
Peres, M. B., Klippel, S. (2003). Reproductive Biology of Southwestern Atlantic Wreckfish, Polyprion...
11
Environmental Biology of Fishes 68: 163–173, 2003. © 2003 Kluwer Academic Publishers. Printed in the Netherlands. Reproductive biology of southwestern Atlantic wreckfish, Polyprion americanus (Teleostei: Polyprionidae) Mˆ onica B. Peres a & Sandro Klippel b a Servic ¸o da Regi˜ ao Litoral, DQA, FEPAM Rua Carlos Chagas 55 sala 707, Centro Porto Alegre, RS 90.030-020, Brazil (e-mail: [email protected]) b Talha-mar Projetos Ambientais, Cons. d’Avila 190 Porto Alegre, RS 91040-450, Brazil Received 5 February 2003 Accepted 12 July 2003 Key words: sexual maturity, fecundity, spawning aggregation, spawning season, migration, deep-sea fish, life-cycle closure, southern Brazil Synopsis We report on the reproductive biology of southwestern Atlantic wreckfish. Females mature first at 77.9cm total length (TL) (10.4 years) and all are mature by 90 cm TL (15.2 years). Males mature first at 74.9 cm (9 years) and all are mature by 80 cm TL (10.9 years). The wreckfish is a gonochoristic multiple spawner and the gonadal cycle is synchronized at the population level. Spawning occurs from late July to early October along the continental slope (>300 m). Ovarian fecundity varies from 3 to 11.9 million (135–311 oocytes × g −1 ) and increases exponentially with length. Spawning at western boundary current systems, maintained by homing of adults, is a basic requirement for self-sustaining populations of this species. Introduction The wreckfish, Polyprion americanus (Bloch and Schneider, 1801), is a large, long-lived demersal teleost that inhabits continental and oceanic island slopes of temperate and subtropical waters of the northern and southern Atlantic Ocean, Mediterranean, and south- ern Indian and southwestern Pacific Ocean (Heemstra 1986, Sedberry et al. 1999). Along the southwestern Atlantic the species occurs from 23 ◦ S, near Rio de Janeiro – Brazil, to 46 ◦ S in Argentina (Cousseau & Perrota 1998, Peres 2000). The only published study on reproduction (Roberts 1989), reports southwestern Pacific wreckfish as a gonochoristic teleost. In New Zealand bottom trawl samples, females attained larger body sizes than males and sex ratios were close to 1 : 1. Other related information is that wreckfish eggs and larvae are pelagic (Hardy 1978) and that for southwestern Pacific, both Polyprion species undergo a northern spawning migration (Roberts 1996, Beentjes & Francis 1999). For North Atlantic stocks, 8–12-year-old wreckfish are mature and juveniles may live more than 2 years at the sea surface before recruitment to the bottom (Sedberry et al. 1999). Despite its worldwide distri- bution and high commercial importance, there is no information on reproductive cycle, fecundity, spawn- ing sites and season of these fish. The development of a method to sample whole fish from commercial landings permitted us to determine: (1) size and age at maturity; (2) reproductive cycle; (3) spawning season and sites; (4) ovarian fecundity (OF); (5) sex ratio. Materials and methods The study area was the continental shelf and slope off southern Brazil, from 27 ◦ 56’S to 34 ◦ 49’S (70–590 m deep). It was divided in two sub-areas, north and south of Rio Grande (32 ◦ 13’S) (Figure 1). Southwestern Atlantic wreckfish is landed and marketed ice-cooled and whole, at large body sizes and high prices. Thus, sampling procedures for commercial landings required fast and non-damaging techniques. Fish were measured
Transcript of Peres, M. B., Klippel, S. (2003). Reproductive Biology of Southwestern Atlantic Wreckfish, Polyprion...

Environmental Biology of Fishes 68: 163–173, 2003.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
Reproductive biology of southwestern Atlantic wreckfish,Polyprion americanus (Teleostei: Polyprionidae)
Monica B. Peresa & Sandro Klippelb
aServico da Regiao Litoral, DQA, FEPAM Rua Carlos Chagas 55 sala 707, Centro Porto Alegre,RS 90.030-020, Brazil (e-mail: [email protected])bTalha-mar Projetos Ambientais, Cons. d’Avila 190 Porto Alegre, RS 91040-450, Brazil
Received 5 February 2003 Accepted 12 July 2003
Key words: sexual maturity, fecundity, spawning aggregation, spawning season, migration, deep-sea fish,life-cycle closure, southern Brazil
Synopsis
We report on the reproductive biology of southwestern Atlantic wreckfish. Females mature first at 77.9 cm totallength (TL) (10.4 years) and all are mature by 90 cm TL (15.2 years). Males mature first at 74.9 cm (9 years) and allare mature by 80 cm TL (10.9 years). The wreckfish is a gonochoristic multiple spawner and the gonadal cycle issynchronized at the population level. Spawning occurs from late July to early October along the continental slope(>300 m). Ovarian fecundity varies from 3 to 11.9 million (135–311 oocytes × g−1) and increases exponentiallywith length. Spawning at western boundary current systems, maintained by homing of adults, is a basic requirementfor self-sustaining populations of this species.
Introduction
The wreckfish, Polyprion americanus (Bloch andSchneider, 1801), is a large, long-lived demersal teleostthat inhabits continental and oceanic island slopes oftemperate and subtropical waters of the northern andsouthern Atlantic Ocean, Mediterranean, and south-ern Indian and southwestern Pacific Ocean (Heemstra1986, Sedberry et al. 1999). Along the southwesternAtlantic the species occurs from 23◦S, near Rio deJaneiro – Brazil, to 46◦S in Argentina (Cousseau &Perrota 1998, Peres 2000).
