
Inhibition of nucleolar transcription as a trigger for neuronal apoptosis
Upload
independentCategory
view
4download
0
Nt(
RCS
a
ARRA
KNNMRMA
1
pT(ut(c(ncTiStas
0d
Micron 42 (2011) 87–96
Contents lists available at ScienceDirect
Micron
journa l homepage: www.e lsev ier .com/ locate /micron
ucleolar cycle and chromatoid body formation: Is there a relationship betweenhese two processes during spermatogenesis of Dendropsophus minutusAmphibia, Anura)?
ita Luiza Peruquetti ∗, Sebastião Roberto Taboga, Lia Raquel de Souza Santos,lassius de Oliveira, Maria Tercília Vilela de Azeredo-Oliveira
ao Paulo State University – UNESP/IBILCE, Department of Biology, Rua Cristovao Colombo, 2265 – Jardim Nazareth, CEP 15054 000 Sao Jose do Rio Preto, SP, Brazil
r t i c l e i n f o
rticle history:eceived 11 May 2010eceived in revised form 20 July 2010ccepted 23 July 2010
eywords:ucleolusucleolar cycle
a b s t r a c t
The goals of this study were to monitor the nucleolar material distribution during Dendropsophus minu-tus spermatogenesis using cytological and cytochemical techniques and ultrastructural analysis, as wellas to compare the nucleolar material distribution to the formation of the chromatoid body (CB) in thegerm epithelium of this amphibian species. Nucleolar fragmentation occurred during the pachytene ofprophase I and nucleolus reorganization occurred in the early spermatid nucleus. The area of the sper-matogonia nucleolus was significantly larger than that of the earlier spermatid nucleolus. Ultrastructuralanalysis showed an accumulation of nuages in the spermatogonia cytoplasm, which form the CB before
ale germ cell structureseproductioneiosis
mphibian
nucleolar fragmentation. The CB was observed in association with mitochondrial clusters in the cyto-plasm of primary spermatocytes, as well as in those of earlier spermatids. In conclusion, the nucleolusseems to be related to CB formation during spermatogenesis of D. minutus, because, at the moment ofnucleolus fragmentation in the primary spermatocytes, the CB area reaches a considerable size and isable to execute its important functions during spermatogenesis. The reorganized nucleolus of the earlierspermatids has a smaller area due to several factors, among them the probable migration of nucleolar
us to
fragments from the nucle. Introduction
The nucleolus is a distinct nuclear territory related to the com-artmentalization of nuclear functions (Hernandez-Verdun, 1991).he majority of ribosome biogenesis events, such as ribosomal RNArRNA) synthesis, processing and assembling of the ribosomal sub-nits, take place inside of the nucleolus (Cmarko et al., 2008). Whenhe nucleolus is observed using transmission electron microscopyTEM), the following three main nucleolar domains can be dis-erned: the fibrillar centers (FCs), the dense fibrillar componentDFCs) and the granular component (GCs). In these three differentucleolar compartments, ribosomal RNAs are synthesized, pro-essed and assembled with ribosomal proteins (Sirri et al., 2008).he main function attributed to the nucleolus is its participationn the ribosome biogenesis (Gerbi et al., 2003; Boisvert et al., 2007;
irri et al., 2008). In fact, most nucleolar proteins have several func-ions related to RNAr synthesis and processing, but some of themre related to other functions as well: nucleotide modifications ofeveral small RNAs, biosynthesis of the signal recognition parti-∗ Corresponding author. Tel.: +55 17 3221 2380; fax: +55 17 3221 2390.E-mail address: [email protected] (R.L. Peruquetti).
968-4328/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.micron.2010.07.009
the cytoplasm, and plays a part in the CB chemical composition.© 2010 Elsevier Ltd. All rights reserved.
cle and the phased sequestration and release of proteins involvedin gene silencing, senescence and cell division (Pederson, 2002).Moreover, some proteins that are involved in the control of the cellcycle, such as Net1, Cdc14 and Sir2, are located in the nucleolarsubcompartments (Shou et al., 1999; Straight et al., 1999; Visitinet al., 1999; Garcia and Pillus, 1999), along with the tumor sup-pressor proteins, ARF (Carmo-Fonseca et al., 2000), p53 and Myc(Montanaro et al., 2007). Furthermore, most of the Pch2 protein,which is required for the meiotic checkpoint that prevents chromo-some segregation when recombination and chromosome synapsesare defective, localizes to the nucleolus. These findings link thenucleolus, chromatin silencing, and the pachytene checkpoint (San-Segundo and Roeder, 1999).
Some authors believe that the fragmentation of the nucleolarmaterial is related to chromatoid body (CB) formation (Comingsand Okada, 1972; Andersen, 1978; Andonov, 1990; Peruquetti etal., 2008a,b). The CB is a typical cytoplasmic organelle of spermato-genic haploid cells and has a function related to RNA and protein
accumulation and/or reserves for later germ-cell differentiation(Söderström and Parvinen, 1976a; Saunders et al., 1992). In germcells of various organisms, dense fibrous material accumulatesinto a cytoplasmic structure and it has a cloud-like appearance,inspiring the term nuage, as the germ plasm or polar granules in
8 / Micr
DcivdcacoKtnd(trirCOsec
dtmDtkanyTafw
2
cdBnmDascat(pee
2
ad(sh
8 R.L. Peruquetti et al.
rosophila melanogaster and the P-bodies in the in some somaticells (Vasileva et al., 2009; Meikar et al., 2010). Some recent stud-es indicate that the CB is a highly specialized kind of nuage in theertebrate germ cells that may function as an intracellular focalomain that organizes and controls RNA processing in male germells. These reports suggest a model in which the CB functions assubcellular coordinator of different RNA-processing pathways,
entralizing post-transcriptional mRNA control in the cytoplasmf haploid male germ cells (Parvinen, 2005; Kotaja et al., 2006;otaja and Sassone-Corsi, 2007). However, some authors suggest
hat CBs have some aggresomal features, suggesting that they areot a synthetic site as proposed previously, but instead are a degra-ation site where unnecessary DNA, RNA, and proteins are digestedHaraguchi et al., 2005). There are many uncertainties related tohe function of this germ cell cytoplasmic structure and its originsemain unclear. In addition to the nucleolar origins that have beenndicated by other authors, other origins of this structure are cur-ently being researched. Some authors suggest that the origin of theB is nuclear (Parvinen and Parvinen, 1979; Parvinen et al., 1997).thers authors have proposed that the CB precursor is dense inter-
titial material occurring between mitochondrial clusters (Fawcettt al., 1970) or mitochondrial product that is released in the cellytoplasm (Reunov et al., 2000).
The goals of this study were to follow the nucleolar cycleuring the meiotic division, and verify the relationship betweenhe fragmented nucleolar material and CB formation in the sper-
atogenesis process. The study was completed using adult maleendropsophus minutus, a neotropical tree frog (Hylidae) species
hat exhibits continuous gametogenesis. Species that undergo thisind of gametogenesis have a partial interruption of spermatogenicctivity during some year season; however, primary spermatogo-ia are possibly under gonadotrophic stimulation during the wholeear (Lofts, 1974; Huang et al., 1997; Santos and Oliveira, 2007).he choice to use animals inhibiting continuous gametogenesis wasppropriate, because animals with germ epithelium activity wereound in all field incursions performed to collect the samples thatere evaluated in this study.
. Materials and methods
Testes from 10 adult male D. minutus (Amphibia, Anura) inalling activities collected during nocturnal field work at Talha-os (20◦45′47,5′′S and 49◦19′38,8′′W), in the state of Sao Paulo,razil, were processed for cytochemical analysis, for measuring theuclear and nucleolar areas of the spermatogonia and earlier sper-atids and for ultrastructural analysis. Testes from 15 adult male. minutus in calling activities collected during nocturnal field workt the Morro do Diabo State Park (Municipality of Teodoro Sampaio,tate of Sao Paulo, Brazil – 22◦32′43,7′′S; 52◦14′02,9′′W) were pro-essed for cytogenetic analysis. All the amphibians were collected,cclimatized in plastic bags, and transported to the laboratory athe Zoology and Botany Department of Sao Paulo State UniversityUNESP) in Sao Jose do Rio Preto, SP, Brazil. After the acclimatizingeriod, the animals were anesthetized and euthanized by ethylicther inhalation and their gonads were removed and prepared forach biological analysis.
