
Obtention and characterization of primary astrocyte and microglial cultures from adult monkey brains

Ocular dominance columns in the adultNew World MonkeyCallithrix jacchus
CATHERINE CHAPPERT-PIQUEMAL,1 CAROLINE FONTA,1 FRANÇOIS MALECAZE,2
and MICHEL IMBERT1
1Centre de Recherche Cerveau et Cognition, UMR5549 Centre National de la Recherche Scientifique0Université Paul Sabatier,Faculté de Médecine de Rangueil, 31062 Toulouse Cedex, France
2Service Ophtalmologique Hôpital Purpan, place Dr Baylac, 31059 Toulouse Cedex, France
(Received November 21, 2000;Accepted January 25, 2001)
Abstract
In the marmosetCallithrix jacchus, ocular dominance columns (ODC) have been reported to be present in younganimals, but absent in adults (Spatz, 1989). We have studied in juvenile and adult animals the postnatal organizationof the retino-geniculo-cortical afferents by means of transneuronal labeling. We show in the present work that ODCare present in the primary visual cortex ofCallithrix jacchus, both in the adult and in the juvenile animal. Thepresent work confirms the presence of ODC in the visual cortex of juvenile marmoset before the end of the firstpostnatal month. In 2-month-old animals, ODC are well demarcated in IVca and IVcb. In the adult marmosets,the present data clearly show that the primary visual cortex is also organized with ODC. In horizontal sections,they form a mosaic through the ventral and dorsal calcarine cortex and through the dorso-lateral occipital part of thestriate cortex. In frontal sections, their presence is manifest in IVcb within the calcarine cortex and they only faintlyappear in IVca. These new findings are important since they underline the usefulness of the adult New WorldMonkeys as a model in visual research.
Keywords: Marmoset, Visual cortex, Postnatal development, Transneuronal labeling
Introduction
An important feature of the primary visual cortex is its columnarorganization. The thalamo-cortical afferents relative to each eyeare arranged in alternate bands in layer IVc of the striate cortex,called ocular dominance columns (ODC). This arrangement hasbeen shown in many mammalian species, and is extensivelydescribed in monkeys. Among primates, ODC have been reportedin several species of Old World Monkeys: macaque (Hubel &Wiesel, 1972; LeVay et al., 1975, 1985), baboon (Hendricksonet al., 1978), and chimpanzee (Tigges & Tigges, 1979). In NewWorld Monkeys, the results have been more variable. The anatom-ical organization of ocular segregation has been revealed in theadult spider monkey (Ateles ater) (Florence et al., 1986). In thesquirrel monkey, earlier reports suggested that the afferents serv-ing each eye are intermingled in layer IV (Hendrickson et al.,1978). This has been refuted by a recent report in which theauthors noted that the ODC exist in squirrel monkeys althoughthey are less well segregated than in macaques (Horton & Hock-ing, 1996a). In the capuchin monkey (Cebus apella), the anatom-ical demonstration of the ODC pattern has been described by
cytochrome oxidase histochemistry after monocular enucleation inthe adult animal (Hess & Edwards, 1987; Rosa et al., 1988, 1992).In the marmosetCallithrix jacchus, transneuronal labeling re-vealed no ODC in the adult animal (Spatz, 1979; DeBruyn &Casagrande, 1981). However, the segregation of inputs correspond-ing to each eye has been demonstrated in normal juvenile marmo-sets (2, 3, and 9 months old) (Spatz, 1989; Fonta et al., 2000). Ithas been suggested that the organization of ODC is lost during thefirst year of life (Spatz, 1989), and that it can be stabilized inadulthood by early monocular deprivation (Sengpiel et al., 1996).Despite the apparent loss of ODC in the adult animal, the presenceof “physiological ODC” has been proposed, by way of activity-dependent immediate early-gene expression (Markstahler et al.,1998).
Marmosets are an attractive model for studying the develop-ment of the primate visual system and specially the ODC (Fontaet al., 1997, 2000). The purpose of the present work is to reexam-ine the loss of ODC in adults. The present data clearly shows thatthe primary visual cortex of adult marmosets contain ODC.