The only published study on reproduction (Roberts1989), reports southwestern Pacific wreckfish as agonochoristic teleost. In New Zealand bottom trawlsamples, females attained larger body sizes than malesand sex ratios were close to 1 : 1. Other relatedinformation is that wreckfish eggs and larvae arepelagic (Hardy 1978) and that for southwestern Pacific,both Polyprion species undergo a northern spawningmigration (Roberts 1996, Beentjes & Francis 1999).For North Atlantic stocks, 8–12-year-old wreckfish
are mature and juveniles may live more than 2 yearsat the sea surface before recruitment to the bottom(Sedberry et al. 1999). Despite its worldwide distri-bution and high commercial importance, there is noinformation on reproductive cycle, fecundity, spawn-ing sites and season of these fish. The developmentof a method to sample whole fish from commerciallandings permitted us to determine: (1) size and age atmaturity; (2) reproductive cycle; (3) spawning seasonand sites; (4) ovarian fecundity (OF); (5) sex ratio.
Materials and methods
The study area was the continental shelf and slope offsouthern Brazil, from 27◦56’S to 34◦49’S (70–590 mdeep). It was divided in two sub-areas, north and southof Rio Grande (32◦13’S) (Figure 1). SouthwesternAtlantic wreckfish is landed and marketed ice-cooledand whole, at large body sizes and high prices. Thus,sampling procedures for commercial landings requiredfast and non-damaging techniques. Fish were measured

164
Figure 1. Study area of wreckfish, P. americanus, divided into two sub-areas off southern Brazil, (N) north and (S) south of Rio Grande.Area of capture of 27 females with hydrated oocytes (grayish) and of eight other, at five fishing sites (full circles), is shown.
from the posterior border of the eye orbit to the fork ofthe caudal fin (EF in cm) with a flexible metallic tapeto the nearest cm. EF was converted to total length (TLin cm) by the relationship, TL = 119.5 × EF − 0.0724(n = 125; r2 = 0.997; 44–192 cm TL). Sample weightswere estimated with the total weight–TL relationshipTW (kg) = 6.29 × 10−6 × TL3.21 (n = 207; 44–174 cmTL; r2 = 0.98).
We determined sex for 641 randomly chosen indi-viduals, 420 females (74–174 cm TL) and 221 males(73–126 cm TL), by visual examination of sub-samplesof the posterior end of the gonads. Sub-samples were
removed with a surgical curette through the urogeni-tal pore (0.1–0.3 ml). This was sufficient to assign sexand to provide qualitative information of the macro-scopic aspect of ovarian tissue, registered at dockside.Whenever possible, we preserved an ovary sub-sample(0.5–2 ml) in 2% formalin to assess the frequency dis-tribution of oocyte diameters (OD) and their generalmicroscopic aspect (16×). Sub-sampling of immaturefish was not possible as gonads are small and tis-sues are tightly compacted (n = 82). We removedthe left otolith through the opercular opening (420females, 221 males and 82 unsexed) and estimated

165
age by opaque band counts of transverse otolith sec-tions (Peres 2000). Ice-preserved whole gonads andlengths of other 114 females (45–174 cm TL) and 86males (44–119 cm TL) were provided by fishers. Addi-tional gonads of 67 females (46–148 cm TL) and 71males (44–108 cm TL) were furnished from researchcruises. We used exact binomial tests to determine if sexratios differed from the expected 1 : 1. We computedSpearman’s rho coefficient (rs) to determine the corre-lation between OF and TL. We performed all statisticalanalyses in ‘R’ environment (Ihaka & Gentleman1996).
Wreckfish gonads may weight up to 4 or 5 kg andwe sampled most of them from commercial catches,after being ice-cooled for 10–25 days. Thus, histolog-ical sections were of poor quality and were only usedto confirm the criteria for gonad staging. We did notquantify the atresia rate but its seemed negligible. Wecould determine gonad staging in a five-point scale for462 females (410 adults) (Table 1). Gonad staging formale wreckfish sub-samples was not possible duringcommercial landings and so we did not assess it inthis study. We separated ovarian follicles, here calledoocytes, with diameters greater than 0.05 mm, alignedand measured in a random axis (16×) for 314 femalewreckfish (291 adults). For 181 females and 157 males,we used fresh gonad weight (GW in kg) to calculate thegonad-somatic index, GSI = (GW/EW) ×100, whereEW (kg) is the eviscerated somatic weight, estimatedby the relationship EW = 9 × 10−6 × TL3.12 (n = 141;r2 = 0.98). We estimated the total number of yolked
oocytes (YO, 0.6 < OD < 1.4 mm), referred here asfemale OF, for 73 females without hydrated oocytes,using the gravimetric method. All YO in a sample of0.2–0.4 g of ovarian tissue, preserved in 10% formalin,were separated from the interstitial tissue, counted andextrapolated for preserved whole ovary weight usingthe relationship, OF = n◦YO × (GW/sample weight).We estimated relative ovary fecundity (ROF) using therelationship: ROF (n◦YO × g−1) = OF/EW. We con-sidered the presence of hydrated oocytes as evidenceof spawning within a few hours (Hunter & Macewicz1985) and used this to determine spawning season andspawning sites. We classified adult males by macro-scopic visual aspect of the non-preserved testes orby the presence of sperm in gonad sub-sample. Weclassified females by: (1) macroscopic visual aspectof the non-preserved ovarian and oocytes; (2) micro-scopic general aspect of preserved whole oocytes and,(3) the microscopic aspect of the histological sections,when available. We calculated length at first maturity(Lm = 50% maturity) by PROBIT regression of adultproportion for each length class (5 cm class intervals)during the active reproductive period. We obtainedages at first maturity from the relationship, Age = t0–(1/K) × ln (1−(Lm/L∞)), where t0, K and L∞ are thevon Bertalanffy growth model parameters obtained forfemales (−6.80 years, 0.054 years−1 and 129.5 cm)and males (−4.69 years, 0.084 years−1 and 109.5 cm)wreckfish (Peres 2000). All observed females greaterthan 90 cm TL were clearly mature. As mature rest-ing and immature females are difficult to distinguish,
Table 1. Monthly sample sizes and body length ranges (TL in cm) of wreckfish P. americanus, assessed forgonadal stages (GSt), gonad-somatic index (GSI), oocyte diameter distribution (OD) and ovarian fecundity(OF). In the parenthesis is the number of adult fishes examined.
