Reproductive ecology of Ocotea catharinensis, an ...
10
RESEARCH PAPER Reproductive ecology of Ocotea catharinensis, an endangered tree species T. Montagna 1 , J. Z. Silva 1 , T. G. Pikart 2 & M. S. Reis 1 1N ucleo de Pesquisas em Florestas Tropicais, Universidade Federal de Santa Catarina, Florian opolis, Santa Catarina, Brazil 2 Laborat orio de Entomologia, Centro de Ci^ encias Biol ogicas e da Natureza, Universidade Federal do Acre, Rio Branco, Acre, Brazil Keywords Conservation; floral biology; mating system; phenology; pollination; seed dispersal. Correspondence T. Montagna, Rodovia Admar Gonzaga, 1346. Florian opolis, Santa Catarina, Brazil. E-mail: [email protected] Editor Z.-X. Ren Received: 19 March 2018; Accepted: 16 May 2018 doi:10.1111/plb.12847 ABSTRACT • Ocotea catharinensis (Lauraceae) is an endangered tree species from the Brazilian Atlantic Rainforest. Currently, little is known about the reproductive ecology of this species. Aiming to propose conservation measures, we described aspects related to phenology, floral biology, pollination, seed dispersal and mating system of O. cathari- nensis. • We conducted phenological observations in 62 individuals for 2 years. In one repro- ductive event, we evaluated nectar production, stigmatic receptivity and pollen viabil- ity. Floral visitors were observed, identified and classified on a scale of pollination effectiveness. Seed dispersers were observed and identified using camera traps. Finally, the mating system was evaluated through pollen/ovule ratios, experimental pollination treatments and genetic analysis with molecular markers. • Ocotea catharinensis presented a supra-annual fruiting pattern with a substantial reduction of reproducing individuals from bud phase to ripe fruit phase. Several mechanisms prompting cross-fertilisation were identified, such as attractive, herkoga- mic and protogynic flowers. The main floral visitors and pollinators were from the Diptera order, and all seed dispersers were birds. The species presented a predomi- nantly outcrossed mixed mating system with significant selfing rate (17.3%). • Although based on restricted evidence, we hypothesised that selfing is an escape mech- anism for situations unfavourable to cross-fertilisation. Specifically, for the studied population selfing is a response to reduced population size, which is caused by the non-reproduction of all potentially reproductive individuals and by past exploitation events. Therefore, conservation efforts should be able to enhance population sizes, as well as prevent overexploitation. INTRODUCTION Knowledge of plant reproductive ecology is critical to predict population survival ability and to propose viable conservation efforts in the case of endangered species (Bernardello et al. 2001; Newton 2007). Studies on reproductive ecology are com- prehensive, often reporting on aspects related to phenology, floral biology, pollination and seed dispersal ecology and mat- ing system (e.g. Mariot et al. 2014; Custodio et al. 2017). Such data are fundamental to a complete understanding of repro- ductive processes of a given species, including possible ecologi- cal relationships with other organisms, such as pollinators or seed dispersers. Phenological studies refer to recurrent (cyclic) events and their timing in plant species (Newstrom et al. 1994). Such stud- ies allow us to know the onset, peak, ending, intensities and variations between individuals, populations and years of flow- ering and fruiting periods of a given species (e.g. Grogan & Loveless 2013). Floral biology is concerned with flowering (Percival 1965), and studies in this field can provide informa- tion about nectar production, stigmatic receptivity, pollen via- bility and floral morphology (e.g. Kubitzki & Kurz 1984; Pacini et al. 2014). Pollination and seed dispersal ecology studies can provide valuable information related to gene flow, such as vec- tors, distances and possible limitations (e.g. Dick et al. 2003; Danieli-Silva & Varassin 2013). Mating system studies, in turn, examine how individuals from a given species cross to form the progeny. Consequently, it is possible to estimate, for instance, outcrossing and selfing rates, number of pollen donors and the degree of inbreeding in the progeny (e.g. Conte et al. 2008; Rogalski et al. 2017). All these study fields are deeply connected and provide use- ful knowledge to identify possible factors that can reduce the reproductive ability of a given species. Thus, conservation pro- posals can best be supported when based on reproductive ecol- ogy studies. For instance, one mating system parameter, effective variance size, is commonly used to determine how many seed trees (or seed plants) are necessary for seed collec- tion in order to retain a given effective size (Hmeljevski et al. 2011; Ferreira et al. 2012). Furthermore, pollination vectors can alert researchers to the need for conservation efforts, espe- cially for species with few, or even single, pollinators, such as figs (Janzen 1979) or Ocotea porosa (Danieli-Silva & Varassin 2013). Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 926 Plant Biology ISSN 1435-8603
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Reproductive ecology of Ocotea catharinensis, an ...

RESEARCH PAPER
Reproductive ecology of Ocotea catharinensis, an endangeredtree speciesT. Montagna1 , J. Z. Silva1, T. G. Pikart2 & M. S. Reis1
1 N�ucleo de Pesquisas em Florestas Tropicais, Universidade Federal de Santa Catarina, Florian�opolis, Santa Catarina, Brazil
2 Laborat�orio de Entomologia, Centro de Ciencias Biol�ogicas e da Natureza, Universidade Federal do Acre, Rio Branco, Acre, Brazil
Keywords
Conservation; floral biology; mating system;
phenology; pollination; seed dispersal.
Correspondence
T. Montagna, Rodovia Admar Gonzaga, 1346.
Florian�opolis, Santa Catarina, Brazil.
E-mail: [email protected]
Editor
Z.-X. Ren
Received: 19 March 2018; Accepted: 16 May
2018
doi:10.1111/plb.12847
ABSTRACT
• Ocotea catharinensis (Lauraceae) is an endangered tree species from the BrazilianAtlantic Rainforest. Currently, little is known about the reproductive ecology of thisspecies. Aiming to propose conservation measures, we described aspects related tophenology, floral biology, pollination, seed dispersal and mating system of O. cathari-nensis.
• We conducted phenological observations in 62 individuals for 2 years. In one repro-ductive event, we evaluated nectar production, stigmatic receptivity and pollen viabil-ity. Floral visitors were observed, identified and classified on a scale of pollinationeffectiveness. Seed dispersers were observed and identified using camera traps. Finally,the mating system was evaluated through pollen/ovule ratios, experimental pollinationtreatments and genetic analysis with molecular markers.
• Ocotea catharinensis presented a supra-annual fruiting pattern with a substantialreduction of reproducing individuals from bud phase to ripe fruit phase. Severalmechanisms prompting cross-fertilisation were identified, such as attractive, herkoga-mic and protogynic flowers. The main floral visitors and pollinators were from theDiptera order, and all seed dispersers were birds. The species presented a predomi-nantly outcrossed mixed mating system with significant selfing rate (17.3%).
• Although based on restricted evidence, we hypothesised that selfing is an escape mech-anism for situations unfavourable to cross-fertilisation. Specifically, for the studiedpopulation selfing is a response to reduced population size, which is caused by thenon-reproduction of all potentially reproductive individuals and by past exploitationevents. Therefore, conservation efforts should be able to enhance population sizes, aswell as prevent overexploitation.
INTRODUCTION
Knowledge of plant reproductive ecology is critical to predictpopulation survival ability and to propose viable conservationefforts in the case of endangered species (Bernardello et al.2001; Newton 2007). Studies on reproductive ecology are com-prehensive, often reporting on aspects related to phenology,floral biology, pollination and seed dispersal ecology and mat-ing system (e.g. Mariot et al. 2014; Custodio et al. 2017). Suchdata are fundamental to a complete understanding of repro-ductive processes of a given species, including possible ecologi-cal relationships with other organisms, such as pollinators orseed dispersers.Phenological studies refer to recurrent (cyclic) events and
their timing in plant species (Newstrom et al. 1994). Such stud-ies allow us to know the onset, peak, ending, intensities andvariations between individuals, populations and years of flow-ering and fruiting periods of a given species (e.g. Grogan &Loveless 2013). Floral biology is concerned with flowering(Percival 1965), and studies in this field can provide informa-tion about nectar production, stigmatic receptivity, pollen via-bility and floral morphology (e.g. Kubitzki & Kurz 1984; Pacini
et al. 2014). Pollination and seed dispersal ecology studies canprovide valuable information related to gene flow, such as vec-tors, distances and possible limitations (e.g. Dick et al. 2003;Danieli-Silva & Varassin 2013). Mating system studies, in turn,examine how individuals from a given species cross to form theprogeny. Consequently, it is possible to estimate, for instance,outcrossing and selfing rates, number of pollen donors and thedegree of inbreeding in the progeny (e.g. Conte et al. 2008;Rogalski et al. 2017).
