Obtention and characterization of primary astrocyte and microglial cultures from adult monkey brains
16
Obtention and Characterization of Primary Astrocyte and Microglial Cultures From Adult Monkey Brains G. Guillemin, 1 F.D. Boussin, 1 * J. Croitoru, 1 M. Franck-Duchenne, 1 R. Le Grand, 1 F. Lazarini, 2 and D. Dormont 1 1 Service de Neurovirologie, CEA, DSV/DRM/SSA/IPSC, Fontenay-aux-Roses, France 2 Laboratoire de Neuropathologie Raymond Escourolle, Groupe Hospitalier Pitie ´-Salpe ˆtrie `re, Paris, France Simple methods for obtention of primary cultures of isolated astrocytes and microglia from adult simian brain have been developed. Characterization of these two glial cell populations were performed by morpho- logical observations and by immunocytochemistry. The astroglial cultures were obtained by an indirect method. After L-leucine methyl-ester treatment and trypsinizations, more than 99% of cells expressed glial fibrillary acidic protein (GFAP), whereas no macro- phages or microglia could be detected. Likely, the 1% remaining cells were immature astrocytes or cells that lost their GFAP expression. Cultured simian astro- cytes expressed vimentin, laminin, and fibronectin. We also found a constitutively low expression of major histocompatibility complex (MHC) class II by cul- tured astrocytes which was significantly enhanced by lipopolysaccharide (LPS), interferon gamma (IFN-g), or tumor necrosis factor alpha (TNF-a) treatments. Microglial cultures were obtained by a direct method of isolation using Percoll gradient separations and compared to simian monocyte-derived macrophages or alveolar macrophages. Microglial cells differed from macrophages by their proliferation upon granu- locyte-macrophage colony stimulating factor (GM- CSF) treatment and by their typical morphology when observed by scanning electron microscopy. As macro- phages, they expressed in vitro CD68, CD64, CD14, CD11b, MHC class II, and fibronectin. However, contrary to macrophages, simian cultured microglia expressed laminin. This observation suggests that microglia repre- sent a new potential source of this extracellular matrix protein in the brain. J. Neurosci. Res. 49:576–591, 1997. r 1997 Wiley-Liss, Inc. Key words: astrocytes; microglia; macaque; primary cultures INTRODUCTION Glial cells allow more than brain cohesion (Vir- chow, 1846). Especially, astrocytes play important and multiple roles in brain development and functions which still remain poorly understood. Astrocytes represent the most abundant cell type in the central nervous system (CNS). They are highly heterogeneous upon brain local- ization in their profiles of enzymes, antigenic markers, transporters, receptors, and ion channels (Wilkin et al., 1990). Their shape is not static and can be modified upon physiologic and pathologic stimuli. They have a syncy- tium-like organization that is mediated by gap junctions allowing intense intercell communications (Giaume and McCarthy, 1996). Situated around neurons and their processes, they are implicated in neuronal guidance, neurite outgrowth, synaptogenesis, and synaptic plastic- ity (Muller et al., 1995). They provide energy, nutrient, and growth factor support for neurons (Tsacopoulos and Magistretti, 1996); regulate extracellular concentrations of ions (Sykova et al., 1992) and neurotransmitters (Martin, 1995); and have an important function in brain detoxification. Astrocytes are also involved in the mainte- nance of the blood-brain barrier (BBB) because they provide signals for the formation and the cohesion of the tight junctions between endothelial cells (Janzer and Raff, 1987; Giaume and McCarthy, 1996). One of the promi- nent features of astrocytes is their vigorous response following various neurologic insults (Hertz et al., 1990; Mucke and Eddelston, 1993). Reactive astrocytosis is characterized by an increase in number and size of cells expressing glial fibrillary acidic protein (GFAP), a spe- cific intermediate filament cytoskeletal protein (Bignami et al., 1972). Activated astrocytes also express a large number of various molecules normally not detected in resting astrocytes, allowing them to participate in many Contract grant sponsor: Agence Nationale de Recherches sur le SIDA. Contract grant sponsor: Conseil Re ´gional de Bourgogne. *Correspondence to: F.D. Boussin, Service de Neurovirologie, CEA, DSV/DRM/SSA/IPSC, 60-68, Avenue de la Division Leclerc, BP 6 92 265 Fontenay-aux-Roses Cedex, France. E-mail: [email protected] Received 5 November 1996; Revised 17 April 1997; Accepted 17 April 1997 Journal of Neuroscience Research 49:576–591 (1997) r 1997 Wiley-Liss, Inc.
Transcript of Obtention and characterization of primary astrocyte and microglial cultures from adult monkey brains

Obtention and Characterization of PrimaryAstrocyte and Microglial Cultures From AdultMonkey BrainsG. Guillemin,1 F.D. Boussin,1* J. Croitoru, 1 M. Franck-Duchenne,1 R. Le Grand,1F. Lazarini,2 and D. Dormont11Service de Neurovirologie, CEA, DSV/DRM/SSA/IPSC, Fontenay-aux-Roses, France2Laboratoire de Neuropathologie Raymond Escourolle, Groupe Hospitalier Pitie´-Salpetriere, Paris, France
Simple methods for obtention of primary cultures ofisolated astrocytes and microglia from adult simianbrain have been developed. Characterization of thesetwo glial cell populations were performed by morpho-logical observations and by immunocytochemistry.The astroglial cultures were obtained by an indirectmethod. After L-leucine methyl-ester treatment andtrypsinizations, more than 99% of cells expressed glialfibrillary acidic protein (GFAP), whereas no macro-phages or microglia could be detected. Likely, the 1%remaining cells were immature astrocytes or cells thatlost their GFAP expression. Cultured simian astro-cytes expressed vimentin, laminin, and fibronectin.We also found a constitutively low expression of majorhistocompatibility complex (MHC) class II by cul-tured astrocytes which was significantly enhanced bylipopolysaccharide (LPS), interferon gamma (IFN-g),or tumor necrosis factor alpha (TNF-a) treatments.Microglial cultures were obtained by a direct methodof isolation using Percoll gradient separations andcompared to simian monocyte-derived macrophagesor alveolar macrophages. Microglial cells differedfrom macrophages by their proliferation upon granu-locyte-macrophage colony stimulating factor (GM-CSF) treatment and by their typical morphology whenobserved by scanning electron microscopy. As macro-phages, they expressed invitro CD68, CD64, CD14,CD11b, MHC class II, and fibronectin. However, contraryto macrophages, simian cultured microglia expressedlaminin. This observation suggests that microglia repre-sent a new potential source of this extracellular matrixprotein in the brain. J. Neurosci. Res. 49:576–591,1997. r 1997 Wiley-Liss, Inc.
Key words: astrocytes; microglia; macaque; primarycultures
INTRODUCTIONGlial cells allow more than brain cohesion (Vir-
chow, 1846). Especially, astrocytes play important and
multiple roles in brain development and functions whichstill remain poorly understood. Astrocytes represent themost abundant cell type in the central nervous system(CNS). They are highly heterogeneous upon brain local-ization in their profiles of enzymes, antigenic markers,transporters, receptors, and ion channels (Wilkin et al.,1990). Their shape is not static and can be modified uponphysiologic and pathologic stimuli. They have a syncy-tium-like organization that is mediated by gap junctionsallowing intense intercell communications (Giaume andMcCarthy, 1996). Situated around neurons and theirprocesses, they are implicated in neuronal guidance,neurite outgrowth, synaptogenesis, and synaptic plastic-ity (Muller et al., 1995). They provide energy, nutrient,and growth factor support for neurons (Tsacopoulos andMagistretti, 1996); regulate extracellular concentrationsof ions (Sykova et al., 1992) and neurotransmitters(Martin, 1995); and have an important function in braindetoxification. Astrocytes are also involved in the mainte-nance of the blood-brain barrier (BBB) because theyprovide signals for the formation and the cohesion of thetight junctions between endothelial cells (Janzer and Raff,1987; Giaume and McCarthy, 1996). One of the promi-nent features of astrocytes is their vigorous responsefollowing various neurologic insults (Hertz et al., 1990;Mucke and Eddelston, 1993). Reactive astrocytosis ischaracterized by an increase in number and size of cellsexpressing glial fibrillary acidic protein (GFAP), a spe-cific intermediate filament cytoskeletal protein (Bignamiet al., 1972). Activated astrocytes also express a largenumber of various molecules normally not detected inresting astrocytes, allowing them to participate in many
Contract grant sponsor: Agence Nationale de Recherches sur le SIDA.Contract grant sponsor: Conseil Re´gional de Bourgogne.
*Correspondence to: F.D. Boussin, Service de Neurovirologie, CEA,DSV/DRM/SSA/IPSC, 60-68, Avenue de la Division Leclerc, BP 6 92265 Fontenay-aux-Roses Cedex, France. E-mail: [email protected]
Received 5 November 1996; Revised 17 April 1997; Accepted 17 April1997
Journal of Neuroscience Research 49:576–591 (1997)
r 1997 Wiley-Liss, Inc.

important functions (Norton et al., 1992; Eddelston andMucke, 1993). Thus, together with microglial cells,activated astrocytes play a major role in the generation ofthe intracerebral immune response (Giulian and Baker,1985; Couraud, 1994). They can produce various cyto-kines, express major histocompatibility complex (MHC)class I and II Ags, and present Ag to T cells (Selmaj et al.,1990; Aloisi et al., 1992; Frei et al., 1994). In differentways, astrocytes are implicated in the neuropathologicalpathways of various neurologic diseases (Eng et al.,1992; Norenberg, 1994).