Month Female Male
GSt GSI OD OF TL GSI TL
Jan. 14 (14) — 14 (14) — 91–130 — —Mar. 16 (16) 2 16 (16) — 90–155 3 77–91Apr. 49 (38) 31 48 (38) 20 55–137 51 58–108May. 63 (60) 63 59 (58) 27 71–148 26 88–109Jun. 34 (31) 4 33 (31) 1 79–137 2 73–78Jul. 105 (100) 22 47 (46) 14 46–137 16 57–112Aug. 16 (6) 16 11 (6) 6 47–174 35 48–105Sept. 105 (90) 37 53 (49) 5 48–140 19 56–100Oct. 37 (36) 2 17 (17) — 87–134 2 72–73Nov. 4 (0) 4 — — 47–49 2 44–47Dec. 19 (19) 0 16 (16) — 92–122 1 103
TL 46–174 46–174 55–174 74–174 46–174 44–112
Total 462 (410) 181 314 (291) 73 157

166
monthly GSI analysis included all females greater than90 cm TL as adults.
Results
Gonad structure and development
Wreckfish ovary and testis organization follow thegeneral patterns for a gonochoristic teleost species.Both are paired organs, joined posteriorly and con-nected medially to the coelom wall by a mesentery.Ovaries have circular sections, radial lamellar orga-nization, a central lumen and a blood vessel networkaround the external wall. Testes are compact with atriangular section, tubular organization, and a dorsalinvagination with a central vascular bundle and thespermatic duct. In the ovary, oocytes are produced andripened within the lamellae, ovulated in batches into thelumen, and released externally through the urogenital
pore. OD frequency distribution for each ovarian stageshows the sequence of ovary development (Figure 2).Oocyte color, aspect and diameter change with devel-opment (Table 2). Description of immature, resting,early and late development, running-ripe and spentovaries are given in Table 3. In seven partially spawnedwhole ovaries (Sep.–Oct.), most oocytes were in lateyolked stage and gonads were flaccid with hemorrhagicportions.
The presence of partially spawned females, OD dis-tributions of running-ripe females with three modalgroups (immature, yolked and ripe oocytes) and sev-eral oocyte stages in the same histological section, areevidence that the species is a multiple spawner.
Reproductive cycle
With gamete and gonad development, GSI monthlymean values of adult fish increased from March to July,
Figure 2. OD distribution for each ovary stage (n = 100) of wreckfish, P. americanus.

167
reaching 8.8 for females and 5.5 for males (Figure 3).A gradual decline of GSI and higher SE from Julyto September are evidence that these are the spawn-ing months when successive ripening and liberationof gametes are responsible for the high variabilityand overall decrease in GW. Monthly relative fre-quency of adult female GSt confirmed the timing ofgonad development and spawning, obtained from theGSI data (Figure 4). Early development was frequentfrom March to June and late development from Julyto August. We observed running-ripe females in lateJuly, September and early October. Spent and resting
Table 2. Oocyte stages of wreckfish, P. americanus. Color, diam-eters (OD) and general microscopic aspect (16× magnification).
Immature oocytes transparent, colorless, inconspicuous to thenaked eye (OD 0.15–0.2 mm). Whole oocytes are difficult toseparate, a thin oocyte wrapper maybe visible
Early YO opaque-whitish to yellow (OD 0.3–0.7 mm), have agranular cytoplasm (30–80% of oocyte volume) with two tothree halos and a well-defined wrapper
Late YO bright yellow to coral-orange (OD 0.6–1.4 mm),compact yolk layers and a distinct wrapper. Whole oocytesare easy to separate
Ripe or hydrated oocytes in ovulating females are large(OD 1.5–2.2 mm), uniform-hyaline, whitish afterpreservation, with one to three oil droplets and a thick,well-developed wrapper
females were common from September to Januaryand almost absent from March to August. Despitegathering data over several years, the monthly ODdistribution of pooled adult female weighed by thenumber of measured oocytes showed that the reproduc-tive cycle is quite synchronized at the population level(Figure 5).
The gradual increase of yolked OD throughout theactive reproductive period (Figure 6), and OD monthlydistributions with no more than three modes and agap of OD between non-yolked and YO from Juneon (Figure 5), are evidence that prior to the spawn-ing months (late July–early October) there is no morerecruitment of immature oocytes and that wreckfish OFis determinate (Hunter et al. 1992, Gunderson 1993).