.1. Cytochemical analysis
The testes of each animal were removed and immersed in
Bouin fixative solution for 24 h. The material was embed-ed in glycol–metacrylate historesin (Historesin Leica®). Sections1–3 �m thick) were obtained in Leica RM 2155 microtome. Tis-ue sections were submitted to one ordinary cytological procedure:ematoxylin–eosin – HE (Ribeiro and Lima, 2000); and various
on 42 (2011) 87–96
cytochemical procedures: toluidine blue (TB), modified CriticalElectrolyte Concentration for detecting RNA (CEC) (Mello et al.,1993), silver ion impregnation (AgNOR) (Howell and Black, 1980)and Feulgen reaction (Mello and Vidal, 1980). The sections of germepithelium were evaluated under an Olympus BX 60 photomicro-scope and documented using Image Pro-Plus; Media Cybernetics,version 6.1 for Windows computer software for image analysis.
In addition to the qualitative analysis of the nucleolar mate-rial distribution, the tissue sections that were subjected to silverion impregnation were used for quantitative analysis in order todetermine the number of nucleoli in the spermatogonia and earlierspermatids and also to measure the nuclear and nucleolar areas ofthe spermatogonia and earlier spermatids.
2.1.1. Determination of the number of nucleoli in thespermatogonia and earlier spermatids
The number of nucleoli was determined in the all spermatogoniaand earlier spermatids used in the present study. It were analyzed109.8 ± 6.14 spermatogonia and 133.2 ± 12.91 earlier spermatidsfrom each amphibian studied (n = 5). Due to the different amount ofthe cells analyzed in each cell type, after determining the absolutevalues, the percentage of the number of nucleolus in each cell typewas calculated.
2.1.2. Measuring the nuclear and nucleolar areas of thespermatogonia and earlier spermatids
The cells analyzed in advance were photo-documented usingan Olympus BX 40 photomicroscope and an Image Pro-Plus MediaCybernetics, version 4.5 for Windows computer software. Next,the nuclear and nucleolar areas of these cells were measuredusing Image J–Image Processing and Analysis in Java, Version 1.40(http://rsb.info.nih.gov/ij/) software for image analysis. The cellsthat had one single nucleolus were measured immediately; in cellsthat presented two or more nucleoli, each individual nucleolus wasmeasured before the calculating the total value.
2.1.3. Data analysisNormal distribution of data sets was tested using Skewness and
Kurtosis analysis (Ha and Ha, 2007), and variance homogenity wastested using the F max test (Zar, 1999). The number of nucleoli wascompared between spermatogonia and earlier spermatids, and alsowithin the same cell type using the Two-Factor Analysis of Vari-ance, complemented by the LSD multiple comparisons test (Zar,1999). Nuclear and nucleolar areas of spermatogonias were com-pared with nuclear and nucleolar area of earlier spermatids usingan independent t-test (Zar, 1999). Statistical significance was con-sidered when p ≤ 0.05.
2.2. Cytogenetic analysis
Standard meiotic cells of D. minutus were obtained using themethod introduced by Kligerman and Bloom (1977) and adapted byBertolo and Mestriner (1998). Soon after, the testes were sectionedinto small fragments and submitted to hypotonic treatment with0.075 M KCl for 20–30 min. The material was then treated with afresh fixative solution (methanol:glacial acetic acid, 3:1) for 30 min,and this procedure was performed a minimum of two times perfragment. After fixation, some fragments were placed in the well ofa depression slide with some drops of a 50% glacial acetic acid solu-tion and mixed until a homogeneous cell suspension was obtained.One drop of this suspension was then deposited onto a clean slide,
and heated to about 38 ◦C with the aid of a fine-tipped Pasteurpipette. The drop was then sucked back into the pipette, forminga cell ring measuring about 1 cm in diameter on the slide, withthe cells preferentially deposited on the border of the ring. Theselast two steps were repeated on 2–3 additional fields of the same
/ Micr
s(rpsv
2(
siMttigvwla9uC
2
AB
3
3
utcaTttaanpfRssmmndsiapnhoti
R.L. Peruquetti et al.
lide. Then, the slides were submitted to silver ion impregnationAgNOR) (Howell and Black, 1980) to follow the nucleolar mate-ial distribution during the meiotic division in the germ cells. Thereparations were evaluated under an Olympus BX 60 photomicro-cope and documented using Image Pro-Plus; Media Cybernetics,ersion 6.1 for Windows computer software for image analysis.
.3. Ultrastructural analysis – transmission electron microscopyTEM)
Testes fragments of each animal were removed and sliced intomall pieces, and samples of the germ epithelium were cut andmmersed in 3% glutaraldehyde plus 0.25% tannic acid solution in
illonig’s buffer (pH 7.3) containing 0.54% glucose for 24 h at roomemperature (Cotta-Pereira et al., 1976). After being washed withhe same buffer, samples were post-fixed in 1% osmium tetrox-de for 1 h at 4 ◦C, washed in Millonig’s buffer, dehydrated in araded acetone series and embedded in Araldite resin. Ultrathin sil-er sections (50–75 nm) were cut using a diamond knife and stainedith 2% alcoholic uranyl acetate for 30 min (Watson, 1958), fol-
owed by 2% lead citrate in sodium hydroxide for 10 min (Venablend Coggeshall, 1965). Samples were evaluated using a Leo-Zeiss06 (Cambridge, UK) transmission electron microscope and doc-mented using ITEM (Soft Image System–Veleta 2K x 2K TEM CCDamera) software for image analysis.
.4. Ethical note
The present study was approved by the Ethical Committee fornimal Research (CEEA), of Sao Paulo State University (UNESP) inotucatu, Sao Paulo, Brazil, under protocol no 057/06.
. Results
.1. Cytochemical analysis
The ordinary cytological technique hematoxylin–eosin (HE) wassed to analyze the male germ epithelium, and it revealed a pat-ern of cystic spermatogenesis and the spermatocystes showingells in all development stages (Fig. 1A). Germ cells were classifiedccording to the categories set up by Oliveira et al. (2002, 2003).he toluidine blue (TB) reaction allowed us to observe the cells ofhe male germ epithelium with an intense metachromasy in all ofhe nuclear structures (euchromatin, heterochromatin, nucleolusnd chromosomes) (Fig. 1C–E). The metachromasy degree variedccording to the genetic material compaction, the ploidy of the cellucleus, and the complexity of the nucleic acids with ribonucleo-roteic (RNP) corpuscles. The TB reaction was employed as a controlor the critical electrolytes concentration (CEC) variant method forNA detection (Fig. 1B). Tissue dyed using the CEC method showedpermatogonia with metachromatic nucleoli (Fig. 1F) and primarypermatocytes and earlier spermatids with fragmented nucleolaraterial (Fig. 1G and H). In the primary spermatocytes, the frag-ented nucleolar material was randomly distributed inside the
ucleus. Several ‘residual bodies’ or ‘chromatophilic spheres’ wereetected in the seminiferous epithelium lumen in touch to freepermatozoon (Fig. 1B). Feulgen reaction is a DNA-specific methodn which all germ cell nuclei are dyed purple, and are categorizedccording to ploidy degree, functional stage and chromatin com-action. Heterochromatic regions were more intensely died and
ucleolar material was observed as light regions connected to theseeterochromatic regions. In the spermatogonia (Fig. 1I), the nucle-lar area was larger than in earlier spermatids (Fig. 1K), indicatinghe occurrence of fragmentation and reduction of the nucleolar arean the last cell type. The nucleolar area of the germ cells was stronglyon 42 (2011) 87–96 89
impregnated using the AgNOR technique (Fig. 1L–N). Spermatogo-nia showed an evident nucleolus (Fig. 1L), primary spermatocytespresented a fragmented nucleolar region (Fig. 1M) and earlierspermatids (Fig. 1N) showed reorganized nucleolus, although itpresented a lesser volume than the spermatogonia nucleolus.