Methods
These experiments were carried out on marmosets (Callithrixjacchus) reared in Centre de Recherche Cerveau et Cognition(Toulouse, France).
Address correspondence and reprint requests to: Caroline Fonta, Centrede Recherche Cerveau et Cognition, UMR5549 Centre National de laRecherche Scientifique0Université Paul Sabatier, Faculté de Médecine deRangueil, 31062 Toulouse Cedex, France.
Visual Neuroscience(2001),18, 407–412. Printed in the USA.Copyright © 2001 Cambridge University Press 0952-5238001 $12.50
407
Our procedures for eye injection and for autoradiographicrevelation have been described previously (Fonta et al., 2000).Briefly, the animals were anesthetized with intramuscular injec-tions of a mixture of xylazine (Rompun-BayerAG, Leverkusen,Germany) (5 mg0kg) and ketamine Imalgene 1000 (Rhone Merieux,Lyon, France) (25 mg0kg). Topical application of tetracaine chlor-hydrate was performed before eye injection. About 2 mCi ofL-(2,3,4,5-3H) proline (specific activity 92 Ci0mmol, Amersham,Arlington Heights, IL) diluted in 20ml NaCl 0.9% were injectedin the left eye. After a survival time of 7 to 15 days (Table 1), theanimals were transcardially perfused under deep pentobarbitalanesthesia (50 mg0kg) with 0.9% NaCl containing heparine. This
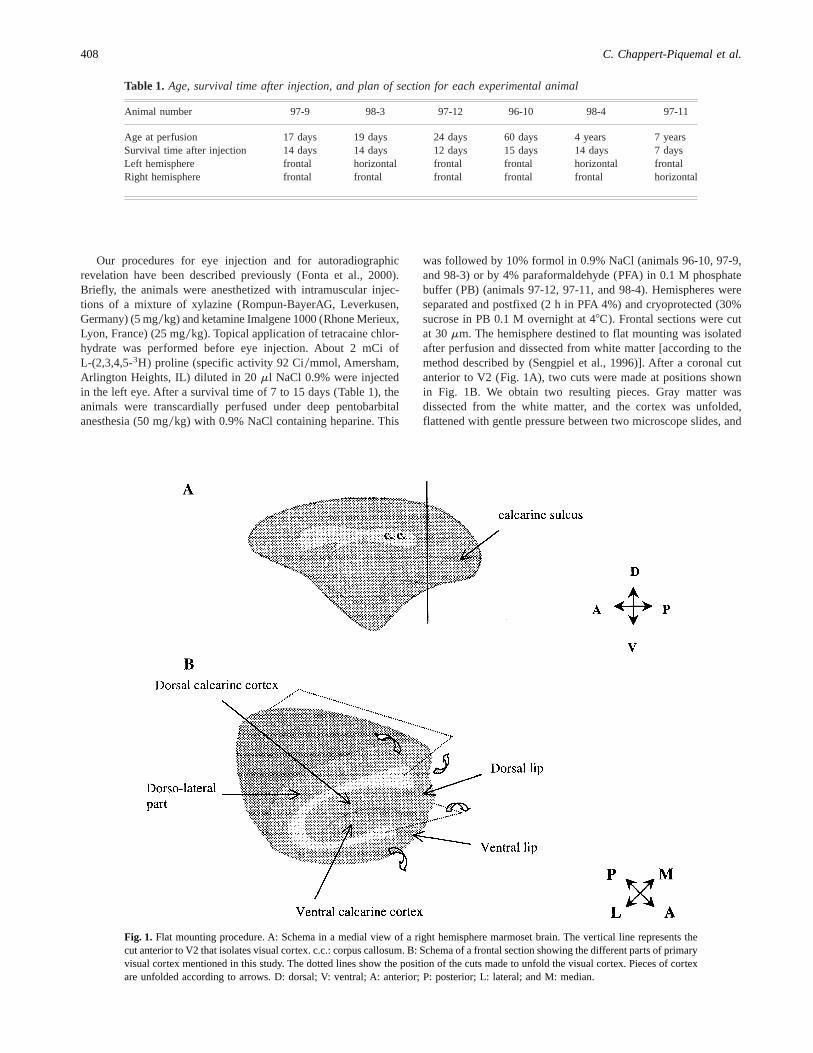
was followed by 10% formol in 0.9% NaCl (animals 96-10, 97-9,and 98-3) or by 4% paraformaldehyde (PFA) in 0.1 M phosphatebuffer (PB) (animals 97-12, 97-11, and 98-4). Hemispheres wereseparated and postfixed (2 h in PFA 4%) and cryoprotected (30%sucrose in PB 0.1 M overnight at 48C). Frontal sections were cutat 30mm. The hemisphere destined to flat mounting was isolatedafter perfusion and dissected from white matter [according to themethod described by (Sengpiel et al., 1996)]. After a coronal cutanterior to V2 (Fig. 1A), two cuts were made at positions shownin Fig. 1B. We obtain two resulting pieces. Gray matter wasdissected from the white matter, and the cortex was unfolded,flattened with gentle pressure between two microscope slides, and
Table 1. Age, survival time after injection, and plan of section for each experimental animal
Animal number 97-9 98-3 97-12 96-10 98-4 97-11
Age at perfusion 17 days 19 days 24 days 60 days 4 years 7 yearsSurvival time after injection 14 days 14 days 12 days 15 days 14 days 7 daysLeft hemisphere frontal horizontal frontal frontal horizontal frontalRight hemisphere frontal frontal frontal frontal frontal horizontal
