
Growth hormone, prolactin and cortisol response to exercise in patients with depression
Upload
khangminh22Category
view
2download
0
Pathophysiology of the Neuroregulation of GrowthHormone Secretion in Experimental Animals
and the Human*
ANDREA GIUSTINA AND JOHANNES D. VELDHUIS
Endocrine Section (A.G.), Department of Internal Medicine, University of Brescia, 25125 Brescia, Italy;and Division of Endocrinology (J.D.V.), Department of Internal Medicine and National ScienceFoundation Center in Biological Timing, University of Virginia, Charlottesville, Virginia 22908
I. IntroductionII. Contemporary Tools for Neuroendocrinological In-
vestigation of the GH AxisA. Genetic, molecular, and transgenic modelsB. Human GH rhythms
III. Neuropeptide Regulation of the GH Axis: Somatosta-tin and GHRHA. Mechanism of somatostatin actions and its recep-
torsB. Human somatostatin receptorC. GHRHD. Feedback regulation of GH secretion
IV. Other GH-Regulating NeuropeptidesA. GH-releasing peptides (GHRPs)B. GalaninC. CalcitoninD. PACAPE. Opioid peptidesF. TRHG. Neuropeptide YH. Substance PI. BombesinJ. Melatonin
K. Other GH secretagoguesV. Neurotransmitter Regulation of GH Secretion
A. Interspecies differencesB. Acetylcholine and catecholaminesC. Other neurotransmitters
VI. Role of Metabolic Substrates in the Regulation of theGH AxisA. Blood glucoseB. Leptin and FFAC. Amino acids
VII. Other Hormonal Regulators of the GH AxisA. Glucocorticoids
B. Gonadal sex hormonesC. Thyroid hormones
VIII. Regulation of the GH Axis Throughout the HumanLifetimeA. Birth and infancyB. PrepubertyC. PubertyD. AdulthoodE. AgingF. GH treatment in older humans
IX. Exercise’s Modulation of the GH AxisA. Experimental animalsB. HumansC. Neural control of GH release during exerciseD. Kinetics of exercise-induced GH release
X. Summary
I. Introduction
MULTIPLE neurotransmitter pathways, as well as avariety of peripheral feedback signals, regulate
GH secretion either by acting directly on the anterior pi-tuitary gland and/or by modulating GH-releasing hor-mone (GHRH) or somatostatin release, or both, from thehypothalamus (Fig. 1). After the isolation and character-ization of GHRH and the cloning of its receptor, as well asthe more recent availability of molecular probes for so-matostatin and its recently cloned receptor family, moredetailed studies of the pathophysiological neuroregula-tion of GH secretion have been performed. In this review,we will update earlier discussions in the Journal (1, 2) bypresenting recent developments in our understanding ofneuroregulatory mechanisms and their relevance to clin-ical alterations in GH control. Primary diseases of thepituitary gland or hypothalamus (e.g., tumors originatingat these loci) resulting in excessive or deficient GH secre-tion and peripheral actions of GH or insulin-like growthfactor-I (IGF-I) on target tissues will not be reviewed here,but are discussed recently elsewhere (3–9). Where possi-ble, major interspecies differences in neuroregulation ofthe GH axis (rat, sheep, and human) will be distinguishedto limit any confusion on this basis.
Address reprint requests to: Johannes D. Velduis, M.D., Division ofEndocrinology, Department of Internal Medicine, Box 202, University ofVirginia Health Sciences Center, Charlottesville, Virginia 22908 USA.E-mail: [email protected]
* This work was partially supported by the Centro Studi e Ricerchedi Neuroendocrinologia (Brescia, Italy), by MURST, and by RegioneLombardia (Project microalbuminuria (A.G.), NIH Grant RR-00847 tothe General Clinical Research Center, the National Science FoundationCenter for Biological Timing, and NIH Grants NIA RO1 AG 147991 andRO3 AG14873 (to J.D.V.).
0163-769X/98/$03.00/0Endocrine Reviews 19(6): 717–797Copyright © 1998 by The Endocrine SocietyPrinted in U.S.A.
717
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

FIG. 1. Summary representation of the putative roles of the principal neuropeptides and neurotransmitters that supervise GH secretion viaGHRH or somatostatin (SS) or by acting directly on the pituitary gland (GH) in the rat (panel A) or human (panel B). Asterisks denote thattwo or more loci of action are recognized. Not shown are numerous other metabolic and hormonal effectors that also act via multiple pathways,e.g., IGF-I, sex steroids, age, glucocorticoids, diabetes mellitus, obesity, T4, etc. (see text). An unproven role for a putative (as yet unidentified)GHRP-like endogenous ligand is also noted, given that receptors for GHRP ligands are expressed in the hypothalamus and pituitary gland.Table 4 gives some further species distinctions among the rat, sheep, and human.
718 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

II. Contemporary Tools for NeuroendocrinologicalInvestigation of the GH Axis
A. Genetic, molecular, and transgenic models
Various highly informative genetic and transgenic, as wellas gene-knockout, animal models have emerged that provideinstructive insights into molecular mechanisms of neuro-regulation of the GH axis (see summary in Table 1). Severalvaluable molecular models, namely, genetically impairedGH secretion in the little (lit) mouse, the dwarf (dw) rat, thefatty (fa) rat, and the high-growth (hg) mouse, were recentlyreviewed in detail by Frohman (10), to which the reader isreferred. Conversely, the GHRH-transgenic mouse exhibitsexcessive GH secretion, pituitary somatotrope hyperplasia,and increased plasma IGF-I levels (11) that are antagonizedby potent GHRH-receptor inhibitors (12). In addition, trans-genic (rodent) models of suppressed secretion have beendeveloped, e.g., utilizing hypothalamically targeted GH orGHRH transgenes (13–15). The experimental notion of tar-geting GH gene expression selectively to the hypothalamus,and thereby producing deficiency of endogenous GH, hasdisclosed alterations in non-rapid-eye-movement sleep inthe induced hyposomatotropic state (16). Targeting of theGH gene in the mouse to the hypothalamus was accom-plished via use of either the tyrosine hydroxylase or GHRH
promoters to drive topographically localized GH excess(13–15). This approach thus allows study of hypothalamic-regulatory peptide responses to localized overexpression ofa selected gene. In the case of GH gene overexpression tar-geted to the hypothalamus, the pathophysiological re-sponses include increased hypothalamic somatostatin anddecreased GHRH accumulation, as predicted by other ex-periments of GH autofeedback (17–19). Conversely, the ge-netic model of the Ames dwarf mouse, which has a recessivedefect with a hypocellular anterior pituitary gland lackingsomatotrophs, lactotropes, and thyrotrophs (20), shows, aspredicted from physiological principles, increased expres-sion of GHRH peptide and mRNA in the hypothalamus (21).This follows from the premise that GH autofeedback nor-mally suppresses GHRH and stimulates somatostatin ex-pression (19). This thesis has recently been corroborated viaimplantable GH-secreting (GC) cells in primary genetic vs.transgenic GH-deficient rat models (22).
Nontargeted GH transgene overexpression, either bovineGH, which is largely free of lactotrophic effects, or humanGH, which has combined lactotrophic and somatotrophicactions, was reviewed recently by Bartke et al. (23). Repro-ductive sequelae of GH excess can also be appraised via suchmolecular strategies and, conversely, GH deficiency can beassessed by GH receptor knockout mice (17, 23–25). IGF-I
TABLE 1. Selected examples of molecular strategies for investigating the GH-IGF-I axis
Molecular strategy Effect Illustrative references
1. Hypothalamic GH transgene overexpression(targeted via the tyrosine hydroxylase or GHRHpromoter)
Endogenous GH deficiency (via GH autonegativefeedback) and dwarfism
(13, 15)
2. GH transgene (human or bovine) Growth excess, reproductive sequelae, etc. (17, 23)
3. GH-receptor antagonist transgene Blocks GH-induced renal glomerulosclerosis andgrowth
(37)
4. IGF-I vs. GH (trans-)gene overexpression GH but not IGF-I increases motoneuron size andinduces renal glomerulosclerosis
(38, 39)
5. Leukemia-inhibitory factor (pituitary-targeted)transgene
Rathke’s cleft cyst (42)
6. Reversible GH transgene activation Reversible obesity (43)
7. Nontargeted GHRH transgene Somatotroph cell hyperplasia/adenomas; GHexcess
(10)
8. IGF-I, and IGF-I receptor knockouts (mouse) Growth failure or death (27, 28)
9. STAT5b knockout (mouse) No response to (male-pattern) GH: dwarfism, lowIGF-I
(32)
10. Antisense GH RNA transgene Growth failure (44)
11. GHF-I overexpression (developmental trap) Dwarfism in mouse (41)
12. GH receptor mutation mouse (human) Dwarfism (Laron) (33, 35, 36)
13. GHRH receptor mutation (human) Dwarfism (Sindh) (47, 48)
14. IGF-I gene deletion Human dwarfism (49)
15. Myocyte-targeted GH or GHRH geneoverexpression (rodent)
Increased serum GH and IGF-I (45, 46)
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 719
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

transgene overexpression has been reviewed recently byD’Ercole (26), and IGF-I and its receptor knockouts have beencreated in mice (27, 28). Concurrent overexpression of IGFs,with or without GH, is a new strategy that will likely helpclarify the more complex interactions among and the relativeroles of GH, IGF-I, and IGF-II; e.g., see recent studies ofcombined GH and IGF-II transgenic mice (29). A transgenicmouse overexpressing IGF binding protein-1 (IGFBP-1)shows growth failure and hyperglycemia, thus suggestingthat this BP limits IGF-I actions to otherwise promote tissuegrowth and glucose uptake (30). Lastly, models of transgenicfarm animals have been developed and applied usefully, asreviewed by Pursel and Rexroad (31). The reader is referredto these individual reviews for an introduction to the mo-lecular-genetic strategies for investigating neuroregulationof the GH axis.
A recent novel gene knockout model, STAT5b gene dis-ruption, which inactivates a specific signal-transductionpathway evokes loss of multiple sexually differentiatedgrowth and cellular responses to GH, akin to those expectedif the sexually dimorphic (male) pattern of pituitary GHsecretion were abolished (32). This reflects the presumptiveintracellular transcriptional factor-signaling role of STAT5bin mediating the cellular and nuclear (gene-transcriptional)actions of pulsatile (but not continuous) GH delivery. Thus,the STAT5b knockout mouse, albeit not measurably GH de-ficient, appears to be GH pulse resistant as reflected bydwarfism, low plasma IGF-I concentrations, and obesity.These features also are recognized in human GH receptormolecular defects causing tissue resistance to GH actions,e.g., Laron-type dwarfism (6, 33–35), and in the GH receptorknockout mouse (36)
Another transgenic strategy, transgenic expression of aGH receptor antagonist, has confirmed an important role ofGH in antagonizing insulin action (37). Moreover, coexpres-sion of the bovine GH gene and a human GH-receptor an-tagonist minigene in mice shows that the GH receptor an-tagonist will oppose the development of severe renalglomerulosclerosis and the increase in body growth other-wise driven by excess GH (38). In contrast, IGF-I overex-pression augments body growth without inducing glomer-ulosclerosis, thus distinguishing certain tissue actions of GHand IGF-I. Indeed, another selective role of GH, but not IGF-I,is to increase motor neuron size in the lumbar spine, asinferred in transgenic mice overexpressing GH vs. IGF-I pep-tide (39).
In relation to pituitary developmental mechanisms, trans-genic ablation (via coupling a relevant glycoprotein a-sub-unit cDNA to diptheria toxin A chain) produces loss of go-nadotrophs and thyrotrophs, as distinct from somatotropeand corticotroph cell lineages (40). Other recent experimentsindicate the potential to “trap” developmental progenitorcells by using transgenic overexpression of regulatory re-gions responsive to cell-specific transcription factors, e.g., thehomeodomain protein, GHF-1. The latter is believed to berequired for the developmental generation of somatotropesand lactotrophs, and hence GH and PRL gene expression(41). This technique of so-called developmental entrapmentcan evaluate the roles of GH progenitor cells by constitutiveoverexpression of GHF-1, and hence “immortalization” of
GH or PRL progenitor cells, which results in dwarfism. Apituitary-targeted transgenic mouse model expressing theleukemia inhibitory factor has suggested the possible role ofaltered pituitary gene transcription and cell replication in thepathogenesis of Rathke’s cysts (42).
Another informative molecular strategy is experimentallycontrolled transgene expression. For example, exogenouslytriggered activation and inactivation of the GH transgene inmice will reversibly reduce and induce obesity in this species(43). GH antisense RNA-transgene transfer in rats hasachieved dose-dependent inhibition of GH gene expression,thus reducing GH secretion to varying degrees in heterozy-gous and homozygous transgenic animals (44). Conversely,expression vectors designed to transfer the GH gene as di-rected by tissue-specific promoters (e.g., muscle) provide anexciting prospect for targeted delivery of GH to, and forregulated expression of GH in, specific cell types, such asmyoblasts in vitro (45) or myocytes in vivo (46). Indeed, in thelatter, in vivo context or single injection of a myocyte-targetedGHRH-minigene expression vector can stimulate GH secre-tion and increase serum IGF-I levels by 3- to 4-fold or morefor 2 weeks.
In brief, a rich and expanding repertoire of genetic, mo-lecular, and transgenic and knockout models of receptors,regulatory peptides, IGF-I or GH itself, and/or signalingmolecules has emerged and continues to unfold. Thisresource will make more specific and compelling studiespossible of the molecular mechanisms of neuroendocrineregulation of the hypothalamo-somatotrophic axis in exper-imental animals. In some circumstances, a clinical counter-part is identifiable, in which a mutation of the correspondinggene is discernible in an individual or family. For example,IGF-I gene deletion, GH receptor, and GHRH receptor mu-tations with dwarfism all are recognized in the human (6, 28,33–35, 47–49). However, GHRH receptor mutations causingisolated GH deficiency in the human are not common (50).
B. Human GH rhythms
1. Introduction. In conjunction with the development of in-creasingly specific pharmacological agents to block or acti-vate individual regulatory receptors, new neuroendocrinetools have been developed that allow improved quantitativemeasures of hormone release over short intervals (ultradianrhythms or episodic peaks), as well as over 24 h (circadianrhythms), and in relation to the patterned orderliness of therelease process (e.g., approximate entropy estimates). Thesenovel technologies often aid in dissecting the neurohormonalmechanisms that underlie pathophysiological states or mo-lecular models (above). Biomathematical advances have alsoenhanced clinical neuroendocrinological investigation, sincein the human direct portal catheterization techniques imple-mented in experimental animals to monitor hypothalamo-pituitary secretion of GH, GHRH, and somatostatin individ-ually and concurrently, e.g., in the rat and sheep (29, 51–56),are not practicable ethically.
2. Background. Even relatively direct studies are not alwaysfree of controversy. For example, a portal vein samplingstudy in the anesthetized male rat indicated that both so-
720 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

matostatin withdrawal and GHRH release tend to coincidewith a GH pulse (56). Other repetitive portal blood samplinginvestigations in the unrestrained and unanesthetized (ovari-ectomized or intact male) sheep indicate that GHRH incre-ments typically precede GH pulses, but somatostatin decre-ments do not necessarily do so (51, 57–60) (Fig. 2). Moreover,in sheep, somatostatin release can actually rise before a GHpulse or remain unchanged (see Table 2). Consequently,GHRH-somatostatin interrelationships in this species may bevariable and quite complex. For example, stress appears tostimulate both GHRH and somatostatin release into portalblood, although the net result is increased GH secretion (61).In addition, technical differences (e.g., anesthesia, samplingfrequency, etc.) may confound a simple mechanistic modeloriginated in the adult male rat consisting of reciprocal so-matostatin withdrawal and GHRH release in initiating a GHpulse. Indeed, taken as a whole, available studies clearlyallow for other input into GH secretory-burst timing, such as1) extra- or intrapituitary feedback by IGF-I (62–64); 2) in-trapituitary paracrine factors; 3) other hypothalamic coregu-lators [e.g., endogenous GH-releasing peptides (GHRPs), ga-lanin, neuropeptide Y (NPY), etc.]; for example, either NPYor galanin can colocalize with GHRH in the hypothalamus(65, 66); and/or 4) other as yet unexplained technical factors(e.g., hydraulic variations in blood flow or sample collection;inconsistencies in GHRH, somatostatin, or GH immunoas-says; etc.).
3. Clinical study methodologies. Neuroendocrine researcherscurrently have several available methodologies for identify-ing, counting, and further quantifying by way of amplitude,duration, and/or area discrete hormone release events (i.e.,pulses) (67–72). Pulse analysis is challenged by short, noisy,and irregular hormone concentration profiles measured in a
body fluid such as peripheral blood (73–77). Such short-termvariations in blood hormone concentrations, e.g., distinct(GH) pulses, are believed to provide a window into thebrain-pituitary interface, whereby organized neural signalsdirect bursts of anterior pituitary hormone secretion (78, 79)Importantly, an adequate schedule of sufficiently frequent(intensively) repetitive and prolonged blood sampling fol-lowed by exemplary assay conditions (below) must be im-plemented to obtain valid, significant, and insightful quan-titative information concerning pulsatile GH release (76, 80).In this setting (GH), pulse and entropy (below) analyses arequite reproducible (81).
Considerable clinical effort and numerous studies haveappraised the endocrine, metabolic, and pharmacologicalregulation of episodic GH release (82–89). For example, inthe case of the GH axis, a burst of GH release presumablyreflects an appropriate stimulus such as endogenous GHRHsecretion (with or without other putative secretagogues) inthe context of responsive somatotropes (i.e., at least not max-imally inhibited by somatostatin), and adequate releasableGH pools within the anterior pituitary gland. Discrete pulseanalysis provides information about the frequency and am-plitude of serum hormone concentration peaks (67, 68, 70)and can disclose insights into neuroendocrine mechanismsthat control this ultradian pituitary release activity. For ex-ample, sampling blood every 30 sec during sleep in youngmen has revealed a strongly correlated pattern of episodicGH release and the onset of slow-wave (stages III and IV)sleep (90). Significantly, in some species such as the rat, thereis abundant evidence that the time profile of (pulsatile) GHrelease strongly conditions the target tissue response antic-ipated, e.g., induction of the LDL receptor and GH receptor(GHBP) genes by a “feminized”/continuous GH release pat-tern, rather than by a “masculinized”/pulsatile GH profile(91) (see Table 3). Hence, substantial physiological effort hasbeen applied to understand the neuroendocrine mechanismsthat supervise and dictate such pulsatile (GH) release pat-terns.
In addition to marking individual GH concentrationpulses in blood as discrete peaks, the newer methodologicalfield of deconvolution (“unraveling”) analysis allows esti-mates of underlying hormone secretion rates and in somecases concurrent calculations of endogenous hormone half-life (72, 73, 85, 92–103). The tools of deconvolution analysishave been reviewed recently and comprise a collection ofwaveform-specific (model-dependent) and waveform-inde-
TABLE 2. Relationships among portal vein GHRH andsomatostatin (SS) and jugular vein GH peaks in intact male sheepand castrate pigs
Condition% of total GH pulses
Rama Pigb
1 GHRH and 2 SS 48 261 GHRH and 1 (or no change) SS 19 222 SS, no change GHRH 13 37No changes SS or GHRH 9 0Other 11 15a Results reflect hypophysial-portal vein sampling in conscious
intact rams (n 5 23): Cataldi et al. (58), and b castrate (male) pigs (n 56): Drisko et al. (1098).
FIG. 2. Presumptive (schematized) individual and joint roles ofGHRH and somatostatin in generating spontaneous GH pulses in vivoin the rat (left panel) or sheep (right panel) based on available hy-pophysial portal blood sampling. GHRH and somatostatin fluctua-tions are illustrated over time. In both species, some, but not all,GHRH pulse trains induce volley-like GH release. In the rat, con-comitant somatostatin withdrawal is deemed prerequisite. In thesheep, episodic GHRH release drives GH pulse without uniform prioror concurrent somatostatin withdrawal. Pulsatile secretion of a pu-tative endogenous GHRP-like ligand is indicated by arrows and as-terisks, to foreshadow this possible (unproven) agonist, or some otherGH cosecretagogue(s), which might synergize with GHRH.
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 721
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

pendent strategies (104). In brief, deconvolution analyseswill provide an estimate of underlying hormone secretionrates from the serially measured serum hormone concentra-tion pulse profiles over time, thus uncovering informationabout regulation of secretory activity independently of, or inaddition to, hormone half-life (e.g., Ref. 105). One such modelof deconvolution analysis calculates both (GH) secretoryburst properties (amplitude, frequency, and mass) and (GH)half-life simultaneously (e.g., Refs. 72, 73, and 93 and see Fig.3A). This general class of analytical technology thereby af-fords insights into the neuroendocrine control of the under-lying secretory events per se (104), which may be regulatedin principle by way of shape, duration, maximal amplitude,and/or mass (106), with or without any attendant changes inhormone half-life or distribution volume (73, 93, 94, 107).Indeed, altered GH half-lives are likely only in subjects withseverely impaired hepatic or renal function [prolonged GHhalf-lives (108–111)], and in obesity [reduced GH life-life(112–114)]. Most recently, a stochastic differential equation,random effects, maximum-likelihood methodology based onfeedback concepts within a neuroendocrine axis, ultradianpulsatility coupled to circadian variations, and biexponentialkinetics has emerged to capture secretory dynamics morefully (115–117).
Additional recent critical developments are ultrahigh sen-sitivity assays of GH, such as afforded by immunofluoro-metric (118) and chemiluminescence principles (119–121).For example, a recent immunofluorometric assay of GH inthe human with sensitivity of 0.013 mg/liter disclosed lowlevels of basal GH release (concentrations of ;0.020–0.05mg/liter). This assay also revealed a pulse amplitude- (andmass-) dependent mechanism for the gender distinction be-tween GH release in healthy middle-aged men and women(118). An even more sensitive chemiluminescence-based GHassay (sensitivity of ;0.002–0.005 mg/liter) confirmed smallamounts of (,15%) basal GH release in healthy men andwomen and illustrated that glucose ingestion reduces serumGH concentrations typically to less than 0.7 mg/liter inwomen and to less than 0.07 mg/liter in men (119–121). Thus,earlier “normal” glucose suppression of serum GH concen-trations assessed by RIA or IRMA has been grossly mises-timated and has also overlooked a strong gender difference.A high-sensitivity enzyme-linked immunosorbent assay(ELISA) also corroborated very low rates of basal GH releasein both normal and hypopituitary adults (122–124). Howsuch basal secretion is generated or regulated is not known(125). To our knowledge, detectable interpulse basal GH
release has not yet been measured in the male rat or othernonhuman species.
Statistically correct treatment of assay data variability, es-pecially at the low end, is also essential (126). Variabilityarises from uncertainties in the zero-dose tubes, among otherreplicates in the standard curve, due to the fit of the standardcurve itself, and from the replicates of the unknown samples(127). Methods have been created recently to address thesefour joint/combined sources of within-assay experimentaluncertainty (126, 128).
Clinical studies now indicate that visceral obesity, age, andother states of relative hyposomatotropism can be attributedmechanistically to decrease in GH-secretory burst mass, asestimated by the combined techniques of deconvolutionanalysis and ultrasensitive GH assays (121). Notably, asmany as 97% of daytime samples for serum GH concentra-tions in older and/or obese subjects can be undetectable byconventional RIA or IRMA methods (125). Immunofluoro-metric, ELISA, and chemiluminescence GH measurementtechniques, combined further in some cases with improvedstatistical assay analyses at the low end (126), have overcomesome earlier limitations in studying dynamics of the humanGH axis (125). For example, an ultrasensitive human GHassay has revealed that somatostatin infusion in young mensuppresses both GH pulse amplitude and frequency (123).This is consistent with somatostatin’s putative inhibition notonly of pituitary GH secretion but also of hypothalamicGHRH release (22, 129, 130). The reciprocal relationship be-tween somatostatin and GHRH has also been inferred re-cently in in vitro studies in bovine hypothalamus (131).
Beyond and complementary to the pulsatile and 24-h (nyc-tohemeral) modes of GH secretion, the subordinate (non-pulsatile) regularity or pattern orderliness of hormone re-lease over time can be quantified now by a so-calledapproximate entropy statistic (132, 133). This is a model-freeand scale-invariant measure different from deterministicchaos (nonlinear dynamical measures). An intuitive notionof the entropy statistic is shown in Fig. 3B. The orderlinessof GH release over time so quantified is markedly altered incertain conditions of health or disease. For example, acro-megalic patients secrete GH with a high degree of quantifi-able disorderliness, i.e., there is a profound loss of or dete-rioration in regularity or reproducibility of point-by-pointsubpatterns recurring across the 24-h GH profile, whetherdefined by the approximate entropy statistic or a network-based predictability measure (134–138). The distinctions be-tween GH-secretory patterns in active and remitted acro-megaly as achieved via the approximate entropy statistic areillustrated in Fig. 4.
A strong gender contrast in the orderliness or regularity ofGH-secretory activity is quantifiable by the approximate en-tropy statistic in the rat and human. Entropy analysis revealsstatistically remarkably greater disorderliness or irregularityof GH release in the intact female than in the male in bothspecies (139). Indeed, in the adult rat, ApEn discriminatesdisorderliness of GH release across the following rank order(from maximally to minimally irregular): intact female,GnRH agonist (triptorelin)-treated female, ovariectomizedrat, orchidectomized rat, GHRH agonist-treated male rat,and intact male rat (140) (Fig. 5). Unlike deterministic chaos
TABLE 3. GH pattern-dependent tissue responses
Pulsatile GH (male) Continuous GH (female)
Linear growth Hepatic GH and LDL receptorsBody weight Hepatic sulfataseSkeletal muscle IGF-I CBGHepatic P450 2C11 steroid
hydroxylaseGlutathione-S-transferase
(several isotypes suppressed)Hepatic EGF receptor Hepatic 5-a-reductaseSTAT 5b tyroxine
phosphroylationHepatic P450 2C12 steroid
hydroxylaseHepatic aldehyde oxidaseCarbonic anhydrase III
LDL, Low-density lipoprotein; CBG, corticosteroid binding globu-lin.
722 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

measures, which require 105 or more observations (and the-oretically noise-free data), approximative entropy values canbe calculated with good statistical replicability in hormoneseries containing as few as 50–300 samples (118, 140–142).Such a small sample size confers significant utility to this newmeasure.
The relevance of approximate entropy as an indirectbarometer of feedback control is also indicated by theability of specific sex steroid hormones, namely, estradiolor testosterone (but not a nonaromatizable androgen), notonly to amplify pulsatile GH secretion but also to reducethe measurable orderliness of the GH release process inprepubertal individuals (142). Concurrently with the nor-mal maximal growth spurt in boys, the pattern of GHrelease also becomes quantifiably more irregular (142) assex steroid concentrations rise. Such studies indicate that
not only the pulsatility but also the orderliness of GHrelease is controlled by sex steroid hormones (143, 144).More remarkable loss of regularity of GH release patternsis evident in acromegaly as assessed by approximate en-tropy (134, 136) or independently via a neural networkmodel (145). Similarly, heightened disorderliness of hor-mone release over time is evident for ACTH-secretingpituitary tumors (146, 147) and for feedback-withdrawnneuroendocrine axes (148 –153).
In addition to the above available tools, other recent stud-ies have begun to model the network or feedback-controllinkages within the GH and other axes, to provide a morequantitative basis for articulating, testing, and revising spe-cific neuroendocrine hypotheses (76, 154, 155). Considerableadditional research will be required to refine the correctconceptual basis and dynamic features of the neuroendo-
FIG. 3. A, Schematized illustration of a model-specific deconvolution concept to quantitate GH secretion. The upper landscape depicts an intuitiveformulation of a hormone-secretory burst, as arising from (multi-)cellular discharge of individual hormone molecules more or less in concerttemporally, each at its own particular secretory rate (velocity). A secretory burst (or pulse) is visualized as an array of such molecular secretoryvelocities centered about some moment in time, and dispersed around this center with a finite standard duration (SD) or half-width (93). Theburst event may or may not be symmetric over time (72, 1097). The lower landscape with the algebraic subheads illustrates the mathematicalnotion, whereby a plasma hormone concentration peak (far right) is viewed as developing from a burst-like secretory process (far left) and afinite hormone-specific removal rate (half-life of elimination) (73, 94). The so-called “convolution” (intertwining or interaction) of the simul-taneous secretory and elimination functions creates a resultant (skewed) plasma concentration pulse. Deconvolution analysis consists ofmathematically estimating the constituent underlying secretory features (and/or associated half-life), given a series of blood hormone con-centration peaks as the starting point. A variety of model-independent (waveform-invariant) deconvolution strategies can also be applied, ifa priori knowledge of the pertinent (biexponential) hormone elimination rate process is available (72, 73, 94, 95).B, Intuitive illustration of concept of approximate entropy to evaluate pattern reproducibility in hormone time series. This statistic iscomplementary to pulse analysis by quantifying the orderliness or regularity of subpatterns in the data (131, 132).
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 723
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

FIG. 4. A, Approximate entropy (ApEn) values for 24-h (5-min sampled) serum GH concentration (IRMA) profiles in acromegalic (male andfemale) patients with active or inactive (treated) disease compared with normal (fed or fasted gender-matched) volunteers. The upper bar graphsillustrate that GH secretion in acromegaly is remarkably more disorderly, as quantified by significantly higher ApEn values, than normalgender-matched GH secretion patterns, even when the latter are augmented by fasting. Higher ApEn denotes greater disorderliness, irreg-ularity, or randomness of the hormone release process. Acromegalic patients in remission have intermediate ApEn values. *, P , 1027 vs. normalfed and fasted subjects; 1, P , 0.02 vs. normal fed volunteers; #, P , 0.001 vs. active acromegalic and normal fed subjects. The bottom scatterplotshows individual subject’s ApEn values vs. mean 24-h serum GH concentrations on a logarithmic scale; the vertical broken line separates ApEnsin all but one acromegalic with active disease from normals (both fed and fasted). Œ, Acromegalic males; �, acromegalic females; f, normalfed males; M, normal fasted males; F, normal fed females; E, normal fasted females; ‚, acromegalic males in remission; ƒ, acromegalic femalesin remission. [Redrawn with permission from M. L. Hartman et al.: J Clin Invest 94:1277–1288, 1994 (134) by copyright permission of TheAmerican Society for Clinical Investigation.]
Panels B, C, and D: Illustrative individual male and female serum GH concentration profiles of control (normal, panel B) and acromegalicpatients with active (panel C) or inactive (remitted, panel D) disease sampled every 10 min for 24 h. GH was measured in duplicate in animmunofluorometric assay (sensitivity 0.013 mg/liter). Profiles are shown for both male and female subjects. Note variable y-axis scales toaccommodate a range of GH secretion rates and concentrations. The upper subpanels show deconvolution-predicted fits of the measured (6intrasample SD) serum GH concentrations, and the lower subpanels give the deconvolution-calculated GH secretory rates. Note increased basal(interpulse) GH release in active acromegalics. [Adapted with permission from G. Van den Berg et al.: J Clin Endocrinol Metab 79:1706–1715,1994 (962). © The Endocrine Society.]
724 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 725
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

crine component of the GH axis in appropriate biomath-ematical models.
III. Neuropeptide Regulation of the GH Axis:Somatostatin and GHRH
The physiology of somatostatin has been discussed in sev-eral reviews (156–159) and thus will only be addressedbriefly here in pertinent sections on pathophysiological reg-ulation of the GH axis. On the other hand, more recently,
multiple somatostatin receptor subtypes have been clonedand their tissue expression and regulation studied. Hence,somatostatin receptor biology will be reviewed first.
A. Mechanism of somatostatin actions and its receptors
Somatostatin binds to a family of specific receptors andinhibits adenylyl cyclase via Gi, with additional actions toreduce net calcium influx. Somatostatin inhibits GH releasebut not its biosynthesis. This concept is important, since it
FIG. 5. A, Illustrative individual serum GH concentration profiles in frequently sampled intact adult female and male rats vs. animals castratedsurgically prepubertally, and rats treated beginning before puberty with a long-acting GnRH agonist (triptorelin). The six subpanels each containan approximate entropy (ApEn) value above the data for that animal’s GH profile. Higher ApEn denotes greater disorderliness, irregularity,or randomness of the GH release process. GH pulse patterns are arranged here from (quantitatively) minimally to maximally disorderly (lowestto highest ApEn values) in the following rank order: (a) intact male . (b) triptorelin-treated male (chemical castration) . (c) orchidectomizedmale . (d) ovariectomized female . (e) triptorelin-treated female . (f) intact female.
B, Bar graph of corresponding group mean (6 SEM) approximate entropy (ApEn) values with median rankings from maximally to minimallydisorderly for the GH profiles in the six groups of animals, as above. *, P , 0.05, and **, P , 0.01 for the indicated comparisons. High ApEndenotes greater irregularity of GH release. [Adapted with permission from E. Gevers et al.: Am J Physiol 274:R437–R444, 1998 (140).]
726 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

may explicate “rebound” GH secretion after somatostatinpriming and withdrawal in both the rat and human (157,160). In addition, somatostatin may play potentially (dual)inhibitory or stimulatory roles in controlling GH secretion byacting on two distinct somatotrope cell populatins at least inporcine pituitary (161).
Five somatostatin receptor subtypes have been cloned andcharacterized to date (162–165). The somatostatin receptor isregulated in a subtype- and tissue-specific manner (166, 167).Furthermore, the cloning of somatostatin receptors andthe development of somatostatin-receptor scintigraphy(168–170) hold significant clinical implications for identify-ing a variety of neuroendocrine and gastroenteropancreatictumors. The clinical tool of somatostatin receptor scintigra-phy has been reviewed in detail elsewhere (168).
Somatostatin peptide and somatostatin receptors are prob-ably important in mediating many feedback and regulatoryactions of neurohormones on the GH axis. For example,glucocorticoids likely influence both hypothalamic GHRHand somatostatin activity (171). In addition, GH autonegativefeedback at the hypothalamic level involves both the GHreceptor and hypothalamic somatostatin expression, inas-much as treatment with antisense RNA to the GH receptoramplifies GH pulsatility (i.e., by abrogating GH receptor-mediated autonegative feedback) and decreases hypotha-lamic somatostatin gene expression in the rat (130). Otherneuronal pathways may also participate in GH autonegativefeedback, such as neuropeptide Y and galanin (172, 173).Indeed, the GH receptor also is expressed in NPY neurons inthe arcuate nucleus (174). Moreover, the GH receptor gene inthe hypothalamus is modulated by sex steroid and glucocor-ticoid hormones under several conditions (175).
Somatostatin receptor subtypes are expressed in individ-ual rat pituitary cells, as defined by double-labeling in situstudies. For example, somatostatin subtypes II and V arepresent in all five major pituitary cell types in the rat (176),with the somatotropes expressing especially subtype V andto a lesser degree II, while thyrotrophs predominantly ex-press subtype IV (177). In the rat hypothalamus, somatostatinreceptor subtypes I and II may modulate GHRH and soma-tostatin release (178).
Somatostatin itself regulates somatostatin receptor sub-type expression in some pituitary cell lines, e.g., GH-3 cellswith differential up-regulation of subtype I vs. subtype II.In addition, food-deprived and diabetic rats show differ-ential pituitary and hypothalamic somatostatin subtypegene expression/regulation (179). The molecular pharma-cology of somatostatin receptors is reviewed further inRef. 180.
Somatostatin receptors also colocalize with GHRH-secret-ing neurons in the rat arcuate nucleus (181, 182). This im-portant synaptic connectivity allows for intrahypothalamicneurohormonal interactions in GH regulation (58, 183), asreviewed further below (see GHRH). Somatostatin receptorsin the locus ceruleus of male rats, seemingly paradoxically,stimulate GH secretion (184), thus illustrating the role ofcentral nervous system (CNS) topography of somatostatinreceptor distribution in mediating differential actions.
B. Human somatostatin receptor
Various somatostatin receptor subtypes are expressedwithin pituitary adenomas, e.g., subtypes II and V in acro-megalic tumors that are responsive to somatostatin (185–187). This observation has significant clinical implications,since available long-acting somatostatin analogs (e.g., oct-reotide and lanreotide) are known to interact especially withthe subtype II somatostatin receptor (187).
All five human somatostatin receptor subtypes are ex-pressed in pituitary tumors as well as in normal (fetal) pi-tuitary tissue (13, 180). In culture, the human pituitary glandpredominantly expresses subtypes II and V whether in tu-moral or normal pituitary cells (176, 188, 189). The humansomatostatin receptor subtype genes are localized on differ-ent chromosomes, e.g., 14, 17, and 22, with simple tandemrepeat DNA polymorphisms in subtypes I and II (190). Allfive cloned human somatostatin receptors subtypes are func-tionally coupled to adenylyl cyclase (191), and subtype Iadditionally stimulates inositol phosphate accumulation(192). The type V receptor also can mediate inhibition of GHsecretion from acromegalic tumor cells in vitro (189). Con-sequently, the development of highly specific somatostatinreceptor subtype agonists by structure/activity-based meth-ods, such as a lanthionine octapeptide with high affinity forthis receptor subtype, may have clinical application (193).Other studies of pituitary adenomas confirm expression ofmultiple subtypes, e.g., as assessed by PCR or other molec-ular methods (185, 194, 195).
Somatostatin itself is expressed in the brain and in theperiphery in two principal forms; namely, somatostatin-14and somatostatin-28 (196) (the latter preferentially binds tothe subtype V receptor) (168). Somatostatin influences notonly GH secretion but also that of numerous other hormones,as well as cognitive and behavioral processes, and impactsthe gastrointestinal tract, the cardiovascular system, and tu-mor growth (156, 158). Thus, somatostatin and its receptorsubtypes are widely distributed and regulated throughoutthe body, rather than solely in the hypothalamus and pitu-itary gland. Considerable complexity exists in the domain ofsomatostatin receptor subtype regulation in health and dis-ease. This should represent a valuable purview for furtherincisive clinical and basic studies.
C. GHRH
1. Isolation, actions, and neuronal distribution. Human GHRHwas isolated originally from two pancreatic tumors in pa-tients with acromegaly from the United States and France as44- and 40-amino acid forms (197, 198). These bioactive pep-tides are derived from either of two larger polypeptide pre-cursors (pre-pro GHRH 107 and 108) (199, 200). The humanGHRH gene resides on chromosome 20. The naturally oc-curring variants of GHRH (1–44 and 1–40) and the syntheticanalog 1–29 are biologically equipotent on a molar basis intheir capacity to stimulate GH release in humans (201). Thebiological half-life of GHRH 1–44 is about 3–6 min. This isbecause GHRH is rapidly inactivated by a plasma dipepti-dylaminopeptidase, producing a more stable metabolite,GHRH 3–44, which is about 1,000 times less potent than theparent compound (202).
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 727
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

Intravenous administration of GHRH in humans evokesdose-related release of GH from the pituitary gland. Ap-proximately 1.0 mg/kg is a maximally stimulating dose (203–207). GH secretion is detectable within 5 min followingGHRH injection, becomes maximal at 15–45 min, and returnsto baseline by 90–120 min (203). GH release induced byintravenous GHRH injection in adults is highly variable in-terindividually (208) and also in the same volunteer stimu-lated on different occasions (207). This may be explained inpart by the diversity of hypothalamic somatostatin and/orcosecretagogue tone that likely characterizes different indi-viduals, and also by time-variable somatostatin (or cosecre-tagogue) release and/or action in the same subject (209). Therelevance of variable hypothalamic somatostatin secretionper se to the nonuniformity of the GH response to single-bolusGHRH injection is suggested by experiments showing that afunctional somatostatin antagonist, i.e., the cholinergic agentpyridostigmine, is able to cause a significant leftward (in-creased sensitivity) and upward (increased efficacy) shift ofthe GHRH dose-response curve in young men (206) (Fig. 6).
GHRH, when given by intranasal or oral routes, is bio-logically active but much less potent than via the intravenousroute (209). GHRH has a marginal effect on the release ofother pituitary hormones in humans (210). The majority ofstudies have also failed to find any major interactions be-tween GHRH and other hypothalamic releasing factors ad-ministered simultaneously (except for the synthetic GHRPs;see below) (211).
Age is a critical factor in influencing the GH response toGHRH in humans. Neonates, children, and young adultshave similar GH responses to GHRH, whereas GH release inmost studies is reduced in older individuals (212). Cells fromthe human fetal pituitary gland become responsive to GHRHin vitro before the appearance of either GHRH-containingneurons or the hypophysial portal vasculature (213). Humanfetal pituitary responsiveness in vitro to GHRH matures fur-ther during gestation (214), which suggests that GH secretionfrom the human pituitary gland is not dependent on hypo-thalamic GHRH’s availability from weeks 7–24 of fetal life(213).
Sex steroids may also influence the GH-secretory responseto GHRH in the human. However, not all workers haveobserved consistent differences between men and women(205), in young women after ovariectomy (215), during pro-gestin-opposed estrogen replacement in the postmenopausalwoman (216), or at different stages of the menstrual cycle(210). This is discussed further in Section VII.B.2.
Prolonged continuous infusion or repeated intravenousbolus administration of GHRH leads to a modest decrease inthe subsequent GH responses to GHRH especially in the ratin vitro (217, 218) and to a lesser extent in the human in vivo(219, 220). Prolonged continuous GHRH infusion in normalhumans causes acute maximal GH release; thereafter, pul-satile GH release continues but tends to decline (albeit re-maining above basal values) by 5 h despite uninterruptedGHRH administration (220). Continued pulsatility of GHsecretion occurs despite an unvarying GHRH stimulus,which suggests intermittent somatostatin action and with-drawal in normal humans or intermittent cosecretagoguerelease (221–223).
Treatment with GHRH in the rat leads to a fall in pituitaryGH content. Concomitant administration of somatostatinwith GHRH in the human or rat largely prevents this atten-uation (human) or loss (rat) of somatotrope responsiveness,which occurs in vivo in normal human subjects (224) and invitro in rat anterior pituitary cells (225). Thus, depletion of aGHRH-sensitive (and somatostatin-antagonized) releasablepool of GH is one factor contributing to the loss of GHresponsiveness to GHRH in the rodent. Other biochemicalmechanisms are also suggested, however, since prior treat-ment with GHRH in vitro in the presence or absence ofsomatostatin causes a decrease in the cAMP response to asubsequent GHRH stimulus as well as an increase in the ED50for the stimulation of both cAMP accumulation and GHsecretion by GHRH (226). Whereas GHRH pretreatment invitro leads to a 48% decrease in the number of GHRH re-ceptors on somatotropes, maximal GH responses to GHRHcan be obtained by occupancy of only 10–20% of the totalnumber of anterior pituitary GHRH receptors. Thus, thephysiological relevance of in vitro receptor desensitization toGHRH observed in the rat is not easily translatable to in vivodown-regulation in the human.
In the human, continuous intravenous infusion of GHRHin normal subjects for 2 weeks evokes a marked increase innocturnal GH pulses and plasma IGF-I levels (227). More-over, 3 days of pulsatile GHRH administration (0.33 mg/kgevery 90 min iv) to men of varying ages and body compo-sitions augments and maintains pulsatile GH secretion andits nyctohemeral rhythm (151, 228). Lastly, GHRH-secretingtumors achieving sustained high serum GHRH concentra-tions elicit pulsatile GH release (229). These clinical dataindicate the robustness of the human (in contradistinction tothe rat) somatotrope to unvarying GHRH stimulation andalso suggest the feasibility of treating selected patients withGHRH on a continuous basis, e.g., via a repository form ofGHRH.
Immunoreactive GHRH exists in high concentrations inneurons of the median eminence and arcuate nuclei of mam-malian and nonmammalian species. GHRH is also expressed
FIG. 6. Schematic summary of proposed loci of mechanistic actions ofGHRP compounds. See text for further discussion.
728 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

in the anterior hypothalamic region as well as the dorsome-dial and ventromedial nuclei (230, 231). GHRH is colocalizedwith other neuropeptides (e.g., galanin or NPY) in the hy-pothalamus (232). Moreover, hypothalamic GHRH neuronsreceive significant afferents from somatostatin neuronal nu-clei (181, 233) that are hypothesized to be important in theGHRH-somatostatin interplay that presumptively directs ul-tradian GH pulsatility (Refs. 22 and 129–131 and Fig. 7).GHRH has also been identified both in the secretory granulesand the nuclei of somatotropes (234). Appreciable quantitiesof immunoreactive material are detected in plasma (235),duodenum (236), and placenta (237). Thus, circulatingGHRH levels are not usually helpful to the endocrinologistclinically, except in the rare condition of ectopic GHRH se-cretion with attendant acromegaly (229).
2. Mechanisms of GHRH action and GHRH receptors. GHRHstimulates both GH synthesis (by increasing the transcriptionrate of the GH gene) and GH release (238). The GHRH pep-tide binds to its specific receptors on the membranes of thesomatotropes (239). The human receptor for GHRH has beencloned and is a member of the seven-transmembrane-span-ning domain G-protein-linked receptor superfamily (240). Inthe rat, desensitization of GHRH receptors by GHRH actioncauses uncoupling of Gs activation (as well as GH depletion)(241), although down-regualtion is far less evident in thehuman (see below). Stimulation of GH release occursthrough a cAMP- and Ca21-dependent mechanism. Indeed,there may be concordance between in vivo GH release afterGHRH injection in acromegalic patients and in vitro cAMPgeneration by tumoral cells stimulated with GHRH (242).
FIG. 7. Network feedback and feedforward linkages within the basic GHRH-somatostatin/GH-IGF-I axis. Somatostatin is abbreviated here asSRIH. “Elim” denotes metabolic elimination; “F” defines selective input functions, e.g., FGHRH indicates relevant input into GHRH neurons viaSRIH, and other neuromodulators; FSRIH defines input into SRIH neurons by GHRH and other neurotransmitters; and subscripts “p” and “s”represent, respectively, particulate (tissue or secretory-granule contained) and secreted hormone or peptide. Red lines denote negative feedback(or feedforward) onto the target node marked by a red dot, whereas green lines mark a positive effector pathway terminating with a green bar.The interconnected dynamic system shown is simplified from a larger family of interrelated parameters anticipated within the full GH-IGF-Iaxis (155). Additional possible secretagogue input via a putative GHRP-like ligand family is not illustrated, although GHRP receptors (see text)are expressed in the hypothalamus and pituitary gland. No endogenous GHRP-receptor ligand(s) has (have) been isolated definitively. [Adaptedwith permission from M. Straume et al.: Methods Neurosci 28:270–310, 1995 (155).]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 729
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

GHRH also increases pituitary phosphatidylinositol turn-over. The latter is antagonized by inhibitors of the lipoxy-genase pathway, thus raising the possibility that epoxidegeneration may also participate in the stimulus-secretingcoupling process mediating GHRH’s actions on the soma-totrope (243, 244).
The human GHRH receptor was cloned from an acrome-galic pituitary cDNA library (240). The porcine and ratGHRH receptors were cloned independently (245, 246). TheGHRH receptor is homologous with that for secretin andvasoactive intestinal polypeptide. Expression of the pituitaryGHRH receptor is developmentally regulated in the rat (247).Moreover, albeit initially unexpected, both the GHRH geneand peptide are expressed in the gonad, e.g., rat ovary (248).There is tissue heterogeneity of GHRH receptor expressionin the human, e.g., kidney compared with pituitary gland(249).
GHRH receptors are critical for GH release and growth,since point mutations of the extracellular peptide-bindingdomain of the GHRH receptor in the dwarf lit/lit mouse (50)disrupt body growth and abrogate GHRH receptor function(250). GH deficiency also occurs in corresponding mutationsof the human GHRH-receptor gene (see Table 1). Moreover,in the human and the rat, overnight GH secretion as well asGH release stimulated via a number of secretagogues can beblocked by a selective GHRH antagonist (251, 252), thussupporting a role for GHRH in endogenous pulsatile GHsecretion.
3. GHRH receptor regulation. The GHRH receptor, its signaltransduction mechanisms, and mediation of GHRH-stimu-lated gene expression were reviewed recently (243, 244). Inprinciple (although not yet described), activating mutationsof this receptor might lead to constitutive cAMP overpro-duction, e.g., in patients with GH-secreting tumors and ac-romegaly. Alternatively spliced mRNA species encodingtruncated nonfunctioning GHRH receptors can be identifiedin human pituitary adenomas (253). The receptor for GHRHis regulated physiologically by glucocorticoids and estrogen(254, 255). Steroid hormone regulation of GHRH receptoractivity will thus require further study.
Lesioning of GHRH neurons or GHRH antagonist ad-ministration diminishes growth length and weight in theexperimental animal, decreases GHRH receptor number,and reduces pituitary GH content (252). The receptor isclearly distinguishable from that of GHRP (below), soma-tostatin, or pituitary adenylate cyclase-activating peptide(PACAP), etc. GHRH receptor mRNA is expressed in thepituitary gland, as well as in the periventricular, arcuate,and ventromedial nuclei of the hypothalamus, thus sug-gesting that these areas are sites of intrahypothalamicGHRH action in the rat (256). Moreover, GH autofeedbacklikely involves, in part, suppression of hypothalamicGHRH expression, since mice expressing a human GHtransgene targeted to the hypothalamus exhibit markedlyreduced expression of GHRH peptide and mRNA in thehypothalamus, of GH in the anterior pituitary gland, andof IGF-I in blood (15).
D. Feedback regulation of GH secretion
Somatostatin, GHRH, GH itself, and its nearly ubiquitoustissue mediator, IGF-I, are maintained homeostatically inhypothalamic loci, the pituitary gland, and the circulation bya complex interplay of feedback signals involving the 4(poly-) peptides themselves (see Fig. 7). So-called GHautofeedback can be demonstrated in normal subjects, sinceGH injection reduces the subsequent GH secretory responseto a GHRH stimulus (257, 258). This inhibitory effect is ob-served as early as 3 h after intraperitoneal GH administrationbefore any rise in the plasma total IGF-I concentration occurs(259, 260). GH autofeedback can be relieved by pyridostig-mine pretreatment (260), which likely acts in part to reducehypothalamic somatostatin secretion and increase GHRHrelease (261, 262). Thus, such clinical experiments supportthe view that GH autonegative feedback stimulates soma-tostatin release from the hypothalamus in vivo. In addition,a less important direct pituitary inhibitory effect and/orconcomitant GHRH withdrawal in response to a GH stim-ulus cannot be excluded. In the rat, gender differences in GHautonegative feedback exist; there is continuing responsive-ness to repeated GHRH stimuli (less sensitivity to GH’s neg-ative feedback) in the female rat, but not in the male animal(see below).
After cloning of the GH receptor gene, studies indicate thatGH feeds back to suppress the hypothalamic expression ofthe GH receptor itself (263). It is noteworthy that GH receptorantisense administration centrally [intracerebroventricularly(icv)] in the rat augments spontaneous GH pulsatility anddiminishes hypothalamic somatostatin expression. This sup-ports a primary role of the GH receptor and secondarily ofsomatostatin in mediating physiological GH autonegativefeedback (130).
In addition to the network-like feedback actions of GH onhypothalamic somatostatin, GHRH, and the GH receptor inthe rat, greater complexity arises since hypothalamic GHRHand somatostatin can each negatively regulate its own se-cretion and reciprocally control secretion of its counterpart(Figs. 1 and 7). In particular, GHRH inhibits its own releasebut increases somatostatin release in vitro (264). Conversely,somatostatin inhibits its own secretion and that of GHRH invitro (131, 265). Intrahypothalamic interactions are also in-dicated by intracerebroventricular administration of soma-tostatin or GHRH to rats, which elicits, respectively, an in-crease or a decrease in blood GH concentrations (266).Moreover, hypothalamic GHRH and somatostatin neuronalsystems are anatomically coupled (183), e.g., somatostatinneurons from the periventricular nuclei synapse on GHRHneurons in the arcuate nucleus (267). Thus, GHRH and so-matostatin may play opposing roles in the control of GHsecretion not only on the pituitary gland but also at thehypothalamic level by acting as neuromodulators.
Presumptively, functional reciprocal intrahypothalamiclinkages between GHRH and somatostatin release also makein vivo studies with GHRH or somatostatin antagonists moredifficult to interpret. For example, in the human, a predom-inant GHRH antagonist with some agonist properties caninhibit spontaneous pulsatile GH secretion (251, 268), as wellas that stimulated by various secretagogues, e.g., GHRH,
730 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

sleep, insulin, pyridostigmine, and l-arginine (269). Suchinhibition might be explicable not only if endogenous GHRHmediates the actions of these secretagogue, but also if theGHRH antagonist serves to block GHRH-stimulated soma-tostatin release.
Feedback actions of IGF-I (and IGF-II) also are inferred atthe hypothalamic-pituitary levels in the rat and human (270).Although somewhat controversial (271), a dominant feed-back action of IGF-I administered peripherally in the human,sheep, or rat probably occurs directly on the pituitary gland(62–64), whereas rapid feedback by GH itself is primarilycentral (except perhaps in the sheep) (272, 273). Evidence alsoexists for hypothalamic actions of IGF-I (271, 274, 275), andhence this issue is not fully established. In earlier studies inthe rat, partially purified preparations of IGF-Is administeredcentrally initially suggested major inhibitory effects of IGF-Iand IGF-II each at hypothalamic loci (270, 276). However,more recent experiments using recombinant human IGF-I orIGF-II indicate that either IGF-I or IGF-II acting alone is notstrongly inhibitory of the GH axis when administered cen-trally or in hypothalamic cultures in vitro (275, 277, 278).Coexposure to IGF-I and -II suppresses the GH axis. Hypo-thalamically targeted IGF-I gene and/or IGF-I receptor an-tisense mRNA expression would likely help clarify this im-portant conceptual issue.
GH autofeedback in the rat may also decrease GHRP re-ceptor expression in the arcuate and ventromedial hypotha-lamic nuclei (279). Whereas a putative GHRP-like endoge-nous ligand has not been identified, its demonstration wouldallow the conjecture that the (endogenous) GHRP-pathwayparticipates in GH autonegative feedback.
IV. Other GH-Regulating Neuropeptides
In addition to GHRH and somatostatin, other neuropep-tides can exert important modulatory effects on GH secre-tion, either by acting directly on the pituitary gland and/orby influencing GHRH and somatostatin release at the hy-pothalamic level. Major neuropeptides that affect GH releasein the human and different experimental models are re-viewed below, although not all have established physiolog-ical roles.
A. GHRPs
1. Introduction. Potent GH-releasing synthetic oligopeptides(so-called GHRPs) were developed mostly in the laboratoryof Bowers (280–282) via conformational energy calculations,solid-phase synthesis, and screening for biological activity.These compounds were first synthesized in the early 1980sas enkephalin derivatives and modified subsequently to non-peptidyl analogs (283–287). Although unproven, GHRP oli-gopeptides likely mimic endogenous effector molecules ofcurrently unknown structure and identity, since they releaseGH via specific receptors expressed in the hypothalamus andpituitary gland (below) in a dose-related and specific mannerboth in vivo and in vitro in several species (280, 288, 289).Furthermore, chronic administration of GHRP to rats, dogs,or humans can promote an increase in body weight, GHsecretion, and plasma IGF-I or BP-3 concentrations (290–
297). The mechanisms of GHRP actions are at least partiallyindependent of those of GHRH, since the combined effectsof GHRP and GHRH in vivo (but not usually in vitro) on GHrelease are typically synergistic at maximal concentrations(282, 298). GHRPs also release in vivo hypothalamic GHRHinto portal blood in the sheep (299, 300), and act directly invitro on functional GHRP receptors in human (fetal) pituitarygland (214). In the rat pituitary in vitro, GHRP but not GHRHreceptor antagonists block GHRP(-2) actions (301). Availabledata thus indicate (plausibly) multiple sites of GHRP actions(Refs. 289, and 302–304 and see below).
In one study in the rat, estrogen (or aromatizable andro-gen) augmented the maximal effect of GHRP in stimulatingGH release (305), and in older women oral estradiol treat-ment amplified the steepness of the GH secretory response(sensitivity change) to increasing doses of GHRP-2 (306),suggesting sex steroid regulation of this pathway. The max-imal GH-secretory response to GHRP (hexarelin) rises 3-foldin mid-late puberal children and falls in elderly individuals(307). In contrast, postpartum hyperprolactinemia and Cush-ing’s syndrome are associated with markedly attenuatedresponsiveness to GHRP (308).
In the human, an intact hypothalamo-pituitary unit is re-quired for maximal GHRP actions (309, 310). Propranololand clonidine (see Section V.B on catecholamines) do notgreatly modify the GH-secretory response driven by GHRP(311). Combined GHRP and GHRH administration typicallyevokes marked GH release in the human (312, 313), andGHRP plus GHRH may be a nearly age-independent test ofGH reserve (314). Most recently, novel GHRP mimetics, suchas the intranasally or orally active nonpeptide (spiropiperi-dine) analog, MK-0677 (283, 315–317), can stimulate andmaintain pulsatile GH release and increase plasma IGF-Iconcentrations during oral treatment over hours to weeks inyoung, older, and obese men and women (290), therebysuggesting possible GHRP mimetic use in GH adjuvant ther-apies (see Fig. 8). Intranasal GHRP stimulates linear growthin children (318). Indeed, both injectable and noninjectable(intranasal or oral) forms of GHRP agonists and mimeticshold promise for enhancing GH release, even in certain con-ditions of GH axis pathophysiology, e.g., obesity (296), star-vation (297), critical illness (319–321), or aging (290, 295, 322).Repeated GHRP or mimetic administration in the human isonly sparingly (or, in some cases, seemingly not at all) down-regulating (290, 293, 296, 297, 323–329), unlike a proclivity todown-regulation in the rat (330).
GHRPs stimulate ACTH and cortisol release acutely in therat and human in vivo (307, 331–333), but longer term oralanalog treatment in the human did not (290). Acute GHRP(hexarelin) administration also elicits transient ACTH secre-tion in Cushing’s disease (334, 335), but not in the ectopicACTH syndrome. In contrast, GHRH has no effect on orslightly blunts cortisol release in normal subjects (336–338).GHRP’s stimulation of ACTH and cortisol secretion is largelyindependent of CRH and arginine vasopressin actions in thehuman (339).
Further studies on the mechanisms of action of GHRPcompounds (283, 287, 324), as well as eventual isolation of theputative endogenous GHRP-like ligands, will clearly be im-portant.
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 731
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

2. GHRP receptor(s) and actions. GHRPs constitute a distin-guishable family of synthetic oligopeptides (e.g., tri-, penta-,hexa-, and heptapeptides) that act presumptively at the hy-pothalamic as well as (probably to a lesser degree) pituitarylevels to drive GH release (340–342). No natural ligand is yetknown, but the receptor family has been cloned (340, 342–347). The type 1a (but not 1b) GHRP receptor is biologicallyactive (345). Stimulation by these novel peptides does notabsolutely require endogenous GHRH activity, since theyremain active after pituitary desensitization to GHRH (348)and can stimulate GH secretion by cells not expressing theGHRH receptor, such as GH1 cells (349). GHRPs act negli-gibly on GH secretion in mice with an 80% reduction insomatotrope cell number associated with a mutated GHRHreceptor, e.g., lit/lit mouse (289). However, in this mutantanimal, the arcuate nucleus c-fos genomic response to GHRPis preserved, indicating that brainstem neuronal activationby GHRP does not require GH or, for that matter, the GHRHreceptor (350). GHRP, unlike GHRH or PACAP-38, does not
activate the pituitary-specific transcription factor, GHF-1/Pit-1, in rat anterior pituitary cells in vitro (351), further dis-tinguishing its biochemical actions from those of GHRH.
The cloned receptor families for GHRP and GHRH aredistinct (352). Purification and later cloning of the GHRPreceptor in pig and rat anterior pituitary membranes re-vealed a magnesium-dependent, GTP-binding, non-GHRH,non-somatostatin receptor that is G-protein coupled and ap-parently mediates the action of multiple GHRPs, e.g.,GHRP-6, hexarelin, and the nonpeptidyl (spiropiperidine)GHRP analog, MK-0677 (340, 342, 343, 353–356). GHRP re-ceptors estimated by binding and functional reactivity havehigh affinity [e.g., dissociation constant (Kd) 5 0.7 nm forMK-0677 (356)], and are expressed in the normal pituitarygland, pituitary tumors, the hippocampus, and ventromedialand arcuate nuclei in the hypothalamus (214, 340, 342, 344,345, 345). GH autonegative feedback reduces (and GH de-ficiency increases) hypothalamic (ventromedial and arcuatenuclear) as well as hippocampal GHRP receptor expression,thus suggesting multisite feedback control mechanisms inregulating the putative endogenous GHRP receptor system(279).
GHRP’s intracellular signaling mechanisms in the rat in-clude the phospholipase C-phosphoinositide pathway (357–359). GHRP also stimulates delayed calcium influx in rat andsheep somatotropes (360, 361), activates protein kinase C(357, 362), heightens GHRH signaling (the latter via cAMP)(302), and depolarizes somatotrope cells (343, 363). Themechanisms of GHRP action are distinctly nonopiate andnon-GHRH receptor dependent (280, 298, 302, 352, 364, 365).However, effects of GHRP are partially antagonized by so-matostatin in the rat and human (339, 366–369), by centrally(icv) infused somatostatin in the rat (370), and (partially) bya GHRH receptor antagonist in the rat and human (371), butnot by an opiate receptor antagonist in the human (372).GHRP peptides are active in vitro and in vivo in multiplespecies, such as the rat, monkey, human, sheep, cow, andchicken (214, 280, 298, 373–379).
Acute intravenous injection of GHRP-6, GHRP-2, hexare-lin, or a nonpeptidyl GHRP mimetic (L-692, 429) rapidlyincreases serum GH concentrations in humans within 5–15min, with the peak GH concentration usually observed 15–30min after infusion (298, 313, 332, 366, 380, 381). The amountof GH released after GHRP is much larger than that secretedafter GHRH (313, 332). Maximally effective doses of GHRP-6and GHRH, when injected simultaneously in man, typicallystimulate GH secretion additively or synergistically (298,382). Both GHRH and GHRPs administered nocturnally in-dividually facilitate slow-wave sleep (332, 336, 383–385) viamechanisms that are not yet established (289, 336). The effectsof GHRH on sleep decline with age (337, 386, 387) and areespecially evident after pulsatile infusion (338, 388). Similardata are not yet available for possible age-related actions ofGHRP on sheep. Daytime GHRH treatment does not modifynighttime sleep or GH release (389), but sleep deprivationmay limit GHRH-stimulated nocturnal GH release (390).This may reflect the increase in somatostatin evoked by sleepdeprivation, at least in the rat (391). Analogous data are notyet available regarding GHRP actions or receptors in sleep-deprived animals.
FIG. 8. The orally active nonpeptidyl (L163, 191, a spiropiperidone)GHRP-receptor agonist, MK0677, administered once daily to men andwomen stimulates pulsatile GH secretion and increases plasma IGF-Iconcentrations over 2 to 4 weeks. Data are percentage changes frombaseline (pretreatment geometric mean 6 SEM). Results are from 24-hserum GH concentration pulse profiles and morning serum IGF-Imeasures, collected before and after 2 or 4 weeks of nighttime treat-ment with MK0677 once daily orally at a dose of zero (placebo), 2, 10,or 25 mg (n 5 10–12 volunteers per group). *, P , 0.05 vs. baseline.[Adapted with permission from I. M. Chapman et al.: J Clin Endo-crinol Metab 81:4249–4257, 1996 (290). © The Endocrine Society.]
732 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

Prolonged infusion of GHRP amplifies pulsatile GH se-cretion over 24 h in men by increasing GH pulse amplitude(323, 392). Partial attenuation of GH release in response to asubsequent GHRP bolus occurs, and is not caused by abso-lute depletion of pituitary GH stores, since the effect ofGHRH persists at this time. Conversely, desensitization toGHRH does not eliminate responsiveness to GHRP, furtherindicating the capacity for independent actions of these pep-tide secretagogues (348).
The synergy between GHRH and GHRP-6 is not enhancedby pyridostigmine, suggesting that somatostatin withdrawalmay participate in their synergy (393), or that pyridostigminealso acts via stimulating GHRH release, as suggested in thesheep (299). Similarly, hexarelin and GHRH do not synergizein another human model (type I diabetes), in which soma-tostatin tone is putatively reduced (394). In combination withGHRH or l-arginine, GHRP evokes substantial GH release inaging animals and humans (314, 379), and combined GHRPand GHRH stimulation will elicit marked GH secretion evenin obese adults (312). Thus, GHRPs constitute among themost effective clinical GH secretagogues known, whetheradministered alone or in combination with physiological orother pharmacological stimuli of GH release.
The effects of GHRP, albeit demonstrably distinct fromthose of GHRH (374), are also influenced by GH autonegativefeedback [see Section III.D]. For reasons not yet established,GHRP’s action may be affected to a lesser degree than thatof GHRH by GH feedback inhibition (395, 396). In the rat, GHautofeedback reduces arcuate and ventromedial nuclearGHRP receptor expression (279), thus offering a speculativelinkage between this (putative) endogenous effector path-way and GH autofeedback.
3. Mechanisms of neuroregulation by GHRPs. Several plausiblemechanisms of GHRP action have been considered by thelaboratory of Bowers and others, including, for example 1) adirect stimulatory action on the anterior pituitary gland,although presumptively relatively minor (281, 340, 349, 353,397); 2) indirect actions on the hypothalamus causing releaseof GHRH (and, less plausibly, a decrease in central soma-tostatin activity), the former being relevant in the sheep andadult rat (299, 300, 340, 398), and possibly human (291, 303);and 3) possible release of an unknown (“U”) hypothalamicfactor, which in both the human and rat is postulated to elicitGH release by acting synergistically with GHRH (302, 399).GHRP may also reduce somatostatin’s feedback inhibition ofGHRH neurons and thus attenuate GH autofeedback (400).Figure 6 summarizes these plausible sites of GHRP actions.We caution that what has been inferred from intravenous oreven icv injections of GHRPs may not fully predict the ul-timately identified (if any) activities of endogenous GHRPreceptor ligands. Natural effectors might act inside or outsidethe blood-brain barrier at very high local concentrations toexert important (new) autocrine and/or paracrine effects inthe hypothalamus, pituitary gland, and/or other sites ofGHRP reception.
In three of four studies in the adult animal, and in twostudies in rat pups, GHRP-6 stimulated GH secretion in vivoeven in the presence of antiserum to GHRH, or somatostatin,or both (350, 398, 400–403). GHRP-6 or mimetics likewise can
enhance pituitary GH gene expression, apparently indepen-dently of GHRH and/or somatostatin, and directly stimulateGH release in vitro from incubated pituitary glands and cul-tured rat anterior pituitary cells (284, 301, 341, 350, 352, 357,358, 367, 404–407). However, the in vitro stimulatory effectof GHRP on either rat or human dispersed GH-secretingpituitary cells is often somewhat diminutive compared withthat of GHRH, except in a recent study of GH-secretingtumors (367). GHRP action in vitro also is typically much lessthan that observed after in vivo GHRP administration (214,349). Of note, early studies utilizing in vitro incubations ofintact pituitary glands from immature rats showed larger (6-to 10-fold) stimulatory effects of GHRPs on GH release thanwere later observed in dispersed (monolayer) pituitary cellculture studies derived from mature animals (1.5- to 4-foldeffects). At the single-cell level, GHRP-6 acts on both GHRH-responsive and nonresponsive somatotroph subpopulationsby increasing the percentage of GH-secreting cells as well asthe amount of GH secreted per cell (404). The foregoingexperiments collectively suggest that, at least in the (adult)rodent, enhancement of GHRH, as well as, conversely, in-hibition of (central) somatostatin, action play modulatoryroles in the GH-stimulating properties of GHRPs. However,these roles are only partial, since GHRPs effectively stimulateGH secretion directly in vitro (above), and in three of fourstudies somatostatin antiserum pretreatment actually aug-mented the GH response to GHRP in both immature femaleand adult male rats (400–402). Augmentation of the actionof GHRPs by experimental neutralization of endogenoussomatostatin supports strongly non-somatostatin-depen-dent actions of GHRPs, which are otherwise impeded par-tially by (endogenous) somatostatin. Indeed, in the malerodent, icv somatostatin administration limits GHRP actions(370).
The actions of GHRP on the CNS are quite likely to berelevant to its overall effects (408). Brain actions are sug-gested by the presence of the GHRP receptor in the arcuateand ventromedial nuclei of the hypothalamus and in thehippocampus, and by the ability of GHRP to induce GHRHmRNA expression in the arcuate nucleus even in GH- (andGHRH receptor)-deficient mice (279, 289, 409). Other CNSactions of GHRPs are also apparent in the rat, since GHRPstimulates eating behavior after icv infusion without alteringGH release (410), increases electrophysiological activity inthe arcuate nucleus, and induces brainstem c-fos mRNA ex-pression in NPY and GHRH-releasing neurons (411–413).These effects are not mimicked by GHRH (408). In the con-scious sheep, GHRP acutely releases GH in association withan increased frequency of pulsatile GHRH release (with noapparent changes in somatostatin secretion) into the hypo-physial portal circulation (299, 300). In the monkey, indirectstudies suggest that GHRH’s involvement may be less con-spicuous (374), whereas in the human recent [but not earlier(414)] studies with a GHRH receptor antagonist indicate amajor (;85%) dependence of GHRP-6’s stimulation of GHsecretion on endogenous GHRH actions (415).
In the rat, sex hormones can increase the effect of GHRPin both males and females (305). In the human, the (single-dose) maximal GHRP-6 effect in early studies was relativelyindependent of age, gender, or menstrual cycle stage (416).
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 733
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

