
Nutrient study for the transition from earthen sedimentation ponds to ones lined with pvc in...
13
Journal of Applied Phycology 16: 341–353, 2004. C 2004 Kluwer Academic Publishers. Printed in the Netherlands. 341 Nutrient study for the transition from earthen sedimentation ponds to ones lined with pvc in integrated mariculture systems, what needs to be done? John J. Lee 1,∗ , Damaris Rodriguez 1 , Amir Neori 2 , Oded Zmora 2 , Andrew Symons 1 & Muki Shpigel 2 1 Department of Biology, City College of CUNY, Convent Ave at 138 St., New York, NY 10031, USA; 2 Israel Oceanographic and Limnological Research, National Center for Mariculture, P.O. Box 1212, Eilat, Israel ∗ Author for correspondence (e-mail: [email protected]. edu; phone +1 212 650 8545; fax +1 212 650 8585) Received 5 August 2003; revised and accepted 18 May 2004 Key words: Amphora cf tenerrima, Amphora luciae, Cylindrotheca closterium, diatoms, integrated mariculture systems, mariculture effluent, microphytobenthos, Nitzschia laevis, Navicula cf lenzii, Navicula cflineola, Navicula salinicola, Navicula cf viminoides, pollution mitigation, Si requirements, trace elements Abstract Stable diatom populations in earthen ponds for fishpond effluent treatment supported fast rates of bivalve growth in integrated mariculture systems. However, when these ponds were lined with PVC plastic sheets to prevent seepage, the populations of benthic diatoms dwindled, and did not support, as before, a commercially acceptable rate of growth by oysters and clams. Experiments were undertaken to understand the problem and restore the diatom productivity of such ponds. Clones of Amphora luciae, A,cftenerrima, Cylindrotheca closterium, Navicula cf lineola, N .cf lenzii, N . salinicola, N . cf viminoides, and Nitzschia laevis were isolated in axenic culture from an earthen sedimentation pond. Their N, P, Si, and trace element requirements for growth in fully defined media, and in media formulated with mariculture effluent, were studied in axenic batch culture. In fully defined batch culture tests, most of the isolates achieved their highest density in media with 32 µM P, 0.7 mM N, 20 µM Fe, 10 nM–20 µM Mn, 10–20 µM Zn and Co, and ≥17.5 µM Si. Enrichment by trace elements and Si stimulated the populations of these diatoms even in media based on nutrient-enriched mariculture effluents. However, in large flow- through agnotobiotic mesocosms (700 L), only Si enrichment was needed. Si concentration >100 µM was required to promote the sustained blooms of diatoms in full-sized and commercial PVC-lined fishpond effluent treatment ponds (300 m 2 , 1 m depth). Except for Si, the requirements of the diatoms for micronutrients were apparently fully satisfied by the fishpond effluents (uneaten food and fish-waste). A molar ratio of 1:1 between Si and N is necessary to sustained dense diatom populations in the pond water. It is therefore recommended to enrich plastic lined mariculture effluent treatment/sedimentation ponds with Si, if the goal is to raise bivalves as a secondary crop. Introduction Mariculture can be an important economic boost to an economy as long as it does not extract a high ecosys- tem price (Stickney, 2002; DeVoe & Hodges, 2002). The gilthead sea bream (Sparus aurata L.) is raised in mariculture ponds at the National Center for Mar- iculture (NCM) of Israel Oceanographic and Limno- logical Research (IOLR) that use sea water from the Gulf of Aqaba (Eilat), the Red Sea. For more than a decade, earthen sedimentation ponds were used to miti- gate pollution from fishpond effluents. The ponds were rich with microalgae, thanks to the plentiful sunlight and the nutrient-rich water. As they grew, the microal- gae removed various forms of nutrients, introduced in to the seawater by the fish culture. The microalgae, in turn, were harvested by clams and oysters, which clarified the water and provided additional cash crops (Hughes-Games, 1977; Shpigel & Fridman, 1990; Sh- pigel & Blaylock, 1991; Shpigel et al., 1993a).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Nutrient study for the transition from earthen sedimentation ponds to ones lined with pvc in...

Journal of Applied Phycology 16: 341–353, 2004.C© 2004 Kluwer Academic Publishers. Printed in the Netherlands.
341
Nutrient study for the transition from earthen sedimentation ponds to oneslined with pvc in integrated mariculture systems, what needs to be done?
John J. Lee1,∗, Damaris Rodriguez1, Amir Neori2, Oded Zmora2, Andrew Symons1
& Muki Shpigel2
1Department of Biology, City College of CUNY, Convent Ave at 138 St., New York, NY 10031, USA; 2IsraelOceanographic and Limnological Research, National Center for Mariculture, P.O. Box 1212, Eilat, Israel∗Author for correspondence (e-mail: [email protected]. edu; phone +1 212 650 8545; fax +1 212 650 8585)
Received 5 August 2003; revised and accepted 18 May 2004
Key words: Amphora cf tenerrima, Amphora luciae, Cylindrotheca closterium, diatoms, integrated mariculturesystems, mariculture effluent, microphytobenthos, Nitzschia laevis, Navicula cf lenzii, Navicula cflineola, Naviculasalinicola, Navicula cf viminoides, pollution mitigation, Si requirements, trace elements
Abstract
Stable diatom populations in earthen ponds for fishpond effluent treatment supported fast rates of bivalve growthin integrated mariculture systems. However, when these ponds were lined with PVC plastic sheets to preventseepage, the populations of benthic diatoms dwindled, and did not support, as before, a commercially acceptablerate of growth by oysters and clams. Experiments were undertaken to understand the problem and restore thediatom productivity of such ponds. Clones of Amphora luciae, A,cftenerrima, Cylindrotheca closterium, Naviculacf lineola, N .cf lenzii, N . salinicola, N . cf viminoides, and Nitzschia laevis were isolated in axenic culture froman earthen sedimentation pond. Their N, P, Si, and trace element requirements for growth in fully defined media,and in media formulated with mariculture effluent, were studied in axenic batch culture. In fully defined batchculture tests, most of the isolates achieved their highest density in media with 32 µM P, 0.7 mM N, 20 µM Fe, 10nM–20 µM Mn, 10–20 µM Zn and Co, and ≥17.5 µM Si. Enrichment by trace elements and Si stimulated thepopulations of these diatoms even in media based on nutrient-enriched mariculture effluents. However, in large flow-through agnotobiotic mesocosms (700 L), only Si enrichment was needed. Si concentration >100 µM was requiredto promote the sustained blooms of diatoms in full-sized and commercial PVC-lined fishpond effluent treatmentponds (300 m2, 1 m depth). Except for Si, the requirements of the diatoms for micronutrients were apparentlyfully satisfied by the fishpond effluents (uneaten food and fish-waste). A molar ratio of 1:1 between Si and N isnecessary to sustained dense diatom populations in the pond water. It is therefore recommended to enrich plasticlined mariculture effluent treatment/sedimentation ponds with Si, if the goal is to raise bivalves as a secondarycrop.
Introduction
Mariculture can be an important economic boost to aneconomy as long as it does not extract a high ecosys-tem price (Stickney, 2002; DeVoe & Hodges, 2002).The gilthead sea bream (Sparus aurata L.) is raisedin mariculture ponds at the National Center for Mar-iculture (NCM) of Israel Oceanographic and Limno-logical Research (IOLR) that use sea water from theGulf of Aqaba (Eilat), the Red Sea. For more than a
decade, earthen sedimentation ponds were used to miti-gate pollution from fishpond effluents. The ponds wererich with microalgae, thanks to the plentiful sunlightand the nutrient-rich water. As they grew, the microal-gae removed various forms of nutrients, introduced into the seawater by the fish culture. The microalgae,in turn, were harvested by clams and oysters, whichclarified the water and provided additional cash crops(Hughes-Games, 1977; Shpigel & Fridman, 1990; Sh-pigel & Blaylock, 1991; Shpigel et al., 1993a).

