
The Cycadofilicales, they formed the dominant fossil plants ...
85
UNIT-2 BSC I YEAR BOTANY-II Page 1 Cycadeoideales Introduction to Cycadeoideales: The Cycadofilicales, they formed the dominant fossil plants during Palaeozoic age. The Cycadofilicales have of course definite affinities with the cycads on one side and ferns on the other, but they had no cones either in the male or in the female part of the plants, so some workers think that the Cycadofilicales form a separate group quite distinct from gymnosperms. In the Mesozoic times, however, we came across fossils plants which had cones and were definitely related to gymnosperms. So in Mesozoic the Cycadofilicales were replaced by true gymnosperms which formed strobili, and the seeds had a naked dicotyledonous embryo in them. The ovule or the seed was never enclosed in closed carpel. The Mesozoic gymnosperms can be placed into two separate groups: 1. Cycadeoidale (Bennettitales) and 2. Cycadales. Pant (1957) has placed the cycadeiods in a distinct class, the Cycadeoideopsida of the division Cycadophyta. The Cycadeoideales (Bennettitales) first appeared in the Permian they reached their highest range during the Jurassic period, after which they disappeared altogether. The second group Cycadales had a world-wide distribution during the Mesozoic period Majority of them had altogether disappeared; only a few types have been left which are confined to special parts of the East. The present day cycads are only the remnants of very large dyeing out group, i.e., they are sometimes described as living fossils, because they are on their way to extinction. The Cycadeoideales (Bennettitales) were very much like the cycads in their general appearance, and as the Mesozoic had these two prominent groups of gymnosperms, so that period sometimes described as age of cycads. These Cycadeoideales are closely related to the Cycadofilicales on one side and to cycads on the other but they have their own characteristic features which distinguish them from all other gymnosperms except the Gnetales. The important feature which separates the Cycadeoideales from other gymnosperms is the presence of bisporangiate strobili.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of The Cycadofilicales, they formed the dominant fossil plants ...

UNIT-2
BSC I YEAR BOTANY-II Page 1
Cycadeoideales
Introduction to Cycadeoideales:
The Cycadofilicales, they formed the dominant fossil plants during Palaeozoic
age. The Cycadofilicales have of course definite affinities with the cycads on
one side and ferns on the other, but they had no cones either in the male or in
the female part of the plants, so some workers think that the Cycadofilicales
form a separate group quite distinct from gymnosperms.
In the Mesozoic times, however, we came across fossils plants which had cones
and were definitely related to gymnosperms. So in Mesozoic the
Cycadofilicales were replaced by true gymnosperms which formed strobili, and
the seeds had a naked dicotyledonous embryo in them. The ovule or the seed
was never enclosed in closed carpel.
The Mesozoic gymnosperms can be placed into two separate groups:
1. Cycadeoidale (Bennettitales) and
2. Cycadales.
Pant (1957) has placed the cycadeiods in a distinct class, the Cycadeoideopsida
of the division Cycadophyta.
The Cycadeoideales (Bennettitales) first appeared in the Permian they reached
their highest range during the Jurassic period, after which they disappeared
altogether.
The second group Cycadales had a world-wide distribution during the Mesozoic
period Majority of them had altogether disappeared; only a few types have been
left which are confined to special parts of the East. The present day cycads are
only the remnants of very large dyeing out group, i.e., they are sometimes
described as living fossils, because they are on their way to extinction.
The Cycadeoideales (Bennettitales) were very much like the cycads in their
general appearance, and as the Mesozoic had these two prominent groups of
gymnosperms, so that period sometimes described as age of cycads.
These Cycadeoideales are closely related to the Cycadofilicales on one side and
to cycads on the other but they have their own characteristic features which
distinguish them from all other gymnosperms except the Gnetales. The
important feature which separates the Cycadeoideales from other gymnosperms
is the presence of bisporangiate strobili.

UNIT-2
BSC I YEAR BOTANY-II Page 2
The plants of this group were diversified in their habit. Some types had short
columnar stems like most of the living cycads. The short columnar stem was
usually un-branched and at the apex of the plant there was a terminal crown of
leaves which in most cases pinnate. Some other forms had branched stems with
multiple crown.
In present day cycads we know that young leaves and megasporophylls are
covered up by unicellular hairy outgrowths known as ramenta.
In Cycadeoideales (Bennettitales) these ramenta were not unicellular; they were
scale like, flattened and were several cells in breadth. Like cycads the plants had
well organized strobili or cones, but in cycads they are monosporangiate
whereas in Cycadeoideales they were usually bisporangiate and they were either
terminal or axiarlly in position.
Majority of Cycadeoideales (Bennettitales) seem to have flowered only once in
their life and after flowering the plant died out as we find in some of present day
angiosperms.
Classification of Cycadeoideales:
According to Sporne (1965), the order Cycadeoideales (Bennettitales) has been
divided into three families.
They are:
1. Cycadeoideaceae.
2.Williamsoniaceae, and
3. Wielandiellaceae.
Here Cycadeoidea (Bennettites) of Cycadeoideaceae and Williamsonia of
Williamsoinaceae have been discussed in detail.
Systematic Position of Cycadeoideales:
Gymnosperms
Class. Cycadopsida
Order. Cycadeoideales
Family. Cycadeoideaceae

UNIT-2
BSC I YEAR BOTANY-II Page 3
Genus. Cycadeoidea
Bennettites, by American workers have been described as Cycadeoidea.
Features of Cycadeoideales:
(A) Morphological features:
In Cycadeoidea the stem was un-branched with a single crown of pinnate leaves
at the tops, but some species had branched stem with a multiple crown. In some
the stem was tuberous. In all cases the stem was covered up by persistent leaf
bases as we find in Cycas.

UNIT-2
BSC I YEAR BOTANY-II Page 4

UNIT-2
BSC I YEAR BOTANY-II Page 5
Bennettitales:
Meaning of Bennettitales:
This group of fossil plants flourished well during the Triassic to Lower
Cretaceous periods of Mesozoic era. As the Carboniferous period is called
the “Ages of Ferns “, the Mesozoic era is called the „Ages of Cycads‟. It is due
to the fact that Cycadeoideales co-existed with Cycadales during Mesozoic era
from Jurassic up to Cretaceous period, and hence this period is called Age of
Cycads.
Bennettitales are found either in the form of compressions or petrifactions. Due
to Cycad-like form of their fronds and the presence of short stems covered with
an armour of presistent leaf bases Bennettitales (Cycadeoideales) have been
treated under Cycadophyta by some workers. However, the two groups are quite
distinct from each other and maintain their independent identity.
2. Origin of Bennettitales:
Palaeobotanists believe that Bennettitales originated from Pteridospermales. But
presence of stalked ovules in Bennettitalean members is a strong evidence
against such a theory.
Some workers have, however, shown resemblance between the fronds of
Pteridospermales and the stalked ovules and inter-seminal scales of
Bennettitales. But it is hard to imagine that stalked ovules and inter-seminal
scales are homologous with the fronds.
Discovery of a fossil plant, Westersheimia, from the Triassic of Austria, is again
a step towards the possible Pteridospermean ancestry of Bennettitales. This

UNIT-2
BSC I YEAR BOTANY-II Page 6
genus occurs along with Bennetticarpus, the seed-bearing organs of
Bennettitales.
In Bennetticarpus wettsteinii and Westersheimia the ovules and inter-seminal
scales were present on the ultimate segments of a pinnate structure. But it is still
not clear how the entire group of Bennettitales evolved from Pteridospermales.
Some peculiar characteristics present in Bennettitales and not in any other
group of gymnosperms, include:
(i) Bisporangiate strobili,
(ii) Synangium-bearing fused microsporophyll‟s,
(iii) Close occurrence of ovules and inter-seminal scales, and
(iv) Production of stalked ovules.
Distinguishing Features of Bennettitales:
1. These extinct Mesozoic plants were present were present on the earth from
Triassic to Cretaceous.
2. Bennettitales were so abundant during Mesozoic era that this period is known
as „Age of Cycads‟.
3. The members of this group are found either as compressions or petrifactions.
4. The stems were stout or slender and had a wide pith.
5. The stem grew very slowly and had manoxylic wood.
6. Resembling living Cycads, the Bennettitalean leaves were mostly pinnately
compound, and only occasionally simple.
7. Venation was open, and only rarely closed.
8. Syndetocheilic type of stomata were present.
9. The wall of the epidermal cells was sinuous.
10. The reproductive organs were organised in the form of hermaphrodite (e.g.
Cycadeoidea) or unisexual (e.g. Wielandiella) “flowers”, which in turn were
protected by many bracts.

UNIT-2
BSC I YEAR BOTANY-II Page 7
11. The „flowers‟ developed in the axil of leaves.
12. Male reproductive organs were borne in a whorl. They were free or fused,
entire or pinnately compound.
13. Microsporangia were present abaxially in the form of synangia.
14. Microsporophyll‟s sometimes surrounded megasporophylls forming
hermaphrodite “flowers”.
15. Ovules were numerous and stalked and borne on a conical, cylindrical or
dome-shaped receptacle.
16. Many inter-seminal bracts were present on the ovule containing receptacle.
17. The scales or bracts were united at end to form shield through which
micropyle protrudes.
18. Seeds were dicotyledonous.
Gnetales
Taxonomic Arrangements of Gnetales:
Foster and Gifford (1959) described Gnetales as “a small group of
gymnosperm-like plants” while Maheshwari and Vasil (1961) Mentioned that ”
the order Gnetales, formerly included three genera, Ephedra, Welwitschia and
Gnetum which were considered to be highest evolved among the gymnosperms
and believed to show an approach to the angiosperms”.
But mainly due to the presence of naked ovules and also because of the absence
of true sty le and stigma in Ephedra, Welwitschia and Gnetum, these members
can only be treated under gymnosperms and not under angiosperms.
General Characteristics of Gnetales:
Gnetales, believed by some botanists to be the ancestors of flowering plants
or angiosperms, are the highly evolved members of gymnosperms and show
following characteristics:
1. These are woody plants, of which some species are trees (Gnetum gnemon),
many are lianes or shrubs and a few. are stumpy turnip-like (e.g. Welwitschia
mirabilis).
2. Leaves are simple elliptical or strap-shaped or sometimes reduced to minute
scales. They are generally opposite or whorled.

UNIT-2
BSC I YEAR BOTANY-II Page 8
3. Vessels are present in the secondary wood.
4. „Flowers‟ are unisexual, usually dioecious and only rarely monoecious as in
some species of Gnetum.
5. „Flowers‟ are arranged in compound strobili or „inflorescences‟.
6. The male flowers are surrounded by a perianth. Each male flower contains an
antherophore with one to eight synangia.
7. A single erect orthotropous ovule is present in each female flower.
8. Nucellus of the ovule remains surrounded by two or three envelopes.
. The micropyle of each ovule remains projected in the form of a long bristle-
like tube.
10. At the time of fertilization the pollen tube contains two male nuclei.
11. A unicellular primary suspensor is present in the embryo.
12. Two cotyledons are present in the embryo.
Ginkgoales
Meaning of Ginkgoales:
The order Ginkgoales is today represented by only one living member, i.e.
Ginkgo biloba. Ginkgoales was, however, very abundantly represented in the
world by several species of about 16 genera during the Triassic period of
Mesozoic age, i.e. about 200,000,000 years ago. Today, all the genera, except
Ginkgo biloba, are extinct.
According to Dallimore and Jackson (1948), G. biloba is represented by five
varieties viz. Ginkgo biloba var. aurea (Nelson) Beisson, G.biloba var. fastigata
Henry, G.biloba var. paciniata Carriere, G. biloba var. pendula Carnere and G.
biloba var. variegata Carriere. Due to the presence of a number of primitive
characters, as well as because of its long geological records, Ginkgo is called
a “living fossil”.
General Characteristics of Ginkgoales:
Some general characteristics of Ginkgoales are under mentioned:
1. Tall, well-branched trees with short and long shoots. However, some earliest
fossil members were without short and long shoots.
2. Wood is pycnoxylic.

UNIT-2
BSC I YEAR BOTANY-II Page 9
3. Leaves are large, leathery and fan-shaped or strap-shaped. They are often
deeply divided.
4. Dichotomous venation is usually present in the leaves.
5. Un-branched, catkin-like male organs are axillary in position.
6. Male organs bear micro-sporangiophores.
7. Each micro-sporangiophore possesses 2-12 pendulous microsporangia.
8. Spermatozoids are motile and contain spiral bands of flagella.
9. Ovules are terminal in position on branched or un-branched axillary axes.
They are 2-10 in number.
10. Seeds are large-sized.
11. Each seed contains a fleshy outer layer and a middle stony layer.
Cycas
General Morphology of Cycas:
Cycas is a palm-like, evergreen plant (Fig. 8.8). Prior to the anatomical studies
of the stem of Cycas revoluta by Brongniart (1829), the Cycas was actually
considered a palm. The plant body consists of a columnar aerial trunk with a
crown of pinnately compound leaves as its top.
According of Eichler (1889), Coulter and Chamberlain (1910), Schuster (1932)
and others, a tap root system persists in the adult plant, but according to
Worsdell (1906) the tap roots are soon replaced by adventitious roots.

UNIT-2
BSC I YEAR BOTANY-II Page 10
Roots:
Roots in Cycas are of two types, i.e., normal tap roots forming a tap root
system, and coralloid roots. Normal tap-roots are positively geotropic, grow
deep into the soil and generally possess no root hairs. Their function is to fix the
plant in the soil and to absorb water and other minerals.
From the normal roots develop some lateral branches near the ground surface.
These lateral roots get infected with some bacteria, fungi and algae, and are
called coralloid roots (Fig. 8.9). They grow- first horizontally in the soil and
become swollen at their tips.
They divide repeatedly to form big bunches of greenish or brownish structures,
which are coral like in appearance. They divide dichotomously, come out of the
soil on the ground surface and are phototrophic in nature. Young plants bear
more coralloid roots than the older ones.

UNIT-2
BSC I YEAR BOTANY-II Page 11
Recently, Pant and Das (1990) reported non-coralloid aerial roots in Cycas
circinalis, C. revoluta and C. rumphii. The charactenstic algal zone of coralloid
roots is absent in these roots. These are positively geotropic, adventitious and
develop from the lower sides of leaf bases or bulbils when they are still attached
to the plant.
Stem:
The stem is thick, woody and usually un-branched. It is tuberous when young
but columnar, erect and stout at maturity. Branching in stem (Fig. 8.10) is also
not rare after the plants have reached a certain age. The aerial part of the trunk
remains covered by a thick armour of large and small rhomboidai leaf bases.
These occur regularly in alternate bands (Fig. 8 .11). The larger leaf bases
represent the bases of foliage leaves, while the smaller ones are the bases of
scaly leaves in male plants and scales and megasporophylls in female plants.
The age of the plant can be calculated by counting the number of crowns of
leaves and megasporophylls which are produced every year.

UNIT-2
BSC I YEAR BOTANY-II Page 12
Among all Cycas species, C. media is tallest, attaining a height up to 20 metres.
Regarding the age of Cycas, the plants can survive for a long period. C.
circinalis, if allowed to grow undisturbed, may attain an age of 100 years or
even more.
Leaves:
Two types of leaves are present in Cycas. These are green, assimilatory ox
foliage leaves, and scaly leaves or cataphylls.
1. Foliage Leaves or Assimilatory Fronds: These are green, large,
pinnately compound and stout leaves with a spiny petiole and large,

UNIT-2
BSC I YEAR BOTANY-II Page 13
strong rachis. They are produced at the apex of the stem in the form of
crown. The rachis bears many leaflets.
With the help of a transversely expanded rhomboidai leaf base, a leaf remains
attached with the stem Two rows of strong and stiff spines are present on the
petiole. These spines gradually transform into two rows of pinnae towards the
upper side of the leaf (Fig. 8.12).
Pant (1953) reported many abnormalities in Cycas leaves. Author, along with
two of his colleagues, also reported many abnormalities in the vegetative parts
of an year-old plant of Cycas circinalis growing in the Botanical Garden of
Meerut College, Meerut.
Cycas leaf is very large and may reach up to 3 metres in length in some species
such as C. thouarsii. Two rows of pinnae on the leaves may be alternate or
opposite. The number of pinnae varies in different species. As many as hundred
pairs of pinnae may be present in a mature leaf.
Each pinna is sessile, elongated, ovate or lanceolate in shape with a spiny or
acute apex. Pinnae are repeatedly and deeply dichotomized in C. micholitzii
(Fig. 8.13). Each pinna or of leaflet contains a midrib without any lateral
branching.
Forking of the midvein of the leaflet has been reported in C. circinalis by author
in 1976. Margins of the leaflets are revolute in C. revoluta and C. beddomei,
while in C. rumphii and C. circinalis they are flat.

