Microglial chimaerism in human xenografts to the rat brain
9
Pergamon 0361-9230(95)02004-B BrainResearchBulletin, Vol. 38, No. 4. pp. 383-391, 1995 Copyright© 1995 Elsevier Science Inc. Printedin the USA.All rights reserved 0361-9230/95$9.50 + .00 Microglial Chimaerism in Human Xenografts to the Rat Brain CHRISTIAN GENY,* SOU,h,D NAIMI-SADAOUI,*~t ABD EL MADJID BELKADI,* ROLAND JENY,1- MOURAD KAMMOUN] ~ AND MARC PESCHANSK1.1 *INSERM U 421, Neuroplasticit~ et Th~rapeutique, Facult~ de M~decine, 8 rue g~n~ral Sarrail, 94010 Cr~teil Cedex, France ?~HSpitalEsquirol, 94410 Saint-Maurice, France .l:Laboratoire de Neurosciences. D~partement de Biologie, Facult~ des Sciences Semlalia-Marrakech, Morocco [Received 24 March 1995; Revised 3 June 1995; Accepted 5 June 1995] ABSTRACT: Neural tissue from human fetuses is currently used for intracerebral transplantation to treat patients with Parkin- son's disease. The development of the human fetal tissue fol- lowing grafting has been considered mostly, up to now, from the neuronal point of view in xonografts. Very little is known, in con- trast, about nonneuronel, glial, or vascular calls in the grafts. Comparison of the data gathered on the development of grafted human neurons with those obtained in comparable studies using rat transplants has demonstrated species-specific features. We have therefore undertaken a series of studies dealing with non- neuronal cells in human-to-rat transplants to reveal other pos- sibie species-specificity of the human tissue. This study has, ac- cordingly, been devoted to the immunohistochemicel analysis of microglia of host and donor origins in a human to rat xenograft paradigm allowing clear distinction of the origin of the cells. Hu- man neural tissue was transplanted as a cell suspension into the thalamus of adult rata. Amoeboid human microglia were ob- served in 1-, 2-, and 3-month-old transplants, but their density, already relatively low at the first stage, decreased further over time. Ramified human microglia were only occasional. In sharp contrast, host rat microglia rapidly invaded the transplant in the absence of any sign of necrosis. The rat cells exhibited first an amoeboid morphology but progressed at the later stages toward a more mature, ramified morphology. These results indicate that donor microglia are quite few in number at first and, at least, do not proliferate actively after transplantation. They seem rather to disappear over time. The parallel migration of a large number of host microglia into the transplant and the apparent maturation of these cells lead to the formation of a cellular chimaera. Ex- trapolation of these results to clinical neural grafting suggests that most, if not all, microglia in fetal transplants may rapidly be of host origin in patients. KEY WORDS: Human microglia, Immunohistochemistry, Brain transplantation, Parkinson's disease, Human fetus. INTRODUCTION Clinical use of human neural tissue for intracerebral transplan- tation has recently provided interesting results in patients with Parkinson's disease [10,15,21-23,30,34,37]. This clinical ex- r To whom requests for reprints should be addressed. perimentation is based upon a number of studies in animals [see refs and discussion in 2,11,16] in which, for most of the cases, fetal rat neural tissue has been transplanted into host brains. How far the development of the human fetal tissue transplanted to the human brain can be extrapolated from the results of the rodent allografts is not clear. Studies specifically exploring the development of human neural cells transplanted into the brain of adult rats [3,4,35,36] have demonstrated, in particular, that the rate of maturation of neurons of interest (mesencephalic dopaminergic neurons) was much slower than that of their ro- dent homologs. Although developing rat neurons may mature over a period of a few weeks after transplantation into adult brain, human neurons take several months to develop. This is- sue may be relevant not only for the neurons that are trans- planted but also for the glial and vascular environment in which they develop, and, up to now, very little is known of the mat- uration of grafted glial and vascular cells. To provide data upon which a better extrapolation would be possible than those provided by grafting of fetal rat tissue, we have undertaken a series of studies dealing with glial and vascular elements in human xenografts into the brain of immunosup- pressed adult rats. We have, thus, demonstrated first that vascu- larization is much delayed in human xenografts into the rat brain. This 1- to 2-month-long delay is likely to be dependent on the rate of maturation of the transplanted tissue [ 12]. As a follow-up, this study analyzes, using specific immuno- cytochemical markers, donor and host microglial cells in human to rat neural transplants. ABBREVIATIONS MHC: Major histocompatibility complex; HLA: Human leu- cocyte antigens. MATERIALS AND METHODS A total of 10 female Sprague-Dawley rats (200-225 g at the beginning of the experiments) were used in this study. Tissue for transplantation was trimmed out of human fetuses at gestational 383
-
Upload
chu-montpellier -
Category
Documents
-
view
2 -
download
0
Transcript of Microglial chimaerism in human xenografts to the rat brain

Pergamon
0361-9230(95)02004-B
Brain Research Bulletin, Vol. 38, No. 4. pp. 383-391, 1995 Copyright © 1995 Elsevier Science Inc. Printed in the USA. All rights reserved
0361-9230/95 $9.50 + .00
Microglial Chimaerism in Human Xenografts to the Rat Brain