3.1.1. Determination of the number of nucleolus in thespermatogonia and earlier spermatids
There was no relation between the cell type and the number ofnucleoli (F = 0.732; p = 0.54), which means that the number of nucle-oli were the same before of the meiosis division (spermatogonia)and after the meiotic division (earlier spermatids). All the sper-matogonia and earlier spermatids analyzed presented 1–4 nucleoli.Nevertheless, there were differences in the number of nucleoli ineach cell type (F = 1536.71; p = 0.00). In both cell types, most of thecells presented single nucleoli (LSD, p < 0.00). The number of cellswith 2 nucleoli was higher than the number of cells with 3 or 4nucleoli (LSD, p < 0.00). The percentage of spermatogonia and ear-lier spermatids showing 3 or 4 nucleoli was similar (LSD, p = 0.52).There were no differences in the number of nucleoli between theanalyzed cell types (F = 0.00; p = 1.00) (Fig. 2A).
3.1.2. Measuring of the nuclear and nucleolar areas of thespermatogonia and earlier spermatids
There were significant differences between the nuclear areasin the spermatogonia and in the earlier spermatids (t = 6.68;p = 0.0001), and between the nucleolar area of the spermatogoniaand earlier spermatids (t = 17.25; p = 0.00). In the both findings, theareas of the spermatogonia were bigger than the areas of earlierspermatids (Fig. 2B).
3.2. Cytogenetics analysis
Standard meiotic preparations impregnated using the silverion method (AgNOR) showed spermatogonia mitotic metaphasewith 2n = 30 chromosomes (Fig. 3A). Some chromosome regionspresented a stronger impregnation with silver ion, indicating thepossible location of the nucleolar organizer regions (NORs). Earlierprimary spermatocytes (Fig. 3B) presented around 4 nucleolar cor-puscles. In primary spermatocytes in the zygotene stage (Fig. 3C)and in pachytene stage (Fig. 3D), a single central nucleolar cor-puscle was observed. In advanced meiotic stages, such as primaryspermatocytes in the initial diplotene stage (Fig. 3E) and in thediplotene stage (Fig. 3F), primary spermatocytes in the diaknesisstage (Fig. 3G) and metaphase II (Fig. 3H), no nucleolar corpuscleswere observed. Earlier and later spermatids were observed pre-senting 1–4 central nucleolar corpuscles (Fig. 3I); however, thesenucleolar areas were smaller than the nucleolar areas of the spe-matogonia.
3.3. Ultrastructural analysis – transmission electron microscopy(TEM)
Ultrastructural analyses were performed to qualitatively followthe fragmentation of the nucleolar material and the chromatoidbody (CB) formation during spermatogenesis of D. minutus. Sper-matogonia presented perfectly organized compact nucleoli andthe formation of some ribonucleoprotein nuages in the cytoplasmcould be discerned (Fig. 4A–C). The origin of these agglomerationsof the ribonucleoprotein material was probably related to somenuclear products that crossed the nuclear pore complex and accu-
mulated in the adjacent regions of the nucleus. In the nucleus ofthe primary spermatocytes the nucleolus was observed during thisnucleolar fragmentation process, and in the cytoplasm of this celltype the earlier CB structure was observed in association with mito-chondria aggregates (Fig. 4D, F and G). In the nucleus of the primary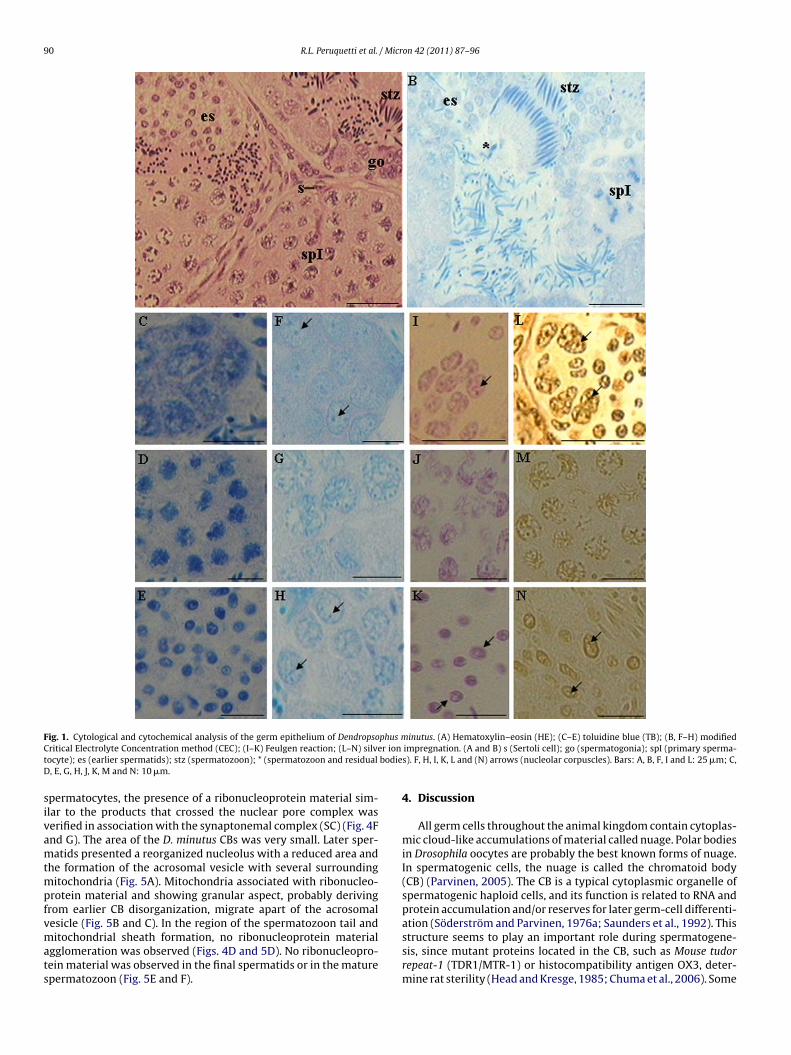
90 R.L. Peruquetti et al. / Micron 42 (2011) 87–96
F phus mC r iont bodieD
sivamtmpfvmats
ig. 1. Cytological and cytochemical analysis of the germ epithelium of Dendropsoritical Electrolyte Concentration method (CEC); (I–K) Feulgen reaction; (L–N) silveocyte); es (earlier spermatids); stz (spermatozoon); * (spermatozoon and residual, E, G, H, J, K, M and N: 10 �m.
