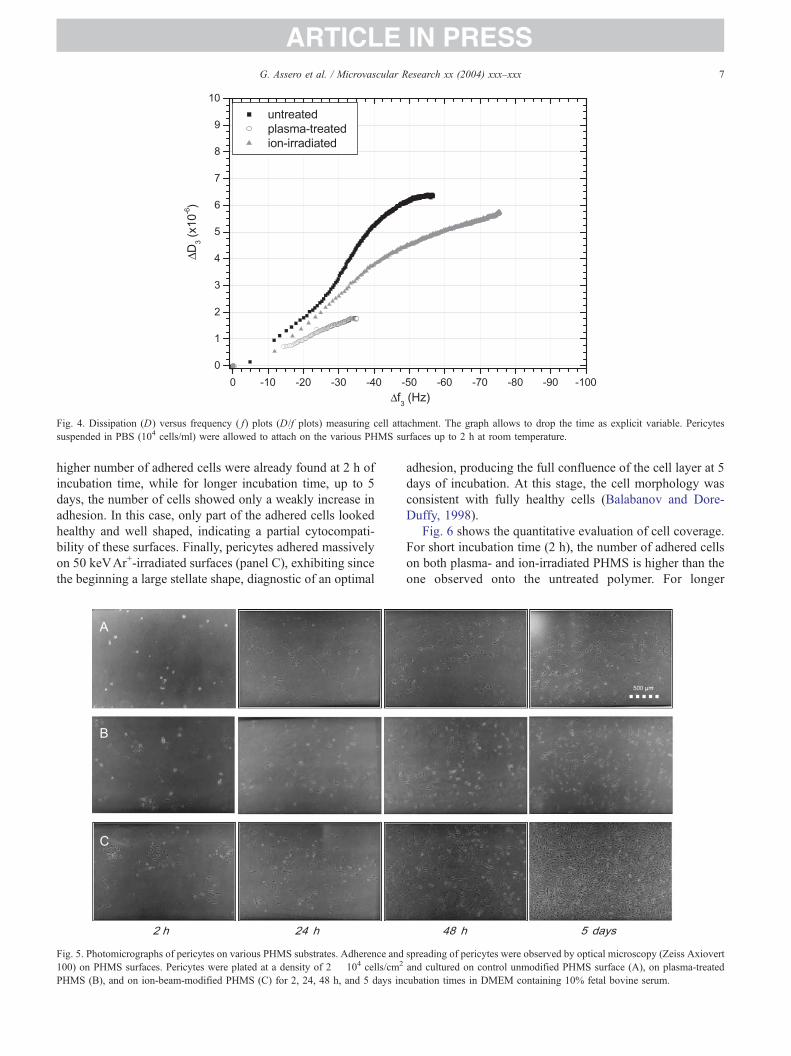
Fig. 4 shows the dissipation versus frequency shifts in
characteristic D/f plots. This graph allows to drop the time as
explicit variable; in turn, it allows to interpret the temporal
homogeneity of the investigated attachment process as due to
fast kinetics for sparse data points and slow attachment
kinetics, reaching the steady state, for overlapping dense
points (Fredriksson et al., 1998). Fig. 4 clearly shows that a
very different cell–surface interaction occurred for the three
investigated surfaces. Thus, the attachment process of
pericytes on the unirradiated PHMS is characterized by a
relative high apparent mass uptake and a relatively soft
adsorbed layer. In this case, the adsorption kinetics consists
in a peculiar curve shape exhibiting an inversion of slope
around the half of the adsorption process and the saturation
of both frequency and dissipation with the time. At variance
of this, the ion-irradiated surfaces exhibit a faster adsorption
kinetics with a large apparent mass uptake and a relatively
stiff adsorbed layer, without slope inversion and slowly
saturating behavior. Finally, for the plasma-treated surfaces
the apparent mass uptake is very low, with a very fast and
linear adsorption kinetics, reaching the saturation in a few
minutes, the adsorbed layer being very rigid.
Long-term events in cell–surface interaction
Fig. 5 reports the optical micrographs for the pericyte
growth on the PHMS substrates treated in the above
discussed way. Only sporadic cells were found on untreated
PHMS surfaces at all investigated incubation times (panel
A); these cells have a characteristic spherical or polygonal
shape. On plasma-treated samples (panel B) a significantly
ARTICLE IN PRESS
Fig. 4. Dissipation (D) versus frequency ( f) plots (D/f plots) measuring cell attachment. The graph allows to drop the time as explicit variable. Pericytes
suspended in PBS (104 cells/ml) were allowed to attach on the various PHMS surfaces up to 2 h at room temperature.
