
Saturnius minutus n. sp. and S. dimitrovi n. sp. (Digenea: Hemiuridae) from Mugil cephalus L....
15
Saturnius minutus n. sp. and S. dimitrovi n. sp. (Digenea: Hemiuridae) from Mugil cephalus L. (Teleostei: Mugilidae), with a multivariate morphological analysis of the Mediterranean species of Saturnius Manter, 1969 I. Blasco-Costa 1, *, P. Pankov 2 , D.I. Gibson 3 , J.A. Balbuena 1 , J.A. Raga 1 , V.L. Sarabeev 4 & A. Kostadinova 2 1 Marine Zoology Unit, Cavanilles Institute of Biodiversity and Evolutionary Biology, University of Valencia, PO Box 22 085, 46071 Valencia, Spain 2 Central Laboratory of General Ecology, Bulgarian Academy of Sciences, 2 Gagarin Street, 1113 Sofia, Bulgaria 3 Department of Zoology, Natural History Museum, London, SW7 5BD UK 4 Department of Biology, Zaporizhzhia National University, 66 Zhukovskogo Street, 69063 Zhaporizhzhia, Ukraine Accepted for publication 3rd December, 2005 Abstract Three species of the bunocotyline genus Saturnius Manter, 1969 are described from the stomach lining of mugilid fishes of the Mediterranean and Black Seas. Two of the species are new: S. minutus n. sp. occurs in Mugil cephalus off the Mediterranean coast of Spain; and S. dimitrovi n. sp., a parasite of M. cephalus off the Bulgarian Black Sea coast and the Spanish Mediterranean coast, was originally described as S. papernai by Dimitrov et al. (1998). In addition, S. papernai Overstreet, 1977 is redescribed from M. cephalus off the Spanish Mediterranean coast and from Liza aurata and L. saliens off the Bulgarian Black Sea coast. The three species are distinguished morphometrically using univariate and multivariate analyses. These results were verified using Linear Discriminant Analysis which correctly allocated all specimens to their species designations based on morphology (i.e. 100% successful classification rate) and assigned almost all spec- imens to the correct population (locality). The following variables were selected for optimal separation between samples: the length of the forebody, ventral sucker and posterior testis, the length and width of the posteriormost pseudosegment, and the width of the muscular flange at ventral sucker level. Introduction Saturnius Manter, 1969 is a small hemiurid genus (subfamily Bunocotylinae Dollfus, 1950) of stom- ach parasites specific to mullets (Mugilidae), which is represented by six species: S. segmentatus Manter, 1969, S. mugilis (Yamaguti, 1970), S. maurepasi Overstreet, 1977, S. belizensis Fischthal, 1977, S. papernai Overstreet, 1977 and S. valam- ugilis Rekharani & Madhavi, 1984. Of these, only S. papernai, originally described from Mugil ceph- alus in Bardawil Lagoon (Sinai coast, Eastern Mediterranean) by Overstreet (1977), has been subsequently recorded in the Mediterranean basin, most records coming from the Black and Azov Seas (see e.g. Dimitrov et al., 1998; Sarabeev, 2000; Domnich & Sarabeev, 2000a-d; Dmitrieva & Gaevskaya, 2001; Merella & Garippa, 2001). Recently, Domnich & Sarabeev (1999) described Bunocotyle constrictus Domnich & Sarabeev, 1999 from Mugil soiuy in the Azov Sea, which they later (see e.g. Domnich & Sarabeev, 2000a,b) consid- ered a misidentification of S. papernai. In a study of the parasites of mullets (Mugil- idae) from two localities off the Spanish Mediter- ranean coast (Ebro Delta and Santa Pola) and one *Author for correspondence (E-mail: [email protected]) Systematic Parasitology (2006) 65:77–91 Ó Springer 2006 DOI 10.1007/s11230-006-9043-9
Transcript of Saturnius minutus n. sp. and S. dimitrovi n. sp. (Digenea: Hemiuridae) from Mugil cephalus L....

Saturnius minutus n. sp. and S. dimitrovi n. sp. (Digenea: Hemiuridae) fromMugil cephalus L. (Teleostei: Mugilidae), with a multivariate morphologicalanalysis of the Mediterranean species of Saturnius Manter, 1969
I. Blasco-Costa1,*, P. Pankov2, D.I. Gibson3, J.A. Balbuena1, J.A. Raga1, V.L. Sarabeev4
& A. Kostadinova21Marine Zoology Unit, Cavanilles Institute of Biodiversity and Evolutionary Biology, University of Valencia,PO Box 22 085, 46071 Valencia, Spain2Central Laboratory of General Ecology, Bulgarian Academy of Sciences, 2 Gagarin Street, 1113 Sofia,Bulgaria3Department of Zoology, Natural History Museum, London, SW7 5BD UK4Department of Biology, Zaporizhzhia National University, 66 Zhukovskogo Street, 69063 Zhaporizhzhia,Ukraine
Accepted for publication 3rd December, 2005
Abstract
Three species of the bunocotyline genus Saturnius Manter, 1969 are described from the stomach lining ofmugilid fishes of the Mediterranean and Black Seas. Two of the species are new: S. minutus n. sp. occurs inMugil cephalus off the Mediterranean coast of Spain; and S. dimitrovi n. sp., a parasite of M. cephalus offthe Bulgarian Black Sea coast and the Spanish Mediterranean coast, was originally described as S. papernaiby Dimitrov et al. (1998). In addition, S. papernai Overstreet, 1977 is redescribed from M. cephalus off theSpanish Mediterranean coast and from Liza aurata and L. saliens off the Bulgarian Black Sea coast. Thethree species are distinguished morphometrically using univariate and multivariate analyses. These resultswere verified using Linear Discriminant Analysis which correctly allocated all specimens to their speciesdesignations based on morphology (i.e. 100% successful classification rate) and assigned almost all spec-imens to the correct population (locality). The following variables were selected for optimal separationbetween samples: the length of the forebody, ventral sucker and posterior testis, the length and width of theposteriormost pseudosegment, and the width of the muscular flange at ventral sucker level.
Introduction
Saturnius Manter, 1969 is a small hemiurid genus(subfamily Bunocotylinae Dollfus, 1950) of stom-ach parasites specific to mullets (Mugilidae), whichis represented by six species: S. segmentatusManter, 1969, S. mugilis (Yamaguti, 1970), S.maurepasi Overstreet, 1977, S. belizensis Fischthal,1977, S. papernai Overstreet, 1977 and S. valam-ugilis Rekharani & Madhavi, 1984. Of these, onlyS. papernai, originally described from Mugil ceph-alus in Bardawil Lagoon (Sinai coast, Eastern
Mediterranean) by Overstreet (1977), has beensubsequently recorded in the Mediterranean basin,most records coming from the Black and AzovSeas (see e.g. Dimitrov et al., 1998; Sarabeev,2000; Domnich & Sarabeev, 2000a-d; Dmitrieva &Gaevskaya, 2001; Merella & Garippa, 2001).Recently, Domnich & Sarabeev (1999) describedBunocotyle constrictus Domnich & Sarabeev, 1999from Mugil soiuy in the Azov Sea, which they later(see e.g. Domnich & Sarabeev, 2000a,b) consid-ered a misidentification of S. papernai.
In a study of the parasites of mullets (Mugil-idae) from two localities off the Spanish Mediter-ranean coast (Ebro Delta and Santa Pola) and one*Author for correspondence (E-mail: [email protected])
Systematic Parasitology (2006) 65:77–91 � Springer 2006DOI 10.1007/s11230-006-9043-9

locality off the Bulgarian Black Sea coast (Sozo-pol), we have collected two different forms ofSaturnius which can be clearly distinguished bysize. The large form agrees well with the originaldescription of S. papernai (see Overstreet, 1977),whereas the small form exhibits enough differencesfrom existing species as to be considered new. Thenewly collected material of S. papernai, however,showed a number of metrical differences (e.g. sizeof body, sinus-sac, pars prostatica, seminal vesicleand vitellarium, and relative size of the ventralsucker) when compared to the redescription of S.papernai given by Dimitrov et al. (1998) based onmaterial from the Black Sea. A re-examination ofthe latter material, and multivariate comparisonsof the three forms plus the type-specimens of S.papernai, convinced us that the set described byDimitrov et al. (1998) also requires a distinctspecies status. This paper presents a descriptionof two new species of Saturnius, new morphomet-ric data for S. papernai and a multivariate analysiswhich discriminates the three forms from theMediterranean basin.