All these study fields are deeply connected and provide use-ful knowledge to identify possible factors that can reduce thereproductive ability of a given species. Thus, conservation pro-posals can best be supported when based on reproductive ecol-ogy studies. For instance, one mating system parameter,effective variance size, is commonly used to determine howmany seed trees (or seed plants) are necessary for seed collec-tion in order to retain a given effective size (Hmeljevski et al.2011; Ferreira et al. 2012). Furthermore, pollination vectorscan alert researchers to the need for conservation efforts, espe-cially for species with few, or even single, pollinators, such asfigs (Janzen 1979) or Ocotea porosa (Danieli-Silva & Varassin2013).
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands926
Plant Biology ISSN 1435-8603

Ocotea catharinensis is a long-lived tree species that occurs inthe Brazilian Atlantic Rainforest. This climax species is dis-tributed between latitudes 19°570 S (Saiter & Thomaz 2014)and 30°150 S (Carvalho 1994). O. catharinensis can reach 30 min height and can surpass 100 cm diameter at breast height(DBH) when adult (Reitz et al. 1978). It was considered as thedominant and most abundant tree in the higher stratum ofOmbrophilous Dense Forest (Veloso & Klein 1959; Klein1980). In spite this dominance, several aspects O. catharinensisreproductive ecology remain poorly understood. For instance,the literature contains little, to no, information regarding nec-tar production, stigma receptivity, pollen viability and floralvisitors. Furthermore, the mating system was only indirectlyestimated (apparent outcrossing rate; Tarazi et al. 2010), andonly one study focused on phenological aspects of this species(Silva et al. 2000).
This species was intensively exploited for timber production(Reitz et al. 1978), and it resulted in severe reductions to popu-lation sizes in Santa Catarina state (Schorn et al. 2012). Becauseof the exploitation processes and the high fragmentation of theAtlantic Rainforest (Ribeiro et al. 2009), O. catharinensis cur-rently figures on the Brazilian List of Endangered Plant Species(MMA 2014) and also on the IUCN Red List (Varty & Guadag-nin 1998), classified as vulnerable. Furthermore, a recent studyhas shown that O. catharinensis presents low recruitment intothe seedling category, potentially harming the populationdynamics in the long term (Montagna et al. 2018).
The endangered condition of O. catharinensis demands stud-ies to better understand aspects of its reproductive ecology. Nev-ertheless, comprehensive knowledge gained from reproductiveecology, although difficult to achieve, is very relevant for the tar-get species of our study. Therefore, in order to propose feasibleconservation actions, we investigated the following aspects ofO. catharinensis: (i) reproductive phenology; (ii) floral biology:nectar production, stigmatic receptivity and pollen viability;(iii) floral visitors and pollinators; (iv) seed dispersers; and(v) mating system. Some of these aspects are described here forthe first time, such as nectar production, stigmatic receptivity,pollen viability, floral visitors and mating system.
MATERIAL AND METHODS
Study area
This study was conducted in a protected area in Santa Catarina,the Floresta Nacional de Ibirama (Ibirama National Forest –FNI; 27°01059.0000 S, 49°27020.0000 W). The FNI comprises570 ha of Ombrophilous Dense Forest in advanced stage of sec-ondary succession (MMA 2008), and the climate is describedas subtropical humid (Cfa), according to K€oppen’s classifica-tion. Studied individuals (DBH > 20 cm) were distributedacross 15.5 ha (350 9 430 m) with a density of 4.1 individu-als�ha�1. Selective logging of O. catharinensis was reported forthe study site in the 1950s (MMA 2008).
Reproductive phenology
Phenological observations were carried out from January 2016to December 2017 (except in February 2016 and 2017 and July2017 due to logistical difficulties). In this period, 62 individu-als, with DBH > 20 cm, were observed monthly for the
flowering (buds and open flowers; Fig. 3) and the fruiting (un-ripe and ripe fruits) using binoculars. We applied the methodproposed by Fournier (1974) to score the intensity of pheno-logical events, attributing 0 for individuals without phenologi-cal activity, and 1, 2, 3 and 4 for individuals with 1–25, 26–50,51–75 and 76–100% of phenological intensity, respectively.Phenological index to each phenophase was obtained by sum-ming all intensity values (6¼0) and averaging by the number ofindividuals presenting phenological activity during a givenmonth, modifying the proposal of Bencke & Morellato (2002).We compared the average DBH between reproductive andnon-reproductive individuals through a confidence interval(CI) (95%) obtained with 1000 bootstrap replicates. Possiblerelationships between phenological events, including monthlyfrequency of individuals presenting buds, open flowers, unripeor ripe fruits and climatological data, including monthly aver-ages of mean temperature, maximum temperature and mini-mum temperature and total monthly rainfall, were investigatedthrough Spearman correlation. Climatological data wereobtained from an official meteorological station 18 km fromthe study area.
Floral biology
Floral biology aspects, such as nectar production, stigmaticreceptivity and pollen viability, were evaluated using threemetallic towers (11–13 m in height) installed near three indi-viduals in order to gain access to flowers. Due to the steeptopography at the study site, we were able to install only threetowers. Standing crop nectar was evaluated every 2 h, from05:00 to 23:00 h, in ten different flowers from three individualsper evaluation using a 1 ll microcapillary tube. In each of theseevaluations, which were conducted over 5 days, the air temper-ature and the presence or absence of floral scent was alsorecorded. Nectar soluble solids concentration (24 flowers fromthree individuals) was measured with a portable refractometeronly when the volume of nectar extracted was >0.5 ll neededto allow refractometer measurement.Stigmatic receptivity was first verified using the Sudan III-
glycerin test (Dafni 1992) on buds (15 stigmas, five of eachindividual) and open flowers (15 stigmas, five of each individ-ual). Later, it was observed that stigmatic receptivity remaineduntil stigma oxidation; therefore, we adopted the criterion ofstigma oxidation to evaluate stigmatic receptivity. To verifywhether the species presents dichogamy (protogyny or protan-dry), 102 flowers were collected from three individuals (43 fullyopened and 59 partially opened) and classified according tostigma oxidation, i.e. not oxidised, partially oxidised or oxi-dised, and anther opening, either open or closed, using a stere-omicroscope (magnification 49). Pollen viability was testedusing the aceto-carmine test (Dafni 1992). Pollen grains werecollected and placed on microscope slides, and then a drop ofaceto-carmine (2%) was added. Using an optical microscope(magnification 129), pollen staining was observed in fourplaces on each microscope slide. This procedure was repeatedfor 15 flowers (five of each individual).
Floral visitors and pollinators
Floral visitors were observed in two of the three individualsfrom the metal towers because of the low flowering intensity of
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 927
Montagna, da Silva, Pickart & dos Reis Reproductive ecology of Ocotea catharinensis

one individual. These observations were made over 9 days inAugust and September 2016, from 07:00 to 00:00 h, using thefollowing methodology. First, only one visitor was observed ata time, even if other visitors were in the tree crown. Second, assoon as the visitor interacted with a flower, it was pho-tographed, and the air temperature was recorded. Third, foreach visitor, the number of visited flowers, time spent in eachflower and whether the visitor touched or not the reproductivestructures (stigma and anthers) were described. Fourth, when-ever possible, visitors were collected with a dip net.The collected visitors were observed and photographed with
a stereomicroscope in order to identify pollen of O. catharinen-sis attached to their bodies. Furthermore, each visitor wasranked on a scale of pollination effectiveness based on whether(i) pollen was attached to the body; (ii) the reproductive struc-tures of flowers had been touched; and (iii) the frequency ofvisits was higher than the median of all frequencies of visits.Therefore, a visitor was classified as “pollinator” if the threeanswers were positive; as “potential pollinator” if two answerswere positive; “eventual pollinator” if only one answer was pos-itive; and “visitor” if all answers were negative.
Seed dispersers
Seed dispersers were observed at tree crown level, aiming torecord the primary dispersers, and at ground level, aiming torecord secondary dispersers. Observations at crown level werecarried out using a camera trap (Bushnell Trophy Cam 8mp�)installed in one of the metallic towers and programmed toshoot 30-s videos with minimum intervals of 2 min. The film-ing started in August 2017, still with unripe fruits in the crown,and was extended until November 2017 when the observedindividual no longer presented ripe fruits. Primary disperserswere also opportunistically observed, with binoculars and pho-tographic camera, along the phenological evaluations. In orderto record secondary dispersers, about 200 ripe fruits were col-lected and deposited grouped in the soil under the crown oftwo individuals of O. catharinensis, about 100 fruits for eachindividual. In each place where fruits were deposited, a cameratrap was installed with programming identical to that previ-ously mentioned. Ground observations began on October 2017and lasted until November 2017.
