
Pattern of FGF2 isoform expression correlated with its biological action in experimental...
11
Acta Neuropathol (2006) 112:491–501 DOI 10.1007/s00401-006-0101-9 123 ORIGINAL PAPER Pattern of FGF-2 isoform expression correlated with its biological action in experimental prolactinomas Jorge H. Mukdsi · Ana L. De Paul · Juan P. Petiti · Silvina Gutiérrez · Agustín Aoki · Alicia I. Torres Received: 26 April 2006 / Revised: 8 June 2006 / Accepted: 8 June 2006 / Published online: 6 July 2006 © Springer-Verlag 2006 Abstract Fibroblast growth factor-2 (FGF-2) synthe- sized in the pituitary is involved in the formation and progression of pituitary tumors. The aim of this study was to analyze the pattern expression of two FGF-2 isoforms at diVerent subcellular levels and to deter- mine its correlation with prolactinoma development. Estrogen administration to male rats for 7, 20, and 60 days generated pituitary tumors, with lactotrophs being the prevalent cell type. Ultrastructural immu- nolabeling showed FGF-2 in the cytosolic and nuclear compartments of somatotrophs, lactotrophs and gonadotrophs, as well as in folliculo-stellate cells of normal rats. Estrogen stimulation increased FGF-2 immunoreactivity in various tumors and enhanced the expression of two FGF-2 isoforms, 18 and 22 kDa, as quantiWed by western blot. The 18 kDa isoform observed in cytosol extracts reached the highest levels after 60 days of hormonal stimulation and this was related to lactotroph proliferation. However, the 22 kDa FGF-2 isoform was only detected in the nuclear compartment and achieved the maximum expression at 7 days of estrogen treatment, without any correlation with lactotroph proliferation. These results suggest that the 18 kDa FGF-2 may play a role in the modulation of lactotroph proliferation in prolactino- mas induced by estrogen. The overproduction of both FGF-2 isoforms appears to be implicated in auto- crine–paracrine–intracrine mitogenic loops; this FGF-2 activity could lead to uncontrolled cell growth, angio- genesis, and tumor formation. Keywords FGF-2 isoforms · Pituitary cells · Prolactinoma · Estrogen · Immunoelectron microscopy Introduction The pituitary gland is a crucial organ for the mainte- nance of several homeostatic functions. The expression of numerous growth factors and cytokines as well as their receptors within the anterior pituitary suggests that these factors could be involved locally in the con- trol of pituitary development, hormone secretion, and cell proliferation. Tumor growth is dependent on angiogenesis; vascular endothelial growth factor and Wbroblastic growth factor-2 (FGF-2) are potent angio- genic factors which act synergistically in vivo and in vitro [30, 38]. FGF-2, which belongs to the FGF family, is found in several molecular isoforms that are translated from a common mRNA by the use of alternative initiation sites [14]. FGF-2 is involved in diVerent processes such as proliferation, migration and cell diVerentiation, besides its role as an angiogenic factor [7]. FGF-2 isoforms have molecular masses of 18, 22, 22.5, 24, and 34 kDa in humans and 18, 21, and 22 kDa FGF2 in mice. The lower molecular weight (LMW) 18-kDa polypeptide is localized primarily in the cytosol before being secreted. Once on the cell surface, it can bind to heparan sulphate proteoglycans (HSPG) and tyrosine kinase receptors (FGFRs) of low and high aYnity [12]. J. H. Mukdsi · A. L. De Paul · J. P. Petiti · S. Gutiérrez · A. Aoki · A. I. Torres (&) Centro de Microscopía Electrónica, Facultad de Ciencias Médicas, Universidad Nacional de Córdoba, Haya de la Torre, esquina Enrique Barros, Pabellón de Biología Celular, Ciudad Universitaria, X5000, HRA, Córdoba, Argentina e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pattern of FGF2 isoform expression correlated with its biological action in experimental...

Acta Neuropathol (2006) 112:491–501
DOI 10.1007/s00401-006-0101-9ORIGINAL PAPER
Pattern of FGF-2 isoform expression correlated with its biological action in experimental prolactinomas
Jorge H. Mukdsi · Ana L. De Paul · Juan P. Petiti · Silvina Gutiérrez · Agustín Aoki · Alicia I. Torres
Received: 26 April 2006 / Revised: 8 June 2006 / Accepted: 8 June 2006 / Published online: 6 July 2006© Springer-Verlag 2006
Abstract Fibroblast growth factor-2 (FGF-2) synthe-sized in the pituitary is involved in the formation andprogression of pituitary tumors. The aim of this studywas to analyze the pattern expression of two FGF-2isoforms at diVerent subcellular levels and to deter-mine its correlation with prolactinoma development.Estrogen administration to male rats for 7, 20, and60 days generated pituitary tumors, with lactotrophsbeing the prevalent cell type. Ultrastructural immu-nolabeling showed FGF-2 in the cytosolic and nuclearcompartments of somatotrophs, lactotrophs andgonadotrophs, as well as in folliculo-stellate cells ofnormal rats. Estrogen stimulation increased FGF-2immunoreactivity in various tumors and enhanced theexpression of two FGF-2 isoforms, 18 and 22 kDa, asquantiWed by western blot. The 18 kDa isoformobserved in cytosol extracts reached the highest levelsafter 60 days of hormonal stimulation and this wasrelated to lactotroph proliferation. However, the22 kDa FGF-2 isoform was only detected in thenuclear compartment and achieved the maximumexpression at 7 days of estrogen treatment, without anycorrelation with lactotroph proliferation. These resultssuggest that the 18 kDa FGF-2 may play a role in themodulation of lactotroph proliferation in prolactino-mas induced by estrogen. The overproduction of
both FGF-2 isoforms appears to be implicated in auto-crine–paracrine–intracrine mitogenic loops; this FGF-2activity could lead to uncontrolled cell growth, angio-genesis, and tumor formation.
Keywords FGF-2 isoforms · Pituitary cells · Prolactinoma · Estrogen · Immunoelectron microscopy
Introduction
The pituitary gland is a crucial organ for the mainte-nance of several homeostatic functions. The expressionof numerous growth factors and cytokines as well astheir receptors within the anterior pituitary suggeststhat these factors could be involved locally in the con-trol of pituitary development, hormone secretion, andcell proliferation. Tumor growth is dependent onangiogenesis; vascular endothelial growth factor andWbroblastic growth factor-2 (FGF-2) are potent angio-genic factors which act synergistically in vivo and invitro [30, 38].
FGF-2, which belongs to the FGF family, is found inseveral molecular isoforms that are translated from acommon mRNA by the use of alternative initiationsites [14]. FGF-2 is involved in diVerent processes suchas proliferation, migration and cell diVerentiation,besides its role as an angiogenic factor [7]. FGF-2isoforms have molecular masses of 18, 22, 22.5, 24, and34 kDa in humans and 18, 21, and 22 kDa FGF2 inmice. The lower molecular weight (LMW) 18-kDapolypeptide is localized primarily in the cytosol beforebeing secreted. Once on the cell surface, it can bind toheparan sulphate proteoglycans (HSPG) and tyrosinekinase receptors (FGFRs) of low and high aYnity [12].
