The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina
60
The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina Richard F. Kay a, * , J.G. Fleagle b , T.R.T. Mitchell a , Matthew Colbert c , Tom Bown d , Dennis W. Powers e a Department of Biological Anthropology and Anatomy, Duke University, Durham, NC 27708-0383, USA b Department of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794, USA c Jackson School of Geological Sciences, University of Texas at Austin, Austin, TX 78712-0254, USA d Erathem-Vanir Geological, 2300 Arapahoe Avenue, No.236, Boulder, CO 80302, USA e 140 Hemley Road, Anthony, TX 79821, USA Received 11 January 2006; accepted 7 September 2007 Abstract Dolichocebus is known from the type skull encased in a concretion, numerous isolated teeth, parts of two mandibles, and a talus. The spec- imens come from the Trelew Member (early Miocene, Colhuehuapian South American Land Mammal Age) of the Sarmiento Formation near the village of Gaiman, Chubut Province, Argentina, dated to about 20 Ma. We describe all Dolichocebus fossil material using conventional surface anatomy and micro-CT data from the cranium. The new material and newly imaged internal anatomy of the skull demonstrate that anatomical characters hitherto supposed to support a phyletic link between Dolichocebus and either callitrichines (marmosets, tamarins, and Callimico) or Saimiri (squirrel monkeys) are either indeterminate or absent. To more fully explore the phyletic position of Dolichocebus, we undertook a com- prehensive phylogenetic analysis. We examined 268 characters of the cranium and dentition of 16 living platyrrhine genera, some late Oligocene and early Miocene platyrrhines, Tarsius, some Eocene and Oligocene stem anthropoids, and several extant catarrhines. These analyses consis- tently indicate that Dolichocebus is a stem platyrrhine, as are late Oligocene Branisella and early Miocene Tremacebus, Soriacebus, and Car- locebus. Platyrrhine evolution often is conceived of as a single ancient adaptive radiation. Review of all available phyolgenetic data suggests a more layered evolutionary pattern, with several independent extinct clades filling modern platyrrhine niche space, and modern platyrrhine families and subfamilies appearing over a nine-million-year interval in the Miocene. The outcome of these analyses highlights the pervasiveness of homoplasy in dental and cranial characters. Homoplasy is a real evolutionary phenomenon that is present at all levels of biological analysis, from amino-acid sequences to aspects of adult bony morphology, behavior, and adaptation. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Argentina; Colhuehuapian; Dolichocebus; Gaiman; Miocene; Platyrrhine origins; Sarmiento Formation Introduction It is now generally agreed that platyrrhines (New World monkeys) are most closely related to catarrhines (Old World monkeys and apes). Fossil evidence indicates that Africa is the most likely origin for the last common ancestor of these taxa, suggesting that the ancestor of extant platyrrhines must have crossed the Atlantic Ocean (Hoffstetter, 1980; Tarling, 1980; Hartwig, 1994; Houle, 1999). The timing of this event is not well constrained. Molecular studies have dated the plat- yrrhine-catarrhine divergence to the middle Eocene (~43 Ma, Eizirik et al., 2004), 37.0 3.0 Ma, or 38.9 4.0 Ma (Poux et al., 2006), while the earliest catarrhines in the fossil record date to the late Eocene. Thus, this migration must have oc- curred sometime before the late Eocene (i.e., earlier than 37 Ma) (Kay et al., 2004c; Seiffert et al., 2004), or a stem * Corresponding author. Tel.: þ1 919 684 2143; fax: þ1 919 684 8034. E-mail address: [email protected] (R.F. Kay). 0047-2484/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.jhevol.2007.09.002 Available online at www.sciencedirect.com Journal of Human Evolution 54 (2008) 323e382
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The anatomy of Dolichocebus gaimanensis, a stem platyrrhine monkey from Argentina

Available online at www.sciencedirect.com
Journal of Human Evolution 54 (2008) 323e382
The anatomy of Dolichocebus gaimanensis, a stemplatyrrhine monkey from Argentina
Richard F. Kay a,*, J.G. Fleagle b, T.R.T. Mitchell a, Matthew Colbert c,Tom Bown d, Dennis W. Powers e
a Department of Biological Anthropology and Anatomy, Duke University, Durham, NC 27708-0383, USAb Department of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794, USA
c Jackson School of Geological Sciences, University of Texas at Austin, Austin, TX 78712-0254, USAd Erathem-Vanir Geological, 2300 Arapahoe Avenue, No.236, Boulder, CO 80302, USA
e 140 Hemley Road, Anthony, TX 79821, USA
Received 11 January 2006; accepted 7 September 2007
Abstract
Dolichocebus is known from the type skull encased in a concretion, numerous isolated teeth, parts of two mandibles, and a talus. The spec-imens come from the Trelew Member (early Miocene, Colhuehuapian South American Land Mammal Age) of the Sarmiento Formation near thevillage of Gaiman, Chubut Province, Argentina, dated to about 20 Ma. We describe all Dolichocebus fossil material using conventional surfaceanatomy and micro-CT data from the cranium. The new material and newly imaged internal anatomy of the skull demonstrate that anatomicalcharacters hitherto supposed to support a phyletic link between Dolichocebus and either callitrichines (marmosets, tamarins, and Callimico) orSaimiri (squirrel monkeys) are either indeterminate or absent. To more fully explore the phyletic position of Dolichocebus, we undertook a com-prehensive phylogenetic analysis. We examined 268 characters of the cranium and dentition of 16 living platyrrhine genera, some late Oligoceneand early Miocene platyrrhines, Tarsius, some Eocene and Oligocene stem anthropoids, and several extant catarrhines. These analyses consis-tently indicate that Dolichocebus is a stem platyrrhine, as are late Oligocene Branisella and early Miocene Tremacebus, Soriacebus, and Car-locebus. Platyrrhine evolution often is conceived of as a single ancient adaptive radiation. Review of all available phyolgenetic data suggestsa more layered evolutionary pattern, with several independent extinct clades filling modern platyrrhine niche space, and modern platyrrhinefamilies and subfamilies appearing over a nine-million-year interval in the Miocene. The outcome of these analyses highlights the pervasivenessof homoplasy in dental and cranial characters. Homoplasy is a real evolutionary phenomenon that is present at all levels of biological analysis,from amino-acid sequences to aspects of adult bony morphology, behavior, and adaptation.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Argentina; Colhuehuapian; Dolichocebus; Gaiman; Miocene; Platyrrhine origins; Sarmiento Formation
Introduction
It is now generally agreed that platyrrhines (New Worldmonkeys) are most closely related to catarrhines (Old Worldmonkeys and apes). Fossil evidence indicates that Africa isthe most likely origin for the last common ancestor of these
* Corresponding author. Tel.: þ1 919 684 2143; fax: þ1 919 684 8034.
E-mail address: [email protected] (R.F. Kay).
0047-2484/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jhevol.2007.09.002
taxa, suggesting that the ancestor of extant platyrrhines musthave crossed the Atlantic Ocean (Hoffstetter, 1980; Tarling,1980; Hartwig, 1994; Houle, 1999). The timing of this eventis not well constrained. Molecular studies have dated the plat-yrrhine-catarrhine divergence to the middle Eocene (~43 Ma,Eizirik et al., 2004), 37.0� 3.0 Ma, or 38.9� 4.0 Ma (Pouxet al., 2006), while the earliest catarrhines in the fossil recorddate to the late Eocene. Thus, this migration must have oc-curred sometime before the late Eocene (i.e., earlier than37 Ma) (Kay et al., 2004c; Seiffert et al., 2004), or a stem
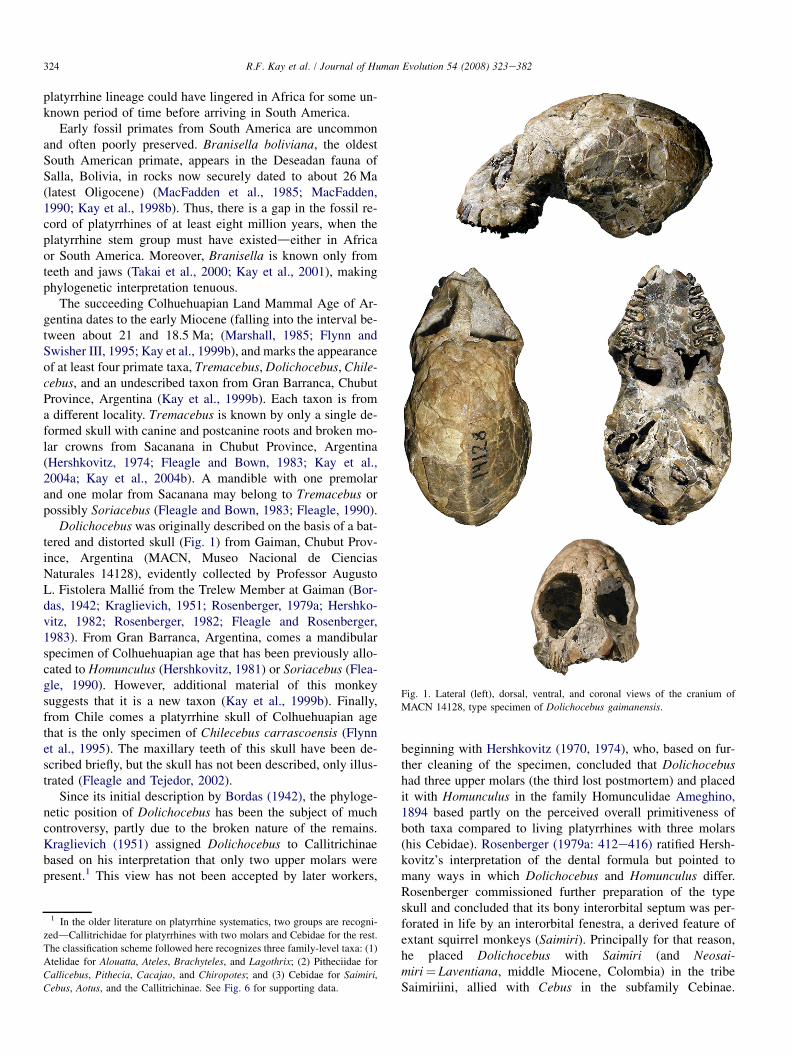
Fig. 1. Lateral (left), dorsal, ventral, and coronal views of the cranium of
MACN 14128, type specimen of Dolichocebus gaimanensis.
324 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
platyrrhine lineage could have lingered in Africa for some un-known period of time before arriving in South America.
Early fossil primates from South America are uncommonand often poorly preserved. Branisella boliviana, the oldestSouth American primate, appears in the Deseadan fauna ofSalla, Bolivia, in rocks now securely dated to about 26 Ma(latest Oligocene) (MacFadden et al., 1985; MacFadden,1990; Kay et al., 1998b). Thus, there is a gap in the fossil re-cord of platyrrhines of at least eight million years, when theplatyrrhine stem group must have existeddeither in Africaor South America. Moreover, Branisella is known only fromteeth and jaws (Takai et al., 2000; Kay et al., 2001), makingphylogenetic interpretation tenuous.
The succeeding Colhuehuapian Land Mammal Age of Ar-gentina dates to the early Miocene (falling into the interval be-tween about 21 and 18.5 Ma; (Marshall, 1985; Flynn andSwisher III, 1995; Kay et al., 1999b), and marks the appearanceof at least four primate taxa, Tremacebus, Dolichocebus, Chile-cebus, and an undescribed taxon from Gran Barranca, ChubutProvince, Argentina (Kay et al., 1999b). Each taxon is froma different locality. Tremacebus is known by only a single de-formed skull with canine and postcanine roots and broken mo-lar crowns from Sacanana in Chubut Province, Argentina(Hershkovitz, 1974; Fleagle and Bown, 1983; Kay et al.,2004a; Kay et al., 2004b). A mandible with one premolarand one molar from Sacanana may belong to Tremacebus orpossibly Soriacebus (Fleagle and Bown, 1983; Fleagle, 1990).
Dolichocebus was originally described on the basis of a bat-tered and distorted skull (Fig. 1) from Gaiman, Chubut Prov-ince, Argentina (MACN, Museo Nacional de CienciasNaturales 14128), evidently collected by Professor AugustoL. Fistolera Mallie from the Trelew Member at Gaiman (Bor-das, 1942; Kraglievich, 1951; Rosenberger, 1979a; Hershko-vitz, 1982; Rosenberger, 1982; Fleagle and Rosenberger,1983). From Gran Barranca, Argentina, comes a mandibularspecimen of Colhuehuapian age that has been previously allo-cated to Homunculus (Hershkovitz, 1981) or Soriacebus (Flea-gle, 1990). However, additional material of this monkeysuggests that it is a new taxon (Kay et al., 1999b). Finally,from Chile comes a platyrrhine skull of Colhuehuapian agethat is the only specimen of Chilecebus carrascoensis (Flynnet al., 1995). The maxillary teeth of this skull have been de-scribed briefly, but the skull has not been described, only illus-trated (Fleagle and Tejedor, 2002).
Since its initial description by Bordas (1942), the phyloge-netic position of Dolichocebus has been the subject of muchcontroversy, partly due to the broken nature of the remains.Kraglievich (1951) assigned Dolichocebus to Callitrichinaebased on his interpretation that only two upper molars werepresent.1 This view has not been accepted by later workers,
1 In the older literature on platyrrhine systematics, two groups are recogni-
zeddCallitrichidae for platyrrhines with two molars and Cebidae for the rest.
The classification scheme followed here recognizes three family-level taxa: (1)
Atelidae for Alouatta, Ateles, Brachyteles, and Lagothrix; (2) Pitheciidae for
Callicebus, Pithecia, Cacajao, and Chiropotes; and (3) Cebidae for Saimiri,Cebus, Aotus, and the Callitrichinae. See Fig. 6 for supporting data.
beginning with Hershkovitz (1970, 1974), who, based on fur-ther cleaning of the specimen, concluded that Dolichocebushad three upper molars (the third lost postmortem) and placedit with Homunculus in the family Homunculidae Ameghino,1894 based partly on the perceived overall primitiveness ofboth taxa compared to living platyrrhines with three molars(his Cebidae). Rosenberger (1979a: 412e416) ratified Hersh-kovitz’s interpretation of the dental formula but pointed tomany ways in which Dolichocebus and Homunculus differ.Rosenberger commissioned further preparation of the typeskull and concluded that its bony interorbital septum was per-forated in life by an interorbital fenestra, a derived feature ofextant squirrel monkeys (Saimiri). Principally for that reason,he placed Dolichocebus with Saimiri (and Neosai-miri¼ Laventiana, middle Miocene, Colombia) in the tribeSaimiriini, allied with Cebus in the subfamily Cebinae.

C
A
B
Dolichocebus
Dolichocebus
Dolichocebus
Saimiri
Saimiri
Saimiri
Marmosets & Tamarins
Marmosets & Tamarins
Marmosets & Tamarins
Cebus
Cebus
Cebus
Aotus
Aotus
Aotus
Pitheciidae
Pitheciidae
Pitheciidae
Atelidae
Atelidae
Atelidae
Fig. 2. Contrasting views of the affinities of Dolichocebus relative to living
platyrrhines. (A) Dolichocebus is a stem platyrrhine (Hershkovitz, 1982;
Fleagle and Bown, 1983). (B) Dolichocebus is a relative of marmosets and
tamarins (Horovitz, 1999). (C) Dolichocebus is the sister taxon of Saimiri
(Rosenberger, 1979a).
325R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
However, this interpretation has been open to question becausethere is not universal agreement as to whether the bony inter-orbital fenestra is anatomical or the result of postmortem dam-age (the specimen was distorted and broken in other waysbefore and after burial) (Hershkovitz, 1982; Rosenberger,1982; Fleagle and Rosenberger, 1983; Ford, 1986; Horovitz,1999).
Between 1983 and 1989, joint Argentine/American expedi-tions collected a number of isolated primate teeth and a talusfrom Gaiman, the type locality of Dolichocebus. The talus andsome of the teeth have already been described (Fleagle andBown, 1983; Gebo and Simons, 1987). Fleagle and Bown(1983) identified several features of isolated upper molarsfrom Gaiman that resemble the extant platyrrhine Saimiri butalso Oligocene African anthropoids Aegyptopithecus and Pro-pliopithecus. They identified these as shared primitive features.Gebo and Simons (1987) called attention to general similaritiesbetween the Dolichocebus talus and that of ‘‘generalized NewWorld monkeys’’ but found no certain shared derived featuresindicating a relationship with any platyrrhine lineage.
In the most recent comprehensive phylogenetic survey ofliving and fossil platyrrhines, Horovitz (1999) did not considerresolvable the arguments about interorbital fenestration inDolichocebus. In her character-taxon matrix, Horovitz scoredDolichocebus for only five cranial characters and none pertain-ing to the talus. In her phylogenetic analysis using these fivecranial and 22 dental characters, she found that Dolichocebuswas a stem callitrichine with three molars (see footnote 1 forinformation about the taxonomic terminology used here). Insummary, there remains little agreement about the place ofDolichocebus within the platyrrhine clade, with the three cur-rently held views being that (1) it is a basal callitrichine (Hor-ovitz, 1999), (2) it is specially related to Saimiri (Rosenberger,1979a), and (3) it is a sister group of all living platyrrhines(Fleagle and Kay, 1989) (Fig. 2).
As already noted, following Fleagle and Bown’s (1983) con-tribution, additional primate material has come to light atGaiman. Expeditions from Stony Brook University in collabo-ration with a group from MACN, Buenos Aires, collected iso-lated teeth, an edentulous mandible, and a talus in the 1980sand early 1990s. Later, Dr. Guiomar Vucetich of Mueso de LaPlata found two mandibles. Taken together, all parts of the per-manent dentition are now represented. In addition, the type skullwas recently CT-scanned in an effort to clarify the controversialquestion of whether there was an interorbital fenestra and toexamine other hitherto inaccessible features of the skull.
The purpose of the present contribution is threefold: (1) tosummarize the available evidence concerning the stratigraphyand geologic age of Colhuehuapian fossils at Gaiman; (2) todescribe and illustrate the dental specimens, as well as to offernew details about the skull based on high-resolution CT scans;and (3) to reconsider the phylogenetic position of Dolichoce-bus and other late Oligocene and early Mocene platyrrhines.
With respect to this last goal, we note that there coexist inthe current literature two radically different hypotheses aboutplatyrrhine cladgenesis. One hypothesis, which we will callthe ‘‘deep-time hypothesis,’’ is that living platyrrhines arose
as single adaptive radiation of great antiquity. Soon afterthey arose, platyrrhines entered adaptive niches that are simi-lar to those occupied by today’s platyrrhine niches. The occu-pants of those niches are the ancestors of the modern clades onSouth American monkeys. This view was foreshadowed by thework of George G. Simpson and Bryan Patterson (Pattersonand Pascual, 1972; Simpson, 1980). Its current proponentsplace all the known late Oligocene-to-Recent South Americanfossil monkeys within the extant platyrrhine familiesdCebi-dae, Atelidae, and Pitheciidae (Rosenberger, 1979b, 1980,1988; Rosenberger et al., 1990; Rosenberger, 1992, 2000;

326 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Rosenberger, 2002; Tejedor et al., 2006). A second hypothesis,which we call the ‘‘layered hypothesis,’’ envisions a morecomplex evolution of platyrrhines and their adaptive niches.Earliest known platyrrhines are stem taxa. Some of thesetaxa followed adaptive strategies similar to those alive today;others entered different niches not represented today. Therewas no single cladogenetic event in platyrrhine evolution. Liv-ing family clades diverged at around 20 Ma. They coexistedthrough the early and middle Miocene with stem taxa (Kay,1990; Fleagle et al., 1997; Kay et al., 1998a). Our phyloge-netic analysis of Dolichocebus and some late Oligocene andearly Miocene platyrrhines revisits this debate.
Sacanana
Gran Barranca
Río Chubut
Gan GanRLago
Musters Lago ColhuéHuapí
ComoRivad
Gaiman
Bryn Gwyn
Dolavon
Angostura a
a'
Río Chubut
Fig. 3. Top, map of Chubut Province, Argentina, showing the geographic location
Sacanana, and Gran Barranca. Bottom, map of the lower valley of the Rıo Chubut, s
(lined), and the location of the Sarmiento section (aea0) with respect to towns an
Geology
The geologic setting of the Gaiman fossils was reviewed inFleagle and Bown (1983), to which we add some informationoriginally mentioned in an unpublished manuscript by two ofthe authors of this paper (TB and DWP). The principal fossillocalities with primates are situated in the Province of Chubut,Argentina, and the Dolichocebus remains come from thesouthern barranca along the lower Rıo Chubut Valley nearGaiman (Fig. 3). Dolichocebus fossils come from the TrelewMember of the Sarmiento Formation. Roth (1899) first re-ported fossils from this area. Simpson (1935) reviewed the
GaimanTrelew
ίo Ch
ico
KILOMETERS
doroavia
0 50 100
Trelew
Drofa Dulog
Rawson
Bahia
Engañ
o
0 5 10
of the Colhuehuapian-aged fossil-primate-bearing localities at Trelew/Gaiman,
howing barrancas (bluffs; in stipple) north and south of the river valley, pampas
d villages.

327R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
geology and assigned this fauna to what is now conventionallyreferred to as the Colhuehuapian Land Mammal Age (see alsoBordas, 1939). The primate teeth, mandibles, and talus werecollected in the Trelew Member of the Sarmiento Formationjust above the Bryn Gwyn Member, which forms a distincttopographic bench in the middle of the barranca (Fig. 4).Figure 5 gives a stratigraphic profile of the fossil occurrencesat Gaiman. The precise stratigraphic level of the type skull ofDolichocebus is not known, but it almost certainly came fromthe Trelew Member at Gaiman because fossil land mammalsare known from no other part of the south baranca in thisregion (personal experience and Bordas, 1939).
The Trelew Member contains a mammalian assemblagetypical of the Colhuehuapian South American Land MammalAge (Pascual and Odreman Rivas, 1971). This similarity im-plies that the Trelew fossils are roughly similar in age toColhuehuapian fossil assemblages reported elsewhere in Pata-gonia, notably at Sacanana, where Tremacebus harringtoni oc-curs, and Gran Barranca, where another early primate is found(Kay et al., 1999b). The reported absolute ages of these faunasare somewhat conflicting. Bown et al. (1988) reported a fis-sion-track age of 15.8� 2.5 Ma at 20 m above the fossil levelin the Trelew Member of the Sarmiento Formation at Gaimanthat contains Colhuehuapian mammals. Kay et al. (1999a,b)mentioned a series of 40Ar/39Ar dates on glass and plagioclaseseparates from samples at Gran Barranca in association withColhuehuapian faunas. In the context of an associated mag-netic-polarity profile, they suggested that the bulk of theGran Barranca Colhuehuapian fauna falls within Chron 6rand 6n (19.1e20.5 Ma). Flynn et al. (1995) reported an40Ar/39Ar date for another apparently Colhuehuapian faunafrom Chile of 20.09� 0.27 Ma where Chilecebus is found,in agreement with the dates of Kay et al. (1999a).
A second independent source of information about the ageof the Colhuehuapian at Gaiman derives from the fact thata marine bed in the Gaiman Formation containing sharks,
Fig. 4. Fossil deposits of the Sarmiento Formation, south of the Chubut River
near Gaiman, Chubut Province, Argentina. The fossils come from the Trelew
Member of the Sarmiento Formation, just above the bench in the middle of the
figure.
whales, and penguins overlies the Colhuehuapian levels atGaiman. These marine faunas are of Aquitanian age (Cozzuolet al., 1991; A.L. Cione, unpubl. data). If so, then the underly-ing Trelew Member (and its Colhuehuapian faunas) at Gaimanmust be older than 20.5 Ma, because the Aquitanian interval isdated to 20.5e23.8 Ma (Berggren et al., 1995). This dateagrees with the published dates for Gran Barranca but conflictswith Bown et al.’s (1988) suggestion of a younger age. Bownet al.’s reported date, if correct, suggests that the Trelew Mem-ber is, at least in part, younger than the Santacrucian SALMAfaunas, which are older than 16 Ma (Fleagle et al., 1995). Thisis exceedingly unlikely, and therefore we accept the prepon-derance of evidence that Dolichocebus and other faunas ofColhuehuapian age are ~20 Ma.
Materials and methods
The type specimen of Dolichocebus gaimanensis is a cra-nium, the subject of our CT-imaging analysis, as described be-low. All remains are summarized in Table 1 and a list of dentaldimensions is provided in Tables 2 and 3. Allocation of unas-sociated specimens to a particular taxon is always problematic.Our reasons for assigning all of the new specimens to Dolicho-cebus are as follows:
(1) There is no evidence that more than one taxon is presentamong the isolated teeth available. The dental remainscompose a harmonious assemblage in terms of size andmorphology.
(2) The isolated upper teeth are of appropriate size to go withthe palate and roots of the type skull (Fleagle, 1985).
(3) Dolichocebus is the only primate taxon definitely knownto occur at Gaiman. The only other known ColhuehuapianPatagonian fossil primates to which these teeth can becompared are Tremacebus and the new monkey from theColhuehuapian of Gran Barranca. The morphology ofthe Gaiman teeth, as described below, definitively rulesout allocation to the new Gran Barranca monkey or, forthat matter, to Santacrucian late early Miocene Soriacebus,Carlocebus, or Homunculus. Possible allocation to Trema-cebus deserves more consideration. Both of these taxacome from the Colhuehuapian, and Sacanana and Gaimanare less than 300 km apart. Available material of Tremace-bus consists only of the type cranium with most of theteeth broken off (only parts of M1e3 crowns are preserved)(see Fig. 1 in Kay et al., 2004a). The upper molars fromGaiman differ in some respects from those of Tremacebus.Although both have strong lingual cingula with well-developed M1e2 hypocones, those of Tremacebus aremuch smaller and more rounded in lingual occlusal profilethan those of Dolichocebus. A mandibular fragment fromSacanana with a broken part of P4 and M1 (Fleagle, 1990)may not belong with Tremacebus, but in any event, it isquite unlike the M1 from Gaiman. Based on the foregoing,we conclude that the isolated teeth from Gaiman belongwith the skull.

Sarm
ient
o Fo
rmat
ion
Río Chico Fm.
Pan
d'Az
ucar
Mem
ber
CAS
AMAY
OR
ANC
OLH
UEH
UAP
IAN
Sarm
ient
o Fo
rmat
ion
Terle
w M
embe
r
Bryn GwynMember
SF ?
Gaiman Formation(marine)
Sandstone tuff
conformable contact
erosionalunconformity
erosionalunconformity
conformablecontact
met
ers
0
5
10
Tuffaceoussandstone
Tuffaceous ss.with large-scaletrough cross-strata
Tuffaceous ss.& sandy mudstone withepsilon cross-strata
Tuffaceoussandy mudstone
Tuffaceousmudstone
Tuffaceousmuddy ss.
Bedded calcretenodules
Bedded & unbeddedcalcium carbonateconcretions
Calcium carbonateveinlets
Bedded & unbeddedsilicate nodules
Irregular massesof opaline silicates
Chalcedonygeodes
Fig. 5. Idealized stratigraphic profile at Gaiman showing the Trelew Member, the principal fossil level containing a Colhuehuapian assemblage. Note the position
of the ash level referred to in the text.
328 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
To determine more precisely the limits of bone and matrixand to better appreciate the structural details of the interior ofthe type skull of Dolichocebus, the skull was scanned at theHigh-Resolution X-ray Computed Tomography (CT) Facilityat the University of Texas at Austin, as described by Ketchamand Carlson (2001). X-ray energies were set to 150 kV and0.16 mA using a FeinFocus� X-ray source. X-ray intensitieswere measured using an Image Intensifier detector employinga 1024� 1024 video camera. Each slice was acquired using1000 views (angular orientations), with four samples takenper view. The specimen, mounted in a plastic cylinder, wasscanned with a centered axis of rotation and a source-to-objectdistance of 135 mm (Ketcham and Carlson, 2001). Slice thick-ness and interslice spacing were 0.0466 mm (one video line).The image field was reconstructed to 43.5 mm, based ona maximum field of view of 44.164 mm, yielding an interpixel
spacing of 0.042 mm. Reconstruction parameters were cali-brated to maximize usage of the 16-bit range of grayscalesavailable in the output images. Twenty-seven slices were ac-quired for each rotation of the turntable, with a resulting ac-quisition time of about 11.1 seconds per slice. The dataconsist of 1430 slices, from the front to the back of the skull.
The coronal slice-by-slice animation, representing the orig-inal CT data, may be viewed on the DigiMorph Web site,http://www.digimorph.org/index.phtml. The coronal movie be-gins at the tip of the rostrum; the slices are on the coronalplane. These original data were digitally resliced to provideslice-by-slice animations on the horizontal and sagittal planes.The horizontal animation starts dorsally and passes ventrally;the slices are oriented in dorsal view. The sagittal animationproceeds from left to right through the specimen; the slicesare in left lateral view.

Table 1
Dolichocebus gaimanensis specimens
Museum number Specimen
MACN 14128 Cranium (type specimen)
MACN CH 356 Left M1 (Figs. 6, 7a in Fleagle and Bown, 1983)
MACN CH 362 Talus (Fig. 4 in Gebo and Simons, 1987)
MACN CH 357 Right M3 (Fig. 7c in Fleagle and Bown, 1983)
MACN CH 358 Lingual part of left upper molar (Fig. 7b in
Fleagle and Bown, 1983)
MACN CH 359 Left I1 (Fig. 8 in Fleagle and Bown, 1983)
MACN CH 361 Right canine (Fig. 8 in Fleagle and Bown, 1983)
MACN CH 864 Right P2
MACN CH 865 Right P2
MACN CH 866 Left M2
MACN CH 868a Right P3
MACN CH 868b Right dp3
MACN CH 870 Right lower canine
MACN CH 871 Left lower canine
MACN CH 872 Right I1 or I2
MACN CH 873 Left P3
MACN CH 876 Left M1
MACN CH 878 Right P4
MACN CH 896 Left M1 or M2 (crown eroded, enamel mostly gone)
MACN CH 897a Upper molar fragment
MACN 2CH 897b Left I2 (?)
MACN CH 898 Left P4
MACN CH 1011 Left dp4
MACN CH 1012 Left I2
MACN CH 1300 Mandible fragment with root socket for I2, roots
for canine, and P2e3
MACN CH 1302 Left canine
MACN CH 1303 Right canine
MPEF 5146 Mandible fragment with left M1e2
MPEF 5147 Mandible fragment with right M1e3
All specimens come from the Trelew Member of the Sarmiento Formation at
Gaiman, Chubut Province, Argentina.
329R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
The platyrrhine skulls that constitute the comparative data-base for cranial anatomical data were scanned with variousprotocols, depending on skull size, in order to maximize reso-lution for each specimen. Details of the CT sample and theresolution of the images are summarized in Table 4.
All dental measurements were made using a Wild M-5binocular microscope using 10� oculars and set at 12� or25�, with a calibrated reticle. Anatomical landmarks for themeasurements are summarized in Kay (1977).
Phylogenetic analysis
2 Recently the rostrum and face of a monkey from the locality of Killik Aike
Norte, Rıo Gallegos, Santa Cruz Province, was described as Killikaike blakei
(Tejedor et al., 2006). The diagnosis of this new genus includes comparisons
to extant platyrrhines, but not with abundant comparative material of Homun-culus patagonicus. Homunculus material comes from the same formation and
same approximate stratigraphic levels as Killikaike (Tauber et al., 2004;
Tejedor et al., 2006). Most of the distinctions drawn between Killikaike and
extant platyrrhines are the same as those between Homunculus and extant plat-
yrrhines (e.g., Tauber, 1991), and the published linear dimensions and anatomy
are very similar between Homunculus and Killikaike (Tauber, 1991; Kay et al.,
2005). Provisionally, we regard Killikaike blakei as the same genus and per-
haps the same species as Homunculus patagonicus.
A series of analyses was undertaken to explore the phyloge-netic placement of Dolichocebus and other late Oligocene andearly Miocene platyrrhines. In addition to Dolichocebus, we in-cluded data from 16 genera of extant platyrrhines, the late Oli-gocene Branisella from Salla, Bolivia, the Colhuehuapian(early Miocene) platyrrhine Tremacebus, and the earliest Santa-crucian taxa Soriacebus and Carlocebus. The sole known Pata-gonian middle Miocene primate Proteropithecia, was alsoincluded. Several taxa are excluded because some or all the per-tinent material is undescribed. These include Chilecebus (Col-huehuapian, Chile) (Flynn et al., 1995), Homunculus from the
Santacrucian (latest early Miocene, Santa Cruz Province)2,and an unnamed taxon from the Colhuehuapian of Gran Bar-ranca, Chubut Province, Argentina (Kay et al., 1999b). As out-groups, we included three extant catarrhines (Miopithecustalapoin, Presbytis melalophos, and Hylobates lar), five of thebetter-known taxa of Eocene/Oligocene African anthropoids(Aegyptopithecus, Proteopithecus sylviae, Apidium phiomense,Simonsius [¼ Parapithecus] grangeri, and Catopithecusbrowni), and the extant haplorhine Tarsius. An analysis of thepossible relationship of platyrrhines to earlier anthropoids ofAfrica would require a more exhaustive sampling of the Africantaxa. For example, inclusion of other lesser known but older andperhaps more primitive taxa (e.g., Qatrania, Abuqatrania, Pro-pliopithecus, Oligopithecus) has the potential to change thebranching pattern among and between outgroups and platyr-rhines. We used the parsimony criterion and utilized the com-puter program PAUP version 4.0b10 (Swofford, 2002).
The list of dental and cranial traits comes mostly from ear-lier studies (Kay et al., 2004c). However, we have added sev-eral cranial characters and revised some cranial and dentalcharacters. We evaluated 268 characters (85 cranial and 183dental). Among the platyrrhines used in this study, we identi-fied 39 characters that exhibit no variability among the an-thropoid taxa studied here (but which are included in thecharacter-taxon matrix so as to enable future comparisonswith nonanthropoid taxa in which they do vary). Thirty vari-able characters in the data set are parsimony-uninformative be-cause they are autapomorphies of terminal taxa. This leaves199 parsimony-informative characters upon which to baseour phylogenetic observations. A list of all cranial and dentalcharacters and their states is given in Appendix 1. The charac-ter-taxon matrix is presented in Appendix 2. A list of speci-mens examined for the phylogenetic analysis is provided inAppendix 3. Missing data can negatively impact the outcomeof an analysis by affecting resolution, making determination ofpolarity uncertain, and underrepresenting homoplasy (Nixonand Davis, 1991; Simmons, 1993; Kay and Williams, 1994).Data for the extant taxa are complete. (However, some charac-ters cannot be scored. For example, the position of the M1 par-aconid relative to protoconid and metaconid is scored as‘‘missing’’ in any taxon where a paraconid is absent.) Datafor the extinct taxa are incomplete. A list of the percentagecompleteness of all taxa is given in Appendix 4.
We considered including postcranial characters based onthe work of Ford, Dagosto, Gebo, and others (Ford, 1986;

Table 2
Dimensions (mm) of the upper teeth of Dolichocebus gaimanensis
MACN
CH 356
MACN
CH 357
MACN
CH 359
MACN
CH 361
MACN
CH 864
MACN
CH 876
MACN
CH 878
MACN
CH 1011
MACN
CH 1012
MACN
CH 1302
MACN
CH 1303
I1
MD 2.98
BL 2.19
C
MD 3.96 3.20 3.45
BL 3.75 2.94 2.94
Height 7.03 4.35 4.16
P2
MD 2.62
BL 2.81
P3
MD 2.56
BL 4.09
P4
MD 2.75
BL 4.73
dp4
MD 3.32
BL 4.48
M1
MD 4.03
BL 5.43
M2
MD 4.22
BL 5.50
M3
MD 2.88
BL 4.41
Table 3
Dimensions (mm) of the lower teeth of Dolichocebus gaimanensis
MACN
CH 865
MACN
CH 866
MACN
CH 868a
MACN
CH 868b
MACN
CH 870
MACN
CH 871
MACN
CH 872
MACN
CH 873
MACN
CH 898
MPEF
5146
MPEF
5147
I1
MD 1.66
BL 2.24
C
MD 3.77 3.32
BL 3.00 2.62
P2
MD 2.88
BL 2.75
P3
MD 2.75 2.75
BL 3.07 3.13
dp3
MD 3.20
BL 2.88
P4
MD 2.88
BL 3.52
M1
MD 3.91
BL (tal) 3.45
BL (trig) 3.46
M2
MD 4.03 3.90
BL (tal) 3.32 3.30
BL (trig) 3.20 3.20
330 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Table 4
Skulls of living platyrrhines examined by CT-imaging and used in the comparative sample
Taxon Specimen Field of
reconstruction
(mm)
Coronal interslice
spacing (mm)
Interslice spacing for
coronal and horizontal
reslicing (mm)
Alouatta guariba female USNM 518255 104 0.11104 0.10156
Aotus lemurinus USNM 464844 52 0.05708 0.0508
Ateles geoffroyi female USNM 291056 93 0.1017 0.0908
Callicebus torquatus USNM 406411 66 0.0723 0.0645
Callimico goeldii USNM 303323 42 0.04635 0.0410
Callithrix (Mico) argentata female USNM 239463 36 0.03778 d
Callithrix jacchus jacchus USNM 503885 35 0.03378 d
Callithrix jacchus female USNM 503895 36 0.04077 dCebuella pygmaea male USNM 337324 28 0.03194 d
Cebus olivaceus USNM 338960 79 0.08683 0.07715
Saguinus fuscicollis USNM 518577 36 0.03778 d
Saguinus fuscicollis Kay collection 142 37.5 0.0412 0.03662
Saimiri sciureus sciureus NMNM 518538 49 0.0551 0.0479
Pithecia sp. Fleagle collection (Suriname?) 65 0.063477 d
331R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Meldrum, 1993; Dagosto and Gebo, 1994; Ford, 1994; Geboet al., 1994) but ruled it out from the present analysis becausenone of the taxa of late Oligocene/early Miocene platyrrhinesevaluated here are known from very much postcranial mate-rial. Just a few isolated specimens are described for Carloce-bus or Soriacebus (allocations uncertain) and only the talusof Dolichocebus is represented. No postcranial elements ofBranisella and Tremacebus are known.
In our PAUP analyses, a number of multistate characterswere used (indicated in Appendix 1). There is disagreementabout whether this sort of character should be ordered or un-ordered (Mickevich, 1982; Mabee, 1989; Mickevich andWeller, 1990; Hauser and Presch, 1991; Slowinski, 1993;Mabee, 2000). Multistate characters should be designated as‘‘ordered’’ if changes from one state to another require passingthrough intermediate states also represented in the data set(e.g., to go from ‘‘absent’’ to‘‘large’’ one must pass throughthe state ‘‘small’’) (Slowinski, 1993). Ordering characters ex-cludes the possibility that characters can change from one endof a morphocline to the other without an intermediate state(Hauser and Presch, 1991). To evaluate the effect of characterordering on tree topologies derived from our data set, we ranour analyses with (1) all multistate characters unordered and(2) some multistate characters ordered.
Aweighting scheme was used in which some multistate char-acters were ordered and their weight scaled. By using scaledweighting, ordered multistate characters are set to have thesame weight regardless of the number of characters states. Thetotal breadth of each morphocline is set to a base weight of100. For a two-state character, it takes one step to cross the mor-phocline (0 to 1 or 1 to 0) and each step is assigned a weight of100. For an ordered three-state character it takes two steps tocross the morphocline (0 to 1, and 1 to 2, or the reverse), soeach step is assigned a value of 50. This weighting scheme elim-inates the situation where ordered multistate characters couldhave more weight (because they use more steps) and thus differ-entially influence tree topologies.
Following the recommendation of Springer et al. (2001),we established a ‘‘molecular scaffold’’ upon which to
superimpose the character distributions using the ‘‘ConstraintsBackbone’’ option of PAUP. Under the ‘‘backbone’’ constraint,extinct taxa are unconstrained and can move about on themolecular phylogenetic scaffold. Springer et al. argued thatclades established by maximum-parsimony analysis of mole-cular data should be assumed to depict a clade accurately ifthey receive �90% bootstrap support. In this case, molecu-lar-sequence data establishes the phylogenetic relationshipsamong many extant platyrrhines to a high degree of probabil-ity (Fig. 6). This branching pattern has been supplemented andverified in several cases with Alu data (see Fig. 6 caption fordetails). For completeness, we also examined the conse-quences of unconstrained analyses.
The distinction between polymorphic and uncertain characterstates was enforced. The tree-bisection-reconnection (TBR)branch-swapping algorithms of PAUP were selected. (Other algo-rithms were tried but never yielded shorter trees). For each set ofcomparisons, starting trees were obtained via stepwise additionwith a random-addition sequence with one tree held at eachstep. The search process was replicated 1000 times.
Abbreviations
Specimens come from the following museums: ANS,Academy of Natural Sciences, Philadelphia; CGM, CairoGovernment Museum, Cairo; DPC, Duke University LemurCenter fossil collections; FMNH, Field Museum of NaturalHistory, mammal collections; MACN, Museo Nacional deCiencias Naturales; MNHN Bol V, Museo Nacional de His-toria Natural, La Paz, Bolivia; MSP, Museu de Zoologia daUniversidade de Sao Paulo, Brazil ; YPM, Yale Peabody Mu-seum paleontology collections; CORD-PZ, Museo de Paleon-tologıa, Facultad de Ciencias Exactas, Fısicas y Naturales dela Universidad Nacional de Cordoba; Tremacebus skull,Rusconi Collection at Museo de Fundacion Miguel Lillo, Tu-cuman, Argentina; MPEF, Museo Paleontologico E. Ferugliode Trelew, Chubut Province, Argentina; and USNM, Smithso-nian Institution, National Museum of Natural History, verte-brate zoology collections.

