
Oxidative metabolism of BDE99 by rat liver microsomes: metabolites formed and CYP enzymes involved
11
TOXICOLOGICAL SCIENCES 123(1), 37–47 (2011) doi:10.1093/toxsci/kfr155 Advance Access publication June 14, 2011 Comparative Oxidative Metabolism of BDE-47 and BDE-99 by Rat Hepatic Microsomes Claudio A. Erratico, 1 Sarah C. Moffatt, 1 and Stelvio M. Bandiera 2 Faculty of Pharmaceutical Sciences, The University of British Columbia, Vancouver, British Columbia, Canada V6T 1Z3 1 These authors contributed equally to this study. 2 To whom correspondence should be addressed at Faculty of Pharmaceutical Sciences, The University of British Columbia, 2146 East Mall, Vancouver, British Columbia, Canada V6T 1Z3. Fax: þ1 (604) 822-3035. E-mail: [email protected]. Received March 4, 2011; accepted June 7, 2011 Polybrominated diphenyl ethers (PBDEs) are flame-retardant chemicals that have become ubiquitous environmental pollutants. 2,2#,4,4#-Tetrabromodiphenyl ether (BDE-47) and 2,2#,4,4#,5- pentabromodiphenyl ether (BDE-99) are among the most prevalent PBDEs detected in humans, wildlife, and abiotic environmental matrices. The purpose of this study was to investigate the oxidative metabolism of BDE-47 and BDE-99 in rat hepatic microsomes by comparing metabolite formation rates, kinetic parameters associ- ated with metabolite formation, and the effects of prototypical cytochrome P450 (CYP) inducers. The CYP enzymes involved were also identified. Incubation of BDE-47 with hepatic microsomes from phenobarbital-treated rats generated a total of five hydroxyl- ated (OH-BDE) metabolites, among which 4#-hydroxy-2,2#,4, 5#-tetrabromodiphenyl ether (4#-OH-BDE-49) and 3-hydroxy-2,2#, 4,4#-tetrabromodiphenyl ether (3-OH-BDE-47) were the major metabolites, as identified using authentic standards and quantified by liquid chromatography/mass spectrometry. Incuba- tions of BDE-99 with hepatic microsomes from dexamethasone- treated rats produced a total of seven hydroxylated metabolites, among which 4-hydroxy-2,2#,3,4#,5-pentabromodiphenyl ether (4-OH-BDE-90) and 6#-hydroxy-2,2#,4,4#,5-pentabromodiphenyl ether (6#-OH-BDE-99) were the major metabolites. Although the overall rate of oxidative metabolism of BDE-99 by hepatic microsomes was greater than that of BDE-47, para-hydroxylation involving a National Institutes of Health shift mechanism represented a major metabolic pathway for both PBDE congeners. Among the rat recombinant CYP enzymes tested, CYP2A2 and CYP3A1 were the most active in BDE-47 and BDE-99 metabolism, respectively. However, CYP1A1 exhibited the highest activity for 4#- OH-BDE-49 and 6#-OH-BDE-99 formation, and CYP3A1 exhibited the highest activity for 3-OH-BDE-47 and 4-OH-BDE- 90 formation. Collectively, the results demonstrate that oxidative metabolism of BDE-47 and BDE-99 is mediated by distinct but overlapping sets of CYP enzymes and represents a key process that determines the bioaccumulation of BDE-47 and BDE-99 in mammals. Key Words: BDE-47; BDE-99; hydroxylated metabolites; hepatic microsomes; cytochrome P450 enzymes. Polybrominated diphenyl ethers (PBDEs), which have been used as flame retardants on a variety of consumer products, have become ubiquitous environmental pollutants. PentaBDE, a commercial PBDE mixture that mainly contains 2,2#,4,4#- tetrabromodiphenyl ether (BDE-47) and 2,2#,4,4#,5-pentabro- modiphenyl ether (BDE-99) (La Guardia et al., 2006), was used predominantly in North America for application to polyurethane foam, textiles, upholstery, and electronic compo- nents (Hale et al., 2003). PBDEs are not chemically bound to the products they are applied to and consequently are released during manufacture, use, and disposal of these products (Hale et al., 2002; Odusanya et al., 2009). As a result, PBDEs have been detected in air, soil, and water and in biota ranging from invertebrates to humans (Hale et al., 2006; Hites, 2004). Although BDE-47 and BDE-99 are present at similar levels in the PentaBDE mixture and in abiotic matrices, concentrations of BDE-99 are lower than those of BDE-47 in biotic samples (Hites, 2004). Different metabolism of BDE-47 versus BDE-99 could account for the greater bioaccumulation of BDE-47 than BDE-99 in biota. In mammals, oxidative metabolism of persistent organic pollutants occurs mainly in the liver and produces polar metabolites that are more easily excreted than the parent compounds. The cytochrome P450 (CYP) enzymes represent the predominant biotransformation system because CYPs catalyze the first step in oxidative metabolism and, therefore, can serve as a significant determinant of bioaccumulation of pollutants such as PBDEs. Oxidative metabolism is of toxicological interest because recent studies have reported that hydroxylated PBDE (OH-BDE) metabolites have greater biological activity than the parent compounds. For example, several OH-BDEs exhibit higher in vitro binding affinities for rat transthyretin (Hamers et al., 2008) and estrogen receptor (Meerts et al., 2001; Mercado-Feliciano and Bigsby, 2008) than PBDE congeners and also inhibit estradiol sulfotransferase in rat hepatic microsomes (Hamers et al., 2008). These adverse effects can contribute to the hormonal, reproductive, and Ó The Author 2011. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For permissions, please email: [email protected] at The University of British Colombia Library on September 7, 2011 toxsci.oxfordjournals.org Downloaded from
-
Upload
uantwerpen -
Category
Documents
-
view
0 -
download
0
Transcript of Oxidative metabolism of BDE99 by rat liver microsomes: metabolites formed and CYP enzymes involved

TOXICOLOGICAL SCIENCES 123(1), 37–47 (2011)
doi:10.1093/toxsci/kfr155
Advance Access publication June 14, 2011
Comparative Oxidative Metabolism of BDE-47 and BDE-99 by RatHepatic Microsomes
Claudio A. Erratico,1 Sarah C. Moffatt,1 and Stelvio M. Bandiera2
Faculty of Pharmaceutical Sciences, The University of British Columbia, Vancouver, British Columbia, Canada V6T 1Z3
1These authors contributed equally to this study.2To whom correspondence should be addressed at Faculty of Pharmaceutical Sciences, The University of British Columbia, 2146 East Mall, Vancouver,
British Columbia, Canada V6T 1Z3. Fax: þ1 (604) 822-3035. E-mail: [email protected].
Received March 4, 2011; accepted June 7, 2011
Polybrominated diphenyl ethers (PBDEs) are flame-retardant
chemicals that have become ubiquitous environmental pollutants.
2,2#,4,4#-Tetrabromodiphenyl ether (BDE-47) and 2,2#,4,4#,5-pentabromodiphenyl ether (BDE-99) are among themost prevalent
PBDEs detected in humans, wildlife, and abiotic environmental
matrices. The purpose of this study was to investigate the oxidative
metabolism of BDE-47 and BDE-99 in rat hepatic microsomes by
comparing metabolite formation rates, kinetic parameters associ-
ated with metabolite formation, and the effects of prototypical
cytochrome P450 (CYP) inducers. TheCYP enzymes involvedwere
also identified. Incubation of BDE-47 with hepatic microsomes
from phenobarbital-treated rats generated a total of five hydroxyl-
ated (OH-BDE) metabolites, among which 4#-hydroxy-2,2#,4,5#-tetrabromodiphenyl ether (4#-OH-BDE-49) and 3-hydroxy-2,2#,4,4#-tetrabromodiphenyl ether (3-OH-BDE-47) were the
major metabolites, as identified using authentic standards and
quantified by liquid chromatography/mass spectrometry. Incuba-
tions of BDE-99 with hepatic microsomes from dexamethasone-
treated rats produced a total of seven hydroxylated metabolites,
among which 4-hydroxy-2,2#,3,4#,5-pentabromodiphenyl ether
(4-OH-BDE-90) and 6#-hydroxy-2,2#,4,4#,5-pentabromodiphenyl
ether (6#-OH-BDE-99) were the major metabolites. Although the
overall rate of oxidative metabolism of BDE-99 by hepatic
microsomes was greater than that of BDE-47, para-hydroxylation
involving a National Institutes of Health shift mechanism
represented a major metabolic pathway for both PBDE congeners.
Among the rat recombinant CYP enzymes tested, CYP2A2 and
CYP3A1 were the most active in BDE-47 and BDE-99 metabolism,
respectively.However,CYP1A1exhibited the highest activity for 4#-OH-BDE-49 and 6#-OH-BDE-99 formation, and CYP3A1
exhibited the highest activity for 3-OH-BDE-47 and 4-OH-BDE-
90 formation. Collectively, the results demonstrate that oxidative
metabolism of BDE-47 and BDE-99 is mediated by distinct but
overlapping sets of CYP enzymes and represents a key process that
determines the bioaccumulation of BDE-47 and BDE-99 in
mammals.