J. H. Mukdsi · A. L. De Paul · J. P. Petiti · S. Gutiérrez · A. Aoki · A. I. Torres (&)Centro de Microscopía Electrónica, Facultad de Ciencias Médicas, Universidad Nacional de Córdoba, Haya de la Torre, esquina Enrique Barros, Pabellón de Biología Celular, Ciudad Universitaria, X5000, HRA, Córdoba, Argentinae-mail: [email protected]
123

492 Acta Neuropathol (2006) 112:491–501
Bound FGF-2 is translocated into the nucleus [24, 43].Although 18 kDa FGF-2 is released in the extracellularmatrix, it lacks a conventional peptide secretory signal.Instead, it is secreted by an alternative pathway whichis independent of the endoplasmic reticulum–Golgicomplex [28]. The higher molecular weight isoforms(HMW: 21, 22, 22.5, 24, and 34 kDa) are found pre-dominantly in the nucleus associated mainly with thenucleolus [21]. The NH2 terminal extension of HMWforms contains a nuclear localization signal that medi-ates their preferential nuclear accumulation [8]. TheLMW and HMW FGF-2 isoforms with diVerent subcel-lular distributions appear to aVect the cell phenotypethrough an autocrine FGFR-dependent, or an intra-crine FGFR-independent pathway [12, 32, 36]. Arecent investigation has suggested that the FGF-2nuclear form may be functionally distinct from the18 kDa cytosolic form in NIH 3T3 Wbroblast cells [35].
FGF-2 is expressed in pituitary folliculo-stellate cells(FSC), which appear to be the major source of thisangiogenic factor [2]. However, other immunocyto-chemical studies at light microscopic level have sug-gested that FGF-2 immunoreactive cells can also besomatotrophs [26], gonadotrophs [39], and cortico-trophs [6].
In normal pituitary glands, FGF-2 regulates thesecretion of PRL, GH, and TSH through paracrinemechanisms and could play a key role initiating lacto-troph diVerentiation and maintaining PRL geneexpression [13, 22]. The participation of FGF-2 in thedevelopment of pituitary adenomas could have a dualrole, acting directly on lactotroph proliferation [10]and as an angiogenic factor [42]. Also, in human pitui-tary adenomas, this cytokine plays an important role inthe prognosis of these tumors as a predictor for recur-rences [15]. However, all these reports have not speci-Wed which of the FGF-2 isoforms are involved in thecell proliferation process responsible for prolactinomadevelopment. In this paper, we have investigated thechanges in the expression proWle of FGF-2 isoformscorrelated with the proliferation rate of lactotroph cellsin experimental prolactinomas. Furthermore, the ultra-structural immunolocalization of FGF-2 could contrib-ute to a better understanding of the subcellular levelswhere this angiogenic factor has its eVects.
Materials and methods
Animals
Male Wistar rats, 3 months old, were maintained undercontrolled temperature (21 § 3°C) and lighting conditions
(14 h light/10 h dark). Intact animals were fed a stan-dard diet. The following experimental groups weredesignated: control rats (C), and rats treated withestradiol benzoate (EB) for 7 (EB-7), 20 (EB-20), and60 (EB-60) days. The number of rats used in eachexperimental group is given in the diVerent methodsapplied, and the protocols were repeated three times inorder to estimate the average for each group. EB(Sigma Chemical Co., St. Louis, MO, USA) wasimplanted subcutaneously in slow-release capsulesmade of silastic tubing (Dow Corning, Medical grade,Midland, MI, USA) Wlled with 10 mg of estrogen crys-tals; the ends were sealed with silastic cement.
The rats were decapitated within 10 s of removalfrom their cage, avoiding any stress or external stimuli.Arterial and venous blood drained from the head andtrunk was collected, allowed to clot, and the serumstored frozen at ¡20°C for subsequent radioimmuno-assay to determine PRL serum levels. Anterior pitui-tary glands were rapidly removed and processed fordiVerent studies. Animal treatment was in compliancewith the Guidelines on the Handling and Training ofLaboratory Animals published by the UniversitiesFederation for Animal Welfare, and the local AnimalCare Committee.
Immunocytochemistry and morphometry
Whole anterior pituitary glands from three rats perexperimental group were Wxed in 4% buVered formal-dehyde, dehydrated and embedded in paraYn.Sections were cut at 3 �m thickness, deparaVinized,rehydrated, and washed in PBS. Before applying theprimary antibody, sections were treated with 3% H2O2for 15 min to inhibit endogenous peroxidase activityfollowed by 1% goat serum for 30 min to block non-speciWc binding of the secondary antibody. Thereafter,pituitary sections were incubated overnight in anti-PRL antiserum (NIH Hormone Program, Bethesda,MD, USA) at 1:1,000 dilution. Slides were then washedwith PBS, exposed to biotin-labeled antibody againstrabbit IgG at room temperature for 30 min, then incu-bated with ABC complex (Vector Labs, Burlingame,CA, USA) for 30 min. The slides were then immersedin a solution containing 3,3�-diaminobenzidine (Sigma)in 0.1 M Tris buVer, pH 7.2, containing 0.03% H2O2 for7 min, counterstained with hematoxylin, dehydratedand coverslipped with Entellan (Merck, Darmstadt,Germany).
For morphometric assessment of the lactotroph pop-ulation, three whole anterior pituitary glands per exper-imental group were embedded in paraYn, and serialsections were cut from the upper, middle, and lower
123

Acta Neuropathol (2006) 112:491–501 493
levels of each pituitary gland. One large section of eachzone was mounted onto glass slides and immunostainedfor PRL. Eight micrographs were taken at random fromeach section at a magniWcation of 400£. The total areaof pituitary scanned was 24 £ 105 um2/gland.
The average number of PRL-immunoreactive cellsfrom a total of 3,000 cells, taken from the sections cor-responding to the three levels of each gland, wasobtained using the ImageJ software (version 1.34s).
Reticulin staining
To provide histological evidence that these animalsdeveloped pituitary tumors and to distinguish hyper-plasia from adenoma, the reticulin network was ana-lyzed in three anterior pituitary glands correspondingto each experimental group, using the Gordon-Sweetreticulin stain.
Prolactin radioimmunoassay
Serum prolactin levels were measured by RIA, byapplying a double antibody technique performed attwo dose levels and the results were expressed in termsof rat prolactin-RP-3 (biological potency equivalent to30 IU/mg). All samples were processed simultaneouslyto avoid interassay variations. Reagents for the RIAused (r-PRL-I-6, anti-rPRL-S-9, and r-PRL-RP-3)were provided by NIDDKD. The intra-assay coeY-cient of variation was lower than 10%.
For statistical purposes, the serum from eight malerats, per experimental group out of a total of threeindependent experiments, was analyzed by RIA.
Electron microscopy and immunocytochemistry
Three anterior pituitary glands from each experimentalgroup were divided in two halves. One half was pro-cessed for ultrastructural studies and the other wasused for immunocytochemistry.
