
Origin of Afferent Projections into Bovine Chromaffin Cell Implants in the Rat Periaqueductal Gray...
18
(C) Freund Publishing House Ltd. 1994 Origin of Afferent Projections into Bovine Chromaffin Cell Implants in the Rat Periaqueductal Gray Determined by Retrograde and Anterograde Tracing John D. Ortega, Jacqueline Sagen and George D. Pappas Department of Anatomy and Cell Biology, University of Illinois at Chicago 808 S. Wood, Chicago, 1L 60612, USA SY We have previously described long-term survival of isolated bovine chromaffin cell suspension grafts in the periaqueductal gray of adult rats. Electron microscopic analysis of the graft sites revealed synapses on the transplanted chromaffin cells. The origin of these synapses is not known, but they are probably derived from the host since the initial grafts were suspensions of chromaffin cells that were essentially free of other cell types. In order to determine the origin of the observed synapses, retrograde and anterograde tracer analyses were performed on grafted rats at 4 and 8 weeks after transplantation. Following injection of the retrograde tracer (Tluoro-Gold) into graft sites, four major host sites were labeled: hindbrain reticular formation, substantia nigra, lateral hypothalamus, and cingulate cortex. Injection of anterograde tracer (rhodamine-conj ugated dextranamine) into the substantia nigra, lateral hypothalamus, and cingulate cortex produced labeled fibers and terminals in and around 4 and 8 week old chromaffin cell graft sites. An increase in both the number of retrogradely labeled cells, as well as in the density of Reprint address: G.D. Pappas Department of Anatomy and Cell Biology University of Illinois at Chicago 808 S. Wood, Chicago, IL. 60612, USA anterogradely labeled fibers and terminals within the graft site, was observed from 4 to 8 weeks. This study shows that graft innervation from the host is primarily from areas that normally project afferent fibers to the periaqueductal gray. The increase in labeled fibers and terminals over $ weeks suggests that de novo synapse formation on grafted bovine chromaffin cells is a continuous process that is dependent on the regenerative capacity and plasticity of the host neuronal network and the grafted bovine chromaffin calls. KEY WORDS adrenal medulla, xenograffs, synapse Fluoro-Gold, transplants, INTRODUCTION Long-term xenograff survival has been achieved in our laboratory using isolated bovine chromaftin cell suspensions accompanied by short-term immunosuppressive treatment of the host rat/9-11/. Electron microscopy of isolated bovine ehromaffin cell gratt sites in the rat pedaqueductal gray (PAG) shows that collagen encapsulation of the gratt and gliotic reaction by the host are significantly less than that observed following transplantation of whole adrenal medullary pieces /12,14/. This minimal boundary formation between host and graft may be VOLUME 5, NO. 1, 1994 31
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Origin of Afferent Projections into Bovine Chromaffin Cell Implants in the Rat Periaqueductal Gray...

(C) Freund Publishing House Ltd. 1994
Origin of Afferent Projections into Bovine Chromaffin CellImplants in the Rat Periaqueductal Gray Determined by
Retrograde and Anterograde Tracing
John D. Ortega, Jacqueline Sagen and George D. Pappas
Department ofAnatomy and Cell Biology, University ofIllinois at Chicago808 S. Wood, Chicago, 1L 60612, USA
SY
We have previously described long-termsurvival of isolated bovine chromaffin cellsuspension grafts in the periaqueductal gray ofadult rats. Electron microscopic analysis of thegraft sites revealed synapses on the transplantedchromaffin cells. The origin of these synapses isnot known, but they are probably derived fromthe host since the initial grafts were suspensionsof chromaffin cells that were essentially free ofother cell types.
In order to determine the origin of theobserved synapses, retrograde and anterogradetracer analyses were performed on grafted ratsat 4 and 8 weeks after transplantation.Following injection of the retrograde tracer(Tluoro-Gold) into graft sites, four major hostsites were labeled: hindbrain reticularformation, substantia nigra, lateralhypothalamus, and cingulate cortex. Injection ofanterograde tracer (rhodamine-conjugateddextranamine) into the substantia nigra, lateralhypothalamus, and cingulate cortex producedlabeled fibers and terminals in and around 4and 8 week old chromaffin cell graft sites. Anincrease in both the number of retrogradelylabeled cells, as well as in the density of
Reprint address:G.D. PappasDepartment of Anatomy and Cell BiologyUniversity of Illinois at Chicago808 S. Wood, Chicago, IL. 60612, USA
anterogradely labeled fibers and terminalswithin the graft site, was observed from 4 to 8weeks.
This study shows that graft innervation fromthe host is primarily from areas that normallyproject afferent fibers to the periaqueductalgray. The increase in labeled fibers andterminals over $ weeks suggests that de novosynapse formation on grafted bovine chromaffincells is a continuous process that is dependent onthe regenerative capacity and plasticity of thehost neuronal network and the grafted bovinechromaffin calls.
KEY WORDS
adrenal medulla,xenograffs, synapse
Fluoro-Gold, transplants,
INTRODUCTION
Long-term xenograff survival has been achievedin our laboratory using isolated bovine chromaftincell suspensions accompanied by short-termimmunosuppressive treatment of the host rat/9-11/.Electron microscopy of isolated bovine ehromaffincell gratt sites in the rat pedaqueductal gray (PAG)shows that collagen encapsulation of the gratt andgliotic reaction by the host are significantly less thanthat observed following transplantation of wholeadrenal medullary pieces /12,14/. This minimalboundary formation between host and graft may be
VOLUME 5, NO. 1, 1994 31

