
Occurrence and Structural Analysis of Highly Sulfated Multiantennary N-linked Glycan Chains Derived...
11
Eur. J. Biochem. 238, 357-367 (1996) 0 FEBS 1996 Occurrence and structural analysis of highly sulfated multiantennary N-linked glycan chains derived from a fertilization-associated carbohydrate-rich glycoprotein in unfertilized eggs of Tribolodon hakonensis Tomohiko TAGUCHI', Mariko IWASAKI', Yutaka MUTO', Ken KITAJIMA I, Sadako INOUE2, Kay-Hooi KHOO', Howard R. MORRIS', Anne DELL' and Yasuo INOUE' ' Department of Biophysics and Biochemistry, Graduate School of Science, University of Tokyo, Japan ' Department of Biochemistry, Imperial College of Science, Technology and Medicine, London, UK (Received 29 JanuaryR7 March 1996) ~ EJB 96 0115/5 School of Pharmaceutical Sciences, Showa University, Tokyo, Japan This study represents the first detailed investigation of the nature of highly sulfated (keratan-sulfate- like) complex-type asparagine-linked glycans having a tetraantennary core structure and shows the effec- tiveness of fast-atom-bombardment mass spectrometric (FAB-MS) methods incorporating derivatization and mild methanolysis for analyzing such complex types of sulfated glycans. The structure of the N-glycan chains was unambiguously established by a combination of composi- tional analysis, methylation analysis, mild methanolysis for desulfation, hydrazinolysis/nitrous acid deamination, enzymatic (endo-P-galactosidase and peptide : N-gl ycosidase F) digestions, and instrumental analyses ('H-NMR spectroscopy and FAB-MS) which revealed the novel repeating sulfated carbohydrate sequences, 2 GalPl +4Gal,81[+4(HS03~6)GlcNAc~1-+3( -C GalPl--+4)Gal~l],-+ (see Structure I; p+q+r+s-14). This sequence is unique in: (a) the skeletal structure is similar to that of keratan sulfate but is completely devoid of 6-0-sulfated Gal residues and (b) the presence of branched Gal residues in the sequence -+4GlcNAc~1+3(Gal,81- 4)GulPl+. +SO;b6 +.Gav-l14 +SO,-b6 \ +Gal/ll+4Ga1~1[+4GlcNAc~1+3Ga~1],+4GlcNAC@l 2Manal 71 L4 2Manal +SO;b6 +Gal/llL4 +SO+ +SO<, +Gav1k4 +SO,-b6 6Ma~l-+4GlcNAc/?l--4GlcNAC@l+Asn +Galp1+4Gal~l[--4GlcNAc~1-+3Galp1],+4GlcNAC@1 Structure I Keywords: highly sulfated N-linked glycan ; keratan-sulfate-like N-linked glycan ; hyosophorin ; tetraan- tennary glycan ; fast-atom-bombardment mass spectrometry. I +Ga~1+4Gal~1[-.4GlcNAc~1+3Ga~l]r+4GlcNA~~l +SO<,6 +Gal/llcq +S03k6 +Galp1~4Ga1~1[+4GlcNAc~l+3Ga~l],+4GlcNAc~1 f The presence of sulfate groups in N-linked oligosaccharides has been reported in several glycoproteins including Dictyostel- ium discoideum lysosomal enzymes [I, 21, human urokinase [3], viral glycoproteins [4], hen egg albumin [S], zona pellucidae Correspondence to Y. hue, Institute of Biological Chemistry Aca- demia Sinica, Taipei I1 5, Taiwan; K. Kitajima, Department of Biophys- ics and Biochemistry, Graduate School of Science, University of Tokyo, Hongo-7, Tokyo, Japan 113 Fuxt +8862 788 9759 (Y. Inoue); +81 3 5684 2394 (K. Kitajima). Abbreviations. FAB, fast atom bombardment; NeuSAc, N-acetylneu- raminic acid; (2,5)-anhMan-ol, 2,5-anhydro-o-mannitol; Gal-ol, D-ga- lactitol ; anhHex-ol, anhydro hexitol ; lD, one-dimensional ; 2D, two-di- mensional. Enzymes. Endo-,8-galactosidase (EC 3.2.1.103); peptide:N-glycosi- dase F (EC 3.5.1.52). glycoproteins [6, 71, thyroid plasma membrane glycoproteins [S], thyroglobulin [9-111, some mammalian cell lines 112-151, and pituitary hormones [16-201. The oligosaccharide chains of keratan sulfate are bound to protein either by an N-glycosidic linkage (type I, as found in cornea) [21-241 or an 0-glycosidic linkage (type 11, associated with skin, cartilage, and bone) [25]. In the type I keratan sulfate derived from cornea, the core structures are restricted to dianten- nary [22-241. The existence of tri-or tetra-antennary complex- type core structures carrying glycosaminoglycan chains has been reported in some mammalian cells [ I41 and virus-transformed baby hamster kidney cells [26], but these reports were mostly based on the results of Iectin-affinity chromatography using radioisotopes and the complete structures of these N-linked gly- cans have not yet been precisely determined.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Occurrence and Structural Analysis of Highly Sulfated Multiantennary N-linked Glycan Chains Derived...

Eur. J. Biochem. 238, 357-367 (1996) 0 FEBS 1996
Occurrence and structural analysis of highly sulfated multiantennary N-linked glycan chains derived from a fertilization-associated carbohydrate-rich glycoprotein in unfertilized eggs of Tribolodon hakonensis Tomohiko TAGUCHI', Mariko IWASAKI', Yutaka MUTO', Ken KITAJIMA I, Sadako INOUE2, Kay-Hooi KHOO', Howard R. MORRIS', Anne DELL' and Yasuo INOUE'
' Department of Biophysics and Biochemistry, Graduate School of Science, University of Tokyo, Japan
' Department of Biochemistry, Imperial College of Science, Technology and Medicine, London, UK
(Received 29 JanuaryR7 March 1996) ~ EJB 96 0115/5
School of Pharmaceutical Sciences, Showa University, Tokyo, Japan
This study represents the first detailed investigation of the nature of highly sulfated (keratan-sulfate- like) complex-type asparagine-linked glycans having a tetraantennary core structure and shows the effec- tiveness of fast-atom-bombardment mass spectrometric (FAB-MS) methods incorporating derivatization and mild methanolysis for analyzing such complex types of sulfated glycans.
The structure of the N-glycan chains was unambiguously established by a combination of composi- tional analysis, methylation analysis, mild methanolysis for desulfation, hydrazinolysis/nitrous acid deamination, enzymatic (endo-P-galactosidase and peptide : N-gl ycosidase F) digestions, and instrumental analyses ('H-NMR spectroscopy and FAB-MS) which revealed the novel repeating sulfated carbohydrate sequences, 2 GalPl +4Gal,81[+4(HS03~6)GlcNAc~1-+3( -C GalPl--+4)Gal~l],-+ (see Structure I ; p+q+r+s-14). This sequence is unique in: (a) the skeletal structure is similar to that of keratan sulfate but is completely devoid of 6-0-sulfated Gal residues and (b) the presence of branched Gal residues in the sequence -+4GlcNAc~1+3(Gal,81- 4)GulPl+.
+SO;b6 +.Gav-l14 +SO,-b6
\ +Gal/ll+4Ga1~1[+4GlcNAc~1+3Ga~1],+4GlcNAC@l
2Manal 7 1 L4 2Manal
+SO;b6 +Gal/llL4 +SO+
+SO<, +Gav1k4 +SO,-b6 6Ma~l-+4GlcNAc/?l--4GlcNAC@l+Asn
+Galp1+4Gal~l[--4GlcNAc~1-+3Galp1],+4GlcNAC@1
Structure I
Keywords: highly sulfated N-linked glycan ; keratan-sulfate-like N-linked glycan ; hyosophorin ; tetraan- tennary glycan ; fast-atom-bombardment mass spectrometry.
I +Ga~1+4Gal~1[-.4GlcNAc~1+3Ga~l]r+4GlcNA~~l
+SO<,6 +Gal/llcq + S 0 3 k 6 +Galp1~4Ga1~1[+4GlcNAc~l+3Ga~l],+4GlcNAc~1 f
The presence of sulfate groups in N-linked oligosaccharides has been reported in several glycoproteins including Dictyostel- ium discoideum lysosomal enzymes [I , 21, human urokinase [3], viral glycoproteins [4], hen egg albumin [S], zona pellucidae
Correspondence to Y. h u e , Institute of Biological Chemistry Aca- demia Sinica, Taipei I1 5, Taiwan; K. Kitajima, Department of Biophys- ics and Biochemistry, Graduate School of Science, University of Tokyo, Hongo-7, Tokyo, Japan 113
Fuxt +8862 788 9759 (Y. Inoue); +81 3 5684 2394 (K. Kitajima). Abbreviations. FAB, fast atom bombardment; NeuSAc, N-acetylneu-
raminic acid; (2,5)-anhMan-ol, 2,5-anhydro-o-mannitol; Gal-ol, D-ga- lactitol ; anhHex-ol, anhydro hexitol ; lD, one-dimensional ; 2D, two-di- mensional.
Enzymes. Endo-,8-galactosidase (EC 3.2.1.103); peptide:N-glycosi- dase F (EC 3.5.1.52).
glycoproteins [6, 71, thyroid plasma membrane glycoproteins [S], thyroglobulin [9-111, some mammalian cell lines 112-151, and pituitary hormones [16-201.