Key Words: BDE-47; BDE-99; hydroxylated metabolites;
hepatic microsomes; cytochrome P450 enzymes.
Polybrominated diphenyl ethers (PBDEs), which have been
used as flame retardants on a variety of consumer products,
have become ubiquitous environmental pollutants. PentaBDE,
a commercial PBDE mixture that mainly contains 2,2#,4,4#-tetrabromodiphenyl ether (BDE-47) and 2,2#,4,4#,5-pentabro-
modiphenyl ether (BDE-99) (La Guardia et al., 2006), was
used predominantly in North America for application to
polyurethane foam, textiles, upholstery, and electronic compo-
nents (Hale et al., 2003). PBDEs are not chemically bound to
the products they are applied to and consequently are released
during manufacture, use, and disposal of these products (Hale
et al., 2002; Odusanya et al., 2009). As a result, PBDEs have
been detected in air, soil, and water and in biota ranging from
invertebrates to humans (Hale et al., 2006; Hites, 2004).
Although BDE-47 and BDE-99 are present at similar levels in
the PentaBDE mixture and in abiotic matrices, concentrations
of BDE-99 are lower than those of BDE-47 in biotic samples
(Hites, 2004). Different metabolism of BDE-47 versus BDE-99
could account for the greater bioaccumulation of BDE-47 than
BDE-99 in biota.
In mammals, oxidative metabolism of persistent organic
pollutants occurs mainly in the liver and produces polar
metabolites that are more easily excreted than the parent
compounds. The cytochrome P450 (CYP) enzymes represent
the predominant biotransformation system because CYPs
catalyze the first step in oxidative metabolism and, therefore,
can serve as a significant determinant of bioaccumulation of
pollutants such as PBDEs. Oxidative metabolism is of
toxicological interest because recent studies have reported that
hydroxylated PBDE (OH-BDE) metabolites have greater
biological activity than the parent compounds. For example,
several OH-BDEs exhibit higher in vitro binding affinities for
rat transthyretin (Hamers et al., 2008) and estrogen receptor
(Meerts et al., 2001; Mercado-Feliciano and Bigsby, 2008)
than PBDE congeners and also inhibit estradiol sulfotransferase
in rat hepatic microsomes (Hamers et al., 2008). These adverse
effects can contribute to the hormonal, reproductive, and
� The Author 2011. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved.For permissions, please email: [email protected]
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

neurobehavioral toxicity observed in laboratory animals treated
with PBDEs (Kuriyama et al., 2005; Viberg et al., 2002; Zhou
et al., 2001).
Accumulated evidence from several studies demonstrates
that PBDEs are biotransformed to hydroxylated metabolites in
mammals. OH-BDEs were identified in blood samples of
women and children who were environmentally exposed to
PBDEs (Athanasiadou et al., 2008; Qiu et al., 2009). In
addition, hydroxylated metabolites have been detected in bile,
urine, and feces of rats and mice treated with BDE-47, BDE-
99, or PBDE mixtures (Chen et al., 2006; Hakk et al., 2002;
Malberg et al., 2005; Marsh et al., 2006; Orn and Klasson-
Wehler, 1998; Qiu et al., 2007; Staskal et al., 2006). More
recently, formation of hydroxylated metabolites of BDE-47 and
BDE-99 by rat and human hepatic preparations has been
reported (Dong et al., 2010; Erratico et al., 2010; Hamers et al.,2008; Lupton et al., 2009, 2010; Stapleton et al., 2009). For
example, three hydroxylated metabolites were detected when
BDE-99 was incubated with rat or human hepatocytes for 72 h
(Dong et al., 2010; Stapleton et al., 2009). Lupton et al. (2010)
detected two hydroxylated metabolites of BDE-47 and BDE-
99, following a 2-h incubation with human liver microsomes,
whereas Hamers et al. (2008) identified six hydroxylated
metabolites of BDE-47, following a 90-min incubation with rat
liver microsomes. Rates of metabolite formation were not
measured, and the CYP enzymes involved in metabolite
formation were not determined in these studies. Characteriza-
tion of hepatic BDE-47 and BDE-99 metabolism in vitro can
provide a better assessment of the role of metabolism as
a determinant of BDE-47 and BDE-99 bioaccumulation,
persistence, and toxicity.
The present study was conducted to characterize CYP-
mediated biotransformation of BDE-47 and BDE-99 by rat
hepatic microsomes. Using our recently developed liquid
chromatography (LC)/mass spectrometry (MS)-based assays
(Erratico et al., 2010; Moffatt et al., 2011), we quantified rates
of OH-BDE metabolite formation, measured kinetic parameters
associated with metabolite formation, and determined the
effects of prototypical CYP inducers. In addition, the hepatic
CYP enzymes responsible for BDE-47 and BDE-99 metabolite
formation were determined using a panel of recombinant CYP
enzymes.
MATERIALS AND METHODS
Chemicals and reagents. BDE-47 (neat, 98.8% or greater purity) and
BDE-99 (neat, 97.7% or greater purity) were purchased from Chiron
AS (Trondheim, Norway). 4-Hydroxy-2,2#,3,4#-tetrabromodiphenyl ether
(4-OH-BDE-42), 3-hydroxy-2,2#,4,4#-tetrabromodiphenyl ether (3-OH-BDE-
47), 5-hydroxy-2,2#,4,4#-tetrabromodiphenyl ether (5-OH-BDE-47),
6-hydroxy-2,2#,4,4#-tetrabromodiphenyl ether (6-OH-BDE-47), 4#-hydroxy-
2,2#,4,5#-tetrabromodiphenyl ether (4#-OH-BDE-49), 4-hydroxy-2,2#,3,4#,5-pentabromodiphenyl ether (4-OH-BDE-90), 5#-hydroxy-2,2#,4,4#,5-pentab
romodiphenyl ether (5#-OH-BDE-99), 6#-hydroxy-2,2#,4,4#,5-pentabromodi-
phenyl ether (6#-OH-BDE-99), 4#-hydroxy-2,2#,4,5,5#-pentabromodiphenyl
ether (4#-OH-BDE-101), and 2,4,5-tribromophenol (2,4,5-TBP) (10 or 50 lg/ml
in acetonitrile, 97.7% grade purity or higher) were obtained from AccuStandard
(New Haven, CT). 4#-Hydroxy-2,2#,4,6#-tetrachlorobiphenyl (4-OH-CB-50)
(neat, 99.9% purity) and 4-hydroxy-2#,3,4#,5,6#-pentachlorobiphenyl
(4-OH-CB-121) (neat, 100% purity), which served as the internal standards
for BDE-47 and BDE-99 biotransformation assay, respectively, were also
purchased from AccuStandard. 2-Hydroxy-2#,3,4,4#,5-pentabromodiphenyl
ether (2-OH-BDE-123; neat) was a generous gift from Dr R.J. Letcher
(Environment Canada, Ottawa, Ontario, Canada). Sodium phenobarbital (PB),
dexamethasone (DEX), and 3-methylcholanthrene (MC) were purchased from
Sigma-Aldrich Canada (Oakville, Ontario, Canada). Hydrochloric acid, sodium
hydroxide, and organic solvents (high performance liquid chromatography
grade or better) were purchased from Fisher Scientific (Ottawa, Ontario,
Canada). Ultra-pure water was obtained using a Millipore Milli-Q system
(Billerica, MA). Baculovirus-insect cell microsomes containing expressed rat CYP
enzyme (CYP1A1, CYP1A2, CYP2A1, CYP2A2, CYP2B1, CYP2C6, CYP2C11,
CYP2C12, CYP2C13, CYP2D1, CYP2D2, CYP2E1, CYP3A1, or CYP3A2)
coexpressed with rat CYP oxidoreductase or with rat CYP oxidoreductase and rat
cytochrome b5 (BD Supersomes) were purchased from BD Biosciences (Oakville,
Ontario, Canada). Baculovirus-insect cell control microsomes containing expressed
rat CYP oxidoreductase and rat cytochrome b5 were also purchased from BD
Biosciences.
Animal treatment and preparation of hepatic microsomes. Male Long-
Evans rats (7–8 weeks of age) were purchased from Charles River Canada
(Saint-Constant, Quebec, Canada). Rats were housed in pairs on corncob
bedding in polycarbonate cages (The Anderson’s, Maumee, OH) with free
access to water and food (Laboratory Rodent Diet; PMI Feeds Inc., Richmond,
IN). Animal quarters were maintained at a constant temperature (23�C) with
controlled light (14 h) and dark (10 h) cycles. Rats were cared for in accordance
with the principles and guidelines of the Canadian Council on Animal Care.
Rats (n ¼ 6–7) were treated with CYP inducers as follows: PB (dissolved in
PBS, 80 mg/kg/day), MC (dissolved in corn oil, 25 mg/kg/day), DEX
(dissolved in corn oil, 100 mg/kg/day), or vehicle (corn oil, 2 ml/kg/day). PB,
MC, and DEX are well-characterized inducers of CYP enzymes and were
chosen because they preferentially induce CYP2B, CYP1A, or CYP3A
enzymes, respectively, in rats, under the dosing regimen described (Edwards
et al., 2007; Hrycay and Bandiera, 2003; Ryan and Levin, 1990). Compounds
were administered by intraperitoneal injection for three consecutive days and
rats were killed by decapitation 24 h after the last treatment. Microsomes were
prepared from pooled livers as described previously (Thomas et al., 1983).
