
Pathogenicity of Alternanthera mosaic virus is affected by determinants in RNA-dependent RNA...
11
Downloaded from www.microbiologyresearch.org by IP: 54.159.222.200 On: Sat, 28 May 2016 14:26:45 Pathogenicity of Alternanthera mosaic virus is affected by determinants in RNA-dependent RNA polymerase and by reduced efficacy of silencing suppression in a movement-competent TGB1 Hyoun-Sub Lim, 1 Anna Maria Vaira, 1,2 Michael D. Reinsel, 1 Hanhong Bae, 3 3 Bryan A. Bailey, 3 Leslie L. Domier 4 and John Hammond 1 Correspondence John Hammond [email protected] 1 USDA-ARS, USNA, Floral and Nursery Plants Research Unit, Beltsville, MD 20705, USA 2 CNR, Istituto di Virologia Vegetale, Torino 10135, Italy 3 USDA-ARS, PSI, Sustainable Perennial Crops Laboratory, Beltsville, MD 20705, USA 4 USDA-ARS, Department of Crop Sciences, University of Illinois at Urbana-Champaign, Urbana, IL 61801, USA Received 10 July 2009 Accepted 24 September 2009 Four biologically active cDNA clones were derived from the Alternanthera mosaic virus (AltMV; genus Potexvirus) isolate, AltMV-SP, which differ in symptoms in infected Nicotiana benthamiana plants. Two clones induced necrosis and plant death; a mixture of all four clones induced milder symptoms than AltMV-SP. Replication of all clones was enhanced by a minimum of fourfold at 15 6C. A mixture of clones 4-7 (severe) and 3-1 (mild) was indistinguishable from AltMV-SP, but the ratio of 4-7 to 3-1 differed at 25 and 15 6C. RNA copy numbers of mixed infections were always below those of 4-7 alone. Determinants of symptom severity were identified in both Pol and TGB1; the mildest (4-1) and most severe (3-7) clones differed at three residues in the ‘core’ Pol domain [R(1110)P, K(1121)R, R(1255)K] and one [S(1535)P] in the C-terminal Pol domain of RNA-dependent RNA polymerase, and one in TGB1 [P(88)L]. Pol [P 1110 ,R 1121 ,K 1255 ]+TGB1 L(88) ] always induced systemic necrosis at 15 6C. Gene exchanges of Pol and TGB1 each affected replication and symptom expression, with TGB1 P(88) significantly reducing silencing suppression. The difference in silencing suppression between TGB1 P(88) and TGB1 L(88) was confirmed by an agroinfiltration assay. Further, co-expression of TGB1 P(88) and TGB1 L(88) resulted in interference in the suppression of silencing by TGB1 L(88) . Yeast two-hybrid analysis confirmed that TGB1 P(88) and TGB1 L(88) interact. These results identify a TGB1 residue that significantly affects replication and silencing suppression, but maintains full movement functions. INTRODUCTION Alternanthera mosaic virus (AltMV) is a potexvirus first described from Alternanthera pungens, closely related to Papaya mosaic virus (Geering & Thomas, 1999). AltMV also infects the species Phlox, and the 6607 nt sequence of AltMV-PA has been determined previously (Hammond et al., 2006). Symptom differences between isolates have been reported; in Nicotiana benthamiana, AltMV-SP (the isolate studied here) and AltMV-PA produce chlorotic lesions on inoculated leaves and systemic chlorotic mosaic, while AltMV-Po produced milder symptoms (Hammond et al., 2006). Occurrence of multiple sequence types, or mixed isolates, within a single plant has been reported for multiple virus groups including potexviruses. For example, six distinct subisolates of Plantago asiatica mosaic virus (PlAMV) were recovered from a single lily (Komatsu et al., 2008). As RNA-dependent RNA polymerase (RdRp) lacks proof- reading activity, RNA virus populations accumulate variations; population variation is limited by selection pressure for variants that interact successfully with host and viral proteins necessary to complete the infection cycle (Garcia-Arenal et al., 2001; Liang et al., 2002; Rico et al., 2006; Schneider & Roossinck, 2001). Changing environmental conditions such as temperature can affect 3Present address: School of Biotechnology, Yeungnam University, Geongsan, 712-749, Republic of Korea. The GenBank/EMBL/DDBJ accession numbers for the sequences reported in this paper are GQ179646 and GQ179647 for AltMV-SP 3-1 and AltMV-SP 4-7, respectively. A supplementary table is available with the online version of this paper. Journal of General Virology (2010), 91, 277–287 DOI 10.1099/vir.0.014977-0 014977 Printed in Great Britain 277
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Pathogenicity of Alternanthera mosaic virus is affected by determinants in RNA-dependent RNA...

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
Pathogenicity of Alternanthera mosaic virus isaffected by determinants in RNA-dependent RNApolymerase and by reduced efficacy of silencingsuppression in a movement-competent TGB1
Hyoun-Sub Lim,1 Anna Maria Vaira,1,2 Michael D. Reinsel,1
Hanhong Bae,33 Bryan A. Bailey,3 Leslie L. Domier4 and John Hammond1
Correspondence
John Hammond
1USDA-ARS, USNA, Floral and Nursery Plants Research Unit, Beltsville, MD 20705, USA
2CNR, Istituto di Virologia Vegetale, Torino 10135, Italy
3USDA-ARS, PSI, Sustainable Perennial Crops Laboratory, Beltsville, MD 20705, USA
4USDA-ARS, Department of Crop Sciences, University of Illinois at Urbana-Champaign, Urbana,IL 61801, USA
Received 10 July 2009
Accepted 24 September 2009
Four biologically active cDNA clones were derived from the Alternanthera mosaic virus (AltMV;
genus Potexvirus) isolate, AltMV-SP, which differ in symptoms in infected Nicotiana benthamiana
plants. Two clones induced necrosis and plant death; a mixture of all four clones induced milder
symptoms than AltMV-SP. Replication of all clones was enhanced by a minimum of fourfold at
15 6C. A mixture of clones 4-7 (severe) and 3-1 (mild) was indistinguishable from AltMV-SP, but
the ratio of 4-7 to 3-1 differed at 25 and 15 6C. RNA copy numbers of mixed infections were
always below those of 4-7 alone. Determinants of symptom severity were identified in both Pol
and TGB1; the mildest (4-1) and most severe (3-7) clones differed at three residues in the ‘core’
Pol domain [R(1110)P, K(1121)R, R(1255)K] and one [S(1535)P] in the C-terminal Pol domain of
RNA-dependent RNA polymerase, and one in TGB1 [P(88)L]. Pol [P1110,R1121,K1255]+TGB1L(88)]
always induced systemic necrosis at 15 6C. Gene exchanges of Pol and TGB1 each affected
replication and symptom expression, with TGB1P(88) significantly reducing silencing
suppression. The difference in silencing suppression between TGB1P(88) and TGB1L(88) was
confirmed by an agroinfiltration assay. Further, co-expression of TGB1P(88) and TGB1L(88)
resulted in interference in the suppression of silencing by TGB1L(88). Yeast two-hybrid analysis
confirmed that TGB1P(88) and TGB1L(88) interact. These results identify a TGB1 residue that
significantly affects replication and silencing suppression, but maintains full movement
functions.