Microglia represent the principal immune cellsresident to the CNS and the major source of braincytokines (Giulian, 1987; Dickson et al., 1993; Davis etal., 1994). They express specific markers for the mono-nuclear phagocytes and are thought to originate fromstem cells common to all classes from mononuclearphagocytes invading the CNS during embryonic develop-ment (Giulian and Baker, 1986; Giulian et al., 1995). Inthe developing brain, microglia have an activated pheno-type, refered to as ameboid microglia or brain macro-phages, including phagocytic behavior, large cell bodywith copious cytoplasm, and upregulation of macrophagemarker expression. In the late fetal and early post-natalperiods maturing microglia differentiate into ramifiedforms, characteristic of the quiescent cells found in adultnervous system. The ramified microglia are predominantin cerebral parenchyma of normal adult CNS. They havea small cell body with tortuous and ramified processesand downregulated expression of macrophage markers:limited expression of MHC class II, reduced capacity topresent antigen, and limited secretory functions. Thisresting state, which differs from that of other tissuemacrophages, appears to be in part regulated by astro-cytes and to prevent deleterious effects of macrophageson brain functions (Giulian et al., 1995). Ameboidmicroglia could reappear during pathological processes.Indeed, these cells react to various injuries or chronicdiseases of the CNS by proliferation and production ofnumerous inflammatory molecules such as nitric oxide,eicosanoids, and cytokines, thus playing an active role inneuropathological pathways (Dickson et al., 1993; Daviset al., 1994; Gendelman et al., 1994; Mallat and Chamak,1994). Attempts to establish a panel of microglia specificmarkers have proved elusive with expression dependingupon age, species, and prior exposure to immunostimu-lant (Giulian and Baker, 1986; Peudenier et al., 1991).Actually, there is no unique histochemical marker thatdistinguishes significantly activated ameboid microgliafrom infiltrated macrophages (Peudenier et al., 1991;Flaris et al., 1993). However, in many aspects microgliacould be regarded as a distinct class of mononuclear
phagocyte (Giulian et al., 1995). Contrary to differenti-ated blood monocytes, microglia may proliferate inpost-natal and adult brain (Giulian and Baker, 1986;Graeber et al., 1988; Raivich et al.,1991; Ganter et al.,1992). Furthermore, primary microglia could adopt in vitroupon various stimuli, particularly surface morphology, whichdistinguishes them from bone marrow precursors, monocytes,or tissue macrophages (Giulian and Baker, 1986; Ganter et al.,1992; Giulian et al., 1995).
The phylogenetic proximity between monkey andhuman and the analogy of their hematopoietic andimmune systems make macaques a unique model forbiomedical research of various diseases (Goudsmit et al.,1980; Axthelm and Shiigi, 1990; Li et al., 1993; Reimannet al., 1994). Particularly, macaques have been shown torepresent relevant models for neuropathogenetic diseasessuch as human immunodeficiency virus (HIV) encepha-lopathy (Brinkmann et al., 1993; Vitkovic et al., 1995)and transmissible subacute spongiform encephalopathy(Lasmezas et al., 1996). Thus it would be of great interestto develop primary cultures of glial cells derived frommonkey brain in order to study neuropathogenic path-ways involved in those diseases. Furthermore, macaquesgive the unique opportunity to easily obtain glial cellsfrom adult primate brain. Indeed, particularly whenconsidering glial cells, numerous in vitro cellular charac-teristics depend on species, culture conditions, and aboveall the differentiation state of cells (fetal, neonatal, oradult). Glial cells derived from adult brain thus appearmore adapted to the study of neuropathological pathwaysaffecting mature brain. The aim of this study was to developand characterize primary pure monocultures of astrocytes andmicroglia isolated from adult macaque brains.
MATERIALS AND METHODSAnimals
Adult rhesus monkeys (Macaca mulatta) wereimported from China (Charles River) and tested negativefor the natural common simian pathogens. Animals weremaintained in single cages housed in P3 facilities inaccordance with EEC guidelines for primate experiments.Monkeys were always handled under ketamine anesthesia(Imalgene, Rhoˆne-Merieux, France). Animal autopsieswere performed in P3 facilities. Monkeys were eutha-nized with sodium pentobarbital (Sanofi, Libourne,France). The cerebral material was obtained during the 4hr after the sacrifice of the animal.
Dissection and DissociationThe meninges were carefully removed and whole
brain was extracted. Several superficial washings wererealized with phosphate buffered saline (PBS) added with
Cultures of Simian Astrocytes and Microglia 577

2% of penicillin, streptomycin, neomycin mix (PSN), and1% of fungisone [washing buffer (WB)] to limit contami-nations. The cerebral hemispheres were divided into twoparts. Two washings were performed with WB on brainportions. Superficial blood vessels were carefully ex-tracted using dissection pliers.
Cell CulturesIsolation and culture of astrocytes. Astrocytes
were prepared from brains of adult monkeys using aprotocol derived from previously described methods(Norton et al., 1988; Selmaj et al., 1990; Levison andMcCarthy, 1991). Previously, the grey matter was split byscrape with scapel and used for microglial cell isolation(see below). Residual portions of cerebral cortex, wherewhite matter was the majority, were washed thoroughlywith WB, minced finely, and incubated with 20 ml of puretrypsin (0.25% w/v; Gibco-BRL, Gaithersburg, MD) for30 min at 37°C after disaggregation through a 10 mlpipet. After 20 min of incubation, 40 µl of DNAse(Boehringer-Mannheim, Mannheim, Germany) at 10 U/µlwas added in the tissular crush for 10 min at 37°C. Thesuspension was centrifuged at 500g for 5 min and thecells located at the top of the resulting pellets wereresuspended in RPMI 1640 medium containing 10%heat-inactivated fetal calf serum (FCS), 2 mM glutamine(Boehringer-Mannheim), 1% PSN mix, 0.5% fungizone(Gibco, Grand Island, NY), and 0.5% glucose.
Cellular suspension was directly plated on 75 cm2
culture flasks (Falcon, Lincoln Park, NJ). Cultures werekept for 3 days at 37°C in a 5% CO2 containingwater-saturated atmosphere. Adherent cells were grownto confluence for 10–15 days. At this time, L-leucinemethyl-ester (LME) at 10 mM (Sigma, St. Louis, MO)was added in culture flasks for 1 hr at room temperature toremove microglial cells and macrophages (Thiele et al.,1983). Then cultures were rinsed twice with PBS at 37°C.After trypsinization (0.05% w/v), cells were cultured inRPMI 1640 medium supplemented with 10% FCS inuncoated flasks. Medium was changed twice a week aftervigorous shaking. Cultures were trypsinized at conflu-ence and cells were transferred in new culture flasks.
Isolation and culture of microglial cells. Microg-lia were prepared using a protocol derived from methodsdescribed previously by Sedgwick et al. (1991) andBrinkmann et al. (1993). Briefly, grey matter was washedwith WB and forced through a 100 µm mesh metal sievewith the plunger of a plastic syringe. The dissociatedmaterial was collected by centrifugation for 10 min at1,000g and washed with WB. After a brief centrifugation,the pellet was incubated with pure trypsin and 40 µl ofDNAse (10 U/µl) for 30 min at 37°C. The digestedhomogenate was pelleted and trypsin was neutralized byaddition of 1 ml of pure FCS. After a washing with PBS,
the cellular pellet was resuspended and mixed in 8 ml ofisotonic Percoll (Pharmacia, Uppsala, Sweden) pH 7.4 at23%. This suspension was delicately layered onto 2 ml ofisotonic Percoll at 70%. After centrifugation at 1,250g for15 min with slow acceleration and deceleration withoutbrake, the cellular ring located at the interface wascollected and washed twice with PBS. Cells were resus-pended in 3 ml of PBS and were layered onto a secondPercoll gradient density composed of four density levelsat 23% (2 ml), 40% (2 ml), 60% (2 ml), 70% (2 ml), and100% (1 ml). Simultaneously, the same Percoll gradientdensity was realized with colored density marker beads(Pharmacia). The gradient was centrifuged at 1,250g for15 min with slow acceleration and deceleration withoutbrake. The cellular ring was collected between 40 and60% at the level of the red density marker beads(d 5 1.062 g/ml), washed twice with RPMI 1640, andresuspended in 5 ml of RPMI 1640 supplemented with10% of heat-inactivated FCS, 2 mM glutamine, 1%glucose, 1% PSN mix, 0.5% fungizone, and 100 U/ml ofgranulocyte-macrophage colony stimulating factor (GM-CSF; Boehringer-Mannheim). Cells were seeded in Per-manox slide flasks or Lab-tek system (Nunc, Roskilde,Denmark) and cultured at 37°C with 5% of CO2.
Other cell types. Monkey blood monocyte-derived macrophages (MdM) were obtained using aclassic adherence method (Gessani et al., 1994). Simianalveolar macrophages were obtained after bronchoalveo-lar lavages (Cheret et al., 1996). Monkey primary fibro-blasts were obtained by tracheal scrape. HUVEC cells,NIH3T3 fibroblastic cell line, and SKNSH cell line camefrom ATCC (Rockville, MD).
AntiseraSo far, few antibodies directed specifically against
monkey determinant cells are available. Thus, numerousantibodies produced with heterologous antigens (human,cow, rat, chicken, mouse, or other) were tested inimmunocytochemical studies on our simian cells. Theywere all used at the concentrations recommended by eachmanufacturer. Monoclonal antibody (mAb) against CD68was provided by Dr. Radzun and has been used at theconcentration of 10 µg/ml (see Table I). All secondaryantibodies conjugated with Texas Red (TR) or fluo-roisothiocyanate (FITC) came from CliniSciences (Bag-neux, France). Three different controls have been performedfor each labeling experiment: 1) isotypic antibody controls, 2)incubation with only the secondary labeled antibodies, and 3)estimation of autofluorescence of unlabeled cells.