32 J.D. ORTEGA ET AL.
due to the homogeneity of the grained bovineehromaffin cell suspension. Cellular contaminants,such as fibroblasts and endothelial cells, that arefound in whole adrenal medullary tissue, arethought to enhance collagen encapsulation of thegraft and reactive gliosis by the host. The lack of aglial-collagen boundary may enhance host-graftintegration, as neuritic processes of host origintypically infiltrate or surround grafted chromaffincells/14/. Furthermore, synapses can be observedon grained bovine chromaftin cells as early as 3weeks post-transplantation/14/. These synapses arepresumably of host origin since no neurons areintroduced with the initial transplant of isolatedbovine ehroman cell suspension.
However, the host origin and functionalrelevance of the observed neuritic processes andsynaptic structures are not known. The purpose ofthis study was to localize potential sites of origin ofhost synaptic inputs using retrograde andanterograde tracing techniques.
minutes in a refrigerated centrifuge (IEC B20, 4C).The band containing healthy chromatn cells, asdefined by morphological analysis, was removedfrom the gradient, washed several times withLocke’s solution, and plated on 100 mm tissueculture dishes in media (1:1 DMEM:F12)supplemented with 5% fetal bovine serum (FBS),antibiotics (penicillin/streptomycin, 100 units/ml;gentamicin, 50 tg/ml) and anti-fungal agents(fungizone, 0.125 tg/ml). The plates were placed inan incubator overnight (37C, 5% CO) to allow fordifferential adherence of unwanted cell types (e.g.fibroblasts and endothelial cells). The next day,chromatn cells were removed from their plates bygentle agitation and once again placed on a Percollgradient to further enhance the purity and viabilityof the final chromaffin cell population. Trypan blueexclusion placed the viability of all chromaffin cellpreparations in excess of95%.
Retrograde analysis
MATERIALS AND METHODS
Bovine chromatn cells were isolated using apreviously described method /10/. Glands werepurchased from a local slaughterhouse, trimmed oftheir fat, and immediately perfused with Ca++- andMg++-free Locke’s solution (1.5 M NaCI, 0.05 MKCI, 0.04 M NaHCO3, 0.06 M glucose, 0.05 MHEPES at pH 7.2, 37C) containing antibiotics (100units/ml penicillin/streptomycin, 25 lag/mlkanamycin) and anti-fungal agents (0.125 lag/mlfungizone). Following a 30 minute incubation inLocke’s solution, the glands were perfused with a0.1% collagenase solution (Boehdnger-Mannheim;0.05% BSA, Sigma; and 0.01% trypsin inhibitor,Sigma) for 30 minutes. After this perfusion, themedullary tissue was dissected free from thesurrounding cortex, minced, filtered through a finenylon mesh, and washed several times with Locke’ssolution before purifying the cell population on aPercoll gradient. In brief, the washed cells wereplaced on a self-generating, buoyant densitygradient (45 ml Percoll, 5 ml of 10 X Locke’ssolution at pH 7.4) and spun at 12,000 RPM for 20
For the accurate injection of retrograde tracerinto the transplants, male Sprague-Dawley ratsweighing 300-350 g were anesthetized (30 mg/kgnembutal), and stereotaxically implanted withstainless steel guide cannulae (22 gauge) placed 3mm above the future transplant site. The guidecannulae were then secured to anchoring screwswith dental cement. After the dental cement haddried, bovine chromatn cells were injected into theventrolateral PAG (A +1.2 mm, L-0.7 mm, H +3.5mm from the intra-aural line, -2.5 mm incisor bar)with a 25 gauge needle using the mounted cannulaas a guide. After the injection of 2 gl of cells(approximately 50,000 cells/lal in sterile Hanks’buffer), the needle was slowly removed, a stainlesssteel wire was fitted into the guide cannula toprevent blockage, and the scalp was closed withwound clips. All animals with grafts received dailycyclosporine injections since we have previouslyshown that immunosuppression enhances thesurvival of transplanted bovine chromatn cells/10/. At 4 and 8 weeks after transplantation (n=8 ateach time point), animals were anesthetized andfitted into the stereotaxic holder. The guide wirewas removed, and the 25 gauge needle used for
JOURNAL OF NEURAL TRANSPLANTATION & PLASTICITY

HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS 33
chromaffm cell injections was filled with theretrograde tracer and lowered into the gratt siteusing the mounted guide cannula. Fluoro-Gold(Fluoroehrome Inc., 0.25 tl of a 2% solution in0.9% saline) was pressure injected over a 2 minuteperiod into the transplant site. After a 3-5 minutewaiting period, the injection needle was slowlywithdrawn and the animal returned to its cage.Several animals, serving as non-transplantedcontrols (n=8), received Fluoro-Gold injections intothe same region of the ventrolateral PAG viaimplanted guide cannulae.
Five days after injection of the retrogradetracers all animals were deeply anesthetized (50mg/kg nembutal) and perfused sequentially throughthe aorta with a 0.9% saline solution, a 4%paraformaldehyde solution, followed by 10%sucrose in a phosphate buffer. For cryoprotection,the brains were removed and placed in 20% sucrosebuffer for 24 hours. Brains were mounted oneryostat chucks and alternate 20 tm frozen sectionsextending from caudal medulla (P -3.5 mm from theintra-aural line) to frontal cortex (A +11.0 mm fromthe intra-aural line) were sectioned with a cryostat(l-Iaeker-Bdght), mounted on gelatin coated slides,and cover-slipped with Fluoromount-G (Fisher).The sections were viewed in an epifluorescencemicroscope (Zeiss Axiophot, 323 nm for Fluoro-Gold) and positively labeled cells were plotted usingan X-Y recorder. Accuracy of the retrograde labelinjection was established by immunocytochemicallystaining alternate sections for dopamine-13-hydroxylase (DBH, Eugene Teeh, diluted to a finalworking concentration of 1:500) or tyrosinehydroxylase (TH, Inestar, diluted to a final workingconcentration of 1:500) immunoreactivity. To dothis, tissue sections were incubated (24 h) inpreimmune sera [phosphate buffered saline (PBS)containing 2% normal goat serum (Cappel) and0.3% Triton X-100 (Sigma)] and then incubated for24 h in primary antibody at 4C. Sections were thenwashed in PBS, and incubated in either afluorescein- (goat anti-rabbit, Cappel, diluted to afinal working concentration of 1:100) orrhodamine- (goat anti-mouse, Cappel, diluted to afinal working concentration of 1"50) conjugatedsecondary antibody for one hour at room
temperature. Stained sections were then washed inPBS, mounted on gelatin coated slides, cover-slipped with Fluoromount-G, and viewed on a ZeissAxiophot microscope using fluorescein (495 nm) orrhodamine (520 nm) filters.
Anterograde tracer
Using the above retrograde tracing studies,possible sites of input into the transplanted cellswere identified. To confirm that these regionsprovide input to the grafts, anterograde traceranalysis was performed using rhodamine-labeleddextranamine (MW 10,000, Molecular Probes, Inc.)/15/. The anterograde tracer was stereotaxicallyinjected in 0.1-0.3 tl volumes into brain areaspreviously identified in the retrograde tracingstudies in animals with and without bovinechromatn cell transplants. These included themedullary reticular formation (P -2.0 mm, L -1.0mm, H +1.0 mm), substantia nigra (A +2.3 mm, L-2.0 mm, H +2.3 mm), lateral hypothalamus (A+6.2 mm, L -2.0 mm, H +1.5 mm), and cingulatecortex (A +10.2 mm, L-0.6 mm, H +7.4 mm). (Allcoordinates were measured from the intra-aural lineand incisor bar at -2.5 mm). In addition, to accountfor non-specific labeling of the grag and nearbyfibers, control injections were made in cerebellarcortical areas that had not contained labeled cellsfollowing previous retrograde tracing studies (P-1.0 nun, L +2.0 mm, H +7.5 mm). Atter 5 days,the animals were perfused as described above. Alltissues were also processed for either dopamine-13-hydroxylase or tyrosine hydroxylase immuno-reactivity using a fluorescein-conjugated secondaryantibody as previously described. In doing so,rhodamine-dextranamine labeled processes werevisualized within positively labeled graft sites.
RESULTS
Retrograde analysis
Figures 1 and 2 are composites made from theanalysis of tissue sections of brain sites containingFluoro-Gold labeled cell bodies from 8 control
VOLUME 5, NO. 1, 1994

34 J.D. ORTEGA ET AL.
GP
Fig, 1 and 2: Composite ofFluoro-Gold labeled cells in the CNS of 8 control animals receiving injections of the retrogradetracer into the ventrolateral PAG.
Fig. 1: Labeled cells were found in numerous sites rostral to the injection, including substantia nigra (SN), zona incerta(ZI), medial habenular nucleus (MHb), lateral hypothalamus (LHy), nuclei of the stria terminalis (ST), medialpreoptic nucleus (MPO), diagonal band ofBroca (DB), and cingulate cortex (Cg).
JOURNAL OF NEURAL TRANSPLANTATION & PLASTICITY

HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS 35
-1.o
-2.o
Core
Fi 2:
i:
Core
Labeled cells were also observed in several sites caudal to or near the level of the injection site, particularlymedullary and pontine reticular formation nuclei [AS, dorsal and lateral paragigantocellular (DPGI, LPGI),gigantocellular (GI), reticular (Rt), pontine reticular (PnC), SUltrier vstibular (SV) and oral pontine (PnO)nuclei]. In addition, local neurons in the pefiaqueductal gray (PAG) surrounding the injection site were heavilylabeled. The majority of labeled cells were found ipsilateral to the injection site, except for regions of the reticularformation and deep mesencephalic (DpMe) areas immediately near the injection site, which were labeled on bothsides. Core cerebellum, Py pyramids, GS central gray, M median eminence, ox optic chiasm, CPucaudate-putamen, GP globus pallidus. Numbers indicate distance (mm) from the intra-aural line.
VOLUME 5, NO. 1, 1994