Microsomal pellets were suspended in 0.25M sucrose and aliquots were stored
at �80�C, until needed. Protein concentration was measured by the method of
Lowry et al. (1951) using bovine serum albumin as a standard. Total CYP
concentration and ethoxyresorufin dealkylase, benzyloxyresorufin dealkylase,
and benzyloxyquinoline dealkylase activities of the microsomal preparations
were measured, as reported in the Supplementary data.
BDE biotransformation assay. In vitro biotransformation assays for BDE-
47 and BDE-99 were performed as previously described (Erratico et al., 2010;
Moffatt et al., 2011). Briefly, reaction mixtures contained BDE-47 or BDE-99
(2.5–200lM), hepatic microsomal protein (0.1, 0.5, or 1.0 mg), and 50mM
potassium phosphate buffer containing 3mM magnesium chloride (pH 7.4) in
a final volume of 0.99 ml in a screw cap tube. Reaction mixtures were
preincubated for 5 min in a shaking water bath. Reactions were initiated by
addition of 0.01 ml of nicotinamide adenine dinucleotide 2’-phosphate reduced,
tetrasodium salt hydrate (NADPH) solution (1mM final concentration) and
terminated after 5 min for BDE-47 or 10 min for BDE-99 by addition of 1 ml
of ice-cold 0.5M sodium hydroxide. A fixed amount (10 ll) of internal standard
was then added to each tube (final concentration 5.0lM for 4-OH-CB-50
or 3.0lM for 4-OH-CB-121). Tubes were vortex mixed for 1 min and
subsequently heated for 10 min in a water bath at 70�C. After cooling to room
temperature, 2 ml of 6M HCl and 1 ml of isopropanol were added to each tube.
Tubes were vortex mixed again for 1 min. An aliquot (2.0 ml) of a mixture of
methyl tert-butyl ether:hexane (1:1 vol/vol) was then added to each tube. Tubes
38 ERRATICO, MOFFATT, AND BANDIERA
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

were vortex mixed for 1 min and then spun in a centrifuge at 2500 revolutions
per minute for 5 min. The top organic layer was transferred to a set of clean
tubes and the extraction procedure was repeated two more times. The three
organic solvent extracts were pooled, dried under nitrogen, reconstituted in 250
ll of methanol, and filtered through a syringe filter (0.45 lm, polytetrafluoro-
ethylene membrane) into 300-ll HPLC vials.
Blank and negative control samples were included in each assay. Blank
samples did not contain substrate and NADPH, whereas negative control
samples were devoid of either NADPH or substrate. To determine if metabolite
formation was enzyme mediated, experiments were conducted using heat-
denatured microsomes. Hepatic microsome samples were boiled for 5 min in
assay buffer prior to use. To determine if metabolite formation was CYP
mediated, carbon monoxide, which is a classic nonspecific CYP inhibitor, was
bubbled for 2 min into an incubation mixture containing assay buffer,
microsomes, and NADPH prior to use.
Preliminary experiments were conducted to determine the most active
microsomal preparation for each substrate. This microsomal preparation was
then used to confirm that both substrate and cofactor concentrations used were
saturating. Samples were prepared in duplicate and three independent
experiments were conducted.
Incubations with rat recombinant CYP enzymes were carried as described
above except that reaction mixtures contained 30 pmoles of recombinant CYP
enzyme instead of rat hepatic microsomes. Insect cell control microsomes
containing expressed rat CYP oxidoreductase and cytochrome b5 were used as
blank controls at an equivalent amount of protein (0.30 mg). To ensure that
product formation was linear with respect to incubation time and CYP
concentration, preliminary experiments were performed using recombinant
CYP3A1 enzyme. Samples were prepared singly and three independent
experiments were performed.
Analytical methods. Formation of hydroxylated BDE metabolites was
analyzed by LC/MS as described previously (Erratico et al., 2010; Moffatt et al.,
forthcoming). Briefly, OH-BDE metabolites were resolved using a Waters
Acquity UPLC BEH C18 (2.1 3 100 mm, 1.7 lm) column and a Waters Acquity
Ultra Performance Liquid Chromatograph System (UPLC; Waters Corp.,
Milford, MA) consisting of a Binary Solvent Manager and Sample Manager.
To extend the validated method for OH-penta-BDEs to two newly available
standards (4#-OH-BDE-101 and 2-OH-BDE-123), a slightly modified elution
gradient was used in which the composition of the mobile phase (water:-
methanol) was increased linearly from 35:65 (vol/vol) to 25.5:74.5 (vol/vol)
between 0 and 38 min. At 38.1 min, methanol was increased to 100% and
maintained for 2 min. The column was then re-equilibrated with water:methanol
35:65 (vol/vol) for 3 min. Flow rate was maintained at 0.2 ml/min. Metabolites
were detected using a Waters Quattro Premier XE triple quadrupole mass
spectrometer (Waters Corp.) and quantified using calibration curves prepared as
previously described (Erratico et al., 2010; Moffatt et al., forthcoming). The mass
spectrometer was operated in negative electrospray ionization mode using single
ion recording mode. Waters MassLynx v4.1 software (Waters Corp.) was used
for data acquisition. Metabolites were identified by comparison of their retention
times and mass to charge ratio (m/z) values with those of authentic standards. The
following m/z values were used: OH-tri-BDEs m/z 421.3, OH-tetra-BDEs m/z500.3, OH-penta-BDEs m/z 578.4, and 2,4,5-TBP m/z 328.7. Because authentic
standards for di-OH-BDEs are not commercially available, m/z values used
to monitor the formation of di-OH-BDEs (di-OH-tetra-BDEs m/z 516.2 and di-
OH-penta-BDEs m/z 594.5) were derived from exact mass values and the
ionization system used. Internal standards for the BDE-47 and BDE-99
biotransformation assays (4-OH-CB-50 and 4-OH-CB-121, respectively) were
monitored at m/z 306.8 and 340.8, respectively. It should be noted that
the present assay was unable to detect nonhydroxylated metabolites or 2,
4-dibromophenol.
Quality control. Along with each set of unknown samples, several quality
control samples were freshly prepared to assess linearity, accuracy, precision,
and recovery values of the assays. To assess linearity, a calibration curve was
prepared by spiking hepatic microsomes (at the same protein concentration as
that used when measuring biotransformation) with OH-BDE authentic
standards. The acceptance criterion for linearity was R2 > 0.95. Accuracy,
precision, and recovery rates were determined using quality control samples
obtained by spiking hepatic microsomes with authentic standards at 30, 150,
and 700nM for OH-tetra-BDEs, at 25 and 500nM for OH-penta-BDEs, and at
0.250 and 5.0lM for 2,4,5-TBP. The acceptance criterion for accuracy was
percent deviation within ±20% of the nominal value, that for precision was
percent relative standard deviation <20%, and that for recovery rates was
values between 80 and 120%. All the acceptance criteria were assessed on an
interday basis. System suitability samples were also used to monitor the
performance of the UPLC/MS system as previously described (Erratico et al.,
2010; Moffatt et al., forthcoming).
Data analysis. Data were analyzed using the SigmaPlot Enzyme Kinetics
Module (v.1.1; Systat Software Inc., Richmond, CA). Metabolite formation as
a function of substrate concentration was analyzed by nonlinear regression
analysis. Apparent Km, K#, and Vmax values were generated using the
Michaelis–Menten equation (Equation 1), the Hill equation (Equation 2), or the
substrate inhibition kinetic equation (Equation 3).
v ¼ Vmax½S�Km þ ½S� ; ð1Þ
v ¼ Vmax½S�n
K# þ ½S�n ; ð2Þ
v ¼ Vmax
1þ Km=½S� þ ½S�=Ki; ð3Þ
where v is initial velocity of the reaction, Vmax is the maximal velocity, [S] is
the substrate concentration, K# is the Hill dissociation constant, n is the Hill
coefficient representing cooperativity in the reaction, Km is the Michaelis–
Menten constant, and Ki is the dissociation constant of substrate binding to the
inhibitory site. The goodness of the fit was assessed according to the following
criteria: residual analysis, coefficient of determination (R2), and Akaike
Information Criterion.
RESULTS
Kinetic Analysis of BDE-47 and BDE-99 Metabolite Formation
Biotransformation of BDE-47 and BDE-99 was assessed
using hepatic microsomes prepared from rats pretreated with
PB or DEX, respectively, because of the low metabolic activity
obtained with hepatic microsomes from corn oil-treated rats, as
discussed in greater detail in the next section.
Formation of hydroxylated metabolites of BDE-47 was
evaluated over a range of substrate concentrations (10–100lM)
using hepatic microsomes from PB-treated rats, an incubation
time of 5 min and a microsomal protein concentration of 1 mg/ml.