INTRODUCTION
Alternanthera mosaic virus (AltMV) is a potexvirus firstdescribed from Alternanthera pungens, closely related toPapaya mosaic virus (Geering & Thomas, 1999). AltMValso infects the species Phlox, and the 6607 nt sequence ofAltMV-PA has been determined previously (Hammondet al., 2006). Symptom differences between isolates havebeen reported; in Nicotiana benthamiana, AltMV-SP (theisolate studied here) and AltMV-PA produce chlorotic
lesions on inoculated leaves and systemic chlorotic mosaic,while AltMV-Po produced milder symptoms (Hammondet al., 2006).
Occurrence of multiple sequence types, or mixed isolates,within a single plant has been reported for multiple virusgroups including potexviruses. For example, six distinctsubisolates of Plantago asiatica mosaic virus (PlAMV) wererecovered from a single lily (Komatsu et al., 2008). AsRNA-dependent RNA polymerase (RdRp) lacks proof-reading activity, RNA virus populations accumulatevariations; population variation is limited by selectionpressure for variants that interact successfully with hostand viral proteins necessary to complete the infectioncycle (Garcia-Arenal et al., 2001; Liang et al., 2002; Ricoet al., 2006; Schneider & Roossinck, 2001). Changingenvironmental conditions such as temperature can affect
3Present address: School of Biotechnology, Yeungnam University,Geongsan, 712-749, Republic of Korea.
The GenBank/EMBL/DDBJ accession numbers for the sequencesreported in this paper are GQ179646 and GQ179647 for AltMV-SP 3-1and AltMV-SP 4-7, respectively.
A supplementary table is available with the online version of this paper.
Journal of General Virology (2010), 91, 277–287 DOI 10.1099/vir.0.014977-0
014977 Printed in Great Britain 277

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
virus RNA replication (Aldaoud et al., 1989; Kaper et al.,1995).
The potexvirus TGB1 is a multifunctional protein, withactivities important for cell-to-cell movement, increasingplasmodesmatal size exclusion limit, RNA silencingsuppression and RNA helicase (Verchot-Lubicz et al.,2007). TGB1 is also the elicitor for resistance mediated bythe Nb gene of potato (Malcuit et al., 1999). Bayne et al.(2005) utilized random mutagenesis to generate two classesof potato virus X (PVX) TGB1 mutants defective insilencing suppression. One class supported cell-to-cellmovement if complemented by a heterologous suppressorof RNA silencing, while the second class was also defectivein a second function required for movement.
We report here multiple distinct biologically active cDNAclones (hereafter ‘infectious clones’) derived from AltMV-SP, and use of these clones and chimeric derivatives toidentify determinants contributing to symptom severityand RNA accumulation. Both the RdRp Pol domain andTGB1 contribute to symptom severity, and significantlyaffect RNA accumulation. Instead of emergence ofvariation and subsequent selection, we observed stableco-existence in N. benthamiana of two dominant sequencevariants within AltMV-SP, resulting in modulation ofsymptom severity and RNA accumulation. Two singleamino acid variants of TGB1 differ significantly in theability to suppress RNA silencing, and interact withinhibitory effects on silencing suppression, differentiatingsilencing suppression from movement functions of TGB1in fully infectious clones of AltMV.
RESULTS
Four distinct clones infectious as transcripts wererecovered from AltMV-SP-infectedN. benthamiana
The AltMV genome was amplified as separate 59 and 39
fragments, each containing a unique MluI site at nt 3125,upstream of the RdRp Pol domain. Four 59 clones (nt 1–3143; RdRp clones 1–4) and seven 39 clones (nt 3117–6607;[Pol+TGB+CP] clones 1–7) were ligated in all combina-tions to produce 28 full-length AltMV clones. Only clones3-1, 3-7, 4-1 and 4-7, combining 59[RdRp] clones 3 or 4and 39[Pol+TGB+CP] clones 1 or 7 (Fig. 1a), wereinfectious. For each infectious clone, 3/3 transcript-inoculated plants were infected as detected by RT-PCRusing primers PP12/PP15 and Western blotting usingAltMV-specific antibodies. No plants were infected by anyother full-length transcripts.
Infectious clones AltMV 3-1 and 4-7 were fully sequencedand submitted to GenBank (accession nos GQ179646 andGQ179647). Clones 3-7 and 4-1 are identical to 3-1 and 4-7from nt 1 to 3125, and to 4-7 and 3-1 from nt 3126 to 6607,respectively.
Symptoms of infectious clones inoculated separately variedfrom very mild to severe depending upon the combinationof 59 and 39 regions; none produced symptoms typical ofAltMV-SP (Fig. 1b). At 25 uC, AltMV 3-1 produced mildsystemic mosaic 10 days post-inoculation (p.i.). However,plants recovered with symptoms rarely visible at 30 daysp.i. No symptoms were observed in 4-1-infected plants at25 uC. When [Pol+TGB+CP] was derived from clone 7(3-7 and 4-7), symptoms were more severe than AltMV-SP;3-7 always induced localized systemic necrosis by 10 daysp.i. and growth was inhibited. Necrosis observed with 4-7was delayed with respect to 3-7; symptoms were initiallymild, but necrosis developed by 20 days p.i., a response notobserved with AltMV-SP at 25 uC. AltMV-SP and mixturesof all four infectious clones produced mild mosaicsymptoms without necrosis (Fig. 1b, upper panels).
All clones induced more severe symptoms at 15 than at25 uC. Clones 3-7 and 4-7 induced necrosis and killedplants by 30 days p.i.; with 3-1 and 4-1, necrosis wasdetected in inoculated and upper leaves and the stem.AltMV-SP and the mixture of all clones developed severesymptoms at 15 uC, but without necrosis until about30 days p.i. (Fig. 1b, lower panels).
Quantification of viral RNA
Differences in AltMV RNA accumulation were observed byquantitative real-time PCR (Q-RT-PCR) using CP-(Fig. 1c) or RdRp-specific primers (Supplementary TableS1, available in JGV Online) in N. benthamiana plantsinfected with each clone, suggesting that both [RdRp] and[Pol+TGB+CP] regions affect replication and/or systemicmovement. Viral RNA levels were significantly increased ininfections including 39 clone 7 compared with clone 1; andwere similarly increased by 59 clone 3 compared with clone4. RNA levels were correlated with symptom severity(Fig. 1).
Viral RNA accumulation of all clones at 15 uC was.fivefold that at 25 uC, as shown by Q-RT-PCR witheither RdRp or CP primers (Fig. 1c and data not shown).
Sequence analysis of mild and severe clones
A total of 240 nt differences (three in the 59 UTR and 237in RdRp) between 59 clones 3 and 4 result in 45 predictedamino acid differences within RdRp. These included five inthe methyl transferase (MT) domain, 24 between MT andAlkB, four in AlkB, eight between AlkB and RNA helicase,and four in the RNA helicase domains (data not shown).The contribution of individual differences was notinvestigated.
There is a difference of 21 nt between 39 [Pol+TGB+CP]
clones 1 and 7 (eight in RdRp-Pol; six in TGB1; one each inTGB3, the intergenic region and the 39 UTR and four inCP). There were only 5 aa differences between 1-[Pol+TGB+CP] and 7-[Pol+TGB+CP]: R(1110)P,
H.-S. Lim and others
278 Journal of General Virology 91

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
K(1121)R, R(1255)K in the ‘core’ Pol domain, S(1535)P inthe ‘variable’ Pol C-terminal domain (pfam00978; Fig. 2a),and TGB1 P(88)L, in the helicase domain (pfam01443; Fig.2b), with no differences in TGB2, TGB3 or CP. A TGB1amino acid alignment of AltMV and closely relatedpotexviruses showed that most had Leu, as for AltMVposition 88 in clones 3-7 and 4-7, but PapMV has Ile andCCMV has Val; no other GenBank sequences examinedhad Pro at this position (Fig. 2c and data not shown).