Mating system
Three methodologies were used to describe the mating systemof O. catharinensis: pollen–ovule ratios (Cruden 1977), experi-mental pollination treatments and genetic analysis of open-pol-linated progeny arrays with molecular markers. Flower budswere collected from three individuals (five buds each), and fiveanthers of each bud were excised and stored in 500 ll lacticacid (85%). Subsequently, the anthers were macerated, andpollen count was performed in a Neubauer chamber, using anoptical microscope (magnification 129). The pollen count wasrepeated four times for each bud. The number of ovules wasdetermined from nine flowers (three of each individual) bylongitudinally cutting the carpels and analysing them under astereomicroscope (magnification 49).Two experimental pollination treatments were implemented:
natural pollination, in which flowers were not manipulated,but marked from the bud phase (1839 buds among three
individuals), and spontaneous self-pollination, in which budswere marked, bagged with voile and after the end of the flower-ing period, bags were removed (1750 buds among three indi-viduals). Buds were marked and bagged in July 2016 andmonitored until September 2017, the time of last ripe fruit. Wewere only able to implement two pollination treatments,because even minimal manipulations caused the oxidation ofbuds. Independence of fruit set and treatments was tested withthe Fisher’s exact test.
Open-pollinated fruits were collected under the crown pro-jection of 12 seed trees in October 2017. Fruit collection wascarried out at ground level, due to the difficulty of climbingseed trees. Fruits were pulped, and then seeds were sown incontainers and maintained under shade net (50% shade) withconstant irrigation. At least 40 seeds from 11 seed trees pluseight seeds from one seed tree were sown, totalling 707 seeds.Open-pollinated progeny arrays (hereinafter progeny: offspringfrom one seed tree) and seed trees were genotyped with allo-zyme markers, suitable for the analysis of mating system(Conte et al. 2008), in maize starch gel (Penetrose 30, 13%).We resolved the following enzymatic systems in a Tris-citratepH 7.5 buffer (Tris 27 g�l�1 and citric acid 16.52 g�l�1): 6-phosphogluconate dehydrogenase (6PGDH, Enzyme Commis-sion 1.1.1.44), a-esterase (aEST, EC 3.1.1.1), diaphorase (DIA,EC 1.8.1.4), malic enzyme (ME, EC 1.1.1.40), phosphogluco-mutase (PGM, EC 5.4.2.2), malate dehydrogenase (MDH, EC1.1.1.37), peroxidase (PO, EC 1.11.1.7) and shikimate dehydro-genase (SKDH, EC 1.1.1.25).
The mating system was analysed under the mixed-matingmodel (Ritland & Jain 1981) and the correlated mating model(Ritland 1989), both implemented in MLTR, version 3.4 (Rit-land 2002). The following parameters were estimated at thepopulation level: multilocus (tm – also estimated at progenylevel) and single locus (ts) outcrossing rates, selfing rate(s ¼ 1� tm), mating among relatives plus selfing rate (1� ts),mating among relatives or biparental inbreeding rate (tm � ts),multilocus paternity correlation (rpðmÞ), and inbreeding coeffi-cient in maternal generation (Fm).
From the aforementioned parameters, we were able to esti-mate other parameters, as follows: effective number of pollendonors (Nep ¼ 1=rpðmÞ) (Ritland 1989) and, according to Seb-benn (2002), the proportions of self-sibs (Pss ¼ s), half sibs(Phs ¼ tmð1� rpðmÞÞ) and full-sibs (Pfs ¼ tm � rpðmÞ). In addi-tion to the inbreeding coefficient in maternal generation, wealso estimated the total inbreeding coefficient in progenies(F0 ¼ F), using the method described by Weir & Cockerham(1984) and implemented in FSTAT (Goudet 2002); inbreedingcoefficient in progenies owing to selfing (Fs ¼ 0:5 � sð1þ FmÞ)(Barrett & Kohn 1991); and inbreeding coefficient in progeniesresulting from mating among relatives (Ftm�ts ¼ F0 � Fs).
Average coancestry within progenies (Hxy) was estimateddividing by two the relatedness correlation within progenies asrxy ¼ 0:250ð1þ FmÞ½4 � sþ ðt2m þ tm � s � rsÞð1þ rpðmÞÞ� (Rit-land 1989), where rs represents the selfing correlation. Basedon the coancestry coefficient, the effective variance size wasestimated as NeðvÞ ¼ 0:5=fHxy½ðn� 1Þ=n� þ ð1þ F0Þ=2ng(Cockerham 1969), in which n is the sample size (assumed tobe 17 – average overall seed trees). Finally, we estimated theminimum number of seed trees as m ¼ NeðrefÞ=NeðvÞ (Sebbenn2002) from which it would be necessary to collect seeds, aimingto retain a reference effective population size (Ne(ref)) of 100. An
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands928
Reproductive ecology of Ocotea catharinensis Montagna, da Silva, Pickart & dos Reis
effective population size of 100 is expected to avoid inbreedingdepression in the short term (five generations; Frankham et al.2014). Confidence intervals (95%) for each parameter wereobtained from 1,000 bootstraps within progenies.
RESULTS
Reproductive phenology
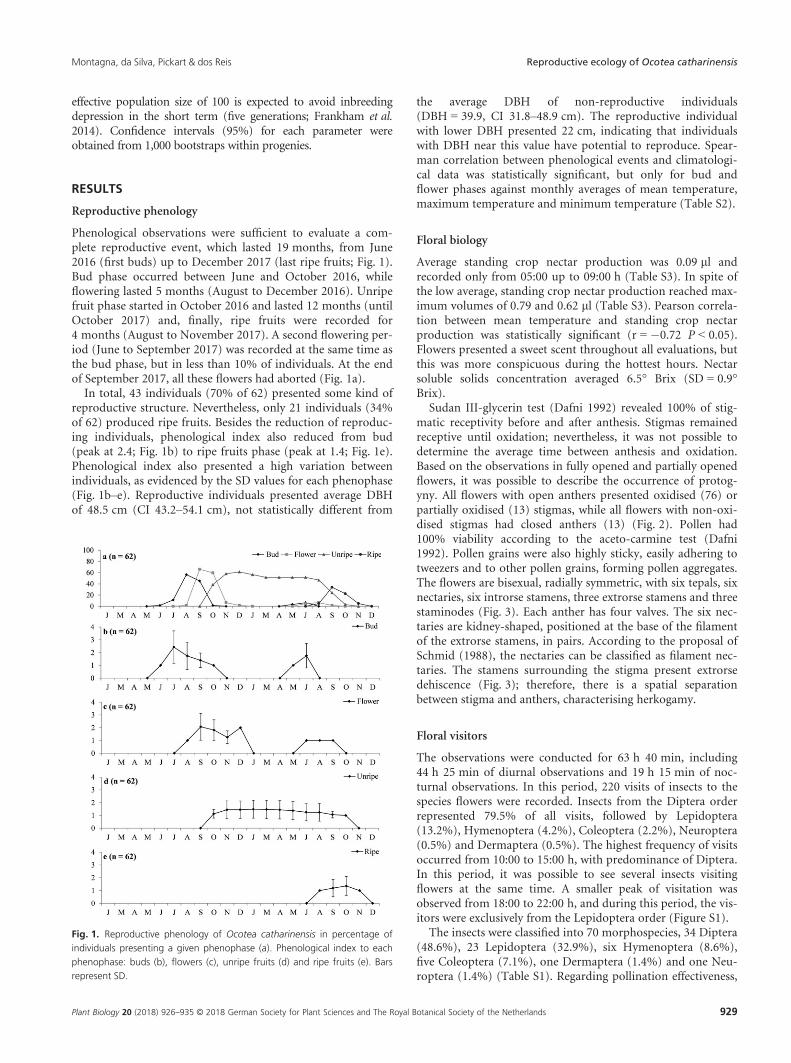
Phenological observations were sufficient to evaluate a com-plete reproductive event, which lasted 19 months, from June2016 (first buds) up to December 2017 (last ripe fruits; Fig. 1).Bud phase occurred between June and October 2016, whileflowering lasted 5 months (August to December 2016). Unripefruit phase started in October 2016 and lasted 12 months (untilOctober 2017) and, finally, ripe fruits were recorded for4 months (August to November 2017). A second flowering per-iod (June to September 2017) was recorded at the same time asthe bud phase, but in less than 10% of individuals. At the endof September 2017, all these flowers had aborted (Fig. 1a).
