
An illustrated key to the genera of Thripinae (Thysanoptera) from South East Asia

Breeding system and thrips (Thysanoptera) pollinationin the endangered tree Ocotea porosa (Lauraceae):implications for conservationpsbi_354 31..40
ALINE DANIELI-SILVA* and ISABELA G. VARASSIN†*Post Graduate Program in Ecology and Conservation and †Department of Botany, Polytechnic Center, Federal University ofParaná, PO Box 19031, Postal Code 81531-980, Curitiba, PR, Brazil
Abstract
Ocotea porosa has been extensively exploited over the past few decades because of thequality of its wood. Today, populations are reduced and the species is now included inthe Red List of threatened species by the International Union for Nature Conservation.For conservation and management purposes, it is extremely important that we understandits reproductive ecology. Floral morphology was described based on field and scanningelectron microscopic examination. The reproductive system was determined throughexperimentally controlled pollination along with observations of pollen tube growth.Pollinators were identified through field observations. Flowers of O. porosa are small,shallow, inconspicuous, asynchronous and grouped in inflorescences. This species pre-sented self-compatibility, but did not reproduce through apomixy and spontaneous self-pollination was very rare (5%). Despite being monoclinal, flowers were protogynic, andthe gynoecium was receptive after the first day of anthesis when the anthers were closed.Spontaneous self-pollination was avoided by the extrorse position of the anthers of theinternal stamens. Frankliniella gardeniae (Thysanoptera) was the only pollinating speciesobserved and, after visiting several asynchronous flowers in the same inflorescence,favors geitonogamy. Together the small distance supposedly achieved by thrips in flightand the small population density of O. porosa can reduce the chances of cross-pollinationin this species. If so, conservation measures must include preservation of the currentpopulation and possibly planting to increase population density. This would reduce thedistance between individuals and increase genetic variability. Thrips as pollinators mustbe included in conservation planning for O. porosa.
Keywords: Imbuia, Ocotea porosa, pollination biology, reproductive biology, self-compatibility.
Received 10 July 2010; revision received 16 May 2011; accepted 17 June 2011
Introduction
Ocotea porosa (Ness & Mart.) Barroso (Lauraceae), knownlocally as Imbuia, has small, shallow, greenish-yellowflowers that give off no strong odor. Trees can reach 30 min height and 320 cm d.b.h. Ocotea porosa is found natu-rally in Araucaria Forests (part of the Atlantic Forest; Cas-tella & Britez 2004), in southeastern and southern Brazil(Reitz et al. 1978). Over the past few decades, this foresthas been subject to exploitation for the lumber industry
at such a scale that it is almost completely devastated(Mittermeier et al. 2005). Imbuia (O. porosa) was intenselyharvested because of the high quality of its wood, which isglobally sought after, and was exported in large quantitiesfor the manufacture of luxury furniture (Carvalho 2003).This pressure has resulted in the inclusion of O. porosa onthe list of threatened species in the Red List of the Inter-national Union for Nature Conservation, as well as theofficial Brazilian list of endangered species (Varty &Guadagnin 1998; Ministério do Meio Ambiente 2008).Population densities of Imbuia in general are not low(20 individuals/ha on average with d.b.h. > 10 cm) asrevealed by two studies of six populations in southern
Correspondence: Aline Danieli-SilvaEmail: [email protected]
Plant Species Biology (2013) 28, 31–40 doi: 10.1111/j.1442-1984.2011.00354.x
bs_bs_banner
© 2012 The AuthorsJournal compilation © 2012 The Society for the Study of Species Biology

Brazil (Amato 2007; Bittencourt 2007), one of whichincluded the populations of the present study area(Amato 2007). Nevertheless, the population structure pre-sents a discontinuity in size distribution, with an absenceof individuals in the intermediate size class (Amato 2007),even in well-preserved areas (Bittencourt 2007). This indi-cates natural discontinuous regeneration (Amato 2007).
Habitat degradation is changing the distribution andabundance of plant populations (Ribas & Kageyama 2004).This may affect the evolutionary history of populationsbecause of the spatial and reproductive isolation of popu-lations and/or individuals. This scenario may alter popu-lation genetic variability because of changes in allelicdispersal. To understand population genetic variability it isnecessary to know the breeding systems of natural popu-lations (Reis 1996) and also the behavior of pollinatorsbecause they are involved in pollen (and genetic) flow, thuscontributing to the genetic structure of plant populations(Fenster 1991; Franceschinelli & Kesseli 1999).
The reproductive system is known to affect the geneticstructure of plant populations (Holsinger 2000). Selferspecies have a smaller effective size and less allelicexchange among individuals (Schoen & Brown 1991).Selfers are more homozygotic and have fewer genotypeswithin populations when compared with outcrossingspecies (Holsinger 2000). Owing to this reduction ingenetic variability, selfers may be less able to respondadaptively to changing environments. In contrast, selfersmay present reproductive assurance when pollinators areabsent or in low densities (Holsinger 2000).
To understand the reproductive scenario under whichImbuia currently lives it is necessary to know its flowerbiology, pollinators and reproductive biology, includingthe chance of selfing. This can be done through manipu-lative studies involving pollen tube growth in manualself-pollination and cross-pollination. As plants from thefamily Lauraceae are usually large trees, their reproduc-tive biology has been poorly studied. The few studies thathave examined sexual characteristics in the family arebased on the population genetics of the genera Crypto-carya, Neolitsea and Ocotea (Gibson & Wheelwright 1996;Moraes & Derbyshire 2002; Moraes & Monteiro 2002;Kageyama et al. 2003; Wang et al. 2005). These studiesmerely provide indications of whether species areself-compatible and do not describe their reproductivesystems, much less evaluate the importance of its pollina-tors. Prior study of the species O. porosa only identifiedpotential pollinators, but did not describe the reproduc-tive system (Daros 2006; Bittencourt 2007; Y. Müller,unpubl. data, 2007). The dominant flower visitors werebeetles (Coleoptera, 35%) and flies (Diptera, 34%; Bitten-court 2007). In addition, although beetles were implicatedin another study as pollinators of O. porosa, they could notbe ruled out as herbivores (Y. Müller, unpubl. data, 2007).
In the present study, we describe for the first timethe reproductive system of the genus Ocotea by attempt-ing to answer the following questions: (i) does O. porosareproduce sexually; (ii) does O. porosa tend towards self-incompatibility, needing pollen vectors to reproduce; and(iii) what are the pollinators of O. porosa?