AltMV-SP exists in infected N. benthamiana as amixture of variants
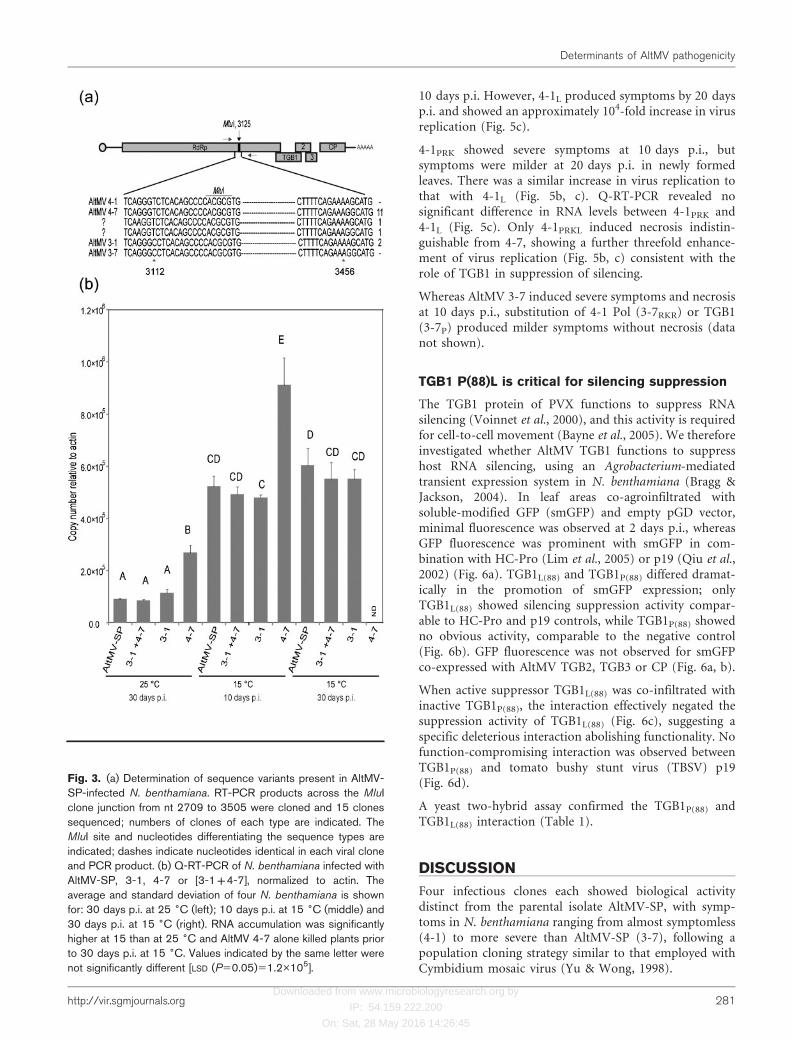
To determine the prevalence of each cloned variant inAltMV-SP, we amplified a region containing changesdistinguishing the 59 and 39 clones (nt 3112 and 3456),using primers PP3/PP20 (Supplementary Table S1).Analysis of 15 cloned RT-PCR products revealed fourtypes of sequences (Fig. 3a); 11 clones identical to the 4-7sequence and two identical to 3-1 were identified.Sequences identical to 3-7 (most severe) and 4-1 (mildest)were not found. Two additional clones had a silent Aupstream from the MluI site where all infectious clones had
a G; downstream one of these clones was identical to 4-7,the other to 3-1. These variants were not studied further.
We also inoculated AltMV-SP to two groups of five N.benthamiana, maintained at 25 and 15 uC. Q-RT-PCR wasperformed at 30 days p.i. with individual plants in order todistinguish 3-1 and 4-7 sequence types. Primer pairs RdRp-F(3-1)/RdRp-R(3-1) and RdRp-F(4-7)/RdRp-R(4-7) have39 nucleotides distinguishing these variants (positions 418and 574; Supplementary Table S1). Q-RT-PCRs wereadjusted to enhance specificity and results were normalizedto actin. At 25 uC, Ct values relative to actin of the (4-7)-like target were 8.61(±3.80) times higher than the (3-1)-like target, consistent with the ratio of cloned RT-PCRproducts. At 15 uC, Ct values for the (4-7)-like target wereonly 1.74(±0.97) times higher than the (3-1)-like target(data not shown).
Mimicking AltMV-SP with infectious clones
Co-inoculation of 4-7 and 3-1 produced symptomsindistinguishable from AltMV-SP. Q-RT-PCR showed thatRNA accumulation of [3-1+4-7] was indistinguishablefrom 3-1 alone or from AltMV-SP at either 15 or 25 uC.
Fig. 1. (a) Full-length clones were derived from AltMV-SP by population cloning, including ligation at the nt 3125 MluI site.Clones 3-1, 3-7, 4-1 and 4-7 were infectious. (b) Each clone produced symptoms distinct from each other and AltMV-SP(shown at 20 days p.i.). Symptoms were more severe at 15 (lower panels) than at 25 6C (upper panels). All clones together(‘Mix’, 3-1+3-7+4-1+4-7) produced symptoms milder than AltMV-SP. (c) Q-RT-PCR with CP-specific primers. RelativeRNA accumulation of 3-1, 3-7, 4-1 and 4-7 was normalized to actin. The average value and standard deviation at 30 days p.i. isshown from four N. benthamiana per clone, at 25 6C (left) and 15 6C (right). RNA accumulation was significantly higher at 15than at 25 6C; 3-7 and 4-7 killed plants prior to 30 days p.i. at 15 6C. Values indicated by the same letter were not significantlydifferent [LSD (P50.05)56.6�104]. RdRp primers yielded equivalent results (data not shown).
Determinants of AltMV pathogenicity
http://vir.sgmjournals.org 279

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
Accumulation of 4-7 alone was significantly higher. In eachcase accumulation was .fourfold higher at 15 than at25 uC (Fig. 3b). Plants infected with 4-7 at 15 uC diedbefore 30 days p.i., while plants infected with AltMV-SP, 3-1 or [3-1+4-7] survived past 30 days p.i. with mosaicsymptoms. At 10 days p.i., accumulation of 4-7 RNA at15 uC was .50 % higher than for AltMV-SP, 3-1 or [3-1+4-7] (Fig. 3b).
Mixed infections of fluorescently labelled clonesreveal differential systemic distribution
To compare cell-to-cell movement and distribution, 3-1 and4-1 were modified to express the enhanced green fluorescentprotein (eGFP), and 3-7 and 4-7 to express DsRed. Equalamounts of 3-1:eGFP and 4-7:DsRed (Fig. 4a) were co-inoculated to N. benthamiana. At 25 uC, 3-1:eGFP and 4-7:DsRed were equally distributed in systemically infectedleaves at 10 days p.i. (data not shown). At 30 days p.i., 4-7:DsRed spread to an area about 10-fold larger than 3-1:eGFP in the uppermost leaves, with many co-infected cells(Fig. 4b). The same pattern occurred when 3-1:eGFP and 3-7:DsRed were co-inoculated, indicating correlation with the39 rather than the 59 region.