In total, 43 individuals (70% of 62) presented some kind ofreproductive structure. Nevertheless, only 21 individuals (34%of 62) produced ripe fruits. Besides the reduction of reproduc-ing individuals, phenological index also reduced from bud(peak at 2.4; Fig. 1b) to ripe fruits phase (peak at 1.4; Fig. 1e).Phenological index also presented a high variation betweenindividuals, as evidenced by the SD values for each phenophase(Fig. 1b–e). Reproductive individuals presented average DBHof 48.5 cm (CI 43.2–54.1 cm), not statistically different from
the average DBH of non-reproductive individuals(DBH = 39.9, CI 31.8–48.9 cm). The reproductive individualwith lower DBH presented 22 cm, indicating that individualswith DBH near this value have potential to reproduce. Spear-man correlation between phenological events and climatologi-cal data was statistically significant, but only for bud andflower phases against monthly averages of mean temperature,maximum temperature and minimum temperature (Table S2).
Floral biology
Average standing crop nectar production was 0.09 ll andrecorded only from 05:00 up to 09:00 h (Table S3). In spite ofthe low average, standing crop nectar production reached max-imum volumes of 0.79 and 0.62 ll (Table S3). Pearson correla-tion between mean temperature and standing crop nectarproduction was statistically significant (r = �0.72 P < 0.05).Flowers presented a sweet scent throughout all evaluations, butthis was more conspicuous during the hottest hours. Nectarsoluble solids concentration averaged 6.5° Brix (SD = 0.9°Brix).Sudan III-glycerin test (Dafni 1992) revealed 100% of stig-
matic receptivity before and after anthesis. Stigmas remainedreceptive until oxidation; nevertheless, it was not possible todetermine the average time between anthesis and oxidation.Based on the observations in fully opened and partially openedflowers, it was possible to describe the occurrence of protog-yny. All flowers with open anthers presented oxidised (76) orpartially oxidised (13) stigmas, while all flowers with non-oxi-dised stigmas had closed anthers (13) (Fig. 2). Pollen had100% viability according to the aceto-carmine test (Dafni1992). Pollen grains were also highly sticky, easily adhering totweezers and to other pollen grains, forming pollen aggregates.The flowers are bisexual, radially symmetric, with six tepals, sixnectaries, six introrse stamens, three extrorse stamens and threestaminodes (Fig. 3). Each anther has four valves. The six nec-taries are kidney-shaped, positioned at the base of the filamentof the extrorse stamens, in pairs. According to the proposal ofSchmid (1988), the nectaries can be classified as filament nec-taries. The stamens surrounding the stigma present extrorsedehiscence (Fig. 3); therefore, there is a spatial separationbetween stigma and anthers, characterising herkogamy.
Floral visitors
The observations were conducted for 63 h 40 min, including44 h 25 min of diurnal observations and 19 h 15 min of noc-turnal observations. In this period, 220 visits of insects to thespecies flowers were recorded. Insects from the Diptera orderrepresented 79.5% of all visits, followed by Lepidoptera(13.2%), Hymenoptera (4.2%), Coleoptera (2.2%), Neuroptera(0.5%) and Dermaptera (0.5%). The highest frequency of visitsoccurred from 10:00 to 15:00 h, with predominance of Diptera.In this period, it was possible to see several insects visitingflowers at the same time. A smaller peak of visitation wasobserved from 18:00 to 22:00 h, and during this period, the vis-itors were exclusively from the Lepidoptera order (Figure S1).The insects were classified into 70 morphospecies, 34 Diptera
(48.6%), 23 Lepidoptera (32.9%), six Hymenoptera (8.6%),five Coleoptera (7.1%), one Dermaptera (1.4%) and one Neu-roptera (1.4%) (Table S1). Regarding pollination effectiveness,
Fig. 1. Reproductive phenology of Ocotea catharinensis in percentage of
individuals presenting a given phenophase (a). Phenological index to each
phenophase: buds (b), flowers (c), unripe fruits (d) and ripe fruits (e). Bars
represent SD.
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 929
Montagna, da Silva, Pickart & dos Reis Reproductive ecology of Ocotea catharinensis

three morphospecies were classified as “pollinator”, 25 as “po-tential pollinator”, 35 as “eventual pollinator” and seven as“visitor” (Table S1). All three pollinators belong to the Dipteraorder: Lauxaniidae sp. 05, Sciaridae sp. 01 and Syrphidae sp.01. Another five morphospecies presented pollen attached totheir bodies, but these were not classified as pollinators becauseof the low frequency of visits (Table S1).
Seed dispersers
Five bird species were recorded dispersing O. catharinensis fruits,four of them at the tree crown (primary dispersers) and theother at ground level (secondary disperser). Primary disperserswere Carpornis cucullata (Contigidae), Turdus albicollis,T. flavipes (Turdidae), and Selenidera maculirostris (Ramphasti-dae). The secondary disperser species was Geotrygon montana(Columbidae). All these species were recorded/sighted swallow-ing O. catharinensis seeds. A rat species (Cricetidae family) was
recorded interacting with the fruits at the ground; however, thisspecies did not swallow, pulp or transport any fruit.
Mating system
The results indicate that O. catharinensis presents a predomi-nantly outcrossed mixed mating system. The species had, on aver-age, 1808 pollen grains (SD = 1693) and one ovule per flower.Thus, the pollen/ovule ratio was 1808, closer to that obtained byCruden (1977) for facultative xenogamy (796.6 � 87.7) than tothe ratio for xenogamy (5859.2 � 963.5). The 1750 buds submit-ted to spontaneous self-pollination yielded five unripe fruits butno ripe fruits. Natural pollinated buds (1839) formed 38 unripefruits and one ripe fruit. Fruit set was not significantly associatedwith treatments according to Fisher’s exact test (P = 0.88),although a trend of higher fruit production was observed in indi-viduals submitted to natural pollination treatment.
Mating system estimates obtained from progeny analysis aregiven in Tables 1 and 2. The multilocus outcrossing rate esti-mate statistically differed from unity (tm = 0.827), thus deter-mining a mixed-mating system and a significant selfing rate(s = 0.173). Mating among relatives rate (tm � ts = 0.086) wasnot statistically significant, unlike the multilocus paternity cor-relation (rpðmÞ = 0.247). Based on rpðmÞ, the average effectivenumber of pollen donors for each progeny array was estimatedto be four. Multilocus outcrossing rate estimates presentedhigh variation among progenies, ranging from 0.452 up to1.091 (�1).
On average, progenies were composed of 17.3% self-sibs,62.3% half-sibs and 20.4% full-sibs. Inbreeding coefficient inmaternal generation was not significant (Fm= 0.017), unlike theinbreeding in progenies (F0 = 0.146). The partition of F0resulted in a significant inbreeding in progenies from selfing(FS = 0.088) and a non-significant inbreeding in progeniesfrom mating among relatives (Ftm�ts = 0.058). Coancestrywithin progenies (Hxy = 0.203) was higher than expected incompletely panmictic progenies (half-sibs, Hxy = 0.125). Inorder to retain effective size of 100 individuals, seeds must becollected from 45 seed trees.
DISCUSSION
Reproductive phenology
Ocotea catharinensis presented a supra-annual fruiting pattern,a characteristic already described for this (Silva et al. 2000) andother Lauraceae species (Wheelwright 1986). The reduction inreproducing individuals from bud phase to ripe fruits phaseand the high variation in flowering and fruiting intensitiesbetween individuals were also described for Lauraceae species,including O. catharinensis (Wheelwright 1986; Silva et al.2000). The occurrence of these features, i.e. reduction of repro-ducing individuals and high variation in phenological indices,can represent a serious restriction to population effective size.
The start of flowering was linked to a drop in the tempera-tures, mean, maximum and minimum, as evidenced by Spear-man correlations. Similar correlations were found forO. porosa, which presented bud phase negatively correlatedwith mean temperature (r = �0.51; Bittencourt 2007). Never-theless, the confirmation of this pattern requires longer pheno-logical observations since flowering can start in different
Table 1. Mating system estimates for a population of Ocotea catharinensis.
parameter
estimates (confidence
interval)
Number of seed-trees [total number of
offsprings]
11 [197]
Multilocus outcrossing rate (tm) 0.827 (0.696/0.953)
Single-locus outcrossing rate (ts) 0.741 (0.574/0.966)
Selfing rate (s) 0.173 (0.047/0.304)
Mating among relatives and selfing (1� ts) 0.259 (0.034/0.426)
Mating among relatives (tm � ts) 0.086 (�0.029/0.150)
Multilocus paternity correlation (rpðmÞ) 0.247 (0.051/0.432)
Effective number of pollen donors (Nep) 4
Self-sibs proportion (Pss) 0.173 (0.047/0.304)
Half-sibs proportion (Phs) 0.623 (0.429/0.865)
Full-sibs proportion (Pfs) 0.204 (0.044/0.333)
Inbreeding in maternal generation (Fm) 0.017 (�0.200/0.170)
Inbreeding in progeny (F0) 0.146 (0.078/0.237)
Inbreeding in progeny from selfing (Fs) 0.088 (0.021/0.154)
Inbreeding in progeny from mating among
relatives (Ftm�ts)
0.058 (�0.008/0.125)
Coancestry within progeny (Hxy) 0.203 (0.136/0.249)
Effective variance size (NeðvÞ) 2.220 (1.864/3.096)
Number of seed-trees for seed collection
(m ¼ 100=NeðvÞ)45
Table 2. Multilocus outcrossing rate (tm) and its respective standard devia-
tion (SE) for each evaluated seed-tree of Ocotea catharinensis.
seed-tree [progeny size] tm SE (tm)
c19 [16] 1.091 0.001
c64 [16] 1.038 0.065
c18 [24] 1.028 0.031
cp3 [20] 0.998 0.032
c57 [17] 0.887 0.074
c62 [24] 0.883 0.079
c58 [24] 0.861 0.093
c46 [10] 0.820 0.105
r15 [15] 0.649 0.110
c23 [15] 0.624 0.151
c66 [10] 0.452 0.414
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands930
Reproductive ecology of Ocotea catharinensis Montagna, da Silva, Pickart & dos Reis

seasons for O. catharinensis, including spring and summer, aswell as winter (Silva et al. 2000).