36 J.D. ORTEGA ET AL.
animals. Histological evaluation of these tissuesections revealed that the majority of controlinjections were restricted to the PAG and areasimmediately surrounding it. These composites wereconstructed from the raw data acquired fromplotting Fluoro-Gold positive cells in tissue sectionswith an X-Y recorder. In brief, individual tissuesections from animals within a given group werematched based on neuroanatomical land-marks. Ofthe matched tissue sections, those containingmaximal labeling of cell bodies were chosen for thecomposite. In doing so, all areas ofthe rat CNS thatcontained labeled cells appear on the compositefigures. Discrete groups of labeled cells wereobserved in several regions rostral to the injectionsite (Fig. 1). The majority of these labeled cellbodies were found in the mesencephalic substantianigra (SN), zona incerta (ZI), and lateral andpreoptie nuclei of the hypothalamus (LHy and PO,respectively). Labeled cell bodies in these areaswere usually on the side ipsilateral to the injection,but occasionally labeled cells were observed incomralateral sites within hypothalamic nuclei andthe central gray. In addition, labeled cells wereobserved in the ipsilateral medial habenula andwithin the bed nuclei of the stria terminalis. Distalsites as far rostral as the diagonal band andeingulate cortex contained numerous Fluoro-Goldlabeled cell bodies. Labeled cells were also found atsites caudal to the injection (Fig. 2), but these werenot as discretely located as in the rostral sites.Caudally, the majority of cells were scatteredbilaterally throughout various nuclei of the reticularformation of the pens and medulla. These includeparagigantocellular, gigantocellular, pontinereticular, and A5 nuclei as well as superiorvestibular and oral pontine nuclei. In addition,labeled cells were found locally in the ventrolateralPAG and deep mesencephalic nuclei (Fig. 2), butthe proximity of this area to the Fluoro-Goldinjection site may have led to uptake of label byneighboring host cell bodies or dendrites.
Fluoro-Gold labeled cells containing a brightblue-white fluorescence were readily apparem. Onoccasion, labeled regions contained widelydispersed labeled cells in small clusters, such asthose found near the vestibular nucleus of the
hindbrain (Fig. 3A), but labeled cells were morecommonly found in larger and more densely packedgroups in specific CNS sites. These larger groups ofpacked cells were found in both the lateralhypothalarnus (Fig. 3B) and the substantia nigra. Inaddition, isolated Fluoro-Gold labeled cells wereobserved in the rat CNS with the majority scatteredthroughout various reticular formation nuclei of thepens and medulla.
Analysis of brain tissue from animals with 4week-old transplants revealed a distribution ofretrogradely labeled cell bodies similar to thatobserved following injection into non-transplantedcontrol animals described above (Figs. 4 and 5).The composite figures reveal that labeled cells weregenerally observed in the same rostral and caudalsites although the total number of specificallylabeled cells in these sites appeared to be fewer.Individual labeled cells were scattered throughoutthe same pontine and medullary reticular formationnuclei labeled in non-transplanted control animals.Discretely labeled groups of cells were morecommon in rostral brain areas; the majority of theselabeled cells were found in the substantia nigra,preoptic and lateral nuclei of the hypothalamus,zona incerta, diagonal band of Broca, and cingulatecortex. Labeled cells were notably absent fromseveral areas previously identified in controlinjections, particularly the medial habenula and bednuclei of the stria terminalis. In addition, severalnew regions, including the medial cortex, thalamus,inferior and superior colliculus, which were notlabeled in animals without transplants, appeared tocontain large numbers of labeled cells in sometransplanted animals. These new labeled regionswere not consistently observed in all animals. Therealso appeared to be larger numbers of labeled cellsin the PAG and surrounding midbrain near the levelof the transplant (approximate transplant areaoutlined in Fig. 5). Labeling within these newregions may be due to the uptake of tracer, whichdiffused along the cannula track, by terminals thatmay not specifically project to the gratt site.
When the distribution of retrogradely labeledcells in animals with 8 week-old transplants wasanalyzed, labeled cells were observed in essemiallythe same areas described at the 4 week time period,
JOURNAL OF NEURAL TRANSPLANTATION & PLASTICITY

HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS
lig. 3: Light micrograph of Fluoro-Gold labeled cells in the CNS of a control animal. Cells were commonly found insmall and large clusters throughout the CNS. A shows a smaller group of" labeled neurons ventral to the vestibularnucleus (Nu). In contrast, large groups of labeled cells were frequently observed, for example, in the lateralhypothalamus, as shown in B.
VOLUME 5. NO. 1.1994

38 J.D. ORTEGA ET AL.
JOURNAL OF NEURAL TRANSPLANTATION & PLASTICITY

HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS 39
.1.0
Cere
Fig. 4 and 5: Composite of Fluoro-Gold labeled cells in theCNS of rats with bovine chromaff]n cell graftsfollowing injection of retrograde tracer into thegraft site in the periaqueductal gray (PAG) 4weeks after transplantation. The location of atypical transplant site is" indicated by dashed linesin the top drawing of Fig. 5. Similar brain sites asin control animals contained Fluoro-Gold labeledcells, including sites rostral to the inje.on in Fig.4: substantia nigra (SN), zona incorta (ZI),diagonal band of Broca (DB), cingulate cortex(Cg), and hypothalamic nuclei (MPO, LHy), andcaudal to the injection site (Fig. 5), distributedthroughout the medullary and pontine reticularformation [dorsal and lateral paragigantocellular(DPGI, LPGI), gigantoceHular (GI), reticular (Rt),caudal pontine reticular (PnC), oral pontine(PnO), and parvocellular reticular (PRCt) nuclei].However, the number ofFluoro-Gold positive cellsat most of these sites appeared to be decreasedcompared to control injections. As in controlinjections, dense labeling was found in cells withinor surrounding the injection site. Many labeledcells were also observed in additional sites notlabeled in controls including superior and inferiorcolliculi (SC, IC), thalamic .nuclei Oh), andmedial cortex (Cx). This may be due to the uptakeof tracer from areas along the cannula track. Cere
cerebellum, Py pyramid, CG central gray,DpMe deep mesencephalic nuclei, M medianeminence, ox optic chiasm, CPucaudate/putamen Numbers indicate distance (mm)from the intra-aural line.
Cere
VOLUME 5, NO. 1, 1994

40 J.D. ORTEGA ET AL.
Fig. 6 and 7: Composite of Fluoro-Gold labeled cells in the CNS of rats with bovine chromaflin cell grafts followinginjection of retrograde tracer into the pefiaqueductal (PAG) graft site 8 weeks after transplantation. The location of a typicaltransplant site is indicated by dashed lines in the top left drawing of Fig. 7. Fluoro-Gold positive cells were observed in thesame brain sites found at the 4 week time point.
Fig. 6: An apparent increase in the number of labeled cells from 4 weeks to 8 weeks was observed in pars compaeta andreticularis of the substantia nigra (SNC, SNR), cingulate cortex (Cg), central gray (CG), medial habenular nucleus(MHb), and in various hypothalamic nuclei (I-Iy, LHy, MPO). Labeled cells were also observed in zona incerta (ZI),and diagonal band ofBroea (DB). As in the 4 week composite, labeled cells were observed in several cortical (Cx)and thalamie (Tit) sites as well as reticular thalamic (RTh) and medial genicular (MGn) sites.

HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS 41
DGi :oGi A5
-5.0
Numerous labeled cells were also found scattered throughout the medullary and pontine reticular formation [A5,dorsal and lateral paragigantocellular (DPGI, LPGI), lateral reticular (LRt), medullary reticular (MdV), caudal andventral pontine reticular (PnC, PnV), reticular (Rt), parvocellular reticular (PRCt), oral portion of pontine (PnO),nuclei], as well as in local regions of the PAG near the transplant site [dorsal tegmental (DTn), deepmesencephalic (DpMe), raphe magnus (RMg) as well as some labeled inferior collicular cells (IC)].Cerefcerebellum, Py=pymmid, DRffidorsal raphe nuclei, M=median eminence, Cpo=caudateYputamen. Numbersindicate distance (ram) from the intra-aural line.

42 HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS
but the total number ofFluoro-Gold positive cells inthese areas appeared to be much greater at this latertime (Figs. 6 and 7). In particular, the density oflabeled cells was markedly increased in the cingulategyms and medial cortex, hypothalamic and thalamicregions, substantia nigra, and the central gray.While the highest density of labeled cells in theseregions was ipsilateral, there was also a dramaticincrease in labeled cells contralaterally as well. Incontrast to the earlier transplants, retrogradeinjection in the 8 week old transplants resulted ingreater numbers and wider areas of labeled cellsthan that found following injections in non-transplanted control animals. Labeled neurons wereagain found in and surrounding the injection site, aswell as scattered throughout the same nuclei of thepontine and medullary reticular formation observedin both non-transplanted control and 4 weektransplanted animals. As in the 4 week group, directlabeling of some cell bodies may have occurred dueto the uptake ofFluoro-Gold by terminals along thecannula track. Evaluation of the injection sitesrevealed that the intensity of the Fluoro-Gold labelwas greatest in the transplant itself, but in mostcases some diffusion of the tracer outside of theimmediate graft site had occurred.
Anterograde analysis
Difsion and uptake of the retrograde traceralong the cannula track, from surrounding regionsor by fibers of passage, can often lead to thelabeling of brain sites that do not send afferentprojections to the PAG. In order to positivelyidentify regions of the CNS that send specificafferent projections into the grafted chromaftin cellsanterograde studies were performed usingrhodamine conjugated dextranamine, a sensitivemarker for anterograde axonal transport/15/. Thelabel was injected into brain sites that haddemonstrated consistent retrograde labeling withFluoro-Gold. Four and 8 week-old transplantedanimals as well as non-grafted animals receivedpressure injections of the anteriograde tracer intoeither caudal reticular formation, substantia nigra,lateral hypothalamus, or cingulate cortex (n=32,4/brain region). Use of TH or DBH immuno-
cytochemistry in combination with the tracer andappropriate fluorescein or rhodamine filters allowedvisualization of both DBH-positive chromaffin cellgrafts and rhodamine labeled processes underepifluorescenee illumination.
Analysis of tissue sections from brainscontaining 4 and 8 week-old bovine chromatfin celltransplants readily revealed TH-positive and DBH-positive grafts located in the ventrolateral PAG. At4 weeks post-transplantation, injection of theanterograde tracer into the reticular formation,substantia nigra, lateral hypothalamus, or cingulatecortex produced labeling throughout the midbrain.The majority of label was located in the PAG tissuearound the graft site. On occasion, rhodaminedextranamine labeled fibers appeared to enterclumps of DBH-positive chromaffin cells, such asthat observed following injection of the tracer intothe substantia nigra (Fig. 8). At the 4 week periodmost labeling observed within the graft siteoccurred following injection ofthe tracer into eitherthe substantia nigra or lateral hypothalamus.Injections into the reticular formation and cingulatecortex did not produce visible labeling within thegraft although labeled fibers could be found outsidethe graft area and at the host-graft interface.
At 8 weeks post-transplantation, injection intoeither the substantia nigra, lateral hypothalamus, orcingulate cortex produced punctate labelingthroughout the graft site with the greatest amountproduced by injections into lateral hypothalarnus.Figure 9A shows the appearance of a typical 8week-old bovine chromaftin cell graft in the PAGstained for DBH. The same site is shown in Figure9B revealing rhodamine-conjugated dextranarninelabeled terminals infiltrating the graft site andsurrounding host tissue following injection of theanterograde tracer into the lateral hypothalarnus. Onoccasion, long processes, presumably of host origin,appeared to infiltrate the graft site extensively (Fig.9B, arrowheads). Labeling of this type was not asfrequent as punctate labeling. Injection into all foursites produced labeling of fibers throughout the hostparenchyma immediately surrounding the graft site.As in 4 week animals, labeled fibers were frequentlyobserved near the host-graft interface. Injection intothe reticular formation did not produce labeling

J.D. ORTEGA ET AE. -
VOLUME 5, NO. 1, 1994

J.D. ORTEGA ET AL.
9; Light micrograph of an 8 week.-old graft site stained for dopamine-13-hydroxylase (DBH) immunoreactivity. Largeclumps of DBH-positive cells were found in clumps within the ventrolateral periaqueductal gray (A). At 8 weeks,fibers and terminals containing rhodamine-conjugated dextranamine were frequently observed throughout the graftsite and surrounding host tissue following injection of anterograde tracer into either substantia nigra, lateralhypothalamus, or cingulate cortex. This labeling was usually punctate in appearance, as can be observedthroughout the graft site following injection of the anterograde tracer into the cingulate cortex (B), but on occasionlong processes (arrowheads) were observed entering the graft site and diverging throughout the tightly clumpedchromaffin cell graft.
JOURNAL OF NEURAL TRANSPLANTAq ION & PLASTICITY