UNIT-2
BSC I YEAR BOTANY-II Page 14
According to Chamberlain (1935) the “vernation is circinate in the midrib
and pinnules of Cycas”. Leaves, when young, have circinately coiled pinnae
like those of ferns (Fig. 8.14). Very young parts of Cycas are also covered by
fern-like hairs or ramenta.
2. Scaly Leaves or Cataphylls:
These are dry, brown-coloured, somewhat triangular leaves with their one end
pointed. They are present at the apex of the stem and remain covered with
several ramental hairs
tal hairs (Fig. 8.15).
UNIT-2
BSC I YEAR BOTANY-II Page 15
3. Anatomy of Vegetative Parts of Cycas:
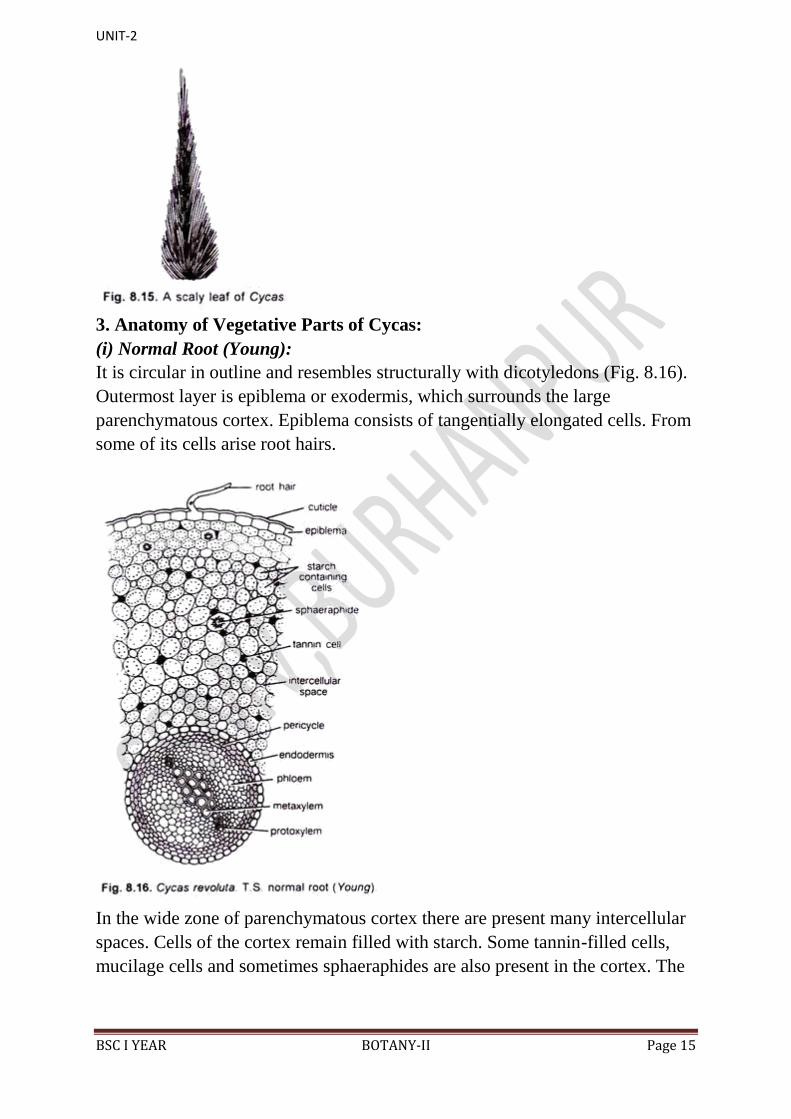
(i) Normal Root (Young):
It is circular in outline and resembles structurally with dicotyledons (Fig. 8.16).
Outermost layer is epiblema or exodermis, which surrounds the large
parenchymatous cortex. Epiblema consists of tangentially elongated cells. From
some of its cells arise root hairs.
In the wide zone of parenchymatous cortex there are present many intercellular
spaces. Cells of the cortex remain filled with starch. Some tannin-filled cells,
mucilage cells and sometimes sphaeraphides are also present in the cortex. The

UNIT-2
BSC I YEAR BOTANY-II Page 16
cortex is delimited by a single- layered endodermis. Casparian steps are present
in the barrel-shaped cells of the endodermis.
Endodermis is followed by multilayered pericycle. Xylem and phloem bundles
in the roots are radially arranged, i.e. present on different radii. The roots are
usually diarch but sometimes the number of protoxylem strands range between
3 to 8.
The protoxylem consists of spiral tracheids while the metaxylem consists of
scalariform tracheids. Vessels are absent. Phloem is present alternately with
xylem groups and consists of sieve tubes and phloem parenchyma. Pith is
generally absent.
(ii) Normal Root (old) Showing Secondary Growth:
The older roots (Fig. 8.17) undergo secondary growth. The cambium cuts
secondary phloem towards the outer side and secondary xylem towards the
inner side. After sometime the pericycle cells also become meristematic and
form a complete cambial ring.
The secondary xylem consists of radial rows of tracheids separated by
parenchymatous cells. The crushed primary phloem is present in the form of
dark streaks outside the secondary phloem. The secondary xylem is manoxvlic
and contains many multiseriate rays.
Periderm starts to develop in the cortex of old roots. Some of the cells of the
outermost cortical region start to become meristematic and function as cork
cambium. It cuts cork towards outer side and secondary cortex towards inner
side. Cork cells are dead and remain filled with subenn. Cycas roots often show
two layers of periderm (Fig. 8.17).
Epiblema is ruptured and there are no root hairs in the older roots.

UNIT-2
BSC I YEAR BOTANY-II Page 17
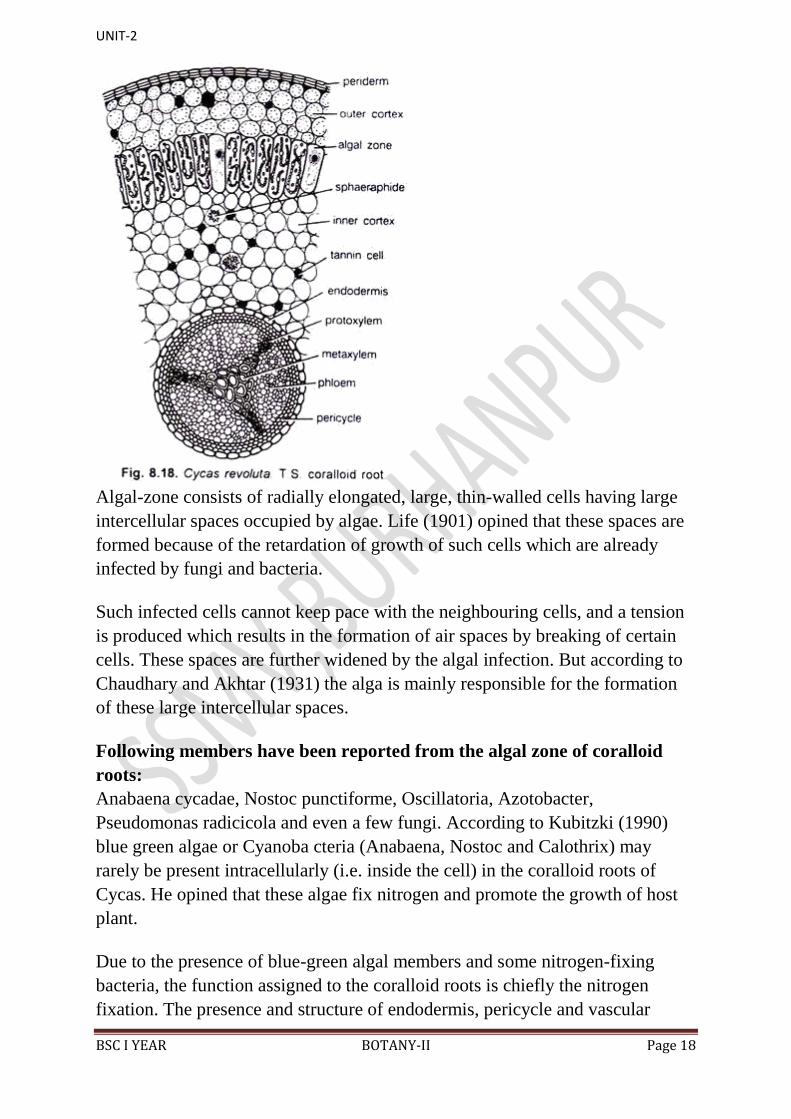
(iii) Coralloid Root:
Anatomically, the coralloid roots (Fig. 8.18) resemble normal roots except
some under mentioned differences:
1. The secondary vascular tissue in coralloid roots is either totally absent or
poorly-developed.
2. The cortex is wider in comparison with the normal root.
3. Presence of a greenish algal-zone in the middle of the cortex. But according
to Chaudhary and Akhtar (1931) the algal-zone is not of universal occurrence in
the coralloid roots of Cycas. It may be absent in such coralloid roots which go
very deep in the soil. According to these workers only those coralloid roots are
negatively geotropic which are infected by algal members.
UNIT-2
BSC I YEAR BOTANY-II Page 18
Algal-zone consists of radially elongated, large, thin-walled cells having large
intercellular spaces occupied by algae. Life (1901) opined that these spaces are
formed because of the retardation of growth of such cells which are already
infected by fungi and bacteria.
Such infected cells cannot keep pace with the neighbouring cells, and a tension
is produced which results in the formation of air spaces by breaking of certain
cells. These spaces are further widened by the algal infection. But according to
Chaudhary and Akhtar (1931) the alga is mainly responsible for the formation
of these large intercellular spaces.
Following members have been reported from the algal zone of coralloid
roots:
Anabaena cycadae, Nostoc punctiforme, Oscillatoria, Azotobacter,
Pseudomonas radicicola and even a few fungi. According to Kubitzki (1990)
blue green algae or Cyanoba cteria (Anabaena, Nostoc and Calothrix) may
rarely be present intracellularly (i.e. inside the cell) in the coralloid roots of
Cycas. He opined that these algae fix nitrogen and promote the growth of host
plant.
Due to the presence of blue-green algal members and some nitrogen-fixing
bacteria, the function assigned to the coralloid roots is chiefly the nitrogen
fixation. The presence and structure of endodermis, pericycle and vascular

UNIT-2
BSC I YEAR BOTANY-II Page 19
bundles in the coralloid roots are similar to that of normal roots. The xylem is
exarch and triarch.
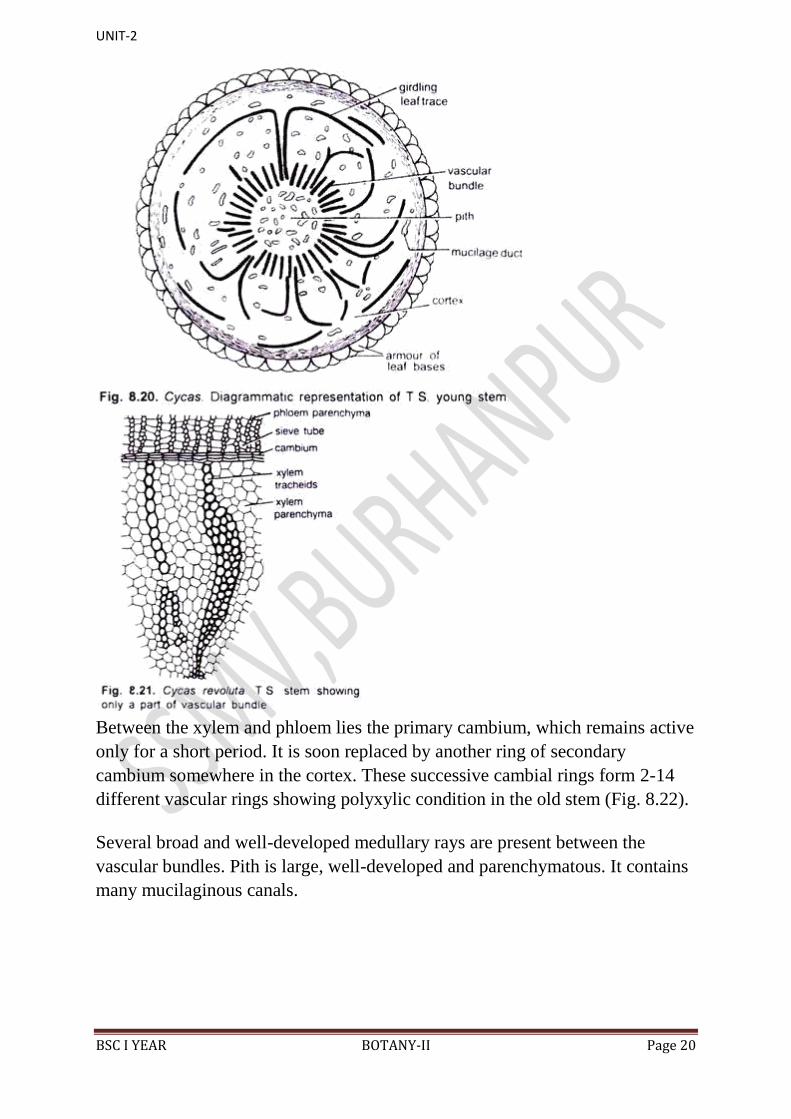
(iv) Stem:
Similar to root, the stem of Cycas also resembles internally with a
dicotyledonous stem.
It shows the following anatomical features:
Epidermis is the outermost layer consisting of compactly arranged thick- walled
cells. Presence of several persistent leaf bases makes the epidermis a
discontinuous and ruptured layer. Cortex is large and consists of thin- walled,
parenchymatous cells, filled densely with starch grains. It contains numerous
mucilaginous canals and girdle traces.
Each mucilage canal is lined with many radially elongated epithelial or
secretory cells (Fig. 8.19). Medullary rays connect the mucilage canals of the
cortex with that of the pith Starch in the parenchymatous cells of the cortex is
the source of „sago‟. Endodermis and pericycle are not clearly demarcated.
Numerous vascular bundles remain arranged in a ring. The stele is ectophloic
siphonostele. Each vascular bundle is conjoint, collateral, open and endarch
(Fig. 8.20). The xylem consists of tracheids and xylem parenchyma (Fig. 8.21).
Protoxylem contains tracheids with spiral thickenings while the metaxylem has
scalariform thickenings with bordered pits. Vessels are absent. The phloem is
located outside the xylem and consists of sieve tubes and phloem parenchyma.
Companion cells are absent.
UNIT-2
BSC I YEAR BOTANY-II Page 20
Between the xylem and phloem lies the primary cambium, which remains active
only for a short period. It is soon replaced by another ring of secondary
cambium somewhere in the cortex. These successive cambial rings form 2-14
different vascular rings showing polyxylic condition in the old stem (Fig. 8.22).
Several broad and well-developed medullary rays are present between the
vascular bundles. Pith is large, well-developed and parenchymatous. It contains
many mucilaginous canals.

UNIT-2
BSC I YEAR BOTANY-II Page 21
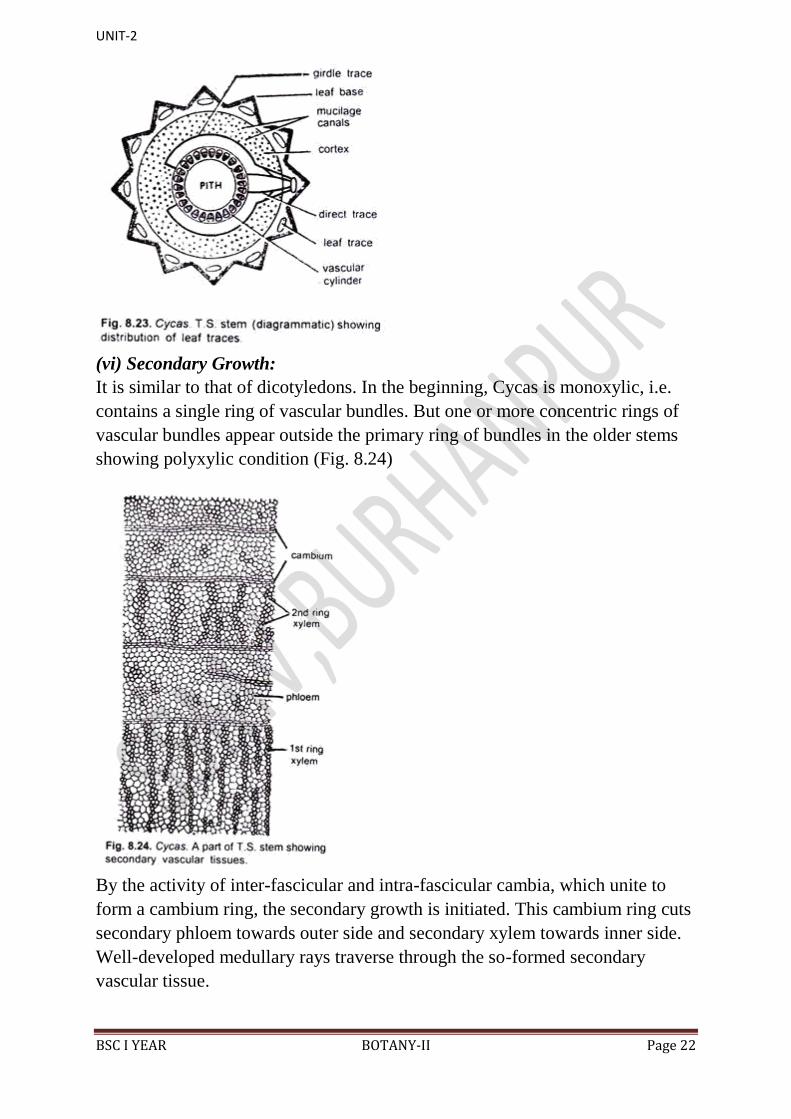
(v) Leaf Traces:
The leaf traces remain scattered in the cortical region of the stem and constitute
the vascular supply to the leaves from the main vascular cylinder. Normally,
there are four leaf traces which form the vascular supply to the leaf. Two of
these are direct traces, while the remaining two axe girdle traces (Fig. 8.23).
The direct traces originate from the vascular cylinder lying in front of the leaf
base while the girdle traces develop from the vascular cylinder lying opposite to
that of direct traces. They proceed together and curve soon in opposite
directions, and by girdling round the vascular cylinder they enter in the leaf
base.
In the cortical region the girdle traces also remain connected with other leaf
traces. At the time of their entrance in the petiole, the leaf trace bundles
subdivide and form many petiole bundles. Such type of unique girdle traces of
Cycas, which also occur in Magnoliaceae. show a close relationship of
Cycadales of Gymnosperms and Magnoliaceae of dicotyledons.
UNIT-2
BSC I YEAR BOTANY-II Page 22
(vi) Secondary Growth:
It is similar to that of dicotyledons. In the beginning, Cycas is monoxylic, i.e.
contains a single ring of vascular bundles. But one or more concentric rings of
vascular bundles appear outside the primary ring of bundles in the older stems
showing polyxylic condition (Fig. 8.24)
By the activity of inter-fascicular and intra-fascicular cambia, which unite to
form a cambium ring, the secondary growth is initiated. This cambium ring cuts
secondary phloem towards outer side and secondary xylem towards inner side.
Well-developed medullary rays traverse through the so-formed secondary
vascular tissue.
UNIT-2
BSC I YEAR BOTANY-II Page 23
After a short while this cambium ring stops functioning and a second cambium
ring develops either in the parenchymatous cortex or in the region of pericycle
This cambium ring also behaves in the similar fashion.
In this fashion, as many as 14 rings of vascular tissue may develop in the stem
of Cycas pectinata of about 20 cm diameter showing polyxylic condition.
Seward (1917) reported 12 such rings in the stem of C. media of about 30 cm
diameter, and Schuster (1932) reported 22 such rings in the stem of C. rumphii
having a diameter of about 85 cm.
Cambial rings towards the periphery of the stem form lesser number of vascular
bundles. The cork cambium develops on the outer region of the cortex and cuts
cork towards outer side and secondary cortex towards inner side.
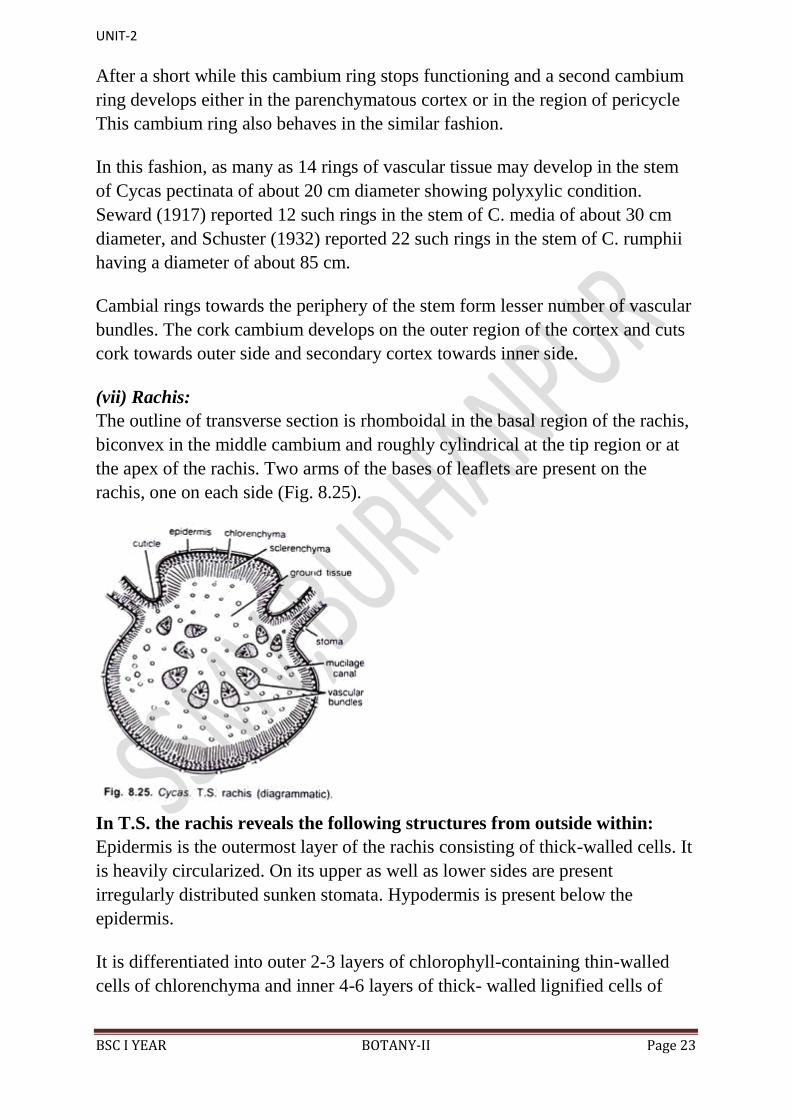
(vii) Rachis:
The outline of transverse section is rhomboidal in the basal region of the rachis,
biconvex in the middle cambium and roughly cylindrical at the tip region or at
the apex of the rachis. Two arms of the bases of leaflets are present on the
rachis, one on each side (Fig. 8.25).
In T.S. the rachis reveals the following structures from outside within:
Epidermis is the outermost layer of the rachis consisting of thick-walled cells. It
is heavily circularized. On its upper as well as lower sides are present
irregularly distributed sunken stomata. Hypodermis is present below the
epidermis.
It is differentiated into outer 2-3 layers of chlorophyll-containing thin-walled
cells of chlorenchyma and inner 4-6 layers of thick- walled lignified cells of