Materials and methods
Study area
Ocotea porosa was studied from September to November2007 at the Experimental Station of the Federal Universityof Paraná (25°34′18″S, 50°05′56″W), in São João do Triunfo,approximately 130 km from the city of Curitiba, in south-ern Brazil. The climate is humid and temperate with rainthroughout the year; Cfb, following the Koeppen classifi-cation (McKnight & Hess 2005). The average temperaturein the hottest month is < 22°C and in the coldest month is> 10°C (Maack 1981). During diurnal observations in thepresent study, the temperature ranged from 8 to 29°C andthe relative humidity ranged from 40 to 90%. This areawas originally dominated by Araucaria Forest. Today thestudy area is imbedded in an agricultural matrix, mostlytobacco cultivation (Fig. 1a).
The study area consists of a single forest fragmentof 32 ha. Although this region was never clear cut, selec-tive logging of Araucaria angustifolia and exploitation ofIlex paraguariensis were carried out. The formation isnow classified as altered primary forest or advancedsecondary forest (Schaaf et al. 2006). Ocotea porosa is adominant species in the fragment with approximately 17individuals/ha (Schaaf et al. 2006). Indicators suggest thatrecruitment rates are very low (Amato 2007).
Floral morphology and biology
Three adult plants were chosen for study, one in the inte-rior and two on the edges of the forest fragment, because
Fig. 1 Location of the study area. In the upper right corner is amap of Brazil showing the location of the study area in the Paranástate. Scale bar = 400 m.
32 A . D A N I E L I - S I LVA A N D I . G . VA R A S S I N
© 2012 The Authors Plant Species Biology 28, 31–40Journal compilation © 2012 The Society for the Study of Species Biology

access to the canopy was difficult and expensive. In addi-tion, a larger sample of trees was unnecessary for thescope of the present study. The high canopy was madeaccessible by a 15 m scaffold built at the tree trunk. For thetwo trees at the edges, a truck equipped with a ‘cherrypicker’ reached the canopy. Plant vouchers collected arenow at the Herbarium UPCB of the Universidade Federaldo Paraná under the following numbers: UPCB 60212(fragment interior) and UPCB 60213 (edge).
Flower morphology was described based on fieldobservations of fresh flowers, and flowers preserved in50% formalin acetic acid (FAA) were examined using ascanning electron microscope (JEOL JSM-5410; JEOL Ltd.,Tokyo 196-8558, Japan). For the microscopy study, theflowers were exposed to an acetone series, dehydratedand dried in a vacuum and then plated with gold-palladium for 4.5 min.
Field observations were used to follow the floweringprocess (anthesis). To determine the right time to do thebreeding manipulations, stigma receptivity was tested bymeans of peroxidase activity using hydrogen peroxide(H2O2), following Zeisler (1938) and Dafni (1992), in 20flowers from each tree. This process was carried outhourly to determine the beginning of receptivity. To deter-mine areas of greatest metabolic activity, possibly indicat-ing osmophores, flowers were dipped in a neutral redsolution (1:10 000) for 15 min, following the procedureoutlined by Vogel (1963).
To check for evidence of pollen malformations (relatedto apomixes; see Goldenberg 2000), pollen viability wasestimated from the material preserved in 50% FAA. Thepreserved pollen was stained with acetic carmine andthen viewed under an optical microscope (Dafni 1992). Asthe pollen from internal anthers is available before thepollen from external stamens, the pollen viability of bothanthers was tested. Internal (three anthers per flower) andexternal stamens (six anthers per flower) were randomlyselected beginning at the bud phase until the third day.Ten flowers and the first 100 pollen grains were countedper plant.
Reproductive system
To determine the reproductive system of O. porosa, expe-rimental pollinations were carried out using seventreatments (adapted from Radford et al. 1974): natural pol-lination (NP), that is, flowers had no manipulation, butwere marked and monitored; apomixy (AP), in whichstamens and styles were cut in the bud phase and bagged;spontaneous self-pollination (SP), in which flowers werebagged; manual self-pollination with pollen from internal(MSI) and from external (MSE) stamens; cross-pollination,with pollen from other plants from internal (CPI) andexternal (CPE) stamens, after which they were bagged. All
bags used in the experimental pollinations were made offine mesh voile. The bags did not prevent access by thrips,so all treatments may include the occurrence of naturalpollination, except the apomixy treatment because thestamens and styles were cut in the bud phase. As thestamens had different probabilities of self-pollen deposi-tion, we tested their eventual differences in reproduction.
Each treatment was carried out with 50 flowers per tree.The treatments NP, AP and SP were carried out on allthree trees, whereas the remaining treatments were onlycarried out on the two trees at the edge for logisticalreasons. To avoid potential high abortion rates as a resultof resource allocation, on average, only four flowers perinflorescence were subjected to controlled pollination.Fruit set in each treatment was evaluated 20 days afterexperimental manipulation.
To test for self-incompatibility we used controlled self-pollinations and cross-pollinations and observation ofpollen tube growth in both treatments (Martin 1959). Afterpollination, the flowers were fixed in 50% FAA and clearedin sodium hypochlorate (NaClO) at 60°C for 40 min. Clear-ing in NaOH results in black pistils. The cleared pistilswere then stained with aniline blue and viewed under anepi-fluorescent microscope (Martin 1959). Flowers werefixed 12, 24, 36, 48, 64 and 72 h after pollination, with fourflowers per treatment per time interval.
Flower visitors
Focal observations (sensu Dafni 1992) were conducted todetermine potential pollinators, and all insects visiting theflowers were captured. Daytime observations, from 06.00to 12.00 hours and from 13.00 to 18.00 hours, included theforest edge and interior. Night observations began at19.00 hours with a red-cellophane covered flashlight andwere only carried out at the edge. Approximately 200flowers were observed, totaling 40 h of diurnal observa-tions and 10 h of nocturnal observations. Considering thesmall size of the pollinators, approximately 10 flowersper tree were removed, fixed in FAA, and the pollinatorswere removed under a microscope. Temperature andrelative humidity were checked every hour using athermo-hygrometer (Incoterm, Porto Alegre, Brazil).
Data analysis
Fruit set, calculated as the per treatment fruit frequency,was compared among treatments using an analysis ofindependence and contingency tables. Pollen viabilitywas compared among treatments using a one-way anova,where the factors were trees or anthesis phase or stamenposition. The percentage data for pollen viability wasarcsin transformed before analysis (Zar 1999).