At 15 uC, 4-7:DsRed and 3-1:eGFP infected similar areas ofthe leaf at 30 days p.i. with many co-infected cells (Fig. 4c, d).
The MluI junction region was amplified at 30 days p.i.from plants co-infected with 3-1:eGFP and 4-7:DsRed,maintained at 25 or 15 uC. Analysis of RT-PCR-derivedclones revealed a 4-7:DsRed to 3-1:eGFP ratio of 8 : 1 at25 uC and 6 : 4 at 15 uC (Fig. 4e).
Mixed infection (at 25 uC) of 3-7:DsRed and 4-1:eGFPresulted in minimal GFP expression at 10 days p.i.; GFPwas not detectable systemically at 30 days p.i., while DsRedwas detected throughout (data not shown).
Substitution of RdRp Pol or TGB1 from 3-7 into4-1 enhances replication and induces necrosis
To determine which regions are important for severesymptoms, we performed gene exchanges between themildest clone 4-1 and the most severe 3-7. We separatelysubstituted the 3-7 ‘core’ Pol [P1110R1121K1255], TGB1L(88)
and [Pol P1110R1121K1255+TGB1L(88)] domains into 4-1 tocreate 4-1PRK, 4-1L and 4-1PRKL, respectively (Fig. 5a). Wealso replaced 3-7 Pol with Pol [R1110K1121R1255] from 4-1 tocreate 3-7RKR, and TGB1L(88) with TGB1P(88) to create 3-7P
(Fig. 5a). In all 4-1 background chimeric clones, RdRpresidue 1535 was changed to S1535, while in the 3-7background P1535 was maintained, in order to examine theeffects of TGB1 residue 88 in a uniform background.Neither 4-1 nor 4-1L developed significant symptoms by
Fig. 2. Amino acid sequence alignments(KAlign, EMBL-EBI) between AltMV 4-7[severe (sev), upper] and 3-1 (mild, lower).(a) Portions of Pol showing the four amino acidchanges at position 1110, 1121, 1255 and1535. Residues highly conserved withinRdRp-2 domain [pfam 00978; NCBIConserved Domain Database (CDD)] areshaded. (b) Portion of TGB1 showing theunique P(88)L difference. Residues conservedwithin helicase domain (pfam01443, viral-heli-case-1; NCBI CDD) are shaded. (c) Portion ofTGB1 alignment of AltMV 4-7 (severe), 3-1(mild) and eight other members of the genusPotexvirus yielding the highest scores (.200)in a BLASTP search. AltMV 4-7 TGB1 Leu 88 ishighly conserved, with chemically similar Ile inPapMV and Val in CCMV; Pro was not foundat the equivalent position in any other potex-virus isolate. PapMV, Papaya mosaic virus;CVX, Cactus virus X; ZVX, Zygocactus virus X;SVX, Schlumbergera virus X; PlAMV, Plantagoasiatica mosaic virus; OVX; Opuntia virus X;TVX, Tulip virus X; CCMV, Cassava commonmosaic virus.
H.-S. Lim and others
280 Journal of General Virology 91
Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
10 days p.i. However, 4-1L produced symptoms by 20 daysp.i. and showed an approximately 104-fold increase in virusreplication (Fig. 5c).
4-1PRK showed severe symptoms at 10 days p.i., butsymptoms were milder at 20 days p.i. in newly formedleaves. There was a similar increase in virus replication tothat with 4-1L (Fig. 5b, c). Q-RT-PCR revealed nosignificant difference in RNA levels between 4-1PRK and4-1L (Fig. 5c). Only 4-1PRKL induced necrosis indistin-guishable from 4-7, showing a further threefold enhance-ment of virus replication (Fig. 5b, c) consistent with therole of TGB1 in suppression of silencing.
Whereas AltMV 3-7 induced severe symptoms and necrosisat 10 days p.i., substitution of 4-1 Pol (3-7RKR) or TGB1(3-7P) produced milder symptoms without necrosis (datanot shown).
TGB1 P(88)L is critical for silencing suppression
The TGB1 protein of PVX functions to suppress RNAsilencing (Voinnet et al., 2000), and this activity is requiredfor cell-to-cell movement (Bayne et al., 2005). We thereforeinvestigated whether AltMV TGB1 functions to suppresshost RNA silencing, using an Agrobacterium-mediatedtransient expression system in N. benthamiana (Bragg &Jackson, 2004). In leaf areas co-agroinfiltrated withsoluble-modified GFP (smGFP) and empty pGD vector,minimal fluorescence was observed at 2 days p.i., whereasGFP fluorescence was prominent with smGFP in com-bination with HC-Pro (Lim et al., 2005) or p19 (Qiu et al.,2002) (Fig. 6a). TGB1L(88) and TGB1P(88) differed dramat-ically in the promotion of smGFP expression; onlyTGB1L(88) showed silencing suppression activity compar-able to HC-Pro and p19 controls, while TGB1P(88) showedno obvious activity, comparable to the negative control(Fig. 6b). GFP fluorescence was not observed for smGFPco-expressed with AltMV TGB2, TGB3 or CP (Fig. 6a, b).
When active suppressor TGB1L(88) was co-infiltrated withinactive TGB1P(88), the interaction effectively negated thesuppression activity of TGB1L(88) (Fig. 6c), suggesting aspecific deleterious interaction abolishing functionality. Nofunction-compromising interaction was observed betweenTGB1P(88) and tomato bushy stunt virus (TBSV) p19(Fig. 6d).
A yeast two-hybrid assay confirmed the TGB1P(88) andTGB1L(88) interaction (Table 1).
DISCUSSION
Four infectious clones each showed biological activitydistinct from the parental isolate AltMV-SP, with symp-toms in N. benthamiana ranging from almost symptomless(4-1) to more severe than AltMV-SP (3-7), following apopulation cloning strategy similar to that employed withCymbidium mosaic virus (Yu & Wong, 1998).
Fig. 3. (a) Determination of sequence variants present in AltMV-SP-infected N. benthamiana. RT-PCR products across the MluIclone junction from nt 2709 to 3505 were cloned and 15 clonessequenced; numbers of clones of each type are indicated. TheMluI site and nucleotides differentiating the sequence types areindicated; dashes indicate nucleotides identical in each viral cloneand PCR product. (b) Q-RT-PCR of N. benthamiana infected withAltMV-SP, 3-1, 4-7 or [3-1+4-7], normalized to actin. Theaverage and standard deviation of four N. benthamiana is shownfor: 30 days p.i. at 25 6C (left); 10 days p.i. at 15 6C (middle) and30 days p.i. at 15 6C (right). RNA accumulation was significantlyhigher at 15 than at 25 6C and AltMV 4-7 alone killed plants priorto 30 days p.i. at 15 6C. Values indicated by the same letter werenot significantly different [LSD (P50.05)51.2�105].
Determinants of AltMV pathogenicity
http://vir.sgmjournals.org 281

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
There were significant differences in RNA accumulationbetween infectious clones when inoculated separately at25 uC, with levels of 3-7 .4-7 .3-1 .4-1. The 59 clone 3resulted in higher replication than clone 4, and the 39 clone7 to higher replication than clone 1, indicating that both 59
and 39 regions are separately involved in pathogenicity, asRNA accumulation was strongly correlated with symptomseverity (Fig. 1).