However, dose-responsive sensitivity of GH release toGHRP has not been appraised in detail as a function of thesex-steroid milieu in the human. With respect to aging whensex steroids decline, GHRP augments the effect of GHRH inboth young and old dogs and potentiates the effects ofclonidine, an a-2 agonist, at least in young dogs (417).
In the human, the potential relative involvement of thehypothalamus vis-a-vis the pituitary gland in the GH-releas-ing action of GHRP-6 has been assessed using differentpathophysiological and pharmacological strategies (418). Di-rect pituitary effects have been evaluated by injecting GHRPin patients with pituitary stalk section (309, 419) or acro-megaly (367) and by in vitro studies (214). Both of the fore-going categories of patients are presumed to have a pituitarygland that is partially or completely disconnected from thehypothalamus, and both show significant, albeit variable,attenuation of GHRP effects. In acromegaly, “functional”disconnection likely reflects tumoral autonomy. The poten-tial role of somatostatin has been appraised in chronicallyglucocorticoid-treated men (382), who are considered to havehigh somatostatin tone (171) and, conversely, in type 1 di-abetic patients, with putatively low somatostatin tone (420).These clinical paradigms indicate that GHRP-6 can increasethe GH response to GHRH in glucocorticoid-treated patients(382), but to a lesser degree in type 1 diabetics (394). Suchobservations suggest indirectly that inhibition of endoge-nous somatostatin tone may play only a subordinate role inGHRP’s stimulatory actions in humans. Conversely, the in-volvement of endogenous GHRH in GHRP’s actions in hu-mans was restudied recently via concurrent GHRH antag-onist administration, which significantly (;85%) attenuatedthe GH-secretory response to GHRP (294). Thus, in the hu-man, available evidence supports at least the importance of(hypothalamic) GHRH release in GHRP’s effects (294). Themarked enhancement of GH secretion by combined GHRHand GHRP administration has led to the suggestion thatthese joint stimuli afford near-maximal stimulation of soma-totrophs and, when used together, offer a plausible test forGH deficiency in obesity, children, and older adults (295, 314,350, 421–425). These suggestions will require further vali-dation in larger diverse normal and hypopituitary popula-tions.
B. Galanin
Galanin is a 29-amino acid peptide initially isolated fromporcine small intestine (426). Galanin-like immunoreactivityis widely distributed in the central and peripheral nervoussystem (427). The hypothalamus is particularly rich in cellbodies and fibers containing galanin-like immunoreactivity,with the highest concentration in the median eminence (427).Specific binding sites for galanin are also demonstrable in themediobasal hypothalamus (428). Taken together, these ob-servations suggest that galanin may have a role in regulatinganterior pituitary function.
The molecular sequences of porcine, rat and, recently,human galanin have been deduced (429). The three peptideforms are identical with respect to their first 15 residues, butdiffer at several positions in the C-terminal part (430). Vir-tually all human experiments have been conducted with
porcine galanin, with fewer studies performed using rat (430,431) or human (432) galanin. Rat galanin, the structure ofwhich is more similar than porcine galanin to that of humanpeptide, is able to cause significantly greater GH release thanporcine galanin in normal humans (430). Therefore, possiblespecies-dependent differences in galanin action should beconsidered in the interpretation of experiments concerningthe physiological role of galanin in man (433).
In the rat, galanin increases GH release when given icv(434–436), subcutaneously (437), or intravenously (434, 436).Although galanin’s interaction with GHRH is not fully de-fined, GHRH-overexpressing transgenic mice show 7-foldand 4-fold increases, respectively, in galanin peptide andmRNA expression in the pituitary gland (438).
In the human, both porcine and human galanin elicit GHsecretion when given alone (432, 439) and facilitate the GH-secretory response to GH-releasing hormone (GHRH) in nor-mal young men (434, 440, 441) (Fig. 9). Although the mech-anism underlying this action of galanin is unknown,evidence in the rat suggests that galanin may act at thehypothalamic level, as the peptide was effective when in-jected into the third ventricle, but does not stimulate pituitarycells in vitro (435, 436). An interaction between galanin andGHRH has been proposed, since the treatment of male ratswith GHRH antiserum markedly inhibits the GH response toeither intravenous or central galanin administration (434).The demonstration of coexistence of galanin and GHRH inthe same neurons in the arcuate nucleus (232) further sug-gests possible interactions between the two neuropeptides.Other authors have hypothesized that galanin may also in-hibit somatostatin release from the hypothalamus (442).However, porcine galanin fails to restore normal GH secre-tion in various pathophysiological conditions thought to becharacterized by increased somatostatin tone in man (441,443, 444). Galanin infusion stimulates GH secretion more inwomen than in men, and peak serum GH responses in thefemale are proportional to the blood estradiol concentration(445), consistent with a hypothesis of gender and sex hor-mone modulation of galanin action. In this regard, a func-tional estrogen-response element has recently been identi-fied within the human galanin gene promoter (446).
GHRPs, discussed above, are synthetic oligopeptides thatpotently stimulate GH secretion in man, apparently acting inlarge part at the hypothalamic level via unknown mecha-nisms (280). Galanin does not enhance the maximal GH re-sponse to hexarelin, a GHRP-6 analog, either in humans orin normal rat somatotropes (A. Giustina, unpublished). Thus,one could speculate that galanin may act through or interfacedistally with the putative endogenous GHRP effector path-way, such that when the latter is activated maximally galanincan exert no further effect to stimulate GH secretion. Al-though not mechanistically explained, galanin has also par-adoxically inhibited GH secretion in acromegalic patientsbearing GH-secretory adenomas (447).
Immunoneutralization of endogenous galanin within theCNS significantly disrupts the normal GH-secretory patternin male rats (448). These results suggest an important phys-iological role for endogenous galanin in the control of spon-taneous pulsatile GH secretion in this species and gender.The major impairment in GH secretion induced by galanin-
734 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
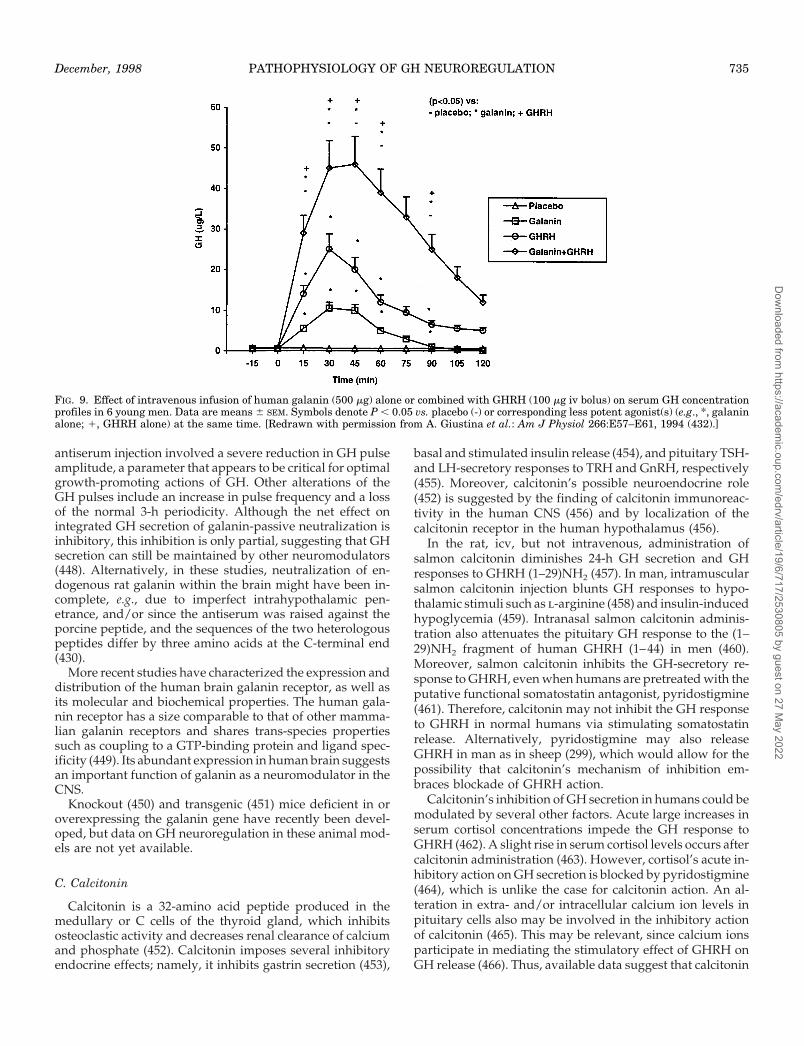
antiserum injection involved a severe reduction in GH pulseamplitude, a parameter that appears to be critical for optimalgrowth-promoting actions of GH. Other alterations of theGH pulses include an increase in pulse frequency and a lossof the normal 3-h periodicity. Although the net effect onintegrated GH secretion of galanin-passive neutralization isinhibitory, this inhibition is only partial, suggesting that GHsecretion can still be maintained by other neuromodulators(448). Alternatively, in these studies, neutralization of en-dogenous rat galanin within the brain might have been in-complete, e.g., due to imperfect intrahypothalamic pen-etrance, and/or since the antiserum was raised against theporcine peptide, and the sequences of the two heterologouspeptides differ by three amino acids at the C-terminal end(430).
More recent studies have characterized the expression anddistribution of the human brain galanin receptor, as well asits molecular and biochemical properties. The human gala-nin receptor has a size comparable to that of other mamma-lian galanin receptors and shares trans-species propertiessuch as coupling to a GTP-binding protein and ligand spec-ificity (449). Its abundant expression in human brain suggestsan important function of galanin as a neuromodulator in theCNS.
Knockout (450) and transgenic (451) mice deficient in oroverexpressing the galanin gene have recently been devel-oped, but data on GH neuroregulation in these animal mod-els are not yet available.
C. Calcitonin
Calcitonin is a 32-amino acid peptide produced in themedullary or C cells of the thyroid gland, which inhibitsosteoclastic activity and decreases renal clearance of calciumand phosphate (452). Calcitonin imposes several inhibitoryendocrine effects; namely, it inhibits gastrin secretion (453),
basal and stimulated insulin release (454), and pituitary TSH-and LH-secretory responses to TRH and GnRH, respectively(455). Moreover, calcitonin’s possible neuroendocrine role(452) is suggested by the finding of calcitonin immunoreac-tivity in the human CNS (456) and by localization of thecalcitonin receptor in the human hypothalamus (456).
In the rat, icv, but not intravenous, administration ofsalmon calcitonin diminishes 24-h GH secretion and GHresponses to GHRH (1–29)NH2 (457). In man, intramuscularsalmon calcitonin injection blunts GH responses to hypo-thalamic stimuli such as l-arginine (458) and insulin-inducedhypoglycemia (459). Intranasal salmon calcitonin adminis-tration also attenuates the pituitary GH response to the (1–29)NH2 fragment of human GHRH (1–44) in men (460).Moreover, salmon calcitonin inhibits the GH-secretory re-sponse to GHRH, even when humans are pretreated with theputative functional somatostatin antagonist, pyridostigmine(461). Therefore, calcitonin may not inhibit the GH responseto GHRH in normal humans via stimulating somatostatinrelease. Alternatively, pyridostigmine may also releaseGHRH in man as in sheep (299), which would allow for thepossibility that calcitonin’s mechanism of inhibition em-braces blockade of GHRH action.
Calcitonin’s inhibition of GH secretion in humans could bemodulated by several other factors. Acute large increases inserum cortisol concentrations impede the GH response toGHRH (462). A slight rise in serum cortisol levels occurs aftercalcitonin administration (463). However, cortisol’s acute in-hibitory action on GH secretion is blocked by pyridostigmine(464), which is unlike the case for calcitonin action. An al-teration in extra- and/or intracellular calcium ion levels inpituitary cells also may be involved in the inhibitory actionof calcitonin (465). This may be relevant, since calcium ionsparticipate in mediating the stimulatory effect of GHRH onGH release (466). Thus, available data suggest that calcitonin
FIG. 9. Effect of intravenous infusion of human galanin (500 mg) alone or combined with GHRH (100 mg iv bolus) on serum GH concentrationprofiles in 6 young men. Data are means 6 SEM. Symbols denote P , 0.05 vs. placebo (-) or corresponding less potent agonist(s) (e.g., *, galaninalone; 1, GHRH alone) at the same time. [Redrawn with permission from A. Giustina et al.: Am J Physiol 266:E57–E61, 1994 (432).]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 735
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

inhibits GHRH actions but not via somatostatin. Interactionsbetween calcitonin and GHRP have not yet, to our knowl-edge, been evaluated.
D. PACAP
Pituitary adenylate cyclase-activating polypeptide con-sists of 38 amino acids that are C-terminally amidated(PACAP 38). This peptide was first isolated from ovine hy-pothalamus, based on to its ability to enhance cAMP accu-mulation in anterior pituitary cells (467). PACAP-like im-munoreactivity is widely distributed in the CNS and highlyconcentrated in the hypothalamus (468). Histochemical stud-ies have revealed dense PACAP-positive fibers in the inter-nal and external layers of the median eminence and in thepituitary stalk (469). The cell bodies are found in the mag-nocellular region of the paraventricular nuclei and the su-prachiasmatic nuclei (470). Dow et al. (471) reported thatPACAP-like immunoreactivity is detectable in the hypoph-ysial-portal plasma of pentobarbital-anesthesized rats in con-centrations of 50–100 pm, which are significantly higher thanthose in the peripheral plasma. These findings suggest thathypothalamic PACAPs, mainly PACAP-38, can be releasedinto the hypophysial portal vessels and reach the anteriorpituitary, where they might play a physiological agonisticrole.
Other indirect evidence further suggests that PACAPplays a role in promoting anterior pituitary GH secretion.PACAP stimulates GH release from the rat pituitary glandboth in vivo and in vitro (472–474). The variable increase inGH release in vitro after stimulation with PACAP-38 prob-ably reflects nonuniform experimental procedures (475). Thepeptide also stimulates pituitary hormone release from ro-dent clonal pituitary cell lines such as ATT-20 and GH3 cells(474). Hormone release from tumoral GH3 cells is stimulatedby PACAP through the type II PACAP receptor (476), whichis common to PACAP and vasoactive intestinal polypeptide(VIP). In contrast, GH release from normal rat somatotropesis stimulated by PACAP, but not by VIP, suggesting that theeffect is mediated by a PACAP-specific (type I) receptor(477). The action of PACAP on rat somatotropes is also dis-tinct from that of GHRH (473). Thus, a synthetic GHRHantagonist fails to impede PACAP-induced GH release inpituitary cell culture and perifusion experiments.
PACAP-38 is a potent secretagogue of GH in consciousmale rats in vivo. In mechanistic studies, Jarry et al. (472)found that PACAP-38 increases plasma GH in hypothala-mus-lesioned anesthetized rats. However, the pulsatile pat-tern of GH release was not affected by a PACAP antagonist(475). If confirmed, the latter observation (assuming ade-quate effective antagonist action at relevant sites) wouldspeak against a primary physiological role of endogenousPACAP in stimulating spontaneous pulsatile GH secretion inthe rat. The role of PACAP in the regulation of GH secretionin man also remains to be elucidated.
E. Opioid peptides
Opioid peptides stimulate GH secretion in both rodentsand humans (478, 479). Opiates can directly inhibit rat me-
diobasal hypothalamic release of somatostatin in vitro (480,481). Opiate effects also can be mediated via GHRH, sincerats pretreated with GHRH antibodies fail to show the ex-pected opioid-mediated rise in GH levels (479). While thereis no clinical evidence at present to suggest that endogenousopiates, e.g., acting through naloxone-sensitive receptors,play a major role in the control of spontaneous basal GHrelease, they may participate in some forms of stress-stim-ulated GH secretion, such as marathon running and calorierestriction (482). In healthy individuals, naloxone and nal-trexone (opiate receptor antagonists) exert few acute effectson GH secretion (482).
F. TRH
In the rat, TRH act as a physiological GH secretagogue(483). In contrast, in the human, TRH stimulates GH secretiononly in certain experimental and pathophysiological condi-tions, such as acromegaly (484), type 1 diabetes mellitus(485), and hepatic and renal failure (486, 487), but not usuallyin normal unmedicated subjects. Conversely, GH responsesto l-dopa, arginine, and insulin-induced hypoglycemia(488), but not to GHRH (489), are reduced or blocked duringTRH administration. In normal subjects these divergent ef-fects of TRH are probably due to two different loci of actionsof TRH. TRH can directly elicit GH secretion from pituitarycells of hypothyroid rats in vitro (490) and can act via specific,high-affinity, low-capacity TRH receptors on GH3 and GH1cells (491, 492). TRH receptors are thyroid hormone inhibitedand consequently increase (reciprocally) in hypothyroidism.This might explain the paradoxical GH response to TRH inhypothyroidism (493), despite the well described reductionin GH responses to other secretagogues including GHRH inthis condition (494). Conversely, the inhibitory actions ofTRH on secretagogue-stimulated GH release are likely ex-erted at the hypothalamic level via enhanced somatostatinrelease.
TRH paradoxically stimulates GH secretion in type 1 di-abetes (484), a condition characterized by low somatostatintone (see below), when the direct stimulating effect of TRHat the pituitary level may be unmasked. Women receiving 30mg ethinyl estradiol and a synthetic progestin daily for 6–24months (495) also show paradoxical GH release after TRHinjection, but the mechanism of this acquired responsivenessto TRH is unknown. In another study in men and midlutealphase women, histamine pretreatment unmasked TRH/GnRH-stimulated GH release (496). The mechanism under-lying this phenomenon has also not been explained. Overall,in harmonizing the discrepant available data, one can hy-pothesize that in the human a direct stimulatory effect ofTRH that is expected (based on rat data) at the pituitary levelcan be overcome by TRH-mediated somatostatin release (thelatter likely does not occur in the rat).
G. NPY
Studies a decade ago indicated that the orexigenic peptide,NPY, can inhibit GH secretion in the male and female rat(497–499). In particular, in the intact male rat, as well as inthe ovariectomized female rat (with or without sex steroid
736 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

hormone replacement), icv infusion of NPY reduces GH se-cretion, and antibody to NPY increases GH release in theseanimals. The suppressive action of NPY appears to be me-diated by both NPY-1 and NPY-2 receptor subtypes ex-pressed in the mediobasal hypothalamus (500). Most plau-sibly, NPY stimulates hypothalamic somatostatin release andthereby inhibits GH secretion, at least in the male rat (498).In addition, NPY may act, in part, directly on the anteriorpituitary gland, since NPY will reduce human somatotrophtumor secretion of GH in vitro (501) and can limit soma-totroph cell proliferation in the rat, possibly by way of go-nadotroph-dependent paracrine mechanisms (502). In con-trast to studies in the rodent, in a single clinical study,infusion of NPY paradoxically evoked GH secretion in 60%of a small group (n 5 15) of patients with prolactinomas(503). NPY also stimulated GH secretion directly from gold-fish pituitary in vitro (504). Thus, the preponderance of ev-idence favors an inhibitory role of NPY on the GH axis in therat, with possible species differences, e.g., stimulation of GHsecretion in the human.
NPY may participate in the negative-feedback actions ofGH on its own secretion in the rat (172, 502). For example, GHreceptors are expressed on NPY neurons in the arcuate nu-cleus (174). GH treatment increases c-fos gene expression inNPY neurons (505) and stimulates hypothalamic NPYmRNA expression in hypophysectomized rats (506). Suchobservations sugggest the thesis that NPY may enlist soma-tostatin release during GH’s negative feedback and/or limitGHRH release in this autofeedback context. Further defini-tive studies with NPY neutralization (e.g., both immunolog-ical and via antisense molecular methods, etc.) will be re-quired to establish or refute this hypothesis definitively.
The activation of GH secretion in the sheep, and inhibitionof GH secretion in the rat, by nutrient restriction might alsobe mediated in part via NPY. In the ovariectomized ewe,NPY immunoreactivity is increased in the arcuate nucleus byprolonged food withdrawal and accompanying weight loss,which resultant selective amplification of GH secretion and,in this particular paradigm, preservation of secretory activityof the reproductive axis (507). Akin to fasting, neuronal glu-coprivation via a metabolic inhibitor of intracellular glucoseutilization increases c-fos gene expression in NPY neurons inthe rat (508). Such studies suggest the hypothesis that NPYparticipates in mediating the impact of dietary manipulationon both the GH and reproductive axes, both of which aresuppressed in the rat by fasting, as reviewed elsewhere (509).However, considerable additional study will be required topinpoint the species specificity and neurohormonal mecha-nisms of NPY’s actions in various nutritionally modifiedcontexts, e.g., how fasting suppresses both LH and GH in therat, but stimulates GH while suppressing LH in the humanand sheep. How NPY might integrate such responses indifferent species is not yet evident.
As noted in Section IV.A, NPY may also participate in theactions of non-GHRH secretagogues. For example, GHRP-6in the rat increases c-fos expression in 51% of NPY neuronsand 23% of GHRH neurons (413). Additional studies will berequired to assess the exact role of NPY in modulatingGHRP‘s actions, and to define whether NPY neurons them-selves express the receptor for GHRP.
In summary, NPY inhibits GH secretion in the male andfemale rat, possibly by stimulating hypothalamic somatosta-tin release, and likely participates in GH autonegative feed-back in this species. In addition, in rodents and ruminants,NPY may have a role in mediating nutritional effects on theGH and/or reproductive axes. The ability of the nutritionalregulatory peptide signal, leptin, to stimulate NPY neuronsin the rat is consistent with this theme, although the acqui-sition of more detailed knowledge of interactions amongleptin, GH, and NPY will be essential (see Section VI.B be-low).
H. Substance P
In male rats, a high percentage of somatotropes colocalizeimmunoreactive substance P and GH. Coexpression of GHand substance P falls in estrogen-treated ovariectomized (fe-male) rats (510). Additional studies are required to clarify therole, if any, of substance P in neuroregulation of the humanGH axis, although this agent will enhance basal and GHRH-stimulated GH release in normal men (511).
I. Bombesin
In the male rat, bombesin inhibits GH secretion, appar-ently (based on antibody studies) independently of soma-tostatin (512). In the female rat, bombesin stimulates GHrelease via mechanisms that are partially antagonized byGHRH antiserum (513). Limited clinical data are availableindicating this peptide’s absence of an effect on basal GHrelease and inhibitory impact on the hypoglycemia-stimu-lated human GH axis in young male volunteers (514, 515).Neuromedin C is a bombesin-like peptide that stimulates GHrelease from perifused (rat) pituitary cells (513). Little, ifanything, is known about neuromedin C’s actions on thesomatotropic axis in the human.
J. Melatonin
The pineal gland, via melatonin, may also modulate GHsecretion. Oral administration of melatonin to normal sub-jects increases basal GH levels and the GH response toGHRH (516), but marginally affects GH responses to hypo-glycemia or apomorphine (517, 518). This suggests that mel-atonin might play a minimal (stimulatory) role in baselineGH secretion, possibly acting at the hypothalamic level viainhibition of somatostatin. The GH response to l-dopa isreduced in blind human subjects (519), who presumptivelylack both light-mediated inhibition of melatonin release andthe normal slow wave sleep-associated rise in plasma GHconcentrations (520). Thus, it is possible, but entirely un-proven, that variations in the release of endogenous mela-tonin could modulate GH secretion in humans.
K. Other GH secretagogues
The cytokine, interleukin-1, stimulates pituitary GH re-lease at the single-cell level by increasing both the number ofsomatotropes secreting GH and the amount of GH secretedper cell (521).
In the fasted male rat, leptin infusion increases GH secre-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 737
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
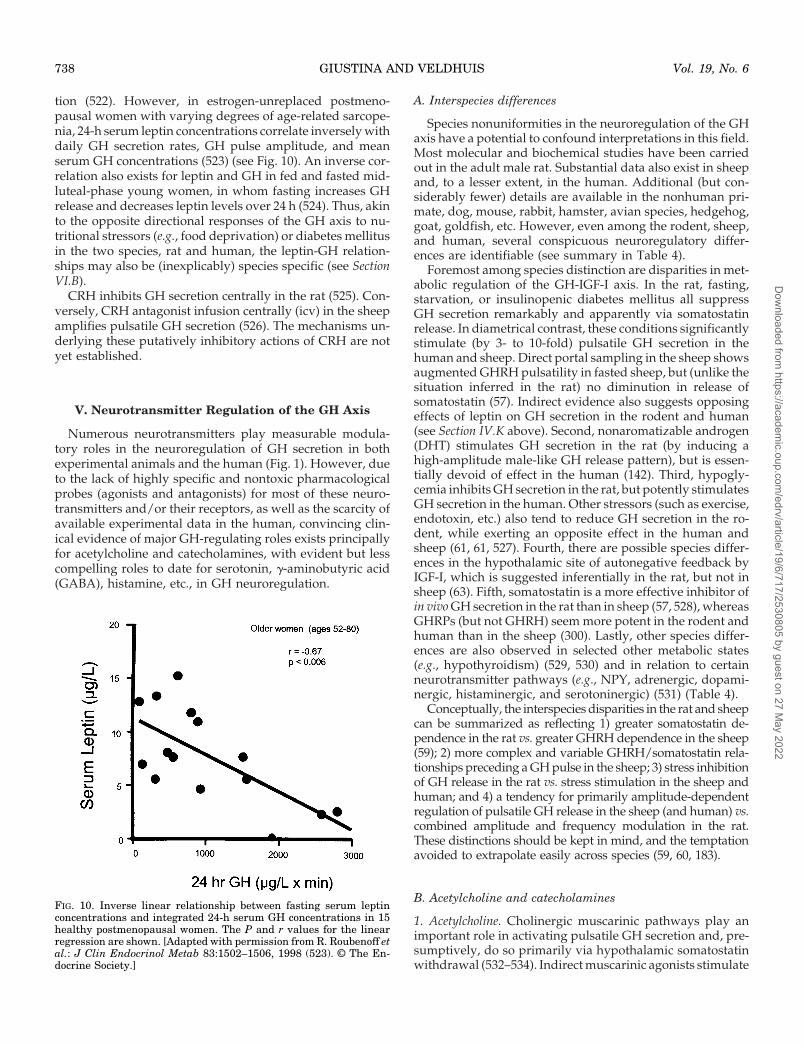
tion (522). However, in estrogen-unreplaced postmeno-pausal women with varying degrees of age-related sarcope-nia, 24-h serum leptin concentrations correlate inversely withdaily GH secretion rates, GH pulse amplitude, and meanserum GH concentrations (523) (see Fig. 10). An inverse cor-relation also exists for leptin and GH in fed and fasted mid-luteal-phase young women, in whom fasting increases GHrelease and decreases leptin levels over 24 h (524). Thus, akinto the opposite directional responses of the GH axis to nu-tritional stressors (e.g., food deprivation) or diabetes mellitusin the two species, rat and human, the leptin-GH relation-ships may also be (inexplicably) species specific (see SectionVI.B).
CRH inhibits GH secretion centrally in the rat (525). Con-versely, CRH antagonist infusion centrally (icv) in the sheepamplifies pulsatile GH secretion (526). The mechanisms un-derlying these putatively inhibitory actions of CRH are notyet established.
V. Neurotransmitter Regulation of the GH Axis
Numerous neurotransmitters play measurable modula-tory roles in the neuroregulation of GH secretion in bothexperimental animals and the human (Fig. 1). However, dueto the lack of highly specific and nontoxic pharmacologicalprobes (agonists and antagonists) for most of these neuro-transmitters and/or their receptors, as well as the scarcity ofavailable experimental data in the human, convincing clin-ical evidence of major GH-regulating roles exists principallyfor acetylcholine and catecholamines, with evident but lesscompelling roles to date for serotonin, g-aminobutyric acid(GABA), histamine, etc., in GH neuroregulation.
A. Interspecies differences
Species nonuniformities in the neuroregulation of the GHaxis have a potential to confound interpretations in this field.Most molecular and biochemical studies have been carriedout in the adult male rat. Substantial data also exist in sheepand, to a lesser extent, in the human. Additional (but con-siderably fewer) details are available in the nonhuman pri-mate, dog, mouse, rabbit, hamster, avian species, hedgehog,goat, goldfish, etc. However, even among the rodent, sheep,and human, several conspicuous neuroregulatory differ-ences are identifiable (see summary in Table 4).
Foremost among species distinction are disparities in met-abolic regulation of the GH-IGF-I axis. In the rat, fasting,starvation, or insulinopenic diabetes mellitus all suppressGH secretion remarkably and apparently via somatostatinrelease. In diametrical contrast, these conditions significantlystimulate (by 3- to 10-fold) pulsatile GH secretion in thehuman and sheep. Direct portal sampling in the sheep showsaugmented GHRH pulsatility in fasted sheep, but (unlike thesituation inferred in the rat) no diminution in release ofsomatostatin (57). Indirect evidence also suggests opposingeffects of leptin on GH secretion in the rodent and human(see Section IV.K above). Second, nonaromatizable androgen(DHT) stimulates GH secretion in the rat (by inducing ahigh-amplitude male-like GH release pattern), but is essen-tially devoid of effect in the human (142). Third, hypogly-cemia inhibits GH secretion in the rat, but potently stimulatesGH secretion in the human. Other stressors (such as exercise,endotoxin, etc.) also tend to reduce GH secretion in the ro-dent, while exerting an opposite effect in the human andsheep (61, 61, 527). Fourth, there are possible species differ-ences in the hypothalamic site of autonegative feedback byIGF-I, which is suggested inferentially in the rat, but not insheep (63). Fifth, somatostatin is a more effective inhibitor ofin vivo GH secretion in the rat than in sheep (57, 528), whereasGHRPs (but not GHRH) seem more potent in the rodent andhuman than in the sheep (300). Lastly, other species differ-ences are also observed in selected other metabolic states(e.g., hypothyroidism) (529, 530) and in relation to certainneurotransmitter pathways (e.g., NPY, adrenergic, dopami-nergic, histaminergic, and serotoninergic) (531) (Table 4).
Conceptually, the interspecies disparities in the rat and sheepcan be summarized as reflecting 1) greater somatostatin de-pendence in the rat vs. greater GHRH dependence in the sheep(59); 2) more complex and variable GHRH/somatostatin rela-tionships preceding a GH pulse in the sheep; 3) stress inhibitionof GH release in the rat vs. stress stimulation in the sheep andhuman; and 4) a tendency for primarily amplitude-dependentregulation of pulsatile GH release in the sheep (and human) vs.combined amplitude and frequency modulation in the rat.These distinctions should be kept in mind, and the temptationavoided to extrapolate easily across species (59, 60, 183).
B. Acetylcholine and catecholamines
1. Acetylcholine. Cholinergic muscarinic pathways play animportant role in activating pulsatile GH secretion and, pre-sumptively, do so primarily via hypothalamic somatostatinwithdrawal (532–534). Indirect muscarinic agonists stimulate
FIG. 10. Inverse linear relationship between fasting serum leptinconcentrations and integrated 24-h serum GH concentrations in 15healthy postmenopausal women. The P and r values for the linearregression are shown. [Adapted with permission from R. Roubenoff etal.: J Clin Endocrinol Metab 83:1502–1506, 1998 (523). © The En-docrine Society.]
738 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

GH release acutely and enhance GH responses to GHRH inthe rat, dog, sheep, and human (206, 535–537). For example,in clinical experiments, the indirect cholinergic agonist, pyri-dostigmine, administered orally repeatedly over 48 h dou-bles the daily pulsatile mass of GH secreted in healthy youngand older men, but evokes lesser GH release in obese subjects(261). Conversely, antagonism of endogenous cholinergicpathways with muscarinic receptor-blocking drugs, such asmethscopolamine, atropine, or pirenzepine, strikingly re-duces sleep-associated GH release (538, 539) and virtuallyabolishes GH secretion otherwise triggered by various se-cretogogues including GHRH (535, 540, 541). An importantexception is that the GH response to insulin-induced hypo-glycemia is spared significantly during cholinergic musca-rinic receptor blockade, although the neuroendocrine mech-
anism of this interaction remains to be elucidated (542). In thedog, pirenzapine (a muscarinic antagonist) inhibited GHRH-stimulated GH release in both fed and fasted animals,whereas somatostatin inhibited this response preferentiallyin the fed state (543). Such findings in the dog tend to speakagainst the facile somatostatin hypothesis (as developed inthe rat) of muscarinic receptor-mediated withdrawal ofsomtatostatin and suggest secondary effects of such drugs onGHRH release and/or a plurality of effects on two or morefunctionally distinct receptor or neuronal populations (543–545).
The inferred mechanism of the GH-releasing action ofacetylcholine is based on direct and indirect in vitro andindirect in vivo experimental evidence in the rat (532–534,546). Acetylcholine can decrease somatostatin release di-
TABLE 4. Interspecies variations in neuroregulation of GH axis
Effector Rat Sheep Human
Adrenergica1 2 2 or 1 No effecta2 1 1 1b2 2 2 2
Age 2 Unknown 2Amino acids 1 (no change) 1 1Autofeedback at hypothalamus
by IGF-IYes No? Uncertain
Bombesin 1 Unknown No effect basally2 Hypoglycemic effect
Dopamine 2 ? 1Muscarinic
(cholinergic) 1 1 1(nicotinic) 2 Unknown 2
Cortisol/glucocorticoids 2or 1 2 2 or 1Diabetes Mellitus
Type I 2 Unknown 1Type II 2 Unknown 2 or 1
Estrogen 2 Amplitude 1 basal 1 Amplitude 1 AmplitudeExcitatory amino acids 1 Unknown UnknownExercise
Acute 2 Unknown 1Chronic 2 Unknown 1
Fatty acids 2 2 2GABA(-B) 1 Basal (2 stim.) 1 1 Basal (2 stim.)Galanin 1 Unknown 1GHRP 11 1 11Glucose No change No change 2Histamine 2 Unknown 1Hypoglycemia 2 No change 1Hypothyroidism 2 2 Stimulated GH (basal no change) 2IGF-I (pituitary inhibition) Yes Yes YesImmunization (or antagonist)
GHRH 2 Amplitude 2 Amplitude 2 AmplitudeSS 1 Valleys No change Unknown
Leptin 1 Unknown Inversely correlated with GHNeuromedin C 1 Unknown UnknownNeuropeptide Y 2 1 ? 1 ?Nitric oxide 1 or 2 Unknown No effectObesity 2 Unknown 2Opiates 1 1 1Senescence/aging 2 2 2Serotonin 1 2 1 (type ID)Starvation 2 1 1Stress (shock, restraint, endotoxin,
psychologic)2 1 1
Testosterone 1 More regular amplitude 1 1TRH 1 Basal, 2 stimulated Unknown No effect (1 in acromegaly)DHT 1 Unknown No effect
The reader is referred to the text and Refs. 51, 58–60, 271, 273, 481, 522, 531, 548, 554, 591, 592, 863, and 1099–1103.
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 739
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

rectly in vitro from rodent hypothalamic slices (547), al-though other experiments have contradicted this observationin the same species (546, 548).
In the rat, pyridostigmine administration stimulates GHrelease in the food-deprived animal (262), which is known tohave increased hypothalamic somatostatin tone (534, 549). Ofmechanistic relevance, pyridostigmine’s effect is diminishedby pretreatment of the animal with somatostatin antibodies(262). Pyridostigmine counteracts the inhibitory effect of glu-cocorticoid treatment for 4 days on the GH response toGHRH in normal male rats, which treatment also is thoughtto increase somatostatin tone (550). In support of the fore-going somatostatin hypothesis, the inhibitory influence ofatropine (cholinergic antagonist) on GH responses to GHRHin normal rats is abolished by pretreatment with somatosta-tin antiserum (551).
A recent hypophysial portal catheterization study showedthat neostigmine, another indirect cholinergic agonist, stim-ulates GHRH without inhibiting somatostatin release in thesheep (531, 546). These ostensible species differences havenot been fully rationalized; however, a plausible notion isthat, due to intrahypothalamic somatostatin-GHRH neuro-nal interconnectivity, central somatostatin withdrawal in-duced by this drug elicits rebound GHRH release (261, 544).This notion would bring several divergent observations intoagreement.
In humans, pyridostigmine partially restores the GH re-sponse to GHRH in physiological states hypothesized toreflect increased hypothalamic somatostatin release, i.e., 2-hrepeated bolus intraperitoneal GHRH injections (536), aswell as GHRH stimulation given 3 h after a bolus intravenousinfusion of recombinant human GH (259, 260). Moreover,pyridostigmine alleviates the acute inhibitory effect of glu-cocorticoid treatment on the GH- secretory response toGHRH in normal adults (465), and partially counteracts in-hibition of GH by glucocorticoid administration in children(552). These findings are relevant to the hypothesis that pyri-dostigmine acts by inhibiting somatostatin release, since,both in the rat and human, glucocorticoids appear to increasehypothalamic somatostatin secretion (171) (as discussed be-low). Conversely, in human GH hypersecretory states char-acterized by putatively low somatostatin tone, e.g., type 1diabetes mellitus, pyridostigmine does not further enhanceexaggerated GH release after GHRH injection (420, 553) (Fig.11). The relatively good clinical tolerability and the favorablepharmacodynamic profile of oral pyridostigmine (comparedwith some other cholinergic agents) have favored the ad-ministration of this substance, together with GHRH, in nu-merous clinical investigations of the somatotropic axis (421).
With the caveats above, available evidence suggests thatmuscarinic cholinergic activation is a dominant inhibitoryregulator of hypothalamic somatostatin release, and therebyparticipates critically in stimulatory GH neuroregulation inboth animals and humans. Based upon our present knowl-edge, the ability of (muscarinic) anticholinergic agents toblock the effects of multiple GH-releasing stimuli in the hu-man can thus be ascribed, in part, to cholinergic control ofsomatostatin secretion (535). We speculate that somatostatinrelease, in turn, likely modulates GHRH release via recip-rocal connectivity between hypothalamic GHRH and soma-
tostatinergic pathways (183, 265, 554, 555). As noted earlier(261), such functional interactions could explain why aGHRH antagonist can impede stimulation of GH release byan indirect muscarinic cholinergic agonist (556) and whysuch an agonist might also release GHRH (531); viz., bylimiting somatostatin’s inhibition of GHRH release intrahy-pothalamically. In brief, an increase in GHRH secretion couldreflect GHRH rebound release after pyridostigmine’s with-drawal of intrahypothalamic somatostatin, which itself re-strains GHRH secretion (557). Moreover, in the human, so-matostatin infusion reduces both GH pulse frequency andamplitude (123), consistent with such bivalent mechanismsof somatostatin’s inhibition of both GH and GHRH release.
Muscarinic and nicotinic cholinergic pathways may playopposing regulatory roles (facilitative and inhibitory, respec-tively), since nicotinic receptor blockers enhance GH releaseduring insulin-induced hypoglycemia or in response to sleep(558).
2. Catecholamines.a. Dopaminergic pathways. In vitro dopamine reduces hu-
man pituitary GH release stimulated by GHRH (559, 560).Since dopamine receptors are present on human somato-tropes, this in vitro inhibitory effect of dopamine is likelyexerted directly. However, most human in vitro experimentaldata derive from the study of GH-secreting pituitary ade-nomas; notably, acromegalics show a paradoxical inhibitoryresponse to l-dopa in vivo compared with normal subjects.Specifically, in acromegalic patients, acute or chronic dopa-mine agonist treatment inhibits GH secretion (560), whereasin normal subjects acute administration of dopamine ago-nists such as l-dopa, apomorphine, dopamine itself, andbromocriptine causes GH release (561, 562). The latter resultsare not in accord with most direct studies of dopamine’s (D2)stimulation of somatostatin secretion in the rat (see Fig. 1A)(548, 563–565). Analagously, in bovine brain explants, in vitrodopamine acting via D1 receptors increases hypothalamicsomatostatin secretion and reduces GHRH release (566). Thismay reflect species differences compared with the human.
Estrogen treatment in one clinical study enhanced theGH-releasing effect of l-dopa in girls with Turner’s syn-drome (567). In prepubertal children, a single oral dose of20–40 mg ethinyl estradiol augmented l-dopa-stimulatedGH release (568). Analogously, and consistent with soma-tostatin withdrawal in the human, significant increases in theGH response to GHRH occur after pretreatment with bro-mocriptine (569, 570). In contrast, dopamine and dopami-nergic drugs fail to enhance the GH-secretory responses toinsulin-induced hypoglycemia and l-arginine (571, 572),both of which are thought to limit somatostatin release. Thus,dopaminergic stimulation of GH release in the human isestrogen sensitive and appears to facilitate GHRH actionpossibly via somatostatin withdrawal (573). Somatostatinwithdrawal per se may evoke or accentuate GHRH release(22, 129–131, 574), which mechanistically likely reflects reliefof inhibitory somatostatinergic synapses on GHRH-secre-tory neurons (Fig. 7). Thus, the rat and human (Fig. 1, A andB) responses to dopamine are apparently opposite, akin tothe major species differences recognized in fasting, diabetes,stress, and hypoglycemia (Table 4).
740 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
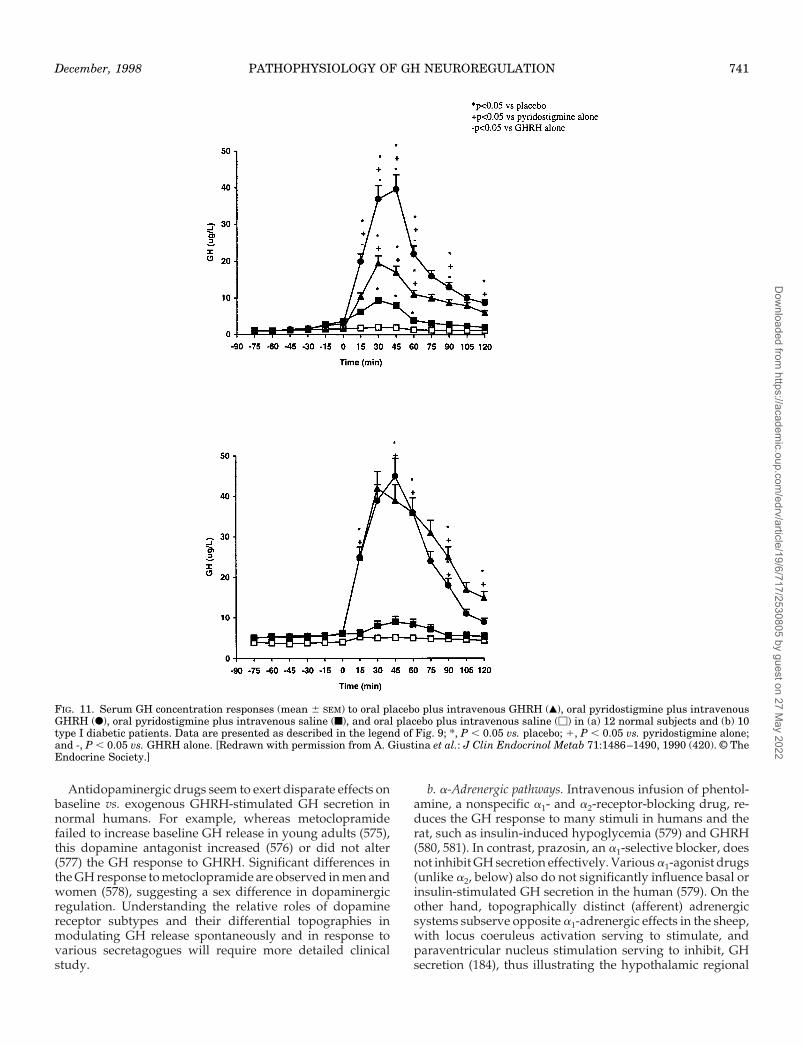
Antidopaminergic drugs seem to exert disparate effects onbaseline vs. exogenous GHRH-stimulated GH secretion innormal humans. For example, whereas metoclopramidefailed to increase baseline GH release in young adults (575),this dopamine antagonist increased (576) or did not alter(577) the GH response to GHRH. Significant differences inthe GH response to metoclopramide are observed in men andwomen (578), suggesting a sex difference in dopaminergicregulation. Understanding the relative roles of dopaminereceptor subtypes and their differential topographies inmodulating GH release spontaneously and in response tovarious secretagogues will require more detailed clinicalstudy.
b. a-Adrenergic pathways. Intravenous infusion of phentol-amine, a nonspecific a1- and a2-receptor-blocking drug, re-duces the GH response to many stimuli in humans and therat, such as insulin-induced hypoglycemia (579) and GHRH(580, 581). In contrast, prazosin, an a1-selective blocker, doesnot inhibit GH secretion effectively. Various a1-agonist drugs(unlike a2, below) also do not significantly influence basal orinsulin-stimulated GH secretion in the human (579). On theother hand, topographically distinct (afferent) adrenergicsystems subserve opposite a1-adrenergic effects in the sheep,with locus coeruleus activation serving to stimulate, andparaventricular nucleus stimulation serving to inhibit, GHsecretion (184), thus illustrating the hypothalamic regional
FIG. 11. Serum GH concentration responses (mean 6 SEM) to oral placebo plus intravenous GHRH (Œ), oral pyridostigmine plus intravenousGHRH (F), oral pyridostigmine plus intravenous saline (f), and oral placebo plus intravenous saline (M) in (a) 12 normal subjects and (b) 10type I diabetic patients. Data are presented as described in the legend of Fig. 9; *, P , 0.05 vs. placebo; 1, P , 0.05 vs. pyridostigmine alone;and -, P , 0.05 vs. GHRH alone. [Redrawn with permission from A. Giustina et al.: J Clin Endocrinol Metab 71:1486–1490, 1990 (420). © TheEndocrine Society.]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 741
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

complexity of noradrenergic control of the GH axis. In themale rat a1-inhibition of GH secretion also can be evoked viaparaventricular nuclear effects (582).
The stimulation of a2-receptors (e.g., with agonists such asclonidine and guanfacine) induces GH release in man andanimals (583–585). Relevant a2-adrenergic receptors areprobably accessible to the blood within the median eminence(586). The postsynaptic a2-adrenergic receptors involved inthe regulation of GH secretion are similar to those mediatinghypotensive effects in the cardiovascular system (587), whichthus may confound experimental interpretations, e.g., in therat (see below). Interestingly, the GH-releasing action ofclonidine is effectively blunted in patients with essentialhypertension, who may harbor hypothalamic alterations inadrenergic tone (588).
Many studies in the rat have shown that, when GHRH isinactived by passive antibody transfer (585), or when thehypothalamic arcuate nucleus that secretes GHRH is le-sioned (589), the GH response to a2-receptor agonists is abol-ished. In contrast, pretreatment of animals with antibodies tosomatostatin does not alter the GH response to a2-agonistdrugs (590). Some other experiments suggest that GH secre-tion induced by a2-receptor stimulation in the rat is notmediated by GHRH release from the median eminence butrather by reduced somatostatin secretion (591). On the otherhand, marked GH release after clonidine treatment in thesheep is temporally associated with or preceded by portalGHRH secretion with no consistent change in somatostatinrelease (592). In bovine hypothalamic slides, guanabenz, ana2-receptor agonist, also releases GHRH but does not affectsomatostatin (566). Caution is required in interpreting someearlier experimental data, since clonidine (an a2 agonist) isnot a very potent GH secretagogue in the rat and inducespotentially confounding hemodynamic, ventilatory, and be-havioral changes in this species (591).
Clinical studies reveal attenuation of clonidine’s stimula-tion of GH rlease in patients with presumptively increasedhypothalamic somatostatinergic tone, e.g., individuals re-ceiving chronic treatment with glucocorticoids (590, 593), orthose with hyperthyroidism (594). However, in humans themechanism of a2-receptor-mediated GH secretion may bemore complex than that in experimental animals. For exam-ple, the hypothesis that a2-adrenergic agonists may act alsovia non-GHRH mechanisms, such as somatostatin release,arises from the finding that pretreatment with GHRH abol-ishes the GH response to a second GHRH stimulus withoutaffecting clonidine’s action (595). These observations are con-sistent with, but not proof of, an ability of clonidine to with-draw somatostatin in the human.
The role of estrogen in regulating a2-adrenergic pathwaysthat impact GH secretion remains unclear. Whereas the mag-nitude of GH release after clonidine (0.15 mg) ingestion inmidluteal phase women is proportionate to the serum es-tradiol concentration (596), conjugated estrogen administra-tion does not alter clonidine’s stimulation of GH release inpostmenopausal women (597). Thus, the nature and extent ofsex-steroidal interactions with a2-receptor-mediated GHneuroregulation are not established in the human. In clinicalstudies in adults, clonidine is a relatively poor GH secreta-
gogue (598), making investigations with this compound dif-ficult.
c. b-Adrenergic pathways. Experiments performed in thehuman and laboratory animals in vivo using several b-adrenergic receptor-blocking agents, such as propranolol(599) or atenolol (600), support the operating hypothesis thatb-adrenergic receptors mediate significant inhibitory effectson GH release. Studies with inhibitors to phenylethano-lamine N-methyl transferase, the enzyme converting norepi-nephrine to epinephrine, indicate that the neurotransmitterprimarily involved in the b-receptor-mediated inhibition ofGH-release is l-epinephrine (601). If b-adrenergic receptorsare responsible for the inhibitory action of catecholamines onGH release in vivo, this effect is not likely to be due to a directaction on the pituitary gland. Although rat somatotropesexpress b-adrenergic receptors, b-agonistic agents actuallystimulate GH secretion in rat pituitary cell cultures (602).Therefore, the inhibitory effect on GH release that followsb-adrenergic activation in vivo is probably mediated by hy-pothalamic actions on GHRH and/or somatostatin release.
Several experiments suggest that b-receptors modulatehypothalamic somatostatin tone: 1) Whereas isoprenaline,a b-agonist, stimulates somatostatin release from the ratpancreas (603), propranolol inhibits somatostatin releasefrom hypothalamic slices in vitro (604); 2) Somatostatinantibodies block inhibition by isoprenaline of GHRH-in-duced GH release in the rat (605); 3) b-Adrenergic antag-onists disrupt the (negative) autofeedback of GH, which islikely mediated by somatostatin in the human (606); 4)Propranolol enhances GH responses to hypoglycemia, ex-ercise, glucagon, and GHRH in humans (599), and thereare ethnic differences in GH responses to this nonspecificb-receptor blocker (607); and 5) clinical studies indicatethat the b2-adrenergic receptor agonist, salbutamol, inhib-its GH secretion and overcomes stimulation by the aminoacid, l-arginine, and the indirect cholinergic agonist, pyri-dostigmine (82). The last two substances are thought toreduce somatostatin release. Thus, b2-adrenergic receptorstimulation of hypothalamic somatostatin secretion seemsable to overcome the ability of l-arginine and acetylcho-line to limit somatostatin release.
Acute administration of either of two different b2-re-ceptor agonist drugs (salbutamol or broxaterol) blunts theGH response to physical exercise in man (608). One mech-anistic explanation is that exercise alters catecholaminer-gic pathway activities in the hypothalamus so as to favoran increase in a2 adrenergic (GH-stimulatory) tone; pre-treatment with b2-agonist (GH-inhibitory) drugs may pre-vent this phenomenon and thus attenuate the serum GHresponse to exercise. Another consideration is that exerciseincreases (via an unknown mechanism) hypothalamicGHRH secretion, which is counteracted by the rise inhypothalamic somatostatin release caused by the b2-re-ceptor agonists. Finally, b2-adrenergic activation andphysical exercise may exert opposing effects on hypotha-lamic somatostatin tone, and/or act on cosecretagogues ormodulators. The neuroregulation of exercise-induced GHrelease is discussed further below.
742 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

C. Other neurotransmitters
1. Serotonin. Serotoninergic pathways are stimulatory to GHrelease in the rat, possibly via promoting GHRH release,since pretreatment with GHRH antiserum inhibits the GH-secretory response to serotonin agonists in this species (609).In contrast, in the sheep, intravenous injection of tianeptine,a serotonin uptake enhancer and inhibitor of serotonin ac-tion, evokes a significant, immediate, and short-lasting (30-min) increase in peripheral GH (1750%) and hypophysialportal GHRH (1180%) concentrations in conscious un-stressed animals. There is no significant concurrent changein the secretion of somatostatin (610). Such data suggest thatendogenous serotoninergic input is inhibitory to GHRH andGH secretion in the ruminant, comparable to indirect infer-ences in the dog (611), but not in the rodent. Thus, speciesdifferences exist in serotoninergic neurotransmitter regula-tion of the GH axis.
In man, administration of 5-hydroxytryptophan, a sero-tonin precursor (which however also releases cat-echolamines from synaptic terminals) increases serum GHconcentrations (612). On the other hand, neither quipazine,a direct serotonin receptor agonist, nor fenfluramine, a drugthat releases serotonin from serotoninergic terminals (andalso interferes with dopamine neurotransmission), altersbasal GH concentrations (613, 614). Fenfluramine abolishesGH release stimulated by l-dopa and propranolol withoutaffecting the response to l-arginine (615). Administration ofthe nominal serotonin receptor antagonist, cyproheptadine(which also has anticholinergic actions) or methysergide(which also appears to be a serotonin agonist), can inhibit GHresponses to presumptive hypothalamic stimuli, such as hy-poglycemia, l-dopa, and clonidine (614, 616). Because of theimperfect specificity of the drugs used in the foregoing clin-ical studies, the precise role of serotonin pathways in theregulation of GH secretion in man has been difficult to es-tablish.
At least four major subtypes of brain serotonin receptorshave been identified recently (617). In this context, a novelselective serotonin-1 (subtype D) receptor agonist,sumatriptan, has been employed to evaluate the relevance ofspecific serotonin receptors in GH secretion (618). The mainadvantage of this agent is that, although it also interacts inhumans with the subtype B serotonin-1 receptor, the latter isnot present in human brain; moreover, this drug does notactivate other serotonin receptors, or adrenergic, dopami-nergic, muscarinic, or GABA receptors (619). Sumatriptanincreases basal GH release in normal adults without affectingcortisol secretion, thus suggesting that this effect is not stressrelated (620). The actions of this serotonin subytpe-1D re-ceptor agonist have also been assessed in normal subjects incombination with GHRH, as well as with compounds that actas presumed functional somatostatin antagonists, e.g., l-arginine and pyridostigmine. Since sumatriptan potentiatesthe GH response to a maximal dose of GHRH (206, 618), onecan postulate that this agent decreases hypothalamic soma-tostatin secretion. The lack of additive effect of sumatriptanand pyridostigmine further implies that the two drugs actthrough the same (final) mechanism, i.e., inhibiting soma-tostatin release. That sumatriptan increases GH responses to
l-arginine is somewhat paradoxical and suggests that thesesecretagogues may act through at least partially independentmechanisms and/or synergize in inhibiting somatostatin re-lease (618). Thus, available clinical data indicate that sub-type-1D serotonin receptors mediate a significant GH-stim-ulating effect in humans, presumably via inhibition ofhypothalamic somatostatin secretion.
2. Histamine. Information regarding the possible role of his-tamine in regulating GH release is relatively sparse. Thecentral histaminergic system seems to mediate inhibition ofphysiological (621) and pharmacologically induced (622) GHsecretion in adult rats. Histamine does not alter GH secretionfrom the pituitary gland in vitro (563), but exerts an inhibitoryeffect on GH secretion in adult rats when injected icv (621,622). This occurs possibly via suppression of GHRH release,since histamine does not appear to increase hypothalamicsomatostatin release (623). Reduction of histaminergic activ-ity by treatment with a-fluoromethylhistidine, an inhibitor ofhistamine synthesis (624), increased GH release as stimulatedby opioids in both neonatal and adult rats (625). An expla-nation is that a-fluoromethylhistidine, by removing inhibi-tory histaminergic control of GHRH release, may facilitateopioid stimulation of GHRH secretion.
Administration of histamine in man does not affect GHsecretion (496). However, an H1-receptor antagonist reducesthe GH response to l-arginine but not to hypoglycemia (626),whereas an H2-receptor antagonist diminishes stimulatedGH release to a much smaller extent (627). An H1-receptoragonist elicited paradoxical GH release in response to a TRHstimulus in normal subjects (496). Thus, in humans, hista-minergic pathways, acting presumptively through H1-receptors, may play a conditional stimulatory role in GHsecretion in man (in contrast to what is observed in the rat).The facilitative effects of histamine in the human are likelymediated at the hypothalamic level, albeit via mechanismsthat are unknown at present.
3. GABA. In humans, administration of sodium valproate, apostsynaptically active drug that increases accumulation ofendogenous GABA, inhibits diazepam-induced GH releasewithout affecting GH release stimulated by hypoglycemia(628, 629) or GHRH. On the other hand, administration of theGABAB receptor agonist, baclofen, stimulates basal GH re-lease and inhibits the GH responses to l-arginine and hy-poglycemia (630). The stimulation by GABA-like agonists ofbasal GH secretion is not likely mediated via GHRH releasefrom the hypothalamus, since, at least in the rat, stimulationis not abolished by passive immunization with GHRH an-tiserum (631). On the other hand, GABA may inhibit exog-enous secretagogue-stimulated GH secretion via an increasein hypothalamic somatostatin tone (632). Of mechanistic in-terest, a GABA agonist stimulates both GH secretion andslow-wave sleep in the human (633), which is consistentwith, but not proof of, GABA pathways interfacing withsomatostatinergic systems (222, 383). Nocturnal withdrawalof somatostatin in the human has been inferred by enhancedGH release in response to GHRH infusions during slow-wave sleep (634).
Although GABA also evokes galanin release from rat hy-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 743
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

pothalamic fragments in vitro (635), and galanin can stimu-late GH release in vivo (above), GABA antagonists actuallyimpede galanin’s stimulation of GH secretion in the rodent(434). Thus, the exact relationship between GABA and ga-lanin’s activation of GH secretion needs to be clarified fur-ther. In this regard, GABA can directly stimulate GH releasein vitro from neonatal rat pituitaries in a chloride and calci-um-dependent manner (636–639). In the sheep, like the rat,either intravenous or icv injection of GABA agonists willstimulate GH secretion (640). In the human, stimulatory ef-fects of GABA agonists are also evident whether assessed viathe use of a GABA-B agonist (baclofen), a GABA-ergic stim-ulus (valproic acid), or a GABA metabolite (641, 642). Themechanisms subserving GABA-stimulated GH secretion inthe human have not been elucidated, although metergoline,a nonspecific serotonin-receptor antagonist, and flumazenil,a benzodiazapine receptor antagonist, both inhibit GABA-stimulated GH secretion (642, 643).
Various clinical conditions are accompanied by dimin-ished GH release (compared with healthy controls) afterbaclofen or valproic acid stimulation of GH secretion. Forexample, in depression, whether or not preceded by 1month’s treatment with tricyclic antidepressants (644),GABA’s stimulation of GH secretion is attenuated (643).Other examples of impaired responsiveness to GABA acti-vation include heroin addicts (645), type I diabetes mellitus(646), Parkinson’s disease (647), and schizophrenia (648).Importantly, there are both sex-specific and age-dependentinfluences on GABA’s stimulation of GH secretion in thehuman, with baclofen (a GABA-B agonist) stimulating GHsecretion in men but not women (649), and in younger butnot older men (650). In addition, in acromegalic patients,valproic acid fails to heighten GH secretion (651), whichlikely reflects the (partial) autonomy of somatotropinomacells.
The production of GABA requires the activity of two keyenzymes: GABA-T (4-aminobutyrate-2-oxaglutarate amino-transferase) and GAD (l-glutamate decarboxylase). GADconverts glutamate to GABA via a decarboxylation reaction,which is the rate-limiting enzymatic step in the biosynthesisof GABA (652). GAD is a major autoantigen in type I diabetesmellitus and a target of both humoral and cell-mediatedautoimmunity in this disease (653). Marked GH hypersecre-tion is characteristic of type I diabetic patients (654), unlikevariably increased or blunted GH secretion in patients withtype II diabetes (655). Whether a putative autoimmune pro-cess is involved in this derangement of GH neuroregulationis unknown. Against this speculation, circulating autoanti-bodies to GAD accompany, but do not correlate with, anincreased GH response to GHRH in long-standing type Idiabetic patients (656). Moreover, neutralizing antibodies orantibodies mirroring destruction of GAD-containing cellswould diminish GABA production and hence be expected toreduce GH secretion.
In summary, the GABA pathway facilitates basal GH se-cretion in the rat, sheep, and human, possibly via mecha-nisms involving galanin, serotoninergic, or benzodiazepinereceptors, albeit no neurotransmitter mechanisms are estab-lished unambiguously. Moreover, in the neonatal rat, GABAstimulates GH release directly in vitro from pituitary cells.
Numerous clinical conditions associated with either exces-sive GH secretion (type I diabetes mellitus or acromegaly) orrelatively decreased GH production (healthy aging in men)are accompanied by impoverished stimulation of GH secre-tion by GABA agonists. Further investigation will be re-quired to clarify the pathophysiological relevance of thisneurotransmitter pathway in impacting the human GH axisin health and disease.
4. Excitatory amino acids. In the rat and guinea pig, excitatoryamino acids, such as N-methyl-d,l-aspartate (NMDA) aswell as glutamate, can stimulate the GH axis (657–659). Al-though the mechanisms mediating excitatory amino acids’stimulation of GH release are not established, GHRH anti-serum blocks excitotoxin-stimulated GH secretion in vivo inthe pig (657), suggesting hypothalamic mediation. However,smaller stimulatory effects (;2-fold and not always dosedependent) are reported in vitro (657, 660). For example,NMDA stimulates single-somatotrope GH secretion in thereverse hemolytic plaque assay, an effect that is blocked bya specific NMDA receptor antagonist, is additive with GHRHactions, and is suppressed by somatostatin (661). Among thearray of known excitatory amino acids, both NMDA andkainic acid stimulate GH secretion in vivo in the rat (659).Much additional investigation will be required to delineatethe express role of excitatory amino acids in the regulatorypathophysiology of GH secretion in the rat and other exper-imental animals, as well as eventually in the human.
5. Role of nitric oxide as a neuromodulator in the GH axis. Theamino acid, l-arginine, is the immediate biological precursorof nitric oxide (662). The enzymes converting l-arginine tonitric oxide were purified and cloned in 1991 (663, 664) andnamed nitric oxide synthase (NOS). This enzyme family re-leases nitric oxide as a mediator gas from the terminal gua-nidine nitrogen group of l-arginine, producing l-citrullineas a byproduct. NOS is inhibited by derivatives modifiedat the terminal guanidino group, such as N-monomethyl-l-arginine (l-NMMA), N-nitro-l-arginine methyl ester (l-NAME) (665, 666). NOS is present in at least three isoformsvariously expressed in vascular, neural, and other tissues. Incentral and peripheral nervous tissues, a constitutive isoformprevails (NOS I) (667). Whereas l-arginine likely promotesGH release via inhibition of somatostatin secretion, neitherthe neurotransmitter intermediates nor the precise biochem-ical basis of this stimulatory effect (e.g., whether via nitricoxide) is known (668).
The possible role of the mediator gas, nitric oxide, in mod-ulating GH secretion and/or action will be important toinvestigate further. GH’s stimulation of target tissues acti-vates a complex intracellular signaling cascade, as reviewedelsewhere recently (5, 669). Whereas GH’s signaling actionsmay involve, in some measure, nitric oxide, the necessaryand/or sufficient roles of nitric oxide in various effects of GHon different target tissues remains to be established. In favorof nitric oxide’s role in GH action, a recent study in GH-deficient patients showed that treatment with recombinanthuman GH stimulated urinary nitrite (a principal metaboliteof nitric oxide) as well as cyclic GMP excretion. This suggestsbut does not prove that nitric oxide generation might account
744 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

for some of the vasodilatory and other hemodynamic actionsof GH clinically (670).
The neuroendocrine role of nitric oxide in mediating ordirecting secretory activity of the GH axis in the rat is unclear,in contrast to its likely agonistic regulation of neuronal CRHand GnRH release (671, 672). For example, recent studies arecontroversial, suggesting either an inhibitory or a stimula-tory role for nitric oxide in GH secretion. In one in vitro study,a nitric oxide scavenger abolished GHRH-stimulated GHsecretion by (prepubertal) rat male pituitary cells in culture(673), whereas in another in vitro study, a nitric oxide syn-thase inhibitor increased GHRH actions (674). In keepingwith the latter inference, nitric oxide seemed to mediate theinhibitory actions of the cytokine, g-interferon, on GH se-cretion in vitro (675). In in vivo experiments, an NOS inhibitorlowered GH secretion in the rat (671). Considerable addi-tional study will be required to clarify these discrepancies,which may arise in large part from the nonspecificity of theexperimental probes of the nitric oxide pathway, and theconfounding complexities of in vivo inhibition of the nitricoxide signal.
In the human, no direct evidence exists for a role of nitricoxide in basal or l-arginine-stimulated GH neuroregulation.In negative clinical studies, intravenous infusion of an NOSinhibitor (l-NAME) did not block the GH response toinsulin-induced hypoglycemia, but did amplify the ACTH-secretory response to this stimulus (676). The converse ex-perimental strategy of administering a nitric oxide donor,molsidomine, also failed to alter GH secretion (668). Thus,available clinical experiments fail to show that nitric oxidedrives GH secretion in the human, although this conclusioncannot be viewed as definitive until highly specific nitricoxide probes become available and are tested with appro-priate positive and negative controls.
VI. Role of Metabolic Substrates in the Regulation ofthe GH Axis
The critical role of GH in promoting somatic growth is wellknown (677). A recent evolving emphasis is that GH alsoserves as an essential regulator of body composition, inter-mediary muscle and bone metabolism (678), and cardiacfunction (679, 680). This more global view of GH as a primemodulator of fuel metabolism, helps explicate why and withwhat physiological significance metabolic substrates (aminoacids, glucose, and lipids) feed back in a complex manner atthe hypothalamic and pituitary levels to regulate GH secre-tion. Such feedback actions are quite species specific, withmany prominent neuroregulatory differences in the GH axisof humans and experimental animals (see Table 4).
A. Blood glucose
1. Hypoglycemia. A mechanistically important observation isthat the clinically well known and marked GH-secretoryresponse to insulin-induced hypoglycemia is not mediatedsolely via GHRH, since GHRH pretreatment abolishes thesubsequent GH response to GHRH, but not to hypoglycemia(681). This finding is compatible also with a true pituitarydesensitization phenomenon specific for GHRH’s stimula-
tion of somatostatin release, or some degree of pituitary GHdepletion (namely, loss of a specific GHRH-releasable pool).On the other hand, combined GHRH stimulation and insu-lin-induced hypoglycemia exert additive effects on GH re-lease (681), which is consistent with proposed somatostatinwithdrawal during hypoglycemia. Alternatively, GHRH andhypoglycemia may act on different releasable pools of GHwithin the somatotrope population. In addition, hypoglyce-mia may inhibit somatostatin release and/or action in man,although the former is not observed by portal vein samplingin the sheep (51). Moreover, the rat responds to the stress ofhyoglycemia with decreased GH secretion, thus highlightingmajor species disparities in the response of the GH axis toglucose deprivation.
2. Hyperglycemia.a. Normal physiology. A rise in serum GH concentrations
occurs 3–5 h after oral glucose administration in man (682,683). This delayed GH increment is preceded by initial sup-pression of plasma GH levels for 1–3 h (682, 683). The clin-ically well known rapid inhibitory effect of glucose on GHrelease may be due to a discharge of somatostatin from thehypothalamus. In accordance with this proposed mecha-nism, acute hyperglycemia blocks GH secretion stimulatedby GHRH (684–690). Conversely, central cholinergic recep-tor activation by pyridostigmine, which is inferred to sup-press hypothalamic somatostatin release, counteracts theacute inhibitory action of glucose on GH release (687, 689,690). Oral glucose does not likely have a direct pituitaryeffect, since glucose does not influence basal GH secretion orGH responses to GHRH in vitro (691, 692). Thus, glucoseappears to modulate GH secretion through one or morehypothalamic mechanisms, probably somatostatinergic, inthe human.
In the human, we infer that oral glucose acts rapidly tosuppress GH release by increasing hypothalamic somatosta-tin release, thus suppressing serum GH for 1–3 h. The re-duced GH release results in an increase in the pituitary storesof GH. When somatostatin release declines, endogenousGHRH secretion is activated reciprocally, and available pi-tuitary stores of GH are released, leading to the “rebound”increase in serum GH levels. Since the administration ofpyridostigmine, which may inhibit somatostatin secretionand release GHRH, potentiates the late GH rise induced bythe oral glucose, we infer that hypothalamic somatostatinrelease may not be shut off completely, and/or GHRH dis-charge may be increased partially, at the time of the 3–5 hdelayed GH rise after oral glucose injection (693).
Studies carried out in vitro using perifused rat hypotha-lamic fragments reveal an inverse relationship between glu-cose concentration and somatostatin (694, 695) or GHRH(695) release. In the rat in vivo, however, basal or GHRH-stimulated GH release is not altered by acute hyperglycemia;therefore, the relevance of (metabolic) data obtained in therodent to man is often limited. Moreover, the control mech-anisms that confer these species differences are not under-stood at present.
b. Diabetes mellitus. GH secretion is markedly altered indiabetes mellitus in both rats and humans, albeit in oppositedirections. In principle, abnormal GH secretion in the dia-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 745
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

betic context could originate via several mechanisms: 1) hy-pothalamic neurotransmitter alterations resulting in changesin GHRH and/or somatostatin secretion; 2) pituitary varia-tions in the affinity and/or number of receptors for GHRHand/or somatostatin; 3) pituitary changes in GH synthesisand release; and 4) altered feedback regulation of GH secre-tion at the hypothalamic and/or pituitary levels (696).
The adverse effects of diabetes on the circulatory, visual,renal, and peripheral nervous systems are commonly rec-ognized; neuroregulatory disruption is also known, as re-viewed in part in 1994 (654). The impact of decreased insulinsecretion or attenuated insulin action on the function of otherendocrine glands is not so well documented. Both clinicaland animal research demonstrate that diabetes mellitus iscommonly associated with altered thyroid, adrenal, and go-nadal function. Some of these changes are reversed by insulinreplacement therapy, but endocrine function is not alwaysnormalized even with rigorous glycemic control. For exam-ple, patients with poorly controlled diabetes mellitus exhibitGH hypersecretion basally and when variously stimulated,while patients with good metabolic control still present withdiurnal and exercise-induced GH hypersecretion (654). Inevident contrast, diabetes suppresses GH secretion in the rat.Clinical and experimental evidence exists for diabetes-asso-ciated changes in GHRH and somatostatin release as well asin the pituitary response to these hypothalamic hormones(697).
Despite GH hypersecretion, basal levels of IGF-I (the prin-cipal tissue mediator of GH activity), are low in type I dia-betes mellitus (698). Serum concentrations of IGFBPs, thecarriers that transport IGFs in serum and modulate theirtissue-specific bioactivity, are also altered. Specifically, bloodconcentrations of IGFBP-3, the major transport protein forIGF-I, are decreased in poorly controlled diabetics (698). Incontrast, IGFBP-1, a carrier protein that also likely functionsin the autocrine/paracrine modulation of IGF bioavailability,is increased in serum of type 1 diabetic patients, and IGFBP-1levels are inversely correlated with glycemic control (698).How such disturbances in circulating IGFBPs influence GHand/or IGF-I autonegative feedback on the GH axis in dia-betic individuals is not known.
Understanding of the mechanisms responsible for the ab-normalities in the GH-IGF-I axis in type I diabetes mellitusis incomplete, although recent evidence strongly suggeststhat portal vein insulinopenia contributes to dysregulation.Down-regulation of hepatic GH receptor expression second-ary to portal insulin deficiency could explain the apparentGH resistance observed in this disease. Consistent with thishypothesis, there is a decrease in circulating concentrationsof GH binding protein, a putative index of GH receptornumber, in patients with type I diabetes mellitus (698). Theabnormalities in the GH/IGF-I axis have been implicated inthe worsening of metabolic control that occurs in some di-abetic patients, as well as in the development of microvas-cular complications, such as retinopathy and nephropathy(699).
Diabetes mellitus in the rat, whether induced by strepto-zotocin administration or occurring spontaneously, mark-edly suppresses pulsatile GH secretion and blunts theGH-secretory response to GHRH (700). Hypothalamic so-
matostatin appears to play a major role in this animal model(701, 702), since 1) there are increases in immunoreactivesomatostatin levels in hepatic portal and peripheral blood inspontaneously diabetic rats (703); 2) the attenuated GH re-sponse to GHRH is normalized after administration of so-matostatin antibodies (702), or deep pentobarbital anesthesia(563); and 3) pituitary GH content is unaltered in spontane-ously diabetic rats (702). Indeed, increased hypothalamicsomatostatin release with unaltered GHRH secretion mightbe expected to increase pituitary GH content in the diabeticrodent. Unexpectedly, direct hypophysial-portal blood sam-pling in the streptozotocin-induced diabetic male rat re-vealed decreased hypothalamic release of both GHRH andsomatostatin (704). Unfortunately, similar data are not yetavailable in other species, such as the sheep or primate, thusrendering this issue indeterminate at present.
The male BB/Worcester rat with insulinopenic diabetesmellitus exhibits a markedly reduced mass of GH secretedper bust and the GH-secretory rate (705), but GH-secretoryburst frequency, secretory burst half-duration, and serumGH half-life are unchanged. In vitro studies of acutely dis-persed somatotropes obtained from rats with diabetes mel-litus show increased sensitivity to GHRH, as quantitated bya greater mean hemolytic plaque area after exposure to anEC50 dose of the secretagogue and diminished sensitivity tosomatostatin’s inhibition of GH release driven by an EC50
dose of GHRH. The numbers of pituitary cells and somato-tropes were indistinguishable in diabetic and normoglyce-mic animals (705). Thus, somatotrope in vitro responsitivityin the diabetic rodent seems to manifest a “denervation-like”hypersensitivity, perhaps reflecting endogenous GHRHwithdrawal (and somatostatin excess).
The GK Wistar rat is a new model of diabetes mellitus innonobese animals with significant fasting hyperglycemia,hyperinsulinemia, and absent insulin response to iv glucose.The GH response to GHRH is reduced at 16 weeks of agecompared with normal, age-matched Wistar rats, but nodifferences are observed at 6 weeks of age. Pretreatment ofolder rats (16 weeks) with somatostatin antibodies signifi-cantly increases the GH response to GHRH in both normalor young (6 weeks old) GK rats (706). These results supportthe thesis (above) that accentuated somatostatin release me-diates the blunted GH response to GHRH in diabetic GK rats.Reduced hypothalamic cholinergic signaling to the soma-tostatinergic neuron might, in turn, permit excess somatosta-tin release. This view is supported by the results of both invitro and in vivo studies. In vitro, cholinergic muscarinicblockade with pirenzepine causes dose-related stimulationof SS release from normal hypothalami, but does not affectGK rat hypothalami (706). In vivo, concentrations of immu-noreactive somatostatin are higher in hepatic portal and pe-ripheral blood in spontaneously diabetic rats (703). Finally,the attenuated GH response to GHRH normalizes after pen-tobarbital anesthesia, which presumably suppresses hypo-thalamic somatostatin (and other neuropeptide) release (563,702). Therefore, increased hypothalamic somatostatin secre-tion may be the primary determinant of the GH suppressionobserved in the diabetic rodent. We emphasize that ampli-fied somatostatin release would also be expected to quash
746 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