R E P R O D U C T I V E E C O L O G Y O F O C O T E A P O R O S A 33
Plant Species Biology 28, 31–40 © 2012 The AuthorsJournal compilation © 2012 The Society for the Study of Species Biology

Results
Floral morphology and biology
Ocotea porosa flowers were in inflorescences (Fig. 2a),monoclinal and trimeric, with actinomorphic symmetryand six tepals that were greenish yellow with goldenyellow trichomes (Fig. 2c,d). Trichomes were diverse, uniand pluricellular, tectorate and secretory in a variety ofshapes.
Flowers were polystaminate and the stamens hadgolden yellow trichomes in two rings, the internal withthree stamens and the external with six smaller stamens(Fig. 2b,c). Internal anthers were extrorse, whereas exter-nal anthers were introrse (Fig. 2d). Dehiscence was valvar,after which the four valves of each anther opened, expos-ing the pollen stuck to the internal face of the valve.Between the internal stamens were three staminodes thatwere completely covered by long tectate trichomes.
Six nectaries were found at the base of the ovary,attached at the base of the internal stamens and alternatingwith the internal tepals (Fig. 2b,c). The nectaries had a thinsecretion layer. The epidermis of the nectaries was intactwhile the flower was a bud (Fig. 2e). The epidermis laterbreaks with anthesis (Fig. 2f). The style was glabrous andthe apex of the stigma was covered with papillae.
Flowers were asynchronous, that is, they opened atdifferent times during the day, with the number of openflowers peaking soon after 10.00 hours. Each inflores-cence lasted from 15 to 20 days. As a bud, both stamenrings were near the stigma, but once anthesis occurredand the flower exposed the stigma, both stamen ringswere farther from the stigma. Subsequently, the valves onthe internal anthers opened after the stigmas had losttheir receptivity.
Within 2 days of anthesis the stigma was apparently nolonger receptive and had darkened in color. At that time,the internal stamens with opened valves were in juxtapo-sition with the no longer receptive stigmas. After approxi-mately 3 days the stigmas became blackened, the flowerwas entirely opened and the outer ring of anthers haddehisced, at approximately 10.00 hours, when the tem-perature was approximately 24°C. Pollen was availableuntil at least 17.00 hours when observations were termi-nated. The pollen was gone after 4 days, when the rest ofthe flower began to dry. The tepals fell after 13–14 dayswhen the fruits began to develop. On the day anthesisbegan, the stigma and part of the nectaries had a positivereaction (metabolism) to neutral red, but after 2–3 daysonly the nectaries reacted.
The average viability of the pollen during anthesisvaried from 84 to 100% (Table 1). Pollen viability waslower in 3-day-old flowers (mean percentage � 95% con-fidence intervals [CI]; bud, 99.3 � 0.6; day 1, 96.5 � 3.3;day 2, 97.8 � 1.2; day 3, 89.7 � 8.0; F3,209 = 19.45, P < 0.05).
Pollen viability (%) was similar from internal (97.5 � 1.8)and external stamens (96.0 � -3.2; F1,209 = 2.31, P > 0.05).The trees presented different pollen viability, thelowest was 94.4 � 4.2%, and this differed from the twohigher values (97.0 � 3.8% and 98.4 � 1.5%; F2,209 = 6.29,P < 0.05).
Reproductive system
Fruit formed in all treatments except the apomixy treat-ment, with the lowest fruit set in the spontaneous self-pollination treatment (5%). Natural pollination (10%) wasalso low and did not differ statistically from the sponta-neous self-pollination treatment (c2 = 3, d.f. = 1, P > 0.05);all other treatments had greater than 50% fruit forma-tion (Table 2). The fruit set varied between treatments(c2 = 221.9, d.f. = 5, P < 0.05), probably because of the lowfruit set in the natural pollination and spontaneous self-pollination treatments. Otherwise, there was no differencebetween the manual self-pollination (AMI + AME) andcross-pollination (PCI + PCE) treatments (c2 = 2.37, d.f. =1, P > 0.05). Pollen from internal stamens in manual self-pollination was the most efficient (c2 = 7.12, d.f. = 1,P < 0.05), but not in the cross-pollination treatment(c2 = 0.51, d.f. = 1, P > 0.05) (Table 2).
In self-pollination the pollen tube reached the ovary(Fig. 3a) and the seminal rudiment 72 h after pollination(Fig. 3b) and fertilization occurred. In cross-pollination wewere unable to see this process, possibly because the clear-ing and staining technique damaged these samples. Usingepi-fluorescent microscopy we were unable to see pollengermination on the stigma in any treatment.
Flower visitors
The only species observed visiting flowers of Ocotea porosawas Frankliniella gardeniae (Thysanoptera). Thrips wereseen on flowers throughout the day (n = 40 h of observa-tions) and night (n = 10 h) and visited several flowers inthe same inflorescence. However, no flight among treeswas observed owing to the small size of F. gardeniae; indi-viduals were only visible within 30 cm from the observer.The thrips foraged on pollen that had fallen from thestamens and were seen consuming pollen in the foldsbetween the stamens rather than directly on the anthers.Frankliniella gardeniae have many setae covering their bodyand it is likely that the pollen grains adhere to these setae.Large quantities of pollen were observed on setae underthe dissecting microscope. After feeding at one flower,the thrips moved to nearby flowers, usually in the sameinflorescence.
