
The spermatogenesis and the sperm structure of Terebrantia (Thysanoptera, Insecta
12
Tissue and Cell 42 (2010) 247–258 Contents lists available at ScienceDirect Tissue and Cell journal homepage: www.elsevier.com/locate/tice The spermatogenesis and the sperm structure of Terebrantia (Thysanoptera, Insecta) Eugenio Paccagnini a , David Mercati a , Fabiola Giusti a , Barbara Conti b , Romano Dallai a,∗ a Department of Evolutionary Biology, University of Siena, Via A. Moro 2, 53100 Siena, Italy b Department of Cultivation and Preservation of Ligneous Species “G. Scaramuzzi” (DCDSL), University of Pisa, Via del Borghetto 80, 56100 Pisa, Italy article info Article history: Received 15 March 2010 Received in revised form 23 April 2010 Accepted 27 April 2010 Available online 9 June 2010 Keywords: Sperm structure Spermiogenesis thrips Electron microscopy Insect ultrastructure abstract Spermatogenesis and the sperm structure of the terebrantian Aeolothrips intermedius Bagnall are described. Spermatogenesis consists of two mitotic divisions; the second is characterized by the loss of half of the spermatids, which have pyknotic nuclei. Early spermatids have two centrioles, but when spermiogenesis starts, a third centriole is produced. The three basal bodies give rise to three flagella; later these fuse into a single flagellum which contains three 9 + 0 axonemes. The basal bodies are surrounded by a large amount of centriole adjunct material. During spermiogenesis this material contributes to the shifting of the three axonemes towards the anterior sperm region parallel to the elongating nucleus, and it is transformed into a dense cylinder. In the mature spermatids the three axonemes amalgamate to create a bundle of 27 doublet microtubules. Near the end of spermiogenesis the dense cylinder of the centriole adjunct lies parallel to the nucleus and the axonemes. It ends where the mitochondrion appears at half-sperm length. We confirm that Terebrantia testes have a single sperm cyst; their sperm are characterized by a cylindrical nucleus, three axonemes fused into one, a small mitochondrion and a short cylindrical centriole adjunct which corresponds to the dense body described in a previous work. The acrosome is lacking. At the midpoint of the anterior half of the sperm the outline of the cross-section is bilobed, with the nucleus contained in a pocket evagination of the plasma membrane. These characters are discussed in light of a comparison between Tubulifera and Terebrantia. © 2010 Published by Elsevier Ltd. 1. Introduction The sperm structure and spermatogenesis of thrips have been well studied recently (Paccagnini et al., 2006, 2007, 2009). These works have confirmed an aberrant spermatogenesis of the haploid males (Bournier, 1956; Bode, 1988; Heming, 1995): two mitotic rather than meiotic divisions, the second of which eliminates half the sperm cells (Paccagnini et al., 2006). The reason for this elimi- nation remains unknown. Another unexpected event is seen early in spermiogenesis: three centrioles, rather than the usual two, are present and each of them gives rise to a flagellum. The result is the production of a tri-flagellate spermatid cell, a unique finding among metazoans. As spermiogenesis progresses, the three flag- ella fuse into one and the three axonemes amalgamate to give rise to a microtubular bundle of 27 units (Paccagnini et al., 2007, 2009), running in a parallel helix with the nucleus. Thysanoptera comprise two suborders, Terebrantia and Tubu- lifera (Mound et al., 1980; Mound and Morris, 2007), which show some differences in sperm structure. Tubulifera testes have several ∗ Corresponding author. Tel.: +39 0577 234 412; fax: +39 0577 234 476. E-mail address: [email protected] (R. Dallai). cysts, each one with germ cell at a different stage of maturation (Heming, 1995). Tubuliferan sperm cells are anchored at the apical region to the cyst cell surrounding the sperm bundle, while Tere- brantia, which contain only a single sperm cyst of synchronously developing germ cells, lack this apical anchorage. Terebrantia also lack an acrosome, a characteristic already mentioned by Bode (1988) and Heming (1995), and their mitochondrion is smaller than in Tubulifera. Finally, Terebrantia have a bilobed sperm cross- section, at the half anterior sperm region (Paccagnini et al., 2009). One unresolved point in Terebrantia sperm structure is the nature of the dense body, which begins in the anterior region and extends posteriorly, remaining parallel to the nucleus and the axoneme, and ends at the level where the mitochondrion starts (Paccagnini et al., 2009). With this study of the spermatogenetic process of Aeolothrips intermedius Bagnall we have followed the entire sequence of germ cell differentiation and have established the nature of the different sperm components, the dense body included. 2. Materials and methods Aeolothrips intermedius Bagnall is a peculiar terebrantian species characterized by the production of a cocoon by the mature larva, 0040-8166/$ – see front matter © 2010 Published by Elsevier Ltd. doi:10.1016/j.tice.2010.04.008
Transcript of The spermatogenesis and the sperm structure of Terebrantia (Thysanoptera, Insecta

T(
Ea
b
a
ARRAA
KSSEI
1
wwmrtniptaetr
ls
0d
Tissue and Cell 42 (2010) 247–258
Contents lists available at ScienceDirect
Tissue and Cell
journa l homepage: www.e lsev ier .com/ locate / t i ce
he spermatogenesis and the sperm structure of TerebrantiaThysanoptera, Insecta)
ugenio Paccagninia, David Mercati a, Fabiola Giusti a, Barbara Contib, Romano Dallai a,∗
Department of Evolutionary Biology, University of Siena, Via A. Moro 2, 53100 Siena, ItalyDepartment of Cultivation and Preservation of Ligneous Species “G. Scaramuzzi” (DCDSL), University of Pisa, Via del Borghetto 80, 56100 Pisa, Italy
r t i c l e i n f o
rticle history:eceived 15 March 2010eceived in revised form 23 April 2010ccepted 27 April 2010vailable online 9 June 2010