Ovarian fecundity
Relative ovarian fecundity increased from April to July,indicating that YO are not totally recruited, at least byApril and May (Figure 7). Decreasing relative fecun-dity from July to September is additional evidence thatthese are the spawning months. The number of late YOincreased with female body length (Figure 8). For 11females caught from 1 June to 15 July, when fecundityis already determined and spawning was not observed,OF varied from 3 million oocytes (116 cm TL) to 11.9million (137 cm TL) and was significantly correlated
Table 3. Ovarian stages of wreckfish, P. americanus. Criteria included: (1) the macroscopic visual aspect of non-preservedovaries; (2) general microscopic aspect of ovarian tissue and oocyte stages present (descriptions in Table 2); (3) GSI range.
Immature ovaries (1) are cylindrical, filiform, compact, pink, translucent, with no visible oocytes. (2) Compact tissue,no spacing between ovarian lamellae, 100% immature oocytes. Thin ovarian capsules. (3) GSI 0.5
Resting ovaries (1) are rounded, opaque-pink, oocytes difficult to see with naked eyes, developing peripheral bloodvessel networks. (2) Compact tissue. Immature and very early YO are present. Ovarian capsule thicker thanimmature. (3) GSI 0.5–1.5
Early developed ovaries (1) are swollen, yellow–orange, with marked peripheral blood vessel networks, oocytesvisible to the naked eye. (2) Immature and early YO, developing radial septa from a thickening ovarian capsule.(3) GSI 1–5
Late developed ovaries (1) are enlarged and brittle, difficult to handle due to extended ovarian walls, easily visibleoocytes. Developed peripheral blood vessel networks with or without hemorrhagic portions. (2) Late YO comprised80–95% of sample volume and the rest, immature oocytes. Some ripe oocytes from earlier batches may be present.(3) GSI 4–15
Running ripe ovaries (1) are very enlarged, part or totally transparent due to ripe oocytes that are easily released.Ovarian capsules are distended to the maximum. There are highly developed peripheral blood vessel networks.(2) Ripe oocytes comprised 40–100% of sample volume; late YO may still be present. (3) GSI 9–18
Spent ovaries (1) are small, emptied, reddish, totally or partly hemorrhagic, visible oocytes may be present.(2) Thickened ovarian walls and septa. Spaced ovarian lamellae with hanging tissue like ‘grape bunches’. Immatureand very early YO are present. Some ovaries contained transparent structures similar to ‘empty wrappers’ (supposedto be postovulatory follicles) and/or remaining ripe oocytes of earlier batches. (3) GSI 2–3
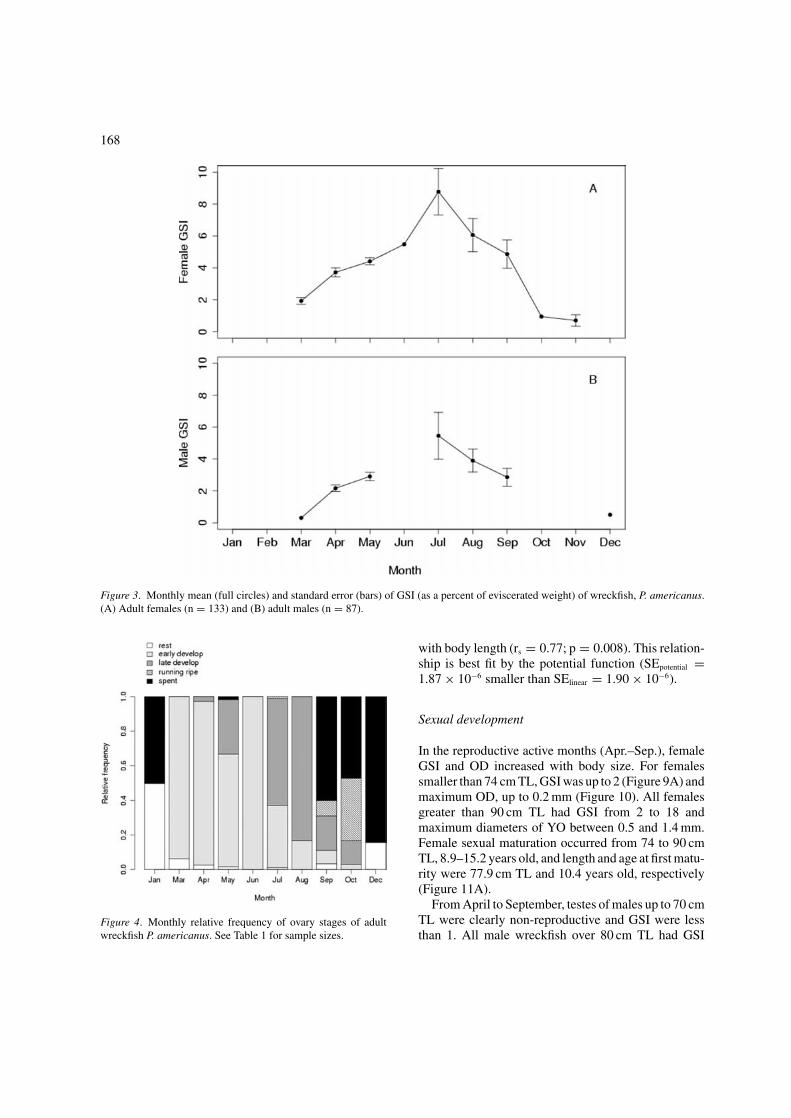
168
Figure 3. Monthly mean (full circles) and standard error (bars) of GSI (as a percent of eviscerated weight) of wreckfish, P. americanus.(A) Adult females (n = 133) and (B) adult males (n = 87).