Because of the many differences between the 59 clones, thecontribution of individual differences was not investigated
further, but amino acid differences in the RdRp Poldomain (residues 1110, 1121 and 1255) in the Pol ‘core’domain (pfam00978) that is conserved among manyflexiviruses were examined. The S(1535)P difference inthe variable Pol C-terminal domain was not furtherinvestigated. The TGB1 P(88)L change is in the relativelyconserved helicase domain (pfam01443), and was interest-ing as the most similar potexvirus TGB1 sequences haveLeu (most common), Ile or Val at that position. Pro wasnot observed at the equivalent position in any otherpotexvirus TGB1 sequence.
Fig. 4. (a) eGFP and DsRed were insertedbetween TGB3 and CP of AltMV 3-1 and 4-7,respectively, under the control of a duplicatedCP sg promoter. AltMV 4-1 was similarlylabelled with eGFP, and 3-7 with DsRed. (b–d) 3-1:eGFP and 4-7:DsRed were co-inocu-lated to N. benthamiana at 25 or 15 6C. Theyoungest expanded leaf was observed byconfocal microscopy at (b) 30 days p.i.(25 6C); and (c) 30 days p.i. (15 6C). (d) Theoutlined region of panel (c) at higher mag-nification; images shown represent: (top)overlay of eGFP and DsRed, (middle) eGFPalone and (bottom) DsRed alone. Bars,100 mm. (e) Relative occurrence of 3-1:eGFPand 4-7:DsRed in upper leaves. PCR productsof the MluI clone junction region amplified fromupper leaves at 30 days p.i. as in (c) and (d)were cloned and sequenced to determineabundance of each sequence type.
H.-S. Lim and others
282 Journal of General Virology 91

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
A single amino acid substitution in the Pol domain of PVXand PlAMV RdRps contributes to necrotic symptoms in N.benthamiana (Kagiwada et al., 2005; Ozeki et al., 2006).Amino acid substitutions between the helicase andmethylase regions of TMV and Pepper mild mottle virusRdRps also affect viral accumulation and symptomexpression (Hagiwara et al., 2002; Lewandowski &Dawson, 1993). For AltMV, not only the Pol domain butalso multiple differences in the upstream RdRp regionscontribute to necrosis. A single residue in TGB1 also hasmajor effects on viral accumulation and necrosis. AlthoughPVX TGB1 has been identified as the elicitor of Nb-mediated HR (Malcuit et al., 1999), we believe this is thefirst identification of TGB1 as a pathogenicity determinantaffecting the severity of disease. The potexvirus TGB1protein has several functions: cell-to-cell movement, RNAhelicase, silencing suppression and increasing PD sizeexclusion limit (Verchot-Lubicz et al., 2007). Bayne et al.(2005) obtained several random TGB1 mutants in whichabrogation of silencing suppressor activity was linked toloss of cell-to-cell movement activity. In contrast, we haveidentified a TGB1 mutant which significantly affectsreplication and silencing suppression, but maintains fullmovement functions.
We have demonstrated that 4-7 (severe) and 3-1 (mild)variants are both maintained in AltMV-SP and in co-inoculated plants and that their prevalence differs withtemperature (Fig. 3). Co-inoculation mimics AltMV-SPinfection in both symptoms and virus replication levels(Fig. 3). Coexistence was visualized in vivo in systemicinfections of eGFP- and DsRed-expressing clones (Fig. 4).3-1:eGFP infected far fewer cells than 4-7:DsRed at 25 uC,but at 15 uC infection by 3-1:eGFP increased from about 10to 50 % of the leaf area, whereas 4-7:DsRed infected almostall cells and co-infection of many cells was evident. Co-infections of 3-1:eGFP with 3-7:DsRed yielded similar data,suggesting that the difference in TGB1 rather than betweenthe 59 regions of 3-1 and 4-7 is most important.
To further understand the involvement of Pol and TGB1differences in viral replication, we made substitutionsbetween the phenotypic extremes of 3-7 and 4-1, neither ofwhich was detected in the AltMV-SP population. Variantsbearing the Pol (4-1PRK) and TGB1 (4-1L) from 3-7 hadreplication levels increased by approximately 1000-foldwith respect to 4-1, and similar effects on symptomseverity. Synergism was observed when Pol and TGB1 wereinserted together (4-1PRKL), resulting in approximately
Fig. 5. Mild clone AltMV 4-1 was substituted with [Pol], [TGB1] and [Pol+TGB1] domains of severe clone 3-7; and 3-7substituted with [Pol] and [TGB1] of 4-1. (a) Five hybrid clones were created, 4-1PRK, 4-1L and 4-1PRKL, and 3-7RKR and 3-7P.(b) 4-1 and each derivative clone were inoculated to N. benthamiana; symptoms are shown at 30 days p.i. in plants inoculatedwith 4-1, 4-1PRK, 4-1L and 4-1PRKL. (c) Q-RT-PCR comparison of 4-1, 4-1PRK, 4-1L and 4-1PRKL accumulation levels at30 days p.i.
Determinants of AltMV pathogenicity
http://vir.sgmjournals.org 283

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
threefold higher RNA levels, together with early necrosisand sustained symptom severity (Fig. 5). These effects wereconfirmed by reciprocal exchanges in the 3-7 background.
Low temperature (15 uC) dramatically increased accu-mulation of each variant inoculated separately, with
symptoms following the same trend (Fig. 1). This wasmost apparent with 3-1 and 4-1. The explanation may bedifferences in plant RNA silencing activity; at lowertemperature the host defence silencing mechanism appearsto be less effective (Chellappan et al., 2005; Jovel et al.,2007; Qu et al., 2005; Siddiqui et al., 2008), allowing aproportionately greater increase in virus replication and/ornumber of cells infected by the challenging virus.
Hypothesizing the involvement of RNA silencing suppres-sion, we tested AltMV CP, TGB3, TGB2 as well asTGB1P(88) and TGB1L(88) variants in a silencing suppressorassay. Only TGB1L(88) had appreciable silencing suppressoractivity and a single point mutation in TGB1P(88) – notobserved in any previously characterized potexvirus isolate– is able to dramatically reduce TGB1 silencing suppressorability, while maintaining viral movement functions.Interactions between TGB1P(88) and TGB1L(88) significantlydiminished functionality of TGB1L(88) (Fig. 6), and maydecrease the proportion of severe variant in a mixedinfection.
In conclusion, differences in virus replication and symp-tom production are likely to be due to interactions betweenRdRp and TGB1 determinants. The previously undescribedTGB1P(88) mutation can explain how 3-1 has lower relativefitness at 25 uC and increased fitness at 15 uC due toreduced effectiveness of host silencing activity. Thereduction of silencing activity demonstrated whenTGB1P(88) and TGB1L(88) interact may explain how 3-1 isable to reduce replication of 4-7 at both high and lowtemperature in mixed infections in which many cells areco-infected, and maintenance of both variants in AltMV-SP. This is consistent with the work of Simon et al. (2005),who have shown that minority genotypes can have animportant influence on the overall pathogenicity of viralpopulations.
METHODS
Plant material. N. benthamiana were grown in 10 cm pots, and 3- to
4-week-old plants were used for virus infection. Inoculated plantswere grown in an air-conditioned greenhouse maintained at 15 uC, or
a greenhouse at 25 uC, under a 14 h light regime. Mechanical
inoculations were performed using 1 % K2HPO4 with celite added as
an abrasive.