eywords:perm structurepermiogenesis thripslectron microscopy
a b s t r a c t
Spermatogenesis and the sperm structure of the terebrantian Aeolothrips intermedius Bagnall aredescribed. Spermatogenesis consists of two mitotic divisions; the second is characterized by the lossof half of the spermatids, which have pyknotic nuclei. Early spermatids have two centrioles, but whenspermiogenesis starts, a third centriole is produced. The three basal bodies give rise to three flagella; laterthese fuse into a single flagellum which contains three 9 + 0 axonemes. The basal bodies are surroundedby a large amount of centriole adjunct material. During spermiogenesis this material contributes to theshifting of the three axonemes towards the anterior sperm region parallel to the elongating nucleus,and it is transformed into a dense cylinder. In the mature spermatids the three axonemes amalgamateto create a bundle of 27 doublet microtubules. Near the end of spermiogenesis the dense cylinder of
nsect ultrastructure the centriole adjunct lies parallel to the nucleus and the axonemes. It ends where the mitochondrionappears at half-sperm length. We confirm that Terebrantia testes have a single sperm cyst; their spermare characterized by a cylindrical nucleus, three axonemes fused into one, a small mitochondrion and ashort cylindrical centriole adjunct which corresponds to the dense body described in a previous work.The acrosome is lacking. At the midpoint of the anterior half of the sperm the outline of the cross-sectionis bilobed, with the nucleus contained in a pocket evagination of the plasma membrane. These characters
com
are discussed in light of a. Introduction
The sperm structure and spermatogenesis of thrips have beenell studied recently (Paccagnini et al., 2006, 2007, 2009). Theseorks have confirmed an aberrant spermatogenesis of the haploidales (Bournier, 1956; Bode, 1988; Heming, 1995): two mitotic
ather than meiotic divisions, the second of which eliminates halfhe sperm cells (Paccagnini et al., 2006). The reason for this elimi-ation remains unknown. Another unexpected event is seen early
n spermiogenesis: three centrioles, rather than the usual two, areresent and each of them gives rise to a flagellum. The result ishe production of a tri-flagellate spermatid cell, a unique findingmong metazoans. As spermiogenesis progresses, the three flag-lla fuse into one and the three axonemes amalgamate to give riseo a microtubular bundle of 27 units (Paccagnini et al., 2007, 2009),
unning in a parallel helix with the nucleus.Thysanoptera comprise two suborders, Terebrantia and Tubu-ifera (Mound et al., 1980; Mound and Morris, 2007), which showome differences in sperm structure. Tubulifera testes have several
∗ Corresponding author. Tel.: +39 0577 234 412; fax: +39 0577 234 476.E-mail address: [email protected] (R. Dallai).
040-8166/$ – see front matter © 2010 Published by Elsevier Ltd.oi:10.1016/j.tice.2010.04.008
parison between Tubulifera and Terebrantia.© 2010 Published by Elsevier Ltd.
cysts, each one with germ cell at a different stage of maturation(Heming, 1995). Tubuliferan sperm cells are anchored at the apicalregion to the cyst cell surrounding the sperm bundle, while Tere-brantia, which contain only a single sperm cyst of synchronouslydeveloping germ cells, lack this apical anchorage. Terebrantia alsolack an acrosome, a characteristic already mentioned by Bode(1988) and Heming (1995), and their mitochondrion is smallerthan in Tubulifera. Finally, Terebrantia have a bilobed sperm cross-section, at the half anterior sperm region (Paccagnini et al., 2009).
One unresolved point in Terebrantia sperm structure is thenature of the dense body, which begins in the anterior regionand extends posteriorly, remaining parallel to the nucleus and theaxoneme, and ends at the level where the mitochondrion starts(Paccagnini et al., 2009). With this study of the spermatogeneticprocess of Aeolothrips intermedius Bagnall we have followed theentire sequence of germ cell differentiation and have establishedthe nature of the different sperm components, the dense bodyincluded.
2. Materials and methods
Aeolothrips intermedius Bagnall is a peculiar terebrantian speciescharacterized by the production of a cocoon by the mature larva,

2 e and
iftmpdwc
awiaiNta
m3atdEmC
3
3
omta(rtcbn(2tcmmsatpno(
3
3
wtwub
48 E. Paccagnini et al. / Tissu
n which it transforms into an adult. Within this cocoon, which isormed by the secretory activity of the Malpighian tubule cells ofhe mature larva (Conti et al., 2010), the individual through three
oults becomes pro-pupa, pupa and adult. The total process ofupation takes about 10 days. Pupation makes it easier to selectifferent stages of spermatogenesis and sperm maturation, whichould be difficult to study in other species of Terebrantia without
ocoons.For our research, larvae of A. intermedius were captured from
lfalfa flowers in the neighbourhoods of Pisa, Italy, and togetherith alfalfa pollen and thrips tabaci larvae (both food sources for A.
ntermedius), were kept under laboratory conditions (20 ◦C, 80% R.H.nd natural photoperiod). Females with their ovipositor clearly vis-ble even inside the puparium can easily be separated from males.umerous males in stages from pro-pupa to adult were used for
he present study. Spermatogenesis usually starts after 2–3 daysfter the mature larva has completed the cocoon.
For transmission electron microscopy (TEM) observations,ales were dissected in phosphate buffer 0.1 M pH 7.2 to which
% sucrose was added (PB) to isolate testes. The material was fixedt 4 ◦C overnight in 2.5% glutaraldehyde in PB. After rinsing in PBhe material was postfixed in 1% osmium tetroxide for 2 h, dehy-rated in a graded series of alcohol and embedded in a mixture ofpon–Araldite resins. Thin sections, obtained with a Reichert ultra-icrotome, were routinely stained and observed with a TEM Philips
M 10 at 80 kV.