For ultrastructural studies of PRL cells, the hemipi-tuitaries were Wxed by immersion in 4% (v/v) glutaral-dehyde, 4% (w/v) formaldehyde in 0.1 M cacodylatebuVer, then treated with 1% osmium tetroxide, dehy-drated and embedded in Araldite. Thin sections werecut with a diamond knife on a JEOL JUM-7 ultrami-crotome (Nikon, Tokio, Japan) and examined using aZeiss Leo 906-E electron microscope (Oberkochen,Germany).
For immunocytochemistry, the hemipituitaries wereWxed in 2% (v/v) glutaraldehyde and 4% (w/v) formal-dehyde in 0.1 M cacodylate buVer pH 7.3, at roomtemperature, for 5–6 h. The hemipituitaries were in a
series of increasing concentrations of ethanol, andembedded in LR White (London Resin Corporation,Berkshire, UK). Rabbit anti-rat FGF-2 (10 ug/ml)(FGF-2 (147): sc-79, Santa Cruz Biotechnology, SantaCruz, CA, USA) was used as the primary antiserum.To identify endocrine cell types, serial thin sectionswere incubated in parallel with antisera raised againstrat prolactin (1:3,000), growth hormone (1:4,000), andluteotrophic � hormone (1:1,000) (provided by theNIH Hormone Program).
The immunoreactive sites were labeled with proteinA–colloidal gold complex. Preparations of the immu-nogold complexes and other details of immunocyto-chemical procedures are described elsewhere [25].
Ultrastructural immunocytochemical controls
To validate the speciWcity of the immunostaining, thefollowing controls were chosen with both absorptionand negative controls being performed. For negativecontrols, the primary antibody was replaced with either1% bovine serum albumin (BSA) in 0.1 M phosphatebuVer or diluted preimmune serum followed by proteinA/colloidal gold complex (Sigma Chemical Co.). Anabsorption (peptide neutralization) control was per-formed to determine the speciWcity of the primary anti-body. Polyclonal rabbit anti-FGF2 antibody wascombined with a Wvefold excess of blocking antigenpeptide [FGF-2 (147-P, Cart. # sc-79 P, Lot # I3002)Santa Cruz Biotechnology] and incubated overnight at4°C [23]. Sections of the pituitary gland were incubatedwith pre-absorbed antibody in parallel with antiseraraised against FGF-2.
Subcellular fractionation
The nuclear and cytoplasmic extracts from male ratpituitary glands were prepared according to themethod of Sugawara et al. [40] with modiWcations [29].BrieXy, Wve anterior pituitary glands per experimentalgroup (out of a total of three independent experi-ments) were minced and homogenized with a teXon-glass Potter-Elvehjem tissue grinder in 2 ml 0.3 Msucrose, 1 mM EDTA, 1 mM PMSF, and 10 mMHepes/KOH (pH 7.5) at 4°C (buVer I). The homoge-nate was centrifuged at 1,000g for 10 min and thesupernatant frozen in aliquots and stored at ¡40°Cuntil required for quantiWcations. The pellet was resus-pended in 2 ml solution of 0.3 M sucrose, 1 mM CaCl2,10 mM Tris–HCl (pH 7.5), 1 mM DTT, and 0.1 mMPMSF (buVer II). Then 75 �l of 10% Igepal CA-630was added, and the solution was incubated on ice for10 min and centrifuged at 250g for 5 min. The pellet
123
494 Acta Neuropathol (2006) 112:491–501
was washed once with buVer II before being resus-pended in 2 vol 20 mM HEPES (pH 7.8), 0.6 M KCl,0.02 mM ZnCl2, 0.2 mM EGTA, 0.5 mM DTT, and0.1 mM PMSF (buVer III) and then incubated on icefor 30 min. The suspension was centrifuged at 20,000gat 4°C for 30 min, and the supernatant containing thehigh salt extracts of nuclei was dialyzed against 20 mMHEPES (pH 7.8), 5 mM 2-mercaptoethanol, 50 mMNaCl, 2 mM EGTA, 10% (vol/vol) glycerol, and0.1 mM PMSF, and then centrifuged at 10,000g for15 min. Aliquots were stored frozen at ¡40°C untilused in protein and western blot assays.
All the reagents used in this protocol were pur-chased from Sigma Chemical Company.
Western blot analysis
The levels of FGF-2 isoform were determined by west-ern blot analysis. Protein concentration was determinedusing a Bio-Rad kit (Bio-Rad Protein Assay, Bio-RadLaboratories, Hercules, CA, USA). Proteins (20 �g/lane) from subcellular fractions were separated by SDS-polyacrylamide gel (12%) under reducing conditionsand then transferred to a nitrocellulose membrane(Hybond, Amersham-Life Science, Bucks, UK). Toassess the corresponding molecular weights, a pre-stained molecular weight marker was used (BenchMarkPre-stained Protein Ladder, Invitrogen, Buenos Aires,Argentina). Membranes were incubated in blockingbuVer containing 5% nonfat dried milk and 0.1% Tween20 for 1 h at room temperature, and then probed with apolyclonal antibody that recognizes FGF-2 (1:700, SantaCruz Biotechnology) for 2 h. After washing in PBS and0.1% Tween 20, the blots were incubated for 1 h with aperoxidase-conjugated (HRP) goat anti-rabbit second-ary antibody diluted 1:5,000 (Jackson ImmunoresearchLabs Inc., West Grove, PA, USA), and then the signalswere visualized using an Enhanced ChemiLuminescencedetection system (ECL, Amershan Biosciences, Bucks,UK), with emitted light being captured on HyperWlm(Amersham-Pharmacia-Biotech, Bucks, UK). Signalswere scanned and quantiWed using Scion Image software(V. beta 4.0.2, Scion Image Corp., Frederick, MD, USA)at three diVerent exposure times.
The expression of �-actin (monoclonal anti �-actin;Sigma Chemical Co. product A 5441, St. Louis, MO,USA, at 1:5,000 dilution) was used as an internal con-trol to conWrm the equivalent total protein loading.
Statistical analysis
Values were expressed as the mean § SEM, eachexperiment was performed in triplicate on diVerent
experimental models. The data obtained were ana-lyzed by one-way ANOVA, followed by the Tukey testfor the number of PRL-immunoreactive cells, serumPRL level, FGF-2 18 kDa, and FGF-2 22 kDa. For �-actinexpression a Fisher post test was applied. A signiW-cance level of P < 0.05 was considered signiWcant for alltests.
Linear correlation analysis was carried out by calcu-lating Pearson’s coeYcient in order to assess the rela-tionship among the following parameters: FGF-218 kDa, FGF-2 22 kDa, and lactotroph cell proliferation.