Materials and methods
A total of 118 mullets was collected and examinedin June, 2004: 60 Mugil cephalus from twolocalities (Ebro Delta and off Santa Pola) off theSpanish Mediterranean coast; and 48 Liza aurataand 10 L. saliens from off Sozopol on theBulgarian Black sea coast. Worms were recoveredfrom fresh fish, fixed in cold 70% alcohol, stainedwith iron acetocarmine (Georgiev et al., 1986),dehydrated through an alcohol series, cleared indimethyl phthalate and examined as permanentmounts in Canada balsam. Comparative materialexamined comprised type-specimens of Saturniuspapernai from the Collection of the NaturalHistory Museum, London [BMNH 1975.9.29.4(holotype) and BMNH 1975.9.29.5 (paratype)],United States National Parasite Collection (US-NPC no. 73271, 1 paratype), the Collection of theHarold Manter Parasitological Laboratory(20248, two paratypes) and additional and vou-cher specimens described as S. papernai anddeposited by Dimitrov et al. (1998) in the Collec-tions of the Central Laboratory of General Ecol-ogy (CLGE 15302) and the Natural HistoryMuseum (BMNH 1997.6.23.2). Measurements
were taken from illustrations, which were madeusing a drawing apparatus at high magnification.All measurements are in micrometres.
Measurements were recorded from a set of 85specimens (23 S. minutus n. sp., 30 S. dimitrovi n.sp. and 32 S. papernai); these include eightS. minutus additional to the type-series, and thetype-material of S. papernai (5 specimens). Thesedata were ln-transformed and subjected to univar-iate (ANOVA, a posteriori contrasts) statisticaltests. Linear Discriminant Analysis (LDA) wasapplied to a set of 69 specimens for which data forall variables measured were available (body length,egg-size and ratios were excluded from theseanalyses) in order to evaluate and visualise themorphometric differences between species. The 69specimens were distributed in six a priori groupsdefined by their species identification and geo-graphical origin, namely S. minutus from off theMediterranean coast of Spain (n=12), S. papernaifrom off the Black Sea coast of Bulgaria (n=7), S.papernai from off the Mediterranean coast ofSpain (n=19), S. dimitrovi from off the Black Seacoast of Bulgaria (n=20) and S. dimitrovi from offthe Mediterranean coast of Spain (n=6), plus thetype-material of S. papernai (n=5) from theEastern Mediterranean.
In the first analysis (A), ln-transformed metri-cal data were subjected to a stepwise LDA to selectthose variables yielding optimal separation be-tween the groups. Since the three Saturnius spp.differed substantially in body size, a secondanalysis (B) was carried out in order to testwhether discrimination between the groups stilloccurred after controlling for this factor. A step-wise LDA was run using a size-adjusting techniqueconsisting in substitution of the original metricvariables by the ratios formed by dividing eachvariable by the geometric mean of all variables(Darroch & Mosimann, 1985). It has been shownthat this approach satisfactorily minimises theeffect of body size in multivariate analyses(Jungers et al., 1995). Univariate and multivariatestatistical analyses were carried out using SPSS�
12.0 (SPSS Inc.; Norusis, 2002) at the Universityof Valencia.
The following abbreviations for ratios are usedin the text and tables: BW/BL, body width as apercent of body length; FO/BL, forebody as apercent of body length; LSL/BL, posteriormostpseudosegment length as a percent of body length;
78

LSW/LSL, posteriormost pseudosegment width asa percent of its length; MFL/VSL, muscular flangelength as a percent of ventral sucker length; LSW/BW, posteriormost pseudosegment width as apercent of body width; VSW/BWVS, ventralsucker width as a percent of body width at levelof the anterior margin of the ventral sucker;MFW/BWVS, muscular flange width as a percentof body width at anterior border of ventral sucker;plus the ratios: VSL/OSL, ventral sucker length asa proportion of oral sucker length, and VSW/OSW, ventral sucker width as a proportion of oralsucker width.
The Host-Parasite Database maintained atthe Natural History Museum, London (http://www.nhm.ac.uk/research-curation/projects/host-parasites/database/) was searched for host-parasiterecords.
Family Hemiuridae Looss, 1899
Genus Saturnius Manter, 1969
Saturnius minutus n. sp.
Type-host: Mugil cephalus L.Type-locality: Off the Mediterranean coast ofSpain (at Santa Pola).Site: Embedded between the grinding stomachlining and the layer of glandular cells.Type-material: Holotype BMNH 2005.11.10.1;paratypes BMNH 2005.11.10.2-5.Voucher material: BMNH 2005.11.10.6-9.
Description (Figures 1, 4A)
[Based on 19 whole-mounted adult specimens;metrical data for 15 adult worms in Tables 1, 2.]Body minute, elongate, cylindrical, with roundedposterior extremity and maximum width at level ofventral sucker flange; width at mid-level of ventralsucker 65–91 (74±5). Tegument unarmed, withfine longitudinal and transverse striations.
Three circular muscular flanges present aroundbody: (i) anterior (oral sucker) flange weaklydeveloped, located at base of oral sucker, seen asfaint thickening in lateral view; (ii) second (ventralsucker) flange strongly muscular, forming 2 lateralsub-conical protuberances provided with concen-tric muscles, with cone length (anterior-posterior)34–59 (47±6) and width (lateral) 17–34 (24±4)[24–47% (32±5%) of body width at mid-level of
ventral sucker], with anterior margin located atlevel of posterior third of ventral sucker; tegumen-tal ridge also distinguishable on ventral surface ofwhole-mounts posterior to posterior margin ofventral sucker; (iii) third flange located in posteriorthird of last pseudosegment, in some specimensrelatively poorly developed.
Body typically with 6 pseudosegments sepa-rated by very faint transverse fibrous septa, diffi-cult to see in many cases. Anteriormost septum atlevel of genital pore, not observed in 6 specimens(32%); 4 other septa (first located just posterior toventral sucker flange) form 4 pseudosegments ofdifferent sizes in hindbody; anterior smallest,middle pair similar in size and posteriormostlargest, 17–29% (22±3%) of body length.
Oral sucker simple, transverse-oval, with ter-minal opening. Ventral sucker strongly muscular,subspherical, larger than oral sucker, at mid-body.Prepharynx absent; pharynx muscular, subspher-ical. Oesophagus very short, in some specimenssurrounded by minute gland-cells. Intestinal bifur-cation at about middle of first pseudosegment.Caeca relatively wide, with several constrictions;most posterior region impossible to observe inadult specimens since it is masked by ovary,vitellarium and uterus; in juvenile specimens,however, caeca can be seen to unite posterior tovitellarium to form cyclocoel (Figure 4A).
Testes 2, subspherical, smooth, in tandem orslightly oblique, occupy most of respective pseudo-segments, in middle of hindbody, contiguous.Seminal vesicle thin-walled, elongate-oval, antero-dorsal to ventral sucker, similar in size to sinus-sac.Pars prostatica external, elongate-oval, vesicular,similar in size to sinus-sac and seminal vesicle, linedwith anuclear blebs, surrounded by large prostaticcells which also partly overlap sinus-sac andanterior part of seminal vesicle. Sinus-sac thin-walled, elongate-oval, contains eversible hermaph-roditic duct lined by small intensely stained cellsand which forms temporary sinus-organ. Genitalpore a transverse slit, median, at level of firstseptum (when visible).
Ovary oval to subtriangular, in anterior part oflast pseudosegment, partly overlaps vitellariumdorsally. Uterine seminal receptacle present; La-urer’s canal not observed. Vitellarium compact,round to elongate-oval, larger than ovary, lengthc.10% of body length. Uterus thin-walled, slightlycoiled between ventral sucker and posterior
79
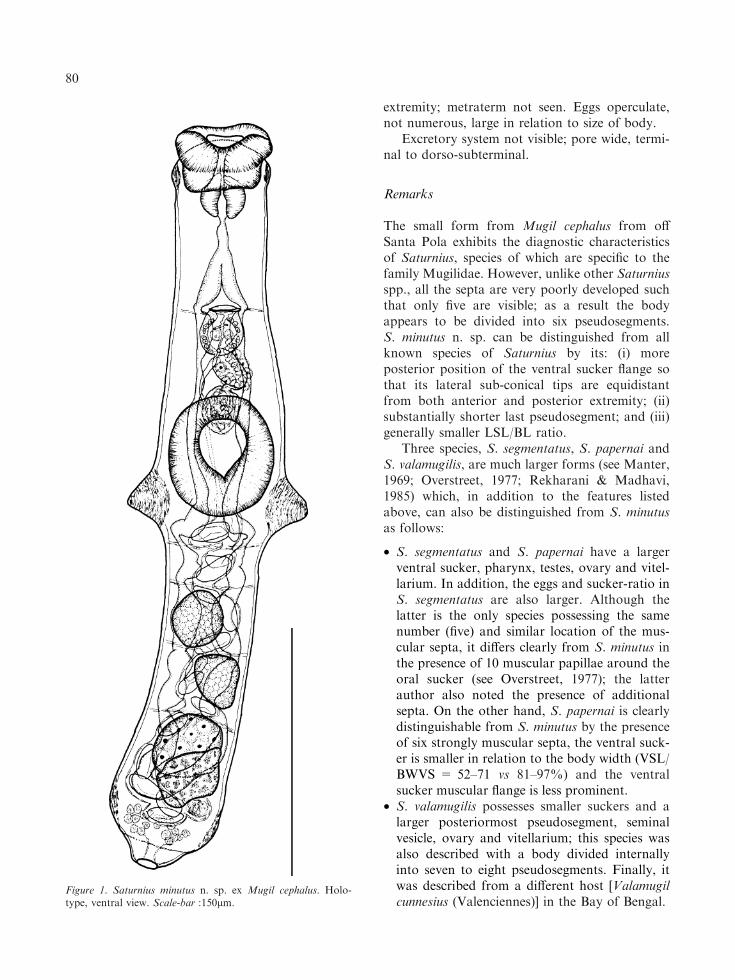
extremity; metraterm not seen. Eggs operculate,not numerous, large in relation to size of body.