J.D. ORTEGA ET AL. 45
within the graft site although labeled fibers wereseen in the surrounding host parenchyma.Inspection of the injection sites revealed that theinjections had indeed been made into the siteslabeled by the retrograde tracer. In addition, controlinjections were made into cerebellar areas that donot project into or through the ventrolateral PAG.The control injections did not label the PAG grattor areas immediately around it. These resultssuggest that labeling in and around PAG gratis,following injection into the substantia nigra, lateralhypothalamus, or eingulate cortex, was due to theanterograde movement of tracer and not due to thenon-specific labeling ofthe gratt and nearby fibers.
For comparison ofthe innervation distribution inthe normal PAG, analysis of control tissues revealedthat injection of anterograde tracer into all of theabove sites produced labeling offibers and terminalsthroughout the ventrolateral PAG (Fig. 10). Denserlabeling was evident in the PAG in tissue sectionsfrom animals receiving injections of the tracer intoeither lateral hypothalamus or substantia nigra. Thislabeling was either in long individual fibers (Fig.10A) or punetate (Fig. 10B), similar to that found intransplant regions. In contrast, sections fromeingulate cortex or reticular formation injectedanimals displayed fewer labeled fibers and terminals.
DISCUSSION
The level of host-graIt integration can beestimated morphologically by the extent ofreciprocal fiber outgrowth and synapse formation.Previous studies have demonstrated that neuralgratis offetal CNS tissue are capable of establishingefferent as well as afferent connections with thehost CNS /2,3,7/. Adrenal medullary grats havealso been reported to exhibit this phenomenon. Inparticular, process outgrowth from intraoculargrafts of adrenal medullary tissue into CNS tissuehas been described/8,18/. Unsieker et al./18/havealso demonstrated by ultrastructural analysis thatautografls of adult guinea-pig adrenal chromancells exhibited synapse-like contacts on one another.Earlier work by different groups gave ultrastructuralevidence for the formation of synapses on adrenal
chromatfin cells grafted into the intraocular space ofrodents/5,17,19/. These synapses were presumablyof host origin from local cholinergic fibers of theciliary ganglion /18/. Previous studies in ourlaboratory have demonstrated that intraparenchymalgrafts of chromatn cells may serve as both post-synaptic and, occasionally, pre-synaptic componentsof synapses/12,13/. More recent studies on bovinechromatn cells implanted into the PAG reveal thatsynapse formation on these gratted xenogeneic cellsoccurs as soon as 3 weeks post-transplantation/14/.These synapses appear more ’mature’ in older grailsites (e.g. 6 or 12 weeks post-transplantation) sincepresynaptic structures are larger, cover a larger areaof the chromatn cell membrane, and contain agreater number ofvesicles than synapses in youngergratis/14/.
The purpose of the present study was tocharacterize further the host-graft integration ofintraparenehymally transplanted bovine chromatfincells by identifying host projections to the graftedcells. Results of the present study suggest that thehost origin of these projections is most likely fromeither local or distal fibers that normally innervatethe PAG. These fibers may arise from neuronal cellbodies as near as the substantia nigra or as far awayas the cingulate cortex, both of which have beenshown to project fibers into the PAG /1,6/. Thedetailed organization of afferent input into the ratPAG has been described by Beitz /1/. The PAGreceives afferent fibers from many medullary,pontine, meseneephalic, diencephalic, and variousmedial and lateral cortical regions /16/. Fibersoriginating from the basal forebrain also constitute alarge proportion of the afferent projections to thePAG. Forebrain sites include septal nuclei and thediagonal band of Broca. The most prominent sitesof afferent input are from various hypothalamicnuclei. In addition, the substantia nigra sends thelargest number of mesencephalic afferent projec-tions to the PAG, while medullary and pontinereticular formation are the greatest sites of afferentinput from the lower brainstem. The PAG itselfappears to have many intrinsic connections andreceives afferents from local sites such as the ventraltegmentum and midline raphe nuclei. Afferents fromsites caudal to the medulla, such as the spinal cord,
VOLUME 5, NO. 1, 1994
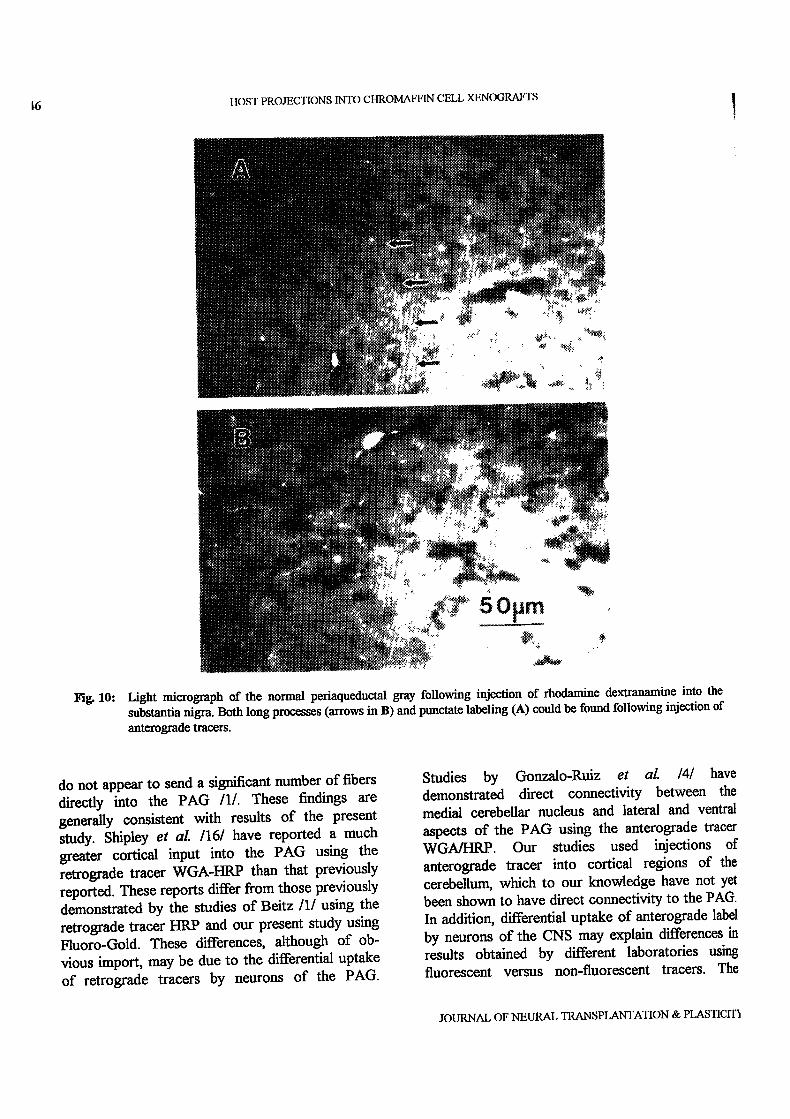
6
Fi. 10:
It()ST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS
Light micrograph of the normal periaqueductal gray following injection of rhodamine dextranamine into the
substantia nigra. Both long processes (arrows in B) and punctate labeling (A) could be found following injection of
anterograde tracers.
do not appear to send a significant number of fibers
directly into the PAG /1/. These findings are
generly consistent with results of the presentstudy. Shipley et al. 1161 have reported a much
greater cortical input into the PAG using the
retrograde tracer WGA-HRP than that previouslyreported. These reports differ from those previouslydemonstrated by the studies of Beitz/1! using the
retrograde tracer HRP and our present study using
Fluoro-Gold. These differences, although of ob-
vious import, may be due to the differential uptakeof retrograde tracers by neurons of the PAG.
Studies by Gonzalo-Ruiz et al. /4/ have
demonstrated direct connectivity between the
medial cerebellar nucleus and lateral and ventral
aspects of the PAG using the anterograde tracer
WGA/HRP. Our studies used injections of
anterograde tracer into cortical regions of the
cerebellum, which to our knowledge have not yetbeen shown to have direct connectivity to the PAG.In addition, differential uptake of anterograde label
by neurons of the CNS may explain differences in
results obtained by different laboratories usingfluorescent versus non-fluorescent tracers. The
JOURNAL OF NEURAL TRANSPLANq’ATION & PLASTICIT5