permatocytes, the presence of a ribonucleoprotein material sim-lar to the products that crossed the nuclear pore complex waserified in association with the synaptonemal complex (SC) (Fig. 4Fnd G). The area of the D. minutus CBs was very small. Later sper-atids presented a reorganized nucleolus with a reduced area and
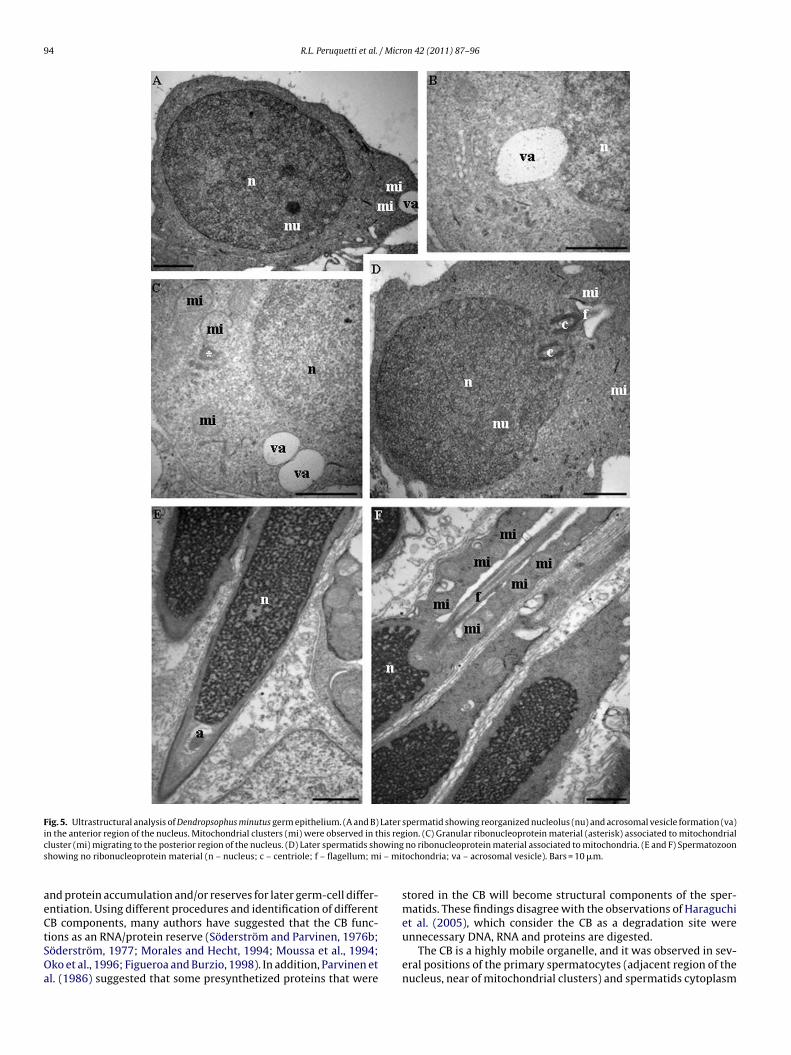
he formation of the acrosomal vesicle with several surroundingitochondria (Fig. 5A). Mitochondria associated with ribonucleo-
rotein material and showing granular aspect, probably derivingrom earlier CB disorganization, migrate apart of the acrosomal
esicle (Fig. 5B and C). In the region of the spermatozoon tail anditochondrial sheath formation, no ribonucleoprotein materialgglomeration was observed (Figs. 4D and 5D). No ribonucleopro-ein material was observed in the final spermatids or in the maturepermatozoon (Fig. 5E and F).
inutus. (A) Hematoxylin–eosin (HE); (C–E) toluidine blue (TB); (B, F–H) modifiedimpregnation. (A and B) s (Sertoli cell); go (spermatogonia); spI (primary sperma-s). F, H, I, K, L and (N) arrows (nucleolar corpuscles). Bars: A, B, F, I and L: 25 �m; C,
4. Discussion
All germ cells throughout the animal kingdom contain cytoplas-mic cloud-like accumulations of material called nuage. Polar bodiesin Drosophila oocytes are probably the best known forms of nuage.In spermatogenic cells, the nuage is called the chromatoid body(CB) (Parvinen, 2005). The CB is a typical cytoplasmic organelle ofspermatogenic haploid cells, and its function is related to RNA andprotein accumulation and/or reserves for later germ-cell differenti-
ation (Söderström and Parvinen, 1976a; Saunders et al., 1992). Thisstructure seems to play an important role during spermatogene-sis, since mutant proteins located in the CB, such as Mouse tudorrepeat-1 (TDR1/MTR-1) or histocompatibility antigen OX3, deter-mine rat sterility (Head and Kresge, 1985; Chuma et al., 2006). Some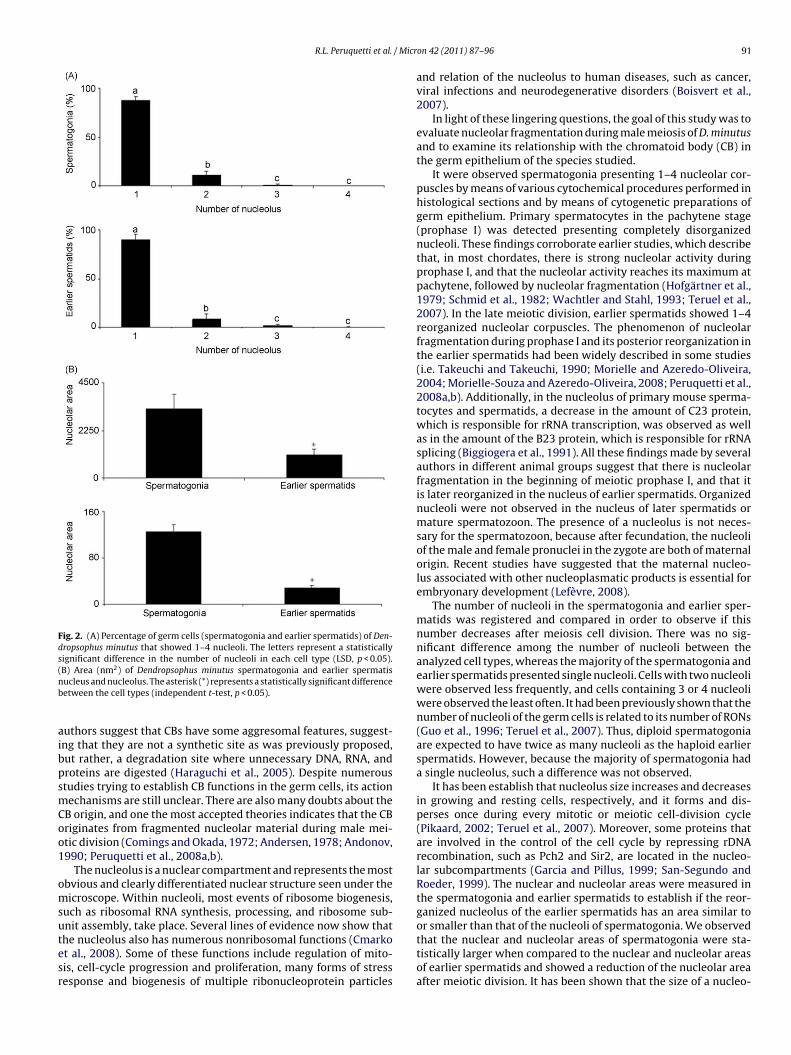
R.L. Peruquetti et al. / Micr
Fig. 2. (A) Percentage of germ cells (spermatogonia and earlier spermatids) of Den-dropsophus minutus that showed 1–4 nucleoli. The letters represent a statisticallysignificant difference in the number of nucleoli in each cell type (LSD, p < 0.05).(nb
aibpsmCoo1
omsutesr
or smaller than that of the nucleoli of spermatogonia. We observed
B) Area (nm2) of Dendropsophus minutus spermatogonia and earlier spermatisucleus and nucleolus. The asterisk (*) represents a statistically significant differenceetween the cell types (independent t-test, p < 0.05).
uthors suggest that CBs have some aggresomal features, suggest-ng that they are not a synthetic site as was previously proposed,ut rather, a degradation site where unnecessary DNA, RNA, androteins are digested (Haraguchi et al., 2005). Despite numeroustudies trying to establish CB functions in the germ cells, its actionechanisms are still unclear. There are also many doubts about the
B origin, and one the most accepted theories indicates that the CBriginates from fragmented nucleolar material during male mei-tic division (Comings and Okada, 1972; Andersen, 1978; Andonov,990; Peruquetti et al., 2008a,b).
The nucleolus is a nuclear compartment and represents the mostbvious and clearly differentiated nuclear structure seen under theicroscope. Within nucleoli, most events of ribosome biogenesis,
uch as ribosomal RNA synthesis, processing, and ribosome sub-nit assembly, take place. Several lines of evidence now show that
he nucleolus also has numerous nonribosomal functions (Cmarkot al., 2008). Some of these functions include regulation of mito-is, cell-cycle progression and proliferation, many forms of stressesponse and biogenesis of multiple ribonucleoprotein particleson 42 (2011) 87–96 91
and relation of the nucleolus to human diseases, such as cancer,viral infections and neurodegenerative disorders (Boisvert et al.,2007).
In light of these lingering questions, the goal of this study was toevaluate nucleolar fragmentation during male meiosis of D. minutusand to examine its relationship with the chromatoid body (CB) inthe germ epithelium of the species studied.