Results
Morphological analysis of pituitary gland. Morphometry of PRL cell population
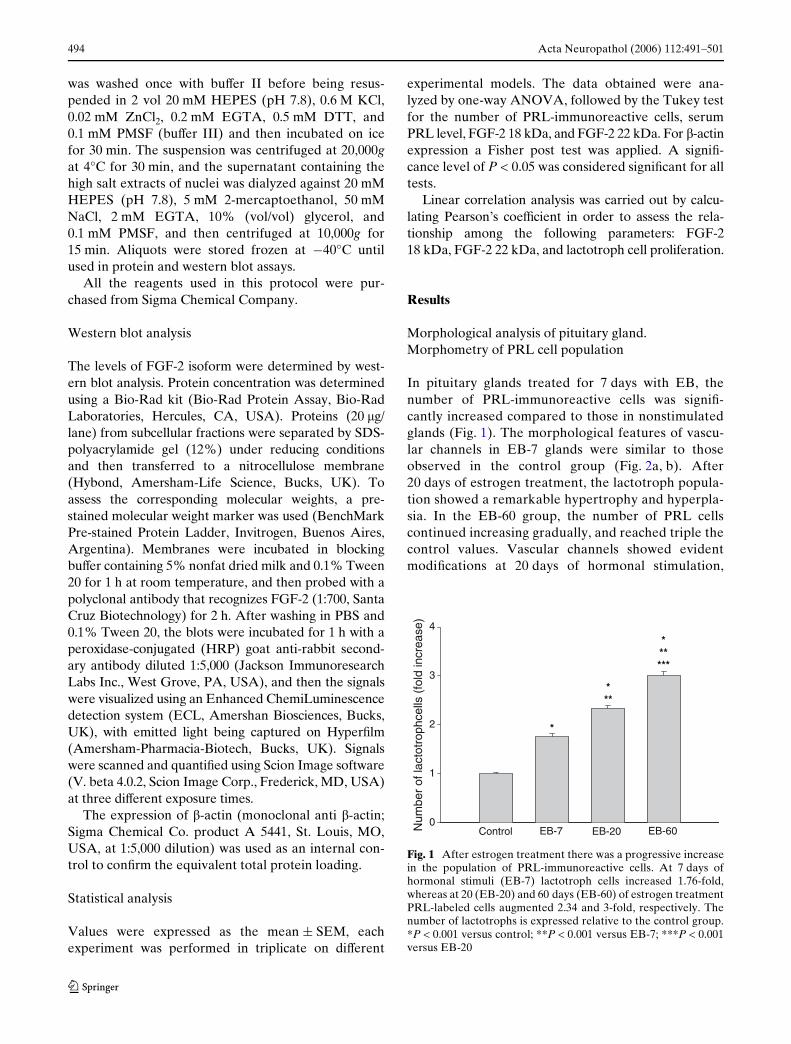
In pituitary glands treated for 7 days with EB, thenumber of PRL-immunoreactive cells was signiW-cantly increased compared to those in nonstimulatedglands (Fig. 1). The morphological features of vascu-lar channels in EB-7 glands were similar to thoseobserved in the control group (Fig. 2a, b). After20 days of estrogen treatment, the lactotroph popula-tion showed a remarkable hypertrophy and hyperpla-sia. In the EB-60 group, the number of PRL cellscontinued increasing gradually, and reached triple thecontrol values. Vascular channels showed evidentmodiWcations at 20 days of hormonal stimulation,
Fig. 1 After estrogen treatment there was a progressive increasein the population of PRL-immunoreactive cells. At 7 days ofhormonal stimuli (EB-7) lactotroph cells increased 1.76-fold,whereas at 20 (EB-20) and 60 days (EB-60) of estrogen treatmentPRL-labeled cells augmented 2.34 and 3-fold, respectively. Thenumber of lactotrophs is expressed relative to the control group.*P < 0.001 versus control; **P < 0.001 versus EB-7; ***P < 0.001versus EB-20
Num
ber
of la
ctot
roph
cells
(fo
ld in
crea
se)
0
1
2
3
4
Control EB-7 EB-20 EB-60
*
***
******
123
Acta Neuropathol (2006) 112:491–501 495
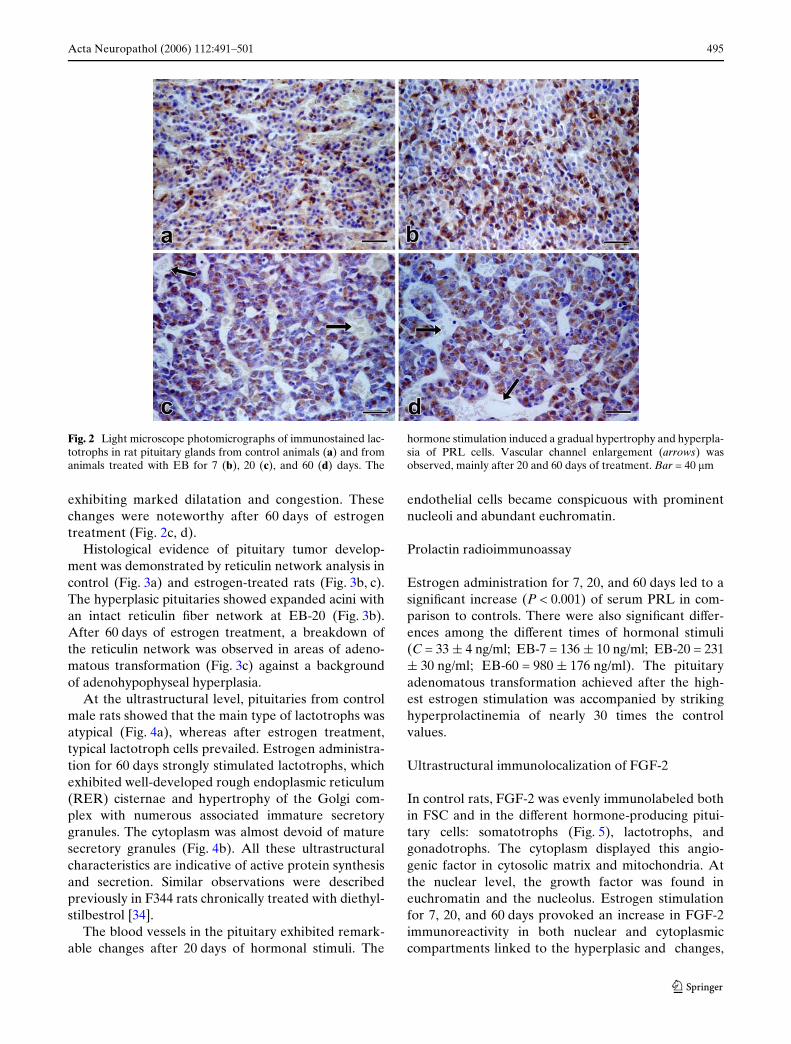
exhibiting marked dilatation and congestion. Thesechanges were noteworthy after 60 days of estrogentreatment (Fig. 2c, d).
Histological evidence of pituitary tumor develop-ment was demonstrated by reticulin network analysis incontrol (Fig. 3a) and estrogen-treated rats (Fig. 3b, c).The hyperplasic pituitaries showed expanded acini withan intact reticulin Wber network at EB-20 (Fig. 3b).After 60 days of estrogen treatment, a breakdown ofthe reticulin network was observed in areas of adeno-matous transformation (Fig. 3c) against a backgroundof adenohypophyseal hyperplasia.