CHRISTIAN GENY,* SOU,h,D NAIMI-SADAOUI,*~t ABD EL MADJID BELKADI,* ROLAND JENY,1- MOURAD KAMMOUN] ~ AND MARC PESCHANSK1.1
*INSERM U 421, Neuroplasticit~ et Th~rapeutique, Facult~ de M~decine, 8 rue g~n~ral Sarrail, 94010 Cr~teil Cedex, France
?~HSpital Esquirol, 94410 Saint-Maurice, France .l:Laboratoire de Neurosciences. D~partement de Biologie, Facult~ des Sciences Semlalia-Marrakech, Morocco
[Received 24 March 1995; Revised 3 June 1995; Accepted 5 June 1995]
ABSTRACT: Neural tissue from human fetuses is currently used for intracerebral transplantation to treat patients with Parkin- son's disease. The development of the human fetal tissue fol- lowing grafting has been considered mostly, up to now, from the neuronal point of view in xonografts. Very little is known, in con- trast, about nonneuronel, glial, or vascular calls in the grafts. Comparison of the data gathered on the development of grafted human neurons with those obtained in comparable studies using rat transplants has demonstrated species-specific features. We have therefore undertaken a series of studies dealing with non- neuronal cells in human-to-rat transplants to reveal other pos- sibie species-specificity of the human tissue. This study has, ac- cordingly, been devoted to the immunohistochemicel analysis of microglia of host and donor origins in a human to rat xenograft paradigm allowing clear distinction of the origin of the cells. Hu- man neural tissue was transplanted as a cell suspension into the thalamus of adult rata. Amoeboid human microglia were ob- served in 1-, 2-, and 3-month-old transplants, but their density, already relatively low at the first stage, decreased further over time. Ramified human microglia were only occasional. In sharp contrast, host rat microglia rapidly invaded the transplant in the absence of any sign of necrosis. The rat cells exhibited first an amoeboid morphology but progressed at the later stages toward a more mature, ramified morphology. These results indicate that donor microglia are quite few in number at first and, at least, do not proliferate actively after transplantation. They seem rather to disappear over time. The parallel migration of a large number of host microglia into the transplant and the apparent maturation of these cells lead to the formation of a cellular chimaera. Ex- trapolation of these results to clinical neural grafting suggests that most, if not all, microglia in fetal transplants may rapidly be of host origin in patients.
KEY WORDS: Human microglia, Immunohistochemistry, Brain transplantation, Parkinson's disease, Human fetus.
INTRODUCTION
Clinical use of human neural tissue for intracerebral transplan- tation has recently provided interesting results in patients with Parkinson's disease [10,15,21-23,30,34,37]. This clinical ex-
r To whom requests for reprints should be addressed.
perimentation is based upon a number of studies in animals [see refs and discussion in 2,11,16] in which, for most of the cases, fetal rat neural tissue has been transplanted into host brains. How far the development of the human fetal tissue transplanted to the human brain can be extrapolated from the results of the rodent allografts is not clear. Studies specifically exploring the development of human neural cells transplanted into the brain of adult rats [3,4,35,36] have demonstrated, in particular, that the rate of maturation of neurons of interest (mesencephalic dopaminergic neurons) was much slower than that of their ro- dent homologs. Although developing rat neurons may mature over a period of a few weeks after transplantation into adult brain, human neurons take several months to develop. This is- sue may be relevant not only for the neurons that are trans- planted but also for the glial and vascular environment in which they develop, and, up to now, very little is known of the mat- uration of grafted glial and vascular cells.
To provide data upon which a better extrapolation would be possible than those provided by grafting of fetal rat tissue, we have undertaken a series of studies dealing with glial and vascular elements in human xenografts into the brain of immunosup- pressed adult rats. We have, thus, demonstrated first that vascu- larization is much delayed in human xenografts into the rat brain. This 1- to 2-month-long delay is likely to be dependent on the rate of maturation of the transplanted tissue [ 12].
As a follow-up, this study analyzes, using specific immuno- cytochemical markers, donor and host microglial cells in human to rat neural transplants.
ABBREVIATIONS
MHC: Major histocompatibility complex; HLA: Human leu- cocyte antigens.
MATERIALS AND METHODS
A total of 10 female Sprague-Dawley rats (200-225 g at the beginning of the experiments) were used in this study. Tissue for transplantation was trimmed out of human fetuses at gestational
383

384 G E N Y ET AL
FIG. I. Nissl-stained coronal section of the rat brain through transplants at two different developmental stages (hi: hippocampus, G: graft). (A) One month after transplantation, the cellular density in the graft is very high. (B) Three months after transplantation, the volume of the graft is larger and the cellular density is lower, as demonstrated by a reduced intensity of staining. Scale bar = 500 #m.
FIG. 2. Transverse section through the cervical spinal cord of an 8-week-old human fetus immunostained with Ki-M7 and counterstained with Cresyl violet. The arrow indicates one of the Ki-M7+ microglial cells. Scale bar ~ 50 ~,m.