ImmunocytochemistryFollowing the cellular purification procedure,
4-week-old astroglial cells were transferred and grown in
578 Guillemin et al.

Permanox chamber slides (slide flasks or Lab-tek system;NUNC, Cergy Pontoise, France) for 2–3 days. Microgliaand MdM were directly cultured in Permanox chamberslides. For intracellular structural protein stainings, cellswere fixed with a cold methanol-acetone (1:1) solutionfor 20 min at220°C, then a membranous permeabiliza-tion was performed by incubation with 0.1% TritonX-100 in PBS for 10 min at room temperature. Formembranous antigen stainings, cells were fixed with aparaformaldehyde (PFA) 4% solution for 30 min at roomtemperature. Then cells were rinsed 3 times with PBS.After washings, cells were incubated with 5% normalgoat serum (NGS) in PBS for 30 min at room tempera-ture, rinsed twice with PBS, and incubated for 1 hr at37°C with primary antibody (Table I) diluted in 5% NGS.Cells were then washed extensively with 5% NGSsolution and incubated for 1 hr at 37°C in the dark withthe appropriately labeled secondary antibodies. In orderto quantify the percentage of labeled cells, nuclearstainings were performed using propidium iodure (IP) orDAPI at 1 µg/ml for 10 min at room temperature. Afterseveral washings with PBS at 37°C, the coverslips werequickly mounted on glass slides with Fluoromount-Gy(SBA, Birmingham, AL) and examined with a Nikonfluorescence microscope.
Scanning Electron Microscopy (SEM)Cells were cultured 2–3 days on Permanox flas-
kettes. Then cells were fixed with 4% PFA for 20 min atroom temperature and washed in 0.2 M cacodylate buffer
at pH 7.3. Following deshydratation in an alcohol series,the coverslip was stripped and the samples were dried bycritical point method (CO2) and then coated with gold/palladium in a sputter coater. The specimens wereexamined with SEM JEOL JSM-840A (Japan).
RESULTSPrimary Astrocyte Cultures
Astrocyte cultures could be maintained approxi-mately 4 months. Cells exhibited heterogeneous morpholo-gies which evolved during subsequent passages, as de-scribed previously for primary astrocytes from otherspecies (Hatten, 1985; Wilkin et al., 1990; Black et al.,1993; Holliday and Gruol, 1993; Perzel’ova and Mares,1993). After mechanical and enzymatic treatments ofcerebral matter followed by 3 days of adherence, about50–100 adhering cells/cm2 were usually found in cultureflasks. Thereafter, cultures were mostly composed of flatand polygonal astrocytes, mainly stellate with slenderprocesses, although some astrocytes can adopt a monopo-lar or bipolar morphology (Table II). Approximately 5%of spindle-shaped cells with broad flared processes,corresponding to probably residual microglia, were alsoobserved (see below). Flat and polygonal astrocytesexhibited a concentric repartition around proliferationcenters, and cultures became totally confluent betweenthe 15th and 20th days (Fig. 1A–C). Then, LME treat-ment and trypsinizations led to the complete disappear-ance of cells resembling microglia. After 4–6 passages
TABLE I. Primary Antibodies and Antisera Used for Immunochemistry
Antibodies and antisera Isotypes Clone Species of immunogens Manufacturers
GFAP Serum Polyclonal Human SigmaGFAP Serum Polyclonal Human EuromedexGFAPa Serum Polyclonal Cow DAKOGFAP IgG1 Clone GA5 Pig BoehringerGFAP IgG1 Clone DP46.10 Bovine ImmunotechVimentin IgG1 Clone V9 Pig SigmaFibronectin Serum Polyclonal Human SigmaLaminin Serum Polyclonal Mouse PasteurLaminin Serum Polyclonal Mouse SigmaLaminin IgG1 Clone 4C12-8 Mouse ImmunotechGal-Cera IgG1 Clone MGal-C Bovine TEBU/CederlaneGal-Cera Serum Polyclonal Rat EuromedexA2B5 IgM Clone 105 Chicken BoehringerNF160a IgG1 Clone NN18 Human BoehringerFactor VIIIa IgG1 Clone F8/86 Human DAKOCD68 IgG1 Clone Ki-M7 Human Dr. RadzunCD11ba IgG1 Clone IOM-1 Human ImmunotechCD14 IgG1 Clone 2.23 Human MedarexCD64 IgG1 Clone 22.2 Human MedarexMHC class II (HLA-DR)a IgG1 Clone B8.12.2 Human ImmunotechCD4 IgG1 Clone SK3 Human Becton Dickinson
aPreviously given by the manufacturer as to be able to recognize simian determinants.
Cultures of Simian Astrocytes and Microglia 579
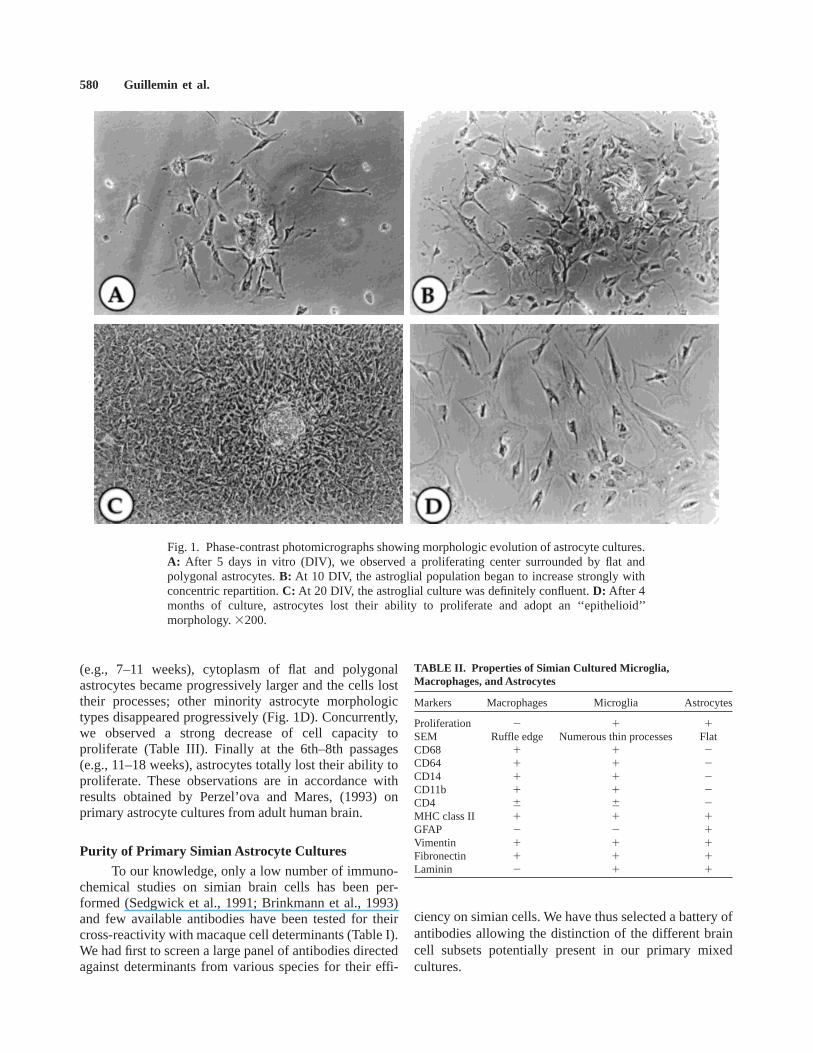
(e.g., 7–11 weeks), cytoplasm of flat and polygonalastrocytes became progressively larger and the cells losttheir processes; other minority astrocyte morphologictypes disappeared progressively (Fig. 1D). Concurrently,we observed a strong decrease of cell capacity toproliferate (Table III). Finally at the 6th–8th passages(e.g., 11–18 weeks), astrocytes totally lost their ability toproliferate. These observations are in accordance withresults obtained by Perzel’ova and Mares, (1993) onprimary astrocyte cultures from adult human brain.
Purity of Primary Simian Astrocyte CulturesTo our knowledge, only a low number of immuno-
chemical studies on simian brain cells has been per-formed (Sedgwick et al., 1991; Brinkmann et al., 1993)and few available antibodies have been tested for theircross-reactivity with macaque cell determinants (Table I).We had first to screen a large panel of antibodies directedagainst determinants from various species for their effi-
ciency on simian cells. We have thus selected a battery ofantibodies allowing the distinction of the different braincell subsets potentially present in our primary mixedcultures.
Fig. 1. Phase-contrast photomicrographs showing morphologic evolution of astrocyte cultures.A: After 5 days in vitro (DIV), we observed a proliferating center surrounded by flat andpolygonal astrocytes.B: At 10 DIV, the astroglial population began to increase strongly withconcentric repartition.C: At 20 DIV, the astroglial culture was definitely confluent.D: After 4months of culture, astrocytes lost their ability to proliferate and adopt an ‘‘epithelioid’’morphology.3200.
TABLE II. Properties of Simian Cultured Microglia,Macrophages, and Astrocytes
Markers Macrophages Microglia Astrocytes
Proliferation 2 1 1SEM Ruffle edge Numerous thin processes FlatCD68 1 1 2CD64 1 1 2CD14 1 1 2CD11b 1 1 2CD4 6 6 2MHC class II 1 1 1GFAP 2 2 1Vimentin 1 1 1Fibronectin 1 1 1Laminin 2 1 1
580 Guillemin et al.
The major specific marker of mature astrocytes isGFAP (Bignami et al., 1972). Detection of this glialintermediate filament is the most frequently used tocharacterize specifically astrocytes or astrogliomas (Gold-man et al., 1978). Thus, we have tested five differentheterologous antibodies or antisera directed against GFAPon monkey astrocytes (Table I). The three polyclonalantisera (Sigma, Saint-Quentin Fallavier, France; Eurome-dex, Soufflelweyersheim, France; and DAKO, Trappes,France) strongly reacted with simian GFAP, whereasmAb clone GA5 (Boehringer-Mannheim) was able torecognize only a few cells (approximately 5%). Con-versely, no staining was observed with mAb cloneDP.46.10 (Immunotech, Marseille, France). Using in aparallel manner Sigma and Euromedex polyclonal anti-sera, immunocytochemical stainings showed that cul-tured astrocytes strongly expressed the astrocyte-specificmarker GFAP with a typical fibrillary intracytoplasmiclocalization (Fig. 2A) concerning 90–95% of cells at day7 after seeding and more than 99% of cells after 1 monthof culture. This staining were observed in all astrocytemorphologic types described previously, and persisted allalong the cultures in the same proportion.