G. Assero et al. / Microvascular Research xx (2004) xxx–xxx 7
higher number of adhered cells were already found at 2 h of
incubation time, while for longer incubation time, up to 5
days, the number of cells showed only a weakly increase in
adhesion. In this case, only part of the adhered cells looked
healthy and well shaped, indicating a partial cytocompati-
bility of these surfaces. Finally, pericytes adhered massively
on 50 keVAr+-irradiated surfaces (panel C), exhibiting since
the beginning a large stellate shape, diagnostic of an optimal
Fig. 5. Photomicrographs of pericytes on various PHMS substrates. Adherence and
100) on PHMS surfaces. Pericytes were plated at a density of 2 � 104 cells/cm2
PHMS (B), and on ion-beam-modified PHMS (C) for 2, 24, 48 h, and 5 days in
adhesion, producing the full confluence of the cell layer at 5
days of incubation. At this stage, the cell morphology was
consistent with fully healthy cells (Balabanov and Dore-
Duffy, 1998).
Fig. 6 shows the quantitative evaluation of cell coverage.
For short incubation time (2 h), the number of adhered cells
on both plasma- and ion-irradiated PHMS is higher than the
one observed onto the untreated polymer. For longer
spreading of pericytes were observed by optical microscopy (Zeiss Axiovert
and cultured on control unmodified PHMS surface (A), on plasma-treated
cubation times in DMEM containing 10% fetal bovine serum.

ARTICLE IN PRESS
Fig. 6. Number of adhered pericytes on various PHMS substrates. Cells viability was determined by trypan blue vital staining. Pericytes were plated on various
PHMS-modified surfaces at a density of 2 � 104 cells/cm2. After 2, 24, and 48 h, the number of cells on control untreated PHMS surface, on plasma-treated
PHMS and on ion-beam-modified PHMS was determined. Data shown are representative of three separate experiments (*P b 0.05; **P b 0.01 compared to
untreated control by Student’s t test).
G. Assero et al. / Microvascular Research xx (2004) xxx–xxx8
incubation times (24 and 48 h), a significant increase in the
number of adhered cells occurred on the Ar+-irradiated, but
not on plasma-treated, samples, indicating that a real
proliferation process was arising only on the ion-modified
surfaces.
The results of adhesion of rat brain endothelial cells, used
for comparison sake, on the same PHMS surfaces are
reported in Fig. 7. No significant cell adhesion was found
for any of the PHMS substrates, and no differences were
observed neither in proliferation nor in typical morpholog-
ical differentiation of EC. In preliminary experiments, we
used endothelial cells isolated from bovine brain micro-
capillaries, but they did not attach as did immortalized cells
from rat.
Fig. 7. Rat brain GP8.39 endothelial cells (EC) growth and adhesion after 2, 24, a
PHMS surfaces (B). No differences were detected over controls as function of time
substrates.
Discussion
The results basically indicate that pericytes do not
significantly adhere onto untreated PHMS surfaces, while
they adhere, but do not proliferate, on plasma-treated
surfaces and adhere and proliferate massively onto ion-
irradiated surfaces. Since all the incubation experiments in
the present work were performed on the bare substrate
surfaces, that is, without the routinely used precoatings of
gelatin, the observed different cell behavior on the various
surfaces must be related to the specific surface properties on
one side, and the response of the complex environment
formed by culture medium and cell membranes on the other
side.
nd 48 h incubation times onto control untreated (A) and ion-beam-modified
. No cell proliferation and phenotypical differentiation were observed in both

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx 9
In particular, the results of the physicochemical charac-
terization indicate that the untreated PHMS surfaces consist
of a hydrophobic layer, formed by methyl groups oriented
towards the solid-water interface, not charged in solution at
pH 7.4. At variance of this, both the plasma treatment and
the Ar+ irradiation induced the formation of an altered
surface layer basically formed of Si-enriched phases. The
XPS results show clearly that two different nanostructures
of the altered layers were produced depending on the type of
treatment. Indeed, the Ar+-irradiation produced a specific
amorphous SiCxOy(Hz) phase, with a high intrinsic content
of carbon-containing domain and Si–C linkages, while the
plasma treatment essentially produced an SiO2-like phase,
that is, without Si–C linkages (Fig. 1). Both surfaces show a
negative charge in solution at pH 7.4 (Satriano et al.,
2003d).