MICROGLIA IN HUMAN TO RAT BRAIN XENOGRAFTS 385
age varying from 6 to 9 weeks, appreciated by measuring crown to rump length between 16 and 42 mm [27].
Lesioning and Grafting Procedures
Rats were anesthetized with chloral hydrate (400 mg/kg) and received stereotaxically a slow pressure injection of kainic acid (5 nmol in 0.15 #1 over l0 mn) using a 1 #1 Hamilton syringe into the right thalamus. Two to four weeks later, they were re- anesthetized and received, according to the same protocol, a slow pressure injection of 10 #1 of the human cell suspension, prepared as described below according to a protocol derived from Brundin et al. [3]. This "lesion-graft ing" experimental paradigm was chosen because it allows transplants to grow larger than when fetal cells are implanted into the intact adult parenchyma [29]. Pairs of rats were allowed to survive 2, 4, 6, 8, and 12 weeks after grafting. They were kept under immunosuppression (cy- closporine A, 10 mg/kg daily; IP) until sacrifice.
Fetuses were obtained from suction abortion according to a protocol described elsewhere [20] that has received the agree- ment of the French National Ethical Committee. Briefly, after informed consent of the woman, suction abortion under deep anesthesia was carried out with ultrasound guidance using a 12 mm diameter wide cannula attached to a 50 ml syringe. After careful dissection of the envelopes overlying the central nervous system, the hindbrain and upper spinal cord were trimmed out under sterile conditions and cut into 1 mm ~ cubes. Pieces were collected in a tube containing 200 #1 of D-glucose saline (0.6%) with 0.1% trypsin (Sigma) and 0.05% DNAse (Sigma). After 20 rain incubation at 37°C, tissue was rinsed with glucose-saline and
TABLE I
SEMIQUANTITATIVE EVALUATION OF THE NUMBER OF LEUCOCYTES IMMUNOSTAINED BY THE VARIOUS MARKERS
I Month 2 Months 3 Months
CD68 (Ki-M7) + + + + + CD4 - - - HLA-ABC - + + HLA-DR - - - CD1 lb (Leu 15) - OX42 + + + + + + + + + ED1 + + + ++ + OX18 + + + ++ ++ OX6 - - OX19 - - -
- , absent; +, small; ++, moderate; +++, high.
then dissociated with DNAse in 200 #1 glucose-saline using fire- polished Pasteur pipettes. Cell survival rate, appreciated using the acridine orange-ethidium bromide fluorescent technique, was over 90%. The final cell concentration ranged between 50.000 and 100.000 per #1.
Histological Procedures
Rats received a lethal dose of sodium pentabarbitone and were perfused transcardially with 100 ml of heparinized phos- phate-buffered saline (pH 7.4; 0.1 M) fol lowed by 400 ml of
o
FIG. 3. Ki-M7 immunostained human microglial cells in transplants at various stages. (A) One month. The mi- croglial cells have a macrophagic phenotype. (B) Two months after transplantation. Note the presence of thick immunostained processes (arrow) of the rare cells present within the graft. Scale bar = 50/~m.
386 GENY ET AL.
°
B
, : .
" x M
C ~ D
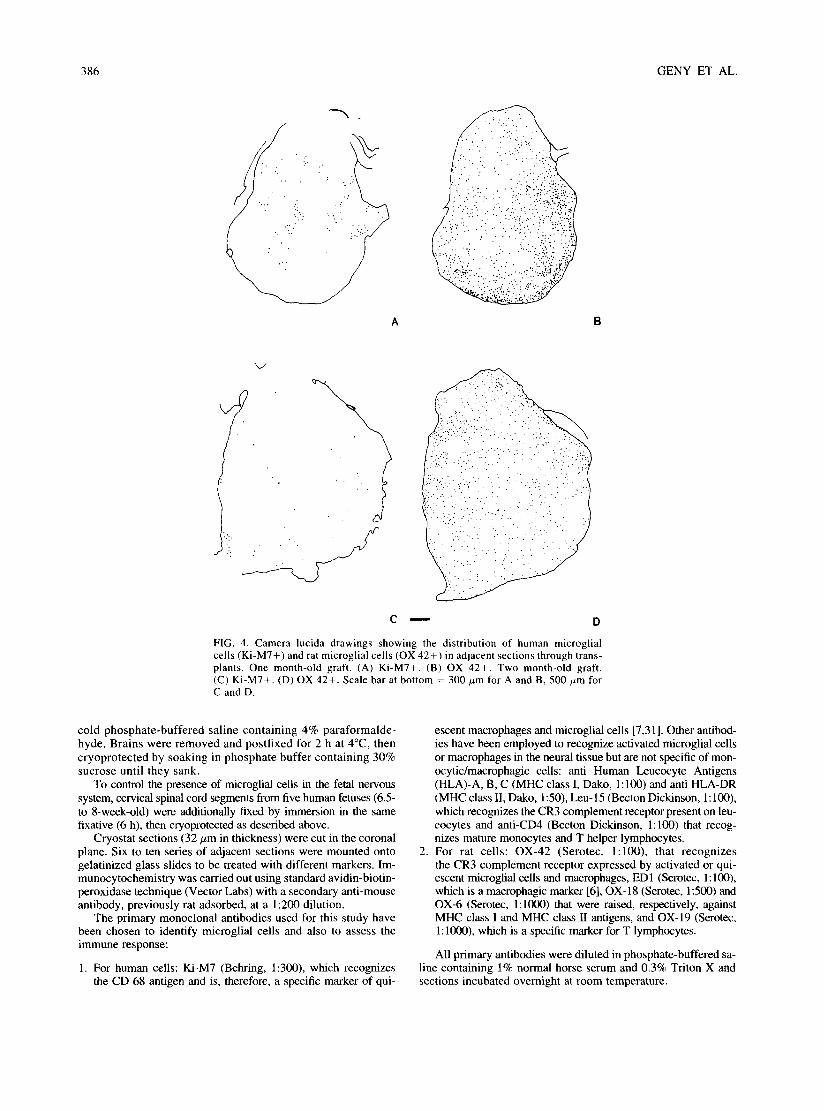
FIG. 4. Camera lucida drawings showing the distribution of human microglial cells (Ki-M7+) and rat microglial cells (OX 42+) in adjacent sections through trans- plants. One month-old graft. (A) Ki-M7+. (B) OX 42+. Two month-old graft. (C) Ki-M7+. (D) OX 42+. Scale bar at bottom = 300/zm for A and B, 500 #m for C and D.
cold phosphate-buffered saline containing 4% paraformalde- hyde. Brains were removed and postfixed for 2 h at 4°C, then cryoprotected by soaking in phosphate buffer containing 30% sucrose until they sank.