342
An unforeseen problem developed when the facilitylined simple earthen ponds with PVC (polyvinyl chlo-ride). The PVC lining prevents seepage of the seawaterinto the brackish aquifer. Oysters raised on water fromthe PVC lined ponds grew much slower than they hadin water from the un-lined earthen ponds (300 m2, 1m depth). Both types of ponds received water from thesame source of mariculture effluent, but their diatomand other flora were different (Shpigel et al., 1993b).Pennate diatoms consistently dominated the flora of theearthen ponds, while blooms of different phytoflagel-lates and centric diatoms fluctuated widely in the plas-tic lined ponds. Levels of nutrients and chlorophyll ain both types of ponds were similar. Apparently, theearthen ponds supplied some limiting nutrient(s) notavailable to the diatoms when the ponds were lined withPVC and covered with a thin layer (1–2 cm) of soil. Thediatom species that grew in these hypertrophic habitatswere uncommon in the flora of the region of the Gulf ofEilat from which they were recruited (Lee et al., 1988,1989; Shpigel et al., 1993b). Since the physical char-acteristics of the earthen and PVC-lined ponds wereidentical (size, depth, flow rates, climate), and the wa-ter flowing into both ponds came from the same source,it was of interest to find out the causes for the change inflora. Boyd’s (1995) thoughtful analysis of interactionsbetween bottom soils and aquaculture ponds had notyet been published, however the literature was repletewith suggestions. It seemed unlikely that any chem-ical resource may limit algal grown in intensive fish-pond effluents, yet Liebig’s “law of the minimum” (Theperformance [survival, growth and reproduction] of anorganism(s) is a function of the most limiting environ-mental factors) couldn’t be ignored. Long-establishedalgal growth media (e.g. Erdschreiber, ASP-7; Prova-soli et al., 1957; Guillard, 1975) were applied to thisquestion. Since C, N and P macronutrients were in ex-cess, the prime suspected chemical resources limitingdiatom growth in these ponds were the macronutrientsilicate (Werner, 1977; Doering et al., 1989; Brzezinskiet al., 1990; Sommer, 1994)—not enriched in fish feed,and the micronutrients Fe, Mn, Co, Mo, Zn and per-haps other trace elements (Green et al., 1991; Patrick,1977).
The aim of the study reported here was to test thehypothesis that micronutrients and/or dissolved Si be-came limiting for the diatoms in the PVC-lined sedi-mentation ponds and to apply the results to intensivemariculture practices. The studies began with batch cul-tures of axenic diatoms, self selected over a decade togrow in the original earthen ponds, then scaled up to
open mesocosms and advanced to trials in a new com-mercial farm.
Materials and methods
Isolation and identification
All diatoms were isolated from the earthen sedimen-tation ponds in the NCM, Eilat, Israel. The primaryisolation media were similar to those described in Leeet al. (1975), except that local seawater (salinity 40 ppt)was used. One per cent (final concentration) Provasoli’santibiotic mixture (Sigma P-8029, containing 12 000u penicillin, 50 µg chloramphenicol, 300 u polymyxinB, 60 µg neomycin mL−1) was added aseptically tothe sterile molten medium just before it was pouredinto the Petri plates. The samples that contained mix-tures of diatoms were aseptically spread on the surfaceof agar-solidified media in Petri plates. These plateswere incubated within plastic bags (to prevent desicca-tion) in front of a bank of fluorescent lights (60 µmolphoton m−2 s−1); 18 h light/6 h dark cycle) for 10days. Differences in diatom colony morphology guidedthe selection of colonies for further study (Lee et al.,1975). Diatom colonies and the underlying agar wereexcised with an alcohol-sterilized micro-spatula andaseptically transferred to Erdschreiber (Lee et al., 1975;Lee, 1993) and F/2 (Guillard, 1975) media. Both thesemedia are suitable for growing benthic diatoms. Af-ter growth of the primary isolation cultures in liquidmedium, the cultures were spread aseptically on agar-solidified medium without the antibiotic mixture tocheck for possible bacterial growth and the uniformityof diatom colonies. The cultures also were examined at1000× by phase contrast microscopy to check that theywere axenic. The pond systems are indeed agnotobiotic(unknown mixtures of organisms), but axenic (pure; notcontaminated with other organisms) cultures were usedso that there were tests of the actual needs of the diatomswithout interactions or competitions that take place inmixtures of organisms in batch cultures. Diatoms wereidentified with the aid of a scanning electron micro-scope (SEM). Diatoms were oxidized with hot H2O2
before filtering them through a 3-µm Nuclepore R© filter.Other oxidized preparations were mounted on micro-scope slides with Hyrax R© and examined by bright fieldand phase contrast microscopy. The extensive literaturecollection at the Diatom Herbarium of the PhiladelphiaAcademy of Natural Sciences was consulted during theidentification phase of this work.

343
We recognize that identification of diatom speciescan be a very thorny problem (Round, 1996). Some“species” have been very narrowly defined by theexamination of a population in one sample; other“species” have been described in broader terms afterexamining many populations. The diatoms isolatedwere from a hypertrophic environment that has notbeen extensively examined from a floristic point ofview. We attempted to link morphological charactersof the isolates to the closest described form withoutregarding ecological factors that might otherwise biastaxonomic decision. The isolates whose identitiesmay be new species are illustrated, but not formallydescribed.
Specimens of each species were mounted in Hyrax R©
on slides and deposited in the permanent collectionof the Philadelphia Academy of Sciences’ DiatomHerbarium. Cultures have been deposited with ATCC(American Type Culture Collection).
Trace requirements of isolates
Fully defined artificial seawater media (holidic; Leeet al., 1970), adjusted for higher salinity, were usedas the basis for designing test media. The ranges ofdissolved salts and nutrients found in various studies ofthe NCM sedimentation ponds by Gordin et al.(1990),Krom (1986, 1991), Krom and Neori (1989), Neori etal. (1989) and Krom et al.(1989) were guides in theconstruction of test media. The ranges of tests were asfollows: P (as NaH2PO4) 0–3 mM; N as (NH4NO3) 0–7 mM; Si (as Na SiO3) 0–70 µM; Fe (as FeSO4) 0–10µM; Mn (as MnCl2·4H2O) 0–30 µM; Co (as CoCl2)0–30 µM; Zn (as ZnCl2) 0–30 µM. Each additive wastested separately in end-point experiments, with theother components of the culture medium maintained atthe standard levels prescribed in the original mediumpreparation protocol.
There were six steps in each test range. Becausethe eventual goal was to demonstrate need in poorlydefined mariculture effluent, there was no attempt totitrate the precise maximum level of stimulation byeach nutrient or trace in these experiments. The mediaused in trace metal experiments were prepared fromreagent or analytical grade chemicals in polyethylenebeakers. Each beaker used was new, soaked with deion-ized water for 3 days, washed with 1N HCl and rinsedthree times with deionized water to remove any traceof metals remaining from the manufacturing process.
The media were prepared with deionized water.Each medium was sterilized by filtration through a
previously sterilized filter (Millipore R© 0.2 µm pore).Aseptic dispensing of the sterilized media into newsterile-disposable polystyrene test tubes was donewithin a laminar-flow hood. After inoculation, the testtubes were placed in racks that were then placed withintransparent plastic bags to retard vapor loss during longincubation. They were incubated at 22 ◦C for 10 days infront of a bank of fluorescent lamps (60 µmole photonsm−2 s−1; 18 L:6 D light–dark cycle). A hemacytometerwas used to quantify each population after batch cul-ture for 10 days. The results of three replicates wereaveraged.