We investigated the presence of contaminating cellsusing polyclonal or mAbs directed against various spe-cific cellular determinants: galactocerebroside (GC) pro-tein (oligodendrocytes), NF160 (neurons), factor VIII(endothelial cells). No cell was stained by these antibod-ies even when assessed in initial cultures at days 7 and 15after seeding.
Finally, the main cellular contaminants for primaryastrocyte cultures are in macrophages and/or microglia.We have investigated their presence using mAbs directedagainst various specific markers like CD11b, CD14,CD64, and CD68. We have found approximately 5% ofcells stained by these mAbs in global initial cultures.These cells adhered directly on plastic flasks, but alsoclung on the top of astrocyte monolayers. Thereafter,LME treatment, vigorous agitations, and successive tryp-sinizations led to their disappearance. After 4–5 weeks,no cell was found positive for CD11b, CD14, CD64, andCD68 in our astrocyte cultures, indicating that macro-phages or microglia, if any, were present below levels ofdetection.
Characterization of Simian Astrocytes inPrimary Cultures
Vimentin is an intermediate filament which is alsooften used to study astrocyte proliferation (Schnitzer etal., 1981; Perzel’ova and Mares, 1993). Vimentin hasmany structural similarities to GFAP and both coexist inactivated astroglial cells in mature brain (Yamada et al.,1992). Using a mAb against pig vimentin, we found thatafter 1 month and till the end of cultures all astrocytesstrongly expressed this intermediate filament. As forGFAP, the staining of vimentin revealed a web ofintracytoplasmic fibrils without any predominant cellularlocalization (Fig. 2B).
As previously described by Liesi et al. (1986),astrocytes may synthesize fibronectin. Using polyclonalantisera against human fibronectin, we found no intracel-lular staining but a web of extracellular deposits offibronectin around astrocytes, which persisted all alongthe cultures (Fig. 2C). This staining differed markedlyfrom the strong intracytoplasmic and membranous label-ing that we observed on both cultured simian primaryfibroblasts (Fig. 2D) and the NIH3T3 murine fibroblasticcell line (data not shown).
We have also observed a characteristic perinucleusintracytoplasmic labeling of astrocytes using either poly-clonal antisera or one mAb directed against murinelaminin (Fig. 2E). As previously reported, astrocytelaminin expression was constant all along the cultures(Liesi et al., 1984, 1986).
Using a mAb against HLA-DR, we have found aconstitutive membranous low MHC class II expressionby primary astrocytes after 1 month of culture. However,this expression is significantly increased by treatments ofastrocyte cultures with interferon gamma (IFN-g; 100U/ml; Fig. 2F), lipopolysaccharide (LPS; 1 mg/ml), ortumor necrosis factor alpha (TNF-a; 100 U/ml) for 24 hr(data not shown).
Microglial Cell CulturesIn culture, adherent microglia are unable to prolifer-
ate without additional activating treatment. Therefore, aspreviously described (Giulian, 1987; Raivich et al., 1991;Lee et al., 1994), we have stimulated in vitro microglia
TABLE III. Time for Obtention of Cellular Confluence and Cellular Doubling Time in Primary Astrocyte Cultures*
Passages
1 2 3 4 5 6 7 8
Time for cellular confluence (days) 19.56 1 8 6 2 8 116 2 126 2 16.56 4.5 186 1 316 2Cumulate time of culture (days) 19.5 27.5 35.5 46.5 58.5 75 93 124Cellular doubling time (hr) 24–36 20–24 20–24 24–36 24–36 24–48 36–48 72–96
*Values are given from three typical primary simian astrocyte cultures. Cellular doubling times have been determined by cellular numeration onMallassez slides after trypsinization.
Cultures of Simian Astrocytes and Microglia 581
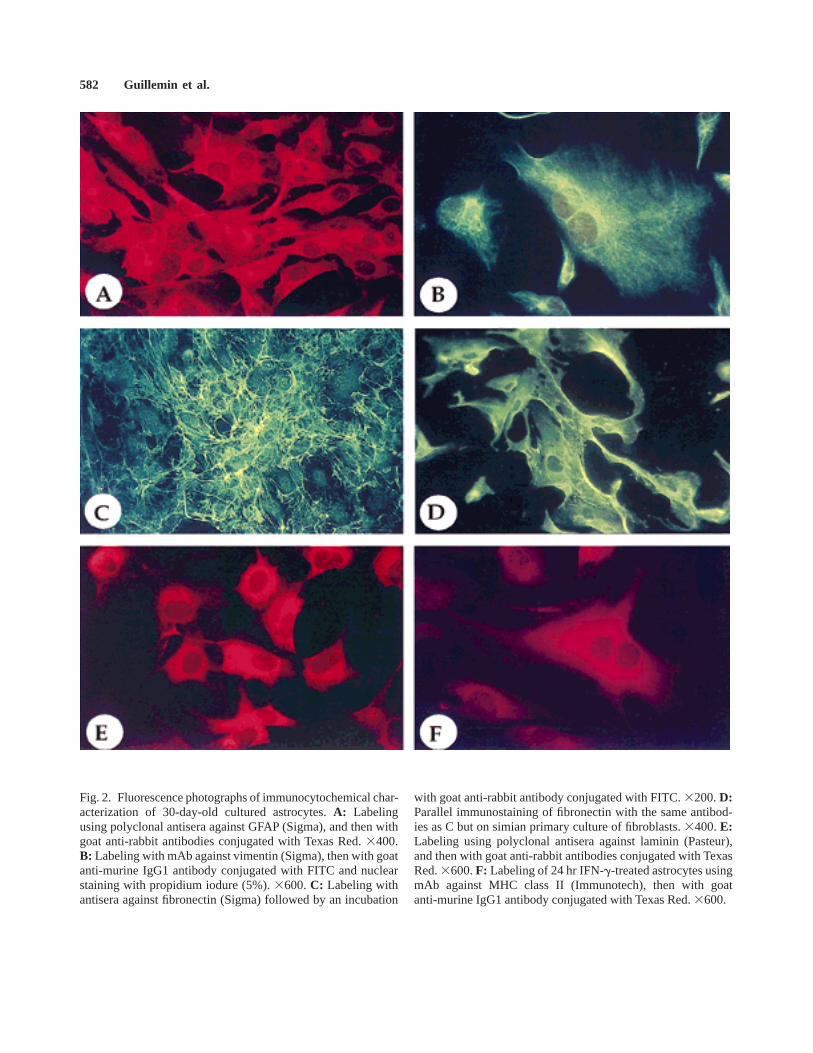
Fig. 2. Fluorescence photographs of immunocytochemical char-acterization of 30-day-old cultured astrocytes.A: Labelingusing polyclonal antisera against GFAP (Sigma), and then withgoat anti-rabbit antibodies conjugated with Texas Red.3400.B: Labeling with mAb against vimentin (Sigma), then with goatanti-murine IgG1 antibody conjugated with FITC and nuclearstaining with propidium iodure (5%).3600.C: Labeling withantisera against fibronectin (Sigma) followed by an incubation
with goat anti-rabbit antibody conjugated with FITC.3200.D:Parallel immunostaining of fibronectin with the same antibod-ies as C but on simian primary culture of fibroblasts.3400.E:Labeling using polyclonal antisera against laminin (Pasteur),and then with goat anti-rabbit antibodies conjugated with TexasRed.3600.F: Labeling of 24 hr IFN-g-treated astrocytes usingmAb against MHC class II (Immunotech), then with goatanti-murine IgG1 antibody conjugated with Texas Red.3600.
582 Guillemin et al.

proliferation by 100 U/ml of h-GM-CSF. After 3 days,primary microglial cell cultures were mainly composedof ameboid and spindle-shaped cells exhibiting longprocesses with highly granulous cytoplasm (Dickson et
al., 1991; Brinkmann et al., 1993) (Fig. 3A). Suchprocesses were never observed in primary cultures ofsimian monocyte-derived and alveolar macrophages,which appeared less granulous and frequently adopt afusiform aspect. After 1 week, proliferation of ameboidmicroglia was evidenced by the presence of numerousmicroglial nodules (Figs. 3B, 4A), when spindle-shapedmicroglia remained isolated. On the contrary, we ob-served no or very low proliferation of monocyte-derivedor alveolar macrophages cultured in the same conditions.Microglial proliferation continued to occur for 2 weeks,and ameboid microglia became the majority. Between the3rd and 4th weeks of culture, some giant multinucleatedgranulous cells appeared (Fig. 3C), then most of theameboid microglia progressively began to die, and onlycells with processes persisted in culture for another 2weeks. It should be pointed out that despite their prolifera-tion, microglia never reached confluence.
Two weeks after seeding, SEM examination re-vealed the characteristic ‘‘sombrero’’-like morphologywith numerous short and thin processes of ameboidmicroglia (Fig. 4B) and long filopods of spindle-shapedcells (Fig. 4C). These specific morphologies markedlydiffer from those of macrophages which exhibit rufflededges but no thin processes when observed on SEM (datanot shown) (Giulian and Baker, 1986; Frei et al., 1987;Rieske et al., 1989; Giulian et al., 1995). In puremicroglia cultures, we never observed highly ramifiedcells resembling resting microglia.