Under these conditions, metabolite formation was directly
proportional to incubation time and protein concentration (Moffatt
et al., 2011). Five hydroxylated metabolites, identified as 4-OH-
BDE-42, 3-OH-BDE-47, 5-OH-BDE-47, 6-OH-BDE-47, and 4#-OH-BDE-49 were formed. A representative chromatogram
showing the hydroxylated metabolites of BDE-47 is presented
in the Supplementary Figure 1. Formation of OH-tri-BDEs or di-
OH-tetra-BDEs was not observed. To further characterize hepatic
microsomal BDE-47 biotransformation, metabolite formation was
quantified and analyzed with respect to kinetic parameters. As
shown in Figure 1, formation of 4-OH-BDE-42, 3-OH-BDE-47,
COMPARATIVE OXIDATIVE METABOLISM OF BDE-47 AND BDE-99 39
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

and 4#-OH-BDE-49 exhibited sigmoidal kinetic profiles and
nonlinear Eadie–Hofstee plots, which signify atypical enzyme
kinetics. Formation of 5-OH-BDE-47 and 6-OH-BDE-47 was
below the limit of quantification at all substrate concen-
trations examined. Apparent K# and Vmax values were calcu-
lated for hepatic microsomal 4-OH-BDE-42, 3-OH-BDE-47, and
4#-OH-BDE-49 formation using the Hill equation (Equation 2).
A Hill coefficient value (n) >2, which is indicative of positive
cooperativity, was obtained for all BDE-47 metabolites quantified
(Fig. 1). The apparent Vmax value for 4#-OH-BDE-49 formation
was approximately 2.5 and seven times higher than the values for
3-OH-BDE-47 and 4-OH-BDE-42 formation, respectively, in-
dicating that 4#-OH-BDE-49 was the major hydroxylated
metabolite of BDE-47 produced by hepatic microsomes from
PB-treated rats. However, apparent K# values were similar
suggesting that all three metabolites would be produced at low
BDE-47 concentrations.
Formation of hydroxylated metabolites of BDE-99 was
similarly evaluated over a range of substrate concentrations
(2.5–200lM) using hepatic microsomes from DEX-treated
rats, an incubation time of 10 min and a microsomal protein
concentration of 0.1 mg/ml. Metabolite formation was
determined previously (Erratico et al., 2010) to be linearly
related to incubation time and protein concentration under
these conditions. Incubation of BDE-99 with liver micro-
somes from DEX-treated rats yielded seven hydroxylated
metabolites, six of which were identified as 2,4,5-TBP,
4-OH-BDE-90, 5#-OH-BDE-99, 6#-OH-BDE-99, 4#-OH-
BDE-101, and 2-OH-BDE-123. A seventh metabolite peak
(M1), corresponding to a OH-penta-BDE based on its m/z value,
was detected but could not be identified because its retention time
did not match that of any of the authentic standards. A repre-
sentative chromatogram showing the hydroxylated metabolites of
BDE-99 is presented in the Supplementary Figure 1. Metabolite
peaks corresponding to OH-tetra-BDEs or di-OH-penta-BDEs
were not detected. Formation of 4-OH-BDE-90, 5#-OH-BDE-99,
6#-OH-BDE-99, and M1 (due to the lack of an authentic standard,
the plot of M1 formation was prepared using relative peak area)
exhibited Michaelis–Menten kinetics, but linear Eadie–Hoftsee
plots were only obtained for 6#-OH-BDE-99 and M1 formation.
Nonlinear Eadie–Hoftsee plots were obtained for 4-OH-BDE-90
and 5#-OH-BDE-99 formation (Fig. 2). Formation of 2,4,5-TBP
was best described by a substrate inhibition kinetic model (Equa-
tion 3). Kinetic analysis of 4#-OH-BDE-101 and 2-OH-BDE-123
formation was not conducted because 4#-OH-BDE-101 and
2-OH-BDE-123 were formed in quantifiable amounts only at
substrate concentrations of 50lM or higher (data not shown).
Apparent Km and Vmax values for hepatic microsomal formation
of 4-OH-BDE-90, 5#-OH-BDE-99, 6#-OH-BDE-99, and M1
were calculated using the Michaelis–Menten equation (Equa-
tion 1), and apparent Km, Ki, and Vmax values for 2,4,5-TBP
formation were calculated using substrate inhibition equation
(Equation 3). The apparent Vmax value for 4-OH-BDE-90
formation was approximately 20 times higher than that for
5#-OH-BDE-99 or 6#-OH-BDE-99 formation and 46 times
greater than that for 2,4,5-TBP formation, indicating that 4-OH-
BDE-90 was the major hydroxylated metabolite of BDE-99
produced by hepatic microsomes from DEX-treated rats (Fig. 1).
Apparent Km values associated with the formation of 2,4,5-TBP
FIG. 1. Enzyme kinetic profiles of 4-OH-BDE-42 (A), 3-OH-BDE-47 (B),
and 4#-OH-BDE-49 (C) formation by rat hepatic microsomes. Rate of
metabolite formation was plotted as a function of substrate concentration
following a 5-min incubation of BDE-47 with hepatic microsomes (1.0 mg
protein/ml) from PB-treated rats. Data points are the mean ± SD of three
separate experiments. Lines represent rates of metabolite formation modeled by
nonlinear regression analysis. The insets depict Eadie–Hofstee plots. Error bars
are not shown on the insets to avoid obscuring the data points, which represent
mean values.
40 ERRATICO, MOFFATT, AND BANDIERA
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

and 6#-OH-BDE-99 were lower than those for 4-OH-BDE-90 and
5#-OH-BDE-99 (Fig. 1), suggesting that 2,4,5-TBP and 6#-OH-
BDE-99 metabolites would be produced preferentially at low
BDE-99 concentrations.
The aggregate BDE-47 and BDE-99 metabolite yields, which
were estimated using Vmax values for quantifiable hydroxylated
metabolites, indicate that approximately 0.05% of BDE-47 and
0.5% of BDE-99 were converted to hydroxylated metabolites/
min/mg protein, under in vitro conditions. This calculation
indicates that BDE-47 and BDE-99 were metabolized relatively
slowly, and that BDE-99 undergoes hepatic microsomal
oxidative biotransformation more readily than BDE-47.
Metabolite formation was not observed when BDE-47 or
BDE-99 was incubated with boiled microsomal preparations,
indicating that metabolite formation was enzyme mediated. In
addition, metabolite formation was not observed when
NADPH or substrate (BDE-47 or BDE-99) was omitted from
the reaction mixture or when carbon monoxide was bubbled for
2 min into an incubation mixture prior to addition of substrate,
suggesting that metabolite formation was CYP mediated.
Effect of CYP Inducers on BDE-47 and BDE-99Biotransformation
To investigate the influence of CYP induction on oxidative
biotransformation of BDE-47 and BDE-99, rates of BDE-47
and BDE-99 metabolite formation were determined using
hepatic microsomes prepared from rats pretreated with corn oil,
DEX, MC, or PB. Based on the enzyme kinetic results reported
above, saturating substrate concentrations (BDE-47 50lM or
BDE-99 100lM) were used. Distinct metabolite profiles were
obtained for BDE-47 (Table 1) and BDE-99 (Table 2) with
each hepatic microsomal preparation. BDE-47 metabolites
were not detected with hepatic microsomes from corn oil-
treated rats even when the microsomal preparation was
incubated with BDE-47 for 60 min at 2 mg protein/ml. Two
metabolites, 4#-OH-BDE-49 and 4-OH-BDE-42, were detected
following incubation with hepatic microsomes from MC-
treated rats, but only 4#-OH-BDE-49 could be quantified
because 4-OH-BDE-42 was formed at a concentration below
the limit of quantification. Three metabolites, 3-OH-BDE-47,
5-OH-BDE-47, and 4#-OH-BDE-49, were formed at quantifi-
able levels by hepatic microsomes from DEX-treated rats, and
four metabolites, 4-OH-BDE-42, 3-OH-BDE-47, 5-OH-BDE-
47, and 4#-OH-BDE-49, were produced by hepatic microsomes
from PB-treated rats (Table 1). Under the experimental
FIG. 2. Enzyme kinetic profiles of 2,4,5-TBP (A), 4-OH-BDE-90 (B),
5#-OH-BDE-99 (C), 6#-OH-BDE-99 (D), and M1 (E) formation by rat hepatic
microsomes. Rate of metabolite formation was plotted as a function of substrate
concentration following a 10-min incubation of BDE-99 with hepatic
microsomes (0.1 mg protein/ml) from DEX-treated rats. Data points are the
mean ± SD of three separate experiments. Lines represent rates of metabolite
formation modeled by nonlinear regression analysis. The insets depict Eadie–
Hofstee plots. Error bars are not shown on the insets to avoid obscuring the data
points, which represent mean values.
COMPARATIVE OXIDATIVE METABOLISM OF BDE-47 AND BDE-99 41
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

conditions used, 3-OH-BDE-47 and 4#-OH-BDE-49 were the
major metabolites produced by hepatic microsomes from DEX-
and PB-treated rats, respectively.