Figure 4. Monthly relative frequency of ovary stages of adultwreckfish P. americanus. See Table 1 for sample sizes.
with body length (rs = 0.77; p = 0.008). This relation-ship is best fit by the potential function (SEpotential =1.87 × 10−6 smaller than SElinear = 1.90 × 10−6).
Sexual development
In the reproductive active months (Apr.–Sep.), femaleGSI and OD increased with body size. For femalessmaller than 74 cm TL, GSI was up to 2 (Figure 9A) andmaximum OD, up to 0.2 mm (Figure 10). All femalesgreater than 90 cm TL had GSI from 2 to 18 andmaximum diameters of YO between 0.5 and 1.4 mm.Female sexual maturation occurred from 74 to 90 cmTL, 8.9–15.2 years old, and length and age at first matu-rity were 77.9 cm TL and 10.4 years old, respectively(Figure 11A).
From April to September, testes of males up to 70 cmTL were clearly non-reproductive and GSI were lessthan 1. All male wreckfish over 80 cm TL had GSI
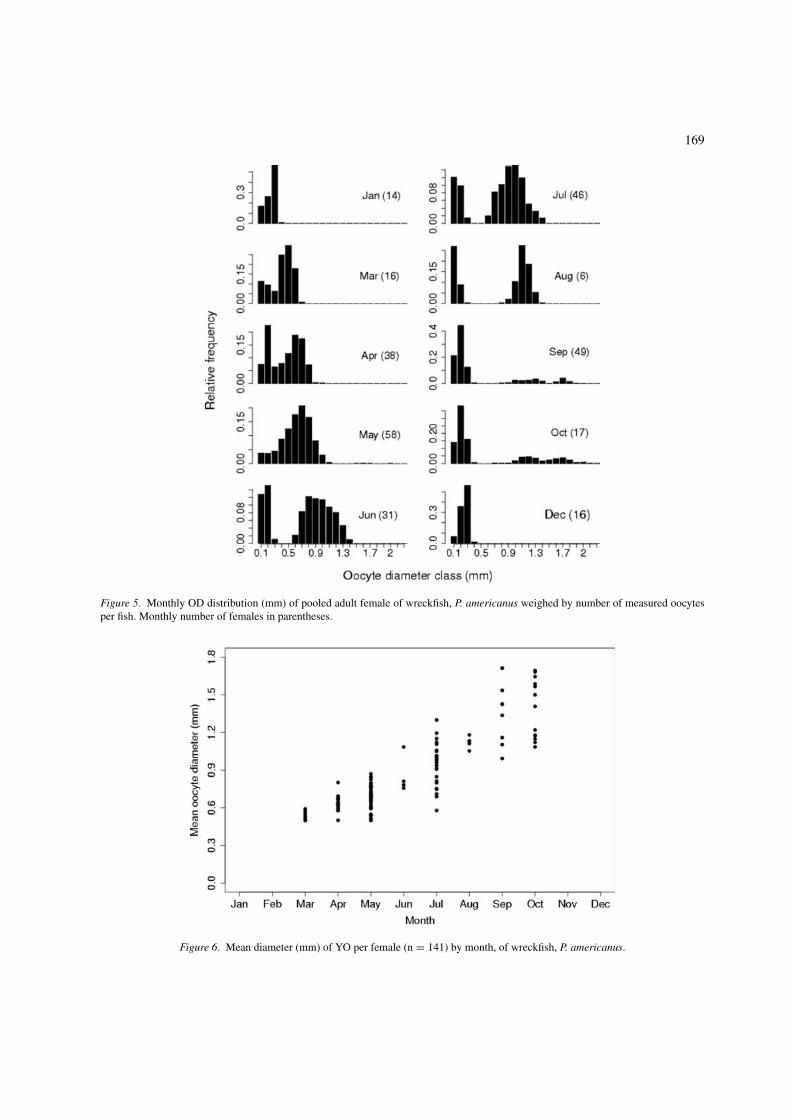
169
Figure 5. Monthly OD distribution (mm) of pooled adult female of wreckfish, P. americanus weighed by number of measured oocytesper fish. Monthly number of females in parentheses.
Figure 6. Mean diameter (mm) of YO per female (n = 141) by month, of wreckfish, P. americanus.

170
from 1 to 14 and were adult (Figure 9B). Male sexualmaturation occurred from 70 to 80 cm TL, 7.4 to 10.9years old, and length and age at first maturity were74.9 cm TL and 9 years old (Figure 11B).
Figure 7. Monthly ROF of wreckfish, P. americanus. ROF as thenumber of late YO (0.6–1.4 mm) per gram of eviscerated bodyweight. Median and interquartile ranges (boxes), minimum andmaximum values (dashed lines) and outliers, values outside 1.5times of the first and third quartile (empty circles). See Table 1for sample sizes.
Spawning
We caught a total of 22 running-ripe females and 13others with remaining hydrated oocytes in their ovariesalong the continental slope, in water depths of 300–500 m (Figure 1). Most of them (n = 34, 97%) werecaptured north of Rio Grande and 77% (n = 27) weresampled from three huge commercial catches: 29 July1995 (13 t of wreckfish), 9 September 1995 (26.6 t)and 4 October 1995 (16.4 t), at a fishing area knownas ‘Tramandaı-Solidao’ (30◦05′S, 048◦W–30◦45′S,048◦30′W, 360–460 m deep). We recorded males (n =30) with distended testicles (GSI > 5) and sperm beingspontaneously released at these same catches.