Purity and Immunophenotyping of MicroglialCultures
Astrocytes constitute a problematic cellular contami-nant of primary microglial cultures. Indeed, the presenceof a small number of astrocytes after seeding is sufficientto lead to a rapid invasion of the whole surface of theculture flasks, which is certainly favored by the presenceof GM-CSF in the culture medium (Guillemin et al.,1996). However, such contamination occurred only rarely,as astrocytes are normally not recovered in the Percollgradient fraction containing microglia. As a result, iso-lated microglia are rapidly drowned under a flood ofastrocytes. We did not detect other types of cellularcontaminants by immunocytochemistry in our microglialcultures. Indeed, no cells were found positive for GC,factor VIII, or NF160 at any time.
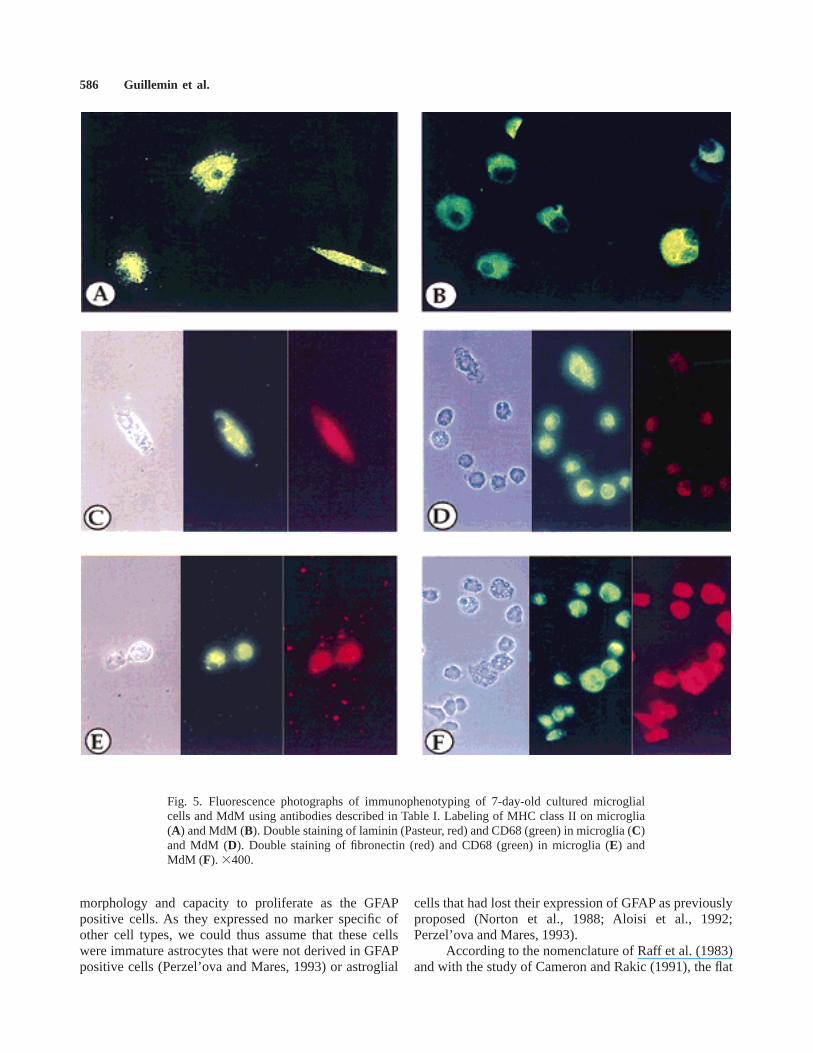
By immunocytochemical staining experiments, mi-croglia showed a significant membranous expression ofCD11b, CD14, CD64, CD68, and intracytoplasmic syn-thesis of vimentin and fibronectin which did not allow usto distinguish these cells from blood MdM or alveolarmacrophages. Furthermore, like cultured macrophages(Fig. 5B), nearly 100% of microglial cells stronglyexpressed membranous MHC class II (Fig. 5A). We alsofound a weak expression of CD4 by both cultured simian
Fig. 3. Phase-contrast photomicrographs showing morphologicevolution of GM-CSF-treated microglial cultures.A: After 3DIV, we observed apparition of a small proliferating center.B:At 7 DIV, the microglial population began to increase but neverreached confluence.C: At 30 DIV, some giant multinucleatedcells appeared in culture.3200.
Cultures of Simian Astrocytes and Microglia 583

Figure 4.

macrophages and microglial cells (data not shown).Furthermore, we have detected a strong intracellularlabeling of cultured microglia with either two differentantisera or one mAb directed against laminin (Fig. 5C),whereas MdM were not significantly stained (Fig. 5D).Regarding labeling of fibronectin, both microglial cellsand macrophages express constitutively this ECM protein(Fig. 5E,F).
DISCUSSIONWe have described here the isolation and character-
ization of both astrocyte and microglia pure primarycultures obtained from simian mature brains.
Our methodology for purification of astrocytes wasan indirect method starting from a mixed brain cellprimary culture (Norton et al., 1988; Shahar et al., 1989).The main issue was thus the eradication of variouspotential cellular contaminants. As previously described(Levison and McCarthy, 1991), some of them were easilyeliminated by an initial careful dissection of brain tissues.Particularly, fibroblasts known to proliferate more rapidlythan astrocytes have been avoided by removal of menin-ges. Elimination of superficial brain blood vessels al-lowed us to discard a majority of endothelial cells.Vigorous shakings of flasks during the first week afterseedings permitted a rapid and easy eradication ofoligodendrocytes and neurons. Moreover, those cells areknown to require particular culture conditions to persistin culture (Levison and McCarthy, 1991). Indeed, oligo-dendrocytes need a cellular matrix, such as a confluentmonolayer of astrocytes, or an artificial matrix, such aspoly-L-lysine, to grow in culture (McCarthy and DeVellis, 1980; Kim et al., 1983). Furthermore, in thepresence of FCS, neurons rarely survive more than 2 or 3days after seeding (Levison and McCarthy, 1991).
Microglia appear as the major contaminating celltype in primary astrocyte cultures. According to theresults of Giulian and Baker (1986), contamination bymicroglia may represent up to 5% of total cells withouttreatments, and we showed that microglia could persistmore than 4 months in cocultures (data not shown). Initialvigorous shakings during the first week after seedingeliminated most of the microglia cling on astrocytes.Then, LME treatment allowed the elimination of most ofthe remaining microglia. However, we have observed that
LME treatment is not able to poison all microglial cells,as previously reported (Giulian and Baker, 1986). Indeed,contrary to MdM or alveolar macrophages, some microg-lia survived this treatment although they contain largevacuoles in their cytoplasm (Nakajima et al., 1989). Thisproblem could not be overcome by prolonged LMEtreatments because long-term exposure to LME is toxicfor astrocytes and induces a strong increase of theiradhesion capacity rendering them totally resistant totrypsinization (data not shown). However, due to the verylow capacity of microglia to readhere after trypsinization,almost complete eradication was achieved by successivetrypsinizations and systematic changes of culture flasks.
By the combination of these successive methods,we have obtained, 1 month after seeding, cultures inwhich no cell expressing microglia/macrophage-specificmarkers could be detected. Cultures were then composedof 99% of astrocytes expressing GFAP. The percentage ofGFAP positive cells then remained constant till the end ofcultures. However, like the morphology (Fischer, 1984),astroglial GFAP expression in primary cultures has beenshown to depend on several factors such as tissue speciesand age and culture conditions (Goldman and Chiu,1984). Indeed, astrocytes expressed GFAP at high percent-age in primary cultures prepared from newborn rodent(Liesi et al., 1986) and fetal human brains (Aloisi et al.,1992). However, GFAP positive cells have been reportedto decrease and disappear rapidly by repeated passagingof cultures (Aloisi et al., 1992). In cultures from maturebrains, elevated percentages of astrocytes appear toexpress GFAP in cultures prepared from rodent or bovineastrocytes only after various delays of cultures (Norton etal., 1988). GFAP positive cells have been reported to berare or even absent in primary cultures from adult humanbrain (Osborn et al., 1981; Rutka, 1986) or to appeartransiently when adult brain cultures spontaneously decel-erated in growth (Rutka, 1986). Conversely, Koka et al.(1995) observed expression of GFAP by proliferatingastrocytes in mixed primary mature brain cultures. In ourprimary monkey astrocyte cultures, we found that GFAPpositive cells also expressed continuously vimentin con-sistently with previous studies showing that primaryastrocyte vimentin expression is detected whatever spe-cies and age of tissue (Bignami et al., 1972; Schnitzer etal., 1981). Vimentin expression in vivo is found inimmature, reactive, and neoplastic astrocytes (Mares etal., 1988; Yamada et al., 1992). In vitro, it has beenproposed that GFAP1 vimentin 1 astrocytes originatefrom GFAP2 vimentin 1 cells both in fetal and adultbrain tissue cultures (Aloisi et al., 1992; Perzel’ova andMares, 1993). In our simian primary cultures, the 1% ofcells lacking GFAP expression occasionally found in ourculture were vimentin positive and had roughly the same
Fig. 4. Scanning electron photomicrographs showing morphol-ogy of isolated microglia from adult macaques in primarycultures.A: After 7 DIV, we observed a typical proliferatingmicroglial nodule.3330. B,C: At 14 DIV, two phenotypesappeared in the majority: sombrero-like morphology (B;31,800)and long filopods of spindle-shaped cells (C;32,300).
Cultures of Simian Astrocytes and Microglia 585
morphology and capacity to proliferate as the GFAPpositive cells. As they expressed no marker specific ofother cell types, we could thus assume that these cellswere immature astrocytes that were not derived in GFAPpositive cells (Perzel’ova and Mares, 1993) or astroglial
cells that had lost their expression of GFAP as previouslyproposed (Norton et al., 1988; Aloisi et al., 1992;Perzel’ova and Mares, 1993).
According to the nomenclature of Raff et al. (1983)and with the study of Cameron and Rakic (1991), the flat
Fig. 5. Fluorescence photographs of immunophenotyping of 7-day-old cultured microglialcells and MdM using antibodies described in Table I. Labeling of MHC class II on microglia(A) and MdM (B). Double staining of laminin (Pasteur, red) and CD68 (green) in microglia (C)and MdM (D). Double staining of fibronectin (red) and CD68 (green) in microglia (E) andMdM (F). 3400.