The related surface free energy measurements are in
agreement with the XPS data, as irradiated surfaces
exhibited a strong modification of water contact angles h,from the hydrophobic character of the unirradiated, methyl-
terminated PHMS surface (h approximately 938) to a mildly
hydrophilic character (h approximately 528) for Ar+-
irradiated surfaces and a strongly hydrophilic one (happroximately 138) for the plasma-treated surfaces. Related
components were also dramatically changed, as the dis-
persive Lifshitz–van der Waals parameter increased from
about 24 mJ/m2 for the hydrophobic untreated PHMS
surface to approximately 36 mJ/m2 for Ar+-irradiated
surfaces and to approximately 40 mJ/m2 for the plasma-
treated ones, respectively, the magnitude being roughly the
same for the two modified samples. In addition, the polar
Lewis base component underwent a dramatic increase due
to the irradiation treatments. In fact, the initial value of
about 5 mJ/m2 becomes about 21 mJ/m2 for Ar+-irradiated
surfaces and about 45 for the plasma-treated one, that is, the
difference in the nanostructure of irradiated surfaces marked
a factor two in the polar Lewis base terms.
In view of the synergic action of culture medium and cell
membranes related to the various surfaces, we discuss the
pericyte behavior discriminating between short-term events,
essentially involving the mere adhesion process (physical
contact) for incubation time up to 2 h, and long-term events
consisting in cell spreading and proliferation phenomena,
for incubation times from 24 h up to 6 days. In short-term
events, we have performed both serum-free and serum-
containing experiments to discriminate between cell mem-
brane–surface interactions and serum-mediated cell adhe-
sion. As shown in the previous section, the short-terms
adhesion events in serum-free experiments could be
analyzed in terms of the QCM-D and optical microscopy
results, whereas the cell response in the serum-containing
experiments has been basically investigated by optical
microscopy only.
In general, QCM-D measurements allow a simultaneous
estimate of the effective mass uptake, given by the measured
frequency shift, and of viscoelastic properties of the sensed
adsorbed layer, that is, the cell–substrate interface reflected
by the energy dissipation factor (Nimeri et al., 1998). In the
case of cell–polymer interactions, the frequency shift is no
longer related in a straightforward way to the mass uptake,
as far as the Sauerbrey equation, stating the proportionality
between quartz frequency shift and the adsorbed mass, is no
longer valid (Fredriksson et al., 1998). However, the
measured frequency shift still contains an indication of the
strength and dynamic character of the interaction between
adhered cells and polymer surfaces, discriminating succes-
sive steps of cell–surface interaction. In fact, the measured
shift is mostly depending on the cell–surface contact area,
that is, both on the rate of cell attachment on the surface and
the spreading process with respect to the initial physical cell
contact. To this purpose, it is to point out that the evanescent
wave produced by the oscillating sensor has a typical
penetration length of approximately 0.25 Am in the over-
laying matter, that is, a sampled thickness much lower than
the typical average vertical size of a single cell.
On the other hand, the energy dissipation measurement
provides valuable information about the viscoelastic proper-
ties, which are originated within the thin cellular region and
the related cell–substrate interface. In particular, the rigidity
of the sampled layer can relate the strength of adhesion to
factors like the number of binding sites, types of expressed
adhesion proteins, changes in the cytoskeleton in proximity
of the cellular wall, etc. (Marx et al., 2003). The data
reported in Fig. 3 have to be interpreted in terms of the
above explanation as due to the characteristics of the cell–
substrate interaction and not to the mere number of cells.
The analysis of D/f plots (see Fig. 4), together with the
optical microscopy (OM) pictures, allows to spot the strong
differences among the features of pericyte adhesion onto
untreated, plasma-treated, and ion-irradiated PHMS. Due to
the fact that OM pictures showed (data not reported) that in
serum-free conditions a comparable number of few cells
was attached on the different surfaces—even if most of them
still did not exhibit the typical pericyte phenotype—the
observed differences in D/f plot must be mostly related to
the differences in the cell–surface interaction features.