UNIT-2
BSC I YEAR BOTANY-II Page 24
sclerenchyma. Sclerenchyma is poorly-developed on the lateral sides. It is also
seen intermixed with chlorenchyma.
Ground tissue is a large region consisting of thin- walled, parenchymatous cells.
Many mucilaginous canals and vascular bundles are present in this region. The
number and arrangement of mucilage canals have no definite relation with that
of vascular bundles. Each mucilage canal is a double-layered structure
consisting of an inner layer of epithelium cells surrounded by an outer layer.
Vascular bundles are arranged in the shape of an inverted Greek letter Omega
(Ω) (Fig. 8.25). Towards the tip of the rachis the bundles are arranged in C-
shaped manner and their number is comparatively less. Each vascular bundle
remains surrounded by a bundle sheath (Fig. 8.26). It is conjoint, collateral and
open.

UNIT-2
BSC I YEAR BOTANY-II Page 25
The xylem in each vascular bundle is present towards inner side. It consists of
tracheids and xylem parenchyma. Cambium separates the xylem from the
phloem. Vessels are absent.
The vascular bundles are diploxylic, i.e. consists of two types of xylem viz.
centripetal xylem and centrifugal xylem. Phloem, present towards the outer side
of the vascular bundle, consists of sieve tubes and phloem parenchyma.
Companion cells are absent.
The vascular bundles show different structure at different levels of rachis
starting from the base up to the apex, especially with regard to their diploxylic
nature.
Their brief description is under mentioned:
(a) Vascular Bundles At the Base of Rachis:
Only the centrifugal xylem is well-developed in the vascular bundles (Fig.
8.27A). Its protoxylem faces towards the centre showing endarch condition.
Centripetal xylem is not developed.
(b) Vascular Bundles In the Middle of Rachis:
Both centripetal as well as centrifugal xylem are present showing diploxylic
condition (Fig. 8.27B). Centripetal xylem is present just opposite to the
protoxylem of the centrifugal xylem.
(c) Vascular Bundles At the Apex of Rachis:
Centripetal xylem is well-developed, triangular and exarch (Fig. 8.27C).
Centrifugal xylem is much reduced and present in the form of two patches lying
one on each side of the protoxylem elements of centripetal xylem. Centrifugal
xylem is totally absent at the extreme tip of the rachis.
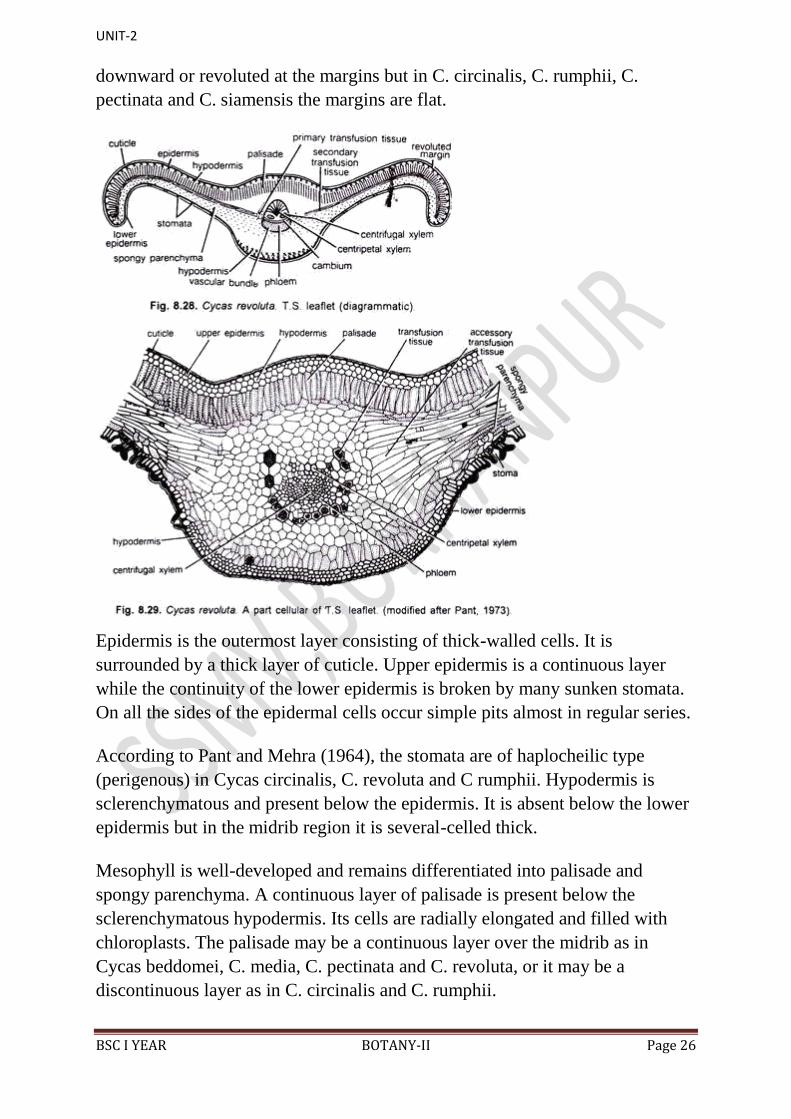
(viii) Leaflet:
Cycas leaflets are large, tough, thick and leathery. In a vertical section the
leaflet is differentiated into a swollen midrib portion and two lateral wings
(Figs. 8.28, 8.29). In C. revoluta and C. beddomei the wings are curved
UNIT-2
BSC I YEAR BOTANY-II Page 26
downward or revoluted at the margins but in C. circinalis, C. rumphii, C.
pectinata and C. siamensis the margins are flat.
Epidermis is the outermost layer consisting of thick-walled cells. It is
surrounded by a thick layer of cuticle. Upper epidermis is a continuous layer
while the continuity of the lower epidermis is broken by many sunken stomata.
On all the sides of the epidermal cells occur simple pits almost in regular series.
According to Pant and Mehra (1964), the stomata are of haplocheilic type
(perigenous) in Cycas circinalis, C. revoluta and C rumphii. Hypodermis is
sclerenchymatous and present below the epidermis. It is absent below the lower
epidermis but in the midrib region it is several-celled thick.
Mesophyll is well-developed and remains differentiated into palisade and
spongy parenchyma. A continuous layer of palisade is present below the
sclerenchymatous hypodermis. Its cells are radially elongated and filled with
chloroplasts. The palisade may be a continuous layer over the midrib as in
Cycas beddomei, C. media, C. pectinata and C. revoluta, or it may be a
discontinuous layer as in C. circinalis and C. rumphii.

UNIT-2
BSC I YEAR BOTANY-II Page 27
Spongy parenchyma is present only in the wings, directly above the lower
epidermis. Its cells are oval, filled with chloroplasts, and loosely arranged
having many air-filled intercellular spaces. Transfusion tissue consists of two
small groups of short and wide tracheid-like cells with reticulate thickenings or
bordered pits on their walls.
These cells have been named as transfusion tissue by Von Mohl (1871), and
were first described by Frank (1864). Few layers of transversely elongated cells
are present in both the wings just in between the palisade and spongy
parenchyma.
This represents the accessory transfusion tissue or secondary transfusion tissue.
The secondary‟ transfusion tissue has also been named as hydrostereom by
Bernard (1904) or radial parenchyma by Pilger (1926). A great phylogenetic
significance has been attributed to the transfusion tissue by Worsdell (1897).
Vascular bundle is one, and present in the midrib region of the leaflet. It is
conjoint, collateral, open and diploxylic. The triangular centrifugal xylem is
well-developed with endarch protoxylem. It is represented by two or sometimes
more small groups on either side of the protoxylem.
Phloem is arc-shaped and remains separated by cambium. Phloem consists of
sieve tubes and phloem parenchyma. Companion cells are absent. The portion
of the midrib in between the palisade layer and lower hypodermal region is
filled with parenchymatous cells. Some of these cells contain calcium oxalate
crystals.
4. Reproduction in Cycas:
(i) Vegetative Reproduction:
The most common method of vegetative propagation in Cycas is by bulbils. The
bulbils develop from the axil of the scaly leaves. They are more or less oval
structures with a broad base narrowing towards the apex. Several scaly leaves
are arranged spirally and compactly over a dormant stem in a bulbil (Fig. 8.30).
On detachment from the stem, a bulbil starts germination by producing many
roots towards the lower side and a leaf towards the upper side. A bulbil from
male plant will develop only into the male plant, while from the female plant
will form only the female plant because Cycas is a strictly dioecious plant.

UNIT-2
BSC I YEAR BOTANY-II Page 28
(ii) Sexual Reproduction:
Cycas is strictly dioecious, i.e. male and female sex organs are borne on
separate plants. After several years of vegetative growth the plants start to form
sex organs. Generally, Cycads of more than 10 years of age produce the sex
organs.
The male plants develop male cones or male strobili bearing microsporophyll‟s,
while the female plants produce a loose collection of megasporophylls. The
male cone is terminal while the megasporophylls are produced in succession
with the leaves at the top of the stem.
Male Reproductive Structures:
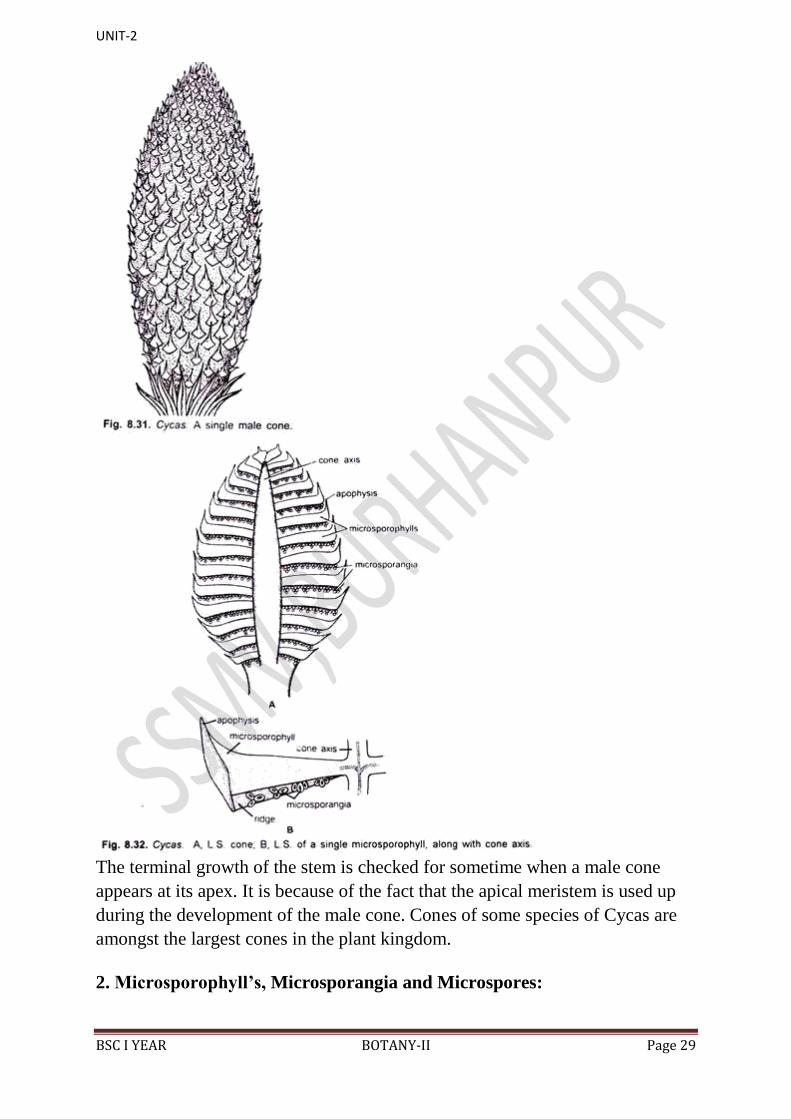
1. Male Cone:
The male cone (Fig. 8.31) or male strobilus is a large, conical or ovoid,
compact, solitary and shortly-stalked structure, which is generally terminal in
position. It sometimes attains a length of as much as 1.5 metre. In the centre of
the cone is present a cone axis (Fig. 8.32).
Several perpendicularly attached microsporophyll‟s are arranged around the
cone axis in closely set spirals. At the base of male cone are present many
young leaves. All the microsporophyll‟s in a male cone are fertile except a few
at its basal and apical parts.
UNIT-2
BSC I YEAR BOTANY-II Page 29
The terminal growth of the stem is checked for sometime when a male cone
appears at its apex. It is because of the fact that the apical meristem is used up
during the development of the male cone. Cones of some species of Cycas are
amongst the largest cones in the plant kingdom.
2. Microsporophyll‟s, Microsporangia and Microspores:

UNIT-2
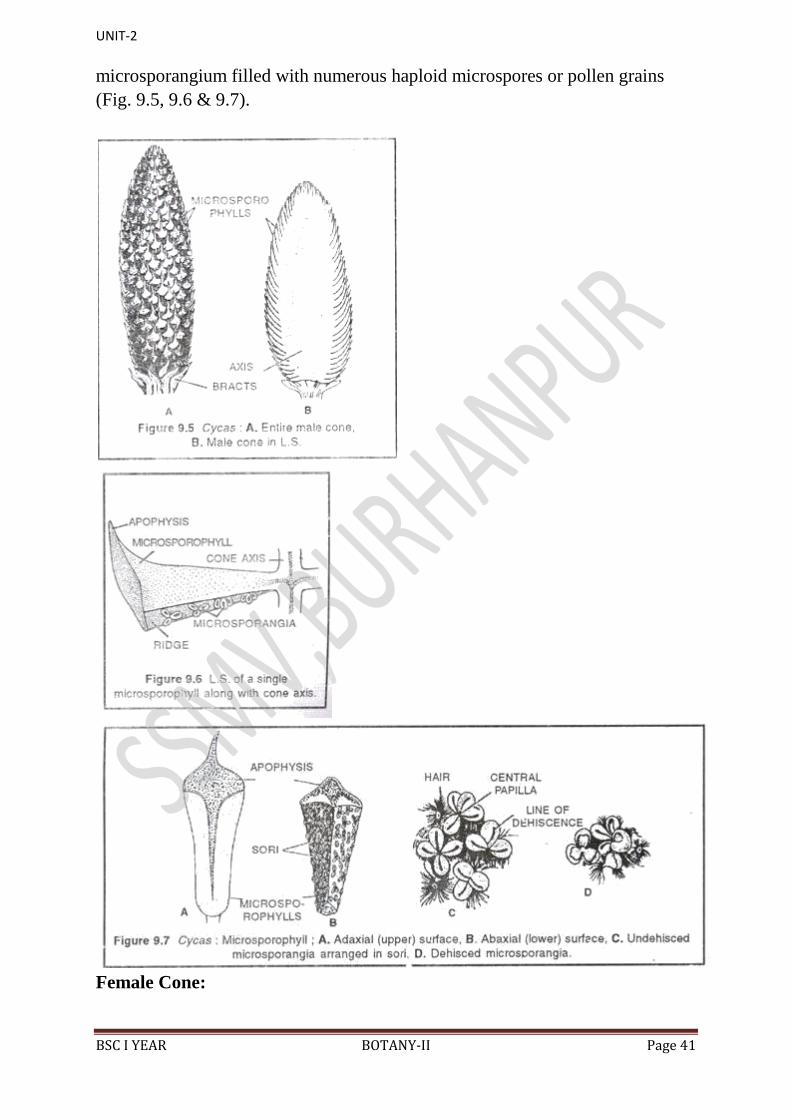
BSC I YEAR BOTANY-II Page 30
Microsporophyll‟s (Fig. 8.33) are flat, leaf-like, woody and brown-coloured
structures with narrow base and expanded upper portion. The upper expanded
portion becomes pointed and is called apophysis. Narrow base is attached to the
cone axis with a short stalk.
Each microsporophyll contains two surfaces, i.e. an adaxial or upper surface and
an abaxial or lower surface. On the adaxial surface is present a ridge-like
projection in the middle and an apophysis at the apex (Fig. 8.33).
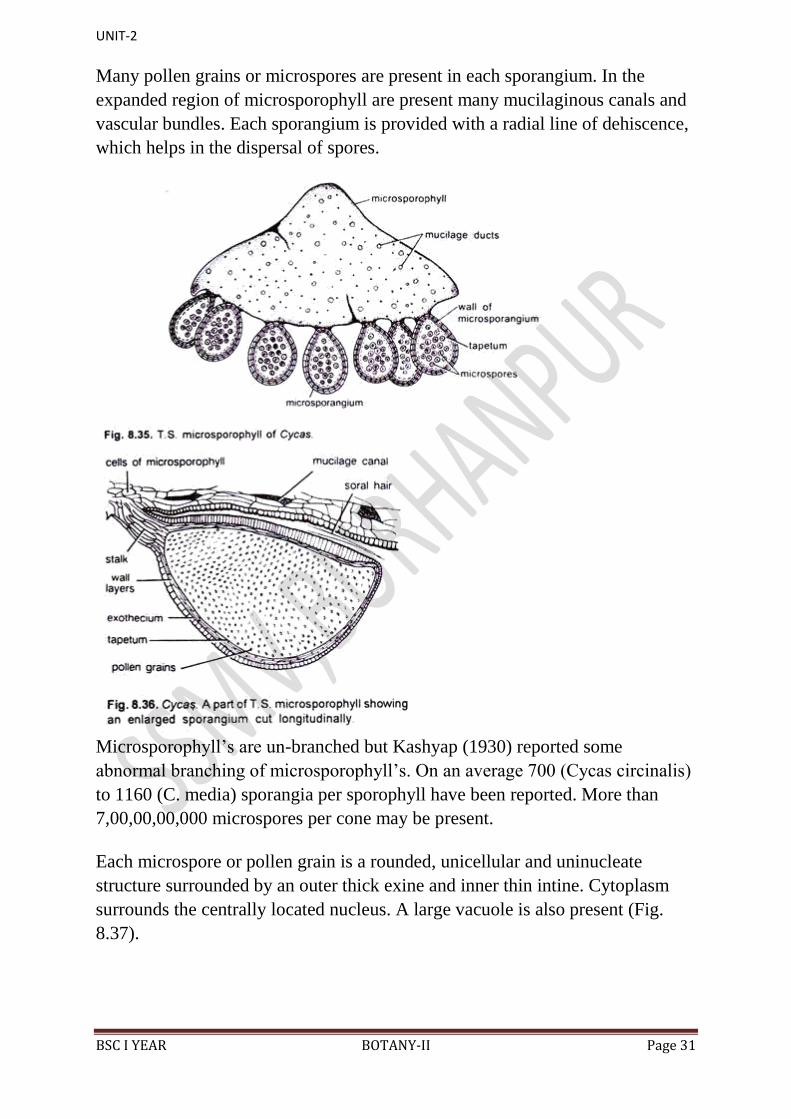
On the abaxial surface (Fig. 8.34A) are present thousands of microsporangia in
the middle region in the groups of 3-5. Each such group is called a sorus. In
between these groups are present many hair-like structures, which are very soft
and one or two- celled structures (Fig. 8.34B).
In T.S. of a microsporophyll, there are present many microsporangia on the
abaxial side (Fig. 8.35). Each shortly-stalked, oval or sac-like microsporangium
is surrounded by 5-6 layers. The wall layers of each sporangium include an
outer thick epidermis or exothecium, middle zone of thin-walled cells and an
innermost layer of tapetum (Fig. 8.36).
UNIT-2
BSC I YEAR BOTANY-II Page 31
Many pollen grains or microspores are present in each sporangium. In the
expanded region of microsporophyll are present many mucilaginous canals and
vascular bundles. Each sporangium is provided with a radial line of dehiscence,
which helps in the dispersal of spores.
Microsporophyll‟s are un-branched but Kashyap (1930) reported some
abnormal branching of microsporophyll‟s. On an average 700 (Cycas circinalis)
to 1160 (C. media) sporangia per sporophyll have been reported. More than
7,00,00,00,000 microspores per cone may be present.
Each microspore or pollen grain is a rounded, unicellular and uninucleate
structure surrounded by an outer thick exine and inner thin intine. Cytoplasm
surrounds the centrally located nucleus. A large vacuole is also present (Fig.
8.37).