At the ultrastructural level, pituitaries from controlmale rats showed that the main type of lactotrophs wasatypical (Fig. 4a), whereas after estrogen treatment,typical lactotroph cells prevailed. Estrogen administra-tion for 60 days strongly stimulated lactotrophs, whichexhibited well-developed rough endoplasmic reticulum(RER) cisternae and hypertrophy of the Golgi com-plex with numerous associated immature secretorygranules. The cytoplasm was almost devoid of maturesecretory granules (Fig. 4b). All these ultrastructuralcharacteristics are indicative of active protein synthesisand secretion. Similar observations were describedpreviously in F344 rats chronically treated with diethyl-stilbestrol [34].
The blood vessels in the pituitary exhibited remark-able changes after 20 days of hormonal stimuli. The
endothelial cells became conspicuous with prominentnucleoli and abundant euchromatin.
Prolactin radioimmunoassay
Estrogen administration for 7, 20, and 60 days led to asigniWcant increase (P < 0.001) of serum PRL in com-parison to controls. There were also signiWcant diVer-ences among the diVerent times of hormonal stimuli(C = 33 § 4 ng/ml; EB-7 = 136 § 10 ng/ml; EB-20 = 231§ 30 ng/ml; EB-60 = 980 § 176 ng/ml). The pituitaryadenomatous transformation achieved after the high-est estrogen stimulation was accompanied by strikinghyperprolactinemia of nearly 30 times the controlvalues.
Ultrastructural immunolocalization of FGF-2
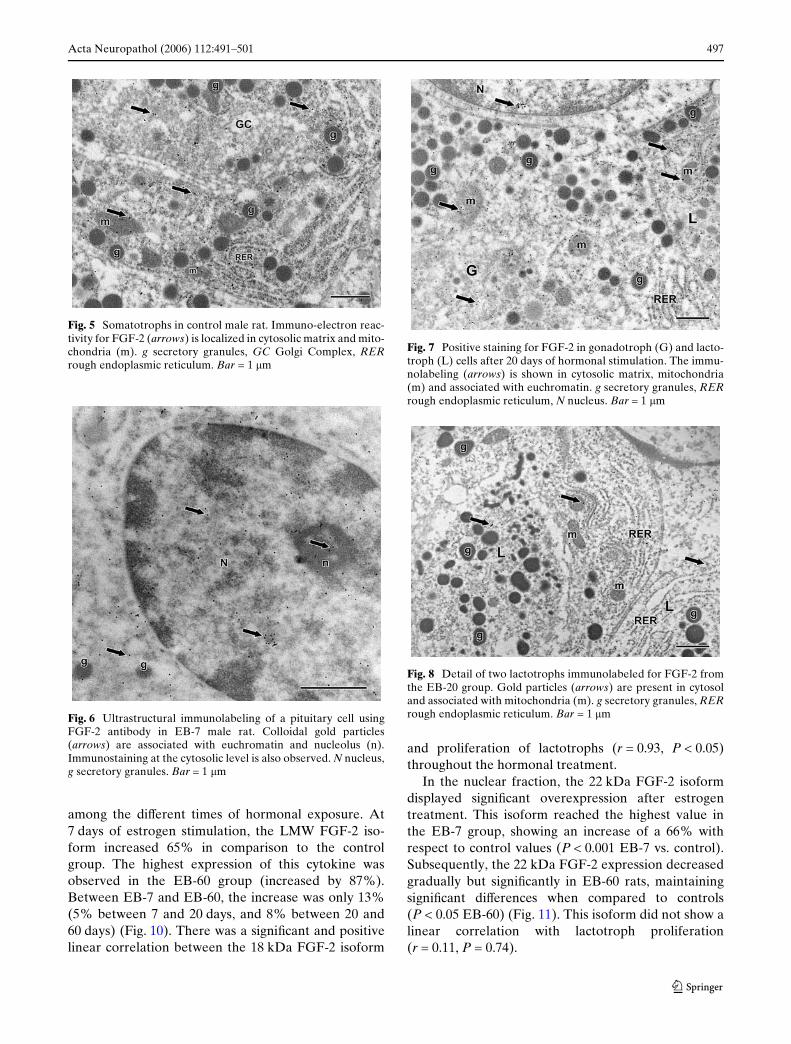
In control rats, FGF-2 was evenly immunolabeled bothin FSC and in the diVerent hormone-producing pitui-tary cells: somatotrophs (Fig. 5), lactotrophs, andgonadotrophs. The cytoplasm displayed this angio-genic factor in cytosolic matrix and mitochondria. Atthe nuclear level, the growth factor was found ineuchromatin and the nucleolus. Estrogen stimulationfor 7, 20, and 60 days provoked an increase in FGF-2immunoreactivity in both nuclear and cytoplasmiccompartments linked to the hyperplasic and changes,
Fig. 2 Light microscope photomicrographs of immunostained lac-totrophs in rat pituitary glands from control animals (a) and fromanimals treated with EB for 7 (b), 20 (c), and 60 (d) days. The
hormone stimulation induced a gradual hypertrophy and hyperpla-sia of PRL cells. Vascular channel enlargement (arrows) wasobserved, mainly after 20 and 60 days of treatment. Bar = 40 �m
123

496 Acta Neuropathol (2006) 112:491–501
where lactotroph cells became the predominant celltype. Figures 6, 7, and 8 illustrate immunolocalizationof FGF-2 in various pituitary cells at diVerent times ofestrogen stimulation.
All immunocytochemical controls validated thespeciWcity of the primary antiserum. The pre-absorp-tion of primary antiserum with its own antigen (Fig. 9)
or omission of the primary antibody resulted in nonoticeable immunolabeling.
QuantiWcation of FGF-2 18 and 22 kDa isoform by Western blotting
The antibody applied in the present work allowed thedetection of the 18 and 22 kDa FGF-2 isoforms, witheach FGF-2 isoform showing a diVerent subcellular dis-tribution. The 18-kDa isoform was only detected incytoplasmic extracts, whereas the 22 kDa wasexpressed in the nuclear fraction for all the experimen-tal models studied.
After EB treatment, the cytoplasmic levels of the18 kDa FGF-2 isoform exhibited signiWcant increases(P < 0.001) compared with the control group and also
Fig. 3 Reticulin pattern in the pituitary gland from control andestrogen treated rats. Untreated rats (a) show the normal reticu-lin network, where a Wber network is deposited around groups ofcells forming an acinar pattern. In EB-20 (b) hyperplasic transfor-mation showed expanded acini, whereas in EB-60 (c) an adeno-matous sector is formed and the reticulin network is completelylost in tumor areas. Sheets of cells are seen with broken fragmentsof the reticulin Wbers. Bar = 40 �m
Fig. 4 a An atypical lactotroph (L) from a control male rat has aquiescent appearance characterized by spherical secretory gran-ules (g), about 200–300 nm, immunolabeled for PRL. The RERand Golgi complex are poorly developed. A somatotroph cell inthe upper right (S) also presents unlabeled secretory granules (g)similar in size to those observed in the lactotroph, and serves as abackground control. Bar = 1 �m. b Electron micrograph of typi-cal lactotrophs from an EB-60 rat pituitary tumor. The cytoplasmof these cells acquired a rather uniform appearance due to RERWlling a large proportion of the cytoplasm. The Golgi complex(GC) also became hypertrophic with numerous immature secre-tory granules (ig). These cells show a small number of large ma-ture secretory granules (mg) (500–900 nm) characteristic oftypical lactotrophs. Bar = 1 �m
123
Acta Neuropathol (2006) 112:491–501 497
among the diVerent times of hormonal exposure. At7 days of estrogen stimulation, the LMW FGF-2 iso-form increased 65% in comparison to the controlgroup. The highest expression of this cytokine wasobserved in the EB-60 group (increased by 87%).Between EB-7 and EB-60, the increase was only 13%(5% between 7 and 20 days, and 8% between 20 and60 days) (Fig. 10). There was a signiWcant and positivelinear correlation between the 18 kDa FGF-2 isoform
and proliferation of lactotrophs (r = 0.93, P < 0.05)throughout the hormonal treatment.