Immature thrips were also observed in flowers. Thethrips collected were deposited at the Entomological
34 A . D A N I E L I - S I LVA A N D I . G . VA R A S S I N
© 2012 The Authors Plant Species Biology 28, 31–40Journal compilation © 2012 The Society for the Study of Species Biology
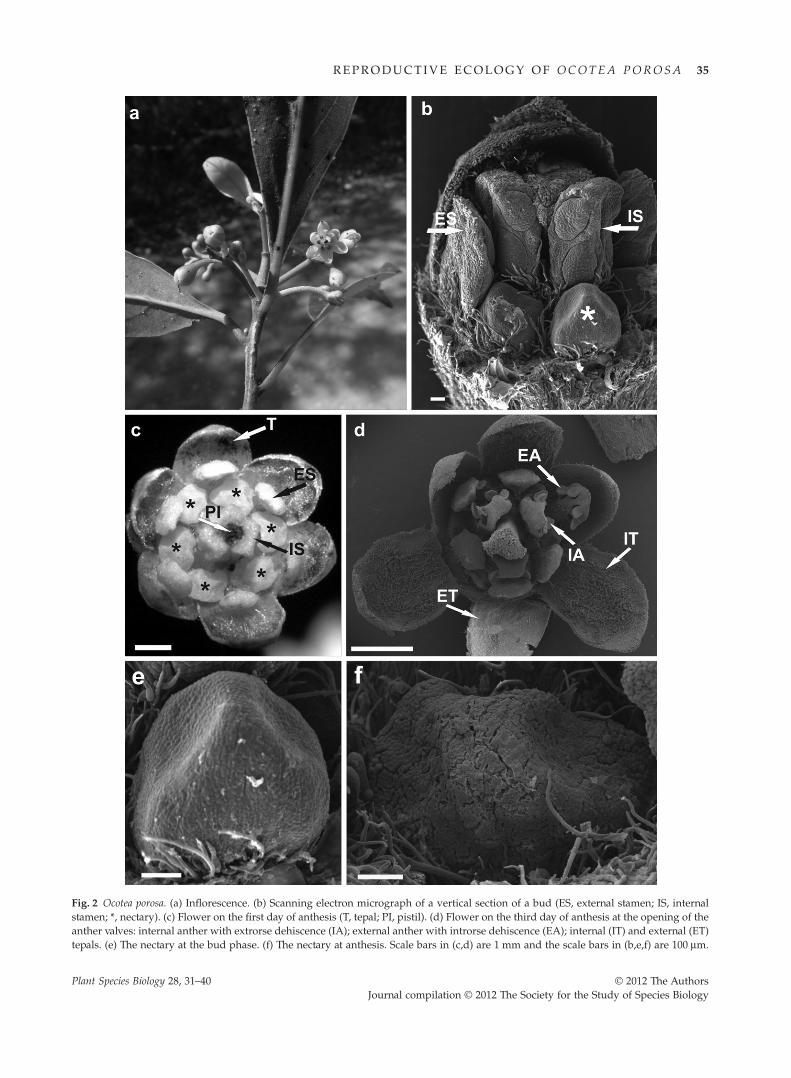
Fig. 2 Ocotea porosa. (a) Inflorescence. (b) Scanning electron micrograph of a vertical section of a bud (ES, external stamen; IS, internalstamen; *, nectary). (c) Flower on the first day of anthesis (T, tepal; PI, pistil). (d) Flower on the third day of anthesis at the opening of theanther valves: internal anther with extrorse dehiscence (IA); external anther with introrse dehiscence (EA); internal (IT) and external (ET)tepals. (e) The nectary at the bud phase. (f) The nectary at anthesis. Scale bars in (c,d) are 1 mm and the scale bars in (b,e,f) are 100 mm.
R E P R O D U C T I V E E C O L O G Y O F O C O T E A P O R O S A 35
Plant Species Biology 28, 31–40 © 2012 The AuthorsJournal compilation © 2012 The Society for the Study of Species Biology

Collection of the Zoology Department of the Univer-sidade Federal do Rio Grande do Sul.
Discussion
Ocotea porosa was self-compatible, but spontaneous self-pollination was very low because of flower herkogamyand dichogamy, with little chance of spontaneous pollenself-transfer. This was supported by the low rates of fruitset in the spontaneous self-pollination treatment, suggest-ing that a pollen vector is important for pollination. Fruitset resulting from natural pollination was double thatfrom spontaneous selfing, but there was no statistical dif-ference between them. In this scenario fruit set in bothtreatments could result from spontaneous selfing or frompollination by thrips, even in bagged flowers.
Despite being monoclinal, flowers of O. porosa wereprotogynic, with the gynoecium only being receptive afterthe first day of anthesis when the anthers were closed.This is also the case in Persea americana and Persea drymi-folia (Lauraceae), although in that case flowers openedtwice; once on the first day during the female phase andagain on the second day during the male phase (Nirody1922; Calvino 1939). Hence, spontaneous self-pollination
in O. porosa was rare and inefficient, with only 5% fruitformation in this experiment. In addition to dichogamy,spontaneous self-pollination was avoided by the extrorseposition of the anthers of the internal stamen, that is,contact with the stigma was avoided. This was true evenon very hot days, when the internal valves of the anthersopened before the stigma had lost its viability. Self-pollination by the external introrse anthers was even moreunlikely because they only opened after the stigma hadlost its viability. Thus, the rarity of self-pollination and theoccurrence of protogyny strongly suggest that thrips mayreach inflorescences even when they are bagged, as seenin Castilla elastica (Moraceae) (Sakai 2001).
Selfing is usually avoided by self-incompatibility, as inCentral American trees (Bawa 1974; Bawa et al. 1985) and inthe Atlantic Forests and Cerrados of Brazil (Barros 1992;Oliveira & Gibbs 1994; Proença & Gibbs 1994; Teixeira &Machado 2000; Goldenberg & Varassin 2001; Moraes &Monteiro 2002; Gusson et al. 2006). Avoiding self-pollination, by means of morphological or genetic mecha-nisms, may have evolved as a result of selective pressuresfor the maintenance of genetic variability in plant popula-tions (Bawa 1974). However, if cross-pollination becomesdifficult or impossible, then self-compatibility will allowself-pollination and continued reproduction (Affre et al.1995; Ortega et al. 1995). Thus, self-compatibility may beassociated with plants in a similar ecological setting toO. porosa, in which isolation and reduced population sizesare common (Stebbins 1957).
Self-compatibility in O. porosa provides anotherexample of reproductive variability in the family Lau-raceae. Other self-compatible species include Ocotea tenera,which is endemic to Costa Rica (Gibson & Wheelwright1996), Cryptocarya moschata (Moraes & Monteiro 2002) andCryptocarya aschersoniana (Moraes & Derbyshire 2002). Inaddition, the family includes obligatory xenogamousspecies (Rohwer 1993), self-incompatible and with syn-chronized dichogamy (Davenport 1986; Kageyama et al.2003; Wang et al. 2005), which effectively prevents self-pollination. However, defining reproductive tendencies
Table 1 Mean pollen viability (%) for eachplant at the different phases of anthesis
Tree
Anthesis phase
BudDay 1
(female phase)
Day 2(male phase, internal
stamens opened)
Day 3(male phase, external
stamens opened)
1-E 98 91 99 841-I 98 91 98 —2-E 100 98 99 972-I 99 99 96 —3-E 100 100 100 863-I 100 98 96 —
E, external stamen; I, internal stamen.