Fig. 1. Flat mounting procedure. A: Schema in a medial view of a right hemisphere marmoset brain. The vertical line represents thecut anterior to V2 that isolates visual cortex. c.c.: corpus callosum. B: Schema of a frontal section showing the different parts of primaryvisual cortex mentioned in this study. The dotted lines show the position of the cuts made to unfold the visual cortex. Pieces of cortexare unfolded according to arrows. D: dorsal; V: ventral; A: anterior; P: posterior; L: lateral; and M: median.
408 C. Chappert-Piquemal et al.

cryoprotected overnight. The cortex was then frozen flat by plac-ing the pial surface on a stand and tangential sections were madeat 14mm. Frontal and horizontal sections were mounted and slideswere coated with Amersham EM1 emulsion, and stored at 48C.Sections were developed with D19 (Kodak) after several months(21
2_–12) exposition. The quality of eye injection was controlled by
verifying the labeling in both lateral geniculate nuclei (LGN).Sections were examined in light-field illumination, and digi-
tized for further analyses by using a Leica microscope, or Leicastereomicroscope, equipped with a 3CCD camera (Sony DXC930P),interfaced with an IBM-compatible computer, Imascan Board(Imagraph), and Optimas software (IMASYS). Montage of serialhorizontal sections was performed with Photoshop 5.0 software(Adobe systems) according to the method described by Hortonand Hocking (1996b). In horizontal sections, the measurementsof proline bandwidths or area occupied by labeled bands weremade in the final montage by using image analysis Optimas soft-ware (IMASYS). Sizes of labeled columns were measured atdifferent levels; because the sizes of bands are irregular only thegreatest and the narrowest widths of the labeled ODC were mea-sured (in animals 97-11n 5 25, in 98-4n 5 37). The areas occu-pied by labeled stripes were measured on images represented inFig. 2. A surface in which ODC are well demarcated was selectedand thus areas occupied by proline bands could be visualized;several measurements were made within a given surface and it isthe average that is given (measurements differing by 6–8%).
All experimental protocols conformed to the National Instituteof Health guidelines.
Results
We made a (3H) proline injection in six marmosets (Table 1). Thepresent results concern four animals in which the injection wassuccessful. We visualized ODC patterns in 24- and 60-day-oldmarmosets and in two adults. The LGN autoradiographies do notshow apparent interlaminar spillover of radioactivity labels inthese animals. We did not succeed in revealing the retino-thalamo-cortical pathway in the two newborn animals, in which LGN wereonly faintly labeled. Visual system immaturity rather than meth-odological factors seems to be the main cause in the failure of thetransneuronal labeling in the very young animals.