The oligosaccharide chains of keratan sulfate are bound to protein either by an N-glycosidic linkage (type I, as found in cornea) [21-241 or an 0-glycosidic linkage (type 11, associated with skin, cartilage, and bone) [25]. In the type I keratan sulfate derived from cornea, the core structures are restricted to dianten- nary [22-241. The existence of tri-or tetra-antennary complex- type core structures carrying glycosaminoglycan chains has been reported in some mammalian cells [ I41 and virus-transformed baby hamster kidney cells [26], but these reports were mostly based on the results of Iectin-affinity chromatography using radioisotopes and the complete structures of these N-linked gly- cans have not yet been precisely determined.

358 Taguchi et al. (Eur: 1. Biochem. 238)
In the course of studying the accumulation of free glycans in unfertilized eggs of Tribolodon haknnensis (a dace), we have discovered a highly acidic glycoprotein, hyosophorin, identified as fertilization-associated [27]. In this study, we have determined the carbohydrate structure of the hyosophorin molecule (molecu- lar mass about 10kDa) derived from these fish eggs, using compositional analysis, methylation analysis, mild methanolysis, digestion with endo-ggalactosidase, hydrazinolysis/nitrous acid deamination, 'H-NMR spectroscopy, and FAB-MS. The results thus obtained showed for the first time the occurrence of highly sulfated huge multiantennary N-linked glycan chains in nature, the backbone of which is similar to that of keratan sulfate but is completely devoid of 6-0-sulfated Gal residues. Substitution by P-Gal residue on C4 of the Gal residues in the poly(iV-acetyllac- tosamine) structure is observed. Analogous branched Gal struc- tures have been found in other hyosophorin molecules [28, 291. This study also shows the effectiveness of FAB-MS methods, including derivatization and desulfation of permethylated gly- cans previously demonstrated by our group [30] for the analysis of highly sulfated large oligosaccharides.
MATERIALS AND METHODS
Chemical analyses. The hexose content was estimated by the phenol/sulfuric acid method 1311. Free and bound sialic acid were quantified or monitored by the thiobarbituric acid method 132, 331 and the resorcinol method 1341, respectively. The molar ratios of component monosaccharides of oligosaccharides and glycoproteins were determined by GLC as previously reported [35]. The sulfate content was analyzed by HPLC on a TSK-gel IC-Anion PW column using a Jasco HPLC system equipped with 980-PU pump, a 970-UV detector, and a Rheodyne injector with a 20.~1 sample loop. The elution pattern was monitored at 265 nm and recorded with a Chromatocorder 12 (System Instru- ments, Tokyo). Samples were evaporated to dryness after hy- drolysis in 6 M HC1 at 110°C for 24 h, dissolved in 30-50 pl water. Samples (1 -10 pl) were injected and eluted isocratically with acetonitrile/l -butanol (4: 1 ) containing 0.5 mM sodium phthalate, 5.82 mM H,BO,, 1.3 mM Na,B,O,, 0.9 mM sodium gluconate at the flow rate of 1.2 ml/min.
Isolation and purification of heavily sulfated hyosopho- rin. The glycoprotein fractions (U5 -U8) [27] were chromato- graphed on Sephacryl S-200 column (1.9 X 176 cm, equilibrated and eluted with 0.1 M NaCl, fraction volume 4.0 ml), respective- ly. The eluant was monitored by absorbance at 230 nm, the thio- barbituric acid method [32, 331 or the resorcinol method [34] for sialic acid, and the phenol/sulfuric acid method [31] for hexose. The phenol/sulfuric acid-positive fraction was collected and desalted by dialysis against distilled water.
Hydrazinolysislnitrous acid deamination. A freeze-dried sample of Ugui egg hyosophorin (1.8 mg as Gal; see Results) was reacted with 2 ml anhydrous hydrazine in the presence of 10 mg hydrazinium sulfate in a commercial hydrazinolysis appa- ratus, Hydraclub (HBnen Corp.), at 110°C for 24 h [28]. After cooling to room temperature, the reaction mixture was dried overnight in a vacuum desiccator over concentrated sulfuric acid. 25 mg sodium nitrite and 2 ml 0.5 M acetic acid were then added. After reaction at room temperature for 1 h, a further 10 mg NaNO, was added and deamination was continued over- night. Ethylamine (70%) was then added to destroy excess nit- rite, and the solution was treated with 10 mg NaBH, at room temperature for 10 h. The pH was adjusted to 5-6 with 1 M acetic acid, and the reaction products were applied to a 15-ml column of DEAE-Sephadex A-25 (HCO; form, equilibrated and eluted with H20). The flow-through fractions were collected and
chromatographed on a column (1.3 X 1 10 cm, 50 mM acetic acid, fraction volume 1.2 ml) of Bio-Gel P-2. The eluant was moni- tored for neutral sugar by the phenol/sulfuric acid method and the positive fractions were collected and analyzed. The DEAE- Sephadex A-25 column was eluted by a linear gradient of 0- 0.4 M NH,HCO, (1 00 ml/l00 ml ; fraction volume 1.7 ml) and the elution profile was monitored by the phenol/sulfuric acid method. The positive fractions were collected, freeze dried to remove NH,HCO, and further purified by preparative TLC.
Preparative TLC. N-1 and A-1 fractions (see Results) were subjected to preparative silica gel TLC (Kiesel Gel 60, Merck; solvent, l-propanol/25 % NH,OH/water, 6: 1 :2.5, by vol., devel- oped for 12 h). A lane from the TLC plate was cut off for monitoring by visualization with 1 % orcinol/50% sulfuric acid reagent (llO°C, 30 min). The materials were eluted from the silica gel plate with 20% ethanol and subjected to Sephadex G- 25 gel chromatography to remove silica gel.
Analytical TLC. Approximately 1 - 10-pg samples were spotted on a silica gel TLC plate and developed under the same conditions used for preparative TLC.
Desulfation. Desulfation for dried samples (10-200 kg as Gal) was performed in 0.5-2.0 ml 0.5 M anhydrous methanol/ HCI at 25°C for 6 h or in 0.05 M anhydrous methanol/HCl at 4°C for 24 h. After the reaction, the samples were dried under vacuum.
Exosialidase digestion of Ugui egg hyosophorin. A sample of the hyosophorin (about 10mg as Gal) was digested with 0.25 U Arthrobacter ureafaciens exosialidase (Nacalai Co., Kyoto) in 2.5 ml 50 mM sodium acetate pH 5.5 at 37°C under slight toluene. After 24 h of incubation, an additional 0.1 U exo- sialidase was added and further incubated for 24 h. Asialo hyo- sophorin was separated from free sialic acid (NeuSAc) by pas- sage of the enzyme digest through a Sephadex G-25 column (1.3 X 105 cm, equilibrated and eluted with 5 % ethanol).
Liberation of glycan chains from hyosophorin by pep- tide :N-glycosidase F. Asialo hyosophorin (about 3 mg as Gal) was incubated with 5 mU peptide:N-glycosidase F (Takara Co., Kyoto) in 2 ml 0.25 M sodium phosphate pH 8.6 under slight toluene for 24 h. Another 5 mU of the enzyme was added and the mixture was incubated for another 24 h. The digest was then applied to a column of Sephadex G-50 (1.8 X 60 cm, equilibrated and eluted with 0.1 M NaCI) and the eluant was monitored by the phenol/sulfuric acid method. The positive fraction (free glycan) was pooled and desalted by Sephadex G-25 column chromatography (1.3 X 105 cm, equilibrated and eluted with 5 LTC ethanol).
Methylation analysis. Oligosaccharides were permethylated by the procedure of either Hakomori [36] for sulfate-containing glycan chains or Ciucanu and Kerek 1371 for neutral ones. Permethylated sulfated glycans were purified with a C18 Sep- Pak cartridge (Waters Ltd) 1301 or Sephadex G-25 column chro- matography (2.2X40 cm, equilibrated and eluted with 10% methanol). Permethylated glycan chains were converted to mix- tures of partially methylated alditol acetates and hexosaminitol acetates according to Levery and Hakomori [38]. Subsequent GLC analyses were performed with the apparatus equipped with a capillary column (CBJS, 30 mX0.32 mm, Shimadzu) at 180- 260°C at 2"C/min.
'H-NMR measurement. The material was treated repeat- edly with 'H,O (99.8 %), with Iyophilization between treatments, finally dissolved in *H,O (99.9%), and subjected to NMR spectroscopy. 1D and 2D 500-MHz 'H-NMR spectra were ob- tained using a Bruker AMX-500 NMR spectrometer at 298 K. For the TOCSY spectrum, a MLEV-17 mixing sequence of 100 ms was used. Proton chemical shifts are expressed relative

Taguchi et al. (Em: J. Biochem. 238) 359
to the methyl proton signal of sodium 3-trimethylsilyl-(2,2,3,3- 'H,)propionate set equal to 0.00 ppm.
Digestion with endo-P-galactosidase. Ugui egg hyosopho- rin (1.2 mg as Gal) was incubated with 0.1 U Escherichiafreun- dii endo-P-galactosidase (Seikagaku Kogyo Co. Ltd, Tokyo) dis- solved in the total volume of 800 pl 100 mM sodium acetate pH 5.5 for 60 h at 37°C under toluene. The reaction mixture was applied to a column ( 1 3 x 6 0 cm, equilibrated and eluted with 0.1 M NaC1, fraction volume 2.0 ml) of Sephadex G-50. The eluant was monitored by the phenolhlfuric acid method.