Trace requirements of axenic cultures in mediaformulated from mariculture effluent water
Mariculture effluent was used as the base for test mediain a second growth test series, to assess the results ob-tained with fully defined medium in batch laboratorycultures when applied to a milieu as rich and undefinedas intensive fishpond water. The effluent was sterilizedby filtration (Millipore R© 0.2 µm) and enriched withthe same nutrients and trace elements that stimulatedthe growth of each of the diatom strains in the fullydefined medium laboratory studies. With the exceptionof Si, the range of trace element supplements brack-eted the optimal concentrations found in the definedmedium study. Since most of the strains grew best atthe highest level of Si we tested (70 µM) we furtherraised the Si level in this test series to 100 µM. Alsotested were the extracts of local soils that might beused to supply the trace elements if they were appliedas a thin layer (1–2 cm) on top of the PVC liner. Thepreparation of soil extracts followed traditional phyco-logical methods (e.g. Provasoli et al., 1957). The soilscame from various sites in Eilat: (1) dredged sedimentand soil from a nearby lagoon excavation site; (2) thebottom of a previously used earthen sediment pond atthe NCM, and (3) fresh surface soil just east of theNCM.
Mesocosm (raceway) experiment
Mesocosm experiments were set up outdoors in sixlarge shallow oval fiberglass tanks (“raceways”). Eachraceway held 700 L of mariculture effluent sea water,had a surface area of 2.8 m2, and was partially dividedin the center, along the longer axis, by a rigid partition.Effluent from the sedimentation pond was pumped infrom above at one end and exited by a standpipe fromthe other end along the long axis. The raceways were

344
not stirred. Trace elements, when added, were drippedin near the water inlet. Six species of diatoms from thepond, relatively easily distinguished morphologicallyfrom each other in the light microscope (Cylindrothecaclosterium (cyc), Navicula cf lineola (9235), Navicu-lar salinicola (E94), Nitzschia laevis (X72), Amphoraluciae (XDA), and Amphora cf tenerrima variant 1(XJ3C)), were used as inocula for the raceways. Af-ter counting in a hemocytometer, 6–11 L of unialgalmass cultures, containing approximately 1 × 108 cellsof each species, were poured into each tank. Silicate(100 µM), Fe (20 µM) and Zn (as ZnSO4·7H2O; 7 nM)were added as enrichments (final concentrations) to theodd-numbered raceways (#1, #3 and #5). These tankswere each paired with an even-numbered (#2, #4 and#6) control raceway, which received no enrichments.The rate of effluent seawater flow through each race-way pair was varied from: 0.5 volume exchange per day(raceways #1 and #2); 1 exchange per day (raceways#3 and #4); and 2 volume exchanges per day (race-ways #5 and #6). The nutrient quantities dripped intothe odd numbered raceways were calculated based onthe mean daily water flow. A mixture of local soil andsand was added to the bottom of all raceways to a depthof 1 cm. After inoculation to the raceways, any diatomswere given two days to attach to the surface of the tank(without flow to prevent wash out).
The raceways were then sampled once a week forthree weeks. Four bottom samples: (A) below efflu-ent seawater inlet; (B & D) middle of each racewayon either side of the divider; (C) near the outlet stand-pipe; (E) water surface of each raceway. Once browndiatom rafts appeared at the surface, usually after afew days, they were also sampled (designated as F).The bottom samples were taken as follows: a plasticcylinder 5 cm in diameter (19.63 cm2) was pushed intothe sediment. Then a 50-mL syringe with a 1-cm di-ameter PVC tubing extension was used to suck off thesurface sediment within the ring formed on the sedi-ment by the cylinder. The contents that were suckedoff were placed in a 20-mL liquid scintillation vial forfurther processing. After allowing the diatoms to settlefor 1 h the overlaying seawater was decanted. Acetonewas added to disrupt cells and extract pigments. Thecells were then rinsed with distilled water, oxidizedwith hot H2O2, and then filtered onto 3-µm porosityPoretics R© or Nuclepore R© filters and mounted on stubs.They were sputter coated with 10 nm Pd, and exam-ined and counted in an SEM. Data were collected onExcel spreadsheets and ANOVA was performed usingstatview v 4.1 for the Macintosh.
Disposable test tubes were used to take 10 mL sam-ples of the water columns in each raceway; sub sampleswere examined alive in a phase contrast microscopeand processed as above for examination in an SEM.The live water samples permitted enumeration of soft-bodied protists and prokaryotes that would have beenoxidized in preparations of diatoms for the SEM.
When super-abundant populations of diatoms devel-oped in the raceways, several days after inoculation, theraceways were stocked with clams (Tapes sp) to controlthe diatom populations. Plastic dividers were placed inthe raceways to create 1-m2 and 1.8-m2 sectors. Smallclams (average wet weight 0.11 g) were evenly dis-tributed in the 1.8-m2 sectors at a density of 3000 m−2
(5400 clams; total wet weight 5.8 g).Three experiments of the same type were done in
the raceways. Flow in the second and third experimentspermitted one exchange per dayin all raceways. In thesecond experiment, two raceways were enriched withZn and Si, two with only Si, while the controls werenot enriched. In the third experiment three racewayswere enriched with Fe, Zn and Si at the same concen-trations as in the first experiment, and three racewayswere enriched only with Si.
Practical field tests
A simple pre-commercial pilot experiment was carriedout directly in a PVC lined and soil-covered sedimen-tation pond (300 m2× 1 m deep). Two pairs of 30-Lplastic cylinders were sunk several centimeters intothe soil at the pond’s up-wind and down-wind ends(north and south). The rims of the cylinders projectedinto the water column about 6 cm above the surface ofthe soil. Each of the cylinders was inoculated with amixture of approximately 1 × 108–1 × 109 cells eachof the six diatom species described above. In addition,an enrichment mixtures was pipetted into one cylinderof each pair with Si, Fe and Zn at concentrations cal-culated to meet the trace requirements of the diatomsin the cylinders with one water exchanges per day.
One week after inoculation, samples were taken asdescribed above from the bottoms within the four cylin-ders, the plastic surfaces of the cylinders, and fromtwo control-bottom sites away from the cylinders. Thesamples were observed fresh by phase microscopy andprepared for later observation in the SEM as describedabove. The samples received only qualitative marks aspoorly or richly populated by the diatom species.
Based on the results of this study, a larger pre-commercial enrichment program was applied twice in

345
a new commercial fish farm (PGP) nearby. Two iden-tical PVC-lined sedimentation ponds (each 300 m2×1 m deep)receiving mariculture effluent were used.Both were inoculated with 20 L each of dense pop-ulations of the diatoms we had been working with plusthe prasinophyte Tetraselmis marina. One of the pondswas also enriched with Si to a final concentration of200 µM.
Diatom growth and recruitment potential
To assess their potential for diatom growth and the rapidestablishment of a stable diatom flora without inocu-lation, fishpond effluent samples (250 mL each) fromthe commercial pond were used in a laboratory exper-iment. Two hundred fifty milliliters in 500-mL flaskswere enriched with 200 µM Si and then inoculatedwith the same diatom mixture. ‘Positive’ controls wereSi enriched but without inoculation with the diatommix. A second set of positive controls was not enrichedwith Si nor was it inoculated with the diatom mixture.‘Negative’ controls were inoculated with the diatom-mix but not Si-enriched. The experimental flasks wereincubated in triplicate in the lab in front of a bank offluorescent lights (60 µmol photons m−2 s−1).
Analytical methods
Silicate was analysed by an automated system (Techni-con Autoanalyzer II), using the method of Glibert andLoder (1977).
Results
Identification
The diatoms were identified with the aid of a scan-ning electron microscope as: A. luciae (Cholnoky)emend. Archibald 1983, clones I (XDA), II(XDB), III(X8A), IV(926), & V (8B13); A. cftenerrima Aleemand Hustedt 1951; clones I (EN), II (XJ3C) & III(AR1); C. closterium (Ehr.) Reimann and Lewin 1964,(CYL);N .cf lineola Grun. 1884 clones I (9235) & II(9249); N .cf lenzii Hustedt 1939 (XPA); N. salinicola(Hustedt, 1939), Clones I (E94) & II (D14); N .cf vimi-noides Giffen, 1975 clones I (XD32) & II (D4); N.laevis Hustedt 1939, clones I (X72) & clone II (939)(Figures 1–13).