GHRH output via intrahypothalamic reciprocal regulation(131, 265).
c. Type 1 diabetes mellitus patients. Available clinical datashow elevated 24-h GH release in untreated and treated typeI diabetic patients (707–710). Detectable serum GH peak fre-quency and interpeak GH concentrations are higher in thesepatients (708), and exaggerated GH release in response toGHRH occurs consistently (420, 711, 712). Mechanistic stud-ies with the acetylcholinesterase inhibitor, pyridostigmine,which is an indirect cholinergic agonist hypothesized to de-crease somatostatin release (262), indicate that type I diabeticpatients with an exaggerated GH response to GHRH fail torespond with further GH release to pyridostigmine treat-ment (Fig. 11). This finding suggests, but does not prove,maximally decreased endogenous somatostatin release in(human) type I diabetes mellitus (420, 558), and/or suggestsmaximal somatotrope output of GH.
Presumed decreased somatostatin release in human typeI diabetes in turn may arise from impaired GH and/or IGF-Iautofeedback. Thus, although GH pretreatment reducesGHRH-stimulated GH secretion in both healthy man and rat,probably by increasing hypothalamic somatostatin secretionvia cholinergic pathways (259), GH pretreatment (with orwithout pyridostigmine) does not limit the GH-secretoryresponse to GHRH in type I diabetic patients (260, 713).Conversely, pirenzepine, a cholinergic muscarinic antago-nist, acutely inhibits spontaneous, sleep-related, and stimu-lated GH release in patients with insulin-dependent diabetesmellitus (714, 715), suggesting the releasability of additionalsomatostatin. Pirenzepine truncates the amplitude of GHpulses, but has no effect on GH peak frequency. However, inmore chronic studies, pirenzepine given in a daily dose of 75or 150 mg for 4 days does not affect GH secretion (716, 717),indicating “escape” of somatostatin withdrawal, drug tachy-phylaxis, etc. Autoimmunity also has been speculated tocontribute to derangement in somatostatin tone, e.g., viaGAD antibodies (718). However, the GH response to GHRHplus pyridostigmine was not significantly correlated withGAD antibodies, which (if anything) would be expected todecrease GH secretion via reduced GABA-ergic stimulationin the human (above). Thus, overall data favor the thesis thatdecreased somatostatinergic tone accounts primarily for el-evated pulsatile GH secretion and exaggerated GH release inresponse to various pharmacological stimuli in type I dia-betic individuals (718). Withdrawal of IGF-I’s negative feed-back actions also likely augments GH secretion in these cat-abolic patients. However, serum IGF-I levels fail to correlatewith lack of responsiveness to pyridostigmine, thus suggest-ing that additional pathophysiological mechanisms may alsooperate (255).
The pathophysiology of the presumptively altered soma-tostatin neuroregulation in type I diabetes mellitus has beenstudied traditionally in three venues: 1) role of metaboliccontrol, 2) neuroendocrine pathways, and 3) altered IGF-Iand BP feedback signaling. In patients with insulin-depen-dent diabetes mellitus, there is an increase in both the am-plitude and frequency of pulsatile GH secretion comparedwith normal subjects (707, 708), which is not affected bymaintenance of overnight normoglycemia. Type I diabeticswith poor metabolic control have significant increases in GH
peak maxima, incremental amplitudes, and pulse durations,when compared with a period of better control (654). Mean24-h GH secretion decreased significantly during improved(insulin-treated) glycemic control, although the fraction ofpulsatile GH secreted per 24 h did not change significantly(719). In contrast, intraportal delivery of insulin nearly nor-malizes the GH/IGF-I/IGFBP axis in recent-onset type Idiabetes mellitus (720).
Chronic subcutaneous recombinant human (rh) IGF-I ad-ministration to children with insulin-dependent diabetesmellitus corrects many of the preexisting abnormalities in theGH/IGF/IGFBP axis. Specifically, 28 days of rhIGF-I therapyto overcome endogenous IGF-I deficiency suppresses bothserum IGFBP-1 and GH concentrations (721). Studies byCheetham et al. (722) further demonstrate that a single sub-cutaneous injection of rhIGF-I increases serum IGF-I levels,decreases overnight GH secretion, and reduces insulin re-quirements in adolescent type I diabetics (n 5 9). Similarly,Bach et al. (723) found that 10-h continuous subcutaneousinfusions of rhIGF-I given on three successive days to eachof four diabetic adolescents increased serum IGF-I, reducedIGF-II, and suppressed GH concentrations (724). Thus, weinfer that feedback responsiveness to IGF-I is preserved intype I diabetes mellitus with GH hypersecretion, and thatIGF-I deficiency contributes to the GH hypersecretory statevia feedback withdrawal. Practically, compared with insulintreatment in type I diabetic children, dual injections of insulinand rhIGF-I tended to lower GH and increase IGFBP-1 con-centrations while improving glycemic control, more thaninsulin treatment alone during 1–4 weeks of therapy (698).
Finally, although not fully established, in principle, sup-pression of endogenous GH secretion with somatostatin an-alogs might ameliorate renal hyperfiltration, an early featureof diabetic nephropathy linked indirectly to GH hypersecre-
FIG. 12. Negative relationship between 24-h mean serum GH con-centration and intraabdominal fat mass as determined by comput-erized axial tomographic scanning of the abdomen, in a cohort ofhealthy middle-aged men and women. GH concentrations were de-termined by 20-min blood sampling for 24 h and subsequent assay byimmunofluorometry. The solid circles denote male subjects, and theopen circles denote females. The regression line shows a stronglynegative relationship between the natural logarithm of intraabdomi-nal adiposity and daily GH secretory activity in both men and women.In multiple linear regression analyses, intraabdominal fat mass ac-counted for the majority of the variability in integrated serum GHconcentrations, exceeding that due to age and gender in this popu-lation [Redrawn with permission from N. Vahl et al.: Am J Physiol272:E1108–E1116, 1997 (750).]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 747
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

tion in type I diabetes, since GH treatment itself tends topromote renal blood flow (725–729).
d. Type II diabetic patients. Clinical investigations of GHsecretion in patients with type II diabetes have yielded con-flicting inferences. Spontaneous GH secretion may be in-creased, normal, or decreased (730–733), and secretagogue-stimulated GH release also may be augmented (734), normal(735), or reduced (654, 655, 736, 737). Many studies of GHsecretion in type II diabetic patients are confounded signif-icantly by major covariates and codeterminants, such as ageand body composition, which profoundly influence GH se-cretion. GH secretion decreases markedly with age and isstrongly inversely related to the degree of obesity (93, 119,738–740) (e.g. see Fig. 12), both of which factors are operativein diabetic (and control) populations.
There is a consensus that obese type II diabetic patients(736, 741), as well as nondiabetic obese subjects (312, 742),have an impaired GH-secretory response to GHRH stimu-lation compared with healthy nondiabetic normal-weightvolunteers. Impaired glucose tolerance and hyperglycemia,in addition to obesity, are relevant pathophysiologically,since even lean type II diabetics exhibit an attenuated GHresponse to GHRH (736). Further clinical investigations in-dicate that pyridostigmine is not able to normalize the GHresponse to GHRH in either lean or obese type II diabeticpatients, unlike its facilitative action in normal subjects (655).Therefore, we hypothesize that chronic hyperglycemia, tis-sue resistance to insulin, and/or elevated circulating insulinlevels in type II diabetics decrease GH secretion by increasinghypothalamic somatostatin release (743). These consider-ations are supported by data obtained in normal subjectsstudied with the oral glucose tolerance test, wherein acutehyperglycemia-mediated GH inhibition can be overcome bypyridostigmine (693), and by studies in the rat showing di-rect pituitary inhibitory actions of insulin or insulinomimet-ics on GH secretion (62, 744). On the other hand, in thehuman, fasting GH hypersecretion is rapidly suppressed bylow-dose IGF-I but not insulin infusion (64). IGF-I also in-hibits pituitary GH secretion in the sheep (63).
An informative recent study demonstrated that sponta-neous GH secretion in type II diabetes mellitus is influencedby two independent factors acting in opposite directions: themarked inhibitory effect of obesity on GH-secretory burstamplitude or mass (119, 733, 740), and the apparent stimu-latory effect of the diabetic state on GH burst frequency (693,708, 743). The observed suppression of GH burst mass inproportion to the degree of obesity in diabetic patients il-lustrates the importance of body composition in the regula-tion of GH secretion in type II diabetes mellitus. In contrastto the reduced GH-secretory burst mass (733), the apparentincrease in GH-secretory burst frequency in the type II di-abetic patients is not explained by obesity. Augmented GHpulse frequency also occurs in adult and adolescent patientswith type I diabetes mellitus, where it leads to an increasedoverall GH production rate (707, 708). Thus, a disruption ofthe hypothalamo-pituitary interplay of GHRH and soma-tostatin that regulates pulsatile GH release may be associatedwith the diabetic state, whether type I or type II. In short,chronic hyperglycemia may elicit variable alterations in pi-tuitary GH secretion in type II diabetics depending on the
degree of obesity (743), viz., on the one extreme, the GH-hypersecretory pattern of type I diabetics (lean young sub-jects with low circulating insulin levels, and frankly elevatedblood glucose) and, on the other extreme, the GH hypose-cretory state of pure obesity (older patients with hyperin-sulinemia and milder hyperglycemia) (693, 708, 733). Thisconcept may explicate many disparities in reported GH axisactivity in type II diabetic patients.
B. Leptin and FFA
In the hyperphagic leptin-deficient (ob/ob) mouse, eithercentral or peripheral infusion of leptin induces weight lossand satiety by reducing hypothalamic NPY levels (745–748).Thus, in the rodent, NPY is one likely link among obesity,eating, and leptin feedback.
The exact metabolic mechanisms subserving reduced GHsecretion in relative adiposity or absolute obesity are notknown (312, 742). Relative hyposomatotropinemia appearsto correlate with total and especially visceral (intraabdomi-nal) fat (121, 749) (Fig. 11). Indeed, visceral fat mass as es-timated by computed tomography scanning is the primary(negative) statistical determinant of GH secretion in middle-aged men and women and accounts for both the gender andage differences in GH secretion in this context (750). Theprecise neuroregulatory basis for the inverse relationshipbetween body fat mass and GH secretion is not fully under-stood, but the recent discovery of a polypeptide, leptin, se-creted by fat cells allows several plausible hypotheses (751).GH secretion in postmenopausal women (fed) as well asyoung midluteal phase women (fed and fasted) is inverselycorrelated with serum leptin concentrations (150, 524) (seeFig. 10). Although direct hypothalamic actions of leptin onsomatostatin and/or GHRH release are imperfectly defined,leptin stimulates GH secretion in the fasted adult male rat(522). Akin to other metabolic antitheses in the rat and hu-man, an opposite action of leptin would be expected in thehuman, viz., to inhibit GH release. Alternatively, serum lep-tin may be a marker of acute nutrient intake (albeit notmacronutrient composition) and longer-term fat depots inthe human, and not a direct regulator of the GH axis (150, 523,524, 752, 753). Thus, rh GH treatment decreases serum leptinconcentrations in proportion to fat loss per se in GH-deficientadults (753).
Full-length leptin receptors are present in rat hypotha-lamic neurons, and leptin administration in the rodentinhibits the expression of the potent orexigenic and GH-suppressing peptide, NPY (748, 754, 755). In addition, leptinadvances puberty in mice by stimulating the hypothalamicneuronal secretion of GnRH and overcomes the suppressiveeffects of fasting on the reproductive axis in the same manner(756, 757). Leptin also acts on the thyroidal axis by stimu-lating TRH-secreting neurons in the paraventricular nucleus(758). Accordingly, leptin, which can enter brain via a sat-urable system (759), can act on multiple hypothalamic neu-rons (754, 760, 761), and may thus directly or indirectly mod-ify somatostatin and/or GHRH release, thereby mediating ormodulating the apparent negative-feedback effects of in-creased body fat on the GH axis in the rodent. However,other (nonleptin) hypothalamically targeted regulators
748 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

might also be released by adipose or other tissues and con-tribute relevant long-term feedback information concerningbody composition. Indeed, the decrease in IGFBP-1 levels inobesity, with possible relative increases in free IGF-I, hasbeen postulated to suppress GH in the human (762), andpossibly exacerbate obesity, whereas IGF-I (or GH) treatmentwill reduce body fat content (763).
The clinical pathophysiology of leptin secretion and thenature of leptin actions in the human are being activelyexplored. From a neuroregulatory perspective, leptin issecreted via both circadian and pulsatile rhythms andachieves higher serum concentrations in women than men(764, 765). Leptin is likely to interface with the GH axis viaGHRH-somatostatin neurons and other putative hypotha-lamic modulators, e.g., NPY-expressing neurons, GnRHneurons, and possibly TRH neurons (748, 754, 756, 760,761). Central actions of leptin are plausible in view of thesaturable transport of this peptide into the brain (759).Since the leptin level is a strongly positive correlate of fatcell mass, and full-length leptin receptors are expressed inthe hypothalamus (above), the observation is not surpris-ing that treatment for 1 yr (but not 1 month) of adultGH-deficient adults with rh GH reduces serum leptin con-centrations significantly (from 16 6 3 to 11 6 2 mg/liter)in direct proportion to the reduction in total percentagebody fat (r 5 0.763, P 5 0.002) (753). In the presence ofconcomitant glucocorticoid treatment, GH treatment for 1week appears to increase serum leptin concentrations fur-ther (above that stimulated by methylprednisolone alone)(766). These preliminary sallies into leptin pathophysiol-ogy indicate the need for further clinical studies to clarifyspecific mechanistic interactions among GH, leptin, andvarious modulators of leptin production.
FFA may well participate in the regulation of pituitaryGH secretion in the human and sheep, since GH secretionis stimulated when plasma FFA levels are reduced,whereas conversely increased FFA levels block GH secre-tion provoked by virtually all stimuli (767–769). For ex-ample, the drug acipimox lowers plasma concentrations ofFFA by blocking their release from adipose tissue (770) andconcomitantly enhances basal GH secretion (771) and theGH-secretory response to GHRH in normal or obese sub-jects (772). Acipimox also potentiates the GH response toGHRH after pyridostigmine pretreatment (773). The syn-ergistic action of acipimox and pyridostigmine can beexplained by hypothesizing a hypothalamic site of actionof the cholinergic agonist (262), and a direct pituitary-inhibitory effect of FFA (774). This concept is supported byin vitro studies showing that fatty acids inhibit basal (775)and GHRH-stimulated (776) GH secretion from culturedrat somatotropes. In obesity, acipimox treatment virtuallynormalizes GH secretion, pointing to an important role ofrelatively increased FFA flux in this relatively hyposoma-totropic state (771). In addition, reportedly elevated freeIGF-I levels (that accompany insulin-driven IGFBP-1 re-ductions) in human obesity may suppress pituitary GHsecretion directly (777), as suggested in the rat in vitro, andthe human and sheep in vivo (62– 64).
C. Amino acids
l-Arginine is an essential amino acid and a well knownpotent GH secretagogue in man (778). In the rat, amino acidsdo not (or less overtly) stimulate GH secretion (563). Themost plausible mechanism through which l-arginine stim-ulates GH secretion is somatostatin withdrawal. Argininedoes not influence either basal or GHRH-induced GH se-cretion from rat anterior pituitary cells in vitro (779), whichspeaks against a direct effect of l-arginine at the somatotropelevel. This interpretation is also concordant with the obser-vation that patients with idiopathic GH deficiency or pitu-itary dwarfism, which is often due to a lack of endogenousGHRH (780), have no GH rise after l-arginine infusion. Inman, several investigators have shown that l-arginine en-hances the GH response to GHRH (779, 781). Since theseexperiments used a maximal dose of GHRH combined withl-arginine, one can infer that this amino acid stimulates GHsecretion by inhibiting endogenous somatostatin, rather thanby promoting endogenous GHRH release. This inference isin agreement with data showing that l-arginine infusionafter an earlier (desensitizing) dose of GHRH will still inducea pronounced GH rise (171). Moreover, l-arginine fails toaugment the GH response to pyridostigmine (781), which isbelieved to act in part via somatostatin withdrawal (above).
The hypothesis that l-arginine suppresses endogenoussomatostatin release is consonant with the ability of l-argi-nine, as well as pyridostigmine (782), to amplify the TSHresponse to TRH (which is at least partially under inhibitorysomatostatin control) in normal humans (779). Furthermore,administration of l-arginine normalizes the GH response toGHRH in patients with nonendocrine diseases receivingchronic immunosuppressive glucocorticoid therapy (783).Indeed, the l-arginine-induced enhancement of the GH re-sponse to GHRH is greater in glucocorticoid-treated thannormal subjects. The ability of l-arginine to reverse glucocor-ticoid inhibition (see below) strengthens the hypothesis thatthis amino acid decreases somatostatin release by the hypo-thalamus. Whether l-arginine, a precursor in the nitric oxide-signaling pathway, works via nitric oxide to stimulate GHrelease in the human is unknown, but seems unlikely basedon a recent clinical study (668).
VII. Other Hormonal Regulators of the GH Axis
Given the relevance of GH in controlling body growth andintermediary metabolism, not surprisingly the secretedproducts of several endocrine glands in turn regulate GHsecretion. The mechanisms underlying these feedback inter-actions and the specific effects of the most important of thesemetabolic effectors on GH secretion are being elucidated. Inthis section, we review the roles of glucocorticoids, gonadalsex hormones, and thyroid hormone in GH neuroregulation.
A. Glucocorticoids
1. In vitro studies. Combined immunocytochemical (below)and mRNA data demonstrate glucocorticoid suppression ofGHRH message and peptide synthesis, which could accountin part for glucocorticoid’s inhibition of GH secretion (784).
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 749
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

As reviewed in part earlier (171, 550), studies from at leastthree laboratories indicate that hypothalamic GHRH mRNAlevels are decreased by chronic glucocorticoid treatment inthe rat (785–787). Glucocorticoids likely significantly inhibitGHRH gene transcription in the arcuate nucleus, plausiblyby acting via a known glucocorticoid-responsive element inthe 59-upstream promoter of the GHRH gene (785). Thisregion of the promoter may mediate repression of GHRHgene transcription by an activated glucocorticoid receptor.Inhibition of GHRH gene expression is dependent on thedose and duration of glucocorticoid treatment and occurseven after pituitary removal (787), which indicates that GHfeedback per se is not required for these brain actions ofglucocorticoid.
Immunologically based studies have shown that totalhypothalamic GHRH peptide content falls in glucocorti-coid-treated rats compared with controls (788). Similarly,Fernandez-Vasquez et al. (789) reported that treating rat hy-pothalamic cells in vitro with high doses of corticosteronedecreased neuronal GHRH release. Other recent immuno-cytochemical data define a reduction in optical density andpercentage area of immunostaining for GHRH only in therostral region of the median eminence of the hypothalamusin glucocorticoid-exposed rats (784). In vitro, corticosteronehas dual effects on hypothalamic neuronal GHRH peptideexpression, i.e., concentrations in the range of the glucocor-ticoid receptor Kd (3 nm) increase GHRH content, whereashigher concentrations (30 and 300 nm) decrease GHRH con-tent in GHRH-secreting cells (789). Biphasic regulation (low-dose stimulation, and higher-dose inhibition) of GH secre-tion by synthetic glucocorticoids is also recognized in thehuman in vivo (below).
Available investigations of glucocorticoid effects on hy-pothalamic somatostatin mRNA accumulation in the rat areconflicting. Nakagawa et al. (790) observed that high-doseglucocorticoid administration (330 mg/day) for 3 days raisedhypothalamic somatostatin mRNA content in adult femalerats. Conversely, Lam et al. (791) found that more prolonged(10 days) administration of dexamethasone in the drinkingwater at a concentration of 5 mg/ml slightly reduced soma-tostatin mRNA content in prepubertal male rats. The latterresults most likely reflect the lower dose, different route ofdelivery, and/or animal age and sex used in that study. Abipotential effect of glucocorticoids on hypothalamic soma-tostatin mRNA levels was observed recently, i.e., a significantincrease in specific somatostatin transcripts in the periven-tricular nucleus after 3 days of dexamethasone treatment (1mg/kg/day), but a reduction after 8 days for all glucocor-ticoid doses tested (786).
The effects of glucocorticoid treatment on the soma-tostatinergic system are tissue specific and apply to bothsomatostatin peptide and mRNA content (792). The rise inhypothalamic somatostatin content due to glucocorticoidsseems to reflect an increase in transcription of the soma-tostatin gene (784). RNAse protection assay also revealed asecond lower molecular weight somatostatin gene-transcrip-tion product in glucocorticoid-treated rats, suggesting pos-sible control of somatostatin gene expression both quantita-tively and qualitatively.
In vitro incubation of fetal hypothalamic cells with corti-
costerone increases the content and release of immunoreac-tive somatostatin (789). Using immunohistochemical tech-niques, others reported in vivo increases in somatostatincontent in the rostral, middle, and caudal regions of themedian eminence of the rat hypothalamus after glucocorti-coid treatment (784). In recent experiments, freely movinganimals received a stereotaxic implant of a push-pull cannulainto the median eminence for 10 days, which was then per-fused with artificial cerebrospinal fluid for 120–150 min. Anintraperitoneal injection of dexamethasone (200 or 300 mg/100 g) induced a mean increase in hypothalamic somatostatinoutput of 63 6 6.2% above basal 15–30 min later (793). Thus,glucocorticoids seem to stimulate hypothalamic somatosta-tin production, storage, and release. Conversely, glucocor-ticoids appear to repress somatostatin receptor expression, atleast by GH-secreting tumoral cell lines, e.g., dexamethasoneinhibits mRNA expression of several somatostatin receptorsubtypes in rat GH4C1 cells (794), whereas sex steroids op-pose this action (below). Thus, multiple actions of glucocor-ticoids are likely in the hypothalamus as well as in the pi-tuitary gland.
Chronic glucocorticoid treatment increases pituitary GHmRNA content in the adult male rat, as recently assessed byRNAse protection assays. This stimulation may reflect directpituitary actions of glucocorticoids to increase GH gene tran-scription (795). Such an interpretation is consistent with thereport of Slater et al. (796), showing the existence of a glu-cocorticoid-regulatory element in the human GH gene pro-moter.
Glucocorticoids can stimulate GH synthesis and secretionin vitro, e.g., by somatotropes obtained from human andnormal monkey pituitaries (797). Similar effects are observedin studies of normal rat pituitaries (798) and pituitary clonalcell lines (799). Enhanced GH secretion occurs after severaldays of incubation and is magnified when the cells are firstcultured for long periods without glucocorticoids. The glu-cocorticoid-induced increase in pituitary GH secretion isabolished by cycloheximide or actinomycin D pretreatment,suggesting requirements for both protein and RNA synthesisin this GH-stimulating effect of glucocorticoid (800).
Glucocorticoids can also increase the GH-secretory re-sponse to GHRH stimulation in vitro. Pretreatment of ratpituitary cells for 24 h with the potent synthetic glucocorti-coid, dexamethasone, increases the GH response to GHRHmore than 5-fold (798). Facilitation of GHRH action by glu-cocorticoids is time dependent; e.g., when pituitary cells areexposed to dexamethasone for less than 4 h, GHRH andforskolin-induced GH release declines, but after 18 h, basaland GHRH-elicited GH secretion rises.
A recent study examined the effects of glucocorticoid onGHRH receptor gene expression in adult male rat anteriorpituitary cells in primary cell culture using a highly sensitiveand quantitative reverse-transcribed PCR methodology(801). Dexamethasone (5 nm) significantly increased GHRHreceptor mRNA levels after 6- and 24 h-incubations, with amaximal effect at 25 nm. The glucocorticoid receptor-specificantagonist, RU 486, completely eliminated dexamethasone’senhancement of GHRH receptor mRNA expression (801).Hydrocortisone also stimulated a marked dose-dependentincrease in GHRH-receptor mRNA (802).
750 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

Dexamethasone increases GH receptor mRNA expressionin the growth plate of long bones and in the liver, targettissues important for body growth, but not in kidney. Glu-cocorticoid also induces liver GH-binding protein expres-sion, but with a biphasic dose-response (803). In other tissues,dexamethasone directly antagonizes GH action at the cellu-lar level; e.g., glucocorticoid inhibits the ability of GH to elicitseveral early events in GH signaling in 3T3-F442A fibroblasts(804).
In conclusion, a plethora of competing actions of glucocor-ticoids are exerted in the hypothalamus, the pituitary gland,and in different peripheral tissues. The complex interplayamong these multiple opposing actions of glucocorticoiddetermines the net in vivo effect of the corticosteroid in theintact individual. Final responses are complicated by thebipotential dose and time dependencies of glucocorticoidactions on the GH axis. Our overall unifying working hy-pothesis is that glucocorticoids are both inhibitors and stim-ulators of regulated GH secretion and action.
2. In vivo studies. Adrenalectomy significantly decreases theGH-secretory response to a submaximal dose of GHRH in therat (805). Since this attenuation is reversed by dexamethasonereplacement, glucocorticoids likely maintain pituitary sen-sitivity to GHRH and/or the releasable pool of GH. In favorof the former, adrenalectomy results in a significant decreasein pituitary GHRH-binding capacity (239). On the otherhand, and typical of bipotential actions of glucocorticoids,the GH response to a physiological dose of GHRH is signif-icantly lower in animals treated with glucocorticoids eithersubacutely or chronically. In contrast, cortisol infusion intocastrated male Suffolk sheep (1 to 1.5 yr old) acutely atten-uates GHRH-stimulated GH release in this species (806).
Nakagawa et al. (788) reported that chronic (several-day)dexamethasone treatment suppresses serum GH concentra-tions, increases hypothalamic somatostatin concentrations,and decreases hypothalamic GHRH concentrations in con-scious female rats. In male rats, glucocorticoids reduce andprolong trough GH values (807). When GH pulses occur,however, they are significantly higher than those seen innormal animals. This resembles a more masculine-like GHrelease pattern. According to the model of reciprocally cou-pled GHRH and somatostatin neuronal systems in the rodent(265), and the ability of somatostatin withdrawal to evokerebound GH release (54, 157, 160), the foregoing dexameth-asone-induced patterns of pulsatile GH release (807, 808)would suggest increased somatostatin tone induced by dexa-methasone, with or without augmented endogenous GHRHrelease (and/or pituitary actions) during somatostatin with-drawal episodes.
Given that deep pentobarbital anesthesia can suppress therelease of several hypothalamic hormones, e.g., somatostatin(563, 809), the dichotomous effects of glucocorticoids in anes-thetized vs. conscious rats suggest multisite actions of glu-cocorticoids, including at hypothalamic loci. A unifying in-terpretation is that at the hypothalamic level glucocorticoidsreduce GHRH and increase somatostatin tone, thus promot-ing inhibition of GH secretion; in contrast, at the pituitarylevel, corticosteroids enhance GH synthesis and the GH re-sponse to GHRH (e.g., via heightened GHRH receptor ex-
pression), thus favoring a stimulatory response (Fig. 13). Insupport of this proposed thesis, experiments in conscious,freely moving rats receiving chronic (4-day) dexamethasonetreatment show a diminished GH-secretory response to anear-physiological dose of GHRH compared with saline-treated rats. However, passive immunization with soma-tostatin antiserum completely alleviated the attenuated GHresponse; indeed, somatostatin-immunoneutralized glu-cocorticoid-treated rats had a significantly higher GH re-sponse to GHRH than control animals (810). These experi-ments further corroborate the notion that glucocorticoidsincrease somatostatin’s inhibitory activity. In this regard,somatostatin antiserum partially counteracts glucocorti-coid’s inhibition of body growth in the rat (811).
Other experiments reveal that selected pharmacologicalmanipulations can interfere with glucocorticoid’s inhibitionof GH secretion in the freely moving conscious rat. Sub-stances believed to decrease hypothalamic somatostatin re-lease (such as pyridostigmine), but not clonidine (which, inthe rat, mainly stimulates GHRH release), partially or com-pletely relieve the inhibitory effects of glucocorticoids on GHsecretion (550, 812).
In recent experiments, three doses of GHRP (1 mg, 4 mg,and 25 mg/kg) and one dose of GHRH (500 ng/kg) wereevaluated alone and together in the glucocorticoid-treatedrat. These studies revealed that GHRP-6 alone increasesplasma GH concentrations in a dose-dependent fashion bothin vehicle- and dexamethasone-treated rats, whereas the GHresponses to GHRP-6 and GHRH are significantly reducedselectively in dexamethasone-pretreated animals. Thus,GHRP-6 is able to stimulate GH secretion in glucocorticoid-treated rats, but only partially overcomes glucocorticoid-induced inhibition of GHRH-stimulated GH secretion (812)(see Fig. 13). This observation may indicate that GHRP onlypartially reverses somatostatin excess induced by glucocor-
FIG. 13. Schematic representation of the authors’ concept of patho-physiological mechanisms of the biphasic dose-dependent effects ofglucocorticoids on the somatotropic axis. Smaller (physiological)amounts of cortisol are required to support pituitary GH gene tran-scription and maintain the GHRH receptor, whereas excessive glu-cocorticoid suppresses GH secretion via augmenting somatostatinrelease, and reducing GHRH secretion, as inferred based on data inthe rat.
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 751
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

ticoids and/or that glucocorticoids inhibit via mechanismsthat do not interface with the GHRP stimulus pathway.
Galanin increases serum GH similarly in both vehicle- anddexamethasone-treated rats. The response to galanin plusGHRH is similar to that of galanin plus saline in vehicle-treated rats, but is significantly enhanced in rats chronicallytreated with dexamethasone. We infer that galanin-mediatedGH release in rats may involve both increased GHRH se-cretion and possibly reciprocally reduced activity of soma-tostatinergic pathways and thereby oppose the inhibitoryeffect of glucocorticoids (813).
Overall, available data are consonant with the hypothesisthat in vivo inhibition of GH secretion by glucocorticoidsoccurs via increased somatostatin release in the rodent, withor without a concurrent (e.g., somatostatin-mediated) declinein GHRH release.
3. Human studies. The impact of untreated primary adrenalfailure (Addison’s disease) on GH secretion in humans isinadequately studied owing to the clinical severity of thisdisease and the urgent need to start glucocorticoid replace-ment promptly. On the other hand, there are more reportsconcerning pituitary function in patients with isolated glu-cocorticoid deficiency secondary to idiopathic ACTH defi-ciency. The first female patient with this rare autoimmunedisease was described in 1988 and exhibited impaired GH-secretory responses to several different pharmacologicalstimuli (l-arginine and insulin). Stimulated GH release wasrestored fully during glucocorticoid replacement therapy(814). Glucocorticoid-remediable reduction in GH secretory-reserve was confirmed in several other (usually female) pa-tients with ACTH deficiency (815) and also in some men(816). In addition, when glucocorticoid treatment is sus-pended in Addisonian patients, there is a progressive loss ofthe GH-secretory response to a GHRH stimulus (817), whichis reversed in a dose-dependent manner by cortisol infusions(818). On the other hand, acute withdrawal of replacementglucocorticoid with resultant prompt biochemical hypocor-tisolemia (cortisol ,55 nmol/liter) does not impair the GH-secretory response to GHRH compared with that observedduring eucortisolemia (serum cortisol, 280–420 nmol/liter)in the same patients. On the basis of these clinical studies, wehypothesize that the impaired GH response to GHRH in thehuman with subacute or chronic adrenal insufficiency is aconsequence of sustained low circulating cortisol concentra-tions. Therefore, physiological amounts of glucocorticoidsprobably play an important long-term role in maintainingbasal somatotrope secretory reserve in humans, as well as therat. Accordingly, small amounts of exogenous synthetic glu-cocorticoids (approximately twice the basal daily cortisolsecretion rate) stimulate pulsatile GH secretion over severaldays in the healthy adult (819), and the serum cortisol con-centrations correlate positively with GH release in fastingwomen (524).
In contrast to the foregoing circumstances of primary orsecondary Addison’s disease, the immediate and short-termeffects of exogenous glucocorticoids in normal humans aresuppression of GH secretion. Hydrocortisone inhibits theGH-secretory response to GHRH within 1 h of its adminis-tration (462), and the GH response is still inhibited 8 h after
treatment (820). The rapid (1-h) inhibitory effect is similar tothat observed in animals (806) and may be somatostatindependent, since pretreatment of the subjects with the cho-linergic agonist, pyridostigmine, an agent thought in part todecrease hypothalamic somatostatin release, relieves glu-cocorticoid’s inhibition (464). In contrast to the effect of cor-tisol per se, treatment with the potent synthetic glucocorti-coid, dexamethasone, for 3 or 4 h enhances both basal andGHRH-stimulated GH secretion (821, 822). Although themechanisms involved in this transient stimulatory responseare not well understood, one possibility is that glucocorticoidtreatment decreases serum IGF bioactivity, perhaps throughpathways involving the IGFBPs, e.g., BP-1, which is rapidlyinsulin-sensitive (823). This would result in decreased freeIGF-I-negative feedback of GH secretion. Alternatively, onecan hypothesize that temporally biphasic somatostatin re-lease may occur after acute dexamethasone administration,and therefore a reciprocally biphasic pattern of GH release.Curiously, 8 h after the administration of dexamethasone, theGH response to GHRH is inhibited, apparently also at leastin part, via a somatostatinergic mechanism (820). Inhibitionof the GH response to GHRH continues after 4 days of ad-ministration of prednisone in normal subjects (824). Thus,acute administration of natural glucocorticoids may stimu-late a rapid burst of somatostatin release. Synthetic glucocor-ticoid may promote rapid rebound GHRH (and GH release)after spontaneous somatostatin withdrawal or act directly tofacilitate pituitary GH gene transcription. A later tonic hy-persomatostatinergic state may follow in the case of long-acting glucocorticoids, such as dexamethasone and pred-nisone. Overall, available clinical data thus indicate thatglucocorticoid administration can stimulate or inhibit GHrelease in a dose- and time-dependent manner, and in anagent-specific fashion, presumably through alterations bothin somatotrope responsiveness to GHRH and in hypotha-lamic somatostatin release.
The chronic effects of glucocorticoids in humans are rec-ognized in patients with spontaneous hypercortisolism(Cushing’s syndrome) and in patients undergoing high-doselong-term immunosuppressive glucocorticoid treatment.The increases in serum GH concentrations in response tocommonly used pharmacological stimuli, such as insulin-induced hypoglycemia, l-arginine, lysine-vasopressin, andGHRH, as well as spontaneous sleep-associated GH secre-tion, all are typically inhibited in patients with significantendogenous hypercortisolism (825–827). In 14 patients withCushing’s disease, blood sampling was performed every 20min over 24 h for later determination of serum GH profilesbefore and 10–11 days and 3, 6, and 12 months after neu-rosurgical cure. Before intervention, Cushing’s disease pa-tients manifested markedly decreased mean 24-h serum GHconcentrations, GH peak heights, and GH peak areas, but GHpulse frequency was similar to values in controls (828). Sev-eral clinical studies have linked this suppression of GH re-lease to hypercortisolemia, since GH secretion tends to nor-malize after the removal of the source of cortisol excess (829).However, surprisingly, a pattern of GH suppression similarto that observed in patients with acute hypercortisolism wasalso evident in Cushing’s disease patients who were studied10–11 days and 3, 6, and 12 months after cure. The basis for
752 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

long-term suppression of the GH axis is not known. In ad-dition, more recent clinical studies have unmasked a moredisorderly pattern of 24-h GH release in Cushing’s disease(830), which on theoretical grounds points to a defect infeedback control of the GH axis (144).
Recent clinical experiments have focused further on theneuroendocrine mechanisms involved in GH axis suppres-sion in Cushing’s disease. Pyridostigmine, used in an at-tempt to decrease hypothalamic somatostatin release, aug-ments, but does not restore fully, GH release in response toGHRH stimulation in patients with Cushing’s disease (831,832). Thus, either this drug only partially opposes the so-matostatin excess presumed to operate in Cushing’s disease,or the inferred glucocorticoid-mediated increase in hypo-thalamic somatostatin tone may not be the sole factor inhib-iting GH secretion in these patients. Indeed, increased IGF-Ifeedback inhibition of the GH axis cannot be excluded, sincein patients with Cushing’s syndrome the marked suppres-sion of endogenous GH secretion is accompanied by normalGHBP, normal to low IGF-I, and low BP-1 and BP-3. Suchbiochemical findings may suggest enhanced tissue sensitiv-ity to GH (high GHBP despite low GH secretion), and normalor increased free IGF-I bioavailability (833). Other relevantcontributing factors in Cushing’s disease could include ele-vated blood glucose and FFA concentrations, and/or obesity(see above), all of which are known to suppress GH secretionin the human. Indeed, obesity also may be marked by in-creased (measurable) free IGF-I concentrations (777) withresultant feedback suppression of pituitary GH secretiondirectly [see Section VI.B above].
The type of glucocorticoid used also may influence theresponse of the GH axis. For example, prednisone bluntedGH release more than deflazacort during an insulin tolerancetest in two matched groups of subjects given nominallyequivalent doses of the two glucocorticoids (834). In otherexperiments, propanolol (40 mg), clonidine (0.3 mg), or pyri-dostigmine (120 mg) was administered orally followed byintravenous GHRH infusion 60 min later with or without aprior nocturnal dose of dexamethasone (8 mg orally). Bothpropanolol and pyridostigmine were able to reverse (par-tially) the inhibitory effect of dexamethasone on GHRH-elicited GH release. These data suggest that the inhibitoryeffect of glucocorticoid excess on GH release is due to in-creased hypothalamic somatostatin secretion, which appearsto be dependent on dexamethasone-enhanced b2-adrenergicactivity or glucocorticoid-suppressed cholinergic tone (835).However, direct pituitary effects are suggested by other stud-ies showing an inhibitory effect of an acute intravenous in-fusion of hydrocortisone (cortisol) on baseline GH release inacromegaly (836). Moreover, bolus intravenous injection ofhydrocortisone hemisuccinate, 100 mg, followed by intrave-nous continuous infusion of 250 mg over 120 min, in acro-megalic patients, blunted the GH-secretory response to thephysiological stimulus GHRH (837) and/or reduced the par-adoxical GH response to TRH (484). In counterpoint, whenacromegalic patients are pretreated with the functional so-matostatin antagonist, l-arginine, acute hydrocortisone in-fusion does not suppress baseline serum GH concentrations,which favors an hypothesis of a cortisol-stimulated soma-tostatin-mediated inhibition of baseline GH release (838).
Other confounding factors in glucocorticoid-treated patients,such as residual organ system disease in posttransplantationuse of glucocorticoids for immunosuppression (839), must beconsidered in interpreting clinical studies.
Overall, available clinical data thus indicate that acute orshort-term glucocorticoid exposure may either inhibit orstimulate GH release in normal subjects or acromegalic pa-tients. In our view, such effects likely occur via several mech-anisms, i.e., namely, changes in somatostatin release, whichmay in turn reflect direct hypothalamic actions of glucocor-ticoids (see above), changes in hypothalamic neurotransmis-sion, and/or altered peripheral feedback signals. Further-more, the magnitude and direction of glucocorticoid effectsare influenced by the nature, potency, and half-life of theparticular glucocorticoid administered, its route of admin-istration, and the time of observation.
Numerous studies have examined GH secretion in pre-pubertal children undergoing long-term immunosuppres-sive glucocorticoid treatment (552, 840). In these children,GH responses to various pharmacological stimuli are re-duced, as expected from the adult paradigm. Similarly, spon-taneous GH secretion is decreased (552). Pyridostigmine sig-nificantly enhances both GHRH and sleep-induced GHrelease in children, but such actions are reduced during long-term glucocorticoid treatment. We note that acetylcholines-terase inhibitors may also act via stimulating GHRH secre-tion, the action of which is impaired during chronicglucocorticoid treatment (552). Thus, attenuated pyridostig-mine actions are consistent with, but not proof of, soma-tostatin’s involvement in glucocorticoid inhibition of GHsecretion in children. Indeed, since neostigmine also releasesGHRH into the sheep hypothalamo-pituitary portal circula-tion (29), and a GHRH antagonist blocks pyridostigmine’saction in the human (269), pyridostigmine’s limited efficacyin chronic glucocorticoid therapy could also reflect restrictedGHRH release and hence reduced stimulation of GH secre-tion by pyridostigmine.
GH secretion in adults receiving chronic immunosuppres-sive therapy with glucocorticoids is also impoverished. Phar-macological agents such as clonidine and galanin, which arepresumed to effect GH release in part via increased hypo-thalamic GHRH release, are less effective in enhancing base-line GH concentrations and facilitating GHRH-induced GHrelease in glucocorticoid-suppressed adults compared withcontrols (441). On the other hand, l-arginine and hexarelinor GHRP-6, which are thought to serve as GH secretagoguesby acting in part as so-called functional somatostatin antag-onists, virtually normalize the GH-secretory response toGHRH and galanin in adults treated with glucocorticoids(783).
Although the natural ligand for the GHRP receptor has notbeen identified, patients with Cushing’s disease showblunted GH release to the otherwise synergistic stimuli ofGHRH and GHRP (334), as reported in the glucocorticoid-treated rat (above). Whereas endogenous hypercortisolismblocks both GHRH and GHRP-6 pathways, exogenous glu-cocorticoids do not always impair GHRP-6- releasing mech-anisms (841). The latter inference was confirmed in otherpatients undergoing long-term (no fewer than 6 months)immunosuppressive glucocorticoid treatment for nonendo-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 753
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

crine disease, who exhibited unimpeded GH release after anintravenous injection of GHRP or hexareliln (without or withGHRH). The suppressive influence on GH secretion of pred-nisolone is reversed by a high dose of L-692,429 (MK0677),a nonpeptidyl mimic of GHRP-6 (842). Thus, the balance ofevidence in the human favors the interpretation that high-dose and/or chronic glucocorticoid treatment inhibits basalGH secretion and that driven by GHRH, but to a lesser extentthat driven by GHRP. In contradistinction, physiologicalamounts of glucocorticoids are required to maintain normalbasal and GHRH-stimulated pituitary GH reserve (Fig. 13).
The clinical significance of the suppressed GH/IGF-I axisin adults treated with glucocorticoids is reinforced by short-term intervention trials. GH administration elicits a signifi-cant increase in nitrogen balance, serum osteocalcin, thecarboxy-terminal propeptide of type I procollagen, andcarboxy-terminal telopeptide of type I collagen. GH treat-ment also improves protein synthesis without alteringprotein breakdown in patients receiving glucocorticoidtreatment (843). Indeed, glucocorticoid-induced proteincatabolism is reversed during coadministration of GH (844),whereas cotreatment with IGF-I and GH elicits net anabolism(845). In patients undergoing long-term glucocorticoid ther-apy for nonendocrine diseases, GH coadministration re-mains able to significantly lower high-density lipoproteinand low-density lipoprotein cholesterol, but increases serumtriglyceride levels (678).
B. Gonadal sex hormones
1. In vitro and animal studies. To establish the necessary foun-dation for interpreting more recent data on gonadal sex hor-mone regulation of the GH axis, we will first highlight issuesintroduced in an earlier review in the Journal in 1992 (1). Thisbackground will be reappraised in view of new insights andtechnologies developed within the last half-decade (76).Moreover, as highlighted in Fig. 14, wherever possible theactions of testosterone will be distinguished as via its aro-matized product estradiol, or by way of its 5-a reducedpotent androgen metabolite, dihydrotestosterone (143, 846–848).
2. GHRH. In situ molecular hybridization studies indicatethat GHRH mRNA expression is approximately 2–3 timeshigher within the arcuate nucleus of male than female rats(849, 850). Developmentally, this difference is recognized onday 10 of age and in adult animals, but not on days 25–35 oflife (851). A gender difference is also evident in the obese(Zucker) rat in GHRH, but not somatostatin, mRNA contentin the hypothalamus (852). Other recent studies show thatGH’s feedback inhibition of GHRH mRNA expression is sexspecific in the rodent. For example, a GH-hypersecretorystate created by MtTW15 tumor implantation in adult ratsprecipitates a 7-fold decrease in hypothalamic GHRH mRNAcontent in males, compared with only a 2-fold decrease infemales (851). Whether GH receptor expression in arcuatenucleus GHRH neurons per se is gender distinct or sex-ste-roid regulated is not yet known, although liver GH receptorsare gonadal steroid inducible (below).
Whereas there is sexual dimorphism of GHRH mRNA
expression, the specific role of gonadal steroids in regulatingthis difference is less clear. For example, hypothalamicGHRH gene expression in adult and aged male rats in onestudy was not influenced by castration or chronic testoster-one replacement (853). In females, GHRH mRNA levels re-main constant throughout the estrous cycle (849) and preg-nancy (850). The latter point is particularly noteworthy, sinceplacentally derived GH (and hence presumably GH negativefeedback) rises during pregnancy (854). In contrast, otherauthors report that brain GHRH mRNA content in the rat isreduced by castration, and this drop is reversed by testos-terone or dihydrotestosterone (but not estradiol) replace-ment (855). Such experiments suggest that in the rodenttestosterone exerts a stimulatory effect on GHRH gene ex-pression via androgen receptors rather than after its aroma-tization (856).
In relation to GHRH action, one study disclosed that in vivotreatment with testosterone increased the subsequent stim-ulation by GHRH of pituitary GH release in vitro (857). Inanother in vivo investigation, gonadectomized male ratstreated with testosterone exhibited enhanced responsivenessto GHRH (858). Such observations suggest that testosteronemay also modify GHRH actions on the pituitary gland, viaone or more mechanisms activated in vivo and persisting invitro (see below).
The role of estrogens in modulating GHRH expression oraction in the rat seems to be minor compared with that
FIG. 14. Schematic representation of possible loci of action(s) of tes-tosterone on the somatostatin/GHRH-GH axis. Testosterone can actdirectly via the androgen receptor or do so after reduction of its 5-aA-ring to DHT. This pathway appears to dominate in the rat. Alter-natively, testosterone may be aromatized to 17b-estradiol and act viathe estrogen receptor, which seems to be most relevant in the human.In principle, the foregoing sex steroids may impact the GH axis by wayof somatostatin and/or GHRH, or via direct effects on GH-secretingcells in the pituitary gland. Direct effects on somatotrope cells havebeen sparingly demonstrable (hence, the “?”). Sites and mechanismsof sex-steroid actions in the rat and human are not identical, asreviewed in the text. [Adapted with permission from J. D. Veldhuiset al.: Somatotropic Axis and Reproductive Process in Health andDisease. Springer-Verlag, New York, 1995 (847).]
754 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

inferred for androgens. For example, treating adult ovariec-tomized rats with estradiol does not alter GHRH-stimulatedGH release (858). In a study of male rats, estrogen admin-istration reduced hypothalamic GHRH mRNA content (859).Although subpopulations of immunoreactive GHRH-con-taining neurons take up [3H]estradiol in the rat (860), howand the extent to which arcuate nucleus GHRH synthesis andrelease are regulated by estrogens, and whether such puta-tive regulation occurs directly or via changes in somatostatinor other neurotransmitter input to or actions on GHRH neu-rons (861), remain largely unknown.
The impact of gender on basal as well as secretagogue-stimulated release of hypothalamic GHRH has been studiedin vitro in rats ranging from 10 days to 14 months of age. Geet al. (862) showed that, although there is no difference inbaseline hypothalamic GHRH secretion in vitro, pharmaco-logically stimulated GHRH release is higher in male thanfemale rats. This is consistent with, but does not necessarilyaccount for, higher serum GH pulses in male rats (863).Indeed, a sensitizing action of somatostatin withdrawal onGHRH release and action, and inferred intrahypothalamicinteractions between GHRH and somatostatin neurons, arelikely to contribute to the biological rhythmicity of GH re-lease in the male rat (140, 157, 265, 555, 864, 865).
Based on experiments employing passive immunizationagainst, and controlled infusions of, GHRH and somatostatinin the rat, a current thesis is that putative sex-specific dif-ferences exist in reciprocal GHRH and somatostatin releasepatterns in male and female animals, which dictate the prom-inent sex-specific features of pulsatile GH secretion that typ-ify male and female rodents (863, 866). To date, to our knowl-edge, no direct evidence documents disparate modes ofhypothalamo-pituitary portal vein GHRH and/or soma-tostatin release in male and female rats. However, indirectstudies conducted in monosodium glutamate-treated ani-mals to lesion GHRH-secreting neurons, and in GHRH-pas-sively immunized rats, suggest that baseline GHRH secre-tion may be elevated with lower amplitude (and possiblymore frequent) GHRH pulses in females, and conversely thatbaseline GHRH release is lower with higher-amplitude (andpossibly less frequent) GHRH secretory bursts in males (863).In the male, more regularly recurrent intervals of significantsomatostatin withdrawal are also inferred to contributemechanistically to the 3- to 3.5-hourly periods of volley-likeGH release (Fig. 2). Further experiments will be necessary toestablish whether such proposed gender differences existwith respect to in vivo hypothalamic GHRH and somatostatinrelease, and if so how they are endowed specifically by sexsteroids (143, 867). It is pertinent that a recent study indicatedthat ovariectomy in rats increases hypothalamic GHRH re-ceptor expression, which is diminished by subsequent es-tradiol supplementation (255). Thus, sex differences in GHrelease in the rat may reflect not only unequal GHRH andsomatostatin release per se, but also differences in GHRHreception at one or more neural levels.
3. Somatostatin. The extent to which somatostatin mRNAcontent in the periventricular nucleus differs in male andfemale rats remains controversial. Werner and colleagues(868) and others (869) failed to observe any sex difference in
hypothalamic somatostatin mRNA concentrations in intactanimals. However, when evaluating pre-pro-somatostatinmRNA, Zorrilla et al. (869) reported that periventricular insitu expression falls after male or female gonadectomy, andis restored by 5a-dihydrotestosterone (DHT), estradiol, or adopamine agonist. Others describe a markedly lower con-centration of periventricular nuclear somatostatin mRNA inproestrous females compared with males (851). In addition,a sex difference in somatostatin gene transcript levels wasevident on the 10th day of life (851), the earliest time studied.Developmentally, male rats exhibited significant increases insomatostatin mRNA between days 10–25 and females be-tween 10–35 days of age. However, such differences in so-matostatin mRNA content in relatively young males andfemales are at variance with the lack of sexual dimorphismin the GH-secretory pattern, somatostatin peptide concen-trations, or growth in animals before 33 days of age (870).Thus, differential mRNA expression taken alone may notaccurately reflect peptide expression and secretory output ofthe GH axis.
There is less ambiguity concerning the effects of gonadalsteroids on hypothalamic somatostatin mRNA concentra-tions in the rat. Castration of adult male rats significantlylowers concentrations of somatostatin mRNA in the periven-tricular nucleus (868, 871). Testosterone or DHT replacementtherapy reverses this effect of castration. With the exceptionof one study, estrogens do not appear to be so effective inaltering somatostatin gene expression in males (851, 868).Similarly, in females, ovariectomy significantly decreases hy-pothalamic somatostatin mRNA content, whereas treatmentwith estradiol, or in one study DHT (872), reverses thesedecrements. Thus, sex steroids likely facilitate somatostatingene expression, with a reduction in gonadal sex steroidsprecipitating similar decreases in somatostatin gene tran-scripts in males and females.
Since present data suggest that no estrogen-receptor bind-ing consensus sequence exists in the somatostatin gene pro-moter (873), indirect mechanisms or sites of sex steroid actionmay be relevant. Of interest, 40–70% of somatostatin-secret-ing neurons, including these in the periventricular area, ex-press nuclear androgen, but not estrogen, receptors, whichmakes these neurons a plausible (albeit not established) tar-get of direct androgen actions (874–876). The male rat showsgreater androgen receptor expression in somatostatin neu-rons than the female (875).
Two of three reports disclosed no differences in hypotha-lamic somatostatin peptide content in the male vs. female rat(877–879). Baseline and stimulated somatostatin secretionfrom hypothalamic fragments in vitro also was similar in thetwo sexes (862, 878). Thus, somatostatin peptide, unlike itsmRNA, shows little or no evident sexual dimorphism in therodent. This disparity attests further to the need to establishgender-specific patterns of somatostatin release and actionby more direct means, e.g., based on hypothalamo-pituitaryportal vein sampling and somatostatin-receptor antisenseknockout.
Other indirect studies also offer insights into plausiblegender-based differences in GHRH/somatostatin secretionand/or action. Wehrenberg et al. (880) inferred that discrep-ant responses to exogenous GHRH stimuli in conscious
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 755
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

freely moving male vs. female rats reflect unequal endoge-nous somatostatin secretion and action. Other experimentsby Clark and Robinson (881) show that female rats respondconsistently with unabated GH secretion to repeated GHRHadministration, whereas male rats respond only briefly andintermittently, further suggesting that patterns of hypotha-lamic somatostatin are different in male and female animals.Somatostatin immunoneutralization also evokes disparateGH release profiles in male and female rats (863, 882). Thus,a plausible inference in the rodent is that in males soma-tostatin is expressed (129) and released (863) in more regular(3- to 3.5-hourly) and higher-amplitude discharges than infemales (870). Conversely, female rats may secrete soma-tostatin approximately continuously, more nearly randomly,and in smaller amounts (53, 883).
Recent approximate entropy analyses of both human andrat GH release profiles in vivo can quantify a notion of de-creased orderliness of GH release in the female (139). Thiscontrast is shown in the human in Fig. 15, wherein serum GHconcentrations in blood collected every 10 min for 24 h weremeasured by high-sensitivity immunofluorometric assay(118, 124, 139). This entropy difference also is consistent with
a postulated reduced regularity of cyclic somatostatin with-drawal in the human and rodent female. An antiestrogenattenuates the rat’s male-like pattern of GH pulsatility, pos-sibly via reducing somatostatin release (884), suggesting arole for estrogen in directing the male rodent’s GH pattern.However, in the human, estrogen (or testosterone, but notDHT) increases the entropy of GH release, thereby inducinga more female pattern (139, 142) (Fig. 15). We speculate thatGHRH-somatostatin-interactions within the hypothalamusfurther modulate such apparent gender differences in GHneuroregulation, but how sex steroids in particular governthese interactive mechanisms remains unclear (183, 861, 863).
With the cloning of multiple somatostatin receptor sub-types (above), more recent studies can evaluate whethergonadal steroids regulate the expression of specific soma-tostatin receptor subtypes. In the rat anterior pituitary gland,the subtype I receptor is regulated by testosterone, and sub-type III is regulated by nonandrogenic testicular factors. Sub-type II appears not to be gonadally modulated (175). Differ-ential topographic distribution of subtype I and subtype II inthe hypothalamus also occurs but is not known to be genderspecific (178). Further studies of sex hormone regulation ofsomatostatin receptor expression and/or function thus willbe helpful in clarifying the specific molecular mechanismsunderlying gender differences in GH neuroregulation.
In relation to sex differences in pituitary GH gene expres-sion, the concentration of GH mRNA is approximately 2 to6 times greater in adult male than female rats (885, 886).Whether this distinction arises from direct actions of gonadalsteroids on pituitary GH gene transcription or mRNA sta-bility, or via indirect sex-steroid effects on hypothalamicinhibitory and releasing hormones, or their receptors, is notestablished. The former indirect mechanisms would includehigher GHRH pulses in males, which in turn induce GH genetranscription (887). Since somatostatin inhibits GH release,but not GH biosynthesis, presumptively greater somatosta-tin release in the male rat would not directly limit GH mRNAaccumulation, but would allow greater GH storage in so-matotropes awaiting the next GHRH pulse (888).
Few studies have evaluated direct actions of gonadal ste-roids on GH gene expression. Estradiol does not directlyinfluence GH mRNA accumulation in the bovine pituitarygland (889) or alter in vitro GH secretion by macaque soma-totropes (890). On the other hand, estrogen stimulates GHmRNA gene expression in MtT/F4 transplantable tumors in40-day-old female rats (891). The latter finding diverges fromthe general theme that estrogen inhibits pituitary responsesto GHRH in the normal rat (see below), and that serum GHconcentrations are lower in the female of this species. Aplausible explanation is that the MtT/F4 tumor is a uniqueestrogen-responsive neoplasm and therefore may not be anappropriate model for evaluating normal physiology. On theother hand, in one study, 80% of normal rat somatotrope cellsexpressed [3H]estradiol binding sites (860), thus allowingpossible direct actions of estrogen on GH-secreting cells. Inthis regard, estrogen treatment of male rats in vivo reducesin situ GH mRNA expression in single somatotropes in vitro(892). Such in vitro observations differ from other in vivostudies in the ovariectomized female rat, in which andro-gens, but not estrogen, can augment pituitary GH content
FIG. 15. A, Approximate entropy (ApEn) contrasts in healthy middle-aged men (open circles) and women (solid circles) in relation to mean24-h serum GH concentrations. ApEn is a statistical measure of rel-ative disorderliness or irregularity. Higher ApEn values (x-axis) de-note greater irregularity or disorderliness of the 24-h GH releaseprocess. ApEn was calculated from 24-h serum GH concentrationprofiles obtained by sampling blood every 10-min followed by im-munofluorometric assay of GH. [Adapted with permission from Pin-cus et al.: Am J Physiol 270:E107–E115, 1996 (139).] B, Ability ofestradiol treatment for 1 (acute, short-term) or 4–5 (longer term)weeks to increase the disorderliness of overnight GH release profilesin girls with Turner’s syndrome. Higher ApEn denotes greater ran-domness of GH release. [Adapted with permission from J. D. Veldhuiset al.: J Clin Endocrinol Metab 82:3414–3420, 1997 (142). © TheEndocrine Society.]
756 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