To control the presence of microglial cells in the fetal nervous system, cervical spinal cord segments from five human fetuses (6.5- to 8-week-old) were additionally fixed by immersion in the same fixative (6 h), then cryoprotected as described above.
Cryostat sections (32/.tm in thickness) were cut in the coronal plane. Six to ten series of adjacent sections were mounted onto gelatinized glass slides to be treated with different markers. Im- munocytochemistry was carried out using standard avidin-biotin- peroxidase technique (Vector Labs) with a secondary anti-mouse antibody, previously rat adsorbed, at a 1:200 dilution.
The primary monoclonal antibodies used for this study have been chosen to identify microglial cells and also to assess the immune response:
1. For human cells: Ki-M7 (Behring, 1:300), which recognizes the CD 68 antigen and is, therefore, a specific marker of qui-
2.
escent macrophages and microglial cells [7,31 ]. Other antibod- ies have been employed to recognize activated microglial cells or macrophages in the neural tissue but are not specific of mon- ocytic/macrophagic cells: anti Human Leucocyte Antigens (HLA)-A, B, C (MHC class I, Dako, 1:100) and anti HLA-DR (MHC class II, Dako, 1:50), Leu-15 (Becton Dickinson, 1:100), which recognizes the CR3 complement receptor present on leu- cocytes and anti-CD4 (Becton Dickinson, 1:100) that recog- nizes mature monocytes and T helper lymphocytes. For rat cells: OX-42 (Serotec, 1:100), that recognizes the CR3 complement receptor expressed by activated or qui- escent microglial cells and macrophages, ED1 (Serotec, 1:100), which is a macrophagic marker [6], OX-18 (Serotec, 1:500) and OX-6 (Serotec, 1:1000) that were raised, respectively, against MHC class I and MHC class II antigens, and OX-19 (Serotec, 1:1000), which is a specific marker for T lymphocytes.
All primary antibodies were diluted in phosphate-buffered sa- line containing 1% normal horse serum and 0.3% Triton X and sections incubated overnight at room temperature.

MICROGL1A IN HUMAN TO RAT BRAIN XENOGRAFI'S 387
• •
) ~ Y
f ~ s ~
FIG. 5. Rat MHC immunoreactivity in a 2-months graft. (A) MHC class II (OX 6). (B) MHC class I (OX18). Note that microglial cells express MHC class I but not MHC class II in the transplants. MHC class II immunoreactivity is present in the lesioned rat parenchyma. Scale bar = 50 #m,
RESULTS
Cresyl violet staining allowed an early identification of the transplants in the host brain (Figs. lA, B). They appeared as large groups of packed well-stained cells. Borders between the graft and the host tissue were well-delineated on a cytoarchitectural basis because of the contrast between the nicely organized host cells respecting specific alignments and the variable polarities of grafted cells. At all stages, human cells also appeared smaller, more rounded, and more tightly packed together than cells in the host parenchyma. Nissl-staining was heterogenous due to differ- ing densities of cells within the graft. Clusters of very tightly packed cells tended to be less numerous and reduced in size over time, suggesting a progressive maturation of the tissue leading to the widening of the neuropil occupied by cell processes. Al- though the small number of cases and a certain variability of the results do not allow us to draw a final conclusion, comparison of sizes seemed to reveal an enlargement of the transplants between l and 3 months. Neither signs of necrosis nor cell-cuffing of blood vessels by mononuclear cells were observed.