586 Guillemin et al.

and polygonal morphologies of cultured simian astro-cytes and their lack of expression of A2B5 (Miller andRaff, 1984; Black et al., 1993), which is reported to bespecific of type 2 astrocytes, suggest that these cellsbelong to the type 1 astrocytes. This is further sustainedby our detection of important extracellular deposits offibronectin. Indeed, among astrocytes only the type 1subset is known to secrete this extracellular matrixprotein (Liesi et al., 1984). This observation is consistentwith previous other reports demonstrating that the pre-dominant source of astrocytes in rodent primary culturesable to proliferate in vitro are type 1 astrocytes (Miller etal., 1985; Hertz et al., 1990; Wilkin et al., 1990; Black etal., 1993; Oh et al., 1993), but differed from observationsin bovine primary cultures in which astrocytes predomi-nantly expressed A2B5 (Norton et al., 1988).
Interestingly, one mAb directed against pig GFAP(clone GA5, Boehringer-Mannheim) reacted stronglyonly with approximately 5% of GFAP positive simianastrocytes, whereas other cells were not stained. Recently,extracellular acidosis has been shown to produce a rapidincrease in GFAP immunoreactivity in astrocytes indepen-dent of de novo protein synthesis (Oh et al., 1995). Ourobservation may thus suggest conformational differencesof GFAP in a particular cultured simian type 1 astrocytesubset (Reeves et al., 1989; Levison and McCarthy, 1991)that should be further investigated.
Reactive astrocytes are involved in multifactorialand multistep responses to CNS injuries and during braindevelopment (for review see Eng et al., 1992; Eddelstonand Mucke, 1993; Mucke and Eddelston, 1993), amongthem, the astroglial secretion of ECM proteins likefibronectin and laminin which participated in neuriteoutgrowth (Liesi et al., 1986; Masuda-Nakagawa et al.,1993; Fawcett, 1994; Muller et al., 1995), and in theinduction of reversible modifications of microglial cellphenotypes (Chamak and Mallat, 1991; Giulian et al.,1995). We found a constant and intense expression oflaminin and fibronectin by cultured simian astrocytes, aspreviously reported for primary astroglial cultures fromother species (Liesi et al., 1984, 1986; Ard and Bunge,1988; Masuda-Nakagawa et al., 1993). Production oflarge amounts of fibronectin and laminin in those culturesmay be in part explained by the fact that culturedproliferating astrocytes from adult brains derived fromastrocyte precursors as generally admitted (Perzel’ovaand Mares, 1993). Fibronectin was found in majority inextracellular deposits, forming a kind of web around andon top of astrocytes, as shown by Liesi et al. (1984) usingthe newborn rodent model. However, unlike these au-thors, we have detected only a weak staining of fibronec-tin in the cytoplasm of astroglial cells. This result couldbe explained by the fact that expression and secretion offibronectin is a rapid metabolic phenomenon (Hatten et
al., 1982). Laminin has been detected in our culturedsimian astrocytes and has a restricted perinucleus intracy-toplasmic localization as observed on early rat astrocytecultures (Liesi et al., 1983). As previously shown, incultures of astrocytes derived from adult rodent brain,expression of laminin was significantly increased inlong-term astroglial cultures compared with short-termcultures (Bignami and Dahl, 1974; Yazaki et al., 1995).An additional factor which could increase the synthesis oflaminin is the medium culture composition. Indeed, FCSstimulates astroglial laminin expression (Ard and Bunge,1988).
Numerous studies have shown that astrocytes areable to express MHC class I and class II antigens aftervarious stimulations (for review see Eddelston and Mucke,1993). Following the addition of IFN-g, TNF-a, or LPSin our simian astroglial cultures, we have obtained asignificant increase of the membranous labeling intensityfor MHC class II determinants compared with untreatedcultures. In addition, according to previous studies (Al-oisi et al., 1992; Frei et al., 1994), we have observed lowconstitutive expression of MHC class II by astrocytes inprimary culture suggesting their weak activation (Eddels-ton and Mucke, 1993).
Contrary to astrocytes, microglial cell purificationand cultures were performed by a direct method involv-ing Percoll gradient centrifugations allowing the elimina-tion of most of the potential cellular contaminants aspreviously described (Brinkmann et al., 1993). The mainissue was thus the exclusion of potential contaminationsby blood-derived monocytes. Several elementary precau-tions have been taken to limit them significantly: at first,monkeys have been bled before removal of the brain;then, the encephalon was extensively washed and superfi-cial blood vessels were precausiously extracted withpliers. However, according to No¨renberg (1994) andGiulian and Baker (1986), no preparation of brain cellscan be absolutely free of blood cells. We thus comparedprimary microglial cultures with monkey MdM andalveolar macrophage cultures.
Two points of the methodology appeared critical forthe in vitro survival and proliferation of primary microg-lia: 1) the initial cellular density should be elevatedsuggesting the occurrence of autostimulation induced bysecretions, and 2) the early addition of GM-CSF isessential for the growth of microglia and strongly in-creased their proliferation rate (Frei et al., 1987; Ganter etal., 1992; Lee et al., 1994). In those conditions, weobserved a significantly higher proliferation rate in pri-mary microglia cultures than in cultures from adherentblood MdM and alveolar macrophages in which weobserved no or occasionally very low proliferation. Thisobservation is in agreement with numerous previousstudies performed on microglia from various species
Cultures of Simian Astrocytes and Microglia 587

demonstrating that the ability of microglial cells toproliferate appears as one major factor of discriminationbetween microglia and macrophages (Giulian and Baker,1986; Frei et al., 1987; Nakajima et al., 1989; Rieske etal., 1989; Ganter et al., 1992; No¨renberg, 1994). How-ever, it should be noted that microglial proliferation ratesalways stay very low compared to those of primaryastrocytes, and that cells never reached confluence.
Morphological examinations allow us to differenti-ate microglia from macrophages and confirm the purity ofmicroglial cultures (Giulian and Baker, 1986; Suzumuraet al., 1991; Giulian et al., 1995), although the phase-contrast light microscopic appearance of freshly isolatedameboid microglia and macrophages could be relativelysimilar. Then, phenotypic differences between these twocellular populations should be evidenced using SEM.Indeed, isolated simian microglia exhibited numerousmembranous thin spinous processes that easily distin-guished them from macrophages, as previously describedfor primary microglia from other species (Giulian andBaker, 1986; Frei et al., 1987; Rieske et al., 1989; Giulianet al., 1995).
According to previous numerous studies performedon various species (Giulian and Baker, 1986; Peudenier etal., 1991; Sedgwick et al., 1991; Brinkmann et al., 1993),we found similar membranous expression levels of CD68(KiM7), CD11b, CD64, and intracytoplasmic synthesis ofvimentin and fibronectin (Graeber et al., 1988; Rieske etal., 1989; Yamada et al., 1992) which did not permit us todistinguish microglial cells from macrophages. In addi-tion, we observed that both simian cell types expressedlow levels of CD4 antigens as reported earlier (Peudenieret al., 1991; Brinkmann et al., 1993). We also found nosignificant differences of CD14 membranous expression;as primary macrophages, nearly 100% of cultured simianmicroglia expressed this antigen at similar levels. How-ever, we observed that, in vitro, all microglia cellsexpressed strongly membranous MHC class II molecules(Hayes et al., 1987; Dickson et al., 1991). This differedmarkedly from observations of Brinkmann et al. (1993)performed on primary cultures from adult rhesus monkeybrains. These authors detected MHC class II expressionon very few microglial cells and a weak staining forCD14 on only 25% of these cells, whereas they found100% of cells expressing these antigens in cultures ofsimian MdM, although expression of these two antigenswas stimulated by SIV infection. Two of the possibleexplanations for these discrepancies may be related to theuse of distinct antibodies for the immunostaining experi-ments and also to some differences in the culture medium.Indeed, Brinkmann et al. (1993) did not confirm anyproliferation of primary microglia despite the fact thattheir cultures were performed like ours in the presence ofGM-CSF, which is known to stimulate microglial prolif-
eration (Frei et al., 1987; Ganter et al., 1992; Lee et al.,1994). We may thus assume that differences between ourresults may originate from their use of 10% of humanAB1 serum in the culture medium instead of 10% ofFCS, which could lead to a better activation of simianmicroglia. In the same way, Peudenier et al. (1991), usingprimary cultures from adult human brains, did not findCD14 labeling either on microglia or on macrophages,and obtained the same results as Brinkmann et al. (1993)on MHC class II expression. However, here again, theculture medium was different since it did not containGM-CSF. These data point out the importance of cultureconditions for the study of neuroglial cells.
One important finding of our study is the observa-tion that 1-week-old cultured microglia contained bothintracellular fibronectin and laminin. Although our resultshave to be again completed with studies on mRNAexpression, they strongly suggest that microglia are apotential source of these two ECM proteins in the CNS(Dickson et al., 1991; Masuda-Nakagawa et al., 1993).These proteins are located on basement membrane and inextracellular spaces, where microglial cells evolve duringdevelopment or after lesions in the CNS (Esiri andMorris, 1991). Microglia as macrophages express lamininand fibronectin receptors (Wright and Meyer, 1985;Huard et al., 1986). Laminin and fibronectin modulatevarious properties of mononuclear phagocytic activityand tumor cytotoxicity (Chamak and Mallat, 1991). Theirsecretion in the CNS has been currently attributed toastroglial cells (Liesi et al., 1984; Price and Hynes, 1985),whereas their expressions by microglia have been poorlyinvestigated. Consistent with our results, unlike bloodmonocytes, MdM and alveolar macrophages have beenshown to express fibronectin in vitro without treatment(Yamauchi et al., 1987). Moreover, Rieske et al. (1989)have described the synthesis of both laminin and fibronec-tin by cultured ramified microglia from rat facial nucleusexplants but they did not detect any expression of theseproteins in the same conditions in cultured macrophages.Astonishingly, these authors did not comment on theirresults, which could theoretically permit an immunocyto-chemical distinction between microglia and macro-phages. Furthermore, in the leech, microglial cells havebeen reported to synthesize laminin both in vivo and invitro (Masuda-Nakagawa et al., 1993). This differentialexpression of laminin could appear as important data todiscriminate microglial cells and macrophages in primaryculture. Moreover, the functional consequences of ECMprotein production by activated microglia may thus playimportant roles during brain development and followingCNS injury (Chamak and Mallat, 1991; Masuda-Nakagawa et al., 1993; Von Bernhardi and Muller, 1995).