Ocular dominance columns (ODC) pattern
The autoradiographies of the primary visual cortex show alternat-ing regions of light and dark staining in layer IVc, both in youngand adult animals. In each animal, the hemisphere ipsilateral to theinjection displays a more distinct pattern of ODC than the contra-lateral hemisphere. The columnar pattern is strictly limited to theregion of V1 but is absent in the monocular segment (which islocated in the anterior region of the primary visual cortex, that is,in the dorsal bank of the calcarine sulcus), and in the regioncorresponding to the blind spot (which is situated in the caudal partof the calcarine cortex). These two regions have the same locationin young and adult animals.
Adult animals (97-11 and 98-4)Horizontal (Fig. 2) and frontal sections (Fig. 3A) clearly show
light and dark columns in layer IVc in both animals.In the horizontal sections, the ODC pattern appears in the
ventral and dorsal parts of the calcarine cortex where patches are
organized in long rows displaying a mosaic of irregular columns(Figs. 2A, 2B, and 2D). This pattern is also observed in thedorso-lateral part of V1 where irregular bands constitute a networkcovering a small region in the occipital cortex (Fig. 2C)
The widths of the dark stripes were measured within the ventralcalcarine cortex. The average width is 211mm (SD 6 51) and257mm (SD6 66) in animals 98-4 and 97-11, respectively. In themontage of horizontal sections and in the sections shown in Fig. 2,the labeled stripes occupy, of the whole measured area, an averageof 57% (in the ipsilateral hemisphere to the injected eye in animal98-4) and of 60% (in the contralateral hemisphere to the injectedeye in animal 97-11).
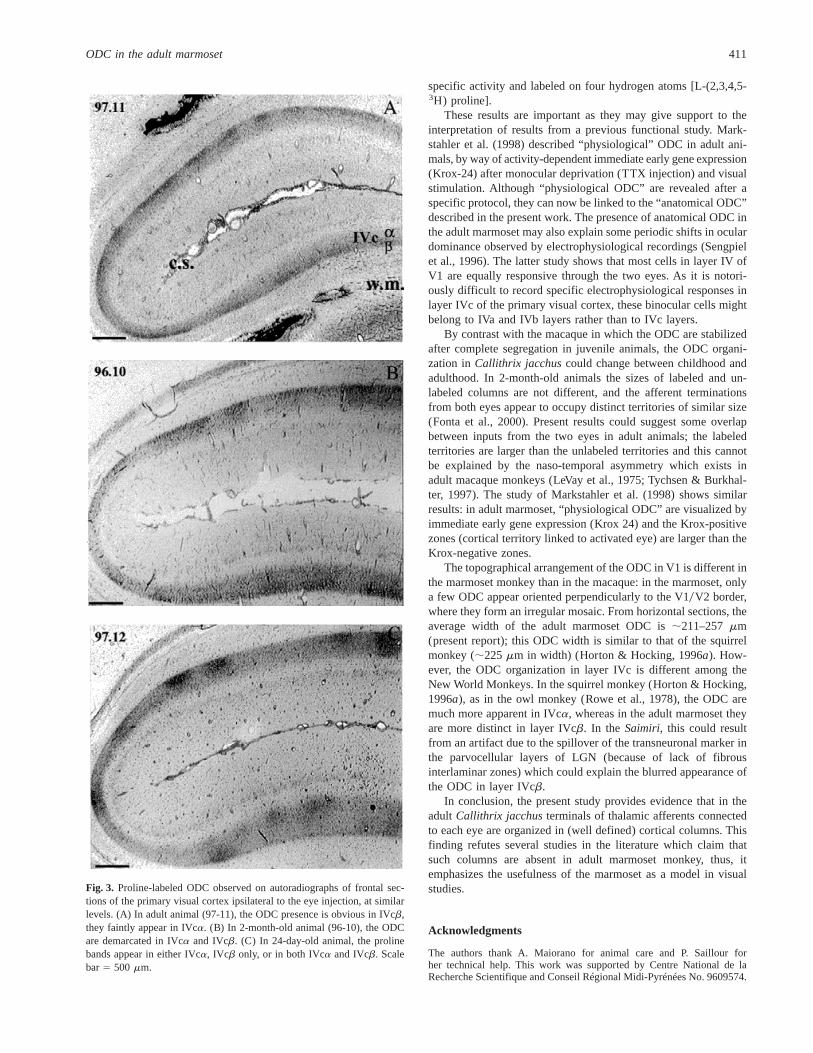
In the frontal sections, clear ocular dominance bands can beseen in the adult marmosets studied. In both animals, the ODC areobvious in IVcb, however, they are less distinct than in thejuveniles (Figs. 3A–3C). In all the sections examined in the adults,the proline bands are faintly labeled in IVca.