It were observed spermatogonia presenting 1–4 nucleolar cor-puscles by means of various cytochemical procedures performed inhistological sections and by means of cytogenetic preparations ofgerm epithelium. Primary spermatocytes in the pachytene stage(prophase I) was detected presenting completely disorganizednucleoli. These findings corroborate earlier studies, which describethat, in most chordates, there is strong nucleolar activity duringprophase I, and that the nucleolar activity reaches its maximum atpachytene, followed by nucleolar fragmentation (Hofgärtner et al.,1979; Schmid et al., 1982; Wachtler and Stahl, 1993; Teruel et al.,2007). In the late meiotic division, earlier spermatids showed 1–4reorganized nucleolar corpuscles. The phenomenon of nucleolarfragmentation during prophase I and its posterior reorganization inthe earlier spermatids had been widely described in some studies(i.e. Takeuchi and Takeuchi, 1990; Morielle and Azeredo-Oliveira,2004; Morielle-Souza and Azeredo-Oliveira, 2008; Peruquetti et al.,2008a,b). Additionally, in the nucleolus of primary mouse sperma-tocytes and spermatids, a decrease in the amount of C23 protein,which is responsible for rRNA transcription, was observed as wellas in the amount of the B23 protein, which is responsible for rRNAsplicing (Biggiogera et al., 1991). All these findings made by severalauthors in different animal groups suggest that there is nucleolarfragmentation in the beginning of meiotic prophase I, and that itis later reorganized in the nucleus of earlier spermatids. Organizednucleoli were not observed in the nucleus of later spermatids ormature spermatozoon. The presence of a nucleolus is not neces-sary for the spermatozoon, because after fecundation, the nucleoliof the male and female pronuclei in the zygote are both of maternalorigin. Recent studies have suggested that the maternal nucleo-lus associated with other nucleoplasmatic products is essential forembryonary development (Lefèvre, 2008).
The number of nucleoli in the spermatogonia and earlier sper-matids was registered and compared in order to observe if thisnumber decreases after meiosis cell division. There was no sig-nificant difference among the number of nucleoli between theanalyzed cell types, whereas the majority of the spermatogonia andearlier spermatids presented single nucleoli. Cells with two nucleoliwere observed less frequently, and cells containing 3 or 4 nucleoliwere observed the least often. It had been previously shown that thenumber of nucleoli of the germ cells is related to its number of RONs(Guo et al., 1996; Teruel et al., 2007). Thus, diploid spermatogoniaare expected to have twice as many nucleoli as the haploid earlierspermatids. However, because the majority of spermatogonia hada single nucleolus, such a difference was not observed.
It has been establish that nucleolus size increases and decreasesin growing and resting cells, respectively, and it forms and dis-perses once during every mitotic or meiotic cell-division cycle(Pikaard, 2002; Teruel et al., 2007). Moreover, some proteins thatare involved in the control of the cell cycle by repressing rDNArecombination, such as Pch2 and Sir2, are located in the nucleo-lar subcompartments (Garcia and Pillus, 1999; San-Segundo andRoeder, 1999). The nuclear and nucleolar areas were measured inthe spermatogonia and earlier spermatids to establish if the reor-ganized nucleolus of the earlier spermatids has an area similar to
that the nuclear and nucleolar areas of spermatogonia were sta-tistically larger when compared to the nuclear and nucleolar areasof earlier spermatids and showed a reduction of the nucleolar areaafter meiotic division. It has been shown that the size of a nucleo-

92 R.L. Peruquetti et al. / Micron 42 (2011) 87–96
Fig. 3. Germ cells of Dendropsophus minutus. Silver ion impregnation (AgNOR). (A) Spermatogonia in metaphase showing karyotype 2n = 30. Chromosomes showing nucleolarorganizer regions (NORs) strongly impregnated by silver ion (arrow). (B) Primary spermatocyte at the beginning of meiotic division presenting several nucleolar corpuscles( , presD laterH
l1ctrlltibgc
wmootaTnosPf
arrow). (C and D) Primary spermatocytes in zigotene and pachytene, respectivelyiakinesis and (H) Metaphases II, presenting no organized nucleolus. (I) Earlier andand I: 10 �m.
us is proportional to the amount of rRNA synthesized (Caspersson,950), that NOR size (the number of rRNA cistrons) is, in general,orrelated with its expression level (Shubert and Künzel, 1990),hat hypertrophy of the nucleolus is a state in which rRNA andibosome synthesis has increased (Nakamoto et al., 2001), and thatarge nucleoli may correlate with cell-division activity and with cel-ular stages having high protein demand (Mosgoeller, 2004). All ofhese findings may be related to the decrease of the nucleolus sizen the earlier spermatids, but its decrease can also be explainedy the migration of the nucleolar fragments to the cytoplasm oferm cells in prophase I, where these fragments may participate inhromatoid body (CB) formation.
In the present study, transmission electron microscopy (TEM)as used to verify the likely relationship between nucleolar frag-entation at the beginning of prophase I during spermatogenesis
f D. minutus and CB formation. Ultrastructural analysis showed therigins of CB ribonucleoproteic materials as they were migrating tohe cytoplasm of spermatogonia across the nuclear pore complexnd then as they accumulated in an adjacent region of the nucleus.he nucleolus of the spermatogonia is still organized; thus, it does
ot participate in initial CB formation. We can observe the CB totallyrganized in the cytoplasm of primary spermatocytes; however, iteems to be smaller than in other species CBs (Peruquetti, 2009,eruquetti et al., 2010), and in this stage it starts to perform itsunctions during spermatogenesis process. Nucleolus fragmenta-enting single nucleolar corpuscle (arrows). (E) Earlier diplotene, (F) Diplotene, (G)spermatids with reorganized nucleolus (arrows). Bars: A, C, E and F: 5 �m; B, D, G,
tion also occurs in the primary spermatocytes and a fraction ofthis fragmented material seems to move to the cytoplasm, andthis ribonucleoprotein material joins with other material that hasalready agglomerated in the primary spermatocyte cytoplasm toform the CB. The CB formation in D. minutus is different from CB for-mation in the freshwater turtle (Phrynops geoffroanus) (Peruquetti,2009) and in some mammals, such as rats (Rattus novergicus), mice(Mus musculus) (Peruquetti et al., 2008a), and Mongolian gerbils(Meriones unguiculatus) (Peruquetti et al., 2008b), because in thesespecies, the process of CB formation starts in the primary spermato-cytes phase rather than in the spermatogonia phase. Other authorshave also revealed the CB formation process occurring in the pri-mary spermatocyte stage (Tandler and Moriber, 1965; Kotaja andSassone-Corsi, 2007).