In turn, the D/f plot for the plasma-treated PHMS, an
indication of a linear slow kinetics of a small mass uptake
providing a rigid layer, can be interpreted as an evidence of
a very strong interaction between the cell membrane and the
very hydrophilic surfaces, blocking the cell and preventing
the subsequent spreading. On the contrary, the initial fast
adsorption processes followed by a second slower step,
involving a large apparent mass uptake and the formation of
viscoelastic layer for the very hydrophobic untreated PHMS
surfaces, can be interpreted as diagnostic of the sticking of
loosely bound cells, ready to detach from the surface and
therefore not suitable for the spreading and proliferation
steps. Finally, the intermediate behavior of the D/f plot for
the ion-irradiated surfaces, characterized also by an inter-
mediate degree of hydrophilicity, is in agreement with the
occurrence of efficient attachment process of cells relatively

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx10
well spread on the surfaces, as it is suggested by dissipative
behavior characteristic of a more rigid adsorbed layer with
respect to the unirradiated surfaces. In this case, the cells
still would have sufficient degrees of freedom to prompt the
subsequent spreading and proliferation processes.
Thus, the whole results for serum-free experiments could
be understood in terms of the different strength of cell
attachment as a function of the surface free energy, which is
in turn related to the strong increase of Lewis base groups.
Accordingly, the very polar nature of the plasma-treated
surfaces prompts pericyte attachment. However, the cell
viability was hindered by the strong sticking, whereas the
almost exclusive dispersive character of the unirradiated
PHMS surfaces prevents any effective cell binding. In
conclusion, only ion-irradiated PHMS surfaces have the
right density of polar groups prompting a suitable attach-
ment of the cells, without preventing the subsequent
proliferation process.
The results obtained in the serum-containing experiments
add more insight to the picture of pericyte behavior on the
various polymer surfaces. In fact, the OM pictures 2 h after
incubation onto plasma- and ion-irradiated surfaces showed
that pericytes were able to reconstruct their phenotype ex-
pression already at this short incubation time, whereas such
an effect was not seen for the unirradiated PHMS. The
average number of adhered cells for different surfaces in this
case was higher than in the corresponding serum-free
experiments, suggesting that the effect of proteins adsorp-
tion on the surfaces from the serum might play a role in
prompting the cell attachment. However, the number of
adhered cells was significantly different among surfaces, the
higher number of adhered cells being observed on the ion-
irradiated surfaces, followed by the plasma-treated and by
the unirradiated ones (see Fig. 6). It is noteworthy that this
quantitative trend corresponded to the above-discussed
findings from D/f plots in terms of relative interaction
strength.
Taking into account long-term events, which surely
involve cell spreading and proliferation processes in
serum-containing culture medium, our results deserve a
special comment. Cell proliferation essentially reflects the
trend already observed for cell attachment. In fact, for
untreated surfaces, there was no proliferation occurring; for
plasma-treated surfaces, the number of attached cells was
almost unchanged, while proliferation took place only for
ion-irradiated surfaces.
Several hypotheses on the role of peculiar cell processes
occurring on a long scale of time contact, that is, protein
adsorption from the serum and cell expression of adhesion
proteins, may be made. A first hypothesis concerns the
differential interaction of adhesion molecules with surfaces.
Integrins, cadherins, intercellular adhesion molecule-1
(ICAM), and vascular cell adhesion molecule-1 (VCAM-
1), some of which are present constitutively and others that
can be up-regulated in response to chemotactic stimuli, play
a key role in cell–cell and cell–extracellular matrix
interactions. Different expression of adhesion molecules
may reflect different culture and experimental conditions
(Wong and Dorovini-Zis, 1995), and it could be influenced
either by wettability or electric charge of solid substrate
available to cells in culture, such as the polymer we used.
However, adhesion molecules are expressed and modulated
in both cultured EC and pericytes (Balabanov et al., 1996;
Ivanov et al., 2001; Daxecker et al., 2002), so this
hypothesis cannot explain the differential behavior of these
two cell types against PHMS.
A second hypothesis concerns the expression of a variety
of extracellular matrix (ECM) components like fibronectin
and laminin (Mandarino et al., 1993; Tilling et al., 2002),
collagen (Cohen et al., 1980), and glycosaminoglycans
(Stramm et al., 1987) by pericytes and EC. These
components enhance, for instance, pericyte adhesion and
growth and play an important role in regulating cell growth,
as demonstrated by coculture (pericytes and EC) experi-
ments (Antonelli-Orlidge et al., 1989) and in the main-
tenance of vessel integrity (Hirschi and D’Amore, 1996).
Expression of ECM components, however, is not able to
explain temporally the early mechanism of cell–polymer
interaction because it occurs within 2 h, while ECM
molecules expression requires longer times.