Alkaline borohydride reduction. Fraction U-p2 (50 pg as Gal; see Results) was reduced in 500 p1 50 mM sodium phos- phate pH 8.0 containing 5 mg NaBH,. The mixture was left at 25 "C for 3 h. The reaction was terminated by addition of glacial acetic acid and the reduced material was desalted by Sephadex G-25 column chromatography (1.3 X 105 cm, equilibrated and eluted with 5 % ethanol).
Perdeuterioacetylation. Base-catalyzed peracetylation was performed using 1-methylimidazole/(2H,)acetic anhydride (1 : 5 , by vol., 30 pl total volume) at room temperature for 1 h. The reagents were dried down under nitrogen.
FAB-MS analysis. FAB-mass spectra were obtained using a VG Analytical ZAB-2SE 2FPD mass spectrometer fitted with a cesium ion gun operated at 30 kV (positive mode) or 20 kV (negative mode). Data were acquired and processed using the VG Analytical OpusQ software. For time-course rnethanolysis, 20 p1 ~ 0 . 1 M methanolic/HCl (prepared by bubbling HC1 gas into 1 ml methanol) was added to the Hakomori methylated sam- ple and 1-p1 aliquots were withdrawn at suitable times prior to and after heating at 60°C and loaded directly onto the probe for FAB-MS analyses [30]. Other derivatized samples were dis- solved in methanol for aliquoting into the matrix. Monothioglyc- erol was used as matrix in both positive and negative ion modes.
RESULTS
Purification of hyosophorin from the unfertilized eggs of T. hakonensis. The glycoprotein fractions (U5 -US) [27], which had previously been separated by DEAE-Sephadex A-25 anion- exchange chromatography of the phenol-treated extract obtained from the unfertilized eggs of 7: hakonensis, were further purified by gel filtration on a Sephacryl S-200 column, respectively. Each chromatography on Sephacryl S-200 showed a single phenol1 sulfuric-acid-positive peak eluted in fractions 46-60, and as- signed as hyosophorin. These compounds showed no differences in carbohydrate compositions including sialic acid residues, and had a high content of sulfate group which accounts for their differences in ionic properties. These purified compounds were then pooled and subjected to structural analysis.
Chemical composition of hyosophorin. The intact hyosophorin contained Man, Gal, GlcNAc, NeuSAc, and sulfate in a molar ratio of 3.0:24:20:2.1: 10. The sialic acid residues were exclusively Neu5Ac. The high content of Gal and GlcNAc resi- dues compared to Man suggests that it has poly(N-acetyllacto- samine) sequences. It should be pointed out here that the major negative charge was based on the sulfate group not on sialic acid residues.
Methylation analysis of the desulfated hyosophorin glycan. Desulfated hyosophorin glycan was prepared by the HCI/ CH,OH treatment of free glycan derived from asialo hyosopho- rin and subjected to methylation analysis (Fig. 1). A number of partially methylated alditol and hexosaminitol acetates was iden- tified and quantified using GLC by comparison of their relative
1
4
2
3 8 I I
I I I I I 5 10 15 20 25
retention time (min)
Fig. 1. Gas-liquid chromatogram of partially methylated alditol ace- tates originating from desulfated Ugui egg hyosophorin glycan. The following peaks were identified as alditol acetates of (1) 2,3,4,6-tetra- 0-methyl Gal; (2) 2,4,6-tri-O-methyl-Gal; (3) 2,h-di-O-methyl-Gal ; (4) 3,6-di-O-rnethyl-Man; ( 5 ) 3,4-di-O-methyl-Man; (6) 2,4-di-0- methyl-Man ; (7) 3,4,6-tri-O-methyI GlcN(Me)Ac ; (8) 3,6-di-O-methyl GlcN(Me)Ac.
retention times with those determined for authentic samples [28, 291. Data for the desulfated glycan showed the presence of 2,3,4,6-tetra-O-Me-Gal, 2,4,6-tri-O-Me-Gal, 2,6-di-O-Me-Gal, 3,6-di-O-Me-Man, 3,4-di-O-Me-Man, 2,4-di-O-Me-Man, 3,4,6- tri-0-Me-GlcN(Me)Ac and 3,6-di-O-Me-GlcN(Me)Ac. These results indicated the presence of terminal Gal, 3-0-substituted Gal, 3,4-di-O-substituted Gal, 2,4-di-O-substituted Man, 2,6-di- 0-substituted Man, 3,6-di-O-substituted Man, terminal GlcNAc and 4-0-substituted GlcNAc residues. The presence of 2,4-di- 0-substituted Man, 2,6-di-O-substituted Man, and 3,6-di-O-sub- stituted Man (2,4,6-tri-O-substituted Man and 3,4,6-tri-O-substi- tuted Man were not detected) in a ratio of nearly 1 : 1 : 1 strongly indicated a tetraantennary core glycan structure for the glycan, which was in good agreement with the evidence from 'H-NMR spectroscopy as described below. Considering that terminal Gal (about 7 mol/mol) is more abundant than the expected value (4 mol/mol) for a non-branched tetraantennary glycan structure and the presence of 3,4-di-O-substituted Gal (about 4 mol), the hyosophorin glycan was expected to have the branched N-acet- yllactosamine sequence, GlcNAcl-, 3(Gall-+4)Gall- 4Glc- NAcl-. This branched structure was confirmed by analyzing the products of hydrazinolysis/nitrous acid deamination and free oligosaccharides released by endo-P-galactosidase digestion (see below).
'H-NMR spectroscopy. The 'H-NMR spectrum of the hyoso- phorin was used to obtain information about the antenna struc- ture and the anomeric configurations (spectrum not shown). By comparison with the previously reported data [28, 39-41 J , the signals appearing at 4.86 pprn and 5.13 ppm were assignable to H1 of Man-4' and Man-4 of a tetraantennary structure, respec- tively, which was consistent with the conclusion from methyl- ation analysis (see above). The signal at 5.03 pprn was assign- able to H1 of the GlcNAc-I residue linked to Asn residue. No other signals were observed in the 4.8-5.2-ppm region indicat- ing that the anomeric configurations of all of the Gal and GlcNAc residues in the hyosophorin glycan are p.
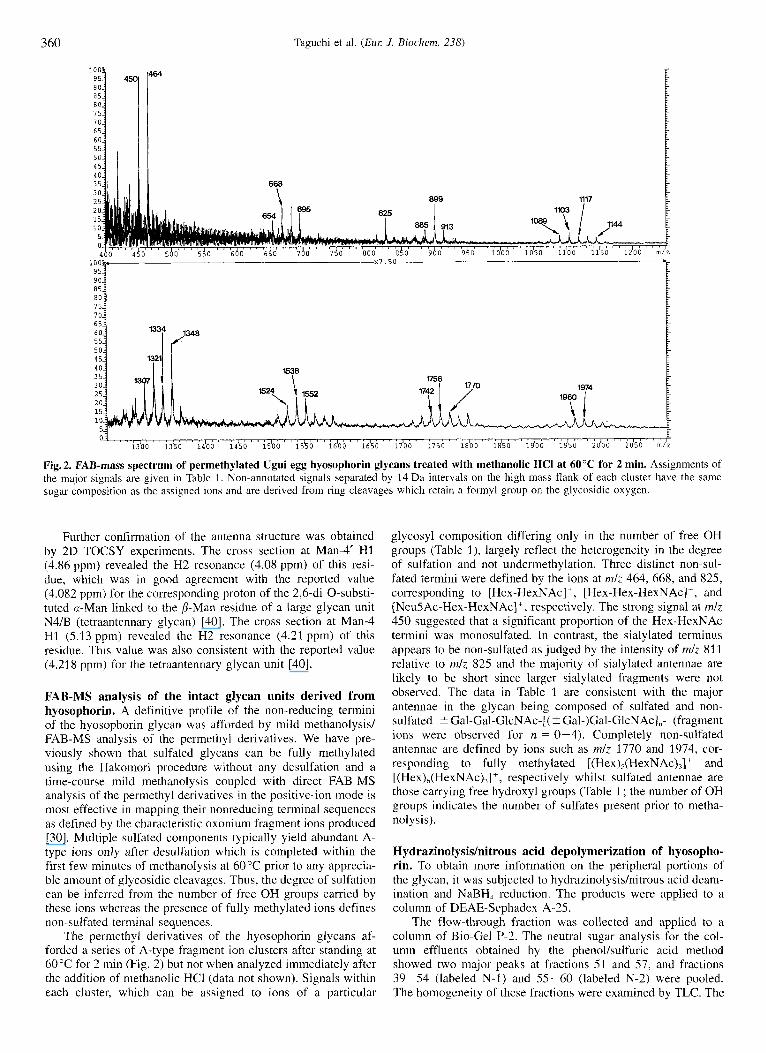
360 Taguchi et al. ( E m J. Biochem. 238)
t
13'00 13'50 14'00 1250 15'00 1250 16100 16150 17'00 17'50 18 '00 18'50 1dOO 19150 20100 20150 ml;
Fig.2. FAB-mass spectrum of permethylated Ugui egg hyosophorin glycans treated with methanolic HCI at 60°C for 2 min. Assignments of the major signals are given in Table 1. Non-annotated signals separated by 14 Da intervals on the high mass flank of each cluster have the same sugar composition as the assigned ions and are derived from ring cleavages which retain a formyl group on the glycosidic oxygen.