J.D. ORTEGA ET AL. 47
results obtained from these control studies revealthat injections into cortical regions of thecerebellum do not produce any significant labelingof fibers within the mesencephalic PAG oftransplanted or non-transplanted animals.
Analysis of animals with bovine ehroman cellimplants revealed that many of the sites thatnormally innervate the PAG are retrogradely labeledfollowing tracer injection into the graft. The majorareas labeled were nuclei within the medullary,pontine, and mesencephalic reticular formation,substantia nigra, lateral hypothalamus, zona incerta,diagonal band ofBroca, septal nuclei, and cingulatecortex. In addition, new labeled sites, includingthalamic nuclei and medial cortex, were found insome transplanted animals. This was not aconsistent finding and may be due to the uptake oftracer that has leaked along the needle track orspread of the tracer outside of the graft region.Interestingly, the numbers of retrogradely labeledcells, even in regions that normally innervate thePAG, appear more numerous in animals withchromaffin cell transplants than in normal rats (e.g.compare Figs. 4 and 5 or Figs. 6 and 7 with Figs. 1and 2). This could indicate an enhanced innervationof the transplant or recruitment of additionalneuronal input from regions normally projecting tothe PAG. Alternatively, the apparent increasedinnervation could be a result of spread of the traceroutside of the actual graft tissue into thesurrounding PAG, or uptake by fibers damaged bythe transplantation procedure. While it appearedthat injection sites were approximately the same sizeand spread as those in transplanted animals, thesepossibilities cannot be eliminated using retrogradetracer injections. Thus, results must be interpretedwith caution, as retrograde labeling from transplantsites can only indicate possible sources of transplantinnervation, rather than definitively identify host-gratt connections or synaptic densities.
Anterograde tracing from possible innervationsources revealed in the retrograde tracing studieswas done in order to more specifically identifyregions sending processes into regions containingtransplanted ehromaffm cells. Of the previouslymentioned sites, midbrain reticular formation,substantia nigra, lateral hypothalamus, and cingulate
cortex were chosen for anterograde analysis, sincethese areas have been shown to be major sites oforigin of afferents to the PAG/1,6/. Analysis oftissue sections revealed that rhodamine-conjugateddextranamine traveled anterogradely into andaround the grat site after injection into thesubstantia nigra, lateral hypothalamus, or cingulatecortex. Injections into the reticular formation didnot produce much labeling within the graft althoughpunctate labeling and labeled fibers could be seennear the graft site. The majority of labeling in 4 and8 week gratis was primarily punctate in appearance,although occasionally long processes could be seenpenetrating the clumps of transplanted cells.Similarly, both long processes and punctate labelingare found in normal PAG sites following injectionsof anterograde tracers in these regions. Inter-estingly, the apparent number of anterogradelylabeled fibers in both the transplant site and thenormal PAG seem small compared to the numbersof cells identified by the previous retrograde tracingstudies. This suggests that some of the retrogradelylabeled cells may be due to uptake from surroundingregions or fibers ofpassage.
Our results suggest that intraparenchymallygratted bovine chromaffin cells integrate into thehost neuronal network since host afferentprojections project into areas that normally containperiaqueductal tissue. One possible explanation isthat the cell suspension disperses around andbetween local host fibers. A second and more likelyexplanation is that fibers are initially disrupted bythe suspension grafts, and that innervation of thegraft is due to sprouting or axonal regrowth. Insupport for this, there was an apparent increase inboth the numbers of retrogradely labeled cells andthe density of anterogradely labeled fibers andterminals from 4 weeks to 8 weeks post-transplantation. Ifthe presence oflabeling in a gratwas due to the chromatn cells surroundingpreviously existing fibers and axonal endings, onewould expect to find the same number ofretrogradely labeled cells at all time points. On theother hand, if labeling of the gratt is dependent onhost tissue plasticity and ability to form de novocircuitry, one would expect to see a gradualincrease in the amount of labeling within a gratover time.
VOLUME 5, NO. 1, 1994