Excretory system not visible; pore wide, termi-nal to dorso-subterminal.
Remarks
The small form from Mugil cephalus from offSanta Pola exhibits the diagnostic characteristicsof Saturnius, species of which are specific to thefamily Mugilidae. However, unlike other Saturniusspp., all the septa are very poorly developed suchthat only five are visible; as a result the bodyappears to be divided into six pseudosegments.S. minutus n. sp. can be distinguished from allknown species of Saturnius by its: (i) moreposterior position of the ventral sucker flange sothat its lateral sub-conical tips are equidistantfrom both anterior and posterior extremity; (ii)substantially shorter last pseudosegment; and (iii)generally smaller LSL/BL ratio.
Three species, S. segmentatus, S. papernai andS. valamugilis, are much larger forms (see Manter,1969; Overstreet, 1977; Rekharani & Madhavi,1985) which, in addition to the features listedabove, can also be distinguished from S. minutusas follows:
• S. segmentatus and S. papernai have a largerventral sucker, pharynx, testes, ovary and vitel-larium. In addition, the eggs and sucker-ratio inS. segmentatus are also larger. Although thelatter is the only species possessing the samenumber (five) and similar location of the mus-cular septa, it differs clearly from S. minutus inthe presence of 10 muscular papillae around theoral sucker (see Overstreet, 1977); the latterauthor also noted the presence of additionalsepta. On the other hand, S. papernai is clearlydistinguishable from S. minutus by the presenceof six strongly muscular septa, the ventral suck-er is smaller in relation to the body width (VSL/BWVS = 52–71 vs 81–97%) and the ventralsucker muscular flange is less prominent.
• S. valamugilis possesses smaller suckers and alarger posteriormost pseudosegment, seminalvesicle, ovary and vitellarium; this species wasalso described with a body divided internallyinto seven to eight pseudosegments. Finally, itwas described from a different host [Valamugilcunnesius (Valenciennes)] in the Bay of Bengal.
Figure 1. Saturnius minutus n. sp. ex Mugil cephalus. Holo-type, ventral view. Scale-bar :150lm.
80
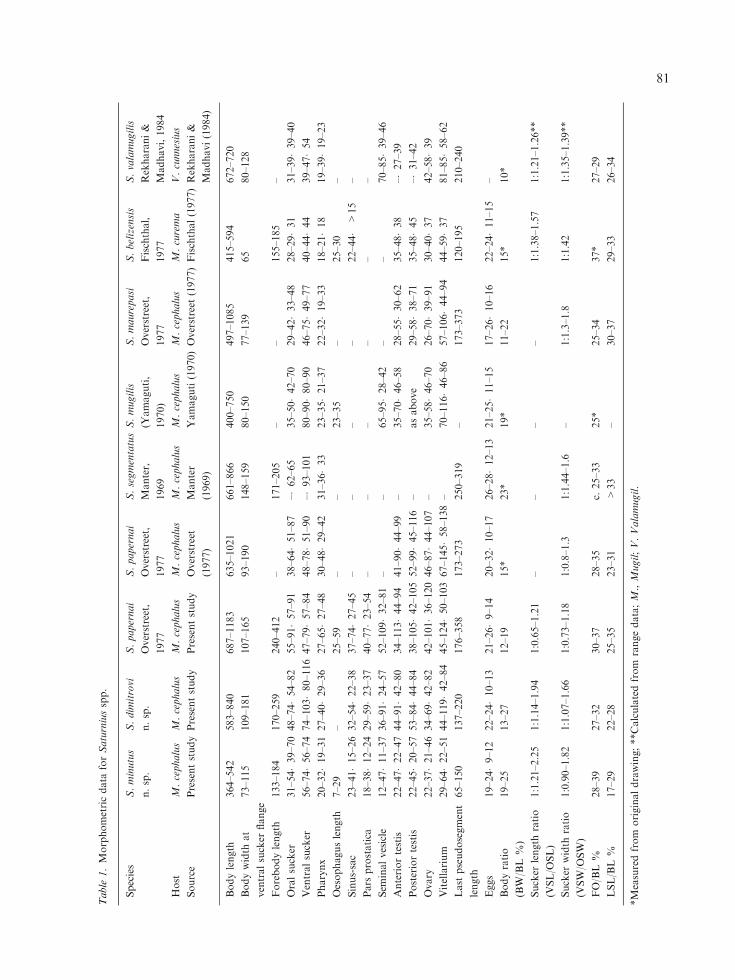
Table
1.Morphometricdata
forSaturniusspp.
Species
S.minutus
n.sp.
S.dim
itrovi
n.sp.
S.papernai
Overstreet,
1977
S.papernai
Overstreet,
1977
S.segmentatus
Manter,
1969
S.mugilis
(Yamaguti,
1970)
S.maurepasi
Overstreet,
1977
S.belizensis
Fischthal,
1977
S.valamugilis
Rekharani&
Madhavi,1984
Host
M.cephalus
M.cephalus
M.cephalus
M.cephalus
M.cephalus
M.cephalus
M.cephalus
M.curema
V.cunnesius
Source
Presentstudy
Presentstudy
Presentstudy
Overstreet
(1977)
Manter
(1969)
Yamaguti(1970)Overstreet
(1977)Fischthal(1977)Rekharani&
Madhavi(1984)
Bodylength
364–542
583–840
687–1183
635–1021
661–866
400–750
497–1085
415–594
672–720
Bodywidth
at
ventralsucker
flange
73–115
109–181
107–165
93–190
148–159
80–150
77–139
65
80–128
Forebodylength
133–184
170–259
240–412
–171–205
––
155–185
–
Oralsucker
31–54�39–70
48–74�54–82
55–91�57–91
38–64�51–87
–�62–65
35–50�42–70
29–42�33–48
28–29�31
31–39�39–40
Ventralsucker
56–74�56–74
74–103�80–116
47–79�57–84
48–78�51–90
–�93–101
80–90�80–90
46–75�49–77
40–44�44
39–47�54
Pharynx
20–32�19–31
27–40�29–36
27–65�27–48
30–48�29–42
31–36�33
23–35�21–37
22–32�19–33
18–21�18
19–39�19–23
Oesophaguslength
7–29
–25–59
––
23–35
–25–30
–
Sinus-sac
23–41�15–26
32–54�22–38
37–74�27–45
––
––
22–44�
>15
–
Pars
prostatica
18–38�12–24
29–59�23–37
40–77�23–54
––
––
––
Sem
inalvesicle
12–47�11–37
36–91�24–57
52–109�32–81
––
65–95�28–42
––
70–85�39–46
Anteriortestis
22–47�22–47
44–91�42–80
34–113�44–94
41–90�44–99
–35–70�46–58
28–55�30–62
35–48�38
–�27–39
Posteriortestis
22–45�20–57
53–84�44–84
38–105�42–105
52–99�45–116
–asabove
29–58�38–71
35–48�45
–�31–42
Ovary
22–37�21–46
34–69�42–82
42–101�36–120
46–87�44–107
–35–58�46–70
26–70�39–91
30–40�37
42–58�39
Vitellarium
29–64�22–51
44–119�42–84
45–124�50–103
67–145�58–138
–70–116�46–86
57–106�44–94
44–59�37
81–85�58–62
Last
pseudosegment
length
65–150
137–220
176–358
173–273
250–319
–173–373
120–195
210–240
Eggs
19–24�9–12
22–24�10–13
21–26�9–14
20–32�10–17
26–28�12–13
21–25�11–15
17–26�10–16
22–24�11–15
–
Bodyratio
(BW/BL%
)
19–25
13–27
12–19
15*
23*
19*
11–22
15*
10*
Sucker
length
ratio
(VSL/O
SL)
1:1.21–2.25
1:1.14–1.94
1:0.65–1.21
––
––
1:1.38–1.57
1:1.21–1.26**
Sucker
width
ratio
(VSW/O
SW)
1:0.90–1.82
1:1.07–1.66
1:0.73–1.18
1:0.8–1.3
1:1.44–1.6
–1:1.3–1.8
1:1.42
1:1.35–1.39**
FO/BL%
28–39
27–32
30–37
28–35
c.25–33
25*
25–34
37*
27–29
LSL/BL%
17–29
22–28
25–35
23–31
>33
–30–37
29–33
26–34
*Measuredfrom
originaldrawing;**Calculatedfrom
rangedata;M.,Mugil;V.Valamugil.