(893). Consequently, far more study will be needed to clarifythe nature of sex steroid actions (if any) directly on soma-totrope cells.
The GH-secretory response to GHRH stimulation in the rattends to be sexually dimorphic. In vitro perifusion studiesreveal that pituitaries from male rats release more GH inresponse to GHRH than those of females. Furthermore, eitherneonatal or prepubertal orchidectomy reduces baseline andGHRH-stimulated secretion of GH in vitro (857). Male ratsgonadectomized on day 22 of age show a decreased in vitroresponse to GHRH, which is reversed by in vivo testosteronereplacement (894). On the other hand, in female rats prepu-bertal ovariectomy does not affect GH secretion in vitro, andestradiol replacement in adult castrate animals lowers base-line and GHRH-stimulated GH secretion in vitro (857).
A gender-appropriate sex-steroid environment is requiredthroughout adult life to maintain physiological GH secretionin the rat in vivo (883). Basal or GHRH-stimulated secretionof GH in vitro is significantly higher by pituitary tissue ob-tained from adult male compared with female animals (878,895). Orchidectomy followed by various sex hormone re-placement therapies suggests that this gender distinction ismediated primarily by testosterone (896). Androgen’s en-hancement of GH release reflects, in part, increased sensi-tivity to GHRH and greater GH secretory capacity of indi-vidual somatotropes, as studied in vitro in the reversehemolytic plaque assay (897, 898). Greater somatotrope GHcontent also has been inferred on a histological basis (877).However, other (non-GHRH) modulatory inputs to soma-totropes may be influenced by androgens, such as soma-tostatin, and possibly the putative endogenous GHRP path-way, galanin, NPY, IGF-I feedback, and GH autofeedback.For example, the exogenous GHRP stimulus is enhanced byboth testosterone and estrogen in the pituitary-transplantedrat (899).
The effects of estrogens on pituitary responsiveness toGHRH in the rodent are controversial. Simard et al. (900)reported that estradiol exposure stimulates in vitro basal andGHRH-stimulated GH secretion by pituitaries obtained fromfemale rats, whereas Fukata and Martin (901) reported noeffect of estrogen on these measures. Other experiments in-dicated that estradiol treatment of castrated male rats leadsto reduced in vitro GH secretion similar to that reported infemale animals (896). Thus, whereas androgens consistentlyfacilitate the pituitary GH response to GHRH in the rat,estrogen effects are inconsistent or inhibitory.
Understanding the mechanistic basis of the distinctive pat-terns of GH secretion in the two sexes, i.e., higher and moreregular GH volleys in males, and lower-amplitude irregularpeaks in females, has been aided by studies in androgenreceptor-deficient rodents (e.g., testicular feminized rat). Thelatter genetically male, but androgen receptor-deficient, ro-dent shows intermediate (to intact male and female, withintact female having the greatest) single somatotrope cellsensitivity to somatostatin’s inhibition in the reverse hemo-lytic plaque assay (902). The testicular feminized rat alsoexhibits an intermediate in vitro GH secretion rate in responseto a maximal GHRH stimulus, i.e., less than of an intact maleand more than that of an adult female (895). In further singlesomatotrope studies, Martha et al. (903) identified more func-
tional somatotropes in the male testicular-feminized rat thanthe normal female, but each somatotrope cell secreted lessGH basally than the wild-type male. These differences couldexplain the more nearly “female” GH release pattern in vivoof the testosterone-producing but androgen receptor-defi-cient testicular-feminized animal (856) and is consistent witha notion of greater somatotrope-secretory activity in themale. How such differences arise mechanistically is not yetestablished.
In nonhuman and nonrodent species, estrogens typicallymodify the GH axis in a stimulatory manner. For example,studies with single bovine pituitary somatotrope cells usingthe reverse hemolytic plaque assay document increasedmammosomatotropes (PRL- and GH-secreting cells) in theluteal phase (904). On the other hand, in this species, bullshave higher mean serum GH concentrations, GH pulse fre-quencies, amplitudes, and GHRH-stimulated GH releasethan heifers (905). In sheep, increased pituitary GH mRNAcontent and serum GH concentrations develop in the latefollicular phase, but the estrogen-induced LH surge 22 h lateroccurs without a further change in GH mRNA concentra-tions (906). In the baboon, estradiol in either physiological orpharmacological amounts delivered via silastic subcutane-ous capsules increases serum IGF-I concentrations and stim-ulates GH release in long-term ovariectomized animals (907).In this primate, basal serum GH concentrations also arehigher in females than in males. These gender differences arelikely mediated via the hypothalamus, since estrogen stim-ulates in vivo GH secretion but has little effect in vitro on GHrelease by primate (macaque) pituitary cells (890).
4. GH receptor and BPs. Sex steroids modify GH receptorexpression in the liver in a sex hormone- and species-specificmanner. For example, in the rabbit (908), estradiol inhibitsGH receptor gene expression. On the other hand, in the rat,estrogen consistently increases liver GH receptor and GHBPexpression (909), as discussed further below. In the human,oral estrogen treatment also augments plasma GHBP levels,which is believed to reflect hepatic GH receptor (extracellulardomain) expression (910). Contrastingly, whether in thepresence or absence of endogenous GH, testosterone treat-ment in men lowers serum GHBP concentrations (911).
Since the demonstration that the GHBP in rat plasma is analternatively spliced transcript of the GH receptor gene (912),the tissue distribution and regulation of GHBP mRNA andprotein have been studied extensively (912–918). Molecularexperiments document species variations in the spectrum ofmRNA transcripts that encode the GH receptor and GHBP,e.g., in the pig, rat, chicken, and human (917), as well as intheir gestational, developmental, and tissue-specific regula-tion in the rodent (916). There is gender-specific (sexuallydimorphic) liver GHBP production in the rat, as reflected bysex steroid and pregnancy’s regulation of GHBP transcriptconcentrations. GHBP gene and protein expression patternsare also controlled by continuous (female-like) vs. pulsatile(male-like) GH exposure (915). For example, the so-calledGHR-1 is an alternatively spliced mRNA class encoding theGHBP, which is GH dependent, signal specific (responsive tocontinuous rather than pulsatile GH), induced by gonadalsteroids, and eliminated by hypophysectomy (915). Such in
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 757
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

vivo evidence of steroidal regulation of specific GHBP ex-pression has been corroborated more directly recently in anin vitro model of primary adult rat hepatocytes (914).
Other hepatic genes that are responsive to a sex-specificpattern of GH secretion, and are subject to androgen im-printing, include the hepatic cytochrome P450 2C11, 3A2,and 5a-reductase enzymes (919) (see Table 3). Gender dif-ferences in GH tissue actions are attributed to pulsatile (male-like) vs. more nearly continuous (female) patterns of GHsecretion (920). Indeed, specific gene induction in varioustarget tissues of GH action (e.g., liver, muscle, etc.) is differ-entially responsive to the GH pulse pattern (921, 922). Sec-ond-messenger intracellular signaling per se is activated dif-ferentially via a pulsatile vs. continuous GH stimulus, withthe former (but not the latter) activating the STAT5b-signal-ing cascade (866). Demonstrating the critical nature of theSTAT5b signaling pathway, STAT5b knockout mice fail torespond with male growth rates or gene induction despite“masculine” GH pulse patterns in the blood (32).
5. Human studies.a. Spontaneous GH secretion. Gender differences in human
GH secretion were reviewed in part by Kerrigan and Rogolin the Journal in 1992 (2), and elsewhere (76). Here, morerecent findings are evaluated, and earlier observations rein-terpreted in the context of new hypotheses and investigativestrategies (76). For example, although androgens clearly acton the GH-IGF-I axis, recent experiments also emphasizetheir capacity to alter the orderliness of GH release and forindependent trophic effects without evident stimulation ofthe GH axis, e.g., to exert anabolic tissue effects (142, 923).
Androgens in the absence of GH are insufficient in thehuman to drive the fully normal tempo of clinical pubertalmaturation, since hypopituitary boys replaced with testos-terone but not GH exhibit a protracted 7- to 8-yr (rather than3- to 4-yr) long pubertal growth period (924). In contrast, amarked androgen-GH synergy unfolds in normal puberty(848), as well as in pubertally delayed boys treated withcombined GH-testosterone replacement therapy. Serum tes-tosterone concentrations in the course of normal pubertyspan a wide spectrum, viz., from prepubertal values (,1nmol/liter) to adult concentrations (20–25 nmol/liter).Cross-sectional and longitudinal studies of healthy pubertalboys have documented the physiological association be-tween rising blood androgen concentrations and augmentedneuroendocrine activity of the somatotropic axis. Initial stud-ies reported GH pulse activity using discrete peak detectionmethodologies and demonstrated that the mean serum GHconcentration peak amplitude in boys increases 2- to 3-foldin the mid- to later stages of puberty, with no change in thenumber of detected GH peaks by cluster analysis (925). Later,a deconvolution technique for estimating endogenous hor-mone secretion rates and/or half-life has been implementedto quantitate 24-h serum GH profiles in males whose pu-bertal development spanned the range of Tanner stages I(prepubertal) through V (fully mature adult). In these cross-sectional analyses, boys in mid-to-late puberty exhibitedhigher mean and pulsatile serum GH concentrations, mech-anistically explained by amplification of GH-secretory burstmass and amplitude due to a rise in the maximal rate of GH
secretion (amplitude) attained within each secretory pulse(926). Linear correlation analysis demonstrated that serumtotal testosterone concentrations in pubertal boys arestrongly positively correlated with plasma total IGF-I levels,the calculated daily pulsatile GH secretion rate, and GH-secretory burst mass (926). In a novel clinical model of leu-prolide-induced gonadotropin down-regulation in normalyoung men (923), Fryburg et al. showed dose-dependentactions of testosterone repletion on each of these measures ofthe GH axis. A nonaromatizable synthetic androgen did notactivate the GH axis (or increase serum IGF-I levels), butpromoted nitrogen retention, thus emphasizing the non-GH-dependent anabolic effects of androgens (923).
Other clinical intervention investigations in boys with con-stitutionally delayed puberty revealed that testosterone ad-ministration significantly increases the amplitude (or mass)of GH-secretory peaks in a selective manner without alteringthe GH half-life, pulse frequency, or pulse duration (927). Aqualitatively similar response was observed in four of fiveboys after administration of oxandrolone, a synthetic non-aromatizable androgen, but this substudy did not includecontrols for possible placebo or order effects of treatmentintervention. More recently and informatively, a longitudi-nal/prospective study of nocturnal pulsatile GH secretionwas performed in six previously untreated boys with iso-lated hypogonadotropic hypogonadism. Deconvolutionanalysis showed that the amount of GH secreted in pulseswas significantly increased by as little as 25 mg of injectedtestosterone (928). Both the number of spontaneous GHpeaks and their mean amplitude rose significantly after dosesof 50 and 100 mg testosterone. This suggests that in younghuman males even minimal increments in serum testoster-one concentrations occurring during the early stages of pu-berty can amplify both the amount and frequency of spon-taneous GH-secretory bursts (928). Moreover, the orderlinessor regularity of GH release, as quantified by approximateentropy (132), decreased rapidly during testosterone treat-ment. The more irregular GH release patterns induced bytestosterone injections in these boys closely mirrors the moredisorderly mode (reduced pattern reproducibility) of GHsecretion that emerges before and during maximal lineargrowth in normal pubertal boys (142) (see Fig. 16). Thus,androgens control not only the quantity but also the quality(orderliness or regularity) of the GH release process, pre-sumably indicate network effects of sex steroids on theintegration of somatostatin-GHRH-GH-IGF-I feedback/feedforward interactions.
In healthy men, as in pubertal boys, a strongly positiveassociation between mean serum GH concentrations andserum total (or free) testosterone concentrations is evident.Relative hypogonadism accompanies healthy aging in menand is closely related statistically to reduced GH secretion(121). Of mechanistic interest is the observation that in bothmen and prepubertal boys serum total testosterone concen-trations are correlated positively with GH-secretory burstmass and amplitude (928, 929). From a neuroendocrine per-spective, this suggests that androgens promote a relativelygreater ratio of GHRH/somatostatin actions on responsivesomatotropes.
Testosterone treatment of primary hypogonadal patients
758 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
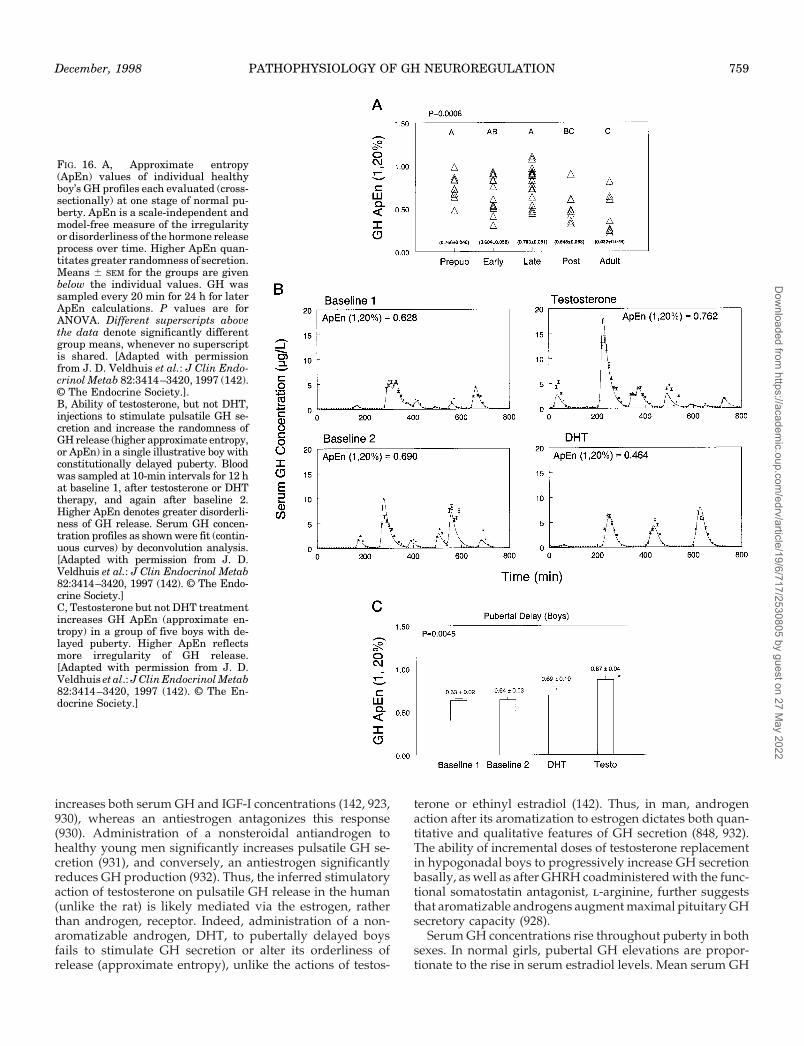
increases both serum GH and IGF-I concentrations (142, 923,930), whereas an antiestrogen antagonizes this response(930). Administration of a nonsteroidal antiandrogen tohealthy young men significantly increases pulsatile GH se-cretion (931), and conversely, an antiestrogen significantlyreduces GH production (932). Thus, the inferred stimulatoryaction of testosterone on pulsatile GH release in the human(unlike the rat) is likely mediated via the estrogen, ratherthan androgen, receptor. Indeed, administration of a non-aromatizable androgen, DHT, to pubertally delayed boysfails to stimulate GH secretion or alter its orderliness ofrelease (approximate entropy), unlike the actions of testos-
terone or ethinyl estradiol (142). Thus, in man, androgenaction after its aromatization to estrogen dictates both quan-titative and qualitative features of GH secretion (848, 932).The ability of incremental doses of testosterone replacementin hypogonadal boys to progressively increase GH secretionbasally, as well as after GHRH coadministered with the func-tional somatostatin antagonist, l-arginine, further suggeststhat aromatizable androgens augment maximal pituitary GHsecretory capacity (928).
Serum GH concentrations rise throughout puberty in bothsexes. In normal girls, pubertal GH elevations are propor-tionate to the rise in serum estradiol levels. Mean serum GH
FIG. 16. A, Approximate entropy(ApEn) values of individual healthyboy’s GH profiles each evaluated (cross-sectionally) at one stage of normal pu-berty. ApEn is a scale-independent andmodel-free measure of the irregularityor disorderliness of the hormone releaseprocess over time. Higher ApEn quan-titates greater randomness of secretion.Means 6 SEM for the groups are givenbelow the individual values. GH wassampled every 20 min for 24 h for laterApEn calculations. P values are forANOVA. Different superscripts abovethe data denote significantly differentgroup means, whenever no superscriptis shared. [Adapted with permissionfrom J. D. Veldhuis et al.: J Clin Endo-crinol Metab 82:3414–3420, 1997 (142).© The Endocrine Society.].B, Ability of testosterone, but not DHT,injections to stimulate pulsatile GH se-cretion and increase the randomness ofGH release (higher approximate entropy,or ApEn) in a single illustrative boy withconstitutionally delayed puberty. Bloodwas sampled at 10-min intervals for 12 hat baseline 1, after testosterone or DHTtherapy, and again after baseline 2.Higher ApEn denotes greater disorderli-ness of GH release. Serum GH concen-tration profiles as shown were fit (contin-uous curves) by deconvolution analysis.[Adapted with permission from J. D.Veldhuis et al.: J Clin Endocrinol Metab82:3414–3420, 1997 (142). © The Endo-crine Society.]C, Testosterone but not DHT treatmentincreases GH ApEn (approximate en-tropy) in a group of five boys with de-layed puberty. Higher ApEn reflectsmore irregularity of GH release.[Adapted with permission from J. D.Veldhuis et al.: J Clin Endocrinol Metab82:3414–3420, 1997 (142). © The En-docrine Society.]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 759
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

peak amplitude increases 2- to 3-fold from prepuberty tomenarche, which increment (especially in the daytime) iscorrelated to serum estradiol concentrations (933). This re-lationship supports the notion that in puberty in girls en-dogenous estradiol, or a pertinent covariate, drives amplifiedGH secretion. Similar GH axis activation by estrogen is de-monstrable in the castrate (female) baboon (907) and ma-caque (890). Moreover, in prepubertal girls with previouslyuntreated Turner’s syndrome, oral administration of a smalldose of ethinyl estradiol (100 ng/kg/day) amplifies the GHsecretion rate 2- to 3-fold and heightens the disorderliness ofGH release, the latter as quantified by the approximate en-tropy statistic (934). Similar responses are achieved in pre-pubertal boys treated with testosterone, but not DHT. Thispoints to an augmentative role of estrogen (or aromatizedandrogen) in GH axis neuroregulation in both boys and girls(142).
Gender differences in GH release in adults were recog-nized in 1965 by Frantz and Rabkin (935), who describedsimilar fasting serum GH concentrations in men and womenin a cohort of 79 individuals (ages 20 to 80 yr) (935), but amarked (6-fold) increase in serum GH concentrations afterambulation in women only. Administration of a potent oralnonsteroidal estrogen, diethylstibestrol, 2.5 mg twice dailyfor 5 days in 4 men evoked a “female pattern” of GH releaseafter exercise. These authors further identified a 2-fold rise inserum GH concentrations across the menstrual cycle andinferred that gender contributes to the regulation of GHrelease and probably does so via estrogen (935). Later studiesby Unger et al. (936) described a sex difference in basal serumGH concentrations. Thompson et al. (92) extended the in-ferred role of estrogen in stimulating higher GH secretionrates in premenopausal than in postmenopausal women.More directly, Dawson-Hughes et al. (937) demonstrated that20 mg ethinyl estradiol administered orally for 15 days elicitsan approximately 45% increase in the 24-h mean serum GHconcentration in older women. Similarly, Dursma et al. (938)showed that 3 weeks of this estrogen regimen will stimulatea rise in serum GH concentrations, while reducing plasmasomatomedin C (IGF-I) levels.
By way of possible physiological regulation, serum GHconcentrations rise 2-fold in the late follicular phase of thenormal menstrual cycle, based on specific GH IRMA inyoung women sampled every 10 min for 24 h (939). Moredramatic increases in endogenous estradiol secretion in-duced by superovulation treatment of infertile women canstimulate serum GH concentrations by up to 4-fold (940).Conversely, suppressing ovarian estrogen secretion byGnRH agonist down-regulation of the gonadotropic axis inpremenopausal women or girls with precocious puberty re-duces basal and GHRH-stimulated GH release (941, 942), aswell as plasma IGF-I concentrations (943). A recent analysisusing longitudinal (within-subject) comparisons in the samemenstrual cycle further disclosed that GH-secretory pulsefrequency and amplitude, and plasma IGF-I concentrations,rise concomitantly in the preovulatory phase with the serumestradiol zenith (944). This physiological reactivity of theGH-IGF-I axis with combined augmentation of GH and IGF-Iconcentrations is not fully recapitulated by oral or transder-mal estrogen delivery in postmenopausal women (below),
but is achieved in the long-term castrate (female) baboon bycrystalline estradiol delivery via subcutaneous silastic cap-sules (907) or parenteral estrogen in the macaque monkey(890).
Weissberger et al. (945) suggested that oral (but not trans-dermal) estrogen delivery augments GH secretion in olderwomen and does so by reducing plasma IGF-I concentra-tions. This would attenuate IGF-I’s negative feedback on thehypothalamic-somatotropic axis. Three different oral estro-gens exerted this effect and concurrently increased serumGHBP concentrations (24, 946). Other studies show that oralestrogen treatment may not always lower plasma IGF-I levels(947), and that higher doses of transdermal estrogen canindeed stimulate GH secretion (204). In contrast to theseparadigms, normal female puberty and the preovulatoryphase of the menstrual cycle are marked by conjoint eleva-tions in circulating estrogen, IGF-I, and GH concentrations(140, 933). Thus, whether during puberty or just before ovu-lation, reduced plasma (total) IGF-I feedback cannot providethe basis for amplified GH release. In addition to physio-logical follicular phase estrogen secretion, parenteral estra-diol treatment of castrate or intact baboons increases serumGH and IGF-I levels concurrently (907). This experimentalparadigm mimics that observed in puberty and in the normalpreovulatory phase of the menstrual cycle and suggests thatestradiol can enhance the net central drive to GH release andthereby concurrently stimulate IGF-I secretion. Thus, theroute and amount of estrogen, as well as the pubertal ormenstrual context, likely govern the nature of estrogen’seffects on the GH-IGF-I axis (204, 910).
The relationship between estrogen and IGF-I may be com-plex, and tissue specific, e.g., different in liver vs. uterus. Forexample, estrogen inhibits liver but stimulates uterine IGF-Iaccumulation in the hypophysectomized rat (948). This couldexplain the ability of low doses of oral estrogen to stimulatelinear growth in patients with Turner’s syndrome, whereasthe same or higher doses of estrogen reduce hepatic IGF-Iproduction (934). Other indirect evidence suggests that es-trogen administration may attenuate plasma IGF-I responsesto fixed doses of exogenous (human recombinant) GH inpostmenopausal women (949); i.e., there is a possible genderdifference in tissue sensitivity to GH actions, with reducedend organ (at least liver) responsiveness to GH in womentreated with estrogen (950). This might reflect the ability, insome species, of estrogen and GH itself to down-regulatehepatic GH receptors (above), or impede GH tissue re-sponses. In contrast, multiple reports demonstrate that par-enteral testosterone stimulates increases in both GH secre-tion and plasma IGF-I concentrations (847, 910, 923, 930). ThisIGF-I response to aromatizable androgen mechanisticallydistinguishes it from oral estrogen action in the human.
Analyses of the responsiveness of the anterior pituitarygland to GHRH injections in women compared with men andin women throughout the menstrual cycle are conflicting. Forexample, the evaluation by Gelato et al. (205) of the GHRHdose-response curve (0.01 to 10 mg/kg) throughout the nor-mal menstrual cycle in young women detected no systematicvariations in the magnitude of GH responses. Although max-imal GH release was similar in men and women, the half-maximally effective dose of GHRH was reduced in women
760 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

at 0.2 mg/kg compared with 0.4 mg/kg in men. Evans et al.(210), utilizing a single maximal dose of 3.3 mg/kg GHRH,also found no GH-secretory differences among the earlyfollicular, late follicular, and midluteal phases. In a laterstudy, there were similar maximal GHRH effects in men andwomen basally and after GnRH agonist-induced down-reg-ulation of the gonadal axis for 40 days (951). However, to ourknowledge, studies of GHRH dose-dependent actions in thesame woman at different stages of the menstrual cycle withvs. without somatostatin withdrawal are not yet available.
Unlike the above findings, other investigators identifylarger effects of GHRH in women than men (952) and greatermaximal GHRH-stimulated (incremental) GH release in thelate follicular phases of the menstrual cycle. The latter risescorrelated with higher serum estradiol concentrations (952).In a large study of 116 men and women ages 18–95 yr, Langet al. (953) unmasked higher incremental release to GHRHstimulation in premenopausal women vs. age-matched men.Serum estradiol concentrations correlated with the magni-tude of maximal GHRH-stimulated GH release. GH incre-ments were equal in postmenopausal women and older men,who have similar estrogen levels.
Greater GH release in women than men is also reported forl-arginine and insulin-stimulated hypoglycemia (954), stim-uli acting at the hypothalamic level (681, 783). Treatmentwith estrogen (but not testosterone’s methylated derivative)augmented l-arginine and insulin-induced GH release inmen, which suggests that these gender differences reflectunequal estrogen exposure. Another peptide also thought tostimulate GH secretion via hypothalamic pathways, i.e., ga-lanin (435), elicits more GH secretion in young women thanmen (432, 445, 955). Whereas the non-GHRH hexapeptide,GHRP-6, had no gender bias in its maximal GH-releasingproperties (efficacy), this study did not evaluate the half-maximally effective GHRH-6 doses (potency of, or GH-axissensitivity to, this potent secretagogue) (291, 416).
Although the mechanism of glucagon’s stimulation of GHrelease is not clear, no sex difference is evident (956). Con-versely, clonidine, an a2-agonist, stimulates more GH releasein men than premenopausal women (597). Most recently,whereas young early follicular phase women were moreresponsive to l-arginine than men, young men exhibitedgreater GH release after GHRP-2 stimulation than compa-rably aged women (957). Combining these two secreta-gogues with an exercise stimulus yielded maximal GH se-cretion, which was equivalent in the two sexes (957).
We infer that the ability of pharmacological agonists tostimulate GH secretion varies as a function of gender anddepends on the specific secretagogue and neurotransmitterpathway employed, the age and the gonadal status of theindividual, the stage of the menstrual cycle, the intensity ofthe stimulus, and possibly the estrogenicity of the endocrinemilieu and route of estrogen delivery.
6. Influence of sex steroids on GH clearance. Steady-state GHinfusions in the rat indicate that the gonadal sex steroidenvironment may influence the MCR of GH. For example,the orchidectomized rat shows a decrease in GH MCR duringestrogen add-back, and an increase after testosterone add-back (958). Since the half-life of GH disappearance from
steady state was similar in male and female animals, thisstudy suggests that sex steroids might alter the volume ofdistribution of GH, which was not measured here. An infu-sion study in the human suggested as much as a 30% meandecrease in the equilibrium GH MCR in women comparedwith men, although the distribution volume and half-life ofGH were not estimated (959). On the other hand, anotherrecent clinical investigation of the half-life of bolus-injectedhuman recombinant GH in young men revealed an inversecorrelation of half-life and serum estradiol concentrations,which would suggest a direct (rather than negative) rela-tionship between estradiol and GH MCR, at least in men(960). In young women, stage of the menstrual cycle had noinfluence on exogenous (bolus) biosynthetic human GH met-abolic clearance, half-life, or distribution volume (961). Menand women had similar GH half-lives and distribution vol-umes (adjusted for body surface area) in this study. Equiv-alent half-lives of endogenous GH were also estimated re-cently by deconvolution analyses in men and women (118).Thus, kinetic differences based on gender are not settled inthe human. However, quantitatively, unequal GH removalrates, if confirmed in women and men, at a difference of only20–30% would account marginally for the 200–300% differ-ences in mean serum GH concentrations between the sexesin young adults and across the menstrual cycle (939, 944,962).
C. Thyroid hormones
1. In vitro and animal studies. Brain GHRH expression in therodent is affected by alterations in circulating levels of thy-roid hormones (963). Propylthiouracil-induced hypothyroid-ism in adult rats increases hypothalamic GHRH mRNA ex-pression (964), while hyperthyroidism acts conversely todecrease GHRH mRNA content (965). The hypothalamiccontent of GHRH peptide is also affected by hypothyroidismand hyperthyroidism in an age-related manner. In rats up to21 days of age, thyroid status does not affect the hypotha-lamic content of GHRH or GHRH positive-staining struc-tures (966). After that age, hypothyroidism decreases GHRHcontent (967). These experiments are consistent with, but notproof of, the hypothesis that there is increased GHRH releasein a homeostatic response by the animal to the low serum GHconcentrations that prevail in hypothyroidism. Hyperthy-roidism reportedly also decreases GHRH content, whichwould concur with the fall in GHRH mRNA under theseconditions (965).
The hypothyroid state enhances both basal and K1-stim-ulated GHRH secretion from in vitro incubated hypothalamicfragments (968), which might represent a compensatorymechanism to the low GH levels. Conversely, excess T4 in-hibits hypothalamic GHRH secretion up to 50% (comparedwith control values) in this model. Using in vitro pulse-labeling techniques, De los Frailes et al. (969) observed thatGHRH synthesis in fetal cortical cells is stimulated by a lowconcentration of T3 (1027 mol/liter), but inhibited by higheramounts (1023 mol/liter). Baseline culture medium con-tained no T3, and thus may have represented a biochemical“hypothyroid” state, while the low and high concentrationsof T3 may resemble euthyroid and hyperthyroid states, re-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 761
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

spectively. Thus, these experiments suggest that hypothy-roidism, as well as hyperthyroidism, may decrease GHRHsynthesis at least in cortical neurons.
In vivo studies indicate that neural somatostatin synthesisis not affected by thyroid hormone during early neonatalrodent development (963). In contrast, in adult rats pharma-cologically induced hypothyroidism decreased hypotha-lamic somatostatin mRNA concentrations (964). The hypo-thalamic content of somatostatin peptide also varies underhypo- and hyperthyroid conditions. In vitro cultures of fetalcortical cells obtained on day 18 of gestation, and treated withT3 or T4, become impoverished in somatostatin (970). How-ever, the neonatal and prepubertal hypothalamic content andsynthesis of somatostatin are not affected by thyroid hor-mones (964). In adult male rats, the somatostatin content inthe median eminence falls with thyroidectomy comparedwith thyroid-intact animals and returns to normal with T4replacement therapy (971). Hyperthyroidism, induced by asubcutaneous implant of T4, does not seem to alter the levelof hypothalamic somatostatin peptide (972). However, pep-tide concentrations in neuronal loci must not be equatedfaciley with their de facto release rates.
Although the secretion of somatostatin from the hypo-thalamus is difficult to assess in vivo, in vitro release of so-matostatin from fetal hypothalamic tissue is stimulated by alow concentration of T3 (10211 mol/liter), but inhibited by ahigh dose (1027 mol/liter) (969). As noted earlier, the base-line medium was devoid of T3 and therefore could reflect a“hypothyroid” condition, while the low and high dose of T3might approach a normal and hyperthyroid state, respec-tively. This argument conforms with the observation that invitro basal and stimulated somatostatin secretion from hy-pothalamic tissue of hypothyroid rats is decreased comparedwith that in euthyroid animals (971). Furthermore, chronicexposure to T3 or T4 suppresses neural somatostatin releaseafter K1-induced depolarization (970). However, one pub-lication suggests that thyroid hormone has no effect on so-matostatin content or release despite altering pulsatile GHsecretion (973).
Direct in vivo quantification of hypothalamic somatostatinsecretion has been reported in pentobarbital-anesthetizedrats. In this model, neither hypothyroidism nor hyperthy-roidism affected hypothalamic-hypophyseal portal bloodconcentrations of somatostatin (974). Although such resultswould suggest that the thyroid environment does not affectsomatostatin secretion, the conclusions must be tempered bythe recognition that anesthesia has inhibitory and possiblyother confounding effects on hypothalamic somatostatin(and/or other neuropeptide) secretion (809).
In somatotrope tumor cell lines, thyroid hormones stim-ulate GH mRNA accumulation (975, 976) by increasing therate of GH gene transcription (977, 978). This effect of T4 alsooccurs in cultured normal rat anterior pituitary cells (979). Ofnote, thyroid hormone’s effects on GH gene expression andGH synthesis vary significantly among mammalian species;e.g., in bovine anterior pituitary cells, T3 fails to augmentbasal and GHRH-stimulated GH secretion (889), and cul-tured human fetal pituitary cells supplemented with T3 showreduced basal GH release and attenuated responses toGHRH (980). Also, in cultured human pituitary tumor cells
obtained at neurosurgery from patients with acromegaly, T3has no major impact on GH secretion (981). Furthermore,after transfection into rat pituitary cells, human GH geneexpression is reduced by T3 administration (982), althoughexpression of the rat and bovine GH promoters are inducedby T3 (983). Thus, although the most prominent action in therat of thyroid hormones on GH secretion at the somatotropelevel is to promote the transcription of the GH gene andhence GH synthesis and accumulation, there are evidentspecies differences.
Thyroid hormones also have important effects on pituitaryGH protein content. In studies conducted in 10- to 21-day-oldrat pups, hypothyroidism decreased pituitary GH content(966). Similarly, thyroidectomy in adult rats profoundly re-duces pituitary GH concentrations with values dropping toless than 1% of those in normal animals (967, 973). T4 re-placement therapy partially restores pituitary GH content.
Another mechanism through which thyroid hormonesmay act is regulation of pituitary sensitivity to GHRH. In fact,the GH response to GHRH is significantly lower in normalpituitary cells maintained in vitro without T3 compared withT3-exposed cells (798). The blunted action of GHRH reflectsprimarily a reduced maximal GH response to GHRH asdistinct from a change in the ED50 of GHRH action. Cellsobtained from hypothyroid animals likewise show a de-creased response to GHRH in vitro. Most recently, decreasesin not only GH and GHRH gene expression, but also insomatostatin and GHRH receptor mRNA, have been de-scribed in hypothyroid animals, thus highlighting an arrayof GH axis disturbances in this metabolic state (984).
2. Human studies.a. Hypothyroidism. In humans, as in other species, hypo-
thyroidism severely impairs postnatal growth (963). More-over, spontaneous nocturnal GH secretion, which is one in-dicator of physiological GH production, is consistently lowin the hypothyroid state and is correlated with reduced cir-culating IGF-I levels (985). Chernausek and Turner (985)reported that mean nocturnal serum GH concentrations inhypothyroid individuals are reduced to 58% of values ob-tained during T4 replacement therapy. Concurrently withreduced spontaneous GH secretion, plasma IGF-I concen-trations are lower in the same patients before, compared withduring, T4 replacement therapy. Replacement T4 therapyreturns GH and IGF-I levels to normal (986).
Consistent with the data derived from studies of sponta-neous GH secretion, acute GH-secretory responses to themost common pharmacological stimuli, such as insulin-induced hypoglycemia, l-arginine and GHRH, are allblunted in hypothyroid patients (494, 987, 988). Whenhypothyroid patients are replaced with T4, plasma IGF-Iconcentrations rise toward normal values. Administration ofT4 to hypothyroid patients also enhances the GH response toGHRH. In fact, in hypothyroid humans, the GH response toGHRH is normalized 2 weeks after T4 replacement therapyis begun (494, 989).
The possible hypothalamic origin of hyposomatotropismand pituitary hyporesponsiveness to GHRH in clinical hy-pothyroidism is supported by investigations using the ace-tylcholinesterase inhibitor, pyridostigmine, and the amino
762 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

acid, l-arginine, both of which are hypothesized to decreasehypothalamic somatostatin release. Indeed, hypothyroid pa-tients manifest reduced GH-secretory responses to putativesomatostatin withdrawal via either pyridostigmine or l-arginine (989). Such observations speak against heightenedsomatostatin tone as the sole cause of GH deficiency andsuggest that either endogenous GHRH action and/or GHsynthesis in the pituitary gland may also be impaired duringthe chronic hypothyroid state in the human. These mecha-nisms cannot be distinguished based on available data, giventhe reduced GH response to GHRH demonstrated in hypo-thyroid subjects. Indeed, decreased pituitary GHRH-releas-able pools of GH could result secondarily from reducedGHRH release and/or action, which would account for re-duced 24-h GH secretion, as well as impaired acute GH-secretory responses to other pharmacological stimuli.
b. Hyperthyroidism. The 24-h GH-secretory rate and theamount of GH released during each pulse are reportedlyreduced in adolescents with untreated thyrotoxicosis com-pared with normal controls (990). During antithyroid treat-ment, GH secretion rises in all previously hyperthyroidsubjects, but normalizes only in the subjects in whom eu-thyroidism was achieved. Mean and peak sleep-related GHrelease also are low in hyperthyroid adult patients (991).However, 24-h GH secretion was normal or increased inanother 24-h sampling study in men studied in a more sen-sitive GH IRMA (992). In thyrotoxicosis, serum total immu-noreactive IGF-I levels are either elevated (993) or normal(994). However, IGF-I bioactivity is low and rises after treat-ment (995), which speculatively may be due to concomitantalterations in the IGF-I BPs. Thyroid status also may influ-ence IGF-I expression in tissues other than the liver. In fact,IGF-I production is increased in the hypothalamus after T3administration (995) and plausibly may participate in neg-ative feedback by triggering either increased somatostatintone and/or decreased GHRH production. Such postulatedneuromodulatory responses might account for the low-amplitude (but high-frequency) GH-secretory burstsreported in hyperthyroid men (992).
Thyroid hormone excess consistently impairs GH releaseas stimulated by several secretagogues in man (963). Forexample, the GH-secretory response to insulin-induced hy-poglycemia is markedly impeded in hyperthyroid patients(996), and GHRH’s effects are reduced to less than 50% ofnormal (Fig. 17). Moreover, the timing of GH peaks is de-layed after GHRH injection in hyperthyroidism (997). After1 month of antithyroid drug (methimazole) therapy, hyper-thyroid volunteers achieved serum thyroid hormone levelswithin the normal range, but the GH-secretory response toGHRH was still reduced in amplitude and delayed in time.Although not yet explained, this persisting defect could re-flect chronically reduced pituitary GH stores, possibly due toprolonged GHRH deficiency. The latter hypothesis wouldalso explain why clonidine, a drug that is believed to releaseGH through the stimulation of endogenous GHRH, evokesless than the expected amount of GH secretion in hyperthy-roid subjects (594). This putative GHRH-GH deficiency stateresolves when euthyroidism is maintained for at least 3months (997) (Fig. 17).
Further clinical studies have examined the foregoing
hypothesis of deficient GHRH release and/or action inhyperthyroidism. In normal individuals, b-adrenergicblockade enhances GH-secretory responses to variouspharmacological stimuli, including GHRH (599, 600). Inpatients with hyperthyroidism, GH responses to GHRHare still enhanced by pretreatment with the nonspecificb-receptor blocker, propranolol (998). Since many of theclinical symptoms of thyrotoxicosis are attributed to sym-pathetic overactivity, b2-adrenergic hyperstimulation it-self could contribute to the reduced GH response topharmacological stimuli observed in patients with hyper-thyroidism.
Other mechanistic investigations in thyrotoxicosis haveexploited agents presumed to decrease hypothalamic soma-tostatin release. For example, the lack of effect of pyridostig-mine in hyperthyroidism suggests that the diminution in GHsecretion is not due solely to reduction in the activity ofcholinergic pathways (999). On the other hand, l-arginine isable to enhance, but not normalize, the GH response toGHRH in patients with hyperthyroidism (1000). Thus, anyinferred excess in hypothalamic somatostatin release in hy-perthyroidism is not likely to represent the exclusive causeof suppressed GH secretion. Alternatively, we hypothesizethat hyperthyroidism may suppress GH secretion directly atthe pituitary level by impairing GHRH receptor activityand/or inhibiting GH biosynthesis or storage. The latterconjecture may explain both the blunting of the serum GHresponse to GHRH in hyperthyroid patients and the slowrecovery of pituitary responsivity to GHRH after correctionof hyperthyroidism (997).
Hyperthyroidism decreases GH release after GHRH stim-ulation alone or combined with GHRP-6, but not afterGHRP-6 alone (1001). To our knowledge, altered expressionof the recently cloned GHRP receptor in the hypothalamusand/or pituitary gland in hyperthyroidism remains un-studied.
The foregoing studies taken together suggest, but do notestablish, that reduced GH release stimulated by GHRH inhyperthyroidism is explained, at least in part, by an in-crease in hypothalamic somatostatin tone with concurrentGHRH deficiency and suppressed pituitary GH produc-tion. Whether these putative alterations are caused di-rectly by excess thyroid hormone or by the elevated cir-culating IGF-I levels remains to be elucidated.Alternatively, deficient GH responses to acute pharmaco-logical challenges may be due to either sustained GHRHdeficiency leading to depletion of somatotrope GH storesor to direct inhibitory effects of thyroid hormone excess onsomatotropes.
VIII. Regulation of the GH Axis Throughout theHuman Lifetime
Available calculations indicate that a nearly 125- to 150-fold spectrum of daily GH secretion rates operates normallywithin the healthy human adult and childhood population(78). The lowest GH secretion rates are observed in older orobese adults, and those with hypothyroidism or type II (non-insulin-dependent) diabetes mellitus (79, 107). Indeed, total
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 763
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
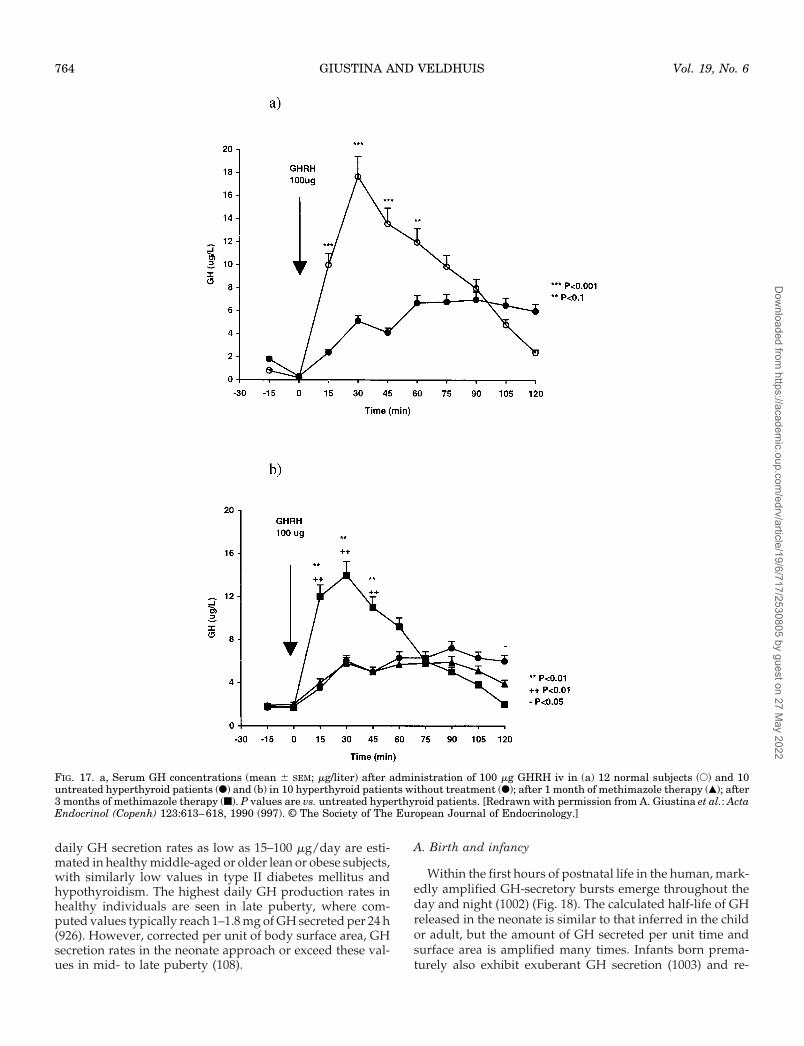
daily GH secretion rates as low as 15–100 mg/day are esti-mated in healthy middle-aged or older lean or obese subjects,with similarly low values in type II diabetes mellitus andhypothyroidism. The highest daily GH production rates inhealthy individuals are seen in late puberty, where com-puted values typically reach 1–1.8 mg of GH secreted per 24 h(926). However, corrected per unit of body surface area, GHsecretion rates in the neonate approach or exceed these val-ues in mid- to late puberty (108).
A. Birth and infancy
Within the first hours of postnatal life in the human, mark-edly amplified GH-secretory bursts emerge throughout theday and night (1002) (Fig. 18). The calculated half-life of GHreleased in the neonate is similar to that inferred in the childor adult, but the amount of GH secreted per unit time andsurface area is amplified many times. Infants born prema-turely also exhibit exuberant GH secretion (1003) and re-
FIG. 17. a, Serum GH concentrations (mean 6 SEM; mg/liter) after administration of 100 mg GHRH iv in (a) 12 normal subjects (E) and 10untreated hyperthyroid patients (F) and (b) in 10 hyperthyroid patients without treatment (F); after 1 month of methimazole therapy (Œ); after3 months of methimazole therapy (f). P values are vs. untreated hyperthyroid patients. [Redrawn with permission from A. Giustina et al.: ActaEndocrinol (Copenh) 123:613–618, 1990 (997). © The Society of The European Journal of Endocrinology.]
764 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
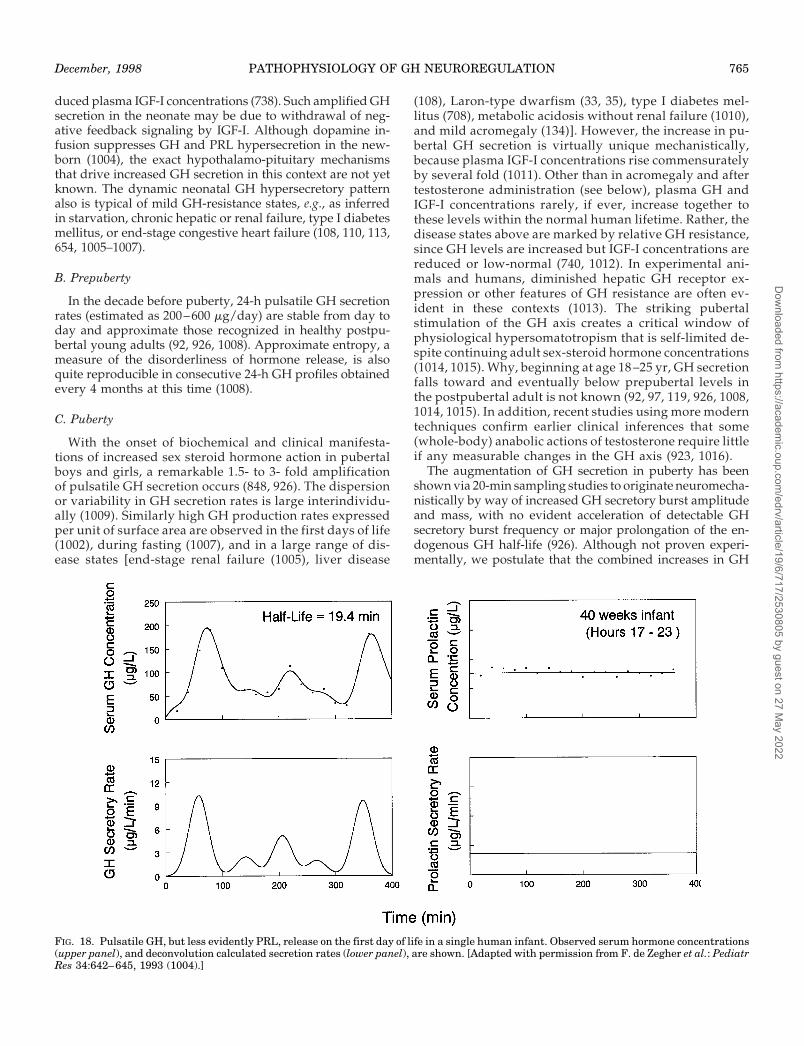
duced plasma IGF-I concentrations (738). Such amplified GHsecretion in the neonate may be due to withdrawal of neg-ative feedback signaling by IGF-I. Although dopamine in-fusion suppresses GH and PRL hypersecretion in the new-born (1004), the exact hypothalamo-pituitary mechanismsthat drive increased GH secretion in this context are not yetknown. The dynamic neonatal GH hypersecretory patternalso is typical of mild GH-resistance states, e.g., as inferredin starvation, chronic hepatic or renal failure, type I diabetesmellitus, or end-stage congestive heart failure (108, 110, 113,654, 1005–1007).
B. Prepuberty
In the decade before puberty, 24-h pulsatile GH secretionrates (estimated as 200–600 mg/day) are stable from day today and approximate those recognized in healthy postpu-bertal young adults (92, 926, 1008). Approximate entropy, ameasure of the disorderliness of hormone release, is alsoquite reproducible in consecutive 24-h GH profiles obtainedevery 4 months at this time (1008).
C. Puberty
With the onset of biochemical and clinical manifesta-tions of increased sex steroid hormone action in pubertalboys and girls, a remarkable 1.5- to 3- fold amplificationof pulsatile GH secretion occurs (848, 926). The dispersionor variability in GH secretion rates is large interindividu-ally (1009). Similarly high GH production rates expressedper unit of surface area are observed in the first days of life(1002), during fasting (1007), and in a large range of dis-ease states [end-stage renal failure (1005), liver disease
(108), Laron-type dwarfism (33, 35), type I diabetes mel-litus (708), metabolic acidosis without renal failure (1010),and mild acromegaly (134)]. However, the increase in pu-bertal GH secretion is virtually unique mechanistically,because plasma IGF-I concentrations rise commensuratelyby several fold (1011). Other than in acromegaly and aftertestosterone administration (see below), plasma GH andIGF-I concentrations rarely, if ever, increase together tothese levels within the normal human lifetime. Rather, thedisease states above are marked by relative GH resistance,since GH levels are increased but IGF-I concentrations arereduced or low-normal (740, 1012). In experimental ani-mals and humans, diminished hepatic GH receptor ex-pression or other features of GH resistance are often ev-ident in these contexts (1013). The striking pubertalstimulation of the GH axis creates a critical window ofphysiological hypersomatotropism that is self-limited de-spite continuing adult sex-steroid hormone concentrations(1014, 1015). Why, beginning at age 18 –25 yr, GH secretionfalls toward and eventually below prepubertal levels inthe postpubertal adult is not known (92, 97, 119, 926, 1008,1014, 1015). In addition, recent studies using more moderntechniques confirm earlier clinical inferences that some(whole-body) anabolic actions of testosterone require littleif any measurable changes in the GH axis (923, 1016).
The augmentation of GH secretion in puberty has beenshown via 20-min sampling studies to originate neuromecha-nistically by way of increased GH secretory burst amplitudeand mass, with no evident acceleration of detectable GHsecretory burst frequency or major prolongation of the en-dogenous GH half-life (926). Although not proven experi-mentally, we postulate that the combined increases in GH
FIG. 18. Pulsatile GH, but less evidently PRL, release on the first day of life in a single human infant. Observed serum hormone concentrations(upper panel), and deconvolution calculated secretion rates (lower panel), are shown. [Adapted with permission from F. de Zegher et al.: PediatrRes 34:642–645, 1993 (1004).]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 765
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

secretion and plasma IGF-I concentrations in boys and girlsin puberty reflect decreased sensitivity of the hypothalamo-pituitary unit to effective feedback inhibition by GH and/orIGF-I during the active growth phase. This could be due toaltered IGFBP, IGF-I clearance, or (more likely) central hy-pothalamo-pituitary changes in GH drive and in autofeed-back sensitivity.
The unique state of physiological GH hypersecretion innormal puberty is driven by sex-steroid hormones and canbe recapitulated in part by administering small doses ofestrogen to girls with ovarian dysgenesis (847, 934) orparenteral testosterone to boys with constitutionally de-layed puberty (925, 927) or hypogonadotropic hypogo-nadism (928). During oral estrogen treatment, plasmaIGF-I concentrations often decrease, whereas in responseto parenteral testosterone administration, plasma IGF-Ilevels typically increase concurrently with GH hyperse-cretion. The latter more closely resembles normal pubertyin boys. Notably, either estrogen or testosterone treatmentselectively augments the mass of GH released per secre-tory burst, which is likely to be the dominant neuroen-docrine mechanism subserving the physiological relativehypersomatotropism of puberty. Although sex steroidsexert many direct effects on the liver, the testosterone-stimulated rise in plasma IGF-I concentrations is believedto require pituitary GH release, since hypopituitary boyswho are markedly GH-deficient fail to show a robust risein plasma IGF-I levels during testosterone repletion (1017,1018).
Serum total and free testosterone concentrations stronglypredict the total daily GH secretion rate in pubertal boys andmen (926, 928). Serum GH concentrations in girls at differentphases of puberty and in menstruating women at differentstages of the cycle are positively correlated with serum es-tradiol levels (939, 1019). In a cross-sectional study in an adultpopulation of both sexes spanning a wide age range, astrongly positive correlation between total or free estradioland peak GH concentrations also pertained (1020). Longi-tudinal studies in boys and girls as they enter and traversepuberty will be required to clarify further the temporal link-ages between endogenous androgen and estrogen concen-trations and increased GH secretion.
D. Adulthood
Although incompletely explained, serum GH concentra-tions and pulsatile GH secretion rates fall in adulthood to lessthan one-quarter or one-half of the maximal values achievedin mid- to late puberty (929, 1021–1023) (see Fig. 19). Indeed,there is an exponential decline (t1/2 of 7 yr) in the 24-h meanserum GH concentration in both men and women beginningat the late pubertal-young adult transition (ages 18–25) andcontinuing into the middle years of life. This decline occursdespite adult levels of sex steroid hormones and is accom-panied by a gradual fall in plasma IGF-I concentrations. Theneurotransmitter mechanism(s) subserving this progressivedecline or “reset” of the hypothalamo-somatotrope-IGF-Iaxis in healthy aging is (are) not known. The term “somato-pause” has been considered to describe this state.
In addition to sex steroids (see above) and age, other im-
portant factors such as body composition (e.g., visceral obe-sity) and exercise capacity substantially influence physio-logically pulsatile GH secretion in the adult. These factorsoperate primarily via an impact on GH-secretory burst am-plitude or mass, without major changes in GH-secretoryburst frequency or GH half-life (112, 749, 929).
Recent studies indicate that gender itself is a strong mod-ifier of the impact of age, obesity, and physical fitness on GHsecretion (749). For example, as shown in Fig. 20, comparedwith men, healthy women of premenopausal age (and thuswith normal premenopausal concentrations of estrogen) aresignificantly protected, by approximately 2-fold, from thenegative impact of increasing age, relative adiposity, anddiminishing physical fitness on daily GH secretion (assum-ing nominally similar GH half-lives in men and women, andsimilar weight-adjusted GH distribution volumes). For ex-ample, mean 24-h serum GH concentrations decline twice asrapidly with increasing age or increasing percentage body fatin men as in women between the ages of 21 and 48 (749).Based on earlier studies, this gender difference vanishes inthe hormone-unreplaced postmenopausal age group (1024).Increasing total adiposity (as quantitated by underwaterweighing to estimate total percentage body fat) also exerts anapproximately 2-fold greater negative impact on mean se-rum GH concentrations in men than premenopausal women.Indeed, in a recent study, the linear relationship betweenpercentage body fat and GH concentrations was significantlynegative in men volunteers (n 5 12), but not in the pre-menopausal women subjects (n 5 32) (749). Furthermore,increased physical fitness, as defined objectively by a highermaximal rate of oxygen consumption, is a positive predictorof serum GH concentrations in men, but the strength of thiseffect (reflected in the standardized regression coefficient) isreduced by approximately 50% in premenopausal women(749). Consequently, gender has a major impact on the ex-pression of several critical determinants of GH secretion,including age, body composition, and physical fitness.
Since sex-steroid hormones can modify body composition,which in turn strongly influences GH secretion, the queryappropriately arises whether gonadal steroid hormones actdirectly or via altered body composition to regulate GHrelease. A recent multilinear regression analysis revealedthat visceral fat mass as quantitated by computerized axialtomography accounted for the majority of the variability inmean 24-h serum GH concentrations in middle-aged menand women (750). However, sex steroids and GH itself altertopographic fat distribution in the long term, thus requiringappropriate interventional studies to resolve the relativeroles of sex steroid concentrations vs. body composition asprimary regulators of the GH axis in older individuals.
Mechanistic clinical studies indicate that testosterone andestradiol both stimulate GH secretion and do so via a com-mon neuroendocrine mechanism of increased GH secretoryburst amplitude or mass (928, 1025). This steroidal effect islikely to be mediated by the estrogen receptor (142, 848, 930)and could in principle be brought about by withdrawal ofsomatostatin’s inhibitory tone, amplification of endogenousGHRH release or its pituitary actions, and/or recruitment ofother mechanisms that stimulate GH release (e.g., via thetheoretical endogenous ligand for the GHRP receptor). Stud-
766 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

ies do not yet clarify whether sex steroid hormones in theadult might also reduce the negative feedback efficacy ofIGF-I and/or GH in suppressing GH release. However,marked changes in the so-called approximate entropy ororderliness of the GH release process at puberty (Fig. 16)strongly suggest that feedback changes occur within the GH-IGF-I axis at this time (142).
In addition to alterations in the pulsatile mode of GHrelease throughout the human lifetime, a recent observationis that a novel regularity statistic, approximate entropy(ApEn), can provide a measure of the regularity, orderliness,
or subpattern reproducibility in hormone release profiles.Orderliness of GH release declines significantly in acromeg-aly and at puberty, is lower in women compared with men,as well as with greater relative or absolute adiposity, withaging, and during estrogen or testosterone (but not DHT)treatment (81, 121, 134, 136, 139, 142, 261). The exact neu-roendocrine mechanisms that govern the moment-to-moment regularity of the GH release process are not known,but probably involve feedback control, and represent animportant focus for further study.
Investigations of spontaneous GH secretion in the human
FIG. 19. Exponential decline with age and/or relative adiposity of 24-h deconvolution-calculated GH secretion rates (mg/liter/day) in a group of 21men of variable ages (top panel) and body mass indices (BMI, a measure of relative adiposity) (middle panel). There was also a strongly negativeinteractive effect of age and BMI (bottom panel). The half-time of decline in the daily GH secretion rate was 7 yr (upper panel) or 1.5 kg/m2 (BMI,middle). [Adapted with permission from A. Iranmanesh et al. (929) and J. D. Veldhuis et al. (1021) with permission from Elsevier Science.]
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 767
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

FIG. 20. A, Impact of gender on the effects of age, adiposity as measured by BMI or percentage body fat, and physical fitness as quantitatedby maximal oxygen consumption (VO2 peak or max) on integrated (24-h) serum GH concentrations in normal men (closed circles, n 5 12) andwomen (open circles, n 5 32). Linear regression plots are given for each sex. The solid lines denote regressions in men, and the interruptedlines depict women’s data. B, Approximately 2-fold greater impact of age, BMI, percentage body fat, and VO2 max on 24-h mean serum GHconcentrations in men than women. Data are means 6 SEM expressed as standardized regression coefficients for the regression lines in panelA. The gender-specific standardized regression coefficient is the slope of the linear relationship (given as a percentage) adjusted per unit SDof the male or female group as pertinent. [Redrawn with permission from A. Weltman et al.: J Clin Endocrinol Metab 78:543–548, 1994 (749).© The Endocrine Society.]
768 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

(and rat) must be understood critically with respect to pre-vailing technical limitations. First, with contemporary meth-odologies, GH secretion rate estimates are stable on succes-sive control days in healthy men (81). Second, the majorityof prior RIA and IRMA estimates of serum GH concentra-tions had an absolute sensitivity of 0.1–0.5 mg/liter. Thus, inmiddle-aged and older, obese, hypothyroid, or fed healthyhumans, daytime serum GH concentrations can fall belowsuch assay thresholds in 20–97% of blood samples (118, 119).Consequently, the recent emergence of immunofluorometricand chemiluminescence GH assays with 10- to 30-fold in-creased sensitivity have provided new insights into the reg-ulation of GH release during daytime hours and in selectedpathophysiologies (118, 119). For example, an ultrasensitivechemiluminescence assay disclosed the existence of a 140-fold range in mean serum GH concentrations in healthy men.In addition, the assay suggested that a low rate of basal GHsecretion exists between distinct GH-secretory bursts. Sim-ilar results were obtained in three independent ELISA-basedstudies (122–124). Notably, low basal rates of GH releasewere correlated to the serum concentration of estrogen aswell as influenced by age and body composition (120). In thehigh-sensitivity chemiluminescence assay, all blood samplesin more than 100 individuals spanning a range of ages andbody compositions and pathophysiologies have containeddetectable GH concentrations (sensitivity limits 0.002 mg/liter at 2 sds and 0.005 mg/liter at 4 sds above blank) (119).This assay further revealed that serum GH concentrations aresuppressed by glucose to less than 0.7 mg/liter in healthywomen and less than 0.07 mg/liter in healthy men, evincinga novel sex distinction in GH axis function (120). Other stud-ies with a high-sensitivity (0.013 mg/liter threshold) immun-ofluorometric assay in middle-aged and perimenopausalwomen and age-matched men show that the quantitativebasis for the gender difference in pulsatile GH release arisesfrom higher GH-secretory burst mass (pulse amplitude andduration) in women compared with men, with no evidentdifferences in the GH half-life or GH-secretory burst fre-quency (118).
Postpartum relative hyposomatotropism in the mother isalso recognized in the human (1026). Although the neuroen-docrine mechanisms are not established, suppression of pi-tuitary GH secretion by the placental GH variant (V) syn-thesized during pregnancy is possible (1027–1029).Interestingly, neither GHRH nor GHRP-1 is an effective GHsecretagogue in this context, until PRL levels decline peri-partally (1028). If GH autonegative feedback is relevant inpostpartum maternal GH deficiency, the reasons for suchsustained inhibition of the GH axis in lactating women re-mains enigmatic.
E. Aging
The manifold changes in the GHRH-somatostatin-GH-IGF-I axis with healthy aging were reviewed (83, 84, 1021,1030–1033). Additional insights have emerged into the mech-anisms and/or treatment issues concerning relative GH de-ficiency in healthy older individuals, as discussed brieflybelow. The reduction in GH secretion with healthy agingbegins after the pubertal maximum and is correlated with
increased percentage total body and visceral fat, decreasedphysical fitness, and decreased testosterone concentrationsin men, or the menopause in women (84, 121). The age effectin younger adults is more evident in men than premeno-pausal women (above) (749).
The response of pituitary GH release as stimulated bymultiple individual secretagogues is decreased in aging, e.g.,sleep, exercise, l-dopa, insulin, l-arginine, GHRH, GHRP,and clonidine (1032, 1034, 1035). However, intravenous (ororal) l-arginine, which is presumed to withdraw somatosta-tin, enhances GHRH’s stimulatory effect on GH release in theelderly by 4-fold, and these combined secretagogues nearlyevoke normal GH secretion acutely (1036). Similarly, l-arginine in combination with GHRP reinstates a virtuallyyoung adult level of acutely stimulated GH release in olderhumans (1037). In the dog, pharmacologically induced so-matostatin withdrawal elevates GH release in older animals(574, 1038), thus also suggesting a partial role for somatosta-tin excess in the hyposomatotropism of aging. A recent clin-ical study in healthy men of varying ages receiving 2 days oforal pyridostigmine treatment (putatively to withdraw so-matostatin and secondarily stimulate GHRH release) sup-port an inference of combined somatostatin excess andGHRH deficiency with aging (261). Such treatment nearlynormalized mean 24-h serum GH and IGF-I concentrationsin older men, compared with levels attained in younger menreceiving the same intervention, but failed to overcome thehyposomatotropism associated with increased total percent-age body fat (see Fig. 21).
Other experiments more directly support a collaborativerole for GHRH deficiency in human aging-associated hypo-somatostropism. For example, a 3-day intravenous GHRHpulsatile infusion fails to fully normalize GH secretion ineither relatively obese or older men (228) (Fig. 22). An in-dependent earlier investigation showed that 15 days ofGHRH treatment alone once daily and/or combined withl-arginine failed to fully normalize GH secretion in oldervolunteers (1039). Another study examined the effects ofhealthy aging on the GH response to acute GHRH stimula-tion alone or in combination with either the neuropeptide,galanin, or the acetylcholinesterase inhibitor, pyridostig-mine. Pretreatment with either galanin or pyridostigmineenhanced, but did not normalize, the GH response to GHRHin aged subjects (443), which is consistent (as above) withcombined GHRH deficiency and somatostatin excess inhealthy aging humans (228, 261, 1036, 1037). This notion issupported by recent studies showing diminished somatosta-tin-withdrawal-induced “rebound” GH secretion (thoughtto be driven by endogenous GHRH activation) in older menand women (1040).
Overall, we believe that available clinical data supportboth excessive somatostatin release and diminished GHRHsecretion in the aging human, as also suggested earlier in theaging dog (574, 1038, 1041). A deficiency of the putativeligand for GHRP is also a compelling speculation at present,since l-arginine and GHRP (or l-arginine and GHRH) vir-tually normalize acute GH release in older individuals(above). Impoverished GH secretion in aging is not likelyattributable to augmented IGF-I negative feedback since, if
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 769
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

anything, older men and women show relative resistance toinfused IGF-I’s inhibition of GH release (1042).
Of analogous interest to the aging phenomenon, pro-longed critical illness is associated with deficiencies of var-ious neuroendocrine axes, e.g., gonadotropic, somatotropic,and thyrotropic (1043). Recent studies suggest that a hypo-thalamic component of relative deficiency of releasing factorsplays a role (319, 320). Indeed, combined GHRP and GHRHinfusions in extended critical illness activate GH, IGF-I, andBP-3 production significantly (1044).
F. GH treatment in older humans
Since this topic has been reviewed earlier, the reader isreferred to relevant background (315, 1032, 1045–1047), andonly more recent issues are alluded to here. Our commentsare restricted to GH treatment in healthy volunteers ratherthan obese (763) or ill patients [e.g., congestive heart failure(1012)].
Twice daily injections of GHRH in healthy older men canincrease serum GH and IGF-I concentrations (1045). Short-term GHRH treatment can enhance muscle strength (1048),and long-term treatment can favorably modified body com-
FIG. 21. Inability of 48 h of treatment with the indirect cholinergicagonist, pyridostigmine (60 mg orally every 6 h), to overcome thesuppressive impact of increased body mass index (BMI) on eitherGH-secretory burst mass (panel A) or the 24-h GH secretion rate(panel B) in men. Pyridostigmine putatively withdraws endogenoussomatostatin and concurrently facilitates GHRH release (see text).“M1-P1” denotes the incremental effect of pyridostigmine (Mestinon,M1) vs. placebo (P1) treatment on GH-secretory burst mass or thedaily GH production rate. Corresponding P and r values are given forthe linear (panel A) or exponential (panel B) regressions. By way ofcontrast, the decline of GH secretion with age did not remain signif-icant during pyridostigmine treatment in this healthy male cohort.[Adapted with permission from K. Friend et al.: Eur J Endocrinol137:337–386, 1997 (261).]
FIG. 22. Failure of 3 days of pulsatile intravenous GHRH treatment(0.33 mg/kg iv every 90 min) to overcome fully the negative impact ofage (panel A), BMI (panel C), or percentage body fat as determinedby hydrostatic weighing (panel D) and/or enhance the positive effectof testosterone (panel B) on the mean 24-h serum GH concentrationin 19 healthy men. Linear or exponential regression fits with P andr values are shown. [Adapted with permission from A. Iranmanesh etal. (151, 228).]
770 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

position in elderly people (1049). Single nightly doses ofGHRH for 6 weeks in older men are less effective than mul-tiple daily doses (1048). The role of GHRH therapy remainslargely unexplored otherwise in the biochemical and clinicalhyposomatotropism of aging.
GH replacement in healthy older individuals improvesbody composition (see below) and enhances some measuresof functional capability. For example, in a recent study of 52men aged 70 yr or older, 30 mg/kg of human GH given 3times weekly for 6 months augmented lean body mass by4.3%, reduced body fat mass by 13%, but did not improvestrength or physical endurance, or enhance mentation. Ap-proximately 50% of the subjects experienced adverse inci-dents on this dose, and the GH dose was reduced in a quarterof the patients (1050). Treatment over 4 weeks with eitherrecombinant human GH or IGF-I also decreased total fatmass, and increased lean body mass and nitrogen retentionin elderly women (GH dose 25 mg/kg and effective IGF-Idose 60 mg/kg twice daily) (1051). However, with both ofthese treatments, patients often had side effects, e.g., head-aches, edema, lethargy, joint swelling and pain, and a bloatedsensation. In addition to the difficulties inherent in cost andrepeated injections, parenteral GH treatment can elicit theabove side effects, as well as carpal tunnel syndrome andgynecomastia, and cause mildly decreased insulin sensitivity(1046).
GH treatment objectively improved muscle mass andstrength, but did not rejuvenate myofibrillar protein synthe-sis in men over the age of 60 yr, who received 30 mg/kg threetimes weekly for 3 months (1052). In another treatment study,fat mass decreased and lean body mass increased, but nosystematic increases in muscle strength occurred in an ex-ercise trial in men over the age of 65 yr (GH treatment 20mg/kg/day for 10 weeks, after a 14-week training interval)(1053).
Older men release GH in response to exercise, albeit at areduced level, and can benefit from exercise training withincreased strength, increased muscle mass, and decreasedtotal body fat over 4 months of a resistive exercise program(1054). Nonetheless, resistive exercise in older men andwomen is less effective in stimulating GH release than inyounger individuals (1055). To our knowledge, exactly howGH, testosterone, and/or exercise interact at the muscle levelis not yet known; e.g., the local IGF-I-BP system is likelyinvolved, and testosterone itself stimulates human muscleIGF-I expression, while reciprocally reducing BP-4 levels inmuscle (1056, 1057). However, less is understood concerningother muscle paracrine factors, which also might be regu-lated by GH and/or androgen, such as the recently clonedmyostatin gene/protein, deletion of which markedly aug-ments muscle growth in mice and cows (Belgian Blue andPiedmontese double-muscled cattle) (1058–1061).
Adult-onset organic GH deficiency, which should be dis-tinguished from healthy aging per se, provides a context inwhich GH replacement has demonstrably favorable effectson body composition; e.g., in one recent study of 32 men withadult-onset GH-deficiency, lean body mass increased by 2.5kg, percentage body fat decreased by 4%, and bone mineraldensity rose by 2.4–5.1% over 1.5-yr treatment interval. Thebone mineral density improvement was 5.1% in the lumbar
spine and 2.4% in the femoral neck (1062). How and whetherthese changes translate into enhanced functional quality oflife with consequent employment benefits, reduced bonefracture rates, etc., is not yet established (3, 4, 1033, 1063–1069).
Although GH treatment in variably GH-deficient humansis capable of increasing plasma IGF-I concentrations, a gen-der difference exists in that serum IGF-I levels in hypopitu-itary women are almost 2-fold less responsive to any givenweight-adjusted dose of GH (1033), especially when the pa-tients are receiving estrogen therapy (949). A similar infer-ence was made recently in a large 6-month multicenter (Aus-tralian) GH treatment trial in GH-deficient adults (3), inanalogous patients in Holland (1070), and in a comparable2-year study in Sweden (1063). After a single intravenouspulse of rhGH, greater acute lipolysis occurs in young males,whereas greater suppression of IGFBP-I occurs in older fe-males (1071), thus indicating that both age and gender dif-ferences exist in tissue actions of GH. Moreover, sex differ-ences further depend on the specific endpoint of GH actionstudied.
Given the costs, inconvenience, and side effects of GHreplacement via conventional once-daily injections, long-act-ing novel secretagogues of endogenous GH that maintainpulsatile GH release and presumptively remain susceptibleto negative feedback control (thus, possibly not elicitingpharmacologically increased IGF-I secretion) are therefore ofmajor interest, as summarized recently elsewhere (289, 291,295, 315). For example, a nonpeptidyl mimetic of GHRP,MK0677, administered once daily orally for days or weeks toyoung, older, or obese humans can stimulate sustained pul-satile GH secretion and increase (double) plasma IGF-I con-centrations commensurately (290) (see Fig. 8). Substantialfurther study and discussion will be required to clarify theclinical roles of GHRP-mimetics in boosting endogenous(pulsatile) GH secretion in elderly healthy or frail individ-uals.
Intranasal peptide delivery also requires further explora-tion (see Sections III.C and IV.A). Recent studies suggest thatintranasal peptide administration might evoke some direct(nonsystemically mediated) brain effects. For example, a“nose-brain pathway” has been inferred in recent clinicalinvestigations, which show that vasopressin and cholecys-tokinin elicit rapid CNS electrical changes when given in-tranasally in the human (1072, 1073). CRH may also exertCNS effects (1074), as discussed above for GHRH and GHRP(e.g., Refs. 338, 384–386, and 388).
IX. Exercise-Induced Modulation of the GH Axis
A. Experimental animals
The hamster is one animal model of exercise-inducedmodulation of the GH axis. Borer and Kelch (1075) observedthat increases in serum GH concentrations accompany ex-ercise in the adult female hamster. Rapid growth in thehamster was related to exercise-induced GH secretion.
A more comprehensive study in the rat compared GHrelease in aerobically exercised vs. sedentary male and femaleanimals. Nonexercising male rats exhibited typical pulses of
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 771
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

serum GH concentrations with values of more than 100 mg/liter every 3–4 h with intervening trough periods of less than10 mg/liter (1076). The acutely and chronically exercisedmales manifested a decline in serum GH concentrations dur-ing the actual exercise bout. In principle, such suppression ofGH release could be attributed to a decrease in hypothalamicGHRH and/or an increase in somatostatin secretion. Anal-ysis revealed that GH, GHRH, and somatostatin mRNAs allfell in acutely exercised male rats, suggesting that both hy-pothalamic and pituitary biosynthetic capacities declinewhen untrained animals are challenged with the energy re-quirements of acute exercise. In trained male rats, more pro-longed decreases in plasma GH concentrations and GH andsomatostatin mRNA levels occurred. A concomitant increasein GHRH mRNA occurred, which may reflect the hypotha-lamic response to somatostatin withdrawal (see Fig. 1).
The sedentary female rat (control) exhibited typical “fem-inized” GH secretory patterns, wherein peak serum GH con-centrations before exercise did not exceed 100 ng/ml, andtrough concentrations averaged 5–15 ng/ml. As in the maleanimals, the acute and chronically exercised female exhibiteda decline in GH secretion during the 30-min exercise bout. Atthe molecular level, the acutely exercised female showed a5-fold increase in pituitary GH mRNA concentrations,whereas chronically exercised females exhibited a 2-fold de-crease. Messages for GHRH (increased) and somatostatin(decreased) also changed in chronically exercised females, asin their male counterparts. Thus, in contrast to the human(below), GH secretion in the rat is inhibited in response toacute exercise in both untrained and trained animals of eithersex. However, compelling interspecies comparisons are lim-ited, and the foregoing experiment focused only on low-intensity (aerobic) exercise. However, this work provides thefirst evidence that trained female rats show more rapid GHadaptive responses to exercise than trained males (1076).
B. Humans
Exercise of appropriate intensity and duration serves as apotent physiological stimulus to GH secretion in humans(1077, 1078). In one study, an increase in core body temper-ature was required to elicit GH release with exercise (1079).However, the neuroendocrine mechanisms of exercise-induced GH release remain incompletely elucidated, e.g.,whether the final pathway involves GHRH stimulationand/or decreased somatostatin release, and/or possiblynon-GHRH and nonsomatostatin modulators, such as anendogenous GHRP ligand, galanin, etc. Exercise-inducedGH release is delayed until approximately 15 min into ex-ercise (1080, 1081) and peaks by the end of short-term exer-tion (1080, 1081) or shortly thereafter (1082). Felsing et al.(1077) concluded that high-intensity exercise must exceed 10min to reliably stimulate GH release.
Intersubject variability in peak exercise-induced GH con-centrations is large (1081), some of which variation is attrib-utable to differences in the age, sex, and body compositionof subjects participating in different studies. In addition,exercise-induced GH release may be intensity dependent(1083, 1084). Vanhelder et al. (1083) demonstrated that high-intensity anaerobic work evoked greater GH release than
lower intensity aerobic work, despite similar duration andtotal work effort in the two sessions. In addition, exceedinga threshold intensity of exercise is required for the release ofGH (1083).
Hartley et al. (1084) reported that extremely high-intensityexercise paradoxically attenuates GH release compared withmoderate intensity exercise. However, in this study the high-intensity exercise bout was completed on the same day as themoderate- and low-intensity bouts and was always per-formed last. Others have suggested that multiple bouts ofexercise in a single day decrease exercise-induced GH releaseafter several sessions (1081). The inferred ad seriatim decreasein exercise-induced GH release was alleviated when longerperiods of rest (2–3 h) were introduced between exercisesessions (1081). Conversely, Felsing et al. (1077) showed thatthree bouts of exercise each separated by 1 h evoked esca-lating amounts of GH release. However, the exercise boutsalso increased in duration in the latter study. Recently, Ka-naley et al. (1086) reported that, independently of the lengthof recovery, three bouts of exercise in a single day progres-sively augmented GH secretion. These clinical data suggestto us the hypothesis that high-intensity aerobic exercise canovercome GH autonegative feedback, for which thesis pre-liminary evidence has been obtained recently (1087).
Differences in GH release during exercise between gen-ders are incompletely defined, e.g., whether the mechanismand amount of exercise-induced GH release are similar inboth sexes. Some authors have noted that women tend tohave higher serum GH levels before exercise that do notreturn to baseline within 1 h, but the response pattern toexercise is similar to that of men (1080, 1086). Exercise-induced GH release was similar during all stages of themenstrual cycle in another analysis (1086).
Hartley et al. (1084) reported that 7 weeks of physicaltraining decreased exercise-induced GH release in associa-tion with mild, moderate, and high intensity work. Thisstudy measured only the GH response to acute exercise andevaluated a small number of “recovery” blood samples. Asexpected, there was large variability in resting and exercise-induced GH concentrations both before and after training(1081). Long-term exercise training in women approximatelydoubles integrated 24-h serum GH concentrations on thenonexercising day, and GH pulse amplitude specifically, ifthe training is performed at an intensity greater than theindividual’s lactate threshold (1088).
C. Neural control of GH release during exercise
Epinephrine, norepinephrine, acetylcholine, and opioidsmay modulate exercise-induced GH release. Many clinicalstudies have attempted to discern the individual contribu-tions of these general neurotransmitters to the exercise-stim-ulated release of GH, but no one mechanism is agreed to beprimary. Possibly several mechanisms are operative or in-teractive during exercise-induced GH release. Regardless ofthe neurotransmitters implicated, the final common pathwaylikely involves either increases in GHRH and/or decreasesin somatostatin release and/or action. In addition, a role forputative endogenous GHRP-receptor ligands and other neu-romodulators cannot be excluded.
772 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