The water column also contained some of thesesame species of diatoms as well as phytoflagel-lates, some introduced by mariculture activities (e.g.
Isochrysis sp, Tetraselmis sp) and others recruited fromthe seawater drawn in to the facility. Of the latter,Amphidinium, Gymnodinium, Dunaliella and Eutrep-tia were the most common genera.
Axenic fully-defined laboratory experiments
The level of test nutrient in these end-point experimentsthat had the highest cell population (average of threereplicates) after 10 days incubation was noted (typi-cally at the end of exponential growth phase as ob-served after initial isolations and transfers in ERD orF/2). There were marked differences betweeen clonesin their response to stimulation by a particular element;10–100-fold differences in the concentration leadingto the highest population size were common (e.g. A.luceae and N . cf viminoides) (Table 1).
P
All strains benefited from P enrichment. The highestcell densities of most (15 clones: XJ3C, XDA, XDB,926, AR1, 8B13, X72, 929, E94, A201, D4, D14, CYL)were found in media supplemented with 32 µM P.(Table 1). Two other isolates (X8A, 9235) grew bestat the lowest P enrichment (0.32 µM P) tested. Oneisolate (XJA1) grew best 3.2 µM P. A 10-fold higherP enrichment (3.2 mM P) inhibited three isolates (926,En, 9249), while 32 mM inhibited the growth of all theisolates.
N (as NH4NO3)
All but three isolates needed N enrichment. Eleven ofthem (XJ3C, XDA, 8B13, 939, XJA1, 9235, E94, 9249,A201, D14, CYL) grew best in medium supplementedwith 714 µM N (Table 1). Three (926, X8A, E94) hadhigher densities with ten times more (7.14 mM) N.Highest density of another isolate (XPA) was found at7.14 µM (Table 1). Two isolates (AR1, D4) grew bestat the lowest N enrichment tested (0.07 µM), and three(XDB, X72, XD32) preferred no enrichment.
Fe
The isolates varied in their ability to grow with differentlevels of enrichment with Fe (Table 1). Cell densitiesof one isolate (X8A) were highest in the medium with10 µM Fe. Most isolates grew better at higher concen-trations. For 11 isolates (XJ3C, XPA, AR1, X72, 939,XJA1, 9235, E94, A201, D14, CYL) enrichment with

346
Figure 1. (1) N .cf viminoides variant 2, scale = 1 µm; (2) N .cf viminoides, scale = 2 µm; (3) N. salinicola; scale = 5 µm; (4)N .cf lineola;scale = 2 µm; (5) N .cf lenzii, scale = 2 µm; (6) N. laevis, scale = 1 µm; (7) C. closterium; scale = 10 µm; (8) A.cf tenerrima variant 2;(9) A.cftenerrima variant 1; ;(10) A.cf tenerrima variant III;(11) A. luciae variant. II; (12) A. luciae variant, I; (13) A. luciae variant III,scale = 2 µm.
20 µM Fe was best (Table 1). For six other isolates(XDA, 926, 8B13, XD32, 9249, D4) enrichment with30 µM Fe was best (Table 1). Three isolates (XDB,EN, A201) grew best with no Fe enrichment.
Mn
The diatoms tested differed greatly in their responsesto the addition of Mn (as MnCl2·4H2O) to the medium(Table 1). Populations of four isolates (X8A, 8B13, D4,D14) were highest in media supplemented with 10 nMMn. Two isolates (AR1, A201) needed 10 µM Mn toreach their highest population sizes. Largest popula-tions of four isolates (XJ3C, XDA, X72, 9235) werein media supplemented with 20 µM Mn. Three iso-lates (En, E94, XD 32) had even greater requirements
(30 µM Mn.) Populations of four isolates (XPA, XJA1,9249, D4) were highest at 40 µM Mn.
Zn
The growth of the diatoms was also stimulated by zinc(Table 1). A low concentration (1 µM) was sufficientfor three isolates (XPA, XD32, 9249). Populations offive isolates (AR1, 8B13, 9235, D4, D14) were highestin 10 µM Zn. The population growth of three isolates(XJ3C, A201, CYL) was enhanced by 20 µM Zn inthe medium and three (XDB, EN, 939) were enhancedby 30 µM Zn. 100 nM Zn stimulated three isolates(XJA1, X72, E94), while 10 nM Zn stimulated twoothers (XDA, X8A).
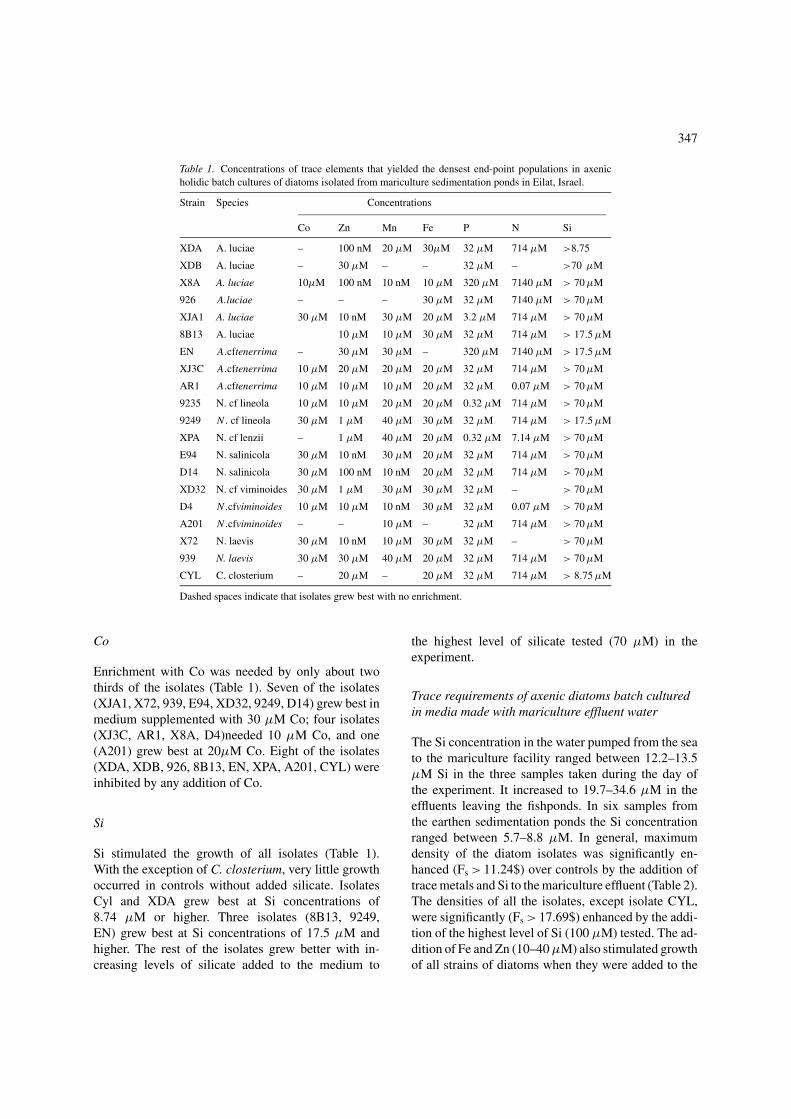
347
Table 1. Concentrations of trace elements that yielded the densest end-point populations in axenicholidic batch cultures of diatoms isolated from mariculture sedimentation ponds in Eilat, Israel.