A further and more consistent hypothesis is that the
different response of pericytes and EC to the interaction
with PHMS could be probably due to their cellular
properties of in vitro growth. EC need an extracellular
matrix to proliferate, usually collagen, and a medium with
high serum percentage to grow and to spread out. Collagen
is designed as a glycoprotein since it contains significant,
but highly variable, amounts of covalently linked carbohy-
drates. All carbohydrate units are linked O-glycosidically to
hydroxylysine residues in a unique way. The absence of this
right substrate in culture dish determines the loss of the
phenotype and makes EC very susceptible of detachment
from polymeric surfaces. Pericytes are instead able to grow
even in absence of matrix support, usually gelatin, which is
an heterogeneous mixture of water-soluble proteins present
in collagen, maintaining cellular characteristics and pheno-
type. The presence of carbohydrate groups in collagen could
play an essential role in EC adhesion to culture dishes in in
vitro conditions.
Based on this premises, we believe that adhesion
molecules or secretion of ECM components appear not to
contribute to the control of cell–substrate interaction on
PHMS slides. We thus propose that surface properties (for
instance, the presence of specific glycoconjugates) can
mediate and modulate cell–polymer matrix adhesion
through the establishment of stereospecific chemical inter-
actions and/or electrostatic repulsion.
In conclusion, our findings highlight the ability of
untreated and irradiated PHMS surface to act as a selective
modulator of cell adhesion, inducting or inhibiting
adhesive interactions depending on the specific properties
of microvascular cells and the matrix around them. In

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx 11
addition, the present study establishes an experimental
framework to analyze adhesive mechanism controlling
cell–surface interactions and provides a general strategy
of surface-directed control to manipulate cellular spreading
in biomaterial and biotechnological applications.
Acknowledgments
The authors wish to acknowledge the financial support of
COFIN 2002 (University of Catania) and CIB (University of
Catania). The authors are also grateful to Dr. F. Rossi (IHCP,
Joint Research Centre, European Commission, Ispra) for the
ion-beam irradiation facilities.
References
Anfuso, C.D., Lupo, G., Alberghina, M., 1999. Amyloid-beta but not
bradykinin induces phosphatidylcholine hydrolysis in immortalized rat
brain endothelial cells. Neurosci. Lett. 271, 151–154.
Antonelli-Orlidge, A., Saunders, K.B., Smith, S.R., D’Amore, P.A.,
1989. An activated form of transforming growth factor beta is
produced by cocultures of endothelial cells and pericytes. Proc. Natl.
Acad. Sci. U. S. A. 86, 4544–4548.
Balabanov, R., Dore-Duffy, P., 1998. Role of the CNS microvascular
pericyte in the blood–brain barrier. J. Neurosci. Res. 53, 637–644.
Balabanov, R., Washington, R., Wagnerova, J., Dore-Duffy, P., 1996. CNS
microvascular pericytes express macrophage-like function, cell surface
integrin alpha M, and macrophage marker ED-2. Microvasc. Res. 52,
127–142.
Brighton, C.T., Lorich, D.G., Kupcha, R., Reilly, T.M., Jones, A.R.,
Woodbury II, R.A., 1992. The pericytes as a possible osteoblast
progenitor cell. Clin. Orthop. 275, 287–299.
Cenni, E., Ciapetti, G., Cavedagna, D., Dileo, A., Pizzoferrato, A., 1993.
Production of prostacyclin and fibrinolysis modulators by endothelial
cells cultured in the presence of polyethylene terephthalate. J. Biomed.
Mater. Res. 27, 1161–1164.
Cerroni, L., Filocamo, R., Fabbri, M., Piconi, C., Caropreso, S., Condo,
S.G., 2002. Growth of osteoblast-like cells on porous hydroxyapatite
ceramics: an in vitro study. Biomol. Eng. 19, 119–124.
Cohen, M.P., Frank, R.N., Khalifa, A.A., 1980. Collagen production by
cultured retinal capillary pericytes. Invest. Ophthalmol. Visual Sci. 19,
90–94.
Couch, J.A., 1990. Pericyte of a teleost fish: ultrastructure, position, and
role in neoplasia as revealed by a fish model. Anat. Rec. 228, 7–14.
Craighead, H.G., James, C.D., Turner, A.M.P., 2001. Chemical and
topographical patterning for directed cell attachment. Curr. Opin. Solid
State Mater. Sci. 5, 177–184.
Curtis, A., Wilkinson, C., 1997. Topographical control of cells. Biomate-
rials 18, 1573–1583.