UNIT-2
BSC I YEAR BOTANY-II Page 32
Scanning electron microscopic studies of Sahashi and Ueno (1986) on the
pollen grains of Cycas revoluta suggest that they are oblong with 1-sulcate
shrunken aperture. Reticulum-like sculpting‟s are present on the inner layer of
exine, and in this character Cycas resembles with Ginkgo biloba.
3. Development of Microsporangium:
It is of eusporangiate type (Fig. 8 .38). Few hypodermal sporangial initials
divide penclinally to form outer primary wall cells and inner primary
sporogenous cells. Primary wall cells divide and re-divide periclinally as well as
anticlinally to form 5-7 cells thick wall of the sporangium while the primary
sporogenous cells divide to form many sporogenous cells.
By further divisions the sporogenous cells develop into microspore mother
cells. The latter divide reductionally to form haploid microspores or pollen
grains arranged tetrahedrally.
The tapetum, which is utilized for the spore formation, develops either from the
outermost layer of the sporogenous tissue or from the innermost layer of the
wall tissue. Microspore is the first cell of the male gametophyte having haploid
number of chromosomes.

UNIT-2
BSC I YEAR BOTANY-II Page 33
The haploid chromosome number in Cycas is 11. But in C. revoluta it is also
sometimes 12. The female plants are homogametic with XX-type of
chromosomes while the male plants are heterogametic having chromosomes of
XY-type.
Female Reproductive Organs:
True female cone or strobilus is absent Cycas. Female reproductive organs are
present in the form of megasporophylls. Many megasporophylls are present
around the apex of the monopodial trunk of the female plant above each crown
of foliage and scaly leaves (Fig. 8.39).
Similar to foliage leaves, megasporophylls also remain spirally arranged at the
apex of the stem but their number is very large and thus they appear like a
rosette. Vegetative leaves and fertile megasporophylls are produced in an
alternate succession without showing any effect on apical men stem.
Pant (1953) observed that usually the megasporophylls in Cycas are produced
only once in a year. From the apex of the main stem the megasporophylls arise
in an acropetal succession.

UNIT-2
BSC I YEAR BOTANY-II Page 34
1. Megasporophyll:
Each megasporophyll is considered a modification of foliage leaf. It reaches up
to 30 cm or more in length in different species. It is a flat body consisting of an
upper dissected or pinnate leafy portion, middle ovule-bearing portion and
proximal petiole. Petiole varies in length in different species.
The middle part is comparatively wider than petiole and bears ovules arranged
in two pinnate rows. The number of ovules varies between 2-12 in different
species. The ovules are green when young but at maturity they are fleshy and
bright orange or red-coloured structures.
The upper, conical sterile part of the megasporophyll is pinnately divided in
Cycas revoluta (Fig. 8.40), C. pectinata (Fig. 8.41 B) and C. siamensis (Fig.
8.41 A). But the margin of the upper part is variously serrate with a tapering
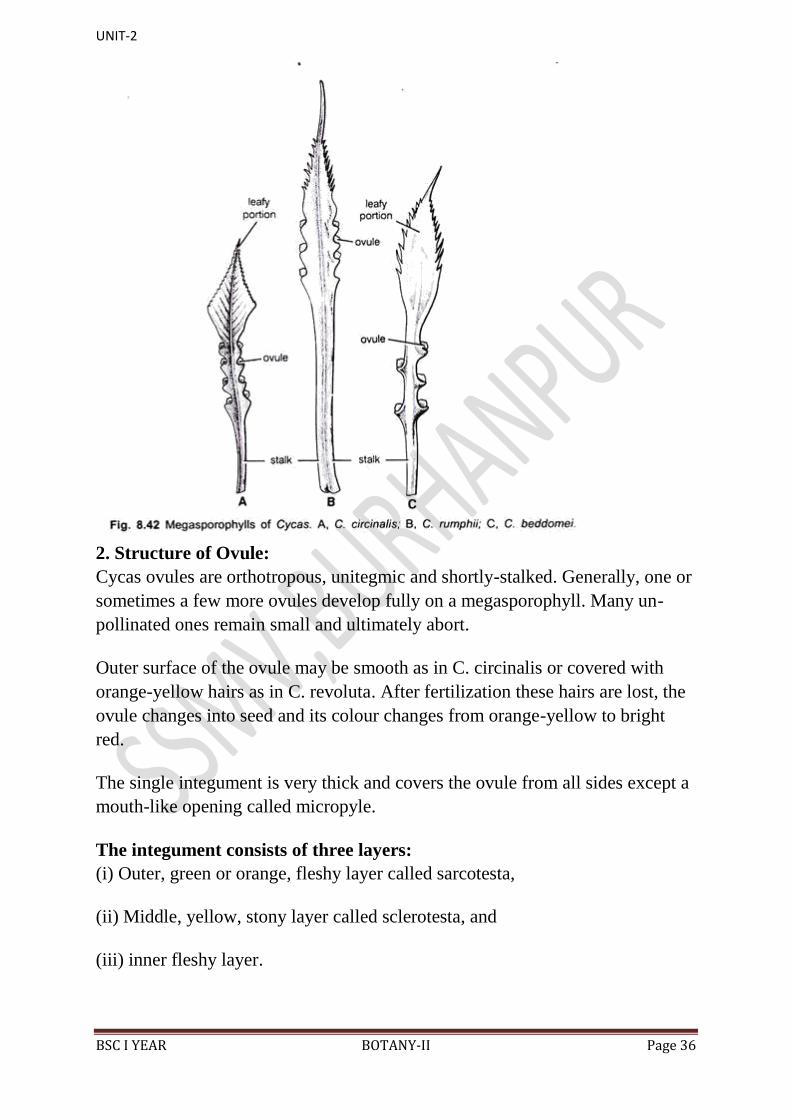
acute apex in C. beddomei (Fig. 8.42C), C. circinalis (Fig. 8.42A) and C.
rumphii (Fig. 8.42B).
Cycas thouarsi contains the largest ovule amongst the living gymnosperms
measuring about 7 cm in length. The megasporophylls remain covered by many
yellow or brown-coloured hairs.

UNIT-2
BSC I YEAR BOTANY-II Page 35
UNIT-2
BSC I YEAR BOTANY-II Page 36
2. Structure of Ovule:
Cycas ovules are orthotropous, unitegmic and shortly-stalked. Generally, one or
sometimes a few more ovules develop fully on a megasporophyll. Many un-
pollinated ones remain small and ultimately abort.
Outer surface of the ovule may be smooth as in C. circinalis or covered with
orange-yellow hairs as in C. revoluta. After fertilization these hairs are lost, the
ovule changes into seed and its colour changes from orange-yellow to bright
red.
The single integument is very thick and covers the ovule from all sides except a
mouth-like opening called micropyle.
The integument consists of three layers:
(i) Outer, green or orange, fleshy layer called sarcotesta,
(ii) Middle, yellow, stony layer called sclerotesta, and
(iii) inner fleshy layer.

UNIT-2
BSC I YEAR BOTANY-II Page 37
Several tannin cells and mucilage canals are present in the parenchymatous
region of sarcotesta. Some pigments are also present in sarcotesta and
epidermis. The sclerotesta consists of lignified thick-walled cells. The inner
fleshy layer consists of parenchymatous cells, and it remains in close
association with the nucellus.
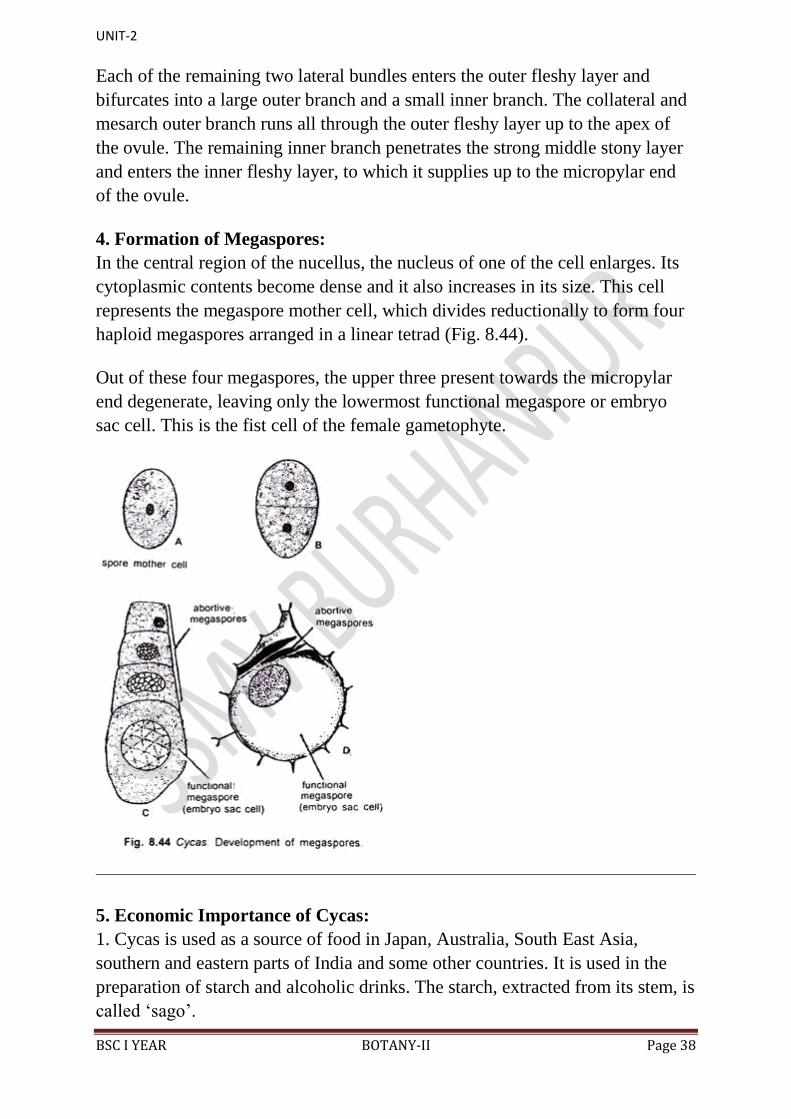
The nucellus grows out into a beak-like portion called nucellar beak. The latter
protrudes into the micropylar canal. Certain cells at the top of the nucellus
dissolve and form a cavity like structure called pollen chamber (Fig. 8.43).
Pollen grains are received in the pollen chamber after pollination.
The nucellus gets reduced in the form of a thin papery layer in mature seeds and
encloses the massive female gametophyte (endosperm). An enlarged megaspore
or the embryo-sac is present within the nucellus. The endosperm is formed by
the repeated divisions of the megaspore nucleus followed by free cell formation.
Just below the pollen chamber is present an archegonial chamber. 3-6
archegonia are present in the female gametophyte near the archegonial chamber.
The latter remains filled with a fluid.
3. Vascular Supply of the Ovule:
Stopes (1904) has worked on the vascular supply of Cycas seed. Out of several
bundles of the megasporophyll only three enter the base of the ovule (Fig. 8.43).
Out of these three bundles, the central one entefs into the base of the inner
fleshy layer of the integument. After its entrance it divides into number of
branches, all of which reach up to chalazal end of the nucellus. But none of
them penetrates the nucellus.
UNIT-2
BSC I YEAR BOTANY-II Page 38
Each of the remaining two lateral bundles enters the outer fleshy layer and
bifurcates into a large outer branch and a small inner branch. The collateral and
mesarch outer branch runs all through the outer fleshy layer up to the apex of
the ovule. The remaining inner branch penetrates the strong middle stony layer
and enters the inner fleshy layer, to which it supplies up to the micropylar end
of the ovule.
4. Formation of Megaspores:
In the central region of the nucellus, the nucleus of one of the cell enlarges. Its
cytoplasmic contents become dense and it also increases in its size. This cell
represents the megaspore mother cell, which divides reductionally to form four
haploid megaspores arranged in a linear tetrad (Fig. 8.44).
Out of these four megaspores, the upper three present towards the micropylar
end degenerate, leaving only the lowermost functional megaspore or embryo
sac cell. This is the fist cell of the female gametophyte.
5. Economic Importance of Cycas:
1. Cycas is used as a source of food in Japan, Australia, South East Asia,
southern and eastern parts of India and some other countries. It is used in the
preparation of starch and alcoholic drinks. The starch, extracted from its stem, is
called „sago‟.

UNIT-2
BSC I YEAR BOTANY-II Page 39
„Sago‟ is prepared in the following way:
The bark of the trunk is removed, and the trunk is cut into thin discs. These are
dried, ground and a paste is prepared by adding water Excess of water is added,
and the paste is left for some time in a standstill position.
The starch settles down, and the clear upper liquid is drained off. Between the
boards, the starch is rolled. This gives the starch a characteristic round shape. It
is finally dried and sold as „sago‟ in the market.
2. In Japan, seeds and stem of Cycas revoluta are used for preparing wine.
3. The juice obtained from young leaves of Cycas circinalis is used in skin
diseases, vomiting of blood and stomach disorders.
4. The decoction of young red seeds of C. circinalis is used as a purgative and
emetic.
5. To relieve the headache, giddiness and sore throat, the seeds of Cycas
revoluta are prepared in the form of a tincture and used.
6. Cycas revoluta and C. circinalis plants are grown for ornamental purposes in
various parts of the world.
7. The wood of Cycas revoluta is used for preparing small boxes and dishes.
8. Cycas leaves, being very large, are used for preparing baskets, mats, etc.
9. Cycas circinalis seeds are used in Democratic Kampuchea as a fish-poison.
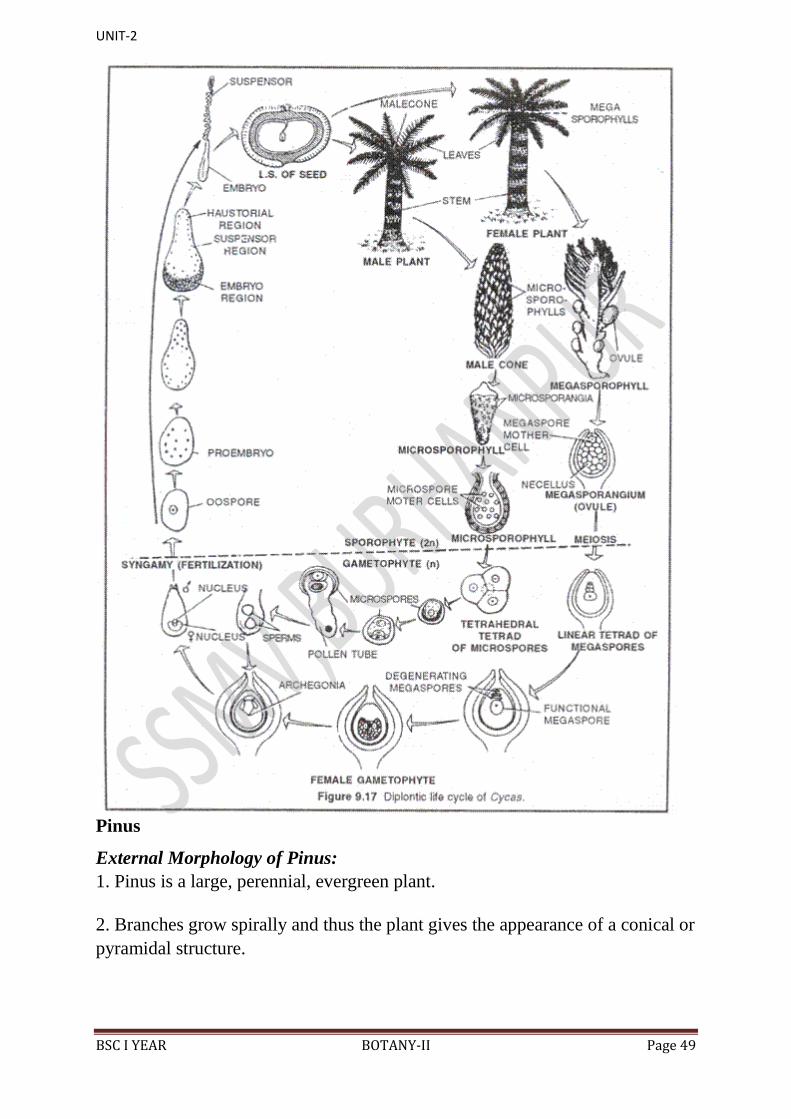
Life Cycle of Cycas
[I] Vegetative Cycle:
This is a deviation of regular alternation of generations between sporophyte and
gametophyte.
In this type of cycle, a sporophyte gives rise to a sporophyte of the same sex.
Vegetative Cycle takes place with the formation of adventitious buds called
bulbils in the basal part of stem
Bulbils are protected by scale leaves. During favourable period bulbils detach
from the parent and grow into an independent sporophyte.