Floral biology
The floral characteristics described for O. catharinensis demon-strated that the species uses several mechanisms to promotecross-fertilisation. The flowers presented typical traits of gener-alised pollination, such as radially symmetrical flowers andexposed reproductive organs (Fenster et al. 2004). Furthermore,flowers produced nectar and scent, which are attractive to insects(Fægri & van der Pijl 1979). Flowers also presented barriers toself-fertilisation, such as protogyny, already described for other
Lauraceae species (Kubitzki & Kurz 1984; Danieli-Silva & Varas-sin 2013), and herkogamy. Nevertheless, none of these traits wasefficient in completely preventing self-fertilisation, as evidencedby the significant estimated selfing rate.Standing crop nectar production was detected only from
05:00 up to 09:00 h. After this period, increases in mean tem-perature and frequency of floral visitors, especially dipterans,were observed. If nectar is produced after 09:00 h, these obser-vations suggest that it is evaporated due to the increase in tem-perature or is collected by visitors. It should be emphasisedthat the volume of nectar produced by O. catharinensis flowerscan be considered high in comparison to other Lauraceae spe-cies. For instance, the amount of nectar produced by Laurusazorica was too small to be extractable with 1 ll micropipettes(Forfang & Olesen 1998). A similar situation was described forL. nobilis (Pacini et al. 2014). The production of nectar repre-sents a large investment of energy (Pyke 1991; Pacini & Nepi2007); therefore, the secretion of nectar in O. catharinensis canbe linked to a dependence on biotic vectors to perform pollina-tion. Regarding pollen presentation, although not evaluated,stickiness may result from the presence of pollenkit, an adhe-sive substance commonly present in pollen grains of Lauraceaespecies (Hesse & Kubitzki 1982; Pacini et al. 2014).According to Kubitzki & Kurz (1984), wide open flowers
with synchronous dichogamy and nectar production wouldrepresent a basic state in Lauraceae. In synchronised dicho-gamy, self-fertilisation is not possible because all flowers of agiven individual are simultaneously in the same sexual phase,with receptive stigma and closed anthers or with wilted stigmaand open anthers – protogyny; nevertheless, the occurrence ofreceptive stigmas and open anthers is synchronised betweenindividuals, allowing cross-fertilisation to be mediated by polli-nators (Kubitzki & Kurz 1984). From these hermaphroditicnectar-producing ancestors, two lines of specialisation haveevolved, according to the same authors: (i) species with lowpollen/ovule ratios, hermaphroditic flowers, synchronousdichogamy, but no nectar production, and (ii) dioecious spe-cies, with high pollen/ovule ratios and nectar production.Although we were not able to describe the occurrence of
Fig. 2. Schematic illustration representing sampled flowers (open and par-
tially open) and the condition of their anthers (open or closed) and stigmas
(oxidized, partially oxidised or not oxidised).
Fig. 3. Ocotea catharinensis. (a) Partially open flower
(ST, oxidized stigma; #, extrorse stamens). (b) Open
flower (TE, tepals; *, nectaries; +, introrse stamens). (c)
Flowering sequence of O. catharinensis: 1, 2, and 3,
buds; 4, partially open flower already with oxidised
stigma; 5 and 6, open flowers; 7, open flower with non-
functional stamens, stigma, and nectaries.
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 931
Montagna, da Silva, Pickart & dos Reis Reproductive ecology of Ocotea catharinensis

synchronised dichogamy, O. catharinensis seems to better fitinto the ancestral group. However, even if the species exhibitssynchronised dichogamy, this mechanism was not completelyefficient in preventing self-fertilisation.
Floral visitors
Flowers of Lauraceae species are visited by insects belonging toseveral orders, such as Hymenoptera (Kubitzki & Kurz 1984),Coleoptera (Forfang & Olesen 1998; Dupont 2002), Lepidoptera(Pacini et al. 2014), Diptera (N�obrega et al. 2016), Hemiptera(Souza & Moscheta 1999) and Thysanoptera (Danieli-Silva &Varassin 2013). Nevertheless, hymenopterans seem to be the morefrequent and diverse floral visitors to Lauraceae species. The pre-dominance of Diptera as major floral visitors, herein describedfor O. catharinensis, was also recorded to O. duckeii (N�obregaet al. 2016). Pollinator type is often related to nectar sugar com-position, either sucrose or hexoses (Baker & Baker 1983; Pacini &Nepi 2007), and Diptera insects are linked to sucrose-poor/hex-ose-rich nectar (Baker & Baker 1983; Wolff 2006).The pollination effectivity of dipterans is limited by their
body size, i.e. small insects carry small pollen loads, but theirnumber can compensate for this (Fægri & van der Pijl 1979).Moreover, Diptera is considered the second most importantorder of flower-visiting and -pollinating insects (Larson et al.2001). According to visiting behaviour and frequency and thepresence or absence of pollen attached to the body, threedipterans were classified as pollinators of O. catharinensis:Lauxaniidae sp. 05, Sciaridae sp. 01 and Syrphidae sp. 01. Addi-tionally, from the 25 potential pollinators, 19 belong to theDiptera order. Insects belonging to Lauxaniidae, Sciaridae andSyrphidae families are recognised as pollinators of severalangiosperms (see Herrera 1987; Ollerton & Liede 1997; Larsonet al. 2001). Also, insects of these families can carry pollen overlong distances, such as 200 m (Wratten et al. 2003) or even400 m (Rader et al. 2011). Nevertheless, it should be stressedthat the mentioned studies were conducted in open-field envi-ronments, which present fewer barriers to insect flights com-pared to forests. Furthermore, flight distance distributions arestrongly leptokurtic, i.e. shorter flights are much more com-mon than longer flights (Herrera 1987; Wratten et al. 2003).Lepidoptera was the second most frequent and diverse order
of flower-visiting insects of O. catharinensis. Nevertheless, lepi-dopterans do not seem to be efficient pollinators for the follow-ing reasons. None of captured insects presented pollenattached to their body, and only three insects visited the flowersmore than once. Furthermore, lepidopterans often acted asnectar thieves, inserting their long proboscides when clingingto the flower pedicel or tepals without touching anthers orstigma. None of collected hymenopterans presented pollenattached to their body, and five of the six identified morphos-pecies visited flowers only once. Therefore, in spite of theirrecognised importance as pollinators (Fægri & van der Pijl1979), hymenopterans cannot be considered effective pollina-tors of O. catharinensis. Finally, among the other sampledinsects, it is worth mentioning Chrysopidae spp. 01. This mor-phospecies was classified as a potential pollinator because itpresented pollen attached to its body. Insects belonging to theChrysoperla genus are described as pollenophagous when adults(Li et al. 2008). Thus, pollination could occur by chance whilethese insects feed on O. catharinensis pollen.
Seed dispersers
All the birds identified as seed dispersers of O. catharinensiswere also recorded feeding on fruits of several other forest spe-cies. Fruits from 35 and 11 species were consumed by T. albi-collis and T. flavipes, respectively (Castro et al. 2012). Carporniscucullata was recorded feeding on fruits from 45 species (Fran-cisco & Galetti 2002), while S. maculirostris was recorded feed-ing on fruits from at least 40 species (Galetti et al. 2000).Geotrygon montana feeds on fruits from Euterpe edulis (Are-caceae; Galetti et al. 2015) and Margaritaria nobilis (Euphor-biaceae; Cazetta et al. 2008).
At the study site, seedlings from O. catharinensis present anaggregated spatial pattern; however, this pattern is independent ofthe spatial distribution of reproductive individuals (Montagnaet al. 2018). Furthermore, a large quantity of deteriorated seedswere found rotten and infested by insect larvae under the crownof the few individuals that had high seed production. These factshighlight the importance of seed dispersers in transporting seedsto appropriate sites for germination and seedling establishment. Itis worth noting that neither of the species cited in the literature asO. catharinensis seed dispersers (Brachyteles arachnoides and Pipilejacutinga) were recorded in the present study.