The relative number of running-ripe females at thesethree landings increased from 2% in July (one out of 53sampled adult females), to 10% in September (six of60) and to 36% in October (13 of 36). Late developmentstage frequency was high in July (98%) and dropped inthe following two landings (18% in Sep., 17% in Oct.).Spent females were completely absent in July, were72% in September and 47% in October. Of 179 sampledadult wreckfish, 149 (83%) were female. The sex ratiowas significant different from 1 : 1 (p �0.001).
Wreckfish spawning occurs at the continental slope(300–500 m deep) off southern Brazil, especially atTramandaı-Solidao fishing area. Commercial catchesat spawning sites and season were highly female biased(p � 0.001).
Figure 8. Relationship of (OF ×10−6) and body TL (in cm) of wreckfish, P. americanus. OF as the number of late YO (0.6–1.4 mm).Crosses for April and May samples (n = 47), open circles for 15 July to September (n = 15) and full circles for 1 June to 15 July(n = 11). Equation, SE and df of the relationship (line) for 1 June to 15 July samples are shown.
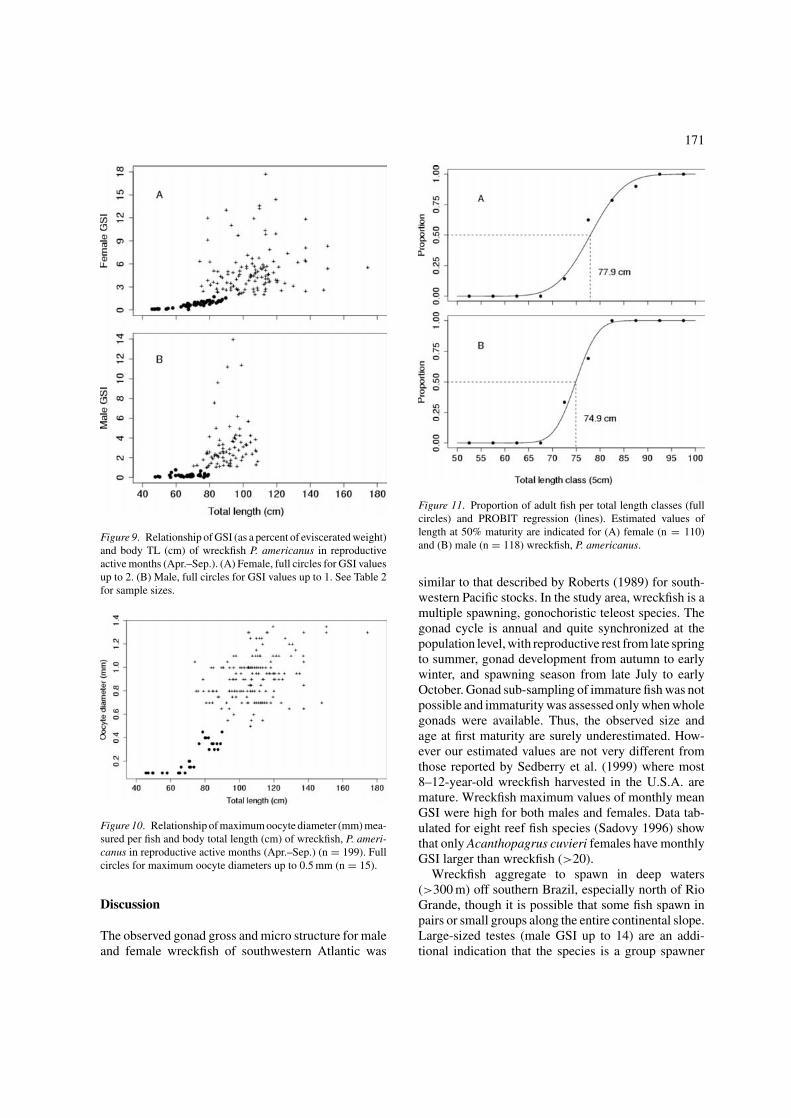
171
Figure 9. Relationship of GSI (as a percent of eviscerated weight)and body TL (cm) of wreckfish P. americanus in reproductiveactive months (Apr.–Sep.). (A) Female, full circles for GSI valuesup to 2. (B) Male, full circles for GSI values up to 1. See Table 2for sample sizes.
Figure 10. Relationship of maximum oocyte diameter (mm) mea-sured per fish and body total length (cm) of wreckfish, P. ameri-canus in reproductive active months (Apr.–Sep.) (n = 199). Fullcircles for maximum oocyte diameters up to 0.5 mm (n = 15).
Discussion
The observed gonad gross and micro structure for maleand female wreckfish of southwestern Atlantic was
Figure 11. Proportion of adult fish per total length classes (fullcircles) and PROBIT regression (lines). Estimated values oflength at 50% maturity are indicated for (A) female (n = 110)and (B) male (n = 118) wreckfish, P. americanus.
similar to that described by Roberts (1989) for south-western Pacific stocks. In the study area, wreckfish is amultiple spawning, gonochoristic teleost species. Thegonad cycle is annual and quite synchronized at thepopulation level, with reproductive rest from late springto summer, gonad development from autumn to earlywinter, and spawning season from late July to earlyOctober. Gonad sub-sampling of immature fish was notpossible and immaturity was assessed only when wholegonads were available. Thus, the observed size andage at first maturity are surely underestimated. How-ever our estimated values are not very different fromthose reported by Sedberry et al. (1999) where most8–12-year-old wreckfish harvested in the U.S.A. aremature. Wreckfish maximum values of monthly meanGSI were high for both males and females. Data tab-ulated for eight reef fish species (Sadovy 1996) showthat only Acanthopagrus cuvieri females have monthlyGSI larger than wreckfish (>20).