Some ribonucleoproteic material was observed near the synap-tonemal complex (SC) of the primary spermatocytes. It was similarto the material that moves from the nucleus to the cytoplasm,and plays a part in CB formation. This finding suggests that thisribonucleoprotein material that participates in the CB formationprocess may be related to the RNA reserve that is transcribed dur-
ing prophase I and will be translated during later spermatogenesis.This is a very important function proposed for the CB, because RNAtranscription occurs in the earlier spermatids, but protein trans-lation is required until the last steps of spermiogenesis (Monesi,1965). Because of this, the CB may have a function related to RNA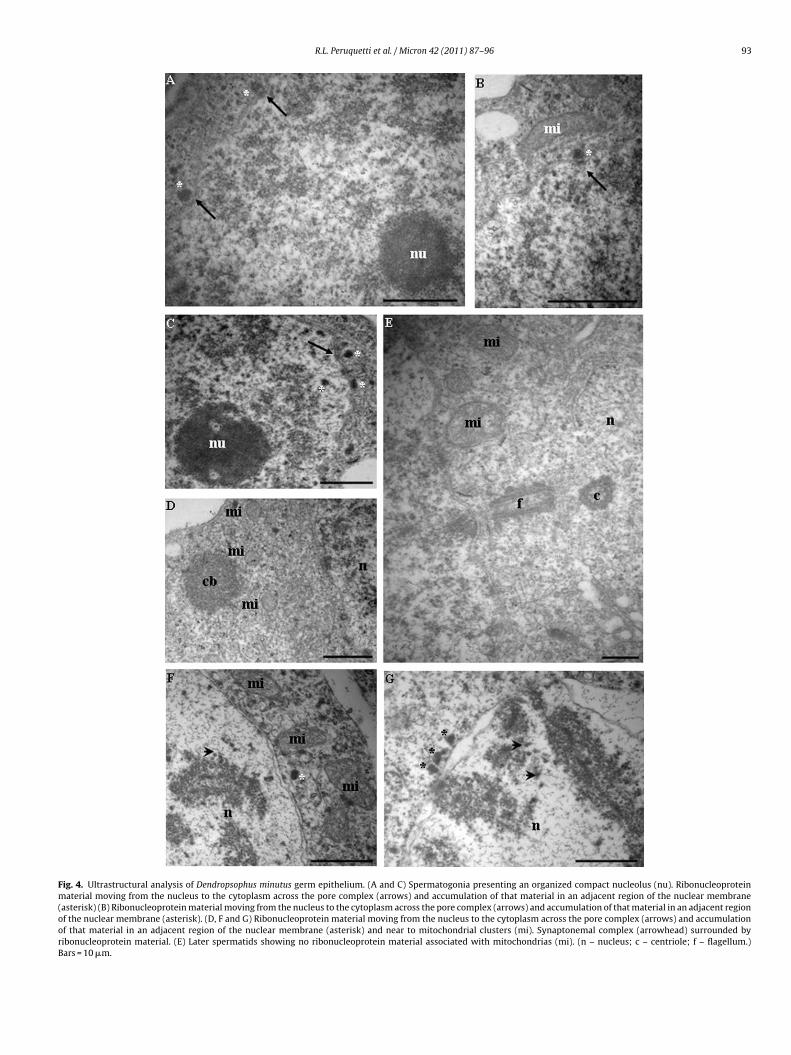
R.L. Peruquetti et al. / Micron 42 (2011) 87–96 93
Fig. 4. Ultrastructural analysis of Dendropsophus minutus germ epithelium. (A and C) Spermatogonia presenting an organized compact nucleolus (nu). Ribonucleoproteinmaterial moving from the nucleus to the cytoplasm across the pore complex (arrows) and accumulation of that material in an adjacent region of the nuclear membrane(asterisk) (B) Ribonucleoprotein material moving from the nucleus to the cytoplasm across the pore complex (arrows) and accumulation of that material in an adjacent regionof the nuclear membrane (asterisk). (D, F and G) Ribonucleoprotein material moving from the nucleus to the cytoplasm across the pore complex (arrows) and accumulationof that material in an adjacent region of the nuclear membrane (asterisk) and near to mitochondrial clusters (mi). Synaptonemal complex (arrowhead) surrounded byribonucleoprotein material. (E) Later spermatids showing no ribonucleoprotein material associated with mitochondrias (mi). (n – nucleus; c – centriole; f – flagellum.)Bars = 10 �m.
94 R.L. Peruquetti et al. / Micron 42 (2011) 87–96
Fig. 5. Ultrastructural analysis of Dendropsophus minutus germ epithelium. (A and B) Later spermatid showing reorganized nucleolus (nu) and acrosomal vesicle formation (va)i his regc owings i – mi
aeCtSOa
n the anterior region of the nucleus. Mitochondrial clusters (mi) were observed in tluster (mi) migrating to the posterior region of the nucleus. (D) Later spermatids shhowing no ribonucleoprotein material (n – nucleus; c – centriole; f – flagellum; m
nd protein accumulation and/or reserves for later germ-cell differ-ntiation. Using different procedures and identification of different
B components, many authors have suggested that the CB func-ions as an RNA/protein reserve (Söderström and Parvinen, 1976b;öderström, 1977; Morales and Hecht, 1994; Moussa et al., 1994;ko et al., 1996; Figueroa and Burzio, 1998). In addition, Parvinen etl. (1986) suggested that some presynthetized proteins that wereion. (C) Granular ribonucleoprotein material (asterisk) associated to mitochondrialno ribonucleoprotein material associated to mitochondria. (E and F) Spermatozoon
tochondria; va – acrosomal vesicle). Bars = 10 �m.
stored in the CB will become structural components of the sper-matids. These findings disagree with the observations of Haraguchi
et al. (2005), which consider the CB as a degradation site wereunnecessary DNA, RNA and proteins are digested.The CB is a highly mobile organelle, and it was observed in sev-eral positions of the primary spermatocytes (adjacent region of thenucleus, near of mitochondrial clusters) and spermatids cytoplasm

/ Micr
((1hacpt
cnt(obnbatcmiCtaorm(
torftatc
A
ThavhFJnFRs
R
A
A
A
B
B
B
R.L. Peruquetti et al.
anterior region of the nucleus) in this study. The presence of actinWalt and Armbruster, 1984) and calcium (Andonov and Chaldakov,989) may be related to the high mobility of this organelle. Theigh mobility of this organelle also was detected by Parvinen etl. (1997), who traced the path of the CB and analyzed its rapidlyhanging positions in relation to the nuclear envelope, Golgi com-lex and nuclear pale chromatin areas in living early spermatids ofhe rat.
The results of the present study also demonstrated a strongonnection between the CB and mithocondrial clusters. This con-ection had been previously described, and some authors proposedhat the origins of the CB could be from intermithocondrial materialFawcett et al., 1970). Other authors suggested that the CB has anrigin from nuclear genome products, and it is then supplementedy mitochondrial genome products (Reunov et al., 2000). The con-ection between the CB and mitochondria may also be explainedy the participation of the CB in the synthesis and transport of thepocytochrome cr, a cytochrome c isoform, which is expressed inhe testis tissue (Hess et al., 1993). In addition, the presence of theytochrome oxydase I (COXI) was detected, and encoded by theitochondrial genome in the CB (Haraguchi et al., 2005). Therefore,
n the present study, we suggest that the relationship between theB and mitochondria can be related to the migration of these struc-ures to the caudal nuclear region where the mithocondrial sheathnd spermatozoa tail are formed. This has also been suggested byther authors (Soley, 1994; Peruquetti et al., 2008a,b). CB mate-ial was not observed in the later spermatids, suggesting that thisaterial is dispersed and, possibly dissolved in the residual bodies
Sud, 1961; Yokota, 2008).In conclusion, the nucleolus seems to be related to CB forma-
ion during spermatogenesis of D. minutus, because, at the momentf nucleolus fragmentation in the primary spermatocytes, the CBeaches a considerable area and is able to execute its importantunctions during spermatogenesis. The reorganized nucleolus ofhe earlier spermatids has a smaller area due to several factors,mong them being the likely migration of nucleolar fragments fromhe nucleus to the cytoplasm, therefore playing a part in the CB’shemical composition.
cknowledgements
Special thanks to Dr. Tiago da Silveira Vasconcelos and to Dr.iago Gomes dos Santos (UNESP/IB – Rio Claro – SP – Brazil) for theirelp with the collection of the specimens used in the present study,nd to Dr. Tiago da Silveira Vasconcelos and Dr. Thaís Billalba Car-alho (UNESP/IBILCE – Sao Jose do Rio Preto – SP – Brazil) for theirelp with statistical analysis. Special thanks also to Mr. Luis Robertoaleiros, Jr. and M.Sc. Rosana Silistino de Souza (UNESP/IBILCE – Saoose do Rio Preto – SP – Brazil) for their help with laboratory tech-iques. The authors are indebted to FAPESP (Sao Paulo Researchundation – Grant number: 2007/04521-4) and CNPq (Brazilianesearch Council – Grant number: 141375/2006-0) for financialupport and fellowships.
eferences
ndersen, K., 1978. Fine structure of spermatogonia and spermatocytes in the bluefox (Alopex lagopus). Acta Veter. Scand. (Denmark) 19 (2), 229–242.
ndonov, M., 1990. Further study of the chromatoid body in rat spermatocytes andspermatids. Z. Mikrosk. Anat. 104, 46–54.
ndonov, M.D., Chaldakov, G.N., 1989. Morphological evidence for calcium storagein the chromatoid body of rat spermatids. Experientia 45, 377–378.
ertolo, L.A.C., Mestriner, C.A., 1998. The X1X2Y sex chromosome system in the fish
Hoplias malabaricus. II. Meiotic analyses. Chromosome Res. 6, 141–147.iggiogera, M., Kaufmann, S.H., Shaper, J.H., Gas, N., Amalric, F., Fakan, S., 1991.Distribuition of nucleolar proteins B23 and nucleolin during mouse spermato-genesis. Chromosoma 100, 162–172.
oisvert, F.M., van Koningsbruggen, S., Navascués, J., Lamnond, A.I., 2007. The mul-tifunctional nucleolus. Nat. Rev., Mol. Cell. Biol. 8, 574–585.
on 42 (2011) 87–96 95
Carmo-Fonseca, M., Mendes-Soares, L., Campos, I., 2000. To be or not to be in thenucleolus. Nat. Cell Biol. 2, E107–E112.