81
The three other Saturnius spp., i.e. S. mugilis, S.maurepasi and S. belizensis, which show overlap-ping lower ranges for body length (see Yamaguti,1970; Overstreet, 1977; Fischthal, 1977), differfrom S. minutus n. sp. as follows:
• S. mugilis has a larger ventral sucker, seminalvesicle, ovary and vitellarium. This species alsolacks a septum at the level of the genital pore.
• S. maurepasi possesses a pars prostatica whichis distinctly smaller in relation to the seminalvesicle, which occupies a large proportion ofthe second pseudosegment, and a larger vitel-larium, the length of which represents a greaterproportion of the body length.
• S. belizensis is a much thinner form, with smal-ler suckers and pharynx. Furthermore, thisspecies was described from Mugil curema fromthe Carribean Sea off Belize and possesses onlythree septa located in the hindbody.
The above comparisons justify, in our opinion,the erection of S. minutus n. sp.
Saturnius dimitrovi n. sp.Syn. S. papernai Overstreet, 1977 of Dimitrovet al. (1998)
Type-host: Mugil cephalus L.Type-locality: Off the Black Sea coast of Bulgaria(at Sozopol).
Table 2. Means for the variables for Saturnius spp. from mullet subjected to univariate statistical analyses.
Species S. minutus n. sp. S. dimitrovi n. sp. S. papernai
Locality Off Santa Pola Off Sozopol & Ebro Delta Off Sozopol & Ebro Delta
Mean ± SD Mean ± SD Mean ± SD
Body length 431±42 691±74 994±149
Body width (at ventral sucker flange) 92±11 130±14 141±16
Body width (at anterior margin of ventral sucker) 74±6 106±9 119±12
Forebody length 155±16 205±23 326±45
Oral sucker 41±6 � 56±7 58±7 � 67±7 71±9 � 78±10
Ventral sucker 67±4 � 66±4 87±6 � 88±7 66±8 � 73±7
Pharynx 25±2 � 25±3 35±3 � 33±2 44±8 � 40±5
Oesophagus length 19±5 28±9 37±8
Sinus-sac 32±4 � 21±4 40±6 � 32±3 54±11 � 37±5
Pars prostatica 27±5 � 17±3 42±7 � 29±4 62±9 � 38±7
Seminal vesicle 33±9 � 22±4 54±14 � 39±9 82±17 � 57±14
Anterior testis 33±8 � 33±6 65±10 � 55±9 72±19 � 67±12
Posterior testis 34±5 � 35±8 65±8 � 61±10 73±17 � 70±17
Ovary 30±5 � 33±7 46±8 � 60±10 62±13 � 71±20
Vitellarium 42±9 � 37±7 76±16 � 62±10 87±22 � 76±14
Last pseudosegment 86±14 � 63±8 169±23 � 109±8 278±45 � 141±24
Muscular flange cone length (anterior-posterior) 46±5 60±6 65±11
Muscular flange cone width (lateral) 23±3 29±3 28±5
Eggs 21±1 � 10±1 23±1 � 11±1 24±2 � 12±1
Body ratio (BW/BL) % 21±2 19±3 14±2
Sucker length ratio (VSL/OSL) 1:1.66±0.25 1:1.51±0.18 1:0.93±0.11
Sucker width ratio (VSW/OSW) 1:1.19±0.13 1:1.32±1.26 1:0.95±0.12
FO/BL % 36±1 30±1 33±2
LSL/BL % 20±3 24±2 28±2
LSW/LSL % 73±11 66±9 51±9
MFL/VSL % 69±8 69±8 99±13
LSW/BW % 68±6 84±6 101±13
VSW/BWVS % 89±7 83±5 61±5
MFW/BWVS % 31±4 27±4 23±4
82
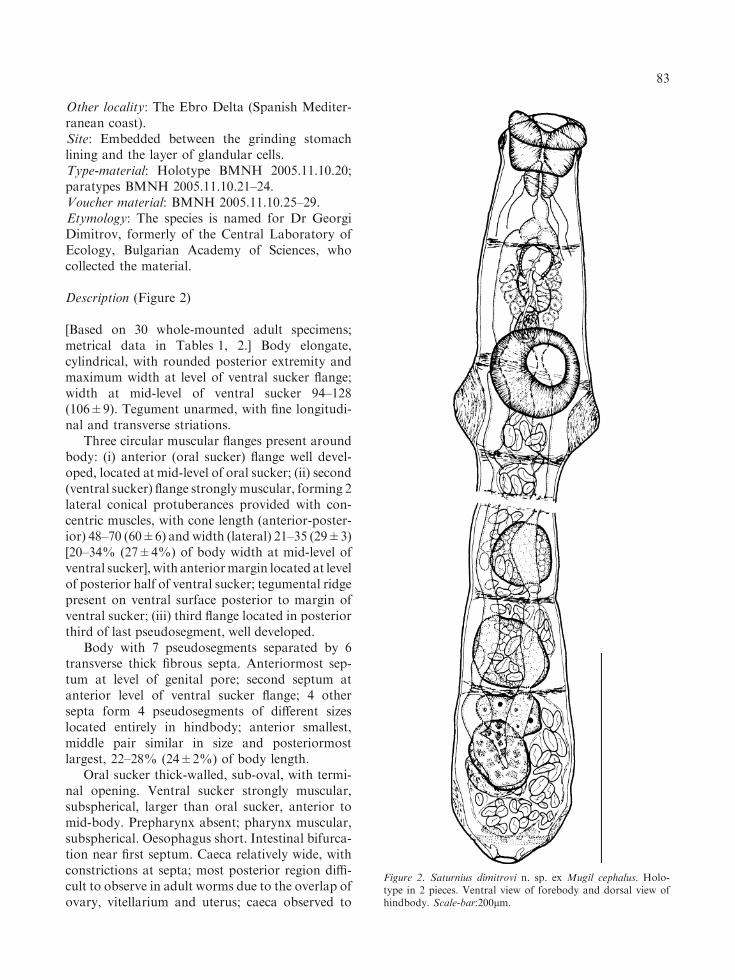
Other locality: The Ebro Delta (Spanish Mediter-ranean coast).Site: Embedded between the grinding stomachlining and the layer of glandular cells.Type-material: Holotype BMNH 2005.11.10.20;paratypes BMNH 2005.11.10.21–24.Voucher material: BMNH 2005.11.10.25–29.Etymology: The species is named for Dr GeorgiDimitrov, formerly of the Central Laboratory ofEcology, Bulgarian Academy of Sciences, whocollected the material.
Description (Figure 2)
[Based on 30 whole-mounted adult specimens;metrical data in Tables 1, 2.] Body elongate,cylindrical, with rounded posterior extremity andmaximum width at level of ventral sucker flange;width at mid-level of ventral sucker 94–128(106±9). Tegument unarmed, with fine longitudi-nal and transverse striations.
Three circular muscular flanges present aroundbody: (i) anterior (oral sucker) flange well devel-oped, located at mid-level of oral sucker; (ii) second(ventral sucker) flange stronglymuscular, forming 2lateral conical protuberances provided with con-centric muscles, with cone length (anterior-poster-ior) 48–70 (60±6) and width (lateral) 21–35 (29±3)[20–34% (27±4%) of body width at mid-level ofventral sucker], with anteriormargin located at levelof posterior half of ventral sucker; tegumental ridgepresent on ventral surface posterior to margin ofventral sucker; (iii) third flange located in posteriorthird of last pseudosegment, well developed.
Body with 7 pseudosegments separated by 6transverse thick fibrous septa. Anteriormost sep-tum at level of genital pore; second septum atanterior level of ventral sucker flange; 4 othersepta form 4 pseudosegments of different sizeslocated entirely in hindbody; anterior smallest,middle pair similar in size and posteriormostlargest, 22–28% (24±2%) of body length.
Oral sucker thick-walled, sub-oval, with termi-nal opening. Ventral sucker strongly muscular,subspherical, larger than oral sucker, anterior tomid-body. Prepharynx absent; pharynx muscular,subspherical. Oesophagus short. Intestinal bifurca-tion near first septum. Caeca relatively wide, withconstrictions at septa; most posterior region diffi-cult to observe in adult worms due to the overlap ofovary, vitellarium and uterus; caeca observed to
Figure 2. Saturnius dimitrovi n. sp. ex Mugil cephalus. Holo-type in 2 pieces. Ventral view of forebody and dorsal view ofhindbody. Scale-bar:200lm.
83
unite posterior to vitellarium to form cyclocoel in 5specimens (see e.g. holotype, Figure 2).
Testes 2, subspherical to elongate-oval,smooth, in tandem, occupy most of respectivepseudosegments in middle of hindbody, contigu-ous or slightly separated. Seminal vesicle thick-walled, elongate-oval, antero-dorsal to ventralsucker, usually larger than sinus-sac. Pars prost-atica external, elongate-oval, vesicular, similar insize to sinus-sac, lined with anuclear blebs, sur-rounded by large prostatic cells which overlapsinus-sac and anterior half of seminal vesicle.Sinus-sac thin-walled, round to elongate-oval,contains eversible hermaphroditic duct lined bysmall intensely stained cells which forms tempo-rary sinus-organ. Genital pore a wide transverseslit, median, at level of first septum.