Strain Species Concentrations
Co Zn Mn Fe P N Si
XDA A. luciae – 100 nM 20 µM 30µM 32 µM 714 µM >8.75
XDB A. luciae – 30 µM – – 32 µM – >70 µM
X8A A. luciae 10µM 100 nM 10 nM 10 µM 320 µM 7140 µM > 70 µM
926 A.luciae – – – 30 µM 32 µM 7140 µM > 70 µM
XJA1 A. luciae 30 µM 10 nM 30 µM 20 µM 3.2 µM 714 µM > 70 µM
8B13 A. luciae 10 µM 10 µM 30 µM 32 µM 714 µM > 17.5 µM
EN A.cftenerrima – 30 µM 30 µM – 320 µM 7140 µM > 17.5 µM
XJ3C A.cftenerrima 10 µM 20 µM 20 µM 20 µM 32 µM 714 µM > 70 µM
AR1 A.cftenerrima 10 µM 10 µM 10 µM 20 µM 32 µM 0.07 µM > 70 µM
9235 N. cf lineola 10 µM 10 µM 20 µM 20 µM 0.32 µM 714 µM > 70 µM
9249 N . cf lineola 30 µM 1 µM 40 µM 30 µM 32 µM 714 µM > 17.5 µM
XPA N. cf lenzii – 1 µM 40 µM 20 µM 0.32 µM 7.14 µM > 70 µM
E94 N. salinicola 30 µM 10 nM 30 µM 20 µM 32 µM 714 µM > 70 µM
D14 N. salinicola 30 µM 100 nM 10 nM 20 µM 32 µM 714 µM > 70 µM
XD32 N. cf viminoides 30 µM 1 µM 30 µM 30 µM 32 µM – > 70 µM
D4 N .cfviminoides 10 µM 10 µM 10 nM 30 µM 32 µM 0.07 µM > 70 µM
A201 N .cfviminoides – – 10 µM – 32 µM 714 µM > 70 µM
X72 N. laevis 30 µM 10 nM 10 µM 30 µM 32 µM – > 70 µM
939 N. laevis 30 µM 30 µM 40 µM 20 µM 32 µM 714 µM > 70 µM
CYL C. closterium – 20 µM – 20 µM 32 µM 714 µM > 8.75 µM
Dashed spaces indicate that isolates grew best with no enrichment.
Co
Enrichment with Co was needed by only about twothirds of the isolates (Table 1). Seven of the isolates(XJA1, X72, 939, E94, XD32, 9249, D14) grew best inmedium supplemented with 30 µM Co; four isolates(XJ3C, AR1, X8A, D4)needed 10 µM Co, and one(A201) grew best at 20µM Co. Eight of the isolates(XDA, XDB, 926, 8B13, EN, XPA, A201, CYL) wereinhibited by any addition of Co.
Si
Si stimulated the growth of all isolates (Table 1).With the exception of C. closterium, very little growthoccurred in controls without added silicate. IsolatesCyl and XDA grew best at Si concentrations of8.74 µM or higher. Three isolates (8B13, 9249,EN) grew best at Si concentrations of 17.5 µM andhigher. The rest of the isolates grew better with in-creasing levels of silicate added to the medium to
the highest level of silicate tested (70 µM) in theexperiment.
Trace requirements of axenic diatoms batch culturedin media made with mariculture effluent water
The Si concentration in the water pumped from the seato the mariculture facility ranged between 12.2–13.5µM Si in the three samples taken during the day ofthe experiment. It increased to 19.7–34.6 µM in theeffluents leaving the fishponds. In six samples fromthe earthen sedimentation ponds the Si concentrationranged between 5.7–8.8 µM. In general, maximumdensity of the diatom isolates was significantly en-hanced (Fs > 11.24$) over controls by the addition oftrace metals and Si to the mariculture effluent (Table 2).The densities of all the isolates, except isolate CYL,were significantly (Fs > 17.69$) enhanced by the addi-tion of the highest level of Si (100 µM) tested. The ad-dition of Fe and Zn (10–40 µM) also stimulated growthof all strains of diatoms when they were added to the

348
Table 2. Nutrient enrichments to mariculture effluent that yielded the densest end-pointpopulations in axenic batch cultures of diatoms isolated from sedimentation ponds.Dashes indicate that isolates grew best with no enrichment.
Soil Fe Zn Mn Co SiDiatom strain extract (µM) (µM) (µM) (µM) (µM)
XJ3C Amph Bottom 30 30 40 10 100
8B13 East 30 10 10 – 100
9235-Nav Bottom 30 10 – 10 100
A201-Nav East 10 20 – 30 100
9249-Nav Canal 40 20 – – 100
D4-Nav East 20 10 30 30 100
X72-Nit. L – 30 30 – 20 100
939-Nit East 40 30 40 40 100
XJA1-Amph East 30 20 30 – 100Cyl bottom 30 40 – – 100
effluent (Fs > 3.89). Extracts of the surface soil justeast of the NCM was the most effective stimulant forhalf of the strains tested. Extract of soil from the bot-tom of a dried earthen sedimentation pond was the beststimulant for three isolates and the population size ofone isolate was not enhanced by any of the soil ex-tracts. Four of the isolates (9235, A201, 9249, X72) nolonger were stimulated by the addition of Mn, and two(9249 and XJA1) were no longer were stimulated bythe addition of Co when it was added to maricultureeffluent.
Mesocosm (raceway) experiment—visual andqualitative light microscopic observations
Two days after inoculation the sediment layers inall of the tanks had a golden-brown color and therewere large (>2 cm2) golden-brown floating rafts ofdiatoms on the surface of the water. In the early af-ternoon, bubbles (most probably photosynthetically-generated oxygen) lifted sections of the diatom layersfrom the benthos and floated then to the water surface,where they formed brown floating ’rafts’. These wereabundant in the fertilized tanks (#1, #3, and #5) andsparse in the unfertilized tanks (#2, #4, and #6), andwere larger with higher rates of water exchange. Allthe six species inoculated were found in the rafts, butthe most common were species of Navicula and Am-phora (later observations in the TEM showed that onlyone of the two species of Amphora inoculated repro-duced in the raceways). Some A. eilatiana (Lee et al.,2003a,b) and non-pigmented flagellates were found inthe water column and the benthos.
After a week of growth in the raceways, there werevisible clumps or heaps of diatoms under the spotswhere the fishpond effluent and Si/trace metals solu-tions were dripped in the fertilized tanks (#1, #3, and#5). There was also a large amount of granular materialaround the diatoms, identified by an electron diffractionX-rays (EDX) attachment of the SEM (data not shown)as Si-rich, with presence of Ca, Al, Fe, and a smallamount of K. The clams we had introduced into theraceways were dug into the sediment with siphons onthe surface. Tracks marking clam movement and possi-ble diatom siphoning were observed on the surfaces ofraceways #2, #4 and #6; they were less conspicuous inthe fertilized raceways #1, #3 and #5. Apparently, withlarger standing crops of diatoms in the fertilized race-ways, the animals needed to forage over smaller areas.Six clams from each raceway were sacrificed and thecontents of their guts were examined by phase contrastmicroscopy. The gut contents of the clams in unfer-tilized raceway #2 (unfertilized, 1/2 exchange per day)were largely bacteria. In addition to bacteria there wereincreasing numbers of diatoms in the guts of clams har-vested from unfertilized raceways #4 and #6, respec-tively, in apparent correspondence with the increasingwater exchange rates in these raceways (1 and 2 ex-changes per day, respectively). The guts of the clamsharvested from the fertilized raceways #1, #3 and #5were all filled with diatoms.
Changes in the benthic assemblages took placeduring the study. The microalgal assemblage in thebottom of raceway #2 was dominated by A. eila-tiana,Tetraselmis and Dunaliella sp. The side of thesame tank had stalked Tetraselmis and the same

349
flagellates as on the bottom of the tank. In tank #4a population of C. closterium formed a mat on thewalls of the tank. The flora on the benthos of race-way #6 was mainly Dunaliella sp., A. eilatiana and C.closterium; small populations of Navicula sp, Amphorasp and Nitzschia sp also were present. The bottom andwalls of the fertilized raceways #3 (1 exchange per day)and #5 (2 exchanges per day), on the other hand, werecarpeted with large populations of Navicula, Amphoraand Nitzschia. Small numbers of harpacticoid cope-pods, nematodes, ciliates and amphipods were also ob-served in the fresh samples.