Table 2 Number of flowers tested and the percentage of fruitsformed in the different pollination treatments in Ocotea porosa
Treatment FlowersFruit(%)
Natural pollination (NP) 150 10Apomixy (AP) 150 0Spontaneous self-pollination (SP) 150 5Manual self-pollination, external stamen (MSE) 100 56Manual self-pollination, internal stamen (MSI) 100 74Cross-pollination, external stamen (CPE) 100 60Cross-pollination, internal stamen (CPI) 100 55Manual self-pollination (MS) total (MSI + MSE) 200 65Cross-pollination (CP) total (CPI + CPE) 200 58
36 A . D A N I E L I - S I LVA A N D I . G . VA R A S S I N
© 2012 The Authors Plant Species Biology 28, 31–40Journal compilation © 2012 The Society for the Study of Species Biology

for this genus is complex, in part because of a lack ofinformation about reproduction in the family (Rohwer1993; Werff & van der Richter 1996; Chanderbali et al. 2001).In addition, Ocotea is one of the least defined genera in thefamily and is known to be quite variable, most likelybecause this is the genus into which species that cannot beotherwise identified are placed (Rohwer 1993; Chander-bali et al. 2001).
Fruit set in O. porosa was low in natural conditions com-pared with experimental cross and manual self-pollination. This suggests that plants could spend moreenergy on fruit production than they do in nature. Nectan-dra oppositifolia is in strong contrast, with low fruit set innature (3%) and even lower fruit set with experimentalcross and self pollination (0 and 2.3%, respectively;Crestana & Baitello 1988). In fact, tropical trees tend tohave low fruit set in nature (< 20%; Bawa 1974; Oliveira &Sazima 1990; Oliveira et al. 1992, 2002; Oliveira & Gibbs1994; Proença & Gibbs 1994; Gibbs et al. 1999; Teixeira &Machado 2000; Neto et al. 2007).
Tropical trees are often relatively rare and scattered,which means that pollination may be limited because ofthe distance between reproductive plants and becausepollinators must fly to get from one to another (Fedorov1966). Ocotea porosa has a clumped distribution in naturewith plants on average 9 m away from one another (Bit-tencourt 2007). A clumped distribution may favor cross-pollination by thrips, which have been shown to travel10–25 m (Bawa 1980; Moog et al. 2002). Thus, clumpeddistributions in some species may be associated with pol-lination by thrips. For example, in the self-incompatible,thrips-pollinated Popowia pisocarpa (Annonaceae), fruitformation is greater in plants that are closer together(Momose et al. 1998). Fruit production in the self-
incompatible Linnaea borealis (Caprifoliaceae), which ispollinated by very small flies, declines rapidly with dis-tance between plants (Scobie & Wilcock 2009). Closerplants may have greater fruit formation when small,weakly flying insects are the most important pollinators(Eriksson & Bremer 1993; Momose et al. 1998; Wolf & Har-rison 2001; Aigner 2004; Scobie & Wilcock 2009). Thissame pattern is found in Rubus saxatilis, Rosaceae (Eriks-son & Bremer 1993); Calystegia collina, Convolvulaceae(Wolf & Harrison 2001) and Dithyrea maritima, Brassi-caceae (Aigner 2004). The flights of thrips are usually con-sidered to be short-distance flights (Norton 1984; Cornaraet al. 2005), but it is possible that long-distance flights orwindblown flights occur. These events may cause cross-pollination that may be important in maintaining geneticvariation in the population.
The thrips F. gardeniae was the only pollinator ofO. porosa. After feeding at one flower, individuals movedto nearby flowers, usually in the same inflorescence. As aresult of this activity, flowers at the beginning of anthesiswere visited as well as flowers well past anthesis andoffering pollen, which ensured pollination. Although theywere seen on flowers of O. porosa during the day and thenight, thrips are more active during the day (Kudô 1971),when O. porosa flowers can be pollinated. As F. gardeniacarried pollen and moved among flowers, they are con-sidered to be the only pollinators of O. porosa. This is notnecessarily because of an absence of potential pollinatorsin the study area as many species of insects were observedboth on the edge and in the interior of the forest (A.Danieli-Silva and I. G. Varassin, pers. obs., 2007). In addi-tion to the O. porosa tree in the interior of the forest, aflowering tree of Nectandra megapotamica (Lauraceae),which shares many floral characteristics with O. porosa,
Fig. 3 Epi-fluorescent micrography of pollen tube growth after manual self-pollination 72 h after pollination. (a) Pollen tube reaching theovary (arrow indicates the beginning of the ovary). Scale bar = 50 mm. (b) Pollen tube at the seminal rudiment (arrow). Scale bar = 10 mm.
R E P R O D U C T I V E E C O L O G Y O F O C O T E A P O R O S A 37
Plant Species Biology 28, 31–40 © 2012 The AuthorsJournal compilation © 2012 The Society for the Study of Species Biology

was observed being visited by many species of bees andwasps (A. Danieli-Silva and I. G. Varassin, pers. obs.,2007). It is possible that the beetles and flies observed onflowers of O. porosa in other areas (Bittencourt 2007; Y.Müller, unpubl. data, 2007) occur only occasionally on theflowers and may be considered to be herbivores of veg-etative whorls. Thrips were the only flower visitor inNectandra oppositifolia and they were not considered to actas pollinators, but rather the species was believed tobe spontaneously pollinated (Crestana & Baitello 1988).However, thrips as pollinators should be tested becausethat study did not specifically test the possibility of polli-nation by thrips. Thrips are very common flower visitorsand are considered to be cosmopolitan. Frankliniella iscommonly observed on the flowers of many plant species(Pinent et al. 2003a), particularly Asteraceae (Monteiro2002), where F. rodeos has been observed feeding onpetals, gynoecium, androecium and pollen (Cavalleri et al.2006). Frankliniella is known to breed on flowers ofApiaceae, Asteraceae, Boraginaceae, Fabaceae, Iridaceae,Lamiaceae, Malpighiaceae, Myrtaceae, Polygalaceae,Rubiaceae, Sapindaceae and Verbenaceae (Pinent et al.2003a) and the fact that immature thrips were found in theflowers of O. porosa suggests that the species is a goodreproductive host for F. gardeniae. The fact that pollenfeeding contributes to population increases in thrips (Tsaiet al. 1996; Leskey et al. 1997), and the possible use offlowers as a refuge, may explain the high abundance ofFrankliniella in flowers (Cavalleri 2005). Furthermore,most species in this genus tend to be polyphagous(Mound & Teulon 1995; Pinent et al. 2003b), which is inter-esting considering that their major feeding resource,flowers, is ephemerous (Cavalleri 2005). Despite their highfrequency in flowers they are usually considered general-ist pollinators of low quality because of their weak flyingabilities (Cornara et al. 2005) and because they may onlybe vectors for self-pollination (Norton 1984). This perspec-tive should be re-evaluated. Thrips may be the principalpollinator of many species and even aid cross-pollination(Norton 1984; Momose et al. 1998; Yi-Bo & Zhen-Yu 1999;Sakai 2001; Moog et al. 2002; Zerega et al. 2004). Thripswere effective pollinators of O. porosa and it is clear thatreproduction was through pollination by thrips, ratherthan through spontaneous pollination. Macaranga hullettii(Euphorbiaceae) has also been shown to be a species effi-ciently pollinated by thrips (Moog et al. 2002).