Virus isolates and construction of AltMV clones infectious as
transcripts. AltMV-SP was isolated from Phlox stolonifera ‘SherwoodPurple’ by mechanical transmission (Hammond et al., 2006), and
serially passaged in N. benthamiana over several years. Infected N.
benthamiana tissue stored at 270 uC from various time points (serial
passages) was used separately to reinitiate AltMV-SP, and to examine
the presence of distinct sequence types over time. Total RNA was
isolated from N. benthamiana leaves using the RNeasy Mini kit
(Qiagen); 2 mg total RNA was used to generate cDNA using
SuperScript III RNase H2 Reverse Transcriptase (Invitrogen)
separately with an oligo(dT)20 primer and internal reverse primer
MluI-R, containing an MluI site that is unique in the sequence ofAltMV-PA (Hammond et al., 2006) at nt 3125. Two cDNAs were
amplified by (i) a 59 non-coding region primer including a T7
Fig. 6. Agroinfiltration was used for comparing gene silencingsuppressor function. For silencing suppression assays, pGD:smGFP was co-infiltrated with (a) pGD (empty vector), pGD:TGB3, pGD:HC-Pro and pGD:p19; (b) pGD:Coat, pGD:TGB2,pGD:TGB1L(88) and pGD:TGB1P(88); (c) [pGD (empty vector)+pGD:TGB1L(88)] and [pGD:TGB1P(88)+pGD:TGB1L(88); (d) [pGD(empty vector)+pGD:p19] and [pGD:TGB1P(88)+pGD:p19].
Table 1. Yeast two-hybrid interactions of TGB1L(88) andTGB1P(88)
TGB1 sequences were fused to the activation domain (AD) or binding
domain (BD) and interactions assessed by growth at 48 h. ‘+’
indicates strong growth by 48 h; ‘2’ indicates no growth.
Construct Interaction
BD AD
TGB1L(88) TGB1L(88) +
TGB1P(88) TGB1P(88) +
TGB1L(88) TGB1P(88) +
TGB1P(88) TGB1L(88) +
BD TGB1L(88) 2
TGB1P(88) AD 2
H.-S. Lim and others
284 Journal of General Virology 91

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
promoter sequence upstream of AltMV nt 1 (PstI-F) paired withMluI-R, and (ii) MluI-F and XbaI-R1, respectively (SupplementaryTable S1) using Pfu polymerase (Stratagene). The two PCR productswere separately cloned into a TOPO vector (Invitrogen), yielding fourclones containing the 59 portion of the genome (nt 1–3143), andseven clones containing the 39 region [nt 3117 to the poly(A) tail];each contained the nt 3125 MluI site. The seven 39 region clones weredigested individually with MluI and XbaI, and each insert individuallyligated into each of the 59 region clones similarly cleaved with MluIand XbaI to create 28 distinct full-length AltMV clones. Each full-length clone was linearized with XbaI, and transcribed in vitro (seebelow); three young N. benthamiana plants per clone were inoculatedwith the transcripts. Inoculated plants were assayed by RT-PCR usingAltMV-specific primers PP12 and PP15 (Supplementary Table S1)and Western blotting. The infectious clones obtained were namedAltMV 3-1 and 4-1 (mild clones: 59 clones 3 and 4, 39 clone 1), andAltMV 3-7 and 4-7 (severe clones: 59 clones 3 and 4, 39 clone 7)(Fig. 1).
Substitution of Pol and TGB1 in chimeric AltMV clones. The 39
region clones 1 and 7 were used to prepare chimeric constructs. ThePol domain of 39 clone 1 was digested with MluI (at nt 3125) andBamHI (at nt 4484) and exchanged with MluI/BamHI Pol fragment of39 clone 7. To replace [Pol+TGB1], clone 1 was digested with MluI(nt 3125) and XmaI (nt 5480) and this fragment replaced with theclone 7 MluI/XmaI [Pol+TGB1] fragment. The clone 1 TGB1 regionwas substituted with the BamHI/XmaI TGB1 fragment of clone 7, andTGB1 of clone 7 substituted by the BamHI/XmaI fragment of clone 1.Because the BamHI/XmaI fragment also included the Pol P(1535)Sdifference between clones 1 and 7, P1535 was altered to S1535 in theclone 1 background so that differences due to TGB1 residue 88 couldbe evaluated independently. Overlap PCR (Wurch et al., 1998) wasused to substitute S1535 into clone 1 using primers Pol-F/(P to S)-Rand (P to S)-F/Pol-R (Supplementary Table S1). Clone 7 Pol andTGB1 domains were similarly substituted into clone 1, and S1535 waschanged to P1535 in the clone 7 background by overlap PCR usingprimers Pol-F/S(1535)P-F and S(1535)P-R/Pol-R (SupplementaryTable S1) such that the only amino acid change was TGB1 P(88).Structures of chimeric clones were confirmed by sequence analysis.The chimeric 39 subclones were then digested with MluI and XbaI andcombined with appropriate MluI/XbaI cleaved 59 region clones tocreate chimeric full-length clones 4-1PRK (Pol from clone 7); 4-1L
(TGB1 from clone 7); 4-1PRKL ([Pol+TGB1] from clone 7); 3-7RKR
(Pol from clone 1) and 3-7P (TGB1 from clone 1).
In vitro transcription of AltMV clones. Twenty micrograms ofXbaI-linearized plasmid DNA were used per 50 ml transcriptionreaction for each of the full-length clones, AltMV 3-1, 4-1, 3-7 and 4-7 and transcript RNA generated using T7 RNA polymerase in parallelreactions to obtain equivalent amounts of transcripts (Petty et al.,1989). Transcripts were ethanol precipitated, resuspended in 20 mlGKP buffer (50 mM glycine, 30 mM KHPO4, pH 9.2, 1 % bentonite,1 % celite) per 50 ml transcription reaction and 10 ml inoculated toeach of two leaves per N. benthamiana plant (Petty et al., 1989).
Agrobacterium infiltration, AltMV constructs and silencing
suppression assays. All binary vectors were derived from pGD aspreviously described (Goodin et al., 2002). PCR primers foramplification of each gene are shown in Supplementary Table S1.TGB1 of AltMV 3-1 and 3-7, and TGB2, TGB3, and CP of 3-7 wereseparately amplified and inserted between the XhoI and BamHI sitesof pGD. Constructs pGD:smGFP and pGD:p19 (TBSV p19) were agift of Andy Jackson (Department of Plant and Microbial Biology,University of California, Berkeley, USA; Bragg & Jackson, 2004).pGD:HC-Pro (Soybean mosaic virus HC-Pro) was amplified usingHC-Pro XhoI-F/HC-Pro XmaI-R from pG5-HC-Pro (Lim et al.,2005).
An smGFP silencing suppression assay was used as previously
described (Bragg & Jackson, 2004). Because 35S promoter-controlled
GFP induces silencing of GFP expression (Shiboleth et al., 2007;Silhavy & Burgyan, 2004), we used only pGD:smGFP co-infiltrated
with the putative silencing suppressor; pGD:smGFP was co-infiltrated
with pGD:TGB1L88 (3-7), and pGD:TGB1P(88) (3-1), pGD:TGB2,
pGD:TGB3, pGD:CP, pGD:HC-Pro and pGD:p19, respectively.