In the nuclear fraction, the 22 kDa FGF-2 isoformdisplayed signiWcant overexpression after estrogentreatment. This isoform reached the highest value inthe EB-7 group, showing an increase of a 66% withrespect to control values (P < 0.001 EB-7 vs. control).Subsequently, the 22 kDa FGF-2 expression decreasedgradually but signiWcantly in EB-60 rats, maintainingsigniWcant diVerences when compared to controls(P < 0.05 EB-60) (Fig. 11). This isoform did not show alinear correlation with lactotroph proliferation(r = 0.11, P = 0.74).
Fig. 5 Somatotrophs in control male rat. Immuno-electron reac-tivity for FGF-2 (arrows) is localized in cytosolic matrix and mito-chondria (m). g secretory granules, GC Golgi Complex, RERrough endoplasmic reticulum. Bar = 1 �m
Fig. 6 Ultrastructural immunolabeling of a pituitary cell usingFGF-2 antibody in EB-7 male rat. Colloidal gold particles(arrows) are associated with euchromatin and nucleolus (n).Immunostaining at the cytosolic level is also observed. N nucleus,g secretory granules. Bar = 1 �m
Fig. 7 Positive staining for FGF-2 in gonadotroph (G) and lacto-troph (L) cells after 20 days of hormonal stimulation. The immu-nolabeling (arrows) is shown in cytosolic matrix, mitochondria(m) and associated with euchromatin. g secretory granules, RERrough endoplasmic reticulum, N nucleus. Bar = 1 �m
Fig. 8 Detail of two lactotrophs immunolabeled for FGF-2 fromthe EB-20 group. Gold particles (arrows) are present in cytosoland associated with mitochondria (m). g secretory granules, RERrough endoplasmic reticulum. Bar = 1 �m
123

498 Acta Neuropathol (2006) 112:491–501
Discussion
The functions of anterior pituitary cells are regulatedby hypothalamic factors, peripheral hormones, andgrowth factors. In the normal pituitary, intraglandularfactors probably play a role by modifying endocrinesignals in hormone-producing cells. However, sincealterations in the production of growth factors and theexpression of receptors have been reported in pituitaryadenomas, these factors may also contribute to thepathophysiology and development of pituitary tumors[38]. Among these growth factors, FGF-2 plays animportant role in the genesis and progression of tumorsin a variety of tissues [1, 16].
In our experimental model, sustained estrogenstimulation induced pituitary tumors in male rats.Light microscopy of estrogen-treated pituitary glandsrevealed striking hyperplastic and hypertrophicchanges in the lactotroph population. Furthermore,changes in the blood vessel architecture werereported, including a change of shape and increase indiameter of the capillaries concurrent with the func-tional alteration of lactotrophs. The intimate associa-tion of the PRL cells with capillaries is essential topromote eYcient and proper secretion of the hormone[19]. In previous investigations we showed that estro-genic stimulation induced a signiWcant increase inserum and pituitary PRL levels when compared to thecontrol group and among diVerent times of hormonalaction [29].
In the present study, we have found importantresults about the involvement of the FGF-2 angiogenic
factor in the development of prolactinomas. The estro-gen-mediated increase of LMW and HMW FGF-2 iso-forms occurred with diVerent expression patterns.There is also a close correlation between lactotrophproliferation and the expression of the 18 kDa FGF-2isoform. The LMW isoform reached the highest levelsin rats treated with estrogen for 60 days (EB-60), whilethe HMW isoform exhibited a swift response, attainingthe maximum expression in the Wrst week of estrogentreatment (EB-7 group).
The diVerential distribution of FGF-2 isoforms indiVerent subcellular compartments observed by west-ern blot is consistent with previous reports suggesting
Fig. 9 Section of the pituitary gland incubated with primary anti-body (FGF-2) pre-absorbed with its speciWc blocking antigen pep-tide. Note the lack of labeling in both, cytoplasm and nucleus of asomatotroph of a control male rat. g secretory granules, RER roughendoplasmic reticulum, m mitochondria, N nucleus. Bar = 1 �m
Fig. 10 Western blots showing the presence of 18 kDa FGF-2protein in cytoplasm extracts of pituitary gland from diVerentexperimental models. The relative intensities are determined bydensitometric analysis. The bands are stronger in the estrogen-treated rats than in the control group. The ratios of 18 kDa FGF-2 normalized with the intensity of �-actin band show a similar re-sponse: C = 0.627; EB-7 = 1.036; EB-20 = 1.088; EB-60 = 1.176.Statistical data are expressed as mean § SEM of Wve anteriorpituitary glands for each group of three independent experi-ments. *P < 0.001 EB-7, EB-20 and EB-60 versus control;**P < 0.001 EB-20 and EB-60 versus EB-7; ***P < 0.001 EB-60versus EB-20. ANOVA followed by Tukey’s test
Arb
itrar
y un
its
0
5000
10000
15000
20000
25000
C EB-7 EB-20 EB-60
*
***
******
FGF-2 18 kDa isoform expression in pituitary
β-actin
cytoplasmic extracts
123

Acta Neuropathol (2006) 112:491–501 499
that HMW and LMW FGF-2 isoforms might playdiVerent biological roles [11].
The presence of the LMW isoform only in the cyto-plasm is in keeping with some reports suggesting thatthe 18-kDa FGF-2 isoform lacks a nuclear localizationsequence [21]. The correlation analysis revealed anassociation between the levels of the 18 kDa isoformand lactotroph proliferation (r = 0.93). It is probablethat this isoform is secreted and activates FGFRs in itstarget cells through an autocrine or paracrine pathway.The induction of lactotroph proliferation through FGF-2 could be mediated by activation of Src kinase, Rasp21, and MAPK p44/42 [10]. It has also been demon-strated that FGF-2 is one of the factors that induces theexpression of c-fos, an oncoprotein also augmented inpituitary adenomas [20]. At variance with the 18 kDa,the 22 kDa isoform occurred only in the nuclear frac-tion in all experimental groups. This molecular isoform
attained the highest concentration at 7 days of estrogenstimulation, and then decreased progressively but with-out reaching the level of the control values. This iso-form was not correlated with lactotroph proliferation(r = 0.11) and our study has not shown a direct associa-tion of this FGF-2 isoform with the development of pro-lactinomas. A recent study has described that HMWFGF-2 has speciWcity in the regulation of genesinvolved in the arrest of cell growth and the suppressionof tumors [35]. The presence of this FGF-2 isoform innuclear extracts has been discussed in various publica-tions and it has been suggested that the existence ofN-terminal nuclear localization signals in the HMWFGF-2 isoforms confer that localization [3].