Mating system
Mating system estimates for O. catharinensis are consistentwith the estimates already obtained for other Lauraceae species.Ocotea porosa presented fruit set of 5% under spontaneousself-pollination treatments (Danieli-Silva & Varassin 2013).Based on molecular markers, O. tenera was classified as pre-dominantly outcrossed (Gibson & Wheelwright 1996), as wasCryptocarya moschata (de Moraes & Monteiro 2002). Highrates of outcrossing are also common to several tropical forestspecies (Ward et al. 2005).
A large portion of the progeny arrays was composed offull-sibs (Pfs = 20.4%) and self-sibs (Pss = 17.3%). These pro-portions resulted from the significant paternity correlation(full-sibs – same father and same mother) and also from thesignificant selfing rate (self-sibs). Self-fertilisation acts byincreasing the frequency of homozygotes, essentially reducinggenetic variability (Wright 1931). Thus, in the long term, self-fertilisation can be detrimental to the adaptive potential of thespecies through the loss of variability (Frankham 2005). On theother hand, the described self-compatibility can be understoodas an escape for situations in which the production of seedsand progenies is maintained, even if it is not possible to cross.This possibility is discussed by other authors for other predom-inantly outcrossed species (Nazareno & dos Reis 2012;Danieli-Silva & Varassin 2013).
Two main lines of evidence support the hypothesis that self-fertilisation is an escape mechanism for situations unfavourablefor cross-fertilisation. The first is related to the population size.Ocotea catharinensis populations from Santa Catarina wentthrough intensive timber exploitation, especially in the last cen-tury (Reitz et al. 1978). This exploitation process caused severereductions in the size of O. catharinensis populations. Accordingto the last estimates for Santa Catarina, the species occurs at anaverage density of 5.86 individuals�ha�1 (DBH > 10 cm; Lingneret al. 2013). However, in the past, higher densities were recorded,such as 23.9 individuals�ha�1 (DBH > 12.7 cm; Veloso & Klein
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands932
Reproductive ecology of Ocotea catharinensis Montagna, da Silva, Pickart & dos Reis

1959). Thus, pollen availability can be affected by the fact thatthe populations nowadays are smaller than in the past. It is worthnoting that O. catharinensis occurs with a density of 4.1 individu-als�ha�1 in the study site. Furthermore, not all potentially repro-ductive individuals are actually reproducing, reducing even morethe population size. Significantly lower outcrossing rates wereestimated for Symphonia globulifera (Clusiaceae) in disturbedsites with lower densities of reproductive individuals (Aldrich &Hamrick 1998), as well as for Carapa procera (Meliaceae) inlogged plots when compared to populations from undisturbedplots (Doligez & Joly 1997).
Second, the multilocus outcrossing rate was highly variableamong evaluated seed trees. This result demonstrates that self-ing rates are not uniform in seed trees. Therefore, in additionto effective size effects, other factors, such as crown position inthe forest stratum, distance to other reproducing individualsand position in the landscape, edge or core, may influence theestimated outcrossing rates. Edge effect, for instance, enhancedselfing rates in individuals of Copaifera langsdorffi (Fabaceae;Tarazi et al. 2013).
Finally, we highlight the high number of flowers producedwhen compared to the number of ripe fruits. Although abor-tion of flowers and fruits is a common phenomenon in plants(Bawa & Webb 1984), it is really surprising that only one ripefruit arose from 1839 buds under natural pollination treat-ment. As several factors can explain abortion of flowers andfruits, such as pollen and resources limitation (Bawa & Webb1984), only further research can clarify this question inO. catharinensis. However, a better understanding regardingflower and fruit abortion could indicate limiting factors to fruitproduction, valuable knowledge for the conservation of anendangered species with low recruitment into the seedling cate-gory (Montagna et al. 2018).
Implications for conservation
Although the species presented several mechanisms to promotecross-fertilisation, a high portion of the progenies was com-posed of full-sibs and self-sibs as a result of correlated matingsand selfing. In the long term, this condition can harm theadaptive potential of the species through the loss of geneticvariability. Furthermore, selfing and correlated matings can be,for the studied population, a direct result of the low density ofreproductive individuals, which is caused by past logging activ-ities and by the fact that not all potentially reproductive indi-viduals are actually reproducing. Consequently, efforts toenhance population sizes are desirable, as well as to continuerestricting exploitation of O. catharinensis. In this sense, theestablishment of local seed collection areas plus the productionand planting of seedlings in natural remnants of OmbrophilousDense Forest should be promoted.
Sticky pollen, when combined with protogynic, herkogamicand nectar-producing flowers, is a clear signal indicating thatO. catharinensis pollination largely depends on insects. Therefore,any effort to create a new protected area or establish new popula-tions, as well as to conserve the existent populations, should takeinto account the presence of pollinating insects. As mentioned, a
large quantity of deteriorated seeds, i.e. rotten and infested byinsect larvae, was observed under the crown of the few individu-als that presented high seed production. Therefore, the conserva-tion of birds, the main seed dispersers of O. catharinensis, is alsoimportant to allow seedling establishment. Regarding ex situ con-servation, the collection of seed from 45 seed trees is suggested inorder to retain an effective size of 100 individuals. The estimateof effective variance size can also be used in combination withother reference effective sizes, if necessary.This report represents the first attempt to understand several
aspects of the reproductive ecology of O. catharinensis. Thus,we firmly believe that this kind of study should be carried outin other populations and continuously in order to better pre-dict reproductive bottlenecks and, consequently, to proposebetter and more effective conservation efforts. Nevertheless,our study allowed inferences regarding in situ and ex situ con-servation measures for O. catharinensis.
ACKNOWLEDGEMENTS
We thank the Instituto Chico Mendes de Conservac�~ao da Biodi-versidade (ICMBio) and the researchers of N�ucleo de Pesquisasem Florestas Tropicais for support with the fieldwork. We alsothank the Laborat�orio de Fisiologia do Desenvolvimento e Gen-�etica Vegetal for providing the necessary infrastructure requiredfor the genetic analysis performed in this work. This study wasfunded by the Fundac�~ao de Amparo �a Pesquisa e Inovac�~ao doEstado de Santa Catarina (FAPESC; grant numbers 11939/2009,18868/2011-9 and TR2013-3558), the Coordenac�~ao de Aper-feic�oamento de Pessoal de N�ıvel Superior (CAPES) to TM andJZS, and the Conselho Nacional de Desenvolvimento Cient�ıficoe Tecnol�ogico (CNPq) to MSR (304724/2010-6). We also thankDavid Martin for editing the manuscript.
SUPPORTING INFORMATION
Additional Supporting Information may be found online in thesupporting information tab for this article:Figure S1. Number of insects visiting the flowers of Ocotea
catharinensis per insect order and daytime hour.Table S1. Floral visitors of Ocotea catharinensis. N, number
of visits; Fr, relative frequency (%); T/V, average time spent pervisit (min:seg); F/V, average number of visited flowers per visit;P, pollen attached to the insect body; TF, insect touched repro-ductive structures of flowers; F>M, frequency of visits washigher than the median of all frequencies of visits; Class., classi-fication; PO, pollinator; PP, potential pollinator; PE, eventualpollinator; V, visitor.Table S2. Spearman correlations between phenological
events of Ocotea catharinensis (monthly frequency of indivi-duals presenting buds, open flowers, unripe or ripe fruits) andclimatological data (monthly averages of mean temperature,maximum temperature, and minimum temperature and totalmonthly rainfall). *P < 0.1; **P < 0.05.Table S3. Standing crop nectar production (lL) of Ocotea
catharinensis per day and daytime hour and mean temperature(�C) per daytime hour.
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 933
Montagna, da Silva, Pickart & dos Reis Reproductive ecology of Ocotea catharinensis

REFERENCES
Aldrich P.R., Hamrick J.L. (1998) Reproductive domi-
nance of pasture trees in a fragmented tropical forest
mosaic. Science, 281, 103–105.
Baker H.G., Baker I. (1983) Floral nectar sugar con-
stituents in relation to pollinator type. In: Jones C.
E., Little R. J. (Eds), The handbook of experimental
pollination biology. Scientific & Academic Editions,
New York, USA, pp 117–141.
Barrett S.C.H., Kohn J.R. (1991) Genetic and evolu-
tionary consequences of small population size in
plants: implications for conservation. In: Falk D. A.
I., Holsinger K. E. (Eds), Genetics and conservation of
rare plants. Oxford University Press, New York,
USA, pp 3–30.