Further confirmation of the antenna structure was obtained by 2D TOCSY experiments. The cross section at Man-4' H1 (4.86 ppm) revealed the H2 resonance (4.08 ppm) of this resi- due, which was in good agreement with the reported value (4.082 ppm) for the correspondlng proton of the 2,6-di-O-substi- tuted a-Man linked to the P-Man residue of a large glycan unit N4/B (tetraantennary glycan) [40]. The cross section at Man-4 HI (5.13 ppm) revealed the H2 resonance (4.21 ppm) of this residue. This value was also consistent with the reported value (4.218 ppm) for the tetraantennary glycan unit [40].
FAB-MS analysis of the intact glycan units derived from hyosophorin. A definitive profile of the non-reducing termini of the hyosophorin glycan was afforded by mild methanolysisl FAB-MS analysis of the permethyl derivatives. We have pre- viously shown that sulfated glycans can be fully methylated using the Hakomori procedure without any desulfation and a time-course mild methanolysis coupled with direct FAB-MS analysis of the permethyl derivatives in the positive-ion mode is most effective in mapping their nonreducing terminal sequences as defined by the characteristic oxonium fragment ions produced [30]. Multiple sulfated components typically yield abundant A- type ions only after desulfation which is completed within the first few minutes of methanolysis at 60°C prior to any apprecia- ble amount of glycosidic cleavages. Thus, the degree of sulfation can be inferred from the number of free OH groups carried by these ions whereas the presence of fully methylated ions defines non-sulfated terminal sequences.
The permethyl derivatives of the hyosophorin glycans af- forded a series of A-type fragment ion clusters after standing at 60°C for 2 min (Fig. 2) but not when analyzed immediately after the addition of methanolic HCI (data not shown). Signals within each cluster, which can be assigned to ions of a particular
glycosyl composition differing only in the number of free OH groups (Table I ) , largely reflect the heterogeneity in the degree of sulfation and not undermethylation. Three distinct non-sul- fated termini were defined by the ions at nzlz 464, 668, and 825, corresponding to [Hex-HexNAc]', [Hex-Hex-HexNAc]', and [NeuSAc-Hex-HexNAc] + , respectively. The strong signal at mlz 450 suggested that a significant proportion of the Hex-HexNAc termini was monosulfated. In contrast, the sialylated terminus appears to be non-sulfated as judged by the intensity of mlz 811 relative to mlz 825 and the majority of sialylated antennae are likely to be short since larger sialylated fragments were not observed. The data in Table 1 are consistent with the major antennae in the glycan being composed of sulfated and non- sulfated -C Gal-Gal-GlcNAc-[( 5 Gal-)Gal-GlcNAc1,- (fragment ions were observed for n = 0-4). Completely non-sulfated antennae are defined by ions such as mlz 1770 and 1974, cor- responding to fully methylated [(Hex),(HexNAc),]+ and [(Hex),(HexNAc),]+, respectively whilst sulfated antennae are those carrying free hydroxyl groups (Table 1 ; the number of OH groups indicates the number of sulfates present prior to metha- nol ysis).
Hydrazinolysishitrous acid depolymerization of hyosopho- rin. To obtain more information on the peripheral portions of the glycan, it was subjected to hydrazinolysis/nitrous acid deam- ination and NaBH, reduction. The products were applied to a column of DEAE-Sephadex A-25.
The flow-through fraction was collected and applied to a column of Bio-Gel P-2. The neutral sugar analysis for the col- umn effluents obtained by the phenol/sulfuric acid method showed two major peaks at fractions 51 and 57, and fractions 39-54 (labeled N-1) and 55-60 (labeled N-2) were pooled. The homogeneity of these fractions were examined by TLC. The

Taguchi et al. (Eur: J. Biochem. 238) 361
Table 1. Assignments of the major signals observed in the FAB-MS spectrum (Fig. 2) of permethylated Ugui egg hyosophorin glycans after mild methanolysis. Signals were detected up to mlz 3000, indicating the presence of at least up to five ( 2 Gal-)Gal-GlcNAc repeating units. For simplicity, only the dominant signals corresponding to ions containing S 3 repeating units are tabulated. The most prominent signal within each cluster is in bold print.
Signal cluster Composition Probable sequence for the non-sulfated component
464 450 668 654 695 825 913 899 885 1117 1103 1089 1144" 1321 1307 1348 1334 1552 1538 1524 1770 1756 1742 1974 1960
Hex,HexNAc: (OH),Hex,HexNAc; Hex,HexNAc: (OH), Hex,HexNAc: (OH), Hex I HexNAc,' Neu Ac, Hex I HexNAc: Hex H e x N A c 2' (OH) ,Hex,HexNAc: (OH),Hex,HexNAc; Hex,HexNAc,' (OH),Hex,HexNAc: (OH),Hex,HexN Ac: (OH),Hex,HexNAc: Hex,HexNAc,' (OH),Hex,HexNAc: (OH),Hex,HexNAc: (OH),Hex,HexNAc: (OH),Hex,HexNAc: (OH),Hex,HexNAc: (OH),Hex,HexNAc: Hex,HexNAc: (OH), Hex,HexNAc: (OH),Hex,HexNAc: Hex,HexNAc: (OH) ,Hex,HexNAc:
Gal-GlcNAc-
Gal-Gal-GlcNAc-
NeuAc-Gal-GlcNAc- Gal-GlcNAc-Gal-GlcNAc-
Gal-Gal-GlcNAc-Gal-GlcNAc or Gal-GlcNAc-(Gal-)Gal-GlcNAc
Gal-Gal-GlcNAc-(Gal-)Gal-GlcNAc
[Gal-GlcN Ac-I,
[Gal-GlcNAc] 2- [(Gal-)Gal-GlcNAc] I
[Gal-GlcNAc] , -[(Gal-)Gal-GlcNAc-1,
[(Gal-)Gal-GlcNAc]
" At early times, fully methylated species were detected for each composition except for those corresponding to Hex,HexNAc2 and Hex,Hex- NAc, which probably resulted from facile secondary cleavages.
Table 2. Carbohydrate compositions of the products obtained by hy- drazinolysis/nitrous acid depolymerization. Values for Gal are given as molar ratios relative to (2,5)-anhMan-ol set equal to 1 .O.
Fraction Gal (2,5)-anhMan-ol
N- 1 1.9 1 .o N-2 0.82 1 .o A- 1 1.9 1 .o
1 2 3 4 5
Fig. 3. TLC analysis of the products derived from hydrazinolysis/ nitrous acid depolymerization. Lane 1 , Gal and lactose; lane 2, N-2; lane 3, N-1 ; lane 4, A-1 ; lane 5, desulfated A-I. Arrow indicates the origin. The upper and the lower spots in lane 1 indicated Gal and lactose, respectively.
N-1 fraction gave one major spot and 2-3 very minor spots with low migration rates. The major component in the N-I frac- tion was purified by preparative TLC (Fig. 3) and renamed as N-1. The N-2 fraction was revealed to be highly pure by TLC (Fig. 3) and used for structural analysis. The results of carbohy- drate composition analysis and methylation analysis are shown in Tables 2 and 3. N-1 was found to have a sugar composition of 2,5-anhydro-~-mannitol [(2,5)-anhMan-ol] and Gal in a ratio of 1 .O: 1.9 and to give 4-0-substituted (2,5)-anhMan-ol, terminal
Gal and 4-0-substituted Gal in 1 .O: 1.0: 1.2 ratio on methylation analysis. The N-2 fraction contained (2,5)-anhMan-ol and Gal in a molar ratio of 1.0:0.82 and was revealed to have 4-0-substi- tuted (2,5)-anhMan-ol and terminal Gal in a ratio of 1 .O: 1 .O on methylation analysis. The 'H-NMR spectrum of hyosophorin showed that the anomeric configuration of all of the Gal residues of these fractions is p (see above). The structure of N-l was thus determined to be Galpl+4Galj?l+4(2,5)-anhMan-ol and that of N-2 to be GaljT1-4(2,5)-anhMan-ol.
Phenolkulfuric acid analysis for the fractions of DEAE- Sephadex A-25 anion-exchange chromatography gave a broad peak (profile not shown), and the fractions 31 -50 (labeled A- 1) and 51-80 (labeled A-2) were collected. No other phenol/ sulfuric-acid-positive fraction was observed. The homogeneity of these fractions was examined by TLC. The A-1 fraction gave one major spot (total amount of this material was about 200 pg as Gal) and very minor spots. The major component in the A-l fraction was purified by preparative TLC (Fig. 3) and renamed as A-1. This was found to have a carbohydrate composition of (2,5)-anhMan-ol and Gal in a ratio of 1.0: 1.9 and to give 4,6-di- 0-substituted (2,5)-anhMan-ol, terminal Gal and 4-0-substituted

362 Taguchi et al. (Eul: J . Biochem. 238)
Table 3. Methylation analyses of the products obtained by hydrazi- nolysislnitrous acid depolymerization. For N-1, N-2, and desulfated A-1, 4-0-substituted (2,5)-anhMan-ol is set equal to 1.0. For A-1, 4,6- di-0-substituted (2,5)-anhMan-ol is set equal to 1 .O.