In comparison, three metabolites, 4-OH-BDE-90, 5#-OH-
BDE-99, and 6#-OH-BDE-99, were detected and quantified
when BDE-99 was incubated with hepatic microsomes from
corn oil- or MC-treated rats. The major metabolite produced
by hepatic microsomes from corn oil- or MC-treated rats was 4-
OH-BDE-90 or 6#-OH-BDE-99, respectively (Table 2). Seven
metabolites, 2,4,5-TBP, 4-OH-BDE-90, 5#-OH-BDE-99, 6’-
OH-BDE-99, 4’-OH-BDE-101, 2-OH-BDE-123, and M1, were
detected using hepatic microsomes from DEX- or PB-treated
rats. The major metabolite produced by both hepatic microsomal
preparations was 4-OH-BDE-90. On the basis of relative peak
areas, M1 formation was greater in hepatic microsomes from
DEX- than PB-treated rats and it appears to be a quantitatively
important metabolite in both microsomal preparations.
Biotransformation Activities of Recombinant CYP Enzymes
A panel of 14 recombinant rat CYP enzymes was used to
identify the individual CYP enzymes involved in the bio-
transformation of BDE-47 and BDE-99 (Fig. 3). Experiments
were initially conducted using CYP3A1 and a saturating
substrate concentration (50lM for BDE-47 or 100lM for
BDE-99) to ensure linearity of product formation with respect
to incubation time and CYP concentration. An incubation time
of 10 min and a recombinant CYP enzyme concentration of 30
pmol/ml were found to be optimal. Hydroxylated metabolites
were not detected when BDE-47 or BDE-99 was incubated
with control insect cell microsomes containing expressed rat
CYP oxidoreductase without CYP enzymes.
As shown in Figure 3A, relatively few recombinant rat CYP
enzymes were involved in the oxidative biotransformation of
BDE-47. Formation of 4-OH-BDE-42 and 6-OH-BDE-47 was
catalyzed solely by CYP2A2, whereas 3-OH-BDE-47 forma-
tion was mediated exclusively by CYP3A1. Formation of
5-OH-BDE-47 was catalyzed at a high rate by CYP2A2 and at
a lower rate by CYP3A1. Formation of 4#-OH-BDE-49 was
catalyzed by CYP1A1, CYP2B1, and CYP3A1. Under the
experimental conditions used, CYP2A2 was the most active
recombinant CYP enzyme in the biotransformation of BDE-47
and 5-OH-BDE-47 was the major metabolite produced.
TABLE 1
Effect of Treatment with CYP Inducers on Formation of Hydroxylated Metabolites of BDE-47 by Rat Hepatic Microsomes
Treatment
Rate of metabolite formation (pmol/min/mg protein)
4-OH-BDE-42 3-OH-BDE-47 5-OH-BDE-47 6-OH-BDE-47 4#-OH-BDE-49
Corn oil N.D.a N.D. N.D. N.D. N.D.
DEX N.D. 3.6 ± 0.3 1.6 ± 0.1 N.D. 1.5 ± 0.2
MC <LOQb N.D. N.D. N.D. 11 ± 2.2
PB 3.6 ± 0.2 6.8 ± 0.1 1.3 ± 0.04 <LOQ 17 ± 0.6
Note. Rat hepatic microsomes (1 mg/ml) were incubated with BDE-47 at saturating substrate concentrations (50lM) for 5 min as described in the Materials and
Methods section. Values represent the mean ± SD of three independent experiments.aN.D., not detected.b<LOQ, below limit of quantification.
TABLE 2
Effect of Treatment with CYP Inducers on Formation of Hydroxylated Metabolites of BDE-99 by Rat Hepatic Microsomes
Treatment
Rate of metabolite formation (pmol/min/mg protein)
2,4,5-TBP 4-OH-BDE-90 5#-OH-BDE-99 6#-OH-BDE-99 4#-OH-BDE-101 2-OH-BDE-123
Corn oil N.D.a 2.7 ± 1.6 0.3 ± 0.1 0.6 ± 0.4 N.D. N.D.
DEX 8.2 ± 2.4 323 ± 51.6 18 ± 3.8 12 ± 3.7 3.6 ± 1.1 3.7 ± 0.6
MC N.D. 6.0 ± 3.6 1.9 ± 0.7 20 ± 4.0 N.D. N.D.
PB 21 ± 1.4 76.5 ± 16 27 ± 5.4 27 ± 3.7 9.0 ± 1.8 7.5 ± 1.0
Note. Rat hepatic microsomes were incubated with BDE-99 at saturating substrate concentrations (100lM) for 10 min. A protein concentration of 0.5 mg/ml was
used with hepatic microsomes from corn oil-treated rats and a protein concentration of 0.1 mg/ml was used with hepatic microsomes from DEX-, MC-, or PB-
treated rats, as described in the Materials and Methods section. Values represent the mean ± SD of three independent experiments. M1 was detected in hepatic
microsomes from DEX- or PB-treated rats only. Rates of M1 formation could not be expressed as pmol/min/mg protein because of the lack of an authentic
standard. Using peak area count values, the rate of M1 formation was estimated to be eight times higher in hepatic microsomes from DEX-treated than PB-treated
rats.aN.D., not detected.
42 ERRATICO, MOFFATT, AND BANDIERA
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from
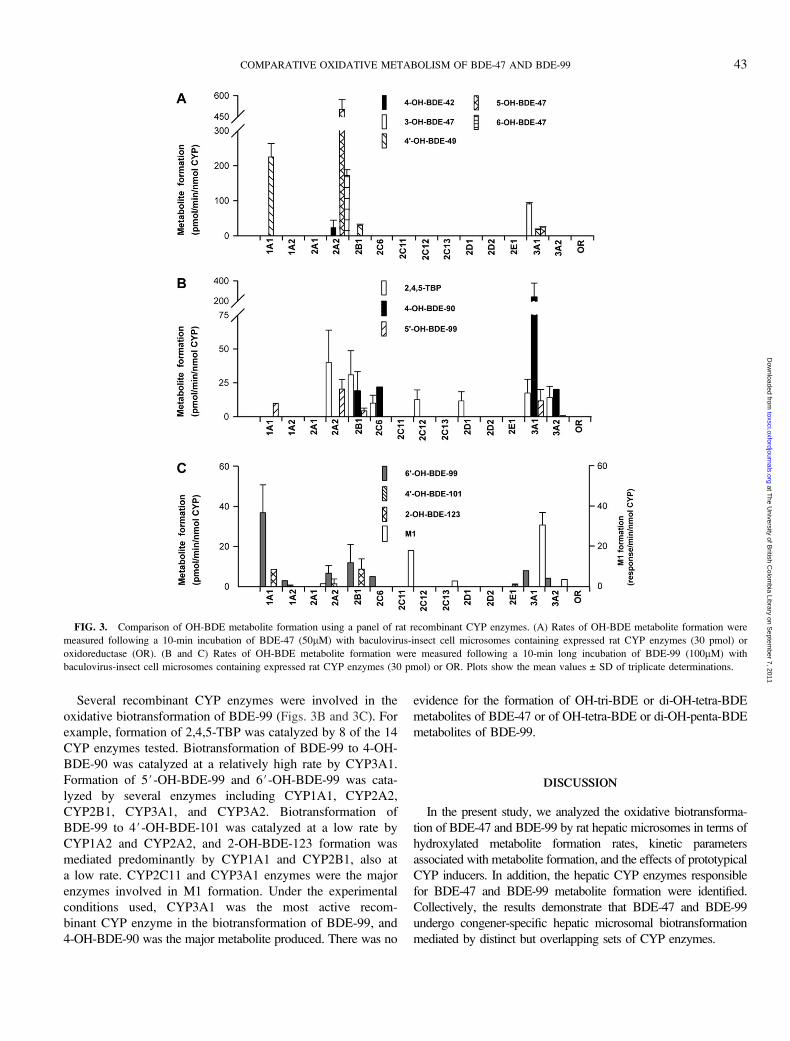
Several recombinant CYP enzymes were involved in the
oxidative biotransformation of BDE-99 (Figs. 3B and 3C). For
example, formation of 2,4,5-TBP was catalyzed by 8 of the 14
CYP enzymes tested. Biotransformation of BDE-99 to 4-OH-
BDE-90 was catalyzed at a relatively high rate by CYP3A1.
Formation of 5#-OH-BDE-99 and 6#-OH-BDE-99 was cata-
lyzed by several enzymes including CYP1A1, CYP2A2,
CYP2B1, CYP3A1, and CYP3A2. Biotransformation of
BDE-99 to 4#-OH-BDE-101 was catalyzed at a low rate by
CYP1A2 and CYP2A2, and 2-OH-BDE-123 formation was
mediated predominantly by CYP1A1 and CYP2B1, also at
a low rate. CYP2C11 and CYP3A1 enzymes were the major
enzymes involved in M1 formation. Under the experimental
conditions used, CYP3A1 was the most active recom-
binant CYP enzyme in the biotransformation of BDE-99, and
4-OH-BDE-90 was the major metabolite produced. There was no
evidence for the formation of OH-tri-BDE or di-OH-tetra-BDE
metabolites of BDE-47 or of OH-tetra-BDE or di-OH-penta-BDE
metabolites of BDE-99.