Caspersson, T., 1950. Cell Growth and Cell Function, a Cytochemical Study. WWNorton, New York.
Chuma, S., Hosokawa, M., Kitamura, K., Kasai, S., Fujioka, M., Hiyoshi, M., Takamune,K., Noce, T., Nakatsuji, N., 2006. Tdrd1/Mtr-1, a tudor-related gene, is essential formale germ-cell differentiation and nuage/germinal granule formation in mice.PNAS 103, 15894–15899.
Cmarko, D., Smigova, J., Minichova, L., Popov, A., 2008. Nucleolus: the ribosomefactory. Histol. Histopathol. 23, 1291–1298.
Comings, D.E., Okada, T.A., 1972. The chromatoid body in mouse spermatogenesis:evidence that it may be formed by the extrusion of nucleolar components. J.Ultrastruct. Res. 39 (1), 15–23.
Cotta-Pereira, G., Rodrigo, F.G., David-Ferreira, J.F., 1976. The use of tannic acidglutaraldehyde in the study of elastic related fibers. Stain Technol. 51, 7–11.
Fawcett, D.W., Eddy, E.M., Phillips, D.M., 1970. Observations on the fine structureand relationships of the chromatoid body in mammalian spermatogenesis. Biol.Reprod. 2 (1), 129–153.
Figueroa, J., Burzio, L.O., 1998. Polysome-like structures in the chromatoid body ofrat spermatids. Cell Tissue Res. 291, 575–579.
Garcia, S.N., Pillus, L., 1999. Net results of nucleolar dynamics. Cell 97, 825–828.Gerbi, S.A., Borovjagin, A.V., Lange, T.S., 2003. The nucleolus: a site of ribonucleopro-
tein maturation. Curr. Opin. Cell Biol. 15, 318–325.Guo, M., Davis, D., Birchler, J.A., 1996. Dosage effects on gene expression in a maize
ploidy series. Genetics 142, 1349–1355.Ha, R.R., Ha, J.C., 2007. Integrative Statistics for Behavioral Science. Pearson Custom
Publishing Boston.Haraguchi, C.M., Mabuchi, T., Hirata, S., Shoda, T., Hoshi, K., Akasaki, K., Yokota,
S., 2005. Chromatoid bodies: aggresome-like characteristics and degradationsites for organelles of spermiogenic cells. J. Histochem. Cytochem. 53 (4),455–465.
Head, J.R., Kresge, C.K., 1985. Reaction of the chromatoid body with a monoclonalantibody to a rat histocompatibility antigen. Biol. Reprod. 33, 1001–1008.
Hernandez-Verdun, D., 1991. The nucleolus today. J. Cell Sci. 99, 465–471.Hess, R.A., Miller, L.A., Kirby, J.D., Margoliash, E., Goldberg, E., 1993. Immunoelec-
tron microscopic localization of testicular and somatic cytochromes c in theseminiferous epithelium of the rat. Biol. Reprod. 48, 1299–1308.
Hofgärtner, F.J., Schmid, M., Krone, W., Zenzes, M.T., Engel, W., 1979. Pattern of activ-ity of nucleolus organizers during spermatogenesis in mammals as analyzed bysilver-staining. Chromosoma 71, 197–216.
Howell, W.M., Black, D.A., 1980. Controlled silver staining of nucleolus organizerregions with protective colloidal developer. I. Step method. Experientia 36,104–105.
Huang, W.S., Lin, J.Y., Yu, J.Y.L., 1997. Male reproductive cycle of the toad Bufo banko-rensis in Taiwan. Can. J. Res.: Zool. Sci. 14, 497–503.
Kligerman, A.D., Bloom, S.E., 1977. Distribuition of F-bodies, heterocromatin andnuclear organizers in the genome of the central mudminnow, Umbralimi. Cyto-genet. Cell Genet. 18, 182–196.
Kotaja, N., Bhattacharyya, S.N., Jaskiewics, L., Kimmins, S., Parvinen, M., Filipowicz,W., Sassone-Corsi, P., 2006. The chromatoid body of male germ cells: similaritywith processing bodies and presence of Dicer and microRNA pathway compo-nents. PNAS 103 (8), 2647–2652.
Kotaja, N., Sassone-Corsi, P., 2007. The chromatoid body: a germ-cell specific RNA-processing centre. Mol. Cell Biol. 8, 85–90.
Lefèvre, B., 2008. The nucleolus of the maternal gamete is essential for life. BioEssays30, 613–616.
Lofts, B., 1974. Reproduction. In: Lofts, B. (Ed.), Physiology of the Amphibians. Aca-demic Press, New York.
Meikar, O., Da Ros, M., Liljenbäck, H., Toppari, J., Kotaja, N., 2010. Accumulation ofpiRNAs in the chromatoid bodies purified by a novel isolation protocol. Exp. CellRes. 316, 1567–1575.
Mello, M.L.S., Vidal, B.C., Dantas, M.N., Monteiro, A.C.P., 1993. Discrimination ofthe nucleolus by a critical eletrolyte concentration method. Acta Histochem.Cytochem. 26, 1–3.
Mello, M.L.S., Vidal, B.C., 1980. Práticas de Biologia Celular. FUNCAMP Editora EdgardBlücher LTDA, Campinas.
Monesi, V., 1965. Synthetic activities during spermatogenesis in the mouse RNA andprotein. Exp. Cell Res. 39 (1), 197–224.
Montanaro, L., Treré, D., Derenzini, M., 2007. Nucleolus, ribosomes, and câncer. Am.J. Pathol. 173 (2), 301–310.
Morales, C.R., Hecht, N.B., 1994. Poly(A)+ Ribonucleic acids are enriched in sperma-tocyte nuclei but not in chromatoid bodies in the rat testis. Biol. Reprod. 50,309–319.
Morielle, A., Azeredo-Oliveira, M.T.V., 2004. Description of the nucleolar activityand karyotype in germinative cell lines of Rhodnius domesticus (Triatominae,Heteroptera). Caryologia 57 (1), 31–37.
Morielle-Souza, A., Azeredo-Oliveira, M.T.V., 2008. Study of the nucleolar cycle andribosomal RNA distribution during meiosis in triatomines (Triatominae, Het-eroptera). Micron 39, 1020–1026.
Mosgoeller, W., 2004. Nucleolar ultrastructure in vertebrates. In: Olson, M.O.J. (Ed.),
The Nucleolus. Kluwer.Moussa, F., Oko, R., Hermo, L., 1994. The immunolocalization of small nuclear ribonu-cleoprotein particles in testicular cells during the cycle of the seminiferousepithelium of the adult rat. Cell Tissue Res. 278, 363–378.