. Results
.1. Early spermatogenesis
Using many male specimens at the early pro-pupa stage webserved the entire sequence of cell division through which sper-atids are formed (Bournier, 1956; White, 1973; Heming, 1995). In
hese haploid males, the divisions are mitotic rather than meiotic,nd are also known as maturation divisions according to Bournier1956). The testes contain only a single sperm cyst, which is sur-ounded by a thin layer of cyst cells. In a cross-section throughestes of an individual 3 days after pupation, the pro-pupa containsells involved in mitotic division (Fig. 1A). Although cytoplasmicridges are present between germ cells, apparently division isot synchronized; cells at different stages of division are visibleFig. 1A and B). Cells of about 3 �m in diameter, with a nucleus of.4 �m diameter showing scattered small patches of chromatin inhe nucleoplasm, are intermingled with cells having a few densehromatin masses in the nuclear centre; cells, in which chro-atin is organized in a large irregular central dense band (possiblyetaphase stage) are also visible (Fig. 1B). Among these cells,
maller cells with a dense compact nucleus of 1.5 �m diameterre also visible (Fig. 1A and B). These pyknotic cells are destinedo degenerate. Both types of cells have two centrioles in parallelosition, confined in the narrow layer of cytoplasm between theuclear envelope and the cell membrane (Fig. 1C and D). Centri-les have the conventional structure with 9 microtubule tripletsFig. 1E).
.2. Spermiogenesis
.2.1. The tri-flagellate spermatid stageEarly spermatids can be found about 4–5 days after pupation,
hen the pro-pupa has already differentiated short pterothecae. Athis stage, a cross-section through the testes shows spermatid cellsith roundish nuclei about 2.4 �m in diameter. One centriole hasndergone replication, and the spermatids now have three, locatedeneath the nucleus (Fig. 2A and B). The three centrioles are arrayed
Cell 42 (2010) 247–258
triangularly (Fig. 1E) or, less frequently, almost linearly (Fig. 2C).The 9 triplets of each centriole are surrounded by a dense mate-rial, which further expands to fuse with that of the neighbouringcentrioles (Fig. 2C). Each centriole (now better called a basal body)extends for a short tract as an intracytoplasmic axoneme with a 9 + 2or 9 + 0 pattern; at a distance of about 0.2–0.3 �m from the nuclearenvelope, it gives rise to a flagellum. Thus, at this stage of spermio-genesis, the spermatid is a tri-flagellate cell (Fig. 2A and B). Lateralto the three axonemes, a circumvoluted mitochondrion is visible,created from the fusion of a few single mitochondria (Fig. 2B).
3.2.2. Differentiation into tri-axoneme spermatidsThe tri-flagellate spermatids are visible only for a short time
(Fig. 3A). They transform into tri-axoneme cells by a process ofexpansion of the plasma membrane of each flagellum. The processis not synchronous for all three flagella, and it is not rare to observethat two axonemes are already in the same cytoplasm, while thethird is still a distinct flagellum (Fig. 3E and F). The plasma mem-brane expansion shows regular small densities at the points wherethe flagellar axoneme exits from the cell and cisterns of smooth orrough endoplasmic reticulum with very few ribosomes surroundthe axoneme (Fig. 3B–D). In longitudinal section, these regions arecharacterized by invaginations of the plasma membrane, which arethickened by dense material (Fig. 3E and F).
The process of cell membrane expansion proceeds up to pointwhere the three flagella are inglobated into the spermatid cyto-plasm. The axonemes show 9 + 0 or 9 + 2 axonemal patterns, withthe two central tubules eccentric. Doublets have short arms. Thespermatid cytoplasm is rich in ribosomes, which diminish only atthe region of the intercellular bridges (Fig. 3E).
At this stage, a longitudinal section through the spermatidshows an elliptic nucleus about 2.6 �m long, with chromatin mate-rial uniformly distributed in small patches. A row of microtubulessurrounds the nucleus. There is a thin layer of cytoplasm around thenuclear region. Beneath the nucleus, a large mass of dense mate-rial (the centriole adjunct) embeds the three basal bodies and alsoexpands laterally (Fig. 4A, B and D). On one side of this material,few small mitochondria with cristae are visible (Fig. 4C). Micro-tubules, running longitudinally, surround the mitochondria; thethree axonemes are still encircled by series of cisterns of endo-plasmic reticulum (Fig. 4C and D).
3.2.3. Spermatid elongationThe next process in spermatid maturation is elongation. This
process is almost synchronous, thus a cross-section through testesof about 6-day old pro-pupa or early pupa shows the whole singlegerm cell cyst filled with elongated spermatid cells. The nucleushas become long and thin (Fig. 5A and B); in cross-section, its widthmeasures 0.4–0.5 �m and the chromatin is more condensed than inthe previous stage. Beneath the nucleus, a large globular dense massof centriole adjunct, 0.7 �m in diameter, embeds the three basalbodies (Fig. 5A). A layer of microtubules surrounds the material. Thethree axonemes are contained within the spermatid cytoplasm upto the most posterior tail region (Figs. 5B and 8C). The few remnanttail ends of spermatid flagella, which are still to be inglobated into acommon cytoplasm, contain only incomplete axonemes, made upof a few microtubule doublets, and the two central tubules (Fig. 5C).