Tarsius
Hylobates
Miopithecus talapoin
Presbytis melalophos
Callicebus
Pithecia
Cacajao
Chiropotes
Saimiri
Cebus
Aotus
Saguinus
Leontopithecus
Callimico
Callithrix
Cebuella
Alouatta
Ateles
Brachyteles
Lagothrix
Fig. 6. Cladogram of extant platyrrhine primates based on molecular-sequence
and Alu data. Molecular-sequence data provide powerful evidence for resolv-
ing most of the nodes within the extant platyrrhine clade; Alu elementsdshort
interspersed nuclear elements (SINEs)dprovide additional evidence for re-
solving platyrrhine cladogenesis because their mode of evolution is predomi-
nantly unidirectional and homoplasy-free (Hillis, 1999; Bashir et al., 2005).
Molecular-sequence data strongly support the monophyly of the Platyrrhini
and recognition of three clades within itdAtelidae, Cebidae, and Pitheciidae
(Harada et al., 1995; Schneider et al., 1996; Barroso et al., 1997; Schneider
et al., 2001; Singer et al., 2002). The rooting of Callimico with the
Callithrix/Cebuella clade is very strongly supported by sequence data despite
an overall lack of morphological support (Hill, 1957). Eighty-seven Alu ele-
ments support platyrrhine monophyly (Singer et al., 2002; Ray et al., 2005).
The tree figured here is resolved at those nodes where bootstrap support of mo-
lecular-sequence data is equal to or greater than 90% and/or when one or more
Alu supports it. Alu data indicate that Atelidae and Cebidae are sister taxa to
the exclusion of Pitheciidae (Ray et al., 2005). Linkage of callitrichines with
Aotus, and a Saimiri/Cebus group is strongly supported by sequence data and
three Alus. Ray et al. (2005) reported that one Alu links cebines with Aotus
(also supported by recent data from the laboratory of T. Disotell, pers.
comm.). That Callicebus is a basal pitheciine is strong supported by many se-
quence studies and three Alus. Aotus consistently is excluded from the Calli-
cebus-pitheciine clade (contra Rosenberger, 1981). Atelidae has strong
molecular support (including six Alus), with Alouatta as the sister to a clade
consisting of Ateles, Brachyteles, and Lagothrix. Meireles et al. (1999a, b) re-
ported a fully resolved tree for extant Atelidae using a combination of g-glo-
bin, e-globin, RBP, G6PD nuclear genomic sequences, and mitochondrial
COII sequences.
MatrixBone
Interorbitalhole
Lacrimalcrest
Lacrimalcrest
A
B
332 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Results
postorbital flange of
Descriptive anatomy of the cranium the zygomaticFig. 7. (A) Left anterolateral CT reconstruction of Dolichocebus gaimanensisshowing the clear separation of gray values between fossil bone and matrix.
Note especially the good preservation of the ventral margin of the left orbit
but absence of bone on its lateral margin and over glabella. (B) Right antero-
lateral CT reconstruction of the same specimen showing better preservation of
the right lateral margin of the orbit, including the postorbital flange of the zy-
gomatic, but poor preservation of the ventral orbital margin. Bone is absent
from the margins of an interorbital hole on the right and left sides. The anterior
crest of the lacrimal is noted on the right and left sides.
CT-imaging of the type skull clearly distinguishes fossilbone from matrix (Fig. 7A, B). The cranium is cementedwithin a soil-derived concretion, the parent material of whichwas volcanic ash. While its overall integrity was maintainedduring diagenesis and subsequent exposure on the surface,the neurocranium was compressed bilaterally, badly crushedin some areas, and some parts were decayed or broken off be-fore and after fossilization. The rostal part of the basicranium
is preserved bilaterally between the foramen magnum and theanteriormost extent of the petrosals. The area between the an-terior apex of the petrosal and the posterior extent of the M2s,including the entire pterygoid region, is poorly preserved ormissing altogether. A part of the caudal margin of the palatalprocess of the palatine appears to be intact. The incisors andpremaxilla are not preserved. The maxilla preserves the rootsof the canine through M2 bilaterally, but the crowns have beenlost. The zygomatic arches are broken away and lost. Althoughthe face has also been badly distorted and damaged, the inter-orbital and orbital regions are preserved in part. Skull length(prosthioneinion) is estimated to be approximately 68.5 mm.Thus, the skull length is similar to that of extant Callicebus,the titi monkey (Hershkovitz, 1990).
An exhaustive summary of anatomical comparisons betweenDolichocebus, extant platyrrhines, Tarsius, representative basalanthropoids, and crown catarrhines is embodied in the list ofcharacters and their states, and the character-taxon matrix(Appendices 1 and 2, respectively). The Nexus-formatted

333R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
character-taxon matrix is available upon request from the seniorauthor. Some phylogenetically or adaptively significant com-parisons are elaborated in the text. Where mentioned in thetext, comparisons with Tremacebus harringtoni are based onthe type specimen (for more details see Kay et al., 2004a). Ex-cept as noted, cranial observations of Homunculus are based onpersonal examination of the Cordoba skull CORD-PZ 1130,collected by A. Tauber (Tauber, 1991).
Orbital and interorbital region. The medial and ventral mar-gin and a small part of the dorsomedial wall of the left orbit arepreserved (Fig. 7). Parts of the left postorbital septum also re-main. A larger portion of the right orbit has been preserved, al-though the right ventral orbital rim is missing. Towards its apex,the right orbit is preserved dorsally but the optic canal and supe-rior orbital fissure are not present (Fig. 7). From anatomicallandmarks preserved on the left side (ventrally) and right side(superiorly, laterally, and medially), the orbit diameter is esti-mated to be about 14.5 mm in a dorsoventral plane and about14.2 mm mediolaterally. Relative to prosthioneinion length,these orbital dimensions are comparable to those of similarlysized platyrrhines, such as Callicebus moloch. Aotus trivirgatusstands apart in having very large orbits (data in Kay and Kirk,2000). Relative orbit size in early Miocene Homunculus resem-bles that of most extant platyrrhines (Tauber, 1991). The orbitsof Tremacebus are somewhat larger than those of most platyr-rhines but not nearly as large as in extant Aotus (Hershkovitz,1974; Fleagle and Rosenberger, 1983; Martin, 1990; Kayet al., 2004a; Kay et al., 2004b).
On superficial inspection, the frontal bone appears to bedepressed at glabella above weak brow ridges. However, theCT images reveal this region to be completely devoid ofbonedwhat appears to be glabella is actually the naturalcast of the inner table of bone over the frontal lobes. Thus, gla-bellar shape is indeterminate (Fig. 7A).
The interorbital region is too poorly preserved to determinethe size and position of the frontal and nasal bones. The max-illary bone contributes to the ventral orbital rim on the left sideof the skull and would therefore have separated the lacrimalfrom the zygomatic. Fragments of the anterior crest delimitingthe lacrimal fossa and a small part of the medial wall of thelacrimal fossa are preserved bilaterally (Fig. 7B). CT crosssections show parts of the nasolacrimal ducts bilaterally aswell. From this, it appears that the lacrimal fossa is situatedin the margin of the orbit. However, it is not clear whetherthe anterior lacrimal crest and anterior part of the lacrimalfossa are composed of the maxillary bone or are part of thelacrimal bone extending onto the face. In Tremacebus, the an-terior crest of the lacrimal fossa is composed of the lacrimalbone. In Homunculus (Cordoba skull), the lacrimal bone ex-tends onto the face anterior to the orbital margin and contactsthe frontal, thereby separating the maxilla from the frontal.Extension of the lacrimal onto the face anterior to the orbitis seen also to varying degrees in Callicebus and atelids.
Postorbital closure is extensive on the right side of the Doli-chocebus skull (Figs. 7B and 8A), but it is impossible to discernthe precise contributions made by individual bones. Neverthe-less, the closure was similar to that of most platyrrhines (e.g.,
Callimico; Fig. 8B) and much more than in Aotus, where a lateralorbital fissure extends laterally to the root of the zygomatic arch(Fig. 8C). Homunculus has an extensive postorbital septum(Tauber, 1991). Although Hershkovitz (1974) claimed thatTremacebus possessed a large infraorbital fissure, our observa-tions on the original specimen in Tucuman indicate that thesize of the opening in the orbital apex is largely due to postmor-tem breakage (see also Fleagle and Rosenberger, 1983).
As in all anthropoids, the orbits are convergent and frontatedand closely approximated beneath the olfactory bulbs. The ex-act amount of convergence is uncertain because the orbitalmargins are incompletely preserved. However, comparison ofthe rostra of Dolichocebus and Saimiri, when distortion is ac-counted for, show that the former is considerably more prog-nathic and that its right ventral orbital margin is oriented lesstransversely (i.e., less convergently) than in Saimiri (Fig. 1,dorsal view, and Fig. 9B). Orbital convergence is notably lessin extant callitrichines (range 58�e63�) than in other livingplatyrrhines (range 66�e75�) or catarrhines (range 70�e82�)(Ross, 1993), and convergence in Dolichocebus was primitive,and that of extant callitrichines is likely to have evolved inde-pendently. Tremacebus also is comparable to callitrichinesand has less orbital convergence than other extant platyrrhinesof similar size (see Fig. 7 in Hershkovitz, 1974). Callitrichine-like less convergent orbits also characterize late Eocene and earlyOligocene African anthropoids Apidium, Simonsius, Proteopithe-cus, and Catopithecus (Simons, 2004), suggesting that the extentof orbital convergence in Dolichocebus and Tremacebus is pri-mitive, and that of extant callitrichines is likely to have evolvedindependently. Aegyptopithecus, at 72� or greater (Ross, 1995;Simons, 2004), had a catarrhine-like level of orbital convergence.
The interorbital region of Dolichocebus appears to be narrow(interorbital breadth, 5.7 mmdalthough we caution that this isa minimum estimate because the skull has been crushed medio-laterally) (Rosenberger, 1979a). In Dolichocebus, the interor-bital region is most notable for a large opening in the matrixconnecting the orbits (Figs. 7, 9B). Coronal interorbital sectionsof Saimiri (Fig. 10A) show that the edges of the interorbital fe-nestra are bounded by a single lamina of bone both dorsally andventrally. In Saguinus, an interorbital septum is composed oftwo laminae of bone; further ventrally in Saguinus, the vomeris sandwiched between the right and left ventromedial surfacesof the orbit (Fig. 10B). The CT images show that the area imme-diately surrounding the fenestra of Dolichocebus is composedmostly of matrix. Preserved interorbital bone ventral to the inter-orbital opening in Dolichocebus (Fig. 11) shows a right and leftparanasal sinus, most probably the sphenoidal (cupular) sinusabove the nasal cavity, and below the interorbital septum. How-ever, the right and left orbital walls on either side of the sinusconverge dorsally, suggesting that the interorbital septum in Do-lichocebus was probably not wide as in Callicebus, but narrow asin Cebus or Saguinus (Fig. 20b,c in Rossie, 2006). Thus, while itseems likely that Dolichocebus had a narrow interorbital sep-tum, the CT scans neither confirm nor disprove the presenceof an interorbital fenestra.
Nuchal region. As in all anthropoids, Dolichocebus hasa pneumatized mastoid bone. The portion of the mastoid

Optic canal
Superior orbitalfissure
Inferior orbitalfissure
Optic canal
Inferior orbitalfissure
Superior orbitalfissure
A
B
C
Fig. 8. Views of the right orbits of platyrrhines showing the optic canal, superior
orbital fissure, and inferior orbital fissure: (A) Dolichocebus gaimanensis, (B)
Callimico goeldii, and (C) Aotus trivirgatus. Note that the postorbital flange of
the zygomatic (indicated by the arrows) is more extensive in Callimico than in
Aotus. The postorbital flange of the zygomatic in Dolichocebus is partially pre-
served. Its extent is also documented in Fig. 7B. The flange of Dolichocebus
more closely resembles that of Callimico in its preserved parts.
3 We measured the length of the postglenoid process in small samples of 15
genera of extant platyrrhines. Relative size of the process is quantified by the
ratio 100� (postglenoid length/prosthioneinion length). The ratio for Doli-
chocebus was estimated to be 242. Four genera and 16 specimens of callitri-
chines range from 67 to 243, with Leontopithecus at the upper end of the
range. Three specimens of Saimiri range from 195 to 282. Atelids have
much larger postglenoid processes: nine specimens representing four genera
range from 525 to 1240.
334 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
closest to the occipital protuberances is flattened as in Trema-cebus, Homunculus, and cebids. In contrast, pitheciids (Rose-nberger, 1979b) and atelids have small paraoccipital processes.Inion of Dolichocebus is placed dorsally relative to the fora-men magnum. Thus, the nuchal plane forms a relatively obtuseangle with the Frankfort horizontal (Fig. 12A). This is typicalof most living platyrrhines (Fig. 12B) (Hershkovitz, 1977), as
well as Tremacebus and Homunculus, but differs from the con-dition in Saimiri, in which the nuchal plane forms a muchmore acute angle with the Frankfurt horizontal (Fig. 12C)(Hershkovitz, 1977).
Zygomatic region. The zygomatic arches are broken awaybilaterally, with only a sliver of the posterior root preservedon the right side. It is impossible to tell from this remnantwhether the entire arch was lightly constructed, as in Saimiriand many callitrichines, or robust, as in, for example, Callice-bus (Fig. 12B, C).
Lateral wall of the braincase. The sutures in the region ofpterion are obscured by breakage, and it is impossible to tellwhether the frontal and alisphenoid are in contact (as in livingcatarrhines) or if the zygomatic and parietal are in contact (asis typical of platyrrhines, with the exception of Alouatta andsome other atelids, in which it is variable) (Ashley-Montague,1933; Mouri, 1988). Notably, the pterionic region in bothTremacebus and Homunculus (Tauber, 1991) shows the ‘‘cat-arrhine’’ condition. Although the lateral wall of the braincaseis also broken in many places, the remnants on the right sidesuggest that Dolichocebus did not possess a temporal emissaryforamen, as reported for atelids (Conroy, 1981; Horovitz andMacPhee, 1999). Neither Tremacebus nor Homunculus hasa temporal emissary foramen.
On the lateral wall of the braincase at the anteroposteriorlevel of the external auditory meatus, a faint right temporalline is present (Fig. 13). This line does not reach the midlineof the skull, so there is no sagittal crest, but, as also notedby Rosenberger (1979a), the temporal lines are more closelyapproximated to the midline of the cranial vault than in Sai-miri. Rosenberger (1979a) suggested the closer approximationof the temporal lines in Dolichocebus to be a consequence ofits larger, more prognathic face and larger teeth than Saimiri.An alternative possibility that needs further investigation isthat Dolichocebus had a smaller brain and braincase, leavingless bony surface for the temporalis muscle to attach. Trema-cebus has the same morphology as Dolichocebus.
Temporomandibular region. The surface of the mandibularfossa, preserved on the right side of the skull, is broad and flat,with no indication of an articular eminence (Fig. 14). The rightpostglenoid process is slightly damaged, but its length can be re-liably estimated as ~1.65 mm. This is relatively very small,within the range of extant Saimiri and callitrichines.3 Tremace-bus likewise has a very small postglenoid process, whereas theprocess is large in Homunculus.
The right postglenoid foramen of Dolichocebus is situatedposterolateral to the postglenoid process and is quite large(Fig. 14)dapproximately 1.8 mm in diameterdindicating

incompletely ossifiedinterorbital septum
?
A
B
more prognathicfacial region
Fig. 9. Anterolateral views of the rostra of (A) Saimiri and (B) Dolichocebus.
The incompletely ossified interorbital septum of Saimiri is mimicked in matrix
by Dolichocebus. Note also the more prognathic face of Dolichocebus.
Fig. 10. Coronal CT sections of the interorbital region: (A) Saimiri sciureus
and (B) Saguinus fuscicollis.
335R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
that a large intracranial venous drainage channel (the petros-quamous sinus) emerged at this point (Saban, 1963; Conroy,1980; Kay et al., 2008). The foramen is smaller in similarlysized Tremacebus (~0.74 mm). Homunculus resembles Doli-chocebus in having a relatively large foramen.
Pterygoid and palatal region. The pterygoid fossa, ptery-goid plates, and the pyramidal processes are broken away. Dis-tally, the tooth rows of Dolichocebus diverge only slightly: theratio of the distance between the lingual sides of the upper ca-nines (9.49 mm) to the distance between the lingual sides ofthe M2s (11.44 mm) is 83%. These proportions of Dolichoce-bus are similar to those of Tremacebus (Hershkovitz, 1974).Eocene/Oligocene anthropoids and many extant catarrhineshave similarly nondivergent tooth rows, whereas many crownplatyrrhines have postcanine teeth that diverge more markedly.
The remnants of the posterior parts of the palatal processesof the palatine bone are slightly thickened to form a weak
posterior palatine torus. We score this feature as present in Do-lichocebus in our character-taxon matrix while recognizingthat its development is far less robust than in some nonanthro-poid primates like Adapis (Ross, 1994).
Facial region. The depth of the maxilla of Dolichocebus iscomparable to that in most anthropoids. The distance from theleft orbital margin to the anterior edge of the canine alveolusis 10.8 mm (preorbital rostrum length; [Ross, 1994]), and the

olfactory fossa andmedial orbital wall
maxillary sinus
nasal cavity
medial orbital wallsand vomer
anteriorcranial fossa
nasal septum
dorsal roofof nasal cavity
Anterior ethmoidor 'sphenoid' sinus
Fig. 11. Coronal CT sections of the interorbital region of Dolichocebus illustrating the maxillary sinus, the nasal cavity, and a large sinus, probably the cupular
sinus, dorsal to the nasal cavity.
336 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
distance from the anterior edge of the canine alveolus to the na-somaxillary suture (maxillary depth [Ross, 1994]) is >9.5 mm.Interpolating these data onto a log-log bivariate plot of preorbi-tal rostrum length versus maxillary depth for 11 extant anthro-poids and 19 extant strepsirrhines (Fig. 17 in Ross, 1994),Dolichocebus fits within the anthropoid cluster (short rostrumand deep maxilla compared with strepsirrhines).
As in other anthropoids, the snout of Dolichocebus is short.With the skull oriented in the Frankfurt horizontal, the left in-fraorbital foramen is located quite far anteriorly on the face,above a point between P2 and P3 (Fig. 12A).
Ear region and braincase. The petrosals are well preservedon both the right and left sides. The right middle ear cavity andits anterior accessory chamber are exposed (Fig. 14). As inmost platyrrhines, excepting atelids and Cacajao (Horovitz,1997), Dolichocebus possesses two prominences on the lateralsurface of the promontorium (as does Tremacebus). These
prominences are the external manifestations of the cochleaon the promontory surface of the middle ear. The dual prom-inences may be a derived feature of crown platyrrhines subse-quently lost in atelid platyrrhines. This is supported by theobservation that Tarsius, Aegyptopithecus, and Simonsiushave a singular prominence (Kay et al., 2008). However, thereare paired prominences on the cochlear housing in the middleear in Apidium (see Fig. 1 in Cartmill et al., 1981). Moreoverwe have observed variation in this feature in extant Saimiri,Cebus, and Aotus.
As in all haplorhines, a transverse septum separates thetympanic cavity proper from a well-developed anterior acces-sory chamber (AAC) (Cartmill and Kay, 1978; MacPhee andCartmill, 1986). The latter develops as a diverticulum fromthe auditory tube (Ross, 1994). The AAC extends more medi-ally and has trabeculae within it, as in other anthropoids (Ross,1994). On the right side of the cranium, a transverse septum is

postorbital closure
angle of nuchalplane
infraorbitalforamen
postorbital closure
infraorbitalforamen
angle ofnuchal plane
angle of nuchalplane
postorbital closure
infraorbitalforamen
A
B
C
Fig. 12. Views of skulls showing angle of the nuchal plane, root of the zygo-
matic arch, and position of the infraorbital foramen: (A) Dolichocebus gaima-
nensis, (B) Callicebus torquatus, and (C) Saimiri sciureus.
Fig. 13. Dorsal view of skull of Dolichocebus gaimanensis showing the posi-
tion of the temporal lines.
337R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
present, as is an anterior accessory chamber. The right petrosalof the specimen is broken ventrally, exposing the middle earcavity, the auditory tube, and the AAC. The AAC extends me-dially and posteriorly onto the promontorium. It appears to belargely devoid of trabeculation dorsally, but is trabeculated lat-erally and ventrally. Medial extension of the anterior accessorychamber is a derived feature of crown and (as far as is known)stem anthropoids. It is a feature of Aegyptopithecus, Apidium,
and Simonsius (¼ Parapithecus), as well as Proteopithecusand Catopithecus (Kay et al., 2008).
The posterior carotid foramen of Dolichocebus (markingthe entrance into the bulla of a large internal carotid artery)is identifiable on the petrosal. Inside the bulla, a ridge onthe ventrolateral surface of the promontorium marks the pres-ence of a bony tube for the internal carotid (promontory) ar-tery (Fig. 14), the presence of which is confirmed in CTscans (Fig. 15B). As in other platyrrhines, the posterior carotidforamen is located posterior to a line joining the midpoints ofthe ectotympanic elements, and medial to the midline of theauditory bulla, here established as a line joining the stylomas-toid foramen and the anteromedial-most point on the petrosal,but far forward of the stylomastoid foramen. The same posi-tion appears to hold for Proteopithecus, Catopithecus, and par-apithecids. In catarrhines and Aegyptopithecus, it is alsopositioned posteriorly but more nearly in the middle of thisline (Kay et al., 2008).
The ectotympanic element is completely visible on theright (Fig. 14), while only a dorsal remnant is preserved onthe left. As is the case among all known Fayum anthropoidsand platyrrhines (Kay et al., 2008), the ectotympanic is extra-bullar, forming a bony ring at the entrance to the tympaniccavity. The ring is ossified outward to form a tube in catar-rhines (only partially so in Pliopithecus) (Szalay and Delson,1979; Fleagle and Kay, 1987) and Tarsius.
On CT images of the skull of Dolichocebus, a narrow canalcan be seen connecting the subarcuate fossa with the sulcus forthe sigmoid sinus (Fig. 16). This is Cartmill’s canal, a venouschannel (Cartmill et al., 1981; Kay et al., 2008) present in allplatyrrhines, including Tremacebus. It is absent in Tarsius andcatarrhines and absent in the Oligocene parapithecids Apidium(Cartmill et al., 1981) and Simonsius (Kay et al., 2008). Thecanal is partially obliterated in the stem catarrhine Aegyptopi-thecus, but is present and well developed in early stem anthro-poids such as Proteopithecus and Catopithecus (Kay et al.,2008).
On coronal CT slices through the braincase, the remnantsof an ossified tentorium cerebelli are seen bilaterally abovethe subarcuate fossa (Fig. 15). Horovitz and MacPhee (Mac-Phee et al., 1995; Horovitz, 1999; Horovitz and MacPhee,

for. magnum
occipital condyle ant. accessorychamber
postglenoidfor. &process
transpromontorialcarotid canal
stylomastoidfor.
dual promomtorialprocesses
jugular for.
basioccipital stem
carotid for.
Fig. 14. Stereopair of right basicranial region of Dolichocebus gaimanensis.
A
338 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
1999) stated that the tentorium is absent in Tarsius and (some-times absent in Saimiri) but present in all other platyrrhines.An ossified tentorium is also absent in Oligocene Fayum an-thropoids Aegyptopithecus and Apidium (Kay et al., 2008).In contrast, Hershkovitz (1977) noted that it is most extensivein Ateles, Lagothrix, and Brachyteles; peripheral in Callicebus,Aotus, and pitheciines; variable in Alouatta; minimal in Sai-miri, Cebus, and Callimico; and absent or rudimentary in othercallitrichines. Our observations on additional specimens sup-port Hershkovitz’s observations for the most part. However,CT images show the tentorium to be quite well developed inCallimico.
petrosquamous
Descriptive anatomy of the dentition sinuscarotid canal
trabeculated anterioracessory chamber
cochlea
middle ear
carotid canal
inferior petrosal sinus
subarcuatefossa
tentoriumcerebelli
B
Fig. 15. Two coronal CT sections of the skull of Dolichocebus gaimanensis;
the upper image (A) is more posterior than the lower image (B). Features iden-
tified include the carotid canal crossing the promontorium, trabeculation in the
anterior accessory chamber, the petrosquamous sinus (venous drainage
through postglenoid foramen [not shown]), the subarcuate fossa, and a partially
ossified tentorium cerebelli.
Lower incisors (MACN CH 872, a right I1 or I2 [not fig-ured]). The crown of MACN CH 872 is broken a short dis-tance above the cervix. The tooth is spatulate. The root hasa broadly oval cross section, not mesiodistally compressedas in Cebuella and Callithrix. The lingual aspect of the crownhas enamel (in Callithrix, the lingual enamel is extremely thinor absent (Rosenberger, 1978)) and a well-developed lingualheel (unlike in Cebuella and Callithrix or pitheciines, in whichthe lingual face is flattened). Mesial and distal lingual crestsare present, but they do not join lingually in the midline. Com-pared to the lower first molar described below, it is a verysmall toothdthe area ratio is 0.27, comparable to ratios inAlouatta, Brachyteles, Callimico, and Callicebus. Among earlyMiocene platyrrhines, Homunculus has similarly small inci-sors (Hershkovitz, 1970), but those of Soriacebus are enlarged(Fleagle et al., 1987; Fleagle, 1990). In all observable featuresof structure and proportions enumerated in our character-taxonmatrix, the structure of the lower incisor of Dolichocebus ismost similar to that of Callimico.
Lower canine (MACN CH 870, a right canine; MACN CH871, a left canine [Fig. 17A,B]). Both of these teeth have
well-preserved crowns, although the distal heel of MACNCH 870 is partly broken away. Specimen MACN CH 870 isa larger tooth with a broadly oval cross section, whereasMACH CH 871 is smaller and slightly more compressed.However, neither tooth is as mesiodistally compressed as in

1156
jugular foramen
Cartmill'scanal
1141
1122
subarcuatefossa
Cartmill'scanal
mastoidair sinus
sigmoid sinussemicircularcanal
Cartmill'scanal
sigmoid sinus
A
B
C
Fig. 16. Three coronal CT sections of the skull of Dolichocebus gaimanensis; the upper image (A) is more posterior than the middle image (B), which is
more posterior than the lower image (C). Features illustrated include mastoid air sinuses and the posterior end of the subarcuate fossa. Cartmill’s canal, a vascular
channel, leads transversely to open into the sigmoid sinus near the junction with the jugular foramen.
339R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Cebuella or Callithrix. The size difference between the twospecimens could be due to sexual dimorphism. If so, the de-gree of dimorphism would have been considerable, withinthe range seen in Saimiri. Canine dimorphism is either un-known or has not been examined among other early Mioceneplatyrrhines. On both teeth, the lingual crest is rounded, notsharp, like pitheciines. A raised and unbroken lingual cingu-lum runs from the large distal heel to the mesial terminus ofthe paracristid. The ratios of canine size (area) to the size ofthe only M1 in the sample are 0.64 and 0.83 for the two spec-imens. Although the specimens come from different individ-uals and the ratio is not necessarily reliable, lower caninesize seems to be comparable to that of Aotus or Callicebus
in our sample of extant taxa and smaller than in Saimiri, inwhich the full range of ratios for a sample of 44 specimensis 0.79 to 1.39. Among early Miocene platyrrhines, the caninesize of Dolichocebus is similar to that of Carlocebus and Ho-munculus, but smaller than in Soriacebus.
Lower premolars (MACN CH 865, P2; MACN CH 868a, rightP3 [Fig. 17C]; MACN CH 868b, right dp3; MACN CH 873, leftP3 [Fig. 17D]; and MACN CH 898, left P4). Specimen MACNCH 865 is tentatively identified as a P2 because of the elongateparacristid, the buccal swelling at the base of the protoconid, andthe simplicity and narrowness of the talonid. If Dolichocebuswere sexually dimorphic, as seems likely from the canines,this tooth would be that of a female. The tooth has a single

340 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
root, as in all living and extinct platyrrhines (including Carloce-bus, Homunculus, and Soriacebus) and EoceneeOligocene Pro-teopithecus and parapithecids. Catarrhines, propliopithecids,and oligopithecids have lost the P2. It has an oval occlusal out-line with a slight buccal flare at the base of the protoconid. Abuccal cingulum is absent, as in Proteopithecus (late Eocene,Africa), catarrhines, Homunculus, Soriacebus, and most extantplatyrrhines. The trigonid has a protoconid but lacks a metaconidand paraconid. A P2 metaconid occurs variably in Eocene/Oli-gocene Fayum anthropoids. A small metaconid appears variablyin Homunculus but is not present in either Carlocebus or Soria-cebus. The protoconid appears not to have projected above theprotoconids of either P3 or P4, as it does in Saguinus and Calli-mico. As in all known platyrrhines, the talonid is small and lacksa hypoconid and entoconid.
Two teeth in our sample, MACN CH 873 and MACN CH868a represent the P3. Each tooth has a single root (as in theP3 and P4 of all other living and fossil platyrrhines but notin any catarrhine or stem anthropoid). The crown is mesiodis-tally short and buccolingually broad. There is a prominent pro-toconid and a smaller metaconid distolingually. The small sizeof the metaconid and its close proximity to the protoconid re-semble the condition in most living callitrichines (but notCallimico). Other extant platyrrhines have a larger metaconidwith greater separation from the protoconid. A small P3 meta-conid also characterizes late Oligocene Branisella and earlyMiocene Carlocebus, Homunculus, and Carlocebus. The disto-lingual position of the cusp is also seen in other early Mioceneplatyrrhines. No paraconid is present (see comments belowconcerning paraconid loss on P4). The trigonid is open lin-gually, unlike that of any extant platyrrhine. The talonid hasa short cristid obliqua and a very small hypoconid positioneddistolingual to the protoconid. No hypocristid is present. Thelingual marginal crest of the talonid is weak and lacks a dis-crete entoconid. The buccal surface of the crown of MACNCH 873 has a poorly developed cingulum, whereas there isno buccal cingulum on MACN CH 868a. The crown is slightlyswollen buccal to the protoconid, although less so than in P2.
Specimen MACN CH 898 is identified as a P4 because of thegreater structural complexity of the root and crown comparedwith the aforementioned premolars. This specimen is single-rooted, but the root is deeply grooved. The crown is mesiodis-tally short and buccolingually broad. The trigonid and talonidare approximately equal in mesiodistal dimensions. The trigo-nid supports a widely spaced protoconid and metaconid; the lat-ter is smaller than the protoconid and placed slightly distal to it.There is no paraconid, which is a resemblance to all living andextinct platyrrhines (except Carlocebus) and catarrhines.4 Thetrigonid is widely open lingually, as in some Homunculus,whereas it is closed in Soriacebus, Carlocebus, and Branisella;P4 trigonids in extant platyrrhines are closed. The talonid isbroad buccolingually with a small hypoconid buccally. No
4 Catopithecus has a paraconid, and thus if oligopithecids are stem catar-
rhines, as advocated by many, then paraconid reduction would have occurred
in parallel in platyrrhines and catarrhines.
hypocristid is present. A lingual marginal crest supports a smallentoconid. There are several obvious distinctions from Soriace-bus, in which P4s are much narrower buccolingually, with themetaconid much closer to the protoconid.
Specimen MACN CH 868b is probably a dp3, a conclusionwe reach for two reasons: (1) this tooth has similar proportionsand shape to the dp3 of Aotus (although it is very differentfrom that taxon in the details of the talonid); and (2) the colorof the enamel on the tooth is distinctly lighter than in the otherspecimens, a phenomenon frequently, but not always, seen inthe deciduous teeth of fossil mammals (Kay and Simons,1983). This tooth is single-rooted and has an elongate oval oc-clusal outline; it is not buccolingually broad, as in MACN CH898, nor is there any buccal swelling mesiobuccally. The trig-onid is relatively short mesiodistally. A paraconid is absent.The metaconid is larger and is spaced further from, andmore directly lingual to, the protoconid. The trigonid is openlingually. The talonid is buccolingually broad with a well-developed hypoconid and a small but distinct entoconid. Thereis no buccal cingulum. There is very little similarity in shapebetween this tooth and the P4 of Soriacebus. The basins ofMACN CH 868b are more squared off, the talonids are broader,the metaconid is bigger and further from the protoconid, and thetalonid is more cuspidate.
Both P3 and P4 have transversely (buccolingually) con-stricted crowns with broadly sloping buccal faces. This is char-acteristic of early Miocene Carlocebus, Homunculus, andSoriacebus. Branisella and most living platyrrhines havemore marginally placed cusps (excepting some callitrichines,Cebus, and Ateles).
Lower molars (MACN CH 866, left M2 [Fig. 17E]; MPEF5147 mandible fragment with right M1e3; MPEF 5146 mandiblefragment with left M1e2 [Fig. 17F,G]). In the collections madebefore 1990, only one poorly preserved and heavily worn lowermolar (MACN CH 866) was identified. The tooth has two rootsbut these are fused over two-thirds of their length. The mesialroot is larger than the distal. The distal root projects posteroin-feriorly from the crown. The distal crown margin is deeplynotched by interproximal wear. The form and structure of thiswear facet shows that the posteriorly adjacent M3 was displacedlaterally. A very weak buccal cingulum is visible in the hypo-flexid. The trigonid and talonid are approximately equal inbreadth. The cristid obliqua is obliquely oriented so that thehypoflexid is moderately incised.
Two mandibular fragments contain molars. SpecimenMPEF 5147 preserves the right M1e3, and MPEF 5146 hasleft M1e2. Neither specimen preserves sufficient parts of themandible to gain an impression as to its depth or morphology.
The occlusal surfaces of MPEF 5147 were broken postmor-tem, with only the distolingual corner of M1, the entire M2,and the buccal half of M3 remaining. Occlusal wear in lifeobliterated the details of the crown morphology. However,the crown proportions can be estimated and root structure dis-cerned. On MPEF 5146, M1 and M2, each with two roots, arewell preserved and the dentin is exposed only at the cusp tips.An interproximal wear facet at the distal margin of M2 indi-cates that three molars were present. The M1 and M2 of

Fig. 17. Lower teeth of Dolichocebus gaimanensis: (A) MACN CH 870, a right canine, in lingual view (mesial is to the left); (B) MACN CH 871, a left canine, in
lingual view (mesial is to the right); (C) MACN CH 868a, a right P3, in occlusolingual view (mesial is to the left); (D) MACN CH 873, a left P3, photographically
reversed for comparison, in occlusolingual view (mesial is to the left); (E) MACN CH 866, a left M2, in occlusobuccal view (mesial is to the left); (F) MPEF 5146,
a mandible fragment of Dolichocebus with left M1e2, in occlusolateral view; (G) MPEF 5146 in occlusal view (mesial is to the left). Scale bars¼ 3 mm. AeE are
at the same scale; F and G are at the same scale.
341R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
both specimens possess two widely separated roots; M3 ofMPEF 5147 has two widely separated roots. The two-rootedcondition is characteristic also of Branisella, Soriacebus, Car-locebus, the unpublished Colhuehuapian species from GranBarranca, and Homunculus, as well as all taxa of African Eo-cene/Oligocene anthropoids. In contrast, all living platyrrhinesexcept Alouatta (and with a small number of exceptional spec-imens of some other species) have single-rooted M3s.
For details of the crown anatomy of the lower first molar,we must rely entirely on MPEF 5146. The M1 crown is‘‘squared,’’ having a low ratio of mesiodistal length to bucco-lingual breadth (Table 3). Low cusp relief and smooth enamelcharacterize the crown. The trigonid is mesiodistally short,constituting about 35% of the mesiodistal length. The overallbreadth of the trigonid exceeds that of the taloniddthe crownbulges buccally and lingually (to a lesser degree) such that the
ratio of protoconid-metaconid breadth to trigonid breadth isonly 50%. Thus, the actual occlusal surface is quite com-pressed and the sides of the marginal cusps bulge, a featureDolichocebus shares with Soriacebus and a few extant platyr-rhines (Cebus and Ateles). Likewise, the occlusal surface ofthe talonid crown is compressed and the sides of the toothbulge. The trigonid is three-cusped, with a small paraconidon the mesiolingual edge of the trigonid. The metaconid is po-sitioned distolingual to the protoconid. A lower first molar par-aconid is unusual for a platyrrhine. We note its presence onlyin some specimens of Homunculus and in an undescribedtaxon of Colhuehuapian monkey from Gran Barranca (Kayet al., 1999b). The paracristid is short and mesiodistally ori-ented. A premetacristid encloses the trigonid lingually. Lin-gual and buccal protocristids form a complete distal trigonidwall. Wear surface ‘‘X’’dfound on the distal trigonid of