Very few (<<1 cm2)A. eilatiana were present inraceway #3 or #5. The clams in raceways #3 and #5added to their wet weight an average of 0.7% perday during the three weeks of the experiment. The re-sults of the second and third series of raceway experi-ments (data not shown) were qualitatively similar to thefirst.
Mesocosm (raceway) experiment-quantitativeexamination of surface samples from each racewaymade by the SEM
Data from SEM of the oxidized diatom frustules (Table3) were in agreement with the qualitative observationsmade on fresh samples with the aid of phase contrastmicroscopy. After three weeks the density of diatomson the surfaces of the fertilized raceways (#1, #3, #5;18–29 × 106 cells cm2) was higher by about an or-der of magnitude than their density in the unfertilizedones (#2, #4, #6; 7 × 105 cells cm−2–18 × 105 cellscm−2) (Table 3). At the same time, the numbers of di-atom species identified were 6, 5 and 4 in the fertilizedraceways #1, #3 and #5, respectively. The overall num-bers of diatoms decreased with time in the unfertilized
Table 3. Average density of diatoms (1 × 106 cm−2) on surface of the raceways after incubation for three weeks*.
Raceway N. salinicola N. viminoides A. tenerrima. A. luciae C. closterium N . laevis Other Total
1 1. 8 ± .8 6 ± 0.2 9.62 ± 0.5 0.12±0.05 0.04 ± 0.3 1.48±0.4 < 0.01 19.162 0.4 ± .2 0 ± 0.01 0.24 ± 0 0 0.04±0.2 < 0.01 0.683 5.24 ± 0.1 3.28 ± 0.3 8.2 ± 0.3 0 0.4±0.1 0.9±0.2 < 0.01 17.644 0.32 ± 0.1 0.72 ± 0.1 0.32 ± 0.01 0 0 0 < 0.01 1.365 10 ± 0.7 0.2 ± 0.02 18 ± 3 0 0 1±0.2 < 0.01 29.26 0.5 ± 0.1 0.04 ± 0.01 0.8 ± 0.1 0 0.24 ± 0.3 0.24±0.1 < 0.01 1.82
∗Average of five 19.63 cm2samples from the bottom surfaces of different sections within each raceway. Samples were filteredon to 3.0-µm Nuclepore R© or Poretics R© filters and observed at 2000 × in an SEM. Fifty randomly selected fields from eachof the five samples from each raceway were counted (mean of 250 fields and SD).The rate of pond sea water inflow was varied for each raceway pair: #1 and 2—1/2 volume exchanged daily; #3 and 4—1exchange daily; #5 and 6—2 exchanges daily.
raceways (from an initial inoculum containing approx-imately 1 × 108 cells per species × six species × 2.8m−2= 2.1 × 105cells cm−2), and increased in the fer-tilized raceways, even though the diatoms were beingheavily cropped by the clams. There were no differ-ences in the diatom species composition between sec-tors with clams, and sectors with out them. Diatom pop-ulations in the fertilized raceways multiplied to a sta-ble level (∼20 × 106 cells cm−2) by the second week.The growth of two species, N. salinicola and A.cf ten-errima, were favored by the fertilization and the rapidexchange of effluent (raceway #5). The opposite wastrue for the isolate of A. luciae. It was found in samplesonly from raceways #1 and #2 (fertilized or unfertil-ized; 1/2 exchange of mariculture effluent per day).During the course of the experiments, no species ofdiatom, which may have been recruited from the mar-iculture effluent, became established in large numbersin the tanks. Small numbers (<< 1 cm2) of Gyrosigmasp. were the most common of the non-inoculated di-atom species.
Practical field tests
Copepods, nematodes, ciliates and amphipods werepresent in large numbers in all samples. One week af-ter starting the experiment, the fertilized cylinders hadabundant populations of diatoms and A. eilatiensis. N.laevis was the dominant diatom present; however therewere also large populations of N. salinicola andN .cflineola. In addition, Amphora spp., Gyrosigma andCylindrotheca were common. Samples taken just out-side the fertilized cylinders contained abundant popula-tions of A. eilatiensis and N. laevis. Inside the unfertil-ized cylinders the flora was dominated by A. eilatiana.All genera of diatoms inoculated were still present, but

350
in only moderate numbers. Samples taken outside, nextto the unfertilized cylinders, were also dominated byA. eilatiana. Some Cylindrotheca and an occasionalNavicula sp were also present. Control samples takenelsewhere in the pond contained mostly detritus withmeiofauna. A. eilatiana, Gyrosigma sp, Oscillatoriaand a large spherical cyanobacterium in modest num-bers. There were some patches of Gyrosigma, Beggia-toa and Oscillatoria.
Tests at commercial scale
The Si-fertilized water in the PVC-lined sedimentationpond (300 m3) of the commercial fish farm containeda 3.6 × 103mL−3 standing stock of diatoms withinthree days. The non-Si-fertilized but similarly inoc-ulated pond, on the contrary, was largely populated(0.4 × 102mL−3) with Gymnodinium sp, and Eutreptiasp. The diatoms in the fertilized pond were pumpedto racks of oysters (Crassostrea gigas Thunberg) thatsteadily cropped them. Fortuitously, from the point ofour research, the manager of the facility suspendedthe application of Si supplement for several months.Relatively rapidly the diatoms disappeared from thewater column and benthos of the sedimentation pondand the water became clear. Samples of the water hadlow numbers of phytoflagellates (Eutreptia, Gymno-dinium, Oxyhurris, and Tetraselmis) and no diatoms.Three days after reinoculating one pond with the di-atom mix and Tetraselmis and enriching the pond withSi it had a noticeable golden-brown color and densitythat proved, after microscopic examination, to be theoutgrowth of the diatoms we had inoculated. By 5 daysthe diatoms in the water column of the enriched pond(which we started pumping to the oysters) reacheddensities of 1 × 103mL−1 in the water column atmidday.
In the diatom-recruitment potential experiment, di-atoms in the flasks inoculated with diatoms and Si-enriched, developed dense populations quite rapidly.The flasks were noticeably brown in 3 days. On thecontrary, in one set of “positive” control flasks (with-out Si enrichment, no diatom inoculation) there wasno detectable diatom population even after a monthof incubation. In one flask there was noticeable pop-ulation of cyanobacteria. In one of the three the other“positive” controls (fertilized, not inoculated with thediatom mixture) there was a noticeable growth of C.closterium. In the “negative control” (inoculated withthe diatom mixture but enriched with silicate) there wasno noticeable growth of diatoms.
Discussion
In addition to growing well in high concentrations ofP and N, most of the diatoms that were self selectedto live in the mariculture effluent earthen maricultureponds, required high concentration of silica and tracemetals when in axenic culture. Although such batchculture laboratory results often do not find direct ap-plicability to agnotobiotic eutrophic mesocosms, thedata demonstrated that the test isolates had Si and tracemetal requirements and that there were ranges of nu-tritional need among the diatoms. The results of thetests to examine trace element needs of cultures grownin mariculture effluent were also as expected. At theoutset it was known that the diatoms from the NCMearthen fish ponds did not grow well in PVC linedponds. The results confirmed our hypothesis that theloss of trace elements and Si mobilized from the sed-iments in earthen ponds was probably underlying thechanges in microflora observed, when a PVC liningseparated the pond water from the earth underneath.The seawater that formed the effluent water introducedinto the sedimentation ponds was enriched with Si andtrace metals as it passes through the fish culture ponds,but not enough for the particular cohort of diatomstested. An earlier NCM investigation of earthen seawa-ter sedimentation ponds suggested that a steady supplyof silicate, at a Si/N ratio of about 2–4, could boostlasting diatom blooms and increase the production ofoysters (Krom, 1986; Shpigel & Blaylock, 1991). But,even without silicate enrichment, the growth rate of di-atoms in these earthen ponds was sufficient to supportthe production of a large yield of diatoms, which inturn, supported significant crops of bivalves (Hughes-Games, 1977).