DISCUSSION
In the present study, we analyzed the oxidative biotransforma-
tion of BDE-47 and BDE-99 by rat hepatic microsomes in terms of
hydroxylated metabolite formation rates, kinetic parameters
associated with metabolite formation, and the effects of prototypical
CYP inducers. In addition, the hepatic CYP enzymes responsible
for BDE-47 and BDE-99 metabolite formation were identified.
Collectively, the results demonstrate that BDE-47 and BDE-99
undergo congener-specific hepatic microsomal biotransformation
mediated by distinct but overlapping sets of CYP enzymes.
FIG. 3. Comparison of OH-BDE metabolite formation using a panel of rat recombinant CYP enzymes. (A) Rates of OH-BDE metabolite formation were
measured following a 10-min incubation of BDE-47 (50lM) with baculovirus-insect cell microsomes containing expressed rat CYP enzymes (30 pmol) or
oxidoreductase (OR). (B and C) Rates of OH-BDE metabolite formation were measured following a 10-min long incubation of BDE-99 (100lM) with
baculovirus-insect cell microsomes containing expressed rat CYP enzymes (30 pmol) or OR. Plots show the mean values ± SD of triplicate determinations.
COMPARATIVE OXIDATIVE METABOLISM OF BDE-47 AND BDE-99 43
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from
Among the BDE-47 metabolites detected, 4#-OH-BDE-49
was the major metabolite produced, as determined by rate of
formation, in hepatic microsomes from MC- and PB-treated
rats, and 3-OH-BDE-47 was the major metabolite produced in
hepatic microsomes from DEX-treated rats. These results
suggest that CYP1A and CYP2B enzymes, which are highly
induced by treatment with MC and PB (Edwards et al., 2007),
respectively, are involved in the hydroxylation of BDE-47 to
4#-OH-BDE-49, whereas CYP3A enzymes, which are induced
by treatment with DEX (Hrycay and Bandiera, 2003), play
a major role in 3-OH-BDE-47 formation. Among the BDE-99
metabolites detected, 4-OH-BDE-90 was the major metabolite
in hepatic microsomes from corn oil-, DEX-, and PB-pretreated
rats, whereas 6#-OH-BDE-99 was the major metabolite in
hepatic microsomes from MC-treated rats. The results suggest
that CYP2B and CYP3A enzymes are involved in 4-OH-BDE-
90 formation, whereas CYP1A enzymes are involved in
6#-OH-BDE-99 formation. CYP2B and CYP3A enzymes are
likely involved in the formation of 2,4,5-TBP, 4#-OH-BDE-101,
2-OH-BDE-123, and M1, as these metabolites were detected
only in hepatic microsomes from PB- and DEX-treated rats. The
contribution of individual CYP enzymes to hydroxylated
metabolite formation was confirmed by experiments with rat
recombinant CYP enzymes. More specifically, among the 14 CYP
enzymes tested, CYP1A1 and CYP3A1 exhibited the highest
activity for 4#-OH-BDE-49 and 3-OH-BDE-47 formation, re-
spectively, from BDE-47 (Fig. 3A). High rate of 4#-OH-BDE-49
formation in liver microsomes from PB-treated rats was due to
CYP2B1 and CYP3A1 activities (Fig. 3A), as PB is known to
induce CYP2B1 and CYP3A1 in rats (Hrycay and Bandiera
2003). The lack of formation of 4#-OH-BDE-49 in liver
microsomes from corn oil-treated rats is due to the low level
of expression of CYP1A1, CYP2B1, and CYP3A1 enzymes in
uninduced rat liver microsomes (Hrycay and Bandiera, 2003).
With BDE-99 as the substrate, CYP3A1 and CYP1A1 were the
most efficient catalysts of 4-OH-BDE-90 and 6-OH-BDE-99
formation, respectively (Figs. 3B and 3C).
Experiments with the recombinant CYP enzymes provided
additional insights into congener-specific differences in CYP-
mediated oxidation of BDE-47 and BDE-99. For example,
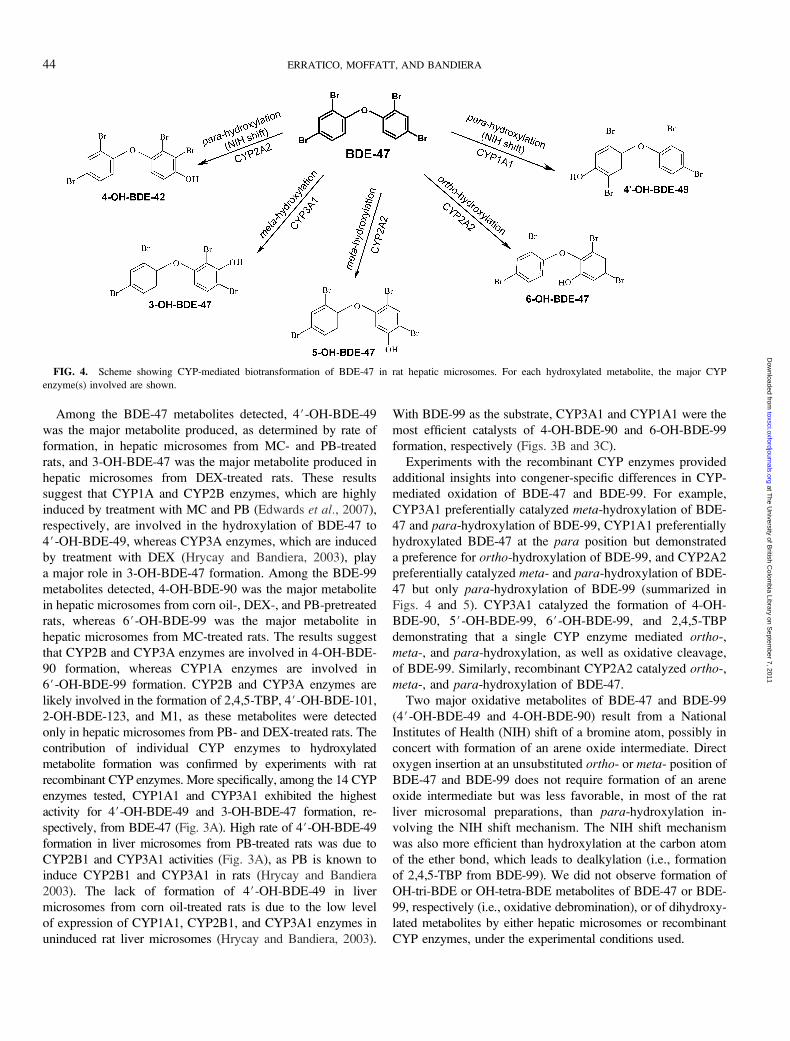
CYP3A1 preferentially catalyzed meta-hydroxylation of BDE-
47 and para-hydroxylation of BDE-99, CYP1A1 preferentially
hydroxylated BDE-47 at the para position but demonstrated
a preference for ortho-hydroxylation of BDE-99, and CYP2A2
preferentially catalyzed meta- and para-hydroxylation of BDE-
47 but only para-hydroxylation of BDE-99 (summarized in
Figs. 4 and 5). CYP3A1 catalyzed the formation of 4-OH-
BDE-90, 5#-OH-BDE-99, 6#-OH-BDE-99, and 2,4,5-TBP
demonstrating that a single CYP enzyme mediated ortho-,
meta-, and para-hydroxylation, as well as oxidative cleavage,
of BDE-99. Similarly, recombinant CYP2A2 catalyzed ortho-,
meta-, and para-hydroxylation of BDE-47.
Two major oxidative metabolites of BDE-47 and BDE-99
(4#-OH-BDE-49 and 4-OH-BDE-90) result from a National
Institutes of Health (NIH) shift of a bromine atom, possibly in
concert with formation of an arene oxide intermediate. Direct
oxygen insertion at an unsubstituted ortho- or meta- position of
BDE-47 and BDE-99 does not require formation of an arene
oxide intermediate but was less favorable, in most of the rat
liver microsomal preparations, than para-hydroxylation in-
volving the NIH shift mechanism. The NIH shift mechanism
was also more efficient than hydroxylation at the carbon atom
of the ether bond, which leads to dealkylation (i.e., formation
of 2,4,5-TBP from BDE-99). We did not observe formation of
OH-tri-BDE or OH-tetra-BDE metabolites of BDE-47 or BDE-
99, respectively (i.e., oxidative debromination), or of dihydroxy-
lated metabolites by either hepatic microsomes or recombinant
CYP enzymes, under the experimental conditions used.
FIG. 4. Scheme showing CYP-mediated biotransformation of BDE-47 in rat hepatic microsomes. For each hydroxylated metabolite, the major CYP
enzyme(s) involved are shown.