1. Adrenergic. Both epinephrine and norepinephrine concen-trations rise significantly in the peripheral blood after high-intensity exercise (1089). However, the temporal patterns ofcirculating GH and catecholamine responses to exercise dif-fer. Although blood catecholamines do not faithfully reflectadrenergic inputs to the hypothalamus, they might stimulatehuman pituitary GH secretion directly, at least as they do inthe rat in vitro (see Section V). More likely, the hypothalamicadrenergic system modulates the exercise-driven release ofGH, since propranolol (a nonselective b-blocker) amplifiesexercise-induced GH secretion (1090). Uusitupa et al. (1090)reported that both a cardioselective b-blocking agent (meto-prolol) and a nonselective b-blocker (propranolol) increasedthe GH response to exercise. Conversely, salbutamol andbroxaterol (b2-agonists) blocked the exercise-induced releaseof GH (608). The consequences of a-receptor blockade areless uniform. Hansen (1091) observed that phentolamine (ana-receptor antagonist) nearly totally suppresses GH releaseduring exercise, whereas Sutton and Lazarus (1092) reportedthat this drug has little effect. The experimental conditionsand the exercise bouts in the two studies were similar, mak-ing the disparity in results difficult to reconcile. However,Hansen (1091) studied only 4 subjects while Sutton and Laza-rus (1092) studied 10. We infer from these data that any roleof a-adrenergic pathways in exercise-stimulated GH releaseis not yet clear, whereas imposed b2-adrenergic neurotrans-mission remains capable of inhibiting the exercise-triggeredsignal(s) directing GH secretion. Thus, b2-receptor input mayoccur either downstream of the primary exercise effectorpathway or be capable of suppressing it.
Exercise-mediated increases in serum GH concentrationsmay also be modulated by the metabolic changes that occurduring maximal exercise. In one study, there were no sig-nificant changes in blood glucose during the four exercisebouts evaluated (608). Although the inhibitory effect of b2-agonists on GH release occurred concomitantly with a slightincrease in preexercise blood glucose levels, the lack of cor-relation between these two variables, as well as the minimalhyperglycemia attained, suggests that a transient glucoseelevation after b2-agonists is not the primary mechanisminvolved in GH suppression. The increase in serum lactate,which normally occurs during maximal exercise (1089), isalso not likely to modify GH secretion, because infusion oflactate in man does not alter GH release (1093). Finally, sincethe b2-agonist-mediated decrease in the GH response to ex-ercise occurs despite an increase in plasma lactate levelsduring the exercise (608), b2-agonists do not inhibit GH re-lease by suppressing lactate production.
2. Cholinergic. Cappa et al. (1094) reported that pyridostig-mine (an indirect cholinergic agonist) administered concom-itantly with an exercise stimulus increased GH releaseadditively. In the rat, pyridostigmine may decrease hypo-thalamic somatostatin release, whereas in the sheep itincreases GHRH secretion (above). Since the combination ofpyridostigmine plus exercise is additive, one could infer thatexercise modifies release of yet an additional (non-GHRH,nonsomatostatin) effector and/or acts more effectively thana submaximally effective dose of pyridostigmine. Thompsonet al. (1095) corroborated that oral pyridostigmine, alone or
in combination with the opiate receptor antagonist, naltrex-one, potentiates exercise-induced GH release. Last, atropine(a muscarinic blocker) inhibits the GH response to exercise(535). In fact, putatively enhanced somatostatinergic tonedue to this cholinergic antagonist impairs GH responsive-ness to several stimuli, including GHRH. Thus, exercise can-not overcome pharmacological antagonism of the cholinergic(muscarinic) pathway or activation of b2-receptor neuro-transmission (above). Observations with atropine suggest,but do not prove, that the exercise signal converges on theGH release pathway at, or proximal to, somatostatinergicsystems normally suppressed by cholinergic inputs.
3. Opioids. The role of opioids in the control of exercise-stimulated GH release is controversial. Experiments inves-tigating the effects of naloxone (an opiate antagonist) onexercise-induced GH release have yielded equivocal results.Moretti et al. (482), using high doses of naloxone and 20 minof high-intensity exercise recorded that naloxone completelyblocked exercise-induced GH release in well trained com-petitive athletes. The ability of naloxone to impede exercise-stimulated GH release in this context may have reflected apropensity of these competitors to perceive greater stress,possibly because of the achievement-oriented task. In con-trast, Coiro et al. (1096) noted that naloxone did not inhibitexercise-induced GH release in subjects who were nottrained athletes. This discrepancy might be explained ifchronic exercise training induces the activity of opioid path-ways regulating GH release during acute exercise. The studyby Coiro et al. (1096) further revealed that the GABA-ergicagonist, sodium valproate, significantly attenuated exercise-elicited GH secretion, and that the combination of sodiumvalproate and naloxone completely abolished GH release.Thus, GABA-ergic mechanism may be able to inhibit the GHresponse to exercise in humans, and an opioid pathway maybe able to relieve the inhibitory effects of GABA.
The interaction of opioid and cholinergic pathways toprompt GH release during exercise was investigated byThompson et al. (1095). Naltrexone (an opioid antagonist) didnot alter resting serum GH concentrations or exercise-stim-ulated GH release. Naltrexone also did not influence the risein mean and integrated serum GH concentrations stimulatedby pyridostigmine, thus disclosing no evident interactionbetween opiatergic and cholinergic pathways in the effect ofexercise. We conclude provisionally that endogenous opi-atergic activation is not crucial for acute exercise-induced GHrelease in untrained individuals, but may be relevant incompetitively trained athletes.
D. Kinetics of exercise-induced GH-release
Lassarre et al. (1080) and Thompson et al. (1095) reportedthat the apparent half-life of endogenous GH during exercisewas lower than that computed at rest (namely ;19 min at restvs. ;16 min after exercise). Since a shorter GH half-life wouldtend to disproportionately reduce the serum GH concentra-tion, the marked rise in blood GH after exercise (assumingno major changes in distribution volume) clearly reflectsamplified GH secretion, rather than decreased GH clearance(1088, 1097). As revealed by deconvolution analyses, the
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 773
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

dominant neuroendocrine mechanism of exercise-inducedGH secretion specifically entails a markedly augmentedmass of GH secreted per burst, which would be consistentwith somatostatin withdrawal and/or amplified GHRH re-lease, or release of one or more cosecretagogues, such as aputative endogenous GHRP receptor ligand, yet to be iden-tified.
X. Summary
During the last decade, the GH axis has become the com-pelling focus of remarkably active and broad-ranging basicand clinical research. Molecular and genetic models, the dis-covery of human GHRH and its receptor, the cloning of theGHRP receptor, and the clinical availability of recombinantGH and IGF-I have allowed surprisingly rapid advances inour knowledge of the neuroregulation of the GH-IGF-I axisin many pathophysiological contexts. The complexity of theGHRH/somatostatin-GH-IGF-I axis thus commends itself tomore formalized modeling (154, 155), since the multivalentfeedback-control activities are difficult to assimilate fully onan intuitive scale. Understanding the dynamic neuroendo-crine mechanisms that direct the pulsatile secretion of thisfundamental growth-promoting and metabolic hormone re-mains a critical goal, the realization of which is challenged bythe exponentially accumulating matrix of experimental andclinical data in this arena.
To the above end, we review here the pathophysiology ofthe GHRH somatostatin-GH-IGF-I feedback axis consistingof corresponding key neurotransmitters, neuromodulators,and metabolic effectors, and their cloned receptors and sig-naling pathways. We propose that this system is best viewedas a multivalent feedback network that is exquisitely sensi-tive to an array of neuroregulators and environmental stres-sors and genetic restraints. Feedback and feedforward mech-anisms acting within the intact somatotropic axis mediatehomeostatic control throughout the human lifetime and aredisrupted in disease. Novel effectors of the GH axis, such asGHRPs, also offer promise as investigative probes and pos-sible therapeutic agents.
Further understanding of the mechanisms of GH neuro-regulation will likely allow development of progressivelymore specific molecular and clinical tools for the diagnosisand treatment of various conditions in which GH secretionis regulated abnormally. Thus, we predict that unexpectedand enriching insights in the domain of the neuroendocrinepathophysiology of the GH axis are likely be achieved in thesucceeding decades of basic and clinical research.
Acknowledgments
A.G. is indebted to Drs. C. Cappelli, F. Negrini, A. Burattin, C. Mas-cadri, and G. Milani for their valuable help, and the support, discussionand scientific guidance of Professor G. Giustina and W. B. Wehrenberg.J.D.V. thanks Patsy Craig for her skillful preparation of the manuscriptand Paula Azimi for the scientific illustrations. J.D.V. is grateful to Dr.Cyril Bowers for insights in the reading of the GHRP section and to thethree anonymous reviewers who made valuable suggestions and cor-rections.
References
1. Wehrenberg WB, Giustina A 1992 Basic counterpoint: mechanisms andpathways of gonadal steroid modulation of growth hormone secretion.Endocr Rev 13:299–308
2. Kerrigan JR, Rogol AD 1992 The impact of gonadal steroid hormone actionon growth hormone secretion during childhood and adolescence. EndocrRev 13:281–298
3. Cuneo RC, Judd S, Wallace JD, Perry-Keene D, Burger H, Lim-Tio S,Strauss B, Stockigt J, Topliss D, Alford F, Hew L, Bode H, Conway A,Handelsman D, Dunn S, Boyages S, Cheung NW, Hurley D 1998 TheAustralian multicenter trial of growth hormone (GH) treatment in GH-deficient adults. J Clin Endocrinol Metab 83:107–116
4. Vance ML 1994 Hypopituitarism. N Engl J Med 330:1651–16625. Campbell GS 1997 Growth-hormone signal transduction. J Pediatr
131[Suppl]:S42–S446. Rosenfeld RG, Rosenbloom AL, Guevara-Aguirre J 1994 Growth hormone
(GH) insensitivity due to primary GH receptor deficiency. Endocr Rev15:369–390
7. Clemmons DR, Underwood LE 1994 Clinical Review 59. Uses of humaninsulin-like growth factor-I in clinical conditions. J Clin Endocrinol Metab79:4–6
8. Baumann G 1995 Editorial: Growth hormone binding protein-errant recep-tor or active player? Endocrinology 136:377–378
9. Shimon I, Melmed S 1997 Genetic basis of endocrine disease: pituitarytumor pathogenesis. J Clin Endocrinol Metab 82:1675–1681
10. Frohman LA 1996 New insights into the regulation of somatotrope functionusing genetic and transgenic models. Metabolism 45:1–3
11. Hammer RE, Brinster RL, Rosenfeld MG, Evans RM, Mayo KE 1985Expression of human growth hormone-releasing factor in transgenic miceresults in increased somatic growth. Nature 315:413–416
12. Kovacs M, Kineman RD, Schally AV, Zarandi M, Groot K, Frohman LA1997 Effects of antagonists of growth hormone-releasing hormone (GHRH)on GH and insulin-like growth factor I levels in transgenic mice overex-pressing the human GHRH gene, an animal model of acromegaly. Endo-crinology 138:4536–4542
13. Flavell DM, Wells T, Wells SE, Carmignanc DF, Thomas GB, Robinson IC1996 Dominant dwarfism in transgenic rats by targeting human growthhormone (GH) expression to hypothalamic GH-releasing factor neurons.EMBO J 15:3871–3879
14. Kineman RD, Aleppo G, Frohman LA 1996 The tyrosine hydroxylase-human growth hormone (GH) transgenic mouse as a model of hypothalamicGH deficiency: growth retardation is the result of a selective reduction insomatotrope numbers despite normal somatotrope function. Endocrinology137:4630–4636
15. Szabo M, Butz MR, Banerjee SA, Chikaraaishi DM, Frohman LA 1995Autofeedback suppression of growth hormone (GH) secretion in transgenicmice expressing a human GH reporter targeted by tyrosine hydroxylase59-flanking sequences to the hypothalamus. Endocrinology 136:4044–4048
16. Zhang J, Obal Jr F, Fang J, Collins BJ, Krueger JM 1996 Non-rapid eyemovement sleep is suppressed in transgenic mice with a deficiency in thesomatotropic system. Neurosci Lett 220:97–100
17. Ikeda A, Matsuyama S, Nishihara M, Tojo H, Takahashi M 1994 Changesin endogenous growth hormone secretion and onset of puberty in transgenicrats expressing human growth hormone gene. Endocr J 41:523–529
18. Hurley DL, Bartke A, Wagner TE, Wee BE, Phelps CJ 1994 Increasedhypothalamic somatostatin expression in mice transgenic for bovine orhuman GH. J Neuroendocrinol 6:539–548
19. Bertherat J, Timsit J, Bluet-Pajot M-T, Mercadier J-J, Gourdji D, KordonC, Epelbaum J 1993 Chronic growth hormone (GH) hypersecretion inducesreciprocal and reversible changes in mRNA levels from hypothalamic GH-releasing hormone and somatostatin neurons in the rat. J Clin Invest 91:1783–1791
20. Gage PJ, Lossie AC, Scarlett LM, Lloyd RV, Camper SA 1995 Ames dwarfmice exhibit somatotrope commitment but lack growth hormone-releasingfactor response. Endocrinology 136:1161–1167
21. Phelps CJ, Dalcik H, Endo H, Talamantes F, Hurley DL 1993 Growthhormone-releasing hormone peptide and mRNA are overexpressed in GH-deficient Ames dwarf mice. Endocrinology 133:3034–3037
22. Pellegrini E, Carmignac DF, Bluet-Pajot MT, Mounier F, Bennett P, Epel-baum J, Robinson IC 1997 Intrahypothalamic growth hormone feedback:from dwarfism to acromegaly in the rat. Endocrinology 138:4543–4551
23. Bartke A, Cecim M, Tang K, Steger RW, Chandrashekar V, Turyn D 1994Neuroendocrine and reproductive consequences of overexpression ofgrowth hormone in transgenic mice. Proc Soc Exp Biol Med 206:345–359
24. Bjorntorp P 1996 Growth hormone, insulin-like growth factor-I and lipidmetabolism: interactions with sex steroids. Horm Res 46:188–191
25. Cecim M, Kerr J, Bartke A 1995 Infertility in transgenic mice overexpressingthe bovine growth hormone gene: luteal failure secondary to prolactindeficiency. Biol Reprod 52:1162–1166
26. D’Ercole AJ 1993 Expression of insulin-like growth factor-I in transgenicmice. Ann NY Acad Sci 692:149–160
774 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

27. Beck KD, Powell-Braxton L, Widmer HR, Valverde J, Hefti F 1995 IGF-Igene disruption results in reduced brain size, CNS hypomyelination, andloss of hippocampal granule and striatal parvalbumin-containing neurons.Neuron 14:717–730
28. Liu J-P, Baker J, Perkins AS, Robertson EJ, Efstratiadis A 1993 Mice car-rying null mutations of the genes encoding insulin-like growth factor I(IGF-I) and a type I IGF receptor (IGF-Ir). Cell 75:59–72
29. Blackburn A, Schmitt A, Schmidt P, Wanke R, Hermanns W, Brem G, WolfE 1997 Actions and interactions of growth hormone and insulin-like growthfactor II: body and organ growth of transgenic mice. Transgenic Res 6:213–222
30. Rajkumar K, Barron D, Lewitt MS, Murphy LJ 1995 Growth retardationand hyperglycemia in insulin-like growth factor binding protein-1 trans-genic mice. Endocrinology 136:4029–4034
31. Pursel VG, Rexroad Jr CE 1993 Status of research with transgenic farmanimals. J Anim Sci 71:10–19
32. Udy GB, Towers RP, Snell RG, Wilkins RJ, Park SH, Ram PA, WaxmanDJ, Davey HW 1997 Requirement of STAT5b for sexual dimorphism of bodygrowth rates and liver gene expression. Proc Natl Acad Sci USA 94:7239–7244
33. Laron Z, Pertzelan A, Karp M 1968 Pituitary dwarfism with high serumlevels of growth hormone. Isr J Med Sci 4:883–894
34. Savage MO, Blum WF, Ranke MB, Postel-Vinay MC, Cotterill AM, HallK, Chatelain PG, Preece MA, Rosenfeld RG 1993 Clinical features andendocrine status in patients with growth hormone insensitivity (Laron syn-drome). J Clin Endocrinol Metab 77:1465–1471
35. Laron Z 1984 Laron-type dwarfism (herediatry somatomedin deficiency): areview. In: Frick P, von Harnack GA, Kochsiek K, Martini GA, Prader A (eds)Advances in Internal Medicine and Pediatrics. Springer-Verlag, Berlin, pp117–150
36. Zhou Y, Xu BC, Maheshwari HG, He L, Reed M, Lozykowski M, OkadaS, Cataldo L, Coschigamok K, Wagner TE, Baumann G, Kopchick JJ 1997A mammalian model for Laron syndrome produced by targeted disruptionof the mouse growth hormone receptor/binding protein gene (the Laronmouse). Proc Natl Acad Sci USA 94:13215–13220
37. Chen NY, Chen WY, Kopchick JJ 1997 Liver and kidney growth hormone(GH) receptors are regulated differently in diabetic GH and GH antagonisttransgenic mice. Endocrinology 138:1988–1994
38. Chen NY, Chen WY, Striker LJ, Kopchick JJ 1997 Co-expression of bovinegrowth hormone (GH) and human GH antagonist genes in transgenic mice.Endocrinology 138:851–854
39. Chen L, Lund PK, Burgess SB, Rudisch BE, McIlwain DL 1997 Growthhormone, insulin-like growth factor I, and motoneuron size. J Neurobiol32:32–202
40. Burrows HL, Birkmeier TS, Seasholtz AF, Camper SA 1996 Targeted ab-lation of cells in the pituitary primordia of transgenic mice. Mol Endocrinol10:1467–1477
41. Lew D, Brady H, Klausing K, Yaginuma K, Theill LE, Stauber C, Karin M,Mellon PL 1993 GHF-1-promoter-targeted immortalization of a somato-tropic progenitor cell results in dwarfism in transgenic mice. Genes Dev7:683–693
42. Akita S, Readhead C, Stefaneanu L, Fine J, Tampanaru-Sarmesiu A, Ko-vacs K, Melmed S 1997 Pituitary-directed leukemia inhibitory factor trans-gene forms Rathke’s cleft cysts and impairs adult pituitary function. A modelfor human pituitary Rathke’s cysts. J Clin Invest 99:2462–2469
43. Pomp D, Oberbauer AM, Murray JD 1996 Development of obesity follow-ing inactivation of a growth hormone transgene in mice. Transgenic Res5:13–23
44. Matsumoto K, Kakidani H, Anzai M, Nakagata N, Takahashi A, TakahashiY, Miyata K 1995 Evaluation of an antisense RNA transgene for inhibitinggrowth hormone gene expression in transgenic rats. Dev Genet 16:273–277
45. Dahler A, Wade RP, Muscat GE, Waters MJ 1994 Expression vectors en-coding human growth hormone (hGH) controlled by human muscle-specificpromoters: prospects for regulated production of hGH delivered by myo-blast transfer or intravenous injection. Gene 145:305–310
46. Draghia-Akli R, Li X, Schwartz RJ 1997 Enhanced growth by ectopic ex-pression of growth hormone releasing hormone using an injectable myo-genic vector. Nat Biotechnol 15:1285–1289
47. Baumann G, Maheshwari H 1997 The Dwarfs of Sindh: severe growthhormone (GH) deficiency caused by a mutation in the GH-releasing hor-mone receptor gene. Acta Pediatr Suppl 432:33–38
48. Wajnrajch MP, Gertner JM, Harbison MD, Chua Jr SC, Leibel RL 1996Nonsense mutation in the human growth hormone-releasing hormone re-ceptor causes growth failure analogous to the little (lit) mouse. Nat Genet12:88–90
49. Woods KA, Camacho-Hubner C, Savage MO, Clark AJ 1996 Intrauterinegrowth retardation and postnatal growth failure associated with deletion ofthe insulin-like growth factor I gene. N Engl J Med 335:1363–1367
50. Cao Y, Wagner JK, Hindmarsch PC, Eble A, Mullis PE 1995 Isolated growthhormone deficiency: testing the little mouse hypothesis in man and exclu-sion of mutations within the extracellular domain of the growth hormone-releasing hormone receptor. Pediatr Res 38:962–966
51. Frohman LA, Downs TR, Clarke IJ, Thomas GB 1990 Measurement ofgrowth hormone-releasing hormone and somatostatin in hypothalamic-portal plasma of unanesthetized sheep: spontaneous secretion and responseto insulin-induced hypoglycemia. J Clin Invest 86:17–24
52. Clark RG, Chambers G, Lewin J, Robinson ICAF 1986 Automated repet-itive microsampling of blood: growth hormone profiles in conscious malerats. J Endocrinol 111:27–35
53. Clark RG, Carlsson LMS, Robinson ICAF 1987 Growth hormone secretoryprofiles in conscious female rats. J Endocrinol 114:399–407
54. Tannenbaum GS, Ling N 1984 The interrelationship of growth hormone(GH)-releasing factor and somatostatin in generation of the ultradianrhythm of GH secretion. Endocrinology 115:1952–1957
55. Tannenbaum GS, Rorstad O, Brazeau P 1979 Effects of prolonged fooddeprivation on the ultradian growth hormone rhythm and immunoreactivesomatostatin tissue levels in the rat. Endocrinology 104:1733–1738
56. Plotsky PM, Vale W 1985 Patterns of growth hormone-releasing factor andsomatostatin secretion into the hypophysial-portal circulation of the rat.Science 230:461–463
57. Thomas GB, Cummins JT, Francis H, Sudbury AW, McCloud PI, ClarkeIJ 1991 Effect of restricted feeding on the relationship between hypophysialportal concentrations of growth hormone (GH)-releasing factor and soma-tostatin, and jugular concentrations of GH in ovariectomized ewes. Endo-crinology 128:1151–1158
58. Cataldi M, Magnan E, Guillaume V, Dutour A, Conte-Devolx B, LombardiG, Oliver C 1994 Relationship between hypophyseal portal GHRH andsomatostatin and peripheral GH levels in the conscious sheep. J EndocrinolInvest 17:717–722
59. Dutour A, Briard N, Guillaume V, Magnan E, Cataldi M, Sauze N, OliverC 1997 Another view of GH neuroregulation: lessons from the sheep. EurJ Endocrinol 136:553–565
60. Bertherat J, Bluet-Pajot MT, Epelbaum J 1995 Neuroendocrine regulationof growth hormone. Eur J Endocrinol 132:12–24
61. Cataldi M, Magnan E, Guillaume V, Dutour A, Sauze N, Mazzocchi L,Conte-Devolx B, Oliver C 1994 Acute stress stimulates secretion of GHRHand somatostatin into hypophysial portal blood of conscious sheep. Neu-rosci Lett 178:103–106
62. Yamashita S, Melmed S 1987 Insulin-like growth factor I regulation ofgrowth hormone gene transcription in primary rat pituitary cells. J ClinInvest 79:449–452
63. Fletcher TP, Thomas GB, Dunshea FR, Moore LG, Clarke IJ 1995 IGFfeedback effects on growth hormone secretion in ewes: evidence for actionat the pituitary but not the hypothalamic level. J Endocrinol 144:323–331
64. Hartman ML, Clayton PE, Johnson ML, Celniker A, Perlman AJ, AlbertiKK, Thorner MO 1993 A low dose euglycemic infusion of recombinanthuman insulin-like growth factor I rapidly suppresses fasting-enhancedpulsatile growth hormone secretion in humans. J Clin Invest 91:2453–2462
65. Ciofi P, Croix D, Tramu G 1987 Coexistence of hGHRF and NPY immu-noreactivities in neurons of the arcuate nucleus of the rat. Neuroendocri-nology 45:425–428
66. Meister B, Scanlon MF, Hokfelt T 1990 Occurrence of galanin-like immu-noreactivity in growth hormone-releasing factor (GRF)-containing neuronsof the monkey (Macaca fascicularis) infundibular nucleus and median emi-nence. Neurosci Lett 119:136–139
67. Veldhuis JD, Rogol AD, Johnson ML 1985 Minimizing false-positive errorsin hormonal pulse detection. Am J Physiol 248:E475–E481
68. Veldhuis JD, Johnson ML 1986 Cluster analysis: a simple, versatile androbust algorithm for endocrine pulse detection. Am J Physiol 250:E486–E493
69. Veldhuis JD, Guardabasso V, Rogol AD, Evans WS, Oerter KE, JohnsonML, Rodbard D 1987 Appraising the nature of luteinizing hormone secre-tory events in man. Am J Physiol 252:E599–E605
70. Urban RJ, Evans WS, Rogol AD, Kaiser DL, Johnson ML, Veldhuis JD 1988Contemporary aspects of discrete peak detection algorithms. I. The para-digm of the luteinizing hormone pulse signal in men. Endocr Rev 9:3–37
71. Veldhuis JD, Johnson ML 1994 Testing pulse detection algorithms withsimulations of episodically pulsatile substrate, metabolite, or hormone re-lease. Methods Enzymol 240:377–415
72. Johnson ML, Veldhuis JD 1995 Evolution of deconvolution analysis as ahormone pulse detection method. Methods Neurosci 28:1–24
73. Veldhuis JD, Johnson ML 1992 Deconvolution analysis of hormone data.Methods Enzymol 210:539–575
74. Veldhuis JD, Faria A, Vance ML, Evans WS, Thorner MO, Johnson ML1988 Contemporary tools for the analysis of episodic growth hormone se-cretion and clearance in vivo. Acta Paediatr Scand Suppl 347:63–82
75. Veldhuis JD, Johnson ML 1990 New methodological aspects of evaluatingepisodic neuroendocrine signals. In: Yen SSC, Vale WW (eds) Advances inNeuroendocrine Regulation of Reproduction. Plenum Publishing Corp.,Philadelphia, PA, pp 123–139
76. Veldhuis JD 1996 New modalities for understanding dynamic regulation ofthe somatotropic (GH) axis: explication of gender differences in GH neu-roregulation in the human. J Pediatr Endocrinol Metab 9:237–253
77. Veldhuis JD 1998 Issues in quantifying pulsatile neurohormone release. In:Van de Kar LD (ed) Methods in Neuroendocrinology: The Cellular and
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 775
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

Molecular Neuropharmacology Series. CRC Press, Boca Raton, FL, pp 181–203
78. Hartman ML, Iranmanesh A, Thorner MO, Veldhuis JD 1993 Evaluationof pulsatile patterns of growth hormone release in man. Am J Hum Biol5:603–614
79. Veldhuis JD 1995 The neuroendocrine regulation and implications of pul-satile GH secretion: gender effects. The Endocrinologist 5:198–213
80. Evans WS, Faria AC, Christiansen E, Ho KY, Weiss J, Rogol AD, JohnsonML, Blizzard RM, Veldhuis JD, Thorner MO 1987 Impact of intensivevenous sampling on characterization of pulsatile GH release. Am J Physiol252:E549–E556
81. Friend K, Iranmanesh A, Veldhuis JD 1996 The orderliness of the growthhormone (GH) release process and the mean mass of GH secreted per burstare highly conserved in individual men on successive days. J Clin EndocrinolMetab 81:3746–3753
82. Ghigo E, Arvat E, Gianotti L, Ramunni J, Maccario M, Camanni F 1994Interaction of salbutamol with pyridostigmine and arginine on both basaland GHRH-stimulated GH secretion in humans. Clin Endocrinol (Oxf) 40:799–802
83. Veldhuis JD, Iranmanesh A 1993 Age-dependent changes in growth hor-mone secretion in adults. In: Scherbaum W, Rossmanith WG (eds) RecentAdvances in the Neuroendocrinology of Aging. Walter De Gruyter & Co.,Berlin, pp 71–93
84. Veldhuis JD, Iranmanesh A, Weltman A 1997 Elements in the pathophys-iology of diminished growth hormone (GH) secretion in aging humans.Endocrine 7:41–48
85. Van Cauter E, Kerkhofs M, Caufriez A, Van Onderbergen A, Thorner MO,Copinschi G 1992 A quantitative estimation of growth hormone secretionin normal man: reproducibility and relation to sleep and time of day. J ClinEndocrinol Metab 74:1441–1450
86. Tonshoff B, Schaefer F, Mehls O 1990 Disturbance of growth hormone-insulin-like growth factor axis in uraemia. Implications for recombinanthuman growth hormone treatment. Pediatr Nephrol 4:654–662
87. Smith WJ, Underwood LE, Clemmons DR 1995 Effects of caloric or proteinrestriction on insulin-like growth factor-I (IGF-I) and IGF-binding proteinsin children and adults. J Clin Endocrinol Metab 80:443–449
88. Snegovskay V, Viru A 1993 Elevation of cortisol and growth hormone levelsin the course of further improvement of performance capacity in trainedrowers. Int J Sports Med 14:202–206
89. Slowinska-Srzednicka J, Zgliczynski W, Makowska A, Jeske W, Brzez-inska A, Soszynski P, Zgliczynski S 1992 An abnormality of the growthhormone/insulin-like growth actor-I axis in women with polycystic ovarysyndrome due to coexistent obesity. J Clin Endocrinol Metab 74:1432–1435
90. Holl RW, Hartman ML, Veldhuis JD, Taylor WM, Thorner MO 1991Thirty-second sampling of plasma growth hormone in man: correlation withsleep stages. J Clin Endocrinol Metab 72:854–861
91. Veldhuis JD 1996 Gender differences in secretory activity of the humansomatotropic (growth hormone) axis. Eur J Endocrinol 134:287–295
92. Thompson RG, Rodriguez A, Kowarski A, Blizzard RM 1972 Growthhormone: metabolic clearance rates, integrated concentrations, and produc-tion rates in normal adults and the effect of prednisone. J Clin Invest 51:3193-
93. Veldhuis JD, Carlson ML, Johnson ML 1987 The pituitary gland secretesin bursts: appraising the nature of glandular secretory impulses by simul-taneous multiple-parameter deconvolution of plasma hormone concentra-tions. Proc Natl Acad Sci USA 84:7686–7690
94. Veldhuis JD, Johnson ML 1995 Specific methodological approaches toselected contemporary issues in deconvolution analysis of pulsatile neu-roendocrine data. Methods Neurosci 28:25–92
95. Veldhuis JD, Evans WS, Johnson ML 1995 Complicating effects of highlycorrelated model variables on nonlinear least-squares estimates of uniqueparameter values and their statistical confidence intervals: estimating basalsecretion and neurohormone half-life by deconvolution analysis. MethodsNeurosci 28:130–138
96. Hindmarsh PC, Matthews DR, Brain C, Pringle PJ, Brook CG 1990 Theapplication of deconvolution analysis to elucidate the pulsatile nature ofgrowth hormone secretion using a variable half-life of growth hormone. ClinEndocrinol (Oxf) 32:739–747
97. Albertsson-Wikland K, Rosberg S, Libre E, Lundberg LO, Groth T 1989Growth hormone secretory rates in children as estimated by deconvolutionanalysis of 24-hr plasma concentration profiles. Am J Physiol 257:E809–E814
98. Oerter KE, Guardabasso V, Rodbard D 1986 Detection and characterizationof peaks and estimation of instantaneous secretory rate for episodic pulsatilehormone secretion. Comput Biomed Res 19:170–191
99. Pilo A, Ferrannini E, Navalesi R 1977 Measurement of glucose-inducedinsulin delivery rate in man by deconvolution analysis. Am J Physiol 233:E500–E508
100. Van Cauter E, Honinchx E 1986 Pulsatility of pituitary hormones. In: SchulzH, Lavie P (eds) Ultradian Rhythms in Physiology and Behavior. Springer-Verlag Publishers, Berlin
101. Rebar R, Perlman D, Naftolin F, Yen SSC 1973 The estimation of pituitaryluteinizing hormone secretion. J Clin Endocrinol Metab 37:917–927
102. Toutain PL, Laurentie M, Autefage A, Alvinerie M 1988 Hydrocortisone
secretion: production rate and pulse characterization by numerical decon-volution. Am J Physiol 255:E688–E695
103. Turner RC, Grayburn JA, Newman GB, Nabarro JDN 1972 Measurementof the insulin delivery rate in man. J Clin Endocrinol Metab 33:279–286
104. Veldhuis JD 1997 How does one get at glandular secretion, when onlyhormone concentrations are measured? (Commentary). Clin Endocrinol(Oxf) 46:397–400
105. Hartman ML, Faria AC, Vance ML, Johnson ML, Thorner MO, VeldhuisJD 1991 Temporal structure of in vivo growth hormone secretory events inman. Am J Physiol 260:E101–E110
106. Sartorio A, De Nicolao G, Pizzini G, Liberati D 1997 Non-parametricdeconvolution provides an objective assessment of GH responsiveness toGH-releasing stimuli innormal subjects. Clin Endocrinol (Oxf) 46:387–395
107. Veldhuis JD, Moorman J, Johnson ML 1994 Deconvolution analysis ofneuroendocrine data: waveform-specific and waveform-independent meth-ods and applications. Methods Neurosci 20:279–325
108. Cuneo RC, Hickman P, Wallace JD, Teh BT, Ward G, Veldhuis JD, WatersMJ 1995 Altered endogenous growth factor (GH) secretory kinetics anddiurnal GH-binding protein profiles in adults with chronic liver disease. ClinEndocrinol (Oxf) 43:265–275
109. Haffner D, Schaefer F, Girard J, Ritz E, Mehls O 1994 Metabolic clearanceof recombinant human growth hormone in health and chronic renal failure.J Clin Invest 93:1163–1171
110. Owens D, Strivastave MC, Tompkins CV, Nabarro JDN, Sonksen PH 1973Studies on the metabolic clearance rate, apparent distribution space andplasma half-disappearance time of unlabelled human growth hormone innormal subjects and in patients with liver disease, renal disease, thyroiddisease and diabetes mellitus. Eur J Clin Invest 3:284–294
111. Rabkin R, Pimstone BL, Marks T, Eales L 1972 Disappearance of humangrowth hormone in the anephric non-uraemic and uraemic rat. Horm MetabRes 4:467–474
112. Veldhuis JD, Iranmanesh A, Ho KKY, Waters MJ, Johnson ML, LizarraldeG 1991 Dual defects in pulsatile growth hormone secretion and clearancesubserve the hyposomatotropism of obesity in man. J Clin Endocrinol Metab72:51–59
113. Schaefer F, Baumann G, Faunt LM, Haffner D, Johnson ML, Mercado M,Ritz E, Mehls O, Veldhuis JD 1996 Multifactorial control of the eliminationkinetics of unbound (free) GH in the human: regulation by age, adiposity,renal function, and steady-state concentrations of GH in plasma. J ClinEndocrinol Metab 81:22–31
114. Dubey AK, Hanukoglu A, Hansen BC, Kowarski AA 1988 Metabolic clear-ance rates of synthetic human growth hormone in lean and obese malerhesus monkeys. J Clin Endocrinol Metab 67:1064–1067
115. Keenan D, Veldhuis JD 1997 Stochastic model of admixed basal and pul-satile hormone secretion as modulated by a deterministic oscillator. Am JPhysiol 273:R1182–R1192
116. Keenan DM, Veldhuis JD 1998 A biomathematical model of time-delayedfeedback in the human male hypothalamic-pituitary-Leydig cell axis. Am JPhysiol 275:E157–E176
117. Keenan DM, Veldhuis JD, Yang R 1998 Joint recovery of pulsatile and basalhormone secretion by stochastic nonlinear random-effects analysis. Am JPhysiol, in press
118. Van den Berg G, Veldhuis JD, Frolich M, Roelfsema F 1996 An amplitude-specific divergence in the pulsatile mode of GH secretion underlies thegender difference in mean GH concentrations in men and premenopausalwomen. J Clin Endocrinol Metab 81:2460–2466
119. Iranmanesh A, Grisso B, Veldhuis JD 1994 Low basal and persistent pul-satile growth hormone secretion are revealed in normal and hyposomato-tropic men studied with a new ultrasensitive chemiluminescence assay.J Clin Endocrinol Metab 78:526–535
120. Chapman IM, Hartman ML, Straume M, Johnson ML, Veldhuis JD, Thor-ner MO 1994 Enhanced sensitivity growth hormone (GH) chemilumines-cence assay reveals lower postglucose nadir GH concentrations in men thanwomen. J Clin Endocrinol Metab 78:1312–1319
121. Veldhuis JD, Liem AY, South S, Weltman A, Weltman J, Clemmons DA,Abbott R, Mulligan T, Johnson ML, Pincus SM, Straume M, IranmaneshA 1995 Differential impact of age, sex-steroid hormones, and obesity on basalvs. pulsatile growth hormone secretion in men as assessed in an ultrasen-sitive chemiluminescence assay. J Clin Endocrinol Metab 80:3209–3222
122. Reutens AT, Veldhuis JD, Hoffman DM, Leung KC, Ho KKY 1996 A highlysensitive growth hormone (GH) ELISA uncovers increased contribution ofa tonic mode of GH secretion in adults with organic GH deficiency. J ClinEndocrinol Metab 81:1591–1597
123. Calabresi E, Ishikawa E, Bartolini L, Delitala G, Fanciulli G, Oliva O,Veldhuis JD, Serio M 1996 Somatostatin infusion suppresses GH secretoryburst number and mass in normal men: a dual mechanism of inhibition. Am JPhysiol 270:E975–E979
124. Reutens AT, Hoffman DM, Leung KC, Ho KKY 1995 Evaluation andapplication of a highly sensitive assay for serum growth hormone (GH) inthe study of adult GH deficiency. J Clin Endocrinol Metab 80:480–485
125. Veldhuis JD 1996 What have we learned so far from ultra-sensitive growthhormone (GH) assays? Clin Chem 42:1731–1732
776 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

126. Straume M, Veldhuis JD, Johnson ML 1994 Model-independent quantifi-cation of measurement error: empirical estimation of discrete variance func-tion profiles based on standard curves. Methods Enzymol 240:121–150
127. Straume M, Johnson ML, Veldhuis JD 1998 Statistically accurate estimationof hormone concentrations and associated uncertainties: methodology, val-idation, and applications. Clin Chem 44:116–123
128. Wright N, Wentz B, Papadea N, Giles B, Veldhuis JD, Pandey JP, Bell NH1998 Growth hormone secretion and bone mineral density in prepubertalblack and white boys. J Bone Miner Res, in press
129. Tannenbaum GS, Farhadi-Jou F, Beaudet A 1993 Ultradian oscillation insomatostatin binding in the arcuate nucleus of adult male rats. Endocrinol-ogy 133:1029–1034
130. Pellegrini E, Bluet-Pajot MT, Mounier F, Bennett P, Kordon C, EpelbaumJ 1996 Central administration of a growth hormone (GH) receptor mRNAantisense increases GH pulsatility and decreases hypothalamic somatostatinexpression in rats. J Neurosci 16:8140–8148
131. West CR, Lookingland KJ, Tucker HA 1997 Regulation of growth hormone-releasing hormone and somatostatin from perifused, bovine hypothalamicslices. III. Reciprocal feedback between growth hormone-releasing hormoneand somatostatin. Domest Anim Endocrinol 14:358–366
132. Pincus SM 1991 Approximate entropy as a measure of system complexity.Proc Natl Acad Sci USA 88:2297–2301
133. Pincus SM 1995 Quantifying complexity and regularity of neurobiologicalsystems. Methods Neurosci 28:336–363
134. Hartman ML, Pincus SM, Johnson ML, Matthews DH, Faunt LM, VanceML, Thorner MO, Veldhuis JD 1994 Enhanced basal and disorderly growthhormone secretion distinguish acromegalic from normal pulsatile growthhormone release. J Clin Invest 94:1277–1288
135. Prank K, Kloppstech M, Nowlan SJ, Sejnowski TJ, Brabant G 1996 Self-organized segmentation of time series: separating growth hormone secretionin acromegaly from normal controls. Biophys J 70:2540–2547
136. Van den Berg G, Pincus SM, Frolich M, Veldhuis JD, Roelfsema F 1998Reduced disorderliness of growth hormone release in biochemically inactiveacromegaly after pituitary surgery. Eur J Endocrinol 138:164–169
137. Prank K, Hartman M, Thorner MO, Vance ML, Kloppstech M, VeldhuisJD, Brabant G, Use of a neural network model to distinguish acromegalicfrom normal pulsatile GH release: effect of sampling interval and duration.Program of the 78th annual meeting of The Endocrine Society, San Francisco,CA, 1996 (Abstract P2–274)
138. Prank K, Kloppstech M, Nowlan SJ, Sejnowski TJ, Brabant G 1996 Ran-dom secretion of growth hormone in humans. Phys Rev Lett 77:1909–1911
139. Pincus SM, Gevers E, Robinson ICAF, Van den Berg G, Roelfsema F,Hartman ML, Veldhuis JD 1996 Females secrete growth hormone with moreprocess irregularity than males in both human and rat. Am J Physiol 270:E107–E115
140. Gevers E, Pincus SM, Robinson ICAF, Veldhuis JD 1998 Differential or-derliness of the GH release process in castrate male and female rats. Am JPhysiol 274:R437–R444
141. Pincus SM, Keefe DL 1992 Quantification of hormone pulsatility via anapproximate entropy algorithm. Am J Physiol 262:E741–E754
142. Veldhuis JD, Metzger DL, Martha Jr PM, Mauras N, Kerrigan JR, KeenanB, Rogol AD, Pincus SM 1997 Estrogen and testosterone, but not a non-aromatizable androgen, direct network integration of the hypothalamo-somatotrope (growth hormone)-insulin-like growth factor I axis in the hu-man: evidence from pubertal pathophysiology and sex-steroid hormonereplacement. J Clin Endocrinol Metab 82:3414–3420
143. Veldhuis JD 1998 Neuroendocrine control of pulsatile growth-hormonerelease in the human: relationship with gender. Growth Hormone IGF Res8:49–59
144. Veldhuis JD, Pincus SM 1998 Orderliness of hormone release patterns: acomplementary measure to conventional pulsatile and circadian analyses.Eur J Endocrinol 138:358–362
145. Prank K, Kloppstech M, Brabant G 1997 Neural networks in the analysisof episodic growth hormone release. Hum Reprod Update 3:215–234
146. Van den Berg G, Pincus SM, Veldhuis JD, Frolich M, Roelfsema F 1997Greater disorderliness of ACTH and cortisol release accompanies pituitary-dependent Cushing’s Disease. Eur J Endocrinol 136:394–400
147. Roelfsema F, Pincus SM, Veldhuis JD 1998 Patients with Cushing’s diseasesecrete adrenocorticotropin and cortisol jointly more asynchronously thanhealthy subjects. J Clin Endocrinol Metab 83:688–692
148. Veldhuis JD, Zwart AD, Iranmanesh A 1997 Neuroendocrine mechanismsby which selective Leydig-cell castration unleashes increased pulsatile LHrelease in the human: an experimental paradigm of short-term ketoconazole-induced hypoandrogenemia and deconvolution-estimated LH secretory en-hancement. Am J Physiol 272:R464–R474
149. Pincus SM, Mulligan T, Iranmanesh A, Gheorghiu S, Godschalk M,Veldhuis JD 1996 Older males secrete luteinizing hormone and testosteronemore irregularly, and jointly more asynchronously, than younger males.Proc Natl Acad Sci USA 93:14100–14105
150. Gravholt CH, Veldhuis JD, Christiansen JS 1998 Increased disorderlinessand decreased mass and daily rate of endogenous growth hormone secretion
in adult Turner’s syndrome: the impact of body composition, physical fitnessand treatment with sex hormones. Metabolism, in press
151. Iranmanesh A, South S, Liem AY, Clemmons D, Thorner MO, WeltmanA, Veldhuis JD 1998 Unequal impact of age, percentage body fat, and serumtestosterone concentrations on the somatotropic, IGF-I, and IGF-bindingprotein responses to a three-day intravenous growth-hormone-releasing-hormone (GHRH) pulsatile infusion. Eur J Endocrinol 139:59–71
152. Veldhuis JD, Iranmanesh A, Urban RJ 1997 Primary gonadal failure in menselectively amplifies the mass of follicle stimulating hormone (FSH) secretedper burst and increases the disorderliness of FSH release: reversibility withtestosterone replacement. Int J Androl 20:297–305
153. Pincus SM, Veldhuis JD, Mulligan T, Iranmanesh A, Evans WS 1997Effects of age on the irregularity of LH and FSH serum concentrations inwomen and men. Am J Physiol 273:E989–E995
154. Straume M, Veldhuis JD, Johnson ML 1995 Realistic emulation of highlyirregular temporal patterns of hormone release: a computer-based pulsesimulator. Methods Neurosci 28:220–243
155. Straume M, Chen L, Johnson ML, Veldhuis JD 1995 Systems-level analysisof physiological regulation interactions controlling complex secretory dy-namics of growth hormone axis: a connectionist network model. MethodsNeurosci 28:270–310
156. Reichlin S 1983 Somatostatin. Part I. N Engl J Med 309:1495–1501157. Tannenbaum GS, Painson JC, Lengyel AM, Brazeau P 1989 Paradoxical
enhancement of pituitary growth hormone (GH) responsiveness to GH-releasing factor in the face of high somatostatin tone. Endocrinology 124:1380–1388
158. Patel YC, Srikant C 1986 Somatostatin mediation of adenohypophysialsecretion. Annu Rev Physiol 48:551–567
159. Reichlin S 1983 Somatosatin. Part II. N Engl J Med 309:1556–1563160. Dickerman Z, Guyda H, Tannenbaum GS 1993 Pretreatment with soma-
tostatin analog SMS 201–995 potentiates growth hormone responsiveness togrowth hormone-releasing factor in short children. J Clin Endocrinol Metab77:652–657
161. Ramirez JL, Torronteras R, Castano JP, Sanchez-Hormigo A, Garrido JC,Garcia-Navarro S, Garcia-Navarro F 1998 Somatostatin plays a dual, stim-ulatory/inhibitory role in the control of growth hormone secretion by twosomatotrope subpopulations from porcine pituitary. J Neuroendocrinol9:841–848
162. Bruns C, Weekbecker G, Raulf F, Lubbert H, Hoyer D 1995 Characteriza-tion of somatostatin receptor subtypes. Ciba Found Symp 190:89–101
163. Holland LJ, Lamberts SW 1996 Somatostatin receptors and disease: role ofreceptor subtypes. Baillieres Clin Endocrinol Metab 10:163–176
164. Patel YC, Greenwood M, Panetta R, Hukovic N, Grigorakis S’LA, SrikantCB 1996 Molecular biology of somatostatin receptor subtypes. Metabolism45:31–38
165. Hoyer D, Bell GI, Epelbaum J, Feniuk W, Humphrey PPA, O’Carrol AM,Patel YC, Schonbrunn A, Taylor JE, Reisine T 1995 Classification andnomenclature of somatostatin receptors. Trends Pharmacol Sci 16:756–768
166. Hukovic N, Panetta R, Kumar U, Patel YC 1996 Agonist-dependent regu-lation of cloned human somatostatin receptor types 1–5 (hSSTR1–5): subtypeselective internalization or upregulation. Endocrinology 137:4046–4049
167. Pscherer A, Dorflinger U, Kirfel J, Gawlas K, Ruschoff J, Buettner R,Schule R 1996 The helix-loop-helix transcription factor SEF-2 regulates theactivity of a novel initiator element in the promoter of the human soma-tostatin receptor II gene. EMBO J 15:6680–6690
168. Lamberts SWJ, van der Lely AJ, De Herder WW, Hofland LJ 1996 Oct-reotide. N Engl J Med 25:246–249
169. Termanini B, Gibril F, Doppman JL, Reynolds JC, Stewart CA, Sutliff VE,Venzon DJ, Jensen RT 1997 Distinguishing small hepatic hemangiomasfrom vascular liver metastases in gastrinoma: use of a somatostatin-receptorscintigraphic agent. Radiology 202:151–158
170. Olsen P, Pozderac RV, Hinkle G, Hill T, O’Dorisio TM, Schirmer WJ,Ellison EC, O’Dorisio MS 1995 Somatostatin receptor imaging of neuroen-docrine tumors with indium-111 pentetreotide (Octreoscan). Semin NuclMed 25:251–261
171. Giustina A, Wehrenberg WB 1992 The role of glucocorticoids in the reg-ulation of growth hormone secretion. Trends Endocrinol Metab 3:306–311
172. Chan YY, Clifton DK, Steiner RA 1996 Role of NPY neurons in GH-dependent feedback signaling to the brain. Horm Res 45:12–14
173. Chan YY, Grafstein-Dunn E, Delemarre-van de Waal HA, Burton KA,Clifton DK, Steiner RA 1996 The role of galanin and its receptor in thefeedback regulation of growth hormone secretion. Endocrinology 137:5303–5310
174. Kamegai J, Minami S, Sugihara H, Hasegawa O, Higuchi H, WakabayashiI 1996 Growth hormone receptor gene is expressed in neuropeptide Yneurons in hypothalamic arcuate nucleus of rats. Endocrinology 137:2109–2112
175. Bennett PA, Levy A, Carmignac DF, Robinson IC, Lightman SL 1996Differential regulation of the growth hormone receptor gene: effects ofdexamethasone and estradiol. Endocrinology 137:3891–3896
176. Day R, Dong W, Panetta R, Kraicer J, Greendwood MT, Patel YC 1995Expression of mRNA for somatostatin receptor (SSTR) types 2 and 5 in
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 777
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

individual rat pituitary cells. A double labeling in situ hybridization analysis.Endocrinology 136:5232–5235
177. O’Carroll AM, Krempels K 1995 Widespread distribution of somatostatinreceptor messenger ribonucleic acids in rat pituitary. Endocrinology 136:5224–5227
178. Beaudet A, Tannenbaum GS 1995 Expression of SSTR1 and SSTR2 in rathypothalamus: correlation with receptor binding and distribution of growthhormone regulatory peptides. Ciba Found Symp 190:142–152
179. Bruno JF, Xu Y, Song J, Berelowitz M 1994 Pituitary and hypothalamicsomatostatin receptor subtype messenger ribonucleic acid expression in thefood-deprived and diabetic rat. Endocrinology 1354:1787–1792
180. Viollet C, Prevost G, Maubert E, Faivre-Bauman A, Gardette R, Kordon C,Loudes C, Slama A, Epelbaum J 1995 Molecular pharmacology of soma-tostatin receptors. Fundam Clin Pharmacol 9:107–113
181. Epelbaum J, Moyse E, Tannenbaum GS, Kordon C, Beaudet A 1989 Com-bined autoradiographic and immunohistochemical evidence for an associ-ation of somatostatin binding sites with growth hormone-releasing factor-containing nerve cell bodies in the rat arcuate nucleus. J Neuroendocrinol1:109–115
182. Bertherat J, Dournaud P, Berod A, Normand E, Bloch B, Rostene W,Kordon C, Epelbaum J 1992 Growth hormone-releasing hormone-synthe-sizing neurons are a subpopulation of somatostatin receptor-labelled cells inthe rat arcuate nucleus: a combined in situ hybridization and receptor light-microscopic radioautographic study. Neuroendocrinology 56:25–31
183. Epelbaum J 1992 Intrahypothalamic neurohormonal interactions in the con-trol of growth hormone secretion. Ciba Found Symp 168:54–64
184. Mounier F, Bluet-Pajot JT, Moinard F, Kordon C, Epelbaum J 1996 Acti-vation of locus coeruleus somatostatin receptors induces an increase ofgrowth hormone release in male rats. J Neuroendocrinol 8:761–764
185. Miller GM, Alexander JM, Bikkal HA, Katznelson L, Zervas NT, Kliban-ski A 1995 Somatostatin receptor subtype gene expression in pituitary ad-enomas. J Clin Endocrinol Metab 80:1386–1392
186. Greenspan Y, Melmed S 1994 Heterogeneous expression of two somatosta-tin receptor subtypes in pituitary tumors. J Clin Endocrinol Metab 78:398–403
187. Giustina A, Zaltieri G, Negrini F, Wehrenberg WB 1996 The pharmaco-logical aspects of the treatment of acromegaly. Pharmacol Res 34:1–22
188. Shimon I, Taylor JE, Dong JZ, Bitonte RA, Kim S, Morgan B, Coy DH,Culler MD, Melmed S 1997 Somatostatin receptor subtype specificity inhuman fetal pituitary cultures. Differential role of SSTR2 and SSTR5 forgrowth hormone, thyroid-stimulating hormone, and prolactin regulation.J Clin Invest 99:789–798
189. Shimon I, Yan X, Taylor JE, Weiss MH, Culler MD, Melmed S 1997Somatostatin receptor (SSTR) subtype-selective analogues differentiallysuppress in vitro growth hormone and prolactin in human pituitary ade-nomas. J Clin Invest 100:2386–2392
190. Yamada Y, Stoffel M, Espinosa III R, Xiang KS, Seino M, Seino S, Le BeauMM, Bell GI 1993 Human somatostatin receptor genes: localization tohuman chromosomes 14, 17, and 22 and identification of simple tandemrepeat polymorphisms. Genomics 15:449–452
191. Law SF, Woulfe D, Reisine T 1995 Somatostatin receptor activation ofcellular effector systems. Cell Signal 7:1–8
192. Kubota A, Yamada Y, Kagimoto S, Yasuda K, Someya Y, Ihara Y, OkamotoY, Kozasa T, Seino S, Seino Y 1994 Multiple effector coupling of soma-tostatin receptor subtype SSTR1. Biochem Biophys Res Commun 204:176–186
193. Osapay G, Prokai L, Kim HS, Medzihradszky KF, Coy DH, Liapakis G,Reisine T, Melacini G, Zhu Q, Wang SH, Mattern RH, Goodman M 1997Lanthionine-somatostatin analogs: synthesis, characterization, biological ac-tivity, and enzymatic stability studies. J Med Chem 40:2241–2251
194. Murabe H, Shimatsu A, Ihara C, Mizuta H, Nakamura Y, Nagata I, KikuchiH, Nakao K 1996 Expression of somatostatin receptor (SSTR) subtypes inpituitary adenomas: quantitative analysis of SSTR2 mRNA by reverse tran-scription-polymerase chain reaction. J Neuroendocrinol 8:605–610
195. Panetta R, Patel YC 1995 Expression of mRNA for all five human soma-tostatin receptors (hSSTR1–5) in pituitary tumors. Life Sci 56:333–342
196. Voltz DM, Giustina A, Gaillard RC, Wehrenberg WB 1993 The biologicalpotency of equimolar amounts of somatostatin-14 and somatostatin-28 inrats. Endocr J 1:235–239
197. Guillemin R, Brazeau P, Bohlen P, Esch F, Ling N, Wehrenberg WB 1982Growth hormone-releasing factor from a human pancreatic tumor thatcaused acromegaly. Science 218:585–587
198. Rivier J, Spiess J, Thorner MO, Vale W 1982 Characterization of a growthhormone-releasing factor from a human pancreatic islet tumor. Nature 300:276–278
199. Gubler U, Monahan JJ, Lomedico PT 1983 Cloning and sequence analysisof cDNA for the precursor of human growth hormone-releasing factor,somatocrine. Proc Natl Acad Sci USA 80:4311–4314
200. Mayo KE, Vale W, Rivier J 1983 Expression, cloning, and sequencing of acDNA encoding human growth hormone-releasing factor. Nature 306:86–88
201. Grossman A, Savage MO, Lytras N 1984 Responses to analogues of growth
hormone releasing hormone in normal subjects; and in growth hormonedeficient children and young adults. Clin Endocrinol (Oxf) 21:321–331
202. Frohman LA, Downs TR 1989 Human growth hormone-releasing hormonemetabolism in vivo and in vivo. In: Casanueva FF, Dieguez C (eds) RecentAdvances in Basic and Clinical Neuroendocrinology. Elsevier Science Pub-lisher B.V., Amsterdam, pp 217–224
203. Thorner MO, Spiess J, Vance ML, Rogol AD, Kaiser DL, Webster JD 1983Human pancreatic growth hormone releasing factor selectively stimulatesgrowth hormone secretion in man. Lancet 1:24–18
204. Friend KE, Hartman ML, Pezzoli SS, Clasey JL, Thorner MO 1996 Both oraland transdermal estrogen increase growth hormone release in postmeno-pausal women – a clinical research center study. J Clin Endocrinol Metab81:2250–2256
205. Gelato MC, Pescovitz OH, Cassorla F, Loriaux DL, Merriam GR 1984Dose-response relationships for the effects of growth hormone releasingfactor-(1–44)-NH2 in young adult men and women. J Clin Endocrinol Metab59:197–201
206. Giustina A, Bossoni S, Bodini C, Doga M, Grelli A, Buffoli MG, SchettinoM, Wehrenberg WB 1992 The role of cholinergic tone in modulating thegrowth hormone response to GH-releasing hormone in normal man. Me-tabolism 40:519–523
207. Mazza E, Ghigo E, Goffi S, Procopio M, Imperiale E, Arvat E, Bellene J,Boghen MF, Mueller EE, Camanni F 1989 Effect of the potentiation ofcholinergic activity on the variability in individual GH responses to GH-releasing hormone. J Endocrinol Invest 12:795–801
208. Giustina A, Bodini C, Bossoni S, Valentini U, Wehrenberg WB 1993Variability in the growth hormone responses to growth hormone-releasinghormone alone or combined with pyridostigmine in type 1 diabetic patients.J Endocrinol Invest 16:585–590
209. Devesa J, Lima L, Lois N, Fraga C, Lechuga MJ, Arce V, Tresguerres JAF1989 Reasons for the variability in growth hormone (GH) responses toGHRH challenge: the endogenous hypothalamic-somatotroph rhythm(HSR). Clin Endocrinol (Oxf) 30:367–377
210. Evans WS, Borges JL, Vance ML, Kaiser DL, Rogol AD, Thorner MO 1984Effects of human pancreatic growth hormone-releasing factor-40 on serumgrowth hormone, prolactin, luteinizing hormone, follicle-stimulating hor-mone, and somatomedin-C concentrations in normal women throughout themenstrual cycle. J Clin Endocrinol Metab 59:1006–1010
211. Sheldon Jr WR, De Bold CR, Evans WS, DeCherney GS, Jackson RV,Island DP, Thorner MO, Orth DN 1985 Rapid sequential intravenous ad-ministration of four hypothalamic releasing hormones as a combined an-terior pituitary function test in normal subjects. J Clin Endocrinol Metab60:623–630
212. Shibasaki T, Shizume K, Nakahara M, Masuda A, Jibiki K, Demura H,Wakabayashi I, Ling N 1984 Age-related changes in plasma growth hor-mone response to growth hormone-releasing factor in man. J Clin Endo-crinol Metab 58:212–214
213. Kaplan SL, Grumbach MM 1976 Development of hormonal secretion by thehuman fetal pituitary gland. In: Martini L, Ganong WF (eds) Frontiers inNeuroendocrinology. Academic Press, San Diego, CA, pp 255–276
214. Shimon I, Yan X, Melmed S 1998 Human fetal pituitary expresses functionalgrowth hormone-releasing peptide receptors. J Clin Endocrinol Metab 83:174–178
215. De Leo V, Lanzetta D, D’Antona D, Danero S 1993 Growth hormonesecretion in premenopausal women before and after ovariectomy: effect ofhormone replacement therapy. Fertil Steril 60:268–271
216. Bellantoni MF, Harman SM, Cho DE, Blackman MR 1991 Effects of pro-gestin-opposed transdermal estrogen administration on growth hormoneand insulin-like growth factor-I in postmenopausal women of different ages.J Clin Endocrinol Metab 72:172–178
217. Bilezikjian LM, Vale WV 1984 Chronic exposure of cultured rat anteriorpituitary cells to GRF causes partial loss of responsiveness to GRF. Endo-crinology 115:2032–2034
218. Ceda GP, Hoffman AR 1985 Growth hormone-releasing factor desensiti-zation in rat anterior pituitary cells in vitro. Endocrinology 116:1334–1340
219. Davis IRE, Sheppard MC, Shakespeare RA, Lynch SS, Clayton RN 1986Does growth hormone releasing factor desensitize the somatotroph? Inter-pretation of responses of growth hormone during and after 10-hour infusionof GRF 1–29 amide in man. Clin Endocrinol (Oxf) 24:135–140
220. Vance ML, Kaiser DI, Evans WS, Thorner MO 1985 Evidence for a limitedgrowth hormone (GH)-releasing hormone (GHRH)-releasable quantity ofGH: effects of 6-hour infusions on GH secretion in normal man. J ClinEndocrinol Metab 60:370–375
221. Jaffe CA, Turgeon DK, Friberg RD, Watkins PB, Barkan AL 1995 Nocturnalaugmentation of growth hormone (GH) secretion is preserved during re-petitive bolus administration of GH-releasing hormone: potential involve-ment of endogenous somatostatin. J Clin Endocrinol Metab 80:3321–3326
222. Thorner MO, Vance ML, Hartman ML, Holl RW, Evans WS, Veldhuis JD,Van Cauter E, Copinschi G, Bowers CY 1990 Physiological role of soma-tostatin on growth hormone regulation in humans. Metabolism 39:40–42
223. Leong DA, Pomes A, Gaylinn BD, Thorner MO, Hellmann PH, VeldhuisJD, A novel hypothalamic hormone measured in hypophyseal portal plasma
778 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

drives rapid bursts of GH secretion. Program of the 80th Annual Meetingof The Endocrine Society, New Orleans, LA, 1998 (Abstract OR9–3)
224. Losa M, Muller OA, Sobieszczyk S, Von Werder K 1985 Interaction be-tween growth hormone releasing factor (GRF) and somatostatin analogue(SMS 201–995) in normal subjects. Clin Endocrinol (Oxf) 23:715–720
225. Clayton RN, Biley LC 1987 Somatostatin partially reverses desensitizationof somatotrophs induced by growth hormone-releasing factor. J Endocrinol112:69–76
226. Bilezikjian LM, Sheifert H, Vale WW 1986 Desensitization to growth hor-mone-releasing hormone factor (GRF) is associated with downregulation ofGRF-binding sites. Endocrinology 118:2045–2052
227. Vance ML, Kaiser DL, Martha PM, Furlanetto R, Rivier J, Vale W, ThornerMO 1989 Lack of in vivo somatotroph densensitization or depletion after 14days of continuous growth hormone (GH)-releasing hormone administra-tion in normal men and a GH-deficient boy. J Clin Endocrinol Metab 68:22–28
228. Iranmanesh A, Liem AY, South S, Clemmons D, Weltman A, Thorner MO,Veldhuis JD, A 3-day exogenous pulsatile GHRH clamp unmasks stronglynegative determination of GH secretory burst mass by age and percentagebody fat and positive control by testosterone in healthy men. Presented atthe Clinical Research Meeting, San Diego, CA, 1995
229. Thorner MO, Perryman RL, Cronin MJ, Rogol AD, Draznin M, JohansonA, Vale W, Horvath E, Kovacs K 1982 Somatotroph hyperplasia: successfultreatment of acromegaly by removal of a pancreatic islet tumor secreting agrowth hormone-releasing factor. J Clin Invest 70:965–977
230. Bloch B, Brazeau P, Ling N, Bohlen P, Esch F, Wehrenberg WB 1983Immunohistochemical detection of growth hormone-releasing factor inbrain. Nature 301:607–608
231. Merchenthaler I, Vigh S, Schally AV, Petrusz P 1984 Immunocytochemicallocalization of growth hormone-releasing factor in the rat hypothalamus.Endocrinology 114:1082–1085
232. Niimi M, Takhara J, Sato M, Kawanishi K 1990 Immunohistochemicalidentification of galanin and growth hormone-releasing factor containingneurons projecting to the median eminence in the rat. Neuroendocrinology51:572–575
233. Horvath S, Palkovits M, Gorcs T, Arimura A 1989 Electron microscopicimmunocytochemical evidence for the existence of bidirectional synapticconnections between growth hormone- and somatostatin-containing neu-rons in the hypothalamus of the rat. Brain Res 481:8–15
234. Frohman LA, Jansson J-O 1986 Growth hormone-releasing hormone. En-docr Rev 7:223–253
235. Saito H, Saito S, Ryuchi Y 1984 Clinical value of radioimmunoassay ofplasma growth hormone releasing factor. Lancet 2:401–402
236. Delvalle J, Yamada T 1990 The gut as an endocrine organ. Annu Rev Med41:447–455
237. Baird A, Wehrenberg WB, Bolhen P, Ling N 1985 Immunoreactive andbiologically active growth hormone releasing factor in the rat placenta.Endocrinology 117:917–921
238. Barinaga M, Bilezikjian LM, Vale WW, Rosenfeld MG, Evans RM 1985Independent effects of growth hormone releasing factor on growth hormonerelease and gene transcription. Nature 314:279–281
239. Seifert H, Perrin M, Rivier J, Vale W 1985 Growth hormone-releasing factorbinding sites in rat anterior pituitary membrane homogenates: modulationby glucocorticods. Endocrinology 117:424–426
240. Gaylinn D, Harrison JK, Zysk JR, Lyons Jr CE, Lynch KR, Thorner MO1993 Molecular cloning and expression of a human anterior pituitary re-ceptor for growth hormone-releasing hormone. Mol Endocrinol 7:77–84
241. Edwards CA, Dieguez C, Ham J, Peters JR, Scanlon MF 1998 Evidence thatgrowth hormone depletion and uncoupling of the regulatory protein ofadenylate cyclase (Ns) both contribute to the desensitization of growthhormone responses to growth hormone-releasing factor. J Endocrinol 116:185–190
242. Natori S, Ikuyama S, Haji M, Ohashi M, Nawata H 1989 Growth hormonereleasing hormone-sensitive adenylate cyclase activity in growth hormone-producing pituitary adenoma: correlation to the response of plasma growthhormone to growth hormone releasing hormone in patients with acromeg-aly. Endocrinol Jpn 36:229–236
243. Mayo KE, Miller TL, DeAlmeida V, Zheng J, Godfrey PA 1996 The growth-hormone-releasing hormone receptor: signal transduction, gene expression,and physiological function in growth regulation. Ann NY Acad Sci 805:184–203
244. Mayo KE, Godfrey PA, Suhr ST, Kulik DJ, Rahal JO 1995 Growth hor-mone-releasing hormone: synthesis and signaling. Recent Prog Horm Res50:35–73
245. Hsiung HM, Smith DP, Zhang XY, Bennett T, Rosteck Jr PR, Lai MH 1993Structure and functional expression of a complementary DNA for porcinegrowth hormone-releasing hormone receptor. Neuropeptides 25:1–10
246. Mayo KE 1992 Molecular cloning and expression of a pituitary-specificreceptor for growth hormone releasing hormone. Mol Endocrinol 6:1734–1744
247. Korytko A, Zeitler P, Cutler L 1996 Developmental regulation of pituitaryGRF-receptor gene expression in the rat. Endocrinology 137:1326–1331
248. Bagnato A, Moretti C, Ohnishi J, Frajese G, Catt KJ 1992 Expression in thegrowth hormone-releasing hormone gene and its peptide product in the ratovary. Endocrinology 130:1097–1102
249. Fujinaka Y, Yokogoshi Y, Zhang CY, Okura T, Kitagawa K, Saito S 1996Tissue-specific molecular heterogeneity of human growth hormone-releas-ing hormone receptor protein. FEBS Lett 394:1–4
250. Godfrey P, Rahal JO, Beamer WG, Copeland NG, Jenkins NA, Mayo KE1993 GHRH receptor of little mice contains a missense mutation in theextracellular domain that disrupts receptor function. Nat Genet 4:227–232
251. Jaffe CA, Friberg RD, Barkan AL 1993 Suppression of growth hormone(GH)secretion by a selective GH-releasing hormone (GHRH) antagonist. Directevidence for involvement of endogenous GHRH in the generation of GHpulses. J Clin Invest 92:695–701
252. Kovacs M, Zarandi M, Halmos G, Groot K, Schally AV 1996 Effects of acuteand chronic administration of a new potent antagonist of growth hormone-releasing hormone in rats: mechanisms of action. Endocrinology 137:5364–5369
253. Hashimoto K, Koga M, Motomura T, Kasayama S, Kouhara H, Ohnishi T,Arita N, Hayakawa T, Sato B, Kishimoto T 1995 Identification of alterna-tively spliced messenger ribonucleic acid encoding truncated growth hor-mone-releasing hormone receptor in human pituitary adenomas. J ClinEndocrinol Metab 80:2933–2939
254. Giustina A, Bussi A, Wehrenberg WB 1994 Dichotomic role of glucocor-ticoids in the regulation of growth hormone secretion in vivo. In: Bercu B,Walker R (eds) Growth Hormone. II. Basic and Clinical Aspects. Springer-Verlag, New York, pp 347–358
255. Lam KS, Lee MF, Tam SP, Srivastava G 1996 Gene expression of thereceptor for growth-hormone-releasing hormone is physiologically regu-lated by glucocorticoids and estrogen. Neuroendocrinology 63:475–480
256. Takahashi T, Okimura Y, Yoshimura K, Shigeyoshi Y, Kaji H, Abe H,Chihara K 1995 Regional distribution of growth hormone-releasing hor-mone (GHRH) receptor mRNA in the rat brain. Endocrinology 136:4721–4724
257. Nakamoto JM, Gertner JM, Press CM, Hintz RL, Rosenfeld RG, Genel M1986 Suppression of the growth hormone (GH) response to clonidine andGH-releasing hormone by exogenous GH. J Clin Endocrinol Metab 62:822–826
258. Rosenthal SM, Hulse JA, Kaplan SL, Grumbach MM 1986 Exogenousgrowth hormone inhibits growth hormone-releasing factor-induced growthhormone secretion in normal men. J Clin Invest 77:176–180
259. Ross RJ, Borges F, Grossman A 1987 Growth hormone pretreatment in manblocks the response to growth hormone-releasing hormone: evidence for adirect effect of growth hormone. Clin Endocrinol (Oxf) 26:117–123
260. Giustina A, Bossoni S, Bodini C, Cimino A, Pizzocolo G, Schettino M,Wehrenberg WB 1991 Efffects of exogenous growth hormone pretreatmenton the pituitary growth hormone response to growth hormone-releasinghormone alone or in combination with pyridostigmine in type 1 diabeticpatients. Acta Endocrinol (Copenh) 125:510–517
261. Friend K, Iranmanesh A, Login IS, Veldhuis JD 1997 Pyridostigmine treat-ment selectively amplifies the mass of GH secreted per burst without alteringthe GH burst frequency, half-life, basal GH secretion or the orderliness ofthe GH release process. Eur J Endocrinol 137:377–386
262. Wehrenberg WB, Wiviott SD, Voltz DM, Giustina A 1992 Pyridostigminemediated growth hormone release: evidence for somatostatin involvement.Endocrinology 130:1445–1457
263. Bennett PA, Levy A, Sophokleous S, Robinson IC, Lightman SL 1995Hypothalamic GH receptor gene expression in the rat: effects of altered GHstatus. J Endocrinol 147:225–234
264. Aguila MC, McCann SM 1985 Stimulation of somatostatin release in vitroby synthetic human growth hormone-releasing factor by a nondopaminergicmechanism. Endocrinology 117:762–765
265. Tannenbaum GS 1994 Multiple levels of cross-talk between somatostatin(SRIF) and growth hormone (GH)-releasing factor in genesis of pulsatile GHsecretion. Clin Pediatr Endocrinol 3:97–110
266. Murakami Y, Kato Y, Kabayama Y, Inoue T, Koshiyama H, Imura H 1987Involvement of hypothalamic growth hormone (GH)-releasing factor in GHsecretion induced by intracerebroventricular injection of somatostatin inrats. Endocrinology 120:311–316
267. Liposits ZS, Merchenthaler I, Paull WK, Flerko S 1988 Synaptic commu-nication between somatostatinergic axons and growth hormone-releasingfactor (GRF) synthesizing neurons in the arcuate nucleus of the rat. Histo-chemistry 89:247–252
268. Ocampo-Lim B, Guo W, DeMott-Friberg R, Barkan AL, Jaffe CA 1996Nocturnal growth hormone (GH) secretion is eliminated by infusion ofGH-releasing hormone antagonist. J Clin Endocrinol Metab 81:4396–4399
269. Jaffe CA, DeMott FR, Barkan AL 1996 Involvement of endogenous growthhormone (GH)-releasing hormone in the GH responses to physiological andpharmacological stimuli. J Clin Invest 97:934–940
270. Berelowitz M, Szabo M, Frohman LA, Firestone S, Chu L 1981 Somato-medin-C mediates growth hormone negative feedback by effects on both thehypothalamus and the pituitary. Science 212:1279–1281
271. Bermann M, Jaffe CA, Tsai W, DeMott-Friberg R, Barkan AL 1994 Negative
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 779
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