The immunoelectron labeling of FGF-2 in the cyto-solic matrix suggests a possible alternative route for acytokine that is devoid of a signal sequence to reachthe extracellular space through a nonconventionalmechanism [5, 31]. In addition, the immunodetectionof FGF-2 in the nucleus and speciWcally in the nucleo-lus supports the notion that it could act through anintracrine pathway [37].
Another interesting observation was the identiWca-tion of FGF-2 in diVerent types of secretory pituitarycells of normal rat in addition to its classical location inFSC. Therefore, our results demonstrated that thiscytokine exhibits ubiquitous distribution in the pitui-tary gland. Some studies have shown that FSC regulatethe cell proliferative activity of estradiol on lactotrophsthrough the release of FGF-2. In this way, FGF-2 couldoriginate loops of cell–cell communications in the pitu-itary [9, 33].
After estrogen treatment, FGF-2 was expressedmainly in lactotrophs and gonadotrophs, the predomi-nant cell types observed under these experimental con-ditions. In this state, lactotroph cells exhibithyperplastic and hypertrophic changes. DiVerent inves-tigators have proposed that FGF-2 is the physiologicalinducer of diVerentiation of the mammotroph lineage,probably maintaining the lactotroph cell phenotype[13, 22]. It is likely, therefore, that FGF-2 remainswithin these cells, where it could act as a direct intra-cellular signal occurring during transdiVerentiation.Throughout this process, monohormonal cells may alsobe multipotential reserve cells, allowing the intercon-version of pituitary cells that could give rise to prolacti-nomas [29].
Likewise, estrogen stimulates the production ofFGF-2, which in turn modulates angiogenesis, tumorformation, and progression in many tissues, includingthe anterior pituitary [7, 18]. In cultured mouse endo-thelial cells, estrogen increases only the HMW FGF-2,conWrming that this molecular isoform is required for
Fig. 11 Western blot analysis of 22 kDa FGF-2 protein in pituitarynuclear extracts of control and estrogen-treated rats. The relativelevels of 22 kDa FGF-2 isoform were determined by densitometricanalysis. The expression of this isoform showed the highest level at7 days of hormonal treatment with a progressive decrease untilEB-60. Data are expressed as mean § SEM of Wve anterior pitui-tary glands for each group of three independent experiments.*P < 0.001 EB-7, EB-20 versus control; +P < 0.05 EB-60 versuscontrol; **P < 0.001 EB-20 and EB-60 versus EB-7; ***P < 0.001EB-60 versus EB-20. ANOVA followed by Tukey’s test
Arb
itrar
y un
its
0
100
200
300
400
C EB-7 EB-20 EB-60
****
*****+
FGF-2 22 kDa expression in pituitary nuclear extracts
123

500 Acta Neuropathol (2006) 112:491–501
the stimulating eVect of this hormone on endothelialcell migration [17]. The highest levels of HMW FGF-2observed at EB-7 under our experimental conditionscould be the switch to activate the angiogenic process[41]. Seven days of estrogen stimulation provoked agradual diminution of the HMW, contrasting with theLMW isoform, which increased progressively untilEB-60. These changes in the pattern of expression ofFGF-2 isoforms are probably related to the angiogenicphenotype observed after 20 days of estrogen treat-ment. Estrogen in fact could be mediating the vascularchanges with the participation of FGF-2 and the PTTGoncogene [4, 27].
The relevant Wndings of this study are the diVeren-tial expression patterns of FGF-2 isoforms quantiWedin diVerent subcellular compartments, and the closecorrelation obtained between 18 kDa FGF-2 and lacto-troph proliferation, suggesting that this molecular iso-form may play a role in the modulation of this processin experimental prolactinomas.
Taken together, the data obtained shed new light onthe possible actions of the 18 and 22 kDa FGF-2 iso-forms and give evidence that the overproduction ofFGF-2 by target cells could result in the establishmentof autocrine–paracrine–intracrine mitogenic loops,whose activity could lead to uncontrolled cell growth,tumor formation, and angiogenesis.
Acknowledgements We are especially grateful to Professor Dr.Sylvia L. Asa from the Department of Laboratory Medicine andPathobiology, University of Toronto for her critical review of themanuscript. The technical assistance of Mrs Mercedes Guevaraand Biochem. Félix D. Roth is gratefully acknowledged. Finally,we would like to thank native speaker Dr Paul Hobson for revis-ing the English of the manuscript. This work was supported bygrants from Consejo Nacional de Investigaciones CientíWcas yTécnicas (CONICET) and the Secretaría de Ciencia y Tecnologíade la Universidad Nacional de Córdoba (SECyT).
References
1. Akashi T, Minami J, Ishige Y, Eishi Y, Takizawa T, Koike M,Yanagishita M (2005) Basement membrane matrix modiWescytokine interactions between lung cancer cells and Wbro-blasts. Pathobiology 72:250–259
2. Amano O, Yoshitake Y, Nishikawa K, Iseki S (1993) Immu-nocytochemical localization of basic Wbroblast growth factorin the rat pituitary gland. Arch Histol Cytol 56:269–276
3. Arnaud E, Touriol C, Boutonnet C, Gensac MC, Vagner S,Prats H, Prats AC (1999) A new 34-kilodalton isoform of hu-man Wbroblast growth factor 2 is cap dependently synthesizedby using a non-AUG start codon and behaves as a survivalfactor. Mol Cell Biol 19:505–514
4. Asa S, Ezzat S (2002) The pathogenesis of pituitary tumours.Nat Rev Cancer 2:1–12
5. Backhaus R, Zehe C, Wegehingel S, Kehlenbach A,Schwappach B, Nickel W (2004) Unconventional protein
secretion: membrane translocation of FGF-2 does not requireprotein unfolding. J Cell Sci 117:1727–1736
6. Baird A, Esch F, Mormede P, Ueno N, Ling N, Bohlen P,Ying SY, Wehrenberg WB, Guillemin R (1986) Molecularcharacterization of Wbroblast growth factor: distribution andbiological activities in various tissues. Recent Prog Horm Res42:143–205
7. Bikfalvi A, Klein S, Pintucci G, Rifkin D (1997) Biologicalroles of Wbroblast growth factor 2. Endocr Rev 18:26–45
8. Bugler B, Amalric F, Prats H (1991) Alternative initiation oftranslation determines cytoplasmic or nuclear localization ofbasic Wbroblast growth factor. Mol Cell Biol 11:573–577