The nucleus continues to elongate and the three axonemes aredisplaced towards the anterior of the cell. This process is likelymediated by the large dense mass of the centriole adjunct embed-ding the basal bodies; this structure elongates to give rise to a dense
cylinder located between the three axonemes and the nucleus(Fig. 5B). A cross-section through an elongating spermatid at thisstage can vary in diameter from about 0.7 to 0.4 �m (Fig. 6A–D). Formost of its anterior region, the spermatid cell consists of a cylindri-cal dense compact nucleus, which reaches the sperm tip (Fig. 6A);
E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258 249
Fig. 1. (A) Cross-section through a testis to show the germ cells at different stages of development of the second mitotic division. sp, spermatids; dc, degenerative cells;asterisk indicates nuclei with chromatin arranged in several blocks; arrow indicates a cytoplasmic bridge between spermatids. (B) Cross-section through germ cells at thesecond mitotic division. sp, spermatids; asterisk, nuclei at anaphase; dc, degenerating cells. (C) Cross-section through a spermatid to show the two parallel centrioles (c) andthe mitochondria (m); N, nucleus. (D) Cross-section through a degenerating cell, also with two parallel centrioles (c) and the mitochondria (m); (N), nucleus. (E) Cross-sectionthrough the centriole just before spermiogenesis. Note the microtubular triplets, running parallel to one another in a triangular array.

250 E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258
Fig. 2. (A and B) Longitudinal sections of early spermatids to show the three flagella. Due to the triangular array of the three centrioles (basal bodies) two flagella arec ). Note( ) in a(
jwi1sb
learly visible, the third being obliquely sectioned at the posterior region (asteriskarrowheads). m, mitochondrion; N, nucleus. (C) The three centrioles (basal bodiesarrowheads).
ust behind, a second dense cylindrical structure appears together
ith the nucleus (Fig. 6C). While the nuclear diameter is only 87 nmn the most anterior sperm end, a little more posteriorly it reaches60 nm. Finally, the axonemes occur in a 9 + 0 pattern. First oneingle axoneme appears, often without all its microtubular dou-lets (Fig. 6A and C); further behind, the second axoneme is evident
also that around the basal bodies a reduced centriole adjunct material is evidentvery early spermatid. They are embedded in the centriole adjunct dense material
and still more posteriorly, the dense cylindrical structure of the
centriole adjunct ends (Fig. 6A–D). Nucleus and centriole adjunctmaterial are surrounded by microtubules. Some microtubules arealso visible in the cytoplasm among the axonemes (Fig. 6D).When the three axonemes are present, together with thenucleus, they are located at the three vertices of a triangle, with

E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258 251
Fig. 3. (A) Cross-section through the three flagella 9 + 2 axonemes of an early spermatid. Note the asymmetry of the two central tubules. (B and C) Cross-section throughthe spermatid flagella during their progressive inclusion in the cytoplasm by expansion of the plasma membrane. The central axoneme in (B) shows doublets with outerradial densities (arrowheads). In (C) two axonemes are already enclosed in the cytoplasm while the third (upper left) is still within a flagellum. Arrowheads indicate thedensities at the point of the plasma membrane expansion. (D) Cross-section through the three axonemes when they are all enclosed in the cytoplasm. Note the long cisternsof endoplasmic reticulum (asterisk) surrounding the axonemes. (E and F) Longitudinal sections through spermatids during axoneme inclusion into the cytoplasm. The plasmamembrane is thicker at the point of the flagellar exit from the cell cytoplasm (arrowheads). Note also the reduction of ribosomes in the area of a cytoplasmic bridge (asterisk).N, nucleus; m, mitochondrion.

252 E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258
Fig. 4. (A) Cross-section through an early spermatid with three basal bodies, each one with triplets embedded in the dense centriole adjunct material. N, nucleus. (B) A shorttime after the spermatid in (A), the material of the centriole adjunct expands and embeds the three centrioles. (C and D) Longitudinal sections through spermatids; (C) lateraland (D) frontal view with the three axonemes (ax). N, nucleus; m, mitochondrion; arrowheads, dense material of the centriole adjunct.

E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258 253
Fig. 5. (A) Longitudinal section through spermatids still showing the very expanded centriole adjunct material (ca) embedding the basal bodies and the initial part of theaxonemes (ax). The nucleus (N) has compact chromatin. (B) Longitudinal section through spermatids in advanced stage of maturation when the dense nucleus (N) helicoidallyruns around the cylinder into which the centriole adjunct material (ca) is transformed. (ax), the three axonemes. (C) Cross-section through a testis during spermatid elongation.Note that spermatids have three axonemes each and that at the most posterior ends the axonemes are incomplete (asterisks).

254 E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258
Fig. 6. (A) Cross-section through the anterior region of almost mature spermatids. N, nucleus; ca, centriole adjunct transformed in a dense cylinder; ax, axonemes. Note thatall the structures are surrounded by microtubules. (B) Cross-section through almost mature spermatids showing their anterior and posterior (asterisks) regions. N, nucleus;ax, axonemes. (C) Cross-section through different levels of the almost mature spermatids. Numbers 1–6 indicate progressive distances from the anterior tip. N, nucleus; ca,centriole adjunct cylinder; ax, axoneme; m, mitochondrion. (D) Higher magnification of almost mature spermatids. N, nucleus; ax, axonemes; ca, centriole adjunct cylinder;m, mitochondrion. The two central spermatids show the amalgamation of the three axonemes. Note that all the structures are still surrounded by microtubules. (E and F)Longitudinal sections through almost mature spermatids with nucleus (N); the axonemes (ax) and the centriole adjunct cylinder (ca) running in helicoidal fashion.

e and
ta(awai
3
wTosspodmmc
rtaSt(btscfpotss
bmTbiam
mbmp
3
ceTtadeas(
E. Paccagnini et al. / Tissu
he nucleus lateral to them. Further behind, the nucleus endsnd a mitochondrion is visible together with the three axonemesFig. 6C). Finally, towards the posterior region only the axonemesre present, but they are fused into a single bundle of microtubules,hich decreases in number of elements towards the tail tip (Fig. 6B
nd C). In longitudinal section, the different sperm components runn helicoidal fashion (Fig. 6E and F).