Ovary oval to subtriangular, in anterior part oflast pseudosegment, overlaps vitellarium some-what dorsally. Uterine seminal receptacle large,dorsal to posterior part of vitellarium; Laurer’scanal not observed. Vitellarium compact, subovalto elongate-oval, larger than ovary, length c.11%of body length. Uterus thin-walled, fills free spacein pseudosegments of hindbody; metraterm notobserved. Eggs operculate, numerous.
Excretory pore wide, terminal to dorsosub-terminal; vesicle elongate-tubular, divides in ante-rior hindbody; arms unite at level of pharynx.
Remarks
Dimitrov et al. (1998) redescribed what theybelieved to be S. papernai from Mugil cephalusoff the Bulgarian Black Sea coast. We have found,however, that our newly collected material ofS. papernai from the Mediterranean showed anumber of differences when compared to thedescription of these authors. Thus, the size of theventral sucker in S. papernai is smaller, the seminalvesicle is longer and the posteriormost pseudoseg-ment is larger in relation to body length (seeDimitrov et al., 1998). After re-examination of thevoucher material, we reached the conclusion thatthis material can be distinguished from both theoriginal description of S. papernai by Overstreet(1977) and our collection of S. papernai in theabove characteristics.
S. dimitrovi n. sp. resembles S. maurepasi,S. valamugilis and S. papernai in the number anddistribution of the muscular septa. However, the
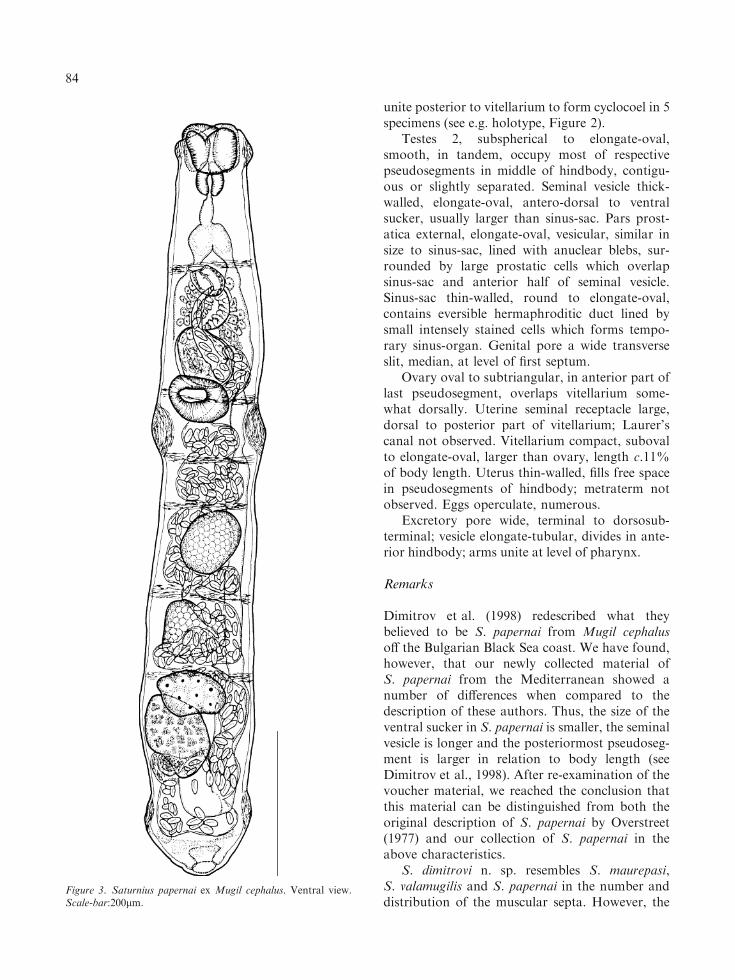
Figure 3. Saturnius papernai ex Mugil cephalus. Ventral view.Scale-bar:200lm.
84
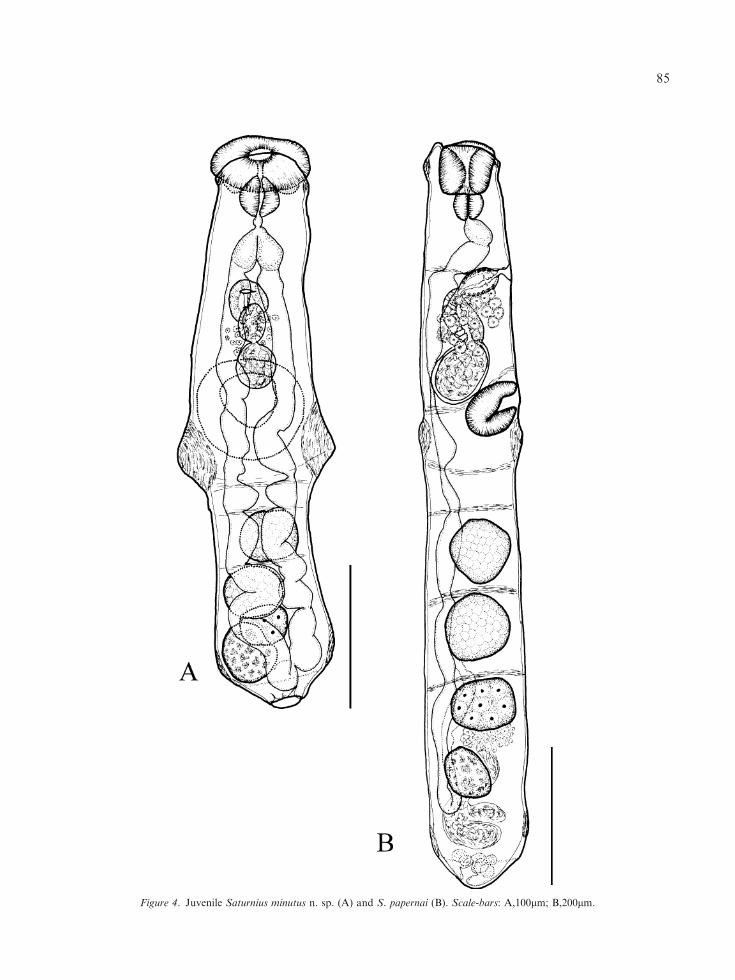
Figure 4. Juvenile Saturnius minutus n. sp. (A) and S. papernai (B). Scale-bars: A,100lm; B,200lm.
85
pars prostatica in S. maurepasi is small in relationto the sinus-sac and seminal vesicle which arelonger; and the posteriormost pseudosegment islonger in relation to the body length and thesuckers are smaller, although the upper range forbody size is higher. S. valamugilis was describedfrom specimens with a similar size of body, which,however, possess smaller suckers, pharynx, testesand ovary; the different host and distant geo-graphical area also tend to support the distinctstatus of the two species.
S. dimitrovi n. sp. appears most similar to thesympatric (both forms occur in mullets from theEbro Delta and the Black Sea) S. papernai (sensustricto), which was originally described fromM. cephalus on the Sinai coast of the EasternMediterranean (Overstreet, 1977). Similarities in-clude: the presence of three clearly distinguishablemuscular flanges; the number of septa, which arestrongly muscular; the somewhat asymmetricalappearance of the anterior margin of the oralsucker; and the overlap in the lower range of mostmetrical features. However, the posteriormostpseudosegment in S. papernai is wider and dis-tinctly more elongate in relation to the body; theventral sucker muscular flange length is similar tothe length of ventral sucker (means for MFL/VSL,99±13 vs 69±8%); and the latter is less promi-nent in S. papernai. Furthermore, although almostall measurements of S. dimitrovi exhibit an overlapwith the lower range of S. papernai (see Table 1),the univariate analyses of variance revealed thatthe two species are distinguishable with respect to
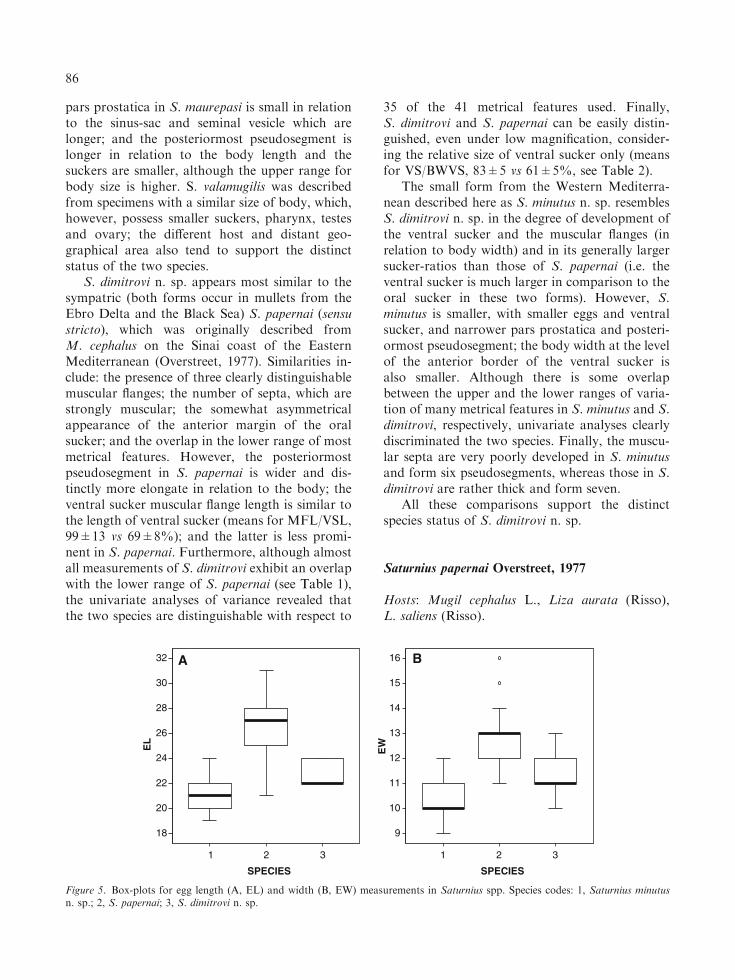
35 of the 41 metrical features used. Finally,S. dimitrovi and S. papernai can be easily distin-guished, even under low magnification, consider-ing the relative size of ventral sucker only (meansfor VS/BWVS, 83±5 vs 61±5%, see Table 2).