The large number of flowers produced by O. porosaresults in a fruit set that appears large enough to supportlong-term, self-sustaining populations. However, recentexploitation for lumber has had a great impact on popu-lation size and density, causing the species to be includedin the list of endangered species in Brazil (Ministério doMeio Ambiente 2008). Reduction in population densityor even the isolation of some individuals may have
decreased the chances of cross-pollination by thrips,resulting in an increased proportion of autogamy. In thisscenario, high autogamy over a long period of time maychange the population genetic structure, reduce geneticdiversity and may even lead to further population decline(Holsinger 2000). This possibility receives some supportfrom a genetic study of O. porosa in southern Brazil.Young plants have greater allele fixation than adults,which suggests greater homozygosity in the young plants(Bittencourt 2007). Conservation measures must be alertto the possibility of inbreeding depression caused byautogamy, although inbreeding depression was notdetected in our self-pollination experiments.
Inbreeding depression may explain why natural regen-eration of the species is low (Amato 2007). If so, conserva-tion measures must include preservation of the currentpopulation and increases in population density wheredensities are low. One alternative is planting trees that areat least 6 years old, which is when Imbuia trees are able toreproduce, at least under controlled conditions (Carvalho1994). Thus, the distance between individuals would bereduced and genetic variability would be increased.Thrips as pollinators must be included in conservationplanning for O. porosa.
Acknowledgments
We thank Adriano Cavalleri for identifying the thrips.Maurício Sedrez dos Reis, Paulo Eugênio M. de Oliveira,Pedro Luis Rodrigues de Moraes and Renato Goldenbergall provided constructive criticism of the manuscript. Wededicate this paper to the memory of our Caboclo Imbuia,the late José Plínio Baptista. This study was supported bythe Brazilian Agricultural Research Company (EmpresaBrasileira de Pesquisas Agropecuárias—Embrapa/Florestas,project 438/04, action plan number 02.03.02.500/03)and by CAPES through a student fellowship to AlineDanieli-Silva.
References
Affre L., Thompson J. D. & Debussche M. (1995) The reproductivebiology of the Mediterranean endemic Cyclamen balearicumWillk. (Primulaceae). Botanical Journal of the Linnean Society118: 309–330.
Aigner P. A. (2004) Ecological and genetic effects on demographicprocesses: pollination, clonality and seed production inDithyrea maritima. Biological Conservation 116: 27–34.
Amato C. M. (2007) Ecologia de populações de Ocotea porosa emáreas submetidas a diferentes graus de perturbação (Master’sthesis Dissertation). Universidade Federal do Paraná,Curitiba.
Barros M. A. G. (1992) Fenologia da floração, estratégias repro-dutivas e polinização de espécies simpátricas do gênero
38 A . D A N I E L I - S I LVA A N D I . G . VA R A S S I N
© 2012 The Authors Plant Species Biology 28, 31–40Journal compilation © 2012 The Society for the Study of Species Biology

Byrsonima Rich (Malpighiaceae). Revista Brasileira De Biologia52: 343–353.
Bawa K. S. (1974) Breeding systems of tree species of a lowlandtropical community. Evolution 28: 85–92.
Bawa K. S. (1980) Evolution of dioecy in flowering plants. AnnualReview of Ecology and Systematics 11: 15–39.
Bawa K. S., Perry D. R. & Beach J. H. (1985) Reproductive biologyof tropical lowland rain forest trees. I. Sexual systems andincompatibility mechanisms. American Journal of Botany 72:331–345.
Bittencourt R. (2007) Caracterização da estrutura genéticainterna e aspectos da autoecologia de uma população naturalde imbuia (Ocotea porosa—Lauraceae) (Master’s thesis).Universidade Federal de Santa Catarina, Florianópolis.
Calvino E. M. (1939) Floral biology of the Persea drymifolia(Mexican Avocado) cultivated in San Remo, Italy. CaliforniaAvocado Society Yearbook 24: 79–86.
Carvalho P. E. R. (1994) Espécies florestais Brasileiras: recomendaçõessilviculturais, potencialidades e uso da madeira. Embrapa, CNPF,Colombo.
Carvalho P. E. R. (2003) Espécies arbóreas Brasileiras. EmbrapaInformação Tecnológica; Colombo, PR, Embrapa Florestas,Brasília.
Castella P. R. & Britez R. M. (2004) A floresta com Araucária noParaná: conservação e diagnóstico de remanescentes florestais.PROBIO, MMA, Brasília.
Cavalleri A. (2005) Comunidades de Tripes (Insecta: Thysan-optera) em flores e ramos com ênfase em Asteraceae, noParque Estadual de Itapuã, Viamão, RS (Master’s thesis).Universidade Federal do Rio Grande do Sul.
Cavalleri A., Romanowski H. P. & Redaelli L. R. (2006) Thripsspecies (Insecta: Thysanoptera) inhabiting plants of theParque Estadual de Itapuã, Viamão, Rio Grande do Sul State,Brazil. Revista Brasileira De Zoologia 23: 367–364.
Chanderbali A. S., van der Werff H. & Renner S. S. (2001)Phylogeny and historical biogeography of Lauraceae: evi-dence from the chloroplast and nuclear genomes. Annals of theMissouri Botanical Garden 88: 104–134.
Cornara L., Borghesi B., Caporali B. E., Casazza G., Roccotiello E.,Troiano G. & Minuto L. (2005) Floral features and reproduc-tive ecology in Thymelaea hirsuta (L.) Endl. Plant Systematicsand Evolution 250: 157–172.
Crestana C. S. M. & Baitello J. B. (1988) Biologia floral deNectandra mollis (H. B. K.) Nees ssp oppositifolia (Nees) Rohwer(Lauraceae) na Estação Experimental de Moji Guaçu, Estadode São Paulo. Boletim Técnico Do Instituto Florestal 42: 121–137.