.3. Amalgamation of the three axonemes
In 7–8 days from pupation, the testes of the pupae are filledith sperm, and there is an amalgamation of the three axonemes.
he regular axonemal pattern is lost and the microtubular doubletsf each axoneme are fused into a bundle. The process follows theame scheme of development already described in another thripsperm (Paccagnini et al., 2007). Either the three axonemes are dis-laced longitudinally (not beginning and ending at the same points)r they are of different lengths, because in cross-sections throughifferent levels of the sperm length there are different numbers oficrotubular doublets; only at half-sperm length the expected 27icrotubular units, from the amalgamation of all three axonemes,
an be counted (Fig. 6D).The sperm has a cross-section of about 0.1 �m at the most ante-
ior end, where only the nucleus is present. Immediately behind,he section increases to 0.3 �m and shows the nucleus and the par-llel cylindrical dense body of the centriole adjunct (Fig. 7A and B).till further, the bundle of microtubules also appears in the sec-ion, which now has an elongated shape. As in Paccagnini et al.2009), a cross-section through the anterior half-sperm length isilobed, with the nucleus on one side and the bundle of micro-ubular doublets and the small mitochondrion on the other; theection measures 0.37 �m × 0.23 �m (Fig. 7A and B). The cylindri-al dense body has gradually shortened until it is no longer visibleor all the anterior sperm length. In contrast, a cross-section of theosterior half-sperm length has a circular outline, with a diameterf about 0.28 �m. This region contains only the bundle of micro-ubular doublets and the small mitochondrion. This latter, however,oon disappears and the most posterior sperm region shows onlyome remnant doublets (Fig. 7A).
As we said, when observed at half-sperm length the axonemalundle consists of 27 microtubular doublets in an orderly arrange-ent, as formerly described by Bode (1988) and Dallai et al. (1991).
he bundle contains 3 rows of four doublets, 3 rows of three dou-lets and 3 rows of two doublets. The first microtubular element
n each of these classes is a singlet A-microtubule with two dyneinrms; in contrast, the last microtubular doublet in each class oficrotubular elements lacks the dynein arms (Fig. 7A and B).Due to the reciprocal shifting of the axonemes, the entire 27
icrotubular elements can be counted only for a short tract, whileoth anteriorly and posteriorly the number of microtubular ele-ents is less than 27, as there are only two or even a single axoneme
resent.
.4. The cyst cell wall
Terebrantia are characterized by testes with a single germell cyst (Heming, 1995). This consists of a very thin (0.2 �m)xternal pigmented wall, with a muscle layer (0.8 �m) beneath.he striated muscle fibers have a sarcomer 3 �m long. Beneathhis muscle wall a layer of parietal cells is evident. They shown elliptical nucleus 5 �m × 0.6 �m and a cytoplasm containing
ense polymorphic bodies and large mitochondria. The pari-tal cells often expand their cytoplasm into the cyst lumennd sometimes surround a bundle of sperm. Adjacent cells areeparated by long and circumvoluted cell junctional complexesFig. 8A–C).Cell 42 (2010) 247–258 255
4. Discussion
4.1. The two spermatogenetic divisions
This study of the terebrantian A. intermedius spermatogenesisyielded several important results. We confirmed, in this haplo-diploid species, that the spermatogenesis of the haploid maledevelops in a quite similar way to that described in the tubuliferanHaplothrips simplex, following two mitotic (maturation) divisions(Paccagnini et al., 2006). While the first division is a conventionalmitosis and gives rise to two equivalent cells, the second is aberrant,producing both a normal spermatid and a small cell character-ized by a condensed nucleus, which later degenerates. The causeof this degeneration is not yet known; we can exclude centrosomaldefects, since two centrioles are evident both in the spermatids andthe degenerating cells. The chromosomes of the species are verysmall and it was quite impossible to establish their number and thepresence of one heterochromosome, as hypothesized by Bournier(1956). He speculated that the spermatogonial cells (better calledgerm cells) would have a 2n + x karyotype; after meiosis, a set of n + xchromosomes would be inherited by the sperm, while the secondset of autosomes alone would be eliminated through the pyknoticnucleus. This interesting hypothesis assumes that spermatogoniaare diploid cells and that meiosis occurs; neither of these eventshas been verified. Spermatogenesis proceeds as in Apis melliferaand other hymenopteran species with haploid males generated byarrhenotoky parthenogenesis (White, 1973).
Another interesting observation, already described byPaccagnini et al. (2006), is the duplication of one of the twocentrioles inherited by young spermatids. In most insects, thespermatids formed by a normal meiosis of diploid primary sper-matocytes have only one centriole, due to the lack of a centriolereplication at the second meiotic division (Gonzáles et al., 1998;Callaini et al., 1999). In the haploid thrips male, since the seconddivision is mitotic, it produces the expected two centrioles. Whatis not easy to explain is why a third centriole is formed and whyonly one of the two centrioles duplicates. Perhaps the mechanismcontrolling the formation of centrioles does not function well,and a centriole that has already duplicated is unable to repeat theprocess (Wong and Stearns, 2003; Tsou and Stearns, 2006). If so,only the new centriole formed at the second mitotic division wouldbe able to duplicate. The result is the formation of a spermatidcell with three parallel centrioles, disposed at the vertices of atriangle or in linear array. Each of these centrioles (basal bodies)gives rise to a flagellum and a tri-flagellate cell is formed. Thispeculiar situation is shared only by some protistan Eimeria sp.microgametes (Scholtysek, 1965; Melhorn, 1972) and by Carpe-diomonas membranifera (Simpson and Patterson, 1999). This stage,however, is only ephemeral, as quite soon the three flagella arefused together into a single flagellum with three axonemes. Nofusions between spermatids, as suggested by Bode (1983), wereobserved, each cell maintaining its own cytoplasm. Sometimescytoplasmic bridges between close cells can be seen.