Bawa K.S., Webb C.J. (1984) Flower, fruit and seed
abortion in tropical forest trees: implications for the
evolution of paternal and maternal reproductive
patterns. American Journal of Botany, 71, 736–
751.
Bencke C.S.C., Morellato L.P.C. (2002) Comparac�~aode dois m�etodos de avaliac�~ao da fenologia de plan-
tas, sua interpretac�~ao e representac�~ao. Revista Brasi-
leira de Botanica, 25, 269–275.
Bernardello G., Anderson G.J., Stuessy T.F., Crawford
D.J. (2001) A survey of floral traits, breeding
systems, floral visitors, and pollination systems of
the angiosperms of the Juan Fern�andez islands
(Chile). Botanical Review, 67, 255–308.
Bittencourt R. (2007) Caracterizac�~ao da estrutura gen-
�etica interna e aspectos da autoecologia de uma pop-
ulac�~ao natural de imbuia (Ocotea porosa -
Lauraceae). M.Sc. Thesis. Universidade Federal de
Santa Catarina, Florian�opolis, Santa Catarina, Brazil.
Carvalho P.E.R. (1994) Esp�ecies florestais brasileiras:
recomendac�~oes silviculturais, potencialidades e uso da
madeira. EMBRAPA/CNPF, Colombo, Brazil.
Castro E.R., Correa Cortes M., Navarro L., Galetti M.,
Morellato L.P.C. (2012) Temporal variation in the
abundance of two species of thrushes in relation to
fruiting phenology in the Atlantic rainforest. Austral
Ornitolhology, 112, 137–148.
Cazetta E., Zumstein L.S., Melo-J�unior T.A., Galetti M.
(2008) Frugivory on Margaritaria nobilis L.f.
(Euphorbiaceae): poor investment and mimetism.
Revista Brasileira de Botanica, 31, 303–308.
Cockerham C.C. (1969) Variance of gene frequencies.
Evolution, 23, 72–84.
Conte R., Reis M.S., Mantovani A., Vencovsky R.
(2008) Genetic structure and mating system of
Euterpe edulis Mart. populations: a comparative
analysis using microsatellite and allozyme markers.
Journal of Heredity, 99, 476–482.
Cruden R.W. (1977) Pollen-ovule ratios: a conservative
indicator of breeding systems in flowering plants.
Evolution, 31, 32–46.
Custodio T., Comtois P., Araujo A.C. (2017) Repro-
ductive biology and pollination ecology of Triplaris
gardneriana (Polygonaceae): a case of ambophily in
the Brazilian Chaco. Plant Biology, 19, 504–514.
Dafni A. (1992) Pollination ecology: a practical
approach. Oxford University Press, Oxford, UK.
Danieli-Silva A., Varassin I.G. (2013) Breeding system
and thrips (Thysanoptera) pollination in the endan-
gered tree Ocotea porosa (Lauraceae): implications
for conservation. Plant Species Biology, 28, 31–40.
Dick C.W., Etchelecu G., Austerlitz F. (2003) Pollen
dispersal of tropical trees (Dinizia excelsa: Fabaceae)
by native insects and African honeybees in pristine
and fragmented Amazonian rainforest. Molecular
Ecology, 12, 753–764.
Doligez A., Joly H.I. (1997) Mating system of Carapa
procera (Meliaceae) in the French Guiana Tropical
forest. American Journal of Botany, 84, 461–470.
Dupont Y.L. (2002) Evolution of apomixis as a strategy
of colonization in the dioecious species Lindera
glauca (Lauraceae). Population Ecology, 44, 293–297.
Fægri K., van der Pijl L. (1979) The principles of pollina-
tion ecology, 3rd edn. Pergamon Press, Oxford, UK.
Fenster C.B., Armbruster W.S., Wilson P., Dudash
M.R., Thomson J.D. (2004) Pollination syndromes
and floral specialization. Annual Review of Ecology,
Evolution, and Systematics, 35, 375–403.
Ferreira D.K., Nazareno A.G., Mantovani A., Bitten-
court R., Sebbenn A.M., Reis M.S. (2012) Genetic
analysis of 50-year old Brazilian pine (Araucaria
angustifolia) plantations: implications for conserva-
tion planning. Conservation Genetics, 13, 435–
442.
Forfang A.S., Olesen J.M. (1998) Male-biased sex ratio
and promiscuous pollination in the dioecious island
tree Laurus azorica (Lauraceae). Plant Systematics
and Evolution, 212, 143–157.
Fournier L. (1974) Un m�etodo cuantitativo para la
medici�on de caracter�ısticas fenol�ogicas en �arboles.
Turrialba, 24, 422–423.
Francisco M.R., Galetti M. (2002) Aves como potenci-
ais dispersoras de sementes de Ocotea pulchella Mart.
(Lauraceae) numa �area de vegetac�~ao de cerrado do
sudeste brasileiro. Revista Brasileira de Botanica, 25,
11–17.
Frankham R. (2005) Genetics and extinction. Biological
Conservation, 126, 131–140.
Frankham R., Bradshaw C.J.A., Brook B.W. (2014)
Genetics in conservation management: revised rec-
ommendations for the 50/500 rules, Red List criteria
and population viability analyses. Biological Conser-
vation, 170, 56–63.
Galetti M., Laps R.R., Pizo M.A. (2000) Frugivory by
toucans (Ramphastidae) at two altitudes in the
Atlantic Forest of Brasil. Biotropica, 32, 842–850.
Galetti M., Bovendorp R.S., Guevara R. (2015) Defau-
nation of large mammals leads to an increase in seed
predation in the Atlantic forests. Global Ecology and
Conservation, 3, 824–830.
Gibson J.P., Wheelwright N.T. (1996) Mating system
dynamics of Ocotea tenera (Lauraceae), a gynodioe-
cious tropical tree. American Journal of Botany, 83,
890–894.
Goudet J. (2002) FSTAT, a program to estimate and test
gene diversities and fixation indices (version 2.9.3.2).
Grogan J., Loveless M.D. (2013) Flowering phenology
and its implications for management of big-leaf
mahogany Swietenia macrophylla in Brazilian Ama-
zonia. American Journal of Botany, 100, 2293–2305.
Herrera C.M. (1987) Components of pollinator “qual-
ity”: comparative analysis of a diverse insect assem-
blage. Oikos, 50, 79–90.
Hesse M., Kubitzki K. (1982) The sporoderm ultra-
structure in Persea, Nectandra, Hernandia, Gomor-
tega and some other Lauralean genera. Plant
Systematics and Evolution, 141, 299–311.
Hmeljevski K.V., Reis A., Montagna T., Reis M.S.
(2011) Genetic diversity, genetic drift and mixed
mating system in small subpopulations of Dyckia ibi-
ramensis, a rare endemic bromeliad from Southern
Brazil. Conservation Genetics, 12, 761–769.
Janzen D.H. (1979) How to be a fig. Annual Review of
Ecology and Systematics, 10, 13–51.
Klein R.M. (1980) Ecologia da flora e vegetac�~ao do
Vale do Itaja�ı. Sellowia, 32, 164–369.
Kubitzki K., Kurz H. (1984) Synchronized dichogamy
and dioecy in Neotropical Lauraceae. Plant System-
atics and Evolution, 147, 253–266.
Larson B.M.H., Kevan P.G., Inouye D.W. (2001) Flies
and flowers: taxonomic diversity of anthophiles and
pollinators. The Canadian Entomologist, 133, 439–465.
Li Y., Meissle M., Romeis J. (2008) Consumption of Bt
maize pollen expressing cry1 Ab or Cry3Bb1 does
not harm adult green lacewings, Chrysoperla carnea.
PLoS One, 3, e2909.
Lingner D.V., Schorn L.A., Vibrans A.C., Meyer L.,
Sevegnani L., Gasper A.L., De Sobral M.G., Kr€uger
A., Klemz G., Schmitt R., Junior C.A. (2013) Fitosso-
ciologia do componente arb�oreo/arbustivo da Flor-
esta Ombr�ofila Densa em Santa Catarina. In:
Vibrans A. C., Sevegnani L., Lingner D. V., De Gas-
per A. L. (Eds), Invent�ario flor�ıstico florestal de Santa
Catarina, Floresta Ombr�ofila Densa. Edifurb, Blume-
nau, Brazil, pp 159–248.
Mariot A., Mantovani A., Bittencourt R., Reis M.S.
(2014) Aspectos da biologia reprodutiva de Drimys
brasiliensisMiers (Winteraceae) em Floresta Ombr�ofila
Mista, Sul do Brasil. Ciencia Florestal, 24, 877–888.