feedback regulation of pulsatile growth hormone secretion by insulin-likegrowth factor I: involvement of hypothalamic somatostatin. J Clin Invest94:138–145
272. Minami S, Suzuki N, Sugihara H, Tamura H, Emoto N, Wakabayashi I1997 Microinjection of rat GH but not human IGF-I into a defined area of thehypothalamus inhibits endogenous GH secretion in rats. J Endocrinol 153:283–290
273. Sato M, Frohman LA 1993 Differential effects of central and peripheraladministration of growth hormone (GH) and insulin-like growth factor onhypothalamic GH-releasing hormone and somatostatin gene expression inGH-deficient dwarf rats. Endocrinology 133:793–799
274. Aguila MC, Boggaram V, McCann SM 1993 Insulin-like growth factor-Imodulates hypothalamic somatostatin through a growth hormone releasingfactor increased somatostatin release and messenger ribonucleic acid levels.Brain Res 625:213–218
275. Harel Z, Tannenbaum GS 1992 Synergistic interaction between insulin-likegrowth factors-I and -II in central regulation of pulsatile growth hormonesecretion. Endocrinology 131:758–764
276. Tannenbaum GS, Guyda HJ, Posher BI 1983 Insulin-like growth factors: arole in growth hormone negative feedback and body weight regulation viabrain. Science 220:77–79
277. Harel Z, Tannenbaum GS 1992 Centrally administered insulin-like growthfactor II fails to alter pulsatile growth hormone secretion or food intake.Neuroendocrinology 56:161–168
278. Korbonits M, Little JA, Camacho-Hubner C, Trainer PJ, Besser GM, Gross-man AB 1996 Insulin-like growth factor-I and -II in combination inhibit therelease of growth hormone-releasing hormone from the rat hypothalamusin vitro. Growth Regul 6:110–120
279. Bennett PA, Thomas GB, Howard AD, Feighner SD, van den Ploeg LH,Smith RG, Robinson IC 1997 Hypothalamic growth hormone secretagogue-receptor (GHS-R) expression is regulated by growth hormone in the rat.Endocrinology 138:4552–4557
280. Bowers CY, Momany FA, Reynolds A, Hong A 1984 On the in vitro and invivo activity of a new synthetic hexapeptide that acts on the pituitary tospecifically release growth hormone. Endocrinology 114:1537–1545
281. Bowers CY, Momany FA, Chang D, Hong A, Chang K 1980 Structure-activity relationships of a synthetic pentapeptide that specifically releasesGH in vitro. Endocrinology 106:663–667
282. Momany F, Bowers CY, Reynolds GA, Chang D, Hong A, Newland K 1985Conformational energy studies and in vitro and in vivo activity data ongrowth hormone-releasing peptides. Endocrinology 114:1531–1536
283. Patchett AA, Nargund RP, Tata JR, Chen MH, Barakat KJ, Johnston DB,Cheng K, Chan WW, Butler B, Hickey G, Jacks T, Schleim K, Pong S-S,Chaunt L-YP, Chen HY, Frazier E, Leung KH, Chiu S-HL, Smith RG 1995Design and biological activities of L-163,191 (MK-0677): a potent, orallyactive growth hormone secretagogue. Proc Natl Acad Sci USA 92:7001–7005
284. Cheng K, Chan WWS, Butler B 1993 Stimulation of growth hormone releasefrom rat primary pituitary cells by L-692,429, a novel non-peptidyl GHsecretagogue. Endocrinology 132:2729–2731
285. Schoen WR, Pisano JM, Prendergast K, Wyvratt Jr MJ, Fisher MH, ChengK, Chan WW, Butler B, Smith RG, Ball RG 1994 A novel 3-substitutedbenzazepinone growth hormone secretagogue (L-692, 429). J Med Chem37:897–906
286. Smith RG, Cheng K, Schoen WR, Pong S-S, Hickey G, Jacks T, Butler B,Chan WWS, Chaung L-YP, Taylor J, Wyvratt MJ, Fisher MH 1993 Anon-peptidyl growth hormone secretagogue. Science 260:1640–1643
287. McDowell RS, Elias KA, Stanley MS, Burdick DJ, Burnier JP, Chan KS,Fairbrother WJ, Hammonds RG, Ingle GS, Jacobsen NE 1995 Growthhormone secretagogues: characterization, efficacy, and minimal bioactiveconformation. Proc Natl Acad Sci USA 92:11165–11169
288. Hartman ML, Farello G, Pezzoli SS, Thorner MO 1992 Oral administrationof growth hormone (GH)-releasing peptide stimulates GH secretion in nor-mal men. J Clin Endocrinol Metab 74:1378–1384
289. Korbonits M, Grossman AB 1995 Growth hormone-releasing peptide andits analogues. Novel stimuli to growth hormone release. Trends EndocrinolMetab 6:43–49
290. Chapman IM, Bach MA, Cauter EV, Farmer M, Krupa D, Taylor AM,Schilling LM, Cole KY, Skiles EH, Pezzoli SS, Hartman ML, Veldhuis JD,Gormley GJ, Thorner MO 1996 Stimulation of the growth hormone (GH)-insulin-like growth factor I axis by daily oral administration of a GH secre-tagogue (MK-0677) in healthy elderly subjects. J Clin Endocrinol Metab81:4249–4257
291. Ghigo E, Arvat E, Muccioli G, Camanni F 1997 Growth hormone releasingpeptides. Eur J Endocrinol 136:445–460
292. Ghigo E, Arvat E, Gianotti L, Grottoli S, Rizzi G, Ceda GP, Boghen MF,Deghenghi R, Camanni F 1996 Short-term administration of intranasal ororal hexarelin, a synthetic hexapeptide, does not desensitize the growthhormone responsiveness in human aging. Eur J Endocrinol 135:407–412
293. Hickey GJ, Jacks TM, Schleim KD, Frazier E, Chen HY, Krupa D, FeeneyW, Nargund RP, Patchett AA, Smith RG 1997 Repeat administration of theGH secretagogue MK-0677 increases and maintains elevated IGF-I levels inbeagles. J Endocrinol 152:183–192
294. Sartor AO, Bowers CY, Chang D 1985 Parallel studies of His-DTrp-Ala-Trp-DPhe-Lys-NH2 and hpGRF-44NH2 in rat pituitary cell monolayer cul-ture. Endocrinology 117:97–105
295. Merriam GR, Buchner DM, Prinz PN, Schwartz RS, Vitiello V 1997 Po-tential applications of GH secretagogues in the evaluation and treatment ofthe age-related decline in growth hormone secretion. Endocrine 7:49–52
296. Svensson J, Lonn L, Jansson J-O, Murphy G, Wyss D, Krupa D, CerchioK, Polvino W, Gertz B, Boseaus I, Sjostrom L, Bengtsson B-A 1998 Two-month treatment of obese subjects with the oral growth hormone (GH)secretagogue MK-677 increases GH secretion, fat-free mass, and energyexpenditure. J Clin Endocrinol Metab 83:362–369
297. Murphy MG, Plunkett LM, Gertz BJ, He W, Wittreich J, Polvino WM,Clemmons DR 1998 MK-677, an orally active growth hormone secreta-gogue, reverses diet-induced catabolism. J Clin Endocrinol Metab 83:320–325
298. Bowers CY, Reynolds GA, Durham D, Barrera CM, Pezzoli SS, ThornerMO 1990 Growth hormone (GH)-releasing peptide stimulates GH release innormal men and acts synergistically with GH-releasing hormone. J ClinEndocrinol Metab 70:975–982
299. Fletcher TP, Thomas GB, Clarke IJ 1996 Growth hormone-releasing andsomatostatin concentrations in the hypophysial portal blood of conscioussheep during the infusion of growth hormone-releasing peptide-6. DomestAnim Endocrinol 13:251–258
300. Guillaume V, Magnan E, Cataldi M, Dutour A, Sauze N, Renard M,Razafindraibe H, Conte-Devolx B, Deghenghi R, Lenaerts V 1994 Growthhormone (GH)-releasing hormone secretion is stimulated by a new GH-releasing hexapeptide in sheep. Endocrinology 135:1073–1076
301. Cheng J, Wu TJ, Butler B, Cheng K 1997 Growth hormone releasing pep-tides: a comparison of the growth hormone releasing activities of GHRP-2and GHRP-6 in rat primary pituitary cells. Life Sci 608:1385–1392
302. Bowers CY, Sartor AO, Reynolds GA, Badger TM 1991 On the actions ofthe growth hormone-releasing hexapeptide. Endocrinology 128:2027–2035
303. Bowers CY, Granda-Ayala R 1996 GHRP-2, GHRH and SRIF interrelation-ships during chronic administration of GHRP-2 to humans. J Pediatr En-docrinol Metab 9:261–270
304. Argente J, Pozo J, Chowen JA 1996 The growth hormone axis: control andeffects. Horm Res 45:9–11
305. Mallo F, Alvarez CV, Benitez L 1993 Regulation of His-D-Trp-Ala-Trp-D-Phe-Lys-NH2 (GHRP-6)-induced GH secretion in the rat. Neuroendocrinol-ogy 57:247–256
306. Shah N, Anderson SM, Azimi P, Evans WS, Bowers CY, Veldhuis JD,Estradiol augments basal GH pulse mass and amplifies GH-secretory re-sponsiveness to low doses of GHRP-2. Program of the Fifth InternationalPituitary Congress, Naples, FL, 1998, p 60
307. Arvat E, Ramunni J, Bellone J, Di Vito L, Baffoni C, Broglio F, DeghenghiR, Bartolotta E, Ghigo E 1997 The GH, prolactin, ACTH and cortisol re-sponses to hexarelin, a synthetic hexapeptide, undergo different age-relatedvariations. Eur J Endocrinol 1237:635–642
308. de Zegher F, Spitz B, Van den Berghe G, Lemmens D, Vanweser K,Keppens K, Bowers CY 1998 Postpartum hyperprolactinemia and hypore-sponsiveness of growth hormone (GH) to GH-releasing peptide. J ClinEndocrinol Metab 83:103–106
309. Popovic V, Damjanovic S, Micic D, Djurovic M, Dieguez C, Casanueva FF1995 Blocked growth hormone-releasing peptide (GHRP-6)-induced GHsecretion and absence of the synergic action of GHRP-6 plus GH-releasinghormone in patients with hypothalamopituitary disconnection: evidencethat GHRP-6 main action is exerted at the hypothalamic level. J Clin En-docrinol Metab 80:942–947
310. Pombo M, Barreiro J, Penalva A, Peino R, Dieguez C, Casanueva FF 1995Absence of growth hormone (GH) secretion after the administration of eitherGH-releasing hormone (GHRH), GH-releasing peptide (GHRP-6), or GHRHplus GHRP-6 in children with neonatal pituitary stalk transection. J ClinEndocrinol Metab 80:3180–3184
311. Penalva A, Pombo M, Carballo A, Barreiro J, Casanueva FF, Dieguez C1993 Influence of sex, age and adrenergic pathways on the growth hormoneresponse to GHRP-6. Clin Endocrinol (Oxf) 38:87–91
312. Cordido F, Penalva A, Dieguez C, Casanueva FF 1993 Massive growth (GH)discharge in obese subjects after the combined administration of GH-re-leasing hormone and GHRP-6: evidence for a marked somatotroph secretorycapability in obesity. J Clin Endocrinol Metab 76:819–823
313. Tiulpakov AN, Brook CG, Pringle PJ, Peterkova VA, Volevodz NN, Bow-ers CY 1995 GH responses to intravenous bolus infusions of GH releasinghormone and GH releasing peptide 2 separately and in combination in adultvolunteers. Clin Endocrinol (Oxf) 43:347–350
314. Micic D, Popovic V, Kendereski A, Macut D, Casanueva FF, Dieguez C1995 Growth hormone secretion after the administration of GHRP-6 orGHRH combined with GHRP-6 does not decline in late adulthood. ClinEndocrinol (Oxf) 42:191–194
315. Saenger P 1996 Editorial: Oral growth hormone secretagogues – better thanAlice in Wonderland’s growth elixir? J Clin Endocrinol Metab 81:2773–2775
316. Hartman ML, Farello G, Pezzoli SS, Thorner MO 1992 Oral administration
780 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

of growth hormone (GH)-releasing peptide stimulates GH secretion in nor-mal men. J Clin Endocrinol Metab 74:1378–1384
317. Bowers CY 1994 Editorial: On a peptidomimetic growth hormone-releasingpeptide. J Clin Endocrinol Metab 79:940–942
318. Laron Z, Frenkel J, Deghenghi R, Anin S, Klinger B, Silbergeld A 1995Intranasal administration of the GHRP hexarelin accelerates growth in shortchildren. Clin Endocrinol (Oxf) 43:631–635
319. Van den Berghe G, de Zegher F, Baxter RC, Veldhuis JD, Wouters P, SchetzM, Verwaest C, Van Der Vorst E, Lauwers P, Bouillon R, Bowers CY 1998Neuroendocrinology of prolonged critical illness: effects of exogenous thy-rotropin-releasing hormone infusion and its combination with growth hor-mone-secretagogues. J Clin Endocrinol Metab 83:309–319
320. Van den Berghe G, de Zegher F, Veldhuis JD, Wouters P, Verbruggen W,Awouters M, Schetz M, Verwaest C, Lauwers P, Bouillon R, Bowers CY1996 The somatotropic axis in critical illness: effects of continuous GHRHand GHRP-2 infusion. J Clin Endocrinol Metab 82:590–599
321. Van den Berghe G, de Zegher F, Veldhuis JD, Wouters P, Gouwy S,Stefaan G, Stockman W, Weekers F, Schetz M, Lauwers P, Bouillon R,Bowers CY 1998 Thyrotropin and prolactin release in prolonged criticalillness: dynamics of spontaneous secretion and effects of growth hormone-secretagogues. Clin Endocrinol 4:599–612
322. Aloi JA, Gertz BJ, Hartman ML, Huhn WC, Pezzoli SS, Wittreich JM,Krupa DA, Thorner MO 1994 Neuroendocrine responses to a novel growthhormone secretagogue, L-692,429, in healthy older subjects. J Clin Endocri-nol Metab 79:943–949
323. Huhn WC, Hartman ML, Pezzoli SS, Thorner MO 1993 Twenty-four-hourgrowth hormone (GH)-releasing peptide (GHRP) infusion enhances pulsa-tile GH secretion and specifically attenuates the response to a subsequentGHRP bolus. J Clin Endocrinol Metab 76:1202–1208
324. Jacks T, Hickey G, Judith F, Taylor J, chen H, Krupa D, Feeney W, SchoenW, Ok D, Fisher M, Wyvratt M, Smith R 1994 Effects of acute and repeatedintravenous administration of L-692,585, a novel non-peptidyl growth hor-mone secretagogue, on plasma growth hormone, IGF-I, ACTH, cortisol,prolactin, insulin, and thyroxine levels in beagles. J Endocrinol 143:399–406
325. Jaffe CA, DeMott Friberg R, Barkan AL 1993 Effects of prolonged GHRPinfusion on pulsatile GH secretion in normal men. J Clin Endocrinol Metab77:1641–1647
326. Fairhall KM, Mynett A, Smith RG, Robinson ICAF 1995 Consistent GHresponses to repeated injections of GH-releasing hexapeptide (GHRP-6) andthe non-peptide GH secretagogue, L-692,585. Endocrinology 145:417–426
327. Ghigo E, Arvat E, Rizzi G, Goffi S, Grottoli S, Mucci M, Boghen MF,Camanni F 1994 Growth hormone-releasing activity of growth hormone-releasing peptide-6 is maintained after short-term oral pretreatment with thehexapeptide in normal aging. Eur J Endocrinol 131:499–503
328. Hayashi S, Okimura Y, Yagi H, Uchiyama T, Takeshima Y, Shakuoui S,Oohashi S, Bowers CY, Chihara K 1991 Intranasal administration of His-dTrp-Ala-Trp-d-Phe-Lys-NH2 (growth hormone releasing peptide) increasedplasma growth hormone and insulin-like growth factor-I levels in normalmen. Endocrinol Jpn 38:15–21
329. Rahum A, O’Neill PA, Shalt SM 1998 Growth hormone status duringlong-term hexarelin therapy. J Clin Endocrinol Metab 83:1644–1649
330. Conley LK, Gaillard RC, Giustina A, Brogan RS, Wehrenberg WB 1998Effects of repeated doses and continuous infusions of the growth hormone-releasing peptide hexarelin in conscious male rats. J Endocrinol 158:367–375
331. Clark RG, Thomas GB, Mortensen DL, Won Wb Ma YH, Tomlinson EE,Fairhall KM, Robinson ICAF 1997 Growth hormone secretagogues stim-ulate the hypothalamic-pituitary-adrenal axis and are diabetogenic in theZucker diabetic fatty rat. Endocrinology 138:4316–4323
332. Massoud AF, Hindmarsh PC, Brook CS 1996 Hexarelin-induced growthhormone, cortisol, and prolactin release: a dose-response study. J Clin En-docrinol Metab 81:4338–4341
333. Thomas GB, Fairhall KM, Robinson IC 1997 Activation of the hypo-thalamo-pituitary-adrenal axis by the growth hormone (GH) secretagogue,GH-releasing peptide-6 in rats. Endocrinology 138:1585–1591
334. Leal-Cerro A, Pumar A, Garcia-Garcia E, Dieguez C, Casanueva FF 1994Inhibition of growth hormone release after the combined administration ofGHRH and GHRP-6 in patients with Cushing’s syndrome. Clin Endocrinol(Oxf) 41:649–654
335. Ghigo E, Arvat E, Ramunni J, Colao A, Gianotti R, Deghenghi G, Lom-bardi G, Camanni F 1997 Adrenocorticotropin- and cortisol-releasing effectof hexarelin, a synthetic growth hormone-releasing peptide, in normal sub-jects and patients with Cushing’s syndrome. J Clin Endocrinol Metab 82:2439–2444
336. Friebos R-M, Murck H, Maier P, Schier T, Holsboer F, Steiger A 1995Growth hormone-releasing peptide-6 stimulates sleep, growth hormone,ACTH and cortisol release in normal men. Neuroendocrinology 61:584–589
337. Steiger A, Guldner J, Hemmeter U, Rothe B, Wiedemann K, Holsboer F1992 Effects of growth hormone-releasing hormone and somatostatin onsleep EEG and nocturnal hormone secretion in male controls. Neuroendo-crinology 56:566–573
338. Marshall L, Molle M, Boschen G, Steiger A, Fehm HL, Born J 1996 Greaterefficacy of episodic than continuous growth hormone-releasing hormone
(GHRH) administration in promoting slow-wave sleep (SWS). J Clin En-docrinol Metab 81:1009–1013
339. Arvat E, Maccagno B, Ramunni J, Di Vito L, Broglio F, Deghenghi R,Camanni F, Ghigo E 1997 Hexarelin, a synthetic growth-hormone releasingpeptide, shows no interaction with corticotropin-releasing hormone andvasopressin on adrenocorticotropin and cortisol secretion in humans. Neu-roendocrinology 66:432–438
340. Smith RG, Pong SS, Hickey G, Jacks T, Cheng K, Leonard R, Cohen CJ,Arena JP, Chang CH, Drisko J, Wyvratt M, Fisher M, Nargund R, PatchettA 1996 Modulation of pulsatile GH release through a novel receptor inhypothalamus and pituitary gland. Recent Prog Horm Res 51:261–285
341. Elias KA, Ingle GS, Burnier JP, Hammonds RG, McDowell RS, RawsonTE, Somers TC, Stanley MS, Cronin MJ 1995 In vitro characterization of fournovel classes of growth hormone-releasing peptide. Endocrinology 136:5694–5699
342. Howard AD, Feighner SD, Cully DF, Arena JP, Liberator PA, RosenblumCI, Hamelin M, Hreniuk DL, Palyha OC, Anderson J, Paress PS, Diaz C,Chou M, Liu KK, McKee KK, Pong SS, Chaung LY, Elbrecht A, Dash-kevicz M, Heavens R, Rigby M, Sirinathsinghji DJS, Dean DC, MelilloDG, van der Ploeg LH 1996 A receptor in pituitary and hypothalamus thatfunctions in growth hormone release. Science 273:974–977
343. Pong SS, Chaung LY, Dean DC, Nargund RP, Patchett AA, Smith RG 1996Identification of a new G-protein-linked receptor for growth hormone secre-tagogues. Mol Endocrinol 10:57–61
344. Sethumadhavan K, Veeraragavan K, Bowers CY 1991 Demonstration andcharacterization of the specific binding of growth hormone-releasing pep-tide to rat anterior pituitary and hypothalamic membranes. Biochem Bio-phys Res Commun 178:31–37
345. Adams EF, Huang B, Buchfelder M, Howard A, Smith RG, Feighner SD,Van Der Ploeg LHT, Bowers CY, Fahlbusch R 1998 Presence of growthhormone secretagogue receptor messenger ribonucleic acid in human pi-tuitary tumors and rat GH3 cells. J Clin Endocrinol Metab 83:638–642
346. Yokote R, Sato M, Matsubara S, Ohye H, Miimi M, Murao K, TakaharaJ 1998 Molecular cloning and gene expression of growth hormone-releasingpeptide receptor in rat tissues. Peptides 19:15–20
347. Codd EE, Shu AYL, Walker RF 1989 Binding of a growth hormone releasinghexapeptide to specific hypothalamic and pituitary binding sites. Neuro-pharmacology 28:1139–1144
348. Robinson BM, Friberg RD, Bowers CY, Barkan AL 1992 Acute growthhormone (GH) response to GH-releasing hexapeptide in humans is inde-pendent of endogenous GH-releasing hormone. J Clin Endocrinol Metab75:1121–1124
349. Giustina A, Bonfanti C, Licini M, Ragni G, Stefana B 1997 Hexarelin, anovel GHRP-6 analog, stimulates growth hormone (GH) release in a GH-secreting rat cell line (GH1) insensitive to GH-releasing hormone. Regul Pept70:49–54
350. Locatelli V, Grilli R, Torsello A, Cella SG, Wehrenberg WB, Mueller EE1994 Growth hormone-releasing hexapeptide is a potent stimulator ofgrowth hormone gene expression and release in the growth hormone-re-leasing hormone-deprived infant rat. Pediatr Res 36:169–174
351. Sota JL, Castrillo JL, Dominguez F, Dieguez C 1995 Regulation of thepituitary-specific transcription factor GHF-1/Pit-1 messenger ribonucleicacid level by growth hormone-secretagogues in rat anterior pituitary cellsin monolayer culture. Endocrinology 136:3863–3870
352. Blake AD, Smith RG 1991 Desensitization studies using perifused rat pi-tuitary cells show that growth hormone-releasing hormone and His-D-Trp-Ala-Trp-D-Phe-Lys-NH2 stimulate growth hormone release through dis-tinct receptor sites. J Endocrinol 129:11–19
353. Solian EB, Hashizume T, Kanemasui S 1994 Effect of growth hormone(GH)-releasing peptide (GHRP) on the release of GH from cultured anteriorpituitary cells in cattle. Endocr J 41:585–591
354. Pomes A, Pong SS, Schaeffer JM 1996 Solubilization and characterizationof a growth hormone secretagogue receptor from porcine anterior pituitarymembranes. Biochem Biophys Res Commun 225:939–945
355. Ong H, McNicoll N, Escher E, Collu R, Deghenghi R, Locatelli V, GhigoE, Muccioli G, Boghen M, Nilsson M 1998 Identification of a pituitarygrowth hormone-releasing peptide (GHRP) receptor subtype by photoaf-finity labeling. Endocrinology 139:432–435
356. McKee KK, Palyha OC, Feighner SD, Hreniuk DL, Tan CP, Phillips MS,Smith RG, van der Ploeg LH, Howard AD 1997 Molecular analysis of ratpituitary and hypothalamic growth hormone secretagogue receptors. MolEndocrinol 11:415–423
357. Cheng K, Chan WWS, Butler P, Barreto A, Smith RG 1991 Evidence for arole of protein kinase-C in His-D-Trp-Ala-Trp-D-Phe-Lys-NH2-inducedgrowth hormone release from rat pituitary cells. Endocrinology 129:3337–3342
358. Lei T, Buchfelder M, Fahlbusch R, Adams EF 1995 Growth hormone re-leasing peptide (GHRP-6) stimulates phosphatidylinositol (PI) turnover inhuman pituitary somatotroph cells. J Mol Endocrinol 14:135–138
359. Mau SE, Witt MR, Bjerrum OJ, Saermark T, Vilhardt H 1995 Growthhormone releasing hexapeptide (GHRP-6) activates the inositol (1,4,5)-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 781
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

trisphosphate/diacylglycerol pathway in rat anterior pituitary cells. J ReceptSignal Transduct Res 15:311–323
360. Herrington J, Hille B 1994 Growth hormone-releasing hexapeptide elevatesintracellular calcium in rat somatotropes by two mechanisms. Endocrinol-ogy 1135:1100–1108
361. Arimura A 1994 Regulation of growth hormone secretion. In: Imura H (ed)The Pituitary Gland, Raven Press, New York, pp 217–259
362. Adams EF, Lei T, Buchfelder M, Bowers CY, Fahlbusch R 1996 Proteinkinase C-dependent growth hormone releasing peptides stimulate cyclicadenosine 39,59-monophosphate production by human pituitary soma-totropinomas expressing gsp oncogenes: evidence for cross-talk betweentransduction pathways. Mol Endocrinol 10:432–438
363. Pong SS, Chaung LY, Smith RG, GHRP-6 (His-D-Trp-Ala-Trp-D-Phe-Lys-NH2) stimulates growth hormone secretion by depolarization in rat pituitarycell cultures. Program of the 73rd Meeting of The Endocrine Society, Wash-ington DC, 1991 (Abstract 230)
364. Akman MS, Girard M, O’Brien LF, Ho AK, Chik CL 1993 Mechanisms ofaction of a second generation growth hormone-releasing peptide (Ala-His-D-BNal-Ala-Trp-D-Phe-Lys-NH2) in rat anterior pituitary cells. Endocrinol-ogy 132:1286–1291
365. Goth MI, Lyons CE, Canny BJ, Thorner MO 1992 Pituitary adenylatecyclase activating polypeptide, growth hormone (GH)-releasing peptide andGH-releasing hormone stimulate GH release through distinct pituitary re-ceptors. Endocrinology 130:939–944
366. Bercu BB, Yang SW, Masuda R, Walker RF 1992 Role of selected endog-enous peptides in growth hormone-releasing hexapeptide activity: analysisof growth hormone-releasing hormone, thyroid hormone-releasing hor-mone, and gonadotropin-releasing hormone. Endocrinology 130:2579–2586
367. Renner U, Brockmeier S, Strasburger C, Lange M, Schopohl J, Muller O1994 Growth hormone (GH)-releasing peptide stimulation of GH releasefrom human somatotroph adenoma cells: interaction with GH-releasinghormone, thyrotropin-releasing hormone, and octreotide. J Clin EndocrinolMetab 78:1090–1096
368. Iranmanesh A, Gupta PM, Bowers CY, Veldhuis JD, GHRH and GHRP-2combined stimulate supra-additive GH secretion in men in the presence oflow-dose synthetic somatostatin (octreotide): evidence for a novel interac-tion between GHRH and GHRP. Program of the Fifth International PituitaryCongress, Naples, FL, 1998, p 61
369. Massoud AF, Hindmarsh PC, Brook CGD 1997 Interaction of the growthhormone releasing peptide hexarelin with somatostatin. Clin Endocrinol(Oxf) 47:537–547
370. Dickson SL, Viltart O, Bailey AR, Leng G 1997 Attenuation of the growthhormone secretagogue induction of Fos protein in the rat arcuate nucleus bycentral somatostatin action. Neuroendocrinology 66:188–194
371. Fairhall KM, Mynett A, Robinson IC 1995 Central effects of growth hor-mone-releasing hexapeptide (GHRP-6) on growth hormone release are in-hibited by central somatostatin action. J Endocrinol 144:555–560
372. Korbonits M, Trainer PJ, Besser GM 1995 The effect of an opiate antagoniston the hormonal changes induced by hexarelin. Clin Endocrinol (Oxf) 43:365–371
373. Badger TM, Millard WJ, McCormick GF, Bowers CY, Martin JB 1984 Theeffects of growth hormone (GH)-releasing peptides on GH secretion inperifused pituitary cells of adult male rats. Endocrinology 115:1432–1438
374. Malozowski S, Hao EH, Ren Sg Marin G, Liu L, Southers JL, Merriam GR1991 Growth hormone (GH) responses to the hexapeptide GH-releasingpeptide and GH-releasing hormone (GHRH) in the cynomolgus macaque:evidence for non-GHRH-mediated responses. J Clin Endocrinol Metab 73:314–317
375. Wu D, Chen C, Katoh K, Zhang J, Clarke IJ 1994 The effect of GH-releasingpeptide-2 (GHRP-2 or KP 102) on GH secretion from primary cultured ovinepituitary cells can be abolished by a specific GH-releasing factor (GRF)receptor antagonist. J Endocrinol 140:R9–R13
376. Wu D, Chen C, Zhang J, Bowers CY, Clarke IJ 1996 The effects of GH-releasing peptide-6 (GHRP-6) and GHRP-2 on intracellular adenosine 39,59-monophosphate (cAMP) levels and GH secretion in ovine and rat soma-totrophs. J Endocrinol 148:197–205
377. Torsello A, Grilli R, Luoni M, Guidi M, Ghigo MC, Wehrenberg WB,Deghenghi R, Mueller EE, Locatelli V 1996 Mechanism of action of hexare-lin. I. Growth hormone-releasing activity in the rat. Eur J Endocrinol 135:481–488
378. Arvat E, Gianotti L, Divito L, Imbimbo BP, Lenaerts V, Deghenghi R,Camanni F, Ghigo E 1995 Modulation of growth hormone-releasing activityof hexarelin in man. Neuroendocrinology 61:51–56
379. Walker RF, Yang SW, Bercu BB 1991 Robust growth hormone (GH) secre-tion in aged female rats co-administered GH-releasing hexapeptide(GHRP-6) and GH-releasing hormone (GHRH). Life Sci 49:1499–1504
380. Kirk SE, Gertz BJ, Schneider SH, Hartman ML, Pezzoli SS, Wittreich JM,Krupa DA, Seibold JR, Thorner MO 1997 Effect of obesity and feeding onthe growth hormone (GH) response to the GH secretagogue L-692,429 inyoung men. J Clin Endocrinol Metab 82:1154–1159
381. Imbimbo BP, Mant T, Edwards M, Amin D, Boutignon F, Lanaerts V,
Wuthrich p Deghenghi R 1994 Growth hormone-releasing activity ofhexarelin in humans. Eur J Clin Pharmacol 46:421–425
382. Giustina A, Bussi AR, Deghenghi R, Imbimbo B, Licini M, Poiesi C,Wehrenberg WB 1995 Comparison of the effects of growth hormone re-leasing hormone and hexarelin, a novel growth hormone releasing peptide-6analog, on growth hormone secretion in humans with or without glucocor-ticoid excess. J Endocrinol 146:227–232
383. Steiger A, Holsboer F 1997 State of the art review. Neuropeptides andhuman sleep. Sleep 20:1038–1052
384. Obal F, Floyd R, Kapas L, Bodosi B, Krueger JM 1996 Effects of systemicGHRH on sleep in intact and hypophysectomized rats. Am J Physiol 270:E230–E237
385. Obal F, Payne L, Opp M, Alfoldi P, Kapas L, Krueger JM 1992 Growthhormone-releasing hormone antibodies suppress sleep and prevent en-hancement of sleep after sleep deprivation. Am J Physiol 263:R1078–R1085
386. Guldner J, Schier T, Friess E, Colla M, Holsboer F, Steiger A 1997 Reducedefficacy of growth hormone-releasing hormone in modulating sleep endo-crine activity in the elderly. Neurobiol Aging 18:491–495
387. Kern W, Dodt C, Fehm HL 1996 Changes in cortisol and growth hormonesecretion during nocturnal sleep in the course of aging. J Gerontol 51A:M3–M9
388. Kerkhofs M, Van Cauter E, Van Onderbergen A, Caufriez A, Thorner MO,Copinschi G 1993 Sleep-promoting effects of growth hormone-releasinghormone in normal men. Am J Physiol 264:E594–E598
389. Garry P, Roussel B, Cohen R, Biot-Laporte S, Charfi AE, Jouvet M, SassolasG 1985 Diurnal administration of human growth hormone-releasing factordoes not modify sleep and sleep-related growth hormone secretion in nor-mal young men. Acta Endocrinol (Copenh) 110:158–163
390. Spath-Schwalbe E, Hudeborn C, Kern W, Fehm HL, Born J 1995 Nocturnalwakefulness inhibits growth hormone (GH)-releasing hormone-induced GHsecretion. J Clin Endocrinol Metab 80:214–219
391. Toppila J, Alanko L, Asikainen M, Tobler I, Stenberg D, Porkka-Heis-kanen T 1997 Sleep deprivation increases somatostatin and growth hor-mone-releasing hormone messenger RNA in the rat hypothalamus. Neu-rosci Lett 220:97–100
392. Jaffe CA, Ho PJ, DeMott-Friberg R, Bowers CY, Barkan AL 1993 Effects ofa prolonged growth hormone (GH)-releasing peptide infusion on pulsatileGH secretion in normal men. J Clin Endocrinol Metab 77:1641–1647
393. Cordido F, Penalva A, Peino R, Casanueva F, Dieguez C 1995 Effect ofcombined administration of growth hormone (GH)-releasing hormone, GH-releasing peptide-6, and pyridostigmine in normal and obese subjects. Me-tabolism 44:745–748
394. Giustina A, Desenzani P, Perini P, Deghenghi R, Bugari G, WehrenbergWB, Giustina G 1996 Hypothalamic control of growth hormone secretionin type 1 diabetic men: effects of the combined administration of GHRH andhexarelin, a novel GHRP-6 analog. Endocr Res 22:159–174
395. Cappa M, Setzu S, Bernardini S, Carta D, Federici G, Grossi A, Loche S1995 Exogenous growth hormone administration does not inhibit the growthhormone response to hexarelin in normal men. J Endocrinol Invest 18:762–766
396. Massoud AF, Hindmarsh PC, Brook CG 1995 Hexarelin induced growthhormone release is influenced by exogenous growth hormone. Clin Endo-crinol (Oxf) 43:617–621
397. Fletcher TP, Thomas GB, Willoughby JO, Clarke IJ 1994 Constitutivegrowth hormone secretion in sheep after hypothalamo pituitary disconnec-tion and the direct in vivo pituitary effect of growth hormone releasingpeptide 6. Neuroendocrinology 60:76–86
398. Conley LK, Teik JA, Deghenghi R, Imbimbo BP, Giustina A, Locatelli V,Wehrenberg WB 1995 Mechanism of action of hexarelin and GHRP-6: anal-ysis of the involvement of GHRH and somatostatin in the rat. Neuroendo-crinology 61:44–50
399. Bowers CY 1993 GH releasing peptides–structure and kinetics. J PediatrEndocrinol 6:21–31
400. Yagi H, Kaji H, Sato M, Okimura Y, Chihara K 1996 Effect of intravenousor intracerebroventricular injections of His-D-Trp-Ala-Trp-D-Phe-Lys-NH2on GH release in conscious, freely moving male rats. Neuroendocrinology63:198–206
401. McCormick GF, Millard WJ, Badger TM, Bowers CY, Martin JB 1985Dose-response characteristics of various peptides with growth hormone-releasing activity in the unanesthetized male rat. Endocrinology 117:97–105
402. De Marinis L, Mancini A, Valle D, Bianchi A, Gentilella R, Zaltieri G,Giustina A, GH response to hexarelin after meal in normal subjects: pre-liminary data. Program of the 10th International Congress of Endocrinology,1996 (Abstract P2-459)
403. Locatelli V, Grilli R, Torsello A, Cella SG, Wehrenberg WB, Muller EE1994 Growth hormone-releasing hexapeptide is a potent stimulator ofgrowth hormone gene expression and release in the growth hormone-releasing hormone-deprived infant rat. Pediat Res 36:169–174
404. Mitani M, Kaji H, Abe H, Chihara K 1996 Growth hormone (GH)-releasingpeptide and GH releasing hormone stimulate GH release from subpopu-lations of somatotrophs in rats. J Neuroendocrinol 8:825–830
405. Wehrenberg WB, Giustina A Neuroendocrine regulation of growth hor-
782 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

mone secretion. In: Conn M, Freeman M (eds) Neuroendocrinology, Hu-mana Press, Totowa, NJ, chapt. 5, in press
406. Cheng K, Chan WS, Barreto A, Convey EM, Smith RG 1989 The synergisticeffects of His-D-Trp-Ala-Trp-D-Phe-Lys-NH2 on growth hormone (GH)-releasing factor-stimulated GH release and intracellular adenosine 39, 59-monophosphate accumulation in rat primary pituitary cell culture. Endo-crinology 124:2791–2798
407. Edwards CA, Dieguez C, Scanlon MF 1989 Effects of hypothyroidism,tri-iodothyronine and glucocorticoids on growth hormone responses togrowth hormone-releasing hormone and His-D-Trp-Ala-Trp-D-Phe-Lys-NH2. J Endocrinol 121:31–36
408. Dickson SL, Leng G, Dyball RE, Smith RG 1995 Central actions of peptideand non-peptide growth hormone secretagogues in the rat. Neuroendocri-nology 61:36–43
409. Sirinathsinghji DJ, Chen HY, Hopkins R, Trumbauer M, Heavens R,Rigby M, Smith RG, van der Ploeg LH 1995 Induction of c-fos mRNA inthe arcuate nucleus of normal and mutant growth hormone-deficient miceby a synthetic non-peptidyl growth hormone secretagogue. Neuroreport6:1989–1992
410. Locke W, Kirgis HD, Bowers CY, Abdoh AA 1995 Intracerebroventiculargrowth-hormone-releasing peptide-6 stimulates eating without affectingplasma growth hormone responses in rats. Life Sci 56:1347–1352
411. Dickson SL, Doutrelant-Viltart O, Dyball RE, Leng G 1996 Retrogradelylabelled neurosecretory neurones of the rat hypothalamic arcuate nucleusexpress Fos protein following systemic injection of GH-releasing peptide-6.J Endocrinol 151:323–331
412. Dickson SL, Luckman SM 1997 Induction of c-fos messenger ribonucleicacid in neuropeptide Y and growth hormone (GH)-releasing factor neuronsin the rat arcuate nucleus following systemic injection of the GH secreta-gogue, GH-releasing peptide-6. Endocrinology 1238:771–777
413. Dickson SM, Luckman SM 1997 Induction of c-fos messenger ribonucleicacid in neuropeptide Y and growth hormone (GH)-releasing factor neuronsin the rat arcuate nucleus following systemic injection of the GH secreta-gogue, GH-releasing peptide-6. Endocrinology 138:771–777
414. Robinson BM, Friberg RD, Bowers CY, Barkan AL 1992 Acute growthhormone (GH) response to GH-releasing hexapeptide in humans is inde-pendent of endogenous GH-releasing hormone. J Clin Endocrinol Metab75:1121–1124
415. Pandya N, DeMott-Friberg R, Bowers CY, Barkan AL, Jaffe CA 1998Growth hormone (GH)-releasing peptide-6 requires endogenous hypotha-lamic GH-releasing hormone for maximal GH stimulation. J Clin EndocrinolMetab 83:1186–1189
416. Penalva A, Pombo M, Carballo A, Barreiro J, Casanueva FF, Dieguez C1993 Influence of sex, age and adrenergic pathways on the growth hormoneresponse to GHRP-6. Clin Endocrinol (Oxf) 38:87–91
417. Cella SG, Locatelli V, Poratelli M, De Gennaro Colonna V, Imbimbo BB,Deghenghi R, Mueller EE 1995 Hexarelin, a potent GHRP analogue: in-teractions with GHRH and clonidine in young and aged dogs. Peptides16:81–86
418. Thorner MO, Vance ML, Rogol AD, Blizzard RM, Veldhuis JD, VanCauter E, Copinschi G, Bowers CY 1990 Growth hormone-releasing hor-mone and growth hormone-releasing peptide as potential therapeutic mo-dalities. Acta Paediatr Scand [Suppl] 367:29–32
419. Casanueva FF, Micic D, Pombo M, Leal A, Bokser L, Zugaza JL, DieguezC 1996 Role of the new growth hormone-releasing secretagogues in thediagnosis of some hypothalamopituitary pathologies. Metabolism 45:123–126
420. Giustina A, Bossoni S, Cimino A, Pizzocolo G, Romanelli G, WehrenbergWB 1990 Impaired growth hormone (GH) response to pyridostigmine intype I diabetic patients with exaggerated GH-releasing hormone-stimulatedGH secretion. J Clin Endocrinol Metab 71:1486–1490
421. Ghigo E, Aimartti G, Gianotti L, Bellone J, Arvat E, Camanni F 1996 Newapproach to the diagnosis of growth hormone deficiency in adults. Eur JEndocrinol 134:352–356
422. Leal-Cerro A, Garcia E, Astorga R, Casanueva FF, Dieguez C 1995 Growthhormone (GH) responses to the combined administration of GH-releasinghormone plus GH-releasing peptide 6 in adults with GH deficiency. Eur JEndocrinol 132:712–715
423. Micic D, Popovic V, Kendereski A, Peino R, Dieguez C, Casanueva FF 1996The sequential administration of growth hormone-releasing hormone fol-lowed 120 minutes later by hexarelin, as an effective test to assess thepituitary GH reserve in man. Clin Endocrinol (Oxf) 45:543–551
424. Pombo M, Leal-Cerro A, Barreiro J, Penalva A, Peino R, Mallo F, DieguezC, Casanueva FF 1996 Growth hormone releasing hexapeptide-6 (GHRP-6)test in the diagnosis of GH-deficiency. J Pediatr Endocrinol Metab 3:333–338
425. Aimaretti G, Corneli G, Razzore P, Bellone S, Baffoni C, Arvat E, CamanniF, Ghigo E 1998 Comparison between insulin-induced hypoglycemia andgrowth hormone (GH)-releasing hormone plus arginine as provocative testsfor the diagnosis of GH deficiency in adults. J Clin Endocrinol Metab 83:1615–1618
426. Tatemoto K, Rokaeus A, Jornvall H, McDonald T, Mutt V 1983 Galanin–a
novel biologically active peptide from porcine intestine. FEBS Lett 164:128–128
427. Skofitsch G, Jacobowitz D 1985 Quantitative distribution of galanin-likeimmunoreactivity in the central nervous system. Peptides 7:609–613
428. Skofitsch G, Sills M, Jacobowitz D 1986 Autoradiographic distribution of125I-galanin binding sites in the central nervous system. Peptides 7:1029–1042
429. Evans HF, Shine J 1996 Human galanin: molecular cloning reveals a uniquestructure. Endocrinology 129:1682–1686
430. Giustina A, Girelli A, Bodini C, Bonfanti C, Doga M, Schettino M, Negro-Vilar A 1992 Comparative effect of porcine and rat galanin on growthhormone secretion in normal adult men. Horm Metab Res 24:90–91
431. Giustina A, Girelli A, Licini M, Schettino M, Negro-Vilar A 1992 Thyro-tropin and prolactin secretion are not affected by porcine and rat galanin innormal subjects. Horm Metab Res 24:351–352
432. Giustina A, Licini M, Schettino M, Doga M, Pizzocolo G, Negro-Vilar A1994 Physiological role of galanin in the regulation of anterior pituitaryfunction in humans. Am J Physiol 266:E57–E61
433. Negro-Vilar A, Lopez F, Merchenthaler I, Liposits Z, Giustina A 1993 Roleof galanin in pituitary control. In: Melmed S (ed) Molecular and ClinicalAdvances in Pituitary Disorders. Endocrine Research and Education, Inc.,Los Angeles, CA, pp 99–105
434. Murakami Y, Kato Y, Shimatsu A, Koshiyama H, Hattori NY, YanaiharaN, Imura H 1989 Possible mechanisms involved in growth hormone secre-tion induced by galanin in the rat. Endocrinology 124:1224–1229
435. Ottlecz A, Samson W, McCann S 1985 Galanin: evidence for a hypothalamicsite of action to release growth hormone. Peptides 7:51–53
436. Ottlecz A, Snyder GG, McCann SM 1988 Regulatory role of galanin incontrol of hypothalamic pituitary function. Proc Natl Acad Sci USA 85:9861–9865
437. Cella SG, Locatelli V, Gennaro V 198 Epinephrine mediates the growthhormone-releasing effect of galanin in infant rats. Endocrinology 122:855–859
438. Moore Jr JP, Morrison DG, Hyde JF 1994 Galanin gene expression is in-creased in the anterior pituitary gland of the human growth hormone-releasing hormone transgenic mouse. Endocrinology 134:2005–2010
439. Bauer FE, Ginsberg L, Venetikou M, McKay D, Burrin J, Bloom S 1987Growth hormone release in man induced by galanin, a new hypothalamicpeptide. Lancet 11:192–195
440. Davis TM, Burrin JM, Bloom SR 1987 Growth hormone (GH) release inresponse to GH-releasing hormone in man is 3-fold enhanced by galanin.J Clin Endocrinol Metab 65:1248–1252
441. Giustina A, Grelli A, Bossoni S, Legati F, Schettino M, Wehrenberg WB1992 Effect of galanin on growth hormone-releasing hormone-stimulatedgrowth hormone secretion in adult patients with nonendocrine diseases onlong-term daily glucocorticoid treatment. Metabolism 41:548–551
442. Loche S, Vista N, Ghigo E, Vannelli S, Arvat E, Benso L, Corda R, CellaS, Mueller EE, Pintor C 1990 Evidence for involvement of endogenoussomatostatin in the galanin-induced growth hormone secretion in children.Pediatr Res 27:405–407
443. Giustina A, Bussi A, Conti C, Doga M, Legati F, Macca C, Zuccato F,Wehrenberg WB 1992 Comparative effect of galanin and pyridostigmine onthe growth hormone response to growth hormone-releasing hormone innormal aged subjects. Horm Res 37:165–170
444. Giustina A, Bussi A, Legati F, Bossoni S, Licini M, Schettino M, ZuccatoF, Wehrenberg WB 1992 Galanin does not affect the growth hormone-releasing hormone-stimulated growth hormone secretion in patients withhyperthyroidism. Acta Endocrinol (Copenh) 127:504–508
445. Giustina A, Licini M, Bussi AR, Girelli A, Pizzocolo G, Schettino M 1993Effects of sex and age on the growth hormone response to galanin in healthyhuman subjects. J Clin Endocrinol Metab 76:1369–1372
446. Howard G, Peng L, Hyde JF 1997 An estrogen receptor binding site withinthe human galanin gene. Endocrinology 138:4649–4656
447. Giustina A, Bresciani E, Bussi AR, Bollati A, Bonfanti C, Bugari G, ChiesaL, Giustina G 1995 Characterization of the paradoxical growth hormoneinhibitory effect of galanin in acromegaly. J Clin Endocrinol Metab 80:1333–1340
448. Maiter DM, Hooi SC, Koenig JI, Martin JB 1990 Galanin is a physiologicalregulator of spontaneous pulsatile secretion of growth hormone in the malerat. Endocrinology 126:1216–1222
449. Walli R, Schaefer H, Morys-Wortmann C, Paetzold G, Nustede R, SchmidtWE 1994 Identification and biochemical characterization of the human braingalanin receptor. J Mol Endocrinol 13:347–356
450. Wynic D, Small C, Ghatei M, Bloom SR, Pachnis V, Targeted disruptionof the murine galanin gene lowers serum prolactin levels and abolisheslactation. Program of the 10th International Congress of Endocrinology, SanFrancisco, CA, 1996 (Abstract 99)
451. Vrontakis E, Perumal P, Faracl C, Galanin overexpression in the pituitaryof transgenic mice. Program of the 10th International Congress of Endocri-nology, San Francisco, CA, 1996 (Abstract 957)
452. Austin LA, Hunter III H 1981 Calcitonin. Physiology and pathophysiology.N Engl J Med 304:269–278
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 783
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

453. Fahrenkrung J, Hornum I, Rehfeld JF 1975 Effects of calcitonin on serumgastrin concentration component pattern in man. J Clin Endocrinol Metab41:149–152
454. Passariello N, Giugliano D, Sgambato S, Torella R, D’Onofrio F 1981Calcitonin, a diabetogenic hormone? J Clin Endocrinol Metab 53:318–323
455. Leicht E, Biro’ G, Weinges KF 1994 Inhibition of releasing-hormone inhealthy human subjects. J Clin Endocrinol Metab 76:1369–1372
456. Fisher JA, Tobler PH, Kauffman M, Born M, Cooper PE, Sagar SM, MartinJB 1981 Calcitonin regional distribution of the hormone and its binding sitesin the human brain and pituitary. Proc Natl Acad Sci USA 78:7801–7805
457. Lengyel AMJ, Tannenbaum GS 1987 Mechanism of calcitonin-inducedgrowth hormone suppression: roles of somatostatin and GH-releasing fac-tor. Endocrinology 120:1377–1383
458. Cantalamessa L, Catania A, Reschini E, Peracchi M 1978 Inhibitory effectof calcitonin on growth hormone and insulin secretion in man. Metabolism27:987–992
459. Petralito A, Lunetta M, Liuzzo A, Mangiafico RA, Fiore CE 1979 Effects ofsalmon calcitonin on insulin-induced growth hormone release in man. HormMetab Res 11:641–643
460. Giustina A, Romanelli G, Doga M, Pizzocolo G, Giustina G 1989 Effectsof intranasal calcitonin administration on pituitary GH response to hGHRH(1–29)NH2 in normal adult subjects. Clin Endocrinol (Oxf) 30:699–704
461. Giustina A, Bodini C, Bossoni S, Doga M, Grelli A, Pizzocolo G,Wehrenberg WB 1990 Effect of calcitonin on GH response to pyridostigminein combination with hGHRH (1–29)NH2 in normal adult subjects. ClinEndocrinol (Oxf) 33:375–380
462. Giustina A, Doga M, Bodini C, Grelli A, Legati F, Bossoni S, RomanelliG 1990 Acute effects of cortisone acetate on growth hormone response togrowth hormone-releasing hormone in normal adult subjects. Acta Endo-crinol (Copenh) 122:206–210
463. Laurian L, Oberman Z, Graf E, Gilad S, Hoerer E, Simantov R 1986 Cal-citonin induced increase in ACTH, beta-endorphin and cortisol secretion.Horm Metab Res 18:268–271
464. Giustina A, Grelli A, Doga M, Bodini C, Bossoni S, Romanelli G,Wehrenberg WB 1990 Pyridostigmine blocks the inhibitory effect ofglucocorticoids on growth hormone releasing hormone stimulated growthhormone secretion in normal men. J Clin Endocrinol Metab 71:580–584
465. Borle AB 1975 Regulation of cellular calcium metabolism and calcium trans-port by calcitonin. J Membr Biol 21:125–146
466. Thorner MO, Holl RW, Leong DA 1988 The somatotrope: an endocrine cellwith functional calcium transients. J Exp Biol 139:169–179
467. Miyata A, Arimura A, Dahl RR, Minamino N, Uehara A, Yiang L, CullerMD, Coy DH 1989 Isolation of a novel 38 residue-hypothalamic polypeptidewhich stimulates adenylate cyclase in pituitary cells. Biochem Biophys ResCommun 164:567–574
468. Arimura A, Somogywari-Vigh A, Mizuno K, Coy DH, Kitada C 1991 Tissuedistribution of PACAP as determined by RIA: highly abundant in the ratbrain and testis. Endocrinology 129:2787–2789
469. Tamada Y, Tanaka M, Ichitani Y, Okamura H, Yanaihara N, Ibata Y 1994Pituitary adenylate cyclase activating polypeptide (PACAP)-like immuno-reactive neuronal elements in rat hypothalamus and median eminence withspecial references to morphological background of its effects on anteriorpituitary-light and electron microscopic immunocytochemistry. NeurosciLett 180:105–108
470. Kimura S, Ohshige Y, Lin L, Okamura T, Yanaiara C, Yanaiara N, ShiotanyY 1994 Localization of pituitary adenylate cyclase activating polypeptide(PACAP) in the hypothalamus-pituitary system in rats: light and electronmicroscopic immunocytochemical studies. J Neuroendocrinol 6:503–507
471. Dow RC, Bennie J, Fink G 1994 Pituitary adenylate cyclase activatingpeptide-38 (PACAP) 38 is released into hypophysial portal blood in thenormal male and female rat. J Endocrinol 142:R1–R4
472. Jarry A, Esmhardt S, Schmidt WE, Creutzfedt W, Wuttke W 1992 Con-cerning effects of pituitary adenylate cyclase activating peptide (PACAP) onin vivo and in vitro prolactin and growth hormone release in male rats. LifeSci 5:823–830
473. Goth MI, Lyons CE, Canny BJ, Thorner MO 1992 Pituitary adenylatecyclase activating polypeptide (PACAP), growth hormone releasing peptideand GH releasing hormone stimulate GH release through distinct pituitaryreceptors. Endocrinology 130:939–944
474. Propato-Mussafiri R, Kanse SM, Ghatei MA, Bloom SR 1992 Pituitaryadenylate cyclase activating polypeptide (PACAP) release, adrenocortico-tropin, growth hormone and prolactin from the mouse and rat clonal pi-tuitary cell lines ATT-20 and GH3. J Endocrinol 132:107–113
475. Yamauchi K, Murakami Y, Koshimura K, Nishiki J, Tanaka J, Kato Y 1996Involvement of pituitary adenylate cyclase activating polypeptide in growthhormone secretion induced by serotoninergic mechanisms in the rat. En-docrinology 137:1693–1697
476. Murakami Y, Koshimura K, Yamauchi K, Nishiki M, Tanaka J, Furuya H,Myake T, Kato Y 1995 Pituitary adenylate cyclase activating polypeptidestimulates growth hormone release from GH3 cells through type II PACAPreceptor. Regul Pept 56:35–40
477. Pisegna JR, Wank SA 1993 Molecular cloning and functional expression of
the pituitary adenylate cyclase activating polypeptide type I receptor. ProcNatl Acad Sci USA 90:6345–6349
478. Delitala G, Grossman A, Besser GM 1983 Opiate peptides control growthhormone through a cholinergic mechanism in man. Clin Endocrinol (Oxf)18:401–405
479. Murakami Y, Kato Y, Kabayama Y, Inhoue T, Kojo K, Otha H, Imhura H1985 Inhibition by antiserum to rat growth hormone release factor of growthhormone secretion induced by a met-enkephalin-analog, FK33 824, in rats.Proc Soc Exp Biol Med 178:151–154
480. Drouva SV, Epelbaum J, Tapia-Arancibia L, Laplante E, Kordon C 1980Met-enkephalin inhibition of K1-induced LHRH and SRIF release from ratmediobasal hypothalamic slices. Eur J Pharmacol 61:411–412
481. Drouva SV, Epelbaum J, Tapia-Arancibia L, Laplante E, Kordon C 1981Opiate receptors modulate LHRH and SRIF release from mediobasal hy-pothalamic neurons. Neuroendocrinology 32:163–167
482. Moretti C, Fabbri A, Gnessi L, Cappa M, Calzolari A, Fraioli F, GrossmanA, Besser GM 1983 Naloxone inhibits exercise-induced release of PRL andGH in athletes. Clin Endocrinol (Oxf) 18:135–138
483. Mueller EE, Panerai A, Cocchi D, Gil-Ad I, Rossi G, Olgiati V 1977 Growthhormone releasing activity of thyrotropin-releasing hormone in rats withhypothalamic lesions. Endocrinology 100:1663–1667
484. Giustina A, Doga M, Bresciani E, Bussi AR, Chiesa L, Misitano V, GiustinaG 1995 Effect of glucocorticoids on the paradoxical growth hormone re-sponse to thyrotropin-releasing hormone in patients with acromegaly. Me-tabolism 44:379–383
485. Valentini U, Cimino A, Rotondi A, Pelizzari R, Giustina A, Marchetti C,Romanelli G 1989 Growth hormone response to thyrotropin releasing hor-mone and placebo in a group of insulin-dependent diabetic patients. JEndocrinol Invest 12:643–646
486. Czernichow P, Dauzet MC, Broyer M, Rappaport R 1976 Abnormal TSH,PRL and GH responses to TSH-releasing factor in chronic renal failure. J ClinEndocrinol Metab 43:630–637
487. Scanlon MF, Peters JR, Foord SM, Dieguez C, Hall R 1983 Clinical appli-cation of TRH. In: Griffiths EC, Bennet W (eds) Thyrotropin ReleasingHormone. Raven Press, New York, pp 303–314
488. Maeda K, Chihara K, Kato Y, Ohgo S, Iwasaki Y, Hiromi A, Imura H 1976Suppression by thyrotropin releasing hormone of growth hormone releaseinduced by arginine and insulin-induced hypoglycemia. J Clin EndocrinolMetab 43:453–456
489. Jordan V, Dieguez C, Valcavi R, Artioli C, Portioli I, Rodriguez-Arnao MdGomez-Pan A, Scanlon MF 1986 Lack of effect of muscarinic cholinergicblockade on the GH responses to GRF 1–29 and TRH in acromegalic subjects.Clin Endocrinol (Oxf) 24:291–298
490. Szabo M’PC, Kramer DE 1985 Growth hormone response to thyrotropinreleasing hormone in the urethane anesthetized rat: effect of thyroid status.Endocrinology 117:330–337
491. Giustina A, Ragni G, Bollati A, Cozzi R, Licini M, Poiesi C, turazzi S,Bonfanti C 1997 Inhibitory effects of galanin on growth hormone release incultured GH-secreting adenoma cells: comparative study with octreotide,GH-releasing hormone, and thyrotropin-releasing hormone. Metabolism46:1–5
492. Gershengorn ML 1978 Bihormonal regulation of the thyrotropin releasinghormone receptor in mouse thyrotropic tumor cells in culture. J Clin Invest62:937–943
493. Dieguez C, Page MD, Scanlon MF 1988 Growth hormone neuroregulationand its alterations in disease states. Clin Endocrinol (Oxf) 28:109–143
494. Williams T, Maxon H, Thorner MO, Frohman LA 1985 Blunted growthhormone (GH) response to GH-releasing hormone in hypothyroidism re-solves in the euthyroid state. J Clin Endocrinol Metab 61:454–456
495. Liukko P, Erkkola R, Lammintausta R, Gronroos M, Kloosterboer HJ 1987Blood glucose, serum insulin, serum growth hormone and serum glycosy-lated proteins during two years’ oral contraception with low-estrogen com-binations. Ann Chir Gynaecol Suppl 202:45–49
496. Knigge U, Thuesen B, Dejgaard A, Svenstrup B, Bennett P 1990 Histamine-induced paradoxical GH response to TRH/GnRH in men and women:dependence on gonadal steroid hormones. Acta Endocrinol (Copenh) 122:354–360
497. Harfstrand A, Eneroth P, Agnati L, Fuxe K 1987 Further studies on theeffects of central administration of neuropeptide Y on neuroendocrine func-tion in the male rat: relationship to hypothalamic catecholamines. Regul Pept17:167–179
498. Rettori V, Milenkovic L, Aguila MC, McCann SM 1990 Physiologicallysignificant effect of neuropeptide Y to suppress growth hormone release bystimulating somatostatin discharge. Endocrinology 126:2296–2301
499. Catzeflis C, Pierroz DD, Rohner-Jeanrenaud F, Rivier JE, Sizonenko PC,Aubert ML 1993 Neuropeptide Y administered chronically into the lateralventricle profoundly inhibits both the gonadotropic and the somatotropicaxis in intact adult female rats. Endocrinology 132:224–234
500. Suzuki N, Okada K, Minami S, Wakabayashi I 1996 Inhibitory effect ofneuropeptide Y on growth hormone secretion in rats is mediated by both Y1-and Y2-receptor subtypes and abolished after anterolateral deafferentationof the medial basal hypothalamus. Regul Pept 65:145–151
784 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