4.2. Spermiogenesis: amalgamation of the three axonemes andthe origin of the dense body in the anterior sperm region
Early in spermiogenesis, the spermatids have roundish or ellip-tical nuclei with a large dense mass of material beneath. This“centriole adjunct material” embeds the three basal bodies, withan axoneme emerging from each. The axoneme has a 9 + 0 pattern,
often also with two central tubules. A long, narrow, conventionalmitochondrion is lateral to the three axonemes. All the above struc-tures are surrounded by a layer of microtubules, which contributeto their elongation. Two main events characterize the followingphase of spermiogenesis: the fusion of the three axonemes in a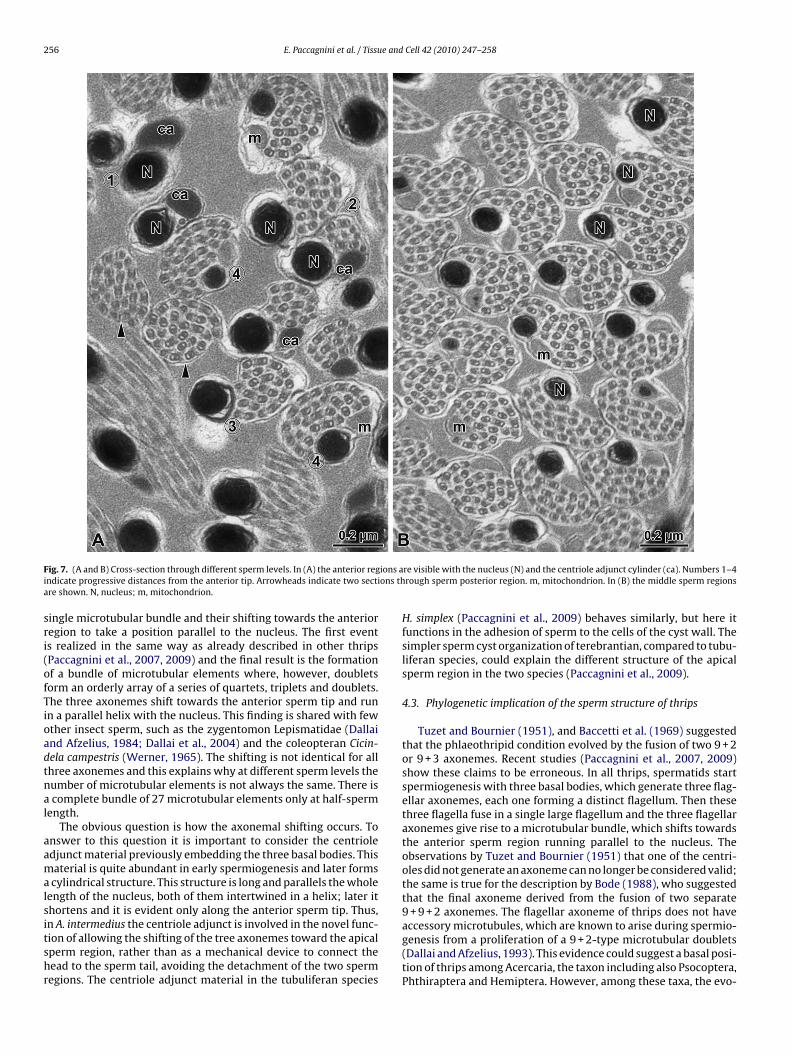
256 E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258
F ions ai ons tha
sri(ofTioadtnal
aamalsitshr
ig. 7. (A and B) Cross-section through different sperm levels. In (A) the anterior regndicate progressive distances from the anterior tip. Arrowheads indicate two sectire shown. N, nucleus; m, mitochondrion.
ingle microtubular bundle and their shifting towards the anterioregion to take a position parallel to the nucleus. The first events realized in the same way as already described in other thripsPaccagnini et al., 2007, 2009) and the final result is the formationf a bundle of microtubular elements where, however, doubletsorm an orderly array of a series of quartets, triplets and doublets.he three axonemes shift towards the anterior sperm tip and runn a parallel helix with the nucleus. This finding is shared with fewther insect sperm, such as the zygentomon Lepismatidae (Dallaind Afzelius, 1984; Dallai et al., 2004) and the coleopteran Cicin-ela campestris (Werner, 1965). The shifting is not identical for allhree axonemes and this explains why at different sperm levels theumber of microtubular elements is not always the same. There iscomplete bundle of 27 microtubular elements only at half-sperm
ength.The obvious question is how the axonemal shifting occurs. To
nswer to this question it is important to consider the centrioledjunct material previously embedding the three basal bodies. Thisaterial is quite abundant in early spermiogenesis and later formscylindrical structure. This structure is long and parallels the whole
ength of the nucleus, both of them intertwined in a helix; later ithortens and it is evident only along the anterior sperm tip. Thus,
n A. intermedius the centriole adjunct is involved in the novel func-ion of allowing the shifting of the tree axonemes toward the apicalperm region, rather than as a mechanical device to connect theead to the sperm tail, avoiding the detachment of the two spermegions. The centriole adjunct material in the tubuliferan speciesre visible with the nucleus (N) and the centriole adjunct cylinder (ca). Numbers 1–4rough sperm posterior region. m, mitochondrion. In (B) the middle sperm regions
H. simplex (Paccagnini et al., 2009) behaves similarly, but here itfunctions in the adhesion of sperm to the cells of the cyst wall. Thesimpler sperm cyst organization of terebrantian, compared to tubu-liferan species, could explain the different structure of the apicalsperm region in the two species (Paccagnini et al., 2009).