48 HOST PROJECTIONS INTO CHROMAFFIN CELL XENOGRAFTS
The results of this study provide further evid-ence that chromaffin cell grafts from xenogeneicdonors can become integrated within the host ratCNS circuitry. However, while host CNS appearsto send afferent projections into graft sites, defini-tive identification of synaptic input to grafted cellsawaits ultrastmctural analysis.
ACKNOWLEDGEMENTS
Supported in part by NIH grants NS28931 andNS25054 and an APA Minority Fellowship (toJ.D.O.).
REFERENCES
1. Beitz AJ. The organization of afferent projections to themidbrain periaqueductal gray of the rot. Neuroscience1982; 7: 133-159.
2. Bolam JP, Freund TF, Bj6rklund A, Dunnett SB, SmithAD. Synaptic input and local output of dopaminergicneurons in grafts that functionally reirmervate the hostneostriatum. Exp Brain Res 1987; 68:131-146.
3. Castro AJ, Tonder N, Sunde N, Zimmer J. Fetalneocortical transplants grafted to the cerebral cortex ofnewborn rats receive afferents from basal forebrain,locus coenfleus, and midline raphe. Exp Brain Res1988; 69: 613-622.
4. Gonzalo-Ruiz A, Leiehnetz GR, Hardy SGP.Projections of the medial cerebellar nucleus tooculomotor-related midbrain areas in the rot: ananterograde and retrograde HRP study. J Comp Neurol1990; 296: 427-436.
5. Kondo H. Reirmervation of the rat adrenal medullatransplanted in the anterior eye chamber. J Anat 1978;127:323-331.
6. Marchand JE, Hagino N. Atferents to theperiaqueduetal gray in the rat. A horseradishperoxidase study. Neuroscience 1983; 9: 95-106.
7. Murata Y, Chiba T, Brundin P, Bj6rklund A, LindvallO. Formation of synaptic graft-host connections bynoradrenergie locus coeruleus neurons transplantedinto the adult rat hippocampus. Exp Neurol 1990; 110:258-267.Olson L, Seiger A, Freedman R, Hoffer B. Chromaffincells can innervate brain tissue: Evidence from
intraocular double grafts. Exp Neurol 1980; 70: 414-426.
9. Ortega JD, Sagen J, Pappas GD. Survival andintegration of chromaffin cell transplants in the CNS.In: Stein GD, ed, Restorative Neurology andNeuroscience, IIIrd International Symposium on NeuralTransplantation, Amsterdam: Elsevier, 1989; 70.
10. Ortega JD, Sagen J, Pappas GD. Short-termimmunosuppression enhances long-term survival ofbovine chromattin cell xenografts in rat CNS. CellTransplant 1990; 1: 33-41.
11. Ortega JD, Sagen J, Pappas GD. Survival andintegration ofbovine chromatfin cells transplanted intorat CNS without exogenous trophic factors. J CompNeurol 1992; 323: 13-24.
12. Pappas GD, Sagen J. The fine structure of chromaffincell implants in the pain modulatory regions of the ratpedaqueductal gray and spinal cord. Prog Brain Res1988; 78: 513-520.
13. Sagen J, Pappas GD, Perlow MJ. Fine structure ofadrenal medullary grafts in the pain modulatoryregions of the rat periaqueductal gray. Exp Brain Res1987; 67: 380-390.
14. Sagen J, Pappas GD, Ortega JD. Host-graftrelationships of isolated bovine chromaffin cells in ratpedaqueductal gray. J Neurocytol 1990; 19: 697-707.
15. Schrnued L. Kyriakidid K, Heimer L. In vivoanterograde and retrograde axonal transport of thefluorescent rhodamine-dextran-amine, Fluoro-Ruby,within the CNS. Brain Res 1990; 526: 127-134.
16. Shipley MT, Ennis M, Rizvi A, Behbehani MM.Topographical specificity of forebrain inputs to themidbrain periaqueduetal gray: Evidence for discretelongitudinally organized input columns. In: DepaulisA, Bander R, eds, The Midbrain Periaqueduetal GrayMatter: Functional, Anatomical, and NeurochemicalOrganization, New York: Plenum Press, 1990; 417-448.
17. Unsieker K, Tschechne B, Tseheclme D. Formation ofeholinergic synapses on adrenal ehromaffin cells inanterior eye chamber transplants. Brain Res 1978; 152:334-340.
18. Unsieker K, Tscheclme B, Tseheclme D. Differentia-tion and tmnsdifferentiation of adrenal chromattin cellsofthe guinea-pig. Cell Tissue Res 1981; 215: 341-367.
19. Unsieker K, Zwarg U, H_abura O. Electron microscopicevidence for the formation of synapses and synaptoidcontacts in adrenal medullary grafts. Brain Res 1977;120: 533-539.
JOURNAL OF NEURAL TRANSPLANTATION & PLASTICITY




