In the rostral calcarine cortex of both adult animals, the col-umns are difficult to appreciate in the dorsal and ventral lipsbecause the contrast is low and the borders are indistinct.
Two-month-old animal (96-10)The ODC pattern has been previously described (Fonta et al.,
2000). The columns are less well demarcated in IVca than inIVcb, but are nevertheless clearly present in both sublaminae(Fig. 3B).
24-day-old animal (97-12)The ODC pattern is clearly pronounced in the calcarine cortex.
In this animal, the alternate stripes exhibit different patterns thanthose found in older animals. The columns are clearly present inboth sublaminae IVca and IVcb. On a same section, labeledstripes can be seen only in IVca, or only in IVcb, or in both IVcaand IVcb (Fig. 3C). Moreover, we observed large unlabeled zonesin both hemispheres. In the two juvenile animals 96-10 and 97-12,the ODC pattern is more distinct in the rostral part than in thecaudal part of the visual cortex.
Discussion
In the present paper, we show ODC organization in adult andjuvenile marmosets. The ODC pattern in very young marmosetshave not yet been described. These results complete Spatz’s dataconcerning the presence of ODC before 3 postnatal months. Thepresence of ODC pattern in the adult marmousets that we havedemonstrated in this study refutes previous studies reporting thatsuch adults exhibit no ODC (DeBruyn & Casagrande, 1981; Spatz,1979, 1989). After (3H) proline and horseradish peroxidase (HRP)eye injection, the authors found a continuous band of label in layerIVc, suggesting a complete intermixing of the crossed and un-crossed retino-cortical pathways.
We saw unequivocal presence of ODC in two adult animalswith a transneuronal transport of tritiated amino acid, both infrontal and horizontal sections. The difference between the previ-ous published works and the present data may be explained bymethodological considerations. The continuous band of labelingpreviously seen in adult marmoset could result from several tech-nical artifacts (spillover, quality of eye injection, amount of radio-activity, emulsion . . . ). The presence of spillover in the LGN hasbeen mentioned (Spatz, 1979), thus it might contribute to blur theODC. Our eye-injection technique was similar to that used in theprevious reports, but we employed a radioactive tracer with great
ODC in the adult marmoset 409

Fig. 2. Ocular Dominance Columns (ODC) in adult animals observed in unfolded visual cortex from autoradiographs through layerIVcb. Left eye was injected with tritiated proline and labeled geniculo-striate fibers were found in layer IVc as alternating bands. Inbright-field illumination, the [3H] proline-labeled columns appear as dark regions. A: Montage of ODC from tangential sections of adultanimals dorsal (D) calcarine gyrus (left) and ventral (V) calcarine gyrus (right) in the ipsilateral visual cortex of animal 98-4; B: ventralcalcarine gyrus in contralateral visual cortex of animal 97-11. Asterisks indicate the representation of the blind spot; it is visualizedin both adults, in the posterior part of the ventral calcarine gyrus; this region is dark in the ipsilateral hemisphere (Fig. 2A) and lightin the contralateral hemisphere (Fig. 2B). Arrowheads limit V1 area. C and D: Individuals sections showing ODC bands in righthemisphere of 97-11 animal, in the occipital dorso-lateral part (C), in the dorsal calcarine gyrus (D); these two sections show the largestpart of layer IVcb occupied by ODC. Scale bar5 1 mm.
410 C. Chappert-Piquemal et al.
specific activity and labeled on four hydrogen atoms [L-(2,3,4,5-3H) proline].