Microglia/Macrophages in the Human Fetal Spinal Cord
Positive results, that is, cellular immunostaining, were ob- tained in all fetuses with only one antibody, Ki-M7 (Fig. 2). Immunostained cells were present mostly in the proximity of the meningeal surface, close to the periventricular germinative zone, and along the medial septum of the primitive cord. Immuno- staining was cytoplasmic and granular. Two morphological types of cells were labeled, large rounded cells without processes and
smaller cells displaying amoeboid processes. No labeled cells exhibited the small soma and thin ramified processes character- istic of quiescent mature microglia.
Human Microglia in the Transplants
Among the battery of antibodies against human leukocyte an- tigens used in this study (Table 1), Ki-M7 was the only one that labeled cells displaying morphologies compatible with microg- lial/macrophagic elements in the transplants (Fig. 3). Anti-HLA A, B, C, (Class I) antibodies stained rare endothelial cells in capillary walls. Other antibodies (Leu 15, anti-CD4, and anti- HLA-DR) did not stain cellular elements either in the transplant or in the host.
Ki-M7 immunoreactive cells were all contained in the trans- plants. They were observed at all stages, from 2 weeks to 3 months after transplantation. Their density diminished, however, over time (compare Figs. 4A, C), indicating at least that the hu- man microglial cells did not proliferate actively after transplan- tation and suggesting even that they disappeared progressively, although the experimental procedures precluded precise quanti- fication of such a phenomenon.
Up to 1 month, most Ki-M7 immunoreactive cells had a mac- rophagic morphology with a large round cell body (Fig. 3A). At later stages (Fig. 3B), immunostained cells resembled amoeboid microglia with one or two thick poorly ramified processes. Cells resembling mature microglia with a small cell body and thin highly ramified processes were not observed.

388 GENY ET AL.
~i!ii~ ~̧ !i~
FIG. 6. Rat macrophages immunostained with EDI in a 1-month-old graft. Labeled cells are numerous in host areas lesioned by the injection of kainate such as the hippocampus (hi) or the area bordering ventrally the transplant (across the dashed line, which indicates the graft border). The density of immunostained cells is higher in the outer than in the inner portions of the graft, but rat macrophages are already present at this stage in its entire volume. This section has been lightly counter- stained with cresyl violet to visualize the host-graft border. Scale bar = 500 #m.
Rat Microglia in the Transplants
Rat microglial cells could be observed in the transplant as early as 2 weeks after grafting, and they were widespread in the entire transplant already at 1 month (Fig. 4B). In the battery of antibodies used to reveal the rat cells (Table 1), two did not stain cells in the transplant, namely OX-6, which recognizes MHC class II antigens (Fig. 5A), and OX-19, which is a lymphocytic marker. OX-42 was the most stable marker, whereas staining with ED-I and OX-18 decreased progressively over time (Fig. 5B).
At 2 weeks and at 1 month (Figs. 4B, 6), rat microglial cells were more numerous in some outer zones of the transplant, close to the host-graft border, than in its more central zones. There was, however, no large area totally devoid of microglial cells. This differential distribution did not persist at later stages (Fig. 4D). At 3 months, the density of labelled cells remained slightly lower than in the normal host parenchyma.
All markers stained cells with different morphological char- acteristics. ED-I and OX-18 stainings were, however, more in- tense in "macrophagic-type" or "amoeboid microglia-type"
cells displaying a large round cell body and few, if any, thick short processes. OX-42 labeled both these cells and others of a "quiescent microglia-type" with a smaller cell body and thinner more ramified processes. One month after transplantation, most of the cells exhibited an amoeboid microglial morphology with a large soma and thick processes (Fig. 7A), whereas no quiescent microglial cells were observed yet. At 3 months, amoeboid mi- croglial cells were still observed, but many cells displayed the morphology of quiescent microglia with a smaller soma and thin- ner, longer processes that were highly ramified (Fig. 7C).
DISCUSSION
A human-to-rat neural xenograft paradigm and a battery of antibodies recognizing epitopes presented by either human or rat microglial/macrophagic cells at various stages of differentiation have been used to study in parallel donor and host cells over time. Two major results were obtained: first, that the density of human microglia was low and further decreased over time, and second, that rat microglial cells rapidly invaded the transplant in large numbers and settled there to acquire a mature quiescent microglia phenotype.
Scarcity and Decreased Density of Donor Microglial Cells
Microglial cells are mesenchymal derivatives that enter the central nervous system progressively during development [see refs and discussion in 24]. In fact, it has been suggested out of studies in rodents that most of these cells enter the brain relatively late during ontogenesis. In agreement with these data, well-dif- ferentiated microglia have only been observed in the human fetus at 35 weeks of gestation [7]. Our results in 6.5- to 8 week-old human fetuses are coherent with these suggestions because the density of microglial cells was low and no cell displayed the morphology of mature quiescent microglia. Conversely, our re- sults show that a few cells immunoreactive for a microglial marker are present at a very early stage of development.