UNIT-2
BSC I YEAR BOTANY-II Page 40
This cycle is more prevalent in Northern India where male plants of Cycas
revolute are not found.
[II] Sexual cycle (Sexual reproduction):
The sexual life cycle of Cycas is diplohaplontic. It shows heterologous or
heteromorphic type of alternation of generations because the sporophyte (2n)
and gametophyte (n) generations exhibit morphological differences. In Cycas,
the sporophyte (2n=22) is a complicated, independent and dominant generation
whereas the gametophytes (n=ll) are inconspicuous and endosporic. The
gametophytes of Cycas are of 2 types: male or microgametophyte and female or
magagametophyte. Female gametophyte is retained whereas male gametophyte
is transfer during pollination.
Reproductive organs:
After many years of vegetative growth sex organs develop on Cycas
sporophytes in the form of cones or strobili. Cycas is dioecious i.e. male cone
and female cone (lax) born on separate sporophytes. The male sporophyte is
heterogametic having sex chromosomes-XY while the female sporophyte is
homogametic having sex chromosomes-XX.
Male Cone:
Male cone is ovoid or conical and grow up to 1.5 m. It has a central axis or cone
axis surround by compactly and spirally arranged micro-sporophylls. Each
microsporophyll has adaxial (upper) and abaxial (lower) surfaces. The adaxial
surface is ridge like with sterile apophysis at the apex. The adaxial surface
contains numerous microsporangia in group of 3-6 called sori. Each
UNIT-2
BSC I YEAR BOTANY-II Page 41
microsporangium filled with numerous haploid microspores or pollen grains
(Fig. 9.5, 9.6 & 9.7).
Female Cone:

UNIT-2
BSC I YEAR BOTANY-II Page 42
In Cycas true and compact female cone (ovulate strobilus) is absent, instead it is
a lax where megasporophylls are loosely arranged at the stem apex that appears
like a rosette. Each megasporophyll is a modified foliage leaf ranging from 15-
30 cm. in length. It has a proximal petiole, middle ovule bearing part and upper
pinnately dissected sterile region. The middle fertile part bears 2-12 sessile
rounded ovules in two rows (Fig. 9.8 & 9.9).
Ovules:
The ovules are orthotropous, unitegmic and sessile or shortly stalked. The
Cycas ovule is largest in plant kingdom with 6-7 cm in diameter. In young stage
ovules are green covered with brown hairs but after fertilization hairs are lost
and appear orange to red in colour. The body of ovule is called nucellus
(megasporangium), covered by a thick integument in all sides except an opening
called micropyle. The apex of the nucellus has a pollen chamber and a nucellar
beak. The integument consists of three distinct layers: outer and inner fleshy
layers and middle stony layer.

UNIT-2
BSC I YEAR BOTANY-II Page 43
(a) Development of female gametophyte (Endosperm):
Inside the nucellus, one cell differentiated into megaspore mother cell. It
undergoes reduction division (meiosis) to form a linear tetrad of four haploid
megaspores. Usually, the upper 3 megaspores towards micropyle degenerate
while the lower most functional megaspore (embryo sac cell) undergoes free
nuclear division followed by wall formation to form a cellular female
gametophyte or endosperm.
Hence, the formation of female gametophyte is monosporic, i.e develops from a
single megaspore. During formation of endosperm nucellus is utilized. It should
be noted that in gymnosperms the endosperm develop before fertilization and is
haploid (n) while in angiosperms it is triploid (3n) and formed after fertilization
(Fig. 9.10).
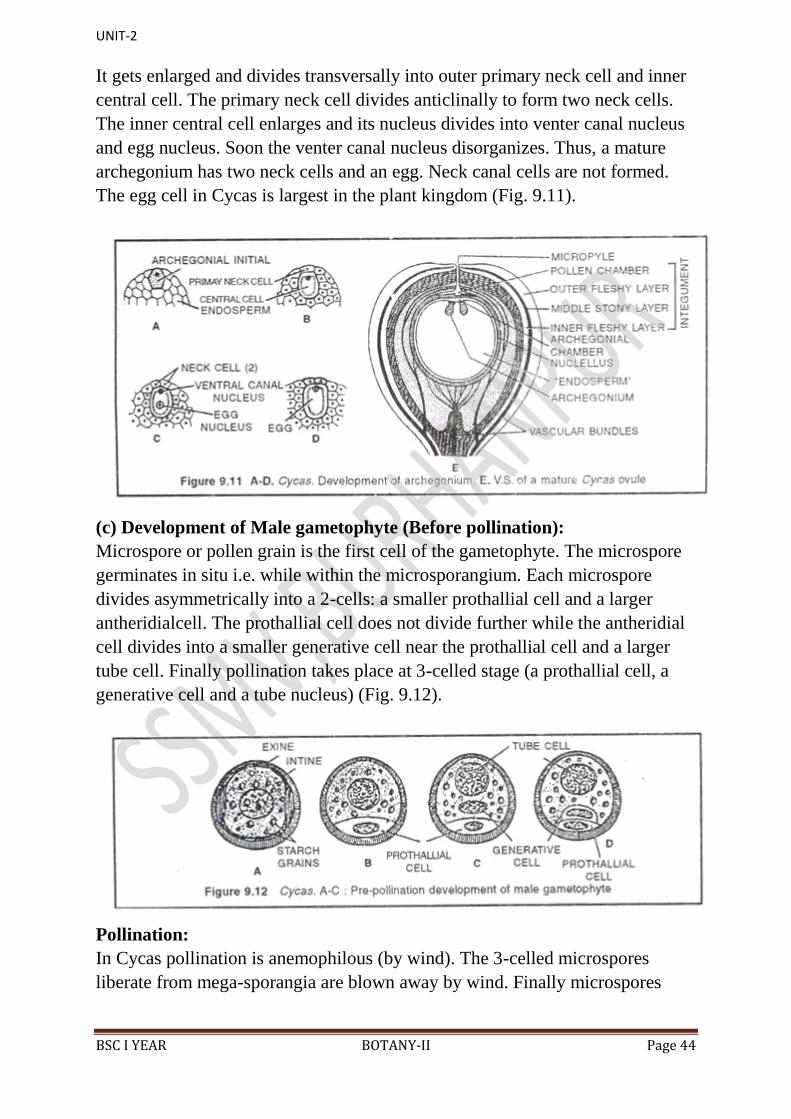
(b) Development of archegonium:
At the micropylar end of female gametophyte 2-8 archegonia develop. All the
necks of archegonia open into an archegonial chamber formed by a depression
in female gametophyte (Fig. 9.11). Each archegonium develops from single
superficial cell called archegonial initial.
UNIT-2
BSC I YEAR BOTANY-II Page 44
It gets enlarged and divides transversally into outer primary neck cell and inner
central cell. The primary neck cell divides anticlinally to form two neck cells.
The inner central cell enlarges and its nucleus divides into venter canal nucleus
and egg nucleus. Soon the venter canal nucleus disorganizes. Thus, a mature
archegonium has two neck cells and an egg. Neck canal cells are not formed.
The egg cell in Cycas is largest in the plant kingdom (Fig. 9.11).
(c) Development of Male gametophyte (Before pollination):
Microspore or pollen grain is the first cell of the gametophyte. The microspore
germinates in situ i.e. while within the microsporangium. Each microspore
divides asymmetrically into a 2-cells: a smaller prothallial cell and a larger
antheridialcell. The prothallial cell does not divide further while the antheridial
cell divides into a smaller generative cell near the prothallial cell and a larger
tube cell. Finally pollination takes place at 3-celled stage (a prothallial cell, a
generative cell and a tube nucleus) (Fig. 9.12).
Pollination:
In Cycas pollination is anemophilous (by wind). The 3-celled microspores
liberate from mega-sporangia are blown away by wind. Finally microspores
UNIT-2
BSC I YEAR BOTANY-II Page 45
reach on ovules and get enlarged in the pollination drop (ooze) of micropyle. As
the ooze dries up, the microspores are drawn into the pollen chamber.
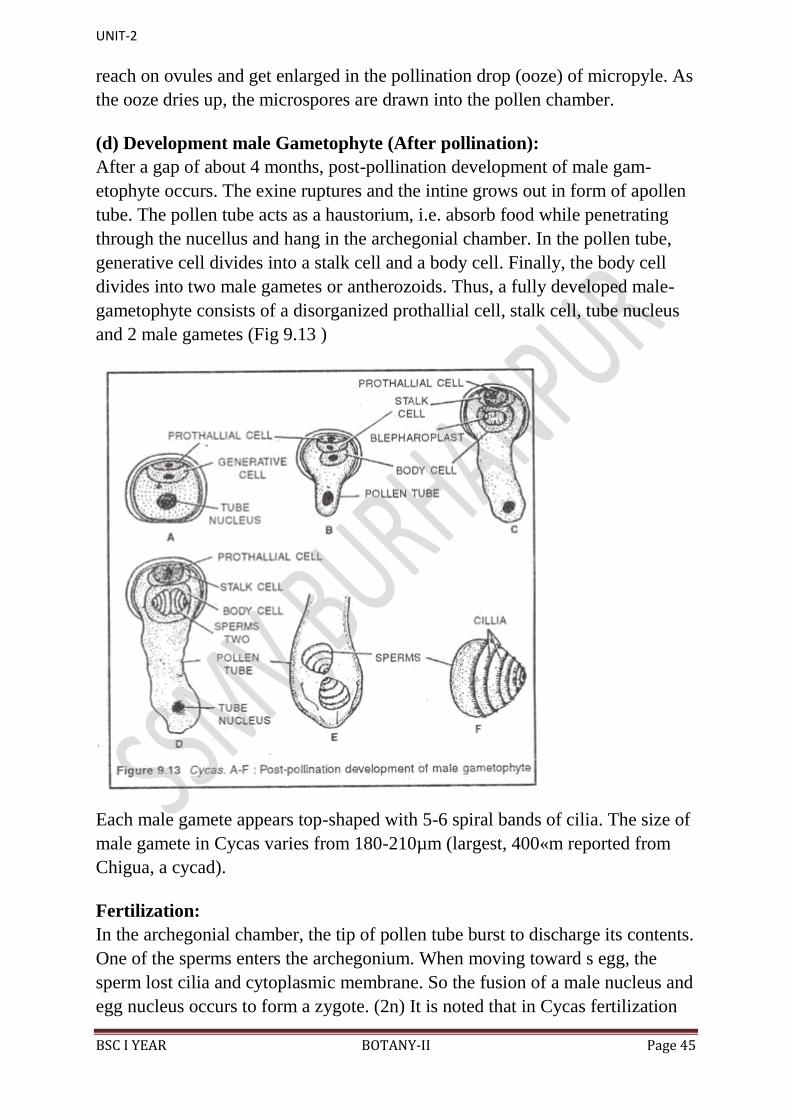
(d) Development male Gametophyte (After pollination):
After a gap of about 4 months, post-pollination development of male gam-
etophyte occurs. The exine ruptures and the intine grows out in form of apollen
tube. The pollen tube acts as a haustorium, i.e. absorb food while penetrating
through the nucellus and hang in the archegonial chamber. In the pollen tube,
generative cell divides into a stalk cell and a body cell. Finally, the body cell
divides into two male gametes or antherozoids. Thus, a fully developed male-
gametophyte consists of a disorganized prothallial cell, stalk cell, tube nucleus
and 2 male gametes (Fig 9.13 )
Each male gamete appears top-shaped with 5-6 spiral bands of cilia. The size of
male gamete in Cycas varies from 180-210µm (largest, 400«m reported from
Chigua, a cycad).
Fertilization:
In the archegonial chamber, the tip of pollen tube burst to discharge its contents.
One of the sperms enters the archegonium. When moving toward s egg, the
sperm lost cilia and cytoplasmic membrane. So the fusion of a male nucleus and
egg nucleus occurs to form a zygote. (2n) It is noted that in Cycas fertilization

UNIT-2
BSC I YEAR BOTANY-II Page 46
exhibits both siphonogamy (i. e. formation of pollen tube) and zoidogamy (i.e.
participation of ciliated male gametes).
Embryogeny:
The zygote (2n) secretes cell wall and becomes the oospore. The zygote or
oospore is the first cell of sporophyte generation. The oospore undergoes free
nuclear division followed by wall formation to form a small cellular mass called
pro-embryo. The pro-embryo differentiated into a basal embryonalzone, middle
suspensor and upper haustorium. The haustorial region remains in contact with
the free-nuclear region and soon disappear (Fig. 9.15).
The cells of embryonal zone divide and re-divide to form embryo proper which
is differentiated into two cotyledons, plumule and radicle. The suspensor
becomes enlarged and coiled to push the embryo into the nutritive endosperm.

UNIT-2
BSC I YEAR BOTANY-II Page 47
The endosperm froms a pad like tissue called coleorhiza which protects the tip
of radicle.
Since, a single Cycas ovule contains 2-8 archegonia; the same number of
embryo develops. All degenerate except one embryo that reaches maturity.
Seed formation:
As a result of post-fertilization changes the entire ovule becomes a seed.
Following changes take place in the process:
Seed Germination:
The testa of Cycas seed emits pleasant odour and sweet in taste. This causes its
dispersal by birds. The seed remain variable for a few months. Under favourable
condition, the seed germinates into a sporophyte. The seed germination in
Cycas is hypogeal i.e. the cotyledon remains underground enclosed in the
endosperm and absorb food for the growing embryo. The plumule form leafy
shoot and radicle elongates into a tap root (Fig. 9.16).

UNIT-2
BSC I YEAR BOTANY-II Page 48
Three Generations Locked in Seed:
A seed contains three generations locked one within another.
The following three generations present in a Cycas Seed are:
1. Parent Sporophyte: Seed coat and nucellus.
2. Female Gametophyte: Endosperms
3. Future Sporophyte: Embryo (radicle, cotyledons and plumule).
UNIT-2
BSC I YEAR BOTANY-II Page 49
Pinus
External Morphology of Pinus:
1. Pinus is a large, perennial, evergreen plant.
2. Branches grow spirally and thus the plant gives the appearance of a conical or
pyramidal structure.

UNIT-2
BSC I YEAR BOTANY-II Page 50
3. Sporophytic plant body is differentiated into roots, stem and acicular (needle-
like) leaves (Fig. 26).
4. A tap root with few root hair is present but it disappears soon. Later on many
lateral roots develop, which help in absorption and fixation.
5. The ultimate branches of these roots are covered by a covering of fungal
hyphae called ectotrophic mycorrhiza.
6. The stem is cylindrical and erect, and remains covered with bark. Branching
is monopodial.
7. Two types of branches are present: long shoots and dwarf shoots. These are
also known as branches of unlimited and limited growth, respectively.
8. Long shoots contain apical bud and grow indefinitely. Many scaly leaves are
present on the long shoot.

UNIT-2
BSC I YEAR BOTANY-II Page 51
9. Dwarf shoots are devoid of any apical bud and thus are limited in their
growth. They arise on the long shoot in the axil of scaly leaves.
10. A dwarf shoot (Fig. 27) has two scaly leaves called prophylls, followed by
5-13 cataphylls arranged in 2/5 phyllotaxy, and 1-5 needles.
11. The leaves are of two types, i.e., foliage and scaly.
12. Scaly leaves are thin, brown-coloured and scale like and develop only on
long as well as dwarf shoots.
13. Foliage leaves are present at the apex of the dwarf shoots only.
14. Foliage leaves are large, needle-like, and vary in number from 1 to 5 in
different species.
15. A spur (Fig. 28) is called unifoliar if only one leaf is present at the apex of
the dwarf shoot, bifoliar if two leaves are present, trifoliar if three leaves are
present, and so on.

UNIT-2
BSC I YEAR BOTANY-II Page 52
Some of the species with different types of spurs are as follows:
(i) Pinus monophylla-unifoliar (having only one needle);
(ii) P. sylvestris-bifoliar (having two needles);
(iii) P. gerardiana-trifoliar (having three needles);
(iv) P. quadrifolia-quadrifoliar (having four needles);
(v) P. wallichiana-pentafoliar (having five needles).
Anatomy of Different Parts of Pinus:
Cut thin sections of different parts of the plant (Young root, old root, young
long shoot, old long shoot, T.L.S. wood, R.L.S. wood, young dwarf shoot, old
dwarf shoot and needle), stain them separately in a safranin-fast green
combination, mount in glycerine and study. Also compare your preparations
with the permanent slides shown to you in the laboratory.
T.S. Young Root:
1. Outermost layer of the circular roots is thick-walled epiblema with many root
hair.
2. Epiblema is followed by many layers of parenchymatous cortex.
3. Inner to the cortex is present a layer of endodermis and many layers of
pericycle.
4. Vascular bundles are radially arranged and diarch to tetrarch with exarch
protoxylem.