44 ERRATICO, MOFFATT, AND BANDIERA
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

The present study indicates that oxidative metabolism of
PBDEs by rat hepatic microsomes is relatively slow compared
with many other xenobiotics such as drugs and polyaromatic
hydrocarbons (pmoles vs. nmoles of product formed/min/mg
protein). The rate and extent of biotransformation of halogenated
aromatic compounds such as polychlorinated biphenyls and
PBDEs was thought to be dependent on the number and position
of halogen atoms in the molecule, so that the greater the number
of halogens, the slower the rate of metabolism (Bandiera, 2001).
In addition, the availability of neighboring unsubstituted carbon
atoms on one ring, especially at the meta- and para- positions,
was thought to facilitate metabolism (Bandiera, 2001). The
results of the present study are inconsistent with this paradigm
because oxidative metabolism of a penta-brominated diphenyl
ether (i.e., BDE-99) by rat hepatic microsomes was greater, in
terms of metabolite formation rates, than that of tetra-brominated
diphenyl ether (i.e., BDE-47). Moreover, hydroxylation of
the tri-brominated phenyl ring of BDE-99 was favored over
the dibrominated ring, and hydroxylation at the substituted
para position of BDE-99 was favored over the unsubstituted
ortho- and meta- positions.
The lower rate of BDE-47 metabolism by rat hepatic
microsomes compared with BDE-99 (Tables 1 and 2) can be
partially explained by differences in the CYP enzymes
involved (Fig. 3). The rate of xenobiotic metabolism depends
on the microsomal level and the intrinsic catalytic activity of
the individual CYP enzymes involved. Our data show that
BDE-47 was mainly metabolized by CYP1A1 and CYP2A2 at
a relatively high rate (Fig. 3A). However, CYP1A1 and
CYP2A2 together represent approximately 5%, or less, of total
CYP content in adult male rat liver microsomes (Thummel
et al., 1988). Although CYP1A1 is extensively induced by MC
treatment (Hrycay and Bandiera, 2003), CYP2A2 is not
induced by MC, PB, or DEX treatments (Matsunaga et al.,1988; Thummel et al., 1988). Conversely, BDE-99 is
metabolized by several CYP enzymes including CYP2B,
CYP2C, and CYP3A enzymes, which together represent 45–
70% of the total CYP content in adult male rat liver
microsomes (Bandiera, 2001; Ryan and Levin, 1990).
Furthermore, CYP3A1, which is induced by PB or DEX
treatment, exhibits higher intrinsic activity toward BDE-99
than BDE-47 (approximately 400 vs. 60 pmol/min/mg protein;
Fig. 3). Thus, differences in hepatic levels of individual CYP
enzymes can account for differences in hepatic microsomal
metabolite profiles and metabolite formation rates for BDE-47
and BDE-99. The lower in vivo excretion rates of BDE-99
compared with BDE-47 reported in mice (Staskal et al., 2006)
support the role of hepatic metabolism in explaining the
difference in bioaccumulation between BDE-99 and BDE-47 in
mammals (Hites, 2004).
The BDE-47 and BDE-99 metabolites quantified in the
present study are consistent with previous in vitro and in vivometabolism studies. All five hydroxylated metabolites of BDE-
47 detected in our study were identified as in vitro metabolites
of BDE-47 produced by rat hepatic microsomes (Hamers et al.,2008) and as in vivo metabolites of rats exposed to BDE-47 by
oral administration (Marsh et al., 2006). We did not observe
formation of 2#-OH-BDE-66 or of OH-tri-BDE metabolites of
BDE-47, which were identified in the study by Marsh et al.(2006). Of the seven hydroxylated metabolites of BDE-99
detected in our study, only two (i.e., 2,4,5-TBP and 5#-OH-
BDE-99) were identified in recent in vitro studies involving rat
and human hepatocytes incubated with BDE-99 (Dong et al.,2010; Stapleton et al., 2009). OH-BDE and di-OH-BDE
FIG. 5. Scheme showing CYP-mediated biotransformation of BDE-99 in rat hepatic microsomes. For each hydroxylated metabolite, the major CYP
enzyme(s) involved are shown.
COMPARATIVE OXIDATIVE METABOLISM OF BDE-47 AND BDE-99 45
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

metabolites of BDE-47 and BDE-99 were also reported in
earlier in vivo rodent studies (Chen et al., 2006; Hakk et al.,2002; Staskal et al., 2006), but the lack of structure elucidation
precludes comparison with the OH-BDE metabolites identified
in our study. Differences in hepatic preparations and the
incubation conditions used may account for discrepancies in
metabolite profiles between our study and other studies.
Several of the OH-BDE metabolites identified in our study
are toxicologically relevant because in vitro and in vivo studies
have shown that they have greater biological activity than the
parent compound. For example, 4-OH-BDE-42, 3-OH-BDE-
47, and 4#-OH-BDE-49 exhibited greater potency than BDE-
47 in displacing 17b-estradiol from the estrogen receptor
(Mercado-Feliciano and Bigsby, 2008), inhibiting aromatase
and 17b-estradiol sulfotransferase activities (Canton et al.,2008; Hamers et al., 2008), binding to transthyretin (Hamers
et al., 2008), and altering intracellular calcium homeostasis
(Dingemans et al., 2008). Another metabolite of BDE-47, 6-
OH-BDE-47, was found to be acutely toxic in developing and
adult zebrafish through uncoupling oxidative phosphorylation
(Van Boxtel et al., 2008). In addition, 4-OH-BDE-90, 5#-OH-
BDE-99, and 6#-OH-BDE-99 exhibited significantly higher
thyroid hormone activities than BDE-99 or other PBDEs (Li
et al., 2010). Thus, oxidative metabolism can enhance PBDE
toxicity and should be an important consideration in the
toxicological risk assessment of PBDEs.
In conclusion, the oxidative metabolism of BDE-47 and
BDE-99 was thoroughly characterized in vitro, and the CYP
enzymes catalyzing the formation of BDE-47 and BDE-99
metabolites in rat liver microsomes were determined for the
first time. BDE-99 undergoes more extensive CYP-mediated
metabolism than BDE-47 because of the metabolic involve-
ment of a larger number of constitutively expressed CYP
enzymes that have higher catalytic activity for BDE-99 than
BDE-47. Extrapolation of our in vitro findings suggests that
BDE-99 is more likely to undergo hepatic CYP-mediated
oxidation in vivo, consistent with the relatively lower levels of
BDE-99 found in mammals. The present study strongly
suggests that CYP-mediated metabolism is a major determinant
of the bioaccumulation and toxicity of BDE-47 and BDE-99 in
rats and, possibly, in other mammalian species.
SUPPLEMENTARY DATA
Supplementary data are available online at http://toxsci.
oxfordjournals.org/.
FUNDING
Natural Sciences and Engineering Research Council of Canada
(RGPIN 138733-01 to S.M.B.); A graduate student fellowship
from The University of British Columbia to C.A.E.
ACKNOWLEDGMENTS
The authors thank Mr Patrick Edwards for under-
taking development of the BDE-47 biotransformation assay,
Mr Andras Szeitz for technical help with the LC/MS analysis,
and Dr Robert J. Letcher, Environment Canada, for providing
the 2-OH-BDE-123 standard.
REFERENCES
Athanasiadou, M., Cuadra, S. N., Marsh, G., Bergman, A., and Jakobsson, K.
(2008). Polybrominated diphenyl ethers (PBDEs) and bioaccumulative
hydroxylated PBDE metabolites in young humans from Managua,
Nicaragua. Environ. Health Perspect. 116, 400–408.
Bandiera, S. M. (2001). Cytochrome P450 enzymes as biomarkers of PCB
exposure and modulators of toxicity. In PCBs: Recent Advances in
Environmental Toxicology and Health Effects (L. W. Robertson and
L. G. Hansen, Eds.), pp. 185–192. Kentucky University Press, Lexington,
KY.
Canton, R. F., Scholten, D. E., Marsh, G., de Jong, P. C., and van den Berg, M.
(2008). Inhibition of human placental aromatase activity by hydroxylated
polybrominated diphenyl ethers (OH-PBDEs). Toxicol. Appl. Pharmacol.
227, 68–75.
Chen, L. J., Lebetkin, E. H., Sanders, J. M., and Burka, L. T. (2006).
Metabolism and disposition of 2,2’,4,4’,5-pentabromodiphenyl ether
(BDE99) following a single or repeated administration to rats or mice.
Xenobiotica 36, 515–534.
Dingemans, M. M., de Groot, A., van Kleef, R. G., Bergman, A., van den
Berg, M., Vijverberg, H. P., and Westerink, R. H. (2008). Hydroxylation
increases the neurotoxic potential of BDE-47 to affect exocytosis and
calcium homeostasis in PC12 cells. Environ. Health Perspect. 116, 637–643.
Dong, H., Li, Z., Man, X., Zhou, J., Lu, H., and Wang, S. (2010). Identification
of the metabolites of polybrominated diphenyl 99 and its related P450s. J.
Biomed. Res. 24, 223–232.
Edwards, P. R., Hrycay, E. G., and Bandiera, S. M. (2007). Differential
inhibition of hepatic microsomal alkoxyresorufin O-dealkylation activities by
tetrachlorobiphenyls. Chem. Biol. Interact. 169, 42–52.