The demonstration of a requirement for Si andsome trace metal requirements in laboratory holidicmedia and then in as ill-defined a milieu as maricul-ture effluent in batch culture, was then confirmed intwo open systems, mesocosms (700 L) and sedimen-tation ponds (300 m3). Our results suggested that sil-icate limited the growth of the benthic diatoms iso-lated from the earthen ponds in the facility. Evenwhen the PVC was covered with a 1–2 cm layer ofsandy soil on the surface, the Si enrichment in thefishpond was not sufficient in relation to enrichmentin the other macronutrients. Some mitigation of thisdeficit was achieved by acceleration of water exchange.This should be understood in light of the high Sirequirements of diatoms, equal to their requirementfor N.

351
Trace elements
The generally high requirements for trace elements bythe cohort of benthic diatom species we worked with isconsistent with present understanding of biogeochem-ical mineral cycling which takes place at the sediment-water column interface of earthen mariculture ponds(Boyd, 1995).. It is unlikely that the patterns of trace nu-tritional requirements of the benthic diatoms we stud-ied developed by chance (Brand et al., 1983). They areprobably adaptations to the highly specialized habi-tat(s) developed in the shallow, well-illuminated, buthypertrophic, mariculture waste treatment sedimenta-tion ponds.
The benthic diatoms tested had high (20–40 µM)Mn requirements. This fits with Patrick’s (1977) obser-vation that cyanobacteria will overgrow diatom com-munities if Mn content is maintained below 40 µgL−1 (0.73 µM). She found that Mn concentrationsabove 7.3 µM were toxic to diatoms. The high tol-erance to Mn exhibited by the diatoms from the sed-imentation ponds in the present study may suggestinternal detoxifying mechanisms such as phytoche-leatins (Ahner et al., 1995). The results of Co ex-periments are always difficult to evaluate. Cobalt isneeded not only for the synthesis of cyanocobalaminbut it can substitute for Zn (Price, 1990). Perhapsthe diatoms can synthesize the cyanocobalamin theyneed if they are supplied with Co, but they might takeup B12, as do some other algae (Venkatarman, 1965;Fries, 1961), as a preformed vitamin. In an environ-ment as rich as the mariculture sedimentation ponds,it is also possible for the diatoms to get this vitaminfrom uneaten fish food in the effluent or from bacterialsources.
Diatom growth enhancement by Si
Si, an essential element for the growth of diatoms (Dar-ley and Volcani, 1969), was clearly the main significantlimiting factor in the growth of the diatoms of the PVC-lined ponds, as demonstrated by the agnotobiotic con-tinuous flow mesocosm experiments. It often becomesthe first to be depleted in eutrophic coastal waters (It-tekot et al., 2000), because of vertical transport follow-ing diatom blooms. A number of studies have suggestedthat proper manipulation of N:Si ratios in experimentalsewage effluent treatment systems could produce phy-toplankton communities dominated by diatoms ratherthan less desirable algae (Officer and Ryther, 1980; Ry-ther and Officer, 1981). They regarded the latter com-
munities as being associated with the undesirable ef-fects of eutrophication, while the diatoms could be em-ployed as food for grazers in economically useful foodchains. Diatoms require N and Si in a ratio of approx-imately 1:1 (Redfield et al., 1963). Without Si enrich-ment, the growth of phytoplankton in well-illuminatedand aerated– diluted sewage will deplete Si well beforeN, leading to non-diatomaceous blooms (Dugdale andWilkerson, 1998). Enrichments by Si in un-enrichednatural waters (e.g. Menzel et al., 1963) also causes in-creases in diatom abundance. The extensive (48 trials)laboratory competition experiments with 11 species ofphytoplankton by Sommer (1994) seem to clarify someof the controlling factors in diatom and non-diatomcompetition. In general, light (as varied in the experi-ments reported) did not influence the competitive out-come of diatoms and non-siliceous flagellates. Diatomswere dominant at Si:N ratios > 25:1 while flagellateswere dominant at much lower ratios. These ratios weremuch higher than those reported in the sea (e.g. GermanBight; Radach et al., 1990) in the winter before springblooms, but as noted by Sommer (1994), the nutrientconditions there before spring blooms (approximately40 µM Si and 40µM N) are far from limiting at thestart of competition. Recent experiments with meso-cosms show the practical application of this type ofknowledge (Doering et al., 1989). Lowering the N:Siration from 14:1 to 1:1 in nutrient (N, P, Si) manipu-lated large (13 m3) mesocosms with, or without intactbenthic communities resulted in higher standing stocksof diatoms. A. eilatiensis, a toxic species that was notcompetitive with the diatoms used in this study at ratiosof Si:N > 1:1 (Lee et al., 2003a,b), developed in non Si-enriched mesocosms. This finding alone is a very im-portant reason for those who use sedimentation pond-grown algae to feed shellfish in red-tide prone habitatsto always consider adding Si to their integrated sys-tems. It is a matter for further consideration, whether Sienrichment could be applicable to combat HAB’s andpromote open water shellfish culture in relatively en-closed bays that are enriched by N and P from cage fishfarms.
The data suggest that soluble silica is the most nec-essary supplement for the promotion of healthy di-atom populations in seawater aquaculture effluents,particularly in ‘environmentally friendly’ PVC linedponds. This does not imply, of course, that the vari-able quality of the mariculture effluent with its or-ganic chelators and microbial flora can never lead totrace metal or vitamin limitations of diatom growth.In our experiments, the addition of trace elements

352
boosted the populations of certain species of diatomsby only 5–20% more, in the presence of high lev-els of Si. This may, or may not, be worth the ex-penditure of time and supplies in the overall eco-nomics of the secondary crop (clams and oyster)production.
The small recruitment experiment suggests that thisis an interesting area for future research. How longdid it take for the population of diatoms found in theearthen sedimentation ponds to be recruited? Consider-ing that the system is open, will the diatom assemblagesbe stable in the plastic-lined ponds over time? While itseems reasonable, that in time, chance may favor therecruitment of diatoms from the sea which will live inSi enriched mariculture effluent, it seems most prac-tical when starting up diatom production in a systemto inoculate it with the diatoms known to grow in thistype of system.
With the exception of C. closterium, none of thealgae we isolated are common on the shallow well illu-minated benthos, or as epiphytes, in the north westernGulf of Eilat. Interestingly, two of the species, N. laevisand Amphora cf tenerrima, have been isolated as en-dosymbionts from a number of diatom-bearing largerforaminiferan hosts (e.g. Heterostegina depressa, As-silina ammonoides, and Amphistigina spp.) living in thebenthos near the NCM and elsewhere on the NorthernGulf of Eilat coastline (Lee et al., 1989).
Acknowledgments
Supported by the Israeli Ministry for National In-frastructures (A.N., M.S. and O.Z.) and NIH/NIGMSgrant 08168-22 (J.J.L.). D. R. was supported in partby NIH/GMS GM MBRS grant 08168 (to J.J.L.). Wewish to thank Dr. Charles Reimer, Diatom Herbarium,Philadelphia Academy of Science, for his generousconsultations on the identification of the organismswith which we worked. He gave us complete accessto the extensive library and specimen collections of theDiatom Herbarium with out which we could not havecompleted this paper.
References
Ahner BA, Kong S, Morel FMM (1995) Phytochelatin production inmarine algae. 1. An intraspecies comparison. Limnol. Oceanogr.40: 649–657.
Boyd CE (1995) Bottom Soils, Sediment and Pond Aquaculture.Chapman and Hall, New York, 420 pp.