UNIT-2
BSC I YEAR BOTANY-II Page 53
5. Protoxylem is bifurcated (Y-shaped) towards the periphery, and in between
each bifurcation is present a resin cannal (Fig. 29).
6. Phloem is present alternate to the protoxylem.
7. Pith is poorly-developed or absent.
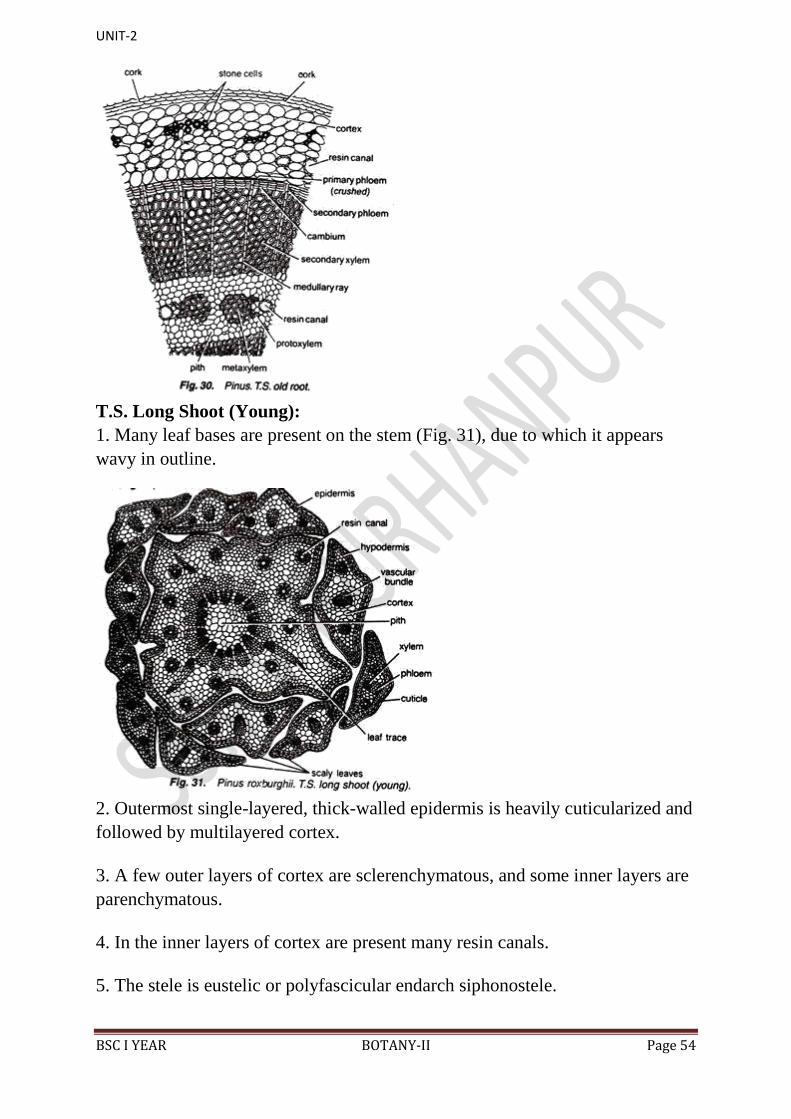
T.S. Old Root Showing Secondary Growth:
1. On the outer side are present a few layers of cork, formed by the meristematic
activity of the cork cambium.
2. Cork cambium cuts secondary cortex towards inner side.
3. Many resin canals and stone cells are present in the secondary cortex, the
cells of which are separated with the intercellular spaces.
4. Below the phloem patches develop cambium, which cuts secondary phloem
towards outer side and secondary xylem towards inner side.
5. Crushed primary phloem is present outside the secondary phloem (Fig. 30).
6. Many uniseriate medullary rays are present in the secondary xylem.
7. Primary xylem is the same as in young roots, i.e., each group is bifurcated
(Y-shaped) and a resin canal is present in between the bifurcation.
UNIT-2
BSC I YEAR BOTANY-II Page 54
T.S. Long Shoot (Young):
1. Many leaf bases are present on the stem (Fig. 31), due to which it appears
wavy in outline.
2. Outermost single-layered, thick-walled epidermis is heavily cuticularized and
followed by multilayered cortex.
3. A few outer layers of cortex are sclerenchymatous, and some inner layers are
parenchymatous.
4. In the inner layers of cortex are present many resin canals.
5. The stele is eustelic or polyfascicular endarch siphonostele.

UNIT-2
BSC I YEAR BOTANY-II Page 55
6. Vascular bundles are conjoint, collateral, open and endarch, and resemble
greatly with that of a dicot stem. 5-10 vascular bundles are arranged in a ring.
7. Endodermis and pericycle are indistinguishable.
8. Narrow xylem rays connect the cortex and pith.
9. Endarch xylem consists of only tracheids.
10. Phloem is present on the ventral side and consists of sieve cells, sieve plates,
phloem parenchyma and some albuminous cells.
11. Intrafascicular cambium is present in between the xylem and phloem.
12. Many leaf traces are also present.
13. A small parenchymatous pith is present in the centre of stem.
T.S. Long Shoot (Old):
1. Secondary growth, similar to that of a dicotyledonous stem, is present in the
old stem of Pinus.
2. Cork cambium cuts cork towards outer side and a few layers of secondary
cortex towards inner side.
3. Many tannin-filled cells and resin canals are distributed in the primary cortex.
4. Cambium cuts secondary phloem towards outer side and secondary xylem
towards inner side (Fig. 32).

UNIT-2
BSC I YEAR BOTANY-II Page 56
5. Primary phloem is crushed and pushed towards outer side by the secondary
phloem.
6. In the secondary xylem, annual rings of thin-walled spring wood (formed in
spring season) and thick- walled autumn wood (formed in autumn season) are
present alternately. Such a compact wood is called pycnoxylic (Age of the plant
can be calculated by counting the number of these annual rings).
7. Below the secondary xylem are present a few groups of endarch primary
xylem.
8. Some of the medullary rays connect the pith with the cortex and called
primary medullary rays while the others run in between secondary xylem and
secondary phloem and called secondary medullary rays.
9. Central part of the stem is filled with the parenchymatous pith.
10. Resin canals are present in cortex, secondary xylem, primary xylem and
rarely in the pith.
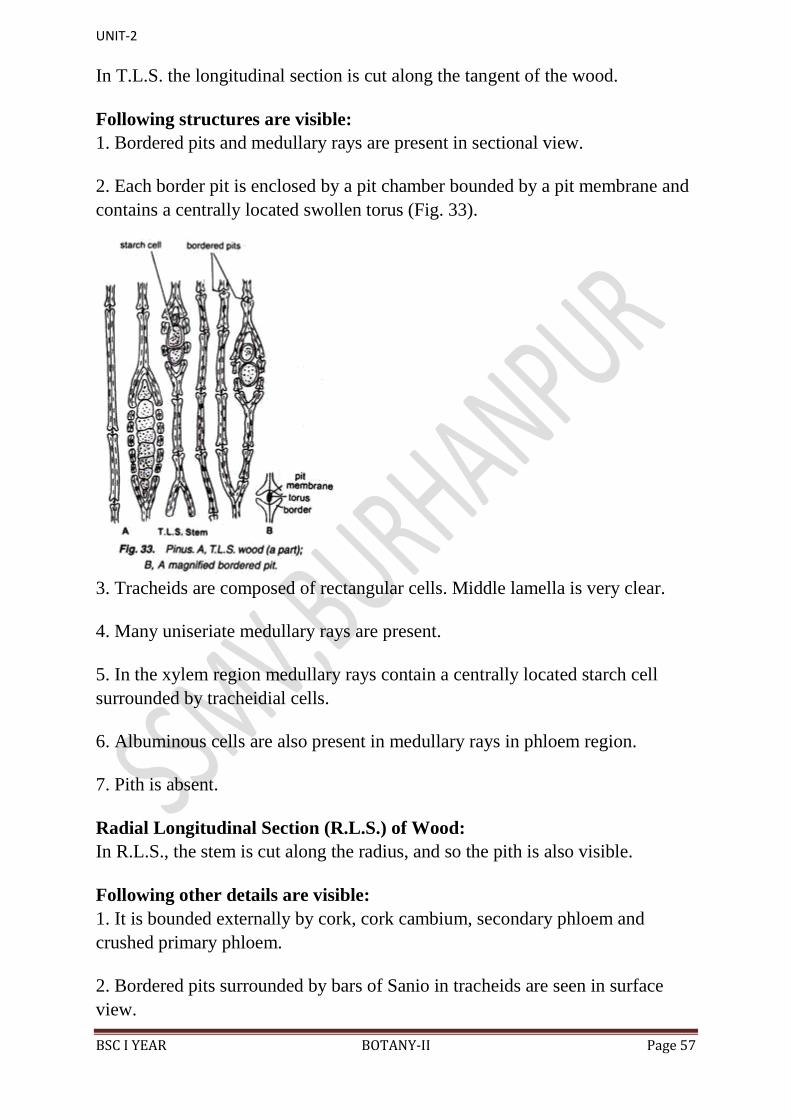
Tangential Longitudinal Section (T.L.S.) of Wood:
UNIT-2
BSC I YEAR BOTANY-II Page 57
In T.L.S. the longitudinal section is cut along the tangent of the wood.
Following structures are visible:
1. Bordered pits and medullary rays are present in sectional view.
2. Each border pit is enclosed by a pit chamber bounded by a pit membrane and
contains a centrally located swollen torus (Fig. 33).
3. Tracheids are composed of rectangular cells. Middle lamella is very clear.
4. Many uniseriate medullary rays are present.
5. In the xylem region medullary rays contain a centrally located starch cell
surrounded by tracheidial cells.
6. Albuminous cells are also present in medullary rays in phloem region.
7. Pith is absent.
Radial Longitudinal Section (R.L.S.) of Wood:
In R.L.S., the stem is cut along the radius, and so the pith is also visible.
Following other details are visible:
1. It is bounded externally by cork, cork cambium, secondary phloem and
crushed primary phloem.
2. Bordered pits surrounded by bars of Sanio in tracheids are seen in surface
view.

UNIT-2
BSC I YEAR BOTANY-II Page 58
3. Uniseriate medullary rays run horizontally.
4. In the xylem region thick medullary ray cells are surrounded by ray tracheids
(Fig. 34).
5. Thin-walled ray parenchyma is also present.
6. Xylem is separated from phloem with the help of cambium.
7. Albuminous cells are present in medullary ray in the phloem region.
8. Phloem consists of sieve tubes, sieve plates and phloem parenchyma.
9. Pith is present.
T.S. Dwarf Shoot (Young):
It is exactly similar to that of T.S. of young long shoot except following
differences:
1. The number of the resin canals present in the cortex is not indefinite but
generally six (Fig. 35).
Though it is variable in different species.

UNIT-2
BSC I YEAR BOTANY-II Page 59
2. The number of the vascular bundles is also generally six. However, it is also
variable in different species.
3. Pith in dwarf shoot is comparatively smaller than the long shoot.
4. Structure of the vascular bundles is same, i.e., conjoint, collateral, open and
endarch.
T.S. Dwarf Shoot (Old):
1. It is also similar to old long shoot in many aspects.
2. Cork, cork cambium and secondary cortex are not normally present, but the
epidermis surrounded externally by scaly leaves and followed internally by
multilayered cortex is present.
3. Inner to the cortex is crushed primary phloem, secondary phloem, cambium
and secondary xylem with medullary rays (Fig. 36). Protoxylem is endarch.

UNIT-2
BSC I YEAR BOTANY-II Page 60
4. A small pith with some tannin cells is present in the centre.
If a section of distal end of dwarf shoot is cut, the needles get separated, each
having the same structure. In a bifoliar spur two needles are present while in a
trifoliar spur there are present three foliage leaves or needles (Fig. 37).
T.S. Needle (Foliage Leaf):
1. It is circular in outline in Pinus monophylla, semicircular in P. sylvestris and
triangular (Fig. 38) in P. longifolia, P. roxburghii, etc.
2. Outermost layer is epidermis, which consists of thick-walled cells. It is
covered by a very strong cuticle.
3. Many sunken stomata are present on the epidermis (Fig. 38).

UNIT-2
BSC I YEAR BOTANY-II Page 61
4. Each stoma opens internally into a substomatal cavity and externally into a
respiratory cavity or vestibule.
5. Below the epidermis are present a few layers of thick-walled
sclerenchymatous hypodermis. It is well-developed at ridges.
6. In between the hypodermis and endodermis is present the mesophyll tissue.
7. Cells of the mesophyll are polygonal and filled with chloroplasts. Many peg-
like infoldings of cellulose also arise from the inner side of the wall of meso-
phyll cells.
8. Few resin canals are present in the mesophyll, adjoining the hypodermis.
Their number is variable but generally they are two in number.
9. Endodermis is single-layered with barrel-shaped cells and clear casparian
strips.
10. Pericycle is multilayered and consists of mainly parenchymatous cells and
some sclerenchymatous cells forming T-shaped girder, which separates two
vascular bundles (Fig. 38). Transfusion tissue consists of tracheidial cells.
11. Two conjoint and collateral vascular bundles are present in the centre. These
are closed but cambium may also present in the sections passing through the
base of the needle.
12. Xylem lies towards the angular side and the phloem towards the convex side
of the needle.

UNIT-2
BSC I YEAR BOTANY-II Page 62
Reproductive Structures of Pinus:
1. Plant body is sporophytic.
2. Pinus is monoecious, and male and female flowers are present in the form of
cones or strobili on the separate branches of the same plant.
3. Many male cones are present together in the form of clusters, each of which
consists of many microsporophylls. The female cones consist of
megasporophylls.
4. The male cones on the plant develop much earlier than the female cones.
Male Cone:
Separate a male cone from the cluster, study its structure, cut its longitudinal
section, study the structure of a single microsporophyll, and also prepare a slide
of pollen grains and study.
1. The male cones develop in clusters (Fig. 39) in the axil of scaly leaves on
long shoot.
2. They replace the dwarf shoots of the long shoot.
3. Each male cone is ovoid in shape and ranges from 1.5 to 2.5 cm. in length
(Fig. 40).

UNIT-2
BSC I YEAR BOTANY-II Page 63
4. A male cone (Fig. 41) consists of a large number of microsporophylls
arranged spirally on the cone axis.
5. Each microsporophyll is small, membranous, brown-coloured structure.
6. A microsporophyll (Fig. 41) is comparable with the stamen of the flower of
angiosperms because it consists of a stalk (=filament) with a terminal leafy
expansion (= anther), the tip of which is projected upwards and called
apophysis.
7. Two pouch-like microsporangia (= pollen sacs) are present on the abaxial or
undersurface of each microsporophyll. In each microsporangium are present
many microspores (= pollen grains).

UNIT-2
BSC I YEAR BOTANY-II Page 64
8. Each microspore or pollen grain is a rounded and yellow-coloured, light,
uninucleate structure with two outer coverings, i.e., thick outer exine and thin
inner intine (Fig. 42).
9. The exine protrudes out on two sides in the form of two balloon-shaped
wings. Wings help in floating and dispersal of pollen grains.
10. Wings help in floating and dispersal of pollen grains.
11. A few microsporophylls of lower side of cone are sterile. Sporangia are also
not present on the adaxial surface of each microsporophyll of the male cone.
Female cone:
Observe the external features and longitudinal section of a young female cone
and also study 1st year, 2nd year and 3rd year female cones.
1. Female cone develops either solitary or in groups of 2 to 4.
2. They also develop in the axil of scaly leaves on long shoots (Fig. 43) like
male cones.
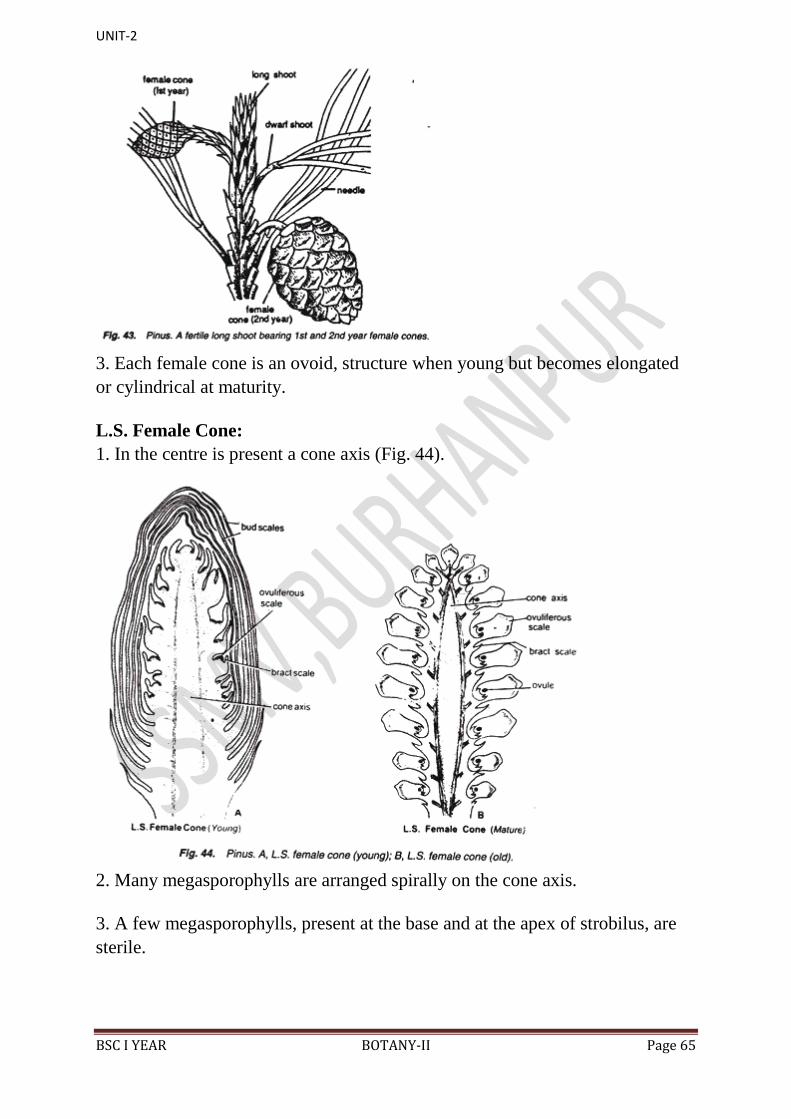
UNIT-2
BSC I YEAR BOTANY-II Page 65
3. Each female cone is an ovoid, structure when young but becomes elongated
or cylindrical at maturity.
L.S. Female Cone:
1. In the centre is present a cone axis (Fig. 44).
2. Many megasporophylls are arranged spirally on the cone axis.
3. A few megasporophylls, present at the base and at the apex of strobilus, are
sterile.

UNIT-2
BSC I YEAR BOTANY-II Page 66
4. Megasporophylls present in the middle of the strobilus are very large and
they decrease in size towards the base and apex.
5. Each megasporophyll consists of two types of scales, known as bract scales
and ovuliferous scales.
6. Bract scales are thin, dry, membranous, brown- coloured structures having
fringed upper part. These are also called carpellary scales.
7. An ovuliferous scale is present on the upper surface of each bract scale.
8. Each oruliferous scale is woody, bigger and stouter than bract scale and it is
triangular in shape. A broad sterile structure, with pointed tip, is present at the
apex of these scales. This is called apophysis.
9. At the base of upper surface of each ovuliferous scale are present two sessile
and naked ovules.
10. Micropyle of each ovule faces towards the cone axis.
11. Each ovule is orthotropous, and it remains surrounded by a single
integument, consisting of an outer fleshy, a middle stony and an inner fleshy
layer. It opens with a mouth opening called micropyle.
12. Integument surrounds the megasporangium or nucellus.
13. Just opposite the micropyle is present a pollen chamber.
14. In the endosperm or female gametophyte are present 2 to 5 archegonia.
Female Cone of 1st Year:
1. It is oval (Fig. 45) in shape.
2. It ranges from 1 to 4 cm. in length.

UNIT-2
BSC I YEAR BOTANY-II Page 67
3. It is green to reddish-green in colour.
4. It is attached with the help of a short stalk on the long shoot.
5. Megasporophylls are arranged very close to each other, and so the cone is a
compact structure.
Female Cone of 2nd Year:
1. It is elongated and larger than the first year cone.
2. It ranges from 5 to 15 cm. or more in length.
3. It is red-coloured structure.
4. It is woody in nature.
5. Megasporophylls are compactly arranged (Fig. 46) but not so compact as in
1st year cone.
6. Seeds are present inside in the later stages (Fig. 46).
Female Cone of 3rd Year:
1. It is elongated or roughly rounded in shape.
2. It is also woody in nature like the 2nd year cone.
3. Megasporophylls (Fig. 47) are loosely arranged.
4. Seeds are dispersed from 3rd year cone.