4.3. Phylogenetic implication of the sperm structure of thrips
Tuzet and Bournier (1951), and Baccetti et al. (1969) suggestedthat the phlaeothripid condition evolved by the fusion of two 9 + 2or 9 + 3 axonemes. Recent studies (Paccagnini et al., 2007, 2009)show these claims to be erroneous. In all thrips, spermatids startspermiogenesis with three basal bodies, which generate three flag-ellar axonemes, each one forming a distinct flagellum. Then thesethree flagella fuse in a single large flagellum and the three flagellaraxonemes give rise to a microtubular bundle, which shifts towardsthe anterior sperm region running parallel to the nucleus. Theobservations by Tuzet and Bournier (1951) that one of the centri-oles did not generate an axoneme can no longer be considered valid;the same is true for the description by Bode (1988), who suggestedthat the final axoneme derived from the fusion of two separate9 + 9 + 2 axonemes. The flagellar axoneme of thrips does not have
accessory microtubules, which are known to arise during spermio-genesis from a proliferation of a 9 + 2-type microtubular doublets(Dallai and Afzelius, 1993). This evidence could suggest a basal posi-tion of thrips among Acercaria, the taxon including also Psocoptera,Phthiraptera and Hemiptera. However, among these taxa, the evo-
E. Paccagnini et al. / Tissue and Cell 42 (2010) 247–258 257
Fig. 8. (A) Cross-section through the testis wall. This consists of a pigment layer (p); muscle fibers (ms) and underneath an epithelial layer (ep). Beneath the testis wall thesperm bundles are visible (sp). (B) Detail of (A) to show the muscle fibers (ms) and the epithelial wall (ep) adapted to surround the sperm bundles (sp). Epithelial cells rich inlarge mitochondria (m) and dense bodies (b) are joined by long and circumvoluted junctional complexes (arrowheads). (C) Detail of the testis wall to show the pigmentedlayer (p), the muscle fiber (ms) and the epithelial cells (ep) with nucleus (N), the dense bodies (b) and the large mitochondria (m). The testis wall surrounds a single germcell cyst here represented by spermatids (spd) which still have three separate axonemes. The nuclei (asterisks) of these spermatids contact the cyst wall.

2 e and
ls1NctvetndAa
trmb
eWttaTbB2HcgatfTiwmiatoa
ttalccatsfe
A
w
Werner, G., 1965. Untersuchungen über die Spermiogenese beim Sandläufer Cicin-
58 E. Paccagnini et al. / Tissu
utionary relationship seems to be Psocoptera + Phthiraptera on oneide and Thysanoptera + Hemiptera on the other side (Kristensen,981; Lyal, 1985). Nevertheless, the ontogeny of germ cells in theeoheegeria verbasci (Osborn) embryo resembles that in the pso-id Liposcelis divergens (Goss, 1952, 1953; see Heming, 1995). Tohis purpose, it is interesting to remember that psocids show quiteariable sperm models, with some taxa having sperm with two flag-llar axonemes (King and Ahmed, 1989). Moreover, according tohese Authors, Psocoptera and Thysanoptera originated from para-eopteran stock in the Permian era, since fossils have been recordeduring the Mesozoic, Tertiary and Recent eras. Mallophaga andnoplura have arisen only in more recent times (see Jamieson etl., 1999).
In conclusion, spermatological data indicate a relationship ofhrips with Phthiraptera and Psocoptera. The presence of threeather than two sperm flagellar axonemes is clearly an autapo-orphy of thrips, due to the duplication of one centriole at the
eginning of the spermiogenesis.Within the taxon, the two suborders Terebrantia and Tubulif-
ra can be better distinguished on the basis of our current results.hile it is still valid that Tubulifera have an acrosome, our observa-
ions clearly establish that the cylindrical structure running alonghe whole sperm and associated with a flattened vesicle is not thecrosome, but rather a remnant of the centriole adjunct material.hus the acrosome in the sperm of these species is represented onlyy a very thin cistern, amply visible in the giant tubuliferan sperm ofolothrips insularis and Compsothrips albosignatus (Paccagnini et al.,009), but very reduced in all the other Tubulifera (see for instance. simplex in the quoted paper by Paccagnini et al. (2009)). This con-lusion reconciles the fate of the centriole adjunct in the two thripsroups. In both Tubulifera and Terebrantia the centriole adjunct,fter having shifted towards the apical sperm region, contributeso transfer the three axonemes to the sperm tip; further it trans-orms in a cylindrical dense structure parallel to the nucleus. Inubulifera, as explained and illustrated by Paccagnini et al. (2009),t serves to anchor sperm cells to the cyst wall and runs along the
hole sperm length, perhaps to prevent the fluttering of the spermembrane and the disorganization of the microtubular bundle dur-
ng sperm beating. In Terebrantia, in contrast, after the centrioledjunct helps shift the three axonemes to the sperm tip, it reduceso a short cylinder running parallel to the nucleus apparently with-ut other function, if not that to give a certain resistance to thenterior sperm region.
Summarizing the two taxa of thrips, Tubulifera and Terebran-ia can be also differentiated on the basis of their sperm structure,he former showing a thin acrosome, a large mitochondrion andlong cylindrical centriole adjunct, which extends for the whole
ength of the sperm. Moreover, the transverse sperm profile is cir-ular or slightly elliptical, with the nucleus close to the other spermomponents. Terebrantia, on the contrary, lack an acrosome, havesmall mitochondrion and a cylindrical centriole adjunct only in
he anterior sperm region. The transverse profile of a terebrantianperm has a bilobed shape, with the nucleus positioned laterallyor the anterior half-sperm length, contained in a pocket-shapedxpansion of the plasma membrane.
cknowledgements
We thank Dr. E. Malatesta for technical assistance. This workas performed with a grant by Ministry of University and Research
Cell 42 (2010) 247–258
(MUR) to R.D. Thanks also to Dr. Ellen Swanson for the Englishrevision of the manuscript.
References
Baccetti, B., Dallai, R., Rosati, F., 1969. The spermatozoon of Arthropoda. IV. Corro-dentia, Mallophaga and Thysanoptera. J. Microsc. 8, 249–262.