Methylated product Amount in
N-1 N-2 A-1 desul- fated A-1
4-0-substituted (2,5)-anhMan-ol 1.0 1.0 - 1.0 4,6-Di-O-substituted (2,5)-anhMan-ol ~ - 1.0 -
Terminal Gal 1.0 1.0 1.2 1.2 4-0-substituted Gal 1.2 - 0.8 1.2
[21 t11 Gaip1-4Gaipl-.4(SO<-.6)-(2,5)anhMan-ol
i
I I I I I I I I , I I ! l )
4.5 4.0 3.5
chemical shift (ppm)
Fig.4. The 500-MHz 'H-NMR spectrum of A-1 in 'H,O at 25OC. The residue numberings are given in the inserted structures.
Gal in nearly equal ratio on methylation analysis (Tables 2 and 3). To determine the substitution position of the sulfate group, A-1 was subjected to desulfation (0.5 M anhydrous methanol/ HCI, 25 "C, 6 h). In this treatment, no glycosidic cleavage was observed and the migration position of desulfated A-1 was completely identical to that of N-1 [GalPl-4GaI~l-4(2,5)- anhMan-ol] (Fig. 3). Methylation analysis of desulfated A-1 gave 4-0-substituted (2,5)-anhMan-ol, terminal Gal and 4-0- substituted Gal in a molar ratio of 1.0: 1.2: 1.2 (Table 3). From these results, A-1 was concluded to have the structure GalPI- 4Gal~1-4(HSO,-6)-(2,5)-anhMan-o1. In addition, the sulfate group in A-l was partially released under the following condi- tions (0.05 M anhydrous methanoliHC1, 4"C, 24 h). The 'H- NMR spectrum of A-I gave supporting evidence for the above structure (shown in Fig. 4). The spectrum showed two ,!-anom- eric proton resonances at 4.59 ppm and 4.51 ppm. The resonance at 4.51 ppm was assignable to HI of the penultimate ,!-Gal resi- due if we considered that (a) the resonance of H1 (4.52 ppm) of the P-Gal residue in the GalPl-4(HSO,-6)GlcNAc,!l-+ struc- ture was downfield shifted by 0.04 ppm compared to that (4.48 ppm) of Gal~l-4GlcNAc~l- sequence [42] and (b) the resonance of H1 of the penultimate P-Gal residue in the Gal~l-4Gal~l-4(2,5)-anhMan-o1 structure was known to appear at 4.47 ppm [43]. Moreover, it is noteworthy that the
Table 4. Proton chemical shift values of the products obtained by hydrazinolysishitrous acid depolymerization. HN2 = Gal/Il+ 4Gal/I1+4(2,5)-anhMan-ol. This data was reported by Taguchi et al. [43].
P-Gal residue Proton Chemical shift in
A-I HN2
PPm
Terminal H1 4.59 4.58 H4 3.90 3.90
Penultimate HI 4.51 4.41 H4 4.18 4.18
resonance of H1 (4.51 ppm) of the penultimate ,&Gal residue in A-1 was downfield shifted by 0.02ppm compared to the reported value (4.49 ppm) of the P-Gal residue in the GalPl-4(HS03-6)(2,5)-anhMan-ol sequence [44]. The reso- nance at 4.59 ppm was then assigned to HI of the terminal P-Gal residue, which showed that the sulfate group on the 6 position of (2,5)-anhMan-ol has little effect on the chemical shift of HI of the terminal P-Gal residue (the resonance of H1 of the terminal P-Gal residue in the Gal~1-4GalP1-4(2,5)-anhMan-o1 was 4.58 ppm [43]). The resonances at 4.18 pprn and 3.90 ppm were assignable to H4 of the penultimate ,!-Gal and the terminal P- Gal residues, respectively, by comparing the reported values [43]. The results are summarized in Table 4.
The A-2 fraction (about 50 pg as Gal) was shown to consist of two components, one of which had the same migration rate as A-I, the other migrating further. Although the amount of the latter material (about 20 pg) was too small to be purified by preparative TLC, some structural information was obtained by FAB-MS and 'H-NMR spectroscopy. The (*H,)acetyl deriva- tive of A-2 afforded two molecular ions [M-HI- at mlz 675 [ (HSO,) ,Hex, an hHex-ol,] and at mlz 972 [(HSO,),Hex,anhHex- ol,] in the negative mode. The 'H-NMR spectrum of A-2 showed three assignable anomeric proton resonances at 4.59 ppm, 4.51 ppm, and 4.48 ppm (spectrum not shown). The resonances at 4.59 ppm and 4.51 ppm were completely identical to those of A-1, while the resonance at 4.48 ppm was close to the value (4.49 ppm) reported in the literature [44] for GalPl - 4(HSO, - 6)(2,5)-anhMan-ol. These data led us to conclude that A-2 consisted of two components: GalPl - 4GalP1 - 4(HSO, - 6)(2,5)-anhMan-ol and GalP1 - 4(HSO, - 6)(2,5)- anhMan-01.
In summary, hydrazinolysis/nitrous acid deamination of the hyosophorin gave the oligosaccharide products : Gal,!l - 4(2,5)-anhMan-ol, GalPl - 4GalP1 - 4(2,5)-anhMan-ol, GalPl - 4(HS03 - 6)(2,5)-anhMan-ol, and GalP1 - 4GalP1 - 4(HSO, - 6)(2,5)-anhMan-ol. The data obtained thus far sug- gested that peripheral portion of the glycan has poly(Wacetyl1ac- tosamine) sequences which were partially P-galactosylated at position 4 of the P-Gal residue and also partially sulfated at posi- tion 6 of the P-GlcNAc residue (Schemes A and B). Scheme A [linear poly(N-acetyllactosamine) sequence] :
[- 3GalP1-4( 2 HS0,-6)Gl~NAc/31-],~ - GalP1-4( 2 HS03-6)(2,5)-anhMan-ol
Scheme B [branched poly(N-acetyllactosamine) sequence] :
[- 3(GalPl-4)Gal/ll-4( 2 HS0,- 6)GlcNAcPI-], - Gal/31-+4Gal/l1-4( 2 HS0,- 6)(2,5)-anhMan-o1 These structures were further confirmed by digestion of the
hyosophorin with endo-P-galactosidase (described below).
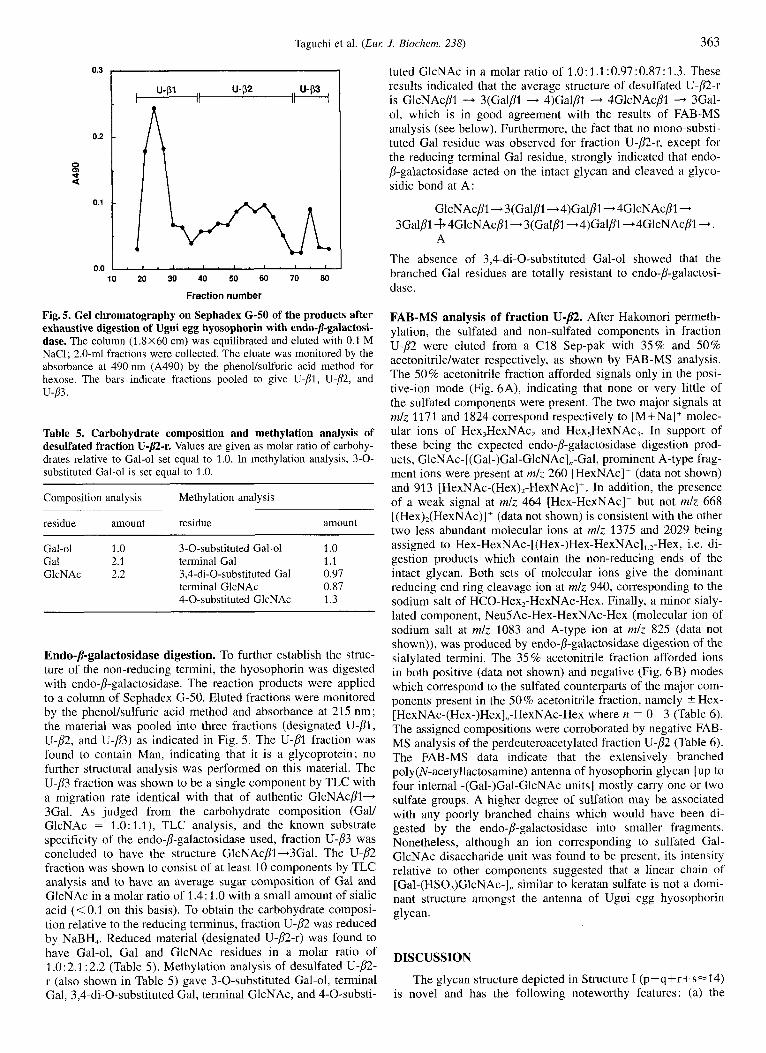
Taguchi et al. (Eur: J . Biochem. 238) 363
0.3 , I
0.2 -
0.1 -
10 20 30 40 50 60 70 80
Fraction number
Fig. 5. Gel chromatography on Sephadex G-SO of the products after exhaustive digestion of Ugui egg hyosophorin with endo-P-galactosi- dase. The column (13x60 cm) was equilibrated and eluted with 0.1 M NaC1; 2.0-ml fractions were collected. The eluate was monitored by the absorbance at 490 nm (A490) by the phenollsulfuric acid method for hexose. The bars indicate fractions pooled to give U-pl, U-P, and u-83.
Table 5. Carbohydrate composition and methylation analysis of desulfated fraction U-p-r. Values are given as molar ratio of carbohy- drates relative to Gal-ol set equal to 1.0. In methylation analysis, 3-0- substituted Gal-ol is set equal to 1.0.