UNIT-2
BSC I YEAR BOTANY-II Page 68
Seed:
1. Both the ovules of each ovuliferous scale develop into seeds (Fig. 48).
2. Each seed contains a large membranous wing formed from the ovuliferous
scale.

UNIT-2
BSC I YEAR BOTANY-II Page 69
Anatomy of seed shows following (Fig. 48C) details:
1. It is enveloped by a seed coat developed from the middle stony layer of the
ovule.
2. Inner fleshy layer may survive in the form a thin membrane. Outer fleshy
layer disappears.
3. A thin, membranous and papery structure, called perisperm, develops inner to
the seed coat.
4. Well-developed endosperm is present.
5. In the centre is present the embryo consisting of a hypocotyle, radicle,
plumule and 2 to 14 or more cotyledons.
Identification:
(i) Sporophytic plant body differentiated into roots, stem and leaves.
(ii) Ovules naked.
(iii) Xylem lacks vessels.
(iv) Phloem lacks companion cells.
(v) Sex organs are present in the form of cones…………. Gymnosperms
(b)(i) Leaves needle shaped.
(ii) Pycnoxylic wood.
(iii) Seeds show bilateral symmetry.
(iv) Male cones in clusters ………………. Coniferopsida
(c) (i) Presence of scaly and foliage leaves.
(ii) Foliage leaves are needle like.
(iii) Wood pycnoxylic and xylem contains bordered pits.
(iv) Pollen grains are winged.
(v) Resin canals present……………. coniferales

UNIT-2
BSC I YEAR BOTANY-II Page 70
(d)(i) Plant is monoecious.
(ii) Female cone is woody.
(iii) Presence of bract and ovuliferous scales in female cone.
(iv) Seeds are winged
(v) Polyembryony present………………………………………….. Pinaceae
(e) (i) Plant conical in appearance.
(ii) Presence of ectotrophic mycorrhiza on roots.
(iii) Presence of monofoliar to pentafoliar spurs.
(iv) Leaves needle-like.
(v) Presence of resin canals in pith, cortex and wood.
(vi) Clusters of male cones present.
(vii) Winged pollen grains and winged seeds.
(viii) Many cotyledons are present………… Pinus.
Life Cycle of Pinus
Sporophyte of Pinus:
Pinus is a tall evergreen tree giving rise to a series of widespread horizontal
branches (Fig. 1.57A). In each year, a whorl of branches is produced in the axil
of scale leaves. The branching is restricted to the upper part of the stem, thus
giving the tree a pyramid-like appearance. The main stem is cylindrical and
covered with scaly bark.
The branches are dimorphic, bearing two types of shoots: long shoots and dwarf
shoots, or spurs or brachyblasts (Fig. 1.57B). Pinus exhibits two types of leaves,
the scale leaves and the green acicular foliage leaves called needles (Fig.
1.57B). The plant has a tap root system which becomes elongated at maturity
and possesses strong lateral roots.

UNIT-2
BSC I YEAR BOTANY-II Page 71
The plants are monoecious where the male and female cones are borne on
separate branches in the same plant.
Reproduction:
Pinus reproduces sexually. Pinus is monoecious, but the male and the female
cones are produced on separate branches of the same plant. The male cones
develop on the lower branches, while the female cones are formed on the upper
branches. The male cones, which replace the dwarf shoots, develop in clusters
on the base of the current year‟s long shoot at early spring (Fig. 1.61 A).
The number of male cones in a cluster varies considerably from 15 (P.
wallichiana) to 140 (P. roxburghii). At the onset of spring, the male cones fall
off and simultaneously the young female cones are borne in pairs or in clusters
round the tip of the long shoot (Fig. 1.61 B).
The female cones grow very slowly and the growth may persist for several
years. Thus, the female cones of different ages may be seen in acropetal
succession in the long shoot (Fig. 1.61B).

UNIT-2
BSC I YEAR BOTANY-II Page 72
i. Male Cone:
The male cone is small (2-4 cm in length) and oval in shape that develops in the
axil of scale leaves. The male cone has a central axis on which 60-150
microsporophylls are spirally arranged around the axis (Fig. 1.62A).
A single microsporophyll is a membranous stalked structure with a distal
expanded roughly triangular sterile part called apophysis (Fig. 1.62B). Each
microsporophyll bears two sac-like microsporangia on the abaxial surface.
The development of microsporangia is of eusporangiate type, i.e., it develops
from a group of hypodermal cells of the microsporophyll. A mature
microsporangium consists of a multilayered wall, tapetum and microspore
mother cells (Fig. 1.62D). Each microspore mother cell — by meiotic division
— produces four microspores or pollen grains.
Thus, at maturity, a single microsporangium contains numerous pale yellow
pollen grains. The pollen grains are boat-shaped with monosulcate apertures and
are bounded by two concentric wall layers: the outer thick exine and the inner
thin intine (Fig. 1.62C).
The exine on the lateral sides of the pollen is expanded to form two wings
(sacci). Thus the pollen grains of Pinus are bisaccate indicating their
anemophilous mode of pollination. The dehiscence of sporangia takes place by
longitudinal slit in dry and warm environment.
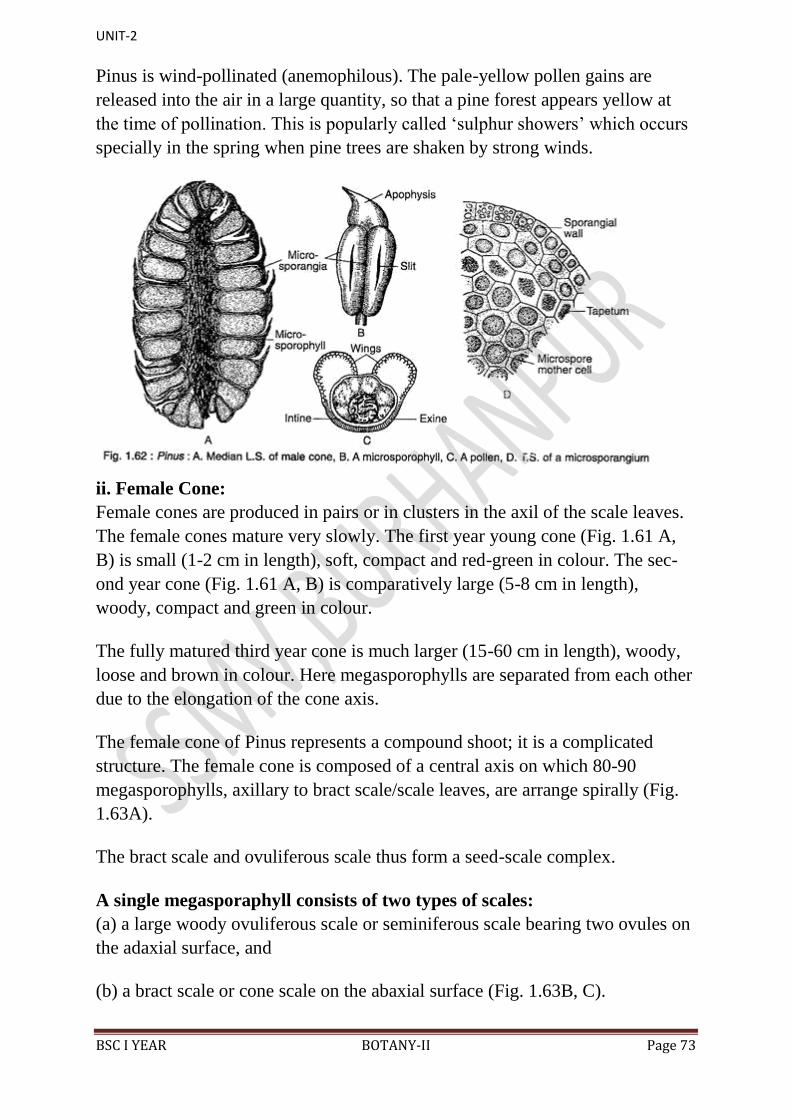
UNIT-2
BSC I YEAR BOTANY-II Page 73
Pinus is wind-pollinated (anemophilous). The pale-yellow pollen gains are
released into the air in a large quantity, so that a pine forest appears yellow at
the time of pollination. This is popularly called „sulphur showers‟ which occurs
specially in the spring when pine trees are shaken by strong winds.
ii. Female Cone:
Female cones are produced in pairs or in clusters in the axil of the scale leaves.
The female cones mature very slowly. The first year young cone (Fig. 1.61 A,
B) is small (1-2 cm in length), soft, compact and red-green in colour. The sec-
ond year cone (Fig. 1.61 A, B) is comparatively large (5-8 cm in length),
woody, compact and green in colour.
The fully matured third year cone is much larger (15-60 cm in length), woody,
loose and brown in colour. Here megasporophylls are separated from each other
due to the elongation of the cone axis.
The female cone of Pinus represents a compound shoot; it is a complicated
structure. The female cone is composed of a central axis on which 80-90
megasporophylls, axillary to bract scale/scale leaves, are arrange spirally (Fig.
1.63A).
The bract scale and ovuliferous scale thus form a seed-scale complex.
A single megasporaphyll consists of two types of scales:
(a) a large woody ovuliferous scale or seminiferous scale bearing two ovules on
the adaxial surface, and
(b) a bract scale or cone scale on the abaxial surface (Fig. 1.63B, C).
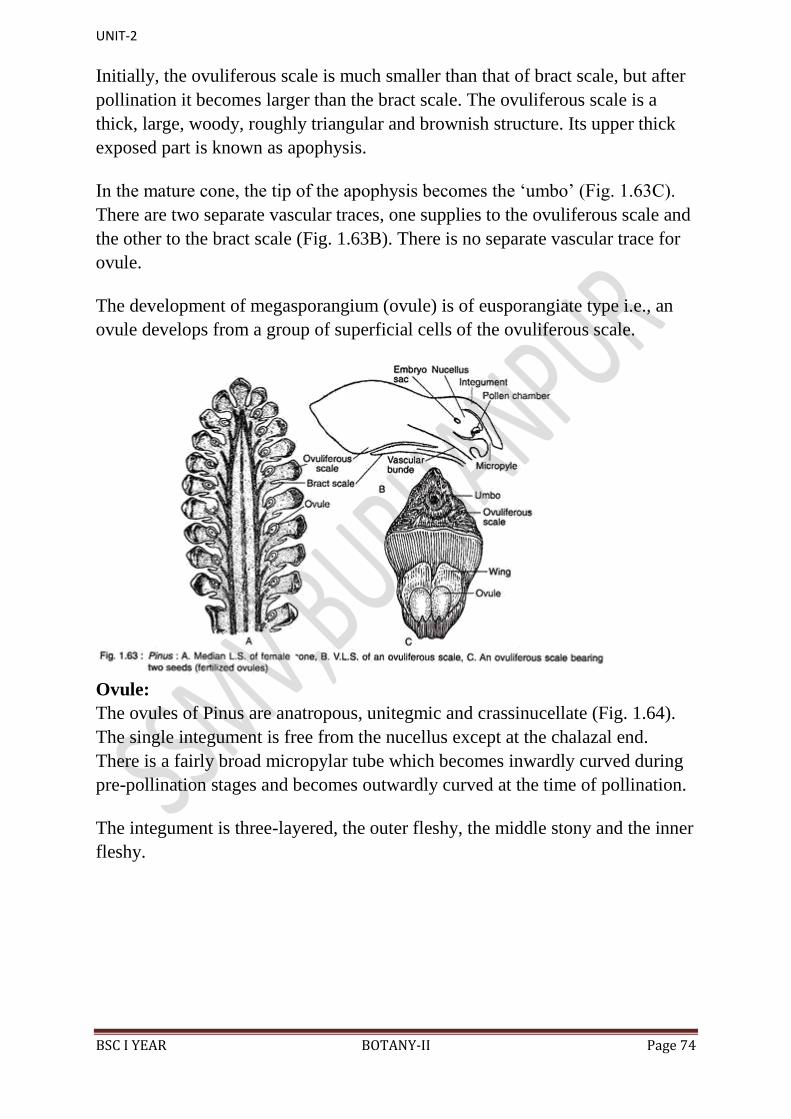
UNIT-2
BSC I YEAR BOTANY-II Page 74
Initially, the ovuliferous scale is much smaller than that of bract scale, but after
pollination it becomes larger than the bract scale. The ovuliferous scale is a
thick, large, woody, roughly triangular and brownish structure. Its upper thick
exposed part is known as apophysis.
In the mature cone, the tip of the apophysis becomes the „umbo‟ (Fig. 1.63C).
There are two separate vascular traces, one supplies to the ovuliferous scale and
the other to the bract scale (Fig. 1.63B). There is no separate vascular trace for
ovule.
The development of megasporangium (ovule) is of eusporangiate type i.e., an
ovule develops from a group of superficial cells of the ovuliferous scale.
Ovule:
The ovules of Pinus are anatropous, unitegmic and crassinucellate (Fig. 1.64).
The single integument is free from the nucellus except at the chalazal end.
There is a fairly broad micropylar tube which becomes inwardly curved during
pre-pollination stages and becomes outwardly curved at the time of pollination.
The integument is three-layered, the outer fleshy, the middle stony and the inner
fleshy.
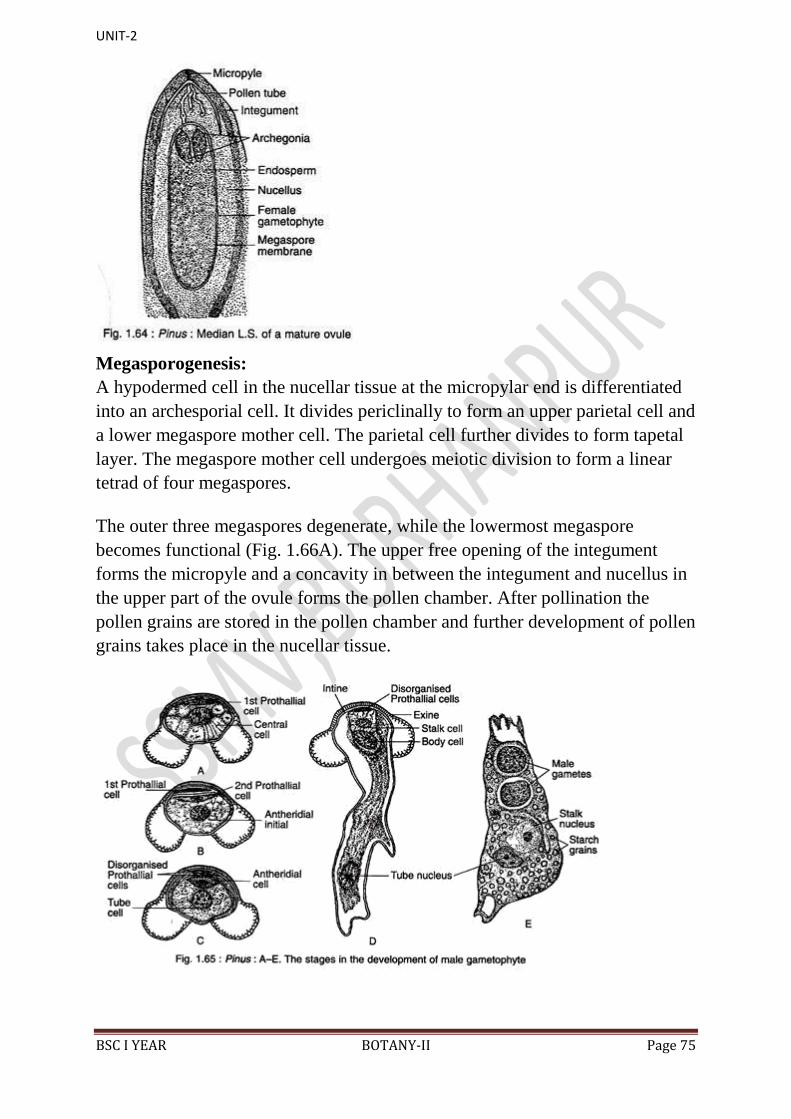
UNIT-2
BSC I YEAR BOTANY-II Page 75
Megasporogenesis:
A hypodermed cell in the nucellar tissue at the micropylar end is differentiated
into an archesporial cell. It divides periclinally to form an upper parietal cell and
a lower megaspore mother cell. The parietal cell further divides to form tapetal
layer. The megaspore mother cell undergoes meiotic division to form a linear
tetrad of four megaspores.
The outer three megaspores degenerate, while the lowermost megaspore
becomes functional (Fig. 1.66A). The upper free opening of the integument
forms the micropyle and a concavity in between the integument and nucellus in
the upper part of the ovule forms the pollen chamber. After pollination the
pollen grains are stored in the pollen chamber and further development of pollen
grains takes place in the nucellar tissue.
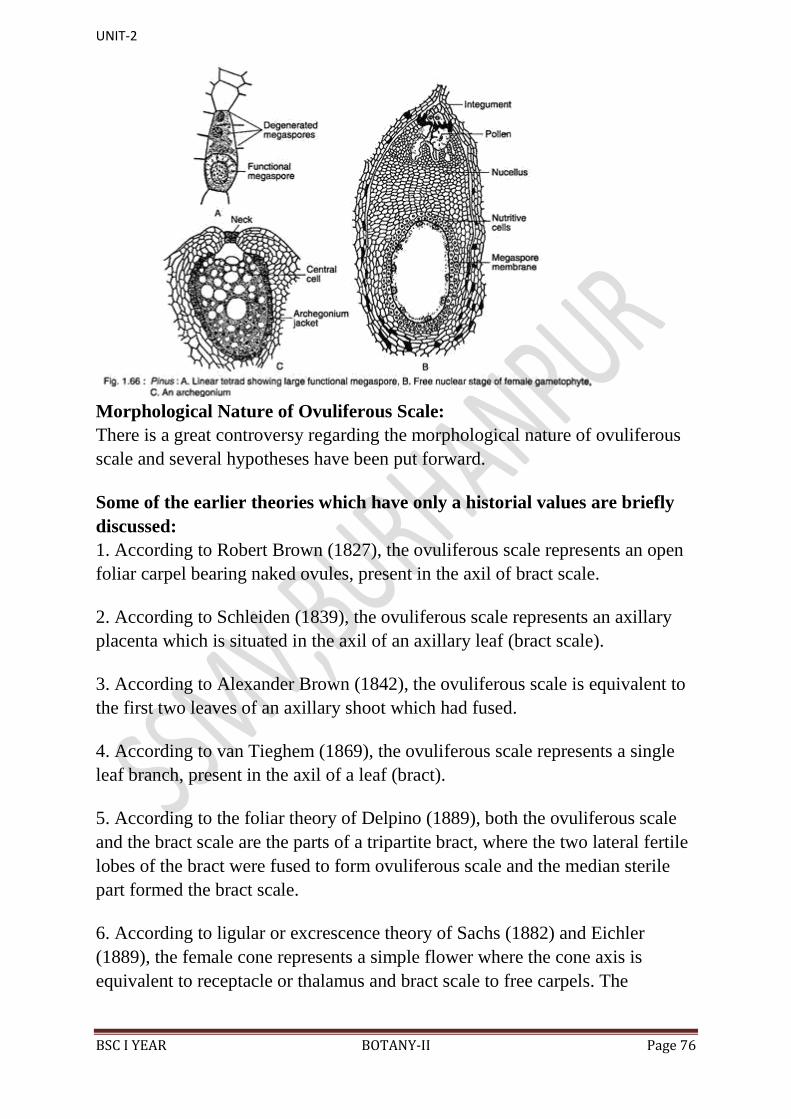
UNIT-2
BSC I YEAR BOTANY-II Page 76
Morphological Nature of Ovuliferous Scale:
There is a great controversy regarding the morphological nature of ovuliferous
scale and several hypotheses have been put forward.
Some of the earlier theories which have only a historial values are briefly
discussed:
1. According to Robert Brown (1827), the ovuliferous scale represents an open
foliar carpel bearing naked ovules, present in the axil of bract scale.
2. According to Schleiden (1839), the ovuliferous scale represents an axillary
placenta which is situated in the axil of an axillary leaf (bract scale).
3. According to Alexander Brown (1842), the ovuliferous scale is equivalent to
the first two leaves of an axillary shoot which had fused.
4. According to van Tieghem (1869), the ovuliferous scale represents a single
leaf branch, present in the axil of a leaf (bract).
5. According to the foliar theory of Delpino (1889), both the ovuliferous scale
and the bract scale are the parts of a tripartite bract, where the two lateral fertile
lobes of the bract were fused to form ovuliferous scale and the median sterile
part formed the bract scale.
6. According to ligular or excrescence theory of Sachs (1882) and Eichler
(1889), the female cone represents a simple flower where the cone axis is
equivalent to receptacle or thalamus and bract scale to free carpels. The