Bode, W., 1983. Spermienstruktur und Spermiohistogenese bei Thrips validus(Insecta, Thysanoptera). Zool. Jahrb. Anat. 109, 301–318.
Bode, W., 1988. The spermatozoa of Thysanoptera and their relevance for system-atics. Acta Phytopathol. Entomol. Hungarica 23, 267–273.
Bournier, A., 1956. Contribution à l’étude de la parthénogenèse des Thysanoptèreset sa cytologie. Arch. Zool. Exp. Gén. 93, 220–317.
Callaini, G., Riparbelli, M.G., Dallai, R., 1999. Centrosome inheritance in insects:fertilization and parthenogenesis. Biol. Cell. 91, 355–366.
Conti, B., Berti, F., Mercati, D., Giusti, F., Dallai, R., 2010. The ultrastructure ofMalpighian tubules and the chemical composition of the cocoon of Aeolothripsintermedius Bagnall (Thysanoptera). J. Morphol. 271, 244–254.
Dallai, R., Afzelius, B.A., 1984. Paired spermatozoa in Thermobia domestica (Insecta,Thysanura). J. Ultrastr. Res. 86, 67–74.
Dallai, R., Afzelius, B.A., 1993. Development of the accessory tubules of insect spermflagella. J. Submicrosc. Cytol. Pathol. 25, 499–504.
Dallai, R., Afzelius, B.A., Lanzavecchia, S., Bellon, P.L., 1991. Bizarre flagellum of thripsspermatozoa (Thysanoptera, Insecta). J. Morphol. 209, 343–347.
Dallai, R., Carapelli, A., Nardi, F., Fanciulli, P.P., Lupetti, P., Afzelius, B.A., Frati, F., 2004.Sperm structure and spermiogenesis in Coletinia sp. (Nicoletiidae, Zygentoma,Insecta) with a comparative analysis of sperm structure in Zygentoma. TissueCell 36, 233–244.
Gonzáles, C., Tavosanis, G., Molinari, C., 1998. Centrosomes and microtubule orga-nization during Drosophila development. J. Cell Sci. 111, 2697–2706.
Goss, R.J., 1952. The early embryology of the book louse, Liposcelis divergens Badonnel(Psocoptera: Liposcelidae). J. Morphol. 91, 135–167.
Goss, R.J., 1953. The advanced embryology of the book louse, Liposcelis divergensBadonnel (Psocoptera: Liposcelidae). J. Morphol. 92, 157–191.
Heming, B.S., 1995. History of the germ line in male and female thrips(Thysanoptera). In: Parker, B.L:, Skinner, M., Lewis, T. (Eds), Thrips Biol-ogy and Management. Proceedings of the 1993 International Conference onThysanoptera. Towards Understanding Thrips Management. 28–30 September1993, Burlington, VT. Plenum Publishing Corp. pp. 505–535.
Jamieson, B.G.M., Dallai, R., Afzelius, B.A., 1999. Insects: Their Spermatozoa andPhylogeny, 555. Science Publishers, Enfield, NH, 105–112.
King, P.E., Ahmed, K.S., 1989. Sperm structure in Psocoptera. Acta Zool. (Stockholm)70, 57–61.
Kristensen, N.P., 1981. Phylogeny of insect orders. Ann. Rev. Entomol. 26, 135–157.Lyal, C.H.C., 1985. Phylogeny and classification of the Psocodea, with particular ref-
erence to the lice (Psocodea: Phthiraptera). Syst. Entomol. 10, 145–165.Melhorn, H., 1972. Elektronenmikroskopische Untersuchungen an Entwicklungssta-
dien von Eimeria maxima aus dem Haushuhn. Z: Parasitenk 40, 243–260.Mound, L.A., Morris, D.C., 2007. The insect order Thysanoptera: classification and
systematics. Zootaxa 1668, 395–411.Mound, L.A., Heming, B.S., Palmer, J.M., 1980. Phylogenetic relationships between
the families of recent Thysanoptera (Insecta). Zool. J. Linn. Soc. 69, 111–141.Paccagnini, E., De Marzo, L., Giusti, F., Dallai, R., 2006. The aberrant spermatogenesis
of the Haplothrips simplex (Buffa) (Thysanoptera): ultrastructural study. TissueCell 38, 177–180.
Paccagnini, E., Mencarelli, C., Mercati, D., Afzelius, B.A., Dallai, R., 2007. Ultrastruc-tural analysis of the aberrant axoneme morphogenesis in thrips (Thysanoptera,Insecta). Cell Motil. Cytoskeleton 64, 645–661.
Paccagnini, E., Lupetti, P., Afzelius, B.A., Dallai, R., 2009. New findings on spermultrastructure in thrips (Thysanoptera, Insecta). Arthropod Struct. Develop. 38,70–83.
Simpson, A.G.B., Patterson, D.J., 1999. The ultrastructure of Carpediemonas membran-ifera: (Eukaryota), with reference to the “excavate hypothesis”. Eur. J. Protist. 35,353–370.
Scholtysek, E., 1965. Die Mikrogameten Entwicklung von Eimeria perforans. Z. Zell-forsch. 66, 625–642.
Tuzet, O., Bournier, A., 1951. Le spermatozoïde de Neoheegeria verbasci (Osborn)(Insecte Thysanoptéres). C.R. Acad. Sci. Paris 233, 1316–1318.
Tsou, M.F., Stearns, T., 2006. Controlling centrosome number: licenses and bloks.Curr. Opin. Cell Biol. 18, 74–78.
dela campestris L. Zeitschr. Zellforsch. Mikrosk. Anat. 66, 255–275.White, M.J.D., 1973. Animal Cytology and Evolution, Third ed. Cambridge University
Press, 1–961.Wong, C., Stearns, T., 2003. Centrosome number is controlled by a centrosome-
intrinsic block to reduplication. Nature Cell Biol. 5, 539–544.




