These results are important as they may give support to theinterpretation of results from a previous functional study. Mark-stahler et al. (1998) described “physiological” ODC in adult ani-mals, by way of activity-dependent immediate early gene expression(Krox-24) after monocular deprivation (TTX injection) and visualstimulation. Although “physiological ODC” are revealed after aspecific protocol, they can now be linked to the “anatomical ODC”described in the present work. The presence of anatomical ODC inthe adult marmoset may also explain some periodic shifts in oculardominance observed by electrophysiological recordings (Sengpielet al., 1996). The latter study shows that most cells in layer IV ofV1 are equally responsive through the two eyes. As it is notori-ously difficult to record specific electrophysiological responses inlayer IVc of the primary visual cortex, these binocular cells mightbelong to IVa and IVb layers rather than to IVc layers.
By contrast with the macaque in which the ODC are stabilizedafter complete segregation in juvenile animals, the ODC organi-zation inCallithrix jacchuscould change between childhood andadulthood. In 2-month-old animals the sizes of labeled and un-labeled columns are not different, and the afferent terminationsfrom both eyes appear to occupy distinct territories of similar size(Fonta et al., 2000). Present results could suggest some overlapbetween inputs from the two eyes in adult animals; the labeledterritories are larger than the unlabeled territories and this cannotbe explained by the naso-temporal asymmetry which exists inadult macaque monkeys (LeVay et al., 1975; Tychsen & Burkhal-ter, 1997). The study of Markstahler et al. (1998) shows similarresults: in adult marmoset, “physiological ODC” are visualized byimmediate early gene expression (Krox 24) and the Krox-positivezones (cortical territory linked to activated eye) are larger than theKrox-negative zones.
The topographical arrangement of the ODC in V1 is different inthe marmoset monkey than in the macaque: in the marmoset, onlya few ODC appear oriented perpendicularly to the V10V2 border,where they form an irregular mosaic. From horizontal sections, theaverage width of the adult marmoset ODC is;211–257mm(present report); this ODC width is similar to that of the squirrelmonkey (;225 mm in width) (Horton & Hocking, 1996a). How-ever, the ODC organization in layer IVc is different among theNew World Monkeys. In the squirrel monkey (Horton & Hocking,1996a), as in the owl monkey (Rowe et al., 1978), the ODC aremuch more apparent in IVca, whereas in the adult marmoset theyare more distinct in layer IVcb. In the Saimiri, this could resultfrom an artifact due to the spillover of the transneuronal marker inthe parvocellular layers of LGN (because of lack of fibrousinterlaminar zones) which could explain the blurred appearance ofthe ODC in layer IVcb.
In conclusion, the present study provides evidence that in theadult Callithrix jacchusterminals of thalamic afferents connectedto each eye are organized in (well defined) cortical columns. Thisfinding refutes several studies in the literature which claim thatsuch columns are absent in adult marmoset monkey, thus, itemphasizes the usefulness of the marmoset as a model in visualstudies.
Acknowledgments
The authors thank A. Maiorano for animal care and P. Saillour forher technical help. This work was supported by Centre National de laRecherche Scientifique and Conseil Régional Midi-Pyrénées No. 9609574.
Fig. 3. Proline-labeled ODC observed on autoradiographs of frontal sec-tions of the primary visual cortex ipsilateral to the eye injection, at similarlevels. (A) In adult animal (97-11), the ODC presence is obvious in IVcb,they faintly appear in IVca. (B) In 2-month-old animal (96-10), the ODCare demarcated in IVca and IVcb. (C) In 24-day-old animal, the prolinebands appear in either IVca, IVcb only, or in both IVca and IVcb. Scalebar5 500 mm.
ODC in the adult marmoset 411

References
DeBruyn, E.J. & Casagrande, V.A. (1981). Demonstration of oculardominance columns in a New World primate by means of monoculardeprivation.Brain Research207, 453–458.
Florence, S.L., Conley, M. & Casagrande, V.A. (1986). Ocular dom-inance columns and retinal projections in New World Spider Monkeys(Ateles ater). Journal of Comparative Neurology243, 234–248.
Fonta, C., Chappert, C. & Imbert, M. (1997). N-Methyl-D-AspartateSubunit R1 Involvement in the postnatal organization of the primaryvisual cortex ofCallithrix jacchus. Journal of Comparative Neurology386, 260–276.