Daxecker, H., Raab, M., Markovic, S., Karimi, A., Griesmacher, A.,
Mueller, M.M., 2002. Endothelial adhesion molecule expression in an
in vitro model of inflammation. Clin. Chim. Acta 325, 171–175.
De Santis, R., Mollica, F., Ambrosio, L., Nicolais, L., Ronca, D., 2003.
Dynamic mechanical behaviour of PMMA based bone cements in wet
environment. J. Mater. Sci.: Mater. Med. 14, 583–594.
Dee, K.C., Bizios, R., 1996. Mini-review: proactive biomaterials and bone
tissue engineering. Biotechnol. Bioeng. 50, 438–442.
Dewez, J.L., Doren, A., Schneider, Y.J., Rouxhet, P.G., 1999. Competitive
adsorption of proteins: key of the relationship between substratum
surface properties and adhesion of epithelial cells. Biomaterials 20,
547–559.
Diaz-Flores, L., Gutierrez, R., Lopez-Alonso, A., Gonzalez, R., Varela, H.,
1992. Pericytes as a supplementary source of osteoblasts in periosteal
osteogenesis. Clin. Orthop. 275, 280–286.
Dong, J., Kojima, H., Uemura, T., Kikuchi, M., Tateishi, T., Tanaka, J.,
2001. In vivo evaluation of a novel porous hydroxyapatite to sustain
osteogenesis of transplanted bone marrow-derived osteoblastic cells.
J. Biomed. Mater. Res. 57, 208–216.
Erbil, H.Y., McHalem, G., Rowan, S.M., Newton, M.I., 1999. Determi-
nation of the receding contact angle of sessile drops on polymer
surfaces by evaporation. Langmuir 15, 7378–7385.
Fredriksson, C., Khilman, S., Kasemo, B., Steel, D.M., 1998. In vitro real-
time characterization of cell attachment and spreading. J. Mater. Sci.:
Mater. Med. 9, 785–788.
Grayson, A.C.R., Shawgo, R.S., Johnson, A.M., Flynn, N.T., Li, Y.W.,
Cima, M.J., Langer, R.A., 2004. BioMEMS Review: MEMS technol-
ogy for physiologically integrated devices. Proc. IEEE 92, 6–21.
Greenwood, J., Pryce, G., Devine, L., Male, D.K., dos Santos, W.L.,
Calder, V.L., Adamson, P., 1996. SV40 large T immortalised cell lines
of the rat blood–brain and blood–retinal barriers retain their phenotypic
and immunological characteristics. J. Neuroimmunol. 71, 51–63.
Hirschi, K.K., D’Amore, P.A., 1996. Pericytes in the microvasculature.
Cardiovasc. Res. 32, 687–698.
Hongbing, L., Hamers, R.J., 1998. An X-ray photoelectron spectroscopy
study of the bonding of unsaturated organic molecules to the Si(001)
surface. Surf. Sci. 416, 354–362.
Hunkeler, D., 1997. Polymers for bioartificial organs. Trends Polym. Sci. 5,
286–293.
Ivanov, D., Philippova, M., Antropova, J., Gubaeva, F., Iljinskaya, O.,
Tararak, E., Bochkov, V., Erne, P., Resink, T., Tkachuk, V., 2001.
Expression of cell adhesion molecule T-cadherin in the human
vasculature. Histochem. Cell Biol. 115, 231–242.
Kilpadi, K.L., Chang, P.L., Bellis, S.L., 2001. Hydroxyapatite binds more
serum proteins, purified integrins, and osteoblast precursor cells than
titanium or steel. J. Biomed. Mater. Res. 57, 258–267.
Kusakabe, M., Suzuki, Y., Kaibara, M., Iwaki, M., Sasabe, H., 1995. Cell
adhesion control by ion implantation into polymeric materials and
extra-cellular matrix. Radiat. Phys. Chem. 46, 263–267.
Lupo, G., Anfuso, C.D., Ragusa, N., Strosznajder, R.P., Walski, M.,
Alberghina, M., 2001. t-Butyl hydroperoxide and oxidized low
density lipoprotein enhance phospholipid hydrolysis in lipopolysac-
charide-stimulated retinal pericytes. Biochim. Biophys. Acta 1531,
143–155.
Mandarino, L.J., Sundarraj, N., Finlayson, J., Hassel, J.R., 1993.