501. Adams EF, Venetikou MS, Woods CA, Lacoumenta S, Burrin JM 1987Neuropeptide Y directly inhibits growth hormone secretion by human pi-tuitary somatotropic tumors. Acta Endocrinol (Copenh) 115:149–154
502. Tilemans D, Andries M, Denef C 1992 Luteinizing hormone-releasinghormone and neuropeptide Y influence deoxyribonucleic acid replication inthree anterior pituitary cell types. Evidence for mediation by growth factorsreleased from gonadotrophs. Endocrinology 130:882–894
503. Watanobe H, Tamura T 1996 Stimulation by neuropeptide Y of growthhormone secretion in prolactinoma in vivo. Neuropeptides 30:429–432
504. Peng C, Huang YP, Peter RE 1990 Neuropeptide Y stimulates growth hor-mone and gonadotropin release from the goldfish pituitary in vitro. Neu-roendocrinology 52:28–34
505. Kamegai J, Minami S, Sugihara H, Higuchi H, Wakabayashi I 1994 Growthhormone induces expression of the c-fos gene on hypothalamic neuropep-tide-Y and somatostatin neurons in hypophysectomized rats. Endocrinology135:2765–2771
506. Chan YY, Steiner RA, Clifton DK 1996 Regulation of hypothalamic neu-ropeptide-Y neurons by growth hormone in the rat. Endocrinology 137:1319–1325
507. Angel P, Karin M 1991 The role of Jun, Fos and the AP-1 complex incell-proliferation and transformation. J Biol Chem 1072:129–157
508. Minami S, Kamegai J, Sugihara H, Suzuki N, Higuchi H, Wakabayashi I1995 Central glucoprivation evoked by administration of 2-deoxy-D-glucoseinduces expression of the c-fos gene in subpopulation of neuropeptide Yneurons in the rat hypothalamus. Brain Res Mol Brain Res 33:305–310
509. Bergendahl M, Veldhuis JD 1995 Altered pulsatile gonadotropin signalingin nutritional deficiency in the male. Trends Endocrinol Metab 6:145–159
510. Brown E, Roth K, Krause J 1991 Sexually dimorphic distribution of sub-stance P in specific anterior pituitary cell populations. Proc Natl Acad SciUSA 88:1222–1226
511. Coiro V, Volpi R, Capretti L, Speroni G, Bocchi R, Caffarri G, Colla R,Rossi G, Chiodera P 1992 Intravenously infused substance P enhances basaland growth hormone (GH) releasing hormone-stimulated GH secretion innormal men. Peptides 13:843–846
512. Benitez L, Mallo F, Alvarez CV, Burguera B, Sanchez-Franco F, DieguezC 1990 Estrogen-dependent effects of bombesin on in vivo growth hormonesecretion in the rat. Neuroendocrinology 52:608–611
513. Houben H, Denef C 1991 Effect of the bombesin receptor blockers [Leu13,psi CH2NH-Leu14]bombesin and N-pivaloyl GRP(20–25) alkylamide(L 686,095–001C002) on basal and neuromedin C-stimulated PRL andGH release in pituitary cell aggregates. Peptides 12:371–374
514. Scarpignato C, Tirelli F, Pontiroli AE 1986 Bombesin inhibits growth hor-mone response to insulin-induced hypoglycemia in humans. Brain Res 371:187–189
515. Pontiroli AE, Scarpignato C 1986 Effect of bombesin on basal and stimulatedsecretion of some pituitary hormones in humans. Horm Res 23:129–135
516. Valcavi R, Dieguez C, Azzarito C, Edwards CA, Dotti C, Page MD, PortioliI, Scanlon MF 1987 Effect of oral administration of melatonin on GH re-sponse to GRF 1–44 in normal subjects. Clin Endocrinol (Oxf) 26:453–458
517. Smythe GA, Lazarus L 1974 Suppression of human growth hormone se-cretion by melatonin and cyproheptadine. J Clin Invest 54:116–121
518. Koulu M, Lammintausta R 1979 Effect of melatonin on L-tryptophan andaphomorphine stimulated growth hormone secretion in man. J Clin Endo-crinol Metab 49:70–72
519. Bellastella A, Colucci C, D’Alessandro B, Cicero ML 1977 L-Dopa stimu-lated growth hormone release in the blind. J Clin Endocrinol Metab 44:194–195
520. Krieger DT, Glick S 1971 Absent sleep peak of growth hormone relrase inblind subjects: correlation with sleep EEG stages. J Clin Endocrinol Metab3:847–850
521. Niimi M, Sato M, Wada Y, Tamaki M, Takahara J, Kawanishi K 1994Analysis of growth hormone release from rat anterior pituitary cells byreverse hemolytic plaque assay: influence of interleukin-1. Life Sci 55:1807–1813
522. Carro E, Senaris R, Considine RV, Casanueva FF, Dieguez C 1997 Regu-lation of in vivo growth hormone secretion by leptin. Endocrinology 139:2203–2206
523. Roubenoff R, Rall LC, Veldhuis JD, Kehayias JJ, Rosen C, Nicolson M,Lundgren N, Reichlin S 1998 The relationship between growth hormonekinetics and sarcopenia in postmenopausal women: the role of fat mass andleptin. J Clin Endocrinol Metab 83:1502–1506
524. Bergendahl M, Evans WS, Iranmanesh A, Veldhuis JD, 1998 Fasting trig-gers profound hypoleptinemia with reciprocal activation of disorderly pul-satile GH secretion in mid-luteal phase women. Program of the 80th AnnualMeeting of The Endocrine Society, New Orleans, LA, 1998 (Abstract P2–180)
525. Rivier C, Vale W 1984 Corticotropin-releasing factor (CRF) acts centrally toinhibit growth hormone secretion in the rat. Endocrinology 114:2409–2411
526. Mounier F, Pellegrini E, Kordon C, Epelbaum J, Bluet-Pajot MT 1997Continuous intracerebroventricular administration of a corticotropin releas-ing hormone antagonist amplifies spontaneous growth hormone pulses inthe rat. J Endocrinol 152:431–436
527. Coleman ES, Elsasser TH, Kemppainen RJ, Coleman DA, Sartin JL 1993
Effect of endotoxin on pituitary hormone secretion in sheep. Neuroendo-crinology 58:111–122
528. Magnan E, Mazzocchi L, Cataldi M, Guillaume V, Dutour A, Dadoun F1995 Effect of actively immunizing sheep against growth hormone-releasinghormone or somatostatin on spontaneous pulsatile and neostigmine-induced growth hormone secretion. J Clin Endocrinol Metab 144:83–90
529. Fletcher TP, Clarke IJ 1994 Effects of hypothyroidism on the growth hor-mone axis in sheep. J Endocrinol 140:495–502
530. Van Kessel AG, Laarveld B 1993 Acute or chronic immunoneutralizationof somatostatin does not affect growth hormone or thyroid hormone secre-tion in sheep. J Endocrinol 136:261–269
531. Magnan E, Cataldi M, Guillaume V, Mazzocchi L, Dutour A, Conte-Devolx B, Giraud P, Oliver C 1993 Neostigmine stimulates growth hor-mone-releasing hormone release into hypophysial portal blood of conscioussheep. Endocrinology 132:1247–1251
532. Locatelli V, Torsello A, Redaelli M, Ghigo E, Massara F, Camanni F,Mueller EE 1986 Cholinergic agonist and antagonist drugs modulate thegrowth hormone response to growth hormone-releasing hormone in the rat:evidence for mediation by somatostatin. J Endocrinol 111:271–278
533. Torsello A, Panzeri G, Cermenati P, Caroleo MC, Ghigo E, Camanni F,Mueller EE, Locatelli V 1988 Involvement of the somatostatin and cholin-ergic systems in the mechanism of growth hormone autofeedback regulationin the rat. J Endocrinol 117:273–281
534. Arce V, Cella SG, Locatelli V, Mueller EE 1991 Studies of growth hormonesecretion in calorically restricted dogs: effect of cholinergic agonists andantagonist, glucose and thyrotropin-releasing hormone. Neuroendocrinol-ogy 53:467–472
535. Casanueva FF, Villaneuva L, Cabranes JA, Cabezas-Carrato J, Fernandez-Cruz A 1984 Cholinergic mediation of growth hormone secretion elicited byarginine, clonidine and physical exercise in man. J Clin Endocrinol Metab59:526–530
536. Massara F, Ghigo E, Molinatti GM, Mazza E, Locatelli V, Mueller EE,Caminni F 1986 Potentiation of cholinergic tone by pyridostigmine bromidere-instates and potentiates the growth hormone responsiveness to intermit-tent administration of growth hormone-releasing factor in man. Acta En-docrinol (Copenh) 113:12–16
537. Penalva A, Muruais C, Casanueva FF, Dieguez C 1990 Effect of enhance-ment of endogenous cholinergic tone with pyridostigmine on the dose-response relationship of growth hormone (GH)-releasing hormone-inducedGH secretion in normal subjects. J Clin Endocrinol Metab 70:324–327
538. Taylor BJ, Smith PJ, Brook CGD 1985 Inhibition of physiological growthhormone secretion by atropine. Clin Endocrinol (Oxf) 22:497–501
539. Peters JR, Evans PJ, Page MD, Hall R, Gibbs JT, Dieguez C, Scanlon MF1986 Cholinergic muscarinic receptor blockade with pirenzepine abolishesslow wave sleep-related growth hormone release in normal adult males. ClinEndocrinol (Oxf) 25:213–217
540. Delitala G, Maouli M, Pacificio A, Briauda SA, Palermo M, Manerelli M1993 Cholinergic receptor control mechanisms for L-dopa, apomorphine,and clonidine-induced growth hormone secretion in man. J Clin EndocrinolMetab 57:1145–1149
541. Massara F, Ghigo E, Emislis K, Tangolo D, Mazza E, Locatelli V, MuellerEE, Molinatti GM, Caminni F 1986 Cholinergic involvement in the growthhormone releasing hormone-induced growth hormone release: studies innormal and acromegalic subjects. Neuroendocrinology 43:679–675
542. Evans PJ, Dieguez C, Foord S, Peters JR, Hall R, Scanlon MF 1985 The effectof cholinergic blockade on the growth hormone and prolactin response toinsulin hypoglycemia. Clin Endocrinol (Oxf) 22:733–737
543. Rigamonti AE, Marazzi N, Cella SG, Cattaneo L, Mueller EE 1998 Growthhormone responses to growth hormone-releasing hormone and hexarelin infed and fasted dogs: effect of somatostatin infusion or pretreatment withpirenzepine. J Endocrinol 156:341–348
544. Mueller EE 1997 Cholinergic function and neural control of GH secretion.A critical re-appraisal. Eur J Endocrinol 137:338–342
545. Casanueva FF, Betti R, Cella SG, Mueller EE, Mantegazza P 1983 Effectsof agonists and antagonists of cholinergic neurotransmission on growthhormone release in the dog. Acta Endocrinol (Copenh) 103:15–20
546. Peterfreund RA, Vale WW 1983 Muscarinic cholinergic stimulation of so-matostatin secretion from long term dispersed cell cultures of fetal rathypothalamus: inhibition by gamma-aminobutyric acid and serotonin. En-docrinology 112:526–534
547. Richardson SB, Hollander CS, De’Eletto R 1980 Acetylcholine inhibits therelease of somatostatin from rat hypothalamus in vitro. Endocrinology 107:1837–1842
548. Chihara K, Arimura A, Schally AV 1979 Effect of intraventricular injectionof dopamine, noreprinephrine, acetylcholine, and 5-hydroxytryptamine onimmunoreactive somatostatin release into rat hypophyseal portal blood.Endocrinology 104:1656–1662
549. Brogan RS, Fife SK, Conley LK, Giustina A, Wehrenberg WB 1997 Effectsof food deprivation on the GH axis: immunocytochemical and molecularanalysis. Neuroendocrinology 65:129–135
550. Giustina A, Misitano V, Voltz D, Piering A, Wehrenberg WB 1995 Ad-renergic and cholinergic involvement in basal and growth-hormone-releas-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 785
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

ing hormone-stimulated growth hormone secretion in glucocorticoid-treated rats. Endocr Rev 21:719–732
551. Locatelli V, Torsello A, Redaelli M, Ghigo E, Massara F, Mueller EE 1987Cholinergic agonist and antagonist drugs modulate the growth hormoneresponse to growth hormone-releasing hormone in the rat: evidence formediation by somatostatin. J Endocrinol 111:271–278
552. Giustina A, Girelli A, Alberti D, Bossoni S, Buzi F, Schettino M, We-hrenberg WB 1991 Effects of pyridostigmine on spontaneous and growth-hormone-releasing hormone stimulated growth hormone secretion in chil-dren on daily glucocorticoid therapy after liver transplantation. ClinEndocrinol (Oxf) 35:391–498
553. Ismail IS, Scanlon MF, Peters JR 1993 Cholinergic control of growth hor-mone (GH) responses to GH-releasing hormone in insulin dependent dia-betics: evidence for attenuated hypothalamic somatostatinergic tone anddecreased GH autofeedback. Clin Endocrinol (Oxf) 38:149–157
554. Wood TL, Berelowitz M, Gelato MC, Roberts CTJ, LeRoith D, Millard WJ,McKelvy JF 1991 Hormonal regulation of rat hypothalamic neuropeptidemRNAs: effect of hypophysectomy and hormone replacement on growthhormone-releasing factor, somatostatin and the insulin-like growth factors.Neuroendocrinology 53:298–305
555. Chowen JA, Argente J, GonzalezParra S, Garcia Segura LM 1993 Differ-ential effects of the neonatal and adult sex steroid environments on theorganization and activation of hypothalamic growth hormone-releasinghormone and somatostatin neurons. Endocrinology 133:2792–2802
556. Janowski BA, Ling NC, Giustina A, Wehrenberg WB 1993 Hypothalamicregulation of growth hormone secretion during food deprivation in the rat.Life Sci 52:981–987
557. Miki N, Ono M, Shizume K 1988 Withdrawal of endogenous somatostatininduces secretion of growth hormone-releasing factor in rats. J Endocrinol117:245–252
558. Mendelson WB, Lantigua Ra Wyatt RJ, Gillin JC, Jacobs LS 1981 Piperidineenhances sleep-related and insulin-induced growth hormone secretion: fur-ther evidence for a cholinergic secretory mechanism. J Clin EndocrinolMetab 52:409–415
559. Peillon F, Cesselin F, Bression B 1979 In vitro effect of dopamine and L-dopaon prolactin and growth hormone release from human pituitary adenomas.J Clin Endocrinol Metab 49:737–741
560. Liuzzi A, Chiodini PG, Botalla L, Cremascoli G, Mueller EE, Silvestrini F1974 Decreased plasma growth hormone (GH) levels in acromegalics fol-lowing CB 154 (2-Br-alpha-ergocryptide) administration. J Clin EndocrinolMetab 38:910–912
561. Page MD, Dieguez C, Valcavi R, Edwards C, Hall R, Scanlon MF 1988Growth hormone (GH) responses to arginine and L-dopa alone and afterGHRH pretreatment. Clin Endocrinol (Oxf) 28:551–558
562. Lal S, Nair NP, Thavundayil JX, Tawar V, Tesfaye Y, Dastoor D, GauthierS, Guyda H 1989 Growth hormone response to apomorphine, a dopaminereceptor agonist, in normal aging and in dementia of the Alzheimer type.Neurobiol Aging 10:227–231
563. Mueller EE 1987 Neural control of somatotropic function. Physiol Rev67:962–1053
564. Kitajima N, Chihara K, Abe H, Okimura Y, Fujii Y, Satoa M, ShakutsuiS, Watanabe M, Fujita T 1986 Effects of dopamine on immunoreactivegrowth hormone-releasing factor and somatostatin secretion from rat hy-pothalamic slices perifused in vitro. Endocrinology 124:69–76
565. Lewis BM, Dieguez C, Lewis M, Hall R, Scanlon MF 1986 HypothalamicD2 receptors mediate the preferential release of somatostatin-28 in responseto dopaminergic stimulation. Endocrinology 119:1712–1717
566. West CR, Lookingland KJ, Tucker HA 1997 Regulation of growth hormone-releasing hormone and somatostatin from perifused, bovine hypothalamicslices. II. Dopamine receptor regulation. Domest Anim Endocrinol 14:349–357
567. Schober E, Frisch H, Waldhauser F, Bieglmayr C 1989 Influence of estrogenadministration on growth hormone respnse to GHRH and L-Dopa in pa-tients with Turner’s syndrome. Acta Endocrinol (Copenh) 120:442–446
568. Moll GW, Rosenfield RL, Fang VS 1986 Administration of low-dose es-trogen rapidly and directly stimulates growth hormone production. Am JDis Child 140:124–127
569. Delitala G, Palermo M, Ross R, Coy D, Besser GM, Grossman A 1987Dopaminergic and cholinergic influences on the growth hormone responseto growth hormone releasing hormone in man. Neuroendocrinology 45:243–247
570. Vance ML, Kaiser DL, Frohman LA, River J, Vale WW, Thorner MO 1987Role of dopamine in the regulation of growth hormone secretion: dopamineand bromocriptine augment growth hormone (GH)-releasing hormone-stimulated GH secretion in normal man. J Clin Endocrinol Metab 64:1136–1141
571. Wolf PD, Lantigua R, Lee LA 1979 Dopamine inhibition of stimulatedgrowth hormone secretion: evidence for dopaminergic modulation of in-sulin and L-dopa-induced growth hormone secretion in man. J Clin Endo-crinol Metab 49:326–330
572. Bansal SA, Lee LA, Wolf PD 1981 Dopaminergic modulation of arginine-
mediated growth hormone and prolactin release in man. Metabolism 30:649–653
573. Chihara K, Kashio Y, Kita T, Okimura J, Kaji H, Abe H, Fushita T 1986L-dopa stimulates release of hypothalamic growth hormone releasing hor-mone in humans. J Clin Endocrinol Metab 62:466–473
574. Cella SG, Luceri M, Cattaneo L, Torsello A, Mueller EE 1996 Somatostatinwithdrawal as generator of pulsatile GH release in the dog: a possible toolto evaluate the endogenous GHRH tone? Neuroendocrinology 63:481–488
575. Giustina A, Bussi AR, Doga M, Licini M, Schettino M, Wehrenberg WB1993 Effects of metoclopramide on the growth hormone response to galaninin normal man. Horm Metab Res 25:489–492
576. Arce V, Lima L, Lois N, Rodriguez A, Diaz MJ, Tresguerres JAF, DevesaJ 1991 Role of central dopaminergic pathways in the neural control of growthhormone secretion in normal men: studies with metoclopramide. Neuroen-docrinology 53:143–149
577. Joradn V, Dieguez C, Lafaffian I, Rodriguez-Arnao MD, Gomez-Pan A,Hall R, Scanlon MF 1986 Influence of dopaminergic, adrenergic and cho-linergic blockade and TRH administration on GH responses to GRF 1–29.Clin Endocrinol (Oxf) 24:291–298
578. Chiodera P, Coiro V, Zanardi G, Volpi R, Valenti R, Butturini U 1982 Effectof metoclopramide on serum GH levels in normal women. Horm Metab Res14:103–104
579. Tatar P, Vigas M 1980 Role of alpha-1 and alpha-2 adrenergic receptors inthe GH and prolactin response to insulin-induced hypoglycemia in man.Neuroendocrinology 39:275–281
580. Ghigo E, Bellone J, Arvat E, Mazza E, Cella SG, Brambilla F, Mueller EE,Camanni F 1990 Effect of a and b-adrenergic agonists and antagonists ongrowth hormone secretion in men. J Neuroendocrinol 2:157–159
581. Cella SG, Morgese M, Mantegazza P, Mueller EE 1984 Inhibitory action ofthe a1 adrenergic receptor on growth hormone secretion in the dog. Endo-crinology 114:2046–2048
582. Mounier F, Bluet-Pajot MT, Durand D, Kordon C, Epelbaum J 1994 Alpha-1-nonadrenergic inhibition of growth hormone secretion is mediatedthrough the paraventricular hypothalamic nucleus in male rats. Neuroen-docrinology 59:29–34
583. Lancranjan I, Marback P 1977 New evidence for GH modulation by alpha-adrenergic system in man. Metabolism 26:1225–1229
584. Cella SG, Picotti GB, Mueller EE 1983 Alpha 2-adrenergic stimulationenhances growth hormone secretion in the dog: a presynaptic mechanism?Life Sci 32:2785–2792
585. Miki N, Ono M, Shizume K 1984 Evidence that opiatergic and alpha-adrenergic mechanisms stimulate rat growth hormone release via growthhormone-releasing factor (GRF). Endocrinology 114:1950–1952
586. McWilliam JR, Melldrum BS 1983 Noradrenergic regulation of growthhormone secretion in the baboon. Endocrinology 112:254–259
587. Brown MJ, Struthers AD, Burrin JM, Di Silvio L, Brown DC 1985 Thephysiological and pharmacological role of presynaptic alpha and beta ad-renoceptor in men. Br J Clin Pharmacol 20:649–658
588. Giustina A, Doga M, Bodini C, Bossoni S, Romanelli G 1990 Centralalpha2-adrenergic function in patients with essential hypertension. HormMetab Res 22:451–452
589. Katakami H, Kato Y, Matsushita N, Imura H 1984 Effects of neonataltreatment with monosodium glutamate on growth hormone release inducedby clonidine and prostaglandin E1 in conscious male rats. Neuroendocri-nology 38:1–5
590. Giustina A, Buffoli MG, Bussi AR, Doga M, Girelli A, Pizzocolo G, PozziA, Wehrenberg WB 1992 Comparative effect of clonidine and growth hor-mone (GH)-releasing hormone on GH secretion in adult patients on chronicglucocorticoid therapy. Horm Metab Res 24:240–243
591. Lanzi R, Lapointe M, Gurd W, Tannenbaum GS 1994 Evidence for aprimary involvement of somatostatin in clonidine-induced growth hormonerelease in conscious rats. J Endocrinol 141:259–266
592. Magnan E, Cataldi M, Guillaume V, Mazzocchi L, Dutour A, Razafind-raibe H, Sauze N, Renard M, Oliver C 1994 Role of growth hormone(GH)-releasing hormone and somatostatin in the mediation of clonidine-induced GH release in sheep. Endocrinology 134:562–567
593. Giustina A, Bossoni S, Licini M, Macca C, Milani G, Wehrenberg WB 1994Effect of the combined administration of galanin and clonidine on serumgrowth hormone levels in normal subjects and in patients under chronicglucocorticoid treatment. Endocr Rev 20:151–164
594. Giustina A, Buffoli MG, Bussi AR, Weherenberg WB 1991 Acute affect ofclonidine and growth hormone (GH)-releasing hormone on GH secretion inpatients with hyperthyroidism. Horm Res 36:192–195
595. Valcavi R, Dieguez C, Page MD 1988 Alpha-2-adrenergic pathways releasegrowth hormone via a non-GRF-dependent mechanism in normal humansubjects. Clin Endocrinol (Oxf) 29:309–316
596. Phipps WR, Campbell BF, Pescovitz OH 1989 Effect of naloxone on thegrowth hormone response to clonidine in normal women during the mid-luteal phase. Psychoneuroendocrinol 14:137–143
597. Tulandi T, Lal S, Guyda H 1987 Effect of estrogen on the growth hormoneresponse to the alpha-adrenergic agonist clonidine in women with meno-pausal flushing. J Clin Endocrinol Metab 65:6–10
786 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

598. Toivola PTK, Gale CC, Goodner CJ 1972 Central alpha-adrenergic regu-lation of growth hormone and insulin. Hormones 3:193–196
599. Chihara K, Kodama H, Kaji H, Kita T, Kashio Y, Okimura Y, Abe H, FujitaT 1985 Augmentation by propranolol of growth hormone releasing hormone(1–44) NH2-induced growth hormone releas in normal short and normalchildren. J Clin Endocrinol Metab 61:229–233
600. Mauras N, Blizzard RM, Thorner MO, Rogol AD 1987 Selective B1-adrenergic receptor-blockade with atenolol enhances basal and growthhormone releasing hormone mediated growth hormone release in man.Metabolism 36:369–372
601. Blackard W, Heidingsfelder S 1968 Adrenergic receptor control mechanismfor growth hormone secretion. J Clin Invest 47:1407–1414
602. Perkins SN, Evans WS, Thorner MO, Cronin MJ 1983 Beta-adrenergicstimulation of growth hormone release from perifused rat anterior pituitarycells. Neuroendocrinology 37:473–475
603. Samols E, Weir GC, Ranseur R, Day JA, Patel YC 1978 Modulation ofpancreatic somatostatin by adrenergic and cholinergic agonism and by hy-poglycemic sulfonamides. Metabolism 9:1219–1223
604. Richardson SB, Twente S 1990 Inhibition of hypothalamic somatostatinrelease by beta-adrenergic antagonist. Endocrinology 126:1043–1046
605. Krieg RJ, Perkins SN, Johnson JH 1988 Beta-adrenergic stimulation ofgrowth hormone (GH) release in vivo and subsequent inhibition of GHreleasing factor-induced GH secretion. Endocrinology 122:531–535
606. Kelijman M, Frohman LA 1989 Beta-adrenergic modulation of growthhormone (GH)-autofeedback on sleep-associated and pharmacologically in-duced GH secretion. J Clin Endocrinol Metab 69:1187–1193
607. Massara F, Camanni F 1972 Effect of various adrenergic receptor stimulatingand blocking agents on human growth hormone secretion. J Endocrinol54:195–206
608. Giustain A, Malerba M, Bresciani E, Desenzani P, Licini M, Zaltieri G,Grassi V 1995 Effect of two b2-agonist drugs, salbutamol and broxaterol, onthe growth hormone response to exercise in adult patients wtih asthmaticbronchitis. J Endocrinol Invest 18:847–852
609. Murakami Y, Kato Kabayama Y, Tojo K, Inoue T, Imura H 1986 Involve-ment of growth hormone releasing factor in GH secretion induced by se-rotoninergic mechanism in conscious rat. Endocrinology 119:1089–1092
610. Cataldi M, Magnan E, Guillaume V, Hery F, Dutour A, Rettori MC,Kamoun A, Oliver C 1994 Effect of tianeptine on the hypothalamic soma-totropic axis in the conscious sheep. Eur J Pharmacol 253:149–153
611. Mueller EE, Udeschini G, Secchi C, Zambotti F, Vicentini L, Panterai F,Cocola F, Mantegazza P 1976 Inhibitory role of the serotoninergic system inhypoglycemia-induced growth hormone release in the dog. Acta Endocrinol(Copenh) 82:71–78
612. Imura H, Nakai I, Yoshimi T 1973 Effect of 5-hydrotryptophan on growthhormone and ACTH release in man. J Clin Endocrinol Metab 39:1–5
613. Parati EA, Zanardi P, Cocci D, Caraceni T, Mueller EE 1980 Neuroendocrineeffect of quipazine in man in healthy state and in neurological disorders.J Neural Transm Gen Sect 47:273–297
614. Delitala G, Devilla L, Bionda S, Franca V 1977 Suppression of human GHsecretion by cyproheptadine. Metabolism 26:931–935
615. Casanueva FA, Villanueva LPA, Cabezas-Cerrato J 1984 Depending on thestimulus, central serotoninergic activation by fenfluramine blocks or doesnot alter growth hormone secretion in man. Neuroendocrinology 38:302–308
616. Biwens CH, Lebovitz HE, Feldman JM 1973 Inhibition of hypoglycemia-induced GH secretion by the serotonin antagonists cyproheptadine andmethysergide. N Engl J Med 289:236–238
617. Hoyer D, Clarke DE, Fozard JR 1994 International Union of Pharmacologyclassification of receptors for 5-hydroxytryptamine (serotonin). PharmacolRev 46:157–203
618. Mota M, Bento A, Penalva A, Pombo M, Dieguez C 1995 Role of theserotonin receptor subtype 5-HTD in basal and stimulated growth hormonesecretion. J Clin Endocrinol Metab 80:1973–1977
619. Peroutka SJ, McCarthy BG 1989 Sumatriptan (GR43157) interacts selec-tively with 5-HT1B and 5-HT1D binding sites. Eur J Pharmacol 163:133–136
620. Rolandi E, Franceschini R, Cataldi A, Barreca T 1992 Endocrine effects ofsumatriptan. Lancet 339:1365–1368
621. Netti C, Guidobono F, Olgiati VR, Sibilia V, Pecile A 1981 Histamineagonist and antagonist drugs: interference with CNS control of GH releasein rats. Horm Res 14:180–191
622. Netti C, Guidobono F, Olgiati VR, Sibilia V, Pagani F, Pecile A 1982Influence of brain histaminergic system on episodic growth hormone se-cretion in the rat. Neuroendocrinology 35:43–47
623. Netti C, Guidobono F, Olgiati VR, Sibilia V, Pagani F, Pecile A 1984 Failureof somatostatin antiserum to reverse histamine-induced inhibition of pul-satile growth hormone secretion. Horm Res 19:12–17
624. Duggan DE, Hooke KF, Maycock AL 1984 Inhibition of histamine synthesisin vitro and in vivo by 5-a-fluoromethylhistidine. Biochem Pharmacol 33:4003–4009
625. Grilli R, Sibilia V, Torsello A, Guidi M, Luoni M, Netti C, Mueller EE 1996Role of neuronal histaminergic system in the regulation of somatotropicfunction: comparison between the neonatal and adult rat. J Endocrinol151:195–201
626. Pontiroli AE, Viberti G, Viccari A, Pozza G 1976 Effect of antihistamineagents meclastine and dexchlorpheniramine on the response of humangrowth hormone to arginine infusion and insulin hypoglycemia. J ClinEndocrinol Metab 43:582–586
627. Zanaboni A, Galmozzi G, Marinoni S, Muciaccia WZ 1984 Inhibitory effectof cimetidine on L-dopa stimulated growth hormone release in man. ClinEndocrinol (Oxf) 21:535–540
628. Koulu M, Lamminstausta R, Kangas L, Dahlstrom S 1979 The effect ofmethysergide, pimozide, and sodium valproate on diazepam stimulatedgrowth hormone secretion in man. J Clin Endocrinol Metab 48:119–123
629. Abraham RR, Dornhorst A, Wynn V, Altaher AR, Campbell EA, BeckfordU, Watts SM, Nicholson SA, Gillham B, Thody A 1985 Corticotropin,cortisol, prolactin and growth hormone responses to insulin-induced hy-poglycaemia in normal subjects given sodium valproate. Clin Endocrinol(Oxf) 22:639–644
630. Cavagnini F, Invitti C, Di Landro A 1977 Effects of a GABA derivative,baclofen, on growth hormone and prolactin secretion in man. J Clin Endo-crinol Metab 45:579–584
631. Acs Z, Lonart G, Makara GB 1990 Role of hypothalamic factors (growth-hormone-releasing hormone and gamma-aminobutyric acid) in the regula-tion of growth hormone secretion in the neonatal and adult rat. Neuroen-docrinology 52:156–160
632. McCann SM, Vijayan E, Negro-Vilar A 1984 Gamma aminobutyric acid(GABA), a modulator of anterior pituitary hormone secretion by hypotha-lamic and pituitary action. Psychoneuroendocrinol 9:97–106
633. Van Cauter E, Plat L, Scharf MB, Leproult R, Cespedes S, L’Hermite-Baleriaux M, Copinschi G 1997 Simultaneous stimulation of slow-wavesleep and growth hormone secretion by gamma-hydroxybutyrate in normalyoung men. J Clin Invest 100:745–753
634. Van Cauter E, Caufriez A, Kerkhofs M, Van Onderbergern A, ThornerMO, Copinschi G 1992 Sleep, awakenings, and insulin-like growth factor-Imodulate the growth hormone (GH) secretory response to GH-releasinghormone. J Clin Endocrinol Metab 74:1451–1459
635. Nishiki M, Murakami Y, Sohmiya M, Tanaka J, Yamauchi K, KoshimuraK, Kato Y 1997 Effects of rat galanin and galanin message associated peptide(GMAP) on rat growth hormone secretion and stimulating effect of gamma-aminobutyric acid on galanin release from rat hypothalamus. Neurosci Lett226:199–202
636. Acs Z, Szabo B, Kapocs G, Makara GB 1987 Gamma-aminobutyric acidstimulates pituitary growth hormone secretion in the neonatal rat. A su-perfusion study. Endocrinology 120:1790–1798
637. Mergl Z, Acs Z, Makara GB 1995 Growth hormone secretion and activationof cyclic AMP by growth hormone releasing hormone and gamma-ami-nobutyric acid in the neonatal rat pituitary. Life Sci 56:579–585
638. Acs Z, Zsom L, Makara GB 1992 Possible mediation of GABA inducedgrowth hormone secretion by increased calcium-flux in neonatal pituitaries.Life Sci 50:273–279
639. Acs Z, Zsom L, Mergl Z, Makara GB 1993 Significance of chloride channelactivation in the gamma-aminobutyric acid induced growth hormone se-cretion in the neonatal rat pituitary. Life Sci 52:1733–1739
640. Spencer GS, Berry CJ, Bass JJ 1994 Neuroendocrine regulation of growthhormone secretion in sheep. VII. Effects of GABA. Regul Pept 52:181–186
641. Lucey JV, Butcher G, O’Flynn K, Clare AW, Dinan G 1994 The growthhormone response to baclofen in obsessive compulsive disorder: does theGABA-B receptor mediate obsessive anxiety? Pharmacopsychiatry 27:23–26
642. Gerra G, Caccavari R, Fontanesi B, Marcato A, Fertonani AG, Maestri D,Avanzini P, Lecchini R, Delsignore R, Mutti A 1994 Flumazenil effects ongrowth hormone response to gamma-hydroxybutyric acid. Int Clin Psycho-pharmacol 9:211–215
643. O’Flynn K, Dinan TG 1993 Baclofen-induced growth hormone release inmajor depression: relationship to dexamethasone suppression test result.Am J Psychiat 150:1728–1730
644. Monteleone P, Steardo L, Tanzillo C, Maj M 1990 Chronic antidepressantdrug treatment does not affect GH response to baclofen in depressed sub-jects. J Neural Transm Gen Sect 82:147–152
645. Volpi R, Gerra G, Vourna S, Vescovi PP, Maestri D, Chiodera P, Coiro V1992 Failure of the gamma-aminobutyric acid (GABA) derivative, baclofen,to stimulate growth hormone secretion in heroin addicts. Life Sci 51:247–251
646. Coiro V, Capretti L, Bianconi I, Castelli A, Cerri L, Roberti G, Marcato A,Volpi R, Chiodera P 1991 Reduction of baclofen-, but not sodium valproate-induced growth hormone release in type I diabetic men. Horm Metab Res23:600–604
647. Volpi R, Scalioni A, Marcato A, Caffarra P, Rossi G, Caffarri G, DelsignoreR, Chiodera P, Coiro V 1991 Failure of the gamma-aminobutyric acid(GABA) derivative, baclofen, to stimulate growth hormone secretion inParkinson’s disease. J Neural Transm Park Dis Dement Sect 3:259–264
648. Monteleone P, Maj M, Iovino M, Forziati D, Veltro F, Steardo L 1988Baclofen-induced growth hormone secretion is blunted in chronic schizo-phrenics: neuroendocrine evidence for a GABA disturbance in schizophre-nia. Psychiatry Res 26:1–9
649. Monteleone P, Maj M, Iovino M, Steardo L 1988 Evidence for a sex dif-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 787
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

ference in the basal growth hormone response to GABAergic stimulation inhumans. Acta Endocrinol (Copenh) 119:353–357
650. Monteleone P, Iovino M, Orio F, Steardo L 1987 Impaired growth hormoneresponse to sodium valproate in normal aging. Psychopharmacology 91:10–13
651. Orio F, Iovino M, Monteleone P, Agrusta M, Steardo L, Lombardi G 1988Pharmacological activation of the GABAergic system does not affect GH andPRL release in acromegaly. Horm Metab Res 20:701–704
652. Gale EF 1966 The bacterial amino acid decarboxylase. Adv Enzymol 6:1–32653. Baekkeskov S, Aanstoot HJ, Christgau S 1990 Identification of the 64K
autoantigen in insulin-dependent diabetes as the GABA-synthesizing en-zyme, glutamic acid decarboxylase. Nature 347:151–156
654. Giustina A, Wehrenberg WB 1994 Growth hormone neuroregulation indiabetes mellitus. Trends Endocrinol Metab 5:73–78
655. Giustina A, Bresciani E, Tassi C, Valentini U, Wehrenberg WB 1994 Effectof pyridostigmine on the growth hormone response to growth hormonereleasing hormone in lean and obese and type 2 diabetic patients. Metab-olism 43:893–898
656. Giustina A, Desenzani P, Perini P, Bazzigaluppi E, Bodini C, Bossoni S,Poiesi C, Wehrenberg WB, Bosi E 1997 Glutamate decarboxylase autoim-munity and growth hormone secretion in type 1 diabetes mellitus. Metab-olism 46:382–387
657. Barb CR, Campbell RM, Armstrong JD, Cox NM 1996 Aspartate andglutamate modulation of growth hormone secretion in the pig: possible siteof action. Domest Anim Endocrinol 13:81–90
658. Veneroni O, Cocilovo L, Mueller EE, Cocchi D 1990 Delay of puberty andimpairment of growth in female rats given a noncompetitive antagonist ofNMDA receptors. Life Sci 47:1253–1260
659. Mason GA, Bissette G, Nemeroff CB 1983 Effects of excitotoxic amino acidson pituitary hormone secretion in the rat. Brain Res 289:366–369
660. Lindstrom P, Ohlsson L 1992 Effect of N-methyl-D,L-aspartate on isolatedrat somatotrophs. Endocrinology 131:1903–1907
661. Niimi M, Sato M, Murao K, Takahara J, Kawanishi K 1994 Effect ofexcitatory amino acid receptor agonists on secretion of growth hormone asassessed by the reverse hemolytic plaque assay. Neuroendocrinology 60:173–178
662. Palmer RMJ, Ashton DS, Moncada S 1987 Vascular endothelial cells syn-thesize nitric oxide from L-arginine. Nature 333:664–666
663. Bredt DS, Hwang PM, Glatt CE, Lowenstein C, Reed RR, Snyder SH 1991Cloned and expressed nitric oxide synthetase structurally resembles cyto-chrome P-450 reductase. Nature 351:714–718
664. Pollock JS, Forstermann U, Mitchell JA, Warner TD, Schmidt HHHW,Nakane M, Murad F 1991 Purification and characterization of particulateendothelium-derived relaxing factor synthase from cultured and nativebovine aortic endothelial cells. Proc Natl Acad Sci USA 88:10480–10484
665. Rees DD, Palmer RMJ, Schulz RMS 1990 Characterization of three inhib-itors of endothelial nitric oxide synthase in vitro and in vivo. Br J Pharmacol101:746–752
666. Borger RH, Bode-Borger SM, Gerecke U 1994 Long-term administration ofL-arginine, L-NAME, and the exogenous NO donor molsidomine modulateurinary nitrate and cyclic GMP excretion in rats–correlation with vascularreactivity. Cardiovasc Res 28:494–499
667. Mayer B, John M, Heinzel B, Werner ER, Schultz G, Bohme E 1991 Brainnitric oxide synthase is a biopterin- and flavin-containing multi-functionaloxido-reductase. FEBS Lett 288:187–191
668. Korbonits M, Trainer PJ, Fanciulli G, Oliva O, Pala A, Dettori A, BesserGM, Delitala G, Grossman AB 1996 L-arginine is unlikely to exert neu-roendocrine effects in humans via the generation of nitric oxide. Eur JEndocrinol 135:543–547
669. Argetsinger LS, Carter-Su C 1996 Mechanism of signaling by growth hor-mone receptor. Physiol Rev 76:1089–1107
670. Boger RH, Skamira C, Bode-Boger SM, Brabant G, von zur Muhlen A,Frolich JC 1996 Nitric oxide may mediate the hemodynamic effects ofrecombinant growth hormone in patients with acquired growth hormonedeficiency. A double-blind, placebo-controlled study. J Clin Invest 98:2706–2713
671. Rettori V, Belova N, Yu WH, Gimeno M, McCann SM 1994 Role of nitricoxide in control of growth hormone release in the rat. Neuroimmunomodu-lation 1:195–200
672. Brunetti L 1994 Nitric oxide: a gas as a modulator of neuroendocrine se-cretions. Clin Ter 144:147–153
673. Tena-Sempere M, Pinilla L, Gonzalez D, Aguilar E 1996 Involvement ofendogenous nitric oxide in the control of pituitary responsiveness to dif-ferent elicitors of growth hormone release in prepubertal rats. Neuroendo-crinology 64:146–152
674. Kato M 1992 Involvement of nitric oxide in growth hormone (GH)-releasinghormone-induced GH secretion in rat pituitary cells. Endocrinology 131:2133–2138
675. Vankelecom H, Matthys P, Denef C 1997 Involvement of nitric oxide in theinterferon-gamma-induced inhibition of growth hormone and prolactin se-cretion in anterior pituitary cell cultures. Mol Cell Endocrinol 129:157–167
676. Volpi R, Chiodera P, Caffarri G, Vescovi PP, Capretti L, Gatti C, Coiro V
1996 Influence of nitric oxide on hypoglycemia–or angiotensin II-stimulatedACTH and GH secretion in normal men. Neuropeptide 30:528–532
677. Jansson JO, Albertsson-Wikland K, Eden S, Thorngren KG, Isaksson O1982 Circumstantial evidence for a role of the secretory pattern of growthhormone in control of body growth. Acta Endocrinol (Copenh) 99:24–30
678. Giustina A, Bussi AR, Jacobello C, Wehrenberg WB 1995 Effect of recom-binant human growth hormone on bone and intermediary metabolism inpatients receiving chronic glucocorticoid treatment with suppressed endog-enous GH reponse to GH-releasing hormone. J Clin Endocrinol Metab 80:122–129
679. Sacca L, Cittadini A, Fazio S 1989 Growth hormone and the heart. EndocrRev 15:555–573
680. Volterrani M, Desenzani P, Lorusso R, d’Aloia A, Manelli F, Giustina A1997 Haemodynamic effects of growth hormone in congestive heart failure.Lancet 349:1067–1068
681. Shibasaki T, Hotta M, Masuda A, Imaki T, Obara N, Demura H 1985Plasma responses to GHRH and insulin-induced hypoglycemia in man.J Clin Endocrinol Metab 60:1265–1267
682. Roth A, Cligg SM, Yallow CP, Berson SA 1963 Secretion of human growthhormone: physiological and experimental modification. Metabolism 12:557–559
683. Yalow RS, Goldsmith SJ, Berson SA 1969 Influence of physiologic fluctu-ations in plasma growth hormone on glucose tolerance. Diabetes 18:402–108
684. Davies RR, Turner S, Johnsston VG 1984 Oral glucose inhibits growthhormone secretion induced by human pancreatic growth hormone releasingfactor 1–44 in normal men. Clin Endocrinol (Oxf) 21:477–481
685. Sharp PS, Foley K, Chahal P, Kohner EM 1994 The effect of plasma glucoseon the growth hormone response to human pancreatic growth hormonereleasing factor in normal subjects. Clin Endocrinol (Oxf) 20:497–501
686. Masuda A, Shibasaki T, Nakahara M, Imaki T, Kijhijsawa T, Jibiki K,Demura H, Shizume H, Ling M 1985 The effect of glucose on growthhormone-releasing hormone-mediated GH secretion in men. Clin Endocri-nol Metab 60:523–526
687. Penalva A, Burgurea B, Casabiel X, Tresguerres JAF, Dieguez C, Cas-anueva FF 1989 Activation of cholinergic neurotransmission by pyridostig-mine reverses the inhibitory effect of hyperglycemia on growth hormone(GH) releasing hormone induced GH secretion in men: does acute hyper-glycemia act through hypothalamic relase of somatostatin? Neuroendocri-nology 49:551–554
688. Shibasaki T, Masuda A, Hotta M, Yamauchi N, Hizuka N, Takano K,Demura H, Shizume K 1989 Effects of ingestion of glucose on GH and TSHsecretion: evidence for stimulation of somatostatin release from the hypo-thalamus by acute hyperglycemia in normal man and its impairment inacromegalic patiens. Life Sci 44:431–438
689. Balzono S, Loche S, Murtas ML, Fanni T, Sica V, Pintor C, Martino E 1989Potentiation of cholinergic tone counteracts the suppressive effect of oralglucose administration on the GH response to GHRH in man. Horm MetabRes 21:52–53
690. Delitala G, Tomasi PA, Palermo M, Fresu P 1990 Interaction of glucose andpyridostigmine on the secretion of growth hormone (GH) induced by GH-releasing hormone (GHRH). Endocrinol Invest 13:653–656
691. Paget MB, Koppeschaar HPF, Edwards CA, Dieguez C, Scanlon MF 1987Additive effects of growth hormone releasing factor and insulin hypogly-cemia on growth hormone releasing hormone in men. Clin Endocrinol (Oxf)26:595–598
692. Caldwell G, Hart G, Kohner EM, Burrin JM 1989 Growth hormone re-leasing factor-induced growth hormone secretion from perifused rat ante-rior pituitary cells: lack of influence of glucose concentration, and normalresponses in pituitary cells from diabetic animals. J Clin Endocrinol Metab122:657–660
693. Valcavi R, Zini M, Davoli S, Portioli I 1992 The late growth hormone riseinduced by oral glucose is enhanced by cholinergic stimulation with pyri-dostigmine in normal subjects. Clin Endocrinol (Oxf) 37:360–364
694. Berelowitz M, Ting NC, Murray L 1989 Glucopenia-mediated release ofsomatostatin from incubated rat hypothalamus: monosaccharide specificityand role of glucocorticoid intermediates. Endocrinology 142:826–830
695. Baes M, Vale WW 1990 Characterization of the glucose-dependent releaseof growth hormone-releasing factor and somatostatin from superfused rathypothalami. Neuroendocrinology 51:202–207
696. Giustina A, Ruggeri G, Bodini C, Bossoni S, Grelli A, Schettino M, PozziA, Valentini U, Bolloli S, Albertini A, Giustina G, Insulin-like growthfactor 1 and growth hormone levels in patients with type I diabetes mellitus.Program of the International Symposium on Biotechnology of Growth Fac-tors, Milano, Italy, 1992, pp 122–126
697. Steiger RW, Rabe MB 1997 The effect of diabetes mellitus on endocrine andreproductive function. Proc Soc Exp Biol Med 214:1–11
698. Thrailkill K, Quattrin T, Baker L, Litton J, Dwigun K, Rearson M, Pop-penheimer M, Kotlovker D, Giltinan D, Gesundheit N, Martha Jr PM 1997Dual hormonal replacement therapy with insulin and recombinant humaninsulin-like growth factor (IGF)-I in insulin-dependent diabetes mellitus:effects on the growth hormone/IGF/IGF-binding protein system. Clin En-docrinol Metab 82:1181–1186
788 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

699. Mercado M, Bauman G 1995 Characteristics of the somatotropic axis ininsulin dependent diabetes mellitus. Arch Med Res 26:101–109
700. Bluet-Pajot MT, Durand D, Kordon C 1983 Influence of streptozotocin-induced diabaetes on growth hormone secretion in the rat. Neuroendocri-nology 36:307–309
701. Ndon JA, Giustina A, Wehrenberg WB 1992 Hypothalamic regulation ofimpaired growth hormone secretion in diabetic rats. Studies in streptozo-tocin-induced diabetic rats. Neuroendocrinology 55:501–505
702. Ndon JA, Giustina A, Wehrenberg WB 1992 Hypothalamic regulation ofimpaired growth hormone secretion in diabetic rats. Studies in spontane-ously diabetic BB Worcester rats. Neuroendocrinology 55:506–511
703. Patel YC, Wheatley T, Malaisse-Lagae F 1980 Elevated portal and periph-eral blood concentration of immunoreactive somatostatin in spontaneouslydiabetic (BBL) Wistar rats. Suppression with insulin. Diabetes 29:757–761
704. Plotksy PM 1991 Hypothalamic secretion of somatostatin and growth hor-mone-releasing factor into the hypophysial-portal circulation is reduced instreptozotocin diabetic male rats. Neuroendocrinology 53:433–438
705. Nieves-Rivera F, Kerrigan JR, Kreig Jr RJ, Egan J, Hwang L-J, TruumeesE, Veldhuis JD, Evans WS, Rogol AD 1993 Altered growth hormone (GH)secretion in vivo and in vitro in the diabetes-prone BB/Worcester rat. GrowthRegul 3:235–244
706. Ismail I, Lewis M, Peters JR, Scanlon MF 1995 Hypothalamic mediation ofreduced GH secretion in diabetic rats: evidence for reduced cholinergicinhibition of somatostatin release. J Neuroendocrinol 7:311–318
707. Martha PM, Clarke WL 1993 Alterations in growth hormone secretion andclearance in adolescent boys with insulin-dependent diabetes mellitus. J ClinEndocrinol Metab 77:638–643
708. Asplin CM, Faria ACS, Carlsen EC, Vaccaro VA, Barr RE, Iranmanesh A,Lee MM, Veldhuis JD, Evans WS 1989 Alterations in the pulsatile mode ofgrowth hormone release in men and women with insulin-dependent dia-betes mellitus. J Clin Endocrinol Metab 69:239–245
709. Hayford JT, Danney MM, Hendrix JA, Thompson RG 1980 Integratedconcentration of growth hormone in juvenile-onset diabetes. Diabetes 29:391–398
710. Arias P, Kerner W, De La Fuente A, Pfeiffer FF 1984 Abnormal growthhormone levels in insulin-dependent diabetic patients under continuoussubcutaneous insulin infusion and intensified conventional treatment. ActaEndocrinol (Copenh) 107:250–255
711. Krassowski J, Felber JP, Rogala H, Jeske W, Zgliczynski S 1988 Exagger-ated growth hormone response to growth hormone-releasing hormone intype I diabetes mellitus. Acta Endocrinol (Copenh) 117:225–229
712. Pietschmann P, Schemthaner G, Prskavec F, Gisinger C, Freyler H 1987 Noevidence for increased growth hormone responses to growth hormone re-leasing hormone in patients with diabetic retinopathy. Diabetes 36:159–162
713. Schaper NC, Tamsma JT, Sluiter WJ, Roelse H, Reitsma WD, DoorenbosH 1990 Growth hormone autoregulation in type I diabetes mellitus. ActaEndocrinol (Copenh) 122:32–39
714. Martina V, Tagliabue M, Maccario M, Bertagna S, Ghigo E, Massara F,Camanni F 1987 Pirenzepine blunts the nocturnal growth hormone releasein insulin dependent diabetes. Horm Metab Res 19:449–450
715. Pae MD, Koppeschaar HPF, Dieguez C, Gibbs JT, Hall R, Peters JR,Scanlon MF 1987 Cholinergic muscarinic receptor blockade with pirenz-epine abolishes slow wave sleep-related GH release in young patients withinsulin dependent diabetes-mellitus. Clin Endocrinol (Oxf) 26:355–359
716. Krassowski J, Szulc P, Makowska A, Dodziejewska M, Jeske W, Zglic-zynski S 1993 Short-term pirenzepine treatment is ineffective in suppressing24-h growth hormone secretion in type 1 diabetes mellitus. Diabetes Res ClinPract 19:211–216
717. Atiea JA, Creagh F, Page MD, Owens DR, Scanlon MF, Peters JR, 1989Early morning hyperglycemia in insulin-dependent diabetes mellitus: acuteand sustained effects of cholinergic blockade. J Clin Endocrinol Metab 69:390–395
718. Press M, Tamborlane WV, Thorner MO 1984 Pituitary response to growthhormone-releasing factor in diabetes. Diabetes 33:804–806
719. Salgado LR, Semer M, Nery M, Knoepfelmacher M, Lerario AC, Povoa G,Jana S, Villares SM, Wajchemberg BL, Liberman B, Nicolau W 1996 Effectof glycemic control on growth hormone and IGFBP-1 secretion in patientswith type 1 diabetes mellitus. Endocrinol Invest 19:433–440
720. Shishko PI, Dreval AV, Abugova IA, Zarjarny IU, Goncharov VC 1994Insulin-like growth factors and binding proteins in patients with recent-onset type 1 (insulin dependent) diabetes mellitus: influence of diabetescontrol and intraportal insulin infusion. Diabetes Res Clin Pract 25:1–12
721. Quatttrin T, Thraikill K, Baker L, Litton J, Dkigun K, Rearson M, Pop-penheirme M, Giltinan D, Kotlovker D, Gesundheit N, Martha P 1997Dual hormonal replacement with insulin and recombinant human insulin-like growth factor I in insulin dependent diabetes mellitus: effects of gly-cemic control, IGF-I levels and safety profile. Diabetes Care 20:374–380
722. Cheetham TD, Jones J, Taylor AM, Holly J, Mathews DR, Dunger DB 1993The effects of recombinant insulin-like growth factor I administration ongrowth hormone levels and insulin requirement in adolescents with type 1(insulin dependent) diabetes mellitus. Diabetologia 36:678–681
723. Bach MA, Chin E, Bondy CA 1994 The effects of subcutaneous insulin-like
growth factor-1 infusion in insulin-dependent diabetes mellitus. Clin En-docrinol Metab 79:1040–1045
724. Jacobs ML, Derkx F, Stijnen T 1997 Effect of long-acting somatostatinanalog (Somatulin) on renal hyperfiltration in patients wtih IDDM. DiabetesCare 20:632–636
725. Veldhuis JD, Johnson ML, Wilkowski MJ, Iranmanesh A, Bolton WK 1992Neuroendocrine alterations in the somatotropic axis in chronic renal failure.Acta Paediatr Scand 372:12–22
726. Johansson G, Sietnieks A, Janssens F, Proesmans W, Vanderschueren-Lodeweyckx M, Holmberg C, Sipila I, Broyer M, Rappaport R, Albertsson-Wikland K, Berg U, Jodal U, Rees L, Ridgen SPA, Preece MA 1990 Re-combinant human growth hormone treatment in short children with chronicrenal disease, before transplantation or with functioning renal transplants:an interim report on five European studies. Acta Paediatr Scand Suppl370:36–42
727. Hokken-Koelega AC, Stijnen T, de Muinck Keizer-Schrama SM, Wit JM,Wolff ED, de Jong MC, Donckerwolcke RA, Abbad NC, Bot A, Blum WF,Drop SLS 1991 Placebo-controlled, double-blind, cross-over trial of growthhormone treatment in prepubertal children with chronic renal failure. Lancet338:585–590
728. Blum WF, Ranke MB, Kietzmann K, Tonshoff B, Mehls O 1991 Growthhormone resistance and inhibition of somatomedin activity by excess ofinsulin-like growth factor binding protein in uraemia. Pediatr Nephrol5:539–544
729. Fine RN 1994 Pathophysiology of growth retardation in children withchronic renal failure. J Pediatr Endocrinol 7:79–83
730. Kjeldsen H, Hansen AP, Lundbaeck K 1975 Twenty-four hour serumgrowth hormone levels in maturity onset diabetics. Diabetes 24:977–982
731. Barnes AJ, Kohner EM, Johnston DG, Alberti KGMM 1985 Severe reti-nopathy and mild carbohydrate intolerance: possible role of insulin defi-ciency and elevated circulating growth hormone. Lancet 1:1465–1468
732. Owens DR, Volund A, Jones D, Shannon JD, Jones IR, Birthwell AJ, LuzioS, Williams S, Dolben J, Creagh FN 1988 Retinopathy in newly presentingnon-insulin dependent diabetic patients. Diabetes Res 9:59–65
733. Huhn W, Asplin C, Evans WS, Veldhuis JD, Iranmanesh A, ImpoverishedGH secretion marks the GH secretory derangement in type II diabetesmellitus. Program of the 77th Annual Meeting of The Endocrine Society,Washington DC, 1995 (Abstract P1–174)
734. Hansen AP 1973 Abnormal serum growth hormone response to exercise inmaturity-onset diabetics. Diabetes 22:619–628
735. Copelman PG, Mason AG, Noonan K, Monson JP 1984 Impaired growthhormone responses to growth hormone releasing factor in diabetic men. ClinEndocrinol (Oxf) 28:33–38
736. Richards NT, Wood SM, Christofides ND, Bhuttacharji SC, Bloom SR 1984Impaired growth hormone response to human pancreatic growth hormonereleasing factor (GRF(1–44)) in type 2 (non-insulin dependent) diabetes.Diabetologia 27:529–534
737. Jacobs ML, Nathoe HM, Blankestijn PJ, Stijnen T, Weber RF 1996 Growthhormone responses to growth hormone-releasing hormone and clonidine inpatients with type I diabetes and in normal controls: effect of age, body massindex and sex. Clin Endocrinol (Oxf) 44:547–553
738. Veldhuis JD 1995 Physiological regulation of GH secretion throughout thehuman lifetime. Current Medical Literature - Growth Hormone and GrowthFactors 10:59–68
739. Veldhuis JD 1996 Physiological regulation of the human growth hormone(GH)-insulin-like growth factor type I (IGF-I) axis: predominant impact ofage, obesity, gonadal function, and sleep. Sleep 29[Suppl]:S221–S224
740. Iranmanesh A, Veldhuis JD 1992 Clinical pathophysiology of the somato-tropic (GH) axis in adults. In: Veldhuis JD (ed) Endocrinology and Metab-olism Clinics of North America. WB Saunders, Inc, Philadelphia, vol 21:783–816
741. Kopelman PG, Mason AC, Noonan K 1988 Growth hormone response togrowth hormone releasing factor in diabetic men. Clin Endocrinol (Oxf)28:33–38
742. Williams T, Berelowitz M, Joffe SN, Thorner MO, Rivier J, Vale W,Frohman LA 1984 Impaired growth hormone responses to growth hormone-releasing factor in obesity. A pituitary defect reversed with weight reduc-tion. N Engl J Med 311:1403–1407
743. Sharp P, Toley K, Chahal P 1984 The effect of glucose on the growthhormone response to human pancreatic growth hormone releasing factor innormal subjects. Clin Endocrinol (Oxf) 20:497–501
744. Yamashita S, Melmed S 1986 Effects of insulin on rat anterior pituitary cells.Inhibition of growth hormone secretion and mRNA levels. Diabetes 35:440–447
745. Stanley BG, Kyrokouli SE, Lampbert S, Leibowitz SF 1986 NeuropeptideY chronically injected into the hypothalamus: a powerful neurochemicalinducer of hyperphagia and obesity. Peptides 7:1189–11192
746. Akabayashi A, Wahlestedt C, Alexander JT, Leibowitz SF 1994 Specificinhibition of endogenous neuropeptide Y synthesis in arcuate nucleus byantisense oligonucleotides suppresses feeding behavior and insulin secre-tion. Brain Res 21:55–61
747. Stephens TW, Bainski M, Bristow PK, Bue-Valleskey JM, Burgett SG,
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 789
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

Craft L, Hale J, Hoffmann J, Hsiung HM, Kriausiunas A 1995 The role ofneuropeptide Y in the antiobesity action of the obese gene product. Nature377:530–532
748. Schwartz MW, Baskin DG, Bukowski TR, Kuujper JL, Foster D, Lasser G,Prunkard DE, Porte D, Woods SC, Seeley RJ, Weigle DS 1996 Specificityof leptin action on elevated blood-glucose levels and hypothalamic neu-ropeptide-Y gene-expression in ob/ob mice. Diabetes 45:531–535
749. Weltman A, Weltman JY, Hartman ML, Abbott RA, Rogol AD, Evans WS,Veldhuis JD 1994 Relationship between age, percentage body fat, fitnessand 24 hour growth hormone release in healthy young adults: effects ofgender. J Clin Endocrinol Metab 78:543–548
750. Vahl N, Jorgensen JOL, Skjaerback C, Veldhuis JD, Orskov H, Chris-tiansen J 1997 Abdominal adiposity rather than age and sex predicts themass and patterned regularity of growth hormone secretion in mid-lifehealthy adults. Am J Physiol 272:E1108–E1116
751. Campfield LA, Smith FJ, Guisez Y, Devos R, Burn P 1995 Recombinantmouse ob protein: evidence for a peripheral signal linking adiposity andcentral neural networks. Science 269:546–549
752. Wideman L, Considine RV, Hartman ML, Veldhuis JD, Weltman A 1998Dietary macronutrient composition does not influence 24 hour serum leptinconcentrations. Program of the 80th Annual Meeting of The EndocrineSociety, New Orleans, LA, 1998 (Abstract P3–379)
753. Janssen YJH, Frolich M, Deurenberg P, Roelfsema F 1997 Serum leptinlevels during recombinant human GH therapy in adults with GH deficiency.Eur J Endocrinol 137:650–654
754. Schwartz MW, Seeley RJ, Campfield LA, Burn P, Baskin DG 1996 Iden-tification of targets on leptin action in rat hypothalamus. J Clin Invest98:1101–1106
755. Erickson JC, Hollopeter G, Palmiter RD 1996 Attenuation of the obesitysyndrome of ob/ob mice by the loss of neuropeptide Y. Science 274:1704–1707
756. Ahima RS, Dushay J, Flier SN, Prabakaran D, Flier JS 1997 Leptin accel-erates the onset of puberty in normal female mice. J Clin Invest 99:391–395
757. Ahima RS, Prabakaran D, Mantzoros C, Qu D, Lowell B, Maratos-Flier E,Flier JS 1996 Role of leptin in the neuroendocrine response to fasting. Nature382:250–252
758. Legradi G, Emerson CH, Ahima RS, Flier JS, Lechan RM 1997 Leptinprevents fasting-induced suppression of prothyrotropin-releasing hormonemessenger ribonucleic acid in neurons of the hypothalamic paraventricularnucleus. Endocrinology 138:250–252
759. Banks WA, Kastin AJ, Huang W, Jaspan JB, Maness LM 1996 Leptin entersthe brain by a saturable system independent of insulin. J Biol Chem 271:13939–13942
760. Vaisse C, Halaas JL, Horvath CM, Darnell JE, Stoffel M, Friedman JM 1996Leptin activation of Stat3 in the hypothalamus of wildtype and ob/ob micebut not db/db mice. Nat Genet 14:95–97
761. Mercer JG, Hoggard N, Williams LM 1996 Coexpression of leptin receptorand preproneuropeptide Y mRNA in arcuate nucleus of mouse hypothal-amus. J Neuroendocrinol 8:733–735
762. Attia N, Tamborlane WV, Heptulla R, Maggs D, Grozman A, Sherwin RS,Caprio S 1998 The metabolic syndrome and insulin-like growth factor Iregulation in adolescent obesity. J Clin Endocrinol Metab 83:1467–1471
763. Thompson JL, Butterfield GE, Gylfadottir UK, Yesavage J, Marcus R,Hintz RL, Pearman A, Hoffman AR 1998 Effects of human growth hormone,insulin-like growth factor I, and diet and exercise on body composition ofobese postmenopausal women. J Clin Endocrinol Metab 83:1477–1484
764. Licinio J, Negrao AB, Mantzoros C, Kaklamani V, Wong M-L, BongiornoPB, Mulla A, Cearnal L, Veldhuis JD, Flier JS, McCann SM, Gold PW 1998Synchronicity of frequently-sampled 24-hour concentrations of circulatingleptin, luteinizing hormone, and estradiol in healthy women. Proc Natl AcadSci USA 95:2541–2546
765. Licinio J, Negra AB, Kaklamani V, Wong M-L, Bongiorno PB, Negro PP,Mulla A, Veldhuis JD, Cearnal L, Flier JS, Gold PW 1998 Sex differencesin circulating human leptin pulse amplitude: clinical implications. J ClinEndocrinol Metab, in press
766. Berneis K, Vosmeer S, Keller U 1996 Effects of glucocorticoids and ofgrowth hormone on serum leptin concentrations in man. Eur J Endocrinol135:663–665
767. Imaki T, Shibasaki T, Shizume K, Masuda A, Hotta M, Kiyosawa Y, JibikiK, Demura H, Tsushima T, Ling N 1985 The effect of free fatty acids ongrowth hormone (GH)-releasing hormone-mediated GH secretion in man.J Clin Endocrinol Metab 60:290–293
768. Estienne MJ, Schillo KK, Green MA, Boling JA 1989 Free fatty acidssuppress growth hormone, but not luteinizing hormone, secretion in sheep.Endocrinology 125:85–91
769. Rosenbaum M, Fong Y, Hesse DG, Tracey KJ, Hintz RL, Lowry SF, GertnerJM 1989 Intravenous refeeding blocks growth hormone (GH)-provoked risesin serum free fatty acids and blunting of somatotroph response to GH-releasing hormone in normal men. J Clin Endocrinol Metab 60:310–316
770. Fuccella L, Golaniga G, Lovisolo P 1980 Inhibition of lipolysis by nicotinicacid and acipimox. Clin Pharmacol Ther 28:790–795
771. Cordido F, Peino R, Penalva A, Alvarez CV, Casanueva FF, Dieguez C 1996
Impaired growth hormone secretion in obese subjects is partially reversedby acipimox-mediated plasma free fatty acid depression. J Clin EndocrinolMetab 81:914–918
772. Pontiroli AE, Lanzi R, Monti LD, Pozza G 1990 Effect of acipimox, a lipidlowering drug, on growth hormone (GH) response to GH-releasing hor-mone in normal subjects. J Endocrinol Invest 13:539–542
773. Lee E, Nam S, Kim K, Lee H, Cho J, Nam M, Song Y, Lim S, Huh K 1995Acipimox potentiates growth hormone (GH) response to GH-releasing hor-mone with or without pyridostigmine by lowering serum free fatty acid innormal and obese subjects. J Clin Endocrinol Metab 80:2495–2501
774. Pontiroli AE, Lanzi R, Monti LD, Sandoli E, Pozza G 1991 Growth hormone(GH) autofeedback on GH response to GH-releasing hormone. Role of freefatty acids and somatostatin. J Clin Endocrinol Metab 72:492–495
775. Casanueva FF, Villanueva L, Dieguez C, Diaz Y, Cabranes JA, Szoke B,Scanlon MF, Schally AV, Fernandez-Cruz A 1987 Free fatty acids blockgrowth hormone (GH) release hormone-stimulated GH secretion in mandirectly at the pituitary. J Clin Endocrinol Metab 65:634–642
776. Kennedy J, Nicolson R, Wellby M 1994 The effect of oleic acid on thesecretion of thyrotrophin and growth hormone by cultured rat anteriorpituitary cells. J Endocrinol 143:557–564
777. Frystyk J, Vestbo E, Skjaerbaek C, Mogensen CE, Orskov H 1995 Freeinsulin-like growth factors in human obesity. Metabolism 44:37–44
778. Parker ML, Hammonds JM, Daughaday WH 1967 The arginine provocativetest: an aid in the diagnosis of hyposomatotropinism. J Clin EndocrinolMetab 27:1129–1136
779. Alba-Roth J, Albrecht Muller O, Schopohl J, Von Werder K 1988 Argininestimulates growth hormone secretion by suppressing endogenous soma-tostatin secretion. J Clin Endocrinol Metab 67:1186–1189
780. Schriock EA, Lustig RH, Rosenthal SM, Kaplan SL, Grumbach MM 1984Effect of growth hormone (GH)-releasing hormone (GRH) on plasma GH inrelation to magnitude and duration of GH deficiency in 26 children andadults with isolated GH deficiency or multiple pituitary hormone deficien-cies. Evidence for hypothalamic GHRH deficiency. J Clin Endocrinol Metab58:1043–1049
781. Ghigo E, Bellone J, Mazza E, Imperiale E, Procopio M, Valente F, Lala R,De Sanctis C, Camanni F 1990 Arginine potentiates the GHRH- but no thepyridostigmine-induced GH secretion in normal short children. Furtherevidence for a somatostatin-suppressing effect of arginine. Clin Endocrinol(Oxf) 32:763–766
782. Giustina A, Bossoni S, Buffoli MG, Ferrari C, Pozzi A, Scalvini T, Schet-tino M 1992 Effect of pretreatment with pyridostimine on the thyrotropinresponse to thyrotropin-releasing hormone in patients with Cushing’s dis-ease. Horm Metab Res 24:248–250
783. Giustina A, Bossoni S, Bodini C, Grelli A, Balestrieri GP, Pizzocolo G,Wehrenberg WB 1992 Arginine normalizes the growth hormone (GH) re-sponse to GH-releasing hormone in adult patients receiving chronic dailyimmunosuppressive glucocorticoid therapy. J Clin Endocrinol Metab 74:1301–1305
784. Fife SK, Brogan RS, Giustina A, Wehrenberg WB 1996 Immunocytochem-ical and molecular analysis of the effects of glucocorticoid-treatment on thehypothalamic-somatotropic axis in the rat. Neuroendocrinology 64:131–138
785. Mayo KE, Cerelli GM, Rosenfeld MG, Evans RM 1985 Characterization ofcDNA and genomic clones encoding the precursor to rat hypothalamicgrowth hormone-releasing factor. Nature 314:464–467
786. Lam KS, Srivastava G 1997 Gene expression of hypothalamic somatostatinand growth hormone-releasing hormone in dexamethasone-treated rats.Neuroendocrinology 66:2–8
787. Senaris RM, Lago F, Coya R, Pineda J, Dieguez C 1996 Regulation ofhypothalamic somatostatin, growth hormone-releasing hormone, andgrowth hormone receptor messenger ribonucleic acid by glucocorticoids.Endocrinology 137:5236–5241
788. Nakagawa K, Ishizuka T, Obara T, Matsubara M, Akikawa K 1987 Di-chotomic action of glucocorticoids on growth hormone secretion. Acta En-docrinol (Copenh) 116:165–171
789. Fernandez-Vazquez G, Cacicedo L, Lorenzo MG, Tolon R, Lopez J,Sanchez-Franco F 1995 Corticosterone modulates growth hormone-releas-ing factor and somatostatin in fetal rat hypothalamic cultures. Neuroendo-crinology 61:31–35
790. Nakagawa K, Ishizuka T, Chikara S, Yoshito I, Ichiji W 1992 Increasedhypothalamic somatostatin mRNA following dexamethasone administra-tion in rats. Acta Endocrinol (Copenh) 127:416–420
791. Lam KSL, Srivastava G, Tam S, Chung L, Chan S, Tang F, Chung SH 1993Dexamethasone decreases somatostatin mRNA levels in the periventricularnucleus of the rat hypothalamus. Neuroendocrinology 58:325–331
792. Papachristou DN, Liu J, Patel YC 1994 Glucocorticoids regulate somatosta-tin peptide and steady state messenger ribonucleic acid levels in normal rattissues and in a somatostatin-producing islet tumor cell line (1027B2). En-docrinology 134:2259–2266
793. Estupina C, Belmar J, Tapia-Arancibia L, Astier H, Arancidia S 1997 Rapidand opposite effects of dexamethasone on in vivo and in vitro hypothalamicsomatostatin release. Exp Brain Res 113:337–342
794. Xu Y, Berelowitz M, Bruno JF 1995 Dexamethasone regulates somatostatin
790 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