342 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
some parapithecids and (homoplastically) in catarrhines (Kay,1977)dis absent. The trigonid and talonid are essentially iden-tical in height and breadth. This is typical of platyrrhines gener-ally, with the exception of Soriacebus, in which the trigonid isextremely elongate. The entoconid is large and positioned disto-lingual to the hypoconid. In contrast, Soriacebus has a very re-duced entoconid. The reduced condition is also seen in manycallitrichines and some advanced pitheciines (but not reducedin Pithecia). The distal edge of the talonid supports a raised ridgeconfluent with the hypocristid and adorned with two bulges, oneof which is in the midline of the tooth and the other at its lingualterminus. The latter is interpreted to be a hypoconulid, placedlingual to the midline of the tooth. A larger, more linguallyplaced hypoconulid is present in Branisella. The small, mediallyor lingually placed cusp also appears in Carlocebus, Soriacebus,and, variably, in Homunculus, whereas it is an unusual feature inliving platyrrhines.
Distolingually, the hypoconulid is separated from the ento-conid by a deep sulcus (groove) and there is a small postero-lingual fovea. The presence of a sulcus is unusual in extantplatyrrhines, occurring in some specimens of Callicebus andAlouatta. It is prominently or weakly displayed in Branisella,Homunculus, Carlocebus, and Soriacebus. The posterolingualbasin is variably present in Callicebus alone among extant plat-yrrhines, and also occurs in early Miocene Carlocebus. Thecristid obliqua connects with the trigonid wall at a point disto-lingual to the protoconid and does not extend up the trigonidwall to the protocristid. The lingual margin of the talonid basinhas a distinct notch where the postmetacristid abuts the pre-entocristid. The presence of a lingual talonid notch is unusualin living platyrrhines, appearing only in Callicebus and somespecimens of Alouatta. The notch is present in Branisella andalso in the undescribed Gran Barranca platyrrhine. A strongbuccal cingulum wraps around the protoconid and spans thehypoflexid, but disappears lingual to the hypoconid. Similarcingulum development is found in the Gran Barranca primate(but not in other early Miocene platyrrhines or Branisella)and in Saimiri and Saguinus among extant platyrrhines. Thecingulum is notched in the hypoflexid, where a strong cuspuleis present. Wear facets on the distolingual corner of M1 and con-tinuing onto the trigonid of M2 indicate that M1 had a hypocone.
The description of M2 is based on MPEF 5146 (Fig. 17G,F).The morphology of the tooth is similar to that of the M1. The M2
crown, short mesiodistally and broad buccolingually (Table 3),has low cusp relief and smooth enamel. The trigonid is short,constituting about 33% of the mesiodistal length. The occlusalsurface of the trigonid is quite compressed, with the crown bulg-ing buccally and lingually such that the ratio of protoconid-metaconid breadth to trigonid breadth is 52%. The trigonid istwo-cusped; unlike on M1, there is no paraconid. The metaconidis positioned more lingually and not as much distally to the pro-toconid compared with the configuration on M1. Other aspectsof the trigonid of M2 resemble those of M1. Wear surface ‘‘X’’on the distal trigonid is absent. The trigonid and talonid are es-sentially identical in height but the trigonid is wider than the ta-lonid (versus similar breadths on M1). The entoconid is large andpositioned more nearly lingually to the hypoconid than on M1.
The configuration of the distal margin of the talonid is very sim-ilar to that described on the M1. Distolingually, the hypoconulidis separated from the entoconid by a deep sulcus. The cristid ob-liqua is oriented more mesiodistally than in M1 and reaches theprotoconid, leaving the hypoflexid more shallow than on M1. Aswith M1, the lingual margin of the talonid basin is closed butnotched. The buccal cingulum is much weaker than on M1 andspans only the hypoflexid, disappearing lingual to the protoco-nid and hypoconid. The cingulum is notched at several points,but no discrete cuspules are present. As with M1, wear facetson the distolingual corner of M2 give a clear indication thatM2 had a hypocone.
The M1 is greater in overall size (i.e., area) than M2, but inlength, M1 and M2 are similar. From the broken remnants ofMPEF 5146, it appears that M3 was slightly shorter mesiodis-tally than M2, and substantially narrower.
Upper incisors (MACN CH 359, a left I1 [Fig. 18A]). Noupper incisors from late Oligoceneeearly Miocene platyr-rhines have been reported previously, so our comparisons arerestricted to extant platyrrhines. The I1 has a rounded, ovaloutline in occlusal view. A complete lingual cingulum is pres-ent. Although somewhat obscured by wear, it appears thata basal lingual cusp was absent. If so, this absence is a resem-blance to callitrichines, Callimico, Aotus, and Saimiri.
Upper canine (MACN CH 361, a right canine [Fig. 18B];MACN CH 1302, a left canine [Fig. 18C]; MACN CH 1303,a right canine). The occlusal outline of MACN CH 361 isa tear-drop-shaped oval. The crown is tall and slender. Adeep mesial groove is present on the mesial aspect extendingonto the root, a condition seen on Branisella and all early Mio-cene platyrrhines. There is a complete lingual cingulum, but itis a narrow shelf, not bulging as in some living platyrrhinessuch as Ateles. The shelf is similarly proportioned in Dolicho-cebus, Branisella, Carlocebus, and Homunculus. The Soriace-bus canine has a weaker lingual cingulum. Specimens MACNCH 1302 and 1303 have a similar morphology to the tooth justdescribed, except that the crowns are less projecting andsmaller overall. These two sizes and shapes suggest that Doli-chocebus canines were sexually dimorphic.
Upper premolars (MACN CH 1012, a left P2; MACN CH864, a right P3; MACN CH 878 [Fig. 18D], a right P4;MACN CH 1011, a right dp4). The P2 is single-rooted as in-Branisella, Tremacebus, and most other platyrrhines (exceptSoriacebus and, among extant platyrrhines, Aotus, Alouatta,and Callicebus, which have two-rooted P2s). In buccal viewthe tooth is lancelet-shaped and bulges at the cervix. The lin-gual cingulum is well developed and was probably complete,but lacks a protocone. As in all extant platyrrhines, crests runfrom the paracone to the mesial and distal borders of thecrown. These are asymmetric, with the mesial crest shorterthan the distal.
The P3 (MACN CH 864) is two-rooted. In addition to thesingle lingual root, there appears to be a single buccal rootthat is broken close to the cervix. The occlusal outline isnearly oval, but broader buccally than lingually. A lingualswelling is present, here interpreted as a small protocone.Strikingly, like Branisella but unlike other platyrrhines, the

Fig. 18. Upper incisors, canines, and premolars of Dolichocebus gaimanensis:
(A) MACN CH 359, a left I1, in occlusolingual view; (B) MACN CH 1302,
left canine, in occlusolingual view; (C) MACN CH 361, right canine, in occlu-
solingual view; (D) MACN CH 878, a right P4, in occlusal view (mesial is to
the left). Scale bar ¼ 3 mm.
343R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
protocone is situated on the mesial margin of the crown. Asmall hypocone is present. The distal profile of the tooth issmoothly convex, not concave (‘‘waisted’’) as in Callithrixand Cebuella. The P2 occlusal area is much less than P3 area.
The P4 (MACN CH 878) is also two-rooted, nearly oval inocclusal outline, with a convex distal profile. It has a strongprotocone. A large hypocone is present on a very strong lin-gual cingulum. The large hypocone and pronounced cingulumare features shared with Soriacebus, Dolichocebus, Homuncu-lus, and Carlocebus; the cusp is smaller or absent in extantplatyrrhines. The cingulum begins at the distal crown marginand wraps completely around the lingual base of the proto-cone. This configuration resembles that of Callicebus, whichhas a very strong lingual cingulum and a smaller hypocone,although in some Callicebus the hypocone is often lingual
to the cingulum rather than connected with it. The tooth hasa small parastyle and metastyle but lacks conules. The post-protocrista is weak and reaches toward the base of the hypo-cone. Such a short, weak postprotocrista is characteristic of allOligoceneeearly Miocene platyrrhines but is not seen in extantplatyrrhines, with the exception of Callithrix and Cebuella). Inextant platyrrhines, the crest is strong and reaches distolinguallyto the distal margin of the crown. A groove is present betweenprotocone and hypocone. This morphology resembles that ofCarlocebus and Soriacebus (and Callicebus, although thegroove between the protocone and hypocone is shallower thanin the latter taxa). The P4 is small compared to M1; the ratioof crown areas is 0.59, resembling Oligocene/Miocene Brani-sella, Carlocebus, and Homunculus, as well as Callimico andmarmosets among extant platyrrhines. This condition is dis-tinctly different from that in Saimiri, in which P4 is enlarged.
The dp4 (MACN CH 1011) is a three-rooted tooth. The lin-gual root is splayed lingually, which suggests that this is a de-ciduous tooth. The buccal tooth roots are broken off. There isa strong preprotocrista. The hypocone is small and distolingualto the protocone. A strong prehypocrista encloses the talon ba-sin lingually. A very small swelling on the preprotocrista maybe a paraconule; there is no trace of a metaconule. The para-style and mesostyle are very small and associated with a veryweak buccal cingulum. The strong lingual cingulum, leadingfrom the small hypocone, wraps part way around the proto-cone before disappearing mesiolingual to that cusp. A postpro-tocrista is strong and joins the prehypocrista. A lateraltransverse crista is continuous with the postprotocrista.
Upper molars (MACN CH 876, a left M1; MACN CH 356,a left M2 [Fig. 19C,D,E]; MACN CH 357, a right M3
[Fig. 19A]). In extant platyrrhines, the metacone is smallerthan the paracone on M2, whereas they are similar in size onM1. On this basis, MACN CH 876 is here identified as anM1 and MACN CH 357 as an M2. The M1 and M2 arethree-rooted. The M1 is heavily worn and many of the detailsof its crown morphology are obliterated. Most of the details ofocclusal anatomy will be based on the M2 structure, but wherethe condition is determinate on M1, this will be noted. The M2
has a strong preprotocrista connected with a small paraconule.Paraconules are an unusual feature in extant platyrrhines,found only in Callimico. Early Miocene Homunculus and Car-locebus share this characteristic with Dolichocebus. AfricanEocene/Oligocene parapithecids have paraconules, as do oli-gopithecids and some propliopithecids. A metaconule is ab-sent. This cusp is present in parapithecids and variable inextant platyrrhines. A pericone is absent on both teeth. The hy-pocone is large and is distal and slightly lingual to the proto-cone on both teeth. A prehypocrista is present but weak. OnM2, there is a small mesostyle, but this cusp is not attachedto the premetacrista or postparacrista. A mesostyle is absenton M1. Both teeth have a weak and discontinuous buccal cin-gulum, but both have a strong lingual cingulum. On M1, thelingual cingulum disappears lingual to the protocone. OnM2, it wraps all the way around the lingual aspect of the pro-tocone and reaches to the preprotocrista. On both teeth, astrong postprotocrista runs toward the base of the metacone

Fig. 19. Upper molars of Dolichocebus gaimanensis: (A, B) MACN CH 357, a left
M3, in occlusal and occlusodistal views; (CeE) MACN CH 356, a left M2, in oc-
clusolingual stereopair, occlusal stereopair, and mesial views. Scale bar ¼ 3 mm.
344 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
where it joins the lateral posterior transverse crista. The M2 isslightly smaller than M1.
The M3 has a single lingual root and a twinned buccal root.There is a strong preprotocrista. A paraconule appears at the
lingual end of a crest running from the paracone. A smallmetaconule is present. There is no pericone. The hypoconeis present but much smaller than on the other molars. Thereis no prehypocrista. The tooth lacks a mesostyle and there isno buccal cingulum. A lingual cingulum is present but disap-pears mesially at the base of the protocone. A strong postpro-tocrista runs toward the base of the metacone where it joins thelateral posterior transverse crista.
Body size
Table 5 provides estimates of body mass from lower firstand second molar dimensions based on formulae publishedby Conroy (1987), Kay and Simons (1980), and Kay andMeldrum (1997). Mass determinations range from 1149 to1754 g. Perhaps the most appropriate estimate is derivedfrom M1 areas and body weights of 15 platyrrhine species,yielding the following least-squares regression (r2¼ 0.935):
ln female body weight¼ ln M1 area (1.565)þ 3.272.
Based on this equation, the estimated weight of MPEF 5146would be 1554 g; if MACN CH 866 is an M1, it wouldcome from an animal estimated to weigh 1527 g, aboutthe same size as the living masked titi monkey (Callicebuspersonatus).
Likewise, the length of the skull (prosthioneinion) of Do-lichocebus (~68 mm) falls within the range of skull lengthsof Callicebus personatus (mean of 10 males¼ 71.7 mm;mean of 10 females¼ 68.9 mm) (Hershkovitz, 1990). MeanC. personatus weights are 1270 g (males) and 1378 (females)(Hershkovitz, 1990). Thus, the estimates from lower molardimensions and cranial lengths are concordant.
Phylogenetic analysis
Two runs were undertaken using PAUP and the character-taxon matrix in Appendix 2. In the first run, extant taxawere constrained by a molecular tree topology for extant plat-yrrhines and catarrhines and with the position of all extincttaxa unconstrained. Thus, an extinct taxon may link whereverit is most parsimonious. In the second set of runs, we analyzedthe data set with the topology of extant platyrrhines and catar-rhines unconstrained. The two runs yielded essentially thesame result.
The backbone-constrained tree yields one most-parsimoni-ous tree (Fig. 20A). The tree is 66440 steps. The consistencyindex is 0.343, the retention index is 0.526, and the rescaledconsistency index is 0.180. The tree unites the crown platyr-rhines as a clade to the exclusion of all of the extinct taxa stud-ied except for Proteropithecia, which is linked to the sakis anduakaries, as previously reported by Kay et al. (1998a). Brani-sella is positioned as the most basal stem platyrrhine. All ofthe early Miocene taxa examined form a clade, within whichDolichocebus joins Carlocebus as its sister taxon, with Trem-acebus and Soriacebus more distantly related.

Tarsius
Aegyptopithecus zeuxis
Hylobates
Miopithecus talapoin
Presbytis melalophos
Apidium phiomense
Simonsius grangeri
Callicebus
Pithecia
Chiropotes
Cacajao
Proteropithecia
Aotus
Saimiri
Cebus
Saguinus
Callithrix
Cebuella
Callimico
Leontopithecus
Alouatta
Ateles
Lagothrix
Brachyteles
Dolichocebus gaimanensis
Carlocebus carmeni
Tremacebus harringtoni
Soriacebus ameghinorum
Branisella boliviana
Catopithecus browni
Proteopithecus sylviae
1000 changes
A
Fig. 20. (A) Maximum-parsimony analysis of the cranial and dental matrix, as constrained by the molecular ‘‘backbone.’’ The results are depicted as a ‘‘phylo-
gram’’ in which the branch lengths are proportional to the number of character changes. The scale for 1000 changes is given at the bottom. The character weight for
each character is 100, so the value 1000 is equivalent to 10 changes in a two-state character. (B) 50% majority consensus of the 11 shortest trees, when parsimony is
relaxed by 130 steps.
Table 5
Estimated body mass of Dolichocebus gaimanensis from M1 and M2 measurements
Specimen M1 area
(mm2)
M2 length
(mm)
Body-mass estimate (g)
Conroy’s all-primate
regression for M1
Conroy’s anthropoid
regression for M1
Kay and Simons’
M2 regression
Kay and Meldrum’s
platyrrhine M1 area
MPEF 5146 13.53 3.90 1322 (383e4568) 1754 (778e3953) 1262 1554
MACN CH 866 12.87 4.03 1296 (377e4461) 1724 (766e3879) 1149 1527
Notes: Formulae for M1 area are from Conroy (1987) and Kay and Meldrum (1997); M2 length formula is from Kay and Simons (1980).
345R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Tarsius
Aegyptopithecus zeuxis
Hylobates
Miopithecus talapoin
Presbytis melalophos
Apidium phiomense
Simonsius grangeri
Callicebus
Pithecia
Chiropotes
Cacajao
Proteropithecia
Aotus
Saimiri
Cebus
Saguinus
Callithrix
Cebuella
Callimico
Leontopithecus
Alouatta
Ateles
Lagothrix
Brachyteles
Dolichocebus gaimanensis
Carlocebus carmeni
Soriacebus ameghinorum
Tremacebus harringtoni
Branisella boliviana
Catopithecus browni
Proteopithecus sylviae
100
100
9164
100
55
73
100
B
Fig. 20 (continued).
346 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
By iteratively relaxing parsimony (i.e., by examining theconcurrence of nodes in trees of successively greater lengths,or Bremer support [Bremer, 1994]), we examined the stabilityof this phylogenetic pattern. Six further trees were identifiedup to 66,530 steps that continue to preserve the integrity ofthe crown platyrrhines, and recognize a cluster of late Oligo-ceneeearly Miocene taxa as stem platyrrhines (i.e., Branisella,Tremacebus, Dolichocebus, Soriacebus, and Carlocebus).However, only a Dolichocebus-Carlocebus clade among thestem taxa receives 100 percent support (the 50% majority-rule consensus of these 7 trees is shown in Fig. 20B). Theunity of crown Platyrrhini, excluding any of the fossil taxain our sample (again with the exception of Proteropithecia),breaks down at the fourteenth tree (>66,580 steps).
With respect to the second set of runs (extant taxa uncon-strained by molecular data), trees recovered from the cranialand dental data set are very similar to those found in the anal-ysis in which the extant taxa were constrained by moleculardata. We obtained a single most-parsimonious tree with theunconstrained cranial and dental data. The tree agrees withthe molecular phylogenies in the following features: (1) Tar-sius is sister to anthropoids; (2) a platyrrhine-catarrhine splitis supported; (3) late Oligocene and early Miocene taxa areplaced as stem platyrrhine taxa; (4) a crown platyrrhine clade(with Proteropithecia) is present; (5) a callitrichine clade in-cludes Callimico; (6) a pitheciine clade is supported; and (7)an atelid clade is supported. The morphological tree differsfrom that of the molecular tree in a number of ways as

Tarsius
Aegyptopithecus zeuxis
Hylobates
Miopithecus talapoin
Presbytis melalophos
Apidium phiomense
Simonsius grangeri
Callicebus
Aotus
Cebus
Pithecia
Chiropotes
Cacajao
Proteropithecia
Alouatta
Brachyteles
Lagothrix
Ateles
Saimiri
Saguinus
Callimico
Callithrix
Cebuella
Leontopithecus
Dolichocebus gaimanensis
Carlocebus carmeni
Tremacebus harringtoni
Soriacebus ameghinorum
Branisella boliviana
Catopithecus browni
Proteopithecus sylviae
1000 changes
Fig. 21. Maximum-parsimony analysis of the cranial and dental matrix uncon-
strained by a molecular ‘‘backbone.’’
347R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
indicated in Fig. 21. Thus, while the results are by no meansidentical, we feel secure in assuming that the cranial and den-tal data are sufficiently reliable to support phylogenetic infer-ences for the fossils.
Discussion
Character evolution
Three views offered in the literature as to the affinities ofDolichocebus relative to living platyrrhines are: (1) Dolichoce-bus is a relative of callitrichines (Kraglievich, 1951; Horovitz,1999); (2) Dolichocebus is a sister taxon of Saimiri (Rosen-berger, 1979a); and (3) Dolichocebus is a stem platyrrhine(Hershkovitz, 1982; Fleagle and Bown, 1983). Except forthe study by Horovitz (1999), proponents of these views singleout one character or a small suite of characters in support oftheir hypothesis. Our approach, following and amplifying
that of Horovitz and colleagues, has been to examine morecomprehensively the cranial and dental characters. Before dis-cussing our results, however, it is appropriate to consider thebasis for previous claims about the affinities of Dolichocebus.
The callitrichine hypothesis. All claims that Dolichocebus isa sister to extant Callitrichinae are based on dental evidence.The original basis for the claim that Dolichocebus is relatedto callitrichines is Kraglievich’s (1951) suggestion that Doli-chocebus shared with Callitrichinae the loss of the third molar.Hershkovitz (1970) noted that the pertinent region of the typeskull was broken away but concluded that there probably werethree molars. The new fossil material and reexamination of thetype skull using CT-imaging show that Dolichocebus had threemolars, not two as in stem callitrichines. The palate of the typeskull is broken away in the pertinent area, but the maxillarysinus is continuous and extensive above the roots of M2 andmust have continued behind this tooth, leaving ample roomfor an M3. Fleagle and Bown (1983) described an isolatedM3 of Dolichocebus (MACN CH 357), and one of the mandib-ular specimens described above has three molars and the otherhas an M2 with an interproximal facet for M3.
Horovitz (1999) suggested that Dolichocebus, along withlate Oligocene Branisella and several early to middle Mioceneplatyrrhines, were sister taxa to extant Callitrichinae (see alsoTakai et al.’s [2000] views about Branisella). She cited threecharacters in support of callitrichine affinities of Dolichoce-bus. The first of these is a large P3 protoconid relative to theP4 protoconid (vs. subequal in size). This character cannotbe evaluated based on the known Dolichocebus material, asrecognized by Horovitz (1999). The second character is theabsence of a P4 hypoconid (vs. present). Our examination ofthis character (number 129 in our matrix) does not supportHorovitz’s observation that extant callitrichines lack a P4 hy-poconid. The P4 hypoconids are generally present in extantplatyrrhines, but absent or cristiform in extant atelids. Likeother nonatelid platyrrhines, Dolichocebus possesses a P4
hypoconid (MACN CH 898). The final character cited byHorovitz is the absence of a P4 entoconid (vs. present). Ourexamination of this character (number 123 in our matrix) con-firms Horovitz’s observation that extant callitrichines havea lingual crest bordering the P4 talonid but lack an entoconid.However, the only known P4 of Dolichocebus (MACN CH898) has a discrete P4 entoconid.
The Saimiri/cebine hypothesis. It has been suggested thatDolichocebus is related to cebines (Cebus and Saimiri) andspecifically to Saimiri. Rosenberger proposed several synapo-morphies that link cebines with Dolichocebus: (1) narrownasal bones, an elongate interorbital process of the frontal,and vaulted frontals without a depressed glabella; and (2) anelongate (dolichocephalic) braincase. CT scans reveal thatthe precise location and size of the frontal and nasal bonescannot be determined due to the poor preservation of the inter-orbital region. Additionally, the glabellar region is missingfrom the type skull.
Values of braincase shape (100 � braincase length/brain-case breadth) of Dolichocebus, Tremacebus, and some extantplatyrrhines are given in Table 6. Using this index, the longer

Table 6
Braincase shape in selected living and extinct platyrrhines
Taxa Braincase
length (n)
Braincase
breadth (n)
Shape*
Pitheciinae
Callicebus torquatus lugens1
Male 53.5 (41) 36.8 (40) 145
Female 52.7 (31) 35.5 (31) 149
Callicebus moloch1
Male 52.4 (24) 35.0 (23) 150
Female 52.0 (32) 34.3 (31) 152
Callicebus personatus1
Male 57.2 (11) 36.4 (11) 157
Female 55.7 (9) 35.9 (10) 155
Aotinae
Aotus trivirgatus2 51.5 (10) 33.9 (10) 152
(mixed sex)
Cebinae
Saimiri boliviensis3
Male 54.5 (11) 36.0 (11) 151
Female 52.6 (7) 36.1 (7) 146
Cebus albifrons cesarae4
Male 78.6 (18) 50.5 (18) 156
Female 74.9 (10) 50.8 (10) 147
Callitrichinae
Cebuella pygmaea5 30.9 (62) 21.1 (60) 146
(mixed sex)
Callithrix jacchus5 40.5 (23) 25.6 (24) 158
(mixed sex)
Callithrix argentata5 39.6 (45) 25.7 (45) 154
(mixed sex)
Saguinus fuscicollis5 42.1 (161) 26.1 (164) 161
(mixed sex)
Saguinus mystax5 45.6 (28) 28.7 (28) 159
(mixed sex)
Leontopithecus rosalia5 47.3 (10) 28.4 (13) 167
(mixed sex)
Callimico goeldii5 45.8 (11) 29.8 (10) 154
(mixed sex)
Atelidae
Alouatta seniculus4
Male 74.4 (7) 56.6 (7) 131
Female 67.5 (7) 49.8 (7) 136
Early Miocene taxa
Tremacebus harringtoni2 51.0 (1) 34.0 (1) 150
Dolichocebus gaimanensis2 52.0 (1) 32.0 (1) 163
Data sources: 1 Hershkovitz (1990: 88, 92, 95); 2 Hershkovitz (1974: 30);3 Hershkovitz (1984); 4 Hershkovitz (1949: 391); 5 Hershkovitz (1977: Appen-
dix Table 1).* Shape¼ 100� (brain case length/braincase breadth).
348 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
and narrower the braincase, the larger the index. The values inthe table show that living cebines are not dolichocephalic andthat Dolichocebus, if anything, resembles tamarins in brain-case shape. Representative species of Saimiri and Cebus rangefrom 147 to 151. This range is encompassed by that of marmo-sets and falls within the range of three species of Callicebus.Living cebines are not especially long-headed. The ratio forDolichocebus is 163. However, we suspect that this value is in-flated given that the skull was mediolaterally compressed post-mortem. Even taken as the true value, Dolichocebus resemblesSaguinus and Leontopithecus more closely than either Saimirior Cebus.
Proposed specific similarities shared between Dolichocebusand Saimiri are based on features of a ‘‘lightly built’’ mastica-tory system and the structure of orbit (Rosenberger, 1979a, b;Fleagle and Rosenberger, 1983; Rosenberger et al., 1990).Turning first to the masticatory system, it is difficult to deter-mine whether or not Dolichocebus did in fact have a lightlybuilt masticatory system (Hershkovitz, 1982) and, if it did,whether this is a shared derived similarity with Saimiri. Rosen-berger (1979a) based his claim on the following points thatSaimiri and Dolichocebus are said to share: (1) gracile anteriorand posterior roots of the zygomatic arch, suggesting a weakmasseter muscle; (2) a slender pyramidal process of the pala-tine, suggesting weakly developed pterygoid muscles; (3) tem-poral line on parietal neither deeply etched nor rugose,suggesting a weak temporalis muscle; (4) a transversely shal-low temporal fossa (i.e., the cross-sectional area of the tempo-ral fossa is small); (5) entoglenoid fossa shallow andpostglenoid process absent medially, suggesting reduced me-diolateral component of force in the temporomandibular joint.
With respect to first point, the zygomatic arches, whichserve as the origin for the masseter, are broken away, as areboth roots of the zygomata bilaterally. Regarding the secondpoint, the pterygoid plates, which provide the origin for themedial pterygoid musculature, together with the pyramidalprocesses that support them, are not preserved. (The palateis broken posteriorly so that even the M3s are absent). Onthe third and fourth points, the temporal lines (marking the or-igin of the temporalis muscle from the sides of the skull) areweak, as Rosenberger (1979a) described, but the temporallines are higher on the skull (closer to the midline) suggestingeither that the temporalis was more extensive or that the brain-case was smaller. Moreover, in the absence of zygomaticarches, it is impossible to determine the cross-sectional areaof the temporal fossa, because the arches define its lateral bor-der. The weak temporalis muscle markings (point four) do re-semble Saimiri, but they also resemble all other small-bodiedplatyrrhines. On the last point, Rosenberger (1979a) correctlystated that Dolichocebus and Saimiri both have a shallow gle-noid fossa, but this is true for most platyrrhines, especially allof the small-bodied taxa. In agreement with Rosenberger, thepostglenoid process of Dolichocebus is quite small, as in Sai-miri, but this similarity also extends to Aotus and callitrichines(see footnote 3).
A strong postglenoid process is associated with posteriorexpansion of the articular surface of the temporomandibularjoint in primates and ungulates that have a large transverse ex-cursion of the mandible during the power stroke of mastication(Wall, 1998). Conversely, having a weak process may tell usless about the amount of muscular force engendered by thepower stroke and more about the extent of transverse excur-sion during that stroke. The portions of the mandible that serveas insertions for the muscles of mastication are not preservedin any specimen.
In summary, the extent of similarity between Dolichocebusand Saimiri has been overstated by reference to anatomical de-tails that are not preserved. Similarities that can be docu-mented in the masticatory apparatus of Dolichocebus and

349R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Saimiri can also be observed in other small-bodied platyr-rhines. Some of these similarities may have more to do withthe extent of mandibular excursion and less to do with masti-catory force per se.
Several aspects of the orbital region also led Rosenberger(1979a) to suggests that Saimiri and Dolichocebus might besister taxa. The most impressive features that have been sug-gested to link Saimiri and Dolichocebus are the narrow inter-orbital pillar and the interorbital fenestra.5 After carefulpreparation using a UV light to distinguish bone from matrix,Rosenberger (1979a) concluded that there was no bone be-tween the orbits and inferred that an interorbital fenestrawas present in life. CT scans reveal that, superficially, the in-terorbital region is poorly preserved. However, cross sectionsindicate that much of the interorbital region of Dolichocebusis composed of matrix rather than bone. They also suggestthat Dolichocebus possessed a cupular sinus (as in Cebus,but not Saimiri) and that the interorbital region was probablynarrow. The narrow interorbital region is indeed a possible de-rived similarity shared with Saimiri, as Rosenberger sug-gested. However, the CT scans neither confirm nor precludethe presence of an interorbital fenestra in Dolichocebus.
Saimiri possesses a number of other anatomical peculiari-ties (probably derived features) of the cranium, such as a rela-tively horizontal nuchal plane and the absence of a maxillarysinus. Dolichocebus, in contrast, has a more obliquely orientednuchal plane and a large maxillary sinus. While both of the lat-ter features are probably symplesiomorphic for platyrrhines asa whole and do not exclude Dolichocebus from the ancestry ofSaimiri, neither do they reinforce the impression that Dolicho-cebus was especially Saimiri-like. The orbits of Dolichocebusare less convergent than those of Saimiri and other noncallitri-chine platyrrhines. The more divergent orbits of Dolichocebusare also seen in early Miocene Tremacebus and may be an-other trait of stem platyrrhines that persists in callitrichines.
Broader phylogenetic inferences
To reassess of the phylogenetic position of Dolichocebus,we undertook a broader phylogenetic analysis. As we haveseen in the descriptive anatomy presented above, Dolichoce-bus presents a confusing mix of similarities to many extantand extinct platyrrhine taxa. Put another way, phylogeneticconclusions based upon a single character or on a few selectedcharacters are very likely to be inaccurate. Overall measures ofhomoplasy, such as the consistency index and the retention in-dex (e.g., Maddison and Maddison, 2001), indicate high levelsof homoplasy within our data set of cranial and dental charac-ters. Accordingly, a few characters can be found to support al-most any phylogenetic hypothesis. Our choice among these
5 It should be added that even though possession of an interorbital fenestra is
unusual in primates, it demonstrably does occur in at least one other extant pri-
mate that we have examineddsome specimens of the catarrhine Erythrocebus
(e.g., Duke Lemur Center Osteology specimen 064) have a large fenestra.
Thus, even this feature exhibits homoplasy.
conflicting hypotheses is overtly cladistic and involves severalassumptions:
(1) We used outgroup comparisons to establish polarities ofthe characters under consideration. For this purpose we se-lected a variety of African late Eocene and early Oligo-cene taxa as outgroups.
(2) Where molecular phylogenies are sufficiently well corrob-orated, we used them to constrain our conclusions.
(3) We eschewed character-weighting schemes. There seemsto be no satisfactory reason for character weighting. Forexample, there is no reason to believe that loss of the hy-pocone on the premolars is any more (or less) likely to oc-cur than loss of the same feature on the molars or loss ofthe third molars. Yet historically, phylogenetic assessmenthas placed greater reliance (weight) on loss of molar hypo-cones and third molars and given scant attention to pres-ence or absence of premolar hypocones. However, thereis simply no a priori reason for believing that some fea-tures are better indicators of phylogeny than others.
(4) We did not assume that certain suites of cranial or dentalfeatures found together in a clade of living platyrrhinesevolved together and simultaneously as a single adaptivepattern or developmental pattern. For example, it hasbeen proposed that loss of the third molar and loss of mo-lar hypocones, along with a number of features of postcra-nial anatomy and several life-history traits, are the resultof phyletic dwarfing and the constraints of small bodysize on diet, foraging patterns, and reproductive strategyin callitrichines (Ford, 1980). This hypothesis might leadus to assume that we had simply scored the same characterseveral times. Three things that we know about the evolu-tion of the Anthropoidea run counter to the expectations ofthis particular hypothesis. First, the ancestors of callitri-chines may not have been larger than they are today, andtherefore callitrichines may not be phyletic dwarfs. Whilethe ancestry of callitrichines is too poorly known to be cer-tain, many Eocene/Oligocene African stem anthropoids andsome Miocene noncallitrichine Colombian platyrrhineswere within the size range of living callitrichines. Second,callitrichine-sized Anthropoidea are not constrained tohave this particular pattern of morphology and behavior.The platyrrhine Patasola from the Miocene of Colombia(Kay and Meldrum, 1997) falls within the size range of liv-ing callitrichines but has a hypocone and third molar. Third,several of the dental traits of this ‘‘character complex’’ arenot always linked with small body size in platyrrhines.Lagonimico, another Miocene Colombian primate (Kay,1994), has lost the hypocone and greatly reduced the thirdmolar but is much larger than any living callitrichine. Con-versely, Callimico goeldii is sister to Callithrix among livingplatyrrhines and falls within the size range of living callitri-chines, but it retains (or has regained) the third molar andhypocone.
Character evolution and the place of Dolichocebus withinthe Platyrrhini. A stem anthropoid is an extinct taxon that is

350 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
more closely related to living Anthropoidea than to the haplor-hine Tarsius, but one that is outside the crown-anthropoidclade, which includes all living anthropoids, their last commonancestor (LCA), and all extinct taxa that are descendants ofthat LCA. Our maximum-parsimony phylogeny (Figs. 20,21) places late Eocene and early Oligocene African Proteopi-thecus as a stem anthropoid. Early Oligocene African Aegyp-topithecus is a stem catarrhine. The analysis places theparapithecids Apidium and Simonsius as stem catarrhines.However, a more inclusive analysis with related, more primi-tive parapithecid taxa such as Qatrania suggests that parapi-thecids are a stem anthropoid clade, and it reveals thata number of the molar characters that link Apidium and Simon-sius with catarrhines are homoplasies instead of synapomor-phies (Kay et al., 1997; Ross et al., 1998; Kay et al., 2004c;Seiffert et al., 2004). The analysis is equivocal as to whetherlate Eocene African Catopithecus (Oligopithecidae) is a stemcatarrhine or a stem anthropoid; for reasons given by Kayand Delson (2000), we consider Catopithecus to be a stemanthropoid.
Our maximum-parsimony trees (Figs. 20, 21) suggest thatBolivian late Oligocene Branisella, Patagonian early MioceneDolichocebus and Tremacebus, and Patagonian late early Mio-cene Carlocebus and Soriacebus are stem platyrrhines. Addi-tionally, there is weak evidence for a stem platyrrhine cladeof the Patagonian taxa comprising Dolichocebus, Tremacebus,Carlocebus, and Soriacebus. Latest early Miocene Homuncu-lus, under study elsewhere, may also belong to such a clade(Kay et al., 2005). Likewise, early Miocene Chilecebus maybe a stem platyrrhine (Flynn et al., 1995) but is unavailableto us for study.
Table 7 summarizes the status of 41 cranial and dental fea-tures we consider most informative of the above phylogeneticrelationships. Figure 22 is a simplified version of Figs. 20 and21 for the purposes of our discussion. Before discussing thesecharacters, it should be emphasized that all of the traits beingdiscussed demonstrate examples of independent parallel acqui-sition under virtually every phylogenetic scenario. A measure ofsuch parallelism is the consistency index (CI; (Maddison andMaddison, 2001). A CI of 1.0 indicates that no homoplasy hasoccurred. The characters in Table 7 have CIs ranging from0.17 to 1.00. Homoplasy is present in nearly all informativecharacters in our most-parsimonious phylogeny. This resultshould come as no surprise, considering the pervasiveness ofthis phenomenon in dental, cranial, and postcranial characters(Sanchez-Villagra and Williams, 1998; Williams, 2007).
The character states listed in Table 7 as the platyrrhine andcatarrhine character states represent the reconstructed states ofthe crown platyrrhine and catarrhine nodes (nodes 1e4 inFig. 22), that is, the states of the LCAs of the crown groups,as reconstructed from the parsimony analysis. We use theterms ‘‘shared derived’’ and ‘‘derived’’ in place of synapomor-phy and apomorphy, respectively. The terms ‘‘primitive’’ and‘‘shared primitive’’ are substituted for plesiomorphic or sym-plesiomorphic, respectively. We first consider the overall pat-tern of anthropoid evolution based on a contrast between thecharacter states of stem anthropoids and our hypothetical
reconstructions of the ancestral condition of crown catarrhineand platyrrhine LCAs (nodes 3 and 4 in Fig. 22). Secondly, weevaluate the evidence for whether Aegyptopithecus is a crownor a stem catarrhine (i.e., a branch of the lineage segment be-tween nodes 1 and 3). Thirdly, we consider the evidence forwhether Dolichocebus and the other late Oligocene and earlyMiocene New World taxa Branisella, Tremacebus, Carlocce-bus, and Soriacebus are crown or stem platyrrhines (a branchor branches along the lineage segment between nodes 1 and 2in Fig. 22). Fourthly, we examine the evidence supporting theexistence of distinct clades of stem platyrrhines (support fornode 4 in Fig. 22).
Stem anthropoids, crown catarrhines, and crown platyr-rhines contrasted. A number of the craniodental charactersof the stem anthropoids of late Eocene and early Oligoceneof AfricadApidium, Simonsius, Proteopithecus, and Catopi-thecusddiffer from the crown anthropoid LCA (node 1) orfrom the morphotypes of the crown catarrhine or crown plat-yrrhine LCAs (nodes 2 and 3, respectively):
� The arrangement of bones at pterion (Character 9) variesamong anthropoids (Ashley-Montague, 1933). The frontalcontacts the alisphenoid at pterion in Tarsius and all crowncatarrhines, whereas in the crown platyrrhine LCA, thezygomatic contacts the frontal. Based on the Tarsius out-group, we infer that alisphenoid-frontal contact is theprimitive stem-anthropoid state, although the conditionhas not been established in any stem anthropoid (seealso the discussion of this feature in Fleagle and Kay,1987).� The infraorbital foramen of the maxillary bone is variably
positioned in anthropoids relative to the maxillary teeth(Character 14). In Catopithecus, it is posteriorly posi-tioned, whereas it is more anteriorly positioned in Apidium,Simonsius, and Proteopithecus. The foramen is positionedquite far forward relative to the maxillary cheek teeth, a de-rived condition in the crown platyrrhine LCA (Horovitz,1999), whereas in the crown catarrhine LCA, the foramenis more posteriorly positioned.� The ascending wing of the premaxilla (Character 21) is
also variably developed, being broad in Catopithecus (Si-mons and Rasmussen, 1996) but narrow in Simonsius andProteopithecus (Simons, 2001). Crown platyrrhine and cat-arrhine LCAs have an anteroposteriorly reduced wing of thepremaxilla. This apparently shared derived state of the twocrown groups evolved independently, as evidenced by thebroad ascending wing of the premaxilla in the stem catar-rhine Aegyptopithecus (Fleagle and Kay, 1983).� In the stem anthropoids Proteopithecus, Catopithecus, and
to a lesser degree Simonsius, the maxillary postcaninetooth rows diverge posteriorly (Character 25). This con-dition is also inferred in the crown platyrrhine LCA,whereas the crown catarrhine LCA is derived in havingmore parallel-sided tooth rows.� The stem anthropoid Proteopithecus has an abbreviated
medial pterygoid plate, whereas Simonsius and the crowncatarrhine LCA have broad medial pterygoid plates