Dafni A. (1992) Pollination Ecology: A Practical Approach. OxfordUniversity Press, New York.
Daros T. L. (2006) Sistema reprodutivo e estrutura genética deuma população de Imbuia (Ocotea porosa (NEES & C. MART.)Barroso—Lauraceae) (Master’s thesis). Universidade Federaldo Paraná, Curitiba.
Davenport T. L. (1986) Avocado flowering. Horticultural Reviews8: 257–289.
Eriksson O. & Bremer B. (1993) Genet dynamics of the clonalplant Rubus saxatilis. Journal of Ecology 81: 533–542.
Fedorov A. A. (1966) The structure of the tropical rain forest andspeciation in the humid tropics. Journal of Ecology 54: 1–11.
Fenster C. B. (1991) Gene flow in Chamaecrista fasciculata(Leguminosae). I. Gene dispersal. Evolution 45: 398–409.
Franceschinelli E. V. & Kesseli R. (1999) Population structure andgene flow of the Brazilian shrub Helicteres brevispira. Heredity82: 355–363.
Gibbs P. E., Oliveira P. E. & Bianchi M. B. (1999) Postzygoticcontrol of selfing in Hymenaea stigonocarpa (Leguminosae-Caesalpinioideae), a bat-pollinated tree of the Brazilian cerra-dos. International Journal of Plant Science 160: 72–78.
Gibson J. P. & Wheelwright N. T. (1996) Mating system dynamicsof Ocotea tenera (Lauraceae), a gynodioecious tropical tree.American Journal of Botany 83: 890–894.
Goldenberg R. (2000) Apomixia como alternativa à reproduçãoem Melastomataceae. In: Cavalcanti T. B. & Walter B. M. T.(eds). Tópicos Anuais em Botânica. Embrapa Recursos Genéti-cos e Biotecnologia/Sociedade Botânica do Brasil, Brasília,pp. 225–230.
Goldenberg R. & Varassin I. G. (2001) Sistemas reprodutivos deespécies de Melastomataceae da Serra do Japi, Jundiaí, SãoPaulo, Brasil. Revista Brasileira de Botânica 24: 283–288.
Gusson E., Sebbenn A. M. & Kageyama P. Y. (2006) Sistema dereprodução em populações de Eschweilera ovata (Cambess.)Miers. Revista Árvore 30: 491–502.
Holsinger K. E. (2000) Reproductive systems and evolution invascular plants. Proceedings of the National Academy of Sciencesof the United States of America 97: 7037–7042.
Kageyama P. Y., Cunha G. C., Barreto K. D., Gandara F. B.,Camargo F. R. A. & Sebbenn A. M. (2003) Diversidade eautocorrelação genética espacial em populações de Ocoteaodorifera (Lauraceae). Scientia Forestalis 64: 108–119.
Kudô I. (1971) Observations on relative abundance, phenologyand flower preference of Thysanoptera in Sapporo and thevicinity. Journal of the Faculty of Science Hokkaido University.Ser. VI, Zool. 17: 610–627.
Leskey T. C., Teulon D. A. J. & Cameron E. A. (1997) Effectsof temperature and sugar maple pollen on oviposition andlongevity of pear thrips (Thysanoptera: Thripidae). Environ-mental Entomology 26: 566–571.
Maack R. (1981) Geografia Física do Estado do Paraná, 2 Ed.Secretaria da Cultura e do Esporte do governo do Paraná. J.Olympio, Rio de Janeiro.
Martin F. N. (1959) Staining and observing pollen tubes inthe style by means of fluorescence. Stain Technology 34: 125–128.
Mcknight T. L. & Hess D. (2005) Physical Geography: A LandscapeAppreciation. Prentice Hall, Upper Saddle River.
Ministério do Meio Ambiente (2008) Lista oficial de espécies daflora brasileira ameaçadas de extinção. Instrução Normativan°6 de 23 setembro de.
Mittermeier R. A., Da Fonseca G. A. B., Rylands A. B. & BrandonK. (2005) A brief history of biodiversity conservation in Brazil.Conservation Biology 19: 601–607.
Momose K., Nagamitsu T. & Inoue T. (1998) Thrips cross-pollination of Popowia pisocarpa (Annonaceae) in a lowlanddipterocarp forest in Sarawak. Biotropica 30: 444–448.
Monteiro R. C. (2002) The Thysanoptera fauna of Brasil. In:Marullo R. & Mound L. A. (eds). Thrips and Tospoviruses:Proceedings of the 7th International Symposium on Thysanoptera.Australian National Insect Collection, Canberra, pp. 325–340.
Moog U., Fiala B., Federle W. & Maschwitz U. (2002) Thripspollination of the dioecious ant plant Macaranga hullettii(Euphorbiaceae) in southeast Asia. American Journal of Botany89: 50–29.
R E P R O D U C T I V E E C O L O G Y O F O C O T E A P O R O S A 39
Plant Species Biology 28, 31–40 © 2012 The AuthorsJournal compilation © 2012 The Society for the Study of Species Biology

Moraes P. L. R. & Derbyshire M. T. V. C. (2002) Estrutura genéticade populações naturais de Cryptocarya aschersoniana mez (Lau-raceae) através de marcadores isoenzimáticos. Biota Neotropica2: 183–201.
Moraes P. L. R. & Monteiro R. (2002) Taxas de cruzamento emuma população natural de Cryptocarya moschata nees (Lau-raceae). Biota Neotropica 2: 29–38.
Mound L. A. & Teulon D. A. (1995) Thysanoptera as phytopha-gous opportunist. In: Parker B. L. (ed.). Thrips Biology andManagement. Plenum Press, London, pp. 3–20.
Neto O. C., Lopes A. V. F. & Machado I. C. (2007) Ecologiada Polinização de Inga striata (Benth.) (Leguminosae-Mimosoideae) em um remanescente de Mata Atlântica noNordeste do Brasil. Revista Brasileira Biociências 5: 570–572.
Nirody B. S. (1922) Investigations in avocado breeding. CaliforniaAvocado Association, Annual Report 7: 65–78.
Norton S. A. (1984) Thrips pollination in the lowland forest ofNew Zealand. New Zealand. Journal of Ecology 7: 157–164.