Monkeys have been shown to represent relevantmodels of various CNS injuries, including AIDS encepha-
588 Guillemin et al.

lopathy (Brinkmann et al., 1993; Vitkovic et al., 1995)and transmissible subacute spongiform encephalopathy(Lasmezas et al., 1996). Data presented here providing abetter knowledge of both simian primary astrocyte andmicroglial cultures may be useful for understanding theneuropathogenetic mechanisms involved in these dis-eases.
ACKNOWLEDGMENTS
This work was supported by the Agence Nationalede Recherches sur le SIDA (A.N.R.S.) and the ConseilRegional de Bourgogne. We greatly appreciate the techni-cal assistance of H. Coffigny. We thank C. Chapel, A.Cheret, G. Gras, A. Jaegly, J.M. Perrin, B. Vaslin, and Ms.Guillaumin (SEM, Jussieu). Thanks also to Dr. M. Tardyand Dr. M. Dubois-Dalcq for their councils.
REFERENCES
Aloisi F, Borsellino G, Samoggia P, Testa U, Chelucci C, Russo G,Peschle C, Levi G (1992): Astrocyte cultures from humanembryonic brain: Characterization and modulation of surfacemolecules by inflammatory cytokines. J Neurosci Res 32:494–506.
Ard MD, Bunge RP (1988): Heparan sulfate proteoglycan and lamininimmunoreactivity on cultured astrocytes: Relationship to differ-entiation and neurite growth. J Neurosci 8:2844–2858.
Axthelm MK, Shiigi SM (1990): Nonhuman primate models foracquired immunodeficiency syndrome. J Med Primatol 19:163–165.
Bignami A, Dahl D (1974): Astrocyte-specific protein and neuroglialdifferentiation. An immunofluorescence study with antibodiesto the glial fibrillary acidic protein. J Comp Neurol 153:27–38.
Bignami A, Eng LF, Dahl D, Uyeda CT (1972): Localization of glialfibrillary acidic protein in astrocytes by immunofluorescence.Brain Res 43:429–435.
Black JA, Sontheimer H, Waxman SG (1993): Spinal cord astrocytes invitro: Phenotypic diversity and sodium channel immunoreactiv-ity. Glia 7:272–285.
Brinkmann R, Schwinn A, Muller J, Stahl-Hennig C, Coulibaly C,Hunsmann G, Czub S, Rethwilm A, Dorries R, Ter Meulen V(1993): In vitro and in vivo infection of rhesus monkeymicroglial cells by simian immunodeficiency virus. Virology195:561–568.
Cameron RS, Rakic P (1991): Glial cell lineage in the cerebral cortex:A review and synthesis. Glia 4:124–137.
Chamak B, Mallat M (1991): Fibronectin and laminin regulate the invitro differentiation of microglial cells. Neuroscience 45:513–527.
Cheret A, Le Grand R, Caufour P, Dereuddre-Bosquet N, Matheux F,Neildez O, Vaslin B, Dormont D (1996): Cytokine mRNAexpression in mononuclear cells from different tissues duringacute SIVmac251 infection of macaques. AIDS Res HumRetrovir 12:1263–1272.
Couraud PO (1994): Interactions between lymphocytes, macrophages,and central nervous system cells. J Leukoc Biol 56:407–415.
Davis EJ, Foster TD, Thomas WE (1994): Cellular forms and functionsof brain microglia. Brain Res Bull 34:73–78.
Dickson DW, Mattiace LA, Kure K, Hutchins K, Lyman WD, BrosnanCF (1991): Microglia in human disease, with an emphasis onacquired immune deficiency syndrome. Lab Invest 64:135–156.
Dickson DW, Lee SC, Mattiace LA, Yen SH, Brosnan C (1993):Microglia and cytokines in neurological disease, with specialreference to AIDS and Alzheimer’s disease. Glia 7:75–83.
Eddelston M, Mucke L (1993): Molecular profile of reactive astro-cytes—Implications for their role in neurologic disease. Neuro-science 54:15–36.
Eng LF, Yu ACH, Lee YL (1992): Astrocytic response to injury. ProgBrain Res 94:353–365.
Esiri MM, Morris CS (1991): Immunocytochemical study of macro-phages and microglial cells and extracellular matrix compo-nents in human CNS disease. II. Non-neoplastic diseases. JNeurol Sci 101:59–72.
Fawcett J (1994): Astrocytes and axon regeneration in the centralnervous system. J Neurol 242:S25–S28.
Fischer G (1984): Growth requirements of immature astrocytes inserum-free hormonally defined media. J Neurosci Res 12:543–552.
Flaris N, Densmore T, Molleston M, Hickey W (1993): Characteriza-tion of microglia and macrophages in the central nervous systemof rats: Definition of the differential expression of moleculesusing standard and novel monoclonal antibodies in normal CNSand in four models of parenchyma reaction. Glia 7:34–40.
Frei K, Siepl C, Groscurth P, Bodmer S, Schwerdel C, Fontana A(1987): Antigen presentation and tumor cytotoxicity by inter-feron-g-treated microglial cells. Eur J Immunol 17:1271–1278.
Frei K, Lins H, Schwerdel C, Fontana A (1994): Antigen presentationin the central nervous system. The inhibitory effect of IL-10 onMHC class II expression and production of cytokines dependson the inducing signals and the type of cell analyzed. J Immunol152:2720–2728.
Ganter S, Northoff H, Ma¨nnel D, Gebicke-Ha¨rter PJ (1992): Growthcontrol of cultured microglia. J Neurosci Res 33:218–230.
Gendelman HE, Lipton SA, Tardieu M, Bukrinsky MI, Nottet HSLM(1994): The neuropathogenesis of HIV-1 infection. J LeukocBiol 56:389–398.
Gessani S, Puddu P, Varano B, Borghi P, Conti L, Fantuzzi L, GherardiG, Belardelli F (1994): Role of endogenous interferon-beta inthe restriction of HIV replication in human monocyte/macrophages. J Leukoc Biol 56:358–361.
Giaume C, McCarthy KD (1996): Control of gap-junctional communi-cation in astrocytic networks. TINS 19:319–325.
Giulian D (1987): Ameboid microglia as effectors of inflammation inthe central nervous system. J Neurosci Res 18:155–171.
Giulian D, Baker TJ (1985): Peptides released by ameboid microgliaregulate astroglial proliferation. J Cell Biol 101:2411–2415.
Giulian D, Baker TJ (1986): Characterization of ameboid microgliaisolated from developing mammalian brain. J Neurosci 6:2163–2178.
Giulian D, Li J, Bartel S, Broker J, Li X, Kirkpatrick JB (1995): Cellsurface morphology identifies microglia as a distinct class ofmononuclear phagocyte. J Neurosci 15:7712–7726.
Goldman JE, Chiu F-C (1984): Growth kinetics, cell shape, and thecytoskeleton of primary astrocyte cultures. J Neurochem 42:175–184.
Goldman JE, Schaumburg HH, Norton WT (1978): Isolation andcharacterization of glial filaments from human brain. J Cell Biol78:426–440.
Goudsmit J, Morrow CH, Asher DM, Yanagihara RT, Masters CL,Gibbs CJ, Gajdusek DC (1980): Evidence for and against thetransmissibility of Alzheimer disease. Neurology 30:945–950.
Cultures of Simian Astrocytes and Microglia 589

Graeber MB, Streit WJ, Kreutzberg GW (1988): The microglialcytoskeleton: Vimentin is localized within activated cells in situ.J Neurocytol 17:573–580.
Guillemin G, Boussin FD, Legrand R, Croitoru J, Coffigny H, DormontD (1996): Granulocyte macrophage colony stimulating factorstimulates in vitro proliferation of astrocytes derived fromsimian mature brains. Glia 16:71–80.
Hatten ME (1985): Neuronal regulation of astroglial morphology andproliferation in vitro. J Cell Biol 100:384–396.
Hatten ME, Furie MB, Rifkin DB (1982): Binding of developingmouse cerebellar cells to fibronectin: A possible mechanism forthe foundation of the external granular layer. J Neurosci2:1195–1206.
Hayes GM, Woodroofe MN, Cuzner ML (1987): Microglia are themajor cell type expressing MHC class II in human white matter.J Neurol Sci 80:25–37.
Hertz L, McFarlin DE, Waksman BH (1990): Astrocytes: Auxiliarycells for immune responses in the central nervous system?Immunol Today 11:265–268.
Holliday J, Gruol DL (1993): Cytokine stimulation increases intracellu-lar calcium and alters the response to quisqualate in culturedcortical astrocytes. Brain Res 621:233–241.
Huard TK, Malinoff HL, Wicha MS (1986): Macrophages express aplasma membrane receptor for basement membrane laminin.Am J Pathol 123:365–370.
Janzer RC, Raff MC (1987): Astrocytes induce blood-brain barrierproperties in endothelial cells. Nature 325:253–257.
Kim SU, Sato Y, Silberberg DH, Pleasure DE, Rorke LB (1983): Longterm culture of human oligodendrocytes. J Neurol Sci 62:295–301.