Q-RT-PCR. Total RNA was isolated from young systemically infected
leaves of N. benthamiana (as above), and treated with DNase I per the
manufacturer’s recommendation; 2 mg total RNA was used to
generate cDNA in a 20 ml reaction with 0.5 mg of an oligo(dT)20
primer (as above). N. benthamiana actin (GenBank accession no.AY179605) was utilized as an internal control. Q-RT-PCR was
performed with primer sets targeting AltMV RdRp and CP, and actin
mRNA (Supplementary Table S1) using an Mx3005P QPCR System
and Brilliant SYBR Green QPCR Master Mix (Stratagene) as
previously described (Bae et al., 2006). Each 25 ml reaction contained12.5 ml 26 Brilliant SYBR Green QPCR Master Mix, 5 ml 10-fold
diluted cDNA, 2.5 pM each gene-specific primer and reference dye
(final concentration5300 nM). Reaction conditions were: 95 uC for
10 min; 40 cycles of 95 uC for 30 s, 60 uC for 1 min, 72 uC for 30 s.
Transcript levels (AltMV CP or RdRp) were normalized to actin(copy number relative to actin), a constitutively expressed gene. Mean
values were calculated from four biological replications. After
identification of the dominant variants present in isolate AltMV-SP
(see below), primer pairs RdRp-F(4-7)/RdRp-R(4-7) and RdRp-F(3-1)/RdRp-R(3-1) (Supplementary Table S1) were used in differential
Q-RT-PCR from infected tissues.
Statistical analysis (ANOVA) of Q-RT-PCR results was carried outusing Microsoft Excel to separate reactions into classes that were
significantly different at P50.05.
Identification of sequence variants in AltMV-SP-infected N.
benthamiana. RNA was extracted and cDNA produced as previouslydescribed, from an infection established from tissue stored at 270 uCfor approximately 3 years and multiple transfers before production of
the infectious clones. A PCR product of about 800 bp, spanning the
MluI site junction of the 59 and 39 clones, was amplified with primers
PP3/PP20 (Supplementary Table S1) and cloned; 15 clones weresequenced using primers M13F and M13R (Invitrogen).
Construction of AltMV 3-1:eGFP, 4-1:eGFP, 3-7:DsRed and 4-
7:DsRed. A multiple cloning site (MCS) including NcoI, BamHI,MluI, BglII and NheI restriction sites was inserted upstream of the CP
gene in a 39-terminal subclone of AltMV 3-1 by overlap PCR (Wurch
et al., 1998). The TGB2/TGB3 region was amplified using forward
primer TGB2/3-F introducing a HindIII site (to aid subcloning)
upstream of the nt 5480 XmaI site, and reverse primer MCS-R(Supplementary Table S1) adding NcoI, BamHI, MluI and BglII sites
after the TGB3/CP intergenic region. The CP-39UTR fragment was
amplified with primer MCS-F adding BamHI, MluI, BglII and NheI
sites upstream of the CP gene, and reverse primer XbaI-R1
(Supplementary Table S1). These fragments were combined byamplification using flanking primers TGB2/3-F and XbaI-R1, and
cloned into HindIII/XbaI digested pUC18 yielding pAltMV:MCS. The
CP subgenomic (sg) promoter was duplicated by amplification of a 3-
1 sg promoter/CP/39UTR fragment using NheI-modified primer NheI-
F and reverse primer XbaI-R2 (Supplementary Table S1). The productwas digested with NheI and XbaI, and substituted for the NheI/XbaI
CP-39UTR fragment of pAltMV:MCS to yield pAltMV:MCS:sg. The
eGFP and DsRed genes were amplified from pGDG and pGDR
(Goodin et al., 2002), respectively, introducing 59 NcoI and 39 MluI or
NheI sites (primers eGFP NcoI-F/eGFP MluI-R, DsRed NcoI-F/DsRedNheI-R; Supplementary Table S1), and separately inserted into
pAltMV:MCS:sg. The eGFP-containing XmaI/XbaI 39 fragment was
Determinants of AltMV pathogenicity
http://vir.sgmjournals.org 285

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
substituted into clones 3-1 and 4-1 to form 3-1:eGFP and 4-1:eGFP,
and the DsRed XmaI/XbaI fragment substituted into clones 3-7 and 4-
7, yielding 3-7:DsRed and 4-7:DsRed.
Western blot analysis. AltMV-CP was detected by Western blot
analysis using a 1 : 2000 dilution of AltMV-specific antibody [a gift
from Andrew Geering (Department of Primary Industries,
Queensland Horticultural Institute, Queensland, Australia; Geering
& Thomas, 1999)], alkaline phosphate-conjugated goat anti-rabbit
antibody (Kirkegaard & Perry), and nitro blue tetrazolium (NBT)/5-
bromo-4-chloro-3-indolyl phosphate (BCIP) (Sigma) substrate
(Hammond et al., 2006).
Detection of fluorescence in epidermal cells of N. benthami-
ana. Expression of smGFP in whole plants was detected using a
Fujifilm LAS-1000 imaging system (Fujifilm). Fluorescence in
epidermal cells of N. benthamiana was visualized by laser scanning
confocal microscopy using a Zeiss LSM 410 microscope (Carl Zeiss
MicroImaging). An Argon laser was used to excite eGFP at 488 nm
with the emission band pass detection at 500–520 nm. DsRed was
excited at 543 nm and the emission passed to the PMT detector
through 500–520 nm and LP570 nm emission filters.
Yeast two-hybrid assays. Yeast two-hybrid vectors (James et al.,
1996) were modified to express fusions of the Gal4 activation (AD) or
binding (BD) domains to the N terminus of the TGBL(88) and
TGB1P(88) to test for interactions. Each TGB1 was amplified using the
primers indicated (Supplementary Table S1), and introduced as
BamHI/PstI fragments to the AD (pGAD-TGB1) and the BD
(pGBDU-TGB1) plasmids.
Yeast strain PJ69-4A was transformed with LEU2 selected AD and
URA3 selected BD constructs as previously described (Becker et al.,
1991). Transformants containing both AD and BD plasmids were
selected on SD-glucose medium containing 20 mg Ade, Met, Trp and
His l21, and 30 mg Lys l21, and grown for 3 days at 28 uC. Yeast
colonies were tested for interactions through expression of reporter
genes ADE2 and HIS3 by streaking on SD-glucose medium containing
20 mg Met and Trp l21, 30 mg Lys l21 and grown for 3–5 days at
28 uC or at room temperature.
REFERENCES
Aldaoud, R., Dawson, W. O. & Jones, G. E. (1989). Rapid, random
evolution of the genetic structure of replicating tobacco mosaic virus
populations. Intervirology 30, 227–233.
Bae, H., Kim, M. S., Sicher, R. C., Bae, H. J. & Bailey, B. A. (2006).Necrosis- and ethylene-inducing peptide from Fusarium oxysporum
induces a complex cascade of transcripts associated with signal
transduction and cell death in Arabidopsis. Plant Physiol 141, 1056–
1067.
Bayne, E. H., Rakitina, D. V., Morozov, S. Y. & Baulcombe, D. C.(2005). Cell-to-cell movement of potato potexvirus X is dependent on
suppression of RNA silencing. Plant J 44, 471–482.
Becker, D. M., Fikes, J. D. & Guarente, L. (1991). A cDNA encoding a
human CCAAT-binding protein cloned by functional complementa-
tion in yeast. Proc Natl Acad Sci U S A 88, 1968–1972.
Bragg, J. N. & Jackson, A. O. (2004). The C-terminal region of the
Barley stripe mosaic virus cb protein participates in homologous
interactions and is required for suppression of RNA silencing. Mol
Plant Pathol 5, 465–481.
Chellappan, P., Vanitharani, R., Ogbe, F. & Fauquet, C. M. (2005).Effect of temperature on geminivirus-induced RNA silencing in
plants. Plant Physiol 138, 1828–1841.