Table 7
Selected cranial and dental character distributions
Character Consistency index Character states
in late Eocene
African anthropoids
Aegyptopithecus Character states
of crown Catarrhini
Dolichocebus Character states
of crown Platyrrhini
Character 9: Pterion 0.75 ? Frontal-alisphenoid contact Frontal-alisphenoid contact ? Zygomatic-parietal contact
Charcter 14: Position
of infraorbital foramen
0.29 Posteriorly positioned
(Catopithecus); anteriorly
positioned (Proteopithecus)
Posteriorly positioned Posteriorly positioned More anteriorly positioned More anteriorly positioned
Character 21: Breadth
of ascending wing
of the premaxilla
0.50 Broad (Catopithecus); narrow
(Proteopithecus)
Broad Narrow Narrow Narrow
Character 25: Divergence
of palatal tooth rows
0.45 Diverge slightly Diverge slightly Diverge slightly Diverge slightly Diverge more widely
Character 27: Size
of medial pterygoid plate
0.40 Short medial
plate (Proteopithecus);
unknown (Catopithecus)
Long medial plate Long medial plate ? Short medial plate
Character 43: Orbital
convergence
0.40 Less than 55� (Apidium,
Simonsius, Catopithecus, and
Proteopithecus)
Greater than 65� Greater than 65� Less than 65� Greater than 65�
Character 57: Position
of external carotid foramen
0.67 Medially positioned Ventrally positioned Ventrally positioned Medially positioned Medially positioned
Character 66: Ectotympanic 0.50 Annular Annular Tubular Annular Annular
Character 74: Extent
of ossification of tentorium
cerebelli
0.20 Extensively ossified Minimally ossified Minimally ossified Extensively ossified Extensively ossified
Character 75: Cartmill’s
canal
0.33 Open Partially occluded Absent Open Open
Character 77: Relative
brain size
0.22 Unknown Small Larger Larger Larger
Character 80: Mandibular
depth
0.25 Shallow Deep Deep ? Deep
Character 81: Mandibular
symphyseal fusion
0.50 Unfused Fused Fused ? Fused
Character 82: Position
of coronoid process
0.20 Well above condyle Well above condyle At or slightly
above condyle
? At or slightly
above condyle
Character 111: P2 0.50 Second premolar
present (Proteopithecus);
absent (Catopithecus)
Second premolar absent Second premolar absent Second premolar present Second premolar present
Character 113: P3e4 roots 0.67 P3e4 two-rooted P3e4 two-rooted P3e4 two-rooted P3e4 single-rooted P3e4 single-rooted
Character 122: P3 metaconid 0.56 P3 metaconid absent
or trace
P3 metaconid absent
or trace
P3 metaconid absent
or trace
Small P3 metaconid Large P3 metaconid
Character 128: P3 protocristid 0.33 Distolingual Distolingual Distolingual Distolingual Transverse
Character 130: P4 protocristid 0.33 Transverse Transverse Transverse Distolingual Transverse
Character 139: Premolar
talonid
0.17 P4 talonid and trigonid
lengths z equal
P4 talonid elongate P4 talonid elongate P4 talonid very short P4 talonid very short
Character 144: Relative
heights of P3 vs. P4
0.50 P3 and P4 of similar height P3 projects above P4 P3 projects above P4 P3 and P4 of similar height P3 and P4 of similar height
Character 145: Size of P3 0.38 P3 not enlarged P3 enlarged P3 enlarged P3 not enlarged P3 not enlarged
Character 146: P4 shape
(MD/BL)
0.50 More elongate Short and broad More elongate Short and broad Short and broad
(continued on next page)
35
1R
.F.
Kay
etal.
/Journal
ofH
uman
Evolution
54(2008)
323e382

Table 7 (continued)
Character Consistency index Character states
in late Eocene
African anthropoids
Aegyptopithecus Character states
of crown Catarrhini
Dolichocebus Character states
of crown Platyrrhini
Character 152: M3 root
number
0.33 Two-rooted Two-rooted Two-rooted Two-rooted Single-rooted
Character 162: M3 heel 0.57 Narrow Narrow Narrow ? Absent
Character 164: M1e2
trigonid-to-talonid height
0.40 Trigonid higher
than talonid
Trigonid and talonid
heights z equal
Trigonid and talonid
heights z equal
Trigonid and talonid
heights z equal
Trigonid and talonid
heights z equal
Character 166: M1 trigonid
lingual wall
0.50 Open lingually Closed lingually Closed lingually Closed lingually Closed lingually
Character 169: Protocristid
orientation
0.50 Protocristid runs
towards metaconid
Runs into hypoflexid Runs into hypoflexid Protocristid runs
towards metaconid
Protocristid runs
towards metaconid
Characters 172, 173: Wear
facet X
1.00, 0.50 Absent Present Present Absent Absent
Character 175: Postentoconid
sulcus
0.44 Present Present, large Present, large Present Absent
Character 176: M1
hypoconulid
0.43 Small Large Large Small Absent
Character 177: M2
hypoconulid
0.46 Moderate Large Large Small Absent
Characters 183, 184: M1e2
cristid obliqua terminus
0.33, 0.25 To base of trigonid To base of trigonid To base of trigonid Extends onto
posterior trigonid wall
or reaches protocristid
Extends onto
posterior trigonid wall
or reaches protocristid
Character 190: M1e2 distal
fovea
0.40 Absent Present Present Present Absent
Character 196: M1 absolute
size (estimate of body size)
0.25 Very small Large Large Small Small
Character 213: Upper
canine occlusion
0.50 Upper canine
occludes with P2
(Proteopithecus); with P3
(Catopithecus)
Upper canine
occludes with P3
Upper canine
occludes with P3
Upper canine
occludes with P2
Upper canine
occludes with P2
Character 217: Number of P3
roots
0.36 Two (Catopithecus); three
(Proteopithecus)
Three Three Two Two
Character 218: Number of P4
roots
0.44 Three (Catopithecus); three
(Proteopithecus)
Three Three Two Two
Character 228: P4 hypocone
development
0.57 Hypocone absent Small hypocone present Hypocone absent Hypocone enlarged Hypocone small
or absent
Character 233: Profile
of P3 distal crown margin
0.25 Waist between
protocone and paracone
Straight or convex
distal crown margin
Straight or convex
distal crown margin
Straight or convex
distal crown margin
Straight or convex
distal crown margin
Character 236: Development
of P3e4 buccal cingulum
0.50 Strong Absent Absent Absent Absent
35
2R
.F.
Kay
etal.
/Journal
ofH
uman
Evolution
54(2008)
323e382

Dolichocebus
Tremacebus
Carlocebus
Soriacebus
Branisella
CrownCatarrhines
CrownPlatyrrhines
crownanthropoidnode
Stem catarrhinetaxa (includingAegyptopithecus)
crownplatyrrhinenode
Stem anthropoidtaxa (includes Apidium,Simonsius,Catopithecus,Proteopithecus)
crowncatarrhinenode
Stem platyrrhines
1
2
Stem & Crown Platyrrhini
3
Stem & Crown Catarrhini
4?
Fig. 22. Simplified cladogram illustrating the crown and stem platyrrhines. Labeled nodes are discussed in the text.
353R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
(Character 27). In the crown platyrrhine LCA, the medialpterygoid plate is extremely reduced to a low ridge or evenabsent.� In the stem anthropoids Apidium, Simonsius, Proteopithe-
cus, and Catopithecus, the orbital convergence is less than65� (Character 43), whereas in the crown platyrrhine andcatarrhine LCAs, orbital convergence is less than 65�
(Ross, 1995).� In the stem anthropoids Apidium, Simonsius, Proteopithe-
cus, and Catopithecus, and in the crown platyrrhine LCA,the internal carotid artery enters the middle ear througha foramen positioned ventromedially on the bulla (Char-acter 57), whereas in the crown catarrhine LCA, the canalis more medially positioned relative to bullar landmarks(Ross, 1994).� The ectotympanic is annular in the stem anthropoids Api-
dium, Simonsius, Catopithecus, and Proteopithecus (Char-acter 66), a condition retained in the crown platyrrhineLCA, whereas the crown catarrhine LCA is derived in hav-ing a laterally expanded, collar-shaped ectotympanic.� In the stem anthropoids Simonsius, Catopithecus, and Pro-
teopithecus the tentorium cerebelli is partially ossified(Character 74) (Kay et al., 2008). The crown platyrrhineLCA shares the stem anthropoid condition, whereas thederived condition in the crown catarrhine LCA is for thetentorium to be unossified.
� In the stem anthropoids Apidium and Simonsius, Cartmill’scanal (Character 75) is absent, whereas it is present inCatopithecus, Proteopithecus, and all crown platyrrhines(Kay et al., 2008). A derived condition in crown catar-rhines is for the canal to be absent.� Overall brain size (endocranial volume) cannot be estab-
lished in Catopithecus or Proteopithecus, but it is smalldinthe strepsirrhine rangedin Simonsius (Character 77). Thecrown catarrhine and platyrrhine LCAs both have relativelylarge endocranial volumes compared with Simonsius, Tar-sius, and extant strepsirrhines. This apparently sharedderived state of the two crown groups evolved indepen-dently, as evidenced by the relatively small endocranialvolume in the stem catarrhine Aegyptopithecus (Radinsky,1977; Simons, 1993; Simons et al., 2007).� In the stem anthropoids Apidium, Simonsius, Catopithecus,
and Proteopithecus, the mandibular corpora are shallow(Character 80). The catarrhine and platyrrhine LCAsboth have deep mandibular corpora.� The mandibular symphysis is fused in Apidium and Simon-
sius but unfused in Catopithecus and Proteopithecus (Char-acter 81). All catarrhines and platyrrhines have fusedmandibular symphyses as adults (Ravosa and Hogue, 2003).� In the stem anthropoids Apidium, Catopithecus, and Pro-
teopithecus, the tip of the coronoid process reaches highabove the condyle (Character 82). In the crown catarrhine

354 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
and platyrrhine LCAs, the coronoid process is at orslightly below the level of the mandibular condyles.� The stem anthropoids Apidium, Simonsius, and Proteopi-
thecus retain three permanent premolars (Character111). All crown platyrrhines share the primitive anthro-poid three-premolar condition. Crown catarrhines and,convergently, Catopithecus show a reduction in the num-ber of premolars from three to two.� In the stem anthropoids Apidium, Simonsius, Catopithe-
cus, and Proteopithecus the lower third and fourthpremolars are two-rooted (Character 113). All crowncatarrhines retain the primitive arrangement, but allcrown platyrrhines have the derived single-rooted P3
and P4 (Kay, 1980).� Stem anthropoids Apidium, Simonsius, Catopithecus, and
Proteopithecus have a small or absent P3 metaconid (Char-acter 122). Crown catarrhines also have a reduced P3 meta-conid. This condition appears not to be simply the result ofthe loss of the second premolar and attendant crown simpli-fication in the latter because, as noted, Proteopithecus,which possesses three premolars, also has a simple P3 with-out the metaconid. The crown platyrrhine LCA is derived inhaving a large P3 metaconid (Kay, 1980).� Stem anthropoids Apidium, Simonsius, Proteopithecus, and
Catopithecus and the crown catarrhine LCA have distolin-gually oriented P3 protocristids (Character 128). In con-trast, the crown platyrrhine LCA is derived in havinga transversely oriented P3 protocristid (Kay, 1980).� The stem anthropoids Proteopithecus and Catopithecus
have transversely oriented P4 protocristids, whereas Api-dium and Simonsius have more distolingually oriented P4
protocristids (Character 130). The crown catarrhine andplatyrrhine LCAs have transversely oriented P4 protocris-tids like Proteopithecus and Catopithecus.� In the stem anthropoids Proteopithecus and Catopithecus,
the ratios of P4 talonid to trigonid mesiodistal lengths areapproximately equal (Character 139). The crown catar-rhine LCA has an elongate talonid and short trigonid.The crown platyrrhine LCA has a mesiodistally short P4
talonid. Thus, crown catarrhines, on one hand, and crownplatyrrhines, on the other hand, have attained the two dif-ferent derived states compared with the primitive condi-tion seen in Catopithecus and Proteopithecus.� In the stem anthropoids Apidium, Simonsius, Proteopithe-
cus, and Catopithecus, the overall size and projectiveheights of P3 and P4 are similar (Characters 144, 145).The crown platyrrhine LCA shares these stem anthropoidconditions, whereas the crown catarrhine LCA is derivedin having an enlarged P3, and its prototoconid projectswell above that of P4.� The stem anthropoids Apidium, Simonsius, Proteopithecus,
and Catopithecus have mesiodistally elongate P4s (Char-acter 146), as does the crown catarrhine LCA. In contrast,the crown platyrrhine LCA is derived in having a mesiodis-tally short and buccolingually broad P4.� In the stem anthropoids Apidium, Simonsius, Proteopithe-
cus, and Catopithecus, as well as the crown catarrhine
LCA, M3 has two roots (Character 152) and M3 crownshave a third lobe featuring a hypoconulid (Character162). The crown platyrrhine LCA is derived in havingthe M3 roots reduced to one, often with a figure-eight crosssection, and the M3 hypoconulid and third lobe are absent(Kay, 1980).� The stem anthropoids Proteopithecus and Catopithecus ex-
hibit the presumed anthropoid primitive condition of havingthe molar trigonids raised above the talonids (Character164). The crown catarrhine and platyrrhine LCAs sharethe derived condition in which the M1 and M2 trigonidsare not or are only slightly raised above the talonids. Theparapithecids Apidium and Simonsius are derived in havingthe trigonid and talonid of similar heights; however, moreprimitive parapithecids such as Qatrania exhibit the stem-anthropoid condition, demonstrating that this characterevolved homoplastically in parapithecids and crowncatarrhines.� In the stem anthropoids Catopithecus and Proteopithecus,
the protocristids of M1 and M2 run towards the metaco-nids, and M1e2 lack Phase II wear facet X (Characters169, 172, 173). The crown catarrhine LCA exhibits a de-rived condition of having M1e2 protocristids deflecteddistally into the hypoflexid and in having a distinctM1e2 Phase II wear facet X. The crown platyrrhineLCA retains the primitive condition of these characters.The parapithecids Apidium and Simonsius have M1e2 pro-tocristids deflected distally into the hypoflexid and a dis-tinct M2 Phase II wear facet X (M2 only). More primitiveparapithecids such as Qatrania exhibit the stem-anthro-poid conditions, demonstrating that these charactersevolved homoplastically in parapithecids and crowncatarrhines.� Stem anthropoids Simonsius, Proteopithecus, and Catopi-
thecus have a weak but distinct M1e2 postentoconid sul-cus. The crown catarrhine LCA is derived in havingbroad, open sulci between the entoconids and hypoconul-ids of M1e2 (Character 175). Additionally, M1 and M2
have a pronounced fovea distal to the entoconid for receiptof the enlarged hypocone (Character 190). The postento-conid sulcus and distal fovea are absentdderived condi-tions in the crown platyrrhine LCA (Kay, 1980). Theparapithecid Apidium, in parallel with basal catarrhines,has a broad, open postentoconid sulcus and distal fovea,but these traits are not present in Simonsius or primitiveparapithecids, and thus we infer the resemblance betweenApidium and crown catarrhines to be homoplastic.� Proteopithecus and Catopithecus have M1e2 hypoconulids
of moderate size (Characters 176, 177). The crown catar-rhine LCA has large M1e2 hypoconulids. A derived featureof the crown platyrrhine node is the absence of M1e2 hypo-conulids (Kay, 1980). The parapithecid Apidium, in parallelwith basal catarrhines, has large hypoconulids.� In stem anthropoids and the crown catarrhine LCA, the
M1e2 cristid obliquas run to the base of the posterior trig-onid wall (Characters 183, 184). The crown platyrrhineLCA exhibits a derived condition in having the M1e2

355R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
cristid obliquas extending onto the posterior wall of thetrigonid reaching towards the protoconid (Kay, 1980).� Using M1 size as a surrogate for body size (Character
196), the stem anthropoids Proteopithecus and Catopi-thecus were very small animals compared with allcrown catarrhines and smaller than the crown platyr-rhine LCA.� The stem anthropoids Apidium, Simonsius, and Proteopithe-
cus share with crown platyrrhines an occlusal arrangementwhereby the upper canine occludes with P2 (Character213). Crown catarrhines are derived in having the upper ca-nine in occlusion with P3. Catopithecus shares the C-P3 oc-clusal arrangement with catarrhines, one of a very fewpossible shared derived cranial or dental features that couldsupport a link between oligopithecids and catarrhines(Simons and Rasmussen, 1996).� The stem anthropoids Apidium, Simonsius, Proteopithecus,
and Catopithecus have three-rooted P3e4sdtwo buccal rootsand one lingual root (Characters 217, 218). This primitivecondition is present in the crown catarrhine LCA. The crownplatyrrhine LCA is derived in having two-rooted P3s andP4sdone buccal and one lingual root (Kay, 1980).� In stem anthropoids Apidium, Simonsius, Proteopithecus,
and Catopithecus, as well as the crown platyrrhine and cat-arrhine LCAs, a P4 hypocone is absent or very weakly de-veloped (Character 228).
Crown-anthropoid features. A number of derived characterstates of the crown Anthropoidea LCA have been identified inour analysis (node 1 in Fig. 22). These features are found intaxa identified in our analysis as stem catarrhines (Aegyp-topithecus) and platyrrhines (Branisella, Dolichocebus, Soria-cebus, and Carlocebus) and in the crown catarrhine andplatyrrhine LCAs:
� All crown anthropoids have a fused mandibular symphy-sis and most have deep mandibular corpora. The stemanthropoids Proteopithecus and Catopithecus haveunfused and shallow mandibles (Characters 80, 81).The late Oligocene Branisella has a shallow mandible.This suggests that deep mandibles may have evolved in-dependently in the crown catarrhines and platyrrhineLCAs.� In the crown anthropoid LCA, the coronoid process of the
mandible is close to the level of the mandibular condyle,whereas in stem anthropoids, the tip of the coronoid processprojects high above the mandibular condyle (Character 82).� The crown anthropoid LCA has M1e2 trigonids and talo-
nids of similar height (Character 164), whereas stem an-thropoids have the molar trigonids raised above talonids.� In the stem anthropoids Proteopithecus and Catopithecus,
the M1 trigonids are open lingually (Character 166). Thecrown platyrrhine and catarrhine LCAs have the M1 trigo-nid enclosed lingually by a premetacristid. The parapithe-cids Apidium and Simonsius are derived in having the M1
trigonid enclosed lingually; however, more primitive para-pithecids such as Qatrania exhibit the stem anthropoid
condition, demonstrating that this character evolved ho-moplastically in parapithecids and crown anthropoids.� The P3s of the stem anthropoids Proteopithecus and Cato-
pithecus have concave, ‘‘waisted’’ distal crown margins(Character 233). Crown anthropoids have P3 crownswith straight or slightly convex distal margins. The parapi-thecids Apidium and Simonsius homoplastically have P3
crowns with straight or slightly convex distal margins.� In the stem anthropoids Proteopithecus and Catopithecus,
the buccal cingulum of P3e4 is well developed (Character236). In the crown catarrhine and crown platyrrhine LCAs,the buccal cingulum is reduced or absent. The parapithe-cids Apidium and Simonsius homoplastically have weaklydeveloped buccal cingula.
Aegyptopithecus is a stem catarrhine. Aegyptopithecus ex-hibits an interesting mixture of crown-catarrhine traits andprimitive retentions suggesting that it is a stem catarrhine. Ae-gyptopithecus and the crown catarrhine LCA (node 3 inFig. 22) share thirteen derived character states relative tostem anthropoids and the crown anthropoid LCA:
� Aegyptopithecus and the crown catarrhine LCA have par-allel-sided tooth rows (Character 25). In stem anthro-poids, stem platyrrhines, and the crown platyrrhine LCA,the postcanine tooth rows diverge posteriorly.� Aegyptopithecus and the crown catarrhine LCA have pro-
nounced medial pterygoid plates (Character 27). Stemanthropoids, stem platyrrhines, and the crown platyrrhineLCA have an abbreviated medial pterygoid plate. The con-dition is unknown in Dolichocebus. Tremacebus has a re-duced medial pterygoid plate. (Examination of micro-CTsof the skull of Tremacebus shows that an apparentlyenlarged medial pterygoid plate is reconstructed fromplaster.)� In Aegyptopithecus and the crown catarrhine LCA, the ca-
rotid foramen is positioned ventrally on the bulla (Charac-ter 57), whereas the canal enters ventromedially in stemanthropoids, the stem platyrrhines Dolichocebus and Trem-acebus, and nodal crown platyrrhines.� In Aegyptopithecus and the crown-catarrhine node, the ten-
torium cerebelli has an unossified petrosal root (Charac-ter 74), whereas in stem anthropoids, the tentorium ispartially ossified (Kay et al., 2008). The stem platyrrhinesDolichocebus and Tremacebus and the crown platyrrhineLCA share the stem-anthropoid condition.� Aegyptopithecus, crown catarrhines, and, convergently,
Catopithecus have a reduced number of premolars, fromthree to two (Character 111). Stem anthropoids, stemplatyrrhines, and crown platyrrhines retain P2.� Aegyptopithecus and crown catarrhines have an enlarged
P3 relative to P4, with its prototoconid projecting wellabove that of P4 (Characters 144, 145). Stem anthro-poids, including oligopithecids with two premolars, aremore primitive in having similar projective heights ofthe P3 and P4 protoconids, with P3 not enlarged relativeto P4. Branisella, Dolichocebus, Carlocebus, and the

356 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
crown platyrrhine LCA share these stem-anthropoidconditions.� Aegyptopithecus and all crown catarrhines exhibit a derived
condition of having M1e2 protocristids deflected distallyinto the hypoflexid and in having a distinct M1e2 PhaseII wear facet X (Characters 169, 172, 173). In stem an-thropoids, stem platyrrhines (Dolichocebus, Branisella,Carlocebus, and Soriacebus), and the crown platyrrhineLCA, the M1e2 protocristids run towards the metaconidsand M1e2 Phase II wear facet X is absent.� Aegyptopithecus and the crown catarrhine LCA share the
derived condition in having broad, open sulci betweenthe M1e2 entoconids and hypoconulids (Character 175).Stem anthropoids and the stem platyrrhines Branisella,Dolichocebus, Carlocebus, and Soriacebus have weakerand less distinct postentoconid sulci on M1e2. The latteris a more primitive condition than that inferred for thecrown platyrrhine LCA. At the latter node, the postentoco-nid sulcus is absent.� Aegyptopithecus and its smaller relatives Propliopithecus
and Moeropithecus are similar to the crown catarrhineLCA in being larger in body size (Character 196).� In Aegyptopithecus and crown catarrhines, the upper ca-
nine occludes with P3 (Character 213). In most stem an-thropoids, the stem platyrrhines Branisella, Carlocebus,Dolichocebus, and Soriacebus, and all crown platyrrhines,the upper canine occludes with P2.
We identify three craniodental features where Aegyptopi-thecus still retains the stem anthropoid condition, suggestingthat it is a stem rather than a crown catarrhine:
� All crown catarrhines have expanded, collar-shaped ecto-tympanics (Character 66), whereas the ectotympanic isannular in stem anthropoids, stem platyrrhines, and allcrown platyrrhines. Aegyptopithecus retains the primitiveannular condition.� In crown catarrhines, Cartmill’s venous canal is absent
(Character 75). The stem catarrhine Aegyptopithecushas an intermediate condition in which the canal isblind-ended or incomplete (Kay et al., 2008). In stem an-thropoids, stem platyrrhines, and all crown platyrrhines(Kay et al., 2008) the canal is open. This is another casewhere Aegyptopithecus is less derived than crown catar-rhines, consistent with it being a stem catarrhine.� The crown catarrhine and platyrrhine LCAs both have rel-
atively large endocranial volumes (Character 77), but asnoted, this is clearly an independent evolutionary eventbecause the stem catarrhine Aegyptopithecus has a com-paratively small endocranial volume (Simons et al.,2007).
Branisella, Dolichocebus, Tremacebus, Carlocebus, andSoriacebus are stem platyrrhines. A number of cranial and den-tal characters suggest that late Oligocene Branisella and earlyMiocene Dolichocebus, Tremacebus, Carlocebus, and Soriace-bus branched from the platyrrhine stem before the appearance
of the last common ancestor of living platyrrhines (the lineagesegment between nodes 1 and 2 in Fig. 22). This hypothesisgains support from the presence in these taxa of retainedstem-anthropoid character states found in combination withsome derived crown-platyrrhine character states. EarlyMiocene platyrrhines share six derived features with the crownplatyrrhine LCA.
� The stem platyrrhines Branisella, Dolichocebus, Carloce-bus, and Soriacebus share the derived platyrrhine condi-tion of having single-rooted P3s and P4s (Character 113).� The mesiodistally short, buccolingually broad P4 of the
crown platyrrhine LCA (Character 146) is also found inDolichocebus, Soriacebus, and Branisella.� The crown platyrrhine LCA had the derived condition
where the M1e2 cristid obliqua extends onto the posteriorwall of the trigonid, reaching towards the protoconid(Characters 183, 184). Dolichocebus, Branisella, Carlo-cebus, and Soriacebus share this derived condition.� The crown platyrrhine LCA has two-rooted P3s and P4s
(Characters 217, 218). Branisella, Carlocebus, Tremace-bus, and Dolichocebus also have the typical platyrrhinetwo-rooted third and fourth premolars. Soriacebus hasthree-rooted P3s and P4s.
At the same time, late Oligocene and early Miocene platyr-rhines depart from the crown platyrrhine LCA, and are morelike stem anthropoids, in retaining nine primitive characterstates:
� In the crown platyrrhine LCA, the zygomatic and temporalbones contact at pterion, covering the contact between thethe frontal and the alisphenoid (Character 9). Thearrangement of these bones cannot be determined forDolichocebus, but in Tremacebus, based upon our exami-nation of the type skull, and reportedly in another latestmiddle Miocene platyrrhine, Homunculus (Tauber, 1991),confirmed by personal observations, the primitive fron-tal-alisphenoid contact is retained.� A derived condition in the crown platyrrhine LCA is for
the infraorbital foramen to be positioned quite far forwardrelative to the cheek teeth (Character 14). Dolichocebusshares the derived crown-platyrrhine condition. HoweverTremacebus and Branisella have a more posteriorly posi-tioned infraorbital foramen.� In stem anthropoids Apidium, Simonsius, Proteopithecus,
and Catopithecus, the orbital convergence is less than65� (Character 43), whereas in the crown platyrrhineand catarrhine LCAs, the orbital convergence is less than65� (Ross, 1995).� Overall brain size (endocranial volume) cannot be estab-
lished for Catopithecus or Proteopithecus, but it issmalldin the strepsirrhine rangedin Simonsius (Charac-ter 77). The crown catarrhine and platyrrhine LCAs bothhave relatively large endocranial volumes, but as noted,this apparently shared derived state of the two crown groupsevolved independently, as evidenced by the relatively small

357R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
endocranial volume in the stem catarrhine Aegyptopithecus.There also is evidence that the stem platyrrhine Homunculusalso had a relatively small endocranial volume (Kay et al.,2006).� The derived enlarged P3 metaconid of the crown platyr-
rhine LCA (Character 122) is not observed in Dolichoce-bus, Branisella, Carlocebus, or Soriacebus, which havesmall but distinct P3 metaconidsdintermediate betweenthe stem anthropoid condition and the derived conditionof the crown platyrrhine LCA.� The derived transversely oriented protocristid on the P3 of
crown platyrrhines (Character 128) is not seen in Doli-chocebus, Carlocebus, or Soriacebus, which have distolin-gually oriented P3 protocristids. However, Branisella hasindependently gained (or a Dolichocebus, Carlocebus,and Soriacebus clade has lost) the derived platyrrhinecondition.� The derived condition of the crown platyrrhine LCA of
possessing a single-rooted M3 without a third lobe (Char-acters 152, 162) is not seen in stem platyrrhines. Dolicho-cebus, Carlocebus, and Soriacebus have a single M3 rootwith a figure-eight cross section. A small M3 third lobewith a small hypoconulid is present in Branisella, Carlo-cebus, and Soriacebus. The surface anatomy of the M3
cannot be evaluated in the available Dolichocebusspecimens.� The postentoconid sulcus is derivedly absent in the crown
platyrrhine LCA (Character 175). The stem platyrrhinesBranisella, Dolichocebus, Carlocebus, and Soriacebus stillretain a distinct M1e2 postentoconid sulcus.� The derived absence of an M1e2 hypoconulid in the crown
platyrrhine LCA (Characters 176, 177) differs froma more primitive condition in stem platyrrhines. Branisellahas a hypoconulid comparable in size to those of stem an-thropoids. Dolichocebus, Carlocebus, and Soriacebus pos-sess greatly reduced hypoconulidsdmore derived than inBranisella but less advanced than in crown platyrrhines.
Evidence for cladogenesis among stem platyrrhines. Amongthe taxa examined here, there is some evidence, although notas yet compelling, that Branisella branched first from thestem-platyrrhine lineage segment, followed by a southern orPatagonian clade consisting of Dolichocebus, Tremacebus,Carlocebus, and Soriacebus. Two features suggest that Brani-sella was first to branch from the stem-platyrrhine lineagesegment:
� Dolichocebus, Carlocebus, and Soriacebus are derived inhaving distolingually oriented P4 protocristids (Character130). By contast, the stem anthropoids Proteopithecus andCatopithecus, the stem platyrrhine Branisella, and thecrown platyrrhine LCA have transversely oriented proto-cristids on P4.� Dolichocebus, Carlocebus, and Soriacebus possess the de-
rived mesiodistally reduced P4 talonid of the crown platyr-rhine LCA (Character 139). However, Branisella is moreprimitive in retaining the primitive elongate P4 talonid.
Two additional features suggest that the Patagonian taxaform their own clade:
� The crown anthropoid LCA has marginally placed pre-molar cusps (Character 137), as do Branisella and thecrown platyrrhine and catarrhine LCAs. In contrast,Dolichocebus, Carlocebus, and Soriacebus have basallyinflated P4s.� Carlocebus, Dolichocebus, and Soriacebus are derived in
having strongly developed P4 hypocones. By contrast,crown platyrrhines, stem anthropoids, and stem and crowncatarrhines lack or have very weakly developed P4 hypo-cones (Character 228). Branisella likewise lacks a P4
hypocone.
Contrasting views of platyrrhine evolution
The implications of the phylogenetic analyses are obviouswith respect to the choice between the ‘‘deep time’’ versus ‘‘lay-ered’’ hypotheses of platyrrhine evolution. Our examination ofthe anatomy of Dolichocebus and our broader phylogeneticanalysis supports the hypothesis that Branisella, Dolichocebus,Tremacebus, Carlocebus, and Soriacebus are stem platyrrhines.Branisella exhibits some dental similarities with callitrichines(Takai et al., 2000) and other crown platyrrhines, but it also pos-sesses a number of similarities with stem anthropoids (e.g., thelarge, lingually placed hypoconulid). Autapomorphies of thedentition (high-crowned teeth that were worn rapidly) suggestthat Branisella occupied a possibly terrestrial or scansorialadaptive niche, which are not found among living platyrrhines(Kay et al., 2001). Dolichocebus is not specially related to Sai-miri, nor is it a cebine. Indeed, the proposed similarities betweenDolichocebus and cebines either do not exist (e.g., dolichoceph-aly; see above) or are not established by the fossils (e.g., inter-orbital fenestration and the structure of glabella; the structureof the nasals and interorbital process of the frontal cannot be de-termined for Dolichocebus). Tremacebus, other than havingsomewhat enlarged orbits, shows few derived similarities toAotusdthe orbits are divergent, as as in basal cebids, the olfac-tory lobes are not as enlarged as in Aotus, the palate is shallow(vs. deep), and the tooth rows diverge posteriorly (Hershkovitz,1974; Kay et al., 2004a). Soriacebus exhibits dental and gnathicsimilarities to the most specialized pitheciines (mesiodistallycompressed and procumbent incisors, robust canines, low-crowned molars with poorly developed shearing crests, anda deep jaw with an apparently expanded angle), but many detailsof the dental structure are more primitive, and there are a numberof ‘‘crossing-specializations’’ (Simpson, 1961: 102) that dem-onstrate an independent entry into the seed-predation niche cur-rently occupied by extant Cebus and the Pitheciinae (Kay, 1990;Meldrum and Kay, 1997; Kay et al., 1998a). Thus, all four ofthese late Oligocene and early Miocene taxa possess a combina-tion of plesiomorphic character states that more closely resem-ble the nearest platyrrhine outgroupsdAfrican late EoceneCatopithecus and Proteopithecus and Oligocene-to-Recentcatarrhinesdwhile showing distinctive platyrrhine synapomor-phies. Such a combination of primitive traits and platyrrhine

358 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
synapomorphies is precisely what would be expected in a clusterof stem taxa.
The ‘‘deep time’’ hypothesis posits that most or all of theplatyrrhine family clades extend back at least 20 million yearsor earlier. However, this hypothesis conflates adaptation withphylogeny. For example, it is argued that nocturnality is suchan unusual adaptive pattern for an anthropoid that it couldonly have evolved once in platyrrhine evolution. Tremacebus,like Aotus, has enlarged orbits and must have been nocturnal.Therefore, according to the ‘‘deep time’’ hypothesis, Tremace-bus must be an aotine. Likewise, it is argued that seed predationand seed-pod husking is such an unusual adaptive pattern that itcould only have evolved once in platyrrhine evolution. Thus,because Soriacebus, like pitheciines, is adapted to seed-huskingand seed predation, it must be a pitheciine.
Rosenberger (2002) framed the debate in terms of parsi-mony. He rejected the stem-group or separate-clade phyloge-nies, stating that ‘‘[these hypotheses] require a high degreeof parallel or convergent evolution’’ (Rosenberger, 2002:158). We agree. The ‘‘layered’’ hypothesis supported heredoes require substantial homoplasy, but not as much as the‘‘deep time’’ hypothesis, which is why we prefer our interpre-tation over hisdboth entail a substantial amount of homo-plasy, but the ‘‘layered’’ hypothesis calls for less.
That so much parallel evolution could have occurred in theplatyrrhine clade is quite plausible based upon what we knowabout primate evolution generally. Homoplasy is a very com-mon phenomenon in primate evolution (Lockwood and Fleagle,1999). We find numerous examples of multiple independent oc-cupations of adaptive niches among primates (Heesy and Ross,2004). Eocene North American Omomyidae show repeated in-dependent acquisition of enlarged lower central incisors asfood-acquisition ‘‘tools’’ (Williams and Covert, 1994). Noc-turnality and enlarged orbits occurred independently amongseveral Malagasy strepsirrhines, as well as in Eocene omomyidsand adapids. Indeed, among the extant platyrrhines, prehensiletails evolved twicedin atelids and Cebus (Rosenberger,1983)dand so did seed predation (Cebus and pitheciines).Moreover, folivory evolved at least twice, once in Brachytelesand once in Alouatta. Why then should it be difficult to acceptthe proposition that seed predation occurred independently inpitheciines and Soriacebus or that nocturnality, or at least cre-puscularity, evolved independently in Aotus and Tremacebus?
Our phylogenetic analysis suggests that some extinct cladesof platrrhines independently occupied similar adaptive nichesto those occupied by crown platyrrhines. As Hershkovitz(1977) observed, there was no single adaptive radiation ofplatyrrhines. What we see today is a ‘‘layered’’ pattern of spe-cializations, the consequence of independent evolution at dif-ferent times over a long platyrrhine career on the SouthAmerican continent. Molecular-branch times for the livingplatyrrhine families support the ‘‘layered’’ model. Pitheciineseed predators came on the scene at least three million yearslater than the nocturnal ancestors of Aotus and at least six mil-lion years earlier than the origin of gum feeding in Callithrixand Cebuella (Schneider et al., 2001). The fossil evidence alsosuggests that extinct platyrrhines may have occupied niche
space not present in the living representatives of the clade.For example, there is suggestive evidence that Branisellawas scansorial (see above). Acrecebus, a poorly known cebinefrom the late Miocene of the Amazon Basin, may have at-tained the size of a small chimpanzee (Kay and Cozzuol,2006).
Summary and conclusions
Dolichocebus gaimanensis comes from deposits of the Sar-miento Formation in Chubut Province, Argentina, near thetown of Gaiman. The geology of the Sarmiento Formation inthe type locality was reviewed and a scheme of subdivisionsof the formation presented for that place. All specimens of Do-lichocebus come from the Trelew Member. This primate andits associated mammalian fauna date to the early Mioce-nedabout 20 Madand are attributed to the ColhuehuapianSouth American Land Mammal Age.
All Dolichocebus fossil material was described using con-ventional surface anatomy and micro-CT data from the cra-nium. The new material and newly imaged internal anatomyof the skull demonstrate that most anatomical characters hith-erto supposed to support a phyletic link between Dolichocebusand either callitrichines or squirrel monkeys are either indeter-minate or absent.
A set of phylogenetic analyses using parsimony was under-taken on Dolichocebus, several fossil platyrrhines, all livingplatyrrhine genera, and a suite of better-known living and fos-sil anthropoid outgroups. All parsimony results point to theconclusion that Dolichocebus is a stem platyrrhine, as arelate Oligocene Branisella and early Miocene Tremacebus, Sor-iacebus, and Carlocebus. The outcome of these analyses alsohighlights the pervasiveness of homoplasy in dental and cra-nial characters. This is not surprising because large amountsof homoplasy are present in previous analyses at all levelsof biological organization, from DNA to aspects of adultbony morphology and behavior.
Platyrrhine evolution often is conceived of as a single, an-cient adaptive radiation. Our phylogenetic investigationsuggests a very different pattern, wherein several extinct platyr-rhine clades independently occupied adaptive niches similar tothose occupied by crown platyrrhines. Simply stated, therewas no single adaptive radiation of platyrrhines. What we seein today’s platyrrhine assemblage is the latest in a series of radi-ations over a long platyrrhine career in the Neotropics. Reviewof our own and other available phyolgenetic data suggestsa more layered evolutionary pattern. Several independent ex-tinct clades filled modern platyrrhine niche space in the Oligo-cene and early Miocene. Modern platyrrhine families andsubfamilies appear subsequently over a nine-million-year inter-val in the Miocene.
Acknowledgements
We benefited greatly from discussions with Chris Wall,James Rossie, Walter Hartwig, and input from the editor andseveral unidentified reviewers. We thank the curators at the