9. Chaturvedi K, Sarkar DK (2004) Involvement of protein ki-nase C-dependent mitogen-activated protein kinase p44/42signaling pathway for cross-talk between estradiol and trans-forming growth factor-beta3 in increasing basic Wbroblastgrowth factor in folliculostellate cells. Endocrinology145:706–715
10. Chaturvedi K, Sarkar DK (2005) Mediation of basic Wbro-blast growth factor-induced lactotropic cell proliferation bySrc-Ras-mitogen-activated protein kinase p44/42 signaling.Endocrinology 146:1948–1955
11. Chen CH, Poucher SM, Lu J, Henry PD (2004) Fibroblastgrowth factor 2: from laboratory evidence to clinical applica-tion. Curr Vasc Pharmacol 2:33–43
12. Delrieu I (2000) The high molecular weight isoforms of basicWbroblast growth factor (FGF-2): an insight into an intracrinemechanism. FEBS Lett 468:6–10
13. Ezzat S (2001) The role of hormone, growth factors and theirreceptors in pituitary tumorigenesis. Brain Pathol 11:356–370
14. Florkiewicz RZ, Sommer A (1989) Human basic Wbroblastgrowth factor gene encodes four polypeptides: three initiatetranslation from non-AUG codons. Proc Natl Acad Sci USA86:3978–3981
15. Fukui S, Otani N, Nawashiro H, Yano N, Miyazaqwa T, Oh-nuki A, Tsuzuki N, Katoh H, Ishihara S, Suzuki T, Shima K(2002) Nuclear accumulation of basic Wbroblast growth factoras a predictor for the recurrence of pituitary adenomas. JNeurooncol 57:221–229
16. Fukui S, Otani N, Nawashiro H, Yano N, Miyazaqwa T, Oh-nuki A, Tsuzuki N, Katoh H, Ishihara S, Suzuki T, Shima K(2003) Nuclear accumulation of basic Wbroblast growth factorin human astrocytic tumors. Cancer 97:3061–3067
17. Garmy-Susini B, Delmas E, Gourdy P, Zhou M, Bossard C,Bugler B, Bayard F, Krust A, Prats AC, Doetschman T, PratsH, Arnal JF (2004) Role of Wbroblast growth factor-2 iso-forms in the eVect of estradiol on endothelial cell migrationand proliferation. Circ Res 94:1301–1309
18. Heaney A, Horwitz G, Wang Z, Singson R, Melmed S (1999)Early involvement of estrogen-induced pituitary tumourtransforming gene and Wbroblast growth factor expression inprolactinoma pathogenesis. Nat Med 5:1317–1321
19. Itoh J, Serizawa A, Kawai K, Ishii Y, Teramoto A, OsamuraYoshiyuki R (2003) Vascular networks and endothelial cellsin rat experimental pituitary glands and in the human pitui-tary adenomas. Microsc Res Tech 60:231–235
20. Kang HB, Kim JS, Kwon HJ, Nam KH, Youn HS, Sok DE,Lee Y (2005) Basic Wbroblast growth factor activates ERKand induces c-fos in human embryonic stem cell line Miz-hES1. Stem Cells Dev 14:395–401
21. Keresztes M, Boonstra J (1999) Importance of growth factorsinto the nucleus. J Cell Biol 145:421–424
22. López J, Fernández I, Palacios D, Castillo A, Tolón R, Aran-da A, Karin M (2000) DiVerentiation of lactotrope precursorGHFT cells in response to Wbroblast growth factor-2. J BiolChem 275:21653–21660
123

Acta Neuropathol (2006) 112:491–501 501
23. Madan AK, Kramer B (2005) Immunolocalization of Wbroblastgrowth factor-2 (FGF-2) in the developing root and supportingstructures of the murine tooth. J Mol Histol 36:171–178
24. Maher PA (1996) Nuclear translocation of Wbroblast growthfactor (FGF) receptors in response to FGF-2. J Cell Biol134:529–536
25. Maldonado C, Aoki A (1986) Improvement of prolactin im-muno-labeling in osmium-Wxed acrylic-embedded pituitarygland. Basic Appl Histochem 30:301–305
26. Marin F, Boya J (1995) Immunocytochemical localization ofbasic Wbroblast growth factor in the human pituitary gland.Neuroendocrinology 62:523–529
27. Melmed S (2003) Mechanisms for pituitary tumorigenesis:the plastic pituitary. J Clin Invest 112:1603–1618
28. Mignatti P, Morimoto T, Rifkin DB (1991) Basic Wbroblastgrowth factor released by single, isolated cells stimulates theirmigration in an autocrine manner. Proc Natl Acad Sci USA88:11007–11111
29. Mukdsi JH, De Paul AL, Munoz S, Aoki A, Torres AI (2004)Immunolocalization of Pit-1 in gonadotroph nuclei is indica-tive of the transdiVerentiation of gonadotroph to lactotrophcells in prolactinomas induced by estrogen. Histochem CellBiol 121:453–462
30. Mukdsi JH, De Paul AL, Gutiérrez S, Roth F, Aoki A, TorresAI (2006) Subcellular localisation of VEGF in diVerent pitu-itary cells. Changes of its expression in estrogen induced pro-lactinomas. J Mol Histol 36:447–454
31. Nickel W (2003) The mystery of nonclassical protein secre-tion. A current view on cargo proteins and potential exportroutes. Eur J Biochem 270:2109–2119
32. Okada-Ban M, Thiery JP, Jouanneau J (2000) Fibroblastgrowth factor-2. Int J Biochem Cell Biol 32:263–267
33. Oomizu S, Chaturvedi K, Sarkada DK (2004) Folliculostel-late cells determine the susceptibility of lactotropes to estra-diol’s mitogenic action. Endocrinology 145:1473–1480
34. Piroli G, Torres AI, Pietranera L, Grillo C, Ferrini M, Lux-Santos V, Aoki A, De Nicola A (2000) Sexual dimorphism indiethylstilbestrol-induced prolactin pituitary tumors in F344rats. Neuroendocrinology 72:80–90
35. Quarto N, Fong KD, Longaker MT (2005) Gene proWling ofcells expressing diVerent FGF-2 forms. Gene 56:49–68
36. Re RN (2003) The intracrine hypothesis and intracellularpeptide hormone action. Bioessays 25:401–409
37. Re RN (2004) A proposal regarding the biology of memory:participation of intracrine peptide networks. Med Hypothe-ses 63:887–894
38. Renner U, Paez-Pereda M, Arzt E, Stalla GK (2004) Growthfactors and cytokines: function and molecular regulation inpituitary adenomas. Front Horm Res 32:96–109
39. Schechter JE (1992) Is cellular disruption the mechanism ofrelease of basic Wbroblast growth factor from anterior pitui-tary gonadotropes? Tissue Cell 24:791–802
40. Sugawara A, Yen PM, Darling DS, Chin W (1993) Character-ization and tissue expression of multiple triiodothyroninereceptor-auxiliary proteins and their relationship to the reti-noid X-receptor. Endocrinology 133:965–971
41. Turner HE, Harris AL, Melmed S, Wass JA (2003) Angio-genesis in endocrine tumors. Endocr Rev 24:600–632
42. Yamada S, Takada K (2003) Angiogenesis in pituitary adeno-mas. Microsc Res Tech 60:236–243
43. Zhan X, Hu X, Friedman S, Maciag T (1992) Analysis ofendogenous and exogenous nuclear translocation of Wbro-blast growth factor-1 in NIH 3T3 cells. Biochem Biophys ResCommun 188:982–991
123




