Nakamoto, K., Ito, A., Watabe, K., Koma, Y., Asada, H., Yoshikawa, H., Shinomura, Y.,Matsuzawa, Y., Nojima, H., Kitamura, Y., 2001. Increased expression of a nucleo-

9 / Micr
O
O
O
P
P
P
P
P
P
P
P
P
P
R
R
S
S
S
6 R.L. Peruquetti et al.
lar Nop5/Sik family member in metastatic melanoma cells: evidence for its rolein nucleolar sizing and function. Am. J. Pathol. 159, 1363–1374.
ko, R., Korley, R., Murray, M.T., Hecht, N.B., Hermo, L., 1996. Germ cell-specific DNAand RNA binding proteins p48/52 are expressed at specific stages of male germcell development and are present in the chromatoid body. Mol. Reprod. Dev. 44,1–13.
liveira, C., Zanetoni, C., Zieri, R., 2002. Morphological observations on the testes ofPhysalaemus cuvieri (Amphibia Anura). Rev. Chil. Anat. 20 (3), 263–268.
liveira, C., Vicentini, C.A., Taboga, S.R., 2003. Structural characterization of nuclearphenotypes during Scinax fuscovarius spermatogenesis (Anura Hylidae). Cary-ologia 56, 75–83.
arvinen, M., 2005. The chromatoid body in spermatogenesis. Int. J. Androl. 28,189–201.
arvinen, M., Parvinen, L., 1979. Active movements of the chromatoid body: a pos-sible transport mechanism for haploid gene products. J. Cell Biol. 80, 621–628.
arvinen, M., Salo, J., Toivonen, M., Nevalainen, O., Soini, E., Pelliniemi, L., 1997.Computer analysis of living cells: movements of the chromatoid body in earlyspermatids compared with its ultrastructure in snap-frozen preparations. His-tochem. Cell Biol. 108, 77–81.
arvinen, M., Vihko, K.K., Toppari, J., 1986. Cell interactions during the seminiferousepithelial cycle. In: Cytology: A Survey of Cell Biology, 3rd ed. Academic Press,New York.
ederson, T., 2002. Proteomics of the nucleolus: more proteins, more functions?Trends Biochem. Sci. 27 (3), 111–112.
eruquetti, R.L., Assis, I.M., Taboga, S.R., Azeredo-Oliveira, M.T.V., 2008a. Meioticnucleolar cycle and chromatoid body formation during the rat (Rattus novergi-cus) and mouse (Mus musculus) spermiogenesis. Micron 39, 419–425.
eruquetti, R.L., Taboga, S.R., Azeredo-Oliveira, M.T.V., 2008b. Characterizationof Mongolian gerbil chromatoid bodies and their correlation with nucleolarcycle during spermatogenesis. Reprod. Dom. Anim., doi:10.1111/j.1439-0531.2008.01204.x.
eruquetti, R.L., 2009. Caracterizacão do ciclo nucleolar e da formacão do corpocromatóide na espermatogênese de algumas espécies de vertebrados [thesis].São Jose do Rio Preto (SP): São Paulo State University (UNESP/IBILCE).
eruquetti, R.L., Taboga, S.R., Azeredo-Oliveira, M.T.V., 2010. Nucleolar cycle and itscorrelation with chromatoid body formation in the Tilapia rendalli (TeleosteiCichlidae) spermatogenesis. Anat. Rec. 293, 900–910.
ikaard, C.S., 2002. Transcription and tyranny in the nucleolus: the organization,activation, dominance and repression of ribosomal RNA genes. In: Somerville,C.R., Meyerowits, E.M. (Eds.), The Arabidopsis Book. American Society of PlantBiologists, Rockville.
eunov, A., Isaeva, V., Au, D., Wu, R., 2000. Nuage constituents arising from mito-chondria: is it possible? Dev. Growth Differ. 42, 139–143.
ibeiro, M.G., Lima, S.R., 2000. Iniciacão às técnicas de preparacão de material paraestudo e pesquisa em morfologia. SEGRAC Editora e Gráfica Limitada, Belo Hor-izonte.
an-Segundo, P.A., Roeder, G.S., 1999. Pch2 links chromatin silencing to meiotic
checkpoint control. Cell 97, 313–324.antos, L.R.S., Oliveira, C., 2007. Morfometria testicular durante o ciclo reprodu-tivo de Dendropsophus minutus (Peters) (Anura Hylidae). Rev. Bras. Zool. 24 (1),64–70.
aunders, P.T.K., Millar, M.R., Maguire, S.M., Sharpe, R.M., 1992. Stage-specificexpression of rat transition protein 2 mRNA and possible localization to the chro-
on 42 (2011) 87–96
matoid body of step 7 spermatids by in situ hybridization using a nonradioactiveriboprobe. Mol. Reprod. Dev. 33, 385–391.
Schmid, M., Löser, C., Schmidtke, J., Engel, W., 1982. Evolutionary conservation of acommon pattern of activity of nucleolus organizers during spermatogenesis invertebrates. Chromosoma 86, 149–179.
Shou, W., Seol, J.H., Shevchenko, A., Baskerville, C., Moazerd, D., Chen, Z.W., Jang,J., Shevchenko, A., Charbonneau, H., Deshaies, R.J., 1999. Exit from mitosis istriggered by tem1-dependent release of the protein phosphatase Cdc14 fromnucleolar RENT complex. Cell 97, 233–244.
Shubert, I., Künzel, G., 1990. Position dependent NOR activity in barley. Chromosoma99, 352–359.
Sirri, V., Urcuqui-Inchima, S., Roussel, P., Hernandez-Verdun, D., 2008. Nucleolus:the fascinating nuclear body. Histochem. Cell Biol. 129, 13–31.
Söderström, K., 1977. Effect of actinomycin D on the structure of the chromatoidbody in the rat spermatids. Cell Tissue Res. 184, 411–421.
Söderström, K., Parvinen, M., 1976a. Transport of material between the nucleus, thechromatoid body and the Golgi complex in the early spermatids of the rat. CellTissue Res. 168, 335–342.
Söderström, K., Parvinen, M., 1976b. Incorporation of [3H]uridine by the chromatoidbody during rat spermatogenesis. J. Cell Biol. 70, 239–246.
Soley, J.T., 1994. Centriole development and formation of the flagellum duringspermiogenesis in the ostrich (Struthio camelus). J. Anat. 185, 301–313.
Straight, A.F., Shou, W., Dowd, G.J., Turck, C.W., Deshaies, R.H., Johnson, A.D., Moazed,D., 1999. Net1, a Sir2-associated nucleolar protein required for rDNA silencingand nucleolar integrity. Cell 97, 245–256.
Sud, B., 1961. Morphological and histochemical studies of the chromatoid bodyand related elements in the spermatogenesis of rat. Q. J. Microsc. Sci. 102,273–292.
Takeuchi, I.K., Takeuchi, Y.K., 1990. Ethanol-phosphotungstic acid and bismuthstaining of spermatid nucleoli in mouse spermiogenesis. J. Struct. Biol. 103,104–112.
Tandler, B., Moriber, L.G., 1965. Fine structure of the chormatoid body duringspermatogenesis in the Water-Strider, Gerris remigis (Say)*. Z. Zellforsch. 68,301–307.
Teruel, M., Cabrero, J., Perfectti, F., Camacho, J.P.M., 2007. Nucleolus size variationduring meiosis and NOR activity of a B chromosome in the grasshopper Eypre-pocnemis plorans. Chromosome Res. 15, 755–765.
Vasileva, A., Tiedau, D., Firooznia, A., Müller-Reichert, T., Jessberger, R., 2009. Tdrd6is required for spermiogenesis, chromatoid body architecture, and regulation ofmiRNA expression. Curr. Biol. 19, 630–639.
Venable, J.H., Coggeshall, R.A., 1965. A simplified lead citrate stain for use in electronmicroscopy. J. Cell Biol. 25, 407–408.
Visitin, R., Hwang, E.S., Arnon, A., 1999. Cfi1 prevents premature exit from mitosisby anchoring Cdc14 phosphatase in the nucleolus. Nature 398, 818–823.
Wachtler, F., Stahl, A., 1993. The nucleolus: a structural and functional interpretation.Micron 24, 473–505.
Walt, H., Armbruster, B.L., 1984. Actin and RNA are components of the chromatoid
bodies in spermatids of the rat. Cell Tissue Res. 236, 487–490.Watson, M.L., 1958. Staining tissue section of electron microscopy with heavy met-als. J. Biophys. Biochem. Cytol. 4, 475–478.
Yokota, S., 2008. Historical survey on chromatoid body research. Acta Histochem.Cytochem. 41 (4), 65–82.
Zar, J.H., 1999. Biostatiscical Analysis. Prentice Hall, New Jersey.
Copyright © 2022 FDOKUMEN




