359R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Museo Nacional de Ciencias Naturales, Buenos Aires, and theMuseo de La Plata for access to material, and we acknowledgethe contribution of the personnel from these museums to var-ious field projects over the past 25 years. RFK especiallythanks Richard Madden, Guiomar Vucetich, and Alfredo Car-lini for help and inspiration in the laboratory and on the out-crop. This research was supported by grants to JGF andRFK by the National Science Foundation and the NationalGeographic Society. The most recent grant is NSF BCS Award0090255 to RFK.
Appendix 1: Cranial and dental characters andcharacter states used in the phylogentic analyses
Characters followed by an asterisk (*) are ‘‘ordered.’’ Or-dered characters are scaled by the number of character states,such that the sum of the steps in the morphocline equals 100.This weight is indicated as ‘‘weight¼ number.’’ A score of‘‘9’’ is used if information is unavailable due to a lack of ma-terial or if the character does not apply to a particular taxon.Cranial characters adopted from Horovitz (1999) and Kayet al. (2004c) are so designated. Many of these cranial charac-ters ultimately are drawn from the following sources: Forsyth-Major (1901), Pocock (1925), Hill (1957), Le Gros Clark(1959), Du Brul (1965), Hershkovitz (1974, 1977), Cartmill(1978), Rosenberger (1979b), Conroy (1981), MacPhee andCartmill (1986), Ross (1993, 1994, 1998), Kay (1994), Mac-Phee et al. (1995), Horovitz (1997, 1999), Kay et al. (1997,2004c), Horovitz and MacPhee (1999), Kay and Kirk(2000), and Lieberman et al. (2000). Dental characters desig-nated with an ‘‘a’’ suffix are new characters. Other dental char-acters are as in previous analyses (Kay et al., 2004c).
Cranial characters
Paranasal sinuses1. (Weight¼ 100). Ethmofrontal sinus (Type II): 0¼
present; 1¼ absent.2. (Weight¼ 100). Splenofrontal sinus (Type I): 0¼
present; 1¼ absent.3. (Weight¼ 100). Maxillary sinus: 0¼ present; 1¼ absent.4. (Weight¼ 100). Anterior ethmiodal sinus: 0¼ present;
1¼ absent.5. (Weight¼ 100). Sphenoidal sinus: 0¼ present; 1¼
absent.
Zygomatic region6. * (Weight¼ 50). Zygomaticofacial foramen (Horovitz
[1999] character 27): 0¼ small relative to M1 breadth;1¼ large relative to M1 breadth; 2¼ very large relativeto M1 breadth.
7. (Weight¼ 100). Zygomatic arch position (Horovitz[1999] character 23): 0¼ above the alveolar border ofthe maxilla; 1¼ below the alveolar border.
8. (Weight¼ 100). Extent of inferior orbital fissure (Horo-vitz [1999] character 84): 0¼ ventrolateral limit of the
inferior orbital fissure does not reach the zygomaticarch; 1¼ the ventrolateral limit of the inferior orbital fis-sure reaches the zygomatic arch.
9. (Unordered; weight¼ 100). Zygomaticoparietal contactat pterion (Kay et al. [2004c] cranial character 46):0¼ no postorbital closure; 1¼ zygomatic-parietal con-tact; 2¼ alisphenoid-frontal contact.
Lacrimal region10. (Weight¼ 100). Position of lacrimal foramen (Kay et al.
[2004c] cranial character 30): 0¼ outside orbital margin;1¼within the orbit or on the rim.
11. * (Weight¼ 50). Extraorbital exposure of the lacrimal:0¼ lacrimal fossa is completed anteriorly by maxilla;1¼ lacrimal has some facial exposure; 2¼ lacrimal con-tacts nasal (excludes maxilla-frontal contact).
12. (Weight¼ 100). Zygomatic-lacrimal contact (Kay et al.[2004c] cranial character 26): 0¼ present on ventral or-bital rim; 1¼ absent on ventral orbital rim.
13. (Unordered; weight¼ 100). Contact between lacrimaland palatine bones (Kay et al. [2004c] cranial character28): 0¼ contact present; 1¼ lacrimal and palatine sepa-rated, contact between frontal and maxilla (or in sometaxa, by a small os planum of the ethmoid); 2¼ separatedby a large os planum.
Facial region14. * (Weight¼ 50). Position of the infraorbital foramen rel-
ative to the Frankfurt horizontal plane (Horovitz [1999]character 26): 0¼ posterior to P4; 1¼ positioned aboveP3e4; 2¼ positioned above P2.
15. * (Weight¼ 50). Angle of cranial kyphosis (Liebermanet al., 2000): 0¼�140�; 1¼>140�;<155 degrees;2¼�155�.
16. (Weight¼ 100). Nasal fossa width (Horovitz [1999]character 25): 0¼ narrower than the palate width; 1¼broader than the palate width.
17. (Weight¼ 100). Nasal capsule (Maier, 1980): 0¼processus alaris superior present; 1¼ processus alaris su-perior absent.
18. (Weight¼ 100). Snout length (Kay et al. [2004c] cranialcharacter 37): 0¼ long snout; 1¼ short snout.
19. (Weight¼ 100). Maxilla depth (Kay et al. [2004c] cranialcharacter 38): 0¼ deep; 1¼ shallow.
20. (Weight¼ 100). Interincisor diastema width (Kay et al.[2004c] cranial character 42): 0¼ broad and wider thanthat of extant haplorhines; 1¼ narrow, haplorhine-like.
21. (Weight¼ 100). Ascending wing of premaxilla (Kay et al.[2004c] cranial character 49): 0¼ narrow; 1¼ broad.
Temporomandibular region22. * (Weight¼ 50). Postglenoid foramen (Horovitz [1999]
character 12): 0¼ absent; 1¼ small; 2¼ large.23. (Weight¼ 100). Temporomandibular joint morphology
(Kay et al. [2004c] cranial character 40): 0¼ biconcave

360 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
and transversely wide; 1¼ anteroposteriorly orientedtrough.
24. * (Weight¼ 50). Postglenoid process size (100�postglenoid process length/prosthioneinion length) (Kayet al. [2004c] cranial character 41): 0¼weak or absent(<0.39); 1¼ strong (�0.39, <0.69); 2¼ very strong(�0.69).
Pterygoid and palatal region25. * (Weight¼ 50). Palate shape: 0¼V-shaped (the distance
between lingual surfaces of the upper canines divided bythe distance between the lingual surfaces of the uppersecond molars is <0.39); 1¼ intermediate (ratio valuesof �0.39, � 0.64); 2¼ approaches parallel (ratiovalues> 0.64).
26. (Weight¼ 100). Interpterygoid fossa (Du Brul, 1965):0¼ deep; 1¼ shallow.
27. * (Weight¼ 50). Length of medial pterygoid plate (Kayet al. [2004c] cranial character 36): 0¼ long medial pter-ygoid plate extending one-third to one-half of the dis-tance to the anterior surface of the bulla enclosinga large fossa between medial and lateral pterygoids;1¼ short but distinct from lateral pterygoid plate for itsentire dorsoventral extentdventrally there is a hamularprocess, and more dorsally the plate merges with the lat-eral plate or if distinct, the fossa is slitlike; 2¼medialpterygoid plate entirely absent or reduced to a low rugo-sitydonly the hamulus is present.
28. (Unordered; weight¼ 100). Encroachment of the audi-tory bulla on the pterygoid fossa (Kay et al. [2004c]cranial character 17): 0¼ no encroachment; 1¼encroachment by the anterior accessory cavity; 2¼present and formed by the tympanic cavity.
29. (Unordered; weight¼ 100). Nature of contact betweenthe lateral pterygoid plate and the bullar wall (Kayet al. [2004c] cranial character 18): 0¼ absent; 1¼laminar; 2¼ abutting.
30. (Weight¼ 100). Extent of contact between the lateralpterygoid plate and the bullar wall (Kay et al. [2004c]cranial character 19): 0¼ slight; 1¼ very extensive.
31. (Weight¼ 100). Pyramidal process of palate and postal-veolar notch: 0¼ no postalveolar notch between the pyra-midal process and the maxillary tuberosity; 1¼ offset frommaxillary tuberosity by a distinct postalveolar notch.
32. * (Weight¼ 50). Mediolateral position of pyramidal pro-cesses (100 times the ratio of interpyramidal breadth toouter M1 palate breadth) (Kay et al. [2004c] cranial char-acter 35): 0¼medially placed (�43); 1¼ intermediate(>43, �64) 2¼ laterally placed (>64).
33. (Weight¼ 100). Posterior palatine torus (Kay et al.[2004c] cranial character 34): 0¼ present; 1¼ absent.
34. * (Weight¼ 50). Posterior nasal spine (Kay et al. (2004c)cranial character 33): 0¼ reduced or absent; 1¼ smallbut distinct; 2¼ robust and long.
35. (Weight¼ 100). Posterior extent of the turbinates:0¼ extend posterior to the palatine; 1¼ completely ante-rior to the palatine.
36. (Weight¼ 100). Angle of the incisive canal in palate:0¼ obliquely oriented with respect to the plane of thepalate; 1¼more closely resembles a right angle with thepalate.
Temporal fossa37. (Weight¼ 100). Temporal emissary foramen (Horovitz
[1999] character 20): 0¼ present and large; 1¼ smallor absent.
Nucal region38. * (Weight¼ 50). Paroccipital processes: 0¼ forms a dis-
tinct shelf or process; 1¼ forms a raised ridge; 2¼weakor absent.
39. (Weight¼ 100). Pneumatization of mastoid (Kay et al.[2004c] cranial character 3): 0¼ absent; 1¼ present.
Orbital region40. * (Weight¼ 50). Lateral cranial profile at glabella:
0¼ depressed; 1¼ flat; 2¼ convex.41. (Weight¼ 100). Interorbital fenestra: 0¼ absent; 1¼
present.42. * (Weight¼ 50). Size of orbits (Kay et al. [2004c] cranial
character 23): 0¼ small; 1¼ large; 2¼ extremely large.43. (Weight¼ 50). Orbital convergence (Ross, 1995):
0¼ less than 55�; 1¼ 55�e65�, 2¼ greater than 65�.44. * (Weight¼ 50). Interorbital breadth: 0¼ narrow; 1¼
broad; 2¼ extremely broad.45. * (Weight¼ 100). Exposure of vomer in orbit: 0¼
unexposed; 1¼ exposed.46. (Weight¼ 100). Postorbital closure (Kay et al. [2004c]
cranial character 24): 0¼ none; 1¼ postorbital bar pres-ent; 2¼ postorbital septum present.
47. (Weight¼ 100). Composition of the postorbital septum(Kay et al. [2004c] cranial character 25): 0¼ zygomaticforms most of the septum; 1¼ frontal forms most ofthe septum.
48. (Weight¼ 100). Position of interorbital constriction rela-tive to olfactory tract (Kay et al. [2004c] cranial character27): 0¼ absent; 1¼ present below olfactory tract.
49. (Weight¼ 100). Foramen rotundum (Kay et al. [2004c]cranial character 29): 0¼ superior orbital fissure trans-mits maxillary nerve; 1¼ separate foramen (f. rotundum)for maxillary nerve.
50. (Weight¼ 100). Metopic suture in adult (Kay et al.[2004c] cranial character 31): 0¼ unfused; 1¼ fused.
Ear region51. (Weight¼ 100). Cochlear housing as exposed in middle
ear (Horovitz [1999] character 15): 0¼ singular; 1¼ dual.52. (Weight¼ 100). Transbullar septa (Horovitz [1999] char-
acter 14): 0¼middle ear’s ventrolateral region withoutsepta; 1¼ anteroventral region with septa.
53. (Unordered; weight¼ 100). Transverse septum arisingfrom the cochlear housing (Kay et al. [2004c] cranialcharacter 1): 0¼ absent; 1¼ present and forming the lat-eral wall of an anterior accessory cavity pneumatized

361R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
from the tympanic cavity; 2¼ present and forming thelateral wall of an anterior accessory cavity pneumatizedfrom the epitympanic recess.
54. (Weight¼ 100). Extent of pneumatization of anterior ac-cessory cavity (Kay et al. [2004c] cranial character 2):0¼ anterior accessory cavity lies anterior to the tympaniccavity and is not trabeculated; 1¼ anterior accessorycavity extends medial to the tympanic cavity, and istrabeculated.
55. (Weight¼ 100). Presence or absence of perbullar path-way for the internal carotid artery (Kay et al. [2004c] cra-nial character 4): 0¼ absent; 1¼ present and formedexclusively by the petrosal bone.
56. (Weight¼ 100). Anteroposterior location of posterior ca-rotid foramen in bulla (Kay et al. [2004c] cranial charac-ter 5): 0¼ posterior to line joining midpoints of tympanicbones; 1¼ anterior to this line.
57. * (Weight¼ 50). Mediolateral position of posterior ca-rotid foramen in bulla (Kay et al. [2004c] cranial charac-ter 6): 0¼medial; 1¼midline of the bulla; 2¼ lateral.
58. (Weight¼ 100). Ventrodorsal position of the carotid fora-men in the bulla (Kay et al. [2004c] cranial character 7):0¼ dorsal, adjacent to basioccipital or mastoid bone;1¼ ventral.
59. (Weight¼ 100). Position of posterior carotid foramen rel-ative to fenestra cochleae (Kay et al. [2004c] cranialcharacter 8): 0¼ posterior; 1¼ ventral; 2¼ anterior.
60. (Weight¼ 100). Position of the internal carotid canal rel-ative to the fenestra cochleae (Kay et al. [2004c] cranialcharacter 9): 0¼ runs across ventral lip of the fenestracochleae, shielding it from ventral view when a canal ispresent; 1¼ internal carotid canal does not shield the fe-nestra cochleae from ventral view.
61. (Weight¼ 100). Position of the portion of the internal ca-rotid/promontory artery (or its accompanying nerves) ly-ing on the promontorium anterior to the fenestra cochleae(Kay et al. [2004c] cranial character 10): 0¼ on ventro-lateral surface of promontorium; 1¼ contacting onlythe cupula of the cochlea.
62. (Unordered; weight¼ 100). Size of stapedial and prom-ontory canals (Kay et al. [2004c] cranial character 11):0¼ both stapedial and promontory canals are large;1¼ stapedial slightly smaller than promontory;2¼ stapedial highly reduced or absent altogether;3¼ stapedial larger than promontory; 4¼ both promon-tory and stapedial canals absent.
63. (Weight¼ 100). Morphology of promontory canal, whenpresent (Kay et al. [2004c] cranial character 12):0¼ open trough; 1¼ complete canal.
64. (Weight¼ 100). Canal for internal carotid artery ornerves (Kay et al. [2004c] cranial character 13):0¼ absent; 1¼ present.
65. (Weight¼ 100). Position of ventral edge of the tympanicbone (Kay et al. [2004c] cranial character 14):0¼ intrabullar, or aphaneric; 1¼ extrabullar or phaneric.
66. (Weight¼ 100). The shape of the tympanic bone (Kayet al. [2004c] cranial character 15): 0¼ ribbonlike or
only slightly expanded; 1¼ laterally expanded into a col-lar or tube.
67. (Unordered; weight¼ 100). Morphology of annularbridge (Kay et al. [2004c] cranial character 16): 0¼ lineasemicircularis or partial annular bridge formed on a ento-tympanic bulla; 1¼ Linea semicircularis formed on a pe-trosal bulla; 2¼ a complete annular bridge.
68. (Weight¼ 100). Flange of basioccipital overlapping me-dial bulla wall (Kay et al. [2004c] cranial character 20):0¼ absent or minimal; 1¼ extensive.
69. (Weight¼ 100). Basioccipital stem (Beard and McPhee,1994): 0¼ narrow; 1¼ broad.
70. (Weight¼ 100). Suprameatal foramen (Kay et al. [2004c]cranial character 21): 0¼ absent; 1¼ present, small, andin the posterior root of the zygomatic arch; 2¼ present,large, and above the external auditory meatus.
71. (Weight¼ 100). Patent parotic fissure (Kay et al. [2004c]cranial character 22): 0¼ present; 1¼ absent.
72. (Weight¼ 100). Enclosure of intratympanic portion ofthe facial nerve in a bony canal (Kay et al. [2004c] cra-nial character 47): 0¼ no canal, facial runs in a sulcus;1¼ bony canal present.
73. (Weight¼ 100). Epitympanic crest (Kay et al. [2004c]cranial character 48): 0¼ absent; 1¼ present.
Brain and internal cranial characters74. (Weight¼ 100). Tentorium cerebelli ossification (Horo-
vitz [1999] character 13): 0¼ absent; 1¼ present.75. (Weight¼ 100). Vascular canal connecting sigmoid sinus
with subarcuate fossa (Horovitz [1999] character 17):0¼ absent; 1¼ present.
76. * (Weight¼ 50). Size of olfactory bulbs: 0¼ large olfac-tory lobe; 1¼moderate olfactory lobe; 2¼ small olfac-tory lobe.
77. * (Weight¼ 50). Relative brain size: 0¼ small; 1¼large; 2¼ very large.
Mandible78. (Weight¼ 100). Symphyseal orientation: 0¼more hori-
zontal orientation of planum alveolare; 1¼more verti-cally oriented relative to planum alveolare.
79. * (Weight¼ 50). Lateral profile of mandible (ratio ofmandible depth [measured buccally] at P2 and M2):0¼ superior and inferior border of the mandibular corpusare essentially parallel from the premolar to the mandib-ular angle (�1.26); 1¼ the inferior border deepens pos-teriorly (>1.26, �1.72); 2¼ ‘‘hyperdeep’’ (>1.72).
80. (Weight¼ 100). Mandibular corpus depth (Kay et al.[2004c] cranial character 45): 0¼ shallow; 1¼ deep.
81. (Weight¼ 100). Symphyseal fusion in young adult (Kayet al. [2004c] cranial character 39): 0¼ absent; 1¼ present.
82. * (Weight¼ 50). Coronoid height relative to condyle:0¼ very far above condyle; 1¼ above the level of con-dyle; 2¼ slightly above or equal to coronoid.
83. * (Weight¼ 50). Condyle height relative to tooth row(Kay et al. [2004c] cranial character 44): 0¼ at level oftooth row; 1¼ slightly above; 2¼well above tooth row.

362 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
84. * (Weight¼ 50). Angle of the mandible: 0¼ hook-shaped angle; 1¼moderately expanded angle; 2¼extremely expanded angle.
85. (Weight¼ 100). Depth of the coronoid-condylar notch:0¼ deep; 1¼ shallow.
Lower Teeth
Incisors86. i1.* (Weight¼ 33). Lower incisor number: 0¼ three;
1¼ two; 2¼ one: I1 present, I2 absent; 3¼ lower incisorsabsent.
87. i2. (Weight¼ 100). Lower incisor occlusal arrangement:0¼ edges wear flat, producing an arcuate battery fromlateral perspective; 1¼ cusp tips staggered.
88. i3. (Weight¼ 100). Lower incisor crown spacing: 0¼ nospaces; 1¼ spaces present between crowns.
89. i4. (Weight¼ 100). I2-C diastema: 0¼ present; 1¼absent. Comment: The presence of a diastema is variablein platyrrhines and never exceeds in breadth the mesio-distal dimension of the I2.
90. i5.*(Weight¼ 50). I1e2 size (ratio of I1e2 area to M1
area): 0¼ very small (�0.69); 1¼moderately sized(�0.70, �1.07); 2¼ large (>1.07).
91. i6.* (Weight¼ 33). I1:I2 proportions (ratio of I1 area to I2
area): 0¼ I1 much smaller than I2 (<0.71); 1¼ I1 smallerthan I2 (�0.71, <0.78); 2¼ I1 almost as large as I2
(�0.78, <1.00); 3¼ I1 larger than I2 (�1.01).92. i7.* (Weight¼ 50). I1 crown width (spatulate incisors
only): 0¼ considerably wider (MD) than root (spatulate):1¼ narrow at apex, but still wider than root; 2¼ ‘‘styli-form’’ (crown apex approximately the same width asthe cervical margin).
93. i8. (Weight¼ 100). I2 crown cross-sectional shape (ratioof MD to BL): 0¼ rounded oval (�0.64); 1¼mesiodistally compressed (<0.64).
94. i9.* (Weight¼ 50). Lower incisor crown height (crownheights judged from cemento-enamel junction to crowntip on the buccal surface): 0¼ low-crowned; 1¼moderately high-crowned; 2¼ high-crowned.
95. i10. (Weight¼ 100). I1e2 crown buccal outline: 0¼ gentlycurved in lateral perspective; 1¼ acutely curved.
96. i11.* (Weight¼ 50). Lower incisor roots: 0¼ erect orvertical; 1¼ slightly procumbent; 2¼ very procumbent.
97. i12.* (Weight¼ 50). Lower incisor crowns: 0¼ erect orvertical; 1¼ procumbent; 2¼ very procumbent.
98. i13.* (Weight¼ 50). Tooth comb: 0¼ absent; 1¼withthree teeth; 2¼with two teeth.
99. i14. (Weight¼ 100). I1 crown shape: 0¼ spatulate; 1¼lanceolate, pointed.
100. i15. (Weight¼ 100). I2 heel development (a lingual swell-ing at the base of crown): 0¼ heel absent; 1¼ heel present.
101. i16. (Weight¼ 100). Incisor lingual enamel: 0¼welldeveloped; 1¼ poorly developed or absent.
102. i17.* (Weight¼ 50). Lower incisor lingual cingulum:0¼ absent to weak; 1¼ strong but incomplete; 2¼strong and complete.
103. i19.* (Weight¼ 50). I1 area to M1 area: 0¼ I1 very small(ratio� 0.32); 1¼moderately enlarged (>0.32, �0.40);2¼ very enlarged (> 0.40).
Canines104. c1.* (Weight¼ 33). Female canine size (area relative to mo-
lars): 0¼ very small (C/M1< 0.40); 1¼moderate (�0.4,<0.80); 2¼ large (�0.80, �1.20); 3¼ very large (�1.20)
105. c2.* (Weight¼ 50). Canine dimorphism (square root ofmale canine area divided by square root of female caninearea): 0¼ low (<1.07); 1¼moderate (�1.07, <1.17);2¼ high (�1.17).
106. c3. (Weight¼ 100). Canine cross-sectional shape:0¼ rounded oval (MD:BL >1.00, <1.90); 1¼MD compressed (ratio� 1.90).
107. c4. (Weight¼ 100). Canine lingual-crest development:0¼ rounded; 1¼ sharp.
108. c5. (not ordered; Weight¼ 100). Canine paracristid (notscored if species has canine incorporated into a toothcomb): 0¼ oblique to occlusal plane; 1¼ nearly horizon-tal to occlusal plane; 2¼ forms part of cropping mecha-nism with I1e2.
109. c6.* (Weight¼ 50). Canine height (females): 0¼ low,squat; 1¼ narrow, short; 2¼ tall, at or above tooth row.
Premolars110. p1. (Weight¼ 100). P1/P1: 0¼ present; 1¼ absent111. p2. (Weight¼ 100). P2/P2: 0¼ present; 1¼ absent.112. p3. (Weight¼ 100). P2 roots: 0¼ single; 1¼ double.113. p4.* (Weight¼ 50). P3e4 roots: 0¼ P3 single, P4 single;
1¼ P3 single, P4 double; 2¼ P3 double, P4 double.114. p5.* (Weight¼ 50). Premolar crowding (overlapping of
crowns): 0¼ no crowding; 1¼ slightly crowded; 2¼ verycrowded.
115. p6.* (Weight¼ 50). P3 paraconid: 0¼ large; 1¼ small;2¼ absent or extremely small.
116. p7.* (Weight¼ 50). P4 paraconid: 0¼ large; 1¼ small;2¼ absent or extremely small.
117. p9.* (Weight¼ 33). P4 paraconid position: 0¼mesial toprotoconid; 1¼mesiolingual, between protoconid andmetaconid; 2¼mesial to metaconid; widely spacedfrom metaconid; 3¼ twinned with metaconid.
118. p11.* (Weight¼ 50). P3e4 cristid obliqua: 0¼ absent;1¼weak; 2¼ strong.
119. p13. (Unordered; weight¼ 100). P2 protoconid height andshape: 0¼ slender, projects above protoconids of P3e4;1¼massive, projects above protoconids of P3e4; 2¼ notprojecting, in line with P3; 3¼ extremely short, shorterthan P3.
120. p14. (Weight¼ 100). P4 metaconid position: 0¼ close toprotoconid; 1¼widely spaced from protoconid.
121. p15.* (Weight¼ 50). P2 metaconid size: 0¼ absent ortrace; 1¼ small; 2¼ large.
122. p16.* (Weight¼ 50). P3 metaconid size: 0¼ absent ortrace; 1¼ small; 2¼ large.
123. p17.* (Weight¼ 50). P4 metaconid size: 0¼ absent ortrace; 1¼ small; 2¼ large, almost as tall as protoconid.

363R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
124. p18. (Weight¼ 100). P4 trigonid lingual wall: 0¼ basinclosed by a premetacristid; 1¼ open with premetacristidabsent or short.
125. p19.* (Weight¼ 50). P3 entoconid and lingual talonidcrest: 0¼ absent; 1¼ lingual talonid crest present butan entoconid does not stand out above it; 2¼ entoconidis a small, discrete cusp.
126. p20.* (Weight¼ 50). P4 entoconid and lingual talonidcrest: 0¼ absent; 1¼ lingual talonid crest present butan entoconid does not stand out above it; 2¼ entoconidis a small, discrete cusp.
127. p21. (Weight¼ 100). P4 lateral and medial protocristid:0¼ continuous between metaconid and protoconid;1¼ discontinuous between metaconid and protoconid.
128. p22. (Weight¼ 100). P3 lateral protocristid orientation:0¼ transversely oriented; 1¼ distolingually oriented.
129. p23a. (Weight¼ 100). P4 lateral protocristid: 0¼ present;1¼ absent.
130. p23. (Weight¼ 100). P4 lateral protocristid orientation:0¼ transversely oriented; 1¼ distolingually oriented.
131. p24. (Weight¼ 100). P3e4 posterior trigonid wall:0¼ complete (taxa without metaconids are assignedthis character state); 1¼ deeply notched
132. p25. (Weight¼ 100). P3e4 hypoconid size: 0¼ large;1¼ cristiform, small, or absent.
133. p26. (Weight¼ 100). P3e4 hypoconid (or distal terminus ofoblique cristid) position: 0¼ distal to protoconid; 1¼ distalto metaconid, or between protoconid and metaconid.
134. p26a. (Weight¼ 100). P4 talonid breadth: 0¼ narrow;1¼ broad.
135. p27. (Weight¼ 100). P4 hypocristid shearing develop-ment: 0¼weak or absent; 1¼ strong.
136. p28.* (Weight¼ 50). P2 buccal cingulum development:0¼ absent; 1¼ incomplete, broken at protoconid and hy-poconid; 2¼ complete.
137. p29. (Weight¼ 100). Lower premolar inflation: 0¼cusps marginal, not basally inflated; 1¼ crown surfacesconstricted, cusp margins sloping.
138. p30.* (Weight¼ 50). P4 exodaenodonty: 0¼ not exodaeno-dont; 1¼ slightly exodaenodont; 2¼ very exodaenodont.
139. p31.* (Weight¼ 33). P4 talonid length (ratio of midlineMD of trigonid to MD of talonid): 0¼ extremely shortor nonexistent (tri:tal� 1.61); 1¼ short (much shorterthan trigonid) (tri:tal� 1.27, <1.61); 2¼ equal orslightly shorter in length to trigonid (tri:tal� 0.92, <1.27); 3¼ talonid longer than trigonid (tri:tal< 0.91).
140. p34. (Weight¼ 100). P4 anterobuccal cingulum develop-ment: 0¼ absent or trace; 1¼ strong.
141. p36.* (Weight¼ 50). P4 postprotoconid ridge: 0¼weakor absent; 1¼ present; 2¼ very strong.
142. p37.* (Weight¼ 50). P4 postmetaconid ridge: 0¼weakor absent; 1¼moderate; 2¼ very strong.
143. p40.* (Weight¼ 50). P4 paraconid height: 0¼ low;1¼moderate; 2¼ high (nearly as high as protoconid).
144. p41.* (Weight¼ 50). P3e4 protoconid height: 0¼ P3
much lower than P4; 1¼ P3 equal or slightly lower thanP4 ; 2¼ P3 higher than P4 .
145. p42.* (Weight¼ 25). P3 to P4 area: 0¼ 0.45e0.59;1¼ 0.60e0.69; 2¼ 0.70e0.79; 3¼>0.80, <1.10; 4, �1.10.
146. p43.* (Weight¼ 20). P4 MD/BL: 0¼<0.95; 1¼�0.96,<1.14; 2¼�1.15, <1.20; 3¼�1.21, <1.35;4¼�1.36,<1.46; 5¼>1.47.
147. p44.* (Weight¼ 20). P4 to M1 area: 0¼<0.62;1¼�0.63, <0.72; 2¼�0.73, <0.82; 3¼�0.83,<0.92; 4¼�0.93, <1.02; 5¼>1.03.
148. p45. (unordered; weight¼ 100). P3e4 root orientation:0¼ P3e4 roots aligned mesiodistally;1¼ P3 root shiftedlaterally, P4 mesial root aligned mesiodistally; 2¼ P3
roots aligned mesiodistally, P4 mesial root shifted later-ally. Score as missing if roots are single.
Molars149. m1. (Weight¼ 100). M3/M3: 0¼ present; 1¼ absent.150. m2. (Weight¼ 100). M1 root number: 0¼ one; 1¼ two.151. m3. (Weight¼ 100). M2 root number: 0¼ one; 1¼ two.152. m4. (Weight¼ 100). M3 root number: 0¼ one; 1¼ two.153. m6.* (Weight¼ 50). M2 trigonid width (ratio of BL of
trigonid and talonid): 0¼much wider than talonid(�1.11); 1¼widths similar (<1.11, >0.90); 2¼muchnarrower than talonid (�0.90).
154. m6a. (Weight¼ 100). M1 trigonid length: 0¼M1 trigo-nid short on the lingual side; 1¼M1 elongate on the lin-gual side.
155. m7.* (Weight¼ 50). M3 trigonid width (based on relativeBL breadths): 0¼much wider than talonid (�1.20);1¼ trigonid and talonid widths similar (�1.20, >1.05);2¼ trigonid narrower than talonid (<1.05).
156. m8.* (Weight¼ 33). M1 paraconid position: 0¼mesialto protoconid; 1¼mesiolingual, between protoconidand metaconid; 2¼mesial to metaconid but widelyspaced from it; 3¼ twinned with metaconid.
157. m9.* (Weight¼ 33). M2 paraconid position: 0¼mesialto protoconid; 1¼mesiolingual, between protoconidand metaconid; 2¼mesial to metaconid but widelyspaced from it; 3¼ twinned with metaconid.
158. m10.* (Weight¼ 33). M3 paraconid position: 0¼mesialto protoconid; 1¼mesiolingual, between protoconid andmetaconid; 2¼mesial to metaconid but widely spacedfrom it; 3¼ twinned with metaconid.
159. m11. (Weight¼ 100). M1 parastylid: 0¼ absent; 1¼present.
160. m12.* (Weight¼ 50). Molar metastylids: 0¼ absent;1¼ small; 2¼ large.
161. m13. (Weight¼ 100). M3 hypoconulid: 0¼ single; 1¼double
162. m14.* (Weight¼ 50). M3 heel: 0¼ absent; 1¼ narrowerthan talonid; 2¼ approximately equal in width to talonid.
163. m15.* (Weight¼ 50). Molar enamel surface: 0¼ smooth;1¼ slightly crenulated; 2¼ highly crenulated.
164. m16.* (Weight¼ 50). M1 trigonid height (ratio of trigonidheight to talonid height): 0¼ higher than talonid (�1.20);1¼ slightly higher than talonid (�1.10, <1.20);2¼ trigonid and talonid of similar height (<1.10).

364 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
165. m17.* (Weight¼ 50). M1e2 cusp relief (ratio of protoco-nid height to hypoconid height, measured buccally):0¼ low (<1.20); 1¼moderate (�1.20, <1.35); 2¼ high(>1.35).
166. m18. (Weight¼ 100). M1 trigonid lingual configuration:0¼ open; 1¼ closed.
167. m19. (Weight¼ 100). M1 metaconid position: 0¼ lingualto protoconid; 1¼ slightly distolingual to protoconid.
168. m20.* (Weight¼ 50). M1e2 paraconid development:0¼ absent; 1¼ small; 2¼ large.
169. m21. (Weight¼ 100). M1e2 lateral protocristid orienta-tion: 0¼ runs toward metaconid; 1¼ runs towardhypoflexid.
170. m22. (unordered; weight¼ 100). M1 distal trigonid wall:0¼ complete; 1¼ deeply notched by protoconid/metaco-nid sulcus; 2¼medial and lateral protocristid do notmeet but no sulcus is discerned.
171. m23. (Unordered; weight¼ 100). M2 distal trigonid wall:0¼ complete; 1¼ deeply notched by a sulcus betweenprotoconid and metaconid; 2¼medial and lateral proto-cristid do not meet but no sulcus is present.
172. m24a. (Weight¼ 100). M1 wear facet X: 0¼ present;1¼ absent.
173. m24. (Weight¼ 100). M2 wear facet X: 0¼ present;1¼ absent.
174. m25.* (Weight¼ 50). M1e2 entoconid: 0¼ absent orvery low, 1¼ lower than metaconid; 2¼ large.
175. m26.* (Weight¼ 50). M1e2 postentoconid sulcus:0¼ prominent; 1¼ shallow sulcus; 2¼ absent.
176. m27.* (Weight¼ 33). M1 hypoconulid size: 0¼ large;1¼moderate; 2¼ small; 3¼ absent.
177. m28* (Weight¼ 33). M2 hypoconulid size: 0¼ large;1¼moderate; 2¼ small; 3¼ absent.
178. m29.* (Weight¼ 33). M3 hypoconulid size: 0¼ large;1¼moderate; 2¼ small; 3¼ absent.
179. m30.* (Weight¼ 50). M1e2 hypoconulid position:0¼ twinned to entoconid; 1¼ near midline; 2¼ slightlybuccal to midline.
180. m31.* (Weight¼ 50). M1e2 cristid obliqua development:0¼weak (rounded); 1¼ strong (trenchant); 2¼ verystrong (trenchant).
181. m32.* (Weight¼ 50). M1 cristid obliqua orientation:0¼ reaches trigonid wall at a point distal to protoconid;1¼ reaches trigonid wall at a point distolingual to proto-conid; 2¼ reaches trigonid wall at a point distal tometaconid.
182. m33.* (Weight¼ 50). M2 cristid obliqua orientation:0¼ reaches trigonid wall at a point distal to protoconid;1¼ reaches trigonid wall at a point distolingual to proto-conid; 2¼ reaches trigonid wall at a point distal tometaconid.
183. m34. (unordered; weight¼ 100). M1 cristid obliqua ter-minus: 0¼ runs to base of trigonid; 1¼ runs part wayup the distal trigonid wall; 2¼ connects with protoconidtip or protocristid; 3¼ connects with metaconid.
184. m35. (unordered; weight¼ 100). M2 cristid obliqua ter-minus: 0¼ runs to base of trigonid; 1¼ runs part way
up the distal trigonid wall; 2¼ connects with protoconidtip or protocristid; 3¼ connects with metaconid.
185. m36. (unordered; weight¼ 100). M3 cristid obliqua ter-minus: 0¼ runs to base of trigonid; 1¼ runs part wayup the distal trigonid wall; 2¼ connects with protoconidtip or protocristid; 3¼ connects with metaconid.
186. m37. (Weight¼ 100). M1e2 centroconid development:0¼ present; 1¼ absent.
187. m38.* (Weight¼ 50). M1e2 hypocristid development:0¼ absent or seen only as a trace; 1¼weak; 2¼ strong.
188. m39.* (Weight¼ 50). M3 hypocristid development:0¼ absent or seen only as a trace; 1¼weak; 2¼ strong.
189. m40.* (Weight¼ 50). M1e2 talonid lingual configuration:0¼ open; 1¼ closed, notched lingually; 2¼ closed, nonotch.
190. m41. (Weight¼ 100). M1e2 distal fovea: 0¼ absent;1¼ present.
191. m44. (Weight¼ 100). Molar cusp inflation: 0¼ cusps notinflated, marginally positioned; 1¼ very inflated.
192. m45.* (Weight¼ 50). M1e2 buccal cingulum develop-ment: 0¼ absent to trace; 1¼ partial, broken at protoco-nid and hypoconid; 2¼ complete.
193. m46.* (Weight¼ 50). M1 hypoflexid depth: 0¼ veryshallow; 1¼moderate; 2¼ deep.
194. m47.* (Weight¼ 50). M2 hypoflexid depth: 0¼ veryshallow; 1¼moderate; 2¼ deep.
195. m53.* (Weight¼ 25). M2 length/M3 length: 0¼M3
much longer than M2 (0.71e0.80); 1¼M3 longer thanM2 (0.81e0.90); 2¼M3 equal to M2 (0.91e1.00);3¼M3 smaller than M2 (1.01e1.12); 4¼M3 muchsmaller than M2 (�1.13). Score as 5 if M3 is absent.
196. m54.* (Weight¼ 33). M1 length: 0¼<2.5 mm; 1¼�2.5,<3.8 mm; 2¼�3.8, �6.0 mm; 3¼�6.0.
197. m55.* (Weight¼ 33). M1 MD/BL: 0¼ 1.00e1.15;1¼ 1.16e1.22; 2¼ 1.23e1.32; 3¼>1.33.
198. m57. (Weight¼ 100). M1e2 entoconid position relative tohypoconid: 0¼ transverse to hypoconid; 1¼ distal tohypoconid.
Upper teeth
Incisors199. I1.* (Weight¼ 50). I1e2 interstitial contact: 0¼ absent;
teeth widely spaced; 1¼ present as narrow contact; 2¼ I2
tightly packed against I1, I1 preparacrista abbreviated.200. I2. (Weight¼ 100). I1-I1 interstitial contact: 0¼ present;
1¼ absent: a wide space occurs in the midline betweenthese teeth.
201. I3. (Weight¼ 100). I2-C diastema: 0¼ present; 1¼ absent.202. I4.* (Weight¼ 50). I1 area: I2 area: 0¼ areas approxi-
mately equal (�1.00); 1¼ I1 slightly larger than I2
(>1.00, <1.40); 2¼ I1 much larger than I2 (>1.40).203. I5.* (Weight¼ 50). I1 size (I1 area:M1 area): 0¼ small
(�0.50); 1¼moderate (>0.50,<0.56); 2¼ large (�0.56).204. I6.* (Weight¼ 50). I1 occlusal shape (MD/BL):
0¼ rounded oval (<1.05); 1¼ buccolingually compressed(>1.05, <1.30); 2¼ extremely compressed (>1.30).