UNIT-2
BSC I YEAR BOTANY-II Page 77
ovuliferous scale represents an outgrowth of the carpel (bract scale) as in ligule
of Selaginella or the placenta of angiosperms.
7. According to brachyblast theory of Braun, the female cone is equivalent to an
inflorescence, where ovuliferous scale represents a determinate axillary shoot
bearing two fertile leaves with a single ovule on the dorsal surface of each leaf.
8. Modern hypothesis: On the basis of the comparative studies of the fossil
Cordaitales and Voltziales members, Florin (1951) introduced a terminology
„seed-cone complex‟ for ovuliferous scale and bract scale, According to Florin,
the female cone of Pinus represents an inflorescence where cone axis is the
peduncle, bract scale is a true bract and the ovuliferous scale is a rudimentary
female flower i.e., a modified reproductive shoot.
This hypothesis has been substantiated by fossil evidences. The occurrence of
several Upper Carboniferous to Triassic seed-cone complex in the members of
Cordaitales and Voltziales supports the above hypothesis (Fig. 1.56A-D). In
Cordaianthus, Emporia and Ernestiodendron of Upper Carboniferous, the cone
axis bears several secondary female reproductive shoots in the axils of bracts.
Each secondary shoot consists of many spirally-arranged sterile and fertile (with
terminal ovule) scales (showing radial symmetry).
The further evolution took place in the Triassic Voltziales like Glyptolepis,
Wallchiostrobus, etc. where the scales were planated (showing bilateral
symmetry) and tangentially fused to form a rudimentary ovuliferous scale
following Telome hypothesis. Thus, it confirms that the ovuliferous scale is a
modified reduced secondary female reproductive shoot and bract scale is
actually a reduced bract.
Gametophyte of Pinus:
The spore is the first phase of gametophyte generation. The microspore or
pollen grain represents the male gametophyte, while the megaspore represents
the first stage of female gametophyte which develops into a female
gametophyte.

UNIT-2
BSC I YEAR BOTANY-II Page 78
i. Development of Male Gametophyte before Pollination:
The basic pattern of the development of male gametophyte of Pinus is similar to
that of Ginkgo. The pollen grains undergo endosporic development. The pollen
nucleus divides mitotically to produce a small lens-shaped first prothallial cell
towards the proximal end and a large central cell on the distal end (Fig. 1.65A).
The central cell again cuts off a second prothallial cell and an antheridial initial
(Fig. 1.65B). Both the prothallial cells are ephemeral and the second prothallial
cell remains attached to the first prothallial cell.
The antheridial initial divides to form a small antheridial cell and a large tube
cell (Fig. 1.65C). The pollen grains are released from the microsporangium at
the 4- celled stage (2 prothallial cells, an antheridial cell and a tube cell).
ii. Development of Male Gametophyte after Pollination:

UNIT-2
BSC I YEAR BOTANY-II Page 79
After pollination, the 4-celled pollen stores in the pollen chamber and remains
ungerminated for about 11 months. The pollen develops the next spring. The
tube cell of the pollen comes out through the pollen aperture in the form of a
pollen tube. The pollen tube proceeds towards the archegonium, penetrating the
nucellar tissue of the ovule. The antheridial cell within the pollen tube divides to
form a stalk cell and a spermatogenous (body) cell (Fig. 1.65D).
The spermatogenous cell divides to form two male nuclei just prior to
fertilisation (Fig. 1.65E). The male nuclei are actually the male gametes which
are non-motile and ephemeral.
iii. Development of Female Gametophyte:
The female gametophyte of Pinus develops from the functional megaspore
which enlarges considerably (Fig. 1.66A). The nucleus of the megaspore
divides mitotically forming a large number of nuclei unaccompanied by wall
formation.
The number of free nuclei is constant for a particular species, say for example, it
is 2,000 for P. gerardiana and 2,500 for P. roxburghii and P. wallichiana. With
the increase in size, the megaspore develops a vacuole at the centre which
forces by cytoplasm along with nuclei towards the periphery.
Thus, the nuclei lie in a thin film of cytoplasm around the vacuole (Fig. 1.66B).
Thereafter, the cell wall formation starts in a centripetal fashion, from periphery
inwards. At this stage, numerous radially elongated multinucleate tube-like cells
called alveoli are formed and the wall formation takes place through alveoli.
Each alveolus containing a nucleus at its mouth directs its growth.
Then, cross-walls are laid down on each alveolus to form uninucleate cells. In
this way, the entire gametophyte becomes cellular and the tissue thus formed
represents endosperm or female prothallus (Fig. 1.64).
iv. Development of Archegonia:
The development of archegonia in Pinus is similar to that of Ginkgo. Two to
four cells of the female gametophyte at the micropylar end enlarge in size and
have dense cytoplasm and prominent nuclei. These cells function as archegonial
initials. Each archegonial initial divides periclinally to form an outer small
primary neck initial and a large central cell.
The primary neck initial divides by two vertical walls at right angles to each
other forming a neck of four cells. Thus, the four neck cells are arranged in a

UNIT-2
BSC I YEAR BOTANY-II Page 80
single tier as in P. roxburghii and P wallichiana. However, in P. rigida, P.
austriaca the four neck cells again divide transversely to form eight cells which
are arranged in two tiers. The central cell enlarges very rapidly and its cyto-
plasm becomes vacuolated (Fig. 1.66C).
The nucleus of the central cell divides into an upper ephemeral ventral canal
cell and a large egg cell. A nutritive layer called archegonial jacket is
differentiated around the archegonium. The nucellar tissue above the archegonia
disorganises to form an archegonial chamber.
v. Pollination:
Pinus is anemophilous i.e., wind-pollinated. The pollen grains are dispersed and
remain suspended in the air for some time. At the same time, the nucellar beak
in the ovule disorganises forming a viscous sugary liquid containing glucose,
fructose and sucrose. This fluid comes out in a cyclic phenomenon (24 hr.
cycle) through the micropyle in the form of a pollination drop either at night or
in the early hours of morning. The pollen grains are caught in the pollination
drop and are collected in the pollen chamber as a result of drying off the fluid.
The mouth of the micropyle is then sealed from the outer environment.
vi. Fertilisation:
The fertilisation takes place after one year of pollination. The pollen tube enters
the tip of the archegonium by forcing itself between the cells of the nucellus.
The pollen tube wall is disintegrated by the enzymes secreted from the egg and
eventually two male nuclei are released. One of the male nuclei fuses with the
egg cell and thus a zygote is formed.
vii. Development of Proembryo:
The zygote nucleus divides by two mitotic divisions forming four nuclei which
move to the base of zygote (Fig. 1.67A). All the four nuclei are arranged in one
plane and only two nuclei are thus visible in lateral view. A synchronous
division gives rise to eight nuclei arranged in two tiers of four each (Fig.
1.67B).
Thus, the upper group of four cells forms the primary upper tier (these cells
have no wall towards upper side) and lower group of four cells forms the
primary embryonic tier (these cells are bounded by wall all around).
The internal division in both the tiers forms four tiers of four cells each (Fig.
1.67C), and the proembryo thus consists of 16 cells. The lowest tier is known as
embryonal tier, (Fig. 1.67D) which further divides to form embryo. The next
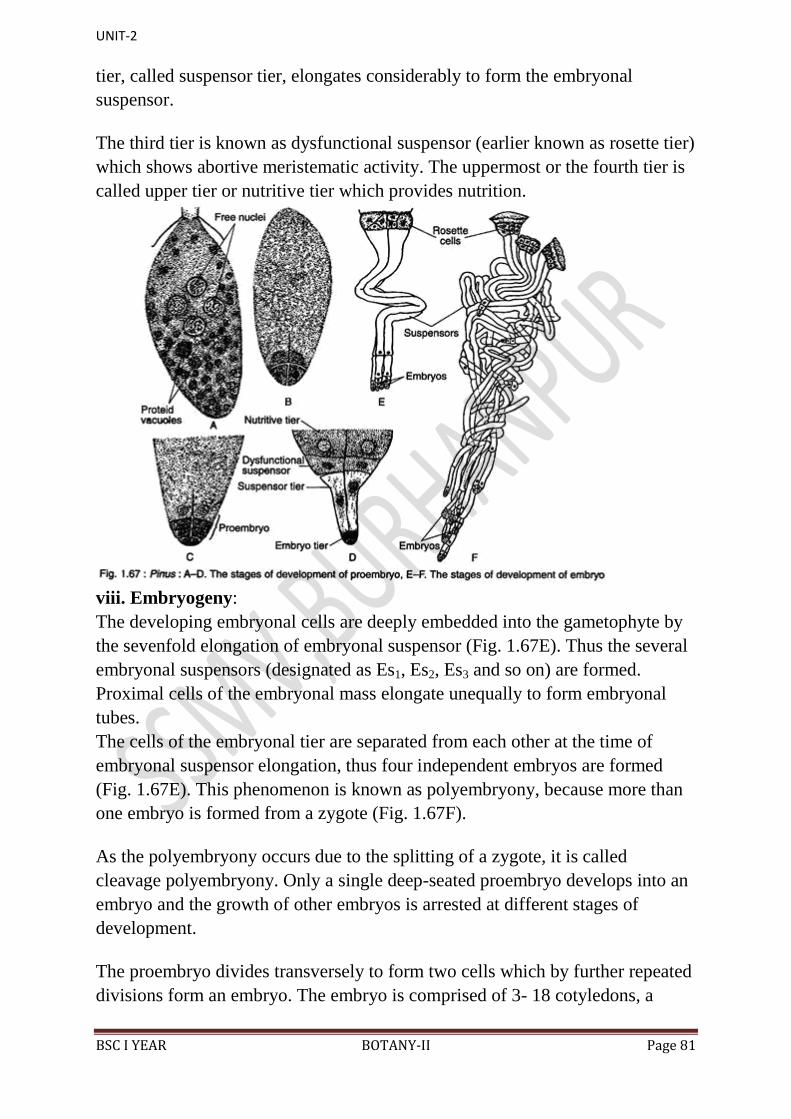
UNIT-2
BSC I YEAR BOTANY-II Page 81
tier, called suspensor tier, elongates considerably to form the embryonal
suspensor.
The third tier is known as dysfunctional suspensor (earlier known as rosette tier)
which shows abortive meristematic activity. The uppermost or the fourth tier is
called upper tier or nutritive tier which provides nutrition.
viii. Embryogeny:
The developing embryonal cells are deeply embedded into the gametophyte by
the sevenfold elongation of embryonal suspensor (Fig. 1.67E). Thus the several
embryonal suspensors (designated as Es1, Es2, Es3 and so on) are formed.
Proximal cells of the embryonal mass elongate unequally to form embryonal
tubes.
The cells of the embryonal tier are separated from each other at the time of
embryonal suspensor elongation, thus four independent embryos are formed
(Fig. 1.67E). This phenomenon is known as polyembryony, because more than
one embryo is formed from a zygote (Fig. 1.67F).
As the polyembryony occurs due to the splitting of a zygote, it is called
cleavage polyembryony. Only a single deep-seated proembryo develops into an
embryo and the growth of other embryos is arrested at different stages of
development.
The proembryo divides transversely to form two cells which by further repeated
divisions form an embryo. The embryo is comprised of 3- 18 cotyledons, a

UNIT-2
BSC I YEAR BOTANY-II Page 82
distinct epicotyl root axis and a hypocotyl shoot axis with remnants of suspen-
sor (Fig. 1.68A). Both the two-year and three- year reproductive cycles are
exemplified by Pinus species.
In two-year type, the pollination and fertilisation take place in late spring of first
and second year, respectively. In three-year type the pollination takes place in
spring of first year, and fertilisation in the spring of third year, a lapse of two
years. The seeds shed in autumn.
Seeds of Pinus:
The seeds are endowed with a well-developed wing which is thin and papery
and is easily detachable at maturity (Fig. 1.68B). The outer fleshy layer of
integument and part of ovuliferous scale contribute to the wing formation (Fig.
1.68C). The seeds are usually dispersed by wind. The embryo remains
embedded within the endosperm.
The seeds of Pinus remain viable for a long time. The germination of seed is
epigeal (Fig. 1.69A, B).

UNIT-2
BSC I YEAR BOTANY-II Page 83
Origin and Relationship of Pinus:
The conifers are large and diversified group of extant gymnosperms. The
Pinaceae is the largest family of the modern conifers. The conifers had evolved
from the members of Permo-Carboniferous Voltziales, commonly known as
“transition conifers”. They reached their climax in mid-Mesozoic forming
extensive forests in Northern Europe.
Among the living families, Pinaceae and Araucariaceae are more primitive,
probably evolved during the Triassic age. On the other hand, Cupressaceae and
Cephalotaxaceae are comparatively younger which have evolved probably
during Upper Jurassic to Lower Cretaceous period.
Pinus is the large and well-representative genus of the family Pinaceae. It
indicates relationship with Cycadales, Ginkgoales and Cordaitales.
Relation to Cycadales:
The similarities between Pinus and Cycadales are:
i. The stem anatomy shows a broad pith, large cortex and centripetal wood.
ii. The presence of haplocheilic, sunken stomata in leaves.
iii. The presence of leaf sclerenchyma.
iv. The seeds are with three-layered integument.
v. The presence of free-nuclear divisions in the development of female gameto-
phyte.
Relation to Ginkgoales:
The resemblance between Pinus and Ginkgoales are:

UNIT-2
BSC I YEAR BOTANY-II Page 84
i. The plants are profusely branched showing excurrent habit.
ii. The shoots are dimorphic, bearing long shoots and dwarf shoots
iii. The wood is pycnoxylic.
iv. The leaves are with haplocheilic sunken stomata.
v. The mature wood shows pitting and Bars of Sanio.
Relation to Cordaitales:
Conifers have derived from the members of Voltziales which are considered as
„transition conifers‟ between Cordaitales and Conifers. Thus, Pinus shows
striking similarities with Cordaitales.
These include:
i. The plants are tall-branched trees.
ii. The leaves are sample with paralled veins.
iii. The presence of sclerenchymatous hypodermis in the leaves.
iv. The presence of pycnoxylic wood.
v. The pollen grains are winged.
vi. The ovules are bilaterally symmetrical.
Figure 1.70 shows the life cycle of Pinus:
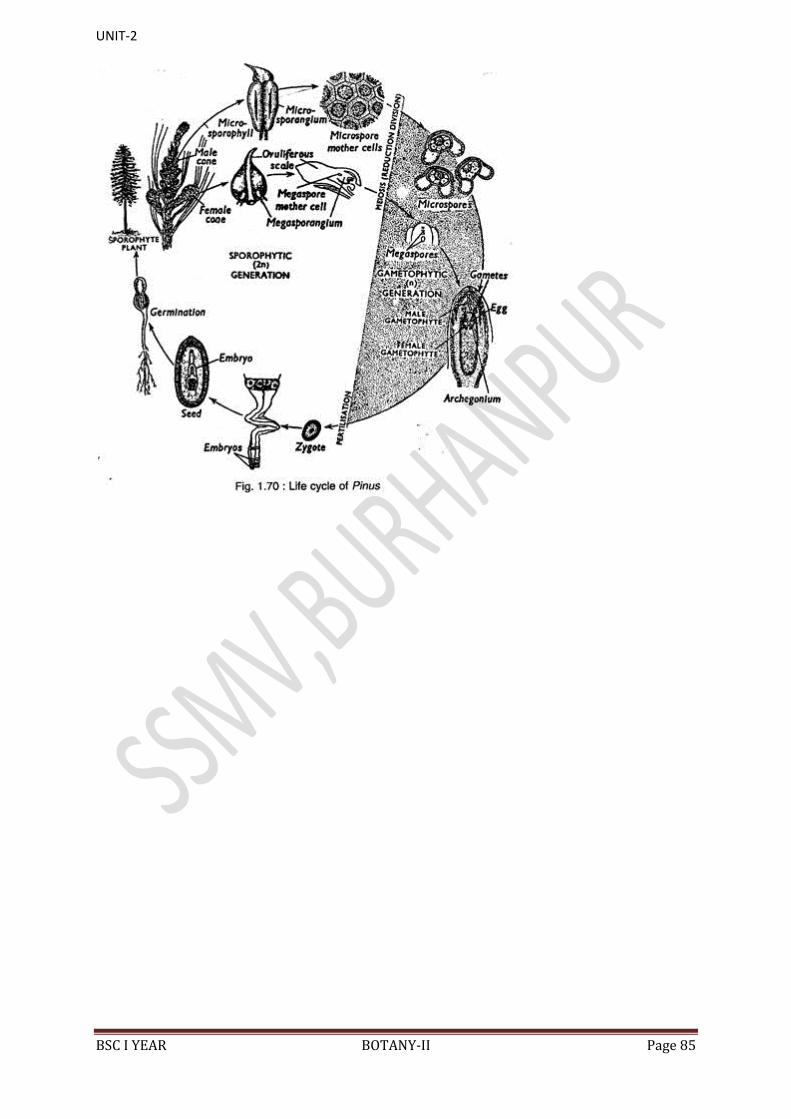
UNIT-2
BSC I YEAR BOTANY-II Page 85




