The small form from the Western Mediterra-nean described here as S. minutus n. sp. resemblesS. dimitrovi n. sp. in the degree of development ofthe ventral sucker and the muscular flanges (inrelation to body width) and in its generally largersucker-ratios than those of S. papernai (i.e. theventral sucker is much larger in comparison to theoral sucker in these two forms). However, S.minutus is smaller, with smaller eggs and ventralsucker, and narrower pars prostatica and posteri-ormost pseudosegment; the body width at the levelof the anterior border of the ventral sucker isalso smaller. Although there is some overlapbetween the upper and the lower ranges of varia-tion of many metrical features in S. minutus and S.dimitrovi, respectively, univariate analyses clearlydiscriminated the two species. Finally, the muscu-lar septa are very poorly developed in S. minutusand form six pseudosegments, whereas those in S.dimitrovi are rather thick and form seven.
All these comparisons support the distinctspecies status of S. dimitrovi n. sp.
Saturnius papernai Overstreet, 1977
Hosts: Mugil cephalus L., Liza aurata (Risso),L. saliens (Risso).
1 3
SPECIES
18
20
22
24
26
28
30
32
EL
1 3
SPECIES
9
10
11
12
13
14
15
16
EW
A
2 2
B
Figure 5. Box-plots for egg length (A, EL) and width (B, EW) measurements in Saturnius spp. Species codes: 1, Saturnius minutusn. sp.; 2, S. papernai; 3, S. dimitrovi n. sp.
86
Localities: M. cephalus: The Ebro Delta (SpanishMediterranean coast); L. aurata and L. saliens: offthe Black Sea coast of Bulgaria (at Sozopol).Site: Embedded between the grinding stomachlining and the layer of glandular cells.Voucher material: BMNH 2005.11.10.10-19.
Description (Figures 3, 4B)
[Based on 27 whole-mounted adult specimens;metrical data in Tables 1, 2]. Body large, elongate,cylindrical, with obtuse posterior extremity andmaximum width at levels of ventral sucker flangeand posterior flange. Width at mid-level of ventralsucker 95–136 (119±12). Tegument unarmed,with fine longitudinal and transverse striations.
Three circular muscular flanges present; (i)anterior (oral) flange well developed, muscular,located approximately at mid-level of oral sucker;(ii) second (ventral sucker) flange strongly muscu-lar, forming 2 lateral circular protuberances,provided with concentric muscles, with cone length(anterior-posterior) 46–91% (65±11%) and width(lateral) 19–36% (28±5%) [17–32% (23±4%) ofbody width at mid-level of ventral sucker], locatedmostly posterior to ventral sucker; tegumental foldcaused by musculature of second flange at somedistance from posterior margin of ventral sucker;(iii) third flange located in posterior third of lastpseudosegment, well developed.
Body with 7 clearly distinguishable pseudoseg-ments separated by thick transverse fibrous septa.Anteriormost septum at level of genital pore;second septum at anterior level of ventral suckerflange; 4 other septa form 4 pseudosegments ofdifferent sizes located entirely in hindbody: ante-rior smallest, middle pair similar in size andposteriormost largest, 25–35% (28±2%) of bodylength.
Oral sucker thick-walled, cup-shaped, withterminal aperture; anterior portion of suckerprotrusible, with flared anterior rim. Ventralsucker muscular, oval to transversely oval, smallerthan oral sucker. Prepharynx absent; pharynxmuscular, elongate-oval. Oesophagus short; intes-tine bifurcates at about middle of first septum.Caeca wide, with constrictions at levels of septa;most posterior portions masked by the ovary,vitellarium and uterus; cyclocoel observed in 1juvenile specimen (Figure 4).
Testes 2, spherical or with slightly irregularshape, smooth, slightly oblique, in fifth and sixthpseudosegments. Seminal vesicle thick-walled,elongate-oval, curves dorsally to anterior portionof ventral sucker, distinctly larger than sinus-sac.Pars prostatica external, large, elongate-oval,vesicular, lined with large anuclear blebs andsurrounded by large prostatic cells which partlyoverlap sinus-sac and anterior part of seminalvesicle. Sinus-sac thick-walled, elongate-oval, con-
Table 3. Classification results of two stepwise LDAs with (A) ln-transformed data and (B) size-adjusted data applied to six sam-ples of 69 specimens of Saturnius defined by species and geographical origin.
Analysis Predicted group
membership
Actual group membership Total
S. minutus
(S)
S. papernai
(S)
S. papernai
(B)
S. dimitrovi
(B)
S. dimitrovi
(S)
S. papernai
(T)
A S. minutus (S) 12 0 0 0 0 0 12
S. papernai (S) 0 19 0 0 0 0 19
S. papernai (B) 0 1 6 0 0 0 7
S. dimitrovi (B) 0 0 0 18 2 0 20
S. dimitrovi (S) 0 0 0 0 6 0 6
S. papernai (T) 0 0 0 0 0 5 5
B S. minutus (S) 12 0 0 0 0 0 12
S. papernai (S) 0 14 5 0 0 0 19
S. papernai (B) 0 3 4 0 0 0 7
S. dimitrovi (B) 0 0 0 18 2 0 20
S. dimitrovi (S) 0 0 0 1 5 0 6
S. papernai (T) 0 0 0 0 0 5 5
S, Spanish Mediterranean coast; B, Bulgarian Black Sea coast; T, type-material of S. papernai.
87
tains eversible hermaphroditic duct lined byintensely stained cells. Genital pore a transverseslit, median, at level of first septum.
Ovary oval to subtriangular, in anterior part oflast pseudosegment, ventral to caeca and partlyoverlapping vitellarium dorsally. Uterine seminalreceptacle large, postero-dorsal to vitellarium;Laurer’s canal not observed. Vitellarium compact,smooth, elongate-oval, larger than ovary, lengthc.9% of body length. Uterus thin-walled, extendsposterior to vitellarium. Eggs very numerous.
Excretory pore wide, terminal to dorso-subter-minal; arms of excretory vesicle unite at level ofpharynx.
Remarks
Of the six species of Saturnius, only S. mugilis andS. papernai have previously been reported from theMediterranean basin. However, only two docu-mented records of the latter species exist (seeOverstreet, 1977; Dimitrov et al., 1998). As wehave shown, the latter record represents a speciesnew to science, i.e. S. dimitrovi n. sp. On the otherhand, the newly collected S. papernai from off theEbro Delta described here fromM. cephalus agreesvery well with the original description of thespecies given by Overstreet (1977) (see Table 1 formetrical data). It also exhibits the main differentialcharacters which distinguish S. papernai from itscongeners: a cup-shaped oral sucker with a flaredanterior margin and a relatively short posterior-most pseudosegment (see Overstreet, 1977). Ourstudy, therefore, represents the second docu-mented record of S. papernai in the Mediterra-nean. L. saliens is a new host record and L. auratais the first documented record for S. papernai inthe Mediterranean basin.