365R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
205. I7.* (Weight¼ 50). I2 occlusal shape (MD/BL):0¼ rounded oval (�1.05); 1¼ slightly compressed(>1.05, <1.30); 2¼ extremely compressed �1.30).
206. I8.* (Weight¼ 50). I1 crown shape: 0¼ spatulate; no ap-parent occlusal cusp, mesial and distal edges continuousand rounded; 1¼ semispatulate, central cusp present butblunt with discernable mesial and distal occlusal crests;2¼ central occlusal cusp pointed, occlusal edges steep.
207. I9. (Weight¼ 100). I1 lingual fovea: 0¼ simple; 1¼ dualwith midcrown pillar.
208. I10. (Weight¼ 100). I1 occlusal edge orientation (spatu-late incisors only): 0¼ occlusal edge orthogonal to longaxis of root; 1¼ occlusal edge wears at a steep angle tolong axis of root; 2¼ crown with pronounced mesialasymmetry (¼mesial process) in unworn state.
209. I11.* (Weight¼ 50). I1e2 lingual cingulum: 0¼weak,discontinuous; 1¼ narrow, continuous; 2¼ strong.
210. I12. (Weight¼ 100). I1 basal lingual cusp: 0¼ absent;1¼ present.
211. I13. (Weight¼ 100). I1e2 buccal cingulum: 0¼ absent;1¼ present.
Canines212. C1. (Weight¼ 100). Canine cross-sectional shape (gap
coded; ratio of maximum length in the occlusal plane tomaximum breadth in the occlusal plane at right angles tomaximum length): 0¼ oval (�1.16); 1¼ rounded (<1.16).
213. C2. (unordered, weight¼ 100). Upper canine occlusion:0¼ canine wears against P1 or P2; 1¼ canine wearsagainst P2; 2¼ canine wears against P2 or P3; 3¼ caninewears against P3.
214. C3. (Weight¼ 100). Canine mesial groove (females):0¼ shallow or absent; 1¼ deep.
215. C4.* (Weight¼ 50). Canine lingual cingulum: 0¼weakor absent; 1¼ strong; 2¼ very strong.
Premolars216. P1.* (Weight¼ 50). P2 root number: 0¼ one; 1¼ two;
2¼ three. If tooth is absent, taxon scored as 9.217. P2.* (Weight¼ 50). P3 root number: 0¼ one; 1¼ two;
2¼ three.218. P3.* (Weight¼ 50). P4 root number: 0¼ one; 1¼ two;
2¼ three.219. P4.* (Weight¼ 50). P2 to P3 area: 0¼ P2 very small
(�0.85); 1¼ P2 small (>0.85, <0.95); 2¼ P2 equal(�0.95). If tooth is absent, taxon scored as 9.
220. P5.* (Weight¼ 33). P4 to M1 area: 0¼ P4<< M1(�0.66); 1¼ P4<M1 (>0.66, �0.76); 2¼ P4¼M1
(0.77e1.05); 3¼ P4>M1 (>1.06).221. P6.* (Weight¼ 50). P2 occlusal shape (MD/BL):
0¼ buccolingually broad (<0.80); 1¼ round (>0.80,<1.05); 2¼mesiodistally elongate (>1.05).
222. P8. (Weight¼ 100). P3e4 trigon/talon proportions:0¼ trigon and talon proportions similar; 1¼ trigonmuch shorter than talon, with the protocone situated onthe mesial aspect of the crown. Comment: Charactermodified from that of Kay et al. (2004c). On P4, some
species have the protocone in a mesial position with ei-ther a long postprotocrista or a short postprotocrista andseparate hypocone. An example of this would be Brachy-teles. Others (e.g., Callithrix) have the protocone morecentrally placed. Concomitantly, this means that the talonis either large, as in Brachyteles, or much smaller, as inCallithrix.
223. P9. (Weight¼ 100). P3 protocone: 0¼ present; 1¼ absent.224. P10. (Weight¼ 100). P4 metacone: 0¼ absent; 1¼ present.225. P11. (Weight¼ 100). P4 protocone: 0¼ low relative to
paracone; 1¼ high relative to paracone.226. P12. (Weight¼ 100). P2 protocone: 0¼ present as dis-
crete cusp; 1¼ absent or indistinguishable from lingualcingular ridge.
227. P13.* (Weight¼ 33). Premolar hypocones: 0¼ absent;1¼ present on P4 only; 2¼ present on P3e4; 3¼ presenton P2e4.
228. P13a.* (Weight¼ 50). P4 hypocone: 0¼ absent or trace;1¼ bump on postprotocone crista or postcingulum;2¼ distinct cusp on distal margin.
229. P14.* (Weight¼ 50). P4 paraconule: 0¼ large;1¼ small; 2¼ absent.
230. P15. (Weight¼ 100). P3e4 parastyles: 0¼ present;1¼weak or absent.
231. P16. (Weight¼ 100). P3e4 metastyles: 0¼weak or ab-sent; 1¼ present.
232. P17. (Weight¼ 100). P3e4 postprotocrista: 0¼ strong,reaches the distal margin, and joins the postcingulum;1¼weak, short.
233. P18. (Weight¼ 100). P2e3 profile of distal crown margin:0¼ convex, smoothly rounded; 1¼ concave, ‘‘waisted’’between buccal and lingual cusps.
234. P19. (Weight¼ 100). P3e4 lingual cingulum: 0¼ absentor weak; 1¼ strong.
235. P20. (Weight¼ 100). P4 metacone: 0¼ absent; 1¼ present.236. P21. (Weight¼ 100). P3e4 buccal cingulum: 0¼ absent
or weak; 1¼ strong.
Molars237. M1.* (Weight¼ 50). M1e2 root count: 0¼ three, three;
1¼ three, two; 2¼ two, two.238. M2.* (Weight¼ 50). M3 root count: 0¼ three; 1¼ two;
2¼ one.239. M3.* (Weight¼ 50). M2 shape (ratio of BL/MD):
0¼ very transverse (>1.65); 1¼ transverse (<1.65,>1.30); 2¼ squared (�1.30).
240. M4.* (Weight¼ 50). Ratio of M1 area to M2 area:0¼M1>> M2 (�1.40); 1¼M1>M2 (<1.40, >1.0);2¼M1�M2 (�1.0).
241. M5.* (Weight¼ 50). M1e2 Nannopithex-fold: 0¼ absent;1¼weak; 2¼ strong.
242. M6.* (Weight¼ 50). M1e2 pseudohypocone: 0¼ absent;1¼ small; 2¼ large.
243. M7. (Weight¼ 100). M1e2 metaconule: 0¼ single (orabsent); 1¼ double.
244. M8.* (Weight¼ 50). M1e2 paraconule: 0¼ absent;1¼ small; 2¼ large.

366 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
245. M9.* (Weight¼ 50). M1e2 preprotoconule: 0¼ absent;1¼weak; 2¼ strong.
246. M10.* (Weight¼ 50). M1 hypocone size: 0¼ large;1¼ small; 2¼ absent.
247. M11.* (Weight¼ 50). M2 hypocone size: 0¼ large;1¼ small; 2¼ absent.
248. M12. (Weight¼ 100). M1e2 hypocone position: 0¼ distal,slightly lingual to protocone; 1¼ distal, far lingual toprotocone.
249. M13.* (Weight¼ 50). M1e2 prehypocrista: 0¼ absent;1¼weak; 2¼ strong, reaches to the postprotocrista, en-closes the talon lingually.
250. M13a. (Weight¼ 100). M1e2 prehypocrista orientation:0¼ buccolingually towards postprotocrista; 1¼ buccallytowards metaconule. This character is an autapomorphyof Cebus among extant platyrrhines but is also seen inAcrecebus (Kay and Cozzuol, 2006).
251. M14.* (Weight¼ 50). M3 prehypocrista development:0¼ absent; 1¼weak; 2¼ strong, reaches to postprotoc-rista to enclose the talon lingually.
252. M15. (Weight¼ 100). M1 or M2 paraconule position:0¼ attached to preprotocrista; 1¼ not attached topreprotocrista.
253. M16.* (Weight¼ 50). M1e2 metaconule size: 0¼ absent;1¼ small to moderate; 2¼ large.
254. M17. (Unordered; weight¼ 100). M1e2 mesostyle size:0¼ absent; 1¼ present, attached to ectocrista; 2¼present on buccal cingulum.
255. M18. (Unordered; weight¼ 100). M1e2 postprotocrista:0¼ strong, runs to base of metaconule or metacone;1¼ strong but short, does not reach base of metacone;2¼ absent.
Appendix 2. Character-taxon matrix for characters described
Taxon 1
CR1 CR2 CR3 CR4 CR5 CR6 CR7 C
Tarsius 1 1 0 1 1 0 1
Aegyptopithecus zeuxis 0 ? 0 0 ? 0 0
Apidium phiomense ? ? ? ? ? 0 0
Catopithecus browni ? ? ? ? ? 0 0
Proteopithecus sylviae ? ? ? ? ? 0 0
Simonsius grangeri ? ? ? ? ? 2 0
Aotus 1 0 0 1 0 {01} 0
Saimiri 1 1 1 1 1 0 0
Cebus 1 0 0 0 {01} 0 0
Callicebus 0 0 0 1 0 1 1
Pithecia 1 1 0 1 1 0 0
Chiropotes 1 ? 1 ? ? 0 0
Cacajao 1 1 1 1 1 0 0
Proteropithecia ? ? ? ? ? ? ?
Saguinus {01} 1 0 0 1 0 0
Callithrix {01} 1 0 1 0 0 0
Cebuella pygmaea 1 1 0 1 0 {12} 0
Callimico goeldii 1 0 0 0 0 1 0
Leontopithecus 0 0 0 1 1 0 0
Alouatta 1 0 0 0 0 1 0
Ateles 1 1 0 {01} 1 1 0
Brachyteles 1 1 0 1 1 2 0
Lagothrix 1 1 0 {01} 0 2 0
256. M19. (Weight¼ 100). M1e2 lateral posterior transversecrista: 0¼ sharp; 1¼ indistinct.
257. M20.* (Weight¼ 50). P4eM1 pericone: 0¼ absent;1¼ small; 2¼ large.
258. M22.* (Weight¼ 50). M1e3 lingual cingulum develop-ment: 0¼ absent; 1¼weak, broken; 2¼ strong,complete.
259. M24.* (Weight¼ 50). M1e2 buccal cingulum develop-ment: 0¼ absent; 1¼weak; 2¼ strong.
260. M27. (Weight¼ 100). M1e2 premetaconule cristae:0¼ absent or weak; 1¼ strong.
261. M28. (Weight¼ 100). M1e2 postmetaconule cristae:0¼ absent or weak; 1¼ strong.
262. M30.* (Weight¼ 50). M3 paraconule: 0¼ absent;1¼ small to moderate; 2¼ large.
263. M31.* (Weight¼ 50). Molar protocone lingual infla-tion: 0¼ not inflated; 1¼ slightly inflated; 2¼ veryinflated.
264. M33.* (Weight¼ 100). M2 buccal expansion of para-cone: 0¼ no expansion; 1¼ expanded.
265. M34.* (Weight¼ 50). M3 metacone: 0¼ absent or verysmall; 1¼moderate (but smaller than paracone); 2¼large (equal to paracone).
266. M36.* (Weight¼ 50). M3 hypocone: 0¼ absent or verysmall; 1¼ small; 2¼ large.
267. M44.* (Weight¼ 50). M1e3 anterior cingulum: 0¼strong, complete, long; 1¼ strong, short; 2¼weak orabsent.
268. M46.* (Weight¼ 50). M1 size relative to M3 (based onthe ratio of areas of each tooth): 0¼M1� 2.5 times thesize of M3 (scored as 0 when M3 is absent);1¼M1< 2.5, � 1.5 times M3; 2¼M1< 1.5 times M3.
in Appendix 1
10 17
R8 CR9 CR10 CR11 CR12 CR13 CR14 CR15 CR16 CR17
1 1 0 0 1 2 1 0 0 ?
0 2 1 0 1 ? 0 2 0 ?
? ? 1 ? ? ? ? ? ? ?
0 ? 1 ? ? ? 1 ? ? ?
0 ? 1 ? 1 ? 2 ? 0 ?
0 ? 1 ? 1 ? 1 2 0 ?
1 1 1 0 1 2 2 0 0 ?
0 1 1 0 1 2 2 0 0 0
0 1 1 0 1 2 2 0 0 ?
0 1 1 1 1 2 2 2 0 ?
0 1 1 0 1 2 2 2 0 1
0 1 1 0 1 2 2 2 0 ?
0 1 1 0 1 2 2 2 1 ?
? ? ? ? ? ? ? ? ? ?
0 1 1 0 1 2 2 0 0 0
0 1 1 0 1 2 2 1 0 ?
0 1 1 0 1 2 2 1 0 ?
0 1 1 0 1 2 2 1 0 ?
0 1 1 0 1 2 2 1 0 ?
0 {12} 1 2 1 2 0 2 0 1
0 1 1 1 1 2 1 2 0 0
0 {12} 1 2 1 2 1 2 0 ?
0 1 1 1 1 2 1 1 0 ?
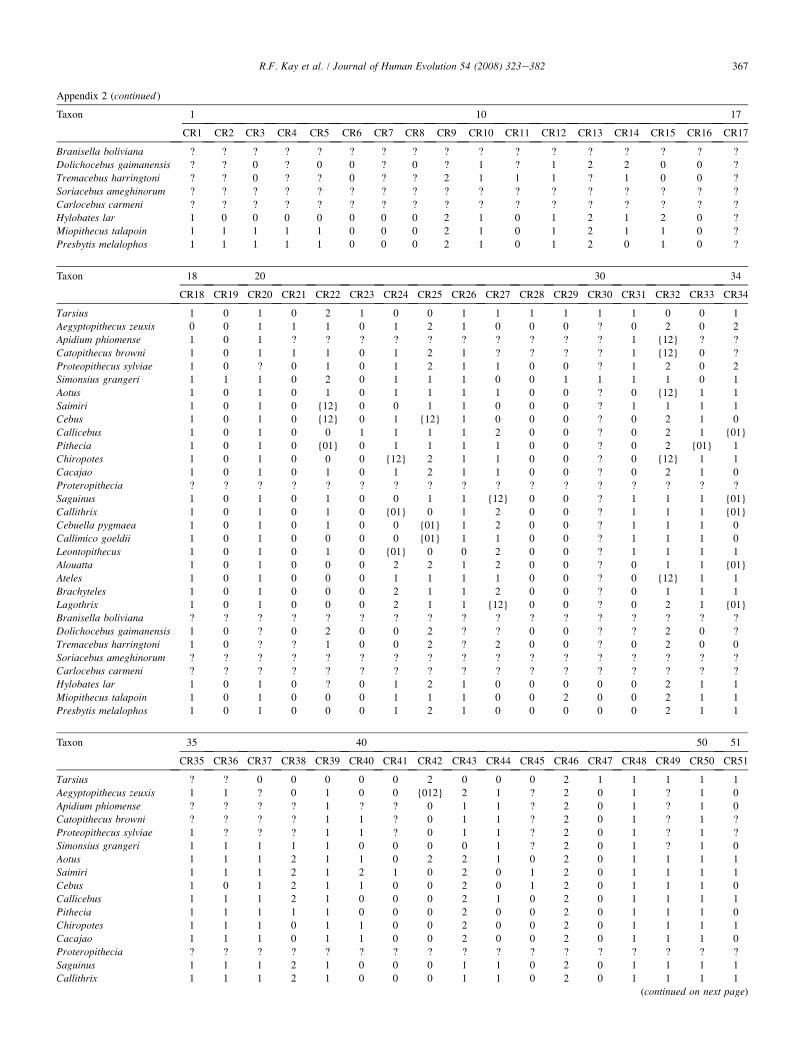
Appendix 2 (continued )
Taxon 1 10 17
CR1 CR2 CR3 CR4 CR5 CR6 CR7 CR8 CR9 CR10 CR11 CR12 CR13 CR14 CR15 CR16 CR17
Branisella boliviana ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Dolichocebus gaimanensis ? ? 0 ? 0 0 ? 0 ? 1 ? 1 2 2 0 0 ?
Tremacebus harringtoni ? ? 0 ? ? 0 ? ? 2 1 1 1 ? 1 0 0 ?
Soriacebus ameghinorum ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Carlocebus carmeni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Hylobates lar 1 0 0 0 0 0 0 0 2 1 0 1 2 1 2 0 ?
Miopithecus talapoin 1 1 1 1 1 0 0 0 2 1 0 1 2 1 1 0 ?
Presbytis melalophos 1 1 1 1 1 0 0 0 2 1 0 1 2 0 1 0 ?
Taxon 18 20 30 34
CR18 CR19 CR20 CR21 CR22 CR23 CR24 CR25 CR26 CR27 CR28 CR29 CR30 CR31 CR32 CR33 CR34
Tarsius 1 0 1 0 2 1 0 0 1 1 1 1 1 1 0 0 1
Aegyptopithecus zeuxis 0 0 1 1 1 0 1 2 1 0 0 0 ? 0 2 0 2
Apidium phiomense 1 0 1 ? ? ? ? ? ? ? ? ? ? 1 {12} ? ?
Catopithecus browni 1 0 1 1 1 0 1 2 1 ? ? ? ? 1 {12} 0 ?
Proteopithecus sylviae 1 0 ? 0 1 0 1 2 1 1 0 0 ? 1 2 0 2
Simonsius grangeri 1 1 1 0 2 0 1 1 1 0 0 1 1 1 1 0 1
Aotus 1 0 1 0 1 0 1 1 1 1 0 0 ? 0 {12} 1 1
Saimiri 1 0 1 0 {12} 0 0 1 1 0 0 0 ? 1 1 1 1
Cebus 1 0 1 0 {12} 0 1 {12} 1 0 0 0 ? 0 2 1 0
Callicebus 1 0 1 0 0 1 1 1 1 2 0 0 ? 0 2 1 {01}
Pithecia 1 0 1 0 {01} 0 1 1 1 1 0 0 ? 0 2 {01} 1
Chiropotes 1 0 1 0 0 0 {12} 2 1 1 0 0 ? 0 {12} 1 1
Cacajao 1 0 1 0 1 0 1 2 1 1 0 0 ? 0 2 1 0
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 1 0 1 0 1 0 0 1 1 {12} 0 0 ? 1 1 1 {01}
Callithrix 1 0 1 0 1 0 {01} 0 1 2 0 0 ? 1 1 1 {01}
Cebuella pygmaea 1 0 1 0 1 0 0 {01} 1 2 0 0 ? 1 1 1 0
Callimico goeldii 1 0 1 0 0 0 0 {01} 1 1 0 0 ? 1 1 1 0
Leontopithecus 1 0 1 0 1 0 {01} 0 0 2 0 0 ? 1 1 1 1
Alouatta 1 0 1 0 0 0 2 2 1 2 0 0 ? 0 1 1 {01}
Ateles 1 0 1 0 0 0 1 1 1 1 0 0 ? 0 {12} 1 1
Brachyteles 1 0 1 0 0 0 2 1 1 2 0 0 ? 0 1 1 1
Lagothrix 1 0 1 0 0 0 2 1 1 {12} 0 0 ? 0 2 1 {01}
Branisella boliviana ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Dolichocebus gaimanensis 1 0 ? 0 2 0 0 2 ? ? 0 0 ? ? 2 0 ?
Tremacebus harringtoni 1 0 ? ? 1 0 0 2 ? 2 0 0 ? 0 2 0 0
Soriacebus ameghinorum ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Carlocebus carmeni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Hylobates lar 1 0 1 0 ? 0 1 2 1 0 0 0 0 0 2 1 1
Miopithecus talapoin 1 0 1 0 0 0 1 1 1 0 0 2 0 0 2 1 1
Presbytis melalophos 1 0 1 0 0 0 1 2 1 0 0 0 0 0 2 1 1
Taxon 35 40 50 51
CR35 CR36 CR37 CR38 CR39 CR40 CR41 CR42 CR43 CR44 CR45 CR46 CR47 CR48 CR49 CR50 CR51
Tarsius ? ? 0 0 0 0 0 2 0 0 0 2 1 1 1 1 1
Aegyptopithecus zeuxis 1 1 ? 0 1 0 0 {012} 2 1 ? 2 0 1 ? 1 0
Apidium phiomense ? ? ? ? 1 ? ? 0 1 1 ? 2 0 1 ? 1 0
Catopithecus browni ? ? ? ? 1 1 ? 0 1 1 ? 2 0 1 ? 1 ?
Proteopithecus sylviae 1 ? ? ? 1 1 ? 0 1 1 ? 2 0 1 ? 1 ?
Simonsius grangeri 1 1 1 1 1 0 0 0 0 1 ? 2 0 1 ? 1 0
Aotus 1 1 1 2 1 1 0 2 2 1 0 2 0 1 1 1 1
Saimiri 1 1 1 2 1 2 1 0 2 0 1 2 0 1 1 1 1
Cebus 1 0 1 2 1 1 0 0 2 0 1 2 0 1 1 1 0
Callicebus 1 1 1 2 1 0 0 0 2 1 0 2 0 1 1 1 1
Pithecia 1 1 1 1 1 0 0 0 2 0 0 2 0 1 1 1 0
Chiropotes 1 1 1 0 1 1 0 0 2 0 0 2 0 1 1 1 1
Cacajao 1 1 1 0 1 1 0 0 2 0 0 2 0 1 1 1 0
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 1 1 1 2 1 0 0 0 1 1 0 2 0 1 1 1 1
Callithrix 1 1 1 2 1 0 0 0 1 1 0 2 0 1 1 1 1
(continued on next page)
367R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
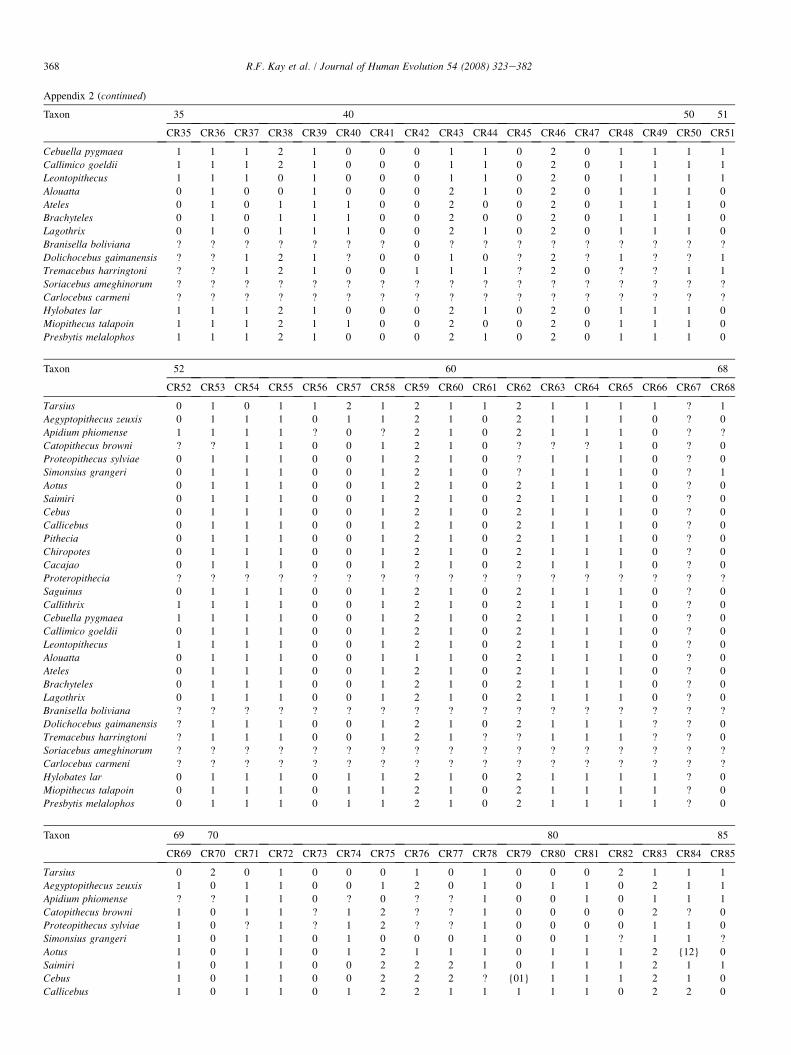
Appendix 2 (continued)
Taxon 35 40 50 51
CR35 CR36 CR37 CR38 CR39 CR40 CR41 CR42 CR43 CR44 CR45 CR46 CR47 CR48 CR49 CR50 CR51
Cebuella pygmaea 1 1 1 2 1 0 0 0 1 1 0 2 0 1 1 1 1
Callimico goeldii 1 1 1 2 1 0 0 0 1 1 0 2 0 1 1 1 1
Leontopithecus 1 1 1 0 1 0 0 0 1 1 0 2 0 1 1 1 1
Alouatta 0 1 0 0 1 0 0 0 2 1 0 2 0 1 1 1 0
Ateles 0 1 0 1 1 1 0 0 2 0 0 2 0 1 1 1 0
Brachyteles 0 1 0 1 1 1 0 0 2 0 0 2 0 1 1 1 0
Lagothrix 0 1 0 1 1 1 0 0 2 1 0 2 0 1 1 1 0
Branisella boliviana ? ? ? ? ? ? ? 0 ? ? ? ? ? ? ? ? ?
Dolichocebus gaimanensis ? ? 1 2 1 ? 0 0 1 0 ? 2 ? 1 ? ? 1
Tremacebus harringtoni ? ? 1 2 1 0 0 1 1 1 ? 2 0 ? ? 1 1
Soriacebus ameghinorum ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Carlocebus carmeni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Hylobates lar 1 1 1 2 1 0 0 0 2 1 0 2 0 1 1 1 0
Miopithecus talapoin 1 1 1 2 1 1 0 0 2 0 0 2 0 1 1 1 0
Presbytis melalophos 1 1 1 2 1 0 0 0 2 1 0 2 0 1 1 1 0
Taxon 52 60 68
CR52 CR53 CR54 CR55 CR56 CR57 CR58 CR59 CR60 CR61 CR62 CR63 CR64 CR65 CR66 CR67 CR68
Tarsius 0 1 0 1 1 2 1 2 1 1 2 1 1 1 1 ? 1
Aegyptopithecus zeuxis 0 1 1 1 0 1 1 2 1 0 2 1 1 1 0 ? 0
Apidium phiomense 1 1 1 1 ? 0 ? 2 1 0 2 1 1 1 0 ? ?
Catopithecus browni ? ? 1 1 0 0 1 2 1 0 ? ? ? 1 0 ? 0
Proteopithecus sylviae 0 1 1 1 0 0 1 2 1 0 ? 1 1 1 0 ? 0
Simonsius grangeri 0 1 1 1 0 0 1 2 1 0 ? 1 1 1 0 ? 1
Aotus 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Saimiri 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Cebus 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Callicebus 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Pithecia 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Chiropotes 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Cacajao 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Callithrix 1 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Cebuella pygmaea 1 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Callimico goeldii 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Leontopithecus 1 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Alouatta 0 1 1 1 0 0 1 1 1 0 2 1 1 1 0 ? 0
Ateles 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Brachyteles 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Lagothrix 0 1 1 1 0 0 1 2 1 0 2 1 1 1 0 ? 0
Branisella boliviana ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Dolichocebus gaimanensis ? 1 1 1 0 0 1 2 1 0 2 1 1 1 ? ? 0
Tremacebus harringtoni ? 1 1 1 0 0 1 2 1 ? ? 1 1 1 ? ? 0
Soriacebus ameghinorum ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Carlocebus carmeni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Hylobates lar 0 1 1 1 0 1 1 2 1 0 2 1 1 1 1 ? 0
Miopithecus talapoin 0 1 1 1 0 1 1 2 1 0 2 1 1 1 1 ? 0
Presbytis melalophos 0 1 1 1 0 1 1 2 1 0 2 1 1 1 1 ? 0
Taxon 69 70 80 85
CR69 CR70 CR71 CR72 CR73 CR74 CR75 CR76 CR77 CR78 CR79 CR80 CR81 CR82 CR83 CR84 CR85
Tarsius 0 2 0 1 0 0 0 1 0 1 0 0 0 2 1 1 1
Aegyptopithecus zeuxis 1 0 1 1 0 0 1 2 0 1 0 1 1 0 2 1 1
Apidium phiomense ? ? 1 1 0 ? 0 ? ? 1 0 0 1 0 1 1 1
Catopithecus browni 1 0 1 1 ? 1 2 ? ? 1 0 0 0 0 2 ? 0
Proteopithecus sylviae 1 0 ? 1 ? 1 2 ? ? 1 0 0 0 0 1 1 0
Simonsius grangeri 1 0 1 1 0 1 0 0 0 1 0 0 1 ? 1 1 ?
Aotus 1 0 1 1 0 1 2 1 1 1 0 1 1 1 2 {12} 0
Saimiri 1 0 1 1 0 0 2 2 2 1 0 1 1 1 2 1 1
Cebus 1 0 1 1 0 0 2 2 2 ? {01} 1 1 1 2 1 0
Callicebus 1 0 1 1 0 1 2 2 1 1 1 1 1 0 2 2 0
368 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Appendix 2 (continued )
Taxon 69 70 80 85
CR69 CR70 CR71 CR72 CR73 CR74 CR75 CR76 CR77 CR78 CR79 CR80 CR81 CR82 CR83 CR84 CR85
Pithecia 1 0 1 1 0 1 2 2 2 1 1 1 1 1 2 2 1
Chiropotes 1 ? 1 1 0 1 2 2 2 1 1 1 1 2 2 2 1
Cacajao 1 ? 1 1 0 1 2 2 2 1 1 1 1 1 2 2 1
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 1 0 1 1 0 0 2 2 0 1 0 1 1 1 2 1 0
Callithrix 1 0 1 1 0 0 2 2 0 0 0 0 1 1 1 0 1
Cebuella pygmaea 1 0 1 1 0 0 2 2 0 0 0 0 1 1 1 0 1
Callimico goeldii 1 ? 1 1 0 1 2 2 1 1 0 1 1 0 2 1 0
Leontopithecus 1 ? 1 1 0 0 2 2 1 0 0 1 1 0 2 1 1
Alouatta 1 0 1 1 0 1 2 2 0 1 1 1 1 2 2 2 1
Ateles 1 {01} 1 1 0 1 2 2 2 1 1 1 1 2 2 2 1
Brachyteles 1 1 1 1 0 1 ? 2 ? 1 1 1 1 2 2 2 1
Lagothrix 1 1 1 1 0 1 2 2 2 1 1 1 1 2 2 2 1
Branisella boliviana ? ? ? ? ? ? ? ? ? ? 0 0 1 ? ? ? ?
Dolichocebus gaimanensis 1 0 1 1 0 1 2 2 ? ? 0 ? ? ? ? ? ?
Tremacebus harringtoni 1 1 ? ? ? 1 ? 2 ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum ? ? ? ? ? ? ? ? ? ? ? 1 ? ? ? ? ?
Carlocebus carmeni ? ? ? ? ? ? ? ? ? ? ? 1 ? ? ? ? ?
Hylobates lar 1 0 1 1 0 0 0 2 2 1 0 0 1 2 2 1 1
Miopithecus talapoin 1 0 1 1 0 0 0 2 2 1 0 1 1 2 2 1 1
Presbytis melalophos 1 0 1 1 0 0 0 2 2 1 0 1 1 2 2 1 1
Taxon 86 90 100 102
i1 i2 i3 i4 i5 i6 i7 i8 i9 i10 i11 i12 i13 i14 i15 i16 i17
Tarsius 2 1 ? 1 ? 0 ? 1 2 0 1 1 0 1 ? 0 0
Aegyptopithecus zeuxis 1 0 0 1 0 ? 0 0 1 0 0 0 0 0 1 0 1
Apidium phiomense 1 0 0 1 0 ? 0 0 0 0 0 0 0 0 1 0 2
Catopithecus browni 1 ? 0 ? ? ? 0 0 1 0 0 0 0 0 1 0 ?
Proteopithecus sylviae 1 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Simonsius grangeri 3 ? ? ? ? 0 ? ? ? ? ? ? 0 ? ? ? ?
Aotus 1 0 0 1 2 2 0 0 0 0 0 0 0 0 1 0 2
Saimiri 1 0 0 1 {12} {01} 0 0 0 0 0 0 0 0 1 0 2
Cebus 1 0 0 1 2 1 0 0 0 0 0 0 0 0 1 0 2
Callicebus 1 0 0 1 0 1 1 0 1 0 0 0 0 0 1 0 0
Pithecia 1 0 0 1 1 1 2 0 2 0 1 1 0 0 0 0 0
Chiropotes 1 0 0 1 1 0 2 1 2 0 1 1 0 0 0 0 0
Cacajao 1 0 0 1 1 0 2 1 2 0 1 1 0 0 0 0 0
Proteropithecia ? 0 0 ? ? ? 2 1 2 0 ? ? 0 0 0 0 0
Saguinus 1 0 0 1 2 1 0 0 1 0 0 0 0 0 1 0 2
Callithrix 1 1 0 1 2 2 2 1 2 1 1 1 0 1 0 1 2
Cebuella pygmaea 1 1 0 1 1 2 2 1 2 1 1 1 0 1 0 1 1
Callimico goeldii 1 0 0 1 0 2 0 0 0 0 0 0 0 0 1 0 2
Leontopithecus 1 0 0 1 1 2 0 0 0 0 0 0 0 0 1 0 2
Alouatta 1 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1
Ateles 1 0 0 1 2 0 0 0 0 0 0 0 0 0 1 0 1
Brachyteles 1 0 0 1 0 0 0 0 0 0 0 0 0 0 1 0 2
Lagothrix 1 0 0 1 2 1 0 0 0 0 0 0 0 0 1 0 2
Branisella boliviana 1 0 0 1 0 {12} ? 0 0 0 1 0 0 0 1 0 2
Dolichocebus gaimanensis 1 ? 0 ? 0 ? 0 0 0 0 ? ? 0 0 1 0 2
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum 1 0 0 1 2 0 2 1 2 0 1 1 0 0 0 0 0
Carlocebus carmeni 1 0 ? 1 ? ? ? ? ? ? ? ? 0 ? ? ? ?
Hylobates lar 1 0 0 1 1 1 0 0 0 0 0 0 0 0 1 0 ?
Miopithecus talapoin 1 0 0 1 2 2 1 0 0 0 0 0 0 0 1 0 ?
Presbytis melalophos 1 0 0 1 1 1 1 0 0 0 0 0 0 0 1 0 0
Taxon 103 110 119
i19 c1 c2 c3 c4 c5 c6 p1 p2 p3 p4 p5 p6 p7 p9 p11 p13
Tarsius 0 1 0 0 0 0 0 1 0 0 1 2 2 2 ? 1 3
Aegyptopithecus zeuxis 0 1 2 0 0 0 2 1 1 ? 2 0 2 2 ? 1 ?
(continued on next page)
369R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
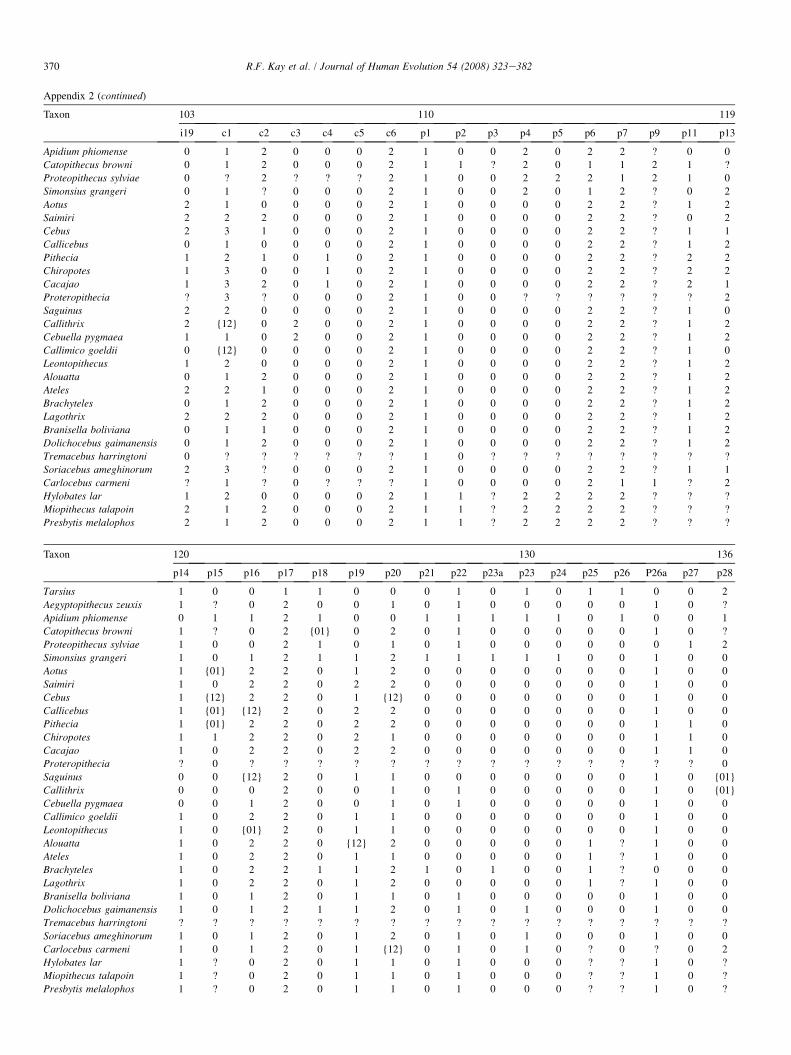
ppendix 2 (continued)
axon 103 110 119
i19 c1 c2 c3 c4 c5 c6 p1 p2 p3 p4 p5 p6 p7 p9 p11 p13
pidium phiomense 0 1 2 0 0 0 2 1 0 0 2 0 2 2 ? 0 0
atopithecus browni 0 1 2 0 0 0 2 1 1 ? 2 0 1 1 2 1 ?
roteopithecus sylviae 0 ? 2 ? ? ? 2 1 0 0 2 2 2 1 2 1 0
imonsius grangeri 0 1 ? 0 0 0 2 1 0 0 2 0 1 2 ? 0 2
otus 2 1 0 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
aimiri 2 2 2 0 0 0 2 1 0 0 0 0 2 2 ? 0 2
ebus 2 3 1 0 0 0 2 1 0 0 0 0 2 2 ? 1 1
allicebus 0 1 0 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
ithecia 1 2 1 0 1 0 2 1 0 0 0 0 2 2 ? 2 2
hiropotes 1 3 0 0 1 0 2 1 0 0 0 0 2 2 ? 2 2
acajao 1 3 2 0 1 0 2 1 0 0 0 0 2 2 ? 2 1
roteropithecia ? 3 ? 0 0 0 2 1 0 0 ? ? ? ? ? ? 2
aguinus 2 2 0 0 0 0 2 1 0 0 0 0 2 2 ? 1 0
allithrix 2 {12} 0 2 0 0 2 1 0 0 0 0 2 2 ? 1 2
ebuella pygmaea 1 1 0 2 0 0 2 1 0 0 0 0 2 2 ? 1 2
allimico goeldii 0 {12} 0 0 0 0 2 1 0 0 0 0 2 2 ? 1 0
eontopithecus 1 2 0 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
louatta 0 1 2 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
teles 2 2 1 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
rachyteles 0 1 2 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
agothrix 2 2 2 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
ranisella boliviana 0 1 1 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
olichocebus gaimanensis 0 1 2 0 0 0 2 1 0 0 0 0 2 2 ? 1 2
remacebus harringtoni 0 ? ? ? ? ? ? 1 0 ? ? ? ? ? ? ? ?
oriacebus ameghinorum 2 3 ? 0 0 0 2 1 0 0 0 0 2 2 ? 1 1
arlocebus carmeni ? 1 ? 0 ? ? ? 1 0 0 0 0 2 1 1 ? 2
ylobates lar 1 2 0 0 0 0 2 1 1 ? 2 2 2 2 ? ? ?
iopithecus talapoin 2 1 2 0 0 0 2 1 1 ? 2 2 2 2 ? ? ?
resbytis melalophos 2 1 2 0 0 0 2 1 1 ? 2 2 2 2 ? ? ?
axon 120 130 136
p14 p15 p16 p17 p18 p19 p20 p21 p22 p23a p23 p24 p25 p26 P26a p27 p28
arsius 1 0 0 1 1 0 0 0 1 0 1 0 1 1 0 0 2
egyptopithecus zeuxis 1 ? 0 2 0 0 1 0 1 0 0 0 0 0 1 0 ?
pidium phiomense 0 1 1 2 1 0 0 1 1 1 1 1 0 1 0 0 1
atopithecus browni 1 ? 0 2 {01} 0 2 0 1 0 0 0 0 0 1 0 ?
roteopithecus sylviae 1 0 0 2 1 0 1 0 1 0 0 0 0 0 0 1 2
imonsius grangeri 1 0 1 2 1 1 2 1 1 1 1 1 0 0 1 0 0
otus 1 {01} 2 2 0 1 2 0 0 0 0 0 0 0 1 0 0
aimiri 1 0 2 2 0 2 2 0 0 0 0 0 0 0 1 0 0
ebus 1 {12} 2 2 0 1 {12} 0 0 0 0 0 0 0 1 0 0
allicebus 1 {01} {12} 2 0 2 2 0 0 0 0 0 0 0 1 0 0
ithecia 1 {01} 2 2 0 2 2 0 0 0 0 0 0 0 1 1 0
hiropotes 1 1 2 2 0 2 1 0 0 0 0 0 0 0 1 1 0
acajao 1 0 2 2 0 2 2 0 0 0 0 0 0 0 1 1 0
roteropithecia ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? 0
aguinus 0 0 {12} 2 0 1 1 0 0 0 0 0 0 0 1 0 {01}
allithrix 0 0 0 2 0 0 1 0 1 0 0 0 0 0 1 0 {01}
ebuella pygmaea 0 0 1 2 0 0 1 0 1 0 0 0 0 0 1 0 0
allimico goeldii 1 0 2 2 0 1 1 0 0 0 0 0 0 0 1 0 0
eontopithecus 1 0 {01} 2 0 1 1 0 0 0 0 0 0 0 1 0 0
louatta 1 0 2 2 0 {12} 2 0 0 0 0 0 1 ? 1 0 0
teles 1 0 2 2 0 1 1 0 0 0 0 0 1 ? 1 0 0
rachyteles 1 0 2 2 1 1 2 1 0 1 0 0 1 ? 0 0 0
agothrix 1 0 2 2 0 1 2 0 0 0 0 0 1 ? 1 0 0
ranisella boliviana 1 0 1 2 0 1 1 0 1 0 0 0 0 0 1 0 0
olichocebus gaimanensis 1 0 1 2 1 1 2 0 1 0 1 0 0 0 1 0 0
remacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
oriacebus ameghinorum 1 0 1 2 0 1 2 0 1 0 1 0 0 0 1 0 0
arlocebus carmeni 1 0 1 2 0 1 {12} 0 1 0 1 0 ? 0 ? 0 2
ylobates lar 1 ? 0 2 0 1 1 0 1 0 0 0 ? ? 1 0 ?
iopithecus talapoin 1 ? 0 2 0 1 1 0 1 0 0 0 ? ? 1 0 ?
resbytis melalophos 1 ? 0 2 0 1 1 0 1 0 0 0 ? ? 1 0 ?
370 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
A
T
AC
P
S
AS
C
C
PC
C
PS
C
C
CL
A
A
BL
B
D
TS
C
HM
P
T
T
AA
C
P
SA
S
C
CP
C
CP
S
C
CC
L
A
AB
L
BD
T
S
CH
M
P