Composition analysis Methylation analysis
residue amount residue amount
Gal-ol 1 .0 3-0-substituted Gal-ol 1 .0 Gal 2.1 terminal Gal 1.1 GlcNAc 2.2 3,4-di-O-substituted Gal 0.97
terminal GlcNAc 0.87 4-0-substituted GlcNAc 1.3
Endo-/I-galactosidase digestion. To further establish the struc- ture of the non-reducing termini, the hyosophorin was digested with endo-/I-galactosidase. The reaction products were applied to a column of Sephadex G-50. Eluted fractions were monitored by the phenollsulfuric acid method and absorbance at 21.5 nm; the material was pooled into three fractions (designated U-PI, U-p2, and U-p3) as indicated in Fig. 5. The U-PI fraction was found to contain Man, indicating that it is a glycoprotein; no further structural analysis was performed on this material. The U-p3 fraction was shown to be a single component by TLC with a migration rate identical with that of authentic GlcNAcPI- 3Gal. As judged from the carbohydrate composition (Gall GlcNAc = 1.0: 1.1), TLC analysis, and the known substrate specificity of the endo-P-galactosidase used, fraction U-p3 was concluded to have the structure GlcNAcpl-3Gal. The U-p2 fraction was shown to consist of at least 10 components by TLC analysis and to have an average sugar composition of Gal and GlcNAc in a molar ratio of 1.4 : 1 .O with a small amount of sialic acid (<0.1 on this basis). To obtain the carbohydrate composi- tion relative to the reducing terminus, fraction U-p2 was reduced by NaBH,. Reduced material (designated U-p2-r) was found to have Gal-ol, Gal and GlcNAc residues in a molar ratio of 1 .O: 2.1 : 2.2 (Table 5). Methylation analysis of desulfated U-p2- r (also shown in Table 5) gave 3-0-substituted Gal-ol, terminal Gal, 3,4-di-O-substituted Gal, terminal GlcNAc, and 4-0-substi-
tuted GlcNAc in a molar ratio of 1.0:1.1:0.97:0.87:1.3. These results indicated that the average structure of desulfated U-@-r is GlcNAcPl - 3(Galp1 - 4)GalPl - 4GlcNAcP1 - 3Gal- 01, which is in good agreement with the results of FAB-MS analysis (see below). Furthermore, the fact that no mono-substi- tuted Gal residue was observed for fraction U-p2-r, except for the reducing terminal Gal residue, strongly indicated that endo- P-galactosidase acted on the intact glycan and cleaved a glyco- sidic bond at A:
GlcNAcpl- 3(Galp1-+4)GalBl -4GlcNAcPl - A
3 G a l p l 4 4GlcNAcP1 -3(Gal~1-4)Gal~1-.4GlcNAc~l- .
The absence of 3,4-di-O-substituted Gal-ol showed that the branched Gal residues are totally resistant to endo-p-galactosi- dase.
FAB-MS analysis of fraction U - P . After Hakomori permeth- ylation, the sulfated and non-sulfated components in fraction U-p2 were eluted from a C18 Sep-pak with 35% and 50% acetonitrilelwater respectively, as shown by FAB-MS analysis. The 50% acetonitrile fraction afforded signals only in the posi- tive-ion mode (Fig. 6A), indicating that none or very little of the sulfated components were present. The two major signals at rnlz 1171 and 1824 correspond respectively to [M+Na]+ molec- ular ions of Hex,HexNAc, and Hex,HexNAc,. In support of these being the expected endo-p-galactosidase digestion prod- ucts, GlcNAc-[(Gal-)Gal-GlcNAc],,-Gal, prominent A-type frag- ment ions were present at rnlz 260 [HexNAc]' (data not shown) and 913 [HexNAc-(Hex),-HexNAc]'. In addition, the presence of a weak signal at rnlz 464 [Hex-HexNAc]' but not mlz 668 [(Hex),(HexNAc)]+ (data not shown) is consistent with the other two less abundant molecular ions at rnlz 137.5 and 2029 being assigned to Hex-HexNAc-[(Hex-)Hex-HexNAc],,,-Hex, i.e. di- gestion products which contain the non-reducing ends of the intact glycan. Both sets of molecular ions give the dominant reducing end ring cleavage ion at mlz 940, corresponding to the sodium salt of HCO-Hex,-HexNAc-Hex. Finally, a minor sialy- lated component, NeuSAc-Hex-HexNAc-Hex (molecular ion of sodium salt at mlz 1083 and A-type ion at rnlz 825 (data not shown)), was produced by endo-l)-galactosidase digestion of the sialylated termini. The 35 % acetonitrile fraction afforded ions in both positive (data not shown) and negative (Fig. 6B) modes which correspond to the sulfated counterparts of the major com- ponents present in the SO% acetonitrile fraction, namely ? Hex- [HexNAc-(Hex-)Hex],,-HexNAc-Hex where n = 0-3 (Table 6). The assigned compositions were corroborated by negative FAB- MS analysis of the perdeuteroacetylated fraction U-p2 (Table 6). The FAB-MS data indicate that the extensively branched poly(N-acetyllactosamine) antenna of hyosophorin glycan [up to four internal -(Gal-)Gal-GlcNAc units] mostly carry one or two sulfate groups. A higher degree of sulfation may be associated with any poorly branched chains which would have been di- gested by the endo-p-galactosidase into smaller fragments. Nonetheless, although an ion corresponding to sulfated Gal- GlcNAc disaccharide unit was found to be present, its intensity relative to other components suggested that a linear chain of [Gal-(HS0,)GlcNAc-1, similar to keratan sulfate is not a domi- nant structure amongst the antenna of Ugui egg hyosophorin glycan.
DISCUSSION
The glycan structure depicted in Structure I (p+q+r+s=14) is novel and has the following noteworthy features: (a) the

364
95:
90:
Taguchi et al. (ELM J . Biochem. 238)
r c
t
L
1003 1940
05
00:
75:
7 0:
6 5:
6 0:
55:
50:
4 5:
4 0:
35:
30
2 5:
20:
15
"i
4
I
1417
1955
1867
I
5 , I I I 'I- 8 b o 960 1doo i l ia0 12'00 1200 1200 15100 16100 17'00 18'00 1400 20'00 21100 22'00 23'00 24 '00 2 5 0 0 , ' ' ~ ~ I " ' ' 2 6 0 0 m/z
Fig. 6. Partial FAB-MS spectra of permethylated fraction U - B : (A) 50 % acetonitrile fraction, in the positive-ion mode; (€3) 35 % acetonitrile fraction in the negative-ion mode. See Table 6 for signal assignments of (B). The signal at m/7 876 is due to a contaminant.
glycan has only a tetraantennary core structure; (b) the structure of the peripheral portion is similar to that of keratan sulfate but is completely devoid of 6-0-sulfated Gal residues ; (c) branched Gal residues are present, -4GlcNAc[~l-3(Gal~l- 4)GalPl-; (d) a sialylated antenna is also present, but it is
notably shorter than the sulfated antennae (Sia -+ Gal -, GlcNAc - Gal - GlcNAc - Man at longest). The occurrence of a tri- or tetra-antennary (multiantennary) complex-type core structures possessing glycosaminoglycan chains has been re- ported i n some mammalian cells [14, 261, but this study presents

Taguchi et al. (EM J. Biochem. 238) 365
Table 6. The major sulfated components in fraction U-p2 as detected by FAB-MS analysis of the permethyl and perdeuteroacetyl deriva- tives in the negative-ion mode. The permethyl derivatives refer to those eluted in the 3.5% acetonitrile fraction (Fig. 6B). Due to the fractionation with a step gradient of acetonitrile/water, the negatively charged di- and tri-saccharides may be under-represented since they were also partially eluted in the earlier 15% acetonitrile fraction. Conversely, some of the mono-sulfated larger components were fractionated into the later 50% acetonitrile fraction, the positive FAB-mass spectrum of which is shown in Fig. 6A. The perdeutroacetyl derivatives were not further fractionated.
Composition assigned mlz for
permethyl perdeutero- acetyl
Hex, HexNAc, (HSO;) Hex,HexNAc,(HSO;) Hex,HexNAc,(HSO, ) Hex,HexNAc,( HSO;) Hex,HexN Ac,(HSO;),Na' Hex,HexNAc,(HSO;) Hex,HexNAc,(HSO;),Na' Hex,HexNAc,( HSO;) Hex,HexN Ac,(HSO;),Na' Hex,HexNAc,(HSO;) Hex,HexNAc,(HSO;),Na '
-
764 1213 1417 1 505 1867 1955
21 60
2609
-
-
732 1029 1619 1917 1914 2508 2565 2805 2862 3395 3453
the precise structural analysis of highly sulfated multiantennary glycan chains in nature.