Statistical comparisons
Univariate analyses of variance revealed thatS. papernai, S. dimitrovi n. sp. and S. minutus n.sp. are morphometrically distinguishable withrespect to the most metrical features used (allp<0.0001, see Table 2 for means). Moreover,although a range overlap was detected in egg-size(see Table 1), the three species differed signifi-cantly with respect to both the length(F1,174=114.6, p<0.0001; multiple comparisonsp<0.001) and width (F1, 174=101.6, p<0.0001;multiple comparisons p<0.0001) of the eggs (seealso Table 2 and Figure 5). Both the conventional
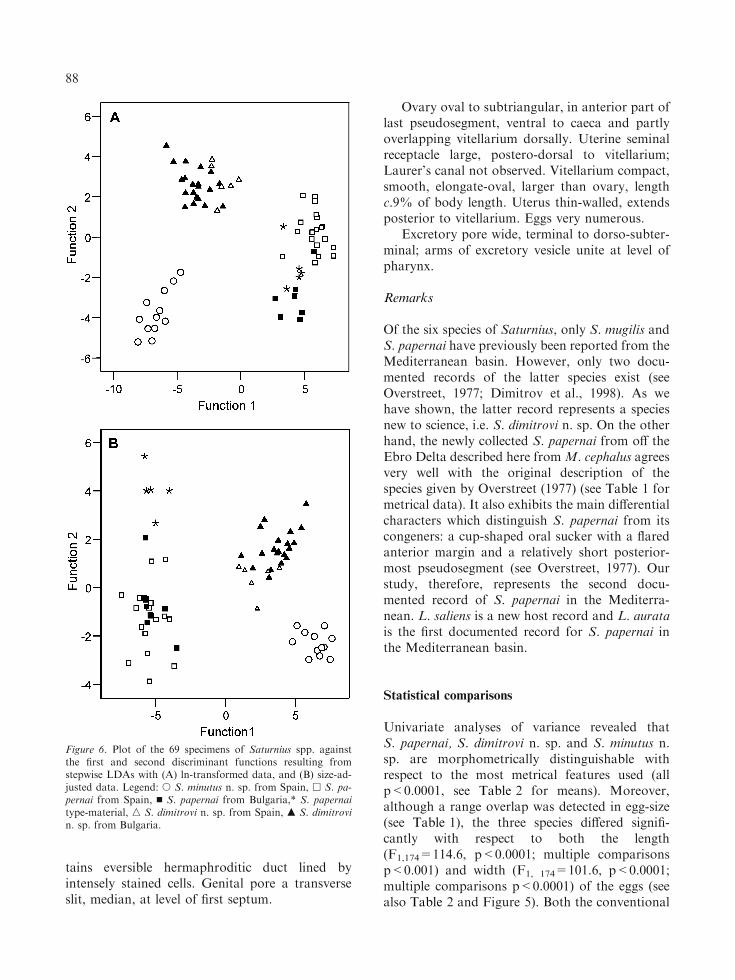
Figure 6. Plot of the 69 specimens of Saturnius spp. againstthe first and second discriminant functions resulting fromstepwise LDAs with (A) ln-transformed data, and (B) size-ad-justed data. Legend: s S. minutus n. sp. from Spain, h S. pa-pernai from Spain, n S. papernai from Bulgaria,* S. papernaitype-material, n S. dimitrovi n. sp. from Spain, m S. dimitrovin. sp. from Bulgaria.
88

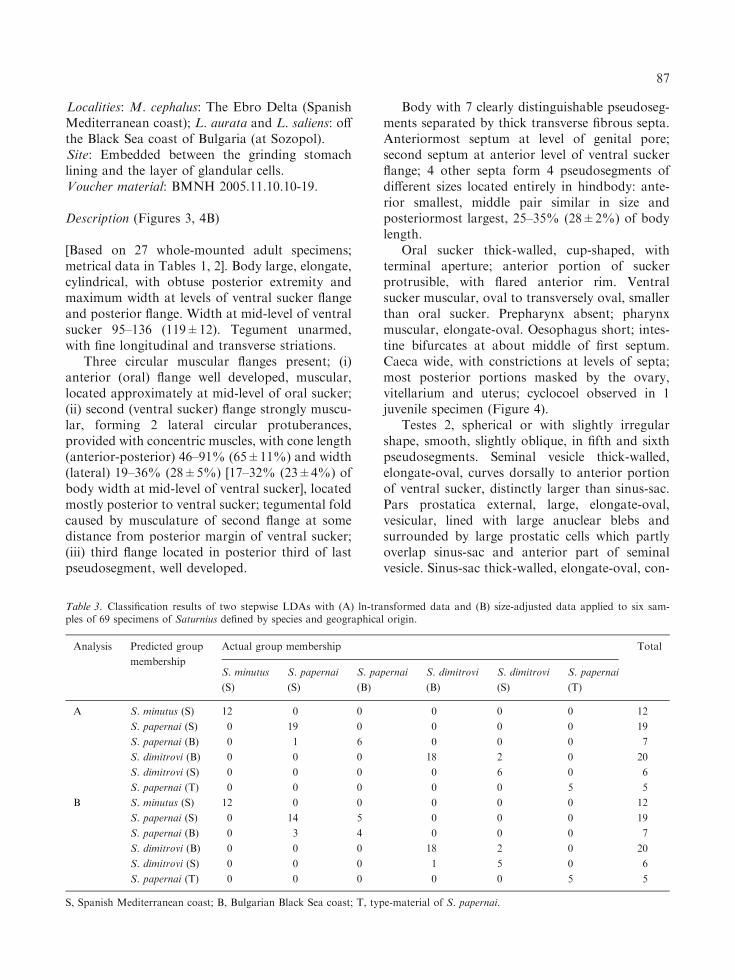
LDA with ln-transformed data (Analysis A) andits size-adjusted counterpart (Analysis B) wereable to correctly allocate all specimens to theirspecies designations based on morphology (i.e.100% successful classification rate, see Figure 6for a plot of the specimens against the first twodiscriminant functions). Furthermore, both mod-els generated by the stepwise procedures selectedthe same set of six variables for optimal separationbetween samples: the length of the forebody,ventral sucker and posterior testis, the length andwidth of the posteriormost pseudosegment, andthe width of the muscular flange at ventral sucker.Almost all specimens were also assigned to thecorrect population (locality) with the exception ofthree (Analysis A, misclassification rate 4.4%) and11 specimens (Analysis B, misclassification rate15.9%) (see Table 3 for details).
Discussion
In his revision of Saturnius, Overstreet (1977)suggested that ‘‘If this report stimulates examina-tion of mullet from different localities, someencountered worms probably will be new speciesand different forms’’. However, although all mullethost species in the Mediterranean basin havebeen subjected to intensive studies, the existingrecords indicate the presence of only two species,S. papernai and S. mugilis. S. papernai has beenreported in Mugil cephalus (see Merella & Gari-ppa, 1998, 2001; Domnich & Sarabeev, 2000a;Dmitrieva & Gaevskaya, 2001; Zhdamirov &Mal’tsev, 2001); M. soiuy (see Sarabeev, 2000;Domnich & Sarabeev, 2000a,b,c,d; Sarabeev &Domnich, 2000; Dmitrieva & Gaevskaya, 2001;Zhdamirov & Mal’tsev, 2001); Liza aurata (seeDomnich & Sarabeev, 2000a; Dmitrieva & Gae-vskaya, 2001; Pronkina, 2001; Gaevskaya & Kor-niychuk, 2003); and L. ramada (see Di Cave et al.,1997). Records of S. mugilis include reports fromM. cephalus (see Gaevskaya & Korniychuk, 2003);L. aurata (see Dmitrieva & Gaevskaya, 2001;Zhdamirov & Mal’tsev, 2001; Gaevskaya & Kor-niychuk, 2003); and L. saliens (see Solonchenko &Tkachuk, 1985; Dmitrieva & Gaevskaya, 2001;Gaevskaya & Korniychuk, 2003). In addition,material unidentified to species level belonging toSaturnius has been reported in Chelon labrosus,
L. aurata, L. ramada and L. saliens in the WesternMediterranean (see D’Amelio et al., 1995; Merella& Garippa, 1998, 2000, 2001).
Although the number of Saturnius spp. recordsfrom the Mediterranean and the Black Seamay appear substantial, most are not original(e.g. Dmitrieva & Gaevskaya, 2001; Gaevskaya &Korniychuk, 2003), whereas the remaining appearto reiterate results of single host populationsamples; virtually none of these provide supportiveevidence for the species identification [only Zhd-amirov & Mal’tsev (2001) provide some measure-ments in an abstract]. It is therefore not surprisingthat this first large-scale morphological study onmullet parasites in the Mediterranean, has in-creased the diversity of the genus by adding twonew species.
In concert, univariate and multivariate analyseshave demonstrated that S. papernai, S. dimitrovin. sp. and S. minutus n. sp. are morphometricallydistinguishable. Our results also verified that usingthe methods of LDA classification, the discrimi-nation of the three closely related and possiblysympatric (S. minutus has not yet been recoveredfrom Black Sea mullet populations) species ofSaturnius may be achieved using a limited numberof metrical variables. In particular, by applying thesize-adjusted LDA, the large difference in sizebetween the three forms is controlled for and, thus,the differentiation observed should be mostlyattributable to shape differences between thespecies (Junger et al., 1995). The LDAs were alsoefficient at assigning the specimens of each speciesto the correct population (locality). Notably, ouranalyses suggest that interpopulational distancesare smaller than interspecific, thus reinforcing thenotion of three distinct morphometric forms ofSaturnius spp. (as shown in Figure 6 and Table 3).Although the intraspecific differences might pointto the existence of geographical morphometricvariants (some of which might be clonal siblings),they might well reflect pseudoreplication derivedfrom samples coming from related hosts or fixed/processed differently (e.g. in the case of thetype-material of S. papernai and S. dimitrovi n.sp. collected by Dimitrov et al., 1998). Our futureefforts, focusing on a detailed morphological studyof large sets of specimens from Liza spp. andM. soiuy, may help testing the prediction (seeabove) of Overstreet (1977), thus revealing theactual diversity of Saturnius in the Mediterranean.