Appendix 2 (continued )
Taxon 137 140 150 153
p29 P30 p31 p34 p36 p37 p40 p41 p42 p43 p44 p45 m1 m2 m3 m4 m6
Tarsius 0 1 0 1 1 1 ? 0 1 1 0 1 0 1 1 1 1
Aegyptopithecus zeuxis 1 0 3 0 1 1 ? 2 4 0 1 1 0 1 1 1 1
Apidium phiomense 1 0 1 0 0 0 ? 1 3 2 {12} 2 0 1 1 1 1
Catopithecus browni 0 0 2 0 1 1 0 1 3 3 1 2 0 1 1 1 1
Proteopithecus sylviae 1 0 2 1 0 1 0 1 1 1 0 0 0 1 1 1 1
Simonsius grangeri 1 0 3 0 0 0 ? 1 3 1 2 0 0 1 1 1 0
Aotus 0 0 1 0 1 1 ? 1 2 0 1 0 0 1 1 0 1
Saimiri 0 0 0 0 1 1 ? 1 3 0 3 0 0 1 1 0 {01}
Cebus 1 0 2 0 1 1 ? 1 3 0 3 0 0 1 1 0 {01}
Callicebus 0 0 2 0 1 1 ? 1 3 0 0 0 0 1 1 0 1
Pithecia 0 0 3 0 1 1 ? 1 3 0 1 0 0 1 1 0 1
Chiropotes 0 0 3 0 1 1 ? 1 3 0 3 0 0 1 1 ? 1
Cacajao 0 0 3 0 1 1 ? 1 3 0 3 0 0 1 1 ? 1
Proteropithecia 1 ? ? ? ? ? ? ? ? ? ? ? ? ? 1 ? 1
Saguinus 0 0 1 0 1 1 ? 1 3 {01} 2 0 {01} 1 0 ? {01}
Callithrix 1 0 2 1 1 1 ? 2 3 {01} 3 0 1 1 0 ? 0
Cebuella pygmaea 1 0 2 0 1 1 ? 2 2 1 2 0 1 1 0 ? 1
Callimico goeldii 0 0 3 0 1 1 ? 1 3 0 2 0 0 1 1 0 1
Leontopithecus 1 0 2 0 1 1 ? 1 3 0 4 0 1 1 0 ? 1
Alouatta 0 0 0 0 1 1 ? 1 3 {01} {12} 0 0 1 1 1 {12}
Ateles 1 0 2 0 1 1 ? 1 3 0 {12} 0 0 0 0 0 1
Brachyteles 0 0 0 0 1 1 0 1 3 0 1 0 0 0 1 ? 1
Lagothrix 0 0 1 0 1 1 ? 1 3 0 1 0 0 1 1 0 1
Branisella boliviana 0 0 3 0 0 1 ? 1 1 1 3 0 0 1 1 1 1
Dolichocebus gaimanensis 1 0 0 0 ? 1 ? 1 3 1 2 ? 0 1 1 1 1
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum 1 0 1 0 1 1 ? 2 3 1 2 0 0 1 1 0 1
Carlocebus carmeni 1 0 2 0 1 1 ? 1 3 ? ? ? 0 1 1 ? 1
Hylobates lar 0 0 3 0 1 1 ? 2 4 2 1 1 0 1 1 1 1
Miopithecus talapoin 0 0 3 0 1 1 ? 2 4 2 0 1 0 1 1 1 1
Presbytis melalophos 0 0 3 0 1 1 ? 2 3 1 1 1 0 1 1 1 1
Taxon 154 160 170
m6a m7 m8 m9 m10 m11 m12 m13 m14 m15 m16 m17 m18 m19 m20 m21 m22
Tarsius 0 1 1 1 1 0 0 0 1 0 0 2 0 1 2 0 0
Aegyptopithecus zeuxis 0 1 ? ? ? 0 1 0 1 0 2 1 1 0 0 1 0
Apidium phiomense 0 1 ? ? ? 0 0 0 2 0 2 1 1 1 0 1 1
Catopithecus browni 0 1 1 ? ? 0 0 0 1 0 0 1 0 0 1 0 0
Proteopithecus sylviae 0 0 1 ? ? 0 0 0 {01} 0 1 2 0 0 1 0 0
Simonsius grangeri 0 0 ? ? ? 0 0 0 1 0 2 1 1 1 0 1 1
Aotus 0 1 ? ? ? 0 0 ? 0 0 2 1 1 0 0 0 0
Saimiri 0 0 ? ? ? 0 0 ? 0 0 2 2 1 1 0 0 0
Cebus 0 0 ? ? ? 0 0 ? 0 0 2 1 1 0 0 0 0
Callicebus 0 1 ? ? ? 0 0 ? 0 0 2 1 1 1 0 0 0
Pithecia 0 1 ? ? ? 0 0 ? 0 1 2 0 1 0 0 0 0
Chiropotes 0 1 ? ? ? 0 0 ? 0 2 2 0 1 0 0 0 2
Cacajao 0 1 ? ? ? 0 0 ? 0 2 2 0 1 0 0 0 2
Proteropithecia ? ? ? ? ? 0 0 ? ? 1 2 ? 1 1 0 0 0
Saguinus 0 ? ? ? ? 0 0 ? ? 0 1 2 1 0 {01} 0 0
Callithrix 0 ? ? ? ? 0 0 ? ? 0 2 2 1 1 0 0 0
Cebuella pygmaea 0 ? ? ? ? 0 0 ? ? 0 2 0 1 1 0 0 0
Callimico goeldii 0 0 ? ? ? 0 0 ? 0 0 2 2 1 0 0 0 0
Leontopithecus 0 ? ? ? ? 0 0 ? ? 0 2 2 1 0 0 0 0
Alouatta 0 {12} ? ? ? 0 0 0 0 0 2 2 {01} 1 0 0 0
Ateles 0 1 ? ? ? 0 0 ? 0 0 2 0 1 0 0 0 0
Brachyteles 0 1 ? ? ? 0 0 0 0 0 2 1 0 1 1 0 0
Lagothrix 0 1 ? ? ? 0 0 ? 0 0 2 0 1 0 0 0 0
Branisella boliviana 0 1 ? ? ? 0 0 0 {01} 0 2 1 1 0 0 0 0
Dolichocebus gaimanensis ? ? 1 ? ? 0 0 ? ? 0 2 1 1 1 1 0 0
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum 1 1 ? ? ? 0 0 ? 2 0 2 1 1 1 0 0 0
(continued on next page)
371R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Appendix 2 (continued)
Taxon 154 160 170
m6a m7 m8 m9 m10 m11 m12 m13 m14 m15 m16 m17 m18 m19 m20 m21 m22
Carlocebus carmeni ? ? ? ? ? 0 0 ? ? 0 2 ? 1 1 0 0 0
Hylobates lar 0 1 ? ? ? 0 0 0 1 0 2 0 1 0 0 1 1
Miopithecus talapoin 0 1 ? ? ? 0 0 ? 0 0 2 0 1 0 0 1 0
Presbytis melalophos 0 1 ? ? ? 0 0 0 1 0 2 0 1 0 0 1 0
Taxon 171 180 187
m23 m24a m24 m25 m26 m27 m28 m29 m30 m31 m32 m33 m34 m35 m36 m37 m38
Tarsius 0 1 1 1 2 3 3 0 ? 2 0 0 0 0 0 1 2
Aegyptopithecus zeuxis 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 1 1
Apidium phiomense 1 1 0 1 0 0 0 0 1 0 2 0 0 0 0 0 1
Catopithecus browni 0 1 1 1 1 1 1 1 0 0 0 0 0 0 0 1 2
Proteopithecus sylviae 0 1 1 1 1 1 1 1 0 0 0 0 0 0 0 1 2
Simonsius grangeri 1 1 0 2 1 0 0 1 1 1 1 0 0 0 0 1 1
Aotus 0 1 1 1 2 3 3 3 ? 1 0 0 2 2 2 1 1
Saimiri 0 1 1 1 2 2 2 1 1 1 1 0 0 0 1 1 2
Cebus 0 1 1 1 2 3 3 2 ? 0 0 0 2 2 2 1 1
Callicebus 0 1 1 1 {12} {23} {23} 1 1 1 1 0 2 2 2 1 1
Pithecia 2 1 1 1 2 3 3 2 ? 0 0 0 2 2 2 1 0
Chiropotes 2 1 1 0 2 3 3 2 ? 0 0 0 2 2 2 1 0
Cacajao 2 1 1 0 2 3 3 2 ? 0 0 0 2 2 2 1 0
Proteropithecia 0 1 1 1 2 3 3 ? ? 1 0 0 2 2 ? 1 1
Saguinus 0 1 1 0 2 3 3 ? ? 1 {01} 0 1 1 ? 1 1
Callithrix 0 1 1 0 2 3 3 ? ? 1 1 0 0 0 ? 1 1
Cebuella pygmaea 0 1 1 1 2 3 3 ? ? 1 1 0 0 0 ? 1 1
Callimico goeldii 0 1 1 0 2 3 3 3 ? 2 0 0 0 0 0 1 2
Leontopithecus 2 1 1 0 2 {23} {23} ? 0 1 0 0 1 1 ? 1 1
Alouatta 0 1 1 1 {12} {23} {23} 1 0 2 1 {01} 1 1 2 1 2
Ateles 0 1 1 1 2 2 3 2 ? 0 0 0 1 2 2 1 1
Brachyteles 0 1 1 2 2 3 3 2 ? 2 1 1 0 0 2 1 2
Lagothrix 0 1 1 1 2 3 3 2 ? 1 0 0 0 2 2 1 1
Branisella boliviana 0 1 1 1 1 1 1 1 0 1 1 0 1 1 0 1 1
Dolichocebus gaimanensis 0 1 1 1 0 2 2 ? 1 1 1 0 1 2 ? 1 1
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum 0 1 1 0 1 2 2 1 1 0 0 0 1 1 1 1 1
Carlocebus carmeni 0 1 1 1 0 2 ? ? 1 1 1 1 1 1 ? 1 2
Hylobates lar 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 1 1
Miopithecus talapoin 0 0 0 2 2 3 3 2 ? 2 0 0 0 0 0 1 2
Presbytis melalophos 0 0 0 2 2 3 3 0 ? 2 0 0 0 0 0 1 2
Taxon 188 190 200 204
m39 m40 m41 m44 m45 m46 m47 m53 m54 m55 m57 I1 I2 I3 I4 I5 I6
Tarsius 2 2 0 0 2 1 1 0 0 0 0 2 0 0 2 0 0
Aegyptopithecus zeuxis 1 1 1 1 {12} 1 1 1 3 0 0 ? ? ? ? ? ?
Apidium phiomense 1 1 1 1 1 2 1 2 1 0 0 ? ? ? ? ? ?
Catopithecus browni 2 2 0 0 0 1 1 3 0 1 0 1 0 0 0 0 1
Proteopithecus sylviae 2 2 0 0 1 1 1 3 0 1 0 ? ? ? 1 ? ?
Simonsius grangeri 1 1 1 1 0 1 1 2 1 0 0 ? ? ? ? ? ?
Aotus 0 2 0 0 0 1 1 3 1 0 0 1 0 0 2 2 2
Saimiri 0 2 0 0 1 1 1 4 1 0 0 1 0 0 2 2 1
Cebus 0 2 0 1 0 1 0 3 2 0 0 1 0 0 1 2 0
Callicebus 0 1 {01} 0 {01} 1 1 3 1 0 0 1 0 0 1 1 1
Pithecia 0 2 0 0 0 0 0 3 2 0 0 1 0 0 2 1 1
Chiropotes 0 2 0 0 0 0 0 3 1 0 0 1 0 0 2 2 1
Cacajao 0 2 0 0 0 0 0 3 2 0 0 1 0 0 2 2 1
Proteropithecia ? 2 0 0 1 1 1 ? 2 0 0 ? ? ? ? ? ?
Saguinus ? 2 0 0 1 1 0 {45} 1 1 0 1 0 0 2 1 1
Callithrix ? 2 0 0 0 1 1 4 0 {01} 0 1 0 0 2 2 1
Cebuella pygmaea ? 2 0 0 0 2 1 5 0 1 0 1 0 0 1 0 2
Callimico goeldii 0 2 0 0 0 1 1 5 1 1 0 1 0 0 1 0 2
Leontopithecus ? 2 0 0 0 1 0 5 1 {01} 0 1 0 0 1 0 1
Alouatta 1 {12} 0 0 0 2 2 1 3 {01} {01} 1 0 0 0 0 0
Ateles 1 2 0 1 0 1 1 3 2 0 0 1 0 0 1 2 1
372 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Appendix 2 (continued )
Taxon 188 190 200 204
m39 m40 m41 m44 m45 m46 m47 m53 m54 m55 m57 I1 I2 I3 I4 I5 I6
Brachyteles 2 0 0 0 0 1 1 3 3 1 0 1 0 0 0 0 0
Lagothrix 1 2 0 0 0 1 1 3 2 0 0 ? 0 0 1 2 1
Branisella boliviana 2 1 0 0 0 1 1 3 1 0 0 ? ? ? ? ? ?
Dolichocebus gaimanensis ? 1 1 1 1 1 0 3 1 1 0 1 0 ? ? ? 1
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Soriacebus ameghinorum 1 2 0 1 0 0 0 3 1 2 0 ? ? ? ? ? ?
Carlocebus carmeni ? 2 1 0 0 1 1 4 2 0 0 ? ? ? ? ? ?
Hylobates lar 1 1 1 1 0 1 1 3 3 ? 0 1 0 0 ? ? ?
Miopithecus talapoin 2 0 0 0 0 1 1 3 3 ? 0 1 0 0 ? ? ?
Presbytis melalophos 2 0 0 0 0 1 1 2 3 ? 0 1 0 0 ? ? ?
Taxon 205 210 220 221
I7 I8 I9 I10 I11 I12 I13 C1 C2 C3 C4 P1 P2 P3 P4 P5 P6
Tarsius 1 2 0 ? 2 0 1 1 1 1 1 0 2 2 0 0 1
Aegyptopithecus zeuxis ? ? ? ? ? ? ? 0 3 1 1 ? 2 2 ? 1 ?
Apidium phiomense ? 0 ? ? ? ? ? ? 1 ? ? 2 2 2 1 1 0
Catopithecus browni 1 0 0 0 2 0 0 0 3 1 0 0 1 2 ? 1 ?
Proteopithecus sylviae ? 0 ? ? ? ? ? 0 1 1 1 ? 2 2 ? 2 0
Simonsius grangeri ? ? ? ? ? ? ? ? 1 ? ? 1 2 2 0 1 0
Aotus 0 0 0 0 2 0 0 {01} 1 0 1 1 1 1 2 {01} 0
Saimiri ? 0 0 0 2 0 0 1 1 1 2 0 0 1 1 {12} 0
Cebus 0 0 0 0 2 1 0 1 1 0 2 ? 1 1 2 2 0
Callicebus 0 0 0 0 2 1 0 1 1 0 2 1 1 1 1 0 0
Pithecia 1 0 0 0 1 1 0 1 1 0 ? 0 0 2 1 1 1
Chiropotes 0 0 0 0 1 1 0 0 1 0 1 ? ? ? 1 2 0
Cacajao 1 0 0 0 1 1 0 0 1 0 1 ? ? ? 1 2 0
Proteropithecia ? ? ? ? ? ? ? 1 ? 1 1 ? ? ? ? ? ?
Saguinus 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 1 0
Callithrix 0 0 0 0 0 0 0 {01} 1 ? 0 0 0 1 2 0 1
Cebuella pygmaea 1 0 0 0 0 0 0 0 1 1 0 0 0 1 0 {01} 1
Callimico goeldii 1 0 0 0 0 0 0 0 1 ? ? 0 1 0 2 0 0
Leontopithecus 1 0 0 0 1 0 0 0 1 1 0 0 1 1 1 1 1
Alouatta 1 0 0 0 1 0 0 0 1 ? 0 1 {12} {012} 0 0 1
Ateles 0 0 1 0 1 1 0 1 1 1 2 0 0 0 1 1 0
Brachyteles 1 0 1 0 ? 1 0 1 1 ? ? ? 1 1 0 {01} 1
Lagothrix 1 0 1 0 1 1 0 1 1 ? ? ? 0 0 2 1 0
Branisella boliviana ? ? ? ? ? ? ? 0 1 1 1 0 1 1 0 0 1
Dolichocebus gaimanensis ? 0 0 0 1 0 0 0 1 1 1 0 1 1 0 0 0
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? 1 ? ? ? ?
Soriacebus ameghinorum ? ? ? ? ? ? ? 0 1 1 0 1 2 2 1 1 0
Carlocebus carmeni ? ? ? ? ? ? ? ? 1 1 1 ? {01} 1 ? 0 ?
Hylobates lar ? 0 ? ? ? ? ? 0 3 1 2 ? 2 2 ? 1 ?
Miopithecus talapoin ? 0 ? ? ? ? ? 0 3 1 0 ? 2 2 ? 0 ?
Presbytis melalophos ? 0 ? ? ? ? ? 0 3 1 0 ? 2 2 ? 0 ?
Taxon 222 230 238
P8 P9 P10 P11 P12 P13 P13a P14 P15 P16 P17 P18 P19 P20 P21 M1 M2
Tarsius 1 1 0 0 1 0 0 2 1 0 0 1 0 0 0 0 0
Aegyptopithecus zeuxis 1 0 0 0 ? 2 1 2 1 0 0 0 1 0 0 0 0
Apidium phiomense 1 0 0 0 0 3 1 0 0 0 0 0 0 0 0 0 0
Catopithecus browni ? 0 0 0 ? 0 0 2 0 0 0 1 0 0 1 0 0
Proteopithecus sylviae ? 0 0 0 0 0 0 ? 0 1 1 1 0 ? 1 0 1
Simonsius grangeri ? 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0
Aotus 1 0 0 0 0 0 0 2 1 0 0 0 0 0 0 1 2
Saimiri 1 0 0 0 0 0 0 2 1 0 0 0 1 0 0 1 2
Cebus 1 0 0 0 0 1 {01} 2 1 1 0 0 0 0 0 1 2
Callicebus 1 0 0 0 0 1 1 2 1 0 0 0 1 0 0 0 1
Pithecia 1 0 0 0 0 {01} 1 2 1 0 0 0 0 0 0 1 2
Chiropotes 1 0 1 0 0 0 1 2 0 1 0 0 0 0 0 ? ?
Cacajao 1 0 1 0 ? 1 1 2 0 1 0 0 0 0 0 ? ?
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
(continued on next page)
373R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Appendix 2 (continued)
Taxon 222 230 238
P8 P9 P10 P11 P12 P13 P13a P14 P15 P16 P17 P18 P19 P20 P21 M1 M2
Saguinus 0 0 0 0 {01} 0 0 2 1 0 0 1 0 0 0 0 ?
Callithrix 0 0 0 0 1 0 0 2 0 0 1 1 0 0 0 0 ?
Cebuella pygmaea 0 0 0 0 1 0 0 2 0 0 1 1 0 0 0 0 ?
Callimico goeldii 0 0 0 0 0 0 0 2 1 0 0 1 0 0 0 0 1
Leontopithecus 1 0 0 0 0 0 0 2 1 0 0 0 1 0 0 0 ?
Alouatta 0 0 0 0 0 {012} {01} 2 1 0 0 0 0 0 {01} 0 2
Ateles 0 0 0 0 0 0 0 2 1 0 0 0 0 0 0 2 2
Brachyteles 0 0 0 0 0 0 0 2 0 1 0 0 0 0 1 ? ?
Lagothrix 0 0 0 0 0 0 0 2 1 0 0 0 0 0 0 ? 2
Branisella boliviana 1 0 0 0 0 0 0 2 0 1 1 0 1 0 0 0 1
Dolichocebus gaimanensis 1 0 0 0 1 2 2 2 0 1 1 0 1 0 0 0 1
Tremacebus harringtoni ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 1 2
Soriacebus ameghinorum 0 0 0 0 0 2 2 2 1 0 1 0 1 0 0 0 ?
Carlocebus carmeni 0 ? 0 0 ? {123} 2 2 1 0 1 ? 1 0 0 {01} ?
Hylobates lar 0 0 0 0 ? 0 0 2 1 ? 0 0 ? 1 0 0 ?
Miopithecus talapoin 0 0 0 0 ? 0 0 2 1 0 0 0 0 1 0 0 ?
Presbytis melalophos ? 0 0 0 ? 0 0 2 1 0 0 0 0 1 0 0 ?
Taxon 239 240 250 255
M3 M4 M5 M6 M7 M8 M9 M10 M11 M12 M13 M13a M14 M15 M16 M17 M18
Tarsius 1 1 0 0 0 2 0 2 2 ? 0 ? 0 0 1 0 0
Aegyptopithecus zeuxis 1 2 0 0 0 1 1 0 0 0 0 ? 0 0 1 0 0
Apidium phiomense 1 2 0 0 0 2 2 0 0 1 1 0 1 1 2 0 2
Catopithecus browni 1 2 0 0 0 1 0 1 1 0 0 ? 0 0 0 0 0
Proteopithecus sylviae 0 1 0 0 0 0 0 1 1 0 0 ? 0 ? {01} 0 0
Simonsius grangeri 2 1 0 0 0 2 0 0 0 0 0 ? 0 1 1 0 2
Aotus 2 1 0 0 0 0 0 0 0 0 2 0 0 ? 0 0 0
Saimiri {01} 1 0 0 0 0 0 0 0 0 1 0 0 ? 0 0 0
Cebus 1 1 0 0 0 0 0 0 0 0 2 1 0 ? 1 0 1
Callicebus {12} 1 0 0 0 {01} 0 0 0 0 2 0 0 ? {01} 0 1
Pithecia 2 1 0 0 0 0 0 0 0 0 2 0 0 ? 0 0 1
Chiropotes 2 1 0 0 0 0 0 0 1 0 1 0 0 ? 0 0 1
Cacajao 2 1 0 0 0 0 0 0 0 0 1 0 0 ? 0 0 1
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 0 0 0 0 0 0 0 2 2 ? ? ? ? ? 0 2 0
Callithrix 1 0 0 0 0 0 0 2 2 ? ? ? ? ? 0 2 0
Cebuella pygmaea 1 0 0 0 0 0 0 2 2 ? ? ? ? ? 0 2 0
Callimico goeldii 1 0 0 0 0 1 0 1 2 0 0 0 0 0 1 2 0
Leontopithecus {12} 0 0 0 0 0 0 2 2 ? ? ? ? ? 0 2 0
Alouatta 2 2 0 0 0 0 0 0 0 0 1 0 0 ? {01} 1 1
Ateles 2 {12} 0 0 0 0 0 0 0 0 1 0 1 ? 0 0 1
Brachyteles 2 1 0 0 0 0 0 0 0 0 0 ? 1 ? {01} 2 1
Lagothrix 2 {12} 0 0 0 0 0 0 0 0 0 ? 0 ? 0 0 1
Branisella boliviana 1 1 0 0 0 0 0 1 1 0 0 ? 0 ? {01} {02} 1
Dolichocebus gaimanensis 0 2 0 0 ? 1 1 0 0 0 1 0 ? 0 0 2 0
Tremacebus harringtoni 1 1 0 0 ? ? ? 1 1 0 ? 0 ? ? ? ? ?
Soriacebus ameghinorum 1 1 0 0 ? 0 0 1 1 0 1 0 0 ? 0 0 1
Carlocebus carmeni 1 ? 0 0 ? 1 ? 0 0 0 {012} 0 0 ? ? 0 0
Hylobates lar 2 2 0 0 0 0 0 0 0 0 2 0 ? ? 0 0 0
Miopithecus talapoin 2 2 0 0 0 0 0 0 0 0 2 0 ? ? 0 0 1
Presbytis melalophos 2 2 0 0 0 0 0 0 0 0 2 0 ? ? 0 0 1
Taxon 256 260 268
M19 M20 M22 M24 M27 M28 M30 M31 M33 M34 M36 M44 M46
Tarsius 0 0 2 2 0 0 0 0 0 2 0 0 2
Aegyptopithecus zeuxis 0 0 2 1 0 0 0 1 1 0 0 1 2
Apidium phiomense 1 2 2 1 0 0 2 1 1 0 1 1 1
Catopithecus browni 1 0 2 2 0 0 0 0 0 1 0 1 2
Proteopithecus sylviae 1 0 2 2 0 0 0 0 0 0 0 1 0
Simonsius grangeri 1 0 2 {12} 0 0 1 1 1 0 0 1 0
Aotus 0 0 {01} 0 0 0 0 0 0 0 1 2 1
374 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
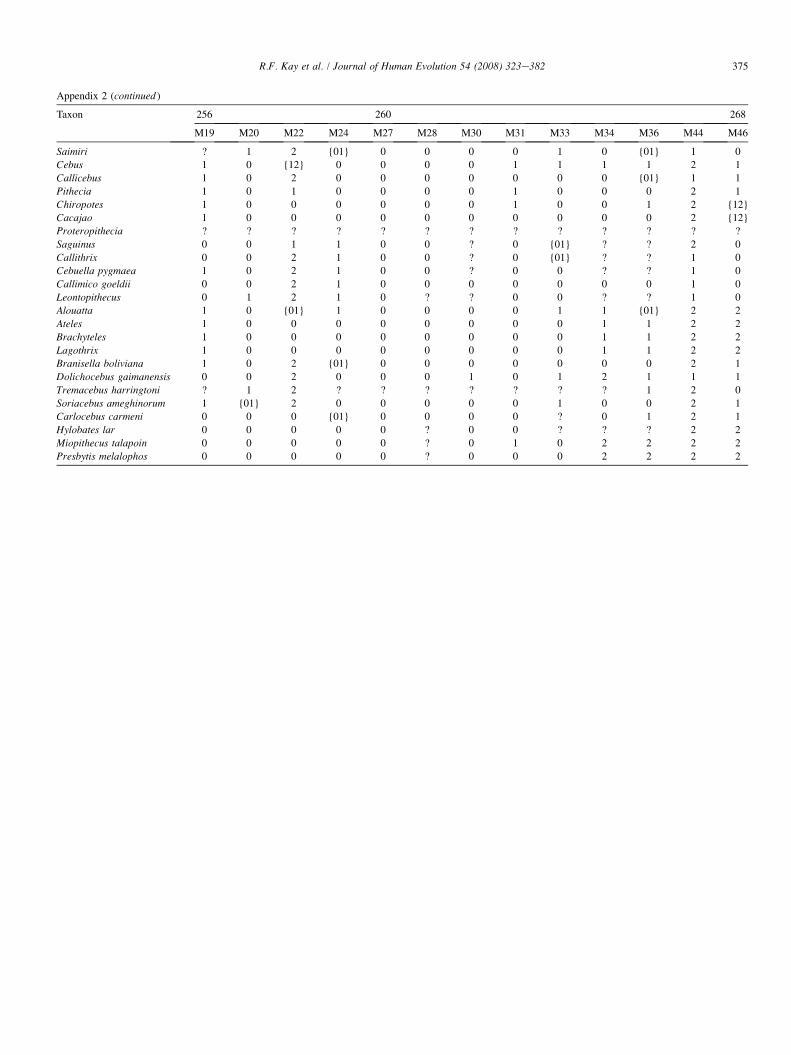
Appendix 2 (continued )
Taxon 256 260 268
M19 M20 M22 M24 M27 M28 M30 M31 M33 M34 M36 M44 M46
Saimiri ? 1 2 {01} 0 0 0 0 1 0 {01} 1 0
Cebus 1 0 {12} 0 0 0 0 1 1 1 1 2 1
Callicebus 1 0 2 0 0 0 0 0 0 0 {01} 1 1
Pithecia 1 0 1 0 0 0 0 1 0 0 0 2 1
Chiropotes 1 0 0 0 0 0 0 1 0 0 1 2 {12}
Cacajao 1 0 0 0 0 0 0 0 0 0 0 2 {12}
Proteropithecia ? ? ? ? ? ? ? ? ? ? ? ? ?
Saguinus 0 0 1 1 0 0 ? 0 {01} ? ? 2 0
Callithrix 0 0 2 1 0 0 ? 0 {01} ? ? 1 0
Cebuella pygmaea 1 0 2 1 0 0 ? 0 0 ? ? 1 0
Callimico goeldii 0 0 2 1 0 0 0 0 0 0 0 1 0
Leontopithecus 0 1 2 1 0 ? ? 0 0 ? ? 1 0
Alouatta 1 0 {01} 1 0 0 0 0 1 1 {01} 2 2
Ateles 1 0 0 0 0 0 0 0 0 1 1 2 2
Brachyteles 1 0 0 0 0 0 0 0 0 1 1 2 2
Lagothrix 1 0 0 0 0 0 0 0 0 1 1 2 2
Branisella boliviana 1 0 2 {01} 0 0 0 0 0 0 0 2 1
Dolichocebus gaimanensis 0 0 2 0 0 0 1 0 1 2 1 1 1
Tremacebus harringtoni ? 1 2 ? ? ? ? ? ? ? 1 2 0
Soriacebus ameghinorum 1 {01} 2 0 0 0 0 0 1 0 0 2 1
Carlocebus carmeni 0 0 0 {01} 0 0 0 0 ? 0 1 2 1
Hylobates lar 0 0 0 0 0 ? 0 0 ? ? ? 2 2
Miopithecus talapoin 0 0 0 0 0 ? 0 1 0 2 2 2 2
Presbytis melalophos 0 0 0 0 0 ? 0 0 0 2 2 2 2
375R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382

Appendix 3. Specimens examined
Taxon Higher-level
taxon
Subtaxon Cranial material examined Dental measurements
(sample size)
Dental material examined
Branisella boliviana From Takai et al.
2000
MNHN Bol V 3447, 3468, 5310, 5311
Tremacebus harringtoni Type specimen, Tucuman,
Argentina
Type specimen, Tucuman, Argentina
Carlocebus carmenensis See Fleagle, 1990 MACN SC 11, 113, 266, 400
Alouatta caraya Atelidae A. caraya group 20 USNM 238675, 390645, 390648,
518237, 518238, 518233
Alouatta palliata Atelidae A. palliata group FMNH 44330, 44331, USNM
240406, 240407, 240408,
Kay lab
50
Alouatta pigra Atelidae A. palliata group 22 BMNH 11.7.27.1
Alouatta belzebul Atelidae A. seniculus group 20 USNM 544508, MSP 8067, 8071
Alouatta guariba Atelidae A. seniculus group 18 USNM 518256, 38392,113436, 518249,
518248, 518246/5, 518253, 518255,
518257, 518247, MSP 5868, 2346
Alouatta macconnelli Atelidae A. seniculus group 22
Alouatta seniculus Atelidae A. seniculus group 51
Ateles fusciceps Atelidae Atelinae 7 USNM 284932, 291240, 291242,
291243
Ateles geoffroyi Atelidae Atelinae USNM 14162, 14128/12128,
USNM 14161, Kay lab
specimen
44 USNM 108529, 244869, 13048, 14161,
14154, 14116, 13297, 171075, 284948,
291056, 291057, 291060, 291064,
315840, 337694, 337698, 337707
Ateles marginatus Atelidae Atelinae MSP 1933019329
Ateles paniscus Atelidae Atelinae FMNH 2138, 21385 29 MSP 5277
Brachyteles cf. arachnoides Atelidae Atelinae USNM 259474 10 ANS 2744, 2685, BMNH 48.10.25.1,
USNM 259474, MSP 1159, 1863, 3533,
1158, 2236, 11100, 11098, 19360,
11104, 1160
Lagothrix cana Atelidae Atelinae USNM 398460, 397775, 194341,
194342, 399302, 269839, 311217, MSP
917, 5533, 11233
Lagothrix lagotricha Atelidae Atelinae USNM 545885, 194342, Kay
lab, FMNH 98053, 98052
ANS 2872, USNM 194342, 398460,
397963, 398460, MSP 917
Lagothrix poeppigii Atelidae Atelinae 23
Callimico goeldii Callitrichinae USNM 303323, 464991,
573934, 528737, 583199,
FMNH 98034, FMNH 60354,
2 USNM 395455, 303323
Cebuella pygmaea Callitrichinae USNM 336310, 336311 37 USNM 319513, 337948, 336302-336314
(12 specimens) 336311, 336313, 336314,
336323, 337325
Leontopithecus chrysomelas Callitrichinae Kay lab skull 4
Leontopithecus rosalia Callitrichinae USNM 546318, 546321,
546317, 546322, 546323,
FMNH 57838, 46165
18 ANS 13497, USNM 395454, 534283,
546320
Callithrix penicillata Callitrichinae Callithrix group 16 USNM 518558-518561 (4 specimens),
518564, 518566e518568
37
6R
.F.
Kay
etal.
/Journal
ofH
uman
Evolution
54(2008)
323e382
,

Callithrix geoffroyi Callitrichinae Callithrix group FMNH 134473, 134474
Callithrix jacchus Callitrichinae Callithrix group UT 0434, USNM 503895,
503886
10 USNM 518554, 518555
Callithrix argentata Callitrichinae Mico group USNM 23459, 239458, 239457, 239561
Saguinus midas Callitrichinae S. midas group 57 USNM 393807
Saguinus mystax Callitrichinae S. mystax group USNM 544383, 544382 USNM 543489, 397242, 397873,
397877, 397930
Saguinus fuscicollis Callitrichinae S. nigricollis group USNM 398731, 336299, UT
0635, FMNH 24194, 24195
34 ANS 16565, USNM 518576, 397302,
skull in Kay collection
Saguinus geoffroyi Callitrichinae S. oedipus group 42 USNM 301653, 336296
Saguinus leucopus Callitrichinae S. oedipus group USNM 281630
Cebus apella Cebidae C. apella group USNM 296639, 361020,
338958, 268935, 33956,
296638, 296640, 338957,
38959, 361019, FMNH
20000, 461384, 46182
USNM 397959, 397979
Cebus libidinosus Cebidae C. apella group 40 USNM 518281, 518283, 518336,
518341, 518347, 518349, 518350,
518466, 518471
Cebus albifrons Cebidae C. capucinus group Kay lab skull USNM 398445
Cebus capucinus Cebidae C. capucinus group USNM 291234, 283945 53
Cebus olivaceus Cebidae C. capucinus group USNM 261913
Saimiri boliviensis Cebidae S. boliviensis group USNM 396265, 397316,
397252, FMNH 55498,
55499, 55500
46
Saimiri oerstedti Cebidae S. sciureus group USNM 291092, 3693149,
3963148, 291045
35
Saimiri sciureus Cebidae S. sciureus group Kay lab skull USNM 194345, 241409, 393795,
398676, 398691, 398704, 398708,
398709, 546267, 393795
Aotus sp. Cebidae Kay lab skull
Aotus lemurinus Cebidae Grey-neck Aotus
group
FMNH 68858, 68860 44
Aotus trivirgatus Cebidae Grey-neck Aotus
group
USNM 514971, 464370,
503709
31 ANS 19813, USNM 171078, 396157,
396429, 396715, 396716, 396720,
396722, 396724, BMNH 45.6.17.6,
73.4.23.2
Callicebus cupreus Pitheciidae C. moloch group FMNH 25335, 25334
Callicebus moloch Pitheciidae C. moloch group 25 BMNH 8.5.9.8, 26.5.5.12, 26.5.5.10,
26.5.5.17
Callicebus ornatus Pitheciidae C. moloch group USNM 283924, 283953
Callicebus personatus Pitheciidae C. moloch group BMNH 3.9.5.2, USNM 518221
Callicebus torquatus Pitheciidae C. torquatus group USNM 406410, 406411,
406412, 406415, 406416,
406417, 406414, 388149,
388149, 406567, 406568,
406569, 406570
20 ANS 14306, USNM 256213
Chiropotes albinasus Pitheciidae Pitheciinae FMNH 94926, 94927 USNM 546262
Chiropotes satanas Pitheciidae Pitheciinae USNM 549519, 338964 34 USNM338962, 406593, 406594
Pithecia monachus Pitheciidae Pitheciinae USNM 461918, 461919, 545891,
518224, 518223
(continued on next page)
Appendix 3 (continued)
Taxon Higher-level
taxon
Subtaxon Cranial material examined Dental measurements
(sample size)
Dental material examined
Pithecia pithecia Pitheciidae Pitheciinae USNM 339658, 339660,
374743, 374744, 374745,
374749, 374748, 374763,
374767, FMNH 50883,
46176, Kay lab skull
25 ANS 1263, USNM 339659
Cacajao calvus Pitheciidae Pitheciinae FMNH 88814, FMNH 88819 19
Cacajao melanocephalus Pitheciidae Pitheciinae USNM 406423, 406422 USNM 256216
Tarsius bancanus borneanus Tarsiidae USNM 488081, 488084,
546331
Tarsius syrichta Tarsiidae FMNH 56763, Duke
osteological collection
USNM 282761, Duke Osteological
collection
Hylobates lar Catarrhini Hylobatidae USNM 111990 40
Miopithecus talapoin Catarrhini Cercopithecidae USNM 395340, 220338 39 Skull in Kay collection
Presbytis melalophos Catarrhini Cercopithecidae USNM 113174 31 Kay collection
Aegyptopithecus zeuxis Catarrhini Propliopithecidae DPC 5401, 6642, CGM
85785
Metrics from Kay t
al., 1981
DPC 1112, CGM 40237, DPC 1112,
CGM 40237, other specimens cited in
Kay et al., 1981.
Apidium phiomense Stem
Anthropoidea
Parapithecidae YPM 23978, 25972, 23968 Metrics from Kay
(unpublished)
DPC 3080, 3871; YPM 21018, 21016
Simonsius (¼ Parapithecus) grangeri Stem
Anthropoidea
Parapithecidae DPC 18651 Metrics from Kay
(unpublished)
DPC 1009, 1091, 2807, type specimen
CGM 26912, 1118, 3110
Catopithecus browni Stem
Anthropoidea
Oligopithecidae DPC 11388 Metrics from Sim s
et al., 1999
CGM 42200, DPC FN 95-1039, FN 95-
771, FN 95-894
Proteopithecus sylviae Stem
Anthropoidea
Proteopithecidae DPC 14095 Metrics from Sim s
et al.,1999
CGM 42200, 42206, DPC 95-1039
37
8R
.F.
Kay
etal.
/Journal
ofH
uman
Evolution
54(2008)
323e382
e
on
on