MMA. (2008) Plano de Manejo da Floresta Nacional de
Ibirama – Informac�~oes Gerais. Available from http://
www.icmbio.gov.br/portal/images/stories/imgs-uni
dades-coservacao/flona_ibiramaaa.pdf%3E (acessed
19 February 2018).
MMA. (2014) Portaria Minist�erio do M�edio Ambiente
no 443, de 17 de dezembro de 2014. Lista Nacional
Oficial de Esp�ecies da Flora Ameac�adas de Extinc�~ao.Available from http://www.mma.gov.br/estruturas/
ascom_boletins/_arquivos/83_19092008034949.pdf
(acessed 15 January 2018).
Montagna T., Lauterjung M.B., Candido-Ribeiro R., da
Silva J.Z., Hoeltgebaum M.P., da Costa N.C.F., Ber-
nardi A.P., Reis M.S. (2018) Spatial genetic struc-
ture, population dynamics and spatial patterns in
the distribution of Ocotea catharinensis Mez. from
southern Brazil: implications for conservation.
Canadian Journal of Forest Research, 48, 506–516.
de Moraes P.L.R., Monteiro R. (2002) Taxas de cruza-
mento em uma populac�~ao natural de Cryptocarya
moschata Nees (Lauraceae). Biota Neotropica, 2, 1–10.
Nazareno A.G., dos Reis M.S. (2012) Linking phenol-
ogy to mating system: exploring the reproductive
biology of the threatened palm species Butia erios-
patha. Journal of Heredity, 103, 842–852.
Newstrom L.E., Frankie G.W., Baker H.G. (1994) A
new classification for plant phenology based on
flowering patterns in lowland tropical rain forest
trees at La Selva, Costa Rica. Biotropica, 26, 141–159.
Newton A.C. (2007) Reproductive ecology and genetic
variation. In: Newton A. C. (Ed), Forest ecology and
conservation: a handbook of techniques. Oxford
University Press, Oxford, UK, pp 235–284.
N�obrega S.R., Fortunato M.E.M., Quirino Z.G.M.
(2016) Biologia reprodutiva, visitantes florais e dis-
pers~ao de Ocotea duckei Vattimo-Gil (Lauraceae) na
Reserva Biol�ogica Guaribas, Para�ıba, Brasil. Gaia Sci-
entia, 10, 252–261.
Ollerton J., Liede S. (1997) Pollination systems in the
Asclepiadaceae: a survey and preliminary analysis.
Biological Journal of the Linnean Society, 62, 593–610.
Pacini E., Nepi M. (2007) Nectar production and pre-
sentation. In: Nicolson S.W., Nepi M., Pacini E.
(Eds), Nectaries and nectar. Springer, Dordrecht, the
Netherlands, pp 167–214.
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands934
Reproductive ecology of Ocotea catharinensis Montagna, da Silva, Pickart & dos Reis

Pacini E., Sciannandrone N., Nepi M. (2014) Floral
biology of the dioecious species Laurus nobilis L.
(Lauraceae). Flora: Morphology Distribution, Func-
tional Ecology of Plants, 209, 153–163.
Percival M. (1965) Floral biology. Pergamon Press,
Oxford, UK.
Pyke G.H. (1991) What does it cost a plant to produce
floral nectar? Nature, 350, 58–59.
Rader R., Edwards W., Westcott D.A., Cunningham
S.A., Howlett B.G. (2011) Pollen transport differs
among bees and flies in a human-modified land-
scape. Diversity and Distributions, 17, 519–529.
Reitz R., Klein R.M.R.M., Reis A. (1978) Projeto
madeira de Santa Catarina. Sellowia, 28–30, 11–
320.
Ribeiro M.C., Metzger J.P., Martensen A.C., Ponzoni
F.J., Hirota M.M. (2009) The Brazilian Atlantic For-
est: how much is left, and how is the remaining for-
est distributed? Implications for conservation.
Biological Conservation, 142, 1141–1153.
Ritland K. (1989) Correlated matings in the partial
selfer Mimulus guttatus. Evolution, 43, 848–859.
Ritland K. (2002) Extensions of models for the estima-
tion of mating systems using n independent loci.
Heredity, 88, 221–228.
Ritland K., Jain S. (1981) A model for the estimation of
outcrossing rate and gene frequencies using n inde-
pendent loci. Heredity, 47, 32–52.
Rogalski J.M., Reis A., Rogalski M., Montagna T., Reis
M.S. (2017) Mating system and genetic structure
across all known populations of Dyckia brevifolia: a
clonal, endemic, and endangered rheophyte brome-
liad. Journal of Heredity, 108, 299–307.
Saiter F.Z., Thomaz L.D. (2014) Revis~ao da lista de
esp�ecies arb�oreas do invent�ario de Thomaz & Mon-
teiro (1997) na Estac�~ao Biol�ogica de Santa L�ucia: o
mais importante estudo fitossociol�ogico em florestas
montanas do Esp�ırito Santo. Boletim do Museu de
Biologia Mello Leit~ao, 34, 101–128.
Schmid R. (1988) Reproductive versus extra-reproduc-
tive nectaries-historical perspective and terminological
recommendations. Botanical Review, 54, 179–227.
Schorn L.A., Gasper A.L., Meyer L., Vibrans A.C.
(2012) S�ıntese da estrutura dos remanescentes flo-
restais em Santa Catarina. In: Vibrans A. C., Seveg-
nani L., Gasper A. L., Lingner D. V. (Eds), Invent�ario
flor�ıstico florestal de Santa Catarina, Diversidade e
Coinservac�~ao dos remanescentes florestais. Edifurb,
Blumenau, pp 125–142.
Sebbenn A.M. (2002) N�umero de �arvores matrizes e
conceitos gen�eticos na coleta de sementes para reflo-
restamentos com esp�ecies nativas. Revista do Insti-
tuto Florestal, 14, 115–132.
Silva A., Aguiar I.B., De Schoffel E.R. (2000) Fenologia
reprodutiva de Canela-Preta (Ocotea catharinensis Mez
- Lauraceae) no Parque Estadual da Cantareira, S~ao
Paulo (SP). Revista do Instituto Florestal, 12, 77–88.
Souza L.A., Moscheta I.S. (1999) Morfo-anatomia da
flor de Ocotea puberula (Rich.) Nees (Lauraceae).
Acta Scientiarum, 21, 343–348.
Tarazi R., Mantovani A., Reis M.S. (2010) Fine-scale
spatial genetic structure and allozymic diversity in
natural populations of Ocotea catharinensis Mez.
(Lauraceae). Conservation Genetics, 11, 965–976.
Tarazi R., Sebbenn A.M., Kageyama P.Y., Vencovsky R.
(2013) Edge effects enhance selfing and seed harvesting
efforts in the insect-pollinated Neotropical tree Copai-
fera langsdorffii (Fabaceae). Heredity, 110, 578–585.
Varty N., Guadagnin D.L. (1998) Ocotea catharinensis.
In: The IUCN Red List of Threatened Species 1998:
e.T33982A9819827. Available from https://doi.org/
10.2305/iucn.uk.1998.rlts.t33982a9819827.en (ac-
cessed 29 January 2018).
Veloso H.P., Klein R.M. (1959) As comunidades e
associac�~oes vegetais da mata pluvial atlantica do sul
do Brasil. II. Dinamismo e fidelidade das esp�ecies
em associac�~oes do munic�ıpio de Brusque, SC. Sel-
lowia, 10, 9–124.
Ward M., Dick C.W., Gribel R., Lowe A.J. (2005) To
self, or not to self.. a review of outcrossing and pol-
len-mediated gene flow in neotropical trees. Hered-
ity, 95, 246–254.
Weir B.S., Cockerham C.C. (1984) Estimating F-Statis-
tics for the analysis of population structure. Evolu-
tion, 38, 1358–1370.
Wheelwright N.T. (1986) A seven-year study of individual
variation in fruit production in tropical bird-dispersed
tree species in the family Lauraceae. In: Estrada A.,
Fleming T.H. (Eds), Frugivores and seed dispersal. Junk
Publishers, Dordrecht, the Netherlands, pp 19–35.
Wolff D. (2006) Nectar sugar composition and volumes
of 47 species of Gentianales from a southern Ecuado-
rian montane forest. Annals of Botany, 97, 767–777.
Wratten S.D., Bowie M.H., Hickman J.M., Evans A.M.,
Sedcole J.R., Tylianakis J.M. (2003) Field boundaries
as barriers to movement of hover flies (Diptera: Syr-
phidae) in cultivated land. Oecologia, 134, 605–611.
Wright S. (1931) Evolution in mendelian populations.
Genetics, 16, 97–159.
Plant Biology 20 (2018) 926–935 © 2018 German Society for Plant Sciences and The Royal Botanical Society of the Netherlands 935
Montagna, da Silva, Pickart & dos Reis Reproductive ecology of Ocotea catharinensis