Fonta, C., Chappert, C. & Imbert, M. (2000). Effect of monoculardeprivation on NMDAR1 immunostaining in ocular dominance col-umns of the marmosetCallithrix jacchus. Visual Neuroscience17,345–352.
Hendrickson, A.E., Wilson, J.R. & Ogren, M.P. (1978). The neuroana-tomical organization of pathways between the dorsal lateral geniculatenucleus and visual cortex in Old World and New World primates.Journal of Comparative Neurology182, 123–136.
Hess, D.T. & Edwards, M.A. (1987). Anatomical demonstration of ocularsegregation in the retino-geniculo-cortical pathway of the New WorldCapuchin Monkey (Cebus apella). Journal of Comparative Neurology264, 409–420.
Horton, J.C. & Hocking, D.R. (1996a). Anatomical demonstration ofocular dominance columns in striate cortex of the squirrel monkey.Journal of Neuroscience16, 5510–5522.
Horton, J.C. & Hocking, D.R. (1996b). Intrinsic variability of oculardominance column periodicity in normal macaque monkeys.Journal ofNeuroscience16, 7228–7239.
Hubel, D.H. & Wiesel, T.N. (1972). Laminar and columnar distributionof geniculo-cortical fibers in the macaque monkey.Journal of Com-parative Neurology146, 421–450.
LeVay, S., Hubel, D.H. & Wiesel, T.N. (1975). The pattern of oculardominance columns in macaque visual cortex revealed by a reducedsilver stain.Journal of Comparative Neurology159, 559–576.
LeVay, S., Conolly, M., Houde, J. & Van Essen, D.C. (1985). Thecomplete pattern of ocular dominance stripes in the striate cortex andvisual field of the macaque monkey.Journal of Neuroscience5,486–501.
Markstahler, U., Bach, M. & Spatz, A. (1998). Transient molecularvisualization of ocular dominance columns (ODCs) in normal adultmarmosets despite the desegregated termination of the retino-geniculo-cortical pathways.Journal of Comparative Neurology393, 118–134.
Rosa, M.G.P., Gattass, R. & Fiorani, M.J. (1988). Complete pattern ofocular dominance stripes in V1 of a New World monkey,Cebus apella.Experimental Brain Research72, 645–648.
Rosa, M.G.P., Gattass, R., Fiorani, M.J. & Soares, J.G.M. (1992).Laminar, columnar and topographic aspects of ocular dominance in theprimary visual cortex ofCebusmonkeys.Experimental Brain Research88, 249–264.
Rowe, M.H., Benevento, L.A. & Rezak, M. (1978). Some observationson the patterns of segregated geniculate inputs to the visual cortex inNew World primates: An autoradiographic study.Brain Research159,371–378.
Sengpiel, F., Troilo, D., Kind, P.C., Graham, B. & Blakemore, C.(1996). Functional architecture of area 17 in normal and monocularlydeprived marmosets (Callithrix jacchus). Visual Neuroscience13,145–160.
Spatz, W.B. (1979). The retino-geniculo-cortical pathway in Callithrix. II.The geniculo-cortical projection.Experimental Brain Research36,401–410.
Spatz, W.B. (1989). Loss of ocular dominance columns with maturity inthe monkey,Callithix jacchus. Brain Research488, 376–380.
Tigges, J. & Tigges, M. (1979). Ocular dominance columns in thestriate cortex of chimpanzee (Pan troglodytes). Brain Research166,386–390.
Tychsen, L. & Burkhalter, A. (1997). Nasotemporal asymmetries inV1: Ocular dominance columns of infant, adult, and strabismic ma-caque monkeys.Journal of Comparative Neurology388, 32–46.
412 C. Chappert-Piquemal et al.
Copyright © 2022 FDOKUMEN


















![Non-amine dopamine transporter probe [3H]tropoxene distributes to dopamine-rich regions of monkey brain](https://static.fdokumen.com/doc/165x107/63224d2f050768990e0fcb6c/non-amine-dopamine-transporter-probe-3htropoxene-distributes-to-dopamine-rich.jpg)