Wreckfish aggregate to spawn in deep waters(>300 m) off southern Brazil, especially north of RioGrande, though it is possible that some fish spawn inpairs or small groups along the entire continental slope.Large-sized testes (male GSI up to 14) are an addi-tional indication that the species is a group spawner

172
(Sadovy 1996). There is evidence that adult fish inhab-iting the southern fishing area perform a northwardspawning migration but there is no data of how long thismight be (Peres 2000). A northward migration of about700 km before spawning, in late winter, was observedfor New Zealand wreckfish (Roberts 1996, Roberts1).Beentjes & Francis (1999), from tag and recapture data,reported a northward seasonal migration up to 1 389 kmfor New Zealand hapuku P. oxygeneios. The straightline distance between the extreme limits of our studyarea is about 480 nautical miles (890 km) and the dis-tance between areas of higher fishing yields (Valaoand Tramandaı-Solidao) is about 300 nautical miles(550 km). It would be reasonable to assume a seasonalmigration, back and forth, of up to 1 000 or 1 500 km.
The North Atlantic wreckfish has a complex life his-tory. The adult demersal phase spawns in deep watersof North American coast (Blake Plateau, 32◦N, 79◦W).The extended pelagic juvenile interval (up to 50 cmTL and 2 years) drifts in the North Atlantic gyre anddescends to the bottom in the eastern North Atlantic(Sedberry et al. 1999). Because oceans are diffusiveand advective environments, a specific oceanographicpattern of spawning location that gives a chance oflife-cycle closure is an important requirement forself-sustaining populations with complex life histo-ries (Sinclair 1988, Sinclair & Iles 1989). The NorthAtlantic wreckfish spawning ground is under the influ-ence of the Gulf Stream, a warm western boundary cur-rent of the North Atlantic Subtropical Gyre (NASG).This would be the oceanographic feature that retainsand passively transports wreckfish early life stages.The time duration of this pelagic interval agrees withthis. As all life stages occur within the NASG, theNorth Atlantic wreckfish is considered a self-sustainedpopulation (sensu Sinclair 1988).
The southern Brazil continental slope is domi-nated by the southward motion of the Brazil current,a warm (>20◦C) western boundary current of theSouth Atlantic Subtropical Gyre (SASG) (Castro &Miranda 1998). While weaker than the Gulf Streamin terms of mass transport, the Brazil current isenergetically comparable to its North Atlantic coun-terpart, particularly in the southern part of the sub-tropical gyre (Campos et al. 1995). The southwesternAtlantic wreckfish is also a self-sustained populationof P. americanus. Although we did not observe pelagic
1 Roberts, C. 2000. Museum of NZ Te Papa, 169 Tory Street,PO Box 467. Wellington, New Zealand. pers. comm.
juvenile stages, the wreckfish recruits to the bottomin shallower waters of the continental shelf, fromsouthern Argentina (35–45 cm TL) (Nakamura 1986)to southern Brazil (44 cm TL, 1.5 years old) (Peres2000). Large, mature fish have not been described forArgentine or Uruguayan waters (Cousseau & Perrota1998, Peres 2000) but they spawn in the continentalslope of southern Brazil. Spawning in warm watersenables wreckfish egg and larvae to develop, even dur-ing winter–spring months, while they are passivelytransported southwards.
The SASG does not seem to be the oceanographicfeature responsible for life-cycle closure of southwest-ern Atlantic wreckfish, because of its larger extent,compared with the NASG. The absence of a well-develop fishery in the eastern Atlantic coast (Sedberryet al. 1999) agrees with this. The northward recircula-tion cell of the Brazil current front (Campos et al. 1995)could be the physical feature responsible for retentionof larvae and juveniles in the region. Vagrancy (sensuSinclair 1988) through the south Atlantic current couldexplain the occurrence of wreckfish in the southeast-ern Atlantic and maybe, in all other places of the worldwithout self-sustained populations.
All wreckfish life stages occur around southeast-ern Australia and New Zealand. With its congenericspecies P. oxygeneios (‘hapuku’), wreckfish (‘bass’)sustain an important recreational and commercial fish-ery (Paul 1995, Roberts 1996) indicating that this is aself-sustaining population of P. americanus too. Alikesouthern Brazil (30◦–31◦S) and southeastern U.S.A.(32◦N), the southeastern Australia continental marginis under the influence of a western boundary system,the east Australia current.
We argue that western boundary current systemsare a basic requirement for self-sustaining wreck-fish populations. This retention mechanism diminisheslosses due to vagrancy. Homing of adults to spe-cific spawning sites generates temporal persistenceof larval distribution in particular geographic spaceand mate assemblage (Sinclair & Iles 1989). Thus,migration of wreckfish spawners to these systems isthe weak link in completing their life cycle. Longmigrations have been observed for all self-sustainingpopulations of wreckfish in the southwestern Atlantic,north Atlantic (Sedberry et al. 1999) and southwest-ern Pacific (Roberts1). If this is true, southwesternPacific wreckfish spawning grounds should be some-where between New Zealand and Australia (30◦S), overcontinental or islands slopes north of the Tasman front.