Appendix 4: Percentage completeness of the data set
Taxon Number of characters missing* Percentage complete (%)
Total Cranial Dental Total Cranial Dental
Tarsius 14 4 10 94.8 95.3 94.5
Aegyptopithecus zeuxis 37 9 28 86.2 89.4 84.7
Apidium phiomense 63 42 21 76.5 50.6 88.5
Catopithecus browni 51 35 16 81.0 58.8 91.3
Proteopithecus sylviae 65 25 40 75.7 70.6 78.1
Simonsius grangeri 53 15 38 80.2 82.4 79.2
Aotus 12 3 9 95.5 96.5 95.1
Saimiri 12 2 10 95.5 97.6 94.5
Cebus 14 4 10 94.8 95.3 94.5
Callicebus 11 3 8 95.9 96.5 95.6
Pithecia 12 2 10 95.5 97.6 94.5
Chiropotes 22 7 15 91.8 91.8 91.8
Cacajao 20 4 16 92.5 95.3 91.3
Proteropithecia 207 85 122 22.8 0.0 33.3
Saguinus 25 2 23 90.7 97.6 87.4
Callithrix 27 3 24 89.9 96.5 86.9
Cebuella 26 3 23 90.3 96.5 87.4
Callimico 10 0 10 96.3 100.0 94.5
Leontopithecus 27 4 23 89.9 95.3 87.4
Alouatta 11 2 9 95.9 97.6 95.1
Ateles 12 2 10 95.5 97.6 94.5
Brachyteles 21 5 16 92.2 94.1 91.3
Lagothrix 19 3 16 92.9 96.5 91.3
Branisella boliviana 102 80 22 61.9 5.9 88.0
Dolichocebus gaimanensis 55 31 24 79.5 63.5 86.9
Tremacebus harringtoni 197 34 163 26.5 60.0 10.9
Soriacebus ameghinorum 108 84 24 59.7 1.2 86.9
Carlocebus carmeni 145 84 61 45.9 1.2 66.7
Hylobates 38 3 35 85.8 96.5 80.9
Miopithecus talapoin 34 2 32 87.3 97.6 82.5
Presbytis melalophos 33 2 31 87.7 97.6 83.1
* ‘‘Missing’’ data includes characters that cannot be scored or are not observed either because pertinent material was not examined or anatomical material is
unknown.
379R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
References
Ameghino, F., 1894. Enumeration synoptique des especes de mammiferes
fossiles des formations Eogenes de Patagonie. Bol. Acad. Nac. Cien.
Cordoba 13, 259e451.
Ashley-Montague, M.F., 1933. The anthropological significance of the pterion
in the primates. Am. J. Phys. Anthropol. 13, 159e336.
Barroso, C.M.L., Schneider, H., Schneider, M.P.C., Sampaio, I., Harada, M.L.,
Czelusniak, J., Goodman, M., 1997. Update on the phylogenetic systematics
of New World monkeys: Further DNA evidence for placing pygmy marmoset
(Cebuella) within the genus Callithrix. Int. J. Primatol. 18, 651e674.
Bashir, A., Ye, C., Price, A.L., Bafna, V., 2005. Orthologous repeats and mam-
malian phylogenetic inference. Genome Res. 15, 998e1006.
Beard, K.C., MacPhee, R.D.E., 1994. Cranial anatomy of Shoshonius and the
antiquity of Anthropoidea. In: Fleagle, J.G., Kay, R.F. (Eds.), Anthropoid
Origins: The Fossil Evidence. Plenum Press, New York, pp. 55e98.
Berggren, W.A., Kent, D.V., Swisher III, C.C., Aubry, M.-P., 1995. A revised
Cenozoic geochronology and chronostratigraphy. In: Berggren, W.A.,
Kent, D.V., Aubry, M.-P., Hardenbol, J. (Eds.), Geochronology, Time
Scales, and Global Stratigraphic Correlation. Society for Sedimentary
Geology, Tulsa, pp. 129e212.
Bordas, A.F., 1939. Diagnosis sobre algunos mamiferos de als capas con
Colpodon del valle del Rio Chubut. Physis 14, 413e433.
Bordas, A.F., 1942. Anotaciones sobre un ‘Cebidae’ fosil de Patagonia. Physis
19, 265e269.
Bown, T.M., Larriestra, C.N., Powers, D.W., Naeser, C.W., Tabbutt, K., 1988.
New information on age, correlation, and paleoenvironments of fossil
platyrrhine sites in Argentina. J. Vert. Paleontol. 8, 9A.
Bremer, K., 1994. Branch support and tree stability. Cladistics 10, 295e304.
Cartmill, M., 1978. The orbital mosaic in prosimians and the use of variable
traits in systematics. Folia Primatol. 30, 89e114.
Cartmill, M., Kay, R.F., 1978. Craniodental morphology, tarsier affinities, and
primate suborders. In: Chivers, D.J., Joysey, K.A. (Eds.), Recent Advances
in Primatology: Evolution. Academic Press, London, pp. 205e214.
Cartmill, M., MacPhee, R.D.E., Simons, E.L., 1981. Anatomy of the temporal
bone in early anthropoids with remarks on the problem of anthropoid
origins. Am. J. Phys. Anthropol. 56, 3e22.
Conroy, G., 1980. Evolutionary significance of cerebral venous patterns in
paleoprimatology. Z. Morphol. Anthropol. 71, 125e134.
Conroy, G., 1981. Cranial asymmetry in ceboid primates: the emissary foram-
ina. Am. J. Phys. Anthropol. 55, 187e194.
Conroy, G.C., 1987. Problems of body-weight estimation in fossil primates.
Int. J. Primatol. 8, 115e137.
Cozzuol, M.A., Fordyce, R.E., Jones, C.M., 1991. La presencia de Eretiscustonnii (Aves, Spheniscidae) en el Mioceno temprano de Nueva Zelandia
y Patagonia. Ameghiniana 28, 406.
Dagosto, M., Gebo, D.L., 1994. Postcranial anatomy and the origin of the
Anthropoidea. In: Fleagle, J.G., Kay, R.F. (Eds.), Anthropoid Origins:
The Fossil Evidence. Plenum Press, New York, pp. 567e594.
Du Brul, E.L., 1965. The skull of the lion marmoset, Leontideus rosalia Lin-
naeus: A study in biomechanical adaptation. Am. J. Phys. Anthropol. 23,
261e276.
Eizirik, E., Murphy, W.J., Springer, M., O’Brien, S.J., 2004. Molecular phy-
logeny and dating of early primate divergences. In: Ross, C., Kay, R.F.
(Eds.), Anthropoid Origins: New Visions. Kluwer Academic/Plenum
Publishers, New York, pp. 45e64.

380 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Fleagle, J.G., 1985. New primate fossils from Colhuehuapian deposits at
Gaiman and Sacanana, Chubut Province, Argentina. Ameghiniana 21,
266e274.
Fleagle, J.G., 1990. New fossil platyrrhines from the Pinturas Formation,
southern Argentina. J. Hum. Evol. 19, 61e85.
Fleagle, J.G., Bown, T.M., 1983. New primate fossils from late Oligocene
(Colhuehuapian) localities of Chubut Province, Argentina. Folia Primatol.
41, 240e266.
Fleagle, J.G., Bown, T.M., Swisher III, C.C., Buckley, G.A., 1995. Age of the
Pinturas and Santa Cruz formations. VI Congreso Argentino de Paleonto-
logia y Bioestratigrafia, Trelew, pp. 129e135.
Fleagle, J.G., Kay, R.F., 1983. New interpretations of the phyletic position of Ol-
igocene hominoids. In: Ciochon, R.L., Corruccini, R.S. (Eds.), New Interpre-
tations of Ape and Human Ancestry. Plenum Press, New York, pp. 181e210.
Fleagle, J., Kay, R.F., 1987. The phyletic position of the Parapithecidae.
J. Hum. Evol. 16, 483e532.
Fleagle, J.G., Kay, R.F., 1989. The dental morphology of Dolichocebus
gaimanensis, a fossil monkey from Argentina. Am. J. Phys. Anthropol.
78, 221.
Fleagle, J.G., Kay, R.F., Anthony, M.R.L., 1997. Fossil New World monkeys.
In: Kay, R.F., Madden, R.H., Cifelli, R.L., Flynn, J.J. (Eds.), Mammalian
Evolution in the Neotropics. Smithsonian Institution Press, Washington,
DC, pp. 473e495.
Fleagle, J.G., Powers, D.W., Conroy, G.C., Watters, J.P., 1987. New fossil plat-
yrrhines from Santa Cruz Province, Argentina. Folia Primatol. 48, 65e77.
Fleagle, J.G., Rosenberger, A.L., 1983. Cranial morphology of the earliest an-
thropoids. In: Sakka, M. (Ed.), Morphologie Evolution Morphogenese du
Crane et Origine de l’Homme. Centre National de Recherche Scientifique,
Paris, pp. 141e155.
Fleagle, J.G., Tejedor, M.F., 2002. Early platyrrhines of southern South Amer-
ica. In: Hartwig, W.C. (Ed.), The Primate Fossil Record. Cambridge
University Press, Cambridge, pp. 161e173.
Flynn, J.J., Swisher III, C.C., 1995. Cenozoic South American land mammal
ages: correlation to global geochronologies. In: Berggren, W.A.,
Kent, D.V., Aubry, M.-P., Hardenbol, J. (Eds.), Geochronology, Time
Scales, and Global Stratigraphic Correlation. Society for Sedimentary
Geology, Tulsa, pp. 317e333.
Flynn, J.J., Wyss, A.R., Charrier, R., Swisher, C.C., 1995. An early Miocene
anthropoid skull from the Chilean Andes. Nature 373, 603e607.
Ford, S.M., 1980. Callitrichids as phyletic dwarfs, and the place of the Calli-
trichidae in Platyrrhini. Primates 21, 31e43.
Ford, S.M., 1986. Systematics of the New World monkeys. In: Swindler, D.R.,
Erwin, J. (Eds.), Comparative Primate Biology, Vol. 1: Systematics, Evolu-
tion, and Anatomy. Alan R. Liss, New York, pp. 73e135.
Ford, S.M., 1994. Primitive platyrrhines? Perspectives on anthropoid origins
from platyrrhine, parapithecid, and preanthropoid postcrania. In:
Fleagle, J.G., Kay, R.F. (Eds.), Anthropoid Origins: The Fossil Evidence.
Plenum Press, New York, pp. 595e676.
Forsyth-Major, C.I., 1901. On some characters of the skull in the lemurs and
monkeys. Proc. Zool. Soc. Lond. 1901, 129e153.
Gebo, D., Simons, E.L., 1987. Morphology and locomotor adaptations of the
foot in early Oligocene anthropoids. Am. J. Phys. Anthropol. 74, 83e101.
Gebo, D.L., Simons, E.L., Rasmussen, D.T., Dagosto, M., 1994. Eocene an-
thropoid postcrania from the Fayum, Egypt. In: Fleagle, J.G., Kay, R.F.
(Eds.), Anthropoid Origins: The Fossil Evidence. Plenum Press, New
York, pp. 203e234.
Harada, M.L., Schneider, H., Schneider, M.P.C., Sampaio, I., Czelusniak, J.,
Goodman, M., 1995. DNA evidence on the phylogenetic systematics of
New World monkeys: Support for the sister grouping of Cebus and Saimiri
from two unlinked nuclear genes. Mol. Phylogenet. Evol. 4, 331e349.
Hartwig, W.C., 1994. Patterns, puzzles, and perspectives on platyrrhine
origins. In: Corruccini, R.S., Ciochon, R. (Eds.), Integrative Paths to the
Past: Paleoanthropological Advances in Honor of F. Clark Howell. Prentice
Hall, Englewood Cliffs, pp. 69e94.
Hauser, D.L., Presch, W., 1991. The effect of ordered characters on phyloge-
netic reconstruction. Cladistics 7, 243e265.
Heesy, C.P., Ross, C., 2004. Mosaic evolution of activity pattern, diet, and
color vision in haplorhine primates. In: Ross, C., Kay, R.F. (Eds.),
Anthropoid Origins: New Visions. Kluwer Academic/Plenum Publishers,
New York, pp. 649e682.
Hershkovitz, P., 1949. Mammals of northern Colombia. Preliminary report
no. 4: monkeys (Primates) with taxonomic revisions of some forms.
Proc. U.S. Nat. Mus. 98, 323e427.
Hershkovitz, P., 1970. Notes on Tertiary platyrrhine monkeys and description
of a new genus from the late Miocene of Colombia. Folia Primatol. 12,
1e37.
Hershkovitz, P., 1974. A new genus of late Oligocene monkey (Cebidae,
Platyrrhini) with notes on postorbital closure and platyrrhine evolution.
Folia Primatol. 21, 1e35.
Hershkovitz, P., 1977. Living New World Monkeys (Platyrrhini) with an Intro-
duction to Primates, vol. 1. University of Chicago Press, Chicago.
Hershkovitz, P., 1981. Comparative anatomy of platyrrhine mandibular cheek
teeth dpm4, pm4, m1, with particular reference to those of Homunculus
(Cebidae), and comments on platyrrhine origins. Folia Primatol. 35,
179e217.
Hershkovitz, P., 1982. Supposed squirrel monkey affinities of the late Oligo-
cene Dolichocebus gaimanensis. Nature 298, 201e202.
Hershkovitz, P., 1984. Taxonomy of squirrel monkeys genus Saimiri: a prelim-
inary report with description of a hitherto unnamed form. Am. J. Primatol.
7, 155e210.
Hershkovitz, P., 1990. Titis, New World monkeys of the genus Callicebus(Cebidae, Platyrrhini): A preliminary taxonomic review. Fieldiana Zool.
55, 1e109.
Hill, W.C.O., 1957. Primates. Comparative Anatomy and Taxonomy III: Pith-
ecoidea, Platyrrhini, Hapalidae. University Press, Edinburgh.
Hillis, D.M., 1999. SINEs of the perfect character. Proc. Natl. Acad. Sci.
U.S.A. 96, 9979e9981.
Hoffstetter, R., 1980. Los monos platirrinos (Primates): origen, extension, filo-
genia, taxonomia. Actas II Congreso Argentino de Paleontologıa y Bioes-
tratigrafıa y I Congreso Latinoamericano de Paleontologıa, 1978. Buenos
Aires, pp. 291e303.
Horovitz, I., 1997. Platyrrhine systematics and the origin of Greater Antilles
monkeys. Ph.D. Dissertation, State University of New York.
Horovitz, I., 1999. A phylogenetic study of living and fossil platyrrhines. Am.
Mus. Novit. 3269, 1e40.
Horovitz, I., MacPhee, R.D.E., 1999. The quaternary Cuban platyrrhine
Paralouatta varonai and the origin of Antillean monkeys. J. Hum. Evol.
36, 33e68.
Houle, A., 1999. The origin of platyrrhines: An evaluation of the Antarctic
scenario and the floating island model. Am. J. Phys. Anthropol. 109,
541e559.
Kay, R.F., 1977. The evolution of molar occlusion in Cercopithecidae and
early catarrhines. Am. J. Phys. Anthropol. 46, 327e352.
Kay, R.F., 1980. Platyrrhine origins: a reappraisal of the dental evidence. In:
Ciochon, R., Chiarelli, B. (Eds.), Evolutionary Biology of the New World
Monkeys and Continental Drift. Plenum Press, New York, pp. 159e188.
Kay, R.F., 1990. The phyletic relationships of extant and fossil Pitheciinae
(Platyrrhini, Anthropoidea). J. Hum. Evol. 19, 175e208.
Kay, R.F., 1994. ‘‘Giant’’ tamarin from the Miocene of Colombia. Am. J. Phys.
Anthropol. 95, 333e353.
Kay, R.F., Campbell, V.M., Rossie, J.B., Colbert, M.W., Rowe, T., 2004a.
The olfactory fossa of Tremacebus harringtoni (Platyrrhini, early Miocene,
Sacanana, Argentina): Implications for activity pattern. Anat. Rec 281A,
1157e1172.
Kay, R.F., Cozzuol, M.A., 2006. A new platyrrhine from the Solim~oes Forma-
tion (late Miocene, Acre State, Brazil) with comments about other
Miocene monkeys from that region. J. Hum. Evol. 50, 673e686.
Kay, R.F., Delson, E., 2000. Oligopithecidae. In: Delson, E., Tattersall, I., Van
Couvering, J.A., Brooks, A. (Eds.), Encyclopedia of Human Evolution
and Prehistory, second ed. Garland Publishing Co., New York, pp. 490e
493.
Kay, R.F., Fleagle, J.G., Simons, E.L., 1981. A revision of the African
Oligocene apes of the Fayum Province. Egypt. Am. J. Phys. Anthropol.
55, 293e322.
Kay, R.F., Johnson, D.J., Meldrum, D.J., 1998a. A new pitheciin primate from
the middle Miocene of Argentina. Am. J. Primatol. 45, 317e336.

381R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
Kay, R.F., Kirk, E.C., 2000. Ostological evidence for the evolution of activity
pattern and visual acuity in primates. Am. J. Phys. Anthropol. 113,
235e262.
Kay, R.F., Kirk, E.C., Malinzak, M., Colbert, M.W., 2006. Brain size, activity
pattern, and visual acuity in Homunculus patagonicus, an early Miocene
stem platyrrhine: the mosaic evolution of brain size and visual acuity in
Anthropoidea. J. Vert. Paleontol. 26, 83Ae84A.
Kay, R.F., MacFadden, B.J., Madden, R.H., Sandeman, H., Anaya, F., 1998b.
Revised age of the Salla beds, Bolivia, and its bearing on the age of the
Deseadan South American Land Mammal ‘Age.’ J. Vert. Paleontol. 18,
189e199.
Kay, R.F., Madden, R.H., Mazzoni, M., Vucetich, M.G., Re, G., Heizler, M.,
Sandeman, H., 1999b. The oldest Argentine primates: first age determina-
tions for the Colhuehuapian South American Land Mammal ‘Age.’ Am. J.
Phys. Anthropol. 28 (Suppl.), 166.
Kay, R.F., Madden, R.M., Vucetich, M.G., Carlini, A.A., Mazzoni, M.M.,
Re, G.H., Heizler, M., Sandeman, H., 1999a. Revised age of the Casa-
mayoran South American Land Mammal ‘‘Age’’dclimatic and biotic
implications. Proc. Natl. Acad. Sci. U.S.A. 96, 13235e13240.
Kay, R.F., Meldrum, D.J., 1997. A new small platyrrhine from the Miocene of
Colombia and the phyletic position of Callitrichinae. In: Kay, R.F.,
Madden, R.H., Cifelli, R.L., Flynn, J.J. (Eds.), Vertebrate Paleontology in the
Neotropics. Smithsonian Institution Press, Washington, DC, pp. 435e458.
Kay, R.F., Ross, C.F., Williams, B.A., 1997. Anthropoid origins. Science 275,
797e804.
Kay, R.F., Ross, J., Simons, E.L., 2008. The basicranial anatomy of African
Eocene/Oligocene anthropoids. Are there any clues for platyrrhine origins?
In: Fleagle, J.G., Gilbert, C.C. (Eds.), Elwyn L. Simons: A Search for
Origins. Springer, New York, pp. 125e158.
Kay, R.F., Rossie, J.B., Colbert, M.W., Rowe, T., 2004b. Observations on the
olfactory system of Tremacebus harringtoni (Platyrrhini, early Miocene,
Sacanana, Argentina) based on high resolution X-ray CT scans. Am. J.
Phys. Anthropol. 35 (Suppl.), 123e124.
Kay, R.F., Simons, E.L., 1980. Comments on the adaptive strategy of the first
African Anthropoidea. Z. Morphol. Anthropol. 71, 143e148.
Kay, R.F., Simons, E.L., 1983. Dental formulae and dental eruption patterns in
Parapithecidae (Primates, Anthropoidea). Am. J. Phys. Anthropol. 62,
363e375.
Kay, R.F., Vizcaino, S.F., Tauber, A., Bargo, M.S., Williams, B.A., Luna, C.,
Colbert, M.W., 2005. Three newly discovered skulls of Homunculus pata-
gonicus support its position as a stem platyrrhine and establish its diurnal
arboreal folivorous habits. Am. J. Phys. Anthropol. 40 (Suppl.), 127.
Kay, R.F., Williams, B.A., 1994. Dental evidence for anthropoid origins. In:
Fleagle, J.G., Kay, R.F. (Eds.), Anthropoid Origins: The Fossil Evidence.
Plenum Press, New York, pp. 361e446.
Kay, R.F., Williams, B.A., Anaya, F., 2001. The adaptations of Branisella
boliviana, the earliest South American monkey. In: Plavcan, J.M., van
Schaik, C., Kay, R.F., Jungers, W.L. (Eds.), Reconstructing Behavior in
the Primate Fossil Record. Kluwer Academic/Plenum Publishers, New
York, pp. 339e370.
Kay, R.F., Williams, B.A., Ross, C.F., Takai, M., Shigehara, N., 2004c. An-
thropoid origins: a phylogenetic analysis. In: Ross, C.F., Kay, R.F.
(Eds.), Anthropoid Origins: New Visions. Kluwer Academic/Plenum
Publishers, New York, pp. 91e135.
Ketcham, R.A., Carlson, W.D., 2001. Acquisition, optimization, and interpre-
tation of x-ray computed tomographic imagery: applications to the geo-
sciences. Comp. Geosci. 27, 381e400.
Kraglievich, J.L., 1951. Contribuciones al conocimiento de los primates fosiles
de la Patagonia. I. Diagnosis previa de un nuevo primat fosil del Oligoceno
superior (Colheuhuapiano) de Gaiman, Chubut. Comm. Inst. Nac. Inv.
Cien. Nat. II, Cien. Zool. 11, 57e82.
Le Gros Clark, W.E., 1959. The Antecedents of Man. Edinburgh University
Press, Edinburgh.
Lieberman, D.E., Ross, C., Ravosa, M.J., 2000. The primate cranial base: on-
togeny, function, and integration. Yearb. Phys. Anthropol. 43, 117e169.
Lockwood, C.A., Fleagle, J.G., 1999. The recognition and evaluation of
homoplasy in primate and human evolution. Yearb. Phys. Anthropol. 42,
189e232.
Mabee, P., 1989. Assumptions underlying the use of ontogenetic sequences for
determining character state order. Trans. Am. Fish. Soc. 118, 151e158.
Mabee, P., 2000. The usefulness of ontogeny in interpreting morphological
characters. In: Wiens, J.J. (Ed.), Phylogenetic Analysis of Morphological
Data. Smithsonian Institution Press, Washington, D. C, pp. 84e114.
MacFadden, B.J., 1990. Chronology of Cenozoic primate localities in South
America. J. Hum. Evol. 19, 7e22.
MacFadden, B.J., Campbell, K.E., Cifelli, R.L., Siles, O., Johnson, N.M.,
Naeser, C.W., Zeitler, P.K., 1985. Magnetic polarity stratigraphy and mam-
malian fauna of the Deseadan (late Oligocene-early Miocene) Salla Beds
of northern Bolivia. J. Geol. 93, 223e250.
MacPhee, R.D.E., Cartmill, M., 1986. Basicranial structures and primate sys-
tematics. In: Swindler, D.R., Erwin, J. (Eds.), Comparative Primate Biol-
ogy, Vol. 1: Systematics, Evolution, and Anatomy. Alan R. Liss, New
York, pp. 219e275.
MacPhee, R.D.E., Horovitz, I., Arredondo, O., Jimenez Vazquez, O., 1995. A
new genus for the extinct Hispaniolian monkey Saimiri bernensis Rımoli,
1977, with notes on its systematic position. Am. Mus. Novit. 3134, 1e21.
Maddison, D.R., Maddison, W.P., 2001. MacClade 4: Analysis of Phylogeny
and Character Evolution. Version 4.03. Sinauer Associates, Sunderland.
Maier, W., 1980. Nasal structures in old and new world primates. In:
Ciochon, R., Chiarelli, B. (Eds.), Evolutionary Biology of the New World
Monkeys and Continental Drift. Plenum Press, New York, pp. 219e241.
Marshall, L.G., 1985. Geochronology and land-mammal biochronology of
the transamerican faunal interchange. In: Stehli, F.G., Webb, S.D.
(Eds.), The Great American Biotic Interchange. Plenum Press, New
York, pp. 49e85.
Martin, R.D., 1990. Primate Origins and Evolution: A Phylogenetic Recon-
struction. Princeton University Press, Princeton, New Jersey.
Meireles, C.M., Czelusniak, J., Ferreira, H.S., Schneider, M.P.C.,
Goodman, M., 1999a. Phylogenetic relationships among Brazilian howler
monkeys, genus Alouatta (Platyrrhini, Anthropoidea), based on g1-globin
pseudogene sequences. Genet. Mol. Biol. 22, 337e344.
Meireles, C.M., Czelusniak, J., Schneider, M.P.C., Muniz, J.A.P.C.,
Brigido, M.C., Ferreira, H.S., Goodman, M., 1999b. Molecular phylogeny
of ateline New World monkeys (Platyrrhini, Atelinae) based on gamma-
globin gene sequences: Evidence that Brachyteles is the sister group of
Lagothrix. Mol. Phylogenet. Evol. 12, 10e30.
Meldrum, D.J., 1993. Postcranial adaptations and positional behavior in fossil
platyrrhines. In: Gebo, D.L. (Ed.), Postcranial Adaptations in Nonhuman
Primates. Northern Illinois University Press, DeKalb, pp. 235e251.
Meldrum, D.J., Kay, R.F., 1997. Nuciruptor rubricae, a new pitheciin seed
predator from the Miocene of Colombia. Am. J. Phys. Anthropol. 102,
407e427.
Mickevich, M.F., 1982. Transformation series analysis. Syst. Zool. 31, 461e478.
Mickevich, M.F., Weller, S.J., 1990. Evolutionary character analysis: tracing
character change on a cladogram. Cladistics 6, 137e170.
Mouri, T., 1988. Intergeneric comparison of ceboid cranial measurements.
Kyoto University Overseas Research Reports of New World Monkeys
VI, pp. 23e31.
Nixon, K.C., Davis, J.I., 1991. Polymorphic taxa, missing values and cladistic
analyses. Cladistics 7, 233e241.
Pascual, R., Odreman Rivas, O., 1971. Evolucion de las comunidades de los
vertebrados del terciario Argentina, los aspectos paleozoogeograficos y
paleoclimaticos relacionados. Ameghiniana 7, 372e412.
Patterson, B., Pascual, R., 1972. The fossil mammal fauna of South America.
In: Keast, A., Erk, F.C., Glass, B. (Eds.), Evolution, Mammals, and South-
ern Continents. State University of New York Press, Albany, pp. 247e309.
Pocock, R.I., 1925. Additional notes on the external characters of some plat-
yrrhine monkeys. Proc. Zool. Soc. Lond. 1925, 27e47.
Poux, C., Chevret, P., Huchon, D., de Jong, W.W., Douzery, E., 2006. Arrival
and diversification of caviomorph rodents and platyrrhine primates in
South America. Syst. Zool. 55, 228e244.
Radinsky, L., 1977. Early primate brains: fact and fiction. J. Hum. Evol. 6,
79e86.
Ravosa, M.J., Hogue, A., 2003. Function and fusion of the mandibular
symphysis in mammals: a comparative and experimental perspective.

382 R.F. Kay et al. / Journal of Human Evolution 54 (2008) 323e382
In: Ross, C., Kay, R.F. (Eds.), Anthropoid Origins: New Visions. Kluwer
Academic/Plenum Publishers, New York, pp. 413e462.
Ray, D.A., Xing, J., Hedges, D.J., Hall, M.A., Laborde, M.E., Anders, B.A.,
White, B.R., Stoilova, N., Fowlkes, J.D., Landry, K.E., Chemnik, L.G.,
Ryder, O.A., Batzer, M.A., 2005. Alu insertion loci and platyrrhine pri-
mate phylogeny. Mol. Phylogenet. Evol. 35, 117e126.
Rosenberger, A.L., 1978. Loss of incisor enamel in marmosets. J. Mammal.
59, 207e208.
Rosenberger, A.L., 1979a. Cranial anatomy and implications of Dolichocebus,
a late Oligocene ceboid primate. Science 279, 416e418.
Rosenberger, A.L., 1979b. Phylogeny, evolution and classification of New
World monkeys (Platyrrhini, Primates). Ph.D. Dissertation, City University
of New York.
Rosenberger, A.L., 1980. Gradistic views and adaptive radiation of platyrrhine
primates. Z. Morphol. Anthropol. 71, 157e163.
Rosenberger, A.L., 1981. Systematics: the higher taxa. In: Coimbra-Filho, A.F.,
Mittermeier, R.A. (Eds.), Ecology and Behavior of Neotropical Primates,
vol. I. Academia Brasileira de Ciencias, Rio de Janeiro, pp. 9e27.
Rosenberger, A.L., 1982. Supposed squirrel monkey affinities of the late Oligo-
cene Dolichocebus gaimanensis, reply to P. Hershkovitz. Nature 298, 202.
Rosenberger, A.L., 1983. Tale of tails: parallelism and prehensility. Am. J.
Phys. Anthropol. 60, 103e107.
Rosenberger, A.L., 1988. Platyrrhini. In: Tattersall, I., Delson, E., Van
Couvering, J., Brooks, A. (Eds.), Encyclopedia of Human Evolution and
Prehistory. Garland Publishing, New York, pp. 456e459.
Rosenberger, A.L., 1992. Evolution of New World monkeys. In: Jones, S.,
Martin, R., Pilbeam, D.R. (Eds.), The Cambridge Encyclopedia of Human
Evolution. Cambridge University Press, Cambridge, pp. 209e216.
Rosenberger, A.L., 2000. Pitheciinae. In: Delson, E., Tattersal, I., Van
Couvering, J., Brooks, A. (Eds.), Encyclopedia of Human Evolution and
Prehistory, second ed. Garland Publishing Co, New York, pp. 562e563.
Rosenberger, A.L., 2002. Platyrrhine paleontology and systematics: the para-
digm shifts. In: Hartwig, W.C. (Ed.), The Primate Fossil Record.
Cambridge University Press, Cambridge, pp. 151e159.
Rosenberger, A.L., Setoguchi, T., Shigehara, N., 1990. The fossil record of
callitrichine primates. J. Hum. Evol. 19, 209e236.
Ross, C.F., 1993. The functions of the postorbital septum and anthropoid
origins. Ph.D. Dissertation, Duke University.
Ross, C.F., 1994. The craniofacial evidence for anthropoid and tarsier relation-
ships. In: Fleagle, J.G., Kay, R.F. (Eds.), Anthropoid Origins: The Fossil
Evidence. Plenum Press, New York, pp. 469e547.
Ross, C.F., 1995. Allometric and functional influences on primate orbit orien-
tation and the origins of the Anthropoidea. J. Hum. Evol. 29, 210e228.
Ross, C., Williams, B.A., Kay, R.F., 1998. Phylogenetic analysis of anthropoid
relationships. J. Hum. Evol. 35, 221e306.
Rossie, J.B., 2006. Ontogeny and homology of the paranasal sinuses in Platyr-
rhini (Mammalia: Primates). J. Morphol. 206, 1e40.
Roth, S., 1899. Aviso preliminary sobre mamiferos mesozoicos encontrados en
Patagonia. Rev. Mus. de La Plata 9, 381e388.
Saban, R., 1963. Contribution a l’etude de l’os temporal des Primates. De-
scription chez l’Homme et les Prosimiens. Anatomie comparee et phyloge-
nie. Mem. Mus. Natl. Hist. Nat. (France) N. Ser. A. Zool. 29, 1e378.
Sanchez-Villagra, M., Williams, B.A., 1998. Levels of homoplasy in the
evolution of the mammalian skeleton. J. Mammal. Evol. 5, 113e126.
Schneider, H., Canavez, F.C., Sampaio, I., Moreira, M.A.M., Tagliaro, C.H.,
Seuanez, H.N., 2001. Can molecular data place each Neotropical monkey
in its own branch? Chromosoma 109, 515e523.
Schneider, H., Sampaio, I., Harada, M.L., Barroso, C.M.L., Schneider, M.P.C.,
Czelusniak, J., Goodman, M., 1996. Molecular phylogeny of the New World
monkeys (Platyrrhini, Primates) based on two unlinked nuclear genes: IRBP
interon 1 and e-globin sequences. Am. J. Phys. Anthropol. 100, 153e179.
Seiffert, E.R., Simons, E.L., Simons, C.V.M., 2004. Phylogenetic, biogeo-
graphic, and adaptive implications of new fossil evidence bearing on crown
anthropoid origins and early stem catarrhine evolution. In: Ross, C.,
Kay, R.F. (Eds.), Anthropoid Origins: New Visions. Kluwer Academic/
Plenum Publishers, New York, pp. 157e182.
Simmons, N., 1993. The importance of methods: Archontan phylogeny and
cladistic analysis of morphological data. In: MacPhee, R.D.E. (Ed.), Pri-
mates and Their Relatives in Phylogenetic Perspective. Plenum Press,
New York, pp. 1e61.
Simons, E.L., 1993. New endocasts of Aegyptopithecus: oldest well-preserved
record of the brain in Anthropoidea. Am. J. Sci. 293-A, 383e390.
Simons, E.L., 2001. The cranium of Parapithecus grangeri, an Egyptian Oligo-
cene anthropoidean primate. Proc. Natl. Acad. Sci. U.S.A. 98, 7892e7897.
Simons, E.L., 2004. The cranium and adaptations of Parapithecus grangeri,
a stem anthropoid from the Fayum Oligocene of Egypt. In: Ross, C.,
Kay, R.F. (Eds.), Anthropoid Origins: New Visions. Kluwer Academic/
Plenum Publishers, New York, pp. 183e217.
Simons, E.L., Plavcan, J.M., Fleagle, J.G., 1999. Canine sexual dimorphism in
Egyptian Eocene anthropoid primates: Catopithecus and Proteopithecus.
Proc. Natl. Acad. Sci. U.S.A. 96, 2559e2562.
Simons, E.L., Rasmussen, D.T., 1996. Skull of Catopithecus browni, an early
Tertiary catarrhine. Am. J. Phys. Anthropol. 100, 261e292.
Simons, E.L., Seiffert, E.R., Ryan, T.M., Attia, Y., 2007. A remarkable female
cranium of the early Oligocene anthropoid Aegyptopithecus zeuxis (Catar-
rhini, Propliopithecidae). Proc. Natl. Acad. Sci. U.S.A. 104, 8731e8736.
Simpson, G.G., 1935. Early and middle Tertiary geology of the Gaiman re-
gion, Chubut, Argentina. Am. Mus. Novit. 775, 1e29.
Simpson, G.G., 1961. Principles of Animal Taxonomy. Columbia University
Press, New York.
Simpson, G.G., 1980. Splendid Isolation: The Curious History of South
American Mammals. Yale University Press, New Haven.
Singer, S.S., Schmitz, J., Schwiegk, C., Zischler, H., 2002. Molecular cladistic
markers in New World monkey phylogeny (Platyrrhini, Primates). Mol.
Phylogenet. Evol. 26, 490e501.
Slowinski, J.B., 1993. ‘‘Unordered’’ versus ‘‘ordered’’ characters. Syst. Biol.
42, 155e165.
Springer, M., Teeling, E., Madsen, O., Stanhope, M., deJong, W.W., 2001.
Integrated fossil and molecular data reconstruct bat echolocation. Proc.
Natl. Acad. Sci. U.S.A. 98, 6241e6246.
Swofford, D.L., 2002. PAUP. Phylogenetic Analysis Using Parsimony, Version
4.0b10 (Altivec). Sinauer Associates, Sunderland.
Szalay, F.S., Delson, E., 1979. Evolutionary History of the Primates. Academic
Press, New York.
Takai, M., Anaya, F., Shigehara, N., Setoguchi, T., 2000. New fossil materials
of the earliest New World monkey, Branisella boliviana, and the problem
of platyrrhine origins. Am. J. Phys. Anthropol. 111, 263e281.
Tarling, D.H., 1980. The geologic evolution of South America with special ref-
erence to the last 200 million years. In: Ciochon, R.L., Chiarelli, B. (Eds.),
Evolutionary Biology of the New World Monkeys and Continental Drift.
Plenum Press, New York, pp. 1e42.
Tauber, A., 1991. Homunculus patagonicus Ameghino, 1891 (Primates,
Ceboidea), Mioceno Temprano, de la costa Atlantica Austral, Prov. de Santa
Cruz, Republica Argentina. Acad. Nac. Cien. Cordoba, Argentina 82, 1e32.
Tauber, A.A., Kay, R.F., Luna, C., 2004. Killik Aike Norte, una localidad
clasica de la Formacion Santa Cruz (Mioceno temprano-medio),
Patagonia, Argentina. Ameghiniana Resumenes 2004, 26.
Tejedor, M.F., Tauber, A.A., Rosenberger, A.L., Swisher III, C.C.,
Palacios, M.E., 2006. New primate genus from the Miocene of Argentina.
Proc. Natl. Acad. Sci. U.S.A. 103, 5437e5441.
Wall, C.E., 1998. The expanded mandibular condyle of the Megaladapidae.
Am. J. Phys. Anthropol. 103, 263e276.
Williams, B.A., 2007. Comparing levels of homoplasy in the primate skeleton.
J. Hum. Evol. 52, 480e489.
Williams, B.A., Covert, H.H., 1994. New early Eocene anaptomorphine pri-
mate (Omomyidae) from the Washakie Basin, Wyoming, with comments
on the phylogeny and paleobiology of anaptomorphines. Am. J. Phys.
Anthropol. 93, 323e340.