receptor subtype messenger ribonucleic acid expression in rat pituitaryGH4C1 cells. Endocrinology 136:5070–5075
795. Evans RM, Bimberg NC, Rosenfeld MG 1992 Glucocorticoid and thyroidhormones transcriptionally regulate growth hormone gene expression. ProcNatl Acad Sci USA 79:7659–7663
796. Slater VEP, Baxer JD, Eberhardt NL 1986 Evolution of the growth hormonegene family. Am Zool 26:939–949
797. Kohler PO, Bridson WE, Rayford PL 1968 Cortisol stimulation of growthhormone production by monkey adenohypophysis in tissue culture. Bio-chem Biophys Res Commun 33:834–840
798. Vale W, Vaughan J, Yamamoto G, Spiess J, Rivier J 1983 Effects of synthetichuman pancreatic (tumor) GH releasing factor and somatostatin, triiodo-thyronine and dexamethasone on GH secretion in vitro. Endocrinology 112:1553–1555
799. Bancroft FC, Levine L, Tashjian AH 1969 Control of growth hormoneproduction by a clonal strain of rat pitutiary cells: stimulation by hydro-cortisone. J Cell Biol 43:432–441
800. Oosterom R, Verleun J, Zuiderwijk J, Lamberts SWJ 1983 Growth hormonesecretion by cultured rat anterior pituitary cells: effects of culture conditionand dexamethasone. Endocrinology 113:735–741
801. Tamaki M, Sato M, Matsubara S, Wada Y, Takahara J 1996 Dexamethasoneincreases growth hormone (GH)-releasing hormone (GHR) receptor mRNAlevels in culture rat anterior pituitary cells. Neuroendocrinology 8:475–480
802. Koyto AI, Cuttler L 1997 Thyroid hormone and glucocorticoid regulation ofpituitary growth hormone-releasing hormone receptor gene expression. JEndocrinol 152:R13–R17
803. Heinrichs C, Yanovski JA, Roth AH, Yu YM, Domene HM, Yano K, CutlerGB, Baron J 1994 Dexamethasone increases growth hormone receptor mes-senger ribonucleic acid levels in liver and growth plate. Endocrinology135:1113–1118
804. King AP, Carter SC 1995 Dexamethasone-induced antagonism of growthhormone (GH) action by down-regulation of GH binding in 3T3–F442Afibroblasts. Endocrinology 136:4796–4803
805. Wehrenberg WB, Baird A, Ling N 1983 Potent interaction between glu-cocorticoids and growth hormone-releasing factor in vivo. Science 221:556–558
806. Thompson K, Coleman S, Hudmon A, Kemppainen RJ, Soyoola EO, SartinJL 1995 Effects of short-term cortisol infusion on growth hormone-releasinghormone stimulation of growth hormone release in sheep. Am J Vet Res56:1228–1231
807. Wehrenberg WB, Baird A, Klepper R, Mormede P, Ling N 1989 Interactionsbetween growth hormone-releasing hormone and glucocorticoids in malerats. Regul Pept 25:147–155
808. Mosier HD, Jansons RA 1985 Increase in pulsatile secretion in growthhormone during catch-up growth following glucocorticoid-induced growthinhibition. Proc Soc Exp Biol Med 178:457–461
809. Martin JB 1973 Studies on the mechanism of pentobarbital-induced GHrelease in the rat. Neuroendocrinology 13:339–350
810. Wehrenberg WB, Janowski BA, Piering AW, Culler F, Jones KL 1990Glucocorticoids: potent inhibitors and stimulators of growth hormone se-cretion. Endocrinology 126:3200–3203
811. Gaillard C, Wehrenberg WB 1996 Glucocorticoids and growth. Curr OpinEndocrinol Diabetes 3:227–232
812. Voltz DM, Piering AW, Magestro M, Giustina A, Wehrenberg WB 1995Effect of GHRP-6 and GHRH on GH secretion in rats following chronicglucocorticoid treatment. Life Sci 56:491–497
813. Giustina A, Voltz DM, Teik J, Wehrenberg WB 1995 Galanin counteractsthe inhibitory effects of glucocorticoids on growth hormone secretion in therat. Metabolism 44:224–227
814. Giustina A, Candrina R, Romanelli G 1988 Growth hormone deficiencyrapidly reversible during glucocorticoid replacement in a case of adreno-corticotrpin deficiency. Neth J Med 33:219–297
815. Giustina A, Romanelli G, Candrina R, Giustina G 1989 Growth hormonedeficiency in patients with idiopathic adrenocorticotropin deficiency re-solves during glucocorticoid replacement. J Clin Endocrinol Metab 68:120–124
816. Vincenzi P, Francia G, Dorizi R, Predrazzoli R, Lazzarin M 1992 IsolatedACTH deficiency and transient GH deficiency. Presentation of a case. Min-erva Endocrinol 17:127–131
817. Giustina A, Romanelli G, Grelli A, Pizzocolo G, Valentini U, Zuccato F1989 Effects of short-term glucocorticoid deprivation on growth hormone(GH) response to GH-releasing hormone in man. Horm Metab Res 21:693–694
818. Giustina A, Bresciani E, Bossoni S, Chiesa L, Misitano V, Wehrenberg WB,Veldhuis JD 1995 Reciprocal relationship between the level of circulatingcortisol and growth hormone secretion in response to growth hormone-releasing hormone in man: studies in patients with adrenal insufficiency.J Clin Endocrinol Metab 79:1266–1272
819. Veldhuis JD, Lizarralde G, Iranmanesh A 1992 Divergent effects of short-term glucocorticoid excess on the gonadotropic and somatotropic axes innormal men. J Clin Endocrinol Metab 74:96–102
820. del Balzo P, Salvatori R, Cappa M, Gertner JM 1990 Pyridostigmine does
not reverse dexamethasone-induced growth hormone inhibition. Clin En-docrinol (Oxf) 33:605–612
821. Casanueva FF, Burguera B, Muruais C, Dieguez C 1990 Acute adminis-tration of corticoids: a new and peculiar stimulus of growth hormone se-cretion in man. J Clin Endocrinol Metab 70:234–237
822. Dieguez C, Mallo F, Senaris R, Pineda J, Martul P, Leal-Cerro A, PomboM, Casanueva FF 1996 Role of glucocorticoids in the neuroregulation ofgrowth hormone secretion. J Pediatr Endocrinol Metab 3:255–260
823. Pralong FP, Miell JP, Corder R, Gaillard RC 1991 Dexamethasone treatmentin man induces changes in 24-hour growth hormone (GH) secretion profilewithout altering total GH released. J Clin Endocrinol Metab 73:1191–1196
824. Kaufmann S, Jones KL, Wehrenberg WB, Culler FL 1988 Inhibition byprednisone of growth hormone (GH) response to GH-releasing hormone innormal men. J Clin Endocrinol Metab 67:1258–1261
825. Demura R, Demura H, Nunokawa T, Baba H, Imura K 1972 Responses ofplasma ACTH, GH, LH, and 11-hydroxycorticosteroids to various stimuli inpatients with Cushing’s syndrome. J Clin Endocrinol Metab 34:852–859
826. Krieger DT, Glick SM 1974 Sleep EEG stages and plasma growth hormoneconcentration in states of endogenous hypercortisolemia or ACTH elevation.J Clin Endocrinol Metab 39:986–1000
827. Smals AE, Pieters GF, Smals AG, Benraad TJ, Kloppenborg PW 1986Human pancreatic growth hormone releasing hormone fails to stimulatehuman growth hormone both in Cushing’s disease and in Cushing’s syn-drome due to adrenocortical adenoma. Clin Endocrinol (Oxf) 24:401–407
828. Magiakow M, Mastorakos G, Gomez M, Rese S, Chrousos G 1994 Sup-pressed spontaneous and stimulated growth hormone secretion in patientswith Cushing’s disease before and after surgical cure. J Clin EndocrinolMetab 78:131–137
829. Whitehead HM, McKnight JA, Sheridan B, Kennedy AL, Hadden DR,Atkinson AB 1990 The growth hormone response to growth hormone re-leasing hormone in patients previously treated with bilateral adrenalectomyalone for Cushing’s disease. J Endocrinol Invest 13:217–220
830. Veldman RG, Frolich M, Veldhuis JD, Roelfsema F, The regularity of GHsecretion is altered in adult patients with ACTH-dependent Cushing’s dis-ease. Program of the 80th Annual Meeting of The Endocrine Society, NewOrleans, LA, 1998 (Abstract P2–458)
831. Giustina A, Bossoni S, Bodini C, Giustina G 1991 Pyridostigmine enhanceseven if it does not normalize the growth responses to growth hormone-releasing hormone in patients with Cushing’s disease. Horm Res 35:99–103
832. Leal-Cerro A, Pereira J, Garcia-Luna P, Astorga R, Coriddo F, CasanuevaF, Dieguez C 1990 Effect of enhancement of endogenous cholinergic tonewith pyridostigmine on growth hormone (GH) responses to GH-releasinghormone in patients with Cushing’s syndrome. Clin Endocrinol (Oxf) 33:291–295
833. Wajchenberg BL, Liberman B, Giannella ND, Morozimatov M, Semer M,Bracco LO, Salgado LR, Knoepfelmacher M, Borges MH, Pinto AC, KaterCE, Lengyel AM 1998 Growth hormone axis in Cushing’s syndrome. HormRes 45:99–107
834. LoCascio V, Francia G, Davi M, Olivieri M, Adami S, Cominacini L, RossiL 1992 Effects of deflazacort on growth hormone response to insulin toler-ance test. Horm Metab Res 24:333–335
835. Lima L, Arce V, Diaz M, Treguerres J, Devesa J 1993 Glucocorticoids mayinhibit growth hormone release by enhancing beta-adrenergic responsive-ness in hypothalamic somatostatin neurons. Clin Endocrinol Metab 76:439–444
836. Giustina A, Bussi AR, Licini M, Pizzocolo G, Schettino M, WehrenbergWB 1992 Acute effects of hydrocortisone on circulating growth hormonelevels in patients with acromegaly. Horm Res 37:212–216
837. Giustina A, Bussi AR, Doga M, Wehrenberg WB 1994 Effect of hydrocor-tisone on the growth hormone (GH) response to GH-releasaing hormone inacromegaly. Horm Res 41:33–37
838. Giustina A, Schettino M, Bossoni S, Bussi AR, Doga M, Licini M,Wehrenberg WB 1993 Arginine blocks the inhibitory effect of hydrocorti-sone on circulating growth hormone levels in patients with acromegaly.Metab Clin Exp 42:664–668
839. Ruzi F, Bontempelli AM, Jones DAJ, Pilotia A, Lombardi A, Giustina A,Preece MA 1998 Growth, insulin-like growth factor I (IGF-I), and IGF-binding proteins 1 and 3 in children with severe liver disease before and afterliver transplantation: a longitudinal and cross-sectional study. Pediatr Res43:478–483
840. Pennisi AJ, Costin G, Phillips LS 1979 Somatomedin and growth hormonestudies in pediatric renal allograft recipients who receive daily prednisone.Am J Dis Child 133:950–954
841. Borges MH, DiNinno FB, Lengyel AM 1997 Different effects of growthhormone releasing peptide (GHRP-6) and GH-releasing hormone on GHreleas in endogenous and exogenous hypercortisolism. Clin Endocrinol(Oxf) 46:713
842. Gertz B, Shiberras B, Yogendran L, Christie K, Bador K, Crupa D, WitreichJ, James I 1994 L-692,429, a nonpeptide growth hormone (GH) secretagogue,reverses glucocorticoid suppressoin of GH secretion. Clin Endocrinol Metab79:745–749
843. Bennet WM, Haymond MW 1992 Growth hormone and lean tissue catab-
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 791
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

olism during long-term glucocorticoid treatment. Clin Endocrinol (Oxf)36:161–164
844. Touati G, Prieur AM, Ruiz JC, Noel M, Czernichow P 1998 Beneficial effectsof one-year growth hormone administration to children with juvenilechronic arthritis on chronic steroid therapy. I. Effects on growth velocity andbody composition. J Clin Endocrinol Metab 83:403–409
845. Bernei K, Ninnis R, Girard J, Frey BM, Keller U 1997 Effects of insulin-likegrowth factor 1 combined with growth hormone on glucocorticoid inducedwhole-body protein catabolism in men. Clin Endocrinol Metab 82:2528–2534
846. Veldhuis JD 1998 Male hypothalamic-pituitary-gonadal axis. In: Yen SSC,Jaffe RB, Barbieri RL (eds) Reproductive Endocrinology. W.B. Saunders Co.,Philadelphia, in press
847. Veldhuis JD, Iranmanesh A, Rogol AD, Urban RJ 1995 Regulatory actionsof testosterone on pulsatile growth hormone secretion in the human: studiesusing deconvolution analysis. In: Adashi EY, Thorner MO (eds) Somato-tropic Axis and the Reproductive Process in Health and Disease. Springer-Verlag, New York, pp 40–57
848. Mauras N, Rogol AD, Haymond MW, Veldhuis JD 1996 Sex steroids,growth hormone, IGF-I: neuroendocrine and metabolic regulation in pu-berty. Horm Res 45:74–80
849. Maiter D, Koenig JI, Kaplan LM 1991 Sexually dimorphic expression of thegrowth hormone-releasing hormone gene is not mediated by circulatinggonadal hormones in the adult rat. Endocrinology 128:1709–1716
850. Mizobuchi M, Frohman MA, Downs TR, Frohman LA 1991 Tissue-specifictranscription initiation and effects of growth hormone (GH) deficiency onthe regulation of mouse and rat GH-releasing hormone gene in hypothal-amus and placenta. Mol Endocrinol 5:476–484
851. Argente J, Chowen BJ, Steiner RA, Clifton DK 1990 Somatostatin mes-senger RNA in hypothalamic neurons is increased by testosterone throughactivation of androgen receptors and not by aromatization to estradiol.Neuroendocrinology 52:342–349
852. Tannenbaum GS, Epelbaum J, Videau C, Dubuis JM 1996 Sex-relatedalterations in hypothalamic growth hormone-releasing hormone mRNA-butnot somatostatin mRNA-expressing cells in genetically obese Zucker rats.Neuroendocrinology 64:186–193
853. De Gennaro V, Zoli M, Cocchi D 1989 Reduced growth hormone releasingfactor (GHRH)-like immunoreactivity and GHRH gene expression in hy-pothalamus of aged rats. Peptides 10:705–709
854. Wehrenberg WB, Corder R, Keher P, Muller AF, Gaillard RC 1987 The roleof growth hormone-releasing factor and growth hormone inhibiting hor-mone in young, neonatal and pubertal somatic growth. Endocrinol Jpn34:1–8
855. Zeitler P, Argente J, Chowen-Breed JA, Clifton DK, Steiner RA 1990Growth hormone-releasing hormone messenger ribonucleic acid in the hy-pothalamus of the adult male rat is increased by testosterone. Endocrinology127:1362–1368
856. Millard WJ, Politch JA, Martin JB, Fox TO 1986 Growth hormone-secretorypatterns in androgen-resistant (testicular feminized) rats. Endocrinology119:2655–2660
857. Ohlsson L, Isaksson O, Jansson JO 1987 Endogenous testosterone enhancesgrowth hormone (GH)-releasing factor-induced GH secretion in vitro. J En-docrinol 113:249–253
858. Wehrenberg WB, Baird A, Ying SY, Ling N 1985 The effect of testosteroneand estrogen on the pituitary growth hormone response to growth hormonereleasing factor. Biol Reprod 32:369–375
859. Senaris RM, Lago F, Lewis MD, Dominguez F, Scanlon MF, Dieguez C1992 Differential effects of in vivo estrogen administration on hypothalamicgrowth hormone releasing hormone and somatostatin gene expression.Neurosci Lett 141:123–126
860. Shirasu K, Stumpf WE, Sar M 1995 Evidence for direct action of estradiolon growth hormone-releasing factor (GRF) in rat hypothalamus: localizationof [3H]estradiol in GRF neurons. Endocrinology 127:344–349
861. McCarthy GF, Beaudet A, Tannenbaum GS 1992 Colocalization of soma-tostatin receptors and growth hormone-releasing factor immunoreactivity inneurons of the rat arcuate nucleus. Neuroendocrinology 56:18–24
862. Ge F, Tsagarakis S, Rees LH, Besser GM, Gorman A 1989 Relationshipbetween growth hormone-releasing hormone and somatostatin in the rat:effects of age and sex on content and in vitro release from hypothalamicexplants. J Endocrinol 123:53–58
863. Painson JC, Tannenbaum GS 1991 Sexual dimorphism of somatostatin andgrowth hormone-releasing factor signaling in the control of pulsatile growthhormone secretion in the rat. Endocrinology 128:2858–2866
864. Turner JP, Tannenbaum GS 1995 In vivo evidence of a positive role forsomatostatin to optimize pulsatile growth hormone secretion. Am J Physiol269:E683–E690
865. Clark RG, Carlsson LMS, Rafferty B, Robinson ICAF 1988 The reboundrelease of growth hormone (GH) following somatostatin infusion in ratsinvolves hypothalamic GH-releasing factor release. J Endocrinol 119:397–404
866. Waxman DJ, Ram PA, Park SH, Choi HK 1995 Intermittent plasma growthhormone triggers tyrosine phosphorylation and nuclear translocation of a
liver-expressed, STAT 5-related DNA binding protein. J Biol Chem 13262–13270
867. Conley LK, Aubert ML, Giustina A, Wehrenberg WB 1996 The role ofgrowth hormone releasing hormone (GHRH) and growth hormone (GH) inthe onset of puberty and during glucocorticoid-altered growth. In: Bercu BB,Walker RF (eds) Growth Hormone Secretagogues, Springer-Verlag, NewYork, pp 185–209
868. Werner H, Koch Y, Baldino F, Gozes I 1988 Steroid regulation of soma-tostatin mRNA in the rat hypothalamus. J Biol Chem 263:7666–7671
869. Zorrilla R, Simard J, Rheaume E, Labrie F, Pelletier G 1990 Multihormonalcontrol of pre-pro-somatostatin mRNA levels in the periventricular nucleusof the male and female rat hypothalamus. Neuroendocrinology 52:527–536
870. Gabriel SM, Roncancio JR, Ruiz NS 1992 Growth hormone pulsatility andthe endocrine milieu during sexual maturation in male and female rats.Neuroendocrinology 56:619–628
871. Chowen-Breed JA, Steiner RA, Clifton DK 1989 Sexual dimorphism andtestosterone-dependent regulation of somatostatin gene expression in theperiventricular nucleus of the rat brain. Endocrinology 125:357–362
872. Hasegawa O, Sugihara H, Minami S, Wakabayashi I 1992 Masculinizationof growth hormone (GH) secretory pattern by dihydrotestosterone is asso-ciated with augmentation of hypothalamic somatostatin and GH-releasinghormone mRNA levels in ovariectomized adult rats. Peptides 13:475–481
873. Goodman R, Rehfuss R, Verhave M, Ventimiglia R, Low M 1990 Soma-tostatin gene regulation — an overview. Metabolism 39:2–5
874. Huang X, Harlan RE 1994 Androgen receptor immunoreactivity in soma-tostatin neurons in the periventricular nucleus but not in the bed nucleusstria terminalis in male rats. Brain Res 652:291–296
875. Herbison AE 1995 Sexually dimorphic expression of androgen receptorimmunoreactivity by somatostatin neurones in rat hypothalamic periven-tricular nucleus and bed nucleus of the stria terminalis. J Neuroendocrinol7:543–553
876. Herbison AE, Augood SJ 1994 Expression of GABA-A receptor alpha 2subunit mRNA by periventricular somatostatin neurones in the rat hypo-thalamus. Neurosci Lett 173:9–13
877. Gross DS 1980 Role of somatostatin in the modulation of hypophysialgrowth hormone production by gonadal steroids. Am J Anat 158:507–519
878. Critchlow V, Dyke A, Kaler LW 1986 Release of growth hormone, prolactinand somatostatin during perifusion of anterior pituitary and preoptic-med-ical basal hypothalamus from male and female rats. Brain Res 398:347–353
879. Maiter DM, Gabriel SM, Koenig JI, Russell WE, Martin JB 1990 Sexualdifferentiation of growth hormone feedback effects on hypothalamic growthhormone-releasing hormone and somatostatin. Neuroendocrinology51:174–180
880. Wehrenberg WB, Ling N, Bohlen P, Esch F, Brazeau P, Guillemin R 1982Physiological roles of somatocrinin and somatostatin in the regulation ofgrowth hormone secretion. Biochem Biophys Res Commun 109:562–567
881. Clark RG, Robinson ICAF 1985 Growth hormone responses to multipleinjections of a fragment of human growth hormone-releasing factor in con-scious male and female rats. J Endocrinol 106:281–289
882. Wehrenberg WB 1986 Continuous infusion of growth-hormone-releasingfactor: effects on pulsatile growth hormone secretion in normal rats. Neu-roendocrinology 43:391–396
883. Painson J-C, Thorner MO, Krieg RJ, Tannenbaum GS 1992 Short term adultexposure to estradiol feminizes the male pattern of spontaneous and growthhormone-releasing factor-stimulated growth hormone secretion in the rat.Endocrinology 130:511–519
884. Tannenbaum GS, Gurd W, Lapointe M, Pollak M 1992 Tamoxifen atten-uates pulsatile growth hormone secretion: mediation in part by somatosta-tin. Endocrinology 130:3395–3401
885. Ahmad I, Finkelstein JA, Downs TR, Frohman LA 1993 Obesity-associateddecrease in growth hormone-releasing hormone gene expression: a mech-anisms for reduced growth hormone mRNA levels in genetically obeseZucker Rats. Neuroendocrinology 58:332–337
886. Nogami H, Takeuchi T, Suzuki K, Okuma S, Ishikawa H 1989 Studies onprolactin and growth hormone gene expression in the pituitary gland ofspontaneous dwarf rat. Endocrinology 125:964–970
887. Morel G, Dihl F, Gossard F 1989 Ultrastructural distribution of growthhormone (GH) mRNA and GH intron I sequences in rat pituitary gland:effects of GH-releasing factor and somatostatin. Mol Cell Endocrinol 65:81–90
888. Birge CA, Peake GT, Mariz IK, Daughaday WH 1967 Radioimmunoas-sayble growth hormone in the rat pituitary gland: effects of age, sex andhormonal state. Endocrinology 81:195–204
889. Silverman BL, Kaplan SL, Grumbach MM, Miller WL 1988 Hormonalregulation of growth hormone secretion and messenger ribonucleic acidaccumulation in cultured bovine pituitary cells. Endocrinology 122:1236–1241
890. Bethea CL 1991 Estrogen action on growth hormone in pituitary cell culturesfrom adult and juvenile macaques. Endocrinology 129:2110–2118
891. Jin L, Song J, Lloyd RV 1989 Estrogen stimulates both prolactin and growthhormone mRNa expression in the MtT/F4 transplantable pituitary tumor.Proc Soc Exp Biol Med 192:225–229
792 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

892. Goth MI, Lyons Jr CE, Ellwood MR, Barrett JR, Thorner MO 1996 Chronicestrogen treatment in male rats reveals mammosomatotropes and allowsinhibition of prolactin secretion by somatostatin. Endocrinology 137:274–280
893. DiPippo WA, Powers CA 1991 Estrogen induction of growth hormone inthe thyroidectomized rat. Endocrinology 129:1696–1700
894. Hertz P, Silbermann M, Even L, Hochberg Z 1989 Effects of sex steroids onthe response of cultured rat pituitary cells to growth hormone-releasinghormone and somatostatin. Endocrinology 125:581–585
895. Batson JM, Krieg Jr RJ, Martha Jr PM, Evans WS 1989 Growth hormone(GH) response to GH-releasing hormone by perifused pituitary cells frommale, female, and testicular feminized rats. Endocrinology 124:444–448
896. Evans WS, Krieg RJ, Limber ER, Kaiser DL, Thorner MO 1985 Effects ofin vivo gonadal hormone environment on in vitro hGRF-40-stimulated GHrelease. Am J Physiol 249:E276–E280
897. Hoeffler JP, Frawley LS 1986 Capacity of individual somatotropes to releasegrowth hormone varies according to sex: analysis by reverse hemolyticplaque assay. Endocrinology 119:1037–1042
898. Ho KY, Leong DA, Sinha YN, Johnson ML, Evans WS, Thorner MO 1986Sex-related differences in GH secretion in rat using reverse hemolytic plaqueassay. Am J Physiol 250:E650–E654
899. Conley LK, Giustina A, Imbimbo BP, Stagg LC, Deghenghi R, Wehren-berg WB 1994 Hexarelin™[His-d-2-methyl Trp-Ala-Trp-d-Phe-Lys-NH2], anew GH-releasing peptide, is biologically active in male and female rats.Endocrine 2:691–695
900. Simard J, Hubert JF, Hosseinzadeh T, Labrie F 1986 Stimulation of growthhormone release and synthesis by estrogens in rat anterior pituitary cells inculture. Endocrinology 119:2004–2011
901. Fukata J, Martin JB 1986 Influence of sex steroid hormones on rat growthhormone-releasing factor and somatostatin in dispersed pituitary cells. En-docrinology 119:2256–2261
902. Kerrigan JR, Martha Jr PM, Krieg Jr RJ, Rogol AD, Evans WS 1989 So-matostatin inhibition of growth hormone secretion by somatotropes frommale, female and androgen receptor-deficient rats: evidence for differingsensitivities. Endocrinology 125:3078–3083
903. Martha Jr PM, Rogol AD, Veldhuis JD, Kerrigan JR, Goodman DW,Blizzard RM 1989 Alterations in the pulsatile properties of circulatinggrowth hormone concentrations during puberty in boys. J Clin EndocrinolMetab 69:563–570
904. Kineman RD, Henricks DM, Faught WJ, Frawley LS 1991 Fluctuations inthe proportions of growth hormone- and prolactin-secreting cells during thebovine estrous cycle. Endocrinology 129:1221–1225
905. Plozek CA, Trenkel A 1991 Growth hormone parameters in intact andcastrated male and female cattle. Domest Anim Endocrinol 8:63–72
906. Landefeld TD, Suttie JM 1989 Changes in messenger ribonucleic acid con-centrations and plasma levels of growth hormone during the ovine estrouscycle and in response to exogenous estradiol. Endocrinology 125:1474–1478
907. Copeland KC, Johnson DM, Kuehl TJ, Castracane VD 1984 Estrogen stim-ulates growth hormone and somatomedin-C in castrate and intact femalebaboons. J Clin Endocrinol Metab 58:698–703
908. Domene HM, Marin G, Sztein J, Yum YM, Baron J, Cassorla FG 1994Estradiol inhibits growth hormone receptor gene expression in rabbit liver.Mol Cell Endocrinol 103:81–87
909. Carmignac DF, Gabrielsson BG, Robinson IC 1993 Growth hormone bind-ing protein in the rat: effects of gonadal steroids. Endocrinology 133:2445–2452
910. Ho KKY, O’Sullivan AJ, Weissberger AJ, Kelly JJ 1996 Sex steroid regu-lation of growth hormone secretion and action. Horm Res 45:67–73
911. Ip TP, Hoffman DM, O’Sullivan AJ, Leung KC, Ho KK 1995 Do androgensregulate growth hormone-binding protein in adult man? J Clin EndocrinolMetab 80:1278–1282
912. Baumbach WR, Horner DL, Logan JS 1989 The growth hormone-bindingprotein in rat serum is an alternatively spliced form of the rat growthhormone receptor. Genes Dev 3:1199–1205
913. Lobie PE, Garcia-Aragon J, Wang BS, Baumbach WR, Waters MJ 1992Cellular localization of the growth hormone binding protein in the rat.Endocrinology 130:3057–3065
914. Ahlgren R, Norstedt G, Baumbach WR, Mode A 1995 Hormonal regulationof the female enriched GH receptor/binding protein mRNA in rat liver. MolCell Endocrinol 113:11–17
915. Baumbach WR, Bingham B 1995 One class of growth hormone (GH) re-ceptor and binding protein messenger ribonucleic acid in rat liver, GHR1,is sexually dimorphic and regulated by GH. Endocrinology 136:749–760
916. Ilkbahar YN, Wu K, Thordarson G, Talamantes F 1995 Expression anddistribution of messenger ribonucleic acids for growth hormone (GH) re-ceptor and GH-binding protein in mice during pregnancy. Endocrinology136:386–392
917. Bingham B, Oldham ER, Baumbach WR 1994 Regulation of growth hor-mone receptor and binding protein expression in domestic species. Proc SocExp Biol Med 206:195–199
918. Sotelo AI, Dominici FP, Engbers C, Bartke A, Talamantes F, Turyn D 1995
Growth hormone-binding protein in normal mice and in transgenic miceexpressing bovine growth gene. Am J Physiol 268:E745–E751
919. Chang TK, Chan MM, Holsmer SL, Bandiera SM, Bellward GD 1996Impact of tamoxifen on peripubertal androgen imprinting of rat hepaticcytochrome P450 2C11, cytochrome P450 3A2, and steroid 5 alpha-reductase.Biochem Pharmacol 51:357–368
920. Waxman DJ, Pampori NA, Ram PA, Agrawal AK, Shapiro BH 1991 In-terpulse interval in circulating growth hormone patterns regulates sexuallydimorphic expression of hepatic cytochrome P450. Proc Natl Acad Sci USA88:6868–6872
921. Waxman DJ, Ram PA, Pampori NA, Shapiro BH 1995 Growth hormoneregulation of male-specific rat liver P450 s 2A2 and 3A2. Mol Pharmacol48:790–797
922. Legraverend C, Mode A, Westin S, Strom A, Eguchi H, Zaphiropoulos PG,Gustafsson J-A 1992 Transcriptional regulation of rat P-450 2C gene sub-family members by the sexually dimorphic pattern of growth hormonesecretion. Mol Endocrinol 6:259–266
923. Fryburg DA, Weltman A, Jahn LA, Weltman JY, Samolijik E, Veldhuis JD1997 Short-term modulation of the androgen milieu alters pulsatile but notexercise or GHRH-stimulated GH secretion in healthy men. J Clin Endo-crinol Metab 82:3710–3719
924. van der Werff ten Bosch JJ, Bot A 1990 Growth of males with idiopathichypopituitarism without growth hormone treatment. Clin Endocrinol (Oxf)32:707–717
925. Link K, Blizzard RM, Evans WS, Kaiser DL, Parker MW, Rogol AD 1986The effect of androgens on the pulsatile release and the twenty-four hourmean concentrations of growth hormone in peripubertal males. J Clin En-docrinol Metab 62:159–165
926. Martha Jr PM, Goorman KM, Blizzard RM, Rogol AD, Veldhuis JD 1992Endogenous growth hormone secretion and clearance rates in normal boysas determined by deconvolution analysis: relationship to age, pubertal statusand body mass. J Clin Endocrinol Metab 74:336–344
927. Ulloa-Aguirre A, Blizzard RM, Garcia-Rubi E, Rogol AD, Link K, ChristieCM, Johnson ML, Veldhuis JD 1990 Testosterone and oxandrolone, a non-aromatizable androgen, specifically amplify the mass and rate of growthhormone (GH) secreted per burst without altering GH secretory burst du-ration or frequency or the GH half-life. J Clin Endocrinol Metab 71:846–854
928. Giustina A, Scalvini T, Tassi C, Desenzani P, Poiesi C, Wehenberg W,Rogol A, Veldhuis JD 1997 Maturation of the regulation of growth hormonesecretion in young males with hypogonadotropic hypogonadism pharma-cologically exposed to progressive increments in serum testosterone. J ClinEndocrinol Metab 82:1210–1219
929. Iranmanesh A, Lizarralde G, Veldhuis JD 1991 Age and relative adiposityare specific negative determinants of the frequency and amplitude of growthhormone (GH) secretory bursts and the half-life of endogenous GH inhealthy men. J Clin Endocrinol Metab 73:1081–1088
930. Weissberger AJ, Ho KK 1993 Activation of the somatotropic axis by tes-tosterone in adult males: evidence for the role of aromatization. J ClinEndocrinol Metab 76:1407–1412
931. Metzger DL, Kerrigan JR 1993 Androgen receptor blockade with flutamideenhances growth hormone secretion in late pubertal males: evidence forindependent actions of estrogen and androgen. J Clin Endocrinol Metab76:1147–1152
932. Metzger DL, Kerrigan JR 1994 Estrogen receptor blockade with tamoxifendiminishes growth hormone secretion in boys: evidence for a stimulatoryrole of endogenous estrogens during male adolescence. J Clin EndocrinolMetab 79:513–518
933. Wennink JM, Delemarre-van de Waal HA, Schoemaker R, Blaauw G, vanden Brakern C, Schoemaker J 1991 Growth hormone secretion patterns inrelation to LH and estradiol secretion throughout normal female puberty.Acta Endocrinol (Copenh) 124:129–135
934. Mauras N, Rogol AD, Veldhuis JD 1989 Specific, time-dependent actionsof low-dose estradiol administration on the episodic release of GH, FSH andLH in prepubertal girls with Turner’s syndrome. J Clin Endocrinol Metab69:1053–1058
935. Frantz AG, Rabkin MT 1965 Effects of estrogen and sex difference onsecretion of human growth hormone. J Clin Endocrinol Metab 25:1470–1480
936. Unger R, Eisentraut A, Madison L Siperstein M 1965 Fasting levels ofgrowth hormone in men and women. Nature 205:804–807
937. Dawson-Hughes B, Stern D, Goldman J, Reichlin S 1986 Regulation ofgrowth hormone and somatomedin-C secretion in postmenopausal women:effect of physiological estrogen replacement therapy. J Clin EndocrinolMetab 63:424–432
938. Dursma SA, Bijlsma JW, Van Paassen HC, van Buul-Offers SC, Skottner-Lundin A 1984 Changes in serum somatomedin and growth hormone con-centrations after 3 weeks oestrogen substitution in post-menopausal women;a pilot study. Acta Endocrinol (Copenh) 106:527–531
939. Faria ACS, Bekenstein LW, Booth Jr RA, Vaccaro VA, Asplin CM,Veldhuis JD, Thorner MO, Evans WS 1992 Pulsatile growth hormonerelease in normal women during the menstrual cycle. Clin Endocrinol (Oxf)36:591–596
940. Wilson EE, Word RA, Byrd W, Carr BR 1991 Effect of superovulation with
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 793
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

human menopausal gonadotropins on growth hormone levels in women.J Clin Endocrinol Metab 73:511–555
941. Friedman AJ, Rein MS, Pandian MR, Barbieri RL 1990 Fasting serumgrowth hormone and insulin-like growth factor-I and -II concentrations inwomen with leiomyomata uteri treated with leuprolide acetate or placebo.Fertil Steril 53:250–253
942. Word RA, Odom MJ, Byrd W, Carr BR 1990 The effect of gonadotropin-releasing hormone agonists on growth hormone secretion in adult pre-menopausal women. Fertil Steril 54:73–78
943. Harris DA, Van Vliet G, Egli CA, Grumbach MM, Kaplan SL, Styne DM,Vainsel M 1985 Somatomedin-C in normal puberty and in true precociouspuberty before and after treatment with potent luteinizing hormone-releas-ing hormone agonist. J Clin Endocrinol Metab 61:152–158
944. Ovesen P, Vahl N, Fisker S, Veldhuis JD, Christiansen JS, Jorgensen JOL1998 Increased pulsatile, but not basal, growth hormone secretion rates andplasma insulin-like growth factor I levels during the preovulatory intervalin normal women. J Clin Endocrinol Metab 83:1662–1667
945. Weissberger AJ, Ho KKY, Lazarus L 1991 Contrasting effects of oral andtransdermal routes of estrogen replacement therapy on 24-hour growthhormone (GH) secretion, insulin-like growth factor I, and GH-binding pro-tein in postmenopausal women. J Clin Endocrinol Metab 72:374–381
946. Kelly JJ, Rajkovic IA, O’Sullivan AJ, Sernia C, Ho KKY 1993 Effects ofdifferent oral estrogen formulations on insulin-like growth factor-1, growthhormone and growth hormone binding protein in post-menopausal women.Clin Endocrinol (Oxf) 39:561–567
947. Hartmann B, Kirchengast S, Albrecht A, Laml T, Bikas D, Huber J 1995Effects of hormone replacement therapy on growth hormone secretion pat-terns in correlation to somatometric parameters in healthy postmenopausalwomen. Maturitas 22:239–246
948. Murphy LJ, Freisen HG 1988 Differential effects of estrogen and growthhormone on uterine and hepatic insulin-like growth factor-I expression inthe ovariectomized hypophysectomised rat. Endocrinology 122:325–332
949. Holloway L, Butterfield G, Hintz R, Gesundheit N, Marcus R 1994 Effectsof recombinant human growth hormone on metabolic indices, body com-position, and bone turnover in healthy elderly women. J Clin EndocrinolMetab 79:470–479
950. Lieberman SA, Mitchell AM, Marcus R, Hintz RL, Hoffman AR 1994 Theinsulin-like growth factor I generation test: resistance to growth hormonewith aging and estrogen replacement therapy. Horm Metab Res 26:229–233
951. Lima L, Arce V, Lois N, Fraga C, Lechuga MJ, Tresquerres JA, Deversa J1989 Growth hormone (GH) responsiveness to GHRH in normal adults is notaffected by short-term gonadal blockade. Acta Endocrinol (Copenh) 120:31–36
952. Benito P, Avila L, Corpas MS, Jimenez JA, Cacicedo L, Sanchez Franco F1991 Sex differences in growth hormone response to growth hormone-releasing hormone. J Endocrinol Invest 14:265–269
953. Lang I, Schernthaner G, Pietschmann P, Kurz R, Stephenson JM, TemplH 1987 Effects of sex and age on growth hormone response to growthhormone-releasing hormone in healthy individuals. J Clin Endocrinol Metab65:535–540
954. Merimee TJ, Rabinowitz D, Fineberg SE 1969 Arginine-initiated release ofhuman growth hormone. Factors modifying the response in normal men.N Engl J Med 280:1434–1438
955. Giustina A, Gastaldi C, Bugari G, Chiesa L, Loda G, Tironi C, Negro-VilarA 1995 Role of galanin in the regulation of somatotrope and gonadotropefunction in young ovulatory woman. Metabolism 44:1028–1032
956. Rao RH, Spathis GS 1987 Intramuscular glucagon as a provocative stimulusfor the assessment of pituitary function: growth hormone and cortisol re-sponses. Metab Clin Exp 36:658–663
957. Wideman L, Weltman JY, Shah N, Story S, Bowers CY, Veldhuis JD,Weltman A, The effects of arginine and GHRP2 on resting and exerciseinduced growth hormone (GH) release. Program of 79th Annual Meeting ofThe Endocrine Society, Minneapolis, MN, 1997 (Abstract P2–197)
958. Badger TM, Millard WJ, Owens SM, Larovere J, O’Sullivan D 1991 Effectsof gonadal steroids on clearance of growth hormone at steady state in therat. Endocrinology 128:1065–1072
959. Rosenbaum M, Gertner JM 1989 Metabolic clearance rates of synthetichuman growth hormone in children, adult women, and adult men. J ClinEndocrinol Metab 69:821–824
960. Holl RW, Schwarz U, Schauwecker P, Benz R, Veldhuis JD, Heinze E 1993Diurnal variation of the elimination of human growth hormone: the serumhalf-life is prolonged in the evening, and affected by the source of thehormone, as well as by body size and serum estradiol. J Clin EndocrinolMetab 77:216–220
961. Shah N, Evans WS, Veldhuis JD, Mode of GH entry into the bloodstream,rather than gender or sex-steroid hormones, determines GH half-life in thehuman. Program of the 79th Annual Meeting of The Endocrine Society,Minneapolis, MN, 1997 (Abstract P2–203)
962. Van den Berg G, Frolich M, Veldhuis JD, Roelfsema F 1994 Growthhormone secretion in recently operated acromegalic patients. J Clin Endo-crinol Metab 79:1706–1715
963. Giustina A, Wehrenberg WB 1995 Influence of thyroid hormones on theregulation of growth hormone secretion. Eur J Endocrinol 133:646–653
964. Levy A, Matovelle MC, Lightman SL, Young III WS 1992 The effects ofpituitary stalk transection, hypophysectomy and thyroid hormone status oninsulin-like growth factor 2-, growth hormone releasing hormone-, andsomatostatin mRNA prevalence in rat brain. Brain Res 579:1–7
965. Jones PM, Burrin JM, Ghatei MA, O’Halloran DJ, Legon S, Bloom SR 1990The influence of thyroid hormone status on the hypothalamo-hypophysealgrowth hormone axis. Endocrinology 126:1374–1379
966. De Gennearo V, Cella SG, Bassetti M, Rizzi R, Cocchi D, Mueller EE 1988Impaired growth hormone secretion in neonatal hypothyroid rats: hypo-thalamic vs. pituitary component. Proc Soc Exp Biol Med 187:99–106
967. Katakami H, Downs TR, Frohman LA 1986 Decreased hypothalamicgrowth hormone-releasing hormone content and pituitary responsiveness inhypothyroidism. J Clin Invest 77:1704–1711
968. Miki N, Ono M, Hizuka N, Aoki T, Demura H 1992 Thyroid hormonemodulation of the hypothalamic growth hormone (GH)-releasing factor-pituitary GH axis in the rat. J Clin Invest 90:113–120
969. De los Frailes MT, Cacicedo L, Lorenzo MJ, Fernandez G, Sanchez-FrancoF 1998 Thyroid hormone action on biosynthesis of somatostatin by fetal ratbrain cells in culture. Endocrinology 123:898–904
970. Peterfreund RA, Sawchenko PE, Vale W 1995 Thyroid hormone reversiblysuppresses somatostatin secretion and immunoreactivity in cultured neo-cortical cells. Brain Res 328:259
971. Berelowitz M, Maeda K, Harris S, Frohman LA 1985 The effect on alter-ations in the pituitary-thyroid axis on hypothalamic content and in vitrorelease of somatostatin-like immunoreactivity. Endocrinology 116:1066–1070
972. Webb SM, Champney TH, Reiter RJ 1995 Nighttime immunoreactive so-matosatin content of the median eminence in hypo- and hyperthyroid rats.Comp Biochem Physiol 80:575–577
973. Martin D, Epelbaum J, Bluet-Pajot MT, Prelot M, Kordon C, Durand D1985 Thyroidectomy abolishes pulsatile growth hormone secretion withoutaffecting hypothalamic somatostatin. Neuroendocrinology 41:476–481
974. Gillioz P, Giraud P, Conte-Devolx B, Jaquet P, Codaccioni JL, Oliver C1979 Immunoreactive somatostatin in rat hypophysial portal blood. Endo-crinology 103:1406–1410
975. Samuels HH, Shapiro LE 1976 Thyroid hormone stimulates de novo growthhormone synthesis in cultured GH1 cells: evidence for the accumulation ofa rate-limiting RNA species in the induction process. Proc Natl Acad Sci USA73:3369–3376
976. Shapiro LE, Samuels HH, Yaffe BM 1978 Thyroid and glucocorticoid hor-mones synergistically control growth hormone mRNA in cultured GH1 cells.Proc Natl Acad Sci USA 75:45–54
977. Spindler SR, Mellon SH, Baxter JD 1982 Growth hormone gene transcrip-tion is regulated by thyroid and glucocorticoid hormones in cultured ratpituitary tumor cells. J Biol Chem 257:11627–11634
978. Diamond DJ, Goodman HM 1985 Regulation of growth hormone messen-ger RNA synthesis by dexamethasone and triiodothyronine. J Mol Biol181:41–52
979. Dobner PR, Kawasaki ES, Yu L, Brancroft FC 1981 Thyroid or glucocor-ticoid hormone induces pre-growth hormone mRNA and its probable nu-clear precursor in rat pituitary cells. Proc Natl Acad Sci USA 78:2230–2238
980. Mulchahey JJ, Di Blasio AM, Jaffe RB 1988 Effects of growth hormone(GH)-releasing hormone and somatostatin on GH secretion from individualhuman and monkey fetal anterior pituitary cells: modulation by thyroidhormones and glucocorticoids. J Clin Endocrinol Metab 66:395–400
981. Adams EF, Brajkovich IE, Masheter K 1981 Growth hormone and prolactinsecretion by dispersed cell cultures of human pituitary adenomas: long-termeffects of hydrocortisone, estradiol, insulin, 3,5,39-triiodothyronine and thy-roxine. J Clin Endocrinol Metab 53:381–386
982. Cattini IA, Anderson TR, Baxter JC, Mellon P, Eberhardt NL 1986 Thehuman growth hormone gene is negatively regulated by triiodothyroninewhen transfected into rat pituitary tumor cells. J Biol Chem 261:13367–13376
983. Brent GA, Hamey JW, Moore DD, Larsen PR 1988 Multihormonal regu-lation of the human, rat, and bovine growth hormone promoters: differentialeffects of 39,59-cyclic adenosine monophosphate, thyroid hormone, and glu-cocorticoids. Mol Endocrinol 2:792–798
984. Tam SP, Lam KS, Srivastava G 1996 Gene expression of hypothalamicsomatostatin, growth hormone releasing factor, and their pituitary receptorsin hypothyroidism. Endocrinology 137:418–424
985. Chernausek SD, Turner R 1989 Attenuation of spontaneous, nocturnalgrowth hormone secretion in children with hypothyroidism and its corre-lation with plasma insulin-like growth factor I concentration. J Pediatr 114:968–972
986. Iwatsubo H, Omori K, Okado Y, Fukuchi M, Miyai K, Abe H 1967 Humangrowth hormone secretion in primary hypothyroidism before and aftertreatment. J Clin Endocrinol Metab 27:1751–1754
987. Bruman H, Smets P, Corvilain J 1973 Comparative study of growth hor-mone response to hypoglycemia in normal subjects and in patients withprimary myxedema or hyperthyroidism before and after treatment. J ClinEndocrinol Metab 36:1162–1174
794 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

988. Katz HP, Youlton R, Kaplan SL, Grumbach MM 1969 Growth and growthhormone. III. Growth hormone release in children with primary hypothy-roidism and thyrotoxicosis. J Clin Endocrinol Metab 29:346–348
989. Valcavi R, Zini M, Portioli I 1992 Thyroid hormones and growth hormonesecretion. J Endocrinol Invest 15:313–330
990. Finkelstein JW, Boyar RM, Hellman L 1974 Growth hormone secretion inhyperthyroidism. J Clin Endocrinol Metab 38:634–638
991. Sasaki N, Tsuyusaki T, Nakamura H, Sanayama K, Niimi H, NakajiamaH 1985 Sleep related growth hormone release in thyrotoxic patients beforeand during propylthiouracil therapy. Endocrinol Jpn 32:39–44
992. Iranmanesh A, Lizarralde G, Johnson ML, Veldhuis JD 1991 Nature ofaltered growth hormone secretion in hyperthyroidism. J Clin EndocrinolMetab 72:108–115
993. Valimaki M, Liewendahl K, Karonen SL, Helenius T, Suikkari AM 1990Concentration of somatomedin-C and triiodothyronine in patients withthyroid dysfunction and non thyroidal illnesses. J Endocrinol Invest 13:155–159
994. Miell JP, Taylor AM, Zini M, Maheshwari H, Ross RM, Valcavi R 1993Effects of hypothyroidism and hyperthyroidism on insulin-like growth fac-tors (IGFs) and growth hormone (GH) and IGF binding proteins. J ClinEndocrinol Metab 76:950–955
995. Binoux M, Faivre-Bauman A, Lassarre C, Tixier-Vidal A 1985 Triiodothy-ronine stimulates the production of insulin-like growth factor I (IGF-I) byfetal hypothalamus cells cultured in serum free medium. Dev Brain Res21:319–323
996. Giustina G, Reschini E, Valentini F, Cantalamessa L 1971 Growth hormoneand cortisol responses to insulin-induced hypoglycemia in thyroxtoxicosis.J Clin Endocrinol Metab 32:130–133
997. Giustina A, Ferrari C, Bodini C, Buffoli MG, Legati F, Schettino M, Zuc-cato F, Wehrenberg WB 1990 Effects of methimazole treatment on growthhormone (GH) response to GH-releasing hormone in patients with hyper-thyroidism. Acta Endocrinol (Copenh) 123:613–618
998. Giustina A, Buffoli MG, Ferrari C, Gazzoli N, Pizzocolo G, Tassi C,Romanelli G 1991 Acute effect of propranolol on the growth hormone j(GH)response to GH-releasing hormone in patients with hyperthyroidism. HormMetab Res 23:506–508
999. Valcavi R, Dieguez C, Zini M, Page MD, Dotti C, Portioli I 1991 Effect ofpyridostigmine and pirenzepine on GH responses to GHRH in hyperthyroidpatients. Clin Endocrinol (Oxf) 35:141–144
1000. Giustina A, Schettino M, Bussi AR, Legati F, Licini M, Zuccato F,Wehrenberg WB 1992 Effect of arginine on the GHRH stimulated GHsecretion in patients with hyperthyroidism. Horm Res 38:259–265
1001. Ramos-Dias JC, Pimentel-Filho F, Reis AF, Lengyel AM 1996 Differentgrowth hormone (GH) response to GH-releasing peptide and GH-releasinghormone in hyperthyroidism. J Clin Endocrinol Metab 81:1343–1346
1002. de Zegher F, Devlieger H, Eggermont E, Veldhuis JD 1993 Properties ofgrowth hormone and prolactin hypersecretion by the human infant on theday of birth. J Clin Endocrinol Metab 76:1177–1181
1003. Wright NM, Northington FJ, Miller JD, Veldhuis JD, Rogol AD 1992Elevated growth hormone secretory rate in premature infants: deconvolu-tion analysis of pulsatile GH secretion in the neonate. Pediatr Res 32:286–290
1004. de Zegher F, Van den Berghe G, Devlieger H, Eggermont E, Veldhuis JD1993 Dopamine inhibits growth hormone and prolactin secretion in thehuman newborn. Pediatr Res 34:642–645
1005. Veldhuis JD, Iranmanesh A, Wilkowski MJ, Samojlik E 1994 Neuroen-docrine alterations in the somatotropic and lactotropic axes in uremic men.Eur J Endocrinol 131:489–498
1006. Giustina A, Lorusso R, Borghetti V, Bugari G, Misitano V, Alfieri O 1996Impaired spontaneous growth hormone secretion in severe dilated cardio-myopathy. Am Heart J 131:629–632
1007. Hartman ML, Veldhuis JD, Johnson ML, Lee MM, Alberti KG, SamojlikE, Thorner MO 1992 Augmented growth hormone (GH) secretory burstfrequency and amplitude mediate enhanced GH secretion during a two-dayfast in normal men. J Clin Endocrinol Metab 74:757–765
1008. Martha Jr PM, Rogol AD, Veldhuis JD, Blizzard RM 1996 A longitudinalassessment of hormonal and physical alterations during normal puberty inboys. II. The neuroendocrine growth hormone axis during late prepuberty.J Clin Endocrinol Metab 81:4068–4074
1009. Roemmich JN, Clark PA, Mai V, Berr SS, Weltman A, Veldhuis JD, RogolAD 1998 Alterations in growth and body composition during puberty. III.Influence of maturation, gender, body composition, fat distribution, aerobicfitness, and energy expenditure on nocturnal growth hormone release. J ClinEndocrinol Metab 83:1440–1447
1010. Challa A, Krieg RJ, Thabet MA, Veldhuis JD, Chan JCM 1993 Metabolicacidosis inhibits growth hormone secretion in the rat. Mechanism of growthretardation. Am J Physiol 265:E547–E553
1011. Veldhuis JD 1996 Neuroendocrine mechanisms mediating awakening of thegonadotropic axis in puberty. Pediatr Nephrol 10:304–317
1012. Giustina A, Volterrani M, Manelli F, Desenzani P, Poiesi C, Lorusso R,Giordano A 1998 Endocrine predictors of the acute hemodynamic efffectsof growth hormone in congestive heart failure. Am Heart J, in press
1013. Brungger M, Hulter HN, Krapf R 1997 Effect of chronic metabolic acidosis
on the growth hormone/IGF-I endocrine axis: new cause of growth hormoneinsensitivity in humans. Kidney Int 51:216–221
1014. Kowarski A, Thompson RG, Migeon CJ, Blizzard RM 1971 Determinationof integrated plasma concentration and true secretion rates of human growthhormone. J Clin Endocrinol Metab 32:356–360
1015. Rudman D, Vintner MH, Rogers CM, Lubin MF, Fleming GH, RaymondPB 1981 Impaired growth hormone secretion in the adult population. J ClinInvest 67:1361–1369
1016. Mauras N, Hayes V, Welch S, Rini A, Helgeson K, Dokler M, Veldhuis JD,Urban RJ 1998 Testosterone deficiency in young men: marked alterations inwhole body protein kinetics, strength and adiposity. J Clin Endocrinol Metab83:1886–1892
1017. Blizzard RM, Thompson RG, Baghdassarian A 1974 The interrelationshipof steroids, growth hormone and other hormones in pubertal growth. In:Grumbach MN, Grave JD, Mayer FE (eds) The Control of the Onset ofPuberty. John Wiley and Sons, New York, pp 342–359
1018. Parker MW, Johanson AJ, Rogol AD, Kaiser DL, Blizzard RM 1984 Effectof testosterone on somatomedin-C concentrations in prepubertal boys. J ClinEndocrinol Metab 58:87–90
1019. Plotnick LP, Thompson RG, Beitins I, Blizzard RM 1974 Integrated con-centrations of growth hormone correlated with stage of puberty and estro-gen levels in girls. J Clin Endocrinol Metab 38:436–439
1020. Ho KY, Evans WS, Blizzard RM, Veldhuis JD, Merriam GR, Samojlik E,Furlanetto R, Rogol AD, Kaiser DL, Thorner MO 1987 Effects of sex andage on the 24-hour profile of growth hormone secretion in man: importanceof endogenous estradiol concentrations. J Clin Endocrinol Metab 64:51–58
1021. Veldhuis JD, Iranmanesh A, Lizarralde G, Urban RJ 1994 Combined def-icits in the somatotropic and gonadotropic axes in healthy older men: anappraisal of neuroendocrine mechanisms by deconvolution analysis. Neu-robiol Aging 15:509–517
1022. Zadik Z, Chalew SA, McCarter Jr RJ, Meistas M, Kowarski AA 1985 Theinfluence of age on the 24-hour integrated concentration of growth hormonein normal individuals. J Clin Endocrinol Metab 60:513–516
1023. Prinz PN, Weitzman ED, Cunningham GR, Karacan I 1983 Plasma growthhormone during sleep in young and aged men. J Gerontol 38:519–524
1024. Cook CB, Nippoldt TB, Kletter GB, Kelch RP, Marshall JC 1991 Naloxoneincreases the frequency of LH (GnRH) pulsatile secretion in hyperprolactine-mia. J Clin Endocrinol Metab 73:1099–1105
1025. Mauras N, Rogol AD, Veldhuis JD 1990 Increased hGH production rateafter low-dose estrogen therapy in prepubertal girls with Turner’s syn-drome. Pediatr Res 28:626–630
1026. Spellacy WN, Buhi WC 1969 Pituitary GH and PL levels measured innormal term pregnancy and at the early and late postpartum periods. Am JObstet Gynecol 15:888–896
1027. Hennen G, Frankenne F, Closset J, Gomez F, Pirens G 1985 A humanplacental GH: increasing levels during second half of pregnancy and pitu-itary GH suppression as revealed by monoclonal antibody radioimmuno-assay. Int J Fertil 30:27–34
1028. de Zegher F, Vanderschueren-Lodeweyckx. M, Spitz B 1990 Perinatalgrowth hormone (GH) physiology: effect of GH-releasing factor on maternaland fetal secretion of pituitary and placental GH. J Clin Endocrinol Metab77:520–522
1029. De Leo V, Lanzetta D, D’Antona D, Latessa AM, Petraglia F 1992 Controlof growth hormone secretion during the postpartum period. Gynecol ObstetInvest 33:31–35
1030. Corpas E, Harman SM, Blackman MR 1993 Human growth hormone andhuman aging. Endocr Rev 14:20–39
1031. Urban RJ, Veldhuis JD 1988 Hypothalamo-pituitary concomitants of aging.In: Sowers JR, Felicetta JV (eds) The Endocrinology of Aging. Raven Press,New York, pp 41–74
1032. Giustina A, Desenzani P, Bossoni S, Perini P 1997 Growth hormone treat-ment in aging: state of the art and perspectives. Aging Clin Exp Res 9:73–74
1033. Jorgensen JO, Christiansen JS 1993 Brave new senescence: GH in adults.Lancet 341:1247–1248
1034. Ghigo E, Arvat E, Gianotti L, Ramunni J, Di Vito L, Maccagno B, GrottoliS, Camanni F 1996 Human aging and the GH-IGF-I axis. J Pediatr EndocrinolMetab 9:271–278
1035. Muggeo M, Fedele D, Tiengo A, Molinari M, Crepaldi G 1975 Humangrowth hormone and cortisol responses to insulin stimulation in aging. JGerontol 30:546–551
1036. Ghigo E, Ceda GP, Valcavi R, Goffi S, Zini M, Mucci M, Valenti G, CocchiD, Mueller EE, Camanni F 1994 Low doses of either intravenously or orallyadministered arginine are able to enhance growth hormone response togrowth hormone releasing hormone in elderly subjects. J Endocrinol Invest17:113–122
1037. Arvat E, Gianotti L, Grottoli S, Imbimbo BP, Lenaerts V, Deghenghi R,Camanni F, Ghigo E 1994 Arginine and growth hormone-releasing hormonerestore the blunted growth hormone-releasing activity of hexarelin in elderlysubjects. J Clin Endocrinol Metab 79:1440–1443
1038. Cella SG, Moiraghi V, Minuto F, Barreca A, Cocchi D, De Gennaro ColonaV, Reina G, Mueller EE 1989 Prolonged fasting or clonidine can restore the
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 795
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

defective growth hormone secretion in old dogs. Acta Endocrinol (Copenh)121:177–184
1039. Ghigo E, Ceda GP, Valcavi R, Goffi S, Zini M, Mucci M, Valenti G,Mueller EE, Camanni F 1995 Effect of 15-day treatment with growth-hormone-releasing hormone alone or combined with different doses ofarginine on the reduced somatotrope responsiveness to the neurohormonein normal aging. Eur J Endocrinol 132:32–36
1040. degli Uberti EC, Ambrosio MR, Cella SG, Margutti AR, Trasforini G,Rigamonti AE, Petrone E, Mueller EE 1997 Defective hypothalamic growthhormone (GH)-releasing hormone activity may contribute to declining GHsecretion with age in man. J Clin Endocrinol Metab 82:2885–2888
1041. Cella SG, Arce VM, Pieretti F, Locatelli V, Settembrini BP, Mueller EE 1993Combined administration of growth-hormone-releasing hormone andclonidine restores defective growth hormone secretion in old dogs. Neu-roendocrinology 57:432–438
1042. Chapman IM, Hartman ML, Pezzoli SS, Harrell Jr FE, Hintz RL, AlbertiKGMM, Thorner MO 1997 Effect of aging on the sensitivity of growthhormone secretion to insulin-like growth factor-I negative feedback. J ClinEndocrinol Metab 82:2996–3004
1043. Van den Berghe G, de Zegher F 1996 Anterior pituitary function duringcritical illness and dopamine treatment. Crit Care Med 24:1580–1590
1044. Van den Berghe G, de Zegher F, Bouillon R 1998 Acute and prolongedcritical illness as different neuroendocrine paradigms. J Clin EndocrinolMetab 83:1827–1834
1045. Shetty KR, Duthie Jr EH 1995 Anterior pituitary function and growthhormone use in the elderly. Endocrinol Metab Clin North Am 24:213–231
1046. Borst SE, Millard WJ, Lowenthal DT 1994 Growth hormone, exercise, andaging: the future of therapy for the frail elderly. J Am Geriatr Soc 42:528–535
1047. Carter WJ 1995 Effect of anabolic hormones and insulin-like growth factor-1on muscle mass and strength in elderly persons. Clin Geriatr Med 11:735–748
1048. Vittone J, Blackman MR, Busby-Whitehead J, Tsiao C, Stewart KJ, TobinJ, Stevens T, Bellantoni MF, Rogers MA, Baumann G, Roth J, Harman SM,Spencer RG 1997 Effects of single nightly injections of growth hormone-releasing hormone (GHRH 1–29) in healthy elderly men. Metab Clin Exp46:89–96
1049. Khorram O, Laughlin GA, Yen SSC 1997 Endocrine and metabolic effectsof long-term administration of [Nle27]growth hormone-releasing hormone-(1–29)-NH2 in age-advanced men and women. J Clin Endocrinol Metab82:1472–1479
1050. Papadakis MA, Grady D, Black D, Tierney MJ, Gooding GA, SchambelanM, Grunfeld C 1996 Growth hormone replacement in healthy older menimproves body composition but not functional ability. Ann Intern Med124:708–716
1051. Thompson JL, Butterfield GE, Marcus R, Hintz RL, Van Loan M, GhironL, Hoffman AR 1995 The effects of recombinant human insulin-like growthfactor-I and growth hormone on body composition in elderly women. J ClinEndocrinol Metab 80:1845–1852
1052. Welle S, Thornton C, Statt M, McHenry B 1996 Growth hormone increasesmuscle mass and strength but does not rejuvenate myofibrillar protein inhealthy subjects over 60 years old. J Clin Endocrinol Metab 81:3239–3243
1053. Taaffe DR, Pruitt L, Reim J, Hintz RL, Butterfield G, Hoffman AR, MarcusR 1994 Effect of recombinant human growth hormone on the muscle re-sponse to resistance to exercise in elderly men. J Clin Endocrinol Metab79:1361–1366
1054. Nicklas BJ, Ryan AJ, Treuth MM, Harman SM, Blackman MR, Hurley BF,Rogers MA 1995 Testosterone, growth hormone and IGF-I responses toacute and chronic resistive exercise in men aged 55–70 years. Int J Sports Med16:445–450
1055. Pyka G, Wiswell RA, Marcus R 1992 Age-dependent effect of resistanceexercise on growth hormone secretion in people. J Clin Endocrinol Metab75:404–407
1056. Giustina A, Cappelli C 1997 Interaction between testosterone and growthhormone replacement therapy. In: Lamberts SWJ (ed) The Diagnosis andTreatment of Pituitary Insufficiency, BioScientific, Ltd, Bristol, UK, pp 181–208
1057. Kraemer WJ, Patton JF, Gordon SE, Harman EA, Deschenes MR, ReynoldsK, Newton RU, Triplett NT, Dziados JE 1995 Compatibility of high-inten-sity strength and endurance training on hormonal and skeletal muscleadaptions. J Appl Physiol 78:976–989
1058. Grobet L, Poncelet D, Royo LJ, Brouwers B, Pirottin D, Michaux C, Menis-sier F, Zanotti M, Dunner S, Georges M 1998 Molecular definition of anallelic series of mutations disrupting the myostatin function and causingdouble-muscling in cattle. Mamm Genome 9:210–213
1059. Grobet L, Martin LJ, Poncelet D, Pirottin D, Brouwers B, Riquet J, Schoe-berlein A, Dunner S, Menissier F, Massabanda J, Fries R, Hanset R,Georges M 1997 A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nat Genet 17:71–74
1060. Kambadur R, Sharma M, Smith TP, Bass JJ 1997 Mutations in myostatin(GDF8) in double-muscled Belgian Blue and Piedmontese cattle. GenomeRes 7:910–916
1061. McPherron AC, Lee SJ 1997 Double muscling in cattle due to mutations inthe myostatin gene. Proc Natl Acad Sci USA 94:12457–12461
1062. Jorgensen JO, Vahl N, Hansen TB, Fisker S, Hagen C, Christiansen JS 1996Influence of growth hormone and androgens on body composition in adults.Horm Res 45:94–98
1063. Johannsson G, Bjarnason R, Bramnert M, Carlsson LM, Degerblad M,Manhem P, Rosen T, Thoren M, Bengtsson BA 1996 The individual re-sponsiveness to growth hormone (GH) treatment in GH-deficient adults isdependent on the level of GH-binding protein, body mass index, age, andgender. J Clin Endocrinol Metab 81:1575–1581
1064. Hodes RJ 1994 Frailty and disability: can growth hormone or other trophicagents make a difference? J Am Geriatr Soc 42:1208–1211
1065. Gargosky SE, Tapanainen P, Rosenfeld RG 1994 Administration of growthhormone (GH), but not insulin-like growth factor-I (IGF-I), by continuousinfusion can induce the formation of the 150-kilodalton IGF-binding pro-tein-3 complex in GH-deficient rats. Endocrinology 134:2267–2276
1066. Roelfsema F, van Dulken H, Frolich M 1985 Long-term results of trans-sphenoidal pituitary microsurgery in 60 acromegalic patients. Clin Endo-crinol (Oxf) 23:555–565
1067. Growth Hormone Research Society 1998 Invited report of a workshop.Consensus guidelines for the diagnosis and treatment of adults with growthhormone deficiency: summary statement of the growth hormone researchsociety workshop on adult growth hormone deficiency. J Clin EndocrinolMetab 83:379–395
1068. Lucidi P, Lauteri M, Laureti S, Celleno R, Santoni S, Volpi E, Angeletti G,Santeusanio F, De Feo P 1998 A dose-response study of growth hormone(GH) replacement on whole body protein and lipid kinetics in GH-deficientadult. J Clin Endocrinol Metab 83:353–357
1069. Wolfe J 1998 Growth hormone: a physiological fountain of youth? JAMA1:9–25
1070. Janssen YJH, Deurenberg P, Roelfsema F 1997 Using dilution techniquesand multi-frequency bioelectrical impedance to assess both total body waterand extracellular water at baseline and during rhGH treatment in GH de-ficient adults. J Clin Endocrinol Metab 82:3349–3355
1071. Vahl N, Moller N, Lauritzen T, Christiansen JS, Jorgensen JOL 1997Metabolic effects and pharmacokinetics of a growth hormone pulse inhealthy adults: relation to age, sex, and body composition. J Clin EndocrinolMetab 82:1–7
1072. Pietrowsky R, Struben C, Molle M, Fehm HL, Born J 1996 Brain potentialchanges after intranasal vs. intravenous administration of vasopressin: ev-idence for a direct nose-brain pathway for peptide effects in humans. BiolPsychiatry 39:332–340
1073. Pietrowsky R, Thiemann A, Kern W, Fehm HL, Born J 1996 A nose-brainpathway for psychotropic peptides: evidence from a brain evoked potentialstudy with cholecystokinin. Psychoneuroendocrinology 21:559–572
1074. Kern W, Schiefer B, Schwarzenburg J, Stange EF, Born J, Fehm HL 1997Evidence for central nervous effects of corticotropin-releasing hormone ongastric acid secretion in humans. Neuroendocrinology 65:291–298
1075. Borer K, Kelch RP 1978 Increased serum growth hormone and somaticgrowth in exercised adult hamster. Am J Physiol 234:E611–E616
1076. Butkus JA, Brogan RS, Giustina A, Kastello G, Sothmann M, WehrenbergWB 1995 Changes in the growth hormone axis due to exercise training inmale and female rats: secretory and molecular responses. Endocrinology136:2664–2670
1077. Felsing NE, Brasel JA, Cooper DM 1992 Effect of low and high intensityexercise on circulating growth hormone in men. J Clin Endocrinol Metab75:157–162
1078. Sutton J, Lazarus L 1976 Growth hormone in exercise: comparison of phys-iological and pharmacological stimuli. J Appl Physiol 41:523–527
1079. Weeke J, Gundersen HJG 1983 The effect of heating and central cooling onserum TSH, GH, and norepinephrine in resting normal man. Acta PhysiolScand 117:33–39
1080. Lassarre C, Girard F, Durand J, Raynaud J 1974 Kinetics of human growthhormone during submaximal exercise. J Appl Physiol 37:826–830
1081. Raynaud J, Capderou A, Martineaud JP, Bordachar J, Durand J 1983 In-tersubject variability in growth hormone time course during different typesof work. J Appl Physiol 55:1682–1687
1082. Parkin JM 1986 Exercise as a test of growth hormone secretion. Acta En-docrinol (Copenh) 279:47–50
1083. Vanhelder WP, Goode RC, Radomski MW 1984 Effect of anaerobic andaerobic exercise of equal duration and work expenditure on plasma growthhormone levels. Eur J Appl Physiol 52:255–257
1084. Hartley LH, Mason JW, Hogan RP, Jones IG, Kotchen TA, Mougey EH,Wherry FE, Pennington LL, Ricketts PT 1972 Multiple hormonal responseto graded exercise in relation to physical training. J Appl Physiol 33:602–606
1085. Kraicer J, Sheppard MS, Luke J, Lussier B, Moor BC, Cowan JS 1988 Effectof withdrawal of somatostatin and growth hormone (GH)-releasing factoron GH release in vitro. Endocrinology 122:1810–1817
1086. Kanaley JA, Boileau RA, Bahr JA, Misner JE, Nelson RA 1992 Substrateoxidation and GH responses to exercise are independent of menstrual phaseand status. Med Sci Sports Exerc 24:873–880
1087. Wideman L, Weltman JY, Weltman A, Patterson M, Mistry DJ, VeldhuisJD, Effect of growth hormone (GH) infusion on GH secretion during rest and
796 GIUSTINA AND VELDHUIS Vol. 19, No. 6
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022

exercise. Program of the 80th Annual Meeting of The Endocrine Society,New Orleans, LA, 1998 (Abstract P1–194)
1088. Weltman A, Weltman JY, Schurrer R, Evans WS, Veldhuis JD, Rogol AD1992 Endurance training amplifies the pulsatile release of growth hormone:effects of training intensity. J Appl Physiol 76:2188–2196
1089. Vigas M, Nemeth S, Jurcovicova JM, Komadel L 1992 The importance oflactate in exercise induced growth hormone release in man. Horm Res166–169
1090. Uusitupa M, Siitonen O, Harkonen M, Gordin A, Aro A, Hersio K, Jo-hansson G, Korhonen T, Rauramaa R 1982 Modification of the metabolicand hormonal response to physical exercise by beta-blocking agents. AnnClin Res 14:165–167
1091. Hansen AP 1971 The effect of adrenergic blockade on the exercise-inducedserum growth hormone rise in normals and juvenile diabetics. J Clin En-docrinol Metab 33:807–812
1092. Sutton J, Lazarus L 1974 Effect of adrenergic blocking agents on growthhormone responses to physical exercise. Horm Metab Res 6:428–429
1093. Luger A, Watschinger B, Deuster P, Svoboda T, Clodi M, Chrousos GP1992 Plasma growth hormone and prolactin responses to graded levels ofacute exercise and to a lactate infusion. Neuroendocrinology 56:112–117
1094. Cappa M, Grossi A, Benedetti S, Drago F, Loche S, Ghigo E 1993 Effect ofthe enhancement of the cholinergic tone by pyridostigmine on the exercise-induced growth hormone release in man. J Endocrinol Invest 16:421–424
1095. Thompson DL, Weltman JY, Rogol AD, Metzger D, Veldhuis JD, WeltmanA 1993 Cholinergic and opioid involvement in release of growth hormoneduring exercise and recovery. J Appl Physiol 75:870–878
1096. Coiro V, Volpi R, Maffei ML, Caoazza A, Caffarri G, Capretti L, Colla R,Chiodera P 1994 Opioid modulation of the gamma-aminobutyric acid-controlled inhibition of exercise-stimulated growth hormone and prolactinsecretion in normal man. Eur J Endocrinol 131:50–55
1097. Veldhuis JD, Lassiter AB, Johnson ML 1990 Operating behavior of dual ormultiple endocrine pulse generators. Am J Physiol 259:E351–E361
1098. Drisko JE, Faidley TD, Chang CH, Zhang D, Nicolich S, Hora Jr DFMcNamara L, Rickes E, Abribat T, Smith RG, Hickey GJ 1998 Hypo-physeal-portal concentrations of growth hormone-releasing factor and so-matostatin in conscious pigs: relationship to production of spontaneousgrowth hormone pulses. Proc Soc Exp Biol Med 217:188–196
1099. Bruno JF, Olchovsky D, White JD, Leidy JW, Song J, Berelowitz M 1990Influence of food deprivation in the rat on hypothalamic expression ofgrowth hormone-releasing factor and somatostatin. Endocrinology127:2111–2116
1100. Jansson J-O, Eden S, Isaksson O 1985 Sexual dimorphism in the control ofgrowth hormone secretion. Endocr Rev 6:128–150
1101. Pavlou EP, Harman SM, Merriam GR, Gelato MC, Blackman MR 1986Responses of growth hormone (GH) and somatomedin-C to GH-releasinghormone in healthy aging men. J Clin Endocrinol Metab 62:595–600
1102. Tannenbaum GS 1991 Neuroendocrine control of growth hormone secre-tion. Acta Paediatr Scand Suppl 372:5–16
1103. Zeitler P, Tannenbaum GS, Clifton DK, Steiner RA 1991 Ultradian oscil-lations in somatostatin and growth hormone-releasing hormone mRNAs insomatostatin and growth hormone-releasing hormone mRNAs in the brainsof adult male rats. Proc Natl Acad Sci USA 88:8920–8924
December, 1998 PATHOPHYSIOLOGY OF GH NEUROREGULATION 797
Dow
nloaded from https://academ
ic.oup.com/edrv/article/19/6/717/2530805 by guest on 27 M
ay 2022
Copyright © 2022 FDOKUMEN




