Oliveira A. F., Carvalho D. & Rosado S. C. S. (2002) Taxa decruzamento e sistema reprodutivo de uma população naturalde Copaifera langsdorffii Desf. na região de Lavras (MG) pormeio de isoenzimas. Revista Brasileira de Botânica 25: 331–338.
Oliveira P. E. & Gibbs P. (1994) Pollination biology and breedingsystems of six Vochysia species (Vochysiaceae) in CentralBrazil. Journal of Tropical Ecology 10: 509–522.
Oliveira P. E., Gibbs P. E., Barbosa A. A. & Talavera S. (1992)Contrasting breeding systems in two Eriotheca (Bombacaceae)species of the Brazilian cerrados. Plant Systematics and Evolu-tion 179: 207–219.
Oliveira P. E. & Sazima M. (1990) Pollination biology oftwo species of Kielmeyera (Guttiferae) from Brazilian cerradovegetation. Plant Systematics and Evolution 172: 35–49.
Ortega A. O., Claver J. P. C. & Alcaraz J. A. D. (1995) Floral andreproductive biology of Drosophyllum lusitanicum (L.) link(Droseraceae). Botanical Journal of the Linnean Society 118: 331–351.
Pinent S. M. J., Romanowski H. P., Redaelli L. R. & Cavalleri A.(2003a) Thysanoptera: plantas visitadas e hospedeiras noParque Estadual de Itapuã, Viamão, RS, Brasil. Iheringia, PortoAlegre: Série Zoologia 95: 9–16.
Pinent S. M. J., Romanowski H. P., Redaelli L. R. & Mound L. A.(2003b) Thrips species (Thysanoptera) collected at ParqueEstadual de Itapuã, RS, Brazil. Neotropical Entomology 32: 619–623.
Proença C. E. B. & Gibbs P. E. (1994) Reproductive biology ofeight sympatric Myrtaceae from Central Brazil. New Phytolo-gist 126: 343–354.
Radford A. E., Dickison W. C., Massey J. R., Bell R. C. & SeilerM. S. (1974) Vascular Plant Systematics. Harper & Row,New York.
Reis M. S. (1996) Dinâmica da movimentação dos alelos: subsí-dios para conservação e manejo de populações naturais emplantas. Revista Brasileira de Genética 19: 37–47.
Reitz R., Klein R. M. & Reis A. (1978) Projeto Madeira de SantaCatarina. Sellowia 28–30: 1–320.
Ribas L. A. & Kageyama P. Y. (2004) Diversidade e estruturagenética em populações naturais de Trema micrantha (L.) B.Scientia Forestalis 66: 66–75.
Rohwer J. G. (1993) Lauraceae. In: Kubitzki K., Rohwer J. G. &Bittrich V. (eds). The Families and Genera of Vascular Plants.Flowering Plants. Dicotyledons. Springer-Verlag, Berlin, pp.366–391.
Sakai S. (2001) Thrips pollination of androdioecious Castilla elas-tica (moraceae) in a seasonal tropical forest. American Journalof Botany 88: 1527–1534.
Schaaf L. B., Figueiredo Filho A., Galvão F., Sanquetta C. R. &Longhi S. J. (2006) Modificações florístico-estruturais de umremanescente de Floresta Ombrófila Mista Montana noperíodo entre 1979 e 2000. Ciência Florestal 16: 271–291.
Schoen D. J. & Brown A. H. D. (1991) Intraspecific variation inpopulation gene diversity and effective population size cor-relates with the mating system in plants. Proceedings of theNational Academy of Sciences, USA 88: 4494–4497.
Scobie A. R. & Wilcock C. C. (2009) Limited mate availabilitydecreases reproductive success of fragmented populations ofLinnaea borealis, a rare, clonal self-incompatible plant. Annalsof Botany 103: 835–846.
Stebbins G. L. (1957) Self-fertilization and population variabilityin higher plants. American Naturalist 91: 337–354.
Teixeira L. A. G. & Machado C. I. (2000) Sistema de polinização ereprodução de Byrsonima sericea dc. (Malpighiaceae). ActaBotanica Brasilica 14: 347–357.
Tsai J. H., Yue B. S., Funderburk J. E. & Webb S. E. (1996) Effectof plant pollen growth and reproduction of Frankliniella bispi-nosa. Acta Horticulturae 431: 535–541.
Varty N. & Guadagnin D. L. (1998) Ocotea porosa, em: 2008 IUCNRed List of Threatened Species. [Cited 14 Jan 2009.] Availablefrom URL: http://www.iucnredlist.org
Vogel S. (1963) Duftdrüsen im Dienste der Bestäubung: Über Bauund Funktion der Osmophoren. Abhandlungen der Akademieder Wissenschaften und der Literatur in Mainz Mathematisch-Naturwissenschaftlichen Klasse 10: 601–763.
Wang Z.-S., An S.-Q., Liu H., Leng X., Zheng J.-W. & Liu Y.-H.(2005) Genetic structure of the endangered plant Neolitseasericea (Lauraceae) from the Zhoushan Archipelago usingRAPD markers. Annals of Botany 95: 305–313.
Werff H. & van der Richter H. G. (1996) Toward and improvedclassification of Lauraceae. Annals of the Missouri BotanicalGarden 8: 419–432.
Wolf A. T. & Harrison S. P. (2001) Effects of habitat size and patchisolation on reproductive success of the serpentine morningglory. Conservation Biology 15: 111–121.
Yi-Bo L. & Zhen-Yu L. (1999) Pollination ecology of Chloranthusserratus (Thunb.) Roem. et Schult. And Ch. fortunei (A. Gray)Solms-Laub. (Chloranthaceae). Annals of Botany 83: 489–499.
Zar J. H. (1999) Biostatistical Analysis, 4th edn. Prentice-Hall,Upper Saddle River.
Zeisler M. (1938) Über die Abgrenzung der eigentlichen Narben-flche mit Hilf von Reaktionen. Beihefte zum Botanischen Zen-tralblatt 58: 308–318.
Zerega N. J. C., Mound L. A. & Weiblen G. D. (2004) Pollination inthe New Guinea endemic Antiaropsis decipiens (Moraceae) ismediated by a new species of thrips, Thrips antiaropsidis sp.nov. (Thysanoptera: Thripidae). International Journal of PlantSciences 165: 1017–1026.
40 A . D A N I E L I - S I LVA A N D I . G . VA R A S S I N
© 2012 The Authors Plant Species Biology 28, 31–40Journal compilation © 2012 The Society for the Study of Species Biology
Copyright © 2022 FDOKUMEN
