Regulation of fibronectin and laminin synthesis by retinal capillary
endothelial-cells and pericytes in vitro. Exp. Eye Res. 57, 609–621.
Marletta, G., Satriano, C., 2004. Irradiation-controlled adsorption and
organization of biomolecules on surfaces: from the nanometric to the
mesoscopic level. In: Buzaneva, E., Scharff, P. (Eds.), Frontiers in
Molecular-Scale Science and Technology of Nanocarbon, Nanosilicon
and Biopolymer Multifunctional Nanosystems. Kluwer Academic
Publishers, pp. 1–26.
Marx, K.A., Zhou, T., Warren, M., Braunhut, S.J., 2003. Quartz crystal
microbalance study of endothelial cell number dependent differences in
initial adhesion and steady-state behaviour: evidence for cell–cell
cooperativity in initial adhesion and spreading. Biotechnol. Prog. 19,
987–999.
McFarland, C.D., Mayer, S., Scotchford, C., Dalton, B.A., Steele, J.G.,
Downes, S., 1999. Attachment of cultured human bone cells to novel
polymers. J. Biomed. Mater. Res. 44, 1–11.
Najman, S., Savic, V., Djordjevic, L., Ignjatovic, N., Uskokovic, D., 2004.
Biological evaluation of hydroxyapatite/poly-l-lactide (HAp/PLLA)
composite biomaterials with poly-l-lactide of different molecular
weights intraperitoneally implanted into mice. Bio-Med. Mater. Eng.
14, 61–70.
Nimeri, G., Fredriksson, C., Elwing, H., Liu, L., Rodahl, M., Kasemo, B.,
1998. Neutrophil interaction with protein-coated surfaces studied by an
extended quartz crystal microbalance technique. Colloids Surf., Bio-
interfaces 11, 255–264.

ARTICLE IN PRESSG. Assero et al. / Microvascular Research xx (2004) xxx–xxx12
Ohgushi, H., Dohi, Y., Yoshikawa, T., Tamai, S., Tabata, S., Okunaga, K.,
Shibuya, T., 1996. Osteogenic differentiation of cultured marrow
stromal stem cells on the surface of bioactive glass ceramics. J.
Biomed. Mater. Res. 32, 341–348.
Ohsawa, K., Neo, M., Matsuoka, H., Akiyama, H., Ito, H., Nakamura, T.,
2001. Tissue responses around polymethylmethacrylate particles
implanted into bone: analysis of expression of bone matrix protein
mRNAs by in situ hybridisation. J. Biomed. Mater. Res. 54, 501–508.
Pu, F.R., Williams, R.L., Markkula, T.K., Hunt, J.A., 2002. Expression of
leukocyte endothelial cell adhesion molecules on monocyte adhesion to
human endothelial cells on plasma treated PET and PTFE in vitro.
Biomaterials 23, 4705–4718.
Ramires, P.A., Giuffrida, A., Milella, E., 2002. Three-dimensional
reconstruction of confocal laser microscopy images to study the
behaviour of osteoblastic cells grown on biomaterials. Biomaterials
23, 397–406.
Reilly, T.M., Seldes, R., Luchetti, W., Brighton, C.T., 1998. Similarities in
the phenotypic expression of pericytes and bone cells. Clin. Orthop.
346, 95–103.
Richardson, R.L., Hausman, G.J., Campion, D.R., 1982. Response of
pericytes to thermal lesion in the inguinal fat pad of 10-day-old rats.
Acta Anat. 114, 41–57.
Rizzi, S.C., Heath, D.T., Coombes, A.G.A., Bock, N., Textor, M., Downes,
S., 2001. Biodegradable polymer/hydroxyapatite composites: surface
analysis and initial attachment of human osteoblasts. J. Biomed. Mater.
Res. 55, 475–486.
Satriano, C., Marletta, G., Conte, E., 1999. Cell adhesion on low-energy ion
beam-irradiated polysiloxane surfaces. Nucl. Instrum. Methods, B 148,
1079–1084.
Satriano, C., Marletta, G., Conte, E., 2001. Surface chemical structure and
cell adhesion onto ion beam modified polysiloxane. Langmuir 17,
2243–2250.
Satriano, C., Carnazza, S., Guglielmino, S., Marletta, G., 2002a. Differ-
ential cultured fibroblast behaviour onto plasma and ion beam-modified
polysiloxane surfaces. Langmuir 18, 9469–9475.