In recent years, several biological roles have been described for the sulfate group in carbohydrates. GlyCAM-1 is reported to be mucin-like glycoprotein with a dense array of 0-linked carbohydrate chains and the majority of 0-linked chains were sulfated in the 6'-sulfated sialyl Le" sequence. The GlyCAM-I molecule purified from lymph nodes treated with chlorate, an inhibitor for sulfation, did not sustain binding activity to the L-selectin-IgG chimera, showing that multiple sulfation of Gly- CAM-1 may form an essential part of the recognition motif for L-selectin [45]. In the course of searching for the major carrier glycoprotein for poly(N-acetyllactosamine) structure, Fukuda's group found the lamp-I ,2 molecule (lysosomal-associated mem- brane glycoprotein) [46] and recently showed that the amount of cell surface lamp-I was directly proportional to the extent of E-selectin-mediated cell-cell adhesion [47], indicating that the amount of sialyl Le" structure (which E-selectin could recog- nize) on the poly(N-acetyllactosamine) sequence of lamp-1 should be involved in this adhesion event. They also found that the highly metastatic cells expressed more lamp-1 and lamp-2 on their cell surface than the low-metastatic cells [48]. These studies suggested that the multivalency of specific epitope structures may possibly be essential for cell -cell interactions. Although the function of this highly sulfated multiantennary N- linked glycan in Ugui egg hyosophorin is unknown, a search for the molecules which interact with this hyosophorin will be now possible.
Recently, we have identified a hyosophorin-like molecule (carbohydrate-rich glycoprotein) in eggs of the Japanese toad Bufo juponicus [49] and the African clawed frog Xenopus laevis (Shimoda et al., unpublished results). The structural features of the former are quite similar to that found in Oryzius lutipes [29], while those of the latter are almost identical to that in Parulich- thys olivuceus [SO]. These glycan structures are basically iden- tical to those expressed in embryonal and oncodevelopmental cells reported by several groups [51-541 in having the poly/ oligo(N-acetyllactosamine) backbone, but critically different in
the properties that these glycan chains were resistant to several endo- and exo-glycosidases which are commonly used for eluci- dating N-linked glycan chains in some cell surfaces. In these studies, structural information was obtained for extremely lim- ited amounts of materials, i.e. radioisotope level, and was mainly based on the experiments using endo- andor exo-glycosidase digestions. In view of this, it is highly possible that such hyoso- phorin-like N-linked glycan chains were overlooked and our studies strongly indicate that such glycan chains may not be confined to fish and amphibian species, but are more widespread in the animal kingdom.
Our study of the structural determination of the T huko- nensis glycan chains demonstrated that the internal glycosidic linkages at the branched Gal residues were not cleaved by endo- P-galactosidase (E. freundii) [28, 291. However, this enzyme was shown to cleave the following site (A):
Galpl '.
4
/" - GlcNAcpl
Consequently, care must be practised in the interpretation of the data for possible diversity in poly(N-acetyllactosamine) units in terms of endo-P-galactosidase digestion. It should also be pointed out that selective chemical degradation methods, such as hydrazinolysis/nitrous acid deamination, have proved to be essential and extremely useful in establishing the precise struc- tures of such complex and large glycan chains as described in this report.
This research was supported in part by Grants-in-Aid for Interna- tional Scientific Research Program: Joint Research (0404405 to YI and 07044228 to KK) and for General Scientific Research (07680658 to SI) from the Ministry of Education, Science, and Culture of Japan. The Im- perial College work was supported by grants (to HRM and AD) from the Medical Research Council (Programme Grant), the Biotechnology and Biological Sciences Research Council and the Wellcome Trust (grant 030826).
REFERENCES 1. Freeze, H. H., Yeh, R., Miller, A. L. & Kornfeld, S. (1983) Struc-
tural analysis of the asparagine-linked oligosaccharides from three lysosomal enzymes of Dictyostelium discoideum. Evidence for an unusual acid-stable phosphodiester, J. Biol. Chem. 258, 14 874- 14 879.
2. Freeze, H. H. & Wolgast, D. (1986) Structural analysis of N-linked oligosaccharides from glycoproteins secreted by Dictyosteliurn discoideum. Identification of mannose 6-sulfate, J. Bid. Cheni.
3. Bergwerff, A. A., van Oostrum, J., Kamerling, J. P. & Vliegenthart, J. F. G. (1995) The major N-linked carbohydrate chains from hu- man urokinase. The occurrence of 4-O-sulfated, (n2-6)-sialylated or (al-3)-fucosylated N-acetylgalactosamine (J'l-4)-N-acetylglu- cosamine elements, Eur: J. Biochenz. 228, 1009-1019.
4. Pinter, A. & Compans, R. W. (1975) Sulfated components of enve- loped viruses, J. Virol. 16, 859-866.
5. Yamashita, K., Ueda, I. & Kobata, A. (1983) Sulfated asparagine- linked sugar chains of hen egg albumin, J . B i d . Chem. 258,
6. Noguchi, S., Hatanaka, Y., Tobita, T. & Nakano, M. (1992) Struc- tural analysis of the N-linked carbohydrate chains of the 55-kDa glycoprotein family (PZP3) from porcine zona pellucida, Eur: J . Biochem. 204, 1089-1100.
7. Noguchi, S. & Nakano, M. (1992) Structure of the acidic N-linked carbohydrate chains of the 55-kDa glycoprotein family (PZP3) from porcine zona pellucida, E m J. Biochem. 209, 883-894.
261, 127-134.
14 144- 14 147.

366 Taguchi et al. (Eur: J. Biochem. 238)
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
Edge, A. S. B. & Spiro, R. G. (1984) Presence of sulfate in N- glycosidically linked carbohydrate units of calf thyroid plasma membrane glycoproteins, J. Biol. Chem. 259, 4710-4713.
Kamerling, J. P., Rijkse, I . , Maas, A. A. M., van Kuik, J. A. & Vliegenthart, J. F. G. (1988) Sulfated N-linked carbohydrate chains in porcine thyroglobulin, FEBS Lett. 241, 246-250.
Spiro, R. G. & Bhoyroo, V. D. (1988) Occurrence of sulfate in the asparagine-linked complex carbohydrate units of thyroglobulin. Identification and localization of galactose 3-sulfate and N-acetyl- glucosamine 6-sulfate residues in the human and calf proteins, J . B id . Chem. 263, 14351-14358.
de Waard, P., Koorevaar, A., Kamerling, J. P. & Vliegenthart, J. F. G. (1991) Structure determination by 'H NMR spectroscopy of (sulfated) sialylated N-linked carbohydrate chains released from porcine thyroglobulin by peptide-N4-(N-acetyl-~-glucosaminyl) asparagine amidase-F, J. Bid. Chem. 266, 4237-4243.
Freeze, H. H. & Varki, A. (1986) Endo-glycosidase F and pep- tide:N-glycosidase F release the great majority of total cellular N-linked oligosaccharides : use in demonstrating that sulfated N- linked oligosaccharides are frequently found in cultured cells, Biochem. Biophys. Res. Cmmmurz. 140, 967-973.
Roux, L., Holojda, S., Sundblad, G., Freeze, H. H. & Varki, A. (1988) Sulfated N-linked oligosaccharides in mammalian cells. I. Complex-type chains with sialic acids and 0-sulfate esters, J , B i d . Chem. 263, 8879-8889.
Sundblad, G., Holojda, S., Roux, L., Varki, A. & Freeze, H. H. (1988) Sulfated N-linked oligosaccharides in mammalian cells. 11. Identification of glycosaminoglycan-like chains attached to com- plex type glycans, J . Biol. Chem. 263, 8890-8896.
Sundblad, G., Kajiji, S., Quaranta, V., Freeze, H. H. & Varki, A. (1988) Sulfated N-linked oligosaccharides in mammalian cells. 111. Characterization of a pancreatic carcinoma cell surface glyco- protein with N- and 0-sulfate esters on asparagine-linked glycans, J. Biol. Chem. 263, 8897-8903.
Green, E. D., van Halbeek, H., Boime, I. & Baenziger, J. U. (1985) Structural elucidation of the disulfated oligosaccharide from bo- vine lutropin, J. B i d . Chem. 260, 1.5 623 - 1.5 630.
Green, E. D. & Baenziger, J. U. (1988) Asparagine-linked oligosac- charides on lutropin, follitropin and thyrotropin. I. Structural elu- cidation of the sulfated and sialylated oligosaccharides on bovine, ovine, and human pituitary glycoprotein hormones, J . Bid. Chem.
Green, E. D. & Baenziger, J. U. (1988) Asparagine-linked oligosac- charides on lutropin, follitropin, and thyrotropin. 11. Distributions of sulfated and sialylated oligosaccharides on bovine, ovine, and human pituitary glycoprotein hormones, J. Bid. Chem. 26.?, 36- 44.
Weisshaar, G., Hiyama, J. & Renwick, A. G. C. (1990) Site specific N-glycosylation of ovine lutropin. Structural analysis by one- and two-dimensional 'H-NMR spectroscopy, Eur: J . Biochem. 192,
Smith, P. L., Bousfield, G. R., Kumar, S., Fiete, D. & Baenziger, J. U. (1993) Equine lutropin and chorionic gonadotropin bear oligo- saccharides terminating with S04-4-GalNAc and Siaa2,3Gal, re- spectively, J. Biol. Chem. 268, 795-802.
Nakazawa, K., Newsome, D. A,, Nilsson, B., Hascall, V. C. & Hassell, J. R. (1983) Purification of keratan sulfate proteoglycan from monkey cornea. Isolation of the keratan sulfate linkage re- gion and the mannose-containing oligosaccharides, J . Biol. Chem.
Nilsson. B., Nakazawa, K., Hassell, J. R., Newsome, D. A. & Hascall, V. C. (1983) Structure of oligosaccharides and the linkage region between kerdtdn sulfate and the core protein on proteoglycans from monkey cornea, J. Biol. Chem. 258, 6056- 6063.
Yamaguchi, H. (1983) Studies on the carbohydrate-protein linkage region in bovine corneal keratan sulfate. 11. Structural studies on linkage region-enriched neutral glycopeptides, J. Biochem.