Our results, and those obtained by others in slightly older human fetuses [7,13,18,31], indicate that the neural tissue such as that currently used for therapeutic purposes does contain a small number of microglial cells. Our results bring two comple- mentary informations about these cells in transplants, first, that their density (already very low 2 weeks after grafting) decreases over time and, second, that they do not express human leucocyte antigens either of Class 1 or II.
The decreased density of human microglia and the scarcity of differentiated cells in the transplant contrast with the early mi- gration and differentiation of numerous microglial cells from the host. These results do not demonstrate an actual loss in number of human microglia because transplants seemed to enlarge over time, and, therefore, proliferation of other cell types may have diluted these cells. They indicate, however, that there is l i t t le - - if any--proliferat ion of human microglia. Perry and Lund [28] have observed a complete lack of donor microglia in mice retinae grafted for 5 weeks to the brain. Although the paradigms are dissimilar, the two studies concur to indicate that donor microglia is not a major component of transplants.
It is interesting to mention that, in the adult brain, microglia play a key role in immune responses by presenting antigens, in a MHC-restricted manner, to activated lymphocytes [17,25,32]. The absence of MHC II expression in the transplant is a likely indication that this phenomenon does not occur in this paradigm.
Constitution (~f a Human-Rat Neural Chimaera
In sharp contrast with the paucity of human microglia and its decreased density over time, host rat microglial cells populate the

M I C R O G L I A IN H U M A N T O R A T B R A I N X E N O G R A F F S 389
FIG. 7. Morphological features of OX42 immunostained ceils in the transplant. (A) 1 month. (B) 2 months. (C) 3 months after grafting. Note that the cell soma tends to become smaller at 3 months, whereas the length and ramification of the processes increase as their caliber decreases. Scale bar = 300/~m.

390 GENY ET AL.
human transplant very rapidly, in agreement with the observation made in retina grafts to the brain by Perry and Lund [28]. Anal- ysis of the distribution and morphology of rat cells in the trans- plant suggests that they migrate from the host parenchyma into the transplant, then settle down there and mature. There is no visible cuffing of blood vessels in the transplants (in which, ac- tually, blood vessels are absent at 1 month and rare afterwards [12]), which would have indicated another source for invasion of the transplant through recruitment of blood-borne cells. This early migration of host microglia raises several questions. The morphology of the cells, that is, amoeboid rather than ramified quiescent microglia, indicates that the cells are in an activated stage [9]. Accordingly, many of them are immunoreactive for ED-1 and MHC class I antigens [9]. Implantation of the human tissue into an excitotoxically lesioned area, in which microglia present an activated phenotype [26], is probably not the only explanation for the phenotype of the migrating cells. This mor- phology was indeed also observed in the outer-zone of the trans- plant in areas abutting to apparently intact host tissue (the internal capsule, for instance). It is therefore likely that microglial cells were induced by the transplant to alter their phenotype and ac- quire the ability to migrate. This implies an interaction between donor and host tissue probably due to diffusible factors, It is interesting to note that, as already indicated, this migration occurs in the absence of a capillary network [12]. This result implies that angiogenesis and microglial migration are, at least largely, independent. It indicates, in particular, that microglial cells find, in the transplant, the substrates necessary for their migration in the absence of the basal lamina of blood capillaries [5]. Inciden- tally, it suggests also a striking capacity of adult rat cells to sur- vive and differentiate in a tissue that is very poor in blood-borne compounds.
The role of the host microglia in the transplants is but a matter of speculation. One can exclude a participation to a rejection process in the absence of T lymphocytes and MHC class lI ex- pression and because microglia evolves toward a quiescent ram- ified phenotype. Whether the activated stage (ED1 +, MHC I +, amoeboid morphology) observed for most cells at l month and a decreasing number of them afterwards is associated to phago- cytic activities and scavenging of debris or simply to the migra- tory process [1,8,9,14,19,33] remains to be determined.
In conclusion, the analysis of donor microglia in a human to rat xenograft paradigm indicates a progressive dilution, or elim- ination, of these cells. Microglial cells from the host are, in con- trast, induced to migrate in massive numbers into the transplant and to differentiate in quiescent microglia.
ACKNOWLEDGEMENTS
This research received the financial support of a CNAMTS/INSERM grant, Agence Nationale de Recherche contre le SIDA, Association Fran- qaise contre les Myopathies, DRED and Assistance Publique des H6pi- taux de Paris (CRC 91-1401). The authors thank V6ronique Poli for technical assistance and Sandoz labs for cyclosporine A.
REFERENCES
1. Ashwell, K. The distribution of microglia and cell death in the fetal rat forebrain. Dev. Brain Res. 58:1-12; 1991.
2. Bj6rklund, A.; Dunnett, S. B.; Nikkhah, G. Nigral transplants in the rat parkinson model functional limitations and strategies to enhance nigrostriatal reconstruction. In: Dunnen, S. B.; Bj~Srklund, A., eds. Functional neural transplantation. New York: Raven Press; 1994:47-69.