Brand LE, Sunda WG, Guillard RRL (1983) Limitation of marinephytoplankton reproduction rate by zinc, manganese and iron.Limnol. Oceanogr. 28: 1182–1198.
Brzezinski MA, Olson RJ, Chisholm SW (1990) Silicon availabilityand cell-cycle progression in marine diatoms. Mar. Ecol. Progr.Ser. 67: 83–96.
Darley M, Volcani B (1969) Role of silicon in diatom metabolism.A silicon requirement for deoxyribonucleic acid synthesis in thediatom Cylindrotheca fusiformis Reimann and Lewin. Exptl. CellRes. 58: 334–342.
DeVoe MR, Hodges CE (2002) Management of marine aquaculture:The sustainability challenge. In: Stickney RR, McVey JP (eds),Responsible Marine Aquaculture. CABI Publishing, Walling-ford, UK: pp. 21–44.
Doering PH, Oviatt CA, Beatty LL, Banzon VF, Rice R, Kelly SP,Sullivan BK, Frithsen JB (1989) Structure and function in a modelcoastal ecosystem: Silicon, the benthos and eutrophication. Mar.Ecol. Progr. Ser. 52: 287–299.
Fries L (1961) Vitamin requirements of Nemalion multifidum. Expe-rientia 17: 75–76.
Gordin H, Krom M, Neori A, Popper D, Porter C, Shpigel M (1990)Intensive integrated seawater fish ponds: Fish growth and waterquality. Eur. Aquaculture Soc. Special Publ. No. 11, pp. 45–65.
Greene RM, Geider RJ, Falkowski PG (1991) Effect of iron limitationon photosynthesis in a marine diatom. Limnol. Oceanogr. 36:1772–1782.
Guillard R (1975) Culture of phytoplankton for feeding marine in-vertebrates. In: Smith WL, Chaney MH (eds), Culture of MarineInvertebrate Animals. Plenum Press, New York: pp. 29–60.
Hughes-Games WL (1977) Growing the Japanese oyster (Cras-sostrea gigas) in sub-tropical seawater fishponds. I. Growth rate,survival and quality index. Aquaculture 11: 217–229.
Ittekot V, Humborg C, Schafer P (2000) Hydrological alterationsand marine biogeochemistry: A silicate issue? BioScience 50:776–782.
Krom MD (1991) Importance of benthic productivity in controllingthe flux of dissolved inorganic nitrogen through the sediment-water interface in a hypertrophic marine ecosystem. Mar. Ecol.Progr. Ser. 78: 163–172.
Krom MD (1986) Sources of water to the seawater well which sup-plies the fish ponds in Eilat: Implications for fishpond manage-ment. Wat. Res. 20: 463–469.
Krom MD, Neori A (1989) A total nutrient budget for an experimen-tal intensive fishpond with circularly moving seawater. Aquacul-ture 88: 345–358.
Krom MD, Neori A, van Rijn J (1989) Importance of water flow ratein controlling water quality in marine and freshwater fishponds.Israeli J. of Aquacult. Bamidgeh 41: 23–33.
Lee, JJ (1993) General techniques for the isolation and cul-ture of marine protists from estuarine, littoral, psammolit-toral, and sublittoral waters. In: Kemp PF, Sherr, BFSherr EB, Cole JJ (eds), Current Methods in Aquatic Mi-crobial Ecology. Lewis Publishers, Bocca Raton. pp. 41–50.
Lee JJ, Shpigel M, Freeman S, Zmora O, McLeod S, Bowen S, Pear-son M, Szostek A (2003a). Physiological ecology and possiblecontrol strategy of a toxic marine dinoflagellate from the benthosof a mariculture pond. Aquaculture 217: 351–371.
Lee JJ, Tietjen JH, Stone RJ, Muller WA, Rullman J, McEnery ME(1970) The cultivation and physiological ecology of members of

353
salt marsh epiphytic communities. Helgolander wiss. Meeresun-ters. 20: 136–156.
Lee JJ, McEnery ME, Kennedy E, Rubin H (1975) A nutritionalanalysis of epiphytic diatoms on Enteromorpha from a LongIsland salt marsh. J. Phycol. 26: 14–49.
Lee JJ, Erez J, ter Kuile B, Lagziel A, Burgos S (1988) Feedingrates of two species of larger foraminifera, Amphistegina lob-ifera and Amphisorus hemprichi from the Gulf of Elat (Red Sea).Symbiosis 5: 61–102.
Lee JJ, McEnery ME, ter Kuile B, Erez J, Rottger R, Rockwell RF,Faber WW Jr, Lagziel A (1989) Identification and distribution ofendosymbiotic diatoms in larger foraminifera. Micropaleontol-ogy 35: 353–366.
Lee, JJ, Olea R, Cevasco M, PochonX, Correia M, Shpigel M,PawlowskiJ (2003b) A marine dinoflagellate, Amphidinium eila-tiensis n. sp., from the benthos of a mariculture sedimentationpond in Eilat, Israel. J. Euk. Microbiol. 50: 439–448.
Menzel DW, Hulbert EM, Ryther JH (1963) The effects of enrichingSargasso Sea water on the production and species compositionof the phytoplankton. Deep Sea Res.10: 209–219.
Neori A, Krom MD, Cohen Y, Gordin H (1989) Water quality con-ditions and particulate chlorophyll aof new intensive seawaterfishponds in Eilat, Israel: Daily and dial variations. Aquaculture80: 63–78.
Officer CB, Ryther JH (1980) The possible importance of silicon inmarine eutrophication. Mar. Ecol. Progr. Ser. 3: 83–91.
Patrick R (1977) Ecology of freshwater diatoms and diatom commu-nities. In: Werner D (ed), The Biology of Diatoms. University ofCalifornia Press, Berkely, pp. 284–330.
Price, NM (1990) Cadmium and cobalt substitution for zinc in marinediatoms. Nature 344: 658–659.
Provasoli L, McLaughlin JJA, Droop MR (1957) The development ofartificial media for marine algae. Arch. Mikrobiol. 25: 392–428.
Radach G, Berg J, Hagmeier E (1990) Long-term changes betweenresource ratios and phytoplankton species composition duringspring in five north German lakes. Limnol.Oceanogr. 38: 846–856.
Redfield AC, Ketchum BH, Richards FA (1963). The influence oforganisms on the composition of seawater. In: Hill NM (ed.),The Sea. Vol. II, John Wiley, New York, pp. 26–77.
Round FE (1996) Opinion: What characters define diatom genera,species, and infraspecific taxa? Diatom Res. 11: 203–218.
Ryther JH, Officer CB (1981) Impact of nutrient enrichment on wateruses. In: Neilson BJ, Cronin LE (eds), Estuaries and Nutrients.Humana Press, Clifton, NJ, pp. 247–261.
Shpigel M, Lee J, Soohoo B, Fridman R (1993) Use of effluent waterfrom fish ponds as a food source for the Pacific oyster Cras-sostrea gigas Thunberg. Aquacult. Fisheries Manage. 24: 529–543.
Shpigel M, Neori A, Popper DM, Gordin H (1993) A proposed modelfor ”environmentally clean” land-based culture of fish, bivalvesand seaweeds. Aquaculture 117: 115–128.
Sommer U (1994) Are marine diatoms favored by high Si:N ratios?Mar. Ecol. Progr. Ser. 115: 309–315.
Stickney RR (2002) Introduction. In: Stickney RR, McVey JP (eds)Responsible Marine Aquaculture. CABI Publishing, Walling-ford, UK, pp. 1–20.
Venkataraman S (1965) The role of vitamin B12 in the metabolismof Euglena gracilis var bacillaria. Biochem. J. 96: 552–556.
Wells ML (1989) The availability of iron in seawater: a perspective.Biol. Ocean. 6: 463–476.




