173
Southwestern Atlantic wreckfish reproductive invest-ment is remarkably high. Besides the presence of along spawning migration and high fecundity, wreckfisheggs are quite large (OD 2 to 2.2 mm) when comparedwith pelagic eggs of other teleosts (Hardy 1978). Sinceestimated natural mortality for the demersal–exploitedinterval (ages 6–9 years) is low, around 0.05–0.09(Peres 2000), losses due to vagrancy during pelagicstages would explain this reproductive investment.
Acknowledgements
Part of these results was submitted in a Ph.D. thesisof the Biological Oceanography Post-Graduation, Uni-versity of Rio Grande (FURG) – Brazil, and wassupported by grants from the Brazilian Ministry of Edu-cation (CAPES). Thanks to southern Brazilian skippersand fishers, specially Antonio Bianchi, for access totheir wreckfish catches, whole gonads and for pro-viding catch and effort data for many years; to LucioWaengertner Pires and George Kotas for field assis-tance; to Maria Paula Mellito da Silveira and JoaoCarlos Cousin for histological assistance; to ManuelHaimovici for whole wreckfish gonads from researchcruises. Many thanks to David Wyanski, Clive Robertsand Dave Japp for unpublished information on wreck-fish from U.S.A., New Zealand and South Africa.Thanks to Beatrice Padovanni Ferreira and CarolusMaria Vooren for critical reviews that greatly improvedearlier versions of this manuscript and George Sedberryfor many fruitful discussions.
References
Beentjes, M. & M. Francis. 1999. Movement of hapuku(Polyprion oxygeneios) determined from tagging studies. N. Z.J. Mar. Freshw. Res. 33: 1–12.
Campos, E., J. Miller, T. Muller & R. Peterson. 1995. Physicaloceanography of the southwest Atlantic Ocean. Oceanography8: 87–91.
Castro, B.M. & L.B. Miranda. 1998. Physical oceanography ofthe Western Atlantic continental shelf located between 4N and34S. Coastal segment (4,W). pp. 209–251. In: A.R. Brink &K.H. Robinson (eds) The Sea, Vol. 11, John Wiley & Sons,New York.
Cousseau, M. & R. Perrota. 1998. Peces marinos de Argentina:Biologıa, distribucion, INIDEP, Mar del Plata, Argentina.000 pp.
Gunderson, D. 1993. Surveys of Fisheries Resources, JohnWiley & Sons, New York. 248 pp.
Hardy, J. 1978. Development of fishes of the Mid-Atlantic Bight.An atlas of egg, larval and juvenile stages, Vol. III. Fish andWildlife Service, U.S. Department of the Interior.
Heemstra, P. 1986. Family No. 165; Polyprionidae. In:M. Smith & P. Heemstra (ed.) Smith’s Sea Fishes, 6th edn,Springer-Verlag, Berlin.
Hunter, J. & B. Macewicz. 1985. Measurement of spawning fre-quency in multiple spawning fishes. pp. 79–94. In: R. Lasker(ed.) An Egg Production Method for Estimating Biomass ofPelagic Fish: Application to the Northern anchovy Engraulismordax. NOAA Technical Report 36, U.S. NMFS.
Hunter, J., B. Macewicz, N. Chyan-Huel Lo & C. Kimbrel.1992. Fecundity, spawning, and maturity of female Dover soleMicrostomus pacificus, with an evaluation of assumptions andprecision. Fishery Bulletin U.S. 90: 101–128.
Ihaka, R. & R. Gentleman. 1996. A language for data analysisand graphics. J. Comput. Graph. Stat. 5: 299–314.
Nakamura, I. 1986. Important Fishes Trawled off Patagonia,Japan Marine Fishery Resource Research Center, Tokyo.369 pp.
Paul, L. 1995. Groper. Seafood New Zealand 3: 26–31.Peres, M.B. 2000. Dinamica populacional e pesca do cherne-
poveiro Polyprion americanus (Bloch e Schneider, 1801)(Teleostei: Polyprionidae) no sul do Brasil. Ph.D. thesis,Fundacao Universidade Federal do Rio Grande, Rio Grande,Brazil. 151 pp.
Roberts, C. 1989. Reproductive mode in the percomorph fishgenus Polyprion Oken. J. Fish Biol. 34: 1–9.
Roberts, C. 1996. Hapuku and bass: The mystery of the missingjuveniles. Seafood New Zealand 4: 17–21.
Sadovy, Y. 1996. Reproduction of reef fishery species. pp. 15–59.In: N. Polunin & C. Roberts (ed.) Reef Fisheries, Chapman &Hall, London.
Sedberry, G., C. Andrade, J. Carlin, R. Chapman, B. Luckhurst,C. Manooch III, G. Menezes, B. Thomsen & G. Ulrich. 1999.Wreckfish Polyprion americanus in the North Atlantic: Fish-eries, biology and management of a widely distributed andlong-lived fish. Am Fish. Soc. Symp. 23: 27–50.
Shapiro, D. 1987. Reproduction in groupers. pp. 295–328. In:J. Polovina & S. Ralston (ed.) Tropical Snappers and Groupers:Biology and Fisheries, Westview Press, London.
Sinclair, M. 1988. Marine populations: An essay on populationregulation and speciation. Washington Sea Grant/University ofWashington Press, Seattle.
Sinclair, M. & T.D. Iles. 1989. Population regulation andspeciation in the oceans. J. Cons. int. Explor. Mer 45:165–175.




