Erratico, C. A., Szeitz, A., and Bandiera, S. M. (2010). Validation of a novel
in vitro assay using ultra performance liquid chromatography-mass
spectrometry (UPLC/MS) to detect and quantify hydroxylated metabolites
of BDE-99 in rat liver microsomes. J. Chromatogr. B Analyt. Technol.
Biomed. Life Sci. 878, 1562–1568.
Hakk, H., Larsen, G., and Klasson-Wehler, E. (2002). Tissue disposition,
excretion and metabolism of 2,2’,4,4’,5-pentabromodiphenyl ether (BDE-99)
in the male Sprague-Dawley rat. Xenobiotica 32, 369–382.
Hale, R. C., Alaee, M., Manchester-Neesvig, J. B., Stapleton, H. M., and
Ikonomou, M. G. (2003). Polybrominated diphenyl ether flame retardants in
the North American environment. Environ. Int. 29, 771–779.
Hale, R. C., La Guardia, M. J., Harvey, E., Gaylor, M. O., and Mainor, T. M.
(2006). Brominated flame retardant concentrations and trends in abiotic
media. Chemosphere 64, 181–186.
Hale, R. C., La Guardia, M. J., Harvey, E., and Mainor, T. M. (2002). Potential
role of fire retardant-treated polyurethane foam as a source of brominated
diphenyl ethers to the U.S. environment. Chemosphere 46, 729–735.
Hamers, T., Kamstra, J. H., Sonneveld, E., Murk, A. J., Visser, T. J., Van
Velzen, M. J., Brouwer, A., and Bergman, A. (2008). Biotransformation of
brominated flame retardants into potentially endocrine-disrupting metabo-
lites, with special attention to 2,2’,4,4’-tetrabromodiphenyl ether (BDE-47).
Mol. Nutr. Food Res. 52, 284–298.
46 ERRATICO, MOFFATT, AND BANDIERA
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from

Hites, R. A. (2004). Polybrominated diphenyl ethers in the environment and in
people: a meta-analysis of concentrations. Environ. Sci. Technol. 38,
945–956.
Hrycay, E. G., and Bandiera, S. M. (2003). Spectral interactions of
tetrachlorobiphenyls with hepatic microsome P450 enzymes. Chem. Biol.Interact. 146, 285–296.
Kuriyama, S. N., Talsness, C. E., Grote, K., and Chahoud, I. (2005).
Developmental exposure to low dose PBDE 99: effects on male fertility and
neurobehavior in rat offspring. Environ. Health Perspect. 113, 149–154.
La Guardia, A. M. J., Hale, R. C., and Harvey, E. (2006). Detailed
polybrominated diphenyl ether (PBDE) congener composition of the widely
used penta-, octa-, and deca-PBDE technical flame-retardant mixtures.
Environ. Sci. Technol. 40, 6247–6254.
Li, F., Xie, Q., Li, X., Li, N., Chi, P., Chen, J., Wang, Z., and Hao, C. (2010).
Hormone activity of hydroxylated polybrominated diphenyl ethers on human
thyroid receptor-beta: in vitro and in silico investigations. Environ. Health
Perspect. 118, 602–606.
Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951). Protein
measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Lupton, S. J., McGarrigle, B. P., Olson, J. R., Wood, T. D., and Aga, D. S.
(2009). Human liver microsome-mediated metabolism of brominated
diphenyl ethers 47, 99, and 153 and identification of their major metabolites.
Chem. Res. Toxicol. 22, 1802–1809.
Lupton, S. J., McGarrigle, B. P., Olson, J. R., Wood, T. D., and Aga, D. S.
(2010). Analysis of hydroxylated polybrominated diphenyl ether metabolites
by liquid chromatography/atmospheric pressure chemical ionization tandem
mass spectrometry. Rapid Commun. Mass Spectrom. 24, 2227–2235.
Malberg, T., Athanasiadou, M., Marsh, G., Brandt, I., and Bergman, A. (2005).
Identification of hydroxylated polybrominated diphenyl ether metabolites in
blood plasma from polybrominated diphenyl ether exposed rats. Environ.
Sci. Technol. 39, 5342–5348.
Marsh, G., Athanasiadou, M., Athanassiadis, I., and Sandholm, A. (2006).
Identification of hydroxylated metabolites in 2,2’,4,4’-tetrabromodiphenyl
ether exposed rats. Chemosphere 63, 690–697.
Matsunaga, T., Nagata, K., Holsztynska, E. J., Lapenson, D. P., Smith, A.,
Kato, R., Gelboin, H. V., Waxman, D. J., and Gonzales, F. J. (1988). Gene
conversion and differential regulation in the rat P-450 IIA gene subfamily. J.
Biol. Chem. 263, 17995–18002.
Meerts, I. A., Letcher, R. J., Hoving, S., Marsh, G., Bergman, A.,
Lemmen, J. G., van der Burg, B., and Brouwer, A. (2001). In vitroestrogenicity of polybrominated diphenyl ethers, hydroxylated PDBEs, and
polybrominated bisphenol A compounds. Environ. Health Perspect. 109,
399–407.
Mercado-Feliciano, M., and Bigsby, R. M. (2008). Hydroxylated metabolites of
the polybrominated diphenyl ether mixture DE-71 are weak estrogen
receptor-alpha ligands. Environ. Health Perspect. 116, 1315–1321.
Moffatt, S., Edwards, P. R., Szeitz, A., and Bandiera, S. M. (2011). A validated
liquid chromatography-mass spectrometry method for the detection and
quantification of oxidative metabolites of 2,2#,4,4#-tetrabromodiphenyl ether
in rat hepatic microsomes. Am. J. Anal. Chem. in press. Available at: http://
www.scirp.org/journal/ajac.
Odusanya, D. O., Okonkwo, J. O., and Botha, B. (2009). Polybrominated
diphenyl ethers (PBDEs) in leachates from selected landfill sites in South
Africa. Waste Manag. 29, 96–102.
Orn, U., and Klasson-Wehler, E. (1998). Metabolism of 2,2’,4,4’-tetrabromo-
diphenyl ether in rat and mouse. Xenobiotica 28, 199–211.
Qiu, X., Bigsby, R. M., and Hites, R. A. (2009). Hydroxylated metabolites of
polybrominated diphenyl ethers in human blood samples from the United
States. Environ. Health Perspect. 117, 93–98.
Qiu, X., Mercado-Feliciano, M., Bigsby, R. M., and Hites, R. A. (2007).
Measurement of polybrominated diphenyl ethers and metabolites in mouse
plasma after exposure to a commercial pentabromodiphenyl ether mixture.
Environ. Health Perspect. 115, 1052–1058.
Ryan, D., and Levin, W. (1990). Purification and characterization of hepatic
microsomal cytochrome P-450. Pharmacol. Ther. 45, 153–239.
Stapleton, H. M., Kelly, S. M., Pei, R., Letcher, R. J., and Gunsch, C. (2009).
Metabolism of polybrominated diphenyl ethers (PBDEs) by human
hepatocytes in vitro. Environ. Health Perspect. 117, 197–202.
Staskal, D. F., Hakk, H., Bauer, D., Diliberto, J. J., and Birnbaum, L. S. (2006).
Toxicokinetics of polybrominated diphenyl ether congeners 47, 99, 100, and
153 in mice. Toxicol. Sci. 94, 28–37.
Thomas, P. E., Reik, L. M., Ryan, D. E., and Levin, W. (1983). Induction of
two immunochemically related rat liver cytochrome P-450 isozymes,
cytochromes P-450c and P-450d, by structurally diverse xenobiotics. J.
Biol. Chem. 258, 4590–4598.
Thummel, K. E., Favreau, L. V., Mole, J. E., and Schenkman, J. B. (1988).
Further characterization of RLM2 and comparison with a related form of
cytochrome P-450, RLM2B. Arch. Biochem. Biophys. 266, 319–333.
Van Boxtel, A. L., Kamstra, J. H., Cenijn, P. H., Pieterse, B., Wagner, M. J.,
Antink, M., Krab, K., Van den Burg, B., Marsh, G., Brouwer, A., et al.
(2008). Microarray analysis reveals a mechanism of phenolic polybromi-
nated diphenylether toxicity in zebrafish. Environ. Sci. Technol. 42,
1772–1779.
Viberg, H., Fredriksson, A., and Eriksson, P. (2002). Neonatal exposure to the
brominated flame retardant 2,2’,4,4’,5-pentabromodiphenyl ether causes
altered susceptibility in the cholinergic transmitter system in the adult mouse.
Toxicol. Sci. 67, 104–107.
Zhou, T., Ross, D. G., DeVito, M. J., and Crofton, K. M. (2001). Effects of
short-term in vivo exposure to polybrominated diphenyl ethers on thyroid
hormones and hepatic enzyme activities in weanling rats. Toxicol. Sci. 61,
76–82.
COMPARATIVE OXIDATIVE METABOLISM OF BDE-47 AND BDE-99 47
at The U
niversity of British C
olombia Library on S
eptember 7, 2011
toxsci.oxfordjournals.orgD
ownloaded from




