89

One important finding of our study is thepresence of a cyclocoel in all three speciesdescribed. It should be stressed that the mostposterior region of the intestine was very difficultto observe in adult specimens due to its dorsallocation; it was obscured by the ovary, vitellariumand uterus which typically overlap it ventrally.Observation was most complicated in the speci-mens of S. papernai due to the large number ofeggs (75–602), a significant proportion of whichwas located in the last body pseudosegment.Examination of the type-material of S. papernairevealed a similar situation, where we also failed toclearly observe blind caeca. However, caeca can beseen to unite posterior to the vitellarium to form acyclocoel in juvenile specimens of S. papernai andS. minutus n. sp., as well as in the holotype ofS. dimitrovi n. sp. (which was mounted dorsallyand also had fewer eggs in the posteriormostpseudosegment). The presence (in BunocotyleOdhner, 1928) and absence (in Saturnius) ofcyclocoel has so far been used as an importantfeature in distinguishing the two genera within theBunocotylinae (e.g. Gibson, 2002). Although atpresent we suggest that the presence of a cyclocoelis a common feature for all Saturnius spp., furthermorphological studies and re-examination of typesof the other members of the genus are required totest this prediction.
Acknowledgements
We are indebted to Patricia Pilitt for the loan ofspecimens from the United States National Para-site Collection and F. Agustin Jimenez for the loanof specimens from the Collection of the Harold W.Manter Parasitological Laboratory. We thank RodBray who kindly read the manuscript. The first au-thor benefits from a doctoral fellowship from theMinistry of Science and Education of Spain (AP-2004–2999). The study was funded by INTASgrant 03–51–5998 and partly supported by a Syn-thesys grant GB-TAF-329 to AK which enabledher to visit the Natural HistoryMuseum, London.
References
D’Amelio, S., Paggi, L., Berrilli, F., Di Cave, D. Orecchia, P.,Gerasi, L., Markus, E.E. & Paperna, I. (1995) Divergence of
the parasite communities of Liza ramada from coastal hab-itats of Israel and Italy. In: Hoffmann, R.W. (Ed.) ‘Programand Abstracts of the Fourth International Symposium of FishParasitology, Munich, October 3–7, 1995’, p. 58.
Darroch, J.N. & Mosimann, J.E. (1985) Canonical and prin-cipal components of shape. Biometrika, 72, 241–252.
Di Cave, D., Kennedy, C.R., D’Amelio, S., Berrilli, F. & Or-ecchia, P. (1997) Struttura della comunita del metazoiparassiti di Liza ramada (Risso, 1826) nel lago di Burano.Biologia Marina Mediterranea, 4, 321–323.
Dimitrov, G.I., Arapoglou, F.K. & Georgiev, B.B. (1998) Firstrecord of Saturnius papernai Overstreet, 1977 in the BlackSea, with a review of the genus Saturnius Manter, 1969(Digenea, Bunocotylidae). Systematic Parasitology, 40,43–48.
Dmitrieva, E.V. & Gaevskaya, A.V. (2001) Parasitological as-pects of mugilids mariculture and of their introduction intothe Sea of Azov and the Black Sea. Ekologiya Morya, 55,73–78. (In Russian).
Domnich, I.F. & Sarabeev, V.L. (1999) Trematodes of thegenus Bunocotyle (Trematoda, Halipegidae) from the haar-der (Mugil so-iuy) acclimatized in the Azov Sea. Para-zitologiya, 33, 67–70. (In Russian).
Domnich, I.F. & Sarabeev, V.L. (2000a) Contemporary faunaof fish parasites in the northern part of the Azov Sea. VisnikZaporiz’kogo Derzhavnogo Universitetu, 1, 224–230. (InUkrainian).
Domnitch, I.F. & Sarabeev, V.L. (2000b) The trematodes of theFar-East-mullet-pelingas (Mugil soiuy) in the Azov Seaaquatories. Vestnik Zoologii, 34, 7–15. (In Russian).
Domnich, I.F. & Sarabeev, V.L. (2000c) Parasitic fauna struc-ture of the pilengas in the Azov Sea. Acta Parasitologica, 45,268.
Domnich, I.F. & Sarabeev, V.L. (2000d) Forming of the par-asitic fauna of the pilengas in the Azov Sea. Acta Parasito-logica, 45, 265–266.
Fischthal, J.H. (1977) Some digenetic trematodes of marinefishes from the barrier reef and reef lagoon of Belize. Zoo-logica Scripta, 6, 81–88.
Gaevskaya, A.V. & Korniychuk, Y.M. (2003) [Parasiticorganisms as a component of ecosystems of the Black Seanear-shore zone of Crimea]. In: Eremeev, V.N. & Gaevs-kaya, A.V. (Eds) [Modern condition of biological diversity innear-shore zone of Crimea (Black Sea sector)]. Sevastopol:EKOSI-Gidrophizika, pp. 425–490.
Georgiev, B.B., Biserkov, V.Y. & Genov, T. (1986) In totostaining method for cestods in iron acetocarmine. Helm-inthologia, 23, 279–281.
Gibson, D.I. (2002) Family Hemiuridae. In: Gibson, D.I.,Jones, A. & Bray, R.A. (Eds) Keys to the Trematoda. Vol. 1.Wallingford: CAB International, pp. 305–340.
Jungers, W.L., Falsetti, A.B. & Wall, C.E. (1995) Shape, rela-tive size and size-adjustments in morphometrics. Yearbook ofPhysical Anthropology, 38, 137–161.
Manter, H.W. (1969) Three new genera of hemiurid trematodesfrom Australian fishes. Journal of the Fisheries ResearchBoard of Canada, 26, 787–792.
Merella, P. & Garippa, G. (1998) Parasites of grey mullets fromthe Oristano lagoons (Sardinia). Parassitologia, 40 (suppl. 1)(EMOP Abstract), p. 106.
Merella, P. & Garippa, G. (2000) Parasite fauna of mugilids(Osteichthyes: Mugilidae) from the Mistras Lagoon (Sardi-nia-Western Mediterranean). Parassitologia, 42 (Suppl. 1),(EMOP Abstract), p. 177.
90

Merella, P. & Garippa, G. (2001) Metazoan parasites of greymullets (Teleostei: Mugilidae) from the Mistras Lagoon(Sardinia, Western Mediterranean). Scientia Marina, 65,201–206.
Norusis, M.J. (2002) SPSS�11.0 Guide to data analysis. NewJersey: Prentice Hall Inc.
Overstreet, R.M. (1977) A revision of Saturnius Manter, 1969(Hemiuridae: Bunocotylinae) with descriptions of two newspecies from the striped mullet. In: Excerta parasitologica enmemoria del doctor Eduardo Caballero y Caballero. Institutode Biologıa, Universidad Nacional Autonoma de Mexico.Publicaciones Especiales, 4, 273–284.
Pronkina, N.V. (2001) Characters of formation of Liza aurata(Risso) fry helminth fauna from the Black Sea. EkologiyaMorya, 58, 50–52. (In Russian).
Rekharani, Z. & Madhavi, R. (1985) Digenetic trematodesfrom mullets of Visakhapatnam (India). Journal of NaturalHistory, 19, 929–951.
Sarabeev, V. (2000) [Seasonal dynamics of the parasitic infec-tion of the pilengas in the northern part of the Azov Sea.] In:
[Materials of the international scientific conference of youngscientists, ‘Aquatic biological resources and ways of their ra-tional use’.] Kiev: Institute of Fish Industry of UAAS, pp.97–99. (In Russian).
Sarabeev, V.L. & Domnich, I.F. (2000) Age-related dynamicsof parasitic infection of the pilengas (Mugil soiuy) in theMolochny Estuary of the Sea of Azov. Vestnik Zoologii, 14,6–12. (In Russian).
Solonchenko, A.I. & Tkachuk, L.P. (1985) Helminth infectionof mullets from Azov-Black Sea basin. Ekologiya Morya, 20,39–43. (In Russian).
Yamaguti, S. (1970) Digenetic trematodes of Hawaiian fishes.Tokyo: Keigaku Publishing Co., 436 pp.
Zhdamirov, V.N. & Mal’tsev, V.N. (2001) [The trematodegenus Saturnius Manter, 1969 (Trematoda, Bunocotylidae) –parasites of mullets in the Azov-Black Sea basin.] In:[‘Problems of ichthyopathology’. Materials of the First All-Ukranian Conference, Kiev, 23–26 October, 2001]. Kiev:Institute of Fish Industry of UAAS, pp.1–4 (In Russian).
91




