Oeben, M., Keller, R., Stuhlsatz, H. W. & Greiling, H. (1987) Con- stant and variable domains of different disaccharide structure in corneal keratan sulphate chains. Biochem. J. 248. 85-93.
263, 25-35.
741 -751.
258, 60.5 1 - 605.5.
(Tokyo) 94, 21.5-221.
25. Hopwood, J. J. & Robinson, H. C. (1974) The alkali-labile linkage between keratan sulphate and protein, Biochem. J. 141, 57-69.
26. Pierce, M. & Arango, J. (1986) Rous sarcoma virus-transformed baby hamster kidney cells express higher levels of asparagine- linked tri- and tetraantennary glycopeptides containing [GlcNAc- p( 1,6)Man-a(1 ,6)Man] and poly(N-acetyllactosamine) sequences than baby hamster kidney cells, J. Biol. Chem. 261, 10772- 10777.
27. Inoue, S., Iwasaki, M., Ishii, K., Kitajima, K. & Inoue, Y. (1989) Isolation and structures of glycoprotein-derived free sialooligo- saccharides from the unfertilized eggs of Tribolodon hakonensis, a dace. Intracellular accumulation of a novel class of biantennary disialooligosaccharides, J. Biol. Chem. 264, 18 520- 18 526.
28. Taguchi, T., Seko, A,, Kitajima, K., Inoue, S., Iwamatsu, T., Khoo, K.-H., Morris, H. R., Dell, A. & Tnoue, Y. (1993) Structural studies of a novel type of tetraantennary sialoglycan unit in a carbohydrate-rich glycopeptide isolated from the fertilized eggs of indian medaka fish, Oryzias melastigma, J. Biol. Chem. 268,
29. Taguchi, T., Seko, A,, Kitajima, K., Muto, Y., Inoue, S., Khoo, K.-H., Morris, H. R., Dell, A. & Inoue, Y. (1994) Structural studies of a novel type of pentaantennary large glycan unit in the fertilization-associated carbohydrate-rich glycopeptide isolated from the fertilized eggs of Oryzias latipes, J. Biol. Chem. 269,
30. Khoo, K.-H., Morris, H. R., McDowell, R. A., Dell, A,, Maccarana, M. & Lindahl, U. (1993) FABMWderivatisation strategies for the analysis of heparin-derived oligosaccharides, Carbohydr: Rex
31. Dubois, M., Gilles, K. A,, Hamilton, J. K., Rebers, P. A. & Smith, F. (1956) Colorimetric method for determination of sugars and related substances, Anal. Chem. 28, 350-356.
32. Aminoff, D. (1961) Methods for the quantitative estimation of N- acetylneuraminic acid and their application to hydrolysates of sialomucoids, Biochem. J . 81, 384-392.
33. Uchida, Y., Tsukada, Y. & Sugimori, T. (1977) Distribution of neuraminidase in Arthrobacter and its purification by affinity chromatography, J. Biochem. (Tokyo) 82, 142.5-1433.
34. Svennerholm, L. (1963) Sialic acids and derivatives: Estimation by the ion-exchange method, Methods Enzymol. 6, 459-462.
35. Nomoto, H., Iwasaki, M., Endo, T., Inoue, S., Inoue, Y. & Matsu- mum, G. (1982) Structures of carbohydrate units isolated from trout egg polysialoglycoproteins : Short-cored units with oligosia- losyl groups, Arch. Biochem. Biophys. 218, 335-341.
36. Hakomori, S. (1964) A rapid permethylation of glycolipid & poly- saccharide catalyzed by methylsulfinyl carbanion in dimethyl sulfoxide, J . Biochem. (Tokyo) 55, 205-208.
37. Ciucanu, I. & Kerek, F. (1984) A simple and rapid method for the permethylation of carbohydrates, Carhohydr: Res. 131, 209- 217.
38. Levery, S. B. & Hakomori, S. (1987) Microscale methylation analy- sis of glycolipids using capillary gas chromatography-chemical ionization mass fragmentography with selected ion monitoring, Methods Enzymol. 138, 13 -25.
39. Vliegenthart, J. F. G., Dorland, L. & van Halbeek, H. (1983) High- resolution, 'H-nuclear magnetic resonance spectroscopy as a tool in the structural analysis of carbohydrates related to glycopro- teins, Adv. Carbohydr: Chern. Biochem. 41, 209-374.
40. Nimtz, M., Martin, W., Wray, V., Kloppel, K.-D., Augustin, J . & Conradt, H. S. (1 993) Structures of sialyldted oligosaccharides of human erythropoietin expressed in recombinant BHK-21 cells, ELK J. Biochem. 213, 39-56.
41. Taguchi, T., Kitajima, K., Muto, Y., Yokoyama, S., Inoue, S. & Inoue, Y. (1995) Proton NMR study of the trimannosyl unit in a pentaantennary N-linked decasaccharide structure. Complete as- signment of the proton resonances and conformational character- ization, Eur. J. Biochem. 225, 822-829.
42. Hokke, C. H., Damm, J. B. L., Penninkhof, B., Aitken, R. J., Kamer- ling, J. P. & Vliegenthart, J . F. G. (1994) Structure of the 0-linked Carbohydrate chains of porcine zona pellucida glycoproteins, Eur: .I. Biochem. 221, 491 -512.
43. Taguchi, T., Kitajima, K., Muto, Y., Inoue, S., Khoo, K.-H., Morris, H. R.. Dell, A., Wallace, R. A,, Selman, K. & Inoue, Y. (1995)
2353 - 2362.
8762 - 877 1.
244, 205-223.

Tdguchi et al. (Eur: J. Biochern. 238) 367
A precise structural analysis of a fertilization-associated carbohy- drate-rich glycopeptide isolated from the fertilized eggs of eury- haline killi fish (Fundulus heteroclitus). Novel penta-antennary N-glycan chains with a bisecting N-acetylglucosaminyl residue, Glycobiology 5, 611 -624.
44. Huckerby, T. N., Brown, G. M., Dickenson, J. M. & Nieduszynski, I. A. (1995) Spectroscopic characterization of disaccharides derived from keratan sulfates, Eur: J. Biochem. 229, 119-131.
45. Imai, Y., Lasky, L. A. & Rosen, S. D. (1993) Sulphation requirement for GlyCAM-1, an endothelial ligand for L-selectin, Nature 361, 555-557.
46. Carlsson, S. R., Roth, J. , Piller, F. & Fukuda, M. (1988) Isolation and characterization of human lysosomal membrane glycopro- teins, h-lamp-l and h-lamp-2. Major sialoglycoproteins carrying polylactosaminoglycan, J. Biol. Chem. 263, 18 91 1 - 18 91 9.
47. Sawada, R., Lowe, J. B. & Fukuda, M. (1993) E-selectin-dependent adhesion efficiency of colonic carcinoma cells is increased by genetic manipulation of their cell surface lysosomal membrane glycoprotein-1 expression levels, J. Bid . Chem. 268, 12675 - 12681.
48. Saitoh, O., Wang, W.-C., Lotan, R. & Fukuda, M. (1992) Dif- ferential glycosylation and cell surface expression of lysosomal membrane glycoproteins in sublines of a human colon cancer ex- hibiting distinct metastatic potentials, J. Biol. Chem. 267, 5700- 571 1.
49. Shimoda, Y., Kitajima, K., inoue, S., Wardrip, N. J., Hedrick, J. L. & Inoue, Y. (1993) Isolation and structural determination of high-molecular-weight glycan units present in egg cortical granu-
lar-derived glycoproteins of Bufo japonicus and Xenopus laevis, Glycoconjugate J . 10, 264.
SO. Seko, A,, Kitajima, K., Iwasaki, M., Inoue, S. & Inoue, Y. (1989) Structural studies of fertilization-associated carbohydrate-rich glycoproteins (hyosophorin) isolated from the fertilized and un- fertilized eggs of Flounder, Paralichthys olivaceus. Presence of a novel penta-antennary N-linked glycan chain in the tandem re- peating glycopeptide unit of hyosophorin, J. Bid . Chem. 264, 15 922- 15 929.
51. Fukuda, M. (1 985) Cell surface glycoconjugates as onco-differentia- tion markers in hematopoietic cells [review], Biochim. Biophys.
52. Hakomori, S., Fukuda, M. & Nudelman, E. (1982) Role of cell sur- face carbohydrates if differentiation: behavior of lactosaminogly- can in glycolipids and glycoproteins, Teratocarcinoma and em- bryonic cell interactions, pp. 179-200, Japan Scientific Societies Press, Tokyo.
53. Muramatsu, H., Ishihara, H., Miyauchi, T., Gachelin, G., Fujisaki, T., Tejima, S. & Muramatsu, T. (1983) Glycoprotein-bound large carbohydrates of early embryonic cells: structural characteristic of the glycan isolated from F9 embryonal carcinoma cells, J . Bio- chern. (Tokyo) 94, 799-810.
54. Kamada, Y., Arita, Y., Ogata, S., Muramatsu, H. & Muramatsu, T. (1987) Receptors for fucose-binding proteins of Lotus tetragono- lobus isolated from mouse embryonal carcinoma cells. Structural characteristics of the poly(N-acetyl1actosamine)-type glycan, Eur. J. Biochem. 163, 497-502.
Acta 780, 119-150.




