Satriano, C., Marletta, G., Conte, E., 2002b. Surface chemical structure and
cell adhesion onto ion beam modified polysiloxane. Langmuir 17,
2243–2250.
Satriano, C., Marletta, G., Carnazza, S., Guglielmino, S., 2003a. Protein
adsorption and fibroblast adhesion on irradiated polysiloxane surfaces.
J. Mater. Sci.: Mater. Med. 14, 663–670.
Satriano, C., Carnazza, S., Guglielmino, S., Marletta, G., 2003b. Surface
free energy and cell attachment onto ion-beam irradiated polymer
surfaces. Nucl. Instrum. Methods, B 208, 287–293.
Satriano, C., Carnazza, S., Licciardello, A., Guglielmino, S., Marletta, G.,
2003c. Cell adhesion and spreading on polymer surfaces micropatterned
by ion beams. J. Vac. Sci. Technol., A 21, 1145–1151.
Satriano, C., Spinella, N., Manso, M., Licciardello, A., Rossi, F., Marletta,
G., 2003d. Ion beam induced nanometric structure and oligopeptides
adsorption on patterned polymer surfaces. Mater. Sci. Eng., C 23,
779–786.
Seah, M.P., Brown, M.T., 1999. Validation and accuracy of peak synthesis
software for XPS. Appl. Surf. Sci. 144–145, 183–187.
Stramm, L.E., Li, W., Aguirre, G.D., Rockey, J.H., 1987. Glycosamino-
glycan synthesis and secretion by bovine retinal capillary pericytes in
culture. Exp. Eye Res. 44, 17–28.
Suzuki, N., Iimura, K., Satoh, S., Saito, Y., Kato, T., Tanaka, A., 1997.
Model for analysis of XPS electron taken-characterized Langmuir–
Blodgett film. Surf. Interface Anal. 25, 650–659.
Teare, D.O.H., Emmison, N., Ton-That, C., Bradley, R.H., 2000. Cellular
attachment to ultraviolet ozone modified polystyrene surfaces. Lang-
muir 16, 2818–2824.
Thomas, W.E., 1999. Brain macrophages: on the role of pericytes and
perivascular cells. Review. Brain Res. 31, 42–57.
Tilling, T., Engelbertz, C., Decker, S., Korte, D., Huwel, S., Galla, H.J.,
2002. Expression and adhesive properties of basement membrane
proteins in cerebral capillary endothelial cell cultures. Cell Tissue Res.
310, 19–29.
van Oss, C.J., 2002. Use of the combined Lifshitz–van der Waals and Lewis
acid-base approaches in determining the apolar and polar contributions
to surface and interfacial tensions and free energies. J. Adhes. Sci.
Technol. 16, 669–677.
Wang, D.A., Ji, J., Sun, Y.H., Shen, J.C., Feng, L.X., Elisseeff, J.H., 2002.
In situ immobilization of proteins and RGD peptide on polyurethane
surfaces via poly(ethylene oxide) coupling polymers for human
endothelial cell growth. Biomacromolecules 3, 1286–1295.
Watanabe, J., Eriguchi, T., Ishihara, K., 2002. Cell adhesion and
morphology in porous scaffold based on enantiomeric poly(lactic acid)
graft-type phospholipid polymers. Biomacromolecules 3, 1375–1383.
Willner, I., Willner, B., 2001. Biomaterials integrated with elecQ
tronic elements: en route to bioelectronics. Trends Biotechnol. 19,
222–230.
Wong, D., Dorovini-Zis, K., 1995. Expression of vascular cell adhesion
molecule-1 (VCAM-1) by human brain microvessel endothelial cells in
primary culture. Microvasc. Res. 49, 325–339.
Yang, Y., Tian, J., Deng, L., Ong, J.L., 2002. Morphological behaviour of
osteoblast-like cells on surface-modified titanium in vitro. Biomaterials
23, 1383–1389.
Zhang, S., Yan, L., Altman, M., L7ssle, M., Nugent, H., Frankel, F.,
Lauffenburger, D.A., Whitesides, G.M., Rich, A., 1999. Biological
surface engineering: a simple system for cell pattern formation.
Biomaterials 20, 1213–1220.
Zreiqat, H., McFarland, C., Howlett, C.R., 1999. The effect of polymeric
chemistry on the expression of bone-related mRNAs and proteins by
human bone-derived cells in vitro. J. Biomater. Sci., Polym. Ed. 10,
199–216.
Copyright © 2022 FDOKUMEN




