3. Brundin, P.; Nilsson, O. G.; Strecker, R. E.; Lindvall, O.; Astedt, B.; BjiSrklund, A. Behavioural effects of human fetal dopamine neu-
rons grafted in a rat model of Parkinson's disease. Exp. Brain Res. 65:235-240; 1986.
4. Brundin, P.; Strecker, R. E.; Widner, H.; Clarke, D. J.; Nilsson, O. G.; Astedt, B.; Lindvall, O.; Bjtirklund, A. Human fetal dopamine neurons grafted in a rat model of Parkinson's disease: Immunolog- ical aspects, spontaneous and drug-induced behaviour, and dopa- mine release. Exp. Brain Res. 70:192-208; 1988.
5. Chamak, B.; Mallat, M. Fibronectine and laminin regulate the in vitro differentiation of microglial cells. Neuroscience 45:513-527; 1991.
6. Dijkistra, C. D.; D6pp, E~ A.; Joling, P.; Kraal, G. The heterogeneity of mononuclear phagocytes in lymphoid organs: Distinct macro- phages subpopulations in the rat recognized by monoclonal antibod- ies EDI, ED2 and ED3. Immunology 54:589-599; 1985.
7. Esiri, M. M.; AI Izzi, M. S.; Reading, M. C. Macrophages, microglial cells, and HLA-DR antigens in fetal and infant brain. J. Clin, Pathol. 44:102-106; 1991.
8. Ferret, l.; Bernet, E.; Soriano, E.; Del Rio, T.; Fonseca, M. Naturally occurring cell death in the cerebral cortex of the rat and removal of cells by transitory phagocytes. Neuroscience 39:451-458; 1990.
9. Flaris, N. A.; Densmore, T. L.; Molleston, M. C.; Hickey, W. F. Characterization of microglia and macrophages in the central ner- vous system of rats: Definition of the differential expression of mol- ecules using standard and novel monoclonal antibodies in normal CNS and in four models of parenchymal reaction. Glia 7:34-40; 1993.
10. Freed, C. R.; Breeze, R. E.; Rosenberg, N. L.; Schneck, S. A.; Kriek, E.; Qi, J. X.; Lone, T.; Zhang, Y. B.; Snyder, J. A.; Wells, T. H.; Olson Ramig, L.; Thompson, L.; Mazziota, J. C.; Huang, S. C.; Grafton, S. T.; Brooks, D.; Sawle, G.; Schroter, G.; Ansari, A. A. Survival of implanted fetal dopamine cells and neurologic improve- ment 12 to 46 months after transplantation for Parkinson's disease. N. Engl. J. Med. 327:1549-1555; 1992.
11. Freed, W. J. Substantia nigra grafts and Parkinson's disease: From animal experiments to human therapeutic trials. Restor. Neurol, Neu- rosci. 3:109-134; 1991.
12. Geny, C.; Naimi-Sadaoui, S.; Jeny, R.; Belkadi, A. M.; Juliano, S.; Peschanski, M. Long-term delayed vascularization of human neural transplants to the rat brain. J. Neurosci. 14:7553-7562; 1994.
13. Gould, S. J.; Howard, S. An immunohistologieal study of macro- phages in the human fetal brain. Neuropathol. Appl. Neurobiol. 17:383-390; 1991.
14. Hamburger, V.; Levi-Montacini, R. Proliferation, differentiation in the spinal ganglia of the chick embryo under normal and experi- mental conditions. J. Exp. Zool. 111:457-501; 1949.
15. Henderson, B. T.; Clough, C. G.; Hughes, R. C.; Hitchkock, E. R.; Kenny, B. G. Implantation of human ventral mesencephalon to the right caudate nucleus in advanced Parkinson's disease. Arch. Neurol. 48:822-827; 1991.
16. Herman, J. P.; Abrous N. Dopaminergic neural grafts after fifteen years: Results and perspectives. Prog. Neurobio. 44:1-35; 1994.
17. Hickey, W. F.; Kimura, H. Perivascular microglial cells of the CNS are bone marrow-derived and present antigen in vivo. Science 239:290-292; 1988.
18. Hutchins, K. D.; Dickson, D. W.; Rashbaum, W. K.; Lyman, W. D. Localization of morphologically distinct microglial populations in the developing human fetal brain: Implications for ontogeny. Dev. Brain Res. 55:95-102; 1990.
19. Innocenti, G. M.; Clarke, S.; Koppel, H. Transitory macrophages in the white matter of the developping visual cortex. II: Development and relations with axonal pathways. Dev. Brain. Res. 11:56-66; 1983.
20. Jeny, R.; Peschanski, M.; Cynober, E.; Kammoun, M.; Bouzaghar, A.; Hammani, M.; Cesaro, P. Apport de 1'6chographie dans les pr~- lbvements embryonnaires h vis6e de greffes intrac6r6brales dans le traitement de la maladie de Parkinson. J. Echo. Med. Ultras. 14:19- 22; 1993.