Koka P, He KY, Zack JA, Kitchen S, Peacock W, Fried I, Tran T, YasharSS, Merrill JE (1995): Human immunodeficiency virus 1envelope proteins induce interleukin 1, tumor necrosis factoralpha, and nitric oxide in glial cultures derived from fetal,neonatal, and adult human brain. J Exp Med 182:941–951.
Lasmezas CI, Deslys J-P, Demaimay R, Adjou KT, Lamoury F,Dormont D (1996): BSE transmission to macaques. Nature381:743–744.
Lee SC, Liu W, Brosnan CF, Dickson DW (1994): GM-CSF promotesproliferation of human fetal and adult microglia in primarycultures. Glia 12:309–318.
Levison S, McCarthy K (1991): Astroglia in culture. In Banker G,Goslin K (eds): ‘‘Culturing Nerve Cells.’’ London: MIT Press,pp 309–336.
Li S-L, Kaaya E, Feichtinger H, Biberfeld G, Biberfeld P (1993):Immunohistochemical distribution of leucocyte antigens inlymphoid tissues of cynomolgus monkeys (Macaca fascicu-laris). J Med Primatol 22:285–293.
Liesi P, Dahl D, Vaheri A (1983): Laminin is produced by early ratastrocytes in primary culture. J Cell Biol 96:920–924.
Liesi P, Kaakkola S, Dahl D, Vaheri A (1984): Laminin is induced inastrocytes of adult brain by injury. EMBO J 3:683–686.
Liesi P, Kirkwood T, Vaheri A (1986): Fibronectin is expressed byastrocytes cultured from embryonic and early postnatal ratbrain. Exp Cell Res 163:175–185.
Mallat M, Chamak B (1994): Brain macrophages: Neurotoxic orneurotrophic effector cells? J Leukoc Biol 56:416–422.
Mares V, Viklicky V, Gerstein LM, Draber P, Ciesielski-Treska J(1988): Immunocytochemistry and heterogeneity of rat brainvimentin. Histochemistry 88:575–581.
Martin DL (1995): The role of glia in the inactivation of neurotransmit-ters. In Kettenmann H, Ransom BR (eds): ‘‘Neuroglia.’’ NewYork: Oxford University Press, pp 732–745.
Masuda-Nakagawa LM, Muller KJ, Nicholls JG (1993): Axonalsprouting and laminin appearance after destruction of glialsheaths. Proc Natl Acad Sci USA 90:4966–4970.
McCarthy KD, De Vellis J (1980): Preparation of separate astroglialand oligodendroglial cell cultures from rat cerebral tissue. J CellBiol 85:890–902.
Miller RH, Raff MC (1984): Fibrous and protoplasmic astrocytes arebiochemically and developmentally distinct. J Neurosci 4:585–592.
Miller RH, David S, Patel R, Abney ER, Raff MC (1985): Aquantitative immunohistochemical study of macroglial celldevelopment in the rat optic nerve: In vivo evidence for twodistinct astrocyte lineages. Dev Biol 111:35–41.
Mucke L, Eddelston M (1993): Astrocytes in infectious and immune-mediated diseases of the central nervous system. FASEB J7:1226–1232.
Muller HW, Junghans U, Kappler J (1995): Astroglial neurotrophic andneurite-promoting factors. Pharmacol Ther 65:1–18.
Nakajima K, Hamanoue M, Shimojo M, Takei N, Kohsaka S (1989):Characterization of microglia isolated from a primary culture ofembryonic rat brain by a simplified method. Biomed Res10:411–423.
Norenberg MD (1994): Astrocyte responses to CNS injury. J Neuro-pathol Exp Neurol 53:213–220.
Norton WT, Farooq M, Chiu F-C, Bottenstein JE (1988): Pure astrocytecultures derived from cells isolated from mature brain. Glia1:403–414.
Norton WT, Aquino DA, Hozumi I (1992): Quantitative aspects ofreactive gliosis: A review. Neurochem Res 17:877–885.
Oh YJ, Markelonis GJ, Oh TH (1993): Effects of interleukin-1b andtumor necrosis factor-a on the expression of glial fibrillaryacidic protein and transferrin in cultured astrocytes. Glia8:77–86.
Oh TH, Markelonis GJ, Von Visger JR, Baik B, Shipley MT (1995):Acidic pH rapidly increases immunoreactivity of glial fibrillaryacidic protein in cultured astrocytes. Glia 13:319–322.
Osborn M, Ludwig-Festl M, Weber K, Bignami A, Dahl D, BayreutherK (1981): Expression of glial and vimentin type intermediatefilaments in cultures derived from human glial material. Differ-entiation 19:161–167.
Perzel’ova A, Mares V (1993): Appearance of GFAP-positive cells inadult human brain cultures spontaneously decelerated in growth.Glia 7:237–244.
Peudenier S, Hery C, Montagnier L, Tardieu M (1991): Humanmicroglial cells: Characterization in cerebral tissue and inprimary culture, and study of their susceptibility to HIV-1infection. Ann Neurol 29:152–161.
Price J, Hynes RO (1985): Astrocytes in culture synthetise and secrete avariant form of fibronectin. J Neurosci 5:2505–2527.
Raff MC, Abney ER, Cohen J, Lindsay R, Noble M (1983): Two typesof astrocytes in cultures of developing rat white matter:Differences in morphology, surface gangliosides and growthcharacteristics. J Neurosci 3:1289–1300.
Raivich G, Gehrmann J, Kreutzberg GW (1991): Increase of macro-phage colony-stimulating factor and granulocyte-macrophagecolony-stimulating factor receptors in the regenerating rat facialnucleus. J Neurosci Res 30:682–686.
Reeves SA, Helman LJ, Allison A, Israel MA (1989): Molecularcloning and primary structure of human glial fibrillary acidicprotein. Proc Natl Acad Sci USA 86:5178–5182.
Reimann KA, Waite BCD, Lee-Parritz DE, Lin W, Uchanska-ZieglerB, O’Connell MJ, Letvin NL (1994): Use of human leukocyte-specific monoclonal antibodies for clinically immunotypinglymphocytes of rhesus monkeys. Cytometry 17:102–108.
590 Guillemin et al.

Rieske E, Graeber MB, Tetzlaff W, Czlonkowska A, Streit WJ,Kreutzberg GW (1989): Microglia and microglia-derived brainmacrophages in culture: Generation from axotomized rat facialnuclei, identification and characterization in vitro. Brain Res492:1–14.
Rutka JT (1986): Effects of extracellular matrix proteins on the growthand differentiation of an anaplastic glioma cell line. Can JNeurol Sci 13:301–306.
Schnitzer J, Franke WW, Schachner M (1981): Immunocytochemicaldemonstration of vimentin in astrocytes and ependymal cells ofdeveloping and adult mouse nervous system. J Cell Biol90:435–447.
Sedgwick JD, Schwender S, Imrich H, Do¨rries R, Butcher GW, TerMeulen V (1991): Isolation and direct characterization ofresident microglial cells from the normal and inflamed centralnervous system. Proc Natl Acad Sci USA 88:7438–7442.
Selmaj KW, Farooq M, Norton WT, Raine CS, and Brosnan CF (1990):Proliferation of astrocytes in vitro in response to cytokines. JImmunol 144:129–135.
Shahar A, de Vellis J, Vernadakis A, Haber B (eds) (1989): ‘‘ADissection and Tissue Culture Manual of the Nervous System.’’New York: Alan R. Liss.
Suzumura A, Marunouchi T, and Yamamoto H (1991): Morphologicaltransformation of microglia in vitro. Brain Res 545:301–306.
Sykova E, Svoboda J, Simonova Z (1992): Role of astrocytes in ionicand volume homeostasis in spinal cord during development andinjury. Prog Brain Res 94:47–56.
Thiele DL, Kurosaka M, Lipsky PE (1983): Phenotype of the accessorycell necessary for mitogen-stimulated T and B cell responses inhuman peripheral blood: Delineation by its sensitivity to the
lysosomotropic agent, L-leucine methyl ester. J Immunol 131:2282–2290.
Tsacopoulos M, Magistretti PJ (1996): Metabolic coupling betweenglia and neurons. J Neurosci 16:877–885.
Virchow R (1846): Ueber das granurlirte Ansehen der Wandungen derGehirnventrikel. In Damerow Flemming, Roller (eds): ‘‘Allge ZPsychiat.’’ Berlin: Hirschwald, pp 242–250.
Vitkovic L, Stover E, Koslow SH (1995): Animal models recapitulateaspects of HIV/CNS disease. AIDS Res Hum Retrovir 11:753–759.
Von Bernhardi R, Muller KJ (1995): Repair of the central nervoussystem. J Neurobiol 27:353–366.
Watkins BA, Dorn HH, Kelly WB, Armstrong RC, Potts BJ, MichaelsF, Kufta CV, Dubois-Dalq M (1990): Specific tropism of HIV-1for microglial cells in primary human brain cultures. Science249:549–553.
Wilkin GP, Marriott DR, Cholewinski AJ (1990): Astrocyte heterogene-ity. TINS 13:43–46.
Wright SD, Meyer BC (1985): Fibronectin receptor of human macro-phages recognizes the sequence Arg-Gly-Asp-Ser. J Exp Med162:762–767.
Yamada T, Kawamata T, Walker DG, McGeer PL (1992): Vimentinimmunoreactivity in normal and pathological human braintissue. Acta Neuropathol 84:157–162.
Yamauchi K, Martinet Y, Crystal RG (1987): Modulation of fibronectingene expression in human mononuclear phagocytes. J ClinInvest 80:1720–1727.
Yazaki T, Asou H, Arimoto K, Toya S, Uyemura K (1995): Decrease ofNCAM expression and astrocyte-neuron interaction in long-term cultured astrocytes. NeuroReport 6:1085–1088.
Cultures of Simian Astrocytes and Microglia 591