Garcia-Arenal, F., Fraile, A. & Malpica, J. M. (2001). Variability and
genetic structure of plant virus populations. Annu Rev Phytopathol 39,157–186.
Geering, A. D. & Thomas, J. E. (1999). Characterisation of a virusfrom Australia that is closely related to papaya mosaic potexvirus.
Arch Virol 144, 577–592.
Goodin, M. M.,Dietzgen, R. G.,Schichnes, D., Ruzin, S. & Jackson, A. O.(2002). pGD vectors: versatile tools for the expression of green and
red fluorescent protein fusions in agroinfiltrated plant leaves. Plant J
31, 375–383.
Hagiwara, K., Ichiki, T. U., Ogawa, Y., Omura, T. & Tsuda, S. (2002).A single amino acid substitution in 126kDa protein of Pepper mildmottle virus associates with symptom attenuation in pepper; the
complete nucleotide sequence of an attenuated strain, C-1421. Arch
Virol 147, 833–840.
Hammond, J., Reinsel, M. D. & Maroon-Lango, C. J. (2006).Identification and full sequence of an isolate of Alternanthera
mosaic potexvirus infecting Phlox stolonifera. Arch Virol 151, 477–493.
James, P., Halladay, J. & Craig, E. A. (1996). Genomic libraries and ahost strain designed for highly efficient two-hybrid selection in yeast.
Genetics 144, 1425–1436.
Jovel, J., Walker, M. & Sanfacon, H. (2007). Recovery of Nicotianabenthamiana plants from a necrotic response induced by a nepovirus
is associated with RNA silencing but not with reduced virus titer.
J Virol 81, 12285–12297.
Kagiwada, S., Yamaji, Y., Komatsu, K., Takahashi, S., Mori, T., Hirata,H., Suzuki, M., Ugaki, M. & Namba, S. (2005). A single amino acidresidue of RNA-dependent RNA polymerase in the Potato virus X
genome determines the symptoms in Nicotiana plants. Virus Res 110,
177–182.
Kaper, J. M., Geletka, L. M., Wu, G. S. & Tousignant, M. E. (1995).Effect of temperature on cucumber mosaic virus satellite-induced
lethal tomato necrosis is helper virus strain dependent. Arch Virol140, 65–74.
Komatsu, K., Yamaji, Y., Ozeki, J., Hashimoto, M., Kagiwada, S.,Takahashi, S. & Namba, S. (2008). Nucleotide sequence analysis
of seven Japanese isolates of Plantago asiatica mosaic virus (PlAMV):
a unique potexvirus with significantly high genomic andbiological variability within the species. Arch Virol 153, 193–
198.
Lewandowski, D. J. & Dawson, W. O. (1993). A single amino acidchange in Tobacco mosaic virus replicase prevents symptom
production. Mol Plant Microbe Interact 6, 157–160.
Liang, X. Z., Lee, B. T. & Wong, S. M. (2002). Covariation in the capsid
protein of hibiscus chlorotic ringspot virus induced by serial
passaging in a host that restricts movement leads to avirulence inits systemic host. J Virol 76, 12320–12324.
Lim, H. S., Ko, T. S., Lambert, K. N., Kim, H. G., Korban, S. S.,Hartman, G. L. & Domier, L. L. (2005). Soybean mosaic virus helpercomponent-protease enhances somatic embryo production and
stabilizes transgene expression in soybean. Plant Physiol Biochem 43,
1014–1021.
Malcuit, I., Marano, M. R., Kavanagh, T. A., De Jong, W., Forsyth, A. &Baulcombe, D. C. (1999). The 25-kDa movement protein of PVX
elicits Nb-mediated hypersensitive cell death in potato. Mol PlantMicrobe Interact 12, 536–543.
Ozeki, J., Takahashi, S., Komatsu, K., Kagiwada, S., Yamashita, K.,Mori, T., Hirata, H., Yamaji, Y., Ugaki, M. & Namba, S. (2006). A single
amino acid in the RNA-dependent RNA polymerase of Plantago
asiatica mosaic virus contributes to systemic necrosis. Arch Virol 151,2067–2075.
H.-S. Lim and others
286 Journal of General Virology 91

Downloaded from www.microbiologyresearch.org by
IP: 54.159.222.200
On: Sat, 28 May 2016 14:26:45
Petty, I. T., Hunter, B. G., Wei, N. & Jackson, A. O. (1989). Infectiousbarley stripe mosaic virus RNA transcribed in vitro from full-lengthgenomic cDNA clones. Virology 171, 342–349.
Qiu, W., Park, J. W. & Scholthof, H. B. (2002). Tombusvirus P19-mediated suppression of virus-induced gene silencing is controlled bygenetic and dosage features that influence pathogenicity. Mol PlantMicrobe Interact 15, 269–280.
Qu, F., Ye, X., Hou, G., Sato, S., Clemente, T. E. & Morris, T. J. (2005).RDR6 has a broad-spectrum but temperature-dependent antiviraldefense role in Nicotiana benthamiana. J Virol 79, 15209–15217.
Rico, P., Ivars, P., Elena, S. F. & Hernandez, C. (2006). Insights intothe selective pressures restricting Pelargonium flower break virusgenome variability: evidence for host adaptation. J Virol 80, 8124–8132.
Schneider, W. L. & Roossinck, M. J. (2001). Genetic diversity in RNAvirus quasispecies is controlled by host-virus interactions. J Virol 75,6566–6571.
Shiboleth, Y. M., Haronsky, E., Leibman, D., Arazi, T., Wassenegger,M., Whitham, S. A., Gaba, V. & Gal-On, A. (2007). The conservedFRNK box in HC-Pro, a plant viral suppressor of gene silencing, isrequired for small RNA binding and mediates symptom development.J Virol 81, 13135–13148.
Siddiqui, S. A., Sarmiento, C., Kiisma, M., Koivumaki, S., Lemmetty,A., Truve, E. & Lehto, K. (2008). Effects of viral silencing suppressors
on tobacco ringspot virus infection in two Nicotiana species. J Gen
Virol 89, 1502–1508.
Silhavy, D. & Burgyan, J. (2004). Effects and side-effects of viral RNA
silencing suppressors on short RNAs. Trends Plant Sci 9, 76–83.
Simon, O., Williams, T., Lopez-Ferber, M. & Caballero, P. (2005).Functional importance of deletion mutant genotypes in an insect
nucleopolyhedrovirus population. Appl Environ Microbiol 71, 4254–
4262.
Verchot-Lubicz, J., Ye, C. M. & Bamunusinghe, D. (2007). Molecular
biology of potexviruses: recent advances. J Gen Virol 88, 1643–1655.
Voinnet, O., Lederer, C. & Baulcombe, D. C. (2000). A viral
movement protein prevents spread of the gene silencing signal in
Nicotiana benthamiana. Cell 103, 157–167.
Wurch, T., Lestienne, F. & Pauwels, P. (1998). A modified overlap
extension PCR method to create chimeric genes in the absence of
restriction enzymes. Biotechnol Tech 12, 653–657.
Yu, H. H. & Wong, S. M. (1998). Synthesis of biologically active cDNA
clones of cymbidium mosaic potexvirus using a population cloning
strategy. Arch Virol 143, 1617–1620.
Determinants of AltMV pathogenicity
http://vir.sgmjournals.org 287




