21. Lindvall, O.; Sawle, G.; Widner, H.; Rothwell, J. C.; Bj6rklund, A.; Brooks, D.; Brundin P., Frackowiack, R.; Marsden, C. D.; Odin, P.; Rehncrona, S. Evidence of long term survival and function of do- paminergic grafts in progressive Parkinson's disease. Ann. Neurol. 35:172-180; 1994.

MICROGLIA IN H U M A N TO RAT BRAIN XENOGRAFTS 391
22. Lindvall, O,; Brundin, P.; Widner, H.; Rehncrona, S.; Gustavii, B.; Frackowiak, R.; Leenders, K. L.; Swale, G.; Rothwell, J. C.; Mars- den, C. D. Grafts of fetal domapine neurons survive and improve motor function in Parkinson's disease. Science 247:574-577; 1990.
23. Lindvall, O.; Widner, H.; Rehncrona, S.; Brundin, P.; Odin, P.; Gus- tavii, B.; Frackowiack, R.; Leenders, K. L.; Sawle, G.; Rothwell, J. C.; Bjrrklund, A. Transplantation of fetal dopamine neurons in Parkinson's disease: One-year clinical and neurophysiological ob- servations in two patients with putaminal implants. Ann. Neurol. 31:155-165; 1992.
24. Ling, E. A.; Wong, W. C. The origin and nature of ramified and amoeboid microglia: A historical review and current concepts. Glia 7:9-18; 1993.
25. Lowe, J.; MacLennan, K. A.; Powe, D. G.; Pound, J. D.; Palmer, J. B. Microglial cells in human brain have phenotypic characteristics related to possible function as dendritic antigen presenting cells. J. Pathol. 159:143-149; 1989.
26. Marty, S.; Dusart, I.; Peschanski, M. Glial changes following an excitotoxic lesion in the CNS -I. Microglial/Macrophages. Neuro- science 45:529-539; 1991.
27. O'Rahilly, R.; Muller, F. Developmental stages in human embryo. Washington, DC: Carnegie Institution; 1987.
28. Perry, V. H.; Lund, R. D. Microglia in retinae transplanted to the central nervous system. Neuroscience 31:453-462; 1989.
29. Peschanski, M.; lsacson, O. Fetal homotypic transplant in the exci- totoxically neuron depleted thalamus: Light microscopy. J. Comp. Neurol. 274:449-463; 1988.
30. Peschanski, M.; Defer, G.; N'guyen, J. P.; Ricolfi, F.; Monfort, J. C.; Remy, P.; Geny, C.; Samson, Y.; Hantraye, Ph.; Jeny, R.; Gaston, A.; Krravel, Y.; Degos, J. D.; Cesaro, P. Bilateral motor improvement and alteration of L-DOPA effect in two patients with
Parkinson's disease following intrastriatal transplantation of fetal ventral mesencephalon. Brain 117:487-499; 1994.
31. Peudenier, S.; Hery, C.; Ng, K. H.; Tardieu M. HIV receptors within the brain, a study of CD4 and MHC-11 on human neurons, astrocytes and microglial cells. Res. Virol. 142:145-149; 1991.
32. Poltorak, M.; Freed, W. J. Immunological reactions induced by in- tracerebral transplantation: Evidence that host mieroglia but not as- troglia are the antigen-presenting cells. Exp. Neurol. 103:222-233; 1989.
33. Sohal, G, S.; Weidman T. A. Ultrastructure and sequence of embry- onic cell death in normal and peripherally deprived trochlear nu- cleus. Exp. Neurol. 61:53-64; 1978.
34. Spencer, D. D.; Robbins, R. J.; Naftolin, F.; Marek, K. L.; Voll- mer, T.; Leranth, C.; Roth, R. H.; Price, L. H.; Gjedde, A.; Bun- hey, B. S.; Sass, K. J.; Elsworth, J. D.; Leon Kier, E.; Makuch, R.; Hoffer, P. B.; Redmond, D. E. Unilateral transplantation of human fetal mesencephalic tissue into the caudate nucleus of pa- tients with Parkinson's disease. N. Engl. J. Med. 327:1541- 1548; 1992.
35. StrOmberg, I.; Bygdeman, M.; Goldstein, M.; Seiger, A.; Olson, L. Human fetal substantia nigra grafted to the dopamine-denervated striatum of immunosuppressed rats: Evidence for functional rein- nervation. Neurosci. Lett. 71:271-276; 1986.
36. Wictorin, K.; Brundin, P.; Gustavii, B.; Lindvall, O.; Bjrrklund, A. Reformation of long axon pathways in adult rat central ner- vous system by human forebrain neuroblasts. Nature 347:556- 558; 1990.
37. Widner, H.; Tetrud, J.; Rehncrona, S.; Snow, B.; Brundin, P.; Gus- tavii, G.; Bjrrklund, A.; Lindvall, O.; Langston W. Bilateral fetal mesencephalic grafting in two patients with parkinsonism induced by 1-Methyl-4-Phenyl-l,2,3,6-Tetrahydropyridine (MPTP). N. Engl. J. Med. 327:1556-1563; 1992.
This article is being published without the benefit of the author 's review of the proofs, which were not available at